4 minute read
Viral Structures


Some suggest that virocells must have predated the current modern cells because they can infect both prokaryotes and eukaryotes. The current data indicate that viruses originated back when cells were so primitive that they lacked ribosomes as well. Without ribosomes, the ability to reproduce independently was impossible as we know it. Capsid proteins were probably acquired after the genetic material was created, indicating that the first genomes were possibly intracellular.
The fact that viruses are parasitic may have also been a late finding in these organisms. For this to be true, the ancient viruses must have had more genes in their genome and must have gradually lost some of these genes that necessitated their parasitic nature. In that sense, the reduction of genomes might have been a driving force in evolution rather than genome expansion. Genome reduction probably started in viruses but also occurred in other organisms later on. The most primordial viruses may have once been able to have their own metabolism.
Advertisement
VIRAL STRUCTURES
As mentioned, all viruses have a genome and a protein capsid, while some have a lipid envelope. In their totality, each viral particle is called a virion. Those virions without a lipid envelope are called naked viruses.
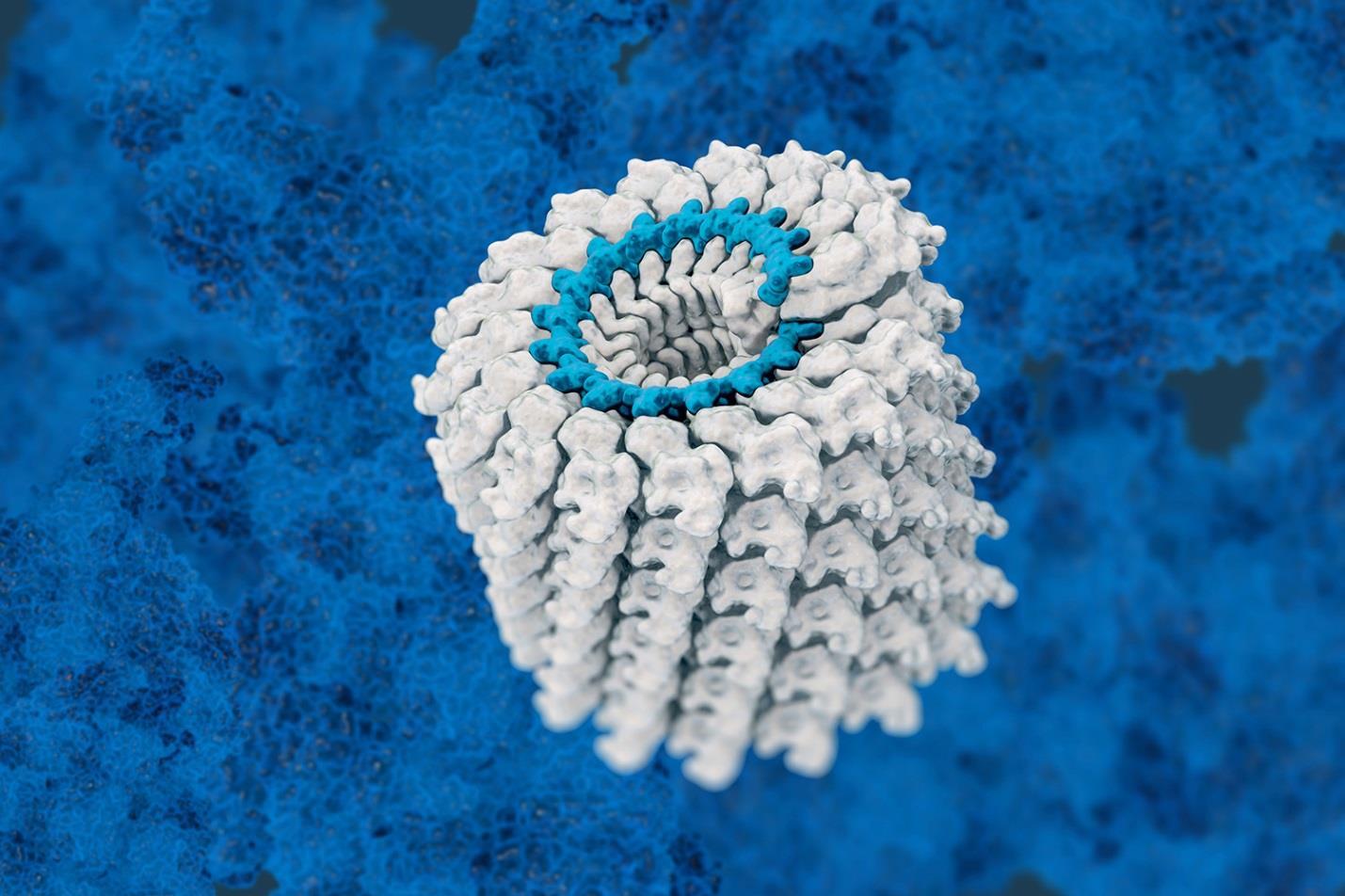
The protein capsid has the capability of recognizing and surrounding just viral DNA. It is suggested that there is a packaging sequence or signal on the viral genome that indicates the necessity of capsids to surround them. Capsids are made of repeating capsomeres that, in most non-phage viruses, are either helical or icosahedral. Each capsomere is usually but not always a single polypeptide chain, also called a structural subunit or protomer.
The first helical capsid to be studied is that of the tobacco mosaic virus, which is shown in figure 1:
Figure 1
This virus has single-stranded RNA with pieces of RNA fitting into one groove of each protein subunit. Some other viruses have more complex helical capsids that involve more than one polypeptide subunit. Helices have two parameters, including an amplitude and pitch. Pitch is the distance covered by each turn of this helix. Many of these viruses are very stable but can come together or dissociate, depending on the environmental circumstances. There are no real covalent bonds between these subunits so they can easily dissociate. Some animal viruses also have helical nucleocapsids and all are enveloped viruses, including the human influenza virus and rabies.
Icosahedral capsids are also made of subunits that form structures that appear to be spherical but are not truly spherical. The genome is contained within this icosahedral structure. One icosahedron has twenty identical equilateral triangle faces shaped into a roughly spherical shape. Figure 2 shows the icosahedral virus shape:

Figure 2.
Because proteins are not shaped like equilateral triangles, each face has more than one strand of polypeptide. The simplest icosahedral virus has three protein subunits per face so there are sixty separate polypeptides per viral particle. Because most viruses have larger genomes than this can support, most of these viruses have some multiple of 60 as their total polypeptide molecule number.
The viral envelope is made from a lipid bilayer, which actually comes from lipids derived from the host cells themselves. It will also contain proteins coded for by the virus themselves. Most of these are trans-membrane glycoproteins. These serve to bind with host receptors and play a role in cell entry and things like membrane fusion or channel formation in the virus membrane itself. Many of these envelopes contain matrix or internal proteins that tack the nucleocapsid to the envelope itself. These are not capsulated. There are also transcription factors and enzymes in low amounts in the viral envelope.
The basis behind enveloped viruses is that they essentially bud out of the cell membranes of the cells they occupy. Rather than causing the cell itself to burst, these viruses take a piece of the cell membrane with them as they leave the cell but it is also
possible that the membrane around them comes from the nucleus, Golgi apparatus, or endoplasmic reticulum. The glycoproteins that will make up the outside of the envelope are largely clustered in one region so that as many of these get onto the envelope during budding.
There are within these structure types many different and diverse morphologies seen. The size variations among viruses are great, with a size range of between 20 nanometers and 300 nanometers, although there are filoviruses that have a length of up to 1400 nanometers but a width of only 80 nanometers. Figure 3 shows a typical filovirus, such as the Ebola virus:

Figure 3.
Scanning electron microscopes are necessary for the visualization of viruses. These are coated with tungsten metal salts that will allow visualization through the scattering of electrons. Some are treated with positive staining that highlights the viral particle or negative staining that stains the background only.
Bacteriophages infect bacteria and these have unique shapes with a head that contains the nucleic acid and a tail plus added structures that allow the nucleic acid alone to be injected directly into the cell itself. Figure 4 shows the basic structure of a bacteriophage:

Figure 4.
There are other complex viruses that do not have the traditional shapes. For example, the poxviruses are large and complex with a central nucleoid consisting of proteins and two lateral bodies that have no known function. It can range in shape from being ovoid to brick-shaped. Mimiviruses are quite large and have protein filamentous structures coming out of their surface. It is large enough to be seen under a light microscope. Figure 5 shows what a mimivirus looks like:


