

Chuyên đề
TÍCH HỢP DI TRUYỀN HOÁ SINH VI SINH VẬT
Đỗ Thuỳ Dung, Trần Thị Loan - Trường THPT Chuyên Lào Cai
Chuyên đề đạt giải Nhất
1. Lí do tiếp cận chuyên đề
Vi sinh vật không phải là một khái niệm phân loại, các vi sinh vật có thể thuộc những nhóm phân loại rất xa nhau. Chúng thường có một số đặc điểm chung như có kích thước hiển vi, cơ thể đơn bào, sinh trưởng nhanh, phân bố rộng, thích ứng cao với môi trường. Thuộc nhóm Vi sinh vật có vi khuẩn (thuộc Giới Vi khuẩn), động vật nguyên sinh và vi tảo (thuộc Giới Nguyên sinh) và Vi nấm (thuộc Giới Nấm), vi sinh vật cổ (thuộc lãnh Giới Vi sinh cổ theo quan điểm phân chia 3 lãnh Giới). Ngoài ra, người ta còn xếp virut vào nhóm Vi sinh vật mặc dù hiện nay virut không được xem là cơ thể sống vì chúng không có cấu tạo tế bào và có đời sống kí sinh bắt buộc trong tế bào. Virut không tồn tại và sống trong môi trường tự nhiên khi ở ngoài tế bào. Vi sinh vật có vai trò quan trọng đối với hệ sinh thái cũng như đời sống con người.
Việc nghiên cứu di truyền học trên các đối tượng vi sinh vật chưa được tiến hành cho mãi đến những năm 40. Tuy ra đời chậm nhưng di truyền học vi sinh vật đã đóng vai trò “cách mạng hoá” tạo ra những bước tiến nhảy vọt cho di truyền học nói riêng và sinh học nói chung về cả phương diện lí thuyết và ứng dụng thực tiễn. Các nghiên cứu di truyền được tiến hành một mặt trên các vi sinh vật nhân thực như nấm men, nấm mốc, vi tảo có sinh sản hữu tính, mặt khác trên các vi khuẩn và virut. Di truyền học của virut và vi khuẩn đã góp phần đáng kể cho sự ra đời của kĩ thuật tái tổ hợp DNA dẫn đến kĩ thuật di truyền làm bùng nổ Công nghệ sinh học.
Hoá sinh học là ngành khoa học nghiên cứu đến cấu trúc và quá trình hoá học diễn ra trong cơ thể sinh vật. Đây là nội dung có liên quan chặt chẽ với các kiến thức hoá học, sinh học, mà phần lớn là sinh học tế bào, sinh học phân tử và di truyền. Kiến thức phần hoá sinh học có ý nghĩa tiên quyết tạo nền tảng đi sâu vào giải quyết các vấn đề đặc biệt quan trọng trong y dược và công nghệ sinh học. Hoá sinh học được chia thành hai lĩnh vực chính: Hoá sinh tĩnh và hoá sinh động. Hoá sinh tĩnh nghiên cứu các vấn đề thuộc về enzyme và các hợp chất hữu cơ. Tế bào chính là đơn vị nhỏ nhất của sự sống - nơi diễn ra hoạt động hoá sinh to lớn được gọi là sự trao đổi chất. Đây là quá trình thay đổi hoá học và vật lí diễn ra liên tục trong sinh vật sống bao gồm quá trình xây dựng mô mới, thay thế mô cũ, chuyển đổi thức ăn thành năng lượng, xử lý chất thải, sinh sản, ...Để quá trình trao đổi chất được diễn ra thuận lợi thì vai trò của enzyme là không thể thiếu. Sự hoạt động của hợp chất có hoạt tính sinh học cao này giúp cho các phản ứng diễn ra trong quá trình trao đổi chất diễn ra dễ dàng và thuận lợi hơn, tạo thành một hệ thống hoàn chỉnh. Các phản ứng hoá học diễn ra trong tế bào có sự tham gia của các hợp chất hữu cơ: protein, cacbonhidrat, lipid và axit nucleic, ... Một khía cạnh khác của hoá sinh học là nghiên cứu về tính chất và các đặc tính sinh học của các đại phân tử đó. Đó là xác định các thành phần tạo nên những cấu trúc lơn hơn phân tử,
mô tả cách lắp ráp, quan sát không gian ba chiều và mô tả vai trò của chúng trong sự sống sinh vật. Hoá sinh động nghiên cứu các quá trình trao đổi saccarit, lipit, tổng hợp DNA, RNA, protein. Một trong những dấu hiệu đặc trưng cơ bản của sinh vật sống là sự trao đổi chất và năng lượng. Như đã nói ở trên, hoá sinh động sẽ xem xét cách thức các tế bào sản xuất ra năng lượng từ các hợp chất hữu cơ. Thông qua nguyên tắc của sự biến đổi năng lượng trong tế bào, các con đường dị hoá phân tử glucose sẽ được mô tả trong sự có mặt hoặc vắng mặt oxi phân tử. Cùng với đó, vai trò của lipit trong cung cấp và dự trữ năng lượng cho sinh vật cũng được làm sáng tỏ, thông qua đó xây dựng mối quan hệ trong con đường trao đổi chất giữa lipit và saccarit. Trong tế bào, quá trình tổng hợp DNA, RNA, protein là yếu tố cơ bản cấu thành nên toàn hệ sinh vật. Hoá sinh động cũng đề cập đến những nét cơ bản của nhân đôi DNA, phiên mã, dịch mã.
Nhân loại đang sống ở thời kì Công nghệ sinh học. Di truyền học đi sâu vào các vấn đề cơ bản của sự tồn tại và lưu truyền sự sống nên nó giữ một vị trí đặc biệt quan trọng, có người ví "Di truyền học là trái tim của Sinh học", vì không ít thì nhiều, nó liên quan và chi phối bất kì lĩnh vực nào của sinh học, từ các cơ chế phân tử của sự sống cho đến sự tiến hóa của toàn bộ thế giới sinh vật trên hành tinh của chúng ta. Những phát minh lớn với số lượng tăng vọt của di truyền học đã có tác dụng cách mạng hóa sinh học, biến sinh học từ mô tả thành thực nghiệm chính xác. Hội nghị Di truyền học thế giới ở Toronto (Canada) năm 1988 đã nêu phương châm "Di truyền học hóa" Sinh học. Các đối tượng vi sinh vật đã góp phần chủ yếu vào các phát minh nền tảng không những của di truyền học phân tử, mà của sinh học phân tử và sinh học hiện đại nói chung.
Đối với các trường THPT Chuyên, việc lựa chọn, phát hiện và bồi dưỡng học sinh giỏi, học sinh năng khiếu là một trong những nhiệm vụ then chốt. Với nguồn học liệu dồi dào và mở như hiện nay, học sinh khối lớp Chuyên Sinh có thể tiếp cận được nguồn tri thức nhanh chóng và hiệu quả, tuy nhiên việc lựa chọn những nội dung kiến thức trọng tâm, có tính chất hệ thống làm cơ sở nền tảng phát huy khả năng tự học, tự đọc của học sinh vẫn đang là vấn đề cần được quan tâm đầu tư.
Qua quá trình tự nghiên cứu, tôi tự nhận thấy mức độ tích hợp các nội dung kiến thức Sinh học trong các câu hỏi ở đề thi học sinh giỏi các cấp, đặc biệt là kì thi học sinh giỏi Quốc gia ngày càng tăng lên về số lượng và độ “khó”. Vì vậy, với mục đích bổ sung thêm nguồn học liệu có tính chất hệ thống, định hướng cho học sinh cách thức tiếp cận và tư duy logic đối với kiến thức phần Hoá sinh, vi sinh và di truyền, tôi lựa chọn và tiếp cận chuyên đề “Tích hợp di truyền hoá sinh vi sinh vật”
2. Mục đích của chuyên đề
- Hệ thống hoá nội dung kiến thức phần hoá sinh, vi sinh và di truyền trên phương diện tiếp cận “sự tích hợp”.
- Biên soạn chuyên đề để nâng cao hiệu quả tự học của cá nhân, cung cấp thêm nguồn tài liệu cho học sinh trong đội tuyển.
- Rèn luyện khả năng tự học, tự đọc tài liệu và khai thác kiến thức.
- Hướng dẫn học sinh cách thức phân tích nội hàm trong các câu hỏi, tránh trả lời không đúng trọng tâm.
3. Khái quát nội dung chuyên đề
Chuyên đề được biên soạn thành 2 phần: Phần A: Cơ sở lý thuyết và phần B:
Câu hỏi, bài tập vận dụng.
Phần Cơ sở lí thuyết chuyên đề nghiên cứu các nội dung phần hoá sinh tế bào vi sinh vật và hiện tượng di truyền ở vi sinh vật.
Phần Câu hỏi, bài tập vận dụng sưu tầm chủ yếu trong các đề thi học sinh giỏi Quốc gia các năm, hướng dẫn phân tích kiến thức lí thuyết và trả lời.
NỘI DUNG
PHẦN A: CƠ SỞ LÝ THUYẾT
I. THÀNH PHẦN HOÁ SINH TẾ BÀO VI SINH VẬT
1. Enzyme
1.1. Cấu trúc enzyme
Enzyme có bản chất là protein (một số ít là RNA - ribozim), có tác dụng xúc tác, làm tăng tốc độ phản ứng một cách đặc hiệu. Enzyme có mặt trong tất cả các hoạt động sống của tế bào, có đầy đủ đặc tính lý hoá của một chất xúc tác.
- Tính đặc hiệu cao: enzyme thường đặc trưng cho một phản ứng nhất định. Ví dụ enzyme ureaza chỉ xúc tác phản ứng phân giải ure. Một số enzyme khác cũng có tính đặc hiệu tương đối, chẳng hạn enzyme lipase có khả năng phân huỷ nhiều loại este khác nhau.
- Hiệu suất xúc tác lớn: tốc độ phản ứng có thể tăng lên 105 - 107 lần so với khi không có chất xúc tác. Ví dụ enzyme catalase xúc tác phản ứng phân huỷ H2O2 -> H2O + O2 chỉ trong một giây nhưng với chất xúc tác là Fe thì cần đến 300 năm.
- Tác dụng xúc tác ở điều kiện bình thường: pH môi trường gần trung tính, áp suất và nhiệt độ bình thường.
Enzyme được chia thành 6 lớp dựa vào kiểu xúc tác (theo thống nhất của Hội hoá sinh học quốc tế năm 1961).
A - Oxidoreductase: xúc tác cho phản ứng oxi hoá khử (vận chuyển điện tử, các ion hydride hoặc nguyên tử hidro)
B - Transferase: xúc tác cho phản ứng chuyển vị các nhóm từ một phân tử này đến một phân tử khác.
C- Hydrolase: Xúc tác các phản ứng thuỷ phân (cũng là phản ứng vận chuyển các nhóm chức năng)
D - Lyase: xúc tác cho các phản ứng thêm các nhóm vào nối đôi, hoặc tạo thành nối đôi bằng cách loại các nhóm.
E - Isomerase: xúc tác cho các phản ứng đồng phân hoá, hoặc chuyển vị các nhóm trong nội bộ phân tử thành các dạng đồng phân.
F - Ligase: xúc tác cho các phản ứng tạo thành liên kết C-C, C-O, C-S và C-N bằng phản ứng ngưng tụ, kèm theo phản ứng cắt đứt các liên kết giàu năng lượng của các nucleoside triphotphate, thường dấu hiệu nhận biết của kiểu enzyme này với lyase có sự xuất hiện của ATP.
Cấu trúc enzyme: Enzyme là các đại phân tử có khối lượng phân tử từ 12000 đến hàng triệu đơn vị Dalton (Da). Cũng như các protein, về thành phần cấu tạo, enzyme chia thành 2 loại: enzyme một thành phần và enzyme hai thành phần. Enzyme
một thành phần: chỉ có một thành phần duy nhất là protein như enzyme pepsin của dạ dày. Enzyme hai thành phần (haloenzyme) gồm 2 thành phần: phần protein gọi là apoenzyme, phần không phải protein là cofactor (hoặc coenzyme). Hầu hết phần ngoại protein có chức năng xúc tác apoenzyme. Coenzyme là các dẫn xuất của vitamin hoà
tan trong nước (ví dụ NAD+) vì vậy khi thiếu một vitamin nào đó, gây nên những bệnh
đặc trưng (chẳng hạn như bệnh pellagra, Scorbut, ...). Cofactor là các ion kim loại với hàm lượng nhỏ (các nguyên tố vi lượng như Cu2+ có trong cytocrom oxidase, carbonic anhydrase, alcohol dehydrogenaza chứa Zn2+, hexokinase, glucose - 6 - phosphatease, pyruvate kinase chứa Mg2+, ....)
Trung tâm hoạt động là một vùng đặc biệt của enzyme có tác dụng gắn với cơ chất để xúc tác cho phản ứng biến đổi cơ chất thành sản phẩm. Mỗi enzyme có thể có một hoặc hai hay nhiều trung tâm hoạt động. Tại đây sẽ chứa những nhóm hoá học và những liên kết tiếp xúc trực tiếp với cơ chất hoặc không tiếp xúc trực tiếp nhưng có chức năng trực tiếp trong quá trình xúc tác.
Về thành phần cấu tạo: Trung tâm hoạt động của enzyme thường bao gồm các axit amin có các nhóm hoá học có hoạt tính cao như serine (có nhóm -OH), cystein (có nhóm SH), glutamate có nhóm y-COO), lysine (có nhóm S-NH3 +) histidine có nhóm imidazol+, tryptophan có nhóm indol+ , ... đây là những nhóm phân cực hoặc ion hoá, có khả năng tạo liên kết hidro hoặc ion với cơ chất.
Để mô tả sự liên kết giữa enzyme và cơ chất, đã có nhiều giả thuyết được đưa ra: Thuyết “mô hình cảm ứng không gian” được Koshland D đưa ra năm 1958 đã giải thích tính đặc hiệu tương đối của enzyme. Theo thuyết này, trung tâm hoạt động của enzyme có tính mềm dẻo và linh hoạt, có thể biến đổi về cấu hình không gian trong quá trình hoạt động tương tác với cơ chất sao cho phù hợp với cấu hình không gian của cơ chất, để có thể hình thành phức hợp enzyme cơ chất (ES).
Enzyme dị lập thể (Enzyme allosteric)
Đây là loại enzyme ngoài trung tâm hoạt động còn có một vài vị trí dị lập thể, trung tâm hoạt động tiếp nhận cơ chất để xúc tác cho phản ứng enzyme trong khi vị trí dị lập thể tiếp nhận yếu tố dị lập thể để điều chỉnh hoạt động xúc tác của enzyme. Phân tử enzyme dị lập thể có thể có thể có vị trí dị lập thể dương, vị trí lập thể âm hoặc chứa cả hai. Khi vị trí dị lập thể dương tiếp nhận yếu tố dị lập thể dương A (chất hoạt hóa) thì cấu hình enzyme thay đổi theo hướng có lợi hơn, enzyme được hoạt hóa, ái lực enzyme với cơ chất tăng lên, tăng tốc độ phản ứng. Khi vị trí dị lập thể âm tiếp nhận yếu tố dị lập thể âm (chất ức chế) thì cấu hình enzyme thay đổi theo hướng có hại, enzyme bị ức chế, ái lực enzyme với cơ chất giảm nên tốc độ phản ứng giảm đi.
hóa bởi yếu tố dị lập thể dương là ADP và AMP, nhưng bị
ứ
1.2. Cơ chế tác động
Enzyme thực hiện chức năng xúc tác làm tăng tốc độ phản ứng bằng cách giảm năng lượng hoạt hóa gồm 2 giai đoạn chính:
- Tạo thành phức ES.
- Phân ly phức ES giải phóng E ở trạng thái ban đầu, giải phóng sản phẩm.
Hình 1: Cơ chế tác động của enzyme

Khi có mặt cơ chất, cơ chất cảm ứng sự định hướng không gian của các nhóm chức trong trung tâm hoạt động của E, tạo nên sự định hướng tương ứng thích hợp giữa các nhóm chức trong TTHĐ của E và các nhóm chức của cơ chất, dọn đường cho S kết hợp vào TTHĐ của E, tạo thành phức ES, đồng thời loại các phân tử nước ra ngoài. Tương tác giữa các gốc amino acid của E với cơ chất, thay đổi đám mây electron, giảm năng lượng hoạt hóa cần thiết để đưa đến trạng thái chuyển tiếp.
Hình 2: Enzyme làm giảm năng lượng hoạt hoá
Phức ES sau khi tạo thành kém bền, chỉ tồn tại trong thời gian ngắn và nhanh chóng chuyển hóa thành sản phẩm và giải phóng E tự do, E lại quay vòng xúc tác như ban đầu.

Năng lượng hoạt hóa là sự sai khác giữa mức năng lượng tự do của trạng thái cơ bản (ground state) và trạng thái chuyển tiếp (transition state), được hình thành do sự tiếp xúc, và chạm giữa các phân tử làm tăng tốc độ phản ứng.
1.3. Các yếu tố ảnh hưởng đến vận tốc của enzyme


1.3.1. Nhiệt độ
Mỗi enzyme hoạt động trong một khoảng nhiệt độ nhất định, đa số các enzyme tối ưu ở nhiệt độ 40 - 45°C. Khi cao hơn nhiệt độ tối ưu, tốc độ phản ứng giảm nhanh vì nhiệt độ cao làm enzyme biến tính, trung tâm hoạt động mất đi cấu hình chuẩn và không còn phù hợp được với cơ chất, vai trò xúc tác bị ảnh hưởng. Cá biệt có những enzyme chịu được nhiệt độ cao như amylase trong công nghiệp dệt chịu được nhiệt độ hơn 100°C. Loài cá băng ở Nam cực có enzyme hoạt động hiệu quả ở -2°C.
1.3.2. Độ pH
Mỗi loại enzyme có một độ pH tối ưu, pH trên hay dưới ngưỡng tối ưu đều làm cho hoạt tính enzyme bị giảm hoặc mất hoạt tính. pH ảnh hưởng đến trạng thái ion hóa các gốc R của các gốc amino acid trong phân tử E, ion hóa các nhóm chức trong trung tâm hoạt động, ion hóa cơ chất, do đó ảnh hưởng đến hai tham số động học Km và V.
Hình 4: Mối liên hệ giữa tốc độ phản ứng và nồng độ cơ chất
1.3.4. Ảnh hưởng của chất kìm hãm
Chất kiềm hãm (Inhibitor = I) là những chất làm giảm tốc độ phản ứng do enzyme xúc tác. Các chất này có thể là ion kim loại, chất hữu cơ hay vô cơ. Các chất kìm hãm có thể chia theo nhiều cách khác nhau: kìm hãm thuận nghịch hay kìm hãm không thuận nghịch.
a. Kìm hãm thuận nghịch
Kìm hãm thuận nghịch là kìm hãm enzyme bằng cách kết hợp với enzyme và tác động đến sự kết hợp của E với S.
* Kìm hãm cạnh tranh
Kìm hãm cạnh tranh: Chất ức chế liên kết vào trung tâm hoạt động của enzyme (cạnh tranh với cơ chất). Nhận biết: Km tăng (ái lực giảm) và Vmax không đổi.
* Kìm hãm không cạnh tranh
Hình 3: Ảnh hưởng của độ pH đến hoạt tính enzyme
1.3.3. Nồng độ cơ chất
Cơ chất là chất mà enzyme xúc tác. Sự thay đổi nồng độ cơ chất khác nhau sẽ ảnh hưởng rất lớn đến hoạt độ của enzyme. Khi nồng độ cơ chất thấp, nhiều phân tử enzyme có trung tâm hoạt động tự do và sự cung cấp hạn chế có chất sẽ xác định tốc độ phản ứng. Ngược lại nồng độ cơ chất cao, hầu hết các trung tâm hoạt động bị chiếm lĩnh do đó lúc này số lượng phân tử enzyme lại là yếu tố quyết định phản ứng. Trong hoạt động trao đổi chất của tế bào mối tương quan này có tầm quan trọng như những phương thức kiểm soát tốc độ phản ứng khác nhau.
- Kìm hãm không cạnh tranh: Chất ức chế liên kết phức hợp enzyme có chất (không phải enzyme tự do) ở vị trí khác trung tâm hoạt động ảnh hưởng đến đến trung tâm hoạt động dẫn đến giảm hoạt tính xúc tác của enzyme. Nhận biết Km không thay đổi và Vmax giảm.
* Kìm hãm hỗn hợp
- Kìm hãm kiểu hỗn hợp: Chất ức chế đồng thời liên kết được vào cả tâm hoạt động và vị trí khác (enzyme tự do và phức hợp enzyme - cơ chất). Nhận biết: đồng thời Km tăng (hoặc ái lực giảm) và Vmax giảm.
b. Các chất kìm hãm không thuận nghịch
Các không thuận nghịch kết hợp với E tạo thành phức EI không hoạt động, không phân ly, hoặc rất khó phân ly, hoặc phân ly với vận tốc không đáng kể. Người ta ứng dụng các chất kìm hãm không thuận nghịch để nghiên cứu cơ chế phản ứng của enzyme.
1.4. Điều hoà hoạt độ của enzyme
Hoạt độ của enzyme, cũng như hoạt độ của các protein có hoạt tính sinh học khác trong tế bào. Chúng phải được điều hòa nghiêm ngặt, tiến hành đúng chỗ, đúng lúc và với mức vừa đủ theo yêu cầu của cơ thể
Hoạt độ của E được điều hòa cả về số lượng và hoạt độ phân tử. Gọi là điều hòa hoạt độ phân tử có nghĩa là số lượng phân tử protein E được tổng hợp là không thay đổi. Hoạt độ của enzyme có thể bị thay đổi bằng cách kết hợp với chất điều hòa (điều hòa allosteric); hoặc cải biến cộng hóa trị thuận nghịch; hoặc thay đổi tỷ lệ giữa các dạng phân tử enzyme có mức độ hoạt động khác nhau, ...
1.4.1. Điều hòa Allosteric
Kiểu điều hòa này hoạt động dựa trên nhu cầu của tế bào. Khi lượng sản phẩm cuối cùng của chu trình đáp ứng đủ cho tế bào, nó sẽ quay ngược trở lại kìm hãm hoạt độ enzyme.
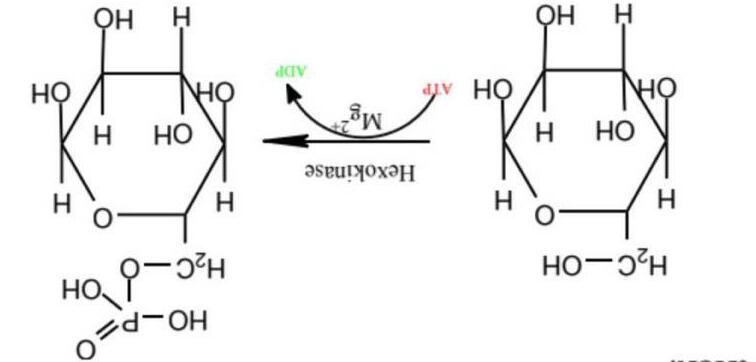
Hình 6: Enzyme hexokinase xúc tác phản ứng
Enzyme hexokinase xúc tác cho phản ứng trên gồm 4 isoenzyme (kí hiệu I, II, III và IV). Khi nồng độ glucose trong mức bình thường, các isoenzyme III hoạt động với tốc độ cực đại, còn IV chỉ làm việc 25%.
1.4. 3. Cải biến cộng hóa trị
Chủ yếu hoạt động cải biến hóa trị là kết hợp thuận nghịch với nhóm phosphate. Với các đặc điểm phổ biến:
- Phản ứng chuyển nhóm phosphoryl từ ATP đến enzyme do protein kinase xúc tác còn phản ứng tách phosphate khỏi enzyme do protein phosphatase xúc tác.
- Các gốc amino acid của enzyme kết hợp với nhóm phosphoryl là: Tyr, Ser, Thr, His.
- Việc kết hợp với nhóm phosphate có thể chuyển enzyme từ dạng có hoạt động thấp thành dạng có dạng hoạt động cao hay ngược lại. Ví dụ: Cơ chế điều hòa của glycogen phosphorylase. Protein kinase (PKA) có vai trò điều hòa hoạt độ enzyme bằng cách phosphoryl hóa. Với cách điều hòa này, tín hiệu điều hòa có thể khuếch đại nhiều lần vì protein kinase có thể tác dụng lên nhiều phân tử mục tiêu khác nhau.
2. Các hợp chất hữu cơ
2.1. Saccharide
Hình 5: Mô hình điều hòa allosteric dưới tác dụng của sản phẩm cuối cùng khi nồng độ các chất này đã đáp ứng nhu cầu.
1.4. 2. Điều hòa tỷ lệ các dạng khác nhau của enzyme có hoạt độ khác nhau
Isozyme (hay còn gọi là isoenzyme) là các enzyme khác nhau trong chuỗi amino acid nhưng xúc tác cho phản ứng hóa học tương tự. Các enzyme này thường hiển thị các thông số động học khác nhau (ví dụ các giá trị K và V khác nhau) hoặc các đặc tính điều chỉnh khác nhau. Sự tồn tại của isozyme cho phép tinh chỉnh sự trao đổi chất để đáp ứng nhu cầu cụ thể của một mô hoặc giai đoạn phát triển.
Saccharide là một chất hữu cơ chứa 3 nguyên tử là carbon (C), oxy (O) và hydro (H). Các đơn vị cơ bản của saccaride được gọi là monosaccharide, ví dụ như glucose, galactose, và fructose. Công thức thu gọn tổng quát của một monosaccaride là (CH2O)n với n lớn hơn hoặc bằng 3; tuy nhiên, không phải tất cả saccharide đều tuân theo công thức này (như acid uronic), hoặc không phải tất cả các chất hóa học có công thức như trên thì được xếp vào nhóm saccharide. Monosaccharide có thể được liên kết với nhau để tạo thành polysaccharide (hay oligosaccharide) theo nhiều cách khác nhau.

Saccharide giữ nhiều vai trò quan trọng trong cơ thể sống như:
- Chúng tồn tại ở dạng dự trữ năng lượng, nhiên liệu và chất trao đổi trung gian (glucose, glycogen, ...).
- Một số loại đường như ribose và deoxyribose tạo thành một phần trong cấu trúc của RNA và DNA (vật chất di truyền của hầu hết sinh vật trên Trái Đất).
- Tham gia cấu tạo các thành phần cấu trúc của thành tế bào vi khuẩn và thực vật (cellulose).
- Saccharide liên kết với một số protein và lipid, các chất này có ý nghĩa trong việc tương tác gián tiếp giữa các tế bào và sự tương tác giữa các tế bào với các thành phần khác trong môi trường của tế bào (proteoglycan, glycoprotein, glycolipid, ...).
a. Đường đơn (Monosaccharide): là dẫn xuất của aldehyde, hoặc ketone và hai hay nhiều nhóm hydroxyl (-OH). Mỗi phân tử có từ 3 đến 7 cacbon và chia thành hai nhóm chính là đường aldose và ketose.
- Đường aldose: là đường chứa nhóm carbonyl là aldehyde (-CHO)
- Đường ketose: là đường chứa nhóm carbonyl là ketone (-CO-)
* Maltose:
- Cấu trúc từ một phân tử anpha-glucose (1->4) glucose
- Có tính khử nhưng yếu hơn so với glucose
- Đường chủ yếu là mạch nha, có ít ở thực vật bậc cao
- Bị thuỷ phân bởi maltase ở hoặc acid loãng nhiệt độ cao
* Lactose
- Cấu tạo từ beta-galactose (1-4) Beta-glucose.
- Đường chủ yếu của sữa, có một ít trong hoa
- Có tính khử
- Bị thuỷ phân bởi beta-galactoside
* Sucarose
- Cấu tạo từ anpha glucose (1->2) beta-fructose.
- Không có tính khử
- Có nhiều trong mía, củ cải đường
- Bị thuỷ phân bởi saccaraza, axit loãng
c. Đường đa (Polysaccharide)
Hình 7: Đường aldose và ketose
- Tính chất: vị ngọt, tan nhiều trong nước, có tính khử mạnh, tham gia các phản ứng oxi hoá-khử
- Vai trò:
+ Cung cấp năng lượng cho tế bào, ví dụ đường glucose.
+ Là nguyên liệu để tạo đường đôi, đường đa; tham gia cấu tạo các thành phần của tế bào, ví dụ đường pentose tham gia cấu tạo DNA, RNA.
+ Dùng trong các sản phẩm chữa bệnh như xylitol hay chẩn đoán bệnh như xác định lượng glucose trong máu.
b. Đường đôi (Disaccharide)
Disaccharide được cấu tạo từ hai monosaccharide, ví dụ như sucrose và lactose. Chúng liên kết với nhau bằng một liên kết cộng hóa trị tạo thành từ phản ứng khử nước (hydrolase), theo đó sẽ tách ra một nguyên tử hydro của một monosaccharide và một nhóm hydroxyl của monosaccharide còn lại. Mặc dù có nhiều disaccharide, chỉ một số ít disaccharide là được chú ý.

Polysaccharide là phân tử carbohydrate cao phân tử gồm chuỗi dài của đơn vị monosaccharide liên kết với nhau bằng liên kết glycosidic và thủy phân cung cấp cho các thành phần hoặc monosaccharide, oligosaccharide. Ví dụ polysaccharide lưu trữ như tinh bột và glycogen, và polysaccharide cấu trúc như cellulose và chitin.
Tinh bột Glicogen Xenlulozo Amilozo Amilopectin
Kích thước 50-500 Có thể lên đến 1 triệu
Liên kết Anpha (14) glicozit Anpha (1>4) glicozit Anpha (1>6) glicozit
Cấu trúc Không phân nhánh, xoắn kiểu lò xo, nhờ liên kết hidro giữa các
Phân nhánh ở khoảng 20-30 gốc
50000
15000
Anpha (1->4) glicozit Anpha (1->6) glicozit
Beta(1>4) glicozit
Phân nhánh nhiều hơn Không phân nhánh, xếp thành bó sợi, có nhiều liên kết
nhóm OH
Nơi tích luỹ chủ yếu Thực vật, mô dự trữ (hạt, củ) Tế bào gan
động vật, vi khuẩn
hidro giữa các
bó sợi
Thành tế bào
thực vật bậc cao, vỏ ngoài
cứng của một
số động vật không xương
sống
Enzyme thuỷ phân Amilaza Amilaza Xenlulaza
Chức năng Dự trữ năng lượng
2.2. Lipid
Dự trữ năng lượng
Bảng 1: Cấu trúc và chức năng polysaccharide
c. Chức năng
Cấu trúc
Lipid là hợp chất sinh học bao gồm các acid béo và các dẫn xuất của chúng có trong tự nhiên.
a. Phân loại dựa theo thành phần cấu tạo
- Lipid đơn giản: là ester của alcohol và acid béo – triacylglycerol, sáp, steride
- Lipid phức tạp: ngoài các thành phần của lipid đơn giản còn chứa các gốc phosphoric, choline, saccharide.
Ví dụ glycolipid, glycerolglycolipid....

b. Tính chất vật lí – hóa học
- Không tan trong nước nhưng tan trong dung môi hữu cơ như benzen, acetone,
…
- Mạch hydrocacbon càng dài, ít nối đôi, độ hòa tan trong nước càng kém, nhiệt độ nóng chảy càng cao do giữa chúng hình thành các tương tác Van der Waals làm bến cấu trúc ở mạch dài.
- Acid béo không no dạng cis thường bị uốn cong nên kém bền hơn so với dạng trans. Vì vậy khi dầu đã rán nhiều lần, lượng acid béo trans tăng cao làm cho lượng cholesterol tỉ trọng thấp (LDL) nhiều gây tăng nguy cơ tim mạch, làm yếu cấu trúc màng, dễ bị nhiễm khuẩn và Cis Trans hóa chất độc.
Hình 8: Cấu hình cis và trans của acid béo
- Thành phần cơ bản của màng sinh học (phospholipid và cholesterol tham gia cấu trúc nên màng sinh học của hầu hết sinh vật trên Trái Đất).
- Là nguồn nguyên liệu dự trữ quan trọng cung cấp năng lượng cho các hoạt động sống của cơ thể (acid béo và glycerol là nguồn năng lượng cũng như chất dự trữ quan trọng cho con người và động vật).
- Là chất mang điện tử, sắc tố hấp thụ ánh sáng, thành phần cấu tạo hormone, chất truyền tin nội bào, hay bảo vệ (glycoprotein tham gia nhận biết đính trên mặt ngoài màng sinh chất; các hormone steroid vận chuyển các chất đến toàn bộ cơ thể).
2.3. Acid nucleic
2.3.1. Nucleotit - đơn phân của axit nucleic
Mỗi nucleotit được cấu tạo từ 3 thành phần: một gốc base nitro gen, một gốc pentose và một gốc photphat. Nucleotit có vai trò tham gia vào cấu trúc nên các hợp chất sống hữu cơ (DNA, RNA), là thành phần cấu tạo của các coenzyme quan trọng như NAD+, cấu trúc nên phân tử ATP, ...
2.3.2. DNA
Mô hình cấu trúc
Theo J. Waston và F. Crick, phân tử DNA là chuỗi xoắn kép gồm hai mạch polinucleotit xoắn đều quanh một trục theo chiều từ trái sang phải như một thang dây xoắn, mà hai tay thang là các phân tử đường (C5H10O4) và axit photphoric sắp xếp xen kẽ nhau, còn mỗi bậc thang là một cặp bazo đối diện và liên kết với nhau bằng các liên kết hidro theo nguyên tắc bổ sung, nghĩa là một bazo lớn (A hoặc G) được bù bằng một base nhỏ (T hoặc X) hay ngược lại. Do đặc điểm cấu trúc, A liên kết với T bằng 2 liên kết hidro, G liên kết với X bằng 3 liên kết hidro. Do các cặp nucleotit liên kết với nhau theo nguyên tắc bổ sung đã đảm bảo cho chiều rộng của chuỗi xoắn kép bằng 20Ao, khoảng cách giữa các bậc thang trên chuối là 3,4Ao, phân tử DNA xoắn theo chu kì xoắn, mỗi chu kì xoắn có 10 cặp nucleotit có chiều cao 34Ao Ở một số loài virus và thực khuẩn thể, DNA của lạp thể, ty thể lại có dạng vòng khép kín. Ngoài mô hình của Waston Crick cho đến nay người ta còn phát hiện ra 4 dạng khác nhau là A, C, D, Z.
Các mô hình này khác với dạng B ở một vài chỉ số: số cặp nucleotit trong một chu kì xoắn, đường kính, chiều xoắn.
Thông tin trong axit nucleic được mã hóa bởi trình tự sắp xếp của các nucleotide dọc theo chuỗi polynucleotide, và trong DNA sợi đôi, trình tự của mỗi sợi xác định trình tự của chuỗi bổ sung phải như thế nào. Mức độ tương đồng về trình tự giữa các DNA từ các vi sinh vật khác nhau là tiêu chí nghiêm ngặt nhất để xác định mức độ liên quan chặt chẽ của chúng.
* Chức năng của DNA:
- Tự nhân đôi trong qúa trình phân chia tế bào, tạo thành 2 bản sao từ một phân tử DNA ban đầu.
- Là khuôn để tạo thành các bản phiên mã khác nhau, các RNA, protein.
-> DNA là vật chất di truyền có chức năng lưu giữ, bảo quản và truyền đạt thông tin di truyền qua các thế hệ, đảm bảo cho sự di truyền giữa các thế hệ đời sau.
2.3.3. RNA
Những nghiên cứu cho thấy, các phân tử RNA xuất hiện sớm hơn DNA và protein trong lịch sử tiến hoá. Các phân tử RNA có nhiều điểm cấu trúc giống với DNA nhưng làm chức năng trung gian trong qúa trình sinh tổng hợp protein với ba dạng khác nhau (rRNA, tRNA, mRNA). Tuy nhiên, trong hai thập niên gần đây, nhiều loại RNA mới được phát hiện cho thấy vai trò đa dạng và quan trọng của chúng trong tế bào. Đáng chú ý có gRNA với vai trò dẫn đường được ứng dụng trong cơ chế chỉnh sửa hệ gen CRISPR/Cas 9 - một bước đột phá của kĩ thuật thao tác DNA bộ gen.
a. Thành phần hoá học và cấu trúc
Các RNA giống với DNA là cấu tạo theo nguyên tắc đa phân, đơn phân là các nucleotit khác nhau ở thành phần đường 5 (pentozo) là ribozo, 4 loại bazo nito là A, U, G, X. Tuy nhiên RNA có cấu trúc 1 mạch đơn thẳng, chiều dài ngắn hơn so với DNA.
RNA có thể hình thành các cấu trúc không gian như kẹp tóc, thuỳ tròn, ...đây là những cấu hình mạch kép do những đoạn nucleotit có trình tự bắt cặp bổ sung theo nguyên tắc G với X và A với U.
b. Cấu trúc và chức năng của các loại RNA
Loại RNA Chức năng
mRN A RNA thông tin, mã hoá cho protein
rRNA RNA ribosome, tạo cấu trúc căn bản của ribosome và xúc tác tổng hợp protein
tRNA RNA vận chuyển, làm cầu nối giữa mRNA và axit amin
snRN A(RNA bé của nhân)
Small nulear RNA có chức năng trong các quá trình ở nhân tế bào, gồm splicing (cắt nối) các tiền mRNA (pre-mRNA)
snoR NA RNA nhỏ hạch nhân (sno RNA - small nucleolar RNA) chế biến và biến đổi hoá học các rRNA
miRN A Điều hoà biểu hiện gen, điển hình là ngăn dịch mã mRNA theo lựa chọn
siRN A RNA nhỏ can thiệp (small interfering siRNA), đóng (dừng) sự biểu hiện gen bằng phân huỷ định hướng các mRNA theo chọn lọc và tạo cấu trúc chất nhiễm sắc cuộn chặt
RNA không mã hoá khác
Có nhiều chức năng trong tế bào như tạo đầu mút NST (telomerase), bất hoạt NST X, vận chuyển các protein vào lưới nội chất
Bảng 2: Cấu trúc và chức năng của các loại RNA
RNA ribosome (rRNA)
rRNA là thành phần cấu tạo, chiếm phân nửa khối lượng riboxom. Tế bào có số lượng lớn ribosome, nên rRNA chiếm tỉ lệ cao, có thể lên tới 75% của tổng số RNA tế bào. Đối với tế bào vi khuẩn, rRNA được tổng hợp ở tế bào chất, ở đây, nó kết hợp với protein thành các tiểu phần của riboxom. Các tiểu phần (rRNA) khác nhau được đánh giá và đặt tên theo hệ số lắng khi siêu li tâm là S (Svedberg - đơn vị đánh giá tốc độ lắng xuống đáy Sedimentation của ống li tâm, S = 1cm3 x 10-13) S càng lớn thì phân tử RNA càng dài. Các riboxom của lục lạp, ti thể và vi khuẩn đều là các riboxom có hệ số lắng li tâm là 70S, gồm hai tiểu đơn vị: Tiểu đơn vị lớn 50S có 1 rRNA 23S và 1 rRNA 5S; tiểu đơn vị nhỏ 30S chỉ có 1 rRNA 16S.
Các ribosome của sinh vật nhân chuẩn thuộc loại 80S, gồm 2 tiểu đơn vị: Tiểu đơn vị lớn 60S có 1 rRNA 28S, 1 rRNA 5,8S và 1rRNA 5S, tiểu đơn vị nhỏ 40S chỉ có 1 rRNA 18S
RNA vận chuyển (transfer RNA)
Trong tế bào có đến hơn 50 loại tRNA khác nhau và ít nhất mỗi loại RNA đặc hiệu cho mỗi loại axit amin. Các tRNA có cấu trúc chung: chiều dài khoảng 70-90 nucleotit, gồm một mạch cuộn lại nhu hình lá chẻ ba nhờ các bắt cặp bổ sung bên trong phân tử, đầu mút 3’ có trình tự kết thúc. Đầu 3’ là nơi gắn vào của axit amin, một thuỳ sẽ mang bộ ba đối mã (anti codon) khớp bổ sung với bộ ba mã hoá trên mRNA là. Bằng cách đó tRNA dịch ngôn ngữ (các codon) trên mRNA thành chuỗi axit amin của mạch polipeptit.
RNA thông tin (mRNA)
mRNA có kích thước rất khác nhau, thời gian sống ngắn, bị phân giải nhanh sau khi dịch mã. Chiều dài trung bình của phân tử mRNA vào khoảng 1,2kilobase. mRNA là một giai đoạn trung gian trong quá trình biểu hiện gen. Các trình tự trên DNA mã hoá cho protein được phiên mã quá tổng hợp phân tử mRNA. Enzyme RNApolymerase dựa vào mạch khuôn mã của DNA mã hoá tạo phân tử mRNA. Bản phiên mã sơ cấp của mRNA tế bào vi khuẩn và cả sinh vật nhân chuẩn chứa trình tự nucleotide nhiều hơn số bộ ba dùng mã hoá protein.
- mRNA vi khuẩn (nhân sơ) mang trình tự mã hoá nhiều gen, mà đầu mỗi đoạn gen có trình tự điển gắn với ribosome, còn gọi là shine-dalgRNAo, để bắt cặp với trình tự bổ sung trên RNA 16S và ở cuối sau dấu dừng có trình tự không mã hoá.
- mRNA trưởng thành của sinh vật nhân chuẩn có cấu trúc phức tạp hơn: đầu 5’ có mũ cap (methyl guanine triphosphate, tiếp theo là các đoạn mã hoá, sau tín hiệu kết thúc là đoạn 3’ không mã hoá, và cuối cùng là đuôi poly-A (150-200 đơn phân adenine). Cấu trúc phức tạp hơn của mRNA nhân chuẩn có lẽ liên quan đến thời gian tồn tại ngắn hay dài để biểu hiện gen
Chức năng: Làm trung gian truyền đạt thông tin di truyền từ DNA đến protein. Xúc tác cho việc cắt đứt các liên kết phosphodieste của các axit nucleic (ribozim).
Ngoài ra, ở virus, RNA còn là các vật chất di truyền.
2.4. Protein
- Thành phần hoá học: bao gồm các nguyên tố C, H, O, N và một số protein ó lượng nhỏ S. Ngoài các nguyên tố trên còn có một lượng rất ít các nguyên tố khác như Fe, Cu, Mn, Ca, …
- Cấu trúc đơn phân: Mỗi acid amin được cấu tạo từ 3 thành phần: Nhóm amino (-NH2), nhóm Carboxyl (-COOH), gốc hữu cơ (R-CH).
- Cấu trúc không gian: Protein gồm 4 bậc cấu trúc, trong đó cấu trúc bậc III, bậc IV là các cấu trúc không gian.
* Chức năng
Protein là cấu trúc sinh học rất đa dạng và linh hoạt nhất trong hệ thống sống. Chính vì vậy, protein có rất nhiều chức năng quan trọng như sau:
- Xúc tác: enzyme tham gia nhiều phản ứng xúc tác trong cơ thể
- Vận tải: hệ thống hemoglobin tham gia vận chuyển oxi, cung cấp cho toàn bộ cơ thể
- Vận động: actin và myosin đảm nhận vai trò vận động.
- Bảo vệ: interferon là một cấu trúc tuyệt vời trong việc nhận biết các tế bào ung thư
- Dự trữ: casein trong sữa mẹ là nguồn cung cấp dinh dưỡng thiết yếu cho trẻ em.
- Điều hòa: hệ thống promoter giúp điều khiển quá trình tổng hợp protein.
* Các liên kết trong từng bậc
- Cấu trúc bậc 1 được tạo ra bởi liên kết peptide là liên kết cộng hóa trị.
- Cấu trúc bậc 2 được hình thành chủ yếu nhờ liên kết hydro giữa các nguyên tử H với N hoặc O là thành phần của các liên kết peptide (khung polipeptide).
- Cấu trúc bậc 3 được hình thành chủ yếu nhờ tương tác kỵ nước giữa các nhóm R không phân cực và nhờ liên kết hydro giữa các nhóm R phân cực hoặc tích điện (các acid amin có tính kiềm và acid) của các acid amin.
- Cấu trúc bậc 4 phổ biến được hình thành chủ yếu do các tương tác Van der Waals giữa các tiểu phần (chuỗi polipeptide với nhau. Cầu disulfide (-S - S-) được hình thành giữa các acid amin cystein là thành phần của các protein có vai trò hình thành ổn định ở các cấu trúc bậc 3 hoặc 4 của các protein nhất định.
3. Các quá trình trao đổi chất
3.1. Trao đổi saccharide
Phân giải Saccharide
Có hai cách phân giải polysaccharide và disaccharide tạo thành monosaccharide; thủy phân và phosphoryl phân.
- Thủy phân là quá trình phân giải các chất nhờ sự có mặt của enzyme và có sự tham gia của nước tạo thành các phân tử nhỏ.
Amylase
Tinh bột + H2O glucose + dextrin
- Phosphoryl phân: acid phosphoric thay thế vai trò của nước trong quá trình thủy phân. Vai trò của phosphorylase là cắt liến kết glycoside của tinh bột hoặc glycogen.
a) Phân giải monosaccaride
Quá trình đường phân
Đường phân hay còn gọi là con đường Embden-Meyerhof xảy ra ở tất cả các sinh vật. Quá trình đường phân xảy ra trong bào tương, sử dụng nguồn nguyên liệu glucose tạo thành 2 phân tử acid pyruvic, tích lũy 2 NADH và kèm theo 2 ATP được giải phóng bằng cách oxy hóa phosphoryl hóa ở mức cơ chất.
Phương trình tổng quát:
C6H12O6 + 2ADP + 2NAD+ + 2P -> 2C3H4O3 + 2ATP + 2H+ + 2H2O
- Giai đoạn tiêu tốn năng lượng gồm 5 phản ứng:
+ Bước 1: là sự phosphoryl hóa glucose, tạo glucose-6-phosphate. Phản ứng cần ATP và Mg2+ tham gia.
+ Bước 2: là sự đổng phân hóa glucose-6-phosphate chuyển thành fructose-6phosphate.
+ Bước 3: là sự phosphoryl hóa lần thứ 2 giữa 2 dạng đường aldose và ketose

+ Bước 4: là phản ứng cắt mạch carbon, biến fructose 1,6-bisphosphate thành dihydroxyacetone phosphate và glyceraldehyde-3-phosphate
+ Bước 5: là phản ứng chuyển hóa nội phân tử giữa dihydroxyacetone phosphate và glyceraldehyde-3-phosphate. Kết quả tạo 2 phân tử glyceraldehyde-3phosphate
- Giai đoạn tái tạo năng lượng
+ Bưóc 6: phản ứng oxy hóa kèm theo sự phosphoryl hóa ở cơ chất biến glyceraldehyde-3- phosphate thành 1,3-bisphosphate.

+ Bước 7: là chuyển gốc phosphate cao năng cho ADP tạo thành 2ATP và 3phosphoglycerate

+ Bước 8: cũng là phản ứng đồng phân hóa, biến đổi 3-phosphoglycerate thành
2-phosphoglycerate
+ Bước 9 là phản ứng loại nước của 2-phosphoglycerate tạo thành
phosphoenolpyruvate
+ Bước 10: là phản ứng tạo pyruvate từ phosphoenolpyruvate và 2 phân tử ATP
enzyme dị lập thể, chúng được tăng cường khi nồng độ ADP, AMP cao và bị ức chế khi nồng độ ATP và citrate cao. Chính vì vậy, khi năng lượng tế bào ở mức thấp, hoạt
độ của các enzyme này tăng cao. Hoặc, với nồng độ citrate cao, phosphofructo kinase bị ức chế vì vậy quá trình đường phân sẽ bị ức chế. Quá trình đường phân bị ức chế glucose sẽ được vận chuyển nhò các đơn chuyển glucose vào các tế bào gan và dự trữ dưới dạng glycogen. Ngoài ra, glucose-6-phosphatase là enzyme xúc tác phản ứng tách glucose khỏi glucose 6-phosphate từ gan vào vòng tuần hoàn. Khi nồng độ glucose 6phosphate tăng, glycogen phosphorylase bị ức chế và glycogen synthase được hoạt hóa, hậu quả là nồng độ glucose trong máu không tăng khi đáp ứng glucagon và epinephrine (Thiếu hụt glucose-6-phosphatase (bệnh Von Gierke)).
Fructose 2,6-bisphosphate là một chất hóa học có khả năng hoạt hóa enzyme phosphofructo kinase-1 bằng cách tăng ái lực giữa enzyme này với fructose 6phosphate. Chẳng hạn, khi nồng độ insulin tăng cao, dưới sự xúc tác của enzyme phosphofructo kinase-2, sản phẩm từ fructose 6-phosphate là fructose 2,6bisphosphate. Quá trình này sử dụng năng lượng ATP, có thể thấy fructose 2,6bisphosphate còn có khả năng giảm hiệu ứng ức chế khi nồng độ ATP cao và thúc đẩy quá trình đường phân diễn ra nhanh chóng một cách gián tiếp. Ngược lại, khi glucagon được giải phóng từ lá lách, chúng sẽ thúc đẩy hoạt tính phosphatase của phosphofructo kinase-2 làm chậm quá trình đường phân.
Biến đổi pyruvate thành các sản phẩm trong điêu kiện thiếu oxy
- Sự tạo thành lactate do xúc tác của lactate dehydrogenase.
Trong mô động vật, lactate được tạo ra trong điều kiện oxy cung cap không đủ. Để duy trì hoạt động sống, lactate được tạo ra nhằm cung cấp một nguồn năng lượng tạm thời cho cơ thể. Ngoài ra, một số vi sinh vật có khả năng tạo lactate như lên men sữa chua, muối dưa, cà, ...
C6H12O6 + 2ADP + 2NAD+ + 2P -> 2 lactate + 2ATP + 2H2O
- Quá trình lên men rượu do một số vi sinh vật khác, chẳng hạn nấm men có khả năng khử nhóm carboxyl thành acetaldehyde và CO2 nhờ xúc tác của pyruvate decarboxylase. Trong bước thứ hai, acetaldehyde bị khử thành ethanol do xúc tác của alcohol dehydrogenase (ADH).
Hình 9: Sơ đồ quá trình đường phân
Điều hòa đường phân
Quá trình đường phân gồm 10 phản ứng với ba phản ứng không thuận nghịch
được xúc tác bởi hexokinase, phosphofructokinase-1, pyruvate kinase. Đây là các
Hình 10: Quá trình khử acetaldehyde thành ethanol
Chu trình CORI
Khi lượng oxy cung cấp cho cơ thể bị hạn chế do hoạt động căng thẳng, mệt mỏi (chạy nhảy, bơi lội, …) thì hệ cơ hầu hết hoạt động trong điều kiện yếm khí, nguồn ATP được cung cấp chủ yếu từ đường phân. Vì vậy, khi mô cơ hoạt động, lượng lactate được tạo ra nhiều sẽ gây độc cho cơ thể (nhiễm toan). Tuy nhiên, một con đường nhằm điều chỉnh nồng độ lactate trong cơ được thực hiện khi lượng lactate cao sẽ vận chuyển đến gan. Tại gan, lactate sẽ được biến đổi thành pyruvate và thực hiện con đường tái tạo glucose (gluconeogenesis). Sau đó glucose lại được đưa đến hệ cơ cung cấp trở lại ATP cho hoạt động. Chu trình này đã giải thích vì sao lượng lactate trong cơ bắp của vận động viên khi tập luyện căng thẳng hay khi thi đầu không ở mức quá cao.
Kết quả của phản ứng: từ pyruvate chứa 3 carbon và kém hoạt động đã biến thành hợp chất 2 carbon ở dạng hoạt hóa là acetyl coenzyme A có chứa liên kết cao năng trong phân tử. Đồng thời, do quá trình oxy hóa nguyên tử hydro của cơ chất được chuyển đến coenzyme NAD+ tạo thành coenzyme dạng khử NADH.
Chu trình acid citric (Chu trình Krebs)
Chu trình acid citric xảy ra ở ty thể thông qua 8 phản ứng đã tạo ra nhiều sản phẩm trung gian là các chất hữu cơ như acid fumarate, acid succinate, acid aketoglutarate, … Các sản phẩm trung gian này sẽ được chuyển hóa thành sản phẩm cuối cùng là CO2, hoặc được vận chuyển ra khỏi ty thể cung cấp cho việc tổng hợp các hợp chất hữu cơ trong tế bào. Phương trình tổng quát:
Acetyl-CoA + 3NAD+ + FAD+ + GDP + P + 2H2O -> 2CO2 + 3NADH + FADH2 + GTP + 2H+ + CoA
Như vậy, từ 2 phân tử acid pyruvic trải qua quá trình phân giải đến sản phẩm cuối cùng là CO2 thì đã tạo được 8NADH, 2FADH2 và 2ATP.


Tính đến đây, 1 phân tử glucose trải qua 2 giai đoạn đường phân và chu trình Krebs, đã tạo ra được 10NADH, 2FADH2 và 4ATP. Chu trình axit citric có ý nghĩa quan trọng:
- Phân tử glucose bị oxy hóa hoàn toàn đến CO2 và H2O và giải phóng toàn bộ năng lượng.
- Tạo ra nhiều coenzyme khử
Từ pyruvate đến acetyl CoA.
Hình 11: Chu trình Cori
Hệ cơ Gan
Oxy hóa acetyl CoA trong chu trình acid citric.
Oxy hóa các coenzyme khử qua chuỗi hô hấp
Hình 12: Sự khử caroboxyl hóa pyruvate tạo thành acetyl CoA
- Nguồn carbon cho các quá trình sinh tổng hợp khác nhau
- Là mắt xích liên hợp, là điểm giao lưu của nhiều đường hướng phân giải và tổng hợp các chất khác nhau trong tế bào, đồng thời cũng là đường hướng chính để phân giải các hợp chất hữu cơ
Hình 13: Chu trình axit citric
Chuỗi vận chuyên điện tử - sự phosphoryl hóa


NADH và FADH2 được sinh ra từ giai đoạn đường phần, chu trình Krebs sẽ
được đưa đến màng trong ty thể để thực hiện việc vận chuyển điện tử cho các chất nh
ận điện tử trên màng ty thể.
Khi một cơ chất bị oxy hóa trong chất nền ty thể, dòng electron sẽ được vận chuyển trong chuỗi hô hấp, tạo sự chênh lệch proton giữa hai bên màng trong ty thể.
Đ
ó là động lực để tổng hợp ATP từ ADP và P thông qua kênh ATP synthase. Khi H+
được vận chuyển từ xoang ty thế vào chất nền (ngược trở lại) theo tỷ lệ: cứ 4H+ sẽ
tổng hợp 1 ATP. Vì vậy NADH cho 10H+ và FADH2 cho 6H+ (do FADH2 đi vào phức hệ II nên bỏ qua phức hệ I dẫn đến lượng proton H+ tạo ra ít hơn).
chylomicron và được chuyển từ ruột non qua hệ bạch huyết và máu đến mô cơ và mô mỡ. Tại mao mạch của các mô này, enzyme ngoại bào lipoprotein lipase sẽ phân giải triacylglycerol thành acid béo và glycerol đi vào trong tế bào để oxy hóa tạo năng lượng (mô cơ), hoặc để tổng hợp triacylglycerol dự trữ (mô mỡ).
Chylomicron vận chuyển triacylglycerol đến các mô ngoại biên như mô mỡ và mô cơ. Phần còn lại của nó sẽ đến gan, VLDL được hình thành tại gan và vận chuyển lipid nội sinh đến các mô ngoại biên. VLDL bị phân giải thành IDL, kết hợp với cholesterol và cholesteryl ester từ HDL tạo nên LDL, mang cholesterol đến các mô khác. HDL vận chuyển cholesterol từ huyết thanh và các mô ngoại biên đến gan. Tại gan, cholesterol có thê biến đổi thành muối mật và đi vào túi mật.

3.2.2. Chuyển hoá glyxerol
Quá trình phân giải triacylglycerol nhờ hệ thống enzyme thủy phân liên kết ester tạo thành glycerol và acid béo. Glycerol được phosphoryl hóa thành L-glycerol3-phosphate (glycerol-3- phosphate ở quá trình đường phân dạng D), rồi oxy hóa thành dihydroxyacetone phosphate và sau đó chuyên thành D-glycerol-3-phosphate. Dglycerol-3-phosphate có thể tiếp tục oxy hóa theo con đường đường phân và chu trình acid citric để tạo thành CO2, H2O và năng lượng, hoặc tham gia tổng hợp glucose.
3.2.3. Oxi hoá axit béo
a. Acid béo được hoạt hoá thành acetyl - CoA ở bào tương
Hình 14: Chuỗi vận chuyển điện tử trên màng trong ty thể
3.2. Trao đổi lipid
Tế bào có thể phân giải từ 3 nguồn: lipid thức ăn, lipid dự trữ và lipid tổng hợp diễn ra ở các cơ quan trong cơ thể.
Sự phân giải lipid bắt đầu là sự thủy phân các liên kết ester, giải phóng các hợp phần cấu tạo như acid béo, glycerol. Sau đó, acid béo và glycerol bị phân giải theo các con đường khác nhau, tạo ra năng lượng hoặc tham gia vào quá trình tổng hợp các chất trung gian trong cơ thể
3.2.1. Thuỷ phân các liên kết ester trong lipid
Ở động vật có xương sống, triacylglycerol từ thức ăn được nhũ tương hóa nhờ muối mật thành các hạt micelle nhỏ dễ khuếch tán và bị thủy phân dưới tác dụng của lipase tuyến tụy cắt liên kết ester, tạo thành 2- monoacylglycerol và hai gốc acid béo.


Các acid béo tự do và monoacylglyecrol được chuyển đến các tế bào niêm mạc ruột để tái tạo triacylglycerol rồi được chuyển đến các mô và bị phân giải tại đó. Các acid béo được biến đổi thành acetyl - CoA. Ba phân tử acetyl - CoA có thể kết hợp với glycerol, hoặc hai phân tử có thể kết hợp với một monoacylglyecrol để tạo thành triacylglycerol. Triacylglycerol kết hợp với cholesterol và các apoprotein tạo thành
Acid béo được hoạt hóa thành acetyl - CoA với sự xúc tác của acetyl - CoA synthetase và sự tham gia của ATP. Pyrophosphate được hình thành sẽ bị thủy phân bởi pyrophosphate từ hai phân tử phosphate.


Hình 15: Hoạt hoá axit béo
b. Acetyl-CoA được vận chuyển vào ty thể
Ở tế bào sinh vật nhân thực, acid béo thường được hoạt hóa ở bào tương gần màng ngoài ty thể và được vận chuyển vào ty thể. Các acetyl có từ 12 carbon trở xuống sẽ trực tiếp đi qua màng ty thể. Acetyl-CoA được ester hóa với cRNAitine thành acetyl-cRNAitine giải phóng HS-CoA dưới sự xúc tác của cRNAitine acyltransferase I và đi qua các lỗ lớn ở màng ngoài ty thể. Acetyl-cRNAitine có thể
được hình thành ngay ở khoang gian màng. Sau đó acyl-cRNAitine được chuyển vào bên trong ty thê nhờ chầ't vận chuyển acetyl-cRNAitine. Gốc acetyl - CoA được tái tạo nhờ cRNAitine acetyltransferase II và cRNAitine và được giải phóng, quay trở lại khoang gian màng đê tham gia vận chuyển tiếp các gốc acid béo khác.
II. ƯU THẾ CỦA CÁC ĐỐI TƯỢNG VI SINH VẬT TRONG NGHIÊN
CỨU HOÁ SINH VÀ DI TRUYỀN HỌC
Trong nghiên cứu di truyền học, các đối tượng vi sinh vật có nhiều ưu thế hơn hẳn các động vật thực vật bậc cao.
1. Thời gian thế hệ ngắn, tốc độ sinh sản nhanh
Trong điều kiện thuận lợi, tế bào E. coli có thể phân chia một lần trong 20 phút, còn bacteriophage có thể trong 30 - 40 phút tạo ra hàng trăm cá thể, nấm men có thể phân chia tế bào trong 2 giờ. Đặc điểm của nghiên cứu di truyền học là theo dõi qua nhiều thế hệ nên các đối tượng vi sinh vật giúp rút ngắn đáng kể thời gian thí nghiệm. Ví dụ, với E. coli, thí nghiệm hôm trước, qua ngày sau đã có thể đánh giá kết quả
2. Số lượng cá thể tăng nhanh
Hình 16: Sự vận chuyển Acetyl-CoA vào hệ thống
Quá trình oxy hoá acetyl - CoA
Acid béo (ROOH) được oxy hóa qua một chuỗi 4 phản ứng tạo sản phẩm Acetyl-CoA và Acetyl-CoA ngắn hơn 2 carbon. Acetyl-CoA di chuyển vào ty thể tham gia vào chu trình citric tiếp tục oxy hóa cung cấp năng lượng cho cơ thể. Trong khi đó, Acetyl-CoA tiếp tục phân cắt theo con đường 3-oxy hóa tạo Acetyl-CoA.
Các vi sinh vật đơn bào, mỗi tế bào là một cá thể. Nếu đủ dinh dưỡng các vi sinh vật sinh sản nhanh tạo quần thể có số lượng cá thể lớn có thể khoảng 1010 - 1012 tế bào/ml. Tế bào E. coli có đường kính 1 µm (micromet) nếu đủ dinh dưỡng và mọi điều kiện thuận lợi khác thì sau 44 giờ có thể tạo ra một lượng sinh khối nặng bằng Trái Đất. Như vậy, số lượng cá thể lớn giúp nâng cao năng suất phân giải di truyền, tức khả năng phát hiện các đột biến và tái tổ hợp có tần số xuất hiện rất nhỏ.
Ưu thế này lại được tăng thêm nhờ môi trường nuôi đơn giản, dễ nuôi cấy, dễ nhân giống, mà điều kiện nuôi cấy không cồng kềnh, ít tốn diện tích, dễ kiểm soát theo công thức chặt chẽ như khi làm thí nghiệm hóa học.
3. Cấu tạo bộ gen đơn giản
Vi khuẩn và virut có bộ gen là DNA trần dễ tiến hành thí nghiệm trực tiếp trên DNA cũng như chiết tách tinh sạch. Số locus cũng ít hơn so với các sinh vật khác. Các vi nấm và vi tảo có thể tồn tại ở dạng đơn bội (n) với thời gian dài, nên các gen lặn có thể biểu hiện ngay, mà không cần phải tiến hành lai phân tích hay đưa về dạng đồng hợp tử lặn. Tuy nhiên, các vi sinh vật kể trên có trạng thái lưỡng bội (2n) nên cũng dễ dàng thực hiện phân tích tái tổ hợp.
Các tính trạng ở vi sinh vật cũng đơn giản hơn, xác định di truyền các tính trạng này ít phức tạp hơn, nên dễ nghiên cứu. Đối với các tính trạng sinh hóa hay tính đề kháng thì có thể dễ dàng sử dụng môi trường chọn lọc để phát hiện.
4. Dễ phát sinh và thu nhận các đột biến
Hình 17: Quá trình oxi hoá của Palmitoy-CoA
Phương trình tổng quát:
Cn-acetyl CoA + CoA + FAD+ + NAD+ + H2O->


Cn2-acetyl CoA + acetyl - CoA + FADH+ + NADH + + H+
Các phân tích di truyền học phần lớn dựa vào những khác biệt của dạng bình thường so với đột biến. Tần số đột biến ở thực vật và động vật khoảng 10–5 – 10–7, khó thu nhận và cần thời gian dài khoảng vài thế hệ để khẳng định đúng là dạng đột biến. Nhiều đột biến ở động vật dễ gây chết nên số lượng đột biến thu nhận được ở động vật rất hạn chế.
- Các đột biến ở vi sinh vật có thể thu nhận dễ dàng, thậm chí có tần số xuất hiện thấp 10–10, mà việc xác nhận dạng đột biến cũng rất nhanh.
- Nhờ ưu thế này mà di truyền học vi sinh vật phát triển rất nhanh hình thành nên di truyền học phân tử và sinh học phân tử
- Dễ nghiên cứu bằng các kỹ thuật vật lý hóa học
Đa số các vi sinh vật có cấu tạo đơn bào nên quần thể của chúng có độ đồng nhất cao hơn so với các tế bào sinh vật đa bào bậc cao bắt nguồn từ nhiều loại mô khác nhau. Cấu tạo tế bào vi sinh vật đơn giản, dễ chiết tách, tinh sạch DNA. Có thể nuôi vi sinh vật đồng nhất (synchronous culture) tức là đa số các tế bào ở những giai đoạn phát triển gần giống nhau. Độ đồng nhất cao của vật liệu thí nghiệm tạo thuận lợi cho việc sử dụng các phương pháp vật lý hóa học trong nghiên cứu di truyền.
Do những ưu điểm kể trên, với việc sử dụng các đối tượng vi sinh vật, di truyền học đã bước vào giai đoạn nghiên cứu di truyền “trong ống nghiệm” (in vitro).
Mặc dù các vi sinh vật có những đặc điểm riêng nhưng chúng vẫn tuân theo các quy luật di truyền chung, các kết quả thu được có thể đối chiếu áp dụng cho các vi sinh vật bậc cao.
III. CÁC ĐẶC ĐIỂM CỦA DI TRUYỀN HỌC VI SINH VẬT
1. Thông tin di truyền trong vi sinh vật
Thông tin di truyền ở vi khuẩn và nhiều loại virus được mã hóa trong DNA, nhưng một số loại virus sử dụng RNA. Sao chép bộ gen là cần thiết để thừa kế các tính trạng do di truyền xác định. Sự biểu hiện gen thường bao gồm phiên mã DNA thành RNA thông tin và dịch mã mRNA thành protein.
2. Tổ chức bộ gen Nhiễm sắc thể vi khuẩn là một phân tử DNA dạng tròn có chức năng như một yếu tố di truyền tự sao chép (bản sao). Các yếu tố di truyền ngoài nhiễm sắc thể như plasmid và vi khuẩn là những bản sao không cần thiết thường xác định khả năng chống lại các chất kháng khuẩn, sản xuất các yếu tố độc lực hoặc các chức năng khác. Nhiễm sắc thể sao chép bán nguyệt; mỗi sợi DNA đóng vai trò như khuôn mẫu để tổng hợp sợi bổ sung của nó.
3. Sự đột biến và chọn lọc
Tập hợp đầy đủ các yếu tố quyết định di truyền của một sinh vật cấu thành kiểu gen của nó và các đặc điểm quan sát được cấu thành kiểu hình của nó. Đột biến là những thay đổi di truyền trong kiểu gen có thể xảy ra một cách tự phát hoặc được gây ra bởi các biện pháp xử lý hóa học hoặc vật lý. Các vi sinh vật được chọn làm dòng tham chiếu được gọi là kiểu hoang dại, và thế hệ con cháu của chúng có đột biến được gọi là thể đột biến. Các phương tiện chọn lọc phân biệt giữa chủng loại hoang dại và chủng đột biến dựa trên tốc độ tăng trưởng; môi trường phân biệt giữa chúng dựa trên các đặc tính kiểu hình khác.
4. Trao đổi thông tin di truyền Trao đổi gen giữa các vi khuẩn xảy ra theo một số cơ chế. Trong quá trình biến nạp, vi khuẩn nhận lấy DNA của người cho ngoại bào. Trong quá trình tải nạp, DNA của vi khuẩn cho được đóng gói trong một thực khuẩn sẽ lây nhiễm sang vi khuẩn nhận. Trong tiếp hợp, vi khuẩn cho truyền DNA cho thể nhận bằng cách giao phối. Tái tổ hợp là sự sắp xếp lại các bộ gen của người cho và người nhận để tạo thành các bộ gen mới, lai. Transposon là các đoạn DNA di động di chuyển từ nơi này sang nơi khác trong hoặc giữa các bộ gen.
5. DNA tái tổ hợp và nhân bản gen
Nhân bản gen là sự kết hợp gen ngoại lai vào vectơ để tạo ra phân tử DNA tái tổ hợp sao chép và biểu hiện gen ngoại lai trong tế bào nhận. Các gen nhân bản được phát hiện bằng kiểu hình mà chúng xác định hoặc bằng trình tự nucleotit cụ thể mà chúng chứa. DNA tái tổ hợp và nhân bản gen là những công cụ cần thiết cho nghiên cứu vi sinh phân tử và y học. Chúng có nhiều ứng dụng y tế, bao gồm phát triển vắcxin mới, sinh học, xét nghiệm chẩn đoán và phương pháp điều trị.
6. Quy chế biểu hiện gen
Các đặc tính kiểu hình của vi khuẩn được quyết định bởi kiểu gen và điều kiện sinh trưởng của chúng. Đối với vi khuẩn trong môi trường nuôi cấy thuần khiết, những thay đổi trong điều kiện sinh trưởng thường dẫn đến sự thích nghi sinh lý có thể dự đoán được ở tất cả các thành viên của quần thể. Thông thường, các sản phẩm gen thiết yếu được tạo ra với số lượng cho phép phát triển nhanh nhất trong môi trường nhất định và các sản phẩm cần thiết trong chỉ được sản xuất khi cần thiết.
Sự thích nghi sinh lý thường gắn liền với những thay đổi trong các hoạt động trao đổi chất. Dòng chảy của các chất chuyển hóa thông qua các con đường sinh hóa cụ thể có thể được kiểm soát bằng cách điều chỉnh quá trình tổng hợp các enzym cụ thể và bằng cách thay đổi hoạt động của các enzym hiện có. Các cơ chế điều chỉnh sự biểu hiện của gen bằng cách ảnh hưởng đến tổng hợp các sản phẩm gen cụ thể được thảo luận ở đây.
Quy chế biểu hiện của một gen hoặc một nhóm gen liên quan đến một quá trình trao đổi chất cụ thể. Cảm ứng và ức chế cho phép vi khuẩn điều chỉnh sản xuất các sản phẩm gen cụ thể để đáp ứng với các tín hiệu thích hợp. Nói chung, các enzyme dị hóa được tạo ra khi có mặt chất nền cho con đường trong môi trường tăng trưởng, và các enzyme sinh tổng hợp bị ức chế bởi sản phẩm của con đường này. Các enzim tham gia vào một con đường sinh hoá đơn lẻ thường chiếm các vị trí liền kề trên nhiễm sắc thể vi khuẩn và được cảm ứng hoặc phối hợp với nhau.
Sự biểu hiện của gen ở vi sinh vật thường do điều kiện nội bào hoặc môi trường quy định. Quá trình điều tiết có thể ảnh hưởng đến bất kỳ bước nào trong biểu hiện gen, bao gồm bắt đầu hoặc kết thúc phiên mã, dịch mã hoặc hoạt động của các sản phẩm gen. Operon là một tập hợp các gen được phiên mã thành một đơn vị duy nhất
và được biểu hiện một cách phối hợp. Cơ chế điều hòa cụ thể tạo ra hoặc kìm hãm một gen hoặc operon cụ thể. Tất cả các operon trong cơ quan điều hòa được điều khiển một cách phối hợp bởi cùng một cơ chế điều chỉnh.
Sự biểu hiện của các yếu tố quyết định di truyền ở vi khuẩn liên quan đến dòng thông tin một chiều từ DNA đến RNA đến protein. Trong thực khuẩn, DNA hoặc RNA đều có thể đóng vai trò là vật chất di truyền. Trong quá trình lây nhiễm vi khuẩn bởi RNA thực khuẩn, các phân tử RNA đóng vai trò như khuôn mẫu để sao chép RNA và như mRNA. Các nghiên cứu với nhóm virus động vật retrovirus cho thấy rằng các phân tử DNA có thể được tổng hợp từ các khuôn mẫu RNA bằng các enzym được chỉ định là các polymerase DNA phụ thuộc RNA (sao chép ngược). Sự đảo ngược hướng thông thường của dòng thông tin di truyền, từ RNA sang DNA thay vì từ DNA sang RNA, là một cơ chế quan trọng để cho phép thông tin từ retrovirus được mã hóa trong DNA và được tích hợp vào bộ gen của tế bào động vật.
7. Vật chất di truyền trong vi sinh vật
Vật chất di truyền của vi khuẩn và plasmid là DNA. Vi rút vi khuẩn (thực khuẩn thể hoặc thể thực khuẩn) có DNA hoặc RNA làm vật liệu di truyền. Hai chức năng thiết yếu của vật chất di truyền là sao chép và biểu hiện. Vật chất di truyền phải
sao chép chính xác để thế hệ con cháu thừa hưởng tất cả các yếu tố quyết định di truyền cụ thể (kiểu gen) của sinh vật bố mẹ. Sự biểu hiện của vật chất di truyền cụ thể trong một tập hợp các điều kiện sinh trưởng xác định các đặc điểm quan sát được (kiểu hình) của sinh vật. Vi khuẩn có ít đặc điểm cấu trúc hoặc phát triển có thể được quan sát dễ dàng, nhưng chúng có một loạt các khả năng sinh hóa và mô hình nhạy cảm với các chất kháng khuẩn hoặc thực khuẩn. Các đặc điểm sau này thường được chọn làm các đặc điểm di truyền để phân tích trong các nghiên cứu về di truyền vi khuẩn.
IV. PHƯƠNG PHÁP NGHIÊN CỨU ĐẶC THÙ CỦA DI TRUYỀN HỌC
VI SINH VẬT
Đối với các vi sinh vật, phân tích di truyền là phương pháp duy nhất để nghiên cứu các đặc tính di truyền và biến dị của chúng. Do các vi sinh vật thường có bộ gen đơn bội, đặc biệt là vi khuẩn chỉ có một nhóm liên kết gen nên sơ đồ phân tích di truyền học ở chúng là đơn giản hơn các sinh vật nhân thực bậc cao, bao gồm các giai đoạn sau:
- Xác định các gen
- Xác định trật tự của các locus trên nhiễm sắc thể
- Xác đinh cấu trúc tinh vi của gen
1. Nghiên cứu trực tiếp ở cấp độ tế bào và cấp độ phân tử
Di truyền học cổ điển sử dụng các mô hình đối tượng như đậu Hà Lan Pisum sativum, cây bắp (ngô) Zea mais, ruồi giấm Drosophila melanogaster và chuột nhắt Mus musculus, là những sinh vật đa bào mà sự biểu hiện tính trạng phải trải qua một quá trình phát triển phức tạp và kèm theo biệt hóa, việc quan sát trên các đối tượng này
được thực hiện gián tiếp. Trong khi đó, nhiều đối tượng vi sinh vật như vi khuẩn Escherichia coli, nấm men Saccharomyces cerevisiae là sinh vật đơn bào nên các quan sát thực hiện trực tiếp hơn ở mức tế bào. Ví dụ, trên môi trường tối thiểu với lượng muối hữu cơ và glucoz cho trước, một tế bào E. coli có thể tổng hợp tất cả các hợp chất cần thiết cho sự tăng trưởng, sống sót và sinh sản. Nếu tế bào bị đột biến mất khả năng tổng hợp một chất nào đó như một loại axit amin thì nó không mọc được trên môi trường tối thiểu. Trên môi trường có bổ sung thuốc kháng sinh thì chỉ có các tế bào đề kháng mới mọc được.
Hơn nữa, các tế bào vi sinh vật có nhu cầu dinh dưỡng đơn giản dễ kiểm soát thành phần môi trường nuôi, dễ nuôi cấy, dễ nhân giống, dễ dàng thu nhận nhiều loại đột biến phục vụ cho nghiên cứu trực tiếp và nhanh chóng ở cấp độ tế bào, phân tử. Do vậy, chúng là những đối tượng lý tưởng cho nghiên cứu di truyền học phân tử
Cấu trúc bộ gen
DNA hiện diện trong bộ gen (Genome) của tất cả các loại tế bào từ vi khuẩn đến người. Giữa các sinh vật nhân sơ Prokaryota và nhân chuẩn Eukaryota có sự khác nhau đáng kể về kích thước, thành phần cấu tạo và tổ chức của DNA trong tế bào.
Bộ gen của vi khuẩn E. coli và đa số các sinh vật nhân sơ là một phân tử DNA có dạng vòng tròn kín. Khái niệm nhiễm sắc thể hiện nay được dùng cho cả vi khuẩn, nên nói nhiễm sắc thể vi khuẩn ta hiểu đó là sợi DNA. Ty thể và lục lạp cũng có DNA riêng, mà cấu trúc bộ gen cũng tương tự vi khuẩn.
Đa phần DNA của Eukaryota như nấm sợi, nấm men, vi tảo được tổ chức thành nhiều nhiễm sắc thể trong nhân tế bào. Mỗi nhiễm sắc thể chứa 1 phân tử DNA thẳng mạch kép kèm theo một số protein như histon. Các nhiễm sắc thể có số lượng và hình dạng đặc trưng cho tế bào của mỗi loài sinh vật nhân chuẩn.
Các virut có bộ gen rất đa dạng: DNA mạch đơn hoặc kép, RNA mạch đơn hoặc kép, nhưng chỉ một loại phân tử.
1.1. Phân tích đột biến
Phương pháp này được áp dụng để xác định các gen và được tiến hành bằng cách đo đếm các kết quả cuối cùng cuả sự biểu hiện gen thành ra sự biến đổi kiểu hình (đặc điểm hình thái, hoá sinh, kháng nguyên hoặc tính mẫn cảm đối với các tác nhân hoá học, vật lí và sinh học khác nhau của các tế bào vi khuẩn). Việc phát hiện một đột biến ngẫu nhiên hay gây tạo sẽ chỉ ra sự tồn tại của một gen cụ thể.
Sự biến đổi hình thái ở vi sinh vật bao gồm các biến đổi về kích thước, hình dạng và sự hình thành sắc tố của các khuẩn lạc do các tế bào bị đột biến tạo nên trên các môi trường dinh dưỡng đặc cũng như sự biến đổi của bản thân các phân tử của tế bào (gia tăng kích thước, thay đổi hình thái, ...). Sự biến đổi hoá sinh bao gồm các biến đổi liên quan đến việc tế bào mất khả năng tổng hợp các amino axit hoặc vitamin hoặc mất khả năng chuyển hoá các hợp chất cacbonhydrat. Các biến đổi về kháng nguyên thể hiện ở chỗ vi khuẩn bị mất đi những kháng nguyên nhất định. Các biến đổi trong
tính bền vững của vi khuẩn đối với các tác nhân khác nhau liên quan đến sự xuất hiện của chúng trong các khả năng đề kháng đối với sự chiếu xạ, với các hoá chất khác nhau (kể cả các loại thuốc kháng sinh) hoặc với phage.
Do tần số đột biến ở vi khuẩn là rất thấp nên việc phân lập các tế bào bị đột biến chỉ có thể thực hiện được trong các thí nghiệm với các quần thể tế bào. Như vậy, về nguyên tắc, trong trường hợp này có thể sử dụng bất kỳ phương pháp nào cho phép tách được các thể đột biến từ các quần thể. Việc xác định số lượng các đột biến dựa trên các phương pháp xác định tần số đột biến. Thông thường, để phân tích di truyền cần có các nòi đột biến mang các đột biến vị trí cho trước, chẳng hạn, đối với B. subtilis, có thể xử lí sơ bộ DNA gây biến nạp bằng các tác nhân gây đột biến. Ở E. coli, có thể gây các đột biến có vị trí xác định bằng cách đưa vào tế bào vi khuẩn các gen đột biến nhờ các phage tải nạp.
1.2. Phân tích tái tổ hợp Đây là phương pháp đặc trưng được dùng để xác định vị trí và trật tự của các gen trên nhiễm sắc thể. Đối với vi khuẩn, việc phân tích di truyền dựa vào các quá trình trao đổi vật liệu di truyền như biến nạp, tải nạp, tiếp hợp hay còn gọi là giao nạp. Ở các vi nấm, việc phân tích di truyền được tiến hành bằng phân tích phép phân tích bộ bốn và dựa trên chu trình cận hữu tính.
Nói chung, sự trao đổi di truyền ở các vi khuẩn và quá trình hữu tính ở các cơ thể bậc cao là không giống nhau. Việc truyền vật liệu di truyền từ vi khuẩn thể cho (donor) sang vi khuẩn thể nhận (recipient) có thể coi như sự kết hợp nhân của các tế bào sinh dục (ở đây là sự tạo thành các thể lưỡng bội từng phần), còn sự sát nhập của vật liệu di truyền vào bộ gen của vi khuẩn thể nhận, và sự hình thành nhiễm sắc thể tái tổ hợp sau đó, có thể so sánh với các kết quả của giảm phân. Chính các hệ thống tái tổ hợp này là cơ sở cho phương pháp phân tích tái tổ hợp và lập bản đồ di truyền ở vi khuẩn. Ví dụ, trật tự của hấu hết các gen trên nhiếm sắc thể E. coli được xác định là nhờ sử dụng tiếp hợp và tải nạp, ở B. subtilis nhờ tải nạp và biến nạp, còn ở Samonela typhimurium chủ yếu là nhờ tải nạp. Ngoài ra, phép phân tích tái tổ hợp này còn dược sử dụng để nghiên cứu cấu trúc tinh vi của gen
1.3. Phân tích sao chép
Phương pháp này cho phép xác định trật tự các gen trên nhiễm sắc thể dựa trên sự tính toán các số liệu về sự bắt đầu sao chép (tái bản) của nhiễm sắc thể từ một điểm xác định. Do thời gian sao chép của một phần nhiễm sắc thể nhất định phụ thuộc vào khoảng cách từ phần đó đến điểm khởi đầu sao chép nên thứ tự sao chép phản ánh trình tự sắp xếp của các gen. Như vậy, bản đồ NST chỉ có thể được xây dựng dựa trên các dẫn liệu về trật tự sao chép của các phần riêng biệt của nhiễm sắc thể
1.4. Phân tích đoạn khuyết
Phép phân tích đoạn khuyết được sử dụng để xác định vị trí của các gen trên nhiễm sắc thể cũng như để nghiên cứu cấu trúc tinh vi của các gen. Nó dựa trên việc
tính toán các đoạn khuyết trên nhiễm sắc thể. Nhờ sự phân tích này người ta đã phát hiện được vị trí cuả hàng loạt gen ở E. coli và S. typhimurium, hiểu biết được cấu trúc tinh vi của các gen trên operon lactose ở vi khuẩn E. coli. Phương pháp này cũng được sử dụng rộng rãi để nghiên cứu cấu trúc tinh vi ở phage.
1.5. Phân tích bổ sung
Phương pháp này được sử dụng để phát hiện chức năng của các gen nhất định tham gia vào việc xác định một đặc tính nào đó của vi khuẩn, dựa trên hiện tượng bổ sung của các gen (nghĩa là sự tương tác của các sản phẩm gen).
Bằng cách lai các thể đột biến của cùng một gen có nguồn gốc độc lập nhau trong khi cho lây nhiễm các phage, đã làm xuất hiện các phage kiểu dại. Điều này chỉ có thể xảy ra bởi sự tái tổ hợp bên trong gene, nếu như các phần nhỏ riêng biệt của gen đều bị đột biến.
Đơn vị chức năng di truyền không chia nhỏ (cistron) có thể xác định được bằng sự phân tích bổ sung (complementation analysis), trong đó gen mà cụ thể là sản phẩm của nó được trắc nghiệm về khả năng bù đắp cho một đột biến tại một gen tương đồng trong cùng tế bào. Sự bổ sung liên tiếp làm phục hồi kiểu hình dại.

Hình 18: Sơ đồ minh hoạ trắc nghiệm cis-trans
1.5. Năng suất phân giải
Năng suất phân giải của di truyền học được xác định bởi khoảng cách giữa các cấu trúc di truyền (gene) cần phân tích trên nhiễm sắc thể Đại lượng này phụ thuộc vào số lượng cá thể đời con nghiên cứu được từ một phép lai cụ thể, số con cháu thu được càng lớn thì khả năng phát hiện các thể tái tổ hợp hiếm càng lớn, tức năng suất phân giải của phân tích di truyền học càng cao. Theo luật số lớn này, các vi khuẩn tỏ ra rất thuận lợi trong phân tích di truyền học, vì trong một thời gian ngắn có thể thu được một số lượng cực kì lớn con cháu từ một tế bào vi khuẩn, cũng như có thể sử dụng các môi trường nuôi cấy khác nhau để chọn lọc các cá thể tái tổ hợp.
2. Dòng tế bào
Đặc điểm của các tế bào vi sinh vật là rất nhỏ bé, phải nhìn dưới kính hiển vi mới thấy được. Do vậy khó có thể quan sát từng tế bào riêng lẻ, hơn nữa, bằng cách này cũng không ghi nhận được các tính trạng biến dưỡng, tính đề kháng và nhiều tính trạng khác. Do vậy, di truyền vi sinh vật không nghiên cứu từng tế bào riêng lẻ mà là dòng của tế bào, tức tập hợp của nhiều tế bào bắt nguồn từ một tế bào ban đầu nhờ
sinh sản vô tính. Thông thường, tế bào vi sinh vật được cấy lên môi trường thạch đặc, rồi trải đều để các tế bào nằm rời xa ra thì mỗi tế bào mọc lên thành một cụm rời gọi là khuẩn lạc (colony). Mỗi khuẩn lạc cũng là một dòng tế bào (clone) gồm các tế bào bắt nguồn tự sự phân chia của một tế bào ban đầu.
4. Kiểu sinh sản Các virut chỉ sinh sản trong tế bào chủ sống với nhiều cơ chế khác nhau phụ thuộc kiểu virut
Các vi khuẩn không có các cơ chế nguyên phân và giảm phân, phân chia tế bào theo cơ chế trực phân, nhưng cũng có sinh sản vô tính và sự trao đổi thông tin di truyền tương tự sinh sản hữu tính được gọi là quá trình cận hữu tính.
Các nấm sợi, nấm men, vi tảo có các quá trình sinh sản vô tính và hữu tính về căn bản giống các sinh vật bậc cao, thực hiện qua nguyên phân và giảm phân.
Hình 19: Khuẩn lạc (colony) của vi sinh vật
Mỗi khuẩn lạc dễ nhìn thấy bằng mắt thường, có tối thiểu 107 tế bào. Như vậy, các tính trạng ở vi sinh vật được ghi nhận qua một quần thể gồm hàng trăm triệu, hàng tỉ tế bào.
Dòng tế bào mang một đặc tính di truyền nào đó gọi là chủng (strain). Ví dụ: chủng vi khuẩn tạo nhiều vitamin B12 hay chủng vi khuẩn mất khả năng tổng hợp một axit amin nào đó.
3. Các tính trạng
Các đột biến ở vi sinh vật thường được phát hiện theo sự biến đổi các tính trạng
V. BIẾN DỊ Ở VI SINH VẬT
Biến dị ở vi sinh vật cũng tuân theo quy luật di truyền chung.
Biến dị và đột biến có ý nghĩa quan trọng trong nghiên cứu di truyền học. Các đột biến gồm nhiều loại khác nhau và chúng được thu nhận dễ dàng từ các vi sinh vật nhờ các phương pháp phát hiện chuyên biệt. Các đột biến gen bắt nguồn từ những biến đổi phân tử trên DNA. Con người có thể sử dụng các tác nhân vật lí và hóa học gây nên các đột biến nhân tạo hay cảm ứng.
sau:
Hình thái: kích thước, hình dạng tế bào hay khuẩn lạc, có màng nhân hay không, khả năng di động, ...
Sinh hóa: sự hiện diện của các sắc tố, màu sắc đặc trưng.
Nuôi cấy: như kiểu hô hấp, kiểu dinh dưỡng (khuyết dưỡng - auxotroph) hoặc nhu cầu đòi hỏi các nhân tố tăng trưởng.
Tính đề kháng: như kháng thuốc, kháng phage, chịu nhiệt, ...

Miễn dịch: như các phản ứng kháng thể, kháng nguyên, ...
Các đột biến có thể xuất hiện ngẫu nhiên hay do gây tạo ra nhờ các tác nhân gây đột biến. Mỗi gen có tần số đột biến đặc trưng.
Các tính trạng ở vi sinh vật được kí hiệu bằng 3 chữ tắt tiếng Anh hoặc đôi khi chữ hoa đầu tiên. Kèm theo kí hiệu còn thêm dấu + hoặc – hoặc chữ tắt để giải thích rõ thêm tính trạng. Ví dụ: lac– để chỉ mất khả năng tổng hợp lactoz; his+ - tổng hợp histidin; strS – nhạy cảm (Sensible) với Streptomycin, strR – đề kháng (Resistant) với Streptomycin.
Để chỉ hai giới tính khác nhau không dùng hai kí hiệu ♀ và ♂ thay vào đó là các chữ như mt (+), mt (-) (mating typ) (ở Chlamydomonas reinhardi), hoặc A, a (ở Neurospora crassa).
Biến dị ở vi sinh vật cũng tuân theo các quy luật di truyền như ở sinh vật bậc cao. Nó có nhiều loại và số lượng rất lớn, cung cấp nguyên liệu thường xuyên cho quá trình tiến hóa và nghiên cứu di truyền học. Nhiều phương pháp phát hiện và chọn lọc các đột biến khác nhau được sử dụng để thu nhận các đột biến có tần số xuất hiện rất thấp. Chúng góp phần đáng kể cho các nghiên cứu di truyền. Các thay đổi thêm, mất hay thay thế các nucleotit trên phân tử DNA dẫn đến các biến đổi phân tử, gây ra các đột biến gen như: lệch khung, sai hay nhầm nghĩa và im lặng. Việc phát minh ra các tác nhân gây đột biến vật lí và hóa học làm tăng đáng kể nguồn đột biến phục vụ cho nghiên cứu và chọn giống. Con người có thể tăng nguồn biến dị bằng lai tạo hay sử dụng các tác nhân gây đột biến.
Vi sinh vật cũng có các loại đột biến như ở sinh vật bậc cao. Ở đây chỉ nêu các đột biến thường gặp ở vi sinh vật và có ý nghĩa quan trọng đối với di truyền phân tử
1. Quá trình đột biến tự nhiên
Đột biến gen là đột biến được hiểu theo nghĩa hẹp, là chỉ những biến đổi xảy ra bên trong cấu trúc gen. Mỗi đột biến gen dẫn đến sự thay đổi trình tự nucleotit tạo ra các alen khác nhau. Đột biến có thể xảy ra do biến đổi nhiều nucleotit, có thể do 1 nucleotit. Đột biến gen không phát hiện được khi quan sát tế bào học.
Các gen khác nhau của cùng 1 sinh vật có thể có tần số đột biến khác nhau.
Nhưng tần số đột biến tự nhiên đối với mỗi gen là một số ổn định.
Tần số đột biến được đánh giá theo các căn cứ khác nhau như: trên 1 lần sao chép, 1 lần phân bào hay trên 1 giao tử và trên 1 tế bào/1 thế hệ
Ví dụ: Ở E. coli đột biến từ nhạy cảm với streptomycin sang kháng tức StrSStrR với tần số 4.10-10 đột biến tính trên 1 tế bào/1 thế hệ Để dễ hiểu ta có thể tính ngược lại tức trong 10 tỉ tế bào của một thế hệ có 4 đột biến StrR (resistance) kháng streptomycin xuất hiện ngẫu nhiên.
Tuy tần số đột biến của từng gen là rất thấp, nhưng tổng các đột biến của nhiều gen là một số đáng kể, có ý nghĩa quan trọng cho tiến hóa. Đột biến ảnh hưởng đến mọi tính trạng khác nhau của sinh vật và tác động theo mọi hướng.
2. Đột biến gen hay đột biến điểm
Đột biến điểm là những biến đổi rất nhỏ trên một đoạn DNA, thường liên quan đến 1 nucleotit hay 1 cặp nucleotit.
Đột biến đồng nghĩa (Samesense) còn gọi là trung tính (neutral) hay im lặng (silent), khi codon mã hóa cho một axit amin bị biến đổi, thường ở bazơ thứ ba nên vẫn mã hóa cho axit amin đó (do tính thoái hoá của mã di truyền).
Đột biến vô nghĩa (Non-sense) khi codon mã hóa cho một axit amin biến thành một trong ba codon UAA, UAG và UGA là các codon kết thúc không mã hóa cho axit amin nào.
Đột biến sai nghĩa (Mis-sense): Khi codon của axit amin này biến thành codon mã hóa cho axit amin khác, làm thay đổi axit amin tương ứng trên phân tử protein.
Đột biến lệch khung: Sự thêm 1 bazơ hay làm mất 1 bazơ dẫn đến các codon sai nghĩa hay vô nghĩa so với codon tương ứng ban đầu từ điểm biến đổi về sau, sự dịch mã bị lệch khung có tính dây chuyền từ bộ ba bị sai.
3. Đột biến nhiễm sắc thể
Các đột biến nhiễm sắc thể hay còn gọi là sai hình nhiễm sắc thể xuất hiện ở vi sinh vật nhân thực.
- Biến đổi trên 1 nhiễm sắc thể: mất đoạn, lặp đoạn, đảo đoạn.
- Biến đổi giữa các nhiễm sắc thể: chuyển đoạn.
- Đột biến bộ gen (genome mutation)
Đa bội thể (Polyploidy) hiểu theo nghĩa rộng là sự thay đổi số lượng nhiễm sắc thể gồm: Đa bội thể nguyên (Polyploidy hay Euploidy) (2n –> 3n, 4n, …), đa bội thể lai còn gọi dị bội thể (Alloploidy) (2nA + 2nB) và đa bội lệch (Aneuploidy), hay đa nhiễm: (ví dụ: 2n + 1 hoặc 2n – 1).
4. Các biến đổi vi cấu trúc
Các thay đổi thành phần nucleotit của gen.
Đột biến thay thế: Thay một nucleotit này bằng nucleotit khác.
- Đồng chuyển (Transition) khi pyrimidin được thay thế bởi pyrimidin hay purin bởi purin. Ví dụ: T thay cho C hoặc ngược lại.
- Đảo chuyển (Transversion) khi pyrimidin được thay thế bởi purin hay purin bởi pyrimidin. Ví dụ: T hay C thay cho A hoặc G và ngược lại.
Mất nucleotit (Deletion): Mất một phần nucleotit của gen.
Đột biến xen nucleotit (Insertion mutant): Thêm 1 hay nhiều nucleotit vào gen.
5. Các đột biến kiểu hình
Các đột biến hình thái: Các biến đổi ảnh hưởng đến hình dạng, màu sắc và kích thước.
Đột biến sinh hóa:
- Các đột biến khuyết dưỡng (Auxotrophe mutation) làm mất khả năng tổng hợp các chất.
- Các đột biến có điều kiện: Các đột biến có thể không có biểu hiện trong những điều kiện giới hạn nhất định (restrictive condition) và có biểu hiện trong các điều kiện cho phép (permissive condition). Ví dụ, các đột biến nhạy cảm với nhiệt độ cao có biểu hiện ở nhiệt độ tương ứng.
- Đột biến đề kháng: Các biến đổi sinh hóa giúp kháng lại được các tác nhân bất lợi.
6. Đột biến thuận và đột biến nghịch
Đột biến thuận hay trực tiếp (Direct mutation): Biến đổi từ kiểu hình hoang dại sang khác thường.
Hồi biến (Reversion): Đột biến từ kiểu hình đột biến quay về kiểu hình hoang dại.
Hồi biến thật hay đột biến nghịch. Đột biến trở lại y như ban đầu.
Ví dụ: adenin -> guanin -> adenin
Đột biến ức chế hay kìm hãm (Supressor): Đột biến ở một điểm khác. Cả hai cùng tạo kiểu hình gần như hoang dại.
Đột biến kìm hãm ngoài gen: Xảy ra ở gen khác với gen bị đột biến.
Đột biến kìm hãm trong gen: Xảy ra ở nucleotit khác trong gen đưa gen trở về trạng thái tạo kiểu hình hoang dại.
Nói chung, các đột biến là các biến đổi rất đa dạng của vật chất di truyền và có nhiều tác động khác nhau.
6.1. Cơ chế phân tử của đột biến
6.1.1. Các biến đổi trên DNA
Tất cả các đột biến đều do những thay đổi trình tự nucleotit trên DNA. Các đột biến có thể xảy ngẫu nhiên (spontaneously) hay gây tạo (cảm ứng - induced) bởi các
tác nhân gây đột biến (mutagens). Các đột biến có thể do thay đổi từng cặp bazơ hay những trình tự dài hơn.
Đột biến điểm (Point mutation) là biến đổi trình tự của một cặp bazơ Đột biến điểm có thể do chuyển đổi hoá học bazơ này thành bazơ khác hoặc do bắt cặp sai trong sao chép. Sự biến đổi gồm nhiều kiểu:
Sự đồng chuyển: thay cặp G-C bằng A-T và ngược lại.
Sự đảo chuyển: thay A-T bằng T-A và ngược lại.
– Sự bắt cặp sai (Bazơ mispairing) là sự bắt cặp không theo đúng nguyên tắc của mô hình Watson-Crick, mà là adenin với cytosin, thymine với guanin.
Xen đoạn (Insertion) là sự thêm vào một đoạn cặp bazơ vào DNA. Tăng đôi đoạn (Duplication) là một dạng đặc biệt của xen đoạn.
Mất đoạn (deletion) là sự mất đi một trình tự DNA, mà trình tự hai bên nối lại với nhau trừ trường hợp mất đầu mút nhiễm sắc thể.
Transposon hay phần tử di động là trình tự DNA có khả năng tự xen vào (hay bản sao chính nó) ở vị trí mới trên bộ gen (genome), mà không cần có quan hệ gì với locus mục tiêu. Xen đoạn là kiểu đột biến phổ biến nhất và do sự di chuyển của các phần tử di động.
Phần lớn đột biến ngẫu nhiên do sự hiện diện của các bazơ bất thường trên DNA. Ngoài ra, một số bazơ bị biến đổi (modified) như thường gặp hơn cả là 5metylcytosin, được tạo ra do enzym metylaz thêm nhóm metyl vào một số cytosin ở những điểm đặc biệt trên DNA.
6.1.2. Các sai hỏng trong sao chép DNA
Các đột biến có thể xảy ra do sai lầm khi sao chép DNA.
Mỗi bazơ tồn tại ở 2 dạng cấu trúc được gọi là tautomer. Ví dụ, adenin bình thường mang nhóm NH2 cung cấp nguyên tử hydro cho sự bắt cặp bổ sung với dạng keto (C = O - keto form) của thymine. Khi có biến đổi tautomer, adenin chuyển sang cấu trúc hiếm là dạng imino NH sẽ bắt cặp bổ sung với cytosin. Thymine có thể chuyển sang dạng enol (COH) không có trong DNA bình thường và bắt cặp với guanin.
Sự bắt cặp sai này có thể là các đột biến đồng chuyển, trong đó purin thay bằng purin khác và pyrimidin thay bằng pyrimidin khác.
Các biến đổi trên, ngoài việc thay thế các nucleotit trên mạch DNA còn có thể làm tăng thêm hay khuyết các nucleotit gây nên các kiểu đột biến ảnh hưởng đến sinh tổng hợp protein.
6.1.3. Ảnh hưởng của đột biến gen đến sinh tổng hợp protein
Đột biến lệch khung
Hai kiểu đột biến có hiệu quả nặng là thêm bazơ (addition) và mất bazơ (delection). Các biến đổi này thường làm enzym mất hoạt tính. Sự thêm 1 bazơ hay làm mất 1 bazơ dẫn đến sự dịch mã lệch khung. Từ điểm biến đổi về sau, từ bộ ba bị sai cái sai sẽ kéo dài liên tục đến cuối mạch polypeptit.
Đột biến thay thế (Bazơ substitution)
Đột biến thay thế bazơ nếu là đột biến sai nghĩa (mis-sense) sẽ có hiệu quả thay đổi từ axit amin này thành axit amin khác trong mạch polypeptit, còn nếu là đột biến vô nghĩa (non-sense) hay đột biến trung tính (hay im lặng) sẽ không ảnh hưởng đến mạch polypeptit.
6.1.4. Sai hỏng ngẫu nhiên
Ngoài các sai hỏng trong sao chép, phân tử DNA còn chịu các sai hỏng ngẫu nhiên có thể dẫn đến đột biến. Hai kiểu sai hỏng ngẫu nhiên thường gặp là mất purin (depurination) và mất amin (desamination). Mất purin là kiểu sai hỏng thường hơn, xảy ra khi liên kết glycosidic giữa C1 của pentoz với bazơ bị đứt và làm mất A hoặc G.
Sự mất amin của cytosin tạo ra uracil. Các gốc U không được sửa sai sẽ bắt cặp bổ sung với A trong sao chép, gây ra đồng chuyển G-C ->A-T. Trong các enzym sửa sai, uracil DNA-glycosylaz nhận biết đặc hiệu uracil trên DNA và cắt rời tạo lỗ hỏng, sau đó được tổng hợp lại đúng theo mạch bổ sung.
Trên phân tử DNA, một số cytosin được metyl hóa thành 5-metyl cytosin, chất này mất nhóm amin biến thành thymine. Sai hỏng này không bị uracil-DNA-glycolaz phát hiện nên không được sửa lại. Sự chuyển C - T do mất amin thường xảy ra ở các điểm có 5-metyl cytosin. Kiểu biến đổi này có ở cả vi khuẩn và tế bào sinh vật bậc cao.
6.1.5. Đột biến nhân tạo hay cảm ứng
Các tác nhân làm tăng tần số đột biến cao hơn mức tự nhiên được gọi là các tác nhân gây đột biến (mutagen). Các tác nhân vật lí như phóng xạ, tia X, tia tử ngoại gây đột biến. Nhiều hóa chất là tác nhân gây đột biến như các đồng đẳng của các bazơ nitric, HNO2 (nitrous axit), các chất alxyl hóa mạch...Các đột biến loại này được gọi là đột biến nhân tạo hay đột biến cảm ứng (induced mutation).
Đối với các vi sinh vật, các tác nhân gây đột biến chủ yếu là tia tử ngoại và một số hóa chất.
6.2. Hồi biến Quá trình đột biến, nói chung, có tính thuận nghịch, nghĩa là nếu một gen A đột biến thành a (A –> a) thì, ngược lại alen a cũng có thể đột biến quay lại thành A (a –>A). Thông thường một dạng được gọi là đột biến khi nó mang kiểu hình khác với dạng hoang dại. Hồi biến là trường hợp từ trạng thái đột biến do biến dị di truyền quay
trở về kiểu hình hoang dại. Hồi biến do đột biến nghịch (back mutation) hoặc do đột biến ức chế hay kìm hãm (supression).
6.2.1. Các đột biến nghịch
Đột biến nghịch có được khi gen đột biến có sự biến đổi quay trở lại có cấu trúc như gen hoang dại ban đầu. Trường hợp này khó xảy ra và khi lai trở lại với dòng hoang dại thì thế hệ con tất cả đều có kiểu hình hoang dại.
Ví dụ: Đột biến nghịch: m– –> m+, khi lai với dạng hoang dại cho thế hệ con đều hoang dại
Đột biến ức chế: m –su+ –> m–su –, khi lai với dạng hoang dại trong thế hệ con sẽ có một ít kiểu hình đột biến.
6.2.2. Đột biến ức chế
Đột biến ức chế (Suppressor mutation) là đột biến có tác động ngược lại hay kìm hãm của một đột biến khác. Các đột biến ức chế có những tính chất sau:
Đột biến ức chế xảy ra ở điểm khác với đột biến bị ức chế. Khi lai thể hồi biến (revertant) với dạng hoang dại sẽ xuất hiện dạng đột biến bị ức chế do tái tổ hợp làm tách rời không bị kìm hãm bởi đột biến ức chế
Đột biến ức chế có thể xảy ra trong cùng một gen, ngoài gen hoặc ở gen khác.
Các đột biến ức chế có thể thực hiện tác động bằng nhiều cách khác nhau. Ví dụ, các đột biến ức chế có thể tác động lên sự phiên mã, dịch mã hay những biểu hiện sinh lí khác của tế bào.
Đột biến kìm hãm thường gặp hơn, nó có được do 1 đột biến thứ hai làm cho biểu hiện kiểu hình của đột biến không biểu hiện ra được nên có kiểu hình hoang dại.
Đột biến kìm hãm có thể xảy ra ngay trên cấu trúc gen. Ví dụ: đột biến thuận mất một nucleotit, đột biến kìm hãm xảy ra gần chỗ đó thêm vào một nucleotit. Đột biến kìm hãm có thể do sự bổ sung trong chu trình trao đổi chất. Sai hỏng do đột biến thứ hai tạo sản phẩm bù trừ được đột biến thứ nhất.
Các biến dị ức chế được dùng để nghiên cứu sâu hơn về cơ chế dịch mã. Ví dụ, các đột biến ảnh hưởng tới anticodon của tRNA, làm thay đổi tính đặc hiệu mã hóa của nó, có thể tạo khả năng ức chế đột biến khác ở mức phiên mã. Các đột biến ức chế đối với codon vô nghĩa (nonsens-suppressor) thường xảy ra trên các tRNA, mà anticodon của nó có thể bị biến thành anticodon bổ sung với codon kết thúc do sự thay thế một nucleotit. Các nonsens - suppressor như vậy thường là trội.
Có thể xảy ra các đột biến ức chế đối với các đột biến nhầm nghĩa. Ví dụ, tRNAgly của axit amin glycine có anticodon CCC thường bắt cặp với GGG (gly) trên mRNA. Sự biến đổi đột biến của anticodon thành CUC dẫn đến chỗ tRNAgly đột biến bắt cặp với GAG (glutamic axit). Như vậy, nếu như đột biến thuận ở một gen cấu trúc nào đó biến codon GGG (glycine) thành GAG (glutamic axit) của đột biến nhầm nghĩa (missens mutation), thì sự ức chế đối với đột biến này có thể do đột biến của tRNAgly
với anticodon CUC sẽ gắn glycine vào chỗ bị đột biến (ở đột biến nhầm nghĩa là glutamic axit).
Các đột biến xảy ra trên tRNA có thể là đột biến ức chế đối với các đột biến lệch khung.
Sự ức chế ở mức phiên mã có thể xảy ra do các đột biến trên các gen mã hóa một số protein của riboxom.
VI. DI TRUYỀN HỌC Ở CÁC NHÓM VI SINH VẬT
1. Di truyền học virrut
Virut có cấu tạo đơn giản và rất nhỏ nên được phát hiện muộn hơn. Việc các nhà vật lí học khi chuyển sang lĩnh vực sinh học đã sử dụng các virut vào nghiên cứu di truyền học cho thấy rõ tầm quan trọng của các đối tượng này. Di truyền học virut đã góp phần đáng kể cho sự phát triển của sinh học phân tử. Sự hiểu biết chi tiết về các virut không những giúp con người biết rõ các cơ nguyên gây nhiều bệnh hiểm nghèo (ung thư, AIDS, ...), mà còn có thể biến chúng thành các công cụ chuyển gen có lợi cho con người
Virut được coi là những thực thể sống chưa có cấu tạo tế bào. Các virut hay các virion là những dạng sinh vật có cấu tạo đơn giản nhất. Xét từ góc độ phân loại học, sinh giới có thể chia thành hai nhóm lớn: các sinh vật có cấu tạo tế bào và virut.
1.1. Đặc điểm chung của virut
Virut có rất nhiều điểm khác vi khuẩn vơi đặc tính rất nhỏ bé, cấu tạo đơn giản. Virut nhỏ nhất có đường kính chỉ 20 nm, nhỏ hơn cả riboxom.
Ngoài ra, virut khác với các sinh vật có cấu tạo tế bào ở nhiều điểm:
Virut là các thể nội kí sinh bắt buộc, không có cấu tạo tế bào. Chúng phát triển và sinh sản chỉ trong các tế bào sống của vật chủ chuyên biệt.
Virut sử dụng bộ máy sinh tổng hợp của tế bào chủ để sinh sản và thường phá hủy tế bào chủ để phóng thích thế hệ virut con tiếp tục gây nhiễm các vật chủ mới.
Virut có bộ gen chỉ gồm một loại nucleic axit (DNA hoặc RNA), trong khi đó các tế bào có cả hai loại, được bao trong vỏ protein. Phần lớn virut gồm 1 loại nucleic axit và một ít loại protein.
Chúng không có hệ thống sinh tổng hợp protein riêng (không có riboxom); không có hệ thống trao đổi chất riêng nên không biến dưỡng năng lượng (không phân hủy thức ăn để tạo ATP), không thực hiện sự lên men, hô hấp và quang hợp.
Virut không tạo màng lipit riêng, mặc dù một số virut có màng ngoài (envelope) được tạo ra bằng biến đổi màng của tế bào chủ trước khi thoát ra khỏi tế bào chủ
Virut không bị tác động bởi các thuốc kháng sinh vì không có cấu trúc vách tế bào và bộ máy sinh hóa riboxom như tế bào vi khuẩn.
Virut không có khung xương tế bào (cytoskeleton) hoặc phương tiện di động ngoài sự khuếch tán.
Virut không “tăng trưởng” theo nghĩa là tăng khối lượng, khi virut đã hình thành nó không tăng kích thước. Hình dạng đặc trưng của virut được xác định bởi vỏ protein.
Virut đã hình thành trọn vẹn được gọi là virion, bộ gen của nó được gói trong vỏ protein và bên ngoài có thể có màng bao bọc.
1.2. Cấu tạo virut
Virut có bộ gen rất đa dạng. Bộ gen (Genome) của virut có thể là DNA mạch kép (double strand - dsDNA), DNA mạch đơn (single strand - ssDNA), RNA mạch kép (double strand - dsRNA), hay RNA mạch đơn (single strand - ssRNA). Các virut có DNA hoặc RNA mạch đơn còn phân biệt thành 2 dạng dương hay cộng (+) và âm hay trừ (–):
Virut mạch dương (Plus (positive)-strand virut): virut với bộ gen RNA hay DNA mà bộ gen có cùng trình tự nucleotit như mRNA của virut.
Virut mạch âm (Minus (negative)-strand virut): virut với bộ gen RNA mà mạch
RNA này có trình tự nucleotit ngược chiều (bổ sung với) với mRNA của virut.
Bộ gen virut thường là một phân tử nucleic axit ở dạng vòng tròn hay
thẳng. Virut nhỏ nhất có chừng 4 gen, virut lớn nhất có chừng vài trăm gen.
Vỏ protein bọc bộ gen được gọi là capsid thường có thể ở dạng hình que, hình
ống xoắn, hình đa diện hay phức tạp. Nucleocapsid là phức hợp nucleic axit và các protein của một virut. Các capsid thường được tạo nên bởi một số lớn tổ hợp các phân tử protein gồm ít loại, được gọi là capsome. Ví dụ, virut đốm thuốc lá có một capsid hình que dài cứng được tạo ra từ hơn 1000 capsome
bao đuôi bằng protein thành ống dài và phần thứ ba là sợi đuôi dài bám vào tế bào vi khuẩn khi gây nhiễm.
Tóm lại, virut được mô tả theo 4 tiêu chuẩn căn bản khác nhau:
- Bộ gen (Genome) chỉ gồm một loại phân tử là DNA hay RNA.
- Nucleic axit bộ gen là mạch đơn (single strand - ss) hay mạch kép (double strand - ds)

- Hình dạng (shape) của virion đơn giản hay phức tạp.
- Virion có hay không màng bao (membrane hay envelope).
1.3. Sao chép của các virut
Các hạt virut (virion) là những vật kí sinh nội bào bắt buộc (obligate intracellular parasite). Chúng chỉ biểu hiện các gen của chúng và sinh sản bên trong 1 tế bào sống khác. Căn cứ vào loại tế bào chủ mà virut kí sinh, người ta gọi tên loại virut: virut kí sinh trên thực vật, virut kí sinh trên động vật, vi rút kí sinh vi sinh vật. Do đặc điểm này, sự sinh sản của virut khác hẳn với sự sinh sản của tế bào. Điểm nổi bật là virut tạo ra hàng trăm hay hàng ngàn virion trong mỗi thế hệ.
Do có sự đa dạng như nêu trên, các bộ gen của virut được sao chép theo nhiều con đường khác nhau:
Các DNA mạch kép –> DNA mạch kép: kiểu bán bảo tồn.
Các RNA mạch kép –> RNA mạch kép: kiểu bán bảo tồn nhờ RNA replicaz do gen tạo enzym sao chép này được mã hóa trong bộ gen của virut RNA.
Các DNA hoặc RNA mạch đơn –> chúng làm khuôn (mạch +) để tổng hợp mạch bổ sung (mạch –) thành dạng mạch kép trung gian (+/–), gọi là dạng sao chép (replicative form), mà từ nó lấy mạch – làm khuôn tạo ra mạch đơn + giống bộ gen ban đầu của virut.
Ở Retrovirut RNA –> nhờ enzym reverse transcriptaz tổng hợp c- DNA mạch đơn từ khuôn RNA virut –> c-DNA kép –> Chèn vào bộ gen tế bào chủ –> tạo ra bộ gen RNA virut mới và mRNA.
Phụ thuộc vào bộ gen DNA hay RNA mạch kép hay đơn, mạch dương (+) hay âm (–), kiểu sao chép bộ gen và tổng hợp mRNA, các virut được chia thành 7 lớp (classes).
Hình 20: Virut đốm thuốc lá
a) Sơ đồ cấu trúc; b) Ảnh hiển vi điện tử.
Trong các loại virut, các virut nhiễm vi khuẩn có capsid phức tạp nhất. Các virut của vi khuẩn được gọi là bacteriophage (thực khuẩn thể - ăn vi khuẩn) hay gọi ngắn là phage. Các phage đầu tiên gây nhiễm E. coli được nghiên cứu mang tên T1, T2, T3...T7 (T từ chữ typ). Các phage T chẵn (T2, T4, T6) có cấu trúc rất giống nhau.
Capsid của chúng gồm một đầu đa diện (20 mặt) bọc chất di truyền. Phần thứ hai là
Một cách sao chép đặc biệt được tế bào vi khuẩn sử dụng trong tiếp hợp để truyền phân tử DNA dạng thẳng sang tế bào khác hoặc được các virut sử dụng để tạo các bộ gen của chúng, mặt khác làm đứt mạch của vòng tròn xoắn kép, tạo đầu hở 3’–OH và 5’–P kết thúc. Helicaz và SSB protein chen vào tạo chẽ 3 sao chép. Sự sao chép được thực hiện không cần mồi (primer) vì mạch 3’–OH sẵn sàng cho việc nối dài như mạch trước (leading strand) nhờ DNA polymeaz I. Đồng thời với sao chép mạch trước, mạch khuôn sau dịch chuyển. Sự dịch chuyển của mạch sau gián đoạn để tổng hợp các
đoạn ngắn Okazki như bình thường và đầu 5’ mạch khuôn duỗi thẳng ra . Sự sao chép có hình giống chữ sigma của Hi Lạp, nên được gọi là sao chép sigma. Kiểu sao chép này còn được gọi là vòng tròn quay (rolling-circle replication), vì mạch khuôn tròn ở giữa không bị đứt và quay tròn làm khuôn cho mạch trước. Sự sao chép kiểu này có thể lặp lại vài lần tạo ra sợi DNA dài, lặp lại nhiều lần bộ gen thẳng của virut, được gọi là concatemer. Enzym endonucleaz cắt ở những điểm khác nhau trên mỗi mạch của DNA tạo ra các đoạn cỡ bộ gen với hai đầu “dính”. Bộ gen thẳng này sẽ tạo thành vòng nhờ bắt cặp bổ sung các đầu “dính”.
Sự nhiễm kép haiphage
Cơ chế tái tổ hợpgen
Hình 22: Tái tổ hợp ở phage
Sự xuất hiện các dạng tái tổ hợp T2 h+r+ và T2 hr chứng tỏ 2 dòng phage T2 đã lai với nhau.
Tái tổ hợp ở virut có các đặc điểm:
Trong quá trình lai ở virut có sự tham gia của cả bộ gen như Eukaryota chứ không từng phần như ở vi khuẩn.
Hình 21: Sao chép kiểu sigma

1.4. Sự tự ráp (Self-assembly) của các virion
Các gen của virut sử dụng các enzym, chất dinh dưỡng, riboxom và các nguồn khác của tế bào chủ để tạo ra nhiều bản sao của bộ gen và các protein của các cấu phần khác (capsid, cổ, bao đuôi...). Khi các sản phẩm riêng lẻ đã tích đủ, chúng tự ráp (selfassembley) nhau thành số lượng lớn các virion rồi phá vỡ tế bào tìm các tế bào chủ mới.
Sự ráp nucleic axit của virut với các protein của capsid để hình thành virion mới diễn ra tự động tương tự các mạch polypeptit tự gắn với nhau để tạo nên các protein có cấu trúc bậc bốn. Các cấu phần được gắn với nhau bằng liên kết yếu nên không cần enzym.

1.5. Tái tổ hợp ở phage
Các phage tuy có kích thước nhỏ bé phải nhìn dưới kính hiển vi điện tử mới thấy được. Nhưng các tính trạng của phage được quan sát dựa theo các vết tan (Plaque) hoặc biên độ chủ. Vết tan (Plaque) là vùng tan hay sự ức chế tế bào gây ra bởi sự nhiễm virut của các tế bào nhạy cảm. Phage T2 có dòng hoang dại r+ tạo vết tan bình thường, còn dòng đột biến r (rapid hoặc ký hiệu r –) làm tan nhanh nên vết tan to.
Lai: T2 h+r x T2 hr+
T2 hr+, T2 h+rT2 h+r+ , T2 hr
Thế hệ phage con: Các dạng cha mẹ Các dạng tái tổ hợp
Quá trình tái tổ hợp có tính quần thể. DNA của 2 dòng phage xâm nhập vào tế bào vi khuẩn, chúng sao chép ra hàng trăm bản sao và giữa hàng trăm DNA bộ gen của phage đã xảy ra tái tổ hợp. Do điều này, ở phage thường thấy hiện tượng nhiễu âm (negative interference) trong tái tổ hợp: tần số tái tổ hợp đôi giữa 2 locus đáng lẽ phải giảm thì lại tăng cao hơn mức bình thường.
2. Di truyền học vi khuẩn
Các sinh vật Prokaryota như vi khuẩn, vi khuẩn lam cũng có các quá trình sinh sản tương đương sinh sản hữu tính, được gọi là cận hữu tính (parazxuality). Sự di truyền nhờ các quá trình cận hữu tính này ở vi khuẩn có những đặc điểm:
Truyền thông tin một chiều từ tế bào cho (donor) sang tế bào nhận(recipient).
Tạo thành hợp tử từng phần (merozygote). Thể cho (donor) chỉ chuyển một đoạn của bộ gen sang thể nhận (recipient) nên chỉ lưỡng bội ở một phần, các phần khác đơn bội.
Bộ gen thường chỉ là một DNA trần nên chỉ có một nhóm liên kết gen và tái tổ hợp thực chất là giữa hai phân tử DNA trần trong biến nạp, hay phage trong tải nạp, hoặc nhờ giao nạp khi 2 tế bào khác giới tính gắn với nhau.
Bản đồ di truyền vi khuẩn được xây dựng nhờ các phương phác khác nhau: giao nạp gián đoạn hoặc không gián đoạn, tái tổ hợp hay dùng mất đoạn.
Plasmid là những DNA vòng tròn có thể tồn tại độc lập hoặc chèn vào DNA tế bào chủ có vai trò quan trọng trong chuyển gen.
Transposition là sự chuyển vị gen gồm IS, Tn và phage Mu. Các phần tử di động có thể gây biến đổi sự biểu hiện gen.

2.1. Kiểu hình và kiểu gen của vi khuẩn
Các đột biến có thể tác động đến 5 loại kiểu hình:
Biến đổi từ nguyên dưỡng (prototrophy) sang khuyết dưỡng (auxotrophy) và ngược lại. Ví dụ mất khả năng tổng hợp một chất trao đổi của chu trình và hồi biến để lại có khả năng tổng hợp chất đó.
Sự mất hay có được khả năng sử dụng chất dinh dưỡng khác. Ví dụ: đột biến làm mất khả năng sử dụng đường lactoz.
Tính nhạy cảm hay đề kháng thuốc như nhạy cảm streptomycin đột biến thành kháng streptomycin.
Nhạy cảm với phage thành kháng phage hoặc ngược lại. Ví dụ: đột biến trên thụ thể ở bề mặt tế bào làm đề kháng với sự nhiễm phage.
Sự mất đi hoặc có lại các thành phần cấu trúc của bề mặt tế bào. Ví dụ, một loại Pneumococcus có vỏ bao polysacarit (polysacarit capsule), trong khi đó dòng khác không có vỏ bao.
2.2. Sinh sản ở vi khuẩn
Tế bào vi khuẩn phân chia theo lối trực phân. Phân tử DNA gắn trực tiếp vào màng sinh chất. Sự sao chép DNA tạo ra hai bản sao gắn chung nhau trên màng sinh chất. Khi tế bào kéo dài ra, các bản sao DNA tách xa nhau do phần màng giữ chúng lớn dần ra. Kiểu sinh sản vô tính này được gọi là “phân đôi” hay “ngắt đôi” (“binary fission”). Tế bào vi khuẩn chia nhanh (20 phút trong điều kiện tốt) hơn rất nhiều so với tế bào Eukaryota (24-48 giờ).
Quá trình sao chép DNA được bắt đầu từ điểm xuất phát oriC kéo dài về hai phía song song với quá trình sao chép màng sinh chất, nơi có điểm gắn vào của DNA bộ gen, mọc dài tách 2 phân tử DNA về 2 tế bào con.
DNA của E. coli cần 40 phút cho 1 vòng sao chép tương ứng với tốc độ 50.000 cặp bazơ/phút. Phụ thuộc vào tốc độ tăng trưởng, thời gian phân chia tế bào trong khoảng từ 18 đến 60 phút. Như vậy ở các tế bào tăng trưởng nhanh, vòng sao chép mới phải được bắt đầu sớm hơn sự phân bào trước đó như tế bào con đầu tiên.
2.3. E. coli là tế bào chủ căn bản của kỹ thuật di truyền
Các dữ liệu di truyền học của vi khuẩn mô hình - E. coli
Escherichia coli (E. coli) là vi khuẩn được nghiên cứu kĩ nhất, rất nhiều chủng khác nhau đã được phân lập. Do những thuận tiện trong nuôi cấy, nhân giống, thu nhận các đột biến và dễ phân tích các sự kiện di truyền hiếm hoi. Đến nay, nó được coi là đối tượng mô hình số một của Sinh học phân tử và công nghệ gen.
Kích thước bộ gen (Genome size): 4,6 Mb, không chứa intron, với số lượng khoảng 4000 gen, trong đó có 8% gen tương đồng với gen người, số lượng transposon khoảng 60 bản sao/bộ gen tuỳ chủng. NST là một phân tử DNA vòng tròn, dạng trần
E. coli có nhiều đóng góp trong việc nghiên cứu các cơ chế di truyền ở cấp độ phân tử tế bào nhân sơ, đồng thời cũng đóng góp cho nghiên cứu các lĩnh vực khác: Trao đổi chất của tế bào (Cell metabolism), gen ức chế vô nghĩa (Nonsense suppressors), sự tuyến tính (Colinearity) giữa gen và polypeptit, các Operon, sự đề kháng thuốc dựa vào plasmid (Plasmid-bazơd drug resistance) và sự vận chuyển tích cực (Active transport).
2.4. Các phương pháp phân tích di truyền
Ngoài các phương pháp lai để phân tích tái tổ hợp (recombination) và bổ trợ (complementation), có nhiều kỹ thuật biến đổi di truyền (Techniques of Gentic Modification):
- Gây đột biến (Mutagensis):
Hóa chất và chiếu xạ: đột biến xoma ngẫu nhiên.
Dùng transposon: xen đoạn (Insertions) xoma ngẫu nhiên.
- Chuyển gen (Transgensis):
Trên vector plasmid: tự do hay chèn vào (integratd).
Trên vector phage: tự do hay chèn vào (integratd).
Biến nạp: chèn vào.
- Làm im lặng gen mục tiêu (Targeted gen knockout):
Alen không (Null alen) trên vector: thay gen bằng tái tổ hợp.
Alen được thiết kế (Engineered alen) trên vector: đột biến điểm định hướng (Site-directed mutagensis) bằng thay gen.
2.5. Quá trình tổng hợp axit nucleic
Các enzym và protein tham gia tổng hợp và phân giải axit nucleic
Enzyme Gen mã hoá Chức năng
DNA polymeaz III polC; DNAE,Q,N,X
Enzym polyme hoá chủ yếu
DNA polymeaz I polA-E, mutD, pol A Cắt mồi RNA, lắp chỗ trống
Helicaz DNAB Tháo xoắn ở chẻ ba sao chép
Primaz DNAG Tạo mồi mạch DNA mới
Protein gắn điểm khởi đầu (Origin
DNAA Gắn điểm khởi đầu sự sao chép (Ori), tạo thuận lợi cho mở tách mạch
binding)
Protein căng
mạch SSB (Singlestrand binding protein)
Ssb Ngăn các mạch đơn đã tách không chập lại
AND ligaza ligA, lig B Nối các đầu hở trên DNA
Bảng 3: Các enzym tham gia sao chép DNA ở vi khuẩn E. coli.

Enzyme Gen mã hoá Chức năng
DNA polimeraza I polC; DNAE,Q,N,X
DNA polimeraza II polA, mutD
Enzyme phục hồi chủ yếu, cắt mồi RNA và lấp kín khoảng trống
Enzyme phục hồi phụ (minor)
DNA polimeraza III polB Replicaz, polyme hoá chủ yếu
DNA polimeraza IV polC Phục hồi cấp cứu (SOS repair)
DNA polimeraza V DNAB , mutD, C Phục hồi cấp cứu (SOS repair)
Bảng 4: Các enzyme DNA polymeraza ở vi khuẩn E. coli
Các enzyme tham gia vào sự phân giải axit nucleic:
Deoxyribonucleaz (DNAaz) là enzym cắt các liên kết trong phân tử DNA. Nó có thể cắt mạch đơn hay kép. Gyraz là một loại enzym topoixomraz cắt DNA làm tháo xoắn.
Ribonucleaz (RNAaz) là các enzym cắt RNA, có thể đặc hiệu đối với RNA mạch đơn hay mạch kép.
Các nucleaz chia thành 2 nhóm:
Exonucleaz cắt từng nucleotit một từ đầu mút của mạch polynucleotit; chúng có thể đặc hiệu đối với đầu mút 5’ hay 3’ của DNA hay RNA.

- Endonucleaz cắt các liên kết bên trong mạch DNA; chúng có thể đặc hiệu đối với RNA hay DNA mạch đơn hoặc mạch kép.
2.6. Các cơ chế di truyền ở vi khuẩn
Trao đổi thông tin di truyền ở vi khuẩn: biến nạp, tải nạp và tiếp hợp khác nhau
ở các phương tiện để đưa DNA từ tế bào cho vào tế bào nhận.
2.6.1. Biến nạp
Hiện tượng biến nạp được Griffith phát hiện ở vi khuẩn Diplococus pneumoniae (nay gọi là Streptococus pneumoniae - phế cầu khuẩn gây sưng phổi ở động vật có vú) vào năm 1928. Phát hiện này và các nghiên cứu về cơ chế biến nạp có ý nghĩa lịch sử cho sự ra đời của Sinh học phân tử.
Vi khuẩn này có 2 dạng khác nhau:
– Dạng SIII, gây bệnh có vỏ bao tế bào (capsule) bằng polysaccharid cản trở bạch cầu phá vỡ tế bào. Dạng này tạo khuẩn lạc trơn (Smooth) trên môi trường agar.
– Dạng RII, không gây bệnh, không có vỏ bao, tạo đốm mọc nhăn (Roughnhăn).
Thí nghiệm tiến hành như mô tả trên hình:
Hình 23: Hiện tượng biến nạp.
Tiêm vi khuẩn S sống gây bệnh cho chuột - chuột chết.
Tiêm vi khuẩn R sống không gây bệnh - chuột sống.
Tiêm vi khuẩn S bị đun chết cho chuột - chuột sống.
Hỗn hợp vi khuẩn S bị đun chết trộn với vi khuẩn R sống đem tiêm cho chuộtchuột chết. Trong xác chuột chết có vi khuẩn S và R.
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các tế bào chết này đã truyền tính gây bệnh cho tế bào R. Nó được gọi là biến nạp (transformation). Năm 1944, T. Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng proteaz (enzym phân hủy protein) hoặc RNA-az (enzym phân hủy RNA) hoạt tính biến nạp vẫn còn, chứng tỏ protein và RNA không phải là tác nhân gây biến nạp. Nhưng nếu tế bào S chết bị xử lý bằng DNA-az (enzym chỉ phân hủy đặc hiệu DNA) thì hoạt tính biến nạp không còn nữa, chứng tỏ DNA là nhân tố biến nạp mang tín hiệu di truyền.
Như vậy, biến nạp là hiện tượng truyền thông tin di truyền bằng DNA. Trong biến nạp, DNA trần từ một tế bào vi khuẩn (thể cho) này được truyền sang tế bào vi khuẩn khác (thể nhận). Biến nạp xảy ra khi vi khuẩn nhận DNA ngoại lai và hấp thu vào trong tế bào. Khi tế bào vi khuẩn bị vỡ do bị tan (lysis), DNA vòng tròn của chúng thoát ra môi trường thành các đoạn thẳng với chiều dài khác nhau, có khả năng gây biến nạp cho các tế bào nhận khác.
Biến nạp được nghiên cứu kĩ nhất ở các vi khuẩn Streptococcus pneumoniae, Bacillus subtilis, Haemophilus influenzae và ở một số nhóm vi khuẩn khác.
Hiệu quả của biến nạp phụ thuộc vào 3 yếu tố:
Tính dung nạp của tế bào nhận.
Kích thước của đoạn DNA ngoại lai.
Nồng độ của DNA
Cơ chế phân tử
Quá trình biến nạp ở cấp độ phân tử gồm các giai đoạn chủ yếu:
Sự phân hủy DNA tế bào cho: DNA tế bào cho có thể là của tế bào tự nhiên bị phân hủy hoặc trong thí nghiệm gây tác động bằng nhiệt độ cao hay tác nhân phá vỡ tế bào.
DNA bám vào bề mặt tế bào: Protein gắn vào DNA.

Thâm nhập của DNA: Sợi DNA mạch kép của dòng vi khuẩn S sau khi chui qua màng tế bào của dòng R thì một mạch S sẽ bị nucleaz của tế bào cắt, còn lại một mạch nguyên (thí nghiệm của Griffith).
Bắt cặp (Synapsis)và tái tổ hợp: Nhờ sự hỗ trợ của protein RecA DNA của thể nhận R sẽ biến tính tách rời 2 mạch ở 1 đoạn dễ bắt cặp với đoạn DNA thể cho S vừa chui vào
Hình 24: Sơ đồ diễn biến của quá trình biến nạp ở cấp độ phân tử a. DNA bám vào. b. Thâm nhập. c. Bắt cặp và tái tổ hợp. d. – Tế bào biến nạp Sao chép: Sau khi bắt cặp tạo đoạn lai R - S, phân tử DNA sao chép tạo ra 2 sợi, một sợi kép R-R và sợi kép khác có mang đoạn DNA thể nhận S-S. Kết quả cuối cùng là đoạn gen của SIII chèn vào bộ gen tế bào nhận. Sau phân bào thì một dòng tế bào nhận được DNA ngoại lai vào bộ gen - tế bào được biến nạp. Tế bào đã được biến nạp sinh sản tạo dòng nhận RII mới.
2.6.2. Tải nạp (Transduction)
Việc tìm ra biến nạp đã thúc đẩy nghiên cứu dẫn đến phát minh ra tải nạp, là hiện tượng truyền DNA qua trung gian virut từ tế bào cho đến tế bào nhận. Có hai kiểu tải nạp: chung và chuyên biệt.
Trong tải nạp (transduction), các virut có chức năng như vật trung gian để đưa DNA từ vi khuẩn cho vào vi khuẩn nhận bằng cách lây nhiễm. Ở chu trình tan (lytic cycle), một số bacteriophage gói nhầm DNA vi khuẩn chủ vào capsid. Tế bào bị nhiễm bởi các virut như vậy nhận đoạn DNA của vi khuẩn A khác, chứ không phải DNA của virut. Do vậy, DNA vi khuẩn A tái tổ hợp với DNA nhiễm sắc thể của tế bào chủ B và biến đổi thành phần di truyền.
Phage là nhân tố chuyển gen
Thí nghiệm được tiến hành trong ống hình chữ U, ở đáy ống được ngăn cách bằng màng lọc vi khuẩn. Màng có lỗ nhỏ vi khuẩn không qua được, nhưng phage qua được (hình 25). Nhánh A của ống chứa vi khuẩn có khả năng tổng hợp tryptophan (trp+), còn nhánh B nuôi các vi khuẩn mất khả năng tổng hợp tryptophan (trp–). Sau
một thời gian nuôi bên nhánh B xuất hiện vi khuẩn có khả năng tổng hợp tryptophan (trp+). Qua nhiều lần thí nghiệm việc phage tải gen trp+ từ nhánh A sang nhánh B được chứng minh.
Hình 25: Thí nghiệm chứng minh có tải nạp do virut
a. Tải nạp chung (Genral transduction)
Tải nạp chung xảy ra khi phage mang bất kì gen nào của vi khuẩn A chuyển sang vi khuẩn B.
Tải nạp chung (genralized transduction) có các đặc điểm:

- Thường do phage kiểu P1 thực hiện.
- Bất kì gen nào của vi khuẩn cũng đều được tải nạp.
- Tải nạp có được do gói nhầm DNA của tế bào chủ khi phage trưởng thành.
- Các thể tái tổ hợp đơn bội được tạo ra.
Do không có sự tương đồng giữa trình tự DNA trên các phage này với trình tự ở tế bào chủ, nên không có điểm gắn vào đặc hiệu cho prophage. Bất kì gen nào cũng được tải nạp vì đầu của phage có thể gói nhầm vào một đoạn DNA của tế bào chủ. Sự đồng tải nạp (cotransduction) là quá trình tải nạp đồng thời 2 gen.
Quá trình phage xâm nhập vào vi khuẩn và sinh sản được mô tả trên hình 26 Đầu tiên phage bám trên bề mặt của vi khuẩn, sau 4 phút bơm DNA của nó vào tế bào, sau đó chúng sinh sản và độ nửa giờ sau thì làm tan vi khuẩn và giải phóng các phage con mới.
Hình 26: DNA phage Nhiễm sắc thể vi khuẩn và


Phage mới phá vỡ tế bào
Quá trình xâm nhập của phage và làm tan vi khuẩn
Khi DNA của phage xâm nhập tế bào vi khuẩn A, chúng cắt DNA của vi khuẩn A thành nhiều đoạn, đồng thời DNA của phage được sao chép thành nhiều phân tử con và các vỏ capsid của phage cũng được tạo thành. Sau đó các vỏ capsid được lắp ruột DNA vào, phá vỡ tế bào vi khuẩn ra ngoài và tiếp tục xâm nhập các vi khuẩn mới. Trong quá trình lắp ráp khoảng 1-2% phage, vô tình mang đoạn DNA của vi khuẩn có chứa gen. Phage mang gen của vi khuẩn A xâm nhập vi khuẩn B, quá trình tái tổ hợp xảy ra làm gen A gắn vào bộ gen B.
b. Tải nạp chuyên biệt (Special transduction)
Tải nạp chuyên biệt hay hạn chế (restricted transduction) là trường hợp chỉ mang một vài gen nhất định, nó có 4 đặc điểm:
- Những gen được chuyển nằm sát chỗ prophage gắn vào.
- Chỉ prophage lamda thực hiện.
- Do kết quả sự cắt sai của prophage khi tách khỏi nhiễm sắc thể của tế bào chủ.
- Các vi khuẩn tái tổ hợp có thể lưỡng bội một phần. Ví dụ: Phage chỉ mang gen gal (đồng hóa đường galactoz) từ vi khuẩn này chuyển sang vi khuẩn khác.
Hình 27: Điểm gắn của phage vào bộ gen vi khuẩn
Điểm gắn của phage vào bộ gen của vi khuẩn nằm giữa 2 gen gal (galactoz) và bio (tổng hợp biotin), tiếp theo xảy ra quá trình tái tổ hợp ở điểm chuyên biệt (sitespecific recombination) chèn DNA của phage vào nhiễm sắc thể vi khuẩn Đầu của phage chỉ có thể chứa một lượng DNA giới hạn, nó chỉ tải nạp được gen gal hoặc bio. Phage tải nạp các gen galactoz được gọi là gal hay dg (d = defective: khuyết, g = galactoz). Nếu tế bào gal– được nhiễm bởi dg (mang gen gal+), sự ráp phage biến dạng vào tế bào chủ sẽ tạo lưỡng bội một phần. Sự cắt sai của phage rất hiếm nên tải nạp hạn chế có tần số thấp.
Tuy nhiên, tải nạp tần số cao có thể nhận được trong điều kiện thí nghiệm. Nếu tế bào vi khuẩn được gây nhiễm kép với phage hoang dại và phage dg, phage hoang dại có thể hỗ trợ chức năng sai sót ở phage biến dạng, và thế hệ con sẽ có cả 2 kiểu với số lượng bằng nhau. Khi dịch tan được dùng tải nạp, quá trình được gọi là tải nạp tần số cao.
Trong nhiều trường hợp, do bộ gen biến dạng dg không gắn được vào bộ gen của tế bào chủ (nên không sao chép được). Sau mỗi lần phân bào, chỉ một trong 2 tế bào có bộ gen của phage biến dạng; quá trình này được gọi là tải nạp sẩy (abortive transduction).
2.6.3. Giao nạp (Conjugation)
Giao nạp ở vi khuẩn là sự kết hợp nhất thời của hai tế bào có kiểu bắt cặp đối nhau, được tiếp nối bằng sự chuyển một phần hoặc toàn bộ vật chất di truyền từ tế bào cho sang tế bào nhận qua cầu tế bào chất, và sau đó các tế bào tách nhau ra (exconjugants). Giao nạp đòi hỏi sự tiếp xúc trực tiếp giữa 2 loại tế bào được khởi sự bằng ống giao nạp hay tính mao (pilus), một sợi ống nhỏ rất dài do tế bào cho (donor cell) tạo ra.
Khả năng cho nhận được xác định bởi các plasmid liên hợp cụ thể được gọi là plasmid khả năng sinh sản hoặc plasmid giới tính.
Plasmid F (còn gọi là yếu tố F) của E. coli là nguyên mẫu cho plasmid khả năng sinh sản ở vi khuẩn Gram âm. Các chủng E. coli có plasmid F ngoài nhiễm sắc thể được gọi là F+ và có chức năng là người cho, trong khi các chủng không có plasmid F là F– và hoạt động như người nhận. Các chức năng liên hợp của plasmid F được quy định bởi một cụm gồm ít nhất 25 gen chuyển xác định sự biểu hiện của F pili, tổng hợp và chuyển ADN trong quá trình giao phối, can thiệp vào khả năng của vi khuẩn F+ để phục vụ như là người nhận, và các chức năng. Mỗi vi khuẩn F+ có 1 đến 3 F pili liên kết với một protein màng ngoài cụ thể (sản phẩm của gen ompA) trên vi khuẩn nhận để bắt đầu giao phối.

DNA được truyền qua một cầu tế bào chất mỏng (a thin cytoplasmic bridge) gọi là ống giao nạp (conjugation tube). Kiểu sao chép sigma được tế bào vi khuẩn sử dụng trong giao nạp để truyền phân tử DNA dạng thẳng sang tế bào khác. Khi đoạn DNA vào bên trong tế bào nhận (recipient cell), nó thực hiện tái tổ hợp với các gen tương đồng (homologous gens).
2.6.4. Hiện tượng lai ở vi khuẩn
Vào năm 1946, J. Lederberg và E. Tatum đã sử dụng các dòng đột biến khuyết dưỡng khác nhau ở E. coli để chứng minh có tái tổ hợp giữa các dòng vi khuẩn khác nhau. Cụ thể dòng A có kiểu gen met–bio–thr+leu+thi+ (có khả năng tổng hợp threonin, leuxin và vitamin B1 (thiamin) và không tổng hợp được metionin và biotin), còn ở dạng B thì ngược lại có khả năng tổng hợp metionin và biotin với kiểu gen met+bio+thr–leu- thi-. Trộn A và B trong ống nghiệm, sau đó cấy lên môi trường tối thiểu (Minimal medium - MM). Các khuẩn lạc mọc trên môi trường tối thiểu chứng tỏ có các dạng lai, chúng chỉ mọc lên được nhờ sự bù đắp cho nhau các nhu cầu dinh dưỡng. Dạng lai có kiểu gen met+bio+thr+leu+thi+ mọc được trên môi trường tối thiểu.
Trong khi đó từng dạng A hoặc B riêng lẻ không mọc được trên MT tối thiểu
Dòng A = met–bio–thr+leu+thi+ x dòng B= met+bio+thr–leu-thi-

Dạng lai met+bio+thr+leu+thi+ mọc thành khuẩn lạc, mỗi dạng riêng lẻ không mọc thành khuẩn lạc.
Hình 29: Sơ đồ lai vi khuẩn (MM: môi trường tối thiểu)
2.6.5. Tái tổ hợp
Ở vi khuẩn có yếu tố giới tính tương tự giống đực và cái ở sinh vật bậc cao: F+ (tương tự giống đực), nó truyền gen sang F- (giống cái). Tần số lai F+ x F- vào khoảng 10-6, tức là lai 1 triệu tế bào thì sẽ có 1 tế bào lai. Khi tiếp xúc với F+ một thời gian tế bào Fsẽ trở thành F-. Dạng Hfr (High frequency of recombination) được phát hiện sau, dạng này có tần số lai với F- cao hơn F+ có thể lên tới 104 lần.
Hình 28: Hai tế bào vi khuẩn giao nạp qua cầu tế bào chất pilus
Plasmid là phân tử DNA vòng tròn nhỏ có khả năng tồn tại độc lập hoặc gắn vào bộ gen vi khuẩn. Plasmid có thể mang một số gen khác nhau như gen kháng kháng sinh (plasmid R), ... Bản chất di truyền của các dòng F+, F- và Hfr được xác định do các plasmid như sau:
F- không chứa plassmid
F+ chứa plasmid ở dạng độc lập
Hfr- plasmid được gắn vào bộ gen của vi khuẩn
Muốn xảy ra tái tổ hợp thì 2 dòng vi khuẩn phải tiếp xúc với nhau (F+ x F-) hoặc (Hfr x F-). Dòng tế bào mang nhân tố F+ được coi là tế bào “đực” và có khả năng tạo protein pilin, từ protein này tạo ống giao nạp là pilus. Sự co lại của pilus đang nối hai tế bào làm chúng tiếp xúc kề nhau. Tế bào F- được coi là cái (female). Sau khi giao nạp tế bào F– trở thành F+
Việc chuyển gen chỉ thực hiện khi plasmid gắn vào bộ gen của vi khuẩn. Trong quá trình chuyển vật chất di truyền sang F- thì DNA của tế bào chủ sao chép và mạch mới có ori đi đầu và F ở cuối Quá trình chuyển DNA từ F+ sang F– có thể bị ngắt quãng.
2.6.6. Lập bản đồ di truyền nhiễm sắc thể của vi khuẩn
a. Giao nạp ngắt quãng (Interrupted conjugation)
Khi trộn dịch tế bào Hfr với F–, sự tiếp hợp có thể bị ngắt quãng vào thời điểm
tùy ý bằng rung lắc mạnh. Mẫu được pha loãng nhanh và cấy lên môi trường chọn lọc. Ngoài ra, dòng Hfr phải có gen đánh dấu hay khuyết dưỡng để chúng không mọc được trên môi trường chọn lọc và chỉ có các dạng tái tổ hợp mọc lên. Sự truyền DNA của Hfr sang F– có tính phân cực, các gen ở đầu sang trước và lần lượt các gen khác theo sau. Nhờ ngắt quãng có thể xác định được thời gian mà một gen được truyền sang F–và khoảng cách giữa chúng.
Ví dụ, dòng Hfr có mang các gen nguyên dưỡng (prototrophic) đánh dấu a+, b+, c+ được trộn với dòng F– mang các alen khuyết dưỡng a, b, c. Sự tiếp hợp được ngắt quãng với khoảng cách 5 phút và pha loãng cấy lên môi trường chọn lọc (ví dụ, môi trường tối thiểu). Kết quả như sau:

Thời gian (phút) Các thể tái tổhợp
5 a b+c 10 a b+ c+
15 a+ b+ c+
Bảng 5: Kết quả của sự tiếp hợp ngắt quãng
Hình 30: Sự truyền DNA từ thể cho sang thể nhận
Hfr X F– Sao chép DNA truyền cho F– Hfr F– tái tổ hợp
Các gen A, B, C được chuyển một chiều từ Hfr sang F– Đọan gen thể cho bắt cặp với các gen tương đồng của DNA tế bào F– và diễn ra trao đổi chéo tạo tế bào F–tái tổ hợp
Dòng Hfr có tần số lai cao hơn nhiều vì plasmid đã nằm sẵn trong bộ gen. Còn F+ phải qua giai đoạn plasmid gắn vào bộ gen rồi mới chuyển gen.

Trong điều kiện thí nghiệm ở 37oC, nguyên bộ gen của tế bào E. coli được chuyển sang tế bào nhận trong vòng 90 phút. Thường sự giao nạp bị ngắt giữa chừng do các pilus bị gãy, nên ít khi bộ gen được chuyển nguyên vẹn vào tế bào nhận. Lúc đó tế bào F– vẫn là F–.
Trình tự gen ở dòng Hfr là b+ – c+ – a+; b cách đầu mút truyền sang ít hơn 5 phút; c ít hơn 10 phút; a ít hơn 15 phút. Ở đây đơn vị thời gian là phút.
b. Giao nạp không ngắt quãng (Uninterrupted conjugation)
Nếu không rung mạnh để ngắt quãng nhân tạo, cầu tế bào chất giữa hai gen càng gần ori (ở đầu của DNA thể cho), xác suất được chuyển sang tế bào nhận càng lớn. Tần số của các gen xuất hiện ở dạng tái tổ hợp trong thể nhận, được phát hiện trên môi trường chọn lọc, tỉ lệ nghịch với khoảng cách so với gen đánh dấu ở đầu. Các gen có nhiều dạng tái tổ hợp xuất hiện thì càng nằm gần phía đầu được chuyển sang tế bào nhận.
Khoảng cách giữa các gen có thể tính bằng 3 loại đơn vị: đơn vị thời gian, đơn vị tái tổ hợp và đơn vị cặp bazơ.
Ví dụ: Nếu một phút giao nạp tương đương 20 đơn vị tái tổ hợp ở E. coli, và nguyên nhiễm sắc thể (DNA) được chuyển sang trong 100 phút, thì tổng chiều dài là 2000 đơn vị tái tổ hợp. Nếu DNA có 107 cặp nucleotit, thì 1 đơn vị tái tổ hợp bằng 107/2000 = 5000 bp.
Hình 31: Đoạn DNA từ thể cho trao đổi chéo với DNA thể nhận tạo F– tái tổ hợp
2.6.7. Lập bản đồ bằng tái tổ hợp
Đặc điểm của tái tổ hợp ở vi khuẩn là chỉ một phần bộ gen được chuyển sang tế bào nhận. Một hay nhiều gen có thể được gắn vào nhiễm sắc thể tế bào nhận nhờ giao nạp và phụ thuộc độ dài đoạn truyền sang. Exogenote phải được gắn vào nhiễm sắc thể nhận thì mới được sao chép và chia về các tế bào trong dòng. Chỉ một đoạn ngắn DNA thường được gắn nhờ biến nạp và tải nạp. Như vậy, nếu hai gen cùng biến nạp (cotransformation) thì hai locus phải liên kết chặt. Tương tự như vậy, hai gen cùng
được tải nạp vào phage sẽ ở gần nhau. Mức độ liên kết giữa các gen có chức năng khác nhau (intercistronic) hay giữa các đột biến bên trong gen chức năng (intracistronic) có thể đánh giá từ các kết quả lai chuyên biệt.
Ở hệ thống hợp tử một phần (merozygote), exogenote muốn gắn vào endogenote phải xảy ra 2 trao đổi chéo ở hai đầu vì nếu chỉ có 1 trao đổi chéo thì cả hai sản phẩm nhận được đều mất cân bằng.
Trong các hệ thống này, tổng cá thể tham gia tái tổ hợp không biết được, nên tần số tái tổ hợp không thể tính như ở ruồi giấm (trên tổng số cá thể). Do đó, tần số tái tổ hợp được trình bày tương đối theo một số tiêu chuẩn chung cho tất cả các tổ hợp lai. Ví dụ, số lượng các cá thể tái tổ hợp nguyên dưỡng được tạo ra khi lai hai dòng đột biến có so sánh với kết quả khi lai dạng hoang dại với mỗi loại đột biến. Tuy nhiên có nhiều sai lệch và cần chỉnh lí.
Xác định trình tự gen ở vi sinh vật thường phức tạp vì có thể xảy ra hiện tượng nhiễu âm cục bộ (localized negative interference). Phương pháp xác định trình tự gen thường dùng là lai 3 điểm hoán đổi (three - factor reciprocall crosses).
nhận biết (recognition sites) của enzym cắt hạn chế (RE – restriction endonucleaz) có tên Not 1. Các mũi tên chỉ điểm khởi sự của các dòng Hfr.
2.6.8. Plasmid

Trong phần nói về quá trình giao nạp (conjugation) hay tiếp hợp ở trên đã đề cặp đến plasmid, là những DNA vòng tròn nhỏ hiện diện ở nhiều vi khuẩn. Plasmid là nhân tố di truyền sao chép độc lập với nhiễm sắc thể (Nhiễm sắc thể) của tế bào chủ. Không giống virut, plasmid không có dạng ngoại bào (extracellular form) và tồn tại trong tế bào đơn giản là DNA tự do và điển hình là dạng vòng tròn. Plasmid và DNA Nhiễm sắc thể của tế bào có vài điểm khác nhau cơ bản: plasmid chỉ mang những gen không thiết yếu (nhưng thường rất có ích), còn những gen thiết yếu nằm trên Nhiễm sắc thể

Plasmid di chuyển giữa các tế bào trong quá trình giao nạp (conjugation), tức sự tiếp xúc giữa hai loại tế bào.
Các kiểu plasmid khác nhau được phân lọai dựa vào các gen mà chúng mang. Một số plasmid, các nhân tố trao đổi chất (metabolic factors) mang các gen mã hóa cho các chức năng trao đổi chất khác thường (unusual metabolic functions) như phân hủy dầu mỏ do tràn dầu.
Đến nay đã biết hàng ngàn plasmid khác nhau. Trên thực tế, người ta đã phân lập hơn 300 plasmid khác nhau trong tự nhiên chỉ từ dòng E. coli duy nhất. Nhiều plasmid được sử dụng làm công cụ chuyển gen trong kỹ thuật di truyền.
Sự chuyển plasmid từ tế bào này sang tế bào khác
Cơ chế chính của việc chuyển plasmid từ tế bào này sang tế bào khác là giao nạp, mà chức năng thực hiện được mã hóa bởi chính một số plasmid. Giao nạp là một quá trình sao chép DNA và plasmid của tế bào cho và truyền bản sao của Nhiễm sắc thể cùng bản sao plasmid sang tế bào nhận.
Bản đồ gen của plasmid F, một plasmid đã được nghiên cứu rất kĩ của Escherichia coli. Khoảng 25 gen trên F plasmid, bao gồm các gen sao chép DNA, các yếu tố thực hiện chức năng của một epixom và cuối cùng là một vòng cung DNA lớn, gọi là vùng tra, chứa những gen giúp nó tự di chuyển từ tế bào này sang tế bào khác.
Hình 32: Bản đồ di truyền E. coli dòng K12
(theo Brock Microbiology 11,2006)
OriC - điểm khởi sự sao chép; RE – restriction endonucleaz Nost1
Bản đồ di truyền liên kết gen vòng tròn của nhiễm sắc thể ở E. coli dòng K12 (hình 32). Bộ gen của E. coli đã được giải ký tự chuỗi (sequencing) năm 1997, nó gồm 4.639.221 bp (cặp bazơ) với khoảng 4288 ORF (khung đọc mở). Vòng ngoài cùng ghi ký hiệu một số gen, mà mặt trong các số từ 0 đến 100 chỉ vị trí tính theo số phút gen được chuyển qua. Các số ở vòng trong cùng chỉ số kilo cặp bazơ (kbp) tại các điểm
Hình 33: Sơ đồ gen của plasmid F của E. coli.Plasmid điều khiển sự truyền gen của chính nó bởi sự tiếp xúc của hai tế bào gọi là có khả năng giao nạp. Không phải tất cả plasmid đều có khả năng tiếp hợp. Khả năng truyền trong tiếp hợp được kiểm soát bởi những gen trong plasmid gọi là vùng tra. Vùng tra chứa những gen mã hóa protein thực hiện chức năng bắt cặp. Sự hiện diện của vùng tra ở plasmid có thể dẫn đến kết quả quan trọng khác nếu plasmid chèn vào nhiễm sắc thể. Trong trường hợp này, plasmid có thể huy động sự chuyển DNA nhiễm sắc thể từ tế bào này sang tế bào khác.
Một vài plasmid tiếp hợp từ Pseudomonas có biên độ tế bào chủ lớn. Điều này có nghĩa là chúng có thể được truyền sang rất nhiều vi khuẩn Gr+ khác. Những plasmid như thé có thể chuyển thông tin di truyền giữa những sinh vật ở gần nhau. Những plasmid tiếp hợp có thể chuyển giữa vi khuẩn Gr- và Gr+, giữa vi khuẩn và tế bào thực vật và giữa vi khuẩn và nấm. Thậm chí nếu plasmid không thể sao chép độc lập trong vật chủ mới, sự chuyển plasmid tự nó có thể có ý nghĩa tiến hóa quan trọng nếu gen từ plasmid nối lại với bộ gen của vật chủ mới.
Vài loại plasmid khác và ý nghĩa sinh học của chúng
Mặc dù plasmid không mang những gen thiết yếu đối với vật chủ, nhưng nó có thể ảnh hưởng đáng kể đến kiểu hình của tế bào. Chẳng hạn, khả năng của Rhizobium tương tác với thực vật và hình thành nốt sần ở rễ cố định đạm là nhờ những plasmid thực hiện. Những plasmid khác mang lại đặc điểm biến dưỡng đặc biệt ở tế bào, như sự phân hủy chất gây ô nhiễm độc hại. Thực ra, plasmid dường như là một bộ máy chính để tạo ra những tính chất riêng biệt của vi khuẩn, và trong nhiều trường hợp, còn “xuất khẩu” (export) những đặc tính này bởi sự chuyển gen theo chiều ngang. Hạn chế duy nhất đối với những kiểu gen này có trong plasmid là chúng không can thiệp vào sự sao chép của chính mình hoặc vào sự tồn tại của vật chủ.
Plasmid đề kháng
Trong số những nhóm plasmid phổ biến nhất và được nghiên cứu kĩ nhất là các plasmid R, có khả năng kháng kháng sinh và nhiều chất ức chế sinh trưởng khác. Bản chất gây nhiễm của plasmid tiếp hợp R cho phép chúng lan rộng nhanh chóng trong quần thể tế bào gây ra một vấn nạn lớn trong y học lâm sàng là sự xuất hiện nhanh các vi khuẩn đề kháng thuốc.
Một plasmid R có thể mang nhiều gen đề kháng. Chẳng hạn, plasmid R100 có 94,3 kbp mang những gen mã hóa cho sự kháng sulfonamid, streptomycin và spectinomycin, fusidic axit, chloramphenicol, tetracylin và có thể một số gen kháng thủy ngân (mercury). Plasmid R100 có thể được chuyển giữa các vi khuẩn đường ruột thuộc chi (Genra) Escherichia, Klebsiella, Proteus, Salmonella và Shigella, có thể chuyển vào vi khuẩn

Gram âm Pseudomonas. Những plasmid R khác có gen kháng kháng sinh cũng
Hình 34: Plasmid R100: RTF là vùng các gen tra chuyển gen; r-determinant là vùng các gen đề kháng (mer - thủy ngân, sul - sulfamid, str - streptomycin, cmlchloramphenicol, tet – tetracyclin).
Plasmid mã hóa chất độc và những độc tính khác
Ở một số vi khuẩn gây bệnh, một số tính chất này được mang trên plasmid. Thí dụ, dòng E. coli gây bệnh đường ruột có tính chất này bởi khả năng sinh sản thành tập đoàn trong ruột non và sản xuất một chất độc gây các triệu chứng của bệnh tiêu chảy. Sự hình thành tập đoàn đòi hỏi sự hiện diện của một protein bề mặt tế bào gọi là yếu tố kháng nguyên tạo quần thể (yếu tố kháng nguyên hình thành tập đoàn), được mã hóa bởi một plasmid. Protein này giúp tế bào có khả năng gắn với tế bào biểu mô của ruột. Ít nhất hai chất độc ở E. coli gây bệnh đường ruột được mã hóa bởi một plasmid: hemolysin làm tan tế bào hồng cầu và enterotoxin gây tiết quá nhiều nước và muối vào ruột và do vậy là chất độc gây tiêu chảy.
Bacteriocin
Nhiều vi khuẩn sản xuất protein ức chế hoặc giết chết tế bào các loài họ hàng gần, thậm chí ngay cả những dòng khác nhau của cùng một loài. Những tác nhân này, được gọi là bacteriocin để phân biệt với chất kháng sinh, có phổ tác động hẹp hơn kháng sinh. Các gen mã hóa bacteriocin và những protein chế biến (processing) và vận chuyển chúng (tạo tính miễn dịch cho sinh vật sản sinh ra nó) thường nằm trên plasmid hay transposon. Bacteriocin được đặt tên theo loài tạo ra nó như ở E. coli có colicin, mã hóa bởi Colplasmid; Bacillus subtilis tạo ra subtilisin, ....
Plasmid Col của E. coli mã hóa nhiều colicin khác nhau. Colicin được giải phóng từ một tế bào gắn với những thụ thể (receptors) đặc hiệu trên màng tế bào dễ cảm nhiễm (susceptible). Colicin giết tế bào bằng cách ngăn cản chức năng quan trọng nào đó của tế bào.
Bacteriocin hoặc tác nhân tương tự bacteriocin của vi khuẩn Gram dương thì hơi khác colicin những cũng thường được mã hóa bởi plasmid, một số thậm chí còn có giá trị thương mại. Chẳng hạn, vi khuẩn lactic tạo ra bacteriocin Nisin A ức chế mạnh sự tăng trưởng của một loạt vi khuẩn Gr+ và được dùng làm chất bảo quản trong công nghệ thực phẩm.
2.6.9. Sự chuyển vị (Transposition)
Các phần tử chuyển vị (transposable elements) hay phần tử di động (mobile elements) là những trình tự DNA có khả năng di chuyển từ vị trí này đến vị trí khác (mục tiêu) của bộ gen (cùng bộ gen hay khác). Sự di chuyển DNA này được gọi là transposition (sự chuyển vị).
Tái tổ hợp (Recombination) trong chuyển vị (transposition) là sự kết nối (junction) giữa các đầu mút của phần tử chuyển vị với các đầu mút bị cắt hở của DNA mục tiêu (target site). Khác với tái tổ hợp tương đồng (homologous recombination), transposition không đòi hỏi sự tương đồng (homologous sequence) giữa các phần tử di động và DNA mục tiêu, và nó độc lập với chức năng được mã hóa bởi recA, vì nó sử dụng enzym đặc hiệu là Transposaz.
Đây là hiện tượng rất phổ biến trong thiên nhiên, các transposon được tìm thấy ở vi khuẩn, nấm, thực vật và động vật
Prokaryota Eukaryota
– Trình tự xen đoạn: IS
Transposon: Tn như Tn3, Tn5, Tn10,...
– Virut: Mu
– Retrotransposon:
Nấm men (Yeast): Ty
Ruồi giấm: copia, P
– Transposon (DNA):
Ngô: hệ thống Ac-Ds
– SINE: ở người (họ Alu)
– Retrovirut: Rous sarcoma, HIV
Bảng 6: Các phần tử chuyển vị có ở Prokaryotae và Eukaryota
Chúng có chức năng như các vector quan trọng thực hiện biến đổi di truyền.
Các xen đoạn, mất đoạn và cấu trúc lại bộ gen thường kèm theo sự di chuyển của các transposon. Sự di chuyển này làm sai hỏng chức năng bình thường của gen. Chúng hoạt hóa hay làm bất hoạt các gen bằng cách xen đoạn vào kề bên hay vào giữa đoạn gen. Các transposon IS, là các phần tử lặp đoạn (repetitive elements), còn tạo các vùng tương đồng rải trong bộ gen, nhờ đó các hệ thống tái tổ hợp tương đồng có thể tác động.
Sự chuyển vị (transposition) gồm các kiểu chính:
IS: trình tự xen đoạn (insertion sequence) là các transposon ngắn (khoảng 7801500 bp), cấu trúc đơn giản và chỉ mã hoá các chức năng cần thiết cho transposition
Transposon (kí hiệu Tn): Các phần tử di động dài (khoảng 5000 bp) có chứa một hoặc vài gen, gây ra đột biến, làm trung gian cho sự sắp xếp lại bộ gen, Nó thực hiện:
+ Transposition không sao chép (nonreplicative): Phần tử chuyển vị được cắt khỏi DNA cho (donor) và được gắn vào phân tử DNA mục tiêu. Thuộc loại này có Tn10, Tn5, Tn7 (trong đó Tn5, Tn10 xác định khả năng kháng kanamycin và tetracycline tương ứng. Các chuyển vị phức tạp có thể phát triển bằng cách chuyển vị các trình tự chèn tương đồng sang các vị trí lân cận trong phân tử DNA)
+ Transposition sao chép (replicative): DNA được nhân đôi và bản sao được xen vào vị trí mới tạo cointergrat (cộng gắn) gồm có thực khuẩn Mu và các phage ôn hoà có liên quan, Tn3 (Transposon kháng ampicillin), Tn1000 (transposon gammadelta) tìm thấy trong plasmid F
Nhóm Transposon Tn917 được phát hiện ở Gram dương bao gồm chuyển vị liên hoàn, khác hoàn toàn với các chuyển vị được mô tả ở trên. Transposon liên hợp không tạo ra sự sao chép của trình tự đích mà nó chèn vào, và ở vi khuẩn Gram dương, dòng vật chủ mang transposon có thể hoạt động như một người cho kết hợp. Vi khuẩn tiếp nhận không cần thiết phải có quan hệ mật thiết với vi khuẩn cho. Transposon được cắt ra khỏi nhiễm sắc thể của người cho và được truyền bằng cách tiếp hợp cho người nhận, nơi nó tích hợp ngẫu nhiên vào nhiễm sắc thể. Tn917 mã hóa kháng tetracycline, nhưng các chuyển vị liên hợp lớn hơn khác có thể mã hóa kháng kháng sinh bổ sung. Các chuyển vị liên hợp dường như là nguyên nhân chính của sự lan truyền tình trạng kháng thuốc kháng sinh ở vi khuẩn Gram dương
Retrotransposition: Sự di chuyển qua trung gian RNA, nhờ reverse transcriptaz tạo thành cDNA, và sự xen đoạn cDNA vào vị trí mới.
2.6.10. Các trình tự xen đoạn IS
Các transposon đơn giản nhất là các trình tự xen đoạn (insertion sequence) và được kí hiệu bởi tiếp đầu ngữ IS kèm số thứ tự như IS4.
Các phần tử IS là các cấu phần bình thường của DNA vi khuẩn và plasmid.
Dòng E. coli chuẩn thường chứa vài bản sao (<10) của bất kì một trong các IS chung thường gặp.
Các IS là những đơn vị tự trị, mỗi một trong chúng mã hóa cho chỉ một protein cần thiết cho sự chuyển vị bản thân chúng. Trình tự của mỗi loại IS có khác nhau, nhưng trong tổ chức cấu tạo có nhiều tính chất chung. Ở giữa có gen transposaz tnp, hai đầu có lặp đoạn đảo ngược (IR – Inverted repeats). Sự xen đoạn của IS ở tiêu điểm (target) được minh họa trên hình 35.
Các đầu mút của transposon có trình tự lặp lại đảo ngược (inverted repeat - IR).
Trong ví dụ này, tiêu điểm có 5 bp. Các đầu mút của transposon gồm các lặp đoạn đảo ngược 9 bp được đánh số từ 1 đến 9.
Cơ chế transposition được nghiên cứu chi tiết ở phage Mu và có thể tóm tắt như sau:
- Nhận biết và bắt cặp 2 đầu mút của transposon nhờ transposaz để hình thành cấu trúc chuyên biệt protein - DNA. Sự hình thành phức hợp được kích thích bởi các protein HU và IHF là các nhân tố tham gia tái tổ hợp điểm chuyên biệt Mu transposaz, giống như Int protein, có 2 vùng gắn độc lập.
- Cắt khấc (nicking) do transposaz tạo ra 3’-OH ở mỗi đầu của Mu.
- Cắt tiêu điểm nhờ transposaz tạo đầu mút so le 5’ - P dài 5 bp.
DNA mục tiêu
I R I R
Hình 35: Cấu trúc IS và sự chèn vào DNA mục tiêu
Ở hai đầu của IS luôn có hai trình tự lặp đoạn trực tiếp (direct repeat) ngắn. Các trình tự này dao động tùy transposon, nhưng cố định đối với mỗi loại IS. Chiều dài của phần lớn lặp đoạn trực tiếp là 9, chúng xác định các đầu mút của transposon.
2.6.11. Transposon không sao chép và sao chép
Sự chuyển vị DNA theo 2 kiểu khác nhau (sao chép và không sao chép hay bảo tồn (conservative)) và có những tính chất chung.
- Nối đầu 5’ - P của tiêu điểm với 3’-OH của transposon tạo cấu trúc trung gian chuyển mạch (strand - transfer intermediate).
2.6.12. Sự chuyển vị ngược (Retrotransposition)
Nhóm các transposon, được phát hiện ở nấm men (Ty) và Drosophila (copia) và có liên quan đến retrovirut, di chuyển đến vị trí mới qua trung gian RNA.
Phương thức phiên mã ngược của các phần tử di động tương tự như của retrovirut. Có sự tương tự về cấu trúc giữa retrovirut và các retrotransposon ở nấm men (Ty) và Drosophila (copia).
Sự chuyển vị ngược tạo bản sao của phần tử ở vị trí mới, trong khi phân tử cho ban đầu vẫn giữ nguyên cấu trúc không biến đổi. Do vậy, retrotransposition tạo nên một ít đứt đoạn và các tái cấu trúc (rearrangement) của bộ gen tế bào chủ. Những biến đổi của bộ gen liên quan với retrotransposition sẽ dẫn đến việc làm ngừng hay hoạt hóa các gen, mà một số có thể gây ung thư
Các transposon đã góp phần vào sự tiến hóa của plasmid, và vài chứng cứ cho thấy các plasmid R có được tính đề kháng kháng sinh thông qua các transposon.

Integron là các transposon có thể thu nhận và biểu hiện các gen từ nhiều nguồn khác nhau. Tuy nhiên, khác với các transposon được chèn vào một cách ngẫu nhiên, các integron chèn vào có tính chọn lọc cao (high selective) trong các điểm xen đoạn (insertion site) của chúng, thường thì đựơc chèn vào trong plasmid.
Hình 36: Sơ đồ về transposition sao chép và không sao chép DNA cho
Cả hai đầu của phân tử mang gần như một trình tự có định hướng đảo ngược nhau.
Các transposon mã hóa ít nhất một protein là transposaz. Transposaz gắn đặc hiệu và cắt các trình tự ở cuối để thực hiện chuyển vị.
Các transposon tạo bản sao ngắn (12bp) của DNA ở điểm mục tiêu trong transposition. Chiều dài của trình tự này đặc trưng và không đổi đối với một phần tử nhất định và được tạo ra nhờ sự cắt so le (staggered cleavage) của DNA mục tiêu nhờ transposaz.
Integron chứa gen mã hóa cho protein được gọi là integraz, cần thiết cho sự tái tổ hợp ở điểm chuyên biệt (site-specific recombination). Bộ gen của phage λ (lambda) chèn vào Nhiễm sắc thể của E. coli ở một vị trí đặc biệt nhờ họat tính của enzym integraz λ. Integron cũng chứa trình tự DNA đặc biệt cho phép integraz chèn các nhóm gen gọi là cassette vào Nhiễm sắc thể, mà trên đó có promoter cho phép các gen cassette mới được biểu hiện.
2.6.13. Tái bản DNA (DNA replication)
a. Các nguyên tắc và đặc điểm chung của tái bản DNA
- Tái bản theo kiểu bán bảo toàn (semi-conservative).
- Tái bản bắt đầu tại một hoặc nhiều khởi điểm (Ori). Từ khởi điểm, DNA mở xoắn tạo thành hai chạc tái bản (replication fork). Cấu trúc như vậy gọi là đơn vị tái bản (replicon). DNA E. coli trong quá trình tái bản như vậy có cấu trúc giống chữ cái theta Hy Lạp (θ) nên gọi là tái bản theta. Đối với các DNA mạch vòng, mỗi phân tử chỉ có một Ori; trong khi đó mỗi DNA nhiễm sắc thể eukaryote có nhiều Ori.
- Tham gia vào sự tái bản DNA có nhiều protein và enzyme.
- Tại mỗi chạc tái bản, trước tiên xảy ra sự tổng hợp các đoạn mồi RNA (primer) bởi vì các DNA polymerase tự nó không thể bắt đầu tổng hợp mới được.
Mặt khác, do hai sợi đơn của mỗi chạc ngược chiều nhau trong khi các DNAvà RNA polymerase chỉ xúc tác theo chiều 3'-5', cho nên sự tái bản DNA trên hai sợi khuôn là không giống nhau: một sợi liên tục gọi là sợi dẫn đầu (leading strand) và một sợi không liên tục gọi là sợi ra chậm (lagging strand). Kiểu tái bản như thế gọi là tái bản nửa gián đoạn (semi-discontinuous), được R. Okazaki phát hiện đầu tiên năm 1969.
Một khởi điểm và hai chạc tái bản sinh trưởng đồng thời theo hai hướng đối lập nhau, mỗi chạc gồm một sợi dẫn đầu và một sợi ra chậm.
sợi dẫn đầu (được tổng hợp liên tục) chạc tái bản chạc tái bản
sợi ra chậm (được tổng hợp không liên tục)
Hình 37: Tái bản AND ở E. coli
Cơ chế tái bản DNA (E. coli)
Đối với nhiễm sắc thể E. coli, sự tái bản bắt đầu tại một khởi điểm đặc thù gọi là Ori. Quá trình diễn biến ở khởi điểm E. coli cho đến lúc hình thành hai chạc có thể tóm tắt như sau: (1) Các protein bám khởi điểm dnaA bám Ori tạo ra cấu trúc nucleoprotein chuyên hoá của khởi điểm; (2) Cấu trúc này mở xoắn vùng DNA giàu
AT để hình thành "phức hợp mở"; và (3) Hai phân tử helicase dnaB chui vào phức hợp mở làm mở xoắn khởi điểm theo cả hai hướng, tạo thành hai chạc tái bản (replication fork). Khi cả hai sợi đơn của mỗi chạc được tách ra thì các protein SSB bám vào
Giai đoạn khởi đầu
Khởi đầu trong tái bản DNA ở E. coli là sự tổng hợp một đoạn mồi ngắn khoảng 10-12 nucleotide bởi primase (xét chung ở các sinh vật là ~ 5 base). Cả ba loại protein dnaB, dnaC và dnaG hợp thành một phức hợp có tên là primasome (hoặc primosome: thể mở đầu)
Giai đoạn kéo dài (elongation)
Một khi primasome tổng hợp xong một mồi, sự kéo dài chuỗi DNA được bắt đầu bằng một phức hợp giữa primasome và DNA polymerase III hoàn chỉnh, gọi là replisome (thể tái bản). Sự tái bản DNA trên mỗi chạc xảy ra theo kiểu nửa gián đoạn (semi-discontinuous)
Trên sợi khuôn dẫn đầu (3'→5'): Sau khi primase (primasome) tổng hợp xong đoạn mồi RNA với đầu 3'-OH tự do, enzyme hoàn chỉnh DNA polymerase III (hay replisome) bắt đầu kéo dài chuỗi DNA mới sinh trưởng theo chiều 5'→3' một cách liên tục.

Trên sợi khuôn ra chậm (5'→3'): Sự kéo dài diễn ra không liên tục dưới dạng các đoạn Okazaki. Kích thước trung bình mỗi đoạn Okazaki ở E. coli là 1.000 - 2.000 nucleotide. Quá trình này đòi hỏi sự "mồi hóa" lặp lại, với sự tham gia lần lượt của bốn enzyme sau: (i) primase tổng hợp một đoạn mồi RNA; (ii) DNA polymerase III kéo dài đoạn Okazaki; (iii) DNA polymerase I vừa cắt bỏ đoạn mồi vừa lấp khoảng trống bằng cách kéo dài dần đoạn Okazaki theo sau; (iv) DNA ligase hàn liền khe hở giữa hai đoạn Okazaki bằng một liên kết phosphodiester.
Sợi mới
Sợi khuôn dẫn đầu
Sợi khuôn ra chậm
DNA polymerase
Protein bám sợi đơn (SSB)
DNA polymerase
Helicase
RNA primer
Đoạn Okazaki
DNA cha mẹ
Primase
Hình 38: Cơ chế tái bản nửa gián đoạn trên một chạc tái bản.
Giai đọan kết thúc (termination)
Do cấu trúc nhiễm sắc thể ở hai nhóm prokaryote và eukaryote là hoàn toàn khác nhau, nên cơ chế kết thúc tái bản của chúng cũng khác nhau.
Cả hai chạc tái bản của DNA E. coli được bắt đầu từ một khởi điểm (Ori), và di chuyển hầu như cùng tốc độ theo hai hướng đối lập nhau xung quanh nhiễm sắc thể mạch vòng cho tới khi chúng gặp nhau tại một điểm kết thúc chung đối diện với ori.
RTP của E. coli có TLPT ~36kD, được mã hóa bởi gene tus (ter) và trong mỗi tế bào có ~80 bản sao của protein này được duy trì hầu như ổn định trong suốt chu kỳ tế bào. Trình tự điều hoà của vùng kết thúc tái bản ở E. coli (R6K), theo Bastia và cs (1997), như sau:
5'NN(A/T)(A/T)(A/T)G(A/T)(A/G)TGTTGTAACTA(A/C)NN3'
2.6.14. Phiên mã (Transcription) ở prokaryote
a. Các nguyên tắc và đặc điểm chung của phiên mã
Phiên mã (transcription) là quá trình tổng hợp các RNA khác nhau từ thông tin di truyền chứa đựng trong DNA. Trừ các gene mã hóa protein trong các operon ở vi khuẩn, nói chung, các RNA mới được tổng hợp chỉ là các bản sao sơ cấp (primary transcript) gọi là các pre-RNA. Các pre-RNA này phải trải qua một quá trình sửa đổi để trở thành các RNA trưởng thành (mature) trước khi tham gia vào quá trình sinh tổng hợp protein của tế bào.
Quá trình phiên mã DNA các đặc điểm chung sau đây:
Hình 39: Sự tổng hợp RNA trên một sợi khuôn của gene (DNA).
- Diễn ra dưới tác dụng của các enzyme RNA polymerase.
- Chỉ một sợi đơn được dùng làm khuôn cho tổng hợp RNA, gọi là sợi khuôn, sợi mã hoá hay sợi có nghĩa (template/ coding / sense strand); còn sợi bổ sung được gọi là sợi đối khuôn, sợi không mã hoá hay sợi đối nghĩa (antitemplate/ non-coding / antisense strand).
- Phản ứng tổng hợp RNA diễn ra theo nguyên tắc bổ sung và được kéo dài theo chiều 5'→3', ngược với chiều của sợi khuôn.
- Nguyên liệu cho tổng hợp gồm: ATP, UTP, GTP và CTP.
- Sản phẩm của phiên mã là các RNA sợi đơn.
- Khởi đầu và kết thúc phiên mã phụ thuộc vào các tín hiệu điều hoà là các trình tự DNA đặc thù nằm trước gen (vùng khởi động) và sau gene.
Ở các prokaryote, RNA polymerase hoàn chỉnh (holoenzyme) là một phức hợp gồm nhân tố sigma và lõi enzyme. Nhân tố sigma giúp RNA polymerase nhận biết và bám vào promoter để có thể bắt đầu phiên mã tại vị trí chính xác, và lõi enzyme đóng vai trò xúc tác tổng hợp RNA.
Vùng khởi động (promoter) nói chung nằm kề trước gene và có chứa các trình tự đặc thù cho phép RNA polymerase nhận biết và bám vào. Trình tự quan trọng nhất
của promoter là hộp TATA hay hộp Pribnow (Pribnow box) ở vị trí "-10". Ngoài ra, còn có trình tự TTGACA ở vị trí ''-35'', gọi là đoạn nhận biết (recognition sequence).
Các vùng này được bảo tồn cao.
Các giai đoạn của quá trình phiên mã ở prokaryote
- Khởi đầu: RNA polymerase holoenzyme nhận biết và bám vào promoter, tháo xoắn một đoạn khoảng 12 cặp base. Sau khi tổng hợp ribonucleotide đầu tiên (Appp hoặc Gppp), nhân tố sigma tách ra để đi vào một chu kỳ phiên mã khác, gọi là chu kỳ sigma (sigma cycle).
- Kéo dài: Enzyme lõi tổng hợp sợi RNA dọc theo sợi khuôn.
- Kết thúc: Khi phiên mã xong hai vùng giàu GC và AT nằm sau gene, ở vùng đuôi của sợi RNA hình thành cấu trúc ''kẹp cài tóc'' (hairpin loop) làm dừng sự phiên mã của lõi RNA polymerase. Cuối cùng, dưới tác dụng của nhân tố rho, sợi RNA và enzyme lõi được giải phóng khỏi DNA khuôn.
2.6.15. Dịch mã tổng hợp protein
Dịch mã (translation) là quá trình sinh tổng hợp protein diễn ra trong tế bào chất, được chia làm hai giai đoạn sau đây:
Hoạt hoá amino acid
Đây là quá trình tạo nguồn các aminoacyl-tRNA cho dịch mã. Mỗi amino acid được đính vào tRNA thích hợp thông qua hai phản ứng nhờ enzyme aminoacyl-tRNA
synthetase đặc thù cùng với ATP và Mg2+.
R−CH(NH2)−COOH + ATP → R−CH(NH2)−CO~AMP + PP
R−CH(NH2)−CO~AMP + tRNA → R−CH(NH2)−CO~tRNA + AMP
Tổng hợp polypeptide
Mở đầu (initiation): Quá trình dịch mã bắt đầu khi một tiểu đơn vị ribosome bé bám vào mRNA tại vị trí của codon khởi đầu AUG. Lúc này một phân tử tRNA khởi đầu đặc thù mang methionine (ở vi khuẩn là formyl-Met) đi vào và khớp anticodon của nó với codon mở đầu của mRNA. Kế đó, tiểu đơn vị ribosome lớn bám vào tiểu đơn vị bé tạo ra một ribosome hoạt động hoàn chỉnh. Lúc này Met-tRNA ở vị trí P và vị trí A để trống; một tRNA thứ hai (ví dụ, tRNAVal) đi vào vị trí A và khớp với codon thứ hai
Kéo dài (elongation): Quá trình kéo dài bắt đầu sau khi liên kết peptide đầu tiên được hình thành. Phản ứng này được xúc tác bởi enzyme peptidyl transferase, và kết quả là tạo ra một peptidyl-tRNA ở vị trí

B Liên kết peptid
D Tách và giải phóng
Chuỗi gồm 8 amino acid
Nhân tố RF
Hình 40: Quá trình tổng hợp chuỗi polypeptide


Sau đó, ribosome lập tức chuyển dịch sang một codon mới dọc theo mRNA theo chiều 5'→3' (C). Phản ứng này đẩy phân tử tRNA tự do vốn ở vị trí P ra ngoài; lúc này peptidyl-tRNA nằm ở vị trí P và vị trí A lại để trống. Một chu kỳ dịch mã mới lại bắt đầu, một aminoacyl-tRNA thứ ba đi vào và khớp anticodon của nó với codon đang để trống ở vị trí A, một liên kết peptide thứ hai được hình thành, và ribosome lại dịch chuyển sang codon kế tiếp. Quá trình nói trên cứ diễn ra một cách tuần tự dọc theo mRNA làm cho chuỗi polypeptide dài dần ra cho đến dịch mã xong codon có nghĩa cuối cùng.

Kết thúc (termination): Quá trình tổng hợp polypeptide sẽ dừng lại khi codon kết thúc của mRNA đối diện với vị trí A. Lúc này nhân tố giải phóng RF (release factor) đi vào (D); chuỗi polypeptide được tách ra và phóng thích cùng với hai tiểu đơn vị ribosome cũng như tRNA ra khỏi mRNA.
3. Di truyền học vi nấm và vi tảo Các vi sinh vật nhân thực Eukaryota như vi nấm, vi tảo có chu trình sống đa dạng và ở nhiều điểm giống với các Eukaryota bậc cao. Cấu tạo đơn bào giúp cho việc sử dụng có hiệu quả các phương pháp phân tích di truyền ở vi sinh vật. Phân tích bộ bốn giúp nghiên cứu được chi tiết từng sản phẩm của giảm phân. Các nghiên cứu di truyền vi nấm tập trung ở 4 vấn đề căn bản: tính không phối hợp, phân tích bộ bốn, tái tổ hợp nguyên phân và sự đơn bội hóa.
Neurospora crassa và Saccharomyces cerevisiae là những đối tượng nghiên cứu quan trọng của di truyền. Ở vi tảo Chlamydomonas reinhardii được nghiên cứu chi tiết hơn cả trong số các vi tảo. Các nghiên cứu ở vi tảo làm sáng tỏ nhiều vấn đề của di truyền quang hợp và sự xác định các sắc tố.
3.1. Nấm men Saccharomyces cerevisiae






Nấm men Saccharomyces cerevisiae là một loài thuộc lớp Nấm Túi (Ascomycetes), còn gọi là “men bánh mì” (baker yeast) hay “men rượu”. Đó là loài nấm men chủ yếu được dùng trong các quá trình lên men rượu. Vì là Eukaryota đơn bào (kích thước khoảng 5 – 10µm, được biết rất chi tiết về di truyền học và sinh lí học, cho nên là đối tượng mô hình nghiên cứu di truyền các sinh vật nhân thực.
Hình 41: Nấm men S. cerevisiae dưới kính hiển vi điện tử và kính hiển vi quanghọc
a. Các đặc tính sinh học
S. Cerevisiae có 2 tính chất: là đơn bào nên có thể tiến hành thí nghiệm như vi khuẩn, đồng thời có những đặc tính chủ yếu điển hình của Eukaryota và có ty thể với bộ gen DNA nhỏ. Giống với vi khuẩn, nó có thể nuôi trên môi trường dịch thể hay đặc và tạo khuẩn lạc trên môi trường thạch. Có thể nuôi tế bào nấm men quy mô lớn trong các nồi lên men và dễ dàng thu nhận sinh khối tế bào.
Nấm men có 2 dạng tế bào đơn bội (n) là a và α có thể tồn tại độc lập nhờ sinh sản vô tính qua nguyên phân (mitosis). Khi 2 dạng a và α gặp nhau bắt cặp, rồi phối hợp tế bào và hợp nhân tạo 1 tế bào lưỡng bội (2n). Tế bào lưỡng bội có thể sinh sản vô tính vô hạng và đây là dạng thường sử dụng trong sản xuất. Trong những điều kiện nhất định, tế bào 2n sinh sản hữu tính qua giảm phân tạo nang có 4 bào tử (2 a và 2 α), mà sự kết hợp a và α tạo tế bào 2n, lặp lai chu trình.
Hình 42: Chu trình sinh sản của nấm men S. cerevisiae
b. Các dữ liệu di truyền học của S. cerevisiae:
Kích thước bộ gen (Genome size): 12 Mb
Nhiễm sắc thể: n = 16
Số lượng gen: 6.000
Phần trăm gen tương đồng với người: 25%
Kích thước trung bình của gen: 1,5 kb, 0,03 intron/gen
Các transposon: một tỷ lệ nhỏ của DNA
Kết thúc giải ký tự chuỗi: 1996
c. Các kĩ thuật biến đổi di truyền trên nấm men:
Gây đột biến (Mutagensis):
Hóa chất và chiếu xạ: đột biến xoma ngẫu nhiên.
Dùng transposon: xen đoạn (Insertions) xoma ngẫu nhiên.
Chuyển gen (Transgensis):
Plasmid tích hợp (integrative): xen đoạn (Inserts) nhờ tái tổ hợp tương đồng (homologous recombination).
Plasmid sao chép (replicative): Có thể sao chép tự lập (autonomously) (Ori sao chép của plasmid 2 hay ARS).
Nhiễm sắc thể nhân tạo của nấm men: sao chép và phân ly như một nhiễm sắc thể.
Vector con thoi (Shuttle vector): Có thể sao chép trong tế bào nấm men hay
E. coli.
Làm im lặng gen mục tiêu (Targeted gen knockout): Thay thế gen (Gen replacement): tái tổ hợp tương đồng thay alen hoang dại bằng alen không (null alen).
S. cerevisiae có nhiễm sắc thể nhân tạo (Yeast artificial chromoxom - YAC) có trình tự sao chép ARS (Autonomously replicative sequence), tâm động (centromere) và 2 telomere (trình tự đầu mút nhiễm sắc thể). YAC tồn tại độc lập trong tế bào nấm men như một nhiễm sắc thể và các gen được chèn vào nó được di truyền như nhiễm sắc thể Mendel. YAC có khả năng mang đoạn gDNA ngoại lai dài 1 – 2 Mb, nên thuận tiện cho tạo dòng và giải ký tự chuỗi (sequencing) những bộ gen lớn như bộ gen người. Nấm men S. cerevisiae được sử dụng rộng rãi làm tế bào chủ trong KTDT.
Ngoài ra, một số protein tái tổ hợp (gốc động vật) như interferon khi biểu hiện trong tế bào Prokaryotae như E. coli thì không có hoạt tính do thiếu biến đổi sau dịch mã, nhưng khi cho biểu hiện trong tế bào nấm men thì sản phẩm có hoạt tính.
Vai trò chủ yếu của S. cerevisiae trong các nghiên cứu:
- Chu trình tế bào (Cell cycle): Sự xác định các gen kiểm soát phân bào nhờ các đột biến nhạy cảm nhiệt (temperature-sensible mutants (cdc mutants)) dẫn đến mô hình rất tốt cho nghiên cứu sự phân bào.
- Tương tác gen (Gen interaction): nghiên cứu sự ức chế gen (suppression). Hệ thống plasmid lai kép (two-hybrid plasmid system) giúp tìm các tương tác giữa các protein ở nấm men đã dẫn đến các bản đồ tương tác phức tạp, mà là khởi đầu cho sinh học các hệ thống (systems biology).
- Di truyền học ty thể: nhờ các đột biến “petite” mất khả năng hô hấp mà phát hiện các gen của ty thể. Nhờ chúng và các đột biến khác của ty thể mà phân tích chi tiết cấu trúc và chức năng bộ gen ty thể.
- Di truyền học kiểu bắt cặp (Gentics of mating typ): Các alen MAT ở nấm men là các gen kiểu bắt cặp đầu tiên được xác định các đặc tính ở mức phân tử.
Đóng góp khác: Sự di truyền của khóa dóng mở (switching) giữa sự tăng trưởng kiểu nấm men (yeast-like) và sợi (filamentous); di truyền học sự thoái hóa (senescense).
3.2. Nấm sợi Neurospora crassa
a. Các đặc tính sinh học
Giống với vi khuẩn và nấm men, nấm sợi này có thể nuôi trên môi trường dịch thể hay đặc và tạo khuẩn lạc trên môi trường thạch. Nó thể hiện các tính chất điển hình của Eukaryota giống như nấm men: bộ gen gồm nhiều nhiễm sắc thể, bào tử túi có trạng thái đơn bội (n) và hợp tử lưỡng bội (2n), sinh sản vô tính qua chu trình nguyên phân (mitotic cycle), sinh sản hữu tính qua giảm phân và có ty thể với bộ gen DNA nhỏ. Nấm thường mọc trên bánh mỳ cũ và có màu vàng tươi.
Hình 43: Nấm sợi Neurospora crassa dưới kính hiển vi quang học Sợi nấm phân đoạn có chứa nhiều nhân đơn bội. Có hai kiểu giới tính được điều khiển bởi một căp alen A và a. Sinh sản hữu tính chỉ thực hiện khi có sự kết hợp của các tế bào khác alen. Như hình vẽ mô tả khi bào tử đính a rơi trên tiền quả thể dạng chai (protoperithecium). Các cơ quan sinh dục cái hay A rơi lên tiền quả thể a thì sự thụ tinh sẽ được xảy ra. Lúc đầu hai nhân đơn bội A và a tồn tại song song và chia đồng thời. Sau đó xảy ra sự hợp nhân và tiền quả thể dạng chai biến thành quả thể

dạng chai (perithecium). Mỗi hợp tử lưỡng bội 2n tạo thành một tế bào kéo dài gọi là nang hay túi (ascus) và lập tức chia giảm nhiễm được 4 nhân, mà 4 nhân này lại chia nguyên nhiễm một lần nữa tạo thành 8 nhân của 8 bào tử nang hay bào tử túi (ascospores).
Hóa chất và chiếu xạ: đột biến xoma ngẫu nhiên.
Dùng transposon: không.
Chuyển gen (Transgensis):
Biến nạp qua trung gian plasmid (Plasmid-mediated transformation): xen
đoạn ngẫu nhiên (Random insertion).
Làm im lặng gen mục tiêu (Targeted gen knockout):
RIP: các đột biến GC –> AT trong các đoạn tăng đôi chuyển gen trước khi lai.
Ức chế (Quelling): bất hoạt xoma sau phiên mã các gen được chuyển vào (Xomatic posttranscriptional inactivation of transgens).
3.3. Các đặc điểm di truyền vủa vi nấm
Hình 44: Chu trình sống của nấm sợi Neurospora crassa
Một đặc điểm độc đáo của Neurospora crassa là các sản phẩm của giảm phân, 8 nang bào tử xếp thẳng hàng trong nang. Nhờ vậy, qua phân tích bộ bốn (tetrade analysis) nghiên cứu hàng loạt các cơ chế di truyền như trao đổi chéo, gen chuyển biến (conversion), tái cấu trúc nhiễm sắc thể (chromosome rearrangement), sự không chia ly trong giảm phân (meiotic nondisjunction) và sự kiểm soát di truyền đối với giảm phân. Các nhiễm sắc thể tuy nhỏ, nhưng có thể nhìn thấy dễ dàng.
N. crassa đạt kỷ lục về tốc độ tăng trưởng trong các loài nấm: mỗi sợi nấm (hypha) mọc dài ra hơn 10 cm/ngày. Sự tăng trưởng nhanh đó kết hợp với chu trình đơn bội và khả năng mọc trên các môi trường nuôi cấy có thành phần xác định làm cho nó thành một đối tượng rất thuận tiện cho nghiên cứu di truyền hóa sinh (biochemical gentics). Trên thực tế, nó nổi tiếng với sự ra đời giả thuyết một gen - một enzyme.
Các dữ liệu di truyền học của Neurospora crassa:
Kích thước bộ gen (Genome size): 43 Mb
Nhiễm sắc thể: 7 Nhiễm sắc thể thường (n = 7)
Số lượng gen: 10.000
Phần trăm gen tương đồng với người: 6 %
Kích thước trung bình của gen: 1,7 kb, 1, 7 intron/gen
Các transposon: rất hiếm
Kết thúc giải ký tự chuỗi: 2003
Phân tích di truyền
Gây đột biến (Mutagensis):
Chu trình sống của các loài nấm khác nhau rất đa dạng, nhưng có thể xếp vào 5 kiểu căn bản khác nhau như: 1. Sinh sản vô tính; 2. Thể đơn bội; 3. Thể đơn bội với giai đoạn dị nhân; 4. Đơn bội - Lưỡng bội; 5. Lưỡng bội.
Ngay trong các nấm sinh sản hữu tính cũng có sự dao động từ hoàn toàn đơn bội như Neurospora đến hoàn toàn lưỡng bội như ở Saccharomyces cerevisiae.
Đối với phần lớn nấm sợi, điểm đặc trưng là nhiều nhân cùng hiện diện trong một tế bào. Các nhân này có thể có kiểu gen khác nhau. Trong trường hợp này được gọi là thể dị nhân (heterokaryon) tương tự dị hợp tử.
Ở các vi sinh vật nói chung, các tính trạng hình thái thường không nhiều, nhưng có sự đa dạng lớn về nhiều loại phản ứng hóa học khác nhau. Sự ra đời của giả thuyết nổi tiếng 1 gen - 1 enzym đã dựa trên các đột biến sinh hóa ở N. crassa. Các môi trường chọn lọc được sử dụng để thu thập các dạng lai hiếm hoi giữa các dòng cha mẹ đơn bội mang các đột biến khuyết dưỡng lặn, khi sai hỏng trong chuỗi phản ứng sinh hóa của chúng bổ sung cho nhau. Ngoài ra tính đề kháng với các tác nhân bất lợi được sử dụng rộng rãi làm tính trạng đánh dấu trong các nghiên cứu di truyền ở nấm.
Nói chung, các vi sinh vật nhân thực có ưu thế đặc biệt là sự kết hợp giữa cấu tạo tế bào và chu trình sống như sinh vật bậc cao với cơ thể đơn bào của vi sinh vật. Nếu như trong những năm 70 của thế kỷ trước, các đối tượng căn bản của di truyền học phân tử là phage và vi khuẩn, thì những từ năm 80 cho đến nay các Eukaryota đơn bào đã thay thế dần.
Bốn vấn đề căn bản được chú ý trong nghiên cứu di truyền ở nấm là: tính không phối hợp (incompatibility), phân tích bộ bốn (tetrade analysis), tái tổ hợp nguyên phân (mitotic recombination), và sự đơn bội hóa (haploidisation).
Nhiều nấm sợi có quả thể nhìn thấy được không phải là vi nấm, nhưng có thể sử dụng các phương pháp dùng cho vi nấm để nghiên cứu ..
a. Lập bản đồ di truyền bằng phân tích bộ bốn
Đối với trường hợp bộ bốn sắp theo thứ tự như ở Neurospora, số phần trăm các nang tái tổ hợp phản ánh khoảng cách giữa gen và tâm động. Trao đổi chéo chỉ liên quan đến hai nhiễm sắc thể, nên chỉ có hai sản phẩm của giảm phân chứa nhiễm sắc thể tái tổ hợp. Do đó để tính phần trăm các nhiễm sắc thể tái tổ hợp, lấy phần trăm mang tái tổ hợp chia đôi.
Trường hợp bộ bốn không theo thứ tự như ở nấm men thì việc tính tần số tái tổ hợp khá phức tạp
mô tả ở các loài nấm khác như Neurospora crassa, Sordaria fimicola và nó có ý nghĩa quan trọng trong giải thích cơ chế tái tổ hợp.
Hình 45: Các kiểu bộ bốn khác nhau: – Kiểu PD parental dityp (không trao đổi chéo) – Kiểu RD recombination dityp hay NPD nonparental dityp (có trao đổi chéo) – Kiểu TT tetratyp (có trao đổi chéo).

b. Hiện tượng chuyển biến (conversion)
Chuyển biến (conversion) là hiện tượng lệch khỏi sự phân li bình thường 2: 2 của các bộ bốn, khi có sự di truyền của 1 gen, được Lindegreen phát hiện năm 1949.
Ví dụ: Dị hợp tử của nấm men a+/+b tạo các bộ bốn bất thường như: (a +) (a
+) (a b) (+ b) và (a +) (+ +) (+ b) (+ b) hoặc (a +) (a b) (+ b) (+ b) và (a +) (a +) (+ +) (+ b). Sự phân li bất thường có thể liên quan đến cả 2 gen như: (a +) (a +) (a +) (+ b) hay (a +) (+ b) (+ b) (+ b).
Các nghiên cứu tiếp theo cho thấy chuyển biến (conversion) là hiện tượng bình thường ở các gen khác nhau của nấm men với tần số 1%. Hiện tượng này cũng được
Hình 46: Trao đổi chéo trong nguyên phân (Mitotic crossing-over) ở đoạn a-b.

a) Trong tứ dị hợp tử theo các gen liên kết, khi alen lặn c (cần được xác định vị trí) nằm trên đoạn tương đồng được đánh dấu.
b) Trong đoạn tương đồng không đánh dấu, trao đổi chéo ở đoạn giữa gen và tâm động đưa đến sự đồng hợp tử của các gen xa điểm trao đổi ở một nửa số nhân của tế bào (thế hệ con của nhân tái tổ hợp). Nhờ đó, có thể sử dụng trao đổi chéo nguyên phân để xác định vị trí gen ở một vai của nhiễm sắc thể so với tâm động.
3.4. Vi tảo
Các tảo đơn bào (Microalgae) cũng được dùng làm đối tượng nghiên cứu của Di truyền học, đặc biệt là di truyền quang hợp và di truyền của bào quan lục lạp. So với nấm, các vi tảo và nói chung là tất cả thực vật, các đột biến khuyết dưỡng ở tảo lục đơn bào chỉ hạn chế trong một phạm vi hẹp như mất khả năng tổng hợp arginin (arg–) và một số vitamin
Loài vi tảo được sử dụng sớm nhất và nghiên cứu di truyền chi tiết hơn cả là Chlamydomonas reinhardii nó có chu trình sinh sản hữu tính. Ưu thế của đối tượng này là có thể tiến hành lai và phân tích bộ bốn không xếp theo thứ tự.
Hình 47: Cấu trúc và hình thái Chlamydomonas
a. Đặc điểm sinh học
Vi tảo Chl. reihardii có thể nuôi cấy trong môi trường dinh dưỡng dịch thể hay môi trường thạch đặc như các vi sinh vật khác. Các tế bào mọc thành khuẩn lạc trên môi trường thạch. Vi tảo có thể nuôi ngoài sáng (ánh sáng mặt trời hay đèn huỳnh quang thông dụng) hoặc trong tối (phải bổ sung axetat natri hoặc glucoz) …
Bình thường các tế bào dinh dưỡng tồn tại lâu dài ở trạng thái đơn bội n với 2 kiểu bắt cặp khác nhau mt+ (mating typ) và mt– tương đương với 2 giới tính, mà về hình thái thì rất giống nhau. Các tế bào dinh dưỡng nay có thể tạo nang bào tử kín hay túi bào tử (sporangia) và sinh sản vô tính tạo ra các tế bào con đơn bội.
Khi được kích thích, như nuôi trên môi trường dinh dưỡng thiếu nitơ, các tế bào dinh dưỡng chuyển thành các động bào tử (zoospores) mt+ và mt–. Sự kết hợp và phối hợp 2 loại tế bào khác kiểu bắt cặp mt+ và mt–sẽ tạo ra hợp tử lưỡng bội 2n. Hợp tử này gặp điều kiện thuận lợi sẽ thực hiện giảm phân tạo ra 4 tế bào đơn bội n, mà chúng sinh sản vô tính tạo ra số lượng lớn tế bào dinh dưỡng.
Ở Chl. reinhardii có thể nhận được nhiều đột biến khác nhau, đặc biệt là các đột biến sắc tố khác với màu khuẩn lạc xanh lục của dạng hoang dại như màu cam, vàng, trắng kem, xanh lợt, …thuận lợi cho nghiên cứu di truyền lục lạp và quang hợp
Các đột biến có biểu hiện kiểu hình như sau:
- Mất khả năng quang hợp; để mọc được ngoài ánh sáng và trong tối cần bổ sung đường khử là axetat.
- Nhạy cảm với nhiệt độ cao hoặc thấp.
- Tính đề kháng với thuốc kháng sinh hoặc có nhu cầu được cung cấp thuốc kháng sinh.
Tất cả các đột biến trên có sự di truyền theo một cha mẹ (uniparental inheritance), có kiểu bắt cặp mt+ (có thể coi là dòng mẹ). Điều này liên quan đến sự hình thành lục lạp trong hợp tử, bằng cách nào đó chỉ nhận DNA từ lục lạp mt+ .
Bảng 7: Tỷ trọng của cpDNA ở thế hệ con từ các tổ hợp lai
Chlamydomonas
Tổ hợp lai
N14 mt+ x
N14 mt–
N15 mt + x
N15mt–
N15 mt + x
N14mt–
N14 mt + x
N15mt–
Tỉ trọng của cpDNA của hợp tử (g/cm3 )
Hình 48: Chu trình sống của vi tảo Chlamydomonas reinhardii
Sự di truyền của lục lạp được nghiên cứu chi tiết hơn cả ở vi tảo Chlamydomonas reinhardii. Tế bào của vi tảo này có một lục lạp lớn với đường kính trung bình 5 micrometer chứa 50 đến 80 bản sao của phân tử DNA vòng tròn dài 196 kb.
Trên thực tế, các kết quả này cho thấy cpDNA của cha mẹ mt– bị mất hoặc bị phân hủy như thế nào đó. Sự mất đó kèm theo mất các đột biến (ví dụ: sm) trên cha mẹ mt–
Các thí nghiệm khác theo nguyên tắc tương tự được tiến hành với việc sử dụng các đoạn RFLP. Mỗi dòng cha mẹ có cpDNA với RFLP khác nhau. Kết quả cũng cho thấy cpDNA ở thế hệ con nhận được từ dòng mt+
Như vậy, có thể kết luận rằng cpDNA của Chlamydomonas được truyền thụ từ một dòng cha mẹ (uniparental) và các gen của nó cũng có sự truyền thụ tương tự.
Lập bản đồ các gen của Chlamydomonas
Trong tổ hợp lai mt+sm –r x mt–sm –s có khoảng 0,1% thế hệ hợp tử con mang cả sm –r và sm–s. Các hợp tử như vậy là hợp tử hai cha mẹ (biparental zygote) và được gọi là cytohet (cytoplasmically heterozygote).

Tần số các cytohet có thể tăng lên 40–100% ở thế hệ hợp tử con nếu mt+ được chiếu tia tử ngoại trước khi lai.
PHẦN B: PHẦN BÀI TẬP
CÂU HỎI VÀ BÀI TẬP CÓ PHÂN TÍCH, HƯỚNG DẪN
học, ví dụ như quá trình diễn ra trong sự sinh trưởng của khối u ung thư, đúng như thực sự nó diễn ra trong cơ thể
Câu 1.
Đánh giá axit amin bằng đồng vị phóng xạ, sau đó theo dõi sự di chuyển của dấu phóng xạ. Lúc đầu người ta thấy dấu phóng xạ xuất hiện trên mạng lưới nội chất hạt, tiếp theo dấu phóng xạ có thể xuất hiện ở những cấu trúc nào của tế bào? Cho biết chức năng của các cấu trúc đó?
Hướng dẫn vận dụng kiến thức: Để trả lời được câu hỏi, yêu cầu học sinh phải nắm chắc kiến thức phần cấu trúc tế bào, sự di chuyển của protein trong tế bào
+ Kiến thức về thành phần hoá học của tế bào (aa là thành phần của protein) vị trí của protein trong tế bào.
+ Kiến thức về hoá sinh tế bào: sự vận chuyển protein trong tế bào
Mặc dù các chất đồng vị phóng xạ rất hữu ích để nghiên cứu sinh học và y học, song sự phóng xạ từ các chất đồng vị bị phân huỷ cũng nguy hiểm cho sự sống do nó phá huỷ các phân tử của tế bào. Tính nghiêm trọng của các tổn thương đó phụ thuộc vào loại và lượng phóng xạ mà cơ thể hấp thụ. Một trong số những mối nguy hiểm nghiêm trọng nhất cho môi trường là đồng vị phóng xạ thải ra từ các sự cố hạt nhân. Tuy nhiên, liều lượng của các chất đồng vị dùng trong chẩn đoán y học là tương đối an toàn.
Tất cả các amino axit đều có cấu trúc chung. Các amino axit là các phân tử hữu cơ có cả nhóm cacboxyl và nhóm amin. Ở trung tâm là Cacbon không đối xứng (Cacbon alpha). Bốn đối tác của nó là nhóm amin, nhóm carboxyl, nhóm hydrogen và nhóm biến động kí hiệu là R, còn gọi là chuỗi phụ, khác nhau đối với mỗi amino axit.
Hệ thống màng nội bào điều hoà sự vận chuyển protein và thực hiện các chức năng chuyển hoá trong tế bào:
Nhiều loại màng khác nhau của tế bào nhân thực là một phần của hệ thống màng nội bào, hệ thống màng thực hiện nhiều nhiệm vụ khác nhau trong tế bào. Các nhiệm vụ đó bao gồm tổng hợp protein và vận chuyển chúng đến màng tế bào và bào quan hoặc ra khỏi tế bào, chuyển hoá và vận động của các lipit, và khử độc các chất độc. Các màng của hệ thống này, hoặc là sự kế tiếp vật lí của nhau một cách trực tiếp, hoặc do tách chuyển một đoạn màng dưới dạng các túi nhỏ
Nhiều loại tế bào tiết các protein do các ribosome gắn với các ER tạo ra. Khi protein mới được tổng hợp vào khoang ER, nó cuộn xoắn thành hình dạng tự nhiên. Hầu hết các protein tiết đều là glycoprotein, loại protein có carbonhydrat liên kết cộng hoá trị với chúng. Carbonhydrat gắn với protein trong ER nhờ các phân tử chuyên hoá được tạo ra ở màng ER.

Các nhà khoa học sử dụng các chất phóng xạ để đánh dấu các hợp chất hoá học nhất định, tạo ra các chất đánh dấu để có thể theo dõi quá trình trao đổi chất hoặc định vị hợp chất trong cơ thể. Trong ví dụ này, các đồng vị phóng xạ còn được dùng như các chất đánh dấu để theo dõi các nguyên tử trong quá trình trao đổi chất - các quá trình hoá học của cơ thể. Tế bào sử dụng được cả các nguyên tử phóng xạ cũng như các chất đồng vị không phóng xạ của một nguyên tố, nhưng các chất đánh dấu phóng xạ thì dễ dàng bị phát hiện.
Các chất đồng vị phóng xạ là phương tiện chẩn đoán quan trọng trong y học. Ví dụ những hội chứng nhất định ở thận có thể được chẩn đoán bằng cách tiêm vào một lượng nhỏ các chất chứa đồng vị phóng xạ vào máu, sau đó đo lượng chất đánh dấu tiết ra ở nước tiểu. Các chất đánh dấu phóng xạ còn được sử dụng phối hợp với các dụng cụ hiện hình tinh vi, như máy nội soi PET- máy có thể theo dõi các quá trình hoá
Sau khi các protein tiết được hình thành, màng ER giữ chúng tách biệt khỏi các protein do các ribosone tự do tạo ra và duy trì chúng trong bào tương. Các protein tiết rời khỏi ER trong các túi có màng bao, giống như các bong bóng, nảy ra từ các vùng chuyên hoá (vùng ER chuyển tiếp). Các túi vận chuyển từ vùng này đến vùng khác của tế bào được gọi là túi vận chuyển (túi tải)
- Sau khi rời ER, nhiều túi vận chuyển đi đến bộ máy golgi là trung tam sản xuất, kho chứa hàng, nơi phân loại và vận chuyển. Ở đây, các sản phẩm của ER, như protein, được sửa đổi, dự trữ và sau đó được gửi tới những chỗ khác nhau.
+ Túi vận chuyển của bộ máy Golgi làm nhiệm vụ đưa các phân tử prôtêin đến các bào quan của tế bào hoặc đến màng sinh chất, hoặc đưa ra bên ngoài tế bào.
+ Màng sinh chất có chức năng vận chuyển các chất qua màng, tiếp nhận thông tin từ bên ngoài tế bào, nhận biết giữa các tế bào, tạo hình dạng tế bào.
Câu 2
Trong tế bào động vật có hai loại bào quan đều thực hiện chức năng khử độc, đó là hai loại bào quan nào? Cơ chế khử độc của hai loại bào quan đó có gì khác nhau?
Hướng dẫn vận dụng kiến thức: phần Sinh học tế bào - cấu trúc và chức năng của các bào quan:
Hóa sinh: cơ chế khử độc của các bào quan:
Lưới nội chất trơn: Chức năng tổng hợp lipit, chuyển hoá các cacbonhydrat, khử độc các thuốc và chất độc. Các enzyme của ER trơn có vai trò quan trọng trong quá trình tổng hợp lipit, bao gồm dầu thực vật, phospholipid và steroid. Cac enzyme khác của ER trơn giúp khử độc thuốc và chất độc, đặc biệt là ở các tế bào gan. Sự khử độc thường là bổ sung nhóm hydroxyl vào các phân tử thuốc làm cho chúng dễ tan hơn và dễ dàng đẩy ra khỏi cơ thể. Thuốc giảm đau và thuốc an thần là những ví dụ về những loại thuốc được trao đổi chất theo cách đó bởi ER trơn ở các tế bào gan. Trong thực tế, các thuốc an thần, alcohol và nhiều loại thuốc khác kích thích sự sinh sôi của ER trơn và các enzyme khử độc liên kết với nó, nhờ vậy làm tăng tốc độ khử độc. Điều đó lại làm tăng sự chịu đựng đối với thuốc, nghĩa là ngày càng cần liều cao hơn mới đạt hiệu quả, ví dụ như để giảm đau.
Ngoài ra, vì một số enzyme khử độc có phổ hoạt động tương đối rộng nên sự sinh sôi của ER trơn để đáp ứng với một loại thuốc có thể làm tăng sự chịu đựng với những loại thuốc khác. Ví dụ, sự lạm dụng thuốc an thần có thể làm giảm hiệu qủa của những thuốc kháng sinh nhất định và những thuốc có ích khác.
Peroxixom là khoang trao đổi chất chuyên hoá, được bao bọc bởi màng đơn.
Peroxixom chứa các enzyme truyền hidro từ các cơ chất khác nhau đến oxygen (O2)
tạo ra hydropeoxit (H2O2) như một sản phẩm phụ. Một số peroxixom dùng oxygen để phá huỷ các axit béo thành các phân tử nhỏ hơn để vận chuyển đến ti thể, nơi chúng
được dung làm nguyên liệu cho hô hấp tế bào. Các peroxixom trong gan khử độc ancohol và các hợp chất độc hại khác bằng cách truyền hydrogen từ các chất độc đến oxygen. H2O2 do các peroxixom tạo ra rất độc nhưng bào quan này còn chứa enzyme chuyển hoá H2O2 thành nước. Đây là một ví dụ tuyệt vời cho thấy cấu trúc phân chia xoang của tế bào có ý nghĩa sống còn với các chức năng của nó: các enyme sinh ra hydrogen peroxide và các enzyme khử bỏ hợp chất độc hại này được cô lập ở cùng 1 nơi, tách khỏi các thành phần khác có thể bị tổn thương của tế bào.
Loại peroxisome chuyên hoá có tên là glyoxisome được tìm thấy ở các mô dự trữ chất béo của hạt cây. Bào quan này chứa các enzyme khởi đầu của chuyển hoá các acid béo thành đường để các cây non mới mọc dung làm nguồn năng lượng và nguồn Carbon cho đến khi nó có thể tự tạo ra nhờ quang hợp.
Không giống như lysosome, các peroxisome không mọc ra từ hệ thống màng nội bào. Chúng to lên nhờ kết hợp thêm các protein được tạo ra ở dịch bào, lipit được
tạo ra ở ER và lipit tự do chúng tự tổng hợp. Số lượng các peroxisome có thể tăng lên bằng cách phân đôi khi chúng đạt kích thước nhất định.
Câu 3 (Trích đề thi HSGQG 2015 ngày 1)
Hãy phân biệt quá trình phiên mã và quá trình sao chép trong tế bào chủ của virut chứa hệ gen RNA (+) với virut chứa hệ gen DNA về: nơi phiên mã, enzyme dùng cho phiên mã, nơi sao chép, enzyme dùng cho sao chép. Quá trình phiên mã có trùng với quá trình sao chép không?
Hướng dẫn vận dụng kiến thức:
Kiến thức phần sinh học vi sinh vật: phân biệt virut RNA+ và virut DNA
Kiến thức phần di truyền: phần sao chép DNA và phiên mã
Hướng dẫn trả lời: Virut RNA (+) Virut DNA
Nơi phiên mã Trong tế bào chất Trong nhân tế bào
Enzyme dùng cho phiên mã
RNA polimeraza phụ thuộc RNA của virut
RNA polimeraza phụ thuộc DNA của tế bào
Nơi sao chép Trong tế bào chất Trong nhân tế bào
Enzyme dùng cho sao chép
RNA polimeraza phụ thuộc RNA của tế bào
DNA polimeraza phụ thuộc DNA của virut
Ở virut chứa hệ gen RNA (+), quá trình phiên mã trùng với quá trình sao chép còn ở virut chứa hệ gen DNA quá trình phiên mã không trùng với quá trình sao chép.
Câu 4
Phân lập từ nước dưa chua thu được vi khuẩn Streptococcus faecalis. Nuôi vi khuẩn này trên môi trường cơ sở gồm các chất sau đây: 1,0 gam NH4Cl; 1,0 gam K2HPO4; 0,2 gam MgSO4; 0,1 gam CaCl2; 5,0 gam glucôzơ; các nguyên tố vi lượng Mn, Mg, Cu, Co, Zn (mỗi loại 2.10-5 gam) và thêm nước vừa đủ 1 lít. Thêm vào môi trường cơ sở các hợp chất khác nhau trong các thí nghiệm từ 1 đến 3 dưới đây, sau đó đưa vào tủ ấm 37oC và giữ trong 24 giờ, kết quả thu được như sau:
Thí nghiệm 1: môi trường cơ sở + axit folic → không sinh trưởng.
Thí nghiệm 2: môi trường cơ sở + pyridoxin → không sinh trưởng.
Thí nghiệm 3: môi trường cơ sở + axit folic + pyridoxin → có sinh trưởng.
a) Dựa theo nguồn cung cấp năng lượng; nguồn cacbon; chất cho electron; các chất thêm vào môi trường cơ sở thì vi khuẩn Streprococcus faecalis có kiểu dinh dưỡng nào?
b) Các chất thêm vào môi trường cơ sở có vai trò như thế nào đối với vi khuẩn Streprococcus faecalis?
Hướng dẫn vận dụng kiến thức: Vi sinh vật nguyên dưỡng, khuyết dưỡng và các nhân tố sinh trưởng
Hướng dẫn trả lời
a) Vi khuẩn có kiểu dinh dưỡng:
Theo nguồn năng lượng: là hóa dưỡng vì vi khuẩn dùng năng lượng được tạo ra từ chuyển hóa glucozơ thành axit lactic
Theo nguồn cacbon: là dị dưỡng vì glucozơ là nguồn cacbon kiến tạo nên các chất của tế bào.
Theo nguồn cho electron: là dinh dưỡng hữu cơ vì glucozơ là nguồn cho electron trong lên men lăctic đồng hình.
Theo các chất thêm vào môi trường cơ sở: là vi khuẩn khuyết dưỡng, thiếu 1 trong 2 chất trên vi khuẩn không phát triển được.
b) Các chất thêm vào môi trường cơ sở có vai trò:
Các chất folic, pyridoxin là các nhân tố sinh trưởng đối với vi khuẩn nêu trên.
Thiếu 1 chất trong 2 chất này thì vi khuẩn không thể tự tổng hợp được và không sinh trưởng.
Axit folic là một loại vitamin giúp hình thành tổng hợp purin và pirimidin. Pyridoxin là vitamin B6 giúp chuyển amin của các axit amin.
Câu 5
Nêu sự khác biệt về quá trình xâm nhập và cởi vỏ giữa virut của vi khuẩn (phagơ) với virut động vật. Nếu bơm prôtôn trong lizôxôm không hoạt động sẽ ảnh hưởng như thế nào đến quá trình xâm nhập và nhân lên của 2 loại virut trên?
Hướng dẫn vận dụng kiến thức:
+ Sinh học vi sinh vật: sự xâm nhập và nhân lên của virut trong tế bào chủ:
+ Sinh học tế bào: Vai trò của lizoxom trong quá trình xâm nhập của virut động vật: lizoxom là túi chứa các enzyme thuỷ phân có màng bao mà các tế bào động vật dung để tiêu hoá các đại phân tử. Các enzyme của lizoxom hoạt động tốt nhất trong môi trường acid. Nếu lizoxom bị phá vỡ hoặc rò rỉ chất chứa của nó thì các enzyme được giải phóng ra hoạt động không mạnh vì dịch bào có pH trung tính. Tuy nhiên, sự
rò rỉ quả nhiều từ một số lượng lướng lizoxom có thể phá huỷ tế bào bằng cách tự tiêu hoá. Cac enzyme thuỷ phân và màng lizoxom do ER hạt tạo ra và sau đó được truyền đến bộ máy Golgi để xử lý tiếp.
Lizoxom thực hiện sự tiêu hoá nội bào trong nhiều hoàn cảnh. Trùng amip và nhiều protisst khác ăn bằng cách nhấn chìm (nuốt) các sinh vật nhỏ hoặc các hạt thức ăn, quá trình đó gọi là sự thực bào (phagocytosis). Sau đó không bào thức ăn được hình thành theo cách này, kết hợp với lizoxom để các enzyme tiêu hoá thức ăn. Các sản phẩm tiêu hoá, bao gồm đường đơn, amino acid và các đơn phân khác, đi vào dịch bào và trở thành chất dinh dưỡng cho tế bào. Một số tế bào người cũng tiến hành thực bào. Trong số đó, có các đại thực bào, loại tế bào bạch cầu bảo vệ cơ thể bằng cách “nuốt” và tiêu huỷ vi khuẩn và các kẻ xâm nhập khác.
Lizoxom cũng dung các enzyme thuỷ phân để tái quay vòng vật chất hữu cơ riêng của tế bào, đó là quá trình tự thực bào. Trong quá trình này, bào quan bị tổn thương hoặc một lượng nhỏ dịch bào bị bao bởi lớp màng kép chưa rõ nguồn gốc và lizoxom kết nối với màng ngoài của túi đó. Các enzyme của lizoxom phá huỷ vật chất được bao bọc bên trong và các monomer được đưa trở lại dịch bào để tái sử dụng. Với sự trợ giúp của lizoxom tế bào liên tục tự đổi mới. Ví dụ, các tế bào gan người, mỗi tuần, tái quay vòng một nửa các đại phân tử của nó.
Tế bào của những người mắc bệnh dự trữ lizoxom di truyền không có enzyme thuỷ phân hoạt động chức năng bình thường trong các lizoxom. Các lizoxom bị nhét đầy các cơ chất không được tiêu hoá bắt đầu ảnh hưởng đến các hoạt động khác của tế bào. Ví dụ, bệnh Tay-sachs, enzyme tiêu hoá lipid bị mất hoặc không hoạt động và não bị hư hại do sự tích luỹ lipit trong tế bào. Rất may là bệnh dự trữ lizoxom rất hiếm trong quần thể.
- Trong quá trình xâm nhập của virut vào tế bào động vật: Nếu bơm prôtôn trong lizoxom không hoạt động, môi trường không bị axit hóa, các enzyme không được hoạt hóa để phân giải capsit thì axit nucleic của virut động vật không được giải phóng khỏi vỏ capsit dẫn đến virut động vật không nhân lên được.
- Quá trình cởi vỏ capsit và nhân nhân lên ở phagơ không sử dụng bơm prôtôn trong lizoxom của tế bào.
Câu 6 (IBO 2013)
Bilirubin là sản phẩm phân huỷ tạo ra từ quá trình dị hoá nhân heme và được đưa đến gan, tại đây nó liên kết với hai phân tử axit glucuronic nhờ enzim UGT (xem hình phía dưới). Bilirubin liên kết sau đó được tiết vào ruột non như là một thành phần của dịch mật:
Hãy chỉ ra các phát biểu sau là đúng hay sai
A. Liên kết với axit glucoronic làm tăng tính hoà tan của bilirubin trong nước
B. Một khối u làm tắc đoạn ống dẫn mật sát với ruột non dẫn đến làm giảm hàm lượng bilirubin liên kết trong máu
C. Một đột biến điểm gây giảm đáng kể hoạt tính của UGT dẫn đến làm giảm hàm lượng bilirubin không liên kết trong máu.
D. Tăng hàm lượng bilirubin liên kết trong máu là triệu chứng của nhiễm trùng sốt rét
Hướng dẫn vận dụng kiến thức: Hoá sinh enzim, đột biến gen
Hướng dẫn trả lời
A. Đúng, B. Sai, C. Sai, D. Đúng Giải thích
A. Đúng - Axit glucoronic là một phân tử hydrpphilic, trong khi bilirubin kỵ nước và không hòa tan trong nước. Liên kết với axit glucoronic làm tăng khả năng hòa tan của nó trong nước
B. Sai - Khi mật không thể vào ruột non, bilirubin liên hợp sẽ tích tụ trong ống mật và di chuyển ngược lên trong đường mật trong gan và đi vào máu. Kết quả là, sự liên hợp bị giảm hoặc thậm chí ngừng gây ra sự tích tụ của bilirubin không liên hợp.
C. Sai - Nếu UGT không hoạt động bình thường (như ở bệnh nhân Morbus Meulengracht), sự liên hợp của bilirubin bị giảm và mức độ bilirubin không liên hợp tăng lên.
D. Đúng - Ở một giai đoạn, Plasmodium falciparum sinh sản trong hồng cầu. Các khi hồng cầu này vỡ ra giải phóng các ký sinh trùng, dẫn đến giải phóng hemoglobin, do đó làm tăng nồng độ bilurubin trong máu, chất này sẽ được liên hợp.
Câu 7 (QG2015 ngày 2)
Một trong các protein đặc thù của phức hệ kết dính các nhiễm sắc tử trong giảm phân là protein Rec8. Protein chỉ được biểu hiện trong giảm phân, không phân rã trong giảm phân I, nhưng bị phân rã trong giảm phân II, nhờ vậy các nhiễm sắc tử chỉ được tách khỏi nhau trong giảm phân II. Từ hiện tượng đó, các nhà khoa học cho rằng trong tế bào có một loại protein ngăn không cho Rec8 phân rã ở giảm phân I. Người ta tạo tế bào nấm men chuyển gen mang nhiễm sắc thể nhân tạo nấm men (YAC) chứa gen Rec8 và cho biểu hiện trong nguyên phân, các tế bào được chuyển gen có thể sống bình thường. Để tìm gen ngăn sự phân rã của Rec8, người ta cho biểu hiện các gen khác trong dòng tế bào biểu hiện Rec8 trong nguyên phân, từ đó tìm ra gen mã hóa protein Shugoshin chính là protein ngăn Rec8 không phân rã ở giảm phân I. Hãy dự đoán:
85
a) Bằng cách nào người ta biết là protein Shugoshin mà không phải là các protein khác ngăn sự phân rã của Rec8?
b) Kiểu hình của tế bào tái tổ hợp biểu hiện đồng thời Rec8 và Shugoshin sẽ như thế nào?
Hướng dẫn vận dụng kiến thức:
- Phân bào giảm phân: Các nhiễm sắc tử chị em đính với nhau trong suốt giảm phân I nhưng lại tách nhau ra trong giảm phân II và trong nguyên phân như thế nào?
Các nhiễm sắc tử được gắn với nhau theo suốt chiều dài của chúng bằng các phức hợp protein được gọi là cohensin. Trong nguyên phân sự gắn kết này kéo dài tới tận cuối kì giữa, khi enzyme phân huỷ cohensin làm cho các nhiễm sắc tử có thể di chuyển về các cực đối lập của tế bào. Trong giảm phân, sự gắn kết các nhiễm sắc tử được giải phóng qua hai bước. Ở kì giữa I, các nhiễm sắc thể được giữ với nhau bởi sự gắn kết giữa các vai của các nhiễm sắc tử trong các vùng mà ở đó DNA đã được trao đổi. Trong kì sau I, cohensin được loại bỏ ở các vai cho phép các NST tương đồng tách nhau ra. Ở kì giữa II, cohensin được loại bỏ ở tâm động cho phép các nhiễm sắc tử tách rời nhau.
Các thí nghiệm được trình bày bởi Yoshinori Watanabe và các cộng sự ở trường đại học Tokyo đã biết rằng có các protein tương tự nhau trong các phức hợp cohensin ở nguyên phân và giảm phân, họ muốn biết rằng cái nào làm nhiệm vụ ngăn cản sự phân huỷ cohensin ở tâm động trong khi sự phân huỷ lại xảy ra ở vài vào cuối kì giữa I. Họ đã tìm thấy một protein có tên là shugosin (tiếng Nhật có nghĩa là “tinh thần vệ sĩ”) bảo vệ cohensin khỏi bị phân huỷ ở tâm động trong suốt giảm phân I. Shugosin tương tự protein của ruồi quả đã được nhận biết cách đó 10 năm bởi Terry OrrWeaver. Giảm phân I được gọi là phân bào giảm nhiễm vì nó làm giảm số bộ nhiễm sắc thể trong mỗi tế bào đi một nửa-giảm từ hai bộ (trạng thái lưỡng bội). Trong giảm phân II, (đôi khi còn gọi là phân bào đồng đều) các nhiễm sắc tử chị em phân ly tạo ra các tế bào đơn bội. Cơ chế phân ly nhiễm sắc tử chị em thực tế cũng y hệt nhau trong giảm phân II và trong nguyên phân. Cơ sở phân tử của hoạt động của các nhiễm sắc thể trong giảm phân còn tiếp tục là tâm điểm thu hút sự quan tâm nghiên cứu của các nhà khoa học.
Yoshinori Watanabe và các cộng sự đã biết rằng trong kì sau I, protein shugosin chỉ có ở xung quanh tâm động. Họ muốn biết liệu nó có bảo vệ cohensin khỏi bị phân huỷ trong giảm phân I để đảm bảo cho các nhiễm sắc tử vẫn gắn với nhau trong khi các NST tương đồng phân ly hay không. Để kiểm tra giả thuyết này, họ đã sử dụng một loài nấm men, trong đó giảm phân tạo ra các bào tử đơn bội nằm thẳng hàng theo một trình tự đặc biệt trong nang bào tử Để theo dõi sự di chuyển của các nhiễm sắc thể, họ đã đánh dấu huỳnh quang vùng gần tâm động của cả hai nhiễm sắc tử của một nhiễm sắc thể tương đồng, còn vùng này của nhiễm sắc thể tương đồng còn lại thì không đánh dấu. Sau đó họ đã làm bất hoạt gen mã hoá shugosin và so sánh dòng nấm men này (shugosin-) với dòng nấm men bình thường (shugosin+). Các nhà nghiên cứu đã kì vọng rằng hai nhiễm sắc thể đã được đánh dấu hình thành từ hai nhiễm sắc tử
trong tế bào bình thường sẽ phân ly về các bào tử khác nhau, nằm mỗi cái ở một đầu của túi bào tử. Tiếp đó, họ cũng tiên đoán rằng nếu shugosin bảo vệ cohensin ở tâm động khỏi bị phân huỷ ở kì sau I thì các nhiễm sắc thể được đánh dấu trong các tế bào shugosin- sẽ phân ly một cách ngẫu nhiên trong giảm phân II, đôi khi chúng lại ở cùng một bào tử
Hướng dẫn vận dụng kiến thức:
Công nghệ sinh học: Phương pháp điện di trên gel:
Ứng dụng: Điện di trên Gel là phương pháp được dùng để phân tách các acid nucleic hoặc protein khác nhau về kích thước, độ tích điện, hoặc các thuộc tính vật lí khác. Các phân tử DNA được phân tách bởi diện di trên gel trong các thí nghiệm phân tích các doạn giới hạn đối với cả các gen được nhân dòng.

Kĩ thuật: Phương pháp này phân tách các đại phân tử trên cơ sở tốc độ di chuyển trên điện trường qua gel polyme. Khoảng cách di chuyển của một phân tử DNA tỷ lệ nghịch với chiều dài của nó. Một hỗn hợp các phân tử, thường là các đoạn cắt giới hạn hoặc là sản phẩm PCR, được phân tách thành các băng. Mỗi băng chứa hàng nghìn phân tử có kích thước giống nhau.
bào shugosin+ hai nhiễm sắc thể đánh dấu đi về các bào tử p. Trong các tế bào shugosin-chừng 50% các trường hợp chúng lại ở cùng với nhau trong cùng một bào tử
Kết luận: Shugosin bảo vệ cohensin ở tâm động trong kì sau I, nhờ vậy duy trì
được sự gắn kết giữa các nhiễm sắc tử chị em và đảm bảo cho chúng phân ly bình thường trong giảm phân II.
Hướng dẫn trả lời
a) Với các tế bào có gen Shugoshin được biểu hiện (có protein Shugoshin) thì sự phân li NST trong nguyên phân diễn ra không bình thường, còn các tế bào có các gen khác được biểu hiện nguyên phân diễn ra bình thường.
b) Vì có cả Shugoshin và protein Rec 8 cùng được biểu hiện trong nguyên phân, nên có nhiễm sắc thể được phân ly, nhiễm sắc thể khác không phân li, do sự ngăn cản của Shugoshin. Kết quả tạo ra nhiều loại tế tế bào con lệch bội khác nhau.
Một số tế bào có thể có 4 bản sao của cùng một 1 NST, một số có 2 bản sao của cùng 1 NST, trong khi một số thiếu (không có) một NST nào đó. Nhiều kiểu gen này gây chết, dẫn đến kiểu hình là khuẩn lạc nấm men chậm phát triển.
Câu 8. (Trích đề thi HSG QG 2015 ngày 2)

Để tìm một loại protein là yếu tố phiên mã được tổng hợp ở một loại mô nhất định, người ta phải tách chiết các yếu tố phiên mã ra khỏi tế bào. Hãy mô tả cách làm tiếp theo để nhận biết được sự có mặt của yếu tố phiên mã cần tìm trong hỗn hợp protein tách chiết (nêu tóm tắt nguyên lý và các bước cần tiến hành).
Phương pháp điện di trên Gel
Phương pháp thẩm tách western-blot
Phương pháp này sử dụng điện di trên gel để phân tách các protein còn nguyên vẹn bằng cấu trúc 3D hay protein đã biến tính theo độ dài của mạch polypeptide.
Protein này sau đó sẽ được chuyển lên màng (thường sử dụng là màng nitrocellulose hay màng PVDF), ở đó chúng được gắn bằng các kháng thể đặc hiệu với protein mục tiêu. Điện di trên gel sẽ được đưa vào hệ thống phân tích Western Blot nhằm giải quyết các vấn đề của các phản ứng chéo giữa các kháng thể.
Phương pháp Western Blot


Western Blot là kỹ thuật lai giữa protein với protein (Kháng nguyên – Kháng thể). Protein kháng nguyên được phát hiện qua phản ứng tạo màu hoặc phát huỳnh quang
2. Các bước thực hiện:
Bước 1: Xử lý mẫu
Lấy mẫu từ toàn bộ mô hay môi trường nuôi cấy tế bào. Đầu tiên, xử lý mô rắn bằng phương pháp cơ học sử dụng máy nghiền, sử dụng máy đồng hóa hoặc siêu âm. Những mẫu virus hay mẫu lấy trừ trong môi trường nuôi cấy cũng có thể là nguồn protein xác định được trong Western Blot. Thêm các loại chất tẩy rửa, muối và dung dịch đệm để ly giải tế bào và hòa tan protein.
Bước 2: Điện di trên gel
Điện di trên gel để phân tách các protein trong mẫu. Sự phân tách này của protein dựa trên điểm đẳng điện, khối lượng phân tử, điện tích hoặc phối hợp các yếu tố trên.
Phương pháp điện di phổ biến nhất là điện di trên gel polyacrylamide và có bổ sung sodium dodecyl sulfate (SDS). Phương pháp SDS – PAGE (điện di trên gel polyacrylamide và có bổ sung SDS) giữ các polypeptide ở trạng thái biến tính sau khi chúng được xử lý với chất khử mạnh và mất đi cấu trúc bậc 2, cấu trúc bậc 3.
Các mẫu được nạp vào trong các giếng của bản gel. Thường để lại một giếng cho chỉ thị (marker) hoặc thang chuẩn ladder, là một sản phẩm thương mại có chứa hỗn hợp các protein với khối lượng phân tử xác định được nhuộm để có thể tạo ra band màu và nhìn thấy được. Khi điện thế được thiết lập trên gel, protein bắt đầu di chuyển với tốc độ khác nhau phụ thuộc vào khối lượng phân tử của chúng. Vận tốc khác nhau của các protein (sự khác biệt về độ di động điện di) sẽ tạo thành vạch khác nhau trên mỗi giếng.
Bước 3: Chuyển protein gel lên màng lai
Để các protein có thể phát hiện được kháng thể thì protein phải được chuyển từ gel lên màng lai. Phương pháp chính để chuyển các protein được gọi là phương pháp thẩm tách điện là sử dụng một dòng điện để kéo các protein từ gel lên màng PVDF hoặc màng nitrocellulose. Các protein được chuyển lên màng mà vẫn duy trì sự sắp xếp như trên bản gel. Kết quả là protein sẽ được phơi ra trên lớp mỏng bề mặt để tiến hành xác định
Bước 4: Xử lý màng lai (Blocking)
Do kháng thể và protein đích đều là protein, nên thường thực hiện các bước để ngăn chặn liên kết giữa màng và kháng thể được sử dụng để xác định protein mục tiêu.
Việc ngăn chặn các liên kết không đặc hiệu bằng cách đặt màng vào dung dịch protein loãng albumin huyết tương bò 3-5 % (BSA) hoặc sữa bột không béo trong Tris –Buffered saline (TBS). Protein trong dung dịch loãng sẽ gắn với màng ở tất cả các vị trí mà protein đích chưa gắn vào. Do đó khi kháng thể được bổ sung vào, sẽ không còn chỗ bám trên màng ngoại trừ các vị trí liên kết đặc hiệu với protein đích. Điều này làm giảm nhiễu cơ bản trong sản phẩm cuối cùng của Western Blot, cho kết quả rõ ràng và ngoại trừ hiện tượng dương tính giả.
Bước 5: Quá trình lai và phát hiện vị trí lai
Để phát hiện các protein chuyên biệt, màng được dò protein quan tâm bằng kháng thể đã biến đổi có khả năng liên kết với enzyme thông báo, enzyme này khi kết hợp với một cơ chất tương ứng sẽ thúc đẩy phản ứng hấp thụ quang và tạo màu. Quá trình này diễn ra gồm 2 bước:
- Màng lai đã cố định protein được lai với kháng thể sơ cấp. Kháng thể sơ cấp là một kháng thể đặc hiệu được tạo ra khi cho vật chủ hay môi trường nuôi cấy tế bào miễn dịch tiếp xúc với protein quan tâm.
- Sau khi rửa màng để loại bỏ kháng thể sơ cấp chưa liên kết, màng tiếp tục gắn kháng thể thứ hai (kháng thể thứ cấp) có gắn enzyme (thường sử dụng horseradish peroxidase). Tiếp tục ủ màng lai trong một hỗn hợp phản ứng đặc hiệu với enzyme. Đặt 1 tấm phim ngạy cảm với tia X lên màng lai để phát hiện các điểm sáng phát ra do enzyme.
Có thể sử dụng nhiều phương pháp để phát hiện kháng thể thứ cấp: sử dụng bức xạ hồng ngoại gần liên kết với kháng thể gắn chất huỳnh quang, sử dụng đánh dấu phóng xạ
3. Ứng dụng:
Xác định sự hoạt động của gel thông qua sự có mặt của protein trong mô
Nhận dạng protein mục tiêu
Đánh giá tính chuyên biệt của kháng thể
Phân tích bệnh do vi khuẩn và virus gây ra: thử nghiệm khẳng định HIV, viêm gan B.
Năm 2002, đã ứng dụng thành công phương pháp Western Blot trong việc xác định kháng nguyên giun đũa chó (Toxocara canis) trên thỏ (Olga Lucia Morales và ctv, 2002), mở ra một hướng đi mới trong việc xét nghiệm chẩn đoán chính xác các bệnh ký sinh trùng trên người.
Hướng dẫn trả lời
- Học sinh cần nói được bước đầu tiên cần tiến hành điện di để tách phân đoạn protein khỏi nhau. Vì các đoạn protein có các phần tích điện trái dấu khác nhau nên
trước khi điện di, các protein phải được xử lý hóa chất để cho các phân đoạn protein đều được tích điện âm (âm hóa) thì chúng mới di chuyển được trong điện trường.
- Tiếp theo cần tiến hành thẩm tách để chuyển các phân đoạn protein từ gel điện di lên màng thẩm tách (thẩm tách Western) để cố định các protein trên màng thẩm tách.
- Nhận biết protein cần tìm dựa trên phản ứng kháng nguyên kháng thể đặc hiệu bằng cách cho kháng thể đánh dấu tiếp xúc với các phân đoạn protein, ở đâu có yếu tố phiên mã liên kết được với kháng thể thì sẽ có thể thì ta có thể nhận ra.
Câu 9 (Trích đề thi HSGQG 2016 ngày 1)
Có hai ống nghiệm bị mất nhãn, trong đó có một ống nghiệm chứa nấm men Saccharomyces cerevisiae (S. cerevisiae) và ống nghiệm còn lại chứa vi khuẩn Escherichia coli (E. coli). Hãy đưa ra 04 phương pháp giúp nhận biết ống nghiệm nào chứa nấm men S. cerevisiae và ống nghiệm nào chứa vi khuẩn E. coli
Hướng dẫn vận dụng kiến thức: hoá sinh tế bào vi sinh vật S. cerevisiae là sinh vật nhân thực có hình bầu dục, kích thước lớn có thể quan sát dưới kính hiển vi với độ phóng đại 400 lần. Trong khi đó, E. coli là vi khuẩn sinh vật nhân sơ, hình que, kích thước của E. coli nhỏ hơn nhiều so với S. cerevisiae nên phải phóng to ít nhất 1000 lần mới nhìn thấy rõ hình thái tế bào. Cấu trúc thành tế bào vi khuẩn E. coli có chứa hợp chất peptidoglycan rất mẫn cảm với kháng sinh. Nấm men có thể sinh sản bằng nảy chồi hoặc sinh sản hữu tính, vi khuẩn phân chia theo lối trực phân.
Nấm men S. cerevisiae có khả năng thực hiện chuyển hoá piruvat trong điều kiện thiếu oxi thành etanol (lên men rượu).
Hướng dẫn trả lời:
- Phương pháp 1: Làm tiêu bản và quan sát tế bào dưới kính hiển vi, S. cerevisiae là sinh vật nhân thực có hình bầu dục, kích thước lớn có thể quan sát dưới kính hiển vi với độ phóng đại 400 lần. Trong khi đó, E. coli là vi khuẩn sinh vật nhân sơ, hình que, kích thước của E. coli nhỏ hơn nhiều so với S. cerevisiae nên phải phóng to ít nhất 1000 lần mới nhìn thấy rõ hình thái tế bào.
- Phương pháp 2: Dùng phương pháp lên men dịch ép hoa quả để phân biệt hai loài: sử dụng dịch chiết hoa quả vô trùng chia đều ra hai bình như nhau rồi cấy vi sinh vật vào. Sau đó bịt kín bình và giữ ở nhiệt độ, thời gian thích hợp. Nếu bình nào sinh ra nhiều CO2, tạo ra nhiều bọt khí có mùi rượu thì bình đó chứa nấm men, bình còn lại là chứa E. coli (do E. coli không có khả năng lên men rượu)
- Phương pháp 3: Bổ sung vào hai ống nghiệm chất kháng sinh có khả năng ức chế sự phát triển của E. coli. ở ống nghiệm chứa E. coli thì E. coli sẽ không sinh trưởng và không phát triển được.
- Phương pháp 4: có thể kiểm tra bằng hình thức quan sát khuẩn lạc trên môi trường nuôi cấy vi khuẩn (MPA) và môi trường nuôi cấy nấm men (Hansen). E. coli
mọc kém hoặc không mọc trên môi trường nấm men, còn nấm men mọc tốt ở cả hai môi trường.
Câu 10 (Trích đề HSG QG 2016 ngày 1)
Một số loài vi khuẩn có thề sử dụng êtanol (CH3-CH2-OH) hoặc axêtat (CH3COOH) làm nguồn cacbon duy nhất trong quá trình sinh trưởng. Tốc độ hấp thụ ban đầu hai loại chất này của tế bào vi khuẩn được trình bày trong bảng dưới đây:
Nồng độ cơ chất (mM) Tốc độ hấp thụ của Vi khuẩn (µmol/phút) Ch
t A Chất B
Vẽ đồ thị biểu diễn mồi quan hệ giữa tốc độ hấp thụ ban đầu và nống độ của hai chất trên
Dưa vào đồ thị đã vẽ, hãy cho biết: Sự vận chuyển của hai chất A và B qua màng tể bào vi khuẩn theo cách nào? Giải thích.
Hai chất A và B, chất nào là êtanol và chất nào là axêtat? Giải thích
Hướng dẫn vận dụng kiến thức:
+ Hoá sinh: Sự vận chuyển các chất qua màng sinh chất
Vận chuyển thụ động: là sự khuếch tán các chất qua màng mà không cần năng lượng. Sự khuếch tán diễn ra khi không có các lực khác tác động thì một chất sẽ khuếch tán từ nơi có nồng độ cao đến nơi có nồng độ thấp hơn. Đây là quá trình vận chuyển thụ động vì tế bào không phải chi phí năng lượng. Gradien nồng độ bản thân nó là thế năng và điều khiển sự khuếch tán. Tuy nhiên, màng có tính thấm chọn lọc vì vậy nó có hiệu ứng khác nhau với tốc độ khuếch tán của các đại phân tử khác nhau
Khuếch tán tăng cường: sự vận chuyển thụ động nhờ protein: Câu hỏi đặt ra là bằng cách nào mà nước và các chất tan ưa nước có thể đi qua màng, vì màng có thành phần chính là photpholipt sẽ cản lại các phân tử phân cực và các ion. Với sự trợ giúp của các protein vận chuyển xuyên màng, nước và các chất tan ưa nước có thể dễ dàng vận chuyển qua màng sinh chất. Hầu hết protein vận chuyển rất đặc hiệu, chúng vận chuyển chất này mà không vận chuyển chất kia. Các protein tham gia vào quá trình này là protein kênh và protein mang. Tốc độ vận chuyển tỉ lệ thuận với số lượng
protein mang và protein kênh, khi tất cả các kênh đã bão hoà, tốc độ vận chuyển sẽ giảm.
Sự vận chuyển các chất qua màng tế bào phụ thuộc vào nhiều yếu tố: bản chất
của chất tan, kích thước, ....
+ Vi sinh: Cấu tạo MSC tế bào vi khuẩn
Hướng dẫn trả lời a. hình vẽ
b. Sự hấp thụ chất B qua đồ thị cho thấy tốc độ vận chuyển các chất vào trong tế
bào lúc đầu tăng cùng với việc tăng nồng độ các chất. Nhưng đến một giai đoạn nhất định thì tốc độ phản ứng gần như không tăng ngay kể cả khi nồng độ chất tan tiếp tục tăng lên. Chất B được vận chuyển qua kênh protein và việc vận chuyển của chất B không tăng ở giai đoạn sau là hiện tượng bão hòa kênh.
Sự hấp thụ chất A qua đồ thị cho thấy tốc độ vận chuyển chất tan phụ thuộc tuyến tính vào nồng độ chất tan. Điều này chỉ ra rằng chất A được khuếch tán trực tiếp qua lớp lipit kép của màng tế bào mà không cần phải qua kênh protein xuyên màng.
Từ đồ thị cho thấy:
- Chất A là etanol vì etanol là chất phân tử nhỏ, không tích điện nên có thể khuếch tán trực tiếp qua lớp lipit kép của màng tế bào dễ dàng hơn rất nhiều so với axêtat.
- Chất B là axêtat vì là chất tích điện nên sẽ khó khuếch tán trực tiếp qua lớp lipit kép của màng tế bào vì lớp phospholipit kép có chứa các đuôi hydrocacbon kị nước (không phân cực).
Câu 11 (Trích đề thi HSGQG 2016-ngày 2)
X là protein có tác dụng ngăn ngừa sự tăng cân ở người. Protein này bất hoạt ở những người béo phì. Các phân tử mRNA trưởng thành của X phân lập được từ một số người béo phì của cùng một gia đình cho thấy, chúng thiếu một đoạn trình tự dài 173 nucleotit so với các phân tử mRNA trưởng thành phân lập được từ những người bình thường. Khi so sánh trình tự gen mã hóa cho protein X của người bình thường và người béo phì, người ta phái hiện ra rằng không có nucleotit nào bị mất mà chỉ có 1 nucleotit bị thay đổi. Sự thay đổi này xảy ra ở vùng intron của gen.
Tại sao việc thay đổi 1 nucleotit lại có thể làm cho mRNA trưởng thành của gen đột biến bị mất một đoạn dài 173 nucleotit? Giải thích và minh họa bằng hình vẽ?
Giải thích hiện tượng bất hoạt của protein X ở người béo phì?
Để sản xuất protein này, người ta sử dụng kỹ thuật di truyền tạo plasmid tái tổ hợp giữa thể truyền bởi gen mã hóa protein X của người bình thường, sau đó chuyển vào vi khuẩn E. coli để sản xuất sinh khối. Sản phẩm protein tạo ra có bị bất hoạt không? Giải thích.

Hướng dẫn vận dụng kiến thức:
+ Di truyền: Đột biến gen thay thế nucleotit này bằng nucleotit khác.
+ Hoá sinh: Cấu tạo của hệ gen sinh vật nhân thực: Gen phân mảnh, xen giữa các đoạn mã hoá các axit amin (exon) là các đoạn không mã hoá (intron).
Điều hoà biểu hiện gen ở sinh vật nhân thực: Các enzyme trong nhân tế bào nhân thực có thể biến đổi phân tử tiền mRNA theo một số cách đặc trưng trước khi bản phiên mã di truyền được chuyển ra khỏi tế bào chất. Trong quá trình hoàn thiện mRNA này cả hai đầu của phân tử mRNA tiền thân đều được biến đổi. Ngoài ra trong phần lớn các trường hợp, một số phần bên trong phân tử RNA cũng được cắt bỏ, trong khi các phần còn lại được nối lại với nhau. Kết quả của những biến đổi này tạo ra một phân tử RNA sẵn sàng cho quá trình dịch mã. Gen ở sinh vật nhân thực là gen phân mảnh. Một giai đoạn đáng chú ý trong quá trình hoàn thiện mRNA trong nhân của sinh vật nhân thực là việc loại bỏ đi một phần lớn các đoạn bên trong phân tử mRNA tiền thân. Chiều dài trung bình của một đơn vị phiên mã dọc theo phân tử DNA của người gồm khoảng 27.000 cặp base, vì vậy, phân tử mRNA tiền thân thường có chiều dài tương ứng. Tuy vậy, để mã hoá một phân tử protein có kích thước trung bình gồm 400 amino acid, chỉ cần một phân tử RNA có kích thước gồm 1200 nucleotit (mỗi amino acid được mã hoá bởi một bộ ba nucleotit). Điều này có nghĩa là phần lớn các gen ở sinh vật nhân thực và các bản phiên mã RNA tiền thân của chúng chứa các đoạn dài không mã hoá, đây là những đoạn không được dịch mã. Những đoạn này thường nằm xen kẽ giữa các đoạn mã hoá của gen và tương ứng là giữa các đoạn mã hoá trên tiền mRNA. Nói cách khác, trình tự các nucleotit DNA mã hoá cho một chuỗi polipeptit thường không liên tục, chúng được phân tán thành các đoạn mã hoá (exon) và đoạn không mã hoá (intron). Để tạo ra một phiên bản tiền thân từ một gen, RNA polimeraza ban đầu tiến hành phiên mã toàn bộ gen, bao gồm cả các đoạn exon và intron. Tuy vậy phân tử mRNA đi vào tế bào chất là phân tử đã được cắt ngắn. Các intron được cắt khỏi phân tử, các exon được nối lại với nhau để hình thành nên một phân tử mRNA mang trình tự mã hoá liên tục. Quá trình này được gọi là sự cắt nối RNA. Sự cắt nối RNA được diễn ra nhờ các tín hiệu, chúng là các trình tự nucleotit ngắn ở hai đầu intron. Các hạt có tên là các ribonucleotitprotein nhân kích thước nhỏ (snRNP) có thể nhận ra các vị trí cắt nối này. Như tên gọi của chúng các snRNP có trong nhân tế bào và có thành phần cấu tạo gồm các phân tử RNA và protein. Các RNA có trong các snRNP là các RNA nhân kích thước nhỏ (snRNA), mỗi phân tử chỉ dài khoảng 150 nucleotit. Một số loại snRNP kết hợp với nhau và với một số protein bổ sung khác hình thành nên bộ máy cắt nối RNA và được gọi là spliceosome (hay thể cắt nối), chúng có kích thước lớn tương đương ribosome. Spliceosome tương tác với những vị
trí nhất định trên intron, giải phóng intron và nối hai exon ở hai đầu của mỗi intron, giải phóng intron và nối hai exon ở hai đầu của mỗi intron lại với nhau. Một số bằng chứng cho thấy, các snRNA, ngoài là thành phần của bộ máy spliceosome và nhận biết các vị trí cắt nối, còn có vai trò trực tiếp xúc tác các phản ứng cắt nối RNA.
Vai trò xúc tác của snRNA bắt nguồn từ việc phát hiện ra các ribozim, đó là các phân tử RNA có chức năng giống như các enzyme. Ở một số sinh vật, sự cắt nối RNA
ở một số gen có thể diễn ra mà không cần có bất cứ một loại protein hay thậm chí một loại RNA bổ sung nào khác. Cac đoạn intron của RNA có chức năng như một loại ribozyme và có khả năng tự xúc tác quá trình cắt-nối. RNA có ba thuộc tính giúp nó có thể biểu hiện chức năng như các enzyme. Thứ nhất, do RNA có cấu trúc mạch đơn nên một vùng trên phân tử có thể bắt cặp bổ sung với một vùng khác trên cùng phân tử đó, điều này giúp cho RNA có cấu trúc không gian đặc thù. Một cấu trúc không gian đặc thù là điều kiện tiên quyết để có chức năng xúc tác của ribosome, cũng giống như ở các enzyme là protein vậy. Thứ hai, giống với một số amino acid trong các enzyme là protein, một số base nucleotit của RNA mang các nhóm chức có thể tham gia vào các hoạt động xúc tác. Thứ ba, các RNA có khả năng hình thành liên kết hydrogen với các phân tử acid khác (RNA hoặc DNA), điều này làm tăng tính đặc hiệu trong quá trình xúc tác của nó. Chẳng hạn như, sự bắt cặp bổ sung giữa các base trên phân tử tiền mRNA giúp định vị chính xác vị trí mà các ribozim sẽ xúc tác phản ứng cắt nối RNA. Vậy câu hỏi đặt ra là chức năng sinh học của các intron và sự cắt nối RNA là gì? Phần lớn các intron hiện nay chưa xác định được rõ chức năng sinh học cụ thể, một số intron đã biết sẽ chứa các trình tự tham gia điều hoà hoạt động của gen. Một trong những hậu quả của việc gen có intron là một gen duy nhất có thể mã hoá cho nhiều hơn một loại chuỗi polypeptit. Đến nay, chúng ta đã biết nhiều gen có thể mã hoá cho hai hay nhiều chuỗi polipeptit khác nhua tuỳ thuộc vào việc những đoạn nào được chọn là exon trong quá trình hoàn thiện mRNA, quá trình này được gọi là sự cắt nối RNA thay thế. Nhờ cơ chế này, số loại sản phẩm protein có thể tạo ra có thể lớn hơn rất nhiều so với lượng gen mà cơ thể đó có. Các protein thường có cấu tạo dạng modun gồm nhiều vùng cấu trúc và chức năng riêng biệt, được gọi là các miền. Chẳng hạn như, một miền của một enzyme có bản chất protein chứa vị trí xúc tác, trong khi một miền khác của nó làm nhiệm vụ liên kết protein với màng tế bào. Trong một số các trường hợp, các exon khác nhau mã hoá cho các miền khác nhau của cùng một protein. Các intron trong gen có thể thúc đẩy sự tiến hoá nhanh của các protein có tiềm năng mới nhờ quá trình được gọi là sự xáo trộn exon. Các intron làm tăng xác suất trao đổi chéo giữa các exon thuộc các gen alen với nhau - đơn giản bởi vì cũng cung cấp thêm “nền” cho sự trao đổi chéo mà không làm gián đoạn các trình tự Đột biến điểm có thể ảnh hưởng đến cấu trúc và chức năng của protein. Đột biến sẽ gây ra những biến đổi trong thông tin di truyền, gây nên các ảnh hưởng về kiểu hình, cũng chính là nguyên nhân dẫn đến sự đa dạng, phong phú của các gen ở mọi loài sinh vật. Đột biến chính là nguồn tạo ra mọi gen mới. Chúng ta xem xét đến các
đột biến điểm, tức là những thay đổi hoá học xảy ra ở một cặp base nucleotide duy
nhất của gen. Nếu đột biến điểm xuất hiện trong các tế bào giao tử hoặc các tế bào phát sinh giao tử thì nó có thể được truyền cho thế hệ con và các thế hệ con cháu sau này. Các kiểu đột biến điểm bao gồm hai nhóm lớn: thay thế cặp base và mất hoặc thêm cặp base. Các đột biến thay thế cặp base là những đột biến mà ở đó một cặp base này được thay thế bằng một cặp nucleotit khác. Một số dạng thay thế được gọi là các đột biến yên lặng, bởi vì do tính thoái hoá của mã di truyền, chúng không gây ảnh hưởng đến kiểu hình và biểu hiện chức năng của protein do gen mã hoá. Nói cách khác, sự thay đổi một cặp base có thể chuyển bộ ba mã hoá này thành bộ ba mã hoá khác, nhưng cả hai bộ ba đều mã hoá chung một axit amin. Các đột biến thay thế cũng có thể dẫn đến các đột biến sai nghĩa. Trường hợp như vậy có thể chỉ gây ảnh hưởng ít đến protein, nếu như amino acid mới có các thuộc tính giống với amino acid mà nó thay thế, hoặc khi nó nằm trong miền cấu trúc ít có tính quyết định đến hoạt động chức năng của protein. Những đột biến được quan tâm hơn cả là các đột biến thay thế cặp base làm thay đổi lớn ở protein. Thay đổi một amino acid trong các miền quan trọng của protein - chẳng hạn như trong phần cấu trúc của hemoglobin hoặc ở vị trí trung tâm hoạt động của enzyme - sẽ làm thay đổi hoạt tính protein một cách đáng kể. Trong một số trường hợp, những đột biến như vậy có thể dẫn đến sự tăng cường hoạt tính hoặc tăng thêm khả năng hoạt động của protein; nhung trong phần lớn các trường hợp, chúng có tác động gây hại, thường làm giảm hoặc mất hoạt tính của protein dẫn đến những sai hỏng trong biểu hiện chức năng của tế bào. Các đột biến thay thế thường là các đột biến sai nghĩa, nghĩa là bộ ba mã hoá bị thay đổi vẫn mã hoá cho một amino axit và vì vậy nó vẫn có nghĩa, nhưng nghĩa của nó “không còn đúng” nữa. Nhưng một đột biến điểm cũng có thể làm thay đổi một bộ ba mã hoá amino acid thành một bộ ba kết thúc dịch mã. Trường hợp này, được gọi là đột biến vô nghĩa, và nó dẫn đến sự kết thúc dịch mã sớm, chuỗi polipeptit được tạo ra thường ngắn hơn chuỗi polipeptit do gen bình thường mã hoá. Hầu hết các đột biến vô nghĩa đều dẫn đến các protein mất chức năng. Các đột biến thêm và mất nucleotit là sự bổ sung thêm vào hoặc mất đi một cặp nucleotit trong gen. Những đột biến này thường gây ảnh hưởng lớn hơn nhiều đến sản phẩm protein do gen mã hoá so với các đột biến thay thế nucleotit. Thêm và mất các nucleotit có thể làm thay đổi khung đọc của một thông điệp di truyền, do các bộ ba mã hoá bị sắp xếp lại trong quá trình dịch mã. Những đột biến như vậy được gọi là đột biến dịch khung, xuất hiện bất cứ khi nào số nucleotit được thêm vào hay mất đi không phải là bội số của ba. Tất cả các nucleotit nằm xuôi dòng sau vị trí đột biến đều bị xếp vào các nhóm bộ ba mã hoá không đúng, dẫn đến kết quả là gen được dịch mã sai nghĩa trầm trọng, ngoài ra, nó thường được kết thúc dịch mã sớm hơn hoặc muộn hơn dạng đột biên vô nghĩ
a. Trừ trường hợp, khung đọc bị thay đổi xuất hiện ở rất gần đầu cuối của gen, còn trong phần lớn các trường hợp đột biến dịch khung, protein tạo ra mất chức năng.
Hướng dẫn trả lời
a. Đột biến thay thế nucleotit này bằng nucleotit khác xảy ra trong vùng intron của gen nhưng lại làm cho phân tử mRNA trưởng thành tổng hợp từ gen đột biến ngắn
hơn phân tử mRNA trưởng thành của gen bình thường 173 nucleotit, chứng tỏ đột biến trên đã xảy ra ở vùng nhận biết và cắt intron làm cho quá trình cắt intron bị biến đổi.
Thay vì chỉ cắt các trình tự intron thì tế bào đã cắt một trình tự gồm 1 đoạn exon xen kẽ 2 đoạn intron và đoạn exon này dài 173 nucleotit nên phân tử mRNA trưởng thành của gen đột biến kém phân từ mRNA trưởng thành của gen bình thường 173 nucleotit.
b. Đoạn exon bị mất dài 173 nucleotit. Đây là một số không chia hết cho 3, có nghĩa là exon này có nucleotit kết hợp với nucleotit của exon khác để tạo thành một bộ ba hoàn chỉnh.
Khi đoạn exon này bị mất sẽ gây ra hiện tượng dịch khung đọc đối với toàn bộ trình tự nucleotit phía sau exon này, nên phân tử protein tổng hợp từ gen đột biến sẽ mất đoạn axit amin do 173 nucleotit mã hóa đồng thời bị thay đồi toàn bộ trình tự axit amin ở đoạn peptide phía sau, làm cho protein bị mất chức năng (bất hoạt).
c. Sản phẩm protein tạo ra vẫn bị bất hoạt vì plasmid tái tổ hợp tạo ra từ gen mã hóa proiein X từ người bình thường mang cả đoạn intron. Đoạn trình tự này vẫn được phiên mã và giải mã bình thường trong cơ thể vi khuẩn. Vì thế, phân tử protein tạo ra có trình tự dài hơn hoặc ngắn hơn trình tự axit amin của protein X. Mặt khác, trình tự axit amin của sản phẩm protein tạo ra sẽ khác vài trình tự các axit amin của protein X.
Câu 12 (Trích đề HSG 2016 ngày 2)
a. Một bệnh nhân nhi là con đẻ cùa một cặp bố mẹ hôn nhân cận huyết mắc bệnh chậm phải triển không rõ nguyên nhân gây bệnh. Xét nghiệm hóa sinh cho thấy bệnh nhân này bị thiểu năng do bất hoạt 4 enzyme ở lizoxom. Hãy nêu các cơ chế để một đột biến lặn trên nhiễm sắc thể thường duy nhất có thể làm giảm hoạt tính hoặc giảm chức năng của 4 enzyme trên.
b. Tại sao các đột biến gen thường là đột biến lặn và nằm trên nhiễm sắc thể thường.
Hướng dẫn vận dụng kiến thức:
Hoá sinh: Cấu trúc và hoạt tính của enzyme: Enzyme là một đại phân tử hoạt động như một chất xúc tác, một tác nhân hoá học làm thay đổi tốc độ phản ứng nhưng không bị phản ứng tiêu thụ. Các enzyme chủ yếu có bản chất là protein. Khi thiếu enzyme thì các phản ứng hoá học trong các con đường chuyển hoá sẽ trở nên bị đình trệ nghiêm trọng do nhiều phản ứng hoá học sẽ tốn nhiều thời gian. Mỗi enzyme có cấu trúc không gian đặc thù, đặc biệt là vùng được gọi là trung tâm hoạt động. Vùng này được cấu tạo bởi một số các axit amin đặc thù làm cho trung tâm có cấu hình không gian đặc thù phù hợp với cơ chất mà enzyme tác động. Hình thù không gian của trung tâm hoạt động có thể bị thay đổi, ví dụ do đột biến sẽ làm thay đổi chức năng của enzyme. Nhiều enzyme, ngoài trung tâm hoạt động còn có thêm phần phụ được gọi là trung tâm điều chỉnh có tác dụng điều chỉnh hình thù của trung tâm hoạt động.
Đối với một số enzyme, ngoài thành phần protein còn có một số chất khác tham gia (được gọi là cofactor ví dụ như một số ion kim loại như sắt, đồng, mangan, kẽm, niken, hoặc các chất hữu cơ như vitamin được gọi là coenzyme...). Cofactor thường liên kết cố định hoặc tạm thời với trung tâm hoạt động của enzyme vầ rất cần thiết cho hoạt động xúc tác. Cấu hình không gian của enzyme có liên quan chặt chẽ đến chức năng của nó. Các rối loạn hoá học sẽ là hậu quả tất yếu nếu tất cả các con đường chuyển hoá của tế bào hoạt động cùng lúc. Bản chất nội tại của quá trình sống là khả năng của tế bào có thể điều hoà một cách chặt chẽ các con đường chuyển hoá thông qua quá trình điều tiết các enzyme khác nhau hoạt động khi nào và ở đâu. Để làm được điều đó, tế bào hoặc đóng mở các gen mã hoá các protein đặc hiệu thông qua điều hoà hoạt động của các enzyme ngay khi chúng được tạo ra.
+ Lizoxom là một bào quan ở tế bào động vật (trừ tế bào hồng cầu), có dạng túi hoặc bóng được giới hạn bởi màng lipoprotein và chứa các enzyme thuỷ phân (hydraza). Lizoxom gồm hai dạng: Lizoxom cấp 1 và lizoxom cấp 2. Lizoxom cấp 1 là lizoxom mới được tạo thành chưa tham gia vào hoạt động phân giải. Lizoxom cấp 2 là lizoxom đang tham gia hoạt động phân giải, bao gồm hai loại: hêtrolizoxom xuất hiện do kết quả kết hợp của lizoxom cấp 1 với phagoxom. Otolizoxom được tạo thành do sự kết hợp của lizoxom cấp 1 với otophagoxom.
Những bệnh liên quan đến lizoxom thường xảy ra do sự bất bình thường trong cấu trúc màng hoặc hệ enzyme của lizoxom. Sự phá huỷ màng lizoxom sẽ làm cho các enzyme trong lizoxom giải phóng gây tiêu huỷ tế bào hoặc giải phóng vi khuẩn vào các tế bào khác gây ra bệnh nhiễm trùng. Màng lizoxom cũng có thể bị sai lệch di truyền dẫn đến biến đổi tính thấm. Sự sai lệch trong hệ enzyme của lizoxom như thiếu một hoặc nhiều loại enzyme, nguyên nhân có thể do di truyền như thiếu hoặc sai lệch gen chịu trách nhiệm tổng hợp enzyme. Sự tích luỹ nhiều chất trong lizoxom do thiếu enzyme phân giải chúng, do nguyên nhân di truyền còn dẫn tới nhiều bệnh khác liên quan đến hệ thần kinh, tim mạch, thận, ...
+ Di truyền: đột biến gen
Hướng dẫn trả lời
a.
- 4 enzyme này cùng dùng chung một co-enzyme và việc tổng hợp hoặc vận chuyển coenzyme này bị ảnh hưởng do đột biến gen mã hóa enzyme tổng hợp hoặc protein vận chuyển
- 4 enzyme này cùng được cấu tạo từ một tiểu phần (chuỗi polypeptit tiền thân) chung. Đột biến gen tổng hợp chuỗi polypeptit tiền thân đã gây ra sự sai khác của 4 enzyme được hình thành từ đoạn polypeptit này.
- 4 enzyme này đều được hoàn thiện (có dạng tiền thân không hoạt tính thành dạng có hoạt tính) bởi một enzyme chung-> đột biến gen này sẽ làm ảnh hưởng tới quá trình hoàn thiện 4 enzyme trên.
- Bình thường cả 4 enzyme đều định vị tại một bào quan (lizoxom) nhưng sai hỏng trong quá trình hoạt động của bào quan đã ảnh hưởng đồng thời hoạt tính của 4 enzyme (ví dụ chúng đều không được vận chuyển vào trong bào quan và bị phân hủy trong tế bào chất)
b.
- Lý do phần lớn đột biến liên quan đến enzyme thiết yếu là đột biến mất chức năng và đều có tính lặn (ngược lại đột biến thêm chức năng là tính trội). Mặt khác, các enzyme thiết yếu được mã hóa bởi các gen đang hoạt động nên dễ bị đột biến hơn các gen không hoặc chưa hoạt động.
- Phần lớn các gen nằm trên nhiễm sắc thể thường, vì thế đột biến lặn trên nhiễm sắc thể thường là phổ biến nhất trong các đột biến mất chức năng của các enzyme thiết yếu.
Câu 13 (trích đề thi HSGQG 2016 ngày 2)
a. Giải thích vì sao phân tử DNA mạch kép có thể tạo phức hợp với protein histon để hình thành nucleoxom?
b. Những đột biến gen nào dẫn đến các cặp nhiễm sắc thể tương đồng ớ kỳ sau của nguyên phân không phân ly bình thường về hai cực của tế bào? Giải thích.
Hướng dẫn vận dụng kiến thức:
Hoá sinh: cấu trúc DNA
- Cấu trúc mạch đơn phân tử DNA: Mỗi nucleotit đơn phân chứa một base nitrogen (A, T, G hoặc C), đường deoxy ribozo và một nhóm photphat. Nhóm photphat của nucleotit này liên kết với đường của nucleotit tiếp theo tạo nên “cột sống” phân tử gồm các nhóm photphat và đường luân phiên, từ đó có các base nhô ra.
Mạch poly nucleotit có tính định hướng từ đầu 5’ (với nhóm phosphate) tới đầu 3’ (với nhóm -OH). 5’ và 3’ là các số chỉ các nguyên tử carbon nằm trên cấu trúc vòng của phần đường.
- Mô hình cấu trúc không gian: Chuỗi xoắn kép bao gồm 2 mạch đơn DNA xoắn song song ngược chiều nhau và xoắn về phía bên phải “xoắn phải” theo hướng đi lên. Hai mạch chuỗi xoắn được giữ lại với nhau qua các liên kết hidro hình thành giữa các base nito kết thành từng cặp bên trong chuỗi xoắn. Liên kết cộng hoá trị mạnh làm nhiệm vụ nối các nucleotit trên một mạch với nhau, trong khi hai mạch đơn được liên kết với nhau bởi liên kết yếu - liên kết hidro. Các cặp base xếp chồng lên nhau và hình thành giữa chúng là lực hấp dẫn Van de Waals - yếu tố chính giúp giữ toàn bộ phân tử với nhau.
Mỗi NST ở sinh vật nhân thực đều chứa một phân tử DNA xoắn kép mạch thẳng duy nhất được đóng gói cùng với các phân tử protein; ở người, kích thước trung bình vào khoảng 1,5 x 108 cặp nucleotit. Lượng DNA là lớn hơn nhiều so với chiều dài
NST khi co xoắn cực đại. Nếu duỗi thẳng, mỗi phân tử DNA ở hệ gen nhân ngừoi có
chiều dài trung bình hơn 4 cm, tức là dài hơn hàng nghìn lần so với đường kính của nhân tế bào - dó là chưa kể đến 45 NST khác còn lại đồng thời có mặt trong nhân tế bào. Trong tế bào, DNA của sinh vật nhân thực kết hợp chính xác với một lượng lớn protein. Phức hợp DNA với proten được gọi là chất nhiễm sắc, có thể gói gọn vào trong nhân tế bào nhờ hệ thống đóng gói DNA ở nhiều cấp độ khác nhau.
Trong chu kì tế bào, NST phải trải qua những thay đổi về cấp độ đóng gói. Khi tế bào chuẩn bị nguyên phân, chất nhiễm sắc cuộn xoắn (cô đặc), cuối cùng ở kì giữa hình thành nên một số lượng đặc trưng các NST có kích thước ngắn và dày có thể phân biệt được bằng kính hiển vi quang học.
NST là một cấu trúc năng động, có thể cô đặc, dãn xoắn, biến đổi thậm chí thay đổi cấu hình theo yêu cầu của các quá trình khác nhau trong tế bào ví dụ như nguyên phân, giảm phân và quá trình biểu hiện của các gen. Các protein histone không chỉ đơn thuần là các lõi cuộn chỉ để DNA có thể quấn quanh trong cấu trúc chất nhiễm sắc. Sự biến đổi hoá học của các protein có vai trò trực tiếp làm thay đổi mức độ tổ chức của chất nhiễm sắc và tham gia điều hoà sự biểu hiện của các gen. Terry Orr - Weaver cùng với cộng sự của mình đã tiến hành các nghiên cứu ở Drosophila và chỉ ra rằng sự photphoryl hoá của một số amino acid đặc thù trong vùng đuôi histone có vai trò quyết định động thái của NST trong kỳ đầu Giảm phân I. Sự photphoryl hoá các biến đổi hoá học của các histone có nhiều tác động đến sự hoạt động của các gen
Các biến đổi của histone
Vùng đầu N của histone (đuôi histone) trong mỗi nucleosome thường lòi ra ngoài nucleosome. Phần đuôi này có thể được tiếp cận và bị biến đổi bởi một số enzyme đặc biệt, chúng xúc tác cho việc bổ sung hoặc loại bỏ một số gốc hoá học đặc thù nào đó. Trong hiện tượng acetyl hoá histone, gốc (-COCH3) được gắn vào các các amino acid ở phần đuôi histone, trong khi đó hiện tượng khử acetyl là hiện tượng loại bỏ những gốc acetyl này. Khi lysine được acetyl hoá, điện tích dương của nó được trung hoà, làm cho đuôi histone không còn liên kết chặt với các nucleosome ở gần nữa (chính sự liên két chặt của đuôi histone vào nucleosome thúc đẩy sự cuộn xoắn của chất nhiễm sắc thành dạng cấu trúc kết đặc hơn).
Một số gốc hoá học khác cũng có thể được gắn thêm vào hoặc loại bỏ khỏi các amino acid thuộc đuôi histone, như các gốc metyl và các gốc photphat. Việc bổ sung gốc methyl (-CH3) vào đuôi histone (methyl hoá) có thể thúc đẩy sự kết đặc hơn của các chất nhiễm sắc. Trong khi đó, việc bổ sung một gốc photphat vào một amino acid (phosphoryl hoá) gần amino acid bị methyl hoá sẽ gây ra hiệu ứng ngược lại.
Di truyền:
Diễn biến kỳ sau Nguyên phân: đây là kì ngắn nhất của nguyên phân, chỉ diễn ra trong vài phút. Kỳ sau được bắt đầu khi các protein cohensin tách nhau ra. Sự kiện đó cho phép các NST chị em của mỗi cặp đột ngột tách nhau ra. Mỗi nhiễm sắc tử trở thành một NST đầy đủ, hai nhiễm thể con đã tách nhau ra đều di chuyển về hai cực đối
100
lập của tế bào khi các vi ống thể động ngắn lại. Do các vi ống này gắn vào vùng tâm động, vùng tâm động sẽ di chuyển trước tiên. Tế bào dài ra khi các vi ống không thể động dài ra. Vào cuối kì sau, hai cực của tế bào có hai bộ NST hoàn chỉnh và tương đương nhau.
Hướng dẫn trả lời:
a. DNA liên kết với protein histon để hình thành nucleoxom:
- Gốc photphat phân bố dọc khung phân tử DNA làm cho phần ngoài phân tử tích điện âm suốt dọc chiều dài phân tử, tạo thuận lợi cho sự hình thành liên kết với các protein histon.
- Các axit amin tích điện dương như lizin hoặc arginin, chiếm hơn 1/5 tổng số các axit amin có trong protein histon giúp hình thành liên kết với gốc photphat trên phân tử DNA.
- Có 14 điểm tương tác khác nhau giữa DNA với protein histon lõi. Ở mỗi tiếp điểm, khe phụ của DNA ở vị trí trực diện với lõi 8 phân tử histon (octamer) có khả năng hình thành gần 140 liên kết hidro với nhau.
b.
Các đột biến làm các NST trong cặp tương đồng không phân ly bình thường về 2 cực tế bào trong kỳ sau của phân bào nguyên phân:
- Đột biến gen mã hóa các protein thể động (kinetochore) gắn kết tâm động của nhiễm sắc thể với thoi phân bào làm cho thoi phân bào không gắn được với nhiễm sắc thể
- Đột biến gen mã hóa protein động cơ dịch chuyển các nhiễm sắc thể dọc theo thoi phân bào về hai cực của tế bào.
- Đột biến gen mã hóa protein cohensin gắn kết giữa hai nhiễm sắc tử của nhiễm sắc thể để hình thành nhiễm sắc thể kép, làm thay đổi cấu trúc, từ đó enzyme phân giải protein cohensin ở đầu kỳ sau không nhận diện được hoặc đột biến gen mã hóa enzyme phân giải protein cohensin làm cho các nhiễm sắc tử không tách ra khỏi nhiễm sắc thể kép đi về 2 cực của tế bào.
Câu 14 (Trích đề HSGQG 2016 ngày 2)
Đa số vi khuẩn không chuyển hóa được xylytol để tạo năng lượng. Người ta tiến hành thí nghiệm chuyển một gen mã hóa enzyme phân giải xylitol vào E. Coli.
Một dòng plasmid làm thể truyền được biển đổi để có một gen kháng ampicilin (ampR) và một gen mã hóa β galactozidaza có chứa trình tự nhận biết của enzyme cắt giới hạn EcoRI (β galactozidaza có khả năng phân giải X- gal từ không màu thành sản phẩm màu xanh dương).
Sau khi đoạn DNA chứa gen mã hóa enzyme phân giải xylitol và thể truvền được xử lý riêng rẽ với EcoRI, chúng được trộn với nhau để tái tổ hợp ngẫu nhiên.
Hỗn hợp DNA này lại tiếp tục được cho biến nạp vào một chủng E. coli đã bị đột biến bất hoạt gen Z trong operon Lac. Sau đó đem cấy trải trên môi trường thạch tối thiểu thu được 3 dòng A, B và C. Tiếp tục nuôi cấy riêng rẽ 3 dòng A, B, C trên môi trường thạch tối thiểu có bổ sung thêm ampiclin và X-gal thu được kết quả như
sau:
Dòng A: Xuất hiện khuẩn lạc và không thay đổi màu
Dòng B: Xuất hiện khuẩn lạc và vùng xung quanh khuẩn lạc chuyển sang màu xanh dương
Dòng C: Không xuất hiện khuẩn lạc.
a. Giải thích kết quả thí nghiệm.
b. Nếu cả 3 dòng A, B, C được nuôi cấy riêng rẽ trong môi trường chỉ chứa xylitol làm nguồn cacbon duy nhất thì khả năng phát triển của mỗi dòng sẽ như thế nào? Giải thích?
Hướng đẫn vận dụng kiến thức
Kĩ thuật di truyền: Kĩ thuật nhân dòng gen bằng plasmid vi khuẩn:
Các phân tử plasmid gốc được gọi là các vector nhân dòng, chúng được định nghĩa là các phân tử DNA có thể mang DNA ngoại lai vào tế bào chủ và sao chép ở đó. Các plasmid vi khuẩn được dùng làm vector nhân dòng vì một số nguyên nhân. Chúng có thể dễ dàng được phân lập từ các tế bào vì khuẩn, được thao tác để tạo nên các plasmid tái tổ hợp bằng việc cài đoạn DNA ngoại lai trong điều kiện invitro, sau đó được đưa trở lại các tế bào vi khuẩn. Hơn nữa các plasmid vi khuẩn tái tổ hợp (và đoạn DNA ngoại lai mà chúng mang) có thể nhân lên nhanh chóng nhờ tốc độ sinh sản cao của các tế bào chủ Ứng dụng nhân dòng gen là quá trình tạo ra nhiều bản sao của một gen được quan tâm. Những bản sao này có thể được dùng trong giải trình tự gen, sản xuất các prtein do gen mã hoá hoặc trong các nghiên cứu cơ bản và ứng dụng khác. Ví dụ, các gen mã háo enzyme phân giải xylitol được cài vào plasmid có nguồn gốc từ E. coli. Kĩ thuật được tiến hành như sau:
1 - Phân lập DNA plasmid từ tế bào vi khuẩn và DNA mã hoá protein phân giải xylitol. Dòng plasmid làm thể truyền được biển đổi để có một gen kháng ampicilin (ampR) và một gen mã hóa β galactozidaza có chứa trình tự nhận biết của enzyme cắt giới hạn EcoRI (β galactozidaza có khả năng phân giải X- gal từ không màu thành sản phẩm màu xanh dương).
2 - Cắt cả hai mẫu DNA bởi cùng một enzyme giới hạn (trong trường hợp này là EcoRI).
3 - Trộn các plasmid đã được mở vòng với các phân đoạn DNA từ gen mã hoá enzym phân giải xylitol. Một số đoạn nối vứoi nhau bởi sự kết cặp giữa các base, DNA ligase nối chúng với nhau. Sản phẩm là các plasmid tái tổ hợp và nhiều plasmid không tái tổ hợp
4 - Trộn DNA với các tế bào vi khuẩn có đột biến ở gen β galactozidaza của riêng nó. Một số tế bào tiếp nhận plasmid tái tổ hợp hoặc phân tử DNA khác qua biến nạp.
5 - Cấy rải vi khuẩn trên môi trường agar chứa ampicillin và X-gal. Nuôi vi khuẩn cho đến khi khuẩn lạc hình thành.
Kết quả: Chỉ các tế bào tiếp nhận plasmid mang gen ampR mới có thể sinh sản và hình thành khuẩn lạc. Các khuẩn lạc mang plasmid không tái tổ hợp sẽ có màu xanh, các khuẩn lạc mang plasmid tái tổ hợp có gen β galactozidaza bị phá vỡ sẽ có màu trắng vì chúng không phân huỷ được X-gal.

Hướng dẫn trả lời:
a. Giải thích kết quả thí nghiệm:
- Dòng A phát triển được trên môi trường có ampicilin => có plasmid chứa ampR. Khi nuôi trên X-gal thì màu của môi trường không đổi => gen β galactozidaza
không được biểu hiện => có hiện tượng biến nạp plasmid tái tổ hợp.
- Dòng B phát triển được trên môi trường có ampicilin và chuyển hóa được Xgal làm môi trường chuyển sang màu xanh => có hiện tượng biến nạp plasmid không tái tổ hợp.
- Dòng C không phát triển được trên môi trường có ampicilin => không có hiện tượng biến nạp.
b. Khả năng phát triển của 3 dòng khi nuôi cấy trên môi trường có xylitol làm nguồn cacbon duy nhất:
- Dòng A có biến nạp plasmid tái tổ hợp tuy nhiên không thể chắc chắn plasmid tái tổ hợp này có gen chuyển hóa xylitol. Do đó:
+ Dòng A có thể phát triển được trên môi trường có xylitol nếu plasmid tái tổ hợp có chứa gen chuyển hóa xylitol hoàn chỉnh.
+ Dòng A sẽ không phát triển được trên môi trường chỉ chứa xylitol nếu plasmid tái tổ hợp không chứa gen chuyển hoá xylitol hoàn chỉnh.
- Dòng B có khả năng sử dụng lactozo (do có gen β galactozidaza trên plasmid hoạt động) nhưng không có khả năng sử dụng xylitol nên không phát triển được trên môi trường chi chứa xilitol.
- Dòng C không có biến nạp => không chuyển hóa được xylitol => không phát triển được trên môi trường có chửa xylitol.
Câu 15 (Trích đề thi IBO năm 2015)
Ở nấm mốc, sự sinh tổng hợp axit amin arginine cần bước chuyển hóa tiền chất thành ornithine; chất này được chuyển hóa tiếp thành citruline là tiền chất trực
tiếp được chuyển hóa thành arginine. Mỗi phản ứng trong 3 bước này được xúc tác bởi một enzym riêng rẽ; mỗi enzym được mã hóa bởi một gen (xem hình)
A: Con đường sinh tổng hợp arginine ở nấm mốc. B: Thiết kế thí nghiệm để xác định chuyển hóa: dải màu nâu đen chỉ Nấm mốc sinh trưởng. Chủng kiểu dại mọc trên môi trường tối thiểu, trong khi các chủng đột biến chỉ mọc trên các môi trường được bổ sung (+) chất dinh dưỡng đặc thù.
Precursor: tiền chất
Ornithine: Ornithine
Citrulline: Citrulline
Arginine: Arginine
Enzyme: enzyme
Gene: Gen
Wild type: Kiểu dại (WT)
Class I mutants: Các đột biến lớp I
Class II mutants: Các đột biến lớp II
Class III mutants: Các đột biến lớp III
+ Ornithine: + Ornithine:
+ Citrulline: + Citrulline:
+ Arginine: + Arginine
Mỗi phát biểu dưới đây là đúng hay sai?
a, Các chủng đột biến lớp III có thể mang đột biến ở nhiều hơn 1 gen liên quan đến con đường sinh hóa này.
b, Các chủng đột biến lớp I thiếu cả 3 enzym.
c, Các thể đột biến thiếu hai enzym A và C có thể mọc nếu môi trường được bổ sung đồng thời Orthnithine và Citruline - (F, thiếu C thì ko mọc được trên môi trường có citruline)
d, Giả sử một chất ức chế enzym B được bổ sung vào môi trường nuôi chủng kiểu dại, gen B sẽ bị bất hoạt.
(Bổ sung chất sau đột biến => chủng vi khuẩn sống được, bổ sung chất trước đột biến, không chuyển hoá được => vi khuẩn không sống được)
Thứ tự đột biến: O -> C -> A
Hướng dẫn vận dụng kiến thức:
Mối quan hệ giữa gen và enzyme: mối quan hệ này được làm sáng tỏ bởi
Beadle và Edward Tatum nghiên cứu ở nấm mốc Neurospora crassa. Trên cơ sở các phương pháp gây tạo đột bién được tìm ra từ những năm 1920, các nhà khoa học đã dùng tia X “bắn phá” các chúng Neurospora để tạo nên các chủng đột biến có nhu cầu dinh dưỡng khác nhau so với kiểu dại. Các chủng nấm mốc Neurospora kiểu dại có nhu cầu dinh dưỡng đơn giản. Chúng có thể dễ dàng sống trên môi trường thạch (agar) được bổ sung một số muối vô cơ, đường glucose và vitamin biotin. Từ môi trường tối thiểu này, các tế bào nấm men có thể dùng các con đường chuyển hoá của chúng để tạo nên tất cả các phân tử cần thiết cho sự sinh trưởng và phát triển của mình. Beadle và Tatum đã xác định được rằng nhiều chủng đột biến không có khả năng sống trên môi trường tối thiểu do mất khả năng tổng hợp một chất thiết yếu nào đó. Để có thể nuôi các chúng đột biến dinh dưỡng này, Beadle và Tatum phải nuôi chúng trong môi trường đủ, gồm các thành phần của môi trường tối thiểu, được bổ sung thêm 20 loại axit amin và một số chất dinh dưỡng khác. Trong môi trường đủ, mọi thể đột biến đều có khả năng sống dù chúng không có khả năng tổng hợp một chất nào đó.
Để phân tích đặc điểm của các dạng sai hỏng chuyển hoá ở các chủng đột biến dinh dưỡng (thể đột biến khuyết dưỡng) Beadle và Tatum đã tiến hành lấy mẫu từ quần thể tế bào đột biến được nuôi trong môi trường đủ, rồi cho các thể đột biến vào các ống nghiệm khác nhau. Trong mỗi ống nghiệm, ngoài môi trường tối thiểu, họ cho thêm một chất dinh dưỡng nhất định. Chất dinh dưỡng bổ sung đặc thù này cho phép nấm mốc đột biến có thể sinh trưởng được chứng tỏ sai hỏng chuyển hoá có liên quan đến chất này. Ví dụ, nếu chủng đột biến được tìm thấy có khả năng phát triển trong môi trường bổ sung amino acid arginine, thì các nghiên cứu kết luận rằng thể đột biến đó bị sai hỏng trong con đường chuyển hoá tổng hợp arginine.
Beadle và Tatum sau đó tiếp tục xác định tính đặc thù của mỗi thể đột biến
Thí nghiệm kiểm tra liệu các gen riêng rẽ quy định các enzyme xúc tác cho các phản ứng trong một con đường hoá sinh? Thí nghiệm được nghiên cứu ở Neurospora crassa, Geogre Beadle và Edward Tatum tại đại học Stanford đã phân lập được các thể đột biến cần bổ sung arginne vào môi trường sinh trưởng của chúng. Các nhà nghiên cứu thấy rằng các đột biến này chia thành 3 nhóm, mỗi nhóm sai hỏng một gen khác nhau. Cân nhắc trên các dữ liệu thí nghiệm, họ dự đoán con đường sinh tổng hợp arginine liên quan đến một tiền chất trong môi trường sinh dưỡng và các phân tử trung gian là ornithine và citrulline. Thí nghiệm nổi tiếng của họ được minh hoạ chứng minh cho giải thiết một gen - một enzyme đồng thời xác nhận con đường tổng hợp arginine. Thí nghiệm được tiến hành bằng cách nuôi ba nhóm thể đột biến nấm mốc trong 4 điều kiện môi trường khác nhau như hình minh hoạ. Trong thí nghiệm, họ đã sử dụng môi trường tối thiểu (MM) làm đối chứng do môi trường này các tế bào kiểu dại có thể sinh trưởng, trong khi các tế bào đột biến thì không.
Kết quả thu được: Chúng kiểu dại có khả năng sinh trưởng trong tất cả các điều kiện thí nghiệm khác nhau, chỉ đòi hỏi môi trường tối thiểu. Trong khi đó, ba nhóm thể đột biến đều cần bổ sung những chất dinh dưỡng đặc thù cho mỗi nhóm. Ví dụ các thể đột biến nhóm II không sinh trưởng được trong môi trường chỉ bổ sung ornithine, mà sinh trưởng trong các môi trường bổ sung citrulline hay arginine.
Kết luận: Từ những yêu cầu về nguồn dinh dưỡng của các thể đột biến, Beadle và Tatum đã suy luận ra rằng mỗi nhóm thể đột biến không thể thực hiện được một bước trong con đường sinh tổng hợp arginine, mà theo giả thiết là do chúng thiếu những enzyme đặc thù. Do mỗi nhóm thể đột biến bị đột biến ở một gen duy nhất, họ kết luận rằng mỗi gen bình thường quy định việc tế bào sản xuất một enzyme. Kết quả nghiên cứu ủng hộ cho giả thiết một gen - một enzyme và đồng thời cũng xác nhận con đường chuyển hoá tổng hợp arginine (chú ý trong phần kết quả là các thể đột biến chỉ sinh trưởng được trong các môi trường bổ sung một hợp chất hình thành sau bước sai hỏng của quá trình chuyển hoá, vì điều này mới có thể giúp khắc phục sai hỏng).
Câu 16 (Trích đề thi HSGQG 2017 - ngày 1)
Nấm men kiểu dại có khả năng phân giải glucose thành etanol và khí cacbonic trong điều kiện thiếu oxi. Khi xử lý đột biến, người ta thu được chủng nấm men mang đột biến suy giảm hô hấp do thiếu xitocrom oxidaza - một thành phần của chuỗi vận chuyển điện tử. Việc sử dụng chủng nấm men này có ưu thế gì so với chủng kiểu dại trong công nghệ lên men rượu? Giải thích?
Ở nấm men mất khả năng lên men, đường phân có thể diễn ra trong điều kiện thiếu oxi không? Tại sao?
Sau đây là hai phản ứng thuộc quá trình đường phân:
Glyxeraldehit-3-photphat + NAD+ + Pi 1,3-Bisphotphoglixerat + NADH 1,3 Bisphotphoglixerat + ADP 3-Photphoglixerat + ATP
Photphat vô cơ (Pi) có vai trò thiết yếu trong quá trình lên men. Khi nguồn cung
cấp Pi cạn kiệt, sự lên men bị dừng lại kể cả khi môi trường có glucose. Asenat (AsO4 3-) tương đồng với photphat (PO4 3-) về cấu trúc hóa học và có thể làm cơ chất thay thế photphat. Este asenat không bền nên dễ thủy phân ngay khi vừa hình thành.
Giải thích tại sao asenat gây độc đối với tế bào?
Hướng dẫn vận dụng kiến thức:
Hoá sinh: quá trình đường phân phân giải glucose
Vi sinh: Dị hoá ở nấm men
Di truyền: biến dị ở vi sinh vật nhân thực
Hướng dẫn trả lời:
Việc sử dụng chủng nấm men đột biến có ưu thế trong việc đơn giản hóa điều kiện lên men vì không cần phải duy trì điều kiện kị khí như đối với nấm men kiểu dại
- Nấm men là vi sinh vật kị khí không bắt buộc. Trong điều kiện thiếu O2, nấm men sẽ lên men rượu. Trong điều kiện có O2, nấm men sẽ tiến hành hô hấp hiếu khí. Do đó, phải duy trì điều kiện kị khí để tiến hành lên men. Trong công nghệ lên men rượu, việc duy trì điều kiện kị khí đòi hỏi chi phí thục hiện.
- Xitocrom oxidaza là một thành phần của chuỗi vận chuyển điện tử. Nếu thiếu enzyme này chuỗi vận chuyển điện tử bị ngừng trệ. Chu trình Crep cũng bị ngừng do thiếu NAD+ từ chuỗi vận chuyển điện tử. Do đó, nấm men sẽ chuyển sang lên men rượu ngay cả khi có O2
b) Quá trình đường phân sẽ không diễn ra ở nấm men mất khả năng lên men trong điều kiện thiếu O2. Vì quá trình đường phân tạo nên một lượng lớn axit pyruvic và NADH làm giảm tỉ lệ [NADH]/[NAD+]. Trong điều kiện thiếu O2, chuỗi vận chuyển điện tử bị ức chế nên không tạo ra NAD+. Nếu không có quá trình lên men, lượng NAD+ không được tái tạo dẫn đến quá trình đường phân không thể diễn ra.
c) Khi có asenat, 1-asenat-3-photphoglixerat (este asenat) được hình thành thay cho bisphotphoglixerat. Khi đó, este asenat bị thủy phân thành 3-photphoglixerat.
(1-asenat-3-photphoglixerat + H2O 3-photphoglixerat + AsO4 3-)
Phân tử 3-photphoglixerat vẫn được tạo thành như trong quá trình đường phân nhưng không kèm theo sự tổng hợp ATP dẫn đến sự giảm năng lượng tạo thành trong các phản ứng tương tự. Vì vậy, asenat độc đối với tế bào.
Câu 17 (Trích đề HSG 2017 ngày 1)
Virut cúm A/H5N1 lây truyền bệnh ở người và gia cầm, còn virut cúm A/H3N2 chỉ lây truyền bệnh ở người. Giả sử, người ta tạo được virut lai bằng cách tách hệ gen
(RNA) của virut cúm A/H5N1 ra khỏi vỏ capsit của nó, rồi chuyển vào đó hệ gen (RNA) của virut A/H3N2
Trình bày giai đoạn sinh tổng hợp (nhân lên) của virut lai vừa tạo ra (thế hệ 0) sau khi xâm nhập vào tế bào người. Biết rằng virut cúm A có hệ gen RNA (-) và phiên mã tổng hợp mRNA từ khuôn RNA hệ gen của nó.
Virut lai thế hệ 1 có khả năng lây truyền bệnh ở gia cầm không? Giải thích
Nếu như gen mã hóa gai glycoprotein H (hemagglutinin) bị đột biến ở chủng gốc A/H5N1 thì phần lớn virut lai vừa tạo ra (thế hệ 0) sẽ thay đổi khả năng lây nhiễm ở người như thế nào? Giải thích?
Hướng dẫn vận dụng kiến thức:
Cấu trúc virut: Virut không có cấu trúc tế bào, cấu tạo đơn giản chỉ gồm vật chất di truyền (DNA hoặc RNA: sợi đơn hoặc sợi kép) được bao bọc bởi một vỏ protein. Trong một số trường hợp có thêm vỏ bọc bên ngoài ...
Vật chất di truyền ở virut RNA: RNA là vật chất di truyền ở một số virut. Mặc dù phage và phần lớn virut thực vật là các virut RNA, nhưng dạng biến thể đa dạng nhất của hệ gen RNA được tìm thấy ở các virut lây nhiễm động vật. Trong ba loại hệ gen RNA mạch đơn được tìm thấy ở các virut động vật, các hệ gen là RNA mạch đơn (ssRNA-RNA+) có thể dùng trực tiếp như mRNA, tức là nó có thể được dùng trực tiếp để dịch mã ngay sau khi lây nhiễm. Một số virut khác hệ gen cũng là phân tử RNA mạch đơn (ssARRN- RNA-) nhưng hệ gen đó RNA của nó được dùng làm khuôn để tổng hợp nên mRNA. Hệ gen này được dùng để phiên mã thành mạch RNA bổ sung; mạch RNA bổ sung này vừa được dùng như mRNA, vừa được dùng làm khuôn để tổng hợp các bản sao RNA mới của virut. Tất cả các virut cần quá trình tổng hợp RNA -> RNA để tạo nên mRNA đều được sử dụng một enzyme của virut để thực hiện quá trình này; không có những enzyme như vậy trong phần lớn các tế bào chủ. Các enzyme của virut được đóng gói cùng với hệ gen bên trong lớp vỏ capsid.
Các virut RNA ở động vật có chu kỳ sinh sản phức tạp nhất là các retrovirus. Những virus này được trang bị một enzyme có tên là enzyme phiên mã ngược (hay reverse trancriptase); enzyme này có thể phiên mã mạch khuôn RNA thành DNA, tạo nên dòng thông tin RNA, ngược chiều với thông thường.
Hướng dẫn trả lời:
Virut cúm sử dụng RNA polymerase của nó và nguyên liệu của tế bào chủ để (phiên mã) tổng hợp mRNA (RNA +) trên khuôn RNA của nó (RNA -).
Các mRNA (RNA +) mớiđược tổng hợp được dùng làm khuôn để tổng hợp các RNA hệ gen mới (RNA -) của virut, đồng thời được dùng làm khuôn để tổng hợp (dịch mã) protein vỏ capsit và vỏ ngoài để lắp ráp thành virut mới.
Virut lai thế hệ 1 không lây truyền bệnh ở gia cầm
Vì, hệ gen của virut lai thế hệ 0 là từ virut cúm A/H3N2 nên sẽ tạo ra thế hệ 1 là
A/H3N2 không lây truyền bệnh ở gia cầm (trừ trường hợp đột biến xảy ra ngay trong lần tái sinh virut thế hệ 0).
Nếu gen mã hóa cho gai H bị đột biến thì phần lớn virut lai không lây nhiễm (hoặc giảm) ở người.
Vì virut không có khả năng đính kết không có khả năng đính kết lên tế bào chủ (qua thụ thể) nên không xâm nhập được vào tế bào vật chủ
Câu 18 (Trích đề thi HSGQG 2018 - ngày 1)
Khi một enzyme có mặt ở một loài vi khuẩn, thì con đường chuyển hoá mà enzyme đó tham gia thường tồn tại trong loài vi khuẩn này. Bảng 1.1 là tên enzyme và phản ứng mà enzyme xúc tác được sử dụng làm chỉ thị cho sự xuất hiện của các con đường chuyển hóa mà nó tham gia. Bảng 1.2 thể hiện sự có mặt hay vắng mặt của một số enzyme ở bốn loài vi khuẩn khác nhau 1,2, 3 và 4.
Bảng 1.1. Enzyme và phản ứng xúc tác tương ứng
Tên enzyme Phản ứng xúc tác
Lactat đêhiđrôgenaza (LDH) Axit piruvic + NADH → axit lactic 4NAD+
Alcohol đêhiđrôgenaza (ADH) Axêtanđêhit 4- NADH → Êtanol + NAD+
Xitôcrôm C oxidaza Vận chuyển electron từ xitôcrôm C tới xitôcrôm a
ATP sintetaza Vận chuyển H+ qua màng tạo ATP từ ADP và Pi
Xitrat sintetaza Axit oxalo axetic + Axetyl coA -> axit xitric
Bảng 1.2. Sự có mặt (+) và vắng mặt (-) của mỗi loại enzyme trong từng loài vi khuẩn
Hãy cho biết:
a) Loài vi khuẩn nào không thể thực hiện được hô hấp hiếu khí? Giải thích.
b) Các sản phẩm chính mỗi loài vi khuẩn tạo ra sau quá trình chuyển hóa glucozo?
Loài vi khuẩn
Tên enzyme LD H ADH Xitô crôm c oxidaza
ATP sintetaza Xitrat sintetaza
Loài 1 - + - + -
Loài 2 + - - + -
Loài 3 + - + + -
Loài 4 - + + + +
Hướng dẫn vận dụng kiến thức:
Các giai đoạn của quá trình hô hấp tế bào:
Đường phân: xảy ra trong bào tương, bắt đầu quá trình phân giải nhờ bẻ gẫy gluocse hình thành hai phân tử có tên là pyruvate.
Chuyển hoá saccharide: Chu trình axit citric xảy ra bên trong chất nền ti thể của tế bào nhân thực hay đơn giản trong bào tương của tế bào nhân sơ, hoàn thành sự phân giải gluocse nhờ oxi hoá một dẫn xuất của pyruvate thành carbon dioxide. Do đó CO2 tạo ra do hô hấp là mảnh vỡ của các phân tử hữu cơ bị oxi hoá. Một số bước của đường phân và chu trình axit citric là các phản ứng oxi hoá khử trong đó dehydrogenaza chuyển electron từ cơ chất đến NAD+ để tạo NADH.
Hướng dẫn trả lời a)
- Các vi khuẩn không thể thực hiện được hô hấp hiếu khí gồm: loài 1, loài 2, loài 3.
- Loài 1 và loài 2 thiếu xitocrom c oxidaza là enzyme chính trong thành phần của quá trình hô hấp hiếu khí (enzym trong chuỗi truyền điện tử).
- Loài 3 thiếu enzyme citrat sintetaza của chu trình Crep
b)
- Loài 1 tạo rượu etanol, CO2 và ATP.
- Loài 2 tạo axit lactic, ATP.
- Loài 3 tạo axit lactic, ATP.
- Loài 4 khi có mặt oxi tạo CO2, H2O và ATP; khi không có mặt oxi tạo rượu etanol, CO2 và ATP.
Câu 19:
Một nhà khoa học tiến hành nuôi cấy nấm men rượu (Saccharomyces cerevisiae) trong một dung dịch dinh dưỡng đơn giản, dùng glucose đánh dấu 14C làm nguồn và năng lượng duy nhất. Cô ta thấy rằng cứ lmol glucose bị oxy hóa hoàn toàn thì tế bào lại tiêu thụ 6mol oxi, và tạo ra 32ATP (1NADH = 2.5 ATP và 1FADH2 = 1.5 ATP).
a, Cô ta đã đo lượng 14C của hợp chất này để có thể nói rằng glucose đã bị oxy hóa hoàn toàn? Tên của quá trình cô ta nghiên cứu?
b, Khi chuyển dung dịch nuôi cấy trên sang môi trường yếm khí và tiếp tục nghiên cứu xem điều gì xảy ra vói glucose đánh dấu phóng xạ, cô ta nhận thây rằng các tế bào tiếp tục sinh trưởng nhưng không tiêu thụ oxi? Theo bạn, quá trình cô ta nghiên cứu đó là quá trình gì? Quá trình này tạo ra bao nhiêu ATP? Cô ấy sẽ đánh dấu 14C trong những hợp chất nào để nghiên cứu quá trình này?
c, So sánh năng lượng của hai quá trình kể trên và giải thích lý do tại sao có sự khác biệt đó?
Hướng dẫn trả lời
a, Hô hấp hiếu khí ở tế bào. Quá trình này diễn ra trong điều kiện hiếu khí. Chất cho electron là glucose và chất nhận electron cuối cùng là O2 để tạo H2O. Tổng số lượng ATP tạo ra là 32ATP với hiệu suất ~ 40%.
b, Khi đưa sang muôi trường yếm khí, các chủng nấm men này sẽ thực hiện quá trình lên men tạo các sản phẩm hữu cơ. Quá trình này tạo chỉ 2ATP từ đường phân. Cô ấy nên đánh dấu vào 14C vào các phân tử hữu cơ như lactacte hoặc ethanol.
c, Năng
lượng (sô ATP/glucose)
Hiếu khí
Yếm khí
electron. Sự giải phóng e làm
động lực vận chuyên H+ từ
chất nền ra xoang sẽ tạo sự
chênh lệch H+ giữa hai bên
màng. Khi có sự chênh lệch,
H+ được quay trở lại chất nền
thông qua phức ATP synthase
và tổng hợp năng lượng ATP.
cũng có thể là ethanol nhờ alcohol dehydrogenase.

Vì thế nên năng lượng chỉ tạo 2ATP hoặc 1 ATP/lglucose.
Câu 20:
Hình dưới đây là thí nghiệm mà các nhà khoa học dựa trên giả thuyết cho rằng giữa sự vận chuyển electron từ NADH tới O2 có mối quan hệ với sự vận chuyển proton qua màng ty thể. Dựa vào biểu đổ với sự thay đổi độ pH trong khoảng thời gian (s) để chứng minh cho giả thuyết trên là đúng?
Giải thích Trong điều kiện hiếu khí, nấm men sẽ thực hiện chu trình Krebs. Năng lượng được tích lũy dưới dạng cơ chất (10NADH, 2FADH2) và được vận chuyển đến chuỗi truyền
Trong điều kiện kị khí, chất vận chuyên điện tử (NADH) sẽ được sử dụng để tạo lactate từ
pyruvate nhờ enzyme lactate dehydrogenase
Hướng dẫn trả lời:
Nếu NADH được đưa từ dịch huyền phù chứa ty thể thiếu oxy NADH sẽ không bị oxy hóa. Khi một lượng nhỏ oxy được thêm vào thí nghiệm, nồng độ proton trong môi trường quanh ty thể tăng đột ngột (pH giảm). Do đó sự oxy hóa NADH bằng oxy được kết họp với chuyển động của proton ra ngoài chất nền. Khi oxy hết, lượng proton dư thừa bên ngoài từ từ quay lại ty thể (cung cấp năng lượng tổng hợp ATP) và pH của môi trường bên ngoài trở về giá trị ban đầu. Điều đó chứng tỏ sự vận chuyển electron từ NADH tới oxy được cặp với vận chuyển proton qua màng ty thể.
Câu 21:
Một liposome nhân tạo là điều kiện lý tưởng để nghiên cứu sự di chuyển electron trong chuỗi truyền điện tử thay vì sử dụng ty thể. Khi một chất cho và một
chất nhận electron phù hợp được cho vào liposome, cùng với phức hệ IV (cytochrome c oxidase) gắn trên màng với mục đích nghiên cứu sự vận chuyển electron từ cytochrome c dạng khử đến O2. K+ gắn với valinomycin được đưa vào với mục đích cân bằng điện thế giữa màng trong và ngoài (K+ thường không thẩm thấu qua màng trong, nhưng valinomycin là một chất vận chuyển ion giúp mang K+ đi qua màng).
Về lý thuyết, có thể thay đổi cơ chế hoạt động của rotor, trục bên trong và núm xúc tác trong ATP synthase (hình) như thế nào để tổng hợp ATP? Giải thích.
Khi thêm O2 vào bên trong, chất nào được tạo ra? Nêu cơ chế của quá trình đó.
Dựa vào đồ thị ở hình (b), giải thích tại sao khi lượng oxy được đưa vào thì có sự thay đổi pH như vậy?
Chất độc được dùng như vũ khí hóa học, để tự tử khi gián điệp phát hiện, để xử tử tù bằng hơi gas, được Đức Quốc Xã dùng để giết hành loạt người Do Thái, ... Chúng bám vào heme cq của cytochrome c oxidase. Giải thích tại sao chúng gây độc?
Hướng dẫn trả lời:
Khi thêm oxy sẽ được nhận electron từ quá trình khử cyt c2+ thành cyt C xúc tác hình thành H2O. PTHH: 2H+ y2O2 H2O
Dựa vào đồ thị ta thấy, lượng oxy khi được đưa vào làm độ pH giảm mạnh vì cytochrome c ở dạng khử (cyt c2+) được oxy hóa thành dạng oxy hóa (cyt c3+), electron được đưa đến oxy để tạo nước. Electron được vận chuyển vào thì đồng thời proton được đưa ra ngoài môi trường làm cho pH giảm mạnh.
Khi cytochrome dạng khử được oxy hóa hoàn toàn, proton lại trở về bên trong túi làm pH dung môi quay trở lại giá trị ban đầu.
Cyanide khi được đưa vào sẽ ngăn chặn vận chuyển electron, quá trình phosphoryl hóa - oxy hóa bị ngăn cản và làm ngừng trệ việc tổng hợp ATP.

Câu 22
Một số vi khuẩn sống trong điều kiện môi trường kiềm (pH = 10) và duy trì được môi trường nội bào trung tính (pH = 7).
Tại sao các vi khuẩn này không thể tận dụng sự chênh lệch về nồng độ ion H+ giữa hai bên màng tế bào cho ATP synthase tổng hợp ATP? Giải thích.
Hướng dẫn trả lời:
Sự chênh lệch nồng độ ion H+ giữa hai bên màng tế bào dẫn đến ion H+ đi từ trong ra ngoài. ATP synthase chỉ tổng hợp ATP khi ion H+ đi từ ngoài vào trong. Do đó, ATP không được tổng hợp.
Khi H+ đi từ ngoài vào, rotor làm trục quay ngược chiều kim đồng hồ (nhìn từ phía tế bào chất) làm núm xúc tác tổng hợp ATP. Do đó, về lý thuyết, có thể thiết kế rotor làm trục vẫn quay ngược chiều kim đồng hồ khi ion H+ đi từ trong ra ngoài để núm xúc tác tổng hợp ATP. Khi ion H+ đi từ trong ra ngoài, trục quay theo chiều kim đồng hồ, ATP bị phân giải. Do đó, thiết kế cơ chế hoạt động của núm xúc tác khi trục quay theo chiều kim đồng hồ vẫn tổng hợp được ATP.
Câu 23:
Tính toán hiệu suất năng lượng trên số nguyên tử carbon giữa glucose và acid palmitic (16C). So sánh hiệu suất chuyển hóa năng lượng với glucose dựa trên số carbon. Acid béo là nguồn dự trữ năng lượng to lớn và hiệu quả của cơ thể. Chúng có gì đặc biệt khi được lựa chọn là nguồn dự trữ mà không phải chất hữu cơ khác?
Hướng dẫn trả lời:
Số vòng oxy hóa cần thiết để oxy hóa hoàn toàn một phân tử acid béo no có n nguyên tử carbon sẽ là: (n/2)-l. Palmitic (16C) sẽ có 7 vòng, phương trình tổng quát quá trình phân giải hoàn toàn một palmitic là:

Acetyl CoA + CoA + FAD+ + NAD+ + H2O->
Cn 2- acetyl Co A + acetyl - CoA + FADH2+ NADH + H+
Số lượng ATP được tạo thành qua sự oxy hóa hoàn toàn một phần tử palmitic (16C)
Giai đoạn phản úng
Beta-oxi hoá
Số coenzyme khử Số ATP tạo thành
7 FADH2 10,5
7 NADH 17,5 Oxy hoá acetylCoA qua chu trình citric
8 FADH2 12 24 NADH 60 8
Hoạt hoá axit béo -2
Tổng cộng 106
Như vậy, quá trình oxy hóa acid béo đã tạo ra một số năng lượng lớn cho tế bào. Nếu tính hiệu suất năng lượng trên số carbon thì quá trình oxy hóa acid béo tạo ra nhiều năng lượng hơn so với quá trình oxy.
Câu 24:
Những trẻ sơ sinh với triệu chứng co bản như giảm đường huyết, kèm theo yếu cơ và chuột rút là dấu hiệu của bệnh do sự khiếm khuyết trong sinh tổng hợp cRNAitine, rối loạn vận chuyển vào tế bào hay bài tiết quá mức ở thận. Dựa vào những kiến thức bạn đã tìm hiểu ở trên, hãy đưa ra một giải thích ngắn gọn cho tình trạng này?
Hướng dẫn trả lời
Sự thiếu cRNAitine xảy ra do khiếm khuyết trong sinh tổng hợp cRNAitine, rối loạn vận chuyển vào tế bào hay sự bài tiết quá mức ở thận. Bệnh này thường xảy ra ở trẻ sơ sinh, đặc biệt là trẻ thiếu tháng. Đây là kết quả của sự suy giảm oxy hóa acid béo khi cRNAitine không đáp ứng, nguồn cung câp năng lượng suy giảm. Đồng thời, quá trình này cũng kéo theo việc lipid tích lũy nhiều làm yếu cơ và chuột rút.
Câu 25: Cấu trúc của một lipoprotein đều bao gồm: một vỏ lưỡng vùng chứa một lớp phospholipid đơn (phospholipid), cholesterol, protein và một lõi kỵ nước chủ yếu là cholesterol ester hay triacylglyceride hoặc cả hai nhưng với lượng nhỏ các châ't béo trung tính. Hình dưới là mô hình lipoprotein mật độ thấp (LDL).
Thí nghiệm đánh dấu phóng xạ thể hiện mối quan hệ giữa tiền tố và sản phẩm trong quá trình hấp thu LDL của tế bào.
Để tạo điều kiện thuận lợi cho việc chuyển giao khối lượng của chất béo giữa các tế bào, các loài động vật đã tiến hóa một cách hiệu quả để gói hàng trăm đến hàng ngàn phân tử lipid trong một phương tiện vận chuyển đại phân tử hòa tan trong nước là lipoprotein. Cấu trúc của phần tử phù hợp như thế nào với vai trò vận chuyển chất béo?
Phương pháp nghiên cứu thực nghiệm đưa các hạt LDL vào tế bào bằng cách đánh dấu nguyên tử phóng xạ 125I bằng liên kết cộng hóa trị vào chuỗi bên của phân tử tyrosine trong apoB-100 trên bề mặt hạt LDL. Sau khi tế bào nuôi cấy được ủ nhiều giờ với các hạt LDL được đánh dấu, chúng ta có thể tính được số lượng các hạt LDL bám vào bề mặt tế bào, số hạt được lấy vào trong và thành phần apoB-100 của LDL bị tiêu enzyme thủy phân thành acid amin. Dựa vào đó, bạn hãy trình bày những sự kiện thay đổi trong quá trình nghiên cứu để chứng minh rằng sự kiện đó diễn ra theo một thứ tự: LDL liên kết với bề mặt tếbào và được lấy vào trong tế bào sau đó bị thủy phân.
Hướng dẫn trả lời Các lipoprotein này có lóp vỏ chứa apoprotein B (thường là B100) đảm nhận vai trò thụ thể giúp vận chuyển phân tử cholesterol vào trong tế bào.

Lớp vỏ lưỡng cực, mặt ngoài ưa nước làm cho các hạt hòa tan trong nước và bê' mặt bên trong kỵ nước.
Tiếp giáp với bề mặt kỵ nước là lõi của chất béo trung tính chủ yếu chứa cholesteron este, triacylglyceride cung cap cho tế bào thực hiện các nhiệm vụ khác nhau (tổng hợp steroid, năng lượng, thụ thể, …)
b. Nhìn vào sơ đồ ta thấy sự di chuyển của các phân tử LDL trong khoảng thời gian nhất định. Ban đầu, LDL liên kết trên bề mặt tế bào nhờ thụ thể là apoprotein B và di chuyển vào bên trong qua kênh vận chuyển. Tuy nhiên, sau khoảng 15-20 phút, thành phần của phân tử LDL bị thủy phân, trong đó apoprotein B bị enzyme thủy phân thành acid amin và được phát hiện bằng số lượng 125I. Sau khi apoprotein B bị tách ra
khỏi cholesterol, các phân tử cholesterol di chuyển thực hiện chức năng sinh học của mình.
Câu 26.
a) Cơ chế gây đột biến tự phát? Tại sao tần số đột biến tự phát lại thấp?
b) Các thể đột biến 1,2,3 được lai với các chủng có mất đoạn a và b, người ta chọn được một số thể tái tổ hợp kiểu dại. Dựa trên kết quả dưới đây, hãy xác định vị trí của mỗi đột biến (+ thể tái tổ hợp kiểu dại; - thể tái tổ hợp đột biến)?

Thể đột biến 1 Thể đột biến 2 Thể đột biến 3
- Đột biến 2 không sinh ra kiểu dại với mất đoạn a, vì vậy nó phải nằm trong vùng tướng ứng với mất đoạn a
- Đột biến 3 cho các kiểu dại với cả hai mất đoạn nên nó phải nằm ngoài cả hai đột biến mất đoạn.
Câu 27
Cho 3 chất dưới đây:
- Chất A có khả năng kháng khuẩn bằng cách ức chế các RNA polymerase trong tế bào vi khuẩn, do đó ức chế sự phiên mã dẫn đến prôtêin không được tổng hợp.
- Chất B là enzym glycoside hydrolase có khả năng phân giải thành peptidoglycan của tế bào vi khuẩn nên gây tan tế bào.
Chủng mất
đoạn a + - +
Chủng mất đoạn b - + +
Hướng dẫn vận dụng kiến thức: biến dị di truyền, cơ chế biến dị tự nhiên (trạng thái bazo nito, hoạt động tự sửa sai, yếu tố di truyền vận động, trao đổi chéo không cân, …)
Hướng dẫn trả lời a) Cơ chế gây đột biến tự phát:
- Sự bắt cặp sai của các bazonito hiếm
- Phản ứng của các bazo, do các gốc tự do có tính oxi hóa mạnh
- Do bộ máy di truyền sửa sai
- Do yếu tố di truyền vận động
- Do trao đổi chéo không cân…
Tần số đột biến tự phát lại thấp vì:
- Đặc tính của DNA bền vững, cấu trúc ổn định và có cơ chế duy trì cấu trúc
- Do có quá trình sữa chữa đột biến trong quá trình tái bản và khi DNA bị tổn thương.
- Enzyme DNA polimeraza hoạt động rất chính xác.
b)
- Đột biến 1 không sinh ra kiểu dại với mất đoạn b, vì vậy nó phải nằm bên trong vùng tương ứng với mất đoạn b.
- Chất C ức chế sự sinh trưởng và phát triển của vi khuẩn do tác động vào quá trình tổng hợp axit folic.
Hình dưới đây biểu diễn đường sinh trưởng của một chủng vi khuẩn được nuôi cấy trong 3 bình chứa môi trường dinh dưỡng dạng lỏng trước và sau khi bổ sung một trong 3 chất A, B, C nói trên tại các thời điểm được thể hiện trên hình (vị trí mũi tên). Đường nét liền biểu diễn số tế bào sống đếm được; đường nét đứt biểu hiện tổng số tế bào chết và sống đếm được.
Dựa vào đường sinh trưởng của vi khuẩn, hãy xác định chất nào đã được bổ sung vào môi trường nuôi cấy? Giải thích?
Hướng dẫn vận dụng kiến thức: Hoá sinh tế bào vi sinh vật, cấu trúc tế bào vi khuẩn và sự mẫn cảm của thành tế bào với enzim glycoside hydrolase.
Hướng dẫn trả lời:
Bình 1 C Bình 2. A Bình 3. B
Giải thích: Bình 1: số lượng tế bào sống tương đương tổng số lượng tế bào sống và chết chứng tỏ trường hợp này các tế bào không bị chết mà chỉ bị ức chế sinh trưởng. Vậy bình 1 là bỏ C vào, không có folic nên vi khuẩn không sinh trưởng
Bình 2: số lượng vi khuẩn chết khá nhiều, nhưng khi đếm tổng số tế bào sống và chết thì không đổi, do các tế bào chế không bị tan ra nên vẫn đếm được, là do bỏ thêm A, không tổng hợp được prôtêin nên vi khuẩn bị chế nhiều.
Bình 3: số lượng tế bào sống giảm nhanh, cũng không đếm được số lượng tế bào chết, chúng tỏ tế bào đã bị tan, do bỏ B vào nên thành bị hòa tan, làm cho vi khuẩn bị tan luôn.
Câu 28: Hình sau mô tả các thí nghiệm của F. Griffith và Chase trên Streptococcus pneumoniae:
Xử lí hỗn hợp vi khuẩn S chết và R sống ở thí nghiệm 4 bằng DNAase, proteinase, RNAase thì thấy chỉ có thỉ nghiệm được xử lí bằng DNAase mới khử được hiệu quả của biến nạp.
- Hiện tượng biến nạp được ứng dụng để chuyển DNA tái tổ hợp vào trong tế bào nhận.
- Hạn chế: DNA tái tổ hợp đi qua các lỗ thủng trên màng tế bào nên hiệu quả biến nạp phụ thuộc vào kích thước DNA tái tổ hợp và kích thước lỗ thủng => Ảnh hưởng tới tỉ lệ sống của tế bào sau biến nạp.
Câu 29:
Allopurinol một chất ức chế xanthine oxydase được tìm ra bởi hai nhà khoa học Gertrude Elion (1918-1999) và George Hitchings (1905-1998), nó sử dụng để điều trị bệnh gout (gút) mãn tính. Giải thích cơ sở hóa sinh cho điều trị bệnh này.
Trình bày vắ
Bằng cách nào có thể xác định được vật chất di truyền trong các thí nghiệm trên là DNA chứ không phải là RNA hay protein?

Trong kĩ thuật di truyền, hiện tượng xảy ra ở thí nghiệm 4 được ứng dụng để làm gì? Em hãy nêu vắn tắt hạn chế của việc ứng dụng hiện tượng này?
Hướng dẫn vận dụng kiến thức: di truyền vi khuẩn - hiện tượng biến nạp
Hướng dẫn trả lời:
* Nội dung thí nghiệm:
- Thí nghiệm 1: Tiêm vi khuẩn R sống vào chuột => chuột sống.
- Thí nghiệm 2: Tiêm vi khuẩn S sống vào chuột => chuột chết.
- Thí nghiệm 3: Tiêm vi khuẩn S đã bị giết bởi nhiệt vào chuột => Chuột sống
- Thí nghiệm 4: Trộn vi khuẩn S chết với R sống tiêm vào chuột => Chuột chết => Phân lập được vi khuẩn S.
* Giải thích:
- Vi khuẩn R không có màng nhầy, không gây bệnh ở chuột trong khi vi khuẩn S có.
- Vi khuẩn S bị giết bởi nhiệt có vật chất di truyền đã truyền vào vi khuẩn R qua con đường biến nạp, biến R thành S và làm chuột chết.
Allopurinol Oxypurinol
Hướng dẫn trả lời:
Gout là bệnh về khớp do nồng độ uric trong máu và mô tăng cao. Khi bị Gout, khớp trở nên viêm, đau do lắng đọng một cách không bình thường các tinh thể urate natri.
Allopurinol là một chất tương tự purine; nó là một đồng phân cấu trúc của hypoxanthine (một purine tự nhiên trong cơ thê) và là một chất ức chế cạnh tranh với enzyme xanthine oxidase. Xanthine oxidase chịu trách nhiệm cho quá trình oxy hóa liên tiếp của hypoxanthine và xanthine, dẫn đến sản xuất acid uric, sản phẩm chuyển hóa purine của con người. Ngoài việc ngăn chặn sự sản xuất axit uric, sự ức chế của xanthine oxidase làm tăng hypoxanthine và xanthine. Khi hypoxanthine và xanthine

lớn, chúng có thể ức chế tổng hợp hoặc chuyển vào tạo adenosine và guanosine monophosphate. Do đó, Allopurinol làm giảm sự hình thành acid uric và cũng có thể ức chế tổng hợp purine. Gout được điều trị hiệu quả bằng kết hợp giữa thuốc và dinh dưỡng. Các thực phẩm giàu nucleic và nucleotide được loại khỏi bữa ăn, kết hợp với việc sử dụng thuốc allopurinol.
Câu 30: Trong một thử nghiệm được thực hiện vào năm 1962, một cysteine gắn với tRNA được thay thế bằng alanine. Những tRNA "lai" sau đó được thêm vào một hệ thống dich mã, đồng thời loại bỏ các cysteine-trNA. Khi kết quả phân tích protein được theo dõi, alanine đã được chèn vào mọi điểm trong chuỗi polypeptide nơi cysteine được cho là có mặt. Thông qua thử nghiệm này, cho biết vai trò của aminoacyl tRNA synthetases trong quá trình dịch mã bình thường?
Hướng dẫn giải
Thử nghiệm này chứng minh rằng ribosome không kiểm tra acid amin được gắn vào tRNA. Khi một acid amin đã được kết họp với một tRNA, ribosome sẽ "tự do" kết hợp acid amin đó vào theo vị trí liên kết codon và anticodon. Do đó, có thể kết luận rằng một phần của việc đọc chính xác mã di truyền, là kết hợp của một codon trong một mRNA với đúng amino acid, được thực hiện bởi các enzyme synthetase một cách chính xác phù hợp với tRNA …
Câu 31: Phép lai giữa các tế bào his+, leu+, thr+, pro+, mẫn cảm với str và các tế bào his-, leu-, thr-, pro-, kháng str, kéo dài 25 phút. Khi ngừng tiếp hợp, các kiểu gen có tái tổ hợp được xác định. Kết quả thu được ở bảng dưới đây. Hãy xác định các gen đầu tiên và trật tự có thể của các gen.
Kiểu gen Số khuẩn lạc
Hướng dẫn vận dụng kiến thức: Quá trình giao nạp (tiếp hợp) ở vi khuẩn Trong quá trình tiếp hợp, một gen có tần số truyền gen cao phải nằm gần với điểm khởi đầu quá trình truyền gen.
Hướng dẫn trả lời:
Chúng ta thấy rằng các tế bào thr+ nhiều nhất. Tần số đứt gãy ống tiếp hợp tăng với sự tăng thời gian tiếp hợp. Vì vậy các gen ở càng xa điểm khởi đầu truyền gen sẽ xuất hiện với tần số càng nhỏ. Chúng ta có thể sắp xếp các gen một cách tương đối dựa trên tần số của các kiểu gen phát hiện được. Trật tự đó là thr leu pro his. Vì không thấy
có his+ và vì chúng ta dừng quá trình tiếp hợp ở 25 phút, nên gen his phải nằm sau 25 phút trên bản đồ thời gian.
Câu 32:
Các chủng bị mất đoạn dưới đây được lai với nhau và chọn ra các thể tái tổ hợp kiểu dại. Kết quả như sau (+: có các thể tái tổ hợp kiểu dại, - không có các thể tái tổ hợp kiểu dại). Hãy xác định trật tự của các mất đoạn. 1 2 3 - ++ -- - -
Hướng dẫn vận dụng kiến thức:
Biến dị ở vi khuẩn: đột biến mất đoạn, nếu hai mất đoạn có chung một phần bị mất (nghĩa là chúng trùm lên nhau) thì chúng không thể cho ra các thể tái tổ hợp kiểu dại.
TH1: Hai mất đoạn không trùm lên nhau:
Sự tái tổ hợp sẽ tạo ra: DNA mang hai mất đoạn: ________/////////___/////////
và DNA bình thường: ______________
TH2: Hai mất đoạn trùm lên nhau: ________///////_______ _________////////_____
Tái tổ hợp sẽ tạo ra các DNA đều có đoạn bị mất.
Hướng dẫn trả lời:
Chúng ta thấy rằng chủng 3 không cho ra thể tái tổ hợp kiểu dại nào. Nó phải mang mất đoạn kéo dài ít nhất đến phần đoạn bị mất ở chủng 1 và 2. Các mất đoạn ở chủng 1 và 2 cho ra các thể tái tổ hợp kiểu dại, vì vậy nó không trùm lên nhau.
Vậy bản đồ có thể là:
Câu 33:
Các thể đột biến điểm 1, 2, 3 được lai với các chủng có mất đoạn a và b; người ta chọn được một số thể tái tổ hợp kiểu dại. Dựa trên kết quả dưới đây, hãy xác định vị trí của mỗi đột biến (+ = thể tái tổ hợp kiểu dại, - = thể tái tổ hợp đột biến).
1 2 3 + - + - + +
Hướng dẫn vận dụng kiến thức: Biến dị - đột biến điểm ở vi khuẩn. Nếu một đột biến điểm cho ra các thể tái tổ hợp khi lai với một mất đoạn, thì đột biến điểm đó nằm ngoài vùng bị mất đoạn.
TH1: điểm đột biến ở bên ngoài đoạn mất
Sự tái tổ hợp sẽ tạo ra một DNA mang đột biến kép:
và một AND bình thường:
TH2: Đột biến nằm bên trong đoạn mất
Sự tái tổ hợp sẽ không xảy ra trong vùng có mất đoạn. Do vậy tất cả các thể tái tổ hợp vẫn là đột biến.
Hướng dẫn trả lời
Đột biến 1 không sinh ra kiểu dại với mất đoạn b, vì vậy nó phải nằm bên trong vùng tương ứng với b.
Lập luận tương tự, ta có đột biến 2 nằm trong vùng tương ứng với mất đoạn a. Vì đột biến 3 cho các dạng dại với cả hai mất đoạn, nên nó phải nằm ngoài cả hai đột biến mất đoạn.
Câu 35:
a. Trong một thực nghiệm biến nạp, thể cho là ara+nic+gal+ và thể nhận là aranic-gal-. Các thể biến nạp ara được chọn và sau đó kiểm tra tiếp. Kết quả cho thấy 40% là ara+gal+ và 5% là ara+nic+. Xác định trật tự có thể của các gen.
b. Bạn có thể chỉ cần làm thêm một thực nghiệm biến nạp để xác định trật tự gen. Bạn phải sử dụng cùng thể cho và thể nhận, nhưng có thể thay đổi quá trình chọn lọc các thể biến nạp đầu tiên. Bạn phải làm gì và kết quả như thế nào khi xác định trật tự gen mà bạn cần?
Hướng dẫn trả lời
a. ara nic gal hoặc nic ara gal.
b. Chọn lọc theo gal, sau đó đếm số khuẩn lạc gal+ara+ và gal+nic+.
Nếu trật tự ara nic gal là đúng thì số khuẩn lạc gal+ ara+< gal+nic+. Vì nic và ara đồng biến nạp 40% thời gian nên chúng phải gần và liền kề nhau. Nếu gal ở giữa thì nic+gal+ phải có tần số đồng biến nạp lớn hơn nic+ara+.
Nếu chúng ta chọn lọc theo gal+ và thứ tự là ara nic gal, thì kiểu gen gal+ara+nic- chỉ cần hai trao đổi chéo đồng thời, trong khi gal+ara+ chỉ đòi hỏi một trao đổi chéo. Nếu trật tự là nic ara gal thì gal+ ara+ chỉ cần một trao đổi chéo và tần số thu được phải lớn hơn gal+nic+.
Câu 36
Trong một thực nghiệm biến nạp, x+y+z+ được sử dụng là thể cho và x-y-z- là thể nhận, thu được 250 thể biến nạp. Sau đó nuôi cấy bản sao để xác định xem có y+ và/ hoặc z+ không. Kiểu gen của các thể biến nạp thu được như dưới đây. Có thể kết luận gì về vị trí tương ứng của các gen?
x+yz - 53
x+yz+ 173
x+y+z - 7
x+y+z+ 17
Hướng dẫn vận dụng kiến thức: biến nạp ở vi khuẩn, lập bản đồ tái tổ hợp
Hướng dẫn trả lời: Chúng ta thấy rằng 76% khuẩn lạc có x+z+. Kết quả này cho thấy x và z gần nhau. x+ và y+ xuất hiện cùng nhau với tỉ lệ 10% cho thấy y cách xa x, và có thể không liên kết với nhau.
Để tính tần số tái tổ hợp giữa x và z, chúng ta xem số lần x+ và z+ không cùng xuất hiện (đó là x+ y- x- và x+ y+ z -).
RF = (53 + 7)/240 = 25 đơn vị bản đồ
Câu 37:
Vi khuẩn E. coli được gây nhiễm một cách lẫn lộn với hai loại phage T4: r+ m+ tu+ và r m tu. Đời con xuất hiện như sau:
+++ 1243
++ tu 322
r m+ 284
r m tu 1158
+ m + 175
+ m tu 54
r + + 58
r + tu 160
Tổng số 3454
Tính khoảng cách giữa các gen và vẽ bản đồ di truyền?
Hướng dẫn vận dụng kiến thức: tái tổ hợp gen ở vi khuẩn và lập bản đồ di truyền ở vi khuẩn bằng tái tổ hợp
Hướng dẫn trả lời:
Xét cặp r-m, hãy xác định các lớp mà trong đó r+m+ không cùng xuất hiện.
RF r-m = (175 + 54 + 58 + 160)/3454 x 100 = 13 đơn vị bản đồ.
Tương tự, RF r-tu = (322 + 284 + 54 + 58) x 100/3454 = 21 đơn vị bản đồ.
RF m-tu = (322 + 284 + 175 + 160)/3454 x 100 = 27 đơn vị bản đồ.
Từ kết quả trên ta có bản đồ di truyền là: ___ m ____ 13 ___ r _________ 21 ___ tu __
Để kiểm tra lại, hãy xác định các trao đổi chéo kép để tạo thành các thể tái tổ hợp r+m tu và r m+tu+. Ta thấy rằng chỉ có các alen r bị thay đổi do sự sắp xếp lại từ bố mẹ, vì vậy r phải nằm giữa.
Câu 38
Có 9 chủng his-, ba trong số đó có những mất đoạn và 6 chủng có những đột biến điểm. Cấy 9 chủng trên môi trường tối thiểu và đếm số khuẩn lạc mọc (+ = mọc được). 1 2 3 4 5 6 7 8 9
- + + - + - + +
Phage được nhiễm vào một trong 9 chủng và sau đó được sử dụng để tải nạp mỗi chủng mất đoạn. Các thể tải nạp từ mỗi phép lai được cấy trên môi trường tối thiểu và đếm các khuẩn lạc mọc (+ = thể tải nạp mọc được). Dựa trên kết quả sau, xác định:
a. Chủng nào là chủng đột biến mất đoạn.
b. Vị trí tương ứng của mỗi đột biến mất đoạn với mất đoạn khác?
Vị trí tương ứng của 6 đột biến điểm. Chủng
Mất đoạn 1 2 3 4 5 6 7 8 9
A + - - - - - - + +
B + - + + - - + + -
C + - + - + + - - +
Hướng dẫn trả lời:
a. Các chủng 2, 5, 7 là những mất đoạn. 5 ////////////////
7 ////////////
2 //////////////////////////
1 9 6 3 4 8
b. Nuôi cấy trong môi trường tối thiểu cho phép phát hiện các thể hồi biến. Vì các mất đoạn không phục hồi được nên chúng không mọc được trên môi trường tối thiểu. Đã biết 2, 5, 7 là những mất đoạn, ta hãy phân biệt các chủng này từ bộ số liệu thứ hai.
2 5 7
A - - -
B - - +
C - + -
Chủng 2 không cho thể tái tổ hợp kiểu dại, vì vậy nó phải trùm lên hai đột biến mất đoạn khác. Các mất đoạn 5 và 7 không trùm lên nhau. Thể đột biến 2 cho kết quả tương tự chủng A nên A=2. Vì B tái tổ hợp với 7, nên B phải là 5; do đó C là 7. Thể đột biến 9 không cho các tái tổ hợp kiểu dại với các mất đoạn 2 và 7. Nó phải nằm ở vùng được định vị duy nhất trong đột biến 5. Lý luận tương tự đối với đột biến 8: 8 ở vùng duy nhất với 7, 3 ở vùng duy nhất với 2. Các đột biến 4 và 6 phải nằm trong vùng mất đoạn chung cho 2 và 7 cũng như cho 2 và 5 tương ứng. Đột biến 1 cho các thể tái tổ hợp kiểu dại với tất cả các mất đoạn, nó phải nằm bên ngoài tất cả các vùng mất đoạn. Nó có thể nằm ở một trong hai đầu tận cùng của bản đồ
Câu 39
Chủng vi khuẩn lys+his+val+ được dùng làm thể cho, còn chủng lys-his-val- là thể nhận. Các thể biến nạp đầu tiên được tách ra trên môi trường tối thiểu có chứa his và val.
Những kiểu gen nào sẽ sinh trưởng trên môi trường này?
Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + his, 75% các khuẩn lạc ban đầu sinh trưởng được. Những kiểu gen nào sẽ sinh trưởng được trên môi trường này?
Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + val, 6% số khuẩn lạc sinh trưởng được. Những kiểu gen nào sinh trưởng được trên môi trường này?
Cuối cùng, cho các khuẩn lạc này sinh sản trên môi trường tối thiểu. Không khuẩn lạc nào sinh trưởng được. Dựa vào thông tin này, xác định xem kiểu gen nào sinh trưởng được trên môi trường tối thiểu + his? trên môi trường tối thiểu + val?
Dựa trên những thông tin trên, tìm xem gen nào nằm gần gen lys.
Thí nghiệm biến nạp được lặp lại, song đĩa nuôi cấy có chứa môi trường tối thiểu + lys + his. 50% khuẩn lạc sống được. Cho các khuẩn lạc này sinh sản trên các môi trường khác nhau để xác định kiểu gen của chúng. Kết quả như sau:
val-his+lys+ 0
val+his-lys+ 37
val+his+lys- 3
Dựa vào kết quả trên, hãy xác định trật tự các gen.
Hướng dẫn vận dụng kiến thức: Biến nạp và tái tổ hợp ở vi khuẩn
Hướng dẫn trả lời:
a. lys+ his+ val+
lys+ his+ val-
lys+ his- val+
lys+ his- val-
Kiểu gen nào có lys+ mới mọc được.
b lys+ val+ his-
lys+ val+ his-
Kiểu gen nào có lys+ và val+ mới mọc được.
c. lys+ val+ his+
lys+ val- his-
Kiểu gen nào có lys+ và his+ mới mọc được.
d lys+ val+ his-
lys+ val- his+
Chúng ta không thấy có kiểu gen lys+ val+ his+ .
e. lys+ và val+ nằm gần nhau; chúng đồng biến nạp 75% thời gian. Trật tự gen có thể là lys val his hoặc val his lys.
f. val lys his. Nếu trật tự gen là val lys his thì kiểu gen val+ lys- his+ phải hiếm gặp vì nó được tạo ra do trao đổi chéo kép.
Câu 40
Bạn phân lập được một số đột biến điểm (ký hiệu từ a đến g) ở vùng rII của phage T4. Các phép lai ghép đôi cho tỷ lệ phần trăm các vết tan kiểu dại như sau:
Phép lai %kiểu dại Phép lai % kiểu dại
a x b 0. 5 a x c 0. 3
a x d 0. 8 a x f 0. 3
b x e 0. 4 b x d 1. 3
b x f 0. 2 c x d 0. 5
c x g 0. 8 e x g 1. 0
b x g 1. 6 g x d 0. 3
Bạn cũng lai mỗi đột biến điểm với mỗi đột biến mất đoạn và tính các thể tái tổ hợp kiểu dại. Kết quả thu được dưới đây (+ = thể tái tổ hợp kiểu dại, - = thể tái tổ hợp đột biến).
Đột biến điểm
Mất đoạn a b c d e f g
a. Vẽ bản đồ các đột biến điểm, tính khoảng cách.
b. Xác định vị trí của các mất đoạn tương ứng với các đột biến điểm.
c. Các thể đột biến a và e bổ trợ với nhau và e được biết là thuộc rIIB. Các đột biến nào ở rIIA và đột biến nào thuộc rIIB?
d:
d. Những tổ hợp của các mất đoạn nào sẽ tạo ra các thể tái tổ hợp kiểu dại?
Hướng dẫn vận dụng kiến thức: Đột biến điểm và tái tổ hợp ở vi khuẩn.
Hướng dẫn trả lời:
a. 0. 2 0. 3 0. 1 0. 2 0. 5 0. 3 | | | | | | | | b f a e c d g
b.
c. a, b, f ở trong rIIA; e, c, d và g ở trong rIIB.
d. Không tổ hợp nào.
Các đột biến b và d phải xa nhau, b và g cũng vậy, e và f phải nằm giữa b và
____ b ___0.2 ___ f __0.4 ___ e ____ d ___ 0.3 __ g
Bây giờ hãy xác định vị trí của a và c. Vì a-b = 0,5 nên a phải cách f là 0. 3 về
bên phải. Đột biến c phải ở bên phải a, hoặc nó có thể cùng vị trí với f. Đột biến c phải ở bên phải a và khoảng cách được cộng kế tiếp. Hãy tìm các dấu (-) trong các phép lai; các đột biến này sẽ bị phủ bởi các mất đoạn riêng. Mất đoạn 1 phải kéo dài dến các đột biến a, c và e vì không có thể tái tổ hợp kiểu dại nào xuất hiện. Tương tự, đột biến 2 phải kéo dài đến c, d, g. Lý luận tương tự cho phép xác định các mất đoạn 3-5. Vì a bổ trợ e, vậy a phải thuộc rIIA, cũng giống như b và f. Tất cả các mất đoạn đều thiếu c, do vậy không thể sinh ra các thể tái tổ hợp kiểu dại.
1. Kết luận
Sau khi thực hiện, chuyên đề đã nghiên cứu các vấn đề lý thuyết của vấn đề Hoá sinh tế bào, di truyền vi sinh vật học. Chuyên đề cũng đã đưa ra được một số các câu hỏi áp dụng kiến thức tích hợp hoá sinh, vi sinh, di truyền trong các đề thi. Từ đó, góp một phần nhỏ vào quá trình nghiên cứu của học sinh và giáo viên, đặc biệt là trong ôn thi đội tuyển học sinh giỏi.
2. Đề xuất
Việc tìm hiểu, nghiên cứu các cơ chế hoá sinh tế bào, di truyền trên các đối tượng vi sinh vật làm nền tảng để giải quyết các vấn đề liên quan đến kĩ thuật di truyền, cách mạng Công nghệ sinh học.
Tuy nhiên, do điều kiện và thời gian viết chuyên đề của tôi còn hạn chế. Vì vậy, những kết quả nghiên cứu trên đây mới chỉ là bước đầu, chúng tôi hy vọng sẽ có những đề tài nghiên cứu sâu sắc hơn về sự tích hợp các kiến thức hoá sinh, di truyền, vi sinh, từ đó ứng dụng những kiến thức này vào thực tiễn đời sống, ứng dụng trong các ngành Sinh học khác nhau. Chuyên đề của chúng tôi chắc hẳn không tránh khỏi những thiếu sót, nên rất mong nhận được sự góp ý tận tình của các bạn đồng nghiệp để chuyên đề hoàn thiện hơn!
TÀI LIỆU THAM KHẢO
1. Campbell & Reece (2015). Sinh học, Nhà xuất bản giáo dục, Hà Nội.
2. Nguyễn Tấn Thành (2018). Bồi dưỡng Học sinh giỏi Hoá sinh - tế bào, Nhà xuất bản Đại học Quốc gia Hà Nội
3. Phạm Thị Trân Châu, Trần Thị Áng. Hoá sinh học, Nhà xuất bản Giáo dục Việt Nam
4. TS. Nguyễn Thị Hồng Vân - TS. Bùi Thị Việt Hà. Giáo trình di truyền học sinh vật nhân sơ và virut, Nhà xuất bản Giáo dục Việt Nam
5. Hoàng Trọng Phán, Trương Thị Bích Phượng, Giáo trình di truyền học vi sinh vật và ứng dụng, Nhà xuất bản Đại học sư phạm Huế.
6. Phạm Thành Hổ, Di truyền học, Nhà xuất bản Giáo dục
7. Urry, Cain & et at (2017). Biology Campbell & Reece 11th edition, Peason, NewYork.
8. Samuel Baron. Medical Microbiology, 4th Edition, Galveston (TX), 1996
9. Các đề thi đề xuất chọn đội tuyển HSGQG của một số tỉnh thành, đề thi chọn học sinh giỏi Quốc gia, chọn học sinh dự thi Quốc tế và đề thi Quốc tế các năm.
129
Chuyên đề:
MỘT SỐ VẤN ĐỀ TÍCH HỢP
HÓA SINH – VI SINH – DI TRUYỀN HỌC VI KHUẨN
Nguyễn Thị Cúc - Trường THPT Chuyên Biên Hòa – Hà Nam
Chuyên đề đạt giải Nhì
1. Lí do chọn đề tài
PHẦN I. MỞ ĐẦU
Sự ra đời của di truyền học phân tử (molecular genetics) gắn liền với các khám phá về DNA (deoxyribonucleic acid) từ giữa thế kỷ XX trên đối tượng nghiên cứu chủ yếu là các vi sinh vật. Tuy nhiên, trước đó Friedrich Miescher (1869) đã khám phá ra một hỗn hợp trong nhân tế bào gọi là nuclein mà thành phần chính của nó sau này được biết là DNA. Về mối quan hệ giữa gene và protein, từ 1902 Archibald Garrod qua nghiên cứu bệnh alcaptonuria ở người đã gợi ý rằng đây là một tính trạng lặn Mendel, có thể liên quan tới sự sai hỏng một enzyme. Bằng các thí nghiệm gây đột biến các gene liên quan đến các con đường sinh hóa trên nấm mốc Neurospora, năm
1941 George Beadle và E.L.Tatum xác nhận mỗi gene kiểm soát sự tổng hợp một enzyme đặc thù. Chính giả thuyết một gene-một enzyme (one gene-one enzyme hypothesis) nổi tiếng này đã mở đường cho sự ra đời của di truyền hóa-sinh, và hai ông đã được trao giải Nobel cùng với Joshua Lederberg năm 1958. Về sau, giả thuyết này được chính xác hóa là một gene xác định chỉ một chuỗi polypeptid - cấu trúc sơ cấp của các protein, trong đó có các enzyme.
Cho đến đầu thập niên 1940 các vi sinh vật, bao gồm các vi khuẩn và virus của chúng và các vi sinh vật nhân chuẩn đơn bào như nấm men, nấm mốc... thực sự trở thành các đối tượng nghiên cứu chính yếu của di truyền học. Từ đây hình thành các
lĩnh vực di truyền học sinh-hoá và di truyền học vi sinh vật, hai nền tảng chính cho sự ra đời của di truyền học phân tử (1953) và công nghệ ADN tái tổ hợp sau này (1978).
Có thể nói, nền tảng của công nghệ DNA tái tổ hợp (recombinant DNA technology) được thành lập từ 1972 khi Paul Berg tạo ra phân tử DNA tái tổ hợp đầu tiên trong ống nghiệm (recombinant DNA in vitro). Một năm sau Herbert Boyer và Stanley Cohen lần đầu tiên sử dụng plasmid để tạo dòng DNA. Lĩnh vực ứng dụng mới này của sinh học phân tử đã tạo ra một cuộc cách mạng mới trong sinh học.
Ở đây vi khuẩn E. coli được xem là một sinh vật mô hình nhất quán tuyệt vời của di truyền học hiện đại. Nó được sử dụng một cách rộng rãi trong các thí nghiệm chứng minh các phương thức tái bản bán của DNA (Meselson và Stahl 1958; John Cairns 1961; Okazaki 1969), phân tích tái tổ hợp và lập bản đồ di truyền, nghiên cứu cấu trúc tinh vi và chức năng sinh hoá của gene (Benzer 1961; Yanofsky 1961); cơ chế điều hoà sinh tổng hợp protein (Jacob và Monod 1961) v.v.
Với sự phát triển vô cùng nhanh chóng của di truyền học trong vài thập niên qua, đặc biệt là sự tiến bộ của công nghệ sinh học (biotechnology) nói chung đã có những tác động mạnh mẽ lên nhiều ngành khoa học và trên mọi mặt của đời sống, kinh tế, chính trị và xã hội ở phạm vi toàn cầu. Di truyền học nói chung và di truyền học vi sinh vật nói riêng được hình dung ở vị trí trung tâm và giao thoa với sinh học, hóa sinh học, công nghệ, y-dược, nông nghiệp, sinh thái học, kinh tế học…..
Chương trình sinh học THPT chuyên sâu hiện nay, di truyền học vi sinh vật chưa có chuyên đề riêng mà gần như được trình bày tích hợp vào nội dung khác. Điều này gây ra những khó khăn nhất định cho cả giáo viên và học sinh trong việc hiểu và vận dụng linh hoạt nội dung kiến thức về hóa sinh – vi sinh và di truyền vi khuẩn. Ngoài ra, hình thức thi của các kì thi cũng rất khác nhau: Kì thi HSG cấp tỉnh, cấp quốc gia thi theo hình thức tự luận, kì thi HSG Olympic quốc tế chủ yếu thi theo hình thức trắc nghiệm. Vì vậy, trong quá trình dạy học, giáo viên phải rèn cho học sinh các kĩ năng học tập và làm bài sao cho phù hợp với mỗi hình thức thi. Và cho dù thi dưới hình thức nào thì vấn đề cốt lõi vẫn là học sinh phải nhớ, hiểu và biết vận dụng kiến thức đã học.
Với những lí do trên, tôi chọn đề tài: Một số vấn đề tích hợp hóa sinh – vi sinh – di truyền học vi khuẩn.
2. Mục đích của đề tài
- Cung cấp những kiến thức cơ bản và chuyên sâu về di truyền học hóa – sinh và di truyền học vi khuẩn
- Xây dựng hệ thống câu hỏi vận dụng liên quan đến mảng kiến thức này.
- Áp dụng trong dạy học lớp chuyên và ôn luyện học sinh giỏi các cấp nhằm nâng cao kết quả học tập của học sinh.
Nội dung của chuyên đề bao gồm 2 phần:
Phần 1: Khái quát kiến thức cơ bản.
Phần 2: Hệ thống câu hỏi ôn tập.
CHƯƠNG I. KHÁI QUÁT VỀ VI SINH VẬT
I. Sơ lược nghiên cứu vi sinh vật
Vi sinh vật (microorganisms, microbials) là tên gọi chung dùng để chỉ tất cả các sinh vật có hình thể bé nhỏ mà chỉ có thể nhìn thấy dưới kính hiển vi quang học hoặc kính hiển vi điện tử. Lĩnh vực nghiên cứu này gọi là Vi sinh học (microbiology), ra đời cách đây hơn 300 năm bởi Antoni van Leeuwenhoek (1676) với khám phá đầu tiên các vi sinh vật bằng kính hiển vi đơn giản.
Các vi sinh vật không phải là một nhóm riêng biệt hoặc một đơn vị phân loại, mà thường bao gồm nhiều nhóm giới khác nhau rất đa dạng, từ các virus, vi khuẩn cho đến các vi sinh vật eukaryote. Dựa vào phân loại học phân tử, năm 1977 Carl Woese chia sinh vật nhân sơ thành 2 nhóm dựa trên trình tự 16S rRNA, gọi là nhóm hay vực (domain) vi khuẩn thực (Eubacteria) và vi khuẩn cổ (Archaebacteria). Ông lý luận rằng hai nhóm này, cùng với sinh vật nhân chuẩn, tiến hóa độc lập với nhau và vào năm 1990 nhấn mạnh thêm quan điểm này bằng cách đưa ra hệ thống phân loại 3 vực, bao gồm vi khuẩn (Bacteria), vi khuẩn cổ (Archaea) và sinh vật nhân chuẩn (Eukarya). Quan điểm này nói chung được chấp nhận rộng rãi giữa các nhà sinh học phân tử
II. Đặc điểm của các vi sinh vật
1. Kích thước bé nhỏ: Các vi sinh vật có kích thước rất bé, đo bằng đơn vị micromet (1μm = 10-6 m) như các vi nấm, vi khuẩn hoặc nanomet (1nm = 10-9 nm) như các virus. Ví dụ: Các tế bào nấm men có đường kính 5 -10 μm. Các vi khuẩn có đường kính × chiều dài cơ thể thay đổi trong khoảng (0,2 - 2,0) × (2,0 -8,0) μm; hay như E. coli chẳng hạn rất bé: 0,5 × 2,0 μm v.v.
2. Hấp thụ nhiều, chuyển hoá nhanh:
Các vi sinh vật tuy nhỏ bé nhất trong sinh giới, nhưng năng lực hấp thu và chuyển hoá của chúng có thể vượt xa các sinh vật bậc cao. Chẳng hạn, vi khuẩn lactic (Lactobacillus) trong 1 giờ có thể phân giải một lượng đường lactose nặng hơn 1.00010.000 lần khối lượng cơ thể chúng...
3. Khả năng sinh sản nhanh:
So với các sinh vật khác thì các vi sinh vật có tốc độ sinh trưởng và sinh sôi nảy nở cực kỳ nhanh. Chẳng hạn, ở E. coli, trong điều kiện thích hợp, thời gian một thế hệ kéo dài khoảng 20 phút. Nếu không bị các điều kiện tự nhiên khống chế, chỉ sau một ngày đêm từ một tế bào ban đầu sẽ sinh sản được 272 tế bào, nặng 4.722 tấn!
4. Khả năng thích ứng rất cao và phát sinh biến dị mạnh:
Nói chung, các vi sinh vật vốn có các cơ chế điều hoà chuyển hoá để thích
ứng được với các điều kiện sống bất lợi. Trong một tế bào vi sinh vật, số lượng các
enzyme thích ứng chiếm tới 10% hàm lượng protein. Nếu có một thay đổi chất dinh dưỡng thì chỉ sau 1/1.000 giây, chúng đã có thể thay đổi để thích ứng rồi. Một số vi khuẩn có thể tiến hành quang hợp dưới tác dụng của ánh sáng, sống không cần oxy; nhưng nếu chuyển vào trong tối lập tức chúng có thể sử dụng oxy để sống. Một số vi sinh vật khi gặp các điều kiện khắc nghiệt thì chuyển sang trạng thái bào tử, ngừng hoạt động. Một số có thể sinh trưởng ngay cả ở nhiệt độ rất cao 2500 C, hoặc sống ở đáy sâu đại dương với áp suất khoảng 1.100 atm, v.v.
Liên quan tới khả năng thích ứng cũng như sự phong phú về chủng loại, các vi sinh vật còn có đặc tính quan trọng nữa đó là dễ phát sinh biến dị, với tần số trung bình 10-5 -10-10. Nguyên do bởi vì cơ thể chúng thường là đơn bào với bộ gene đơn bội, sinh sản nhanh, số lượng nhiều, tiếp xúc trực tiếp với môi trường sống. Hình thức biến dị thường gặp là các đột biến gene và kéo theo các biến đổi về hình thái, cấu tạo, kiểu trao đổi chất, sản phẩm trao đổi chất, tính kháng nguyên, tính đề kháng ...
5. Phân bố rộng, chủng loại nhiều:
Các vi sinh vật phân bố khắp mọi nơi và phát triển nhanh chóng ở những nơi có đủ thức ăn, độ ẩm, và nhiệt độ tối ưu cho sự phân chia và lớn lên của chúng. Chúng có thể được mang đi bởi gió từ nơi này sang nơi khác. Cơ thể người là nơi cư trú của hằng tỷ vi sinh vật; chúng ở trên da, đường ruột, trong mũi, miệng và những chỗ hở khác của cơ thể. Chúng có trong không khí, nước uống và thức ăn.
CHƯƠNG II : KHÁI QUÁT HÌNH THÁI CẤU TRÚC VÀ CẤU TẠO VI
KHUẨN
1. Hình thái - kích thước vi khuẩn
- Vi khuẩn ( Bacteria) Là những sinh vật mà cơ thể chi gồm một tế bào, có hình thái và đặc tính sinh vật riêng, có thể nuôi cấy trong các môi trường nhân tạo và quan sát được trên kính hiển vi bình thường


- Một số có khả năng gây bệnh rộng rãi cho sinh vật nói chung
- Đa số sống họại sinh trong tự nhiên, một số loài có khả nàng tiết ra chất kháng sinh như Bacillus subtilis tiết subtilin, hoặc Bacillus brevis tiết Tirotoxin...
- Vi khuẩn có hình thái nhất định do màng vi khuẩn quyết định, một số không có màng nên không có hình thái nhất định
1.1. Hình thái của vi khuẩn
Hình dạng vi khuẩn khác nhau ở loài này và loài khác, đối với các vi khuẩn đa hình thì hình dạng có thể khác nhau trong các giai đoạn sống khác nhau của chu kỳ sinh trưởng.
- Vi khuẩn có các hình dạng chính:
- Cầu khuẩn (Coccus)
- Trực khuẩn (Bacillus, Bacterium)




- Cầu trực khuẩn (Cocco- bacillus)
- Xoắn khuẩn (Spirillum)
- Phẩy khuẩn (Vibrio)
* Cầu khuẩn
- Nếu TB phân chia theo một phương và dính nhau thì gọi là song cầu khuẩn hoặc chuỗi cầu khuẩn.
- Khi phân chia theo 2 phương và dính nhau thì tạo thành tứ cầu khuẩn
- Khi phân chia theo 3 phương và dính nhau tạo thành liên cầu khuẩn
- Khi phân chia theo nhiều phương và dính nhau tạo thành tụ cầu khuẩn
Hình 2: Một số dạng cầu khuẩn
* Trực khuẩn
- Tương tự cũng có song trực khuẩn, tứ trực khuẩn
Hình 3: Một số dạng trực khuẩn
1.2.Kích thước vi khuẩn
Đơn vị đo kích thước của vi khuẩn là µm
- Kích thước của vi khuẩn khoảng 1- 10µm
- Kích thước tế bào vi khuẩn = 1/10 TB nhân thực
vi khuẩn thường gặp
2. Cấu tạo và chức năng các thành phần của tế bào vi khuẩn
Tế bào vi khuẩn có các thành phần: thành tế bào, màng sinh chất, tế bào chất và các bào quan chưa có màng bao bọc. Ở một số vi khuẩn còn có thêm màng nhày (màng giáp) và lông roi.
2.1. Thành tế bào (Cell wall)
Thành tế bào còn gọi là vách tế bào, chiếm 10-40% trọng lượng khô của tế bào, độ dày thành tế bào vi khuẩn Gram âm là 10nm, Gram dương là 14-18nm. Thành tế bào là lớp cấu trúc ngoài cùng, có độ rắn chắc nhất định để duy trì hình dạng tế bào, có khả năng bảo vệ tế bào đối với một số điều kiện bất lợi. Nồng độ đường muối bên trong tế bào thường cao hơn bên ngoài tế bào (áp suất thẩm thấu tương đương với dung dịch glucose 10-20%) do đó tế bào hấp thu khá nhiều nước từ bên ngoài vào.
Nếu không có thành tế bào vững chắc thì tế bào sẽ bị phá vỡ. Khi thực hiện co nguyên sinh rồi quan sát dưới kính hiển vi, thấy rõ lớp thành tế bào. Quan sát dưới kính hiển vi điện tử thấy rõ hơn.
- Chức năng :
+ Thành tế bào giúp duy trì hình thái của tế bào
+ Hỗ trợ sự chuyển động của tiên mao
+ Giúp tế bào đề kháng với áp suất thẩm thấu, hỗ trợ quá trình phân cắt tế bào
+ Cản trở sự xâm nhập của một số chất có phân tử lớn,
+ Liên quan đến tính kháng nguyên, tính gây bệnh, tính mẫn cảm với
thực khuẩn thể
*So sánh thành tế bào vi khuẩn Gram dương và Gram âm
VK Gram dương VK Gram âm
Đại diện:
- Tụ cầu (Staphylococcus): Tụ cầu vàng (S. aureus), Tụ cầu da (S. epidermidis), ….
- Liên cầu (Streptococcus): S. pyogenes, S. viridans, S. pneumoniae,…
- Liên cầu đường ruột (Enterococcus)
- Peptococcus và Peptostreptococcus
- Lactobacillus,…
- Clostridium tetani,…
- Trực khuẩn mủ xanh (Pseudomonas aeruginosa):
- Bordetela pertusis: Gây bệnh ho gà.:
- Bacteroides: Gây bệnh áp xe đường tiêu hóa và ngoài đường tiêu hóa.
- Escherichia coli: Nhiễm khuẩn đường tiêu hóa và ngoài đường tiêu hóa như đường niệu và đường máu…
- Thành tế bào vi khuẩn Gram dương chứa peptidoglican đầy đủ 4 lớp (chiếm >50% trọng lượng khô của thành).
+ Peptidoglycan là loại polyme xốp, khá bền vững, cấu tạo bởi 3 thành phần:
• N-Acetylglucosamin
• Acid N-Acetylmuramic
• Galactozamin
+ Ngoài ra còn thấy thành phần acid teichoic gắn với PG hay màng tế bào..
- Thành tế bào Gram âm có lớp peptidoglican mỏng và một lớp màng ngoài
- Màng ngoài : lipopolisaccarit (LPS): gồm 3 thành phần:
+ LipitA : 2 phân tử N acetyl glucozamin, 5 chuỗi dài axit béo: lipit A là nội độc tố của vi khuẩn, gây sốt, tiêu chảy, phá hủy hồng cầu…
+ Polisaccarit lõi
+ Kháng nguyên O : phần polisccarit vươn khỏi màng vào môi trường: quyết định nhiều đặc tính huyết thanh của các vi khuẩn có chứa 1 LPS và vị trí gắn thụ thể của thể thực khuẩn
- Màng ngoài còn có thể có 1 số loại prôtein: prôtein cơ chất: ví dụ porin (protein lỗ) ở E. coli, protein màng ngoài có năng lực vận chuyển chuyên biệt các phân tử lớn và lipoprotein : liên kết giữa lớp peptiđôglican bên trong với màng ngoài
- Thành Gram dương có axit teichoicpolime của ribitol và glixerol photphat: vận chuyển các ion dương vào ra tế bào, giúp tế bào dự trữ photphat. có liên quan đến kháng nguyên bề mặt và tính gây bệnh của 1 số vk gram dương.
- Không có axit teichoic
- Có thể không có khoang chu chất - Có khoang chu chất Tỷ lệ % đối với khối lượng khô của thành tế bào
Thành phần Gram dương Gram âm
Peptidoglycan 30 – 95 5 – 20
Axit teicoic (Teichoic acid) Cao 0
Lipid Hầu như không có 20
Protein Không có hoặc có ít Cao
Hình 4: Thành tế bào vi khuẩn Gram dương và Gram âm
*Peptidoglican
Thành tế bào vi khuẩn được cấu tạo từ các đơn phân murein (còn gọi là peptidoglycan hay glucopeptit). Mỗi đơn phân murein bao gồm: N- acetyl glucozamin (G), N- acetyl muramic (M), Alanin, D- glutamic, Di – aninoaxit, với tỉ lệ là: G: M:
Ala: D – glu: Diamin = 1: 1: 2: 1: 1
Mô hình đơn phân murein:
Mối liên kết giữa các chất trong đơn phân: G và M
liên kết với nhau bởi liên kết 1-4 β glucozit. Các axit amin liên kết với nhau và liên kết với M bởi liên kết pettit. Các đơn phân liên kết với nhau để tạo lớp thành vững chắc.
Giữa M của đơn phân này với G của đơn phân kế tiếp cũng liên kết với nhau bởi liên kết 1-4 β glucozit tạo thành liên kết chuỗi dọc của thành tế bào.
Liên kết 1-4 β glucozit bị enzim lyzozim cắt đứt, do đó thành tế bào bị thuỷ phân bởi lyzozim.
Giữa các đơn phân trên hai chuỗi kế tiếp nhau cũng liên kết với nhau tạo thành liên kết ngang của thành tế bào.
-Thành tế bào vi khuẩn có peptidoglican (murein) được cấu tạo từ N- acetyl glucozamin (G), N- acetyl muramic (M) liên kết với nhau bằng liên kết 1-4 β glucozit.

- Enzyme lyzozyme có ái lực rất mạnh với liên kết 1-4 β glucozit. Dưới tác dụng của lyzozyme và trong dung dịch đẳng trương, vi khuẩn G+ bị mất thành, biến thành tế bào trần (Protoplast – có 1 lớp màng); vi khuẩn G- biến thành thể hình cầu (Spheroplast – có 2 lớp màng).
- Kháng sinh nhóm β-lactam ngăn cản sự tổng hợp thành bằng cách ngăn cản sự hình thành mạch peptide => vi khuẩn không phân chia được.
- Các vi khuẩn tiêu giảm thành tế bào có thể thay đổi được hình dạng.
*Khoang chu chất - Không gian chứa chất:
- Chứa các protein tham gia vào sự thu nhận chất dinh dưỡng: vd các enzim proteinaza, nucleaza,các protein vận chuyển qua màng, protein thụ thể (làm hỗ bám của thể thực khuẩn).
- Các vi khuẩn phản nitrát hóa và hóa tự dưỡng vô cơ thường chứa các protein của chuỗi vận chuyển điện tử
- Chứa các enzim tham gia vào sự tổng hợp peptiđôglican và cải biến các hợp chất độc tố có thể gây hại cho tế bào.
Các vi khuẩn gram dương có thể không chứa 1 khoang chu chất rõ rệt chúng tiết ra các enzim ngoại bào (giống với enzim chứa trong khoang chu chất của VK gram âm).
2.2. Các thành phần bên ngoài thành tế bào
2.2.1. Vỏ nhầy
Một số loài vi khuẩn bên ngoài thành tế bào còn có chứa một lớp bao nhầy hay còn gọi là lớp giáp mô (vỏ nhầy, màng nhầy). Đó là một lớp vật chất dạng keo, có độ dày bất định.
Vỏ nhầy có hai loại, vỏ nhầy lớn và vỏ nhầy nhỏ. Vỏ nhầy lớn có chiều dày lớn hơn 0,2 µm nên thấy được dưới kính hiển vi quang học. Còn vỏ nhầy nhỏ có chiều dày dưới 0,2 µm, chỉ quan sát được qua kính hiển vi điện tử.
- Thành phần chủ yếu là nước , polysaccarid, ngoài ra cũng có polypeptid và protein.

- Chức năng chính: bảo vệ Ngoài ra còn có chức năng:
Ở vi khuẩn Gram âm hai đơn phân liên kết trực tiếp ở Diamin và D- ala. Ở vi khuẩn Gram dương hai đơn phân liên kết ở Diamin và D- ala thông qua cầu nối pentaglyxin. 140
+ Cung cấp chất dinh dưỡng cho vi khuẩn khi thiếu thức ăn
+ Là nơi tích luỹ một số sản phẩm trao đổi chất (dextran, xantan...)
+ Giúp vi khuẩn bám vào giá thể
2.2.2. Lông nhung và roi:
Một số loại vi khuẩn có khả năng di động một cách chủ động nhờ những cơ quan đặc biệt gọi là tiên mao (flagella từ tiếng La Tinh có nghĩa là cái roi) hay còn gọi là roi (tricha, trichos).
Vi khuẩn có thể có roi hoặc không có roi tùy từng chi. Roi là những sợi nguyên sinh chất rất mảnh, rộng khoảng 0,01-0,05 µm, cấu tạo từ các sợi protein bện xoắn vào nhau. Nhiệm vụ chính của roi là giúp cho vi khuẩn di dộng một cách chủ động.
Lông nhung
Ngoài roi, một số vi khuẩn còn có sợi pili, đó là những sợi lông rất ngắn và mảnh khoảng 0,3-1µm, đường kính khoảng 0,01 µm và thường có khoảng 100-400 sợi trên một tế bào. Lông không phải là cơ quan di động mà là phương tiện giúp cho vi khuẩn bám được tốt trên bề mặt của cơ chất. Lông còn có thể tham gia vào quá trình dinh dưỡng của vi khuẩn, giúp cho bề mặt tế bào hấp thu chất dinh dưỡng lên rất nhiều lần.
Ngoài ra ở một số vi khuẩn có một số sợi lông (nhung mao) sinh dục có nhiệm vụ tiếp nhận các đoạn ADN từ bên ngoài vào, trong trường hợp có trao đổi tín hiệu di truyền, nhất là trong lúc hai vi khuẩn tiếp hợp nhau. Lông còn là chỗ bám cho thực khuẩn thể. Số lượng lông thông thường có hàng trăm nhưng lông sinh dục thì chỉ có 15 mà thôi.
2.3. Màng sinh chất
2.3.1. Cấu tạo Màng sinh chất còn được gọi là màng tế bào hay màng chất (Cytoplasmic membrane), viết tắt là CM, dày khoảng 7-8 nm.
Có cấu tạo gồm protein (chiếm hơn 50% trọng lượng khô của màng và 10-20% protein tế bào) và một lớp kép photpholipit (20-3% trọng lượng khô của màng) nằm ở giữa. Sự phân bố protein và photpholipit ở màng sinh chất khác nhau ở từng vùng. Sự phân bố đó tạo nên các lỗ hổng trên màng thuận lợi cho sự vận chuyển.
Protein của màng gồm có hai dạng: protein cấu trúcvà enzyme. Enzyme gồm có: permeaza vận chuyển các chất vào tế bào và các enzyme tổng hợp các chất murein, photpholipit, LPS,...
2.3.2. Chức năng
+ Trao đổi chất với môi trường một cách chọn lọc.
+ Chứa các enzyme tham gia tổng hợp lipid của màng và các hợp chất cấu tạo thành tế bào (protein, peptidoglican, axit teicoic, lipopolisaccharide – LPS).
+ Là nơi định vị của nhiều enzyme tham gia tổng hợp ATP, các chuỗi hô hấp của màng sinh chất của vi khuẩn (chức năng tương tự như ở màng trong ty thể).
+ Khi màng sinh chất gấp nếp để biến thành mezoxom thì nó là nơi định vị của ADN – điểm khởi đầu của quá trình nhân đôi ADN vòng và hình thành vách ngăn trong phân bào vô tơ thành tế bào con.
+ Sự gấp nếp màng tạo thành các túi (hình cầu hoặc bản dẹt) – chlososomechứa sắc tố quang hợp còn thấy ở vi khuẩn lưu huỳnh màu tía.
+ Ở các vi khuẩn cố định đạm: hệ enzyme nitrogenase bị ức chế bởi oxi, do đó chúng được bảo vệ trong các nếp gấp của màng sinh chất.
+ Ở các vi khuẩn nitrat hóa, màng sinh chất nếp gấp nhiều làm tăng bề mặt tiếp xúc của các enzyme thực hiện quá trình này.
2.4. Tế bào chất
Tế bào chất toàn bộ phần nằm trong màng tế bào trừ nhân. Đây là vùng dịch thể dạng keo đồng nhất khi tế bào non và có cấu trúc lổn nhổn khi tế bào già. Nguyên sinh chất có hai bộ phận chính:
Cơ chất tương bào: chủ yếu chứa các enzyme.
Các cơ quan con: mesosom, ribosom, không bào, hạt sắc tố, chất dự trữ
2.4.1. Mesosom
Là thể hình cầu giống như cái bong bóng, mesosom có đường kính khoảng 250nm, gồm nhiều lớp bện chặt với nhau, nằm sát vách tế bào chỉ xuất hiện khi tế bào phân chia. Hình thành vách ngăn tế bào trong phân bào và là trung tâm hô hấp của tế bào vi khuẩn hiếu khí.
Trong mesosom có nhiều hệ thống men vận chuyển điện tử.
2.4.2. Ribosom
Ribosom có đường kính 15-20nm, hằng số lắng 70S (tiểu thể lớn 50S và tiểu thể nhỏ 30S). Ribosom chứa 40-60% ARN và 35-60% protein và một ít lipid, một số enzime như ribonucleaza, ... và một ít chất khoáng. Khi thủy phân tiểu đơn vị 50S sẽ được các rARN 5S và 23S, còn thủy phân tiểu đơn vị 30S sẽ được rARN 16S (S là đơn vị Svedberg biểu thị tỉ trọng của một chất thông qua tốc độ lắng khi li tâm siêu tốc).
Phần protein của ribosom làm thành một mạng lưới bao quanh phần ARN. Trong tế bào vi khuẩn phần lớn ribosom nằm tự do trong tế bào chất, phần ít bám trên màng nguyên sinh chất.
Ribosom của vi khuẩn khác với ribosom ở người, vì thế nếu một chất kháng sinh tác động vào ribosom của vi khuẩn để ngăn cản tổng hợp protein thì không gây hại cho người.
Ribosom là trung tâm tổng hợp protein của tế bào. Số ribosom tham gia vào quá trình này chiếm khoảng 5-10% và được liên kết với nhau gọi là polisom hay polyribosom.
2.4.3. Các hạt dự trữ
Trong nguyên sinh chất thấy xuất hiện các hạt có hình dạng, kích thước và thành phần hóa học khác nhau. Sự xuất hiện của các hạt này không thường xuyên, phụ thuộc vào điều kiện môi trường và giai đoạn phát triển của tế bào. Chúng có hình dạng và kích thước khác nhau và sự có mặt của chúng không ổn định, phụ thuộc vào điều kiện ngoại cảnh. Nhiều loại hạt được vi sinh vật sử dụng như các chất dự trữ. Chúng thường được hình thành khi tế bào tổng hợp thừa các chất đó và được sử dụng khi thiếu thức ăn.
Các hạt hydrat carbon: các hạt này chứa tinh bột hoặc glycogen hoặc các chất tương tự nằm trong tế bào chất như những chất dự trữ. Khi thiếu thức ăn vi khuẩn sẽ lấy các hạt này làm nguồn năng lượng hoặc nguồn thức ăn carbon.
Hạt volutin: đây là các chất dị nhiễm sắc (bắt màu đỏ khi nhuộm xanh metylen trong khi tế bào chất bắt màu xạnh). Trừ một số vi khuẩn (như Corynebacbacterium, Mycobacterium,...) thường xuyên chứa các hạt volutin ở giai đoạn sinh trưởng cuối, còn thông thường vi sinh vật chỉ chứa hạt volutin trong điều kiện dinh dưỡng bất thường (thiếu chất nào đó). Volutin là một phức chất, cấu tạo bởi polyphosphat, lipoprotein, ARN và Mg+2. Trong số các hạt volutin có thể có là giọt mỡ, xuất hiện nhiều khi nuôi cấy vi khuẩn trên môi trường chứa nhiều đường, glycerin hoặc các hợp chất carbon đễ đồng hóa khác.
Giọt lưu huỳnh: Một số vi khuẩn ưu lưu huỳnh, có chứa thường xuyên các giọt lưu huỳnh trong tế bào, do kết quả oxy hóa H2S sinh ra. Giọt lưu huỳnh được dùng làm nguồn năng lượng khi đã sử dụng hết H2S của môi trường xung quanh.
H2S + 1/2O2→ S + H2O + Q1
2S + 3O2 + H2O → 2H2SO4 + Q2
Tinh thể diệt côn trùng: Trong một vài vi khuẩn có thể chứa thêm một tinh thể đặc biệt. Vi khuẩn Bacillus thuringiensis hoặc B. dendrolimus,... Các tinh thể đặc biệt này có khả năng giết hại một số côn trùng phá hoại mùa màng. Hiện nay, người ta sử dụng các vi khuẩn này trong công tác bảo vệ thực vật để chống lại một số sâu trong nông nghiệp. Ví dụ sản phẩm BT tức là vi khuẩn B. thuringiensis có khả năng giết sâu tơ trên cải bắp.
Không bào khí: Các vi khuẩn quang hợp thủy sinh không có tiên mao, chứa các không bào khí trong tế bào chất. Chúng được bao bọc bởi một lớp màng protein dày khoảng 2 nm, không bào khí đóng vai trò điều tiết tỷ trọng để tế bào nổi lên những tầng nước thích hợp. Thường gặp ở các chi vi khuẩn như Halobacterium, Penlodictyon, Rhodopseudomonas, các chi vi khuẩn lam như: Anabaena.
2.5. Thể nhân (Vùng nhân -nuclear body)
Nhân là một cấu trúc ADN kép, xoắn lại khép kín thành hình cầu, hình que, hình quả tạ hay hình chữ V. Nhân tiếp xúc trực tiếp với nguyên sinh chất, liên kết với protein HU (tương tự histon) và gắn vào màng tế bào. Nhân phân chia đơn giản bằng
cách thắt lại không có sự gián phân bởi vì nhân vi khuẩn là một nhiễm sắc thể duy nhất. Tế bào vi khuẩn ở thời kỳ ổn định chỉ có một nhân nhưng trong quá trình tiền phân bào thì có từ 2 - 4 nhân.
Thể nhân là nhân nguyên thủy chưa có màng nhân, đặc trưng cho các cơ thể thuộc giới Monera (hay Prokaryota). Thể nhân còn gọi là vùng nhân, thể giống nhân, khi nhuộm bằng thuốc thử Feulgen hoặc Shiff có thể quan sát thấy thể nhân có hình dạng bất định, bắt màu tím. Thể nhân của vi khuẩn là một nhiễm sắc thể duy nhất cấu tạo bởi một sợi ADN xoắn kép, rất dài và cuộn lại thành hình vòng tròn. Nhân tế bào vi khuẩn không phân hóa thành khối rõ rệt còn gắn với màng tế bào chất. Như vậy, phần lớn các tế bào của các vi sinh vật có nhân nguyên thủy là tế bào đơn bội. NST của vi khuẩn có chiều dài thay đổi trong khoảng 0,25-3µm và chứa 6,6-13x106cặp base nitơ, số lượng hệ gen của vi khuẩn thay đổi tùy thuộc vào điều kiện nuôi cấy. Khi nuôi cấy tĩnh tế bào E. coli chứa 1-4 genom (bộ gen).
Trong NST vi khuẩn ngoài ADN còn có protein và ARN. Phần lớn protein để cấu tạo nên enzyme ARN-polymeraza. Phân tử ADN là những chuỗi xoắn kép, khép vòng dính với ARN ở giữa đầu mút của nhiều búi khác nhau.
Ngoài NST, nhiều vi khuẩn còn chứa ADN ngoài NST, đấy cũng là những sợi ADN kép, dạng vòng kín, có khả năng sao chép độc lập gọi là plasmid.
Thể nhân chứa đựng thông tin di truyền của vi khuẩn.
2.6. Nội bào tử (hoặc nha bào)
Nội bào tử là một một kết cấu do sự biến đổi của tế bào sinh dưỡng trong một giai đoạn nào đó của quá trình sinh trưởng của vi khuẩn. Mỗi tế bào chỉ có thể tạo ra một nội bào tử.
Thường gặp nội bào tử ở hai chi trực khuẩn Gram dương là Bacillus và Clotridium. Một số loài trong phẩy khuẩn (Deessulft-vibrio desulfuricans), cầu khuẩn (Sarcina ureae), xoắn khuẩn (Spirillium volutans) cũng có khả năng sinh nội bào tử
2.6.1. Cấu tạo
Dưới kính hiển vi điện tử, nội bào tử có nhiều lớp màng bao bọc, lớp ngoài cùng gọi là lớp màng ngoài của nội bào tử. Kế đó là lớp vỏ của nội bào tử gồm nhiều lớp, có tác dụng ngăn chặn sự thẩm thấu của nước và các chất hòa tan trong nước. Dưới đó là lớp màng trong và trong cùng là lớp khối tế bào chất có cấu tạo đồng nhất.
Các bức ảnh hiển vi điện tử đã chỉ ra rằng cấu trúc nội bào tử rất phức tạp. Bào tử thường được bao quanh bởi một lớp vỏ ngoài màng, mịn được gọi là vỏ ngoài (coat). Một áo bào tử (outer membrane) nằm ở bên dưới vỏ ngoài gồm một vài lớp protein và có thể khá dày. Nó không thấm nước và chịu trách nhiệm đối với tính đề kháng của bào tử đối với các hóa chất. Vỏ bào tử (cortex), có thể chiếm tới một nửa thể tích bào tử, nằm phía dưới áo bào tử. Nó gồm một lớp peptiđoglican chứa ít liên kết chéo hơn so với các tế bào dinh dưỡng. Thành tế bào của bào tử (thành của lõi –
Germ cell wall) nằm bên trong vở bào tử và bao quanh nguyên sinh chất hoặc lõi. Lõi (core) chứa các cấu trúc tế bào bình thường như riboxom và một thể nhân song không hoạt động về mặt trao đổi chất.

2.6.2. Quá trình hình thành nội bào tử
Các tế bào sinh nội bào tử khi gặp điều kiện thiếu thức ăn, hoặc có tích lũy các sản phẩm trao đổi chất có hại sẽ bắt đầu thực hiện quá trình hình nội bào tử. Về mặt hình thái học, có thể chia quá trình hình thành nội bào tử ra làm các giai đoạn:
Hình thành những búi chất nhiễm sắc.
Tế bào bắt đầu phân cắt không đối xứng, tạo ra một vùng nhỏ gọi là tiền bào tử.
Tiền bào tử hình thành hai lớp màng, tăng cao tính kháng bức xạ.
Lớp vỏ sơ khai hình thành giữa hai lớp màng của bào tử sau khi đã tích lũy nhiều PG và tổng hợp ADP, tích lũy canci, tính chiết quang cao.
Kết thúc việc hình thành áo nội bào tử
Kết thúc việc hình thành vỏ nội bào tử. Nội bào tử thành thục, bắt đầu có tính kháng nhiệt.
Bào nang vỡ ra, bào tử thoát ra ngoài.
Sự nẩy mầm của nội bào tử: Quá trình chuyển từ trạng thái nghỉ sang tế bào sinh dưỡng của vi khuẩn được gọi là quá trình nẩy mầm của nội bào tử. Quá trình này gồm 3 giai đoạn: hoạt hóa, nẩy mầm và sinh trưởng.
2.6.3. Vai trò
- Là dạng nghỉ của tế bào
- Một số vi khuẩn khi hình thành nội bào tử có thể tồn tại rất lâu trong tự nhiên.
Nội bào tử là hình thức bảo vệ vi khuẩn vượt qua điều kiện bất lợi nhờ các đặc tính
bền nhiệt, chịu hạn và tồn tại khá lâu trong chất độc.
- Khi đưa vào môi trường thuận lợi, nội bào tử nảy mầm tạo 1 tế bào vi khuẩn.
2.6.4. Nội bào tử không phải là hình thức sinh sản
- 1 tế bào vi khuẩn chỉ tạo 1 nội bào tử
- Nội bào tử khi gặp điều kiện thuận lợi, nảy mầm chỉ tạo ra 1 tế bào vi khuẩn
=> Nội bào tử là dạng nghỉ của vi khuẩn.
2.7. Không bào khí: là túi chứa đầy khí thường gặp ở nhiều loài vi khuẩn quang hợp, giúp vi khuẩn trôi lơ lửng ở nước hoặc nổi trên mặt nước.
Kết luận:
- Vi khuẩn có cấu trúc đơn giản gồm :
+ Thành tế bào và các thành phần bên ngoài có thể có
+ Màng sinh chất
+ Tế bào chất: có ribôxôm và các hạt dự trữ.
+ Vùng nhân: không có màng nhân, thường chỉ có 1 phân tử ADN vòng. Có thể có thêm plasmit.
- Có kích thước rất nhỏ có tỉ lệ S/V lớn trao đổi chất mạnh mẽ, phân chia nhanh , vận chuyển các chất trong tế bào nhanh.
3. Đặc điểm sinh trưởng của vi khuẩn trong môi trường nuôi cấy Các nhà khoa học nghiên cứu vi khuẩn cố gắng tạo ra môi trường nuôi cấy cực thuận trong phòng thí nghiệm, với nguồn năng lượng thiết yếu, các chất dinh dưỡng, pH và nhiệt độ mà khả năng sinh trưởng của vi khuẩn có thể dự đoán được.
(a) (b) (c)
Hình 5. (a) Các khuẩn lạc E. coli sinh trưởng trên đĩa thạch agar. (b) Hai dạng khuẩn lạc nhẵn và thô nhám - trên và dưới - của S. pneumoniae; và (c) phương pháp thu nhận bản sao các khuẩn lạc qua đêm sinh trưởng trên môi trường đặc hiệu: các khuẩn lạc mọc được có màu xanh và không mọc được màu trắng.

Vi khuẩn có thể được nuôi cấy trên môi trường đặc (thường chứa thạch agar) hoặc trong môi trường lỏng. Trong môi trường lỏng, vi khuẩn sinh sản theo hàm số mũ cho đến khi hết chất dinh dưỡng hoặc cho đến khi tích luỹ những sản phẩm độc hại. Số lượng vi khuẩn tồn tại ở mỗi thời điểm trong môi trường lỏng có thể xác định

được một cách dễ dàng. Dùng pipet đưa một mẫu nhỏ lên đĩa petri có môi trường đặc rồi cấy chải đều trên mặt thạch. Sau thời gian ủ 24 - 36 giờ mỗi tế bào vi khuẩn sẽ cho một cụm tế bào có thể dễ dàng nhìn thấy được bằng mắt thường gọi là khuẩn lạc (colonies; Hình 5a) chứa hàng triệu tế bào, ngay cả trong những điều kiện sinh trưởng tương đối nghèo nàn. Do đó, các thí nghiệm ở E. coli thường chỉ mất một ngày, trong khi ở ngô chẳng hạn phải mất hàng tháng.. Khả năng mọc hay không mọc của vi khuẩn trên những môi trường riêng biệt giúp ta xác định kiểu gene của tế bào vi khuẩn (Hình 5c). Các vi khuẩn thường trải qua các pha sinh trưởng trong môi trường nuôi cấy huyền phù như sau (xem Hình 6):
(i) Pha lag: Sinh trưởng thoạt đầu rất chậm, vì chúng phải làm quen với đời sống trong các điều kiện mới.
(ii) Pha log (logarithmic hay exponential): Một khi bộ máy chuyển hoá vận hành, chúng bắt đầu phân chia theo hàm số mũ, gấp đôi số lượng sau vài phút:


(iii) Pha cân bằng (stationary): Khi môi trường sống cạn kiệt, sự sinh trưởng vi khuẩn dừng lại và ổn định về số lượng. Và, cuối cùng,
(iv) Pha suy vong (death): Các sản phẩm độc do bài tiết tích luỹ có thể gây chết vi khuẩn.
được không gian và dưỡng chất không giới hạn, thì chúng sẽ sinh trưởng ở tốc độ hàm số mũ. Như chỉ ra ở Hình 7, cùng các số liệu về đường cong sinh trưởng như nhau được phác hoạ trên một thang tuyến tính (linear scale) hay thang log (log scale).
Mặc dù các tế bào có thể sinh trưởng một cách vô hạn trong một không gian nào đó, nhưng các phòng thí nghiệm thì không thể làm được những cái bình to quá cỡ. Vì vậy các tế bào thường được cho sinh trưởng trong các môi trường pha loãng. Chẳng hạn, thử hình dung 1.000 tế bào được cho vào trong một bình thí nghiệm mới và sau đó có 1.000.000 tế bào dược sinh ra trong đó. Nếu mất độ tế bào quá dày đặc, thì chúng sẽ sinh trưởng chậm lại. Để ngăn chặn sự suy giảm các tế bào, thì một phần mẫu đại diện của các tế bào (ví dụ, một mL chứa 1.000 tế bào) sẽ được chuyển sang một bình mới và sẽ sinh trưởng. Quá trình lấy một số tế bào từ bình này và cho sinh trưởng tiếp tục trong một bình mới cho đến khi nhà nghiên cứu có đủ số lượng tế bào để tiến hành thí nghiệm.
Có nhiều cách khác nhau để đo tốc độ sinh trưởng. Đôi khi, người ta bổ sung các dNTP có đánh dấu đồng vị phóng xạ (radioactive) vào môi trường nuôi cấy các tế bào. Khi các tế bào tái bản các DNA của chúng, chúng sẽ kết hợp các dNTP vào trong DNA của chúng và qua đó có thể định lượng được. Phương pháp tái bản DNA này thường được dùng để dành các mẫu mô sống hoặc các tế bào eukaryote được cho sinh trưởng trong các đĩa nuôi cấy. Các vi khuẩn, nấm men và nhiều vi sinh vật khác thường được đếm một cách trực tiếp.
Hình 6. Các tế bào vi khuẩn E. coli đang phân chia (bên trái), và đường cong sinh trưởng của vi khuẩn trong dịch huyền phù (bên phải; trục tung biểu thị số tế bào tăng theo hàm số mũ, và trục hoành biểu thị thời gian).
Đã nhiều lần các nhà nghiên cứu muốn xác định xem bằng cách nào các tế
bào phân chia hay sinh trưởng một cách nhanh chóng như vậy. Một phương pháp
đơn giản để đo tốc độ sinh trưởng là đếm số tế bào trong một đơn vị thể tích nhỏ (aliquot) tại nhiều thời điểm và biểu diễn một đường cong sinh trưởng (growth curve; Hình 7). Đó là một đường cong không quá cong lắm như như đồ thị về số lượng tế bào (number of cells) đếm được tại các thời điểm khác nhau. Nếu như các tế bào có

Hình 7: Các đường cong sinh trưởng. Đồ thị bên trái cho thấy tốc độ sinh trưởng được biểu thị bằng các chấm trên một thang tuyến tính. Đồ thị bên phải cho thấy cùng số liệu đó trên thang logarith.
4. Sinh sản của vi khuẩn
Vi khuẩn sinh sản bằng cách chia đôi (binary fission) hay trực phân (amitosis). Mặc dù không có hình thức sinh sản hữu tính (chỉ là sinh sản cận hữu tính, parasexual reproduction), các biến đổi di truyền vẫn xảy ra trong từng tế bào vi khuẩn thông qua các hoạt động tái tổ hợp di truyền.

Có ba kiểu tái tổ hợp di truyền đã được phát hiện ở vi khuẩn:
+ Biến nạp (transformation): chuyển DNA trần từ một tế bào vi khuẩn sang tế bào khác thông qua môi trường lỏng bên ngoài, hiện tượng này gồm cả vi khuẩn chết.
+ Tải nạp (transduction): chuyển DNA vi khuẩn từ tế bào sang tế bào khác thông qua thể thực khuẩn (bacteriophage).
+ Tiếp hợp (conjugation): chuyển DNA từ vi khuẩn này sang vi khuẩn khác thông qua ống tiếp hợp hay lông giới tính (pilus). Sau khi nhận được DNA từ một trong những kiểu trao đổi thông tin di truyền nói trên, vi khuẩn sẽ tiến hành phân chia và truyền bộ gene tái tổ hợp cho thế hệ sau.
5. Các quá trình trao đổi chất ở vi khuẩn Có rất nhiều kiểu trao đổi chất khác nhau ở vi khuẩn. Vi khuẩn dị dưỡng (heterotroph) phải dựa vào nguồn carbon hữu cơ bên ngoài, trong khi các vi khuẩn tự dưỡng (autotroph) có khả năng tổng hợp chất hữu cơ từ CO2 và nước. Các vi khuẩn tự dưỡng thu nhận năng lượng từ phản ứng oxy-hóa các hợp chất hóa học gọi là vi khuẩn hóa dưỡng (chemotroph), và những nhóm thu năng lượng từ ánh sáng thông qua quá trình quang hợp được gọi là vi khuẩn quang dưỡng (phototroph). Ngoài ra, các vi khuẩn còn được phân biệt nhờ vào nguồn chất khử mà chúng sử dụng. Những nhóm sử dụng hợp chất vô cơ (như nước, khí hiđrô, sulfua và ammoniac) làm chất khử được gọi là vi khuẩn vô cơ dưỡng (lithotroph) và những nhóm cần hợp chất hữu cơ (như đường, acid hữu cơ) gọi là vi khuẩn hữu cơ dưỡng (organotroph). Những kiểu trao đổi chất dựa vào nguồn năng lượng (quang dưỡng hay hóa dưỡng), nguồn chất khử (vô cơ dưỡng hay hữu cơ dưỡng) và nguồn carbon (tự dưỡng hay dị dưỡng) có thể được kết hợp khác nhau trong từng tế bào, và nhiều loài có thể thường xuyên chuyển từ kiểu trao đổi chất này sang kiểu trao đổi chất khác.
Những chất dinh dưỡng cần thiết cho sự phát triển bình thường gồm nitơ, lưu huỳnh, phospho, vitamin và các nguyên tố kim loại như natri, kali, canxi, ma-nhê, mangan, sắt, kẽm, côban, đồng, nikel... Một số loài cần thêm một số nguyên tố vết khác như tungsten, vanađi hay bo. Vi khuẩn quang vô cơ tự dưỡng bao gồm vi khuẩn lam (cyanobacteria) là một trong những loài cổ nhất được biết đến từ hóa thạch và có lẽ đã đóng một vai trò quang trọng trong việc tạo ra nguồn oxy cho khí quyển. Chúng là những tiên phong trong việc sử dụng nước như là nguồn electron vô cơ (lithotrophic) và là sinh vật đầu tiên dùng bộ máy quang hợp để phân rã nước. Các vi khuẩn quang hợp khác dùng các nguồn electron khác nên không tạo ra oxy.
Dựa vào phản ứng với oxy, hầu hết các vi khuẩn có thể được xếp vào 3 nhóm: một số chỉ có thể mọc khi có oxy được gọi là vi khuẩn hiếu khí (aerobe); một số khác chỉ có thể mọc khi không có oxy được - vi khuẩn kị khí (anaerobe); và một số có thể mọc cả khi có hay không có oxy thì thuộc nhóm vi khuẩn kị khí tùy ý (facultative anaerobe). Các vi khuẩn không sử dụng oxy nhưng vẫn có thể mọc khi có ôxy - vi khuẩn chịu oxy (aerotolerant). Những vi khuẩn có thể mọc tốt trong môi trường khắc nghiệt đối với con người được gọi là extremophile. Một số vi khuẩn sống trong suối nước nóng - vi khuẩn chịu nhiệt (thermophile); một số khác sống trong hồ nước rất
mặn - vi khuẩn chịu mặn (halophile); trong khi đó có loài lại sống trong môi trường acid hay kiềm - vi khuẩn chịu axit (acidophile) hay vi khuẩn chịu kiềm (alkaliphile) và
còn một số sống dưới lớp băng hà trong dãy núi Alpes - vi khuẩn chịu hàn (psychrophile).
6. Di động
Vi khuẩn di động nhờ vào tiên mao (flagellum), trượt (bacterial gliding) hay thay đổi sức nổi (buoyancy). Nhóm xoắn khuẩn (spirochaete) có các cấu trúc tương tự tiên mao gọi là sợi trục (axial filament). Chúng có một thể xoắn ốc đặc biệt quay tròn khi di chuyển.
Tiên mao của vi khuẩn được sắp xếp theo nhiều cách. Vi khuẩn có thể có một tiên mao ở mỗi cực của tế bào, hay có thể có một nhóm nhiều tiên mao ở một đầu. Nhiều vi khuẩn (như E. coli) có hai kiểu di động khác nhau: di động tiến tới (bơi) và quay vòng.
Vi khuẩn di động khi bị thu hút hay đẩy ra bởi một số tác nhân kích thích, hoạt động này được gọi là tính hướng động (taxes), chẳng hạn như: hóa hướng động (chemotaxis), quang hướng động (phototaxis), cơ hướng động (mechanotaxis) và từ hướng động (magnetotaxis).
150
CHƯƠNG III: DI TRUYỀN HỌC VI KHUẨN
1. Đặc điểm và nhân tố di truyền của vi khuẩn
1.1. Đặc điểm trong nghiên cứu di truyền ở vi khuẩn
1. Thời gian thế hệ ngắn, tốc độ sinh sản nhanh
2. Có sự tăng nhanh số lượng cá thể
3. Có cấu tạo bộ máy di truyền đơn giản
4. Dễ nghiên cứu bằng các kỹ thuật vật lý và hóa học
1.2. Nhân tố di truyền của vi khuẩn
Acid nucleic là cơ sở vật chất di truyền của tất cả các dạng sinh vật. Ở tất cả các sinh vật nhân sơ (prokaryote, còn gọi là sinh vật nhân sơ sơ, bao gồm các vi khuẩn) hay nhân chuẩn (eukaryote, còn gọi là sinh vật nhân chuẩn), trừ virus và các yếu tố sinh học đơn giản hơn như viroid và prion, tính trạng được mã hóa và tồn trữ dưới dạng mã hóa là trình tự thẳng của các nucleotide trong thành phần acid deoxyribonucleic (ADN). Vật chất di truyền này được thể hiện thành tính trạng của cá thể thông qua quá trình tổng hợp từ khuôn ADN thành phân tử ARN thông tin (quá trình phiên mã) và sau đó ARN thông tin này lại làm khuôn để tổng hợp protein (cấu trúc, enzyme, thụ thể, kích thích, kìm hãm,...) trong quá trình gọi là dịch mã (transcription) dẫn đến biểu hiện tính trạng. Vật chất di truyền được truyền từ thế hệ tế bào này sang thế hệ tế bào khác (và từ thế hệ này sang thế hệ khác) nhờ quá trình tự sao (replication) của phân tử ADN. Quy tắc này được gọi là quy tắc trung tâm biểu hiện di truyền.
Ở vi khuẩn DNA có thể gặp ở hai dạng: ADN nhiễm sắc thể và ADN plasmid
* ADN nhiễm sắc thể
Nhân (vùng nhân) vi khuẩn là một nhiễm sắc thể vi khuẩn có cấu tạo rất đơn giản: Kích thước nhỏ, toàn bộ genome của vi khuẩn là một phân tử ADN ở dạng vòng, mạch kép và không có màng nhân bao bọc. Các gen đều tồn tại đơn bản và ở trạng thái đơn bội. Các đột biến điểm phát sinh đều có thể được biểu hiện ngay ra kiểu hình. Vì vậy, vi khuẩn sinh sản rất nhanh đồng thời liên tục biến đổi thành những chủng loại mới để có thể thích nghi được với những điều kiện môi trường khác nhau và thường xuyên thay đổi trong quá trình tiến hóa.
Hình 8. Genome của vi khuẩn E.coli.

- Toàn bộ hệ gen của vi khuẩn chỉ chứa các đoạn ADN không lặp lại, và hầu hết các đoạn trình tự nuclêôtit này đều được tham gia mã hóa cho các prôtêin cho tế bào.
- Genome của vi khuẩn chỉ chứa một số ít các gen, trong đó các phân bố sát nhau, không bị gián đoạn bởi các đoạn ADN không chứa mã di truyền (intron). Tuy nhiên, gần đây một số công trình nghiên cứu cho thấy ở nhóm đối tượng sinh vật cổ (Archae) cũng thuộc giới sinh vật nhân sơ thì có hệ gen phân mảnh, xen kẽ các đoạn có ý nghĩa mã hóa (exôn) có các đoạn trình tự nuclêôtit không có ý nghĩa mã hóa (intron).
Ở sinh vật nhân sơ, mỗi gen cấu trúc được chia ra làm hai phần, gồm có vùng điều khiển hoạt động của gen và vùng mang thông tin di truyền.
Vùng điều khiển hoạt động của gen là một đoạn trình tự nuclêôtit nằm trước các đoạn gen mang thông tin mã hóa cho các phân tử prôtêin, bắt đầu từ -1, gồm các vị trí:
- Promoter: Promoter là trình tự nhận biết của enzyme ARN polymeraza và là nơi mà enzyme ARN polymeraza gắn vào để xác định vị trí bắt đầu phiên mã. Do vậy nên promoter có những cấu trúc đặc hiệu giúp enzyme nhận biết chính xác. Khảo sát nhiều promoter khác nhau của các gen, người ta nhận thấy phần tâm của promoter có những trình tự chung giống nhau và gọi là các hộp, ví dụ ở E. coli có hộp TATAAT. Hộp này thường nằm ở vị trí khoảng -10, tức là nằm ở khoảng 10 nuclêôtit phía trước vị trí khởi đầu phiên mã hay trình tự TTGACA nằm ở vị trí -35, tức là khoảng 35 nuclêôtit trước vị trí khởi đầu phiên mã. Ở vi khuẩn có một loại promoter vì chỉ có một loại ARN polymeraza.
Hình 9. Vùng trung tâm của promoter của operon Tryptophan ở vi khuẩn
- Vị trí hoạt hóa (A) hoặc vị trí ức chế (O): được nhận biết bởi các prôtêin điều khiển, chúng có thể liên kết với ADN hoặc ARN polymeraza làm tăng cường hoặc kìm hãm hoạt động của gen trong quá trình phiên mã.
Tiếp theo vùng điều khiển hoạt động của gen là vùng mang thông tin di truyền, là đoạn trình tự nuclêôtit được phiên mã từ ADN sang mARN theo chiều 5’ => 3’ trên sợi đang tổng hợp (bắt đầu từ vị trí +1). Đoạn trình tự này gồm một số vùng như sau:
- Vùng đầu 5’ và 3’ không dịch mã (5’UTR và 3’UTR): Đây là các đoạn trình tự
nuclêôtit chưa được biết rõ các chức năng một cách đầy đủ, tuy nhiên các nghiên cứu đã chỉ ra rằng chúng đóng vai trò quan trọng trong việc quy định tuổi thọ của các phân tử mARN, đồng thời tham gia vào quá trình kiểm soát phiên mã ở trong tế bào. Trong đó, vùng đầu 5’UTR không dịch mã được tính từ nuclêôtit phiên mã đầu tiên đến bộ ba nuclêôtit khởi đầu quá trình dịch mã (AUG hoặc GUG tùy thuộc vào các đối tượng sinh vật). Trình tự các nuclêôtit không dịch mã đầu 3’ (3’ UTR) được tính từ một trong ba codon dừng dịch mã (UAA, UAG, UGA) đến hết trình tự kết thúc quá trình phiên mã.
- Vùng khung đọc mở của gen: Đây là phần ADN sẽ tham gia vào quá trình mã hóa các chuỗi polipeptit trong các phân tử prôtêin, có tính liên tục vì không có sự xen kẽ giữa các đoạn exôn và các đoạn intron. Vùng khung đọc mở của gen được bắt đầu bằng một codon khởi đầu (AUG) và kết thúc bằng một trong 3 mã kết thúc là UAA/UAG/UGA. Quá trình dịch mã được đọc từ đầu 5’ -> 3’, đọc theo phân tử mARN một cách liên tục theo từng mã một, mỗi codon tương ứng với một axit amin trong chuỗi pôlipeptit đang được kéo dài, không có hiện tượng chùm và gối lên nhau (chỉ xảy ra ở vi rút) cho đến tận mã kết thúc thì dừng lại.
Các gen prokaryote thường sắp xếp nằm gần nhau và chịu sự điều khiển chung của một promoter, tức là chúng được phiên mã sang cùng một phân tử mARN. Cấu trúc này được gọi là opêrôn. Như vậy, một opêrôn gồm hai hay nhiều gen nằm cạnh nhau trên một nhiễm sắc thể. Thông thường, đó là các gen cùng tham gia vào một con đường chuyển hóa, ví dụ như các gen mã hóa cho các enzyme cần thiết cho quá trình chuyển hóa glucose. Do có chung promoter điều khiển cho mọi gen nằm trong một opêrôn cho nên chỉ có một loại phân tử mARN được tổng hợp từ một opêrôn (mang thông tin di truyền của tất cả các gen nằm trong đó). Nói cách khác, quá trình phiên mã của các gen trong một opêrôn xảy ra đồng thời và phân tử mARN đặc trưng cho opêrôn được gọi là mARN polycistron.
Hình 10: Vùng 5’UTR và 3’UTR trong gen cấu trúc.
Hình 11. Cấu trúc operon trong genome vi khuẩn. Một operon là một đơn vị phiên mã đơn bao gồm một chuỗi các gen cấu trúc, một promoter và một operator.



Tuy nhiên, điều cần ghi nhớ là quá trình dịch mã trên các phân tử mARN polycistron xảy ra hoàn toàn độc lập với nhau. Mỗi đoạn tương ứng với một gen trên phân tử này đều có vị trí bám của ribôxôm, có mã bắt đầu và kết thúc tổng hợp chuỗi polypeptit riêng biệt. Do đó, tốc độ tổng hợp các prôtêin trên các phân tử mARN polycistron hoàn toàn khác nhau.
*ADN plasmid
Bên cạnh ADN nhiễm sắc thể ở vi khuẩn còn có thể gặp cơ sở vật chất di truyền ngoài nhiễm sắc thể gọi là plasmid. Về bản chất hóa học, plasmid cũng là một phân tử
ADN xoắn kép, khép kín, nhưng có phân tử lượng thấp hơn nhiều (khoảng 106 - 108
Dalton, hay khoảng 1/100 đến 1/200 lần nhiễm sắc thể vi khuẩn). Một vi khuẩn có thể
không mang, hoặc mang một hoặc một số plasmid. Plasmid có thể phân bố trong tế bào chấtở trạng thái tự do và có thể sao chép một cách độc lập với nhiễm sắc thể hoặc nằm trong nhiễm sắc thể (tái tổ hợp), khi đó plasmid được gọi là episome. Ở trạng thái tái tổ hợp trong nhiễm sắc thể (hay episome) plasmid thường được sao chép đồng thời với nhiễm sắc thể và được truyền từ thế hệ này sang thế hệ khác của vi khuẩn.

Plasmid giới tính còn gọi là yếu tố sinh dục hay yếu tố F (fertility factor), là plasmid điều khiển sự hình thành các pili giới tính (nhung mao giới tính), sự tiếp hợp và sự vận chuyển thông tin di truyền (gen plasmid hay gen nhiễm sắc thể) một chiều từ tế bào cho sang tế bào nhận. Trong trường hợp này tế bào mang plasmid F là tế bào cho thường được gọi là tế bào đực hay tế bào F+, còn tế bào nhận được gọi là tế bào cái hay tế bào F-. Yếu tố F có thể nằm ở trạng thái tự do trong tế bào chất hay ở trạng thái liên kết với nhiễm sắc thể hay trạng thái episome. Trong trường hợp sau quá trình vận chuyển thông tin di truyền thường diễn ra với tốc độ và tần suất cao nên những chủng vi khuẩn này thường được gọi là chủng Hfr (bắt nguồn từ tiếng Anh: high frequency of recombination).
Hình 12. Bản đồ enzyme hạn chế của plasmid pBR322
Plasmid pBR322, là một plasmid được sử dụng phổ biến trong kỹ thuật ADN tái tổ hợp. Plasmid cải biến nhân tạo này mang hai gen kháng thuốc (tetR đề kháng tetracycline và ampR đề kháng ampicillin) và một loạt vị trí cắt của enzyme hạn chế trong đó mỗi enzyme này chỉ có một vị trí phân cắt, nhờ vậy plasmid vector này áp dụng được với nhiều ADN khác nhau.
Thêm hoặc mất một plasmid thường dẫn đến mất hoặc bớt một hoặc một số tính trạng, mặc dù nhiều plasmid không thể hiện tính trạng (plasmid "câm"). Tính trạng mà các plasmid điều khiển đa dạng nhưng nhìn chung plasmid không phải là yếu tố thiết yếu đối với sự sống còn của vi khuẩn. Dựa vào sự tương hợp của các sản phẩm của plasmid mà người ta chia các plasmid thành một số nhóm: plasmid giới tính, plasmid đề kháng, plasmid điều khiển sinh tổng hợp (nhóm chất nào đó),... và thường liên quan đến tính gây bệnh của các vi khuẩn. Ví dụ, đã xác định được rằng một số loài (hoặc còn được coi là dạng huyết thanh học) vi khuẩn Salmonella có được độc tính nhờ mang plasmid.
Yếu tố đề kháng, hay yếu tố kháng thuốc, hay plasmid R (resistance factor), là plasmid điều khiển sự đề kháng của vi khuẩn đối với các chất kháng sinh và thuốc chống vi khuẩn khác. Cơ chế đề kháng này là sự điều khiển việc tổng hợp các enzyme (như penicillinase,...) phân 200 hủy thuốc kháng sinh thành các sản phẩm vô hoạt hoặc ức chế sự xâm nhập của thuốc kháng sinh vào tế bào (như chặn các lỗ khổng trên màng tế bào chất đối với tetracycline hoặc thuốc kháng sinh họ macrolide,...). Yếu tố R lan truyền được (transmissible R factor) là yếu tố F kháng thuốc, gồm hai miền gen. Một miền gen điều khiển sự đề kháng. Số lượng yếu tố bị đề kháng có thể là 1 đến 10 hoặc nhiều hơn nữa, trường hợp sau thường gọi là plasmid đề kháng đồng loạt. Miền gen thứ hai (miền RTF) là gen điều khiển sự vận chuyển các plasmid vào tế bào khác. Thông thường sự vận chuyển các plasmid này có tốc độ cao, nhưng yếu tố vận chuyển cũng có thể tự ức chế hoặc bị ức chế bởi yếu tố khác. Yếu tố R không lan truyền được (non-transmissible R factor) chỉ mang miền gen điều khiển tính đề kháng. Các plasmid này không tự vận chuyển sang tế bào khác mà chỉ làm cho tế bào mang chúng có tính đề kháng thuốc kháng sinh. Chúng có thể tồn tại và phổ biến trong quần thể vi khuẩn nhờ quá trình sinh sản của vi khuẩn hoặc nhờ các quá trình vận chuyển vật chất di truyền qua dịch thể tức quá trình biến nạp hoặc sự vận chuyển của virus của vi khuẩn (phage, hay thực khuẩn thể) tức nhờ quá trình tải nạp.
Plasmid thường chứa một số gen có tính đặc thù cao và có ý nghĩa rất quan trọng giúp cho vi khuẩn có thêm những đặc tính khác biệt như gen kháng kháng sinh giúp chúng sinh trưởng và phát triển tốt trong điều kiện môi trường có các chất kháng sinh diệt khuẩn , gen giúp cho vi sinh vật có khả năng tuyển chọn được khoáng trên môi trường tuyển chọn, một số gen chỉ thị màu,… Bên cạnh đó, các vi khuẩn có thể truyền cho nhau những gen đặc biệt này thông qua cầu tiếp hợp giúp cho các quần thể vi khuẩn có thể tồn tại và chống chọi tốt hơn với những điều kiện môi trường xung quanh, đồng thời gây nên những khó khăn nhất định cho con người trong công cuộc chống lại các bệnh truyền nhiễm do vi sinh vật gây ra.
1.3. Các thể đột biến của vi khuẩn Để phân tích di truyền ở vi khuẩn thường dùng các thể đột biến sau :
(i) Đột biến khuyết dưỡng (auxotroph): Các thể đột biến không có khả năng tổng hợp chất cần thiết như kiểu dại (hay thể nguyên dưỡng, prototroph) và do đó, không sinh trưởng được nếu không thêm vào môi trường chất dinh dưỡng đó. Ví dụ, thể đột biến khuyết dưỡng methionin không sống được trên môi trường chỉ chứa muối vô cơ (môi trường tối thiểu, minimal medium) nhưng nếu thêm methionin vào thì nó sống được.
(ii) Đột biến kháng chất kháng sinh: Những đột biến này có thể sinh trưởng được khi có chất kháng sinh trong môi trường, như streptomycin (str) hay tetracyclin (tet). Ví dụ, tế bào mẫn cảm với streptomycin (Strs)là kiểu dại và không mọc trên môi trường có streptomycin nhưng những thể đột biến kháng streptomycin (Strr) lại mọc được.
(iii) Đột biến nguồn carbon: Các thể đột biến này không sử dụng được một cơ chất nào đó làm nguồn năng lượng hay nguồn cung cấp carbon. Ví dụ, thể đột biến lackhông sử dụng đường lactose.
Môi trường mà trên đó mọi vi khuẩn đều mọc được được gọi là môi trường không chọn lọc. Nếu môi trường chỉ cho một kiểu tế bào mọc được, thì gọi là môi trường chọn lọc. Ví dụ, môi trường chứa streptomycin là chọn lọc cho thể đột biến Strr và môi trường tối thiểu chứa lactose là chọn lọc cho Lac+. Để lai vi khuẩn người ta trộn hai thể đột biến khuyết dưỡng khác nhau với nhau, chẳng hạn a b c d+ e+ f+ và a+ b+ c+ d e f, rồi đem cấy hỗn hợp lai lên môi trường tối thiểu, các tế bào nào mọc được trên môi trường này chính là các tế bào lai nguyên dưỡng (a+ b+ c+ d+ e+ f+).
1.4. Kiểu hình và kiểu gene của vi khuẩn
Các đột biến có thể tác động đến 5 loại kiểu hình:
– Biến đổi từ nguyên dưỡng (prototrophy) sang khuyết dưỡng (auxotrophy) và ngược lại.
Ví dụ: mất khả năng tổng hợp một chất trao đổi của chu trình và hồi biến để lại có khả năng tổng hợp chất đó.
– Sự mất hay có được khả năng sử dụng chất dinh dưỡng khác. Ví dụ: đột biến làm mất khả năng sử dụng đường lactose.
– Tính nhạy cảm hay đề kháng thuốc như nhạy cảm streptomycin đột biến thành kháng streptomycin.
– Nhạy cảm với phage thành kháng phage hoặc ngược lại. Ví dụ: đột biến trên thụ thể ở bề mặt tế bào làm đề kháng với sự nhiễm phage.
– Sự mất đi hoặc có lại các thành phần cấu trúc của bề mặt tế bào. Ví dụ, một loại Pneumococcus có vỏ bao polyxacarit (polyxacarit capsule), trong khi đó dòng khác không có vỏ bao.
tắc:
Các kí hiệu dùng biểu hiện kiểu hình và kiểu gen được thống nhất theo nguyên
– Kí hiệu kiểu hình gồm 3 chữ thường (chữ đầu viết hoa) với dấu phía trên góc “+” hay
“–” nhằm chỉ sự hiện diện hay thiếu tình trạng tương ứng, và “s” hay “r” tương ứng chỉ tính mẫn cảm (sensitive) hay đề kháng (resistance).
– Kí hiệu kiểu gen được viết chữ nghiêng, chữ đầu không viết hoa.
Ví dụ 1: Tế bào hoang dại tự tổng hợp được leuxin thì kiểu hình được viết Leu+ . Đột biến khuyết dưỡng mất khả năng tổng hợp leuxin được kí hiệu kiểu hình Leu .
Tương ứng với hai kiểu hình trên kí hiệu kiểu gen là leu+ hay leu và leu-
Nếu cần nhiều hơn một gen để tạo ra một chất nhất định, sau 3 chữ nghiêng kí hiệu gen có thêm chữ nghiêng hoa. Ví dụ: leuA, leuB... là các gen cần thiết cho tổng hợp leuxin khác nhau.
Ví dụ 2: Kiểu hình kháng hoặc nhạy cảm với penixilin được viết là Penr và Pens. Kiểu gen tương ứng là penr và pens .
Nếu lưỡng bội ở một phần thì viết thêm gạch nghiêng. Ví dụ, leu+ /leu.
Kiểu gen Kiểu hình Mô tả kiểu hình
lac- Lac- Không thể chuyển hoá đường lactose
mal- Mal- Không thể chuyển hoá đường maltose
ara- Ara- Không thể chuyển hoá đường arabinose
trp- Trp- Không thể tạo ra amino acid tryptophan
bio- Bio- Không thể tạo ra vitamin biotin
strr Strr Kháng được chất kháng sinh streptomycin
strs Strs Mẫn cảm với chất kháng sinh streptomycin
2. Các phương pháp nghiên cứu đặc thù của di truyền học vi khuẩn Đối với các vi khuẩn, phân tích di truyền học cũng là phương pháp duy nhất để nghiên cứu các đặc tính di truyền và biến dị của chúng. Do các vi sinh vật thường có bộ gene đơn bội, đặc biệt các vi khuẩn chỉ có một nhóm liên kết gene nên sơ đồ phân tích di truyền học ở chúng là đơn giản hơn các eukaryote bâc cao, gồm các giai đoạn sau:
(i) Xác định các gene;
(ii) Xác định trật tự của các locus trên nhiễm sắc thể; và
(iii) Xác định cấu trúc tinh vi của gene.
Tổng quát, có các phương pháp cơ bản được áp dụng cho phân tích di truyền vi khuẩn như sau: phân tích đột biến, phân tích tái tổ hợp, phân tích sao chép, phân tích đoạn khuyết và phân tích bổ sung.
2.1 . Phân tích đột biến
Phân tích đột biến được áp dụng để xác định các gene và được tiến hành bằng cách đo đếm các kết quả cuối cùng của sự biểu hiện gene thành ra sự biến đổi kiểu hình (đặc điểm hình thái, hoá sinh, kháng nguyên hoặc tính mẫn cảm đối với các tác nhân hoá học, vật lý và sinh học khác nhau) của các tế bào vi khuẩn. Việc phát
hiện một đột biến ngẫu nhiên hay gây tạo chỉ ra sự tồn tại của một gene cụ thể
Sự biến đổi hình thái ở vi khuẩn bao gồm các biến đổi về kích thước, hình dạng và sự hình thành sắc tố của các khuẩn lạc do các tế bào bị đột biến tạo nên trên các môi trường dinh dưỡng đặc cũng như sự biến đổi của bản thân các phân tử của tế bào (ví dụ sự tăng kích thước hoặc mất lông tơ trên bề mặt màng tế bào). Sự biến đổi hoá sinh bao gồm các biến đổi liên quan tới việc tế bào mất khả năng tổng hợp các amino acid và vitamin hoặc mất khả năng chuyển hoá các hợp chất hydrat carbon. Các biến đổi về kháng nguyên thể hiện ở chỗ vi khuẩn bị mất đi những kháng nguyên nhất định. Các
biến đổi trong tính bền vững của vi khuẩn đối với các tác nhân khác nhau liên quan tới sự xuất hiện trong chúng các khả năng đề kháng đối với sự chiếu xạ, với các hoá chất khác nhau (kể các các loại thuốc kháng sinh) hoặc với phage v.v.
Do tần số đột biến ở vi khuẩn là rất thấp nên việc phân lập các tế bào bị đột biến chỉ có thể thực hiện được trong các thí nghiệm với các quần thể tế bào. Như thế, về nguyên tắc, trong trường hợp này có thể sử dụng bất kỳ phương pháp nào cho phép tách được các thể đột biến từ các quần thể. Việc xác định số lượng các đột biến dựa trên các phương pháp xác định tần số đột biến. Thông thường, để phân tích di truyền cần có các nòi đột biến mang các đột biến vị trí cho trước. Chẳng hạn, đối với B. subtilis, có thể xử lý sơ bộ DNA gây biến nạp bằng các tác nhân gây đột biến; ở E. coli, có thể gây các đột biến có vị trí xác định bằng cách đưa vào tế bào vi khuẩn các gene đột biến nhờ các phage tải nạp.
2.2. Phân tích tái tổ hợp Phân tích tái tổ hợp là phương pháp đặc trưng được dùng để xác định vị trí và trật tự của các gene trên nhiễm sắc thể Đối với vi khuẩn, việc phân tích di truyền dựa vào các quá trình trao đổi vật liệu di truyền như biến nạp, tải nạp và tiếp hợp.
. Nói chung, sự trao đổi di truyền ở các vi khuẩn và quá trình hữu tính ở các cơ thể bậc cao là khá giống nhau. Việc truyền vật liệu di truyền từ vi khuẩn thể cho (donor) sang vi khuẩn thể nhận (recipient) có thể coi như như sự kết hợp nhân của các tế bào sinh dục (ở đây là sự tạo thành các thể lưỡng bội từng phần), còn sự sát nhập của vật liệu di truyền vào bộ gene của vi khuẩn thể nhận, và sự hình thành nhiễm sắc thể tái tổ hợp sau đó, có thể so sánh với các kết quả của giảm phân. Chính các hệ thống tái tổ hợp này là cơ sở cho phương pháp phân tích tái tổ hợp và lập bản đồ di truyền ở vi khuẩn. Ví dụ, trật tự của hầu hết các gene trên nhiễm sắc thể E. coli được xác định là nhờ sử dụng tiếp hợp và tải nạp; ở B. subtilis nhờ tải nạp và biến nạp; còn ở
2.3. Phân tích sao chép
Phương pháp này cho phép xác định trật tự các gene trên nhiễm sắc thể dựa trên sự tính toán các số liệu về sự bắt đầu sao chép (tái bản) của nhiễm sắc thể từ một điểm xác định. Do thời gian sao chép của một phần nhiễm sắc thể nhất định phụ thuộc vào khoảng cách từ phần đó đến khởi điểm sao chép nên thứ tự sao chép phản ảnh trình tự sắp xếp của các gene. Như vậy, bản đồ nhiễm sắc thể chỉ có thể được xây dựng dựa trên các dẫn liệu về trật tự sao chép của các phần riêng biệt của nhiễm sắc thể. 2.4. Phân tích đoạn khuyết
Phép phân tích đoạn khuyết được sử dụng để xác định vị trí của các gene trên nhiễm sắc thể cung như để nghiên cứu cấu trúc tinh vi của gene. Nó dựa trên việc tính toán các đoạn khuyết trên nhiễm sắc thể. Nhờ sự phân tích này người ta đã phát hiện được vị trí của hàng loạt gene ở E. coli và S. typhimurium, hiểu biết được cấu trúc tinh vi của các gene trên operon lactose ở E. coli. Phương pháp này cũng được sử dụng rộng rãi để nghiên cứu cấu trúc tinh vi của gene ở phage.
2.5. Phân tích bổ sung Phương pháp này được sử dụng để phát hiện chức năng của các gene nhất định tham gia vào việc xác định một đặc tính nào đó của vi khuẩn, dựa trên hiện tượng bổ sung của các gene (nghĩa là sự tương tác giữa các sản phẩm gene). Phương pháp này do Lewis tìm ra năm 1951 trong khi nghiên cứu tính allele ở ruồi giấm. Dưới đây ta hãy xem xét phép thử cistrans (đều-lệch) này qua công trình của Benzer.
Các công trình nghiên cứu của Seymour Benzer (từ 1957 đến 1961) về tái tổ hợp ở phage T4 đã cho thấy rằng, gene theo quan niệm của Morgan có thể chia nhỏ thành các đơn vị nhỏ hơn. Ông đã đưa ra các thuật ngữ muton, recon và cistron để định nghĩa các đơn vị không chia nhỏ tương ứng là đột biến, tái tổ hợp và chức năng. Bằng cách lai các thể đột biến của cùng một gene có nguồn gốc độc lập nhau trong khi cho lây nhiễm phage, đã làm xuất hiện phage kiểu dại. Điều này chỉ có thể xảy ra bởi sự tái tổ hợp bên trong gene, nếu như các phần nhỏ riêng biệt của gene đều bị đột biến. Điều này chứng tỏ rằng gene bị phân chia thành các đơn vị nhỏ hơn thông qua tái tổ hợp và dột biến. Tuy nhiên, vì kích thước của muton và recon được coi là tương đương với một cặp nucleotide, cho nên ngày nay tự thân hai đơn vị này không còn giá trị sử dụng nữa. Thuật ngữ cistron của Benzer có nghĩa là đơn vị chức năng di truyền không chia nhỏ. Điều này có thể xác định bằng sự phân tích bổ sung (complementation analysis), trong đó gene mà cụ thể là sản phẩm của nó được trắc nghiệm về khả năng bù đắp cho một đột biến tại một gene tương đồng trong cùng tế bào. Sự bổ sung liên tiếp làm phục hồi kiểu hình dại.
Salmonella typhimurium chủ yếu nhờ tải nạp. Ngoài ra, phép phân tích tái tổ hợp này còn được sử dụng để nghiên cứu cấu trúc tinh vi của gene. 160
Hình 13: Sơ đồ minh họa trắc nghiệm cis-trans: (a) con đường chuyển hóa bình thường; (b) trắc nghiệm cis; (c) và (d) trắc nghiệm trans.
Chú thích: S-cơ chất (subtrate); I- sản phẩm trung gian (intermediate); P- sản phẩm cuối cùng (product), ở đây là sắc tố đặc trưng cho kiểu hình dại; các mũi tên (↓) chỉ các enzyme sản phẩm sinh ra từ các cistron 1 và cistron 2.
Cơ sở của phân tích bổ sung là trắc nghiệm cis-trans (cis-trans test), mà từ đây nảy sinh ra thuật ngữ cistron, trong đó các cặp đột biến bắt nguồn độc lập được xét ở các cấu hình cis (đều) và trans (lệch). Trắc nghiệm cis được dùng làm đối chứng, vì nếu như cả hai đột biến đều có mặt trong một bộ gene thì bộ gene kia phải là kiểu dại ở cả hai locus và sinh ra các sản phẩm gene bình thường, do đó cho ra kiểu hình dại (hình 13b). Trắc nghiệm trans là phép thử bổ sung và xác định gới hạn của đơn vị chức năng. Nếu như các đột biến nằm trong các gene khác nhau, khi chúng có mặt ở cấu hình trans, mỗi một bộ gene có thể bổ sung sản phẩm mà gene kia không tạo ra được. Khi có đủ tất cả các sản phẩm gene cần thiết thì tế bào là kiểu dại (hình 13c), nghĩa là có sự bổ sung dương tính (positive complementation). Nếu như cả hai đột biến thuộc cùng một gene, khi chúng có mặt ở cấu hình trans, thì mỗi một bộ gene có thể mang một bản sao đột biến của gene đó và không có sản phẩm hoạt động chức năng được tạo ra trong tế bào, nghĩa là không có sự bổ sung (hình 13d). Từ các kết quả nghiên cứu của Benzer cho thấy: Cistron (hay gene cấu trúc) là một đoạn xác định của DNA mang thông tin cấu trúc của một polypeptide cụ thể mà giới hạn của nó được xác định bằng trắc nghiệm cis-trans. Theo đó, kích thước trung bình của một cistron ~1.200 cặp base.

3. Những hiện tượng di truyền của vi khuẩn
3.1. Biến nạp ở vi khuẩn (Transformation)
3.1.1. Định nghĩa, thí nghiệm và đặc điểm chung
* Định nghĩa
Biến nạp chỉ những biến đổi tính trạng của vi khuẩn dưới ảnh hưởng của ADN dung dịch được tách chiết từ vi khuẩn cho xâm nhập vào vi khuẩn nhận.
* Thí nghiệm
Hiện tượng biến nạp được Griffith phát hiện ở vi khuẩn Diplococus pneumoniae (nay gọi là Streptococus pneumoniae - phế cầu khuẩn gây sưng phổi ở động vật có vú) vào năm 1928. Phát hiện này và các nghiên cứu về cơ chế biến nạp có ý nghĩa lịch sử cho sự ra đời của Sinh học phân tử
Vi khuẩn này có 2 dạng khác nhau:
– Dạng SIII, gây bệnh có vỏ bao tế bào (capsule) bằng polysaccharid cản trở bạch cầu phá vỡ tế bào. Dạng này tạo đốm mọc (khuẩn lạc) láng (Smooth- láng) trên môi trường agar.
– Dạng RII, không gây bệnh, không có vỏ bao, tạo đốm mọc nhăn (Roughnhăn).
Thí nghiệm tiến hành như mô tả trên hình 14.
Griffith đã tiêm cho chuột một liều vi khuẩn Diplococcus pneumoniae dạng S (có màng nhày, gây bệnh viêm phôi nặng) làm cho chuột chết. Nếu xử lý bằng nhiệt thì vi khuẩn này không có khả năng gây bệnh cho chuột. Tiêm vi khuẩn dạng
Hình 14. Hiện tượng biến nạp.
R ( không có màng nhày) không gây độc đối với chuột. Nhưng khi ông tiêm cho chuột một hỗn hợp các vi khuẩn dạng R ( không có màng nhày) với vi khuẩn dạng S
(có màng nhày), nhưng đã xử lí bằng nhiệt, thì thấy chuột vẫn bị chết. Từ máu chết ông đã phân lập được Diplococcus pneumoniae dạng S điển hình. Điều đó có nghĩa các vi khuẩn dạng S bị chết vì nhiệt đã truyền khả năng tạo vỏ nhày cho các tế bào dạng R làm cho nó trở thành tế bào dạng S và tính chất này được truyền cho các thế hệ con cháu của tế bào dạng S mới (Hình 14).
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các tế bào chết này đã truyền tính gây bệnh cho tế bào R. Nó được gọi là biến nạp (transformation).Năm 1944, T.Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng proteaza (enzym phân hủy protein) hoặc ARN-aza (enzym phân hủy ARN) hoạt tính biến nạp vẫn còn, chứng tỏ protein và ARN không phải là tác nhân gây biến nạp. Nhưng nếu tế bào S chết bị xử lý bằng ADN-aza (enzym chỉ phân hủy đặc hiệu ADN) thì hoạt tính biến nạp không còn nữa, chứng tỏ ADN là nhân tố biến nạp. Kết quả thí nghiệm có thể tóm tắt như sau:
ADN của S + các tế bào R sống => chuột => chết (có R + S)
Hiện tượng biến nạp là một chứng minh sinh hóa xác nhận rằng ADN mang tín hiệu di truyền.
Như vậy, biến nạp là hiện tượng truyền thông tin di truyền bằng ADN. Trong
biến nạp, ADN trần từ một tế bào vi khuẩn (thể cho) này được truyền sang tế bào vi khuẩn khác (thể nhận). Biến nạp xảy ra khi vi khuẩn nhận ADN ngoại lai và hấp thu vào trong tế bào. Khi tế bào vi khuẩn bị vỡ do bị tan (lysis), ADN vòng tròn của chúng thoát ra môi trường thành các đoạn thẳng với chiều dài khác nhau, có khả năng gây biến nạp cho các tế bào nhận khác.
Biến nạp được nghiên cứu kĩ nhất ở các vi khuẩn Streptococcus pneumoniae, Bacillus subtilis, Haemophilus influenzae và ở một số nhóm vi khuẩn khác.
3.1.2 Cơ chế phân tử của biến nạp Cơ chế biến nạp chủ yếu bao gồm việc vi khuẩn thể nhận tiếp nhận ADN của thể cho (gọi là đoạn ngoại lai, exogenote) và sau đó ADN này có thể trao đổi với đoạn ADN tương đồng của thể nhận (gọi là đoạn nội tại, endogenote) bằng trao đổi chéo. Những tế bào có khả năng tiếp nhận ADN gọi là các tế bào khả biến (competent). Tế bào vi khuẩn nhận đoạn ngoại lai lúc đó có bộ gene ở trạng thái lưỡng bội một phần (merodiploid) hay hợp tử từng phần (merozygote). Quá trình trao đổi thông tin di truyền bằng cách chuyển chỉ một phần vật chất di truyền như thế được gọi là sự giao nạp hay tiếp hợp từng phần (meromixis).
Tương tự như trong tiếp hợp và tải nạp, để lập bản đồ di truyền bằng biến nạp cần có các tế bào thể cho và thể nhận có các kiểu gene khác nhau.
Về mặt thực nghiệm, ADN được tách ra từ các tế bào thể cho, sau đó được đưa
163
vào quần thể các thể bào thể nhận. Các tế bào thể nhận sẽ tiếp nhận các đoạn ADN một cách ngẫu nhiên. Không phải tất cả các loài vi khuẩn đều có khả năng tiếp nhận ADN. Ngay cả những loài có khả năng này cũng chỉ có thể tiếp nhận được ADN ở những pha sinh trưởng nhất định và trong môi trường nuôi cấy cụ thể. Các loài Streptoccocus pneumoniae (tức Diplococcus) và Bacillus subtilis tương đối dễ dàng trở thành khả biến hơn, trong khi E. coli phải mất đi hai loại enzyme exonuclease và phải được nuôi cấy trong môi trường có nồng độ cao của calcium chloride để làm cho màng tế bào của nó có thể thấm được DNA. Do vậy để lập bản đồ gene ở E. coli người ta ưa dùng tiếp hợp và tải nạp hơn. Tuy nhiên, trong công nghệ DNA tái tổ hợp, biến nạp E. coli là một khâu rất quan trọng.
Để biến nạp có thể xảy ra với hiệu quả cao ở vi khuẩn, chẳng hạn B. subtilis, DNA biến nạp phải có mạch kép và phân tử lượng tương đối cao (1.106 dalton). Khi AND xuyên qua màng tế bào của vi khuẩn khả biến thì một trong các sợi của ADN bị phân huỷ. Sau đó sợi đơn ADN chuyển sang có thể trao đổi với nhiễm sắc thể thể nhận ở vùng tương đồng; sự kiện này có thể phát hiện được nhờ những khác biệt di truyền thích hợp giữa các tế bào thể cho và thể nhận.
Tóm lại, hiệu quả của biến nạp phụ thuộc vào ba yếu tố:
(i) Tính dung nạp hay khả biến của tế bào thể nhận;
(ii) Kích thước của đoạn ADN được biến nạp;
(iii) Nồng độ của ADN.
Cơ chế phân tử của biến nạp (trong thí nghiệm Griffith), về cơ bản, có thể giải thích như sau:
Diễn biến của quá trình biến nạp ở cấp độ phân tử được tóm tắt trên sơ đồ hình 14. Để dễ hiểu, các giải thích dựa theo ADN của các dòng vi khuẩn S và R trong thí nghiệm của Griffith. Quá trình gồm các giai đọan chủ yếu:
– Sự phân hủy ADN tế bào cho: ADN tế bào cho có thể là của tế bào tự nhiên bị phân hủy hoặc trong thí nghiệm bị gây chế bằng nhiệt độ cao hay tác nhân phá vỡ tế bào.
– ADN bám vào bề mặt tế bào: Protein gắn vào ADN.
– Thâm nhập của ADN: Sợi ADN mạch kép của dòng vi khuẩn S sau khi chui qua màng tế bào của dòng R thì một mạch S sẽ bị nucleaza của tế bào cắt, còn lại một mạch nguyên.
– Bắt cặp (Synapsis)và tái tổ hợp: Nhờ sự hỗ trợ của protein RecA ADN của thể nhận R sẽ biến tính tách rời 2 mạch ở 1 đoạn dễ bắt cặp với đoạn ADN thể cho S vừa chui vào.
– Sao chép: Sau khi bắt cặp tạo đoạn lai R - S, phân tử ADN sao chép tạo ra 2 sợi, một sợi kép R-R và sợi kép khác có mang đoạn ADN thể nhận S-S. Kết quả
cuối cùng là đọan gen của SIII chèn vào bộ gen tế bào nhận. Sau phân bào thì một dòng tế bào nhận được ADN ngoại lai vào bộ gen - tế bào được biến nạp. Tế bào đã
được biến nạp sinh sản tạo dòng nhận RIImới. Quá trình biến nạp được mô tả ở hình 15a và 15b
Hình 15a-. Sơ đồ diễn biến quá trình biến nạp ở cấp độ phân tử ở vi khuẩn gram dương
a. ADN bám vào. b. Thâm nhập. c. Bắt cặp và tái tổ hợp. d. – Tế bào biến nạp
Hình 15b: Sơ đồ quá trình biến nạp 2 thành công và không thành công
3.1.3. Xác định liên kết gene bằng biến nạp
nồng độ ADN. Nếu hai gene nằm trên cùng một đoạn ADN thì sự biến đổi tần số biến nạp kép (liên kết) sẽ giống như biến nạp đơn. Còn nếu như biến nạp kép là do hai đoạn ADN khác nhau cùng chui vào tế bào (không liên kết) thì đường cong biến đổi của tần số biến nạp kép sẽ có độ dốc rõ rệt hơn so với biến nạp đơn.

Biến nạp chỉ được dùng để lập bản đồ gene cho một số loài. ADN thể cho được tách ra và làm đứt gãy thành những đoạn nhỏ Đối với những tế bào khả biến, tần số biến nạp khoảng 1/ 103 tế bào. Nếu hai gene a và b xa nhau trên nhiễm sắc thể thì nó thường nằm ở 2 đoạn bị đứt ra khác nhau. Khi đó tần số biến nạp cả hai gene này vào thể cho sẽ khoảng 1/103 x 1/103 = 1/106. Nếu hai gene này gần nhau thì tần số đồng biến nạp của chúng xấp xỉ bằng biến nạp đơn: 1/103. Nghiên cứu khả năng đi kèm nhau của các gene biến nạp có thể xác định được trật tự của chúng.
3.2 Tiếp hợp ở vi khuẩn (Conjugation)
3.2.1. Định nghĩa, thí nghiệm và đặc điểm chung
- Định nghĩa: Tiếp hợp là hiện tượng tiếp xúc trực tiếp giữa hai tế bào vi khuẩn và kèm theo việc truyền một phần vật chất di truyền từ tế bào thể cho (donor) sang tế bào thể nhận (recipient, receptor).
- Thí nghiệm: Vào năm 1946, Joshua Lederberg và E.L.Tatum (Hình 16a) đã sử dụng các nòi đột biến khuyết dưỡng khác nhau ở E, coli để chứng minh có tái tổ hợp giữa chúng. Cụ thể, nòi A có kiểu gene met- bio- thr+ leu+ và nòi B có kiểu gene ngược lại, me+ bio+ thr- leu. Nếu đem nuôi cấy riêng rẽ trên các môi trường tối thiểu, các nòi này không sinh trưởng được. Tuy nhiên sau khi trộn lẫn hai nòi A và B trong ống nghiệm và đem cấy lên môi trường tối thiểu, thấy có xuất hiện các khuẩn lạc. Điều đó chứng tỏ có sự tái tổ hợp giữa hai nòi và làm xuất hiện dạng lai hay các thể tái tổ hợp, với kiểu gene met+ bio+ thr+ leu+ , bù đắp sự khiếm khuyết cho nhau trong nhu cầu dinh dưỡng (Hình 16b).
Trong quá trình tách chiết ADN để tiến hành biến nạp, nhiếm sắc thể của vi khuẩn thường bị đứt ra thành khoảng 250 đoạn, tức các phân tử riêng biệt. Các gene nằm ở những vị trí khác nhau trên nhiễm sắc thể vi khuẩn khi đó sẽ bị tách rời nhau và được truyền đi trong quá trình biến nạp một cách độc lập. Vì số gene ở vi khuẩn rất lớn mà số đoạn ADN lại hạn chế nên không loại trừ các trường hợp hai gene khác nhau cùng nằm trên một đoạn ADN, và vì vậy, cùng được chuyển đi. Trên thực tế những trường hợp như thế đã quan sát thấy ở hàng loạt vi khuẩn và gọi là biến nạp liên kết. Có thể phát hiện được biến nạp liên kết bằng cách xác định tần số biến nạp kép (biến nạp đồng thời hai gene, hay đồng biến nạp, cotransformed) và so sánh nó với trị số kỳ vọng khi hai gene được truyền đi một cách độc lập. Trong thí nghiệm người ta xác định sự liên kết gene bằng cách phát hiện hiệu quả pha loãng ADN. Trong một giới hạn nào đó của nồng độ ADN thì tần số biến nạp tỉ lệ tuyến tính với 166
Hình 16: Thí nghiệm kinh điển về tái tổ hợp ở vi khuẩn.
(a) Hai nòi khuyết dưỡng A và B được hỗn hợp với nhau. Mỗi nòi tự nó không thể mọc được trên trên môi trưởng tối thiểu, nhưng khi hỗn hợp hai nòi làm xuất hiện các thể tái tổ hợp có thể mọc được trên môi trường tối thiểu, cho các khuẩn lạc mọc trên đĩa petri ở giữa.
(b) Sự kiện tái tổ hợp làm sản sinh các thể nguyên dưỡng. Một trao đổi chéo xảy ra giữa các gene bio và thr đem các allele kiểu dại về một nhiễm sắc thể và các allele đột biến trên nhiễm sắc thể kia.
Như vậy, theo hiểu biết sau này ta có thể hình dung một số vi khuẩn, ví dụ E. coli, có thể truyền một phần nhiễm sắc thể của chúng cho thể nhận mà chúng tiếp xúc trực tiếp. Khi thể cho tái bản nhiễm sắc thể của nó thì bản sao được tiêm vào thể nhận. Tại thời điểm bất kỳ thể cho và thể nhận tách khỏi nhau thì việc truyền gene dừng lại. Các gene thực hiện "chuyến du hành" thành công sẽ thay chỗ tương đương trong nhiễm sắc thể của thể nhận. DNA được truyền gồm một bộ các gene nằm kề nhau gọi là các gene truyền. Các gene truyền có thể tồn tại ở dạng phân tử DNA mạch vòng gọi là các plasmid hoặc nằm bên trong nhiễm sắc thể thể cho gọi là plasmid lồng ghép trong nhiễm sắc thể
Phân tích: Hình 17 cho thấy cơ chế tiếp hợp ở E. coli, trong đó:
- Thể "cho" (donor) thiếu hẳn các gene chức năng cần thiết cho tổng hợp vitamin biotin và amino acid methionine (Bio-, Met-) vì thế cần phải bổ sung các chất này vào môi trường nuôi cấy của nó.
- Thể "nhận" (recipient) có các gene này (Bio+, Met+) nhưng các gene (đột biến) không hoạt động chức năng ngăn cản không cho nó tổng hợp các amino acid threonine và leucine (Thr-, Leu-), cho nên phải bổ sung các chất này vào môi trường nuôi cấy của nó.
- Khi được nuôi cấy chung với nhau, một số tế bào thể nhận nhận được các gene Thr và Leu có chức năng bình thường từ thể cho.
- Một sự trao đổi chéo kép có thể làm thay thế các allele không hoạt động chức năng bằng các allele có chức năng.
- Bây giờ các tế bào có thể sinh trưởng trên môi trường tối thiểu chỉ chứa glucose và muối.
Hình 17: Cơ chế tiếp hợp và tái tổ hợp gene ở các tế bào E. coli .
- Các đặc điểm
(i) Chỉ có thể xảy ra giữa các tế bào thuộc các kiểu bắt cặp đối diện; trong đó thể cho (male) mang một nhân tố hữu thụ (F+) và thể nhận (female) không có nhân tố này (F-). Lưu ý rằng, hiện tượng phân hoá giới tính này ở vi khuẩn (tương tự các giống đực và cái ở sinh vật bậc cao) đã được Hayes phát hiện từ năm 1953.
(ii) F là một bộ các gene nguyên vẹn nhận được từ một plasmid và bây giờ được sát nhập vào nhiễm sắc thể vi khuẩn; nó xác định khởi điểm tái bản cho nhiễm sắc thể; một phần của F là motor ("đầu máy") đẩy nhiễm sắc thể vào trong tế bào thể nhận; phần còn lại của nó là "toa công nhân".

(iii) Ở E. coli, trung bình một gene truyền qua mất một giây mà các tế bào vẫn được duy trì tiếp hợp với nhau, lúc đó mất khoảng 100 phút để chuyển toàn bộ bộ gene 4.377 gene của nó. Nhưng quá trình này thực ra dễ dàng bị gián đoạn, vì thế có thể các gene vật chủ nằm gần phía sau các gene của plasmid F đi trước sẽ được truyền đi sớm hơn là các gene nằm xa hơn. Phần còn lại "toa công nhân" hiếm khi được truyền đi vì vậy khó mà nhận được một nhân tố F hoàn chỉnh, tế bào thể nhận tiếp tục là F.

(iv) DNA truyền qua sẽ tìm ra vùng tương đồng trên DNA thể nhận và thay chỗ đó bằng một trao đổi chéo kép.
(v) Bằng cách tách các tế bào một cách cẩn thận vào bất cứ lúc nào, trật tự và khoảng cách tương đối của các gene có thể được xác định. Theo đó, ta có thể thiết lập một bản đồ di truyền — tương đương với bản đồ di truyền của các eukaryote. Nhưng ở đây các quãng cách bản đồ được tính bằng giây (hoặc phút), chứ không phải bằng centiMorgan.
Tiếp hợp được gán cho các kiểu yếu tố di truyền nhất định, cụ thể là các plasmid. Việc truyền plasmid từ thể cho sang thể nhận không đòi hỏi ADN của plasmid để tái tổ hợp với ADN của thể nhận. Tiếp hợp là có tính bảo tồn - thể cho vẫn giữ lại bản sao của plasmid sau khi truyền đi, điều đó chỉ ra rằng sự tái bản của plasmid xảy ra trong khi tiếp hợp.
Sự tái bản plasmid đòi hỏi phải có một "cầu tiếp hợp" (mating bridge) giữa các tế bào cho và nhận. Đó là vùng tiếp xúc giữa các tế bào này, trong đó ADN được truyền qua một cái lỗ. Như vậy, trước khi cầu tiếp hợp có thể hình thành, thể cho phải nhận biết tế bào nhận và phải tiếp xúc với thể nhận. Hai sự kiện này không nhất thiết xảy ra đồng thời.
Có nhiều gene được cần đến cho tiếp hợp. Chẳng hạn, đối với plasmid TiC58 các trb operon mã hoá cho các sản phẩm gene cần thiết để nhận biết thể nhận và bắt cặp (giao phối). Các gene trong cụm phức hợp này mã hoá cho sợi lông giới tính (sex pilus), yếu tố thiết yếu cho tiếp hợp. Lông giới tính có thể rất dài (ví dụ lông sinh ra bởi plasmid F) hoặc rất ngắn (ví dụ lông sinh ra bởi plasmid RP4). DNA không phải được truyền qua lông giới tính. Mặc dù thể nhận còn chưa biết về plasmid tiếp hợp bất kỳ nào, nhưng sợi lông giới tính là cần cho việc nhận biết tế bào thể nhận. Các plasmid tương tự F mã hoá cho các lông gấp nếp (flexous pili) bắt cặp tốt trong môi trường lỏng, nhưng các plasmid mã hoá cho các lông ngắn cứng (short stiff pili) thường bắt cặp chỉ trong bề mặt đặc. Sự tiếp xúc như sau, sợi lông dài gấp nếp của F hoạt động như một chiếc motor co rút - các tế bào cho và nhận được kéo lại gần nhau khi các tiểu đơn vị của sợi lông khử polymer thành ra màng tế bào chất của các tế bào cho. Ngược lại, bản chất co rút của các sợi lông khác vẫn chưa được xác định.
Các tra operon mã hoá các sản phẩm gene cần cho các chức năng tái bản DNA. Sự tái bản DNA đòi hỏi nhiều bước.
Hình 18: Sự tiếp hợp giữa các tế bào E. coli F+ (đực) và F- (cái)
Sự tiếp hợp giữa các tế bào E. coli F+ (đực) và F- (cái), với việc truyền nhân tố F- (ở dạng DNA sợi đơn đang tái bản kiểu vòng lăn) từ tế bào F+ sang tế bào F- qua cầu tiếp hợp. Trong điều kiện lý tưởng, tế bào F- nhận được một bản sao của nhân tố F và sau đó tổng hợp sợi DNA bổ sung và trỏ thành tế bào F+.
Như vậy, đặc trưng của việc truyền DNA trong tiếp hợp là đòi hỏi phải có sự tiếp xúc tế bào-tế bào (tiếp hợp được ngăn bởi màng lọc chỉ cho phép pha trộn môi trường nhưng ngăn cản tiếp xúc giữa các tế bào cho và nhận); xảy ra thông qua lỗ tiếp hợp; truyền theo một chiều từ thể cho sang thể nhận mà không ngược lại;
• Khởi đầu tái bản vòng lăn (rolling circle replication) hay tái bản sigma (σ replication) đòi hỏi phải đứt sợi ở DNA của plasmid đóng vòng. DNA plasmid bị đứt tại một vị trí tác động cis đặc thù (specific cis-acting site) gọi là nic bên trong khởi điểm của đoạn truyền (oriT). Enzyme thuỷ phân làm đứt đó được gọi là "nickase" hay "strand transferase". Để làm đứt ADN, nickase vẫn giữ sự kết dính đồng hoá trị nhóm 5' phosphate, để lại nhóm 3'OH tự do. Các protein hỗ trợ tạo thuận lợi cho nickase bám vào oriT, và phức hợp này được gọi là "relaxosome" (thể giãn xoắn).

• Tái bản DNA khởi đầu tại 3'OH, và tiến hành theo chiều 5' đến 3'.
• Phức hợp relaxasome bám vào lỗ truyền.
• Sự tái bản DNA kiểu vòng lăn trong thể nhận đẩy ADN sợi đơn (single stranded ADN = ssDNA) vào trong tế bào thể nhận.
• ssDNA được biến đổi thành ADN sợi kép (double stranded ADN = dsDNA) trong thể nhận bẳng cách tổng hợp ADN sợi ra chậm.
Khi vắng mặt thể nhận, relaxasome làm đứt và nối lại DNA của plasmid. Măc dù cơ chế còn chưa rõ, sự tiếp xúc với tế bào thể nhận bằng cách nào đó kích thích làm đứt và truyền DNA đi sau đó.
- Phạm vi vật chủ Phạm vi vật chủ (host range) có thể được xác định bằng phạm vi các vật chủ mà có thể sửdụng với tư cách là các thể nhận cho việc truyền DNA hoặc phạm vi các vật chủ mà plasmid có thể tái bản bên trong chúng. Phạm vi vật chủ cho tiếp hợp được xác định tiêu biểu bằng khả năng plasmid tái bản nội bên trong vật chủ đặc thù. Chẳng hạn, xét các plasmid tiếp hợp sau:
P lasmid Phạm vi vật chủ tiếp hợp Phạm vi vật chủ tái bản Phạm vi vật chủ truyền
F hẹp (Gram- đường ruột) hẹp rộng
R P4 rộng (Gram- và Gram+ ) rộng rộng
Phạm vi vật chủ của một số plasmid tiếp hợp là rất rộng, bao gồm cả việc truyền sang thể nhận là các vi khuẩn, nấm men, các tế bào thực vật và các tế bào động vật có vú. Tuy nhiên, sự tái bản của plasmid thường bị giới hạn vào các vật chủ nào đ
3.2.2. Các plasmid và sự truyền DNA ở vi khuẩn
Plasmid là những phân tử DNA mạch vòng có khả năng tái bản độc lập và có kích thước 0,05- 10% kích thước của nhiễm sắc thể vi khuẩn.
Chúng có ở nhiều loài vi khuẩn và thường không quan trọng đối với sự sinh trưởng của tế bào. Plasmid rất đa dạng về gene và nhiều gene của chúng không có trong nhiễm sắc thể vi khuẩn và bằng nhiều cách, chúng có thể có được các gene của vi khuẩn. Sự tồn tại của plasmid được chứng minh bằng kiểu hình mà tế bào mang chúng thể hiện. Ví dụ, plasmid mang alen tetr sẽ làm cho vi khuẩn kháng với tetracyclin (Hình 18).

Plasmid dựa vào các enzyme tái bản DNA của tế bào chủ để tái bản, nhưng sự khởi đầu lại do các gene của plasmid kiểm soát. Do vậy số bản sao của chúng có thể khác nhau. Plasmid tái bản mạnh có thể tạo ra 50 bản sao trong một tế bào, trong khi các plasmid khác chỉ cho 1 hoặc 2 bản sao.
Có nhiều loại plasmid ở E. coli. Các plasmid đã được nghiên cứu kỹ là plasmid R, Col và F. Plasmid R gọi là các plasmid kháng thuốc vì chúng có mang các gene kháng với một hay nhiều loại kháng sinh (Hình 18).
Plasmid Col có khả năng tổng hợp colicin - các protein có thể giết chết các vi khuẩn họ hàng không mang plasmid Col.
Hình 18 Các plasmid của vi khuẩn: pSC101 (trái), plasmid siêu xoắn (giữa), và pBR322 - một vector tạo dòng được dùng trong DNA sequencing - với vị trí của khởi điểm tái bản (ori), các gene kháng AmpR và TetR và một số vị trí của các enzyme cắt giới hạn.
Plasmid F (F = fertility), còn gọi là các plasmid giới tính hay nhân tố F có kích thước ~94.500 cặp base (xấp xỉ 1/50 chiều dài của nhiễm sắc thể vi khuẩn), rất được quan tâm vì chúng chứa các gene truyền và xác định tính hữu thụ của vi khuẩn. Nhân tố F có chứa các gene quy định sự hình thành các lông giới tính (pili) mảnh, mềm và có thể rất dài trên mặt tế bào gọi là lông F. Các gene khác trong nhân tố F chuyên trách việc hình thành cầu tiếp hợp nối các tế bào cho và nhận để nhân tố F có thể chuyển qua. F là plasmid tái bản yếu, và một tế bào F+ điển hình mang 1 hoặc 2 bản sao của nhân tố F. Phần tử di truyền F+ này tồn tại ngoài nhiễm sắc thể nên còn gọi là episome
Nhân tố F được truyền từ tế bào F+ sang F- trong quá trình tiếp hợp và xảy ra đồng thời với việc tái bản plasmid. Sự tiếp xúc giữa F+ và F là tín hiệu bắt đầu cho việc tái bản nhân tố F và chỉ một sợi đơn thẳng được truyền sang tế bào thể nhận (Hình 6.6). Quá trình tổng hợp DNA xảy ra đồng thời ở cả hai tế bào cho và nhận để tạo ra DNA sợi kép dạng vòng.
Sau đó cả hai tế bào đều chứa nhân tố F và có thể thực hiện chức năng như là tế bào thể cho (Hình 17).
3.2.3. Nòi Hfr
Một số nòi F+ có thể sinh ra những tế bào "thể cho" có khả năng truyền gene trên nhiễm sắc thể với tần số cao, gọi là các tế bào có khả năng tái tổ hợp cao hay các nòi Hfr (high frequency recombination). Các nòi Hfr cũng có những lông F như các tế bào F+ nhưng giữa chúng có một điểm khác biệt quan trọng ở chỗ, trong các tế bào Hfr nhân tố F được đính vào nhiễm sắc thể vi khuẩn. Vì vậy ngoài những biểu hiện của nhân tố F giống như các tế bào F+, các tế bào Hfr còn có khả năng truyền đi nhiễm sắc thể vi khuẩn qua ống tiếp hợp.
3.2.4. Sự xen plasmid F vào trong nhiễm sắc thể vật chủ
Nhân tố F được đính vào nhiễm sắc thể bằng cơ chế trao đổi chéo đơn và làm cho nhiễm sắc thể này trở nên lớn hơn. Tế bào Hfr có thể biến đổi thành tế bào F+ (mang nhân tố F trong tế bào chất) bằng một quá trình ngược lại.
Hình 19: (a) Sự sát nhập của nhân tố F vào nhiễm săc thể ở vi khuẩn "cho" F+ tạo thành nhiễm sắc thể có kích thước lớn trong vi khuẩn "nhậni" Hfr bằng một trao đổi chéo đơn; hiện tượng này mang tính thuận nghịch. (b) Truyền DNA sợi đơn từ thể cho (Hfr) sang thể nhận (F-)
Bởi vì nhân tố F có khả năng đính vào nhiều vị trí khác nhau trên nhiễm sắc thể vi khuẩn, do đó trình tự gene được chuyển sang các tế bào F- bởi các nòi Hfr khác nhau là rất khác nhau. Vì khi bắt đầu chuyển gene từ Hfr sang F- thì nhân tố F bị tách ra, nên một phần được chuyển đi trước, phần còn lại chính là đuôi của nhiễm sắc thể
E. coli được chuyển đi sau cùng hoặc không được truyền sang nếu qúa trình bị ngắt quãng giữa chừng. Khi đó chỉ một phần nhiễm sắc thể được chuyển sang F-, phần còn lại cùng với nhân tố F vẫn nằm lại trong tế bào Hfr.
Những khác biệt cơ bản giữa việc truyền nhân tố F và Hfr như sau:
(i) Cần 100 phút để truyền toàn bộ nhiễm sắc thể vi khuẩn trong khi chỉ cần khoảng 2 phút để truyền nhân tố F.
(ii) Quá trình truyền DNA Hfr thường bị đứt quãng. Trung bình khoảng vài trăm gene được truyền trước khi hai tế bào tách ra.

(iii) Sau khi được truyền F- vẫn là F- vì đoạn cuối của F thường không được truyền sang (khi tiếp hợp giữa F với Hfr ).
(iv) Một đoạn DNA của Hfr được truyền đi không thể tạo thành mạch vòng được và không tự tái bản được, chúng có thể trao đổi chéo với nhiễm sắc thể của thể nhận để tạo nên các thể tái tổ hợp F- . Ví dụ, cho tiếp hợp Hfr leu+ x F- leu- có thể cho ra F- leu+ .
* Sự hình thành Hfr: Các nòi Hfr có thể hình thành giữa một yếu tố (đoạn xen) IS trên plasmid F và cùng yếu tố IS đó trên nhiễm sắc thể vật chủ (Hình 20).

Hình 20: Sự hình thành Hfr tại đoạn xen IS
Có nhiều đoạn xen IS trong nhiều nhiễm sắc thể vi khuẩn. Chẳng hạn, E. coli K-12 kiểu dại có 8 đoạn xen IS1, 6 đoạn xen IS2 và 5 đoạn xen IS3. Vì các đoạn xen IS là các yếu tố di truyền vận động, mỗi nòi có thể có số lượng các đoạn xen IS khác nhau. Sự định khu của một số đoạn xen IS1, IS2 và IS3 trên nhiễm sắc thể E. coli K-12 và hướng của chúng được cho thấy ở Hình 21 dưới đây.
Hình 21: Vị trí của các đoạn xen IS1, IS2 và IS3 trên nhiễm sắc thể E. coli K12 và hướng của chúng.
Bằng cách đó, các đoạn xen của Hfr có thể được phân lập tại nhiều vị trí ở E. coli và các hướng khác nhau so với nhiễm sắc thể. Một số ví dụ về các đoạn xen của Hfr đã được phân lập ở E. coli được cho thấy ở hình 22 dưới đây. Lưu ý rằng nhiễm sắc thể E. coli ở đây được biểu diễn dưới dạng mạch thẳng. Các con số bên trên hình biểu thị số phút (hay là "centisome") trên bản đồ di truyền của E. coli.
Hình 22: Các đoạn xen của Hfr đã được phân lập ở E. coli.
Ngược lại, Salmonella typhimurium khuyết nhiều yếu tố IS vốn có mặt ở E. coli. Rõ ràng, các đoạn xen của Hfr được gây ra bởi sự tái tổ hợp tương đồng giữa các đoạn xen IS trên plasmid F và nhiễm sắc thể là hiếm gặp ở S. typhimurium.

Một thể đột biến mới được phân lập đó là StrR và không thể sử dụng acetate như một nguồn carbon (ace). Để xác định vị trí các đột biến trên bản đồ, nó được cho giao phối với bốn nòi Hfr StrS ace+ khác nhau được chỉ ra dưới đây. [Các mũi tên chỉ ra sự định khu và hướng truyền từ mỗi Hfr khác nhau.]
các thể tái tổ hợp. Từ đó xác định được khoảng cách giữa các gene và vị trí của chúng trên nhiễm sắc thể đo bằng đơn vị thời gian (phút; Hình 24a). Đó là kỹ thuật ngắt quãng tiếp hợp mà Elie Wollman và Jacob đã sử dụng để thiết lập bản đồ trật tự phân bố các gene.
Hình 23 Vị trí và hướng truyền của các Hfr 1-4.
3.2.5. Lập bản đồ di truyền bằng tiếp hợp ngắt quãng
Bằng các phép giao phối Hfr x F- có thể lập được bản đồ gene. Tuy nhiên, bản đồ này khác với bản đồ liên kết gene thông thường, đó là bản đồ trật tự truyền gene.
Bằng cách ngắt quãng cơ học việc truyền gene trong quá trình giao phối có thể xác định thời gian mà một gene nào đó được truyền sang nhờ việc xác định tần số
Hình 24: (a) Vị trí và khoảng cách của một số gene trên nhiễm sắc thể đo bằng phút. (b) Bản đồ mạch vòng của bộ gene E. coli được thiết lập từ năm 1976 và chỉ chứa một phần các gene mà hiện giờ đã được lập bản đồ

Để lập bản đồ của toàn bộ nhiễm sắc thể mạch vòng ở E. coli người ta đã sử dụng nhiều nòi Hfr khác nhau, mặc dù vậy trật tự của gene và vị trí của chúng trên nhiễm sắc thể, kể cả khoảng cách giữa các gene (đo bằng phút), vẫn giống nhau. Toàn bộ bản đồ nhiễm sắc thể ở E. coli là 100 phút (Hình 24).
3.2.6. Lập bản đồ với E. coli: các plasmid F' và phép thử cis-trans

3.2.6.1. Các plasmid F'
Nhân tố F đôi khi bị cắt khỏi DNA của tế bào Hfr bằng cơ chế trao đổi chéo các đoạn tương đồng giống như khi lồng ghép. Tuy nhiên, trong một vài trường hợp. Sự trao đổi chéo xảy ra không thật chính xác - tại đoạn không tương đồng - và vì
vậy có thể tạo ra một plasmid mang một phần DNA của nhiễm sắc thể vi khuẩn - đó là plasmid F'.
Bằng cách dùng những nòi Hfr có các điểm khởi đầu truyền gene khác nhau, người ta đã tách được những plasmid F' khác nhau. Những plasmid này có mang các đoạn nhiễm sắc thể của tế bào Hfr ở dạng lưỡng bội từng phần nên rất có ích cho việc nghiên cứu sự biểu hiện của gene. Người ta ký hiệu kiểu gene của tế bào, ví dụ mang đột biến lac và mẫn cảm với streptomycin, có chứa plasmid F' lac+ như sau: F' flac+ flac- str.
3.2.6.2. Phép thử bổ sung cis-trans (cis-trans complementation test)

Có thể tóm tắt như sau: Hai đột biến khác nhau ảnh hưởng lên cùng một chức năng thì có thể thuộc về trắc nghiệm cis-trans, hay phép thử bổ sung, để xác định xem liệu chúng xảy ra trong cùng gene hay trong các gene khác nhau. Trong phép thử này, hai gene đột biến được cung cấp cho cùng tế bào ở dạng trans (trên các nhiễm sắc thể riêng biệt). Nếu như các đột biến bổ sung bù trừ cho nhau để cho chức năng kiểu dại, chúng có thể nằm trong các gene riêng biệt. Nếu như các đột biến không bổ sung được cho nhau, chứng tỏ chúng ảnh hưởng cùng một gene như nhau.
3.3 Tải nạp (Transduction)
3.3.1. Định nghĩa, thí nghiệm và đặc điểm chung Tải nạp là quá trình chuyển vật chất di truyền từ vi khuẩn cho sang vi khuẩn nhận thông qua phage. Những phage này được gọi là các hạt tải nạp. Có hai dạng phage tải nạp là phage tải nạp chung và phage tải nạp đặc hiệu. Phage tải nạp chung sản sinh ra các hạt mang những đoạn DNA vi khuẩn từ bất kỳ phần nào của nhiễm sắc thể vi khuẩn và không có DNA phage. Còn phage tải nạp đặc hiệu sản sinh ra các hạt mang cả DNA phage và gene vi khuẩn liên kết thành một sợi đơn, và gene vi khuẩn được lấy từ những vùng đặc biệt của nhiễm sắc thể vi khuẩn.
3.3.2. Tải nạp chung (generalized transduction): Phage P1
Tải nạp chung là trường hợp bất kì gene nào của thể cho cũng có thể được chuyển sang thể nhận bằng phage. Thí nghiệm đầu tiên về tải nạp chung được N.
Zinder và J. Lederberg tiến hành vào năm 1952 trên vi khuẩn Salmonella typhimurium. Các tác giả này đã sử dụng một ống thuỷ tinh hình chữ U có ngăn ở giữa bằng màng lọc vi khuẩn, còn phage vẫn chui qua được. Bên trái của ống có chứa nòi vi khuẩn LA2 với kiểu gene phe+ trp+ met- his- còn bên phải ống mang nòi LA22 với kiểu gene phe- trp- met+ his+ .
Vi khuẩn kiểu dại đã xuất hiện ở bên phải ống nhưng không thấy có ở bên trái ống, chứng tỏ vật trung gian chuyển gene (mà sau này tìm ra là P22) đã được LA2 sinh ra và nó làm xuất hiện các thể tái tổ hợp kiểu dại ở nòi LA22. P22 là một phage ôn hoà có khả năng tiềm tan hoá nòi LA22 và gây tan nòi LA2. Trong quá gây tan một đoạn DNA vật chủ có thể được bọc gói trong vỏ của phage, vì vậy khi tiềm tan hoá các
tế bào LA22 chúng có thể sinh ra các thể tái tổ hợp kiểu dại do kết quả của trao đổi chéo giữa các đoạn DNA của LA2 (do phage P22 đưa sang) và nhiễm sắc thể của LA22.
Hình 25 Tải nạp chung (generalized transduction).
Tải nạp có thể diễn ra ở các vi khuẩn khác như E. coli với sự trung gian của phage P1, ở Bacillus subtilisvowis sự tham gia của phage SP10. Phân tích di truyền bằng tải nạp chung: DNA của phage P22 bằng khoảng 1/100 DNA của Salmonella typhimurium, vì vậy phage chỉ có thể chuyển đi một đoạn rất nhỏ nhiễm sắc thể vật chủ. Do đó tải nạp có thể cung cấp thông tin về hai đột biến nằm rất gần nhau và cũng có thể giúp xác định trình tự tương đối của các gene khi tiến hành nghiên cứu đồng thời ba gene. Ví dụ, dùng phage P1 để tải nạp các gene giữa hai nòi E. coli. Nòi cho là leu+ thr+ azir, nòi nhận là leu- th- azis. Kết quả của thí nghiệm được tổng kết ở bảng dưới đây.

Tần số đồng tải nạp các dấu chuẩn trong thí nghiệm dùng phage P1, nòi cho là leu+ thr+ azir, và nòi nhận là leu- th- azis
Trong thí nghiệm chọn lọc theo leu+ các kết quả cho thấy các gene leu và azi nằm gần nhau và cả hai đều nằm xa gene thr. Kết quả của thí nghiệm chọn lọc theo
178
thr+ cho thấy gene leu nằm gần gene thr hơn so với gene azi. Vậy trình tự các gene là
thr- leu- azi
Đoạn DNA tải nạp thường mang khoảng 50 gene và tải nạp có thể dùng để lập bản đồ gene. Giả sử một quần thể phage P1 được lấy từ vi khuẩn có kiểu gene leu+ gal+ bio+. Trong số các phage này sẽ có các hạt tải nạp chỉ mang hoặc leu+ hoặc gal+. Vì vậy nếu cho chúng xâm nhiễm vi khuẩn có kiểu gene leu-gal- thì có thể có các thể tải nạp leu+gal- hoặc leu-gal+ . Nếu tỷ lệ phage / vi khuẩn rất nhỏ hơn 1 sẽ không có các vi khuẩn lai leu+ gal+ . Rất hiếm khi một đoạn DNA vi khuẩn lại mang cả hai gene leu và gal vì hai gene này khá xa nhau trên nhiễm sắc thể vi khuẩn.
Hai gene gal và bio chỉ cách nhau 2,3 x 104 cặp base nên chúng có thể cùng có mặt trên đoạn DNA tải nạp, vì hạt tải nạp có thể bọc gói một đoạn DNA có kích thước 7,7x 104 cặp base. Tuy nhiên không phải tất cả hạt tải nạp gal+ đều cũng phải là bio+ và ngược lại, vì enzyme nuclease có thể cắt DNA ở điểm gẽưa hai gene này. Hiện tượng tải nạp cả hai gene đánh dấu được gọi là đồng tải nạp. Tần số đồng tải nạp tỷ lệ nghịch với khoảng cách giữa các gene. Sử dụng môi trường chọn lọc cho cả hai gene, ta sẽ phát hiện được các thể đồng tải nạp. Như vậy, việc nghiên cứu hiện tượng đồng tải nạp có thể giúp ta xây dựng bản đồ di truyền.
Có thể sử dụng đoạn DNA tải nạp mang ba gene đánh dấu để xác định trật tự gene. Trong trường hợp này gene nằm giữa sẽ có tần số tải nạp thấp nhất vì nó cần bốn trao đổi chéo để hình thành, trong khi hai gene kia chỉ cần có hai.
3.3.3. Tải nạp chuyên biệt (specialized transduction): Phage λ
Tải nạp chuyên biệt hay đặc hiệu là trường hợp phage chỉ truyền đi những gene nhất định từ thể cho sang thể nhận. Ví dụ, trường hợp phage lambda (λ) thực hiện tải nạp giũa các vi khuẩn E. coli. Phage λ chứa DNA có chiều dài 50.000 cặp base, bằng khoảng 1/4 DNA của các phage T chẵn. Hầu như toàn bộ DNA của λ có mạch kép và bổ sung nhau.
Khi E. coli bị nhiễm λ thì DNA của phage tạo thành vòng tròn, nó có thể sao chép và bắt đầu sinh tan, hoặc có thể xen vào nhiễm sắc thể vật chủ để chuyển sang trạng thái prophage. Việc xen vào này diễn ra giống như đối với nhân tố F: có một điểm dính đặc hiệu cho λ ở trên DNA vật chủ (λ attachment site, viết tắt là attλ ). Đây là một đoạn tương đồng với đoạn trên DNA phage gọi là b2. Sau đó diễn ra trao đổi chéo giữa DNA phage và DNA vi khuẩn tại vị trí nói trên dẫn đến xen bộ gene λ vào giữa các gene gal (galactose) và gene bio (biotin) trên nhiễm sắc thể E. coli.

Hình 26 Tải nạp chuyên biệt hay tải nạp hạn chế (restricted transduction).
3.3.4. Lập bản đồ các đột biến bằng tải nạp
Năm 1956 J. Lederberg đã tiến hành tải nạp gene từ nòi vi khuẩn E. coli
K12(λ) kiểu dại tiềm tan sang nòi E. coli K12 không tiềm tan và có mang nhiều đột biến khuyết dưỡng. Kết quả là chỉ có gene gal+, tức gene nằm kế sát điểm attλ, mới được phage chuyển sang thể nhận, vì vậy gọi là tải nạp đặc hiệu.
Cơ chế tải nạp đặc hiệu nêu trên hình 6.16. Bước đầu tiên là hình thành vòng bộ gene phage sai (vì ngoài bộ gene λ còn có một đoạn nhỏ nhiễm sắc thể vi khuẩn chứa gene gal+ nằm trong vòng tròn). Một trao đổi chéo xảy ra tạo thành vòng DNA có chứa phần lớn bộ gene λ (chứ không phải tất cả) và một đoạn ngắn nhiễm sắc thể vi khuẩn chủ mang gene gal+. DNA mạch vòng (dưới tác dụng của enzyme) chuyển thành mạch thẳng để sau này lắp ráp vào các hạt phage thế hệ con gọi là λ dg (λ defective galactose), chúng có mang gene gal+ của vi khuẩn.
Phage λ dg có thể truyền gene gal+ vào tế bào thể nhận không tiềm tan. Khi nhiễm vào tế bào đó, DNA của λ dg có thể xen vào nhiễm sắc thể thể nhận bằng trao đổi chéo diễn ra ở vùng gal tương đồng. Ở đây, trao đổi chéo tạo thành DNA mạch thẳng có chứa prophage khiếm khuyết (λ def, defective prophage) nằm giữa hai gene gal của vi khuẩn. Vì gal+ là trội so với gal- nên kiểu hình là gal+ .
Tải nạp đặc hiệu có thể sử dụng để nghiên cứu di truyền học. Ví dụ, có thể tiến hành phép thử nghiệm bổ trợ đối với các đột biến nằm trong vùng gal để xác định số lượng đơn vị chức năng (cistron). Trên thực tế, người ta đã xác định được vùng gal có chứa ba cistron.
CHƯƠNG IV. HỆ THỐNG CÂU HỎI VÀ BÀI TẬP
I. Tóm tắt các dạng bài tập cơ bản
1. Tần số đồng biến nạp và đồng tải nạp cao cho thấy 2 gen rất gần nhau.
Ví dụ : Trong một thực nghiệm biến nạp, x+y+z+ được sử dụng là thể cho và xyz - là thể nhận, thu được 250 thể biến nạp. Sau đó nuôi cấy bản sao để xác định xem có y+ và/ hoặc z+ không. Kiểu gen của các thể biến nạp thu được như dưới đây. Bạn có thể kết luận gì về vị trí tương ứng của các gen?
x+yz - 53
x+yz+ 173
x+y+z - 7
x+y+z+ 17
Tần số đồng biến nạp
x+z+ =( 173 + 17 ) / 250 = 76 %
x+y+ =( 7 + 17 ) / 250 = 10 %




Chúng ta thấy rằng 76% khuẩn lạc có x+z+ . Kết quả này cho thấy x và z gần nhau. x+ và y+ xuất hiện cùng nhau với tỉ lệ 10% cho thấy y cách xa x, và có thể không liên kết với nhau.
Để tính tần số tái tổ hợp giữa x và z, chúng ta xem số lần x+ và z+ không cùng xuất hiện (đó là x+ yx - và x+ y+ z -).
RF = (53 + 7)/250 = 24 đơn vị bản đồ
2. Để các gen không liên kết đồng biến nạp và đồng tải nạp cần có 2 trao đổi chéo.
Xét hai gen a và b đủ xa nhau, tốt nhất là hai gen trên hai phân tử ADN riêng biệt:
Để các thể biến nạp là a+b+ , cả hai đọan ADN này cùng phải xâm nhập vào thể nhận. Quá trình này cần hai đứt gãy, tương đương với một trao đổi chéo kép. Vì vậy điều đó là rất hiếm.
thể cho
thể nhận
Lưu ý rằng nếu a và b gần nhau và trên cùng một mạch ADN, thì chỉ một đứt gãy và trao đổi chéo là đủ thể cho thể nhận
Ví dụ: Xét các tần số đồng biến nạp sau:
A và B: 73%
A và C: 7%
B và C: 6%.
Những gen nào không liên kết với nhau?
Chúng ta thấy gen C xuất hiện cùng với A hoặc B với tỉ lệ thấp. Như vậy có thể gen C không liên kết với A và B
3. Để xác định trình tự các gen trong quá trình biến nạp hoặc tải nạp, hãy tìm các trậttự gen cho phépphán đoán về cáctrao đổi chéokép.
Ví dụ: Khi a+b+c - là thể cho và a -bc+ là thể nhận, ta thấy lớp kiểu hình có tần số thấp nhất có kiểu gen a+b+c+ . Hãy xác định trật tự của các gen.
Giả sử trật tự gen là abc, khi đó chúng ta có thể vẽ sơ đồ phép lai như sau:
Một trao đổi chéo kép sẽ tạo ra a+ bc, lớp này không phải có tần số ít nhất. Vì vậy b không thể ở giữa.
Giả sử c ở giữa:
Trao đổi chéo kép sẽ tạo ra a+b+ c+, lớp này có tỉ lệ nhỏ nhất, chứng tỏ rằng c ở giữa.
4. Trong quá trình tiếp hợp, một gen có tần số truyền gen cao phải nằm gần với điểm khởi đầu quá trình truyền gen.
Ví dụ: Phép lai giữa các tế bào his+, leu+, thr+, pro+ , mẫn cảm với str và các tế bào his-, leu-, thr, pro -, kháng str, kéo dài 25 phút. Khi ngừng tiếp hợp, các kiểu gen có tái tổ hợp được xác định. Kết quả thu được ở bảng dưới đây. Hãy xác định các gen đầu tiên và trật tự có thể của các gen.
Nhận xét thấy các tế bào thr+ nhiều nhất. Tần số đứt gãy ống tiếp hợp tăng với sự tăng thời gian tiếp hợp. Vì vậy các gen ở càng xa điểm khởi đầu truyền gen sẽ xuất hiện với tần số càng nhỏ. Chúng ta có thể xắp xếp các gen một cách tương đối dựa trên tần số của các kiểu gen phát hiện được. Trật tự đó là thr leu pro his. Vì không thấy có his+ và vì chúng ta dừng quá trình tiếp hợp ở 25 phút, nên gen his phải nằm sau 25 phút trên bản đồ thời gian.
5. Các gen của virut thường được xác định trật tự bằng cách tương tự như việc lập bản đồ di truyền ở eukaryote.
Ví dụ: E. coli được gây nhiễm một cách lẫn lộn với hai loại phage T4: r+ m+ tu+ và r m tu. Đời con xuất hiện như sau: +++ 1243 ++ tu 322
Để kiểm tra lại, hãy xác định các trao đổi chéo kép để tạo thành các thể tái tổ hợp r+m tu và r m+tu+ Ta thấy rằng chỉ có các alen r bị thay đổi do sự sắp xếp lại từ bố mẹ, vì vậy r phải nằm giữa.
6. Thiếu sự bổ trợ nghiã là hai đột biến thuộc một gen.
Xét hai trường hợp sau: I: II: gen 1 gen 1 gen 2 Trong trường hợp I, cả hai phân tử ADN cùng tạo ra một protein không có chức năng. Tế bào không có protein bình thường cho thấy rằng nó có kiểu hình đột biến. Trong trường hợp II, ADN phía trên có gen 2 bình thường và ADN bên dưới có gen 1 bình thường. Tế bào này sẽ có cả protein 1 và protein 2 bình thường và sẽ xuất hiện kiểu hình bình thường. Sự sinh trưởng được của tế bào, hoặc chính là sự bổ trợ cho nhau, cho thấy rằng các đột biến này thuộc những đơn vị chức năng khác nhau, hay thuộc các gen khác nhau.
Ví dụ: Xét số liệu về hiện tượng bổ trợ sau. Bao nhiêu gen có liên quan đến kiểu hình đột biến và đột biến nào thuộc về gen nào ? (+: bổ trợ, và -: khuyết dưỡng).
Tính khoảng cách giữa các gen và vẽ bản đồ di truyền.
HD : Xét cặp r-m, hãy xác định các lớp mà trong đó r+m+ không cùng xuất hiện.
RF r-m = (175 + 54 + 58 + 160)/3454 x 100 = 13 đơn vị bản đồ
Tương tự, RF r-tu = (322 + 284 + 54 + 58) x 100/3454 = 21 đơn vị bản đồ.
RF m-tu = (322 + 284 + 175 + 160)/3454 x 100 = 27 đơn vị bản đồ.
Từ kết quả trên ta có bản đồ di truyền là:
Hãy tìm các tổ hợp trong đó không có sự bổ trợ. Các lớp này sẽ chứa các đột biến thuộc cùng một gen. 1 và 4 không bổ trợ và vì vậy nó thuộc một gen (gen A). Bằng lập luận tương tự, 2 và 3 thuộc một gen khác (gen B). Đột biến 5 bổ trợ với tất cả các đột biến khác và vì vậy nó phải thuộc một gen khác với gen chứa 1, 4 hoặc 2, 3 (gen C).
7. Nếu một đột biến không bổ trợ cho các đột biến ở hai gen khác nhau thì đột biến mới này chắc chắn là một mất đoạn.
Ví dụ: Xét một đột biến mới số 6 cùng với các đột biến ở khái niệm F. Nó cho những kết quả bổ trợ sau. Bản chất của đột biến số 6 là gì?
Chúng ta thấy đột biến số 6 không bổ trợ cho 1 và 4, cho phép ta nghĩ rằng đột biến số 6 thuộc gen A. Nhưng đột biến 6 cũng không bổ trợ với đột biến 5, đó là đột biến ta đã biết thuộc một gen khác. Đột biến 6 ảnh hưởng đến cả gen chứa đột biến 1 và 4 và cả gen chứa đột biến 5. Điều đó cho thấy đột biến này nằm ở gen khác.
Một cách giải thích cho kết quả này là có một mất đoạn bao trùm những vùng có các đột biến 1 và 4 và vùng có đột biến 5. Hoặc đột biến 6 có thể là một đột biến kép với mỗi đột biến của nó nằm trong một gen.
8. Nếu hai mất đoạn có chung một phần bị mất (nghĩa là chúng trùm lên nhau) thì chúng không thể cho ra các thể tái tổ hợp kiểu dại.
Xét các khả năng sau:
a. Hai mất đoạn không trùm lên nhau:
Sự tái tổ hợp sẽ tạo ra:

ADN mang hai mất đoạn:
và ADN bình thường _____________________
b. Hai mất đoạn trùm lên nhau:
Tái tổ hợp sẽ tạo ra các ADN đều có đoạn bị mất.
Ví dụ: Các chủng bị mất đoạn dưới đây được lai với nhau và chọn ra các thể tái tổ hợp kiểu dại. Kết quả như sau (+: có các thể tái tổ hợp kiểu dại, - không có các thể tái tổ hợp kiểu dại). Hãy xác định trật tự của các mất đoạn.


9. Nếu một đột biến điểm cho ra các thể tái tổ hợp khi lai với một mất đoạn, thì đột biến điểm đó nằm ngoài vùng bị mất đoạn.
Xét phép lai sau giữa một mất đoạn và một đột biến điểm:
Trường hợp 1: điểm đột biến ở bên ngoài đoạn mất
Sự tái tổ hợp sẽ tạo ra một AND mang đột biến kép:
và một AND bình thường:
Trường hợp 2: Đột biến nằm bên trong đoạn mất
Sự tái tổ hợp sẽ không xảy ra trong vùng có mất đoạn. Do vậy tất cả các thể tái tổ hợp vẫn là đột biến.
Ví dụ: Các thể đột biến điểm 1, 2, 3 được lai với các chủng có mất đoạn a và b; người ta chọn được một số thể tái tổ hợp kiểu dại. Dựa trên kết quả dưới đây, hãy xác định vị trí của mỗi đột biến (+ = thể tái tổ hợp kiểu dại, - = thể tái tổ hợp đột biến).
1 2 3
a + - +
b - + +

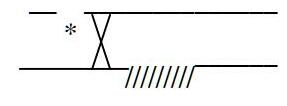

Đột biến 1 không sinh ra kiểu dại với mất đoạn b, vì vậy nó phải nằm bên trong vùng tương ứng với b.
Lập luận tương tự, ta có đột biến 2 nằm trong vùng tương ứng với mất đoạn a.
Chúng ta thấy rằng chủng 3 không cho ra thể tái tổ hợp kiểu dại nào. Nó phải mang mất đoạn kéo dài ít nhất đến phần đoạn bị mất ở chủng 1 và 2. Các mất đoạn ở chủng 1 và 2 cho ra các thể tái tổ hợp kiểu dại, vì vậy nó không trùm lên nhau.
Vậy bản đồ có thể là:
Vì đột biến 3 cho các dạng dại với cả hai mất đoạn, nên nó phải nằm ngoài cả hai đột biến mất đoạn.
10. Các đột biến ở tận cùng của operon hoặc các đột biến ở vùng kiểm soát có thể cho ra những kết quả âm tính giả trong phép thử bổ trợ.
Những đột biến ở tận cùng của operon làm mất chức năng của các gen ở xa trong một operon.
Ví dụ: Xét một operon gồm 3 gen theo thứ tự A, C, B. Một thể lưỡng bội một phần có kiểu gen a - (ở tận cùng của operon) b+ c+/ a+bc+ được thiết kế. Kiểu hình của chủng này là gì?


Đột biến ở cực a - làm cho phân tử ADN thứ nhất có biểu hiện thành a -bc. Chủng thứ 2 có kiểu hình b. Thể lưỡng bội một phần sẽ có kiểu gen b-/b, vậy nó sẽ là thể đột biến.
11. Các mất đoạn thường không phục hồi lại được.
Nếu là một đột biến nhầm nghĩa, thì bazơ bị biến đổi có thể thay đổi thành bazơ bình thường. Trường hợp này sẽ không xảy ra nếu một số bazơ cùng bị mất.
Ví dụ: Có 5 thể đột biến trp- kí hiệu từ 1 đến 5 được nuôi trên môi trường tối thiểu. Một số khuẩn lạc xuất hiện ở chủng 2 và 5. Bản chất của mỗi đột biến này là gì ?
Chắc chắn một số ít khuẩn lạc đã biểu hiện sự phục hồi, do vậy các chủng đột biến 2 và 5 phải là những đột biến điểm. Các chủng đột biến 1, 3 và 4 có thể là những mất đoạn.
12. Có thể tách những thể hồi biến chịu ức chế nhờ tái tổ hợp.
Những thể hồi biến gồm hai loại: những thể hồi biến thật và những thể hồi biến do các đột biến ở vị trí thứ 2 ức chế nó hoặc bù trừ cho đột biến ban đầu.
1. Thể đột biến bị đột biến tiếp, sẽ sinh ra thể hồi biến thật:
2. Thể đột biến bị đột biến tiếp sinh ra đột biến kép: có kiểu hình “bình thường”.
Khi một trong hai chủng này được lai với kiểu dại sẽ cho các kết quả khác nhau.
Trường hợp 1:
nhiễm sắc thể của thể hồi
biến thật
nhiễm sắc thể kiểu dại
Tái tổ hợp, sinh ra các con kiểu dại:
Trường hợp 2:
nhiễm sắc thể của thể hồi biến chịu ức chế
nhiễm sắc thể kiểu dại
Tái tổ hợp, tạo ra hai dạng đột biến:
Cả hai thể tái tổ hợp ở trường hợp 2 vẫn là dạng đột biến. Lưu ý rằng tình huống này cũng sẽ xảy ra, nếu đột biến thứ 2 nằm trong gen gốc hoặc nằm trong gen khác hoàn toàn cách biệt. Đột biến cũ cũng sẽ được khôi phục trong cả hai trường hợp đó.
Ví dụ: Ba thể hồi biến của leu- là r1, r2, r3 được khôi phục bình thường. Mỗi dạng này được lai với kiểu dại và kiểm tra đời con. Thể hồi biến 1 và 3 không cho ra thế hệ con kiểu dại. Bản chất của mỗi thể hồi biến đó ?
Vì các thể hồi biến 1 và 3 không tạo ra các thể tái tổ hợp kiểu dại, nên chúng phải chứa một đột biến ức chế ở vị trí thứ hai . Thể hồi biến 2 là thể hồi biến thật.
II. Bài tập tự luyện có hướng dẫn
Bài 1
a. Trong một thực nghiệm biến nạp, thể cho là ara+nic+gal+ và thể nhận là ara -nic-gal. Các thể biến nạp ara được chọn và sau đó kiểm tra tiếp. Kết quả cho thấy 5 % là ara+gal+ và 40% là ara+nic+ . Xác định trật tự có thể của các gen.
b. Bạn có thể chỉ cần làm thêm một thực nghiệm biến nạp để xác định trật tự gen. Bạn phải sử dụng cùng thể cho và thể nhận, nhưng có thể thay đổi quá trình chọn lọc các thể biến nạp đầu tiên. Bạn phải làm gì và kết quả như thế nào khi xác định trật tự gen mà bạn cần?
Hướng dẫn:
a. Trật tự có thể của các gen. ara nic gal hoặc nic ara gal.
b. Chọn lọc theo gal, sau đó đếm số khuẩn lạc gal+ara+ và gal+nic+ .
Nếu trật tự ara nic gal là đúng thì số khuẩn lạc gal+ ara+< gal+nic+ Vì nic và ara đồng biến nạp 40% thời gian nên chúng phải gần và liền kề nhau. Nếu gal ở giữa thì nic+gal+ phải có tần số đồng biến nạp lớn hơn nic+ara+
Nếu chúng ta chọn lọc theo gal+ và thứ tự là ara nic gal, thì kiểu gen gal+ara+nic- chỉ cần hai trao đổi chéo đồng thời, trong khi gal+ara+ chỉ đòi hỏi một trao đổi chéo. Nếu trật tự là nic ara gal thì gal+ ara+ chỉ cần một trao đổi chéo và tần số thu được phải lớn hơn gal+nic+ .
Bài 2.
ADN từ một chủng a+b+c+ được sử dụng để chuyển sang chủng a -bc - Số lượng từng kiểu gen xuất hiện dưới đây. Hóy xác định vị trí tương đối của các gen.
Có lẽ thể đột biến 3 chứa một mất đoạn trùm lên các gen A và C. Hãy bắt đầu bằng việc tìm ra các đột biến không bổ trợ cho nhau. Các đột biến này phải trong cùng một gen. Các đột biến 2 và 5 trong cùng một gen. Đầu tiên chúng ta ngờ rằng các đột biến 1, 3, 4, 8 trong một gen và 6, 7 trong một gen khác. Nếu thể đột biến 3 thuộc gen A nó phải với 6 và 7. Điều đó không xảy ra.
Cách giải thích khác là đột biến 3 là một mất đoạn trùm lên A và C. Khả năng khác là đột biến 3 có thể thuộc gen A nhưng là một đột biến cực. Cả hai điều đó đều ngụ ý nói rằng trật tự của 3 gen là B A C
Bài 4: Các chủng Hfr sau đây truyền các gen theo trật tự sau:
Hướng dẫn: Vị trí tương đối của các gen: a và b gần nhau, c không liên kết với a và b.
Chúng ta lưu ý rằng c+ ít xuất hiện cùng với các gen khác cho thấy c rất xa a và b. Các gen không liên kết sẽ không nằm cùng một phân tử AND. Sự đồng biến nạp sẽ đòi hỏi 2 trao đổi chất đồng thời.
Bài 3: 8 đột biến trp- độc lập nhau được phân lập. Tiến hành phép thử bổ trợ với các đột biến theo kiểu gép đôi. Dựa trên kết quả thu được dưới đây, bạn có thể xác định được bao nhiêu gen và đột biến nào thuộc gen nào? (+ = bổ trợ, - = không bổ trợ).

Hãy xác định trật tự các gen trên nhiễm sắc thể
Hướng dẫn:
Hãy sắp xếp theo trật tự để chúng trùm lên nhau:
Hướng dẫn: Xác định được 3 gen.
Gen A: các đột biến 1, 4, 8; Gen B: các đột biến 2 và 5; Gen C: các đột biến 6 và



Vì nhiễm sắc thể vi khuẩn có dạng vòng nên ta có trật tự biểu diễn ở hình trên.
Bài 5:
Bạn phân lập được hai thể đột biến rII là rII-1 và rII-2 ở phage T4. RII-1 không cho thể hồi biến kiểu dại và đột biến rII-2 cho 4 thể hồi biến là 2rva, 2rvb,
2rvc, 2rvd. Mỗi thể hồi biến được lai với kiểu dại cho kết quả sau (+ = thể tái tổ hợp hồi biến kiểu dại):


Bài 7
Operon trp ở E. coli cú trật tự sau: p E C D B A. Bạn tách được 3 thể

đột biến trp- độc lập nhau và tạo ra các thể lưỡng bội từng phần cho mỗi đột biến với p+ E+D+C-B+A+; không thể lưỡng bội từng phần nào có thể tổng hợp được tryptophan.
Ba vị trí khác nhau có thể có của các đột biến này là gì?
Hướng dẫn:
Đột biến ở C, đột biến ở promoter, hoặc đột biến cực ở E hoặc D.
a. Bản chất của đột biến rII-1 và rII-2?
b. Bản chất của mỗi thể hồi biến?
Hướng dẫn:
a rII-1 có thể là một mất đoạn và rII-2 là một đột biến điểm.
b. Có lẽ các thể hồi biến biến revb và revc chứa đột biến tại điểm thứ hai.
Sự thiếu vắng các thể hồi biến gợi ý ta nghĩ đến một mất đoạn, do vậy rII-1 có thể là một mất đoạn . Nếu các thể tái tổ hợp kiểu dại nào được phục hồi từ các thể hồi biến, thì các thể hồi biến này có thể là đột biến ức chế hoặc là các đột biến vị trí thứ hai.
Bài 6.
Trong một thí nghiệm tải nạp, thể cho là d+e+f+ , thể nhận là de -f; tiến hành chọn lọc theo f+. Kết quả thu được như sau, hãy xác định trật tự gen:
Vì đột biến không tạo ra sự bổ trợ với C- nên nó có thể thuộc cùng một một gen . Nếu thể đột biến là p – thì thể lưỡng bội là: p- E D C B A/ p+ E D C- B A. ADN bên trên có thể không tạo ra ARN và sẽ không có sự bổ trợ . Một kiểu đột biến khác cũng sẽ không bổ trợ là một đột biến cực trong gen nằm trước C
Bài 8 :
Có chủng his, ba trong số đó có những mất đoạn và 6 chủng có những đột biến điểm.
Cấy 9 chủng trên môi trường tối thiểu và đếm số khuẩn lạc mọc (+ = mọc được). Phage được nhiễm vào một trong 9 chủng và sau đó được sử dụng để tải nạp mỗi chủng mất đoạn. Các thể tải nạp từ mỗi phép lai được cấy trên môi trường tối thiểu và đếm các khuẩn lạc mọc (dấu + biểu thị thể tải nạp mọc được). Dựa trên kết quả sau, xác định:
a. Chủng nào là chủng đột biến mất đoạn.
b. Vị trí tương ứng của mỗi đột biến mất đoạn với mất đoạn khác ?
a. Vị trí tương ứng của 6 đột biến điểm.
Hướng dẫn:
Ta tính tần số đồng tải nạp
d+f+ = 7,95 %
e+f+ = 28,7 %
Trật tự cần tìm là : def.
Chúng ta thấy rằng f và d đòng tải nạp gen ít nhất. Vì vậy d có lẽ nằm xa f và đòi hỏi 2 trao đổi chéo đồng thời (trao đổi chất kép). Chúng ta thấy rằng e và f có tần số đồng tải nạp cao, do vậy e gần f hơn d
Hướng dẫn:
Nuôi cấy trong môi trường tối thiểu cho phép phát hiện các thể hồi biến. Vì các mất đoạn không phục hồi được nên chúng không mọc được trên môi trường tối thiểu (KN9-K). Đã biết 2, 5, 7 là những mất đoạn, ta hãy phân biệt các chủng này từ bộ số liệu thứ hai.
Chủng 2 không cho thể tái tổ hợp kiểu dại, vì vậy nó phải trùm lên hai
đột biến mất đoạn khác (KN 9-H). Các mất đoạn 5 và 7 không trùm lên nhau. Thể
đột biến 2 cho kết quả tương tự chủng A nên A=2. Vì B tái tổ hợp với 7, nên B phải là 5; do đó C là 7.






Bạn cũng lai mỗi đột biến điểm với mỗi đột biến mất đoạn và tính các thể tái tổ hợp kiểu dại. Kết quả thu được dưới đây (+ = thể tái tổ hợp kiểu dại, - = thể tái tổ hợp đột biến).
Thể đột biến 9 không cho các tái tổ hợp kiểu dại với các mất đoạn 2 và 7. Nó phải nằm ở vùng được định vị duy nhất trong đột biến 5 (KN 9-I). Lý luận tương tự đối với đột biến 8: 8 ở vùng duy nhất với 7, 3 ở vùng duy nhất với 2. Các đột biến 4 và 6 phải nằm trong vùng mất đoạn chung cho 2 và 7 cũng như cho 2 và 5 tương ứng. Đột biến 1 cho các thể tái tổ hợp kiểu dại với tất cả các mất đoạn, nó phải nằm
bên ngoài tất cả các vùng mất đoạn. Nó có thể nằm ở một trong hai đầu tận cùng
của bản đồ.
Vậy vị trí tương ứng :
a. Vẽ bản đồ các đột biến điểm, tính khoảng cách.
b. Xác định vị trí của các mất đoạn tương ứng với các đột biến điểm.
c. Các thể đột biến a và e bổ trợ với nhau và e được biết là thuộc rIIB. Các đột biến nào ở rIIA và đột biến nào thuộc rIIB?
d. Những tổ hợp của các mất đoạn nào sẽ tạo ra các thể tái tổ hợp kiểu dại?
Hướng dẫn:
a.b
Dựa vào bảng 1, ta xác định được các đột biến điểm khi được ghép đôi có tỉ lệ phần trăm thể tái tổ hợp kiểu dại càng cao chứng tỏ các gen càng nằm xa nhau
Vị trí của các mất đoạn tương ứng với các đột biến điểm
Bài 9:
Bạn phân lập được một số đột biến điểm (ký hiệu từ a đến g) ở vựng rII của phage T4. Các phép lai ghép đôi cho tỷ lệ phần trăm các vết tan kiểu dại như sau:
c. a, b, f ở trong rIIA; e, c, d và g ở trong rIIB.
d. Không tổ hợp nào.
Các đột biến b và d phải xa nhau, b và g cũng vậy, e và f phải nằm giữa b và d:
____ b ___0.2 ___ f __0.4 ___ e ____ d ___ 0.3 __ g
Bây giờ hãy xác định vị trí của a và c. Vì a-b = 0,5 nên a phải cách f là 0. 3 về bên phải. Đột biến c phải ở bên phải a, hoặc nó có thể cùng vị trí với f. Đột biến c phải ở bên phải a và khoảng cách được cộng kế tiếp. Hãy tìm các dấu (-) trong các phép lai; các đột biến này sẽ bị phủ bởi các mất đoạn riêng. Mất đoạn 1 phải kéo dài dến các đột biến a, c và e vì không có thể tái tổ hợp kiểu dại nào xuất hiện . Tương tự, đột biến 2 phải kéo dài đến c, d, g. Lý luận tương tự cho phép xác định các mất đoạn 3-5. Vì a bổ trợ e, vậy a phải thuộc rIIA, cũng giống như b và f . Tất cả các mất đoạn đều thiếu c, do vậy không thể sinh ra các thể tái tổ hợp kiểu dại .



Bài 11:
Có 3 thể đột biến lac- (ký hiệu từ a đến c). Bạn thực hiện cỏc thớ nghiệm tải nạp và thu được số khuẩn lạc kiểu dại như sau:
Xác định trật tự các đột biến lac so với gen pro
Hướng dẫn:
Cách 1 : trật tự cần tìm là : c a b pro.
Chúng ta phải xây dựng sơ đồ tải nạp, sử dụng 2 trật tự gen và tìm ra trật tự sẽ tạo ra và kiểu dại bằng trao đổi chất kép. Trật tự này sẽ cho ra một số ít thể tải nạp kiểu dại (KN 9-C). Giả sử rằng trật tự đó là b a pro. Những phép đầu là:
Nếu trật tự như trên, phép lai thứ nhất dự đoán có số lượng thể tái tổ hợp kiểu dại nhỏ hơn ở phép lai thứ 2 vì cần 2 cặp trao đổi chéo đồng thời cho phép lai 1. Kết quả này không thấy có, do vậy b phải nằm gần pro hơn a. Lý luận tương tự để tìm ra trật tự của c tương ứng với a và b.
Cách 2 :
Ta tính tần số đồng tải nạp
lac b+ pro+ = 62,2 %
lac a+ pro+ = 29,6 %
lac c+ pro+ = 8,2 %
trật tự cần tìm là: c a b pro.
Bài 12:
Thể đột biến phage 1 gây nhiễm vào chủng vi khuẩn B và D, thể đột biến 2 được gây nhiễm vào A và D. Chỉ phage kiểu dại mới sinh trưởng được ở chủng C. Cả hai chủng phage đột biến được sử dùng để lây nhiễm đồng thời vào D. Phage thế hệ
con được pha loóng 10-7 và cấy vào D, thu được 200 vết tan. Phage thế hệ con được pha loóng 10-6 và cấy vào chủng C thu được 30 vết tan. Sự tái tổ hợp giữa thể đột biến 1 và 2 là gì ?
Hướng dẫn:
Sự tái tổ hợp giữa thể đột biến 1 và 2 là 3%.
Chúng ta phải tính tổng số cá thể đời con và số con tái tổ hợp. Vì tất cả các phage sinh trưởng trong chủng D, nên số khuẩn lạc sinh trưởng được ở đây phải là tổng số cá thể đời con, đó là 200 x 107. Kiểu dại chỉ sinh trưởng ở chủng C, do vậy các vết tan này phải là dạng tái tổ hợp. Tuy nhiên, số lượng thu được chỉ bằng 1/ 2 tổng số thể tái tổ hợp, vì các thể đột biến kép sẽ không sinh trưởng được.
Số thể tái tổ hợp là 2 x 30 x106 = 60 x 106. Suy ra
RF = (60 x 106)/(200 x 107) x 100 = 3% .
III. Bài tập trong các đề thi HSG
Bài 1:
Chủng vi khuẩn lys+his+val+ được dùng làm thể cho, còn chủng lys-his-vallà thể nhận. Các thể biến nạp đầu tiên được tách ra trên môi trường tối thiểu có chứa his và val.
a. Những kiểu gen nào sẽ sinh trưởng trên môi trường này?
b. Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + his, 75% các khuẩn lạc ban đầu sinh trưởng được. Những kiểu gen nào sẽ sinh trưởng được trên môi trường này?
c. Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + val, 6% số khuẩn lạc sinh trưởng được. Những kiểu gen nào sinh trưởng được trên môi trường này?
d. Cuối cùng, cho các khuẩn lạc này sinh sản trên môi trường tối thiểu. Không khuẩn lạc nào sinh trưởng được. Dựa vào thông tin này, xác định xem kiểu gen nào sinh trưởng được trên môi trường tối thiểu + his? trên môi trường tối thiểu + val?
e. Dựa trên những thông tin trên, tìm xem gen nào nằm gần gen lys
f. Thí nghiệm biến nạp được lặp lại, song đĩa nuôi cấy có chứa môi trường tối thiểu + lys + his. 50% khuẩn lạc sống được. Cho các khuẩn lạc này sinh sản trên các môi trường khác nhau để xác định kiểu gen của chúng. Kết quả như sau:
val-his+lys+ 0
val+his-lys+ 37
val+his+lys- 3
Dựa vào kết quả trên, hãy xác định trật tự các gen.
Hướng dẫn:
a. Những kiểu gen sẽ sinh trưởng trên môi trường này là lys+ his+ val+ lys+ his+ vallys+ his- val+ lys+ his- val-
Ki
ểu gen nào có lys+ mới mọc được vì Các thể biến nạp được tách ra trên môi trường tối thiểu có chứa his và val.
b. Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + his, 75% các khuẩn lạc ban đầu sinh trưởng được. Những kiểu gen sẽ sinh trưởng được trên môi trường này là các kiểu gen nào có chứa lys+ và val+
Gồm lys+ val+ his- và lys+ val+ his+
c. Cho các khuẩn lạc này sinh sản trên môi trường tối thiểu + val, 6% số khuẩn lạc sinh trưởng được. Những kiểu gen sinh trưởng được trên môi trường này là kiểu gen phải có có lys+ và his+
Gồm lys+ val+ his+ và lys+ val- his+
d. Khi cho các khuẩn lạc này sinh sản trên môi trường tối thiểu. Không khuẩn lạc nào sinh trưởng được.
+ Kiểu gen nào sinh trưởng được trên môi trường tối thiểu + his là lys+ val+ his-
+ Kiểu gen nào sinh trưởng được trên môi trường tối thiểu + val là lys+ valhis+
Chúng ta không thấy có kiểu gen lys+ val+ his+
e. Ta có tần số đồng biến nạp của lys+ val+ là 75 %,
+ tần số đồng biến nạp của lys+ his+ là 6 %,
Vậy lys+ và val+ nằm gần nhau ( chúng đồng biến nạp 75% thời gian)
Trật tự gen có thể là lys val his hoặc val lys his
f. Ta thấy các khuẩn lạc mọc được trên môi trường này chắc chắn phải có val+ , vậy chọn lọc theo val+
Tính tần số đồng biến nạp có
val+ lys+ 3 7
Val gần lys
Val xa his
Vậy trật tự gen có thể là : Val ly his
Cách 2 . Dựa vào ý e
*Giả sử trật tự là lys val his
Thì khi đó sau khi BN kiểu gen lys- val+ his+ sẽ đc hình thành do trao đổi chéo đơn nên là vô lý vì 3 < 37
*Giả sử trật tự là val lys his.
Thì khi đó sau khi BN kiểu gen val+ lys- his+ sẽ đc hình thành do trao đổi chéo kép nên là hợp lý vì 3 < 37 ( phù hợp là hiếm gặp vì nó được tạo ra do trao đổi chéo kép)
Bài 2
Đột biến RII ở phagơ T4 sẽ sinh trưởng và tạo ra những vết tan lớn trên chủng vi khuẩn B; các đột biến rII sẽ không sinh trưởng trên K12 - một thể tiềm tan lambda. Các phép lai sau đây được thực hiện ở chủng B. Đời con của các phép lai được làm loãng đến nồng độ 10-7 và được sử dụng để tái nhiễm chủng B. Dưới đây là số lượng các vết tan thu được:
Bài 3
Mr Long có 10 chủng vi khuẩn Escherichia coli mang các đột biến khác nhau nằm trên operon lac của các vi khuẩn này. Khi phân tích ADN của vi khuẩn, MrLong thấy rằng mỗi chủng mang một trong 5 đột biến: lacZ- , lacY- , lacI- , lacIs (lacIs mã hóa LacIs là chất ức chế có thể bám vào operator nhưng không thể bám vào chất cảm ứng), hoặc lacOc (Operator của operon lac đột biến làm chất ức chế không thể bám vào nó). MrLong cũng biết rằng chủng số 6 là một thể đột biến lacZ- . Cấu trúc operon lac được mô tả như hình dưới đây.


Đời con của các phép lai trên cũng còn được làm loãng đến nồng độ 10-4 và cấy lên K12. Số lượng các vết tan thu được như sau:
Mr Long phân lập được đoạn ADN mang operon lac từ mỗi chủng (gọi là chủng cho) và biến nạp đoạn ADN này vào các chủng khác, tạo ra chủng lưỡng bội từng phần (gọi là chủng nhận). Sau đó, các chủng nhận được nuôi trên môi trường tối thiểu chứa lactose là nguồn carbon duy nhất . Sự sinh trưởng của các chủng được ghi lại trong ảnh dưới đây (+ biểu thị chủng đang sinh trưởng, - biểu thị chủng không sinh trưởng được).
Hãy cho biết câu sau đây là Đúng hay Sai và giải thích
A. Chủng số 7 là thể đột biến lac Z .
Hãy vẽ bản đồ các đột biến này và chỉ rõ khoảng cách giữa chúng.
Hướng dẫn:
Các vết tan trên B cho ta biết tổng số các khuẩn lạc đời con. Các vết tan trên
K12 là các thể tái tổ hợp kiểu dại vì chỉ có kiểu dại mới mọc được trên K12. Tuy nhiên số lượng
này chỉ là 1/2 số thể tái tổ hợp, 1/2 còn lại là đột biến kép.
200

B. Chủng số 3 là thể đột biến lac Y .
C. Chủng số 2 và số 4 mang cùng loại đột biến.
D. Nếu chủng số 5 nhận được ADN của chính nó thì thể biến nạp này không thể sinh trưởng được.

Hướng dẫn: A, B,C đúng.

- Mỗi chủng
mang 1 loại đột biến
+ Có Lac Is: VK không sinh trưởng
+ Có Lac Oc: VK luôn sinh trưởng
+ Lac I-: Sinh trưởng bình thường
+ Lac Z- không sinh trưởng
+ Lac Y- không sinh trưởng
Nếu ở thể dị hợp: Lac Z Lac Z- và Lac Y Lac Y-: sinh trưởng bình thường
- Chủng 6 đột biến Lac Z
+ Chùng 7 có tác động giống hệt chủng 6, khi biến nạp với chủng 6 => không sinh trưởng => chủng 7 là đột biến Lac Z-
- Chủng 5 và chủng 9 có tác động giống hệt nhau, dù biến nạp với chủng nào vi khuẩn cũng luôn sinh trưởng => chủng 5, 9 đột biến Lac Oc
- Chủng 1 và chủng 8 có tác động giống hệt nhau, biến nạp với các chủng khác thì đều không sinh trưởng (trừ biến nạp với chủng 5, 9) => chủng 1, 8 đột biến Lac Is
- Chủng 2 và chủng 4 có tác động giống hệt nhau, biến nạp với các chủng khác đều sinh trưởng được (trừ biến nạp với chủng 1, 8) và khi biến nạp chủng 2 với chủng 4 thì vẫn sinh trưởng được => chủng 2, 4 đột biến Lac I-
- Chủng 3, 10: có tác động giống nhau, mà khi biến nạp 3 với 10 thì ko sinh trưởng => chủng 3, 10 đột biến Lac Y-
Bài 4:
Mỗi một dòng trong số 5 dòng đột biến của E. coli (1-5) có một đột biến ảnh hưởng đến operon lac . Đột biến 1 có một đột biến trong gen lac Y và các dòng đột biến khác mỗi cái lại có một đột biến như sau:
Đột biến vô nghĩa trong gen lacZ tạo ra β-galactosidase không có chức năng
Đột biến lacOc làm cho chất ức chế không thể liên kết vào operator (vùng vận hành)
Đột biến ở promoter làm cho RNA polymerase không thể liên kết với promoter.
Đột biến siêu ức chế làm lactoz không thể liên kết và làm bất hoạt protein ức chế. Operon lac của mỗi thể đột biến được chèn vào một plasmit rồi cho biến nạp vào tế bào kiểu dại và tất cả các dòng E. coli đột biến cùng khả năng sinh trưởng trong môi
trường chứa lactose đều được đánh giá. Dấu + chỉ khả năng sinh trưởng tốt, còn dấu –chỉ không có khả năng sinh trưởng. Sự sinh trưởng đòi hỏi phải có cả hai gen LacZ và LacY có chức năng.
Hãy cho biết câu nào dưới đây là đúng, câu nào sai dựa trên kiểu đột biến ở thể đột biến 2-5
A. Thể đột biến 2 có đột biến LacOc
B. Thể đột biến 3 có đột biến ở promotor
C. Thể đột biến 4 có đột biến siêu ức chế
D. Thể đột biến 5 có đột biến vô nghĩa ở
LacZ
Hướng dẫn:
A, C đúng
- Mỗi chủng mang 1 loại đột biến
- Có Lac Is: VK ko sinh trưởng
- Có Lac Oc:VK luôn sinh trưởng
- ĐB ở promoter: VK ko sinh trưởng
- Có Lac Z- hoặc Lac Y-: không sinh trưởng
Nếu ở thể dị hợp: Lac Z Lac Z- và Lac Y Lac Y-: sinh trưởng bình thường
- Chủng 1: Lac Y-
- Chủng 2: biến nạp với 1 và 2 đều luôn sinh trưởng => chủng 2 ĐB Lac Oc
- Chủng 5: luôn ko sinh trưởng trừ khi biến nạp với chủng 2 => ĐB ở promoter
4: luôn ko sinh trưởng trừ khi biến nạp với chủng 2 => ĐB siêu ức chế (Is)
- Chủng
- Chủng 3: ĐB ở lac Z
Bài 5.
Hiện tại, bạn đang nghiên cứu về sự tổng hợp threonin ở E.coli. Để tìm các gen mã hóa các protein quan trọng cho tổng hợp threonin, bạn tiến hành gây đột biến để tìm các đột biến yêu cầu bổ sung threonin cho phát triển. Bạn đã tìm thấy có 9 dòng
đột biến và tất cả đều cần bổ sung threonin vào môi trường nuôi cấy để phát triển. Sau đó bạn tiến hành cis-trans test với tất cả các dòng đột biến bạn có. Kết quả thu được như bảng sau (+: phát triển; -: không phát triển):


Hãy xác định trật tự chuyển hóa của các chất A, B, C, D, E và các gen mã hóa các enzym xúc tác sự chuyển hóa tương ứng. Giải thích.
Hướng dẫn:
a) Ta thấy:
- Khi cho các dòng đột biến giống nhau lai với nhau => Không phát triển, không tạo khuẩn lạc.
=> Đột biến trên cùng một gen nên không bù trừ được cho nhau. => Tạo kiểu hình đột biến.
- Khi cho những dòng đột biến khác nhau lai với nhau:
+ Nếu khuẩn lạc phát triển => Hai đột biến thuộc hai gen khác nhau bù trừ cho nhau tạo kiểu gen dị hợp => Tạo kiểu hình bình thường (kiểu dại). + Nếu khuẩn lạc không phát triển
=> Hai đột biến thuộc cùng một gen nên không bù trừ cho nhau
=> Tạo kiểu gen đồng hợp
=> Tạo kiểu hình đột biến.
a. Có bao nhiêu gen mã hóa các enzym tham gia vào quá trình tổng hợp threonin? Giải thích.
b. Khi xem xét quá trình tổng hợp threonin, người ta nhận thấy có 5 cơ chất tham gia vào quá trình đó. Người ta tiến hành nuôi cấy các đột biến ở trên trong các môi trường chứa các cơ chất khác nhau để xác định trình tự quá trình biến đổi. Kết quả thu được trình bày ở bảng sau (+: phát triển; -: không phát triển):
Từ đó, ta có:
- 1,3,5,6,9 thuộc cùng một gen.
- 2,8 thuộc cùng một gen.
- 4 không bù cho mình, bù cho những dòng đột biến còn lại => gen độc lập
- 7 không bù cho mình, bù cho những dòng đột biến còn lại
=> gen độc lập
=> Có 4 gen tham gia tổng hợp threonin.
b) sơ đồ chuyển hóa:
Giải thích
- Các chủng đột biến không sinh trưởng được trong môi trường chỉ chứa
A => A là chất đầu dãy chuyển hóa
- Chỉ có chủng đột biến Mt 1 sinh trưởng được trong môi trường chỉ chứa
D => D là chất thứ 2 của dãy chuyển hóa và chủng Mt 1 bị đột biến ở bước chuyển hóa chất A thành chất D
- Trong môi trường chỉ chứa C, có hai chủng Mt 1 và Mt 7 sống được => chất C là chất thứ 3 của dãy chuyển hóa và chủng Mt 7 là chủng bị đột biến ở bước chuyển hóa chất D thành chất C
- Chủng Mt 4 sống được trong môi trường chỉ chứa B và E => chất B và
E cùng là chất thứ 4 trong dãy chuyển hóa và chủng Mt 2 bị đột biến ở bước chuyển hóa chất C thành chất B hoặc E.
- Chủng Mt 2 chỉ sống được trong môi trường chứa Threonin => chủng này bị đột biến ở bước chuyển hóa chất B hoặc chất E thành Threonin
Bài 6 - V1 năm 2016
Đa số vi khuẩn không chuyển hóa được xylitol để tạo năng lượng. Người ta tiến hành thí nghiệm chuyển một gen mã hóa enzim phân giải xylitol vào E. coli. Một dòng plasmit làm thể truyền được biến đổi để có một gen kháng ampicilin (ampR) và một gen mã hóa β galactozidaza có chứa trình tự nhận biết của enzim cắt giới hạn EcoRI (β galactozidaza có khả năng phân giải X-gal từ không màu thành sản phẩm màu xanh dương).
Sau khi đoạn ADN chứa gen mã hóa enzim phân giải xylitol và thể truyền được xử lý riêng rẽ với EcoRI, chúng được trộn với nhau để tái tổ hợp ngẫu nhiên.
Hỗn hợp ADN này tiếp tục được cho biến nạp vào một chủng E. coli đã bị đột biến bất hoạt gen Z trong opêron Lac. Sau đó đem cấy trải trên môi trường thạch tối thiểu thu được 3 dòng A, B và C. Tiếp tục nuôi cấy riêng rẽ 3 dòng A, B và C trên môi trường thạch tối thiểu có bổ sung thêm ampicilin và X-gal thu được kết quả như
sau:
Dòng A: Xuất hiện khuẩn lạc và không thay đổi màu.
Dòng B: Xuất hiện khuẩn lạc và vùng xung quanh khuẩn lạc chuyển sang màu xanh dương. Dòng C: Không xuất hiện khuẩn lạc.
a) Giải thích kết quả thí nghiệm.
b) Nếu cả 3 dòng A, B và C được nuôi cấy riêng rẽ trong môi trường chỉ chứa xylitol làm nguồn cacbon duy nhất thì khả năng phát triển của mỗi dòng sẽ như thế nào? Giải thích.
Hướng dẫn
a) Giải thích kết quả thí nghiệm:
Dòng
A phát triển được trên môi trường có ampicilin → có plasmit chứa ampR. Khi nuôi trên X- gal thì màu của môi trường không thay đổi → gen β galactosidaza không
được biểu hiện → có hiện tượng biến nạp plasmit tái tổ hợp.
Dòng
B phát triển được trên môi trường có ampicilin và chuyển hóa được X-gal làm môi trường chuyển sang màu xanh → có hiện tượng biến nạp plasmit không tái tổ hợp.
- Dòng C không phát triển trên môi trường có ampiclin → không có hiện tượng biến nạp.
b) Khả năng phát triển của 3 dòng khi nuôi cấy trên môi trường có xylitol làm nguồn cacbon duy nhất:
- Dòng A có biến nạp plasmit tái tổ hợp tuy nhiên không thể chắc chắn plasmit tái tổ hợp này có gen chuyển hóa xylitol. Do đó:
+ Dòng A có thể phát triển được trên môi trường có chứa xylitol nếu plasmit tái tổ hợp có chứa gen chuyển hóa xylitol hoàn chỉnh.
+ Dòng A sẽ không phát triển được trên môi trường chỉ chứa xylitol nếu plasmit tái tổ hợp không chứa gen chuyển hóa xylitol hoàn chỉnh.
- Dòng B có khả năng sử dụng lactozơ (do có gen β galactosidaza trên plasmit hoạt động), nhưng không có khả năng sử dụng xylitol nên không phát triển được trên môi trường chỉ chứa xilytol.
- Dòng C không có biến nạp → không chuyển hóa được xilitol → không phát triển được trên môi trường có chứa xylitol.
Bài 7 - V2 năm 2010
Các câu hỏi thuộc phần này liên quan đến 2 thí nghiệm được mô tả dưới đây. hai thí nghiệm này được tiến hành để kiểm tra khả năng gây đột biến và gây ung thư của ba hợp chất x, y và z.
Thí nghiệm 1 Kiểm tra khả năng gây đột biến
Trong thí nghiệm này, mỗi chất được bổ sung vào môi trường nuôi cấy chủng vi khuẩn mang gen His- bị đột biến làm mất khả năng tổng hợp axit amin histidin. Vì thế, chủng vi khuẩn này chỉ sinh trưởng được trong môi trường có histidin. Tuy vậy, các hợp chất gây đột biến có thể biến đổi gen His- thành gen His+có khả năng tổng hợp histidin. Chất thử nghiệm có hiệu lực gây đột biến càng cao thì càng có nhiều khuẩn lạc hình thành trên môi trường nuôi cấy không bổ sung histidin (vì mật độ vi khuẩn giữa các lô thí nghiệm và đối chứng là như nhau).
Trong thí nghiệm bổ sung 2 mg của mỗi chất X và Y và 3 mg của chất Z vào 2ml dung dịch nuôi cấy vi khuẩn không có histidin rồi cấy vi khuẩn His- lên đĩa Petri (hộp lồng), kết quả thu được như hình dưới đây:
1. 2 mg hợp chất X khi được bổ sung vào môi trường đã kích thích tạo ra bao nhiêu khuẩn lạc?
2. Hệ số gây đột biến (mI) được tính bằng công thức dưới đây
Số khuẩn lạc thí nghiệm – Số khuẩn lạc đối chứng
mI = Liều lượng (mg) × Thể tích dịch tế bào vi khuẩn (ml)
Từ kết quả thí nghiệm, hãy cho biết hệ số gây đột biến của chất Z là bao nhiêu.
3. Hãy xếp thứ tự 3 hợp chất X, Y và Z theo trật tự giảm dần về hiệu lực gây đột biến dựa theo hệ số mI.
Thí nghiệm 2. Kiểm tra khả năng gây ung thư
Trong thí nghiệm này, mỗi chất thử nghiệm được hòa tan vào dung dịch sinh lý chứa 0,9% NaCl rồi được tiêm dưới da riêng từng chất vào các nhóm chuột lấy từ dòng thuần chủng. Tỉ lệ chuột trong mỗi nhóm phát triển thành khối u (tỉ lệ mắc phải)
c ghi nhận sau 6 tháng. Kết quả thí nghiệm nêu ở bảng dưới:
4. Hãy vẽ đồ thị biểu diễn mối quan hệ giữa liều tiêm và tỉ lệ chuột mắc khối u đối với hai chất X và Y trên cơ sở các số liệu thu được từ thí nghiệm 2 cùng với các chú thích cần thiết vào bài làm.
5. Khoảng nồng độ nào thì chất Y có hiệu lực gây phát sinh khối u cao hơn so với chất X?
6. Chất X được chuẩn bị ở nồng độ dung dịch gốc là 20 mg/ml. Cần thể tích dung dịch gốc và dung dịch sinh lý là bao nhiêu để tạo ra liều tiêm 2 ml chứa 16 mg chất X?

7. Phương thức đối chứng trong thí nghiệm 2 này là gì?
8. (0,2 điểm) Có thể đưa ra nhận định gì về chất Z trên cơ sở các kết quả thu được từ hai thí nghiệm 1 và 2?
Hướng dẫn
1. 20 khuẩn lạc
2. Hệ số gây đột biến của chất Z là 4,5
3. Thứ tự là : Y > X > Z
4. Đồ thị
5. Từ 8 đến 16 mg

6. 1,6 ml dd gốc + 0,4 ml dd sinh lý 0,9% NaCl
7. Chuột được tiêm dd sinh lý 0,9% NaCl
8. Chất Z là một chất gây đột biến, song vào cơ thể chuột có thể đã được chuyển hóa rất nhanh thành một hợp chất không độc (được
giải độc) và do vậy mất khả năng gây đột biến và không gây ung thư
Bài 8 : V2 năm 2008
Có 6 chủng vi khuẩn e. coli (kí hiệu 1 - 6) mang đột biến ở các gen khác nhau nhưng đều liên quan đến một con đường chuyển hóa trong tế bào. khi nuôi cấy các chủng vi khuẩn này trên các môi trường bổ sung chọn lọc các chất chuyển hóa trung gian là a, b, c, d, e và f, thu được kết quả như sau
Ch ủng vi khuẩn
Chất chuyển hóa trung gian được bổ sung chọn lọc vào môi trường a b c d e f
1 0 0 + 0 + 0
2 0 0 0 + 0 0
3 0 + 0 0 0 0
4 0 0 + 0 + 0
5 0 0 0 0 + 0
6 + 0 + 0 + 0
Trong đó, 0 là chết, + là sống và sinh trưởng bình thường.
Biết rằng tất cả các chất chuyển hóa trên đều thấm vào tế bào dễ dàng như nhau; mỗi chủng chỉ mang một đột biến gen duy nhất. tất cả các đột biến chỉ ảnh
hưởng đến các bước chuyển hóa sau khi f đã hình thành. sơ đồ nào dưới đây phù hợp nhất để phản ánh quá trình sinh tổng hợp các chất nêu trên ?
Đáp án: D
Bài 9: V2 năm 2012
Để xác định chất X có phải là tác nhân đột biến ở người hay không, một nhà nghiên cứu đã tiến hành xử lý chất X trên vi khuẩn Salmonella. Thí nghiệm được tiến hành như sau: cấy 109 tế bào vi khuẩn Salmonella lấy từ một chủng vi khuẩn không có khả năng tổng hợp histidine lên một đĩa petri thạch đặc chứa môi trường tối thiểu không có histidine, ở giữa đĩa petri có đặt một khoanh giấy thấm nhỏ tẩm dịch chiết gan lợn tươi cùng với chất X. Sau đó đĩa petri được đưa vào buồng nuôi cấy một thời gian để xem có khuẩn lạc nào xuất hiện trên đĩa petri hay không.
a) Cần phải bố trí thí nghiệm đối chứng như thế nào cho thí nghiệm trên? Giải thích.
b) Tại sao người ta lại dùng dịch chiết gan lợn tươi?
c) Nếu X thực sự là tác nhân đột biến thì tần số đột biến được tính bằng cách nào và đột biến xảy ra là loại đột biến gì? Giải thích.
d) Ngoài đặc điểm không có khả năng tổng hợp histidine, chủng vi khuẩn Salmonella dùng trong thí nghiệm trên còn phải có thêm một đột biến gì khác nếu không sẽ khó đánh giá được chất X là tác nhân đột biến khi tần số đột biến do nó gây ra là khá thấp?
TRẢ LỜI:
a) Thí nghiệm đối chứng được bố trí như sau: một đĩa petri thạch chứa môi trường tối thiểu không có histidine, được cấy 109 tế bào vi khuẩn khuyết dưỡng histidine, và ở giữa đĩa petrri cũng được đặt khoanh giấy thấm tẩm một lượng dịch chiết gan lợn tươi y hệt như ở lô thí nghiệm.
b)
- Gan có nhiều enzym chuyển hoá có thể chuyển hoá chất độc khi con người ăn vào thành chất không độc hoặc bớt độc, hoặc cũng có thể biến một chất thành một chất độc.
- Ta có thể dùng gan lợn vì lợn là loài động vật có vú có rất nhiều gen giống với các gen của người.
c) Tần số đột biến:
- Tần số đột biến được tính bằng cách đếm số lượng khuẩn lạc trên đĩa thí nghiệm rồi trừ đi tổng số khuẩn lạc trên đĩa đối chứng.
- Đột biến do chất X tạo ra là các loại đột biến ngược phục hồi khả năng tổng hợp histidine. Loại đột biến này nhiều khả năng là đột biến thay thế nucleotid.
d)
209
- Chủng vi khuẩn cần có thêm đột biến làm ức chế enzym sửa sai trong quá trình tổng hợp ADN. Nhờ vậy tần số đột biến sẽ gia tăng nên chúng ta dễ dàng phát hiện ra các đột biến cho dù chất X có thể là tác nhân đột biến yếu.
Bài 10: V2 năm 2012
Một chủng vi khuẩn pneumoccoci kháng penicilin tồn tại được dù các tế bào của chúng không có gen beta lactamase là gen có vai trò giúp vi khuẩn kháng penicilin. Ngoài ra, một nửa số tế bào của chủng vi khuẩn này không có khả năng chuyển hoá đường đôi sacarozơ và đường lactozơ. Tất cả các tế bào của chủng vi khuẩn này đều bị nhiễm Phagơ I và Phagơ II và cả hai đều chèn ADN của chúng vào nhiễm sắc thể của vi khuẩn. Để xác định xem liệu việc nhiễm Phagơ có thể tạo ra chủng vi khuẩn mới này hay không người ta đã tiến hành gây nhiễm chủng dại với hai Phagơ này.
Thí nghiệm 1: Cho 10 µl Phagơ I và 10 µl của Phagơ II vào 2 ống nghiệm riêng biệt. Mỗi ống nghiệm đều chứa 5 ml nước luộc thịt có các tế bào chủng vi khuẩn pneumoccoci kiểu dại đang sinh trưởng mạnh. Một ống nghiệm khác chứa 5mL nước luộc thịt và chỉ có các tế bào chủng vi khuẩn pneumoccoci kiểu dại mà không có phagơ được sử dụng làm ống đối chứng. Sau 20 phút nuôi cấy ở nhiệt độ phòng, người ta lấy từ mỗi ống nghiệm 1µl dung dịch nuôi cấy và bổ sung thêm 1,999 mL nước cất khử trùng để pha loãng. Dung dịch tế bào sau khi pha loãng được cấy trải trên các đĩa thạch có chứa lần lượt glucozơ, sacarozơ và lactozơ. Các đĩa này được nuôi cấy ở trong tủ ấm có nhiệt độ 37oC trong 24 giờ. Kết quả thu được ở bảng 1.
Bảng 1. Kết quả thí nghiệm 1
Đĩa Tế bào bị gây nhiễm bởi Phage I
Tế bào bị gây nhiễm bởi Phage II
Tế bào chủng vi khuẩn dạng dại
Chứa glucozơ + + +
Chứa sacarozơ + - +
Chứa lactozơ + - -
Ghi chú: (+) Có sự phát triển của vi khuẩn ; (-) không có sự phát triển của vi khuẩn
Thí nghiệm 2: Từ mỗi ống nghiệm trong thí nghiệm, lấy 1µl dung dịch nuôi cấy rồi pha loãng bằng 1,999 mL nước cất khử trùng. Các dung dịch tế bào sau khi pha loãng được cấy trên 3 đĩa thạch: đĩa thứ nhất chứa tetraciline, đĩa thứ hai chứa penicilin và đĩa thạch còn lại không chứa kháng sinh nào. Các đĩa sau đó được nuôi cấy ở trong tủ ấm có nhiệt độ 37oC trong 24 giờ. Kết quả thu được ở bảng 2. Bảng 2. Kết quả thí nghiệm 2
Đĩa Tế bào bị gây nhiễm bởi Phage I
Tế bào bị gây nhiễm bởi Phage II Tế bào chủng vi khuẩn dạng dại
Chứa tetracilin - - -
Chứa penicilin + + -
Không chứa kháng sinh + + +
Ghi chú: (+) Có sự phát triển của vi khuẩn ; (-) không có sự phát triển của vi khuẩn
Câu 10 a.
Dựa trên các kết quả thí nghiệm, câu nào dưới đây là đúng nhất với Phagơ I?
A. Phagơ I chèn ADN vào nhiễm sắc thể của vi khuẩn làm cho vách tế bào vi khuẩn không chịu tác động bởi penicilin.
B. Phagơ I làm giảm penicilin trên đĩa thạch do đó cho phép tế bào vi khuẩn phát triển.
C. Phagơ I mang gen mã hoá cho beta lactamase
D. Phagơ I ức chế sự phát triển của vi khuẩn.
Câu 10 b.
Kết luận nào dưới đây có thể rút ra từ dữ liệu ở bảng 1?
A. Phagơ I chèn đoạn ADN vào nhiễm sắc thể của vi khuẩn ở vùng mã hoá cho các enzym của quá trình đường phân.
B. Phagơ I ngăn các phân tử lớn như lactozơ và sacarozơ có thể đi qua thành tế bào của vi khuẩn.
C. Phagơ II chèn đoạn ADN vào nhiễm sắc thể của vi khuẩn ở vị trí mã hoá cho enzym phân giải đường đôi.
D. Phagơ II sử dụng cả sacarozơ và lactozơ của vi khuẩn làm cho vi khuẩn chết đói.
Câu 10 c.
Kết luận nào dưới đây thể hiện kết quả đúng nhất của thí nghiệm 2?
I. ADN của cả Phagơ I và Phagơ II đều mã hoá cho beta lactamase.
II. ADN của cả Phagơ I và Phagơ II đều mã hoá cho enzym ức chế tác dụng có hại của tetracilin.
III. Cả hai phagơ đều phá huỷ khả năng kháng kháng sinh của tế bào vi khuẩn.
IV. Tế bào vi khuẩn dạng dại trong tự nhiên không có khả năng kháng lại cả penicilin lẫn tetracilin.
A. Chỉ có I.
B. Chỉ có I và IV.
C. Chỉ có III và IV.
D. Chỉ có I, II và IV.
Đáp án:
Câu 10 a Câu 10 b Câu 10 c
C C B
Bài 11- V2 năm 2013
Nấm men lưỡng bội dùng đường glucôzơ làm nguồn năng lượng ưu tiên, song cũng có thể dùng mantôzơ như nguồn năng lượng thay thế khi môi trường không có glucôzơ. Để tìm hiểu con đường chuyển hóa mantôzơ, người ta thường sàng lọc để tìm ra các chủng nấm men đột biến mất khả năng sử dụng mantôzơ. Từ phòng thí nghiệm, một nhà nghiên cứu đã phân lập được 6 chủng nấm men đột biến về khả năng chuyển hóa mantôzơ. Cả 6 đột biến đều là lặn và được đặt tên tương ứng là mal1, mal2, mal3, mal4, mal5 và mal6. Nhằm xác định các đột biến này có alen với nhau (thuộc cùng một gen) hay không, nhà nghiên cứu tiến hành phân tách giao tử từ các chủng nấm men đột biến đồng hợp tử nêu trên rồi tiến hành lai giữa chúng với nhau. Thí nghiệm nuôi tế bào lai trên các môi trường chỉ có mantôzơ và không có glucôzơ thu được kết quả như sau:
ác đột biến al1 al2 al3 al4 al5 al6 al1
Trong đó, “+” là nấm phát triển bình thường, còn “-” là nấm không phát triển.
Các thể đột biến này liên quan đến ít nhất bao nhiêu gen khác nhau?
Các đột biến nào alen với nhau? Thí sinh trả lời bằng cách kẻ bảng như ví dụ dưới đây trong bài làm, rồi điền đáp số hoặc tên đột biến (mal1-6) vào hai cột tương ứng
Số gen có liên quan Các đột biến alen với nhau
(ví dụ: 1, 2, 3....) (ví dụ: Gen 1 gồm mal1 và ....)
(ví dụ: Gen 2 gồm mal1 và ....)
Đáp án :
Số gen có liên quan Các đột biến alen với nhau
3 Gen 1 gồm mal1, mal2 và mal6
Gen 2 gồm mal3 và mal5; Gen 3 gồm mal4
Bài 12- V2 năm 2015
Có hai chủng vi khuẩn Staphylococcus aureus. Chủng I kháng penicilin và kanamixin, chủng II kháng streptomycine và ampixilin. Người ta tiến hành các thí nghiệm sau:
- Thí nghiệm 1: Cấy cả chủng I và II vào một bình đựng môi trường dinh dưỡng lỏng. Sau 24 giờ nuôi, phân lập từ bình này được một chủng mới (III) có khả năng kháng penicilin, kanamixin và streptomycine.
- Thí nghiệm 2: Cũng giống thí nghiệm 1, nhưng ngay từ đầu đã bổ sung enzyme endonuclease. Sau một thời gian, cũng phân lập được chủng III có khả năng kháng penicilin, kanamixin và streptomycine.
- Thí nghiệm 3: Lấy 1 ống hình chữ U đựng môi trường dinh dưỡng như ở thí nghiệm 1, ở đáy có một màng lọc, ngăn không cho các tế bào qua lại giữa hai bên ống nhưng lại cho phép các phân tử nhỏ như ADN hay phage đi qua. Cấy mỗi chủng vào một bên ống. Sau một thời gian cũng
ễ dàng phân lập được chủng III kháng penicilin, kanamixin và streptomycine.
- Thí nghiệm 4: Cũng giống thí nghiệm 3, nhưng người ta bổ sung endonuclease vào hai bên ống. Cuối cùng, cũng phân lập được chủng III kháng penicilin, kanamixin và streptomycine.
Hãy cho biết sự trao đổi vật chất di truyền ở 4 thí nghiệm trên là do kết quả của biến nạp, tải nạp hay tiếp hợp. Biết rằng, trong môi trường
của bốn thí nghiệm trên có thể nhiễm phage
Hướng dẫn:
- Thí nghiệm 1: biến nạp, tải nạp hoặc tiếp hợp.
- Thí nghiệm 2: Do ADN tự do bị phân giải bởi enzyme, nên chủng mới tạo thành có thể là do tải nạp hoặc tiếp hợp.
- Thí nghiệm 3: Do tế bào không thể qua màng lọc, nên chủng mới
tạo thành chỉ có thể do tải nạp hoặc biến nạp.
- Thí nghiệm 4: Chủng mới tạo thành có thể là do tải nạp.
Bài 13 - V2 năm 2018
Các con đường tổng hợp axit amin ở vi sinh vật được khám phá nhờ thí nghiệm nuôi chéo các chủng đột biến khuyết dưỡng về một bước trong con đường đó. Kết quả thí nghiệm nuôi chéo 3 chủng đột biến (TrpB-, TrpD-, và TrpE-) về các gen tương ứng (mã hoá cho các enzim TrpB, TrpD, TrpE) tham gia con đường tổng hợp tryptophan được biểu diễn ở hình bên. Trong thí nghiệm này, 3 chủng đột biến được cấy theo 3 được tạo thành hình tam giác trên đĩa thạch nhưng không dính với nhau. Môi trường nuôi chỉ có một lượng nhỏ tryptophan. Tại một số vị trí đường cấy gần nhau, chủng đột biến sinh trưởng mạnh hơn hẳn tạo nên vết sẫm màu. Điều này cho thấy một chủng đột biến có thể sử dụng sản phẩm chuyển hoá của chủng kia để tạo tryptophan.
a) Từ kết quả ở hình trên, hãy chỉ ra thứ tự tham gia trong con đường tổng hợp tryptophan của các enzim TrpB, TrpD và TrpE. Giải thích.
b) Chủng kiểu dại và 3 chủng đột biến trên được nuôi cấy trong một loại môi trường có tryptophan hoặc một số chất chuyển hoá trung gian thuộc con đường tổng hợp tryptophan. Kết quả sinh trưởng
của các chủng được trình bày trong bảng dưới đây
Chủn g vi sinh vật
Chất có trong môi trường nuôi cấy
Khô ng có Trypt ophan Anthr anilate Choris mate Indol e
Kiểu dại + + + + +
TrpB -
TrpD -

TrpE -
- + - - -
- + - - +
- + + - +
(+: có sinh trưởng; -: không sinh trưởng)
Hãy viết sơ đồ khái quát con đường sinh tổng hợp tryptophan từ các chất trung gian có trong thí nghiệm với sự tham gia của các enzim TrpB, TrpD và TrpE. Giải thích.
Hướng dẫn a)
- Thứ tự tham gia trong con đường sinh tổng hợp tryptophan: TrpE
TrpD TrpB
- Giải thích: tại vị trí gần với các chủng khác, chủng TrpB- không phát triển được chứng tỏ nó không sử dụng được sản phẩm của hai chủng kia để tạo tryptophan nên enzim TrpB tham gia ở bước cuối cùng. Ngược lại, chủng TrpE- có sự phát triển tốt ở vị trí gần chủng TrpB- và TrpD- chứng tỏ nó sử dụng được sản phẩm trung gian của 2 chủng kia để tạo tryptophan hoàn chỉnh, nó bị hỏng enzim cần cho sự chuyển hoá ở bước đầu so với TrpB và TrpD.
b)
Sơ đồ khái quát: TrpE TrpD TrpB Chorismate Anthranilate Indole Tryptophan.
- Giải thích: Cả 3 chủng không sinh trưởng trên môi trường có bổ sung chorismate chứng tỏ chúng đều không thể chuyển hoá chorismate thành tryptophan nên chorismate phải nằm ở đầu con đường sinh tổng hợp (chủng TrpE- thiếu enzim chuyển hoá chorismate). Ngược lại, chỉ có chủng TrpB-
không sinh trưởng được trên môi trường có indole chứng tỏ chủng này thiếu enzim chuyển hoá indole và indole là phân tử được chuyển thành tryptophan. Chủng TrpD- thiếu enzim chuyển hoá sản phẩm ở giữa là anthranilate.
Bài 14 - V2 năm 2019
Có hai chủng E. coli khuyết dưỡng khác nhau : một chủng có kiểu gen thrleu- thi- bio+ phe+cys+, chủng kia thr+ leu+ thi+ bio- phe- cys - ( + : kiểu hình kiểu dại, - : kiểu hình khuyết dưỡng thr: theronin; leu: leucine; thi: thiamine; bio:biotin; phe: phenylalanine và cys: cysteine). Hai chủng này được nuôi trong cùng một môi trường lỏng bổ sung cả 6 chất là theronin; leucine; thiamine; biotin; phenylalanine và cysteine. Sau đó, chúng được cấy trải trên một đĩa thạch chứa môi trường đầy đủ cả 6 chất trên. Đĩa thạch chứa 6 khuẩn lạc được đánh số từ 1 đến 6 là đĩa gốc ( hình dưới).
A- Sai , B- sai , C- sai , D- sai

Hướng dẫn
Chủng thứ nhất : thr- leu- thi- bio+ phe+cys+.: chỉ mọc đc thi bổ sung thr , leu, thi
Chủng thứ 2 : thr+ leu+ thi+ bio- phe- cys -: chỉ mọc đc thi bổ sung bio, phe, cys
A- Sai, KL 1 không thể thể là chủng 1 thr- leu- thi- bio+ phe+cys+:
KL1 vẫn mọc đc trong điều kiện chỉ có thr, leu ( đĩa 1), không cần bổ sung nó vẫn mọc không phải là khuyết dưỡng với threonin không thể là chủng 1 theo đề bài
B- Sai vì đĩa 3 và đĩa 5 ( tính trái sang, trên xuống dưới) nó vẫn mọc khi không bổ sung bio ( dĩa 3) và phe ( đĩa 5 ) nên nó chỉ khuyết dưỡng với 1 axit amin là cys
C- Sai vì 5 mọc được 1 đĩa số 1, mà KL1 mọc được ở 2 đĩa
KL 5 sẽ khuyết dưỡng với thimine
D- Sai vì 6 là nguyên dưỡng vì mọc ở tất cả các đĩa .
Bài 15- Vòng 2 năm 2020
Phép thử bổ sung (complementation test) là một phương pháp nghiên cứu cơ bản nhằm xác định sự tương tác giữa các gen cùng phối hợp thực hiện một chức năng chung hoặc để lập bản đồ di truyền giữa các gen liên kết bằng việc tiến hành lai giữa các thể đột biến mất chức năng.
Người ta sử dụng kĩ thuật đánh dấu khuẩn lạc để chuyển các khuẩn lạc từ đĩa gốc này sang các đĩa khác sao cho các khuẩn lạc không thay đổi gì so với đĩa gốc. Kết quả đã tạo ra 6 đĩa bản sao từ đĩa gốc trong đó mỗi đĩa bản sao chứa môi trường tối thiểu và chỉ bổ sung thêm 2 trong số 6 chất trên ( được ghi tương ứng dưới mỗi đĩa thạch). Khả năng mọc của 6 khuẩn lạc trên mỗi loại môi trường như hình vẽ Mỗi nhận định sau đây là đúng hay sai ?
A. Khuẩn lạc 1 là chủng khuyết dưỡng có kiểu gen thr- leu- thibio+ phe+cys+
B. Khuẩn lạc 3 là chủng khuyết dưỡng có kiểu gen thr+ leu+ thi+ bio- phe- cys -
C. Khuẩn lạc 3 là chủng khuyết dưỡng có kiểu gen khuẩn lạc 1
D. Khuẩn lạc 3 là chủng khuyết dưỡng về cả 6 loại axit amin đã
Virut Herpes (HPV) là một virut lớn có vật chủ sinh vật nhân thực. Sự phát triển của virut trải qua nhiều giai đoạn. Để chuyển từ giai đoạn này sang giai đoạn khác, virut phải hoàn thành giai đoạn trước đó. Để phòng tránh và điều trị bệnh do virut HPV, các nhà khoa học cần tìm hiểu vai trò của các gen virut tham gia vào các giai đoạn phát triển khác nhau. Có 9 chủng virut đột biến bị dừng phát triển ở giai đoạn sớm được kí hiệu lần lượt từ A đến I, và một chủng đột biến làm virut dừng phát triển ở giai đoạn muộn được kí hiệu W. Mỗi chủng virut đều phát triển tự do ở nhiệt độ 34oC, nhưng không phát triển ở nhiệt độ 39oC. Tất cả các chủng, trừ chủng I, đều có tỉ lệ đột biến phục hồi (trở về kiểu dại) ở tần số thấp giống nhau; riêng chủng I cũng có khả năng đột biến phục hồi, song tần số thấp hơn nhiều so với những chủng còn lại.
Phép thử bổ sung đã được tiến hành giữa 9 chủng virut đột biến giai đoạn sớm (từ A đến I) với nhau và giữa mỗi chủng này với chủng giai đoạn muộn (W). Từng cặp đột biến được đồng biến nạp vào tế bào chủ ở 39oC, rồi thu môi trường nuôi tế bào và định lượng virut trước và sau khi lây nhiễm chúng vào tế bào ở 34oC. Trong phép thử bổ sung ở HPV, khả năng tương tác và tái tổ hợp của 2 thể đột biến x và y nhất định được xác định tương ứng thông qua chỉ số tương tác Qxy và chỉ số tái tổ hợp RFxy:
Lượng virut khi đồng biến nạp cả X và Y
Qxy =
Lượng virut khi biến nạp X riêng rẽ + Lượng virut khi biến nạp Y riêng r
Lượng virut khi đồng biến nạp cả X và Y ở 39oC
b) Hệ số Q giữa 9 chủng đột biến giai đoạn sớm đều nhỏ hơn 2 chứng tỏ chúng đều liên quan đến 1 chức năng. Riêng chủng W là 1 chức năng khác (nhờ Q > 2) => 10 chủng đột biến trong thí nghiệm liên quan đến 2 chức năng.
RFxy =
Lượng virut khi đồng biến nạp X và Y ở 34oC
Theo đó, tương tác bổ sung về chức năng được xem là “có” (do các đột biến có chức năng khác nhau) nếu Q > 2 và được xem là “không có” khi Q ≤ 2. Tái tổ hợp được xem là “có” khi RF > 0 và được xem là “không có” khi RF = 0.
Số liệu phép thử bổ sung giữa các chủng virut được trình bày ở các Bảng 2.1 và 2.2.
Bảng2.1. Hệ số Qgiữamỗicặpchủng virut
Chủng B C D E F G H I W
A 1,50,70,80,40,40,60,50,7170
B 0,30,30,80,80,90,30,34,9
C 0,30,21,91,50,60,7 19
D 0,20,40,70,10,5 72
E 1,40,80,30,4580
F 0,50,10,2570
G 0,40,9 22
H 0,4444
30
Bảng2.2. Hệ số RFgiữamỗicặpchủng virut
Dựa trên chỉ số RF, H và I là 2 đột biến ở cùng vị trí, còn các đột biến còn lại thuộc các vị trí khác nhau => 10 chủng đột biến liên quan đến 9 vị trí khác nhau
c) Vì chủng I có tần số đột biến phục hồi thấp hơn nhiều nên dẫn đến số liệu thí nghiệm thiếu nhất quán hơn những chủng khác
d)
a) Xét về cấu trúc gen bị tác động, nhiều khả năng loại đột biến ở mỗi chủng virut nêu trên là loại đột biến gì? Giải thích.
b) Các thể đột biến trong thí nghiệm liên quan đến bao nhiêu chức năng và bao nhiêu vị trí khác nhau trong hệ gen virut? Giải thích.
c) Tại sao chủng I có giá trị RF biến động giữa các lần lặp lại thí nghiệm lớn hơn nhiều so với các chủng còn lại (số liệu không được nêu ở đây)?
d) Vẽ bản đồ di truyền biểu thị thứ tự sắp xếp các đột biến (từ A đến
I) trên hệ gen virut, biết rằng chỉ số RF phản ánh khoảng cách giữa các vị trí đột biến.
Hướng dẫn
a) Vì tất cả các chủng đột biến trên đều có khả năng đột biến phục hồi về kiểu dại, nên nhiều khả năng hơn cả là cả 10 chủng đột biến đều là đột biến điểm (đột biến thay thế các nucleotit)
Bài 16- Vòng 2 năm 2020
Trong một thí nghiệm, chất gây đột biến 5-bromo uracil (5-BU) được bổ sung vào môi trường nuôi vi khuẩn E. coli kiểu dại đang phân chia nhanh. Thành phần môi trường nuôi giàu dinh dưỡng và có bổ sung axit amin Arginine (Arg). Sau khoảng thời gian tương ứng một lần phân bào, các tế bào được rửa sạch chất gây đột biến, rồi được nuôi trên môi trường tối thiểu chỉ bổ sung Arg. Các đĩa nuôi cấy có khuẩn lạc được phân tách tốt, nghĩa là mỗi khuẩn lạc có nguồn gốc chỉ từ một tế bào duy nhất. Các khuẩn lạc sau đó được chuyển lên môi trường tối thiểu bằng phương pháp đóng dấu. Một khuẩn lạc phát triển khi có Arg, nhưng không phát triển trên môi trường tối thiểu được phân lập. Khuẩn lạc này được rửa bằng nước cất vô trùng, rồi phân chia đều vào 20 ống nghiệm chứa môi trường tối thiểu bổ sung Arg. Sau khi tế bào trong mỗi ống nghiệm phát triển tới mật độ ~108 tế bào/mL, người ta thu 0,1 mL từ mỗi ống và cấy trải trên đĩa chứa môi trường thạch tối thiểu không có Arg (mỗi đĩa được đánh số tương ứng với ống nghiệm gốc). Bảng dưới cho thấy số lượng khuẩn lạc quan sát được trên mỗi đĩa.
Mỗi nhận định dưới đây là ĐÚNG hay SAI?
A. Đột biến phục hồi (đột biến đảo ngược) về tổng hợp Arg có lẽ đã không xảy ra ở các dòng 2, 4, 10, 15, 16 và 20.
B. Có lẽ đột biến phục hồi xảy ra sớm nhất ở dòng 19.
C. Nếu 5-BU được thêm vào môi trường ở cả 20 ống, thì số lượng khuẩn lạc ở mỗi đĩa đều tăng.
D. Nếu hóa chất thuộc nhóm acrydine (như ethidium bromide) được
bổ sung vào môi trường ở cả 20 ống (thay vì 5-BU) thì số lượng khuẩn lạc ở mỗi
đĩa đều tăng.
Đáp án
A. Đúng B. Đúng C. Sai D. Sai
Bài 17- Vòng 2 năm 2020
Một số chủng đột biến khuyết dưỡng được phân lập bắt nguồn từ nấm men đơn bội, kiểu dại. Các chủng đột biến này đáp ứng khác nhau với các môi trường tối thiểu được bổ sung các chất chuyển hóa trung gian khác nhau (kí hiệu từ S đến W) biểu hiện bằng khả năng sinh trưởng (+) hoặc không sinh trưởng (0). Bảng sau cho biết kết quả của các chủng nấm men mang các đột biến.
Cá c chủng đột biến
Các chất được bổ sung vào môi trường tối thiểu
Có một số các chủng đột biến dinh dưỡng được phân lập từ các chủng nấm mốc bánh mỳ đỏ kiểu hoang dại Neurospora crassa đáp ứng với việc bổ sung thêm các chất nhất định vào môi trường nuôi cấy, nếu phát triển (+) hoặc không phát triển (0). Bảng dưới đây cho biết các số liệu về các thể đột biến gen đơn.
S train C hủng
Supplements added to minimal culture medium Chất bổ sung vào môi trường tối thiểu Citr ulline Glutamic semialdehyde A rginine Or nithine Gluta mic acid A + 0 + 0 0 B + + + + 0 C + 0 + + 0 D 0 0 + 0 0
1.1. Hãy chỉ ra thứ tự chuyển hóa (1-5) của 5 sản phẩm chuyển hóa trong quá trình trao đổi trong phiếu trả lời
Sản phẩm trao đổi Thứ tự chuyển hóa
Citrulline
Glutamic semialdehyde
Mỗi sơ đồ chuyển hóa dưới đây là ĐÚNG hay SAI khi đối chiếu với dữ liệu ở bảng trên?
Arginine
Ornithine
Glutamic acid
1.2. Hãy chỉ ra chủng nào bị ngăn cản ở mỗi một bước trong 4 bước của quá trình chuyển hóa nêu trong phiếu trả lời
Bước trong quá trình chuyển hóa 1 → 2 2 → 3 3 → 4 4 → 5
Chủng (bị ngừng trệ trong bước)
Sản phẩm trao đổi Thứ tự chuyển hóa
Citrulline 4
Glutamic semialdehyde 2
Arginine 5
Ornithine 3


Glutamic acid 1
Bước trong quá trình chuyển hóa
Chủng (bị ngừng trệ trong bước) B C A D
Bài 17 - IBO 2014
Có 3 dòng vi khuẩn đột biến TrpB-, TrpC- and TrpE-, mỗi dòng đều bị hỏng
ở một bước trong con đường sinh tổng hợp tryptophan. Mỗi dòng đều được cấy thành vệt tam giác trên đĩa petri như trong hình dưới đây. Một nồng độ
tryptophan rất nhỏ có trong môi trường nuôi cấy cho phép chúng sinh trưởng và tạo nên một dải mảnh hơn cho đến khi hết tryptophan. Tuy nhiên, một số vùng trên dải vẫn có tế bào sinh trưởng chậm và lâu dần trở nên dày hơn. Con đường chuyển hóa tryptophan bao gồm các bước chuyển hóa liên tiếp chorismate thành anthranilate, indole và cuối cùng thành tryptophan.
TrpE TrpD TrpB
Chorismate Anthranilate Indole
Tryptophan
Hãy chỉ ra câu đúng, câu sai dưới đây.
A. Kết quả này chứng tỏ rằng các chất trung gian của con đường sinh tổng hợp được chủ động tiết ra khỏi tế bào.
B. Dòng đột biến TrpC- có một đột biến trong gen quy định enzym xúc tác việc chuyển hóa indole thành tryptophan.
C. Dòng đột biến TrpE- có khả năng tổng hợp tryptophan nếu môi trường nuôi cấy chứa anthranilatehoặc indole.
D. Indole sẽ tiếp tục được tích tụ lại trong môi trường nuôi cấy ở vùng các tế bào TrpBnằm cạnh các tế bào TrpC-
Đáp án : A- Đúng , B- Sai, C- đúng , D- sai
Bài 18 - IBO 2014
Vị trí của 6 mất đoạn đã được lập bản đồ ở nhiễm sắc thể ở
Drosophila được trình bày trong sơ đồ dưới đây.
Ghi chú: Deletion = mất đoạn; chromosome = nhiễm sắc thể




Các đột biến lăn a, b, c, d, e và f được biết nằm trong cùng một vùng mất đoạn, nhưng trình tự của các đột biến trên nhiễm sắc thể chưa được biết. Khi các con ruồi đồng hợp tử về các đột biến lặn với các con ruồi đồng hợp tử về các kiểu mất đoạn người ta nhận được kết quả dưới đây. Lưu ý “m” chỉ kiểu hình đột biến và dấu (+) chỉ kiểu hìnhdại Đột biến
Mất đoạn a b c d e f
Hãy chỉ ra câu nào dưới đây là đúng hoặc sai.
A. Đột biến d nằm ở vùng bao trùm giữa mất đoạn 4 và mất đoạn 6.
B. Vị trí của đoạn bị mất nơi chứa đột biến b có thể suy ra bằng thông tin từ vùng bao trùm giữa mất đoạn 3,4 và 5.
C. Trong số 6 đột biến, đột biến d nằm trong trong vùng mất đoạn ngắn nhất.
D. .Trình tự tương đối của 6 đột biến trên nhiễm sắc thể là a, f, d, b, c, e. Đáp án : vị trí của 6 đột biến

Các phát biểu sau đúng hay sai
A. Gen chỉ thị (kháng kanamycin -KM) được chuyển trước gen tổng hợp uracil
B. Số liệu gợi ý rằng toàn bộ hệ gen của vi khuẩn có thể được chuyển thông qua tiếp hợp chỉ trong thời gian ít hơn 15 phút.
C. Số liệu gợi ý rằng một tỷ lệ nhỏ các khuẩn lạc thể nhận có thể mọc trên môi trường có kháng sinh kanamycin (+KM) thậm chí trong điều kiện không có tiếp hợp.
D. Hơn 20% tất cả các khuẩn lạc được ủ sau 15 phút có thể mọc trên môi trường thiếu cả arginin và uracil.
Đáp án
A- Sai , B- Sai , 3- Đúng , 4- Đúng

Bài 20 - IBO 2015
Ở nấm mốc, sự sinh tổng hợp axit amin arginine cần bước chuyển hóa tiền chất thành ornithine; chất này được chuyển hóa tiếp thành citruline là tiền chất trực tiếp được chuyển hóa thành arginine. Mỗi phản ứng trong 3 bước này được xúc tác bởi một enzym riêng rẽ; mỗi enzym được mã hóa bởi một gen (xem Hình)
A- Sai , B- Đúng , C- Đúng , D- Đúng
Bài 19 - IBO 2015
Trong một thí nghiệm, hai chủng vi khuẩn I và II được cho tiếp hợp với nhau. Chủng I chứa các gen cho phép nó mọc trên môi trường thiếu arginin và uracil, với đường galactose là nguồn carbon duy nhất, và thậm chí có mặt chất kháng sinh kanamycin (+KM). Chủng II không thể mọc trên môi trường với các điều kiện vừa nêu. Sau một thời gian ủ, chủng II được nuôi trên môi trường chọn lọc cho tới khi thu được 100 khuẩn lạc. Sự thành công của quá trình tiếp hợp được đánh giá (Xem Bảng)
A: Con đường sinh tổng hợp arginine ở nấm mốc. B: Thiết kế thí nghiệm để xác định chuyển hóa: dải màu nâu đen chỉ Nấm mốc sinh trưởng. Chủng kiểu dại mọc trên môi trường tối thiểu, trong khi các chủng đột biến chỉ mọc trên các môi trường được bổ sung (+) chất dinh dưỡng đặc thù.
Các phát biểu sau đúng hay sai
A. Các chủng đột biến lớp III có thể mang đột biến ở nhiều hơn 1 gen liên quan đến con đường sinh hóa này
B. Các chủng đột biến lớp I thiếu cả 3 enzym
C. Các thể đột biến thiếu hai enzym A và C có thể mọc nếu môi trường được bổ sung đồng thời Orthnithine và Citruline
D. Giả sử một chất ức chế enzym B được bổ sung vào môi trường nuôi chủng kiểu dại, gen B sẽ bị bất hoạt Đáp án : A- Sai , B- Sai , 3- Sai , 4- Sai

Bài 21 - IBO 2016
Các nhà khoa học đã phân lập được 3 chủng vi khuẩn khác nhau ProA-, ProB-, ProC- cần bổ sung proline để sinh trưởng. Một chủng nhạy cảm với điều kiện lạnh, một chủng nhạy cảm với điều kiện nóng, và một chủng có một gen bị mất (deletion).

Các thí nghiệm nuôi chéo được thực hiện bằng cách cấy đồng thời các chủng vi khuẩn trên đĩa thạch chứa môi trường tối thiểu được bổ sung proline ở nồng độ rất thấp. Trong các thí nghiệm nuôi chéo này, sản phẩm trao đổi chất được tiết ra từ một chủng có thể nuôi dưỡng chủng bên cạnh. Sau khi sinh trưởng ở 3 nhiệt độ khác nhau, kết quả được biểu diễn ở Hình Q.3.
Precursor Tiền chất
Ornithine Ornithine
Citrulline Citruline
Class I mutants Các đột biến lớp I
Class II mutants Các đột biến lớp II
Class III mutants Các đột biến lớp III
Arginine Arginine + Ornithine + Ornithrine
Enzyme Enzym + Citrulline + Citruline
Gene Gen + Arginine + Arginine

Wild type Kiểu dại (WT)
Hình Q.3. Kết quả của các thí nghiệm nuôi đồng thời 3 chủng không có khả năng tổng hợp proline trên đĩa thạch. Các vùng màu đen sẫm thể hiện tốc độ sinh trưởng mạnh của tế bào; các vùng xám nhạt thể hiện tốc độ sinh trưởng thấp; wt là chủng kiểu dại.
Hãy xác định mỗi câu sau đây là Đúng hay Sai và viết vào Phiếu trả lời.
A. Chất trung gian tích luỹ trong chủng ProC- xuất hiện sau khi sự trao đổi chất ở chủng ProA- bị dừng lại.
B. Chất trung gian tích luỹ trong chủng ProB- xuất hiện sau khi sự trao đổi chất ở chủng ProA- bị dừng lại.
C. Có ít nhất 3 gen khác nhau ảnh hưởng đến sự tổng hợp proline.
D. Dưới ít nhất một điều kiện, proline được tạo ra sẽ nhanh chóng
được sử dụng để tổng hợp protein và proline sẽ bị dừng tổng hợp nếu dư thừa.
Đáp án: A, C, D đúng; B sai .


Bài 22 - IBO 2018
Các hợp chất X và Y là những tiền chất trong con đường tổng hợp chất Z. Z là hợp chất thiết yếu đối với sự sinh trưởng. Kiểu dại (WT) của Neurospora (nấm men) là dạng nguyên dưỡng, tức là chúng có thể sinh trưởng trên môi trường tối thiểu (MM).
Thí nghiệm Sinh trưởng:
Bốn thể đột biến neurospora Z- được phân lập, và các kết quả thí nghiệm với các thể đột biến này được trình bày dưới đây (+ = sinh trưởng được, - = không sinh trưởng được).
Phép lai % bào tử đơn bội nguyên dưỡng
Thể đột biến 1 x Thể đột biến 2 25%
Thể đột biến 1 x Thể đột biến 3 25%
Thể đột biến 1 x Thể đột biến 4 0% (nhiều bào tử được đếm)
Thể đột biến 2 x Thể đột biến 3 0.004%
Thể đột biến 2 x Thể đột biến 4 0.001%
Thể đột biến 3 x Thể đột biến 4 0.001%
Cho biết mỗi phát biểu dưới đây là Đúng hay Sai.
Các đột biến ở các chủng bị đột biến dường như nằm trên hai gen.
Xét tất cả các chủng đột biến, ít nhất có 5 vị trí khác nhau của hệ gen Neurospora bị đột biến ở một hoặc nhiều chủng.
Có ít nhất hai chủng đột biến là các thể đột biến kép (tức là có nhiều hơn 1 vị trí bị đột biến ở mỗi chủng này).
Phép thử bổ sung (tương tác bổ trợ)
Phép thử bổ sung được thực hiện bằng việc kiểm tra khả năng sinh trưởng trên môi trường tối thiểu của tế bào hợp nhân lưỡng bội tạo thành từ phép lai hai tế bào nấm men đơn bội đột biến. Các kết quả thu được như sau (+ = sinh trưởng được, -: không sinh trưởng được).
Vị trí đột biến của Thể đột biến 3 nằm giữa vị trí đột biến của Thể đột biến 2 và vị trí đột biến của Thể đột biến 4.
Theo lý thuyết, 25% bào tử đơn bội tạo thành từ phép lai giữa Thể đột biến 2 và chủng kiểu dại là thể nguyên dưỡng.
Đáp án: đúng sai sai sai sai
Thí nghiệm lai:
Các chủng đột biến được lai và tỷ lệ % thể nguyên dưỡng trong số các bào tử đơn bội tạo thành từ mỗi phép lai được xác định. Kết quả như sau.
ĐÚNG SAI
1. Kết luận Trên cơ sở tổng hợp những tài liệu khác nhau, chuyên đề trên đã:
- Hệ thống hóa được kiến thức cơ bản và chuyên sâu về di truyền học hóa –sinh và di truyền học vi khuẩn
- Nêu ra một số câu hỏi vận dụng kiến thức trong chuyên đề và hướng dẫn học sinh trả lời, qua đó giúp cho các em có thể hệ thống lại kiến thức của mình về phần này, giúp các em phát huy được tính chủ động, tích cực. Từ đó, góp một phần nhỏ vào quá trình nghiên cứu của học sinh và giáo viên, đặc biệt là trong ôn thi đội tuyển học sinh giỏi.
2. Kiến nghị
Việc nghiên cứu về các nội dung chuyên đề di truyền học hóa – sinh và di truyền học vi khuẩn là vấn đề cần thiết, các đề thi học sinh giỏi hiện nay có xu hướng khai thác triệt để vấn đề này. Với sự phát triển như vũ bão của ngành sinh học phân tử, đặc biệt trong thế kỷ 21, thế kỷ của công nghệ sinh học, để làm chủ kiến thức liên quan đến sự sống thì những hiểu biết về di truyền học vi sinh vật nói chung và di truyền vi khuẩn nói riêng là không thể thiếu.
Mặc dù đã có nhiều cố gắng, nhưng không thể tránh khỏi những hạn chế, thiếu sót. Tôi rất mong nhận được sự đóng góp ý kiến của các đồng nghiệp để chuyên đề hoàn thiện hơn.
Tôi xin chân thành cảm ơn!
TÀI LIỆU THAM KHẢO CHÍNH
1. Campbell & Reece (2015), Sinh học, Nhà xuất bản giáo dục, Hà Nội.
2. Nguyễn Lân Dũng (2019) , Vi sinh vật học, Phần 2, Nhà xuất bản khoa học kĩ thuật, Hà Nội.
3. Đinh Đoàn Long (2013), Cơ sở di truyền học phân tử và tế bào, Nhà xuất bản đại học Quốc gia Hà Nội.
4. Nguyễn Hoàng Lộc (2007), Giáo trình sinh học phân tử, Nhà xuất bản đại học Huế.
5. Hoàng Trọng Phán (2008), Giáo trình di truyền học vi sinh vật và ứng dụng, Nhà xuất bản đại học Huế.
6. Nguyễn Thị Hồng Vân (2011), Giáo trình di truyền học sinh vật nhân sơ và virus, Nhà xuất bản giáo dục Việt Nam, Hà Nội.
7. Harvey Lodish & et at (2019), Molecular cell biology 8th Edition, Pearson, NewYork.
8. Lodish & et at (2019), Sinh học phân tử và tế bào tập 4, Nhà xuất bản trẻ, TP Hồ Chí Minh..
9. Các đề thi đề xuất lớp 10 của khu vực Duyên Hải và Đồng bằng Bắc Bộ các năm.
10. Các đề thi chọn học sinh giỏi Quốc gia, chọn học sinh dự thi Quốc tế và đề thi Quốc tế các năm.
Chuyên đề
TÍCH HỢP DI TRUYỀN HOÁ SINH VI SINH VẬT
Nguyễn Mạnh Quỳnh, Nguyễn Thị Thu Trang - THPT Chuyên Thái Nguyên
Chuyên đề đạt giải Nhì
- Các giáo viên sinh học.
B. NỘI DUNG CHUYÊN ĐỀ
I. LÝ THUYẾT
1. Ưu thế của các đối tượng vi sinh vật.
A. PHẦN MỞ ĐẦU.
I. Lí do chọn đề tài
Những năm 40 của thế kỷ XX, vi sinh vật bắt đầu được sử dụng vào nghiên cứu di truyền học. Đem đến những phát minh quan trọng có tính cách mạng trong sinh học như chứng minh trực tiếp ADN là vật chất di truyền, sao chép ADN, sinh tổng hợp prôtêin, kỹ thuật di truyền... Di truyền học vi sinh vật đóng vai trò “cách mạng hóa”, tạo ra bước tiến nhảy vọt cho di truyền học nói riêng, hình thành Sinh học phân tử và sự phát triển sinh học hiện đại nói chung cả về lí thuyết và ứng dụng thực tiễn.
Di truyền học vi sinh vật đã góp phần chủ yếu cho sự phát triển di truyền học phân tử, mà đỉnh cao là sự ra đời kỹ thuật tái tổ hợp ADN hay kỹ thuật di truyền (KTDT), làm bùng nổ cách mạng công nghệ sinh học.
Việc sử dụng đối tượng vi sinh vật để nghiên cứu hóa sinh tế bào, sinh học phân tử, di truyền biến dị ở sinh vật là tất yếu và đã đạt được rất nhiều thành tựu to lớn, ý nghĩa.
Trong chương trình sinh học cấp trung học phổ thông, phần kiến thức về hóa sinh tế bào, vi sinh vật và di truyền, biến dị là một nội dung rất quan trọng, được đưa nhiều vào đề thi chọn học sinh giỏi Quốc gia, Quốc tế. Mặc dù hiện nay có rất nhiều tài liệu viết về hóa sinh tế bào, vi sinh vật và di truyền, biến dị nhưng ở các tài liệu với mục đích khác nhau, phần kiến thức riêng lẻ, được trình bày theo nhiều hướng vì thế nhiều giáo viên và học sinh còn lúng túng với việc vận dụng phần kiến thức này trong giảng dạy và ôn luyện đội tuyển học sinh giỏi. Xuất phát từ thực tế này, chúng tôi biên soạn chuyên đề: “Tích hợp một số vấn đề về hóa sinh, vi sinh và di truyền” để giúp học sinh có cái nhìn tổng quát hơn, thống nhất hơn. Đồng thời hỗ trợ cho giáo viên kiểm tra kiến thức học sinh thông qua hệ thống các câu hỏi. Đây cũng là nguồn tài liệu ôn tập trong các kì thi trung học phổ thông, học sinh giỏi các cấp, kì thi chọn học sinh giỏi Quốc gia và Quốc tế.
II. Mục tiêu của đề tài
- Hệ thống hóa một số kiến thức về vấn đề tích hợp hóa sinh, vi sinh và di truyền.
- Giới thiệu một số câu hỏi, bài tập vận dụng.
III. Đối tượng áp dụng
- Học sinh ôn thi trung học phổ thông.
- Các đội tuyển ôn thi học sinh giỏi môn sinh học các cấp.
1.1. Thời gian thế hệ ngắn, tốc độ sinh sản nhanh
- Thời gian thế hệ của E. coli là 20 phút, còn bacteriophagơ có thể trong 30 - 40 phút tạo ra hàng trăm cá thể, nấm men có thể phân chia tế bào trong 2 giờ

- Thời gian thế hệ của ruồi giấm là 2 tuần, của chuột là 2 tháng, của người có thể lên đến 20 năm.
1.2. Tăng vọt số lượng cá thể
- Các vi sinh vật đơn bào, mỗi tế bào là một cá thể. Nếu đủ dinh dưỡng, các vi sinh vật sinh sản nhanh tạo quần thể có số lượng cá thể lớn có thể khoảng 1010 - 1012 tế bào/ml. Nhờ số lượng lớn cá thể của vi sinh vật, có thể phát hiện các sự kiện di truyền hiếm hoi (như đột biến hay các dạng tái tổ hợp) với tần số 10–8 - 10–11 .
- Số lượng cá thể lớn giúp nâng cao năng suất phân giải (resolving power) di truyền, tức khả năng phát hiện các đột biến và tái tổ hợp có tần số xuất hiện rất nhỏ
- Ưu thế này lại được tăng thêm nhờ môi trường nuôi đơn giản, dễ nuôi cấy, dễ nhân giống, mà điều kiện nuôi cấy không cồng kềnh, ít tốn diện tích, dễ kiểm soát theo công thức chặt chẽ như khi làm thí nghiệm hóa học.
1.3. Cấu tạo bộ gen đơn giản
Hình
1. Cấu trúc vi khuẩn
- Vi khuẩn và virut có bộ gen là ADN trần dễ tiến hành thí nghiệm trực tiếp trên ADN cũng như chiết tách tinh sạch. Số locut cũng ít hơn so với các sinh vật khác.
- Các vi nấm và vi tảo có thể tồn tại ở dạng đơn bội (n) với thời gian dài, nên các gen lặn có thể biểu hiện ngay. Các vi sinh vật ở trạng thái lưỡng bội (2n) cũng dễ dàng thực hiện phân tích tái tổ hợp. Các tính trạng ở vi sinh vật cũng đơn giản hơn, xác định di truyền các tính trạng này ít phức tạp hơn.
1.4. Dễ thu nhận các đột biến
- Các đột biến ở vi sinh vật có thể thu nhận dễ dàng, thậm chí có tần số xuất hiện thấp 10–10, việc xác nhận dạng đột biến cũng rất nhanh.
- Nhờ ưu thế này mà di truyền học vi sinh vật phát triển rất nhanh hình thành nên di truyền học phân tử và sinh học phân tử
1.5. Dễ nghiên cứu bằng các kỹ thuật vật lý, hóa học
- Cấu tạo tế bào vi sinh vật đơn giản, dễ chiết tách, tinh sạch ADN. Có thể nuôi vi sinh vật đồng nhất (synchronous culture) tức là đa số các tế bào ở những giai đoạn phát triển gần giống nhau. Độ đồng nhất cao của vật liệu thí nghiệm tạo thuận lợi cho việc sử dụng các phương pháp vật lý hóa học trong nghiên cứu di truyền.
- Mặc dù các vi sinh vật có những đặc điểm riêng nhưng chúng vẫn tuân theo các quy luật di truyền chung, các kết quả thu được có thể đối chiếu áp dụng cho các vi sinh vật bậc cao.
2. Các đặc điểm của di truyền học vi sinh vật.
2.1. Nghiên cứu trực tiếp ở cấp độ tế bào và phân tử
- Nhiều đối tượng vi sinh vật như vi khuẩn Escherichia coli, nấm men Saccharomyces cerevisiae là sinh vật đơn bào nên các quan sát thực hiện trực tiếp hơn ở mức tế bào. Ví dụ, trên môi trường tối thiểu với lượng muối hữu cơ và glucose cho trước, một tế bào E. coli có thể tổng hợp tất cả các hợp chất cần thiết cho sự tăng trưởng, sống sót và sinh sản.

- Nếu tế bào bị đột biến mất khả năng tổng hợp một chất nào đó như một loại axit amin thì nó không mọc được trên môi trường tối thiểu. Trên môi trường có bổ sung thuốc kháng sinh thì chỉ có các tế bào đề kháng kháng sinh mới hình thành được khuẩn lạc.
- Các tế bào vi sinh vật có nhu cầu dinh dưỡng đơn giản dễ kiểm soát thành phần môi trường nuôi, dễ nuôi cấy, dễ nhân giống, dễ dàng thu nhận nhiều loại đột biến phục vụ cho nghiên cứu trực tiếp và nhanh chóng ở cấp độ tế bào, phân tử. Do vậy, chúng là những đối tượng lý tưởng cho nghiên cứu di truyền học phân tử
2.2. Dòng tế bào
- Di truyền vi sinh vật không nghiên cứu từng tế bào riêng lẻ mà là dòng của tế bào, tức tập hợp của nhiều tế bào bắt nguồn từ một tế bào ban đầu nhờ sinh sản vô tính. Thông thường, tế bào vi sinh vật được cấy lên môi trường thạch đặc, rồi trải đều để các tế bào nằm rời xa ra thì mỗi tế bào mọc lên thành một cụm rời gọi là khuẩn lạc (colony). Mỗi khuẩn lạc cũng là một dòng tế bào (clone) gồm các tế bào bắt nguồn tự sự phân chia của một tế bào ban đầu.
Hình 2. Khuẩn lạc (colony) của vi sinh vật
- Mỗi khuẩn lạc dễ nhìn thấy bằng mắt thường, có tối thiểu 107 (mười triệu) tế bào. Như vậy, các tính trạng ở vi sinh vật được ghi nhận qua một quần thể gồm hàng trăm triệu, hàng tỉ tế bào.
- Dòng tế bào mang một đặc tính di truyền nào đó gọi là chủng (strain). Ví dụ: chủng vi khuẩn tạo nhiều vitamin B12 hay chủng vi khuẩn mất khả năng tổng hợp một axit amin nào đó.
2.3. Các tính trạng.
Các đột biến ở vi sinh vật thường được phát hiện theo sự biến đổi các tính trạng sau:
- Hình thái: kích thước, hình dạng tế bào hay khuẩn lạc, có màng nhân hay không, khả năng di động...
- Sinh hóa: sự hiện diện của các sắc tố, màu sắc đặc trưng.
- Nuôi cấy: như kiểu hô hấp, kiểu dinh dưỡng (khuyết dưỡng - auxotroph) hoặc nhu cầu đòi hỏi các nhân tố tăng trưởng.
- Tính đề kháng: như kháng thuốc, kháng phagơ, chịu nhiệt...
- Miễn dịch: như các phản ứng kháng thể, kháng nguyên...
Các đột biến có thể xuất hiện ngẫu nhiên hay do gây tạo ra nhờ các tác nhân gây đột biến. Mỗi gen có tần số đột biến đặc trưng.
Các tính trạng ở vi sinh vật được kí hiệu bằng 3 chữ tắt tiếng Anh hoặc đôi khi chữ hoa đầu tiên. Kèm theo kí hiệu còn thêm dấu + hoặc – hoặc chữ tắt để giải thích rõ thêm tính trạng. Ví dụ: lac– để chỉ mất khả năng tổng hợp lactoz; his+/- tổng hợp histidin; strS – nhạy cảm (Sensible) với Streptomycin, strR – đề kháng (Resistant) với Streptomycin.
Để chỉ hai giới tính khác nhau không dùng hai kí hiệu ♀ và ♂ thay vào đó là các chữ như mt (+), mt (-) (mating typ) (ở Chlamydomonas reinhardi), hoặc A, a (ở Neurospora crassa) hay a và α (vi nấm).
Hình 3. Tảo đơn bào Chlamydomonas
2.4. Cấu trúc bộ gen
- Bộ gen của vi khuẩn E. coli và đa số các sinh vật nhân sơ là một phân tử ADN có dạng vòng tròn kín. Khái niệm nhiễm sắc thể hiện nay được dùng cho cả vi khuẩn, nên nói nhiễm sắc thể vi khuẩn ta hiểu đó là sợi ADN. Ty thể và lục lạp cũng có ADN riêng, mà cấu trúc bộ gen cũng tương tự vi khuẩn.
- Đa phần ADN của Eukaryota như nấm sợi, nấm men, vi tảo được tổ chức thành nhiều nhiễm sắc thể trong nhân tế bào. Mỗi nhiễm sắc thể chứa 1 phân tử ADN thẳng mạch kép kèm theo một số prôtêin như histon. Các nhiễm sắc thể có số lượng và hình dạng đặc trưng cho tế bào của mỗi loài sinh vật nhân chuẩn.
- Các virut có bộ gen rất đa dạng: ADN mạch đơn hoặc kép, ARN mạch đơn hoặc kép, nhưng mỗi chủng chỉ mang một loại axit nuclêic.
2.5. Kiểu sinh sản
- Các nấm sợi, nấm men, vi tảo có các quá trình sinh sản vô tính và hữu tính về căn bản giống các sinh vật bậc cao, thực hiện qua nguyên phân và giảm phân.
- Các vi khuẩn không có các cơ chế nguyên phân và giảm phân, phân chia tế bào theo cơ chế trực phân, nhưng cũng có sinh sản vô tính và sự trao đổi thông tin di truyền tương tự sinh sản hữu tính được gọi là quá trình cận hữu tính.
- Các virut chỉ sinh sản trong tế bào chủ sống với nhiều cơ chế khác nhau phụ thuộc kiểu virut.
3. Biến dị ở vi sinh vât.
3.1. Quá trình đột biến tự nhiên
- Trong tự nhiên, dù giữ trong điều kiện nào, tất cả các gen đều có đột biến, được gọi là đột biến tự nhiên hay ngẫu nhiên (spontanous mutation). Các đột biến tự nhiên thường xuất hiện rất ít. Khái niệm tần số đột biến được dùng để đánh giá mức độ xuất hiện nhiều hay ít đột biến ở một gen. Có người phân biệt tần số (frequency) với tốc độ (rat) đột biến.
- Các gen khác nhau của cùng 1 sinh vật có thể có tần số đột biến khác nhau. Nhưng tần số đột biến tự nhiên đối với mỗi gen là một số ổn định.
- Tần số đột biến được đánh giá theo các căn cứ khác nhau như: trên 1 lần sao chép, 1 lần phân bào hay trên 1 giao tử và trên 1 tế bào / 1 thế hệ
- Để dễ hiểu các số trên có thể tính đảo ngược lại. Ví dụ: Ở E. coli đột biến từ nhạy cảm với streptomycin sang kháng tức StrS → StrR với tần số 4.10-10 đột biến tính trên 1 tế bào/1 thế hệ. Để dễ hiểu ta có thể tính ngược lại tức trong 10 tỉ tế bào của một thế hệ có 4 đột biến StrR (resistance) kháng streptomycin xuất hiện ngẫu nhiên.
- Tuy tần số đột biến của từng gen là rất thấp, nhưng tổng các đột biến của nhiều gen là một số đáng kể, có ý nghĩa quan trọng cho tiến hóa.
- Đột biến ảnh hưởng đến mọi tính trạng khác nhau của sinh vật và tác động theo mọi hướng.
3.2. Các loại đột biến thường gặp ở vi sinh vật
3.2.1. Đột biến gen hay đột biến điểm
Biến đổi rất nhỏ trên một đoạn ADN, thường liên quan đến một nucleotit hay một cặp nucleotit.
- Đột biến đồng nghĩa (Samesense) còn gọi là trung tính (neutral) hay im lặng (silent), khi côđôn mã hóa cho một axit amin bị biến đổi, thường ở bazơ thứ ba nên vẫn mã hóa cho axit amin đó (do tính thoái hóa của mã di truyền).
- Đột biến vô nghĩa (Non-sense) khi côđôn mã hóa cho một axit amin biến thành một trong ba côđôn 5′ UAA 3′ , 5′ UAG 3′ và 5′ UGA 3′ là các côđôn kết thúc không mã hóa cho axit amin nào.
- Đột biến sai nghĩa (Mis-sense): Khi côđôn của axit amin này biến thành côđôn mã hóa cho axit amin khác, làm thay đổi axit amin tương ứng trên phân tử prôtêin.
- Đột biến lệch khung: Sự thêm một bazơ hay làm mất một bazơ dẫn đến các côđôn sai nghĩa hay vô nghĩa so với côđôn tương ứng ban đầu từ điểm biến đổi về sau, sự dịch mã bị lệch khung có tính dây chuyền từ bộ ba bị sai.
3.2.2. Đột biến nhiễm sắc thể
Các đột biến nhiễm sắc thể hay còn gọi là sai hình nhiễm sắc thể xuất hiện ở sinh vật nhân thực.
- Biến đổi trên một nhiễm sắc thể: mất đoạn, lặp đoạn, đảo đoạn.
- Biến đổi giữa các nhiễm sắc thể: chuyển đoạn.
- Đột biến bộ gen (genome mutation)
- Đa bội thể (Polyploidy) hiểu theo nghĩa rộng là sự thay đổi số lượng nhiễm sắc thể gồm: Đa bội thể nguyên (Polyploidy hay Euploidy) (2n → 3n, 4n, …), đa bội thể lai còn gọi dị bội thể (Alloploidy)(2n A + 2n B) và đa bội lệch (Aneuploidy), hay đa nhiễm:(ví dụ: 2n + 1 hoặc 2n – 1).

3.2.3. Các biến đổi vi cấu trúc
Các thay đổi thành phần nucleotit của gen.
a) Đột biến thay thế: Thay một nucleotit này bằng nucleotit khác.
- Đồng chuyển (Transition) khi pyrimidin được thay thế bởi pyrimidin hay purin bởi purin. Ví dụ: T thay cho C ; A thay cho G hoặc ngược lại.
- Đảo chuyển (Transversion) khi pyrimidin được thay thế bởi purin hay purin bởi pyrimidin.Ví dụ: T hay C thay cho A hoặc G và ngược lại.
b) Mất nucleotit (Deletion): Mất một phần nucleotit của gen.
c) Đột biến xen nucleotit (Insertion mutant): Thêm 1 hay nhiều nucleotit vào gen.

3.2.4. Các đột biến kiểu hình
a. Các đột biến hình thái: Các biến đổi ảnh hưởng đến hình dạng, màu sắc và kích thước. Ví dụ: một dạng đột biến khuẩn lạc màu đỏ ở Serratia marcescens.
Đột biến thuận Đột biến nghịch.
- Đột biến ức chế hay kìm hãm (Supressor): Đột biến ở một điểm khác. Cả hai cùng tạo kiểu hình gần như hoang dại.
- Đột biến kìm hãm ngoài gen: Xảy ra ở gen khác với gen bị đột biến.
- Đột biến kìm hãm trong gen: Xảy ra ở nucleotit khác trong gen đưa gen trở về trạng thái tạo kiểu hình hoang dại.
4. Di truyền vi khuẩn.
4.1. Kiểu hình và kiểu gen của vi khuẩn.
Các đột biến có thể tác động đến 5 loại kiểu hình:
- Biến đổi từ nguyên dưỡng (prototrophy) sang khuyết dưỡng (auxotrophy) và ngược lại. Ví dụ: mất khả năng tổng hợp một chất trao đổi của chu trình và hồi biến để lại có khả năng tổng hợp chất đó.
- Sự mất hay có được khả năng sử dụng chất dinh dưỡng khác. Ví dụ: đột biến làm mất khả năng sử dụng đường lactose.
- Tính nhạy cảm hay đề kháng thuốc như nhạy cảm streptomycin đột biến thành kháng streptomycin.
- Nhạy cảm với phagơ thành kháng phagơ hoặc ngược lại. Ví dụ: đột biến trên thụ thể ở bề mặt tế bào làm đề kháng với sự nhiễm phagơ.
Hình 4. Đột biến khuẩn lạc màu đỏ ở Serratia marcescens
b. Đột biến sinh hóa:
- Các đột biến khuyết dưỡng (Auxotrophe mutation) làm mất khả năng tổng hợp các chất.
- Các đột biến có điều kiện: Các đột biến có thể không có biểu hiện trong những điều kiện giới hạn nhất định và có biểu hiện trong các điều kiện cho phép. Ví dụ, các đột biến nhạy cảm với nhiệt độ cao có biểu hiện ở nhiệt độ tương ứng.
- Đột biến đề kháng: Các biến đổi sinh hóa giúp kháng lại được các tác nhân bất lợi.
3.2.5. Đột biến và hồi biến
a. Đột biến thuận hay trực tiếp: Biến đổi từ kiểu hình hoang dại sang khác thường.
b. Hồi biến: Đột biến từ kiểu hình đột biến quay về kiểu hình hoang dại.
- Hồi biến thật hay đột biến nghịch. Biến đi trở lại y như ban đầu.
Ví dụ: adenin → guanin → adenin
- Sự mất đi hoặc có lại các thành phần cấu trúc của bề mặt tế bào. Ví dụ: một loại Pneumococcus có vỏ bao polyxacarit (polyxacarit capsule), trong khi đó dòng khác không có vỏ bao.
Các kí hiệu dùng biểu hiện kiểu hình và kiểu gen được thống nhất theo nguyên tắc:
- Kí hiệu kiểu hình gồm 3 chữ thường (chữ đầu viết hoa) với dấu phía trên góc “+” hay “–” nhằm chỉ sự hiện diện hay thiếu tình trạng tương ứng, và “s” hay “r” tương ứng chỉ tính nhạy cảm (sensitive) hay đề kháng (resistance).
- Kí hiệu kiểu gen được viết chữ nghiêng, chữ đầu không viết hoa.
Ví dụ 1: Tế bào hoang dại tự tổng hợp được leuxin thì kiểu hình được viết Leu+ Đột biến khuyết dưỡng mất khả năng tổng hợp leuxin được kí hiệu kiểu hình Leu–. Tương ứng với hai kiểu hình trên kí hiệu kiểu gen là leu+ hay leu và leu–
Nếu cần nhiều hơn một gen để tạo ra một chất nhất định, sau 3 chữ nghiêng kí hiệu gen có thêm chữ nghiêng hoa. Ví dụ: leuA, leuB... là các gen cần thiết cho tổng hợp leuxin khác nhau.
Ví dụ 2: Kiểu hình kháng hoặc nhạy cảm với penixilin được viết là Penr và Pens . Kiểu gen tương ứng là penr và pens .
Nếu lưỡng bội ở một phần thì viết thêm gạch nghiêng. Ví dụ, leu+/leu-
4.2. Các phương pháp phân tích di truyền
Ngoài các phương pháp lai để phân tích tái tổ hợp và bổ trợ, có nhiều kỹ thuật biến đổi di truyền.
* Gây đột biến (Mutagensis):
- Hóa chất và chiếu xạ: đột biến xoma ngẫu nhiên.
- Dùng transposon: xen đoạn (Insertions) xoma ngẫu nhiên.
* Chuyển gen (Transgensis):
- Trên vectơ plasmit: tự do hay chèn vào (integratd).
- Trên vectơ phagơ: tự do hay chèn vào (integratd).
- Biến nạp: chèn vào
* Làm im lặng gen mục tiêu (Targeted gen knockout):
- Alen không (Null alen) trên vectơ: thay gen bằng tái tổ hợp.
- Alen được thiết kế (Engineered alen) trên vectơ: đột biến điểm định hướng (Site-directed mutagensis) bằng thay gen.
4.3. Cơ chế di truyền ở vi khuẩn
4.3.1. Biến nạp.
4.3.1.1. Hiện tượng và điều kiện
Hiện tượng biến nạp được Griffith phát hiện ở vi khuẩn Diplococus pneumoniae vào năm 1928. Vi khuẩn này có 2 dạng khác nhau:

- Dạng SIII, gây bệnh có vỏ bao tế bào (capsule) bằng polysaccharid cản trở bạch cầu phá vỡ tế bào. Dạng này tạo đốm mọc (khuẩn lạc) láng trên môi trường agar.
- Dạng RII, không gây bệnh, không có vỏ bao, tạo đốm mọc nhăn.
Thí nghiệm tiến hành như mô tả trên hình 5.
Hình 5. Hiện tượng biến nạp.
- Tiêm vi khuẩn S sống gây bệnh cho chuột - chuột chết.
- Tiêm vi khuẩn R sống không gây bệnh - chuột sống.
- Tiêm vi khuẩn S bị đun chết cho chuột - chuột sống.
- Hỗn hợp vi khuẩn S bị đun chết trộn với vi khuẩn R sống đem tiêm cho chuột - chuột chết. Trong xác chuột chết có vi khuẩn S và R.
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các tế bào chết này đã truyền tính gây bệnh cho tế bào R. Nó được gọi là biến nạp (transformation). Năm 1944, T.Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng protease (enzym phân hủy prôtêin) hoặc ARN-ase (enzym phân hủy ARN) hoạt tính biến nạp vẫn còn, chứng tỏ prôtêin và ARN không phải là tác nhân gây biến nạp. Nhưng nếu tế bào S chết bị xử lý bằng ADN-ase (enzym chỉ phân hủy đặc hiệu ADN) thì hoạt tính biến nạp không còn nữa, chứng tỏ ADN là nhân tố biến nạp. Kết quả thí nghiệm có thể tóm tắt như sau:
ADN của S + các tế bào R sống → chuột → chết (có R + S)
Hiện tượng biến nạp là một chứng minh sinh hóa xác nhận rằng ADN mang tín hiệu di truyền.
Như vậy, biến nạp là hiện tượng truyền thông tin di truyền bằng ADN. Trong biến nạp, ADN trần từ một tế bào vi khuẩn (thể cho) này được truyền sang tế bào vi khuẩn khác (thể nhận). Biến nạp xảy ra khi vi khuẩn nhận ADN ngoại lai và hấp thu
vào trong tế bào. Khi tế bào vi khuẩn bị vỡ do bị tan (lysis), ADN vòng tròn của chúng thoát ra môi trường thành các đoạn thẳng với chiều dài khác nhau, có khả năng gây biến nạp cho các tế bào nhận khác.
Hiệu quả của biến nạp phụ thuộc vào 3 yếu tố:
- Tính dung nạp của tế bào nhận.
- Kích thước của đoạn ADN ngoại lai.
- Nồng độ của ADN

4.3.1.2. Tính dung nạp của tế bào nhận.
Một điều quan trọng của biến nạp là tế bào nhận phải có trạng thái sinh lí đặc biệt được gọi là khả năng dung nạp hay khả nạp. Tế bào có khả năng hấp thu ADN ngoại lai và được biến nạp gọi là khả nạp và đây là tính trạng di truyền. Tính khả nạp trong phần lớn các vi khuẩn biến nạp tự nhiên được kiểm soát và các prôtêin đặc hiệu tham gia vào hấp thu và tác động đến ADN. Các prôtêin đặc hiệu khả nạp đó gồm prôtêin gắn ADN liên kết màng, autolysin vách tế bào và các nucleaz khác nhau.
Biến nạp tự nhiên hiệu quả cao được phát hiện chỉ ở một ít loài vi khuẩn. Tuy nhiên, có thể gây ra sự dung nạp bằng xử lý hóa chất hay tạo những điều kiện nhất định cho sự tăng trưởng của tế bào. Khi xử lý tế bào E. coli với ion canxi nồng độ cao, chúng trở thành khả nạp và biến nạp các plasmit thực hiện có hiệu quả.
Phần lớn các vi khuẩn chỉ dung nạp trong một giai đoạn giới hạn của chu trình sống. Trong giai đoạn dung nạp, tế bào tổng hợp một hay nhiều prôtêin được gọi là “các nhân tố dung nạp”, chúng biến đổi màng tế bào để có thể gắn với đoạn ADN ngoại lai. Như vậy, các điểm thụ thể chỉ hiện diện trong giai đoạn dung hợp.
4.3.1.3. Sự hấp thu ADN. Trong biến nạp, vi khuẩn khả nạp gắn thuận nghịch với ADN. Các tế bào khả nạp có thể gắn ADN nhiều hơn cả 1000 lần so với tế bào không khả nạp.
Điều kiện quan trọng thứ hai để thực hiện được biến nạp là ADN phải có mạch kép và đoạn biến nạp phải có trọng lượng phân tử tối thiểu là 400.000 dalton. Số lượng tế bào được biến nạp tăng tỉ lệ thuận với nồng độ của ADN cho đến lúc mà các điểm thụ thể bão hòa do các đoạn ADN gắn vào (thường khoảng 10 đoạn/tế bào).
Mật độ ADN tối thiểu để có thể phát hiện biến nạp là 0,01 ng/ml, một con số thấp đến nỗi không thể phát hiện bằng phương pháp hóa học.
4.3.1.4. Đoạn ngoại lai và đoạn nội tại.
mạch đơn không bền vững và bị phân hủy nếu không được gắn vào bộ gen thể nhận.
Quá trình trao đổi thông tin di truyền bằng chuyển chỉ một phần vật liệu di truyền từ tế
bào này sang tế bào khác được gọi là sự tiếp hợp từng phần.
Nếu đoạn ngoại lai chứa một alen với đoạn nội tại, thì mạch kép tái tổ hợp sẽ có một hay nhiều chỗ bắt cặp sai và đoạn ADN này được gọi là heteroduplex. Nếu các tế bào con nhận alen mới, sửa sai được thực hiện bằng cách cắt đoạn nội tại và dùng mạch ngoại lai làm khuôn để thay thế. Sự gắn đoạn ngoại lai vào ADN tế bào nhận được thực hiện nhờ tái tổ hợp tương đồng
Hai hoặc nhiều hơn các gen liên kết chặt có thể nằm trong đoạn ADN biến nạp. Nếu sự xâm nhập của hai hay nhiều gen vào đoạn nội tại, thì tế bào nhận sẽ được đồng biến nạp. Tần số của đồng biến nạp tỉ lệ nghịch tương ứng với khoảng cách giữa các gen cùng biến nạp.
4.3.1.5. Cơ chế phân tử
Diễn biến của quá trình biến nạp ở cấp độ phân tử được tóm tắt trên sơ đồ hình 6. Để dễ hiểu, các giải thích dựa theo ADN của các dòng vi khuẩn S và R trong thí nghiệm của Griffith. Quá trình gồm các giai đọan chủ yếu:
- Sự phân hủy ADN tế bào cho.
- ADN bám vào bề mặt tế bào.
- Thâm nhập của ADN.
- Bắt cặp (Synapsis) và tái tổ hợp.
- Sao chép.
Khi ADN mạch kép xâm nhập vào tế bào, một mạch bị phân hủy. Bất kì đoạn ADN nào từ tế bào cho xâm nhập tế bào nhận, được gọi là đoạn ngoại lai, ADN nguyên vẹn của tế bào nhận gọi là đoạn nội tại. Tế bào vi khuẩn nhận đoạn ngoại lai sẽ lưỡng bội ở một phần bộ gen, được gọi là hợp tử từng phần. Tuy nhiên, đoạn ngoại lai 244
Hình 6. Sơ đồ diễn biến của quá trình biến nạp ở cấp độ phân tử
4.3.2. Tải nạp
Là hiện tượng truyền ADN qua trung gian virut từ tế bào cho đến tế bào nhận. Có hai kiểu tải nạp: tải nạp chung và tải nạp chuyên biệt.
Trong tải nạp, các virut mang các gen từ tế bào này sang tế bào khác. Ở chu trình tan, một số bacteriophagơ gói nhầm ADN vi khuẩn chủ vào capsid. Tế bào bị nhiễm bởi các virut như vậy nhận đoạn ADN của vi khuẩn A khác, chứ không phải ADN của virut. Do vậy, ADN vi khuẩn A tái tổ hợp với ADN nhiễm sắc thể của tế bào chủ B và biến đổi thành phần di truyền.
4.3.2.1. Phagơ là nhân tố chuyển gen.
Thí nghiệm được tiến hành trong ống hình chữ U, ở đáy ống được ngăn cách bằng màng lọc vi khuẩn. Màng có lỗ nhỏ vi khuẩn không qua được, nhưng phagơ qua được. Nhánh A của ống chứa vi khuẩn có khả năng tổng hợp tryptophan (trp+), còn nhánh B nuôi các vi khuẩn mất khả năng tổng hợp tryptophan (trp–). Sau một thời gian nuôi bên nhánh B xuất hiện vi khuẩn có khả năng tổng hợp tryptophan (trp+). Qua nhiều lần thí nghiệm việc phagơ tải gen trp+ từ nhánh A sang nhánh B được chứng minh.
Do không có sự tương đồng giữa trình tự ADN trên các phagơ này với trình tự ở tế bào chủ, nên không có điểm gắn vào đặc hiệu cho prophagơ. Bất kì gen nào cũng được tải nạp vì đầu của phagơ có thể gói nhầm vào một đoạn ADN của tế bào chủ. Sự đồng tải nạp (cotransduction) là quá trình tải nạp đồng thời 2 gen.
Quá trình phagơ xâm nhập vào vi khuẩn và sinh sản được mô tả trên hình 8. Hình 8. Quá trình xâm nhập của phagơ và làm tan vi khuẩn
4.3.2.3. Tải nạp chuyên biệt.
Tải nạp chuyên biệt là trường hợp chỉ mang một vài gen nhất định, có 4 đặc điểm:
- Những gen được chuyển nằm sát chỗ prophagơ gắn vào.
- Chỉ prophagơ kiểu λ thực hiện.
- Do kết quả sự cắt sai của prophagơ khi tách khỏi nhiễm sắc thể của tế bào chủ
- Các vi khuẩn tái tổ hợp có thể lưỡng bội một phần.
Ví dụ: Phagơ λ chỉ mang gen gal (đồng hóa đường galactose) từ vi khuẩn này chuyển sang vi khuẩn khác.
Hình 7. Thí nghiệm chứng minh có tải nạp do virut

4.3.2.2. Tải nạp chung
Tải nạp chung xảy ra khi phagơ mang bất kì gen nào của vi khuẩn A chuyển sang vi khuẩn B.
Tải nạp chung (genralized transduction) có các đặc điểm:

- Thường do phagơ kiểu P1 thực hiện.
- Bất kì gen nào của vi khuẩn cũng đều được tải nạp.
- Tải nạp có được do gói nhầm ADN của tế bào chủ khi phagơ trưởng thành.
- Các thể tái tổ hợp đơn bội được tạo ra.
Hình 9. Điểm gắn của phagơ λ vào bộ gen vi khuẩn
4.3.3. Tiếp hợp
Tiếp hợp ở vi khuẩn là sự kết hợp nhất thời của hai tế bào có kiểu bắt cặp đối nhau, được tiếp nối bằng sự chuyển một phần vật chất di truyền từ tế bào cho sang tế bào nhận qua cầu tế bào chất, và sau đó các tế bào tách nhau ra. Tiếp hợp đòi hỏi sự tiếp xúc trực tiếp giữa 2 loại tế bào được khởi sự bằng ống tiếp hợp hay tính mao (pilus), một sợi ống nhỏ rất dài do tế bào cho tạo ra.
4.3.3.1. Chứng minh có hiện tượng lai ở vi khuẩn
Vào năm 1946, J.Lederberg và E.Tatum đã sử dụng các dòng đột biến khuyết dưỡng khác nhau ở E. coli để chứng minh có tái tổ hợp giữa các dòng vi khuẩn khác nhau. Cụ thể dòng A có kiểu gen met–bio–thr+leu+thi+ (có khả năng tổng hợp threonin, leuxin và vitamin B1 (thiamin) và không tổng hợp được metionin và biotin), còn ở dạng B thì ngược lại có khả năng tổng hợp metionin và biotin với kiểu gen met+bio+thr–leu– thi– Trộn A và B trong ống nghiệm, sau đó cấy lên môt môi trường tối thiểu (Minimal medium MM). Các khuẩn lạc mọc trên môi trường tối thiểu chứng tỏ có các dạng lai, chúng chỉ mọc lên được nhờ sự bù đắp cho nhau các nhu cầu dinh dưỡng.
Dạng lai có kiểu gen met+bio+thr+leu+thi+ mọc được trên môi trường tối thiểu. Trong khi đó từng dạng A hoặc B riêng lẻ không mọc được trên MT tối thiểu (hình 11)
Dòng A = met–bio–thr+leu+thi+ × dòng B= met+bio+thr–leu–thi–


Dạng lai met+bio+thr+leu+thi+ mọc thành khuẩn lạc, mỗi dạng riêng lẻ không mọc thành khuẩn lạc.
Hình 10. Hai tế bào vi khuẩn tiếp hợp qua cầu tế bào chất pilus
ADN được truyền qua một cầu tế bào chất mỏng gọi là ống tiếp hợp. Kiểu sao chép sigma (σ) được tế bào vi khuẩn sử dụng trong tiếp hợp để truyền phân tử ADN dạng thẳng sang tế bào khác. Khi đọan ADN vào bên trong tế bào nhận nó thực hiện tái tổ hợp với các gen tương đồng.
Hình 11. Sơ đồ lai vi khuẩn (MM: môi trường tối thiểu)
4.3.3.2. Sự phân hóa giới tính ở vi khuẩn
Vi khuẩn có các dạng khác nhau tương tự giống “đực” và “cái” ở sinh vật bậc cao. Các dạng đó được kí hiệu F+ và F–. F+ tương tự giống đực có ở sinh vật bậc cao, nó truyền gen sang F–. Tần số lai F+ x F– khoảng 10–6 tức lai 1 triệu tế bào sẽ có 1 tế bào lai.
a. Epixom và plasmit
Khi tiếp xúc với F+ một thời gian tế bào F– trở thành F+ . Về sau dạng Hfr được phát hiện, dạng này có tần số lai với F– cao hơn F+ có thể đến 104 lần.
Khi F+ tiếp xúc với F–một thời gian, F– biến thành F+ do nó nhận được một phần tử di truyền gọi là epixom. Epixom F+ là phần tử di truyền ngoài nhiễm sắc thể, có thể tồn tại hoặc ở dạng phân tử ADN vòng tròn tự sao chép hoặc gắn vào phân tử ADN của tế bào chủ (ví dụ như phagơ λ). Epixom F+ được gọi là nhân tố giới tính.
Plasmit lúc đầu được định nghĩa là phân tử ADN vòng tròn nhỏ có khả năng sao chép độc lập với nhiễm sắc thể tế bào chủ và không có khả năng gắn vào nhiễm sắc thể tế bào chủ. Plasmit có thể mang một số gen khác nhau như đề kháng thuốc (ví dụ, plasmit R đề kháng nhiều thuốc kháng sinh),... Hiện nay “plasmit” được dùng cho cả hai nghĩa là epixom lẫn plasmit. Các plasmit có thể tồn tại độc lập hoặc gắn vào bộ gen vi khuẩn. Về sau người ta phát hiện ở vi khuẩn còn có nhiều plasmit khác. Bản chất di truyền của các dòng F–, F+ và Hfr được xác định do các plasmit như sau:
F– không chứa plasmit.
F+ chứa plasmit ở dạng tự do không gắn vào bộ gen vi khuẩn.
Hfr - plasmit được gắn vào bộ gen vi khuẩn
Hfr X F– Sao chép ADN truyền cho F– Hfr F– tái tổ hợp
Hình 13. Sự truyền ADN từ thể cho sang thể nhận
Các gen A, B, C được chuyển một chiều từ Hfr sang F– Đọan gen thể cho bắt cặp với các gen tương đồng của ADN tế bào F– và diễn ra trao đổi chéo tạo tế bào F–tái tổ hợp (Hình 14).
Dòng Hfr có tần số lai cao hơn nhiều vì plasmit đã nằm sẵn trong bộ gen. Còn F+ phải qua giai đoạn plasmit gắn vào bộ gen rồi mới chuyển gen.
Trong điều kiện thí nghiệm ở 37oC, nguyên bộ gen của tế bào E. coli được chuyển sang tế bào nhận trong vòng 90 phút. Thường sự tiếp hợp bị ngắt giữa chừng do các pilus bị gãy, nên ít khi bộ gen được chuyển nguyên vẹn vào tế bào nhận. Lúc đó tế bào F–vẫn là F–. Bằng cách ngắt quãng tiếp hợp bản đồ di truyền của E. coli được xây dựng và có dạng 1 vòng tròn.
Hình 12. Quá trình chèn ADN plasmit vào ADN Nhiễm sắc thể tế bào vi khuẩn
b. Các nhân tố F’ và tính nạp

Sự cắt rời nhân tố F từ nhiễm sắc thể của dòng Hfr nhiều khi không chính xác và lúc đó một đoạn bộ gen của vi khuẩn thay thế một phần của F. Trong trường hợp này, nhân tố F’ được tạo thành và nó có khả năng chuyển gen của vi khuẩn một cách độc lập, nhưng với các tính trạng của tế bào cho. Hiện tượng này được gọi là tính nạp, có nghĩa là sự chuyển gen kèm theo nhân tố giới tính. Nhờ tính nạp có thể nhận được các thể lưỡng bội từng phần theo các gen được gắn vào F+
4.3.3.3. Tái tổ hợp Muốn xảy ra tái tổ hợp thì 2 dòng vi khuẩn phải tiếp xúc với nhau (F+ x F- ) hoặc (Hfr x F-). Dòng tế bào mang nhân tố F+ được coi là tế bào “đực” và có khả năng tạo prôtêin pilin, từ prôtêin này tạo ống tiếp hợp là pilus. Sự co lại của pilus đang nối hai tế bào làm chúng tiếp xúc kề nhau. Tế bào F– được coi là cái (female). Sau khi tiếp hợp tế bào F– trở thành F+
Việc chuyển gen chỉ thực hiện khi plasmit gắn vào bộ gen của vi khuẩn. Trong quá trình chuyển vật chất di truyền sang F– thì ADN của tế bào chủ sao chép và mạch mới có ori đi đầu và F ở cuối (Hình 13). Quá trình chuyển ADN từ F+ sang F– có thể bị ngắt quãng.
Hình 14. Đoạn ADN từ thể cho trao đổi chéo với ADN thể nhận tạo F– tái tổ hợp


4.4. Lập bản đồ di truyền nhiễm sắc thể của vi khuẩn
4.4.1. Tiếp hợp ngắt quãng
Khi trộn dịch tế bào Hfr với F–, sự tiếp hợp có thể bị ngắt quãng vào thời điểm tùy ý bằng rung lắc mạnh. Mẫu được pha loãng nhanh và cấy lên môi trường chọn lọc. Ngoài ra, dòng Hfr phải có gen đánh dấu hay khuyết dưỡng để chúng không mọc được trên môi trường chọn lọc và chỉ có các dạng tái tổ hợp mọc lên. Sự truyền ADN của Hfr sang F– có tính phân cực, các gen ở đầu sang trước và lần lượt các gen khác theo sau. Nhờ ngắt quãng có thể xác định được thời gian mà một gen được truyền sang F–và khoảng cách giữa chúng.
4.4.2. Tiếp hợp không ngắt quãng
Nếu không rung mạnh để ngắt quãng nhân tạo, cầu tế bào chất giữa hai gen càng gần ori (ở đầu của ADN thể cho), xác suất được chuyển sang tế bào nhận càng lớn. Tần số của các gen xuất hiện ở dạng tái tổ hợp trong thể nhận, được phát hiện trên môi trường chọn lọc, tỉ lệ nghịch với khoảng cách so với gen đánh dấu ở đầu. Các gen
có nhiều dạng tái tổ hợp xuất hiện thì càng nằm gần phía đầu được chuyển sang tế bào nhận.
Khoảng cách giữa các gen có thể tính bằng 3 loại đơn vị: đơn vị thời gian, đơn vị tái tổ hợp và đơn vị cặp bazơ.
4.4.3. Lập bản đồ bằng tái tổ hợp
Đặc điểm của tái tổ hợp ở vi khuẩn là chỉ một phần bộ gen được chuyển sang tế bào nhận. Một hay nhiều gen có thể được gắn vào nhiễm sắc thể tế bào nhận nhờ tiếp hợp và phụ thuộc độ dài đoạn truyền sang. Exogenote phải được gắn vào nhiễm sắc thể nhận thì mới được sao chép và chia về các tế bào trong dòng. Chỉ một đoạn ngắn ADN thường được gắn nhờ biến nạp và tải nạp. Như vậy, nếu hai gen cùng biến nạp (cotransformation) thì hai locut phải liên kết chặt. Tương tự như vậy, hai gen cùng được tải nạp vào phagơ sẽ ở gần nhau. Mức độ liên kết giữa các gen có chức năng khác nhau (intercistronic) hay giữa các đột biến bên trong gen chức năng (intracistronic) có thể đánh giá từ các kết quả lai chuyên biệt.
Ở hệ thống hợp tử một phần (merozygote), exogenote muốn gắn vào endogenote phải xảy ra 2 trao đổi chéo ở hai đầu vì nếu chỉ có 1 trao đổi chéo thì cả hai sản phẩm nhận được đều mất cân bằng.
Trong các hệ thống này, tổng cá thể tham gia tái tổ hợp không biết được, nên tần số tái tổ hợp không thể tính như ở ruồi giấm (trên tổng số cá thể). Do đó, tần số tái tổ hợp được trình bày tương đối theo một số tiêu chuẩn chung cho tất cả các tổ hợp lai.
Ví dụ, số lượng các cá thể tái tổ hợp nguyên dưỡng được tạo ra khi lai hai dòng đột biến có so sánh với kết quả khi lai dạng hoang dại với mỗi loại đột biến. Tuy nhiên có nhiều sai lệch và cần chỉnh lí.
Xác định trình tự gen ở vi sinh vật thường phức tạp vì có thể xảy ra hiện tượng nhiễu âm cục bộ (localized negative interference). Phương pháp xác định trình tự gen thường dùng là lai 3 điểm hoán đổi (three - factor reciprocall crosses).
4.5. Plasmit
4.5.1.Tính chất vật lý và sự sao chép plasmit
Hầu hết các plasmit đã biết đều có ADN mạch kép, mạch vòng. Tuy nhiên vẫn có nhiều plasmit mạch thẳng. Các plasmit tự nhiên có kích thước rất khác nhau, từ 1 kbp đến 1 megabp. Plasmit điển hình là một phân tử ADN vòng tròn, mạch kép nhỏ chưa đến 5 % kích thước nhiễm sắc thể (Hình 15). Phần lớn những plasmit ADN được phân lập từ tế bào có dạng siêu xoắn, dạng nhỏ gọn nhất có thể tồn tại trong tế bào.
Enzim liên quan đến việc sao chép plasmit trên thực tế là những enzim bình thường trong tế bào. Vì thế những gen của plasmit chủ yếu liên quan đến sự kiểm soát quá trình khởi sự sao chép và với sự phân bổ những plasmit được sao chép vào các tế bào con.
Hình 15. Hình hiển vi điện tử các plasmit vòng tròn
Tương tự, những plasmit khác nhau hiện diện trong tế bào với số lượng khác nhau, gọi là số bản sao (copy number). Một số plasmit chỉ có 1-3 bản sao trong tế bào, trong khi những plasmit khác lại có đến 100 bản sao.
Số lượng bản sao được kiểm soát bởi những gen trên plasmit và bởi sự tương tác gen giữa tế bào chủ và plasmit. Phần lớn các plasmit ở vi khuẩn Gram âm sao chép tương tự như nhiễm sắc thể vi khuẩn. Việc này thực hiện từ điểm khởi sự sao chép (Ori) và chép theo hai hướng quanh vòng plasmit, tạo ra dạng trung gian theta (θ). Tuy nhiên một số plasmit không sao chép theo hai hướng. Do kích thích nhỏ của ADN plasmit, toàn bộ quá trình sao chép xảy ra rất nhanh, có lẽ bằng 1/10 tổng thời gian diễn ra chu kì phân bào.
Hầu hết plasmit của vi khuẩn Gr+ sao chép theo cơ chế vòng xoay tương tự như phagơ φx174. Cơ chế này sẽ tạo ra mạch đơn trung gian, và do đó những plasmit này thỉnh thoảng được xem như plasmit ADN mạch đơn. Phần lớn ADN mạch thẳng sao chép dùng cơ chế liên quan đến một prôtêin gắn với đầu 5′ của mỗi mạch mà được dùng trong tổng hợp mồi ADN.
4.5.2. Tính không tương hợp (Incompatibility) và chỉnh sửa (curing)
Một số vi khuẩn có thể chứa vài dạng plasmit khác nhau. Ví dụ, Borrelia burgdorferi (nhân tố gây bệnh Lyme) chứa 17 plasmit mạch thẳng và vòng. Khả năng hai plasmit khác nhau, mà mỗi một sao chép trong cùng một tế bào, được kiểm soát bởi gen của các plasmit tham gia điều hòa sao chép ADN.

Khi một plasmit được chuyển vào trong tế bào đã có sẵn một plasmit khác, thường thì plasmit thứ hai có thể không tồn tại được và mất đi trong quá trình sao chép tế bào sau đó. Nếu điều này xảy ra, hai plasmit trên được gọi là không tương hợp (incompatible). Một số nhóm không tương hợp (Incompatibility - Inc) đã được ghi nhận. Các plasmit thuộc cùng một nhóm không tương hợp sẽ loại trừ nhau khi sao chép trong cùng tế bào nhưng có thể cùng tồn tại với plasmit của nhóm khác.
Plasmit cùng nhóm không tương hợp có chung cơ chế điều hòa sao chép, do đó chúng có họ hàng với nhau. Vì vậy, dù tế bào vi khuẩn có thể chứa nhiều loại plasmit khác nhau, thì mỗi loại vẫn khác biệt nhau về mặt di truyền.
4.5.3. Sự chuyển plasmit từ tế bào này sang tế bào khác
Cơ chế chính của việc chuyển plasmit từ tế bào này sang tế bào khác là tiếp hợp, mà chức năng thực hiện được mã hóa bởi chính một số plasmit. Tiếp hợp là một quá trình sao chép ADN và plasmit của tế bào cho và truyền bản sao của nhiễm sắc thể cùng bản sao plasmit sang tế bào nhận.
Hình 16 cho thấy bản đồ gen của plasmit F, một plasmit đã được nghiên cứu rất kĩ của Escherichia coli. Khoảng 25 gen trên F plasmit, bao gồm các gen sao chép ADN, các yếu tố thực hiện chức năng của một epixom và cuối cùng là một vòng cung
ADN lớn, gọi là vùng tra, chứa những gen giúp nó tự di chuyển từ tế bào này sang tế bào khác.
Plasmit R lần đầu tiên được phát hiện tại Nhật ở dòng vi khuẩn đường ruột ; đó là những vi khuẩn có tính đề kháng đối với nhiều lọai kháng sinh. Một plasmit R có thể mang nhiều gen đề kháng.
Hình 16. Sơ đồ gen của plasmit F của E. coli.
Con số bên trong cho biết kích thước của plasmit (chính xác là 99.159 bp).
Vùng màu xanh sẫm ở dưới chứa những gen chịu trách nhiệm chính để sao chép và tách plasmit F trong tế bào phát triển bình thường. Vàng xanh sáng, vùng tra, chứa những gen chịu trách nhiệm chuyển gen. Chuỗi ori T là điểm khởi đầu của quá trình chuyển plasmit trong tiếp hợp. Mũi tên cho biết hướng chuyển (vùng tra sẽ được chuyển qua cuối cùng). Những vùng màu vàng là những yếu tố có thể hoán đổi vị trí, nơi mà có thể xảy ra sự hợp nhất vào những yếu tố tương đồng trên nhiễm sắc thể vi khuẩn và dẫn đến sự hình thành dòng Hfr.
Plasmit điều khiển sự truyển gen của chính nó bởi sự tiếp xúc của hai tế bào gọi là có khả năng tiếp hợp. Không phải tất cả plasmit đều có khả năng tiếp hợp. Khả năng truyền trong tiếp hợp được kiểm soát bởi những gen trong plasmit gọi là vùng tra.
Vùng tra chứa những gen mã hóa prôtêin thực hiện chức năng bắt cặp. Sự hiện diện của vùng tra ở plasmit có thể dẫn đến kết quả quan trọng khác nếu plasmit chèn vào nhiễm sắc thể. Trong trường hợp này, plasmit có thể huy động sự chuyển ADN nhiễm sắc thể từ tế bào này sang tế bào khác.
4.5.4. Một số loại plasmit khác và ý nghĩa sinh học của chúng
- Plasmit đề kháng
Trong số những nhóm plasmit phổ biến nhất và được nghiên cứu kĩ nhất là các plasmit R, có khả năng kháng kháng sinh và nhiều chất ức chế sinh trưởng khác.
Hình 17. Plasmit R100: RTF là vùng các gen tra chuyển gen ; rdeterminant là vùng các gen đề kháng (mer - thủy ngân, sul - sulfamid, strstreptomycin,cmlchloramphenicol, tet – tetracyclin). (Theo Brock Microbiology 11, 2006)
- Plasmit mã hóa chất độc và những độc tính khác
Ở một số vi khuẩn gây bệnh, một số tính chất này được mang trên plasmit.
Dòng E. coli gây bệnh đường ruột có tính chất này bởi khả năng sinh sản thành tập đoàn trong ruột non và sản xuất một chất độc gây các triệu chứng của bệnh tiêu chảy. Sự hình thành tập đoàn đòi hỏi sự hiện diện của một prôtêin bề mặt tế bào gọi là yếu tố kháng nguyên tạo quần thể (yếu tố kháng nguyên hình thành tập đoàn), được mã hóa bởi một plasmit. prôtêin này giúp tế bào có khả năng gắn với tế bào biểu mô của ruột. Ít nhất hai chất độc ở E. coli gây bệnh đường ruột được mã hóa bởi một plasmit: hemolysin làm tan tế bào hồng cầu và enterotoxin gây tiết quá nhiều nước và muối vào ruột và do vậy là chất độc gây tiêu chảy.
- Bacteriocin.
Nhiều vi khuẩn sản xuất prôtêin ức chế hoặc giết chết tế bào các loài họ hàng gần, thậm chí ngay cả những dòng khác nhau của cùng một loài. Những tác nhân này, được gọi là bacteriocin để phân biệt với chất kháng sinh, có phổ tác động hẹp hơn kháng sinh.

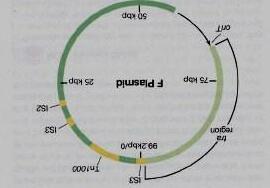
4.6. Sự chuyển vị
Các phần tử chuyển vị hay phần tử di động là những trình tự ADN có khả năng di chuyển từ vị trí này đến vị trí khác (mục tiêu) của bộ gen (cùng bộ gen hay khác).
Sự di chuyển ADN này được gọi là transposition (sự chuyển vị).
Tái tổ hợp trong chuyển vị là sự kết nối giữa các đầu mút của phần tử chuyển vị với các đầu mút bị cắt hở của ADN mục tiêu. Khác với tái tổ hợp tương đồng,
transposition không đòi hỏi sự tương đồng giữa các phần tử di động và ADN mục tiêu, và nó độc lập với chức năng được mã hóa bởi recA, vì nó sử dụng enzim đặc hiệu là Transposase.
Bảng. Các phần tử chuyển vị có ở Prokaryotae và Eukaryota Prokaryota Eukaryota
- Trình tự xen đoạn: IS
-Transposon: Tn như Tn3, Tn5, Tn10,...
- Virut: Mu
- Retrotransposon: Nấm men (Yeast): Ty
Ruồi giấm: copia, P
- Transposon ( ADN)


Ngô: hệ thống Ac-Ds
- SINE: ở người ( họ Alu)
- Retrovirut: Rous sarcoma, HIV
Ở hai đầu của IS luôn có hai trình tự lặp đoạn trực tiếp (direct repeat) ngắn. Các trình tự này dao động tùy transposon, nhưng cố định đối với mỗi loại IS. Chiều dài của phần lớn lặp đoạn trực tiếp là 9, chúng xác định các đầu mút của transposon.
M"ctiêu
Hình 19. Sơ đồ cấu trúc của transposon có các IS, các lặp đoạn đảo ngược IR
(inverted repeat) và tạo lặp đoạn trực tiếp ở hai đầu ADN điểm mục tiêu
4.6.2. Transposon sao chép và không sao chép
4.6.1. Các trình tự xen đoạn IS.
Các transposon đơn giản nhất là các trình tự xen đoạn (insertion sequence) và
được kí hiệu bởi tiếp đầu ngữ IS kèm số thứ tự như IS4.
Các phần tử IS là các cấu phần bình thường của ADN vi khuẩn và plasmit.
Dòng E. coli chuẩn thường chứa vài bản sao (<10) của bất kì một trong các IS chung thường gặp. Để mô tả sự xen đoạn vào điểm đặc biệt, kí hiệu 2 chấm kép được sử dụng như λ:: IS1 chỉ IS1 xen vào phagơ λ.
Các IS là những đơn vị tự trị, mỗi một trong chúng mã hóa cho chỉ một prôtêin cần thiết cho sự chuyển vị bản thân chúng. Trình tự của mỗi loại IS có khác nhau, nhưng trong tổ chức cấu tạo có nhiều tính chất chung. Ở giữa có gen Transposase tnp, hai đầu có lặp đoạn đảo ngược (IR – Inverted repeats).
Các đầu mút của transposon có trình tự lặp lại đảo ngược (inverted repeat - IR).
Trong ví dụ này, tiêu điểm có 5 bp. Các đầu mút của transposon gồm các lặp đoạn đảo ngược 9 bp được đánh số từ 1 đến 9.
Sự chuyển vị ADN theo 2 kiểu khác nhau (sao chép, không sao chép hay bảo tồn (conservative)) và có những tính chất chung (hình 20).
Hình 20. Sơ đồ về transposition sao chép và không sao chép ADN cho
Sơ đồ diễn biến transposition sao chép (transposon Tn3) goomfcacs giai đoạn cắt và nối, sự cộng gắn và phân tách (hình 21).
Hình 21. Sơ đồ transposition sao chép (Tn3)
(A. Sự nối giữa Tn và trình tự mục tiêu (ligation) ADN nhận.
B. Sự cộng gắn (Cointegrat Formation) 2 loại ADN.


C. Sự phân tách (Resolution) ra 2 loại ADN.)
Quá trình transposition có những đặc điểm sau đây:
- Cả hai đầu của phân tử mang gần như một trình tự có định hướng đảo ngược nhau.
- Các transposon mã hóa ít nhất một prôtêin là Transposase. Transposase gắn đặc hiệu và cắt các trình tự ở cuối để thực hiện chuyển vị
- Các transposon tạo bản sao ngắn (≤ 12bp) của ADN ở điểm mục tiêu trong transposition. Chiều dài của trình tự này đặc trưng và không đổi đối với một phần tử nhất định và được tạo ra nhờ sự cắt so le (staggered cleavage) của ADN mục tiêu nhờ Transposase.
4.6.3. Sự chuyển vị ngược (Retrotransposition)
Nhóm các transposon, được phát hiện ở nấm men (Ty) và Drosophila (copia) và có liên quan đến retrovirut, di chuyển đến vị trí mới qua trung gian ARN. Transposon của nấm men Ty, giống với retrovirut, có lặp đoạn cuối dài (long terminal repeat) được gọi là trình tự δ nằm ở 2 phía đoạn mã hóa. Các prôtêin được mã hóa, có trình tự tương đồng với integraz của retrovirut và reverse transcriptaz, rất cần thiết cho transposition.
Phương thức phiên mã ngược của các phần tử di động tương tự như của retrovirut. Có sự tương tự về cấu trúc giữa retrovirut và các retrotransposon ở nấm men (Ty) và Drosophila (copia).
Sự chuyển vị ngược tạo bản sao của phần tử ở vị trí mới, trong khi phân tử cho ban đầu vẫn giữ nguyên cấu trúc không biến đổi. Do vậy, retrotransposition tạo nên một ít đứt đoạn và các tái cấu trúc (rearrangement) của bộ gen tế bào chủ. Những biến đổi của bộ gen liên quan với retrotransposition sẽ dẫn đến việc làm ngừng hay hoạt hóa các gen, mà một số có thể gây ung thư
Các transposon đã góp phần vào sự tiến hóa của plasmit, và vài chứng cứ cho thấy các plasmit R có được tính đề kháng kháng sinh thông qua các transposon.
4.6.4. Gây đột biến bằng transposon
Các ứng dụng của transposon bao gồm:
- Gây đột biến (mutagensis) bằng transposition.
- Công cụ cho kỹ thuật di truyền in vivo (in vivo Gentic Engineering).
- Nhân tố trung gian đề kháng kháng sinh (mediator of antibiotic resistance)
Sự chèn transposon vào giữa gen sẽ dẫn đến đột biến làm mất chức năng bình thường của gen . Transposon cung cấp một phương tiện dễ dàng để tạo các đột biến trên nhiễm sắc thể. Một thuận lợi cho gây đột biến bằng transposon là nó có chứa 1 gen kháng kháng sinh, mà gen này dùng làm marker (dấu chuẩn) để nhận biết transposon được chèn vào. Trước tiên các dòng mang gen kháng kháng sinh được phân lập từ môi trường giàu dinh dưỡng, nơi các đột biến khuyết dưỡng tăng trưởng tạo khuẩn lạc. Sau đó chúng có thể được sàng lọc trên môi trường giới hạn được cung cấp các chất tăng trưởng nào đó để xác định chất nào cần thiết cho đột biến.
Các transposon cũng hữu ích cho sự chèn 1 maker gen khuyết dưỡng (auxotrophic) vào trong dòng sinh vật hoang dại. Thường thì các đột biến khuyết dưỡng khó phân lập trực tiếp. Nhưng nếu marker khuyết dưỡng được đưa vào cùng transposon với marker kháng kháng sinh, thì có thể qua chọn dòng đề kháng kháng sinh mà xác định marker khuyết dưỡng. Hai transposon thường được sử dụng trong gây đột biến là Tn5 (kháng neomycin và kanamycin) và Tn10 (chứa 1 marker kháng tetracycline) (Hình 22).
Hình 22. Transposon Tn10 chèn vào A làm mất chức năng, nhưng kháng tetra.
Integron là các transposon có thể thu nhận và biểu hiện các gen từ nhiều nguồn khác nhau. Tuy nhiên, khác với các transposon được chèn vào một cách ngẫu nhiên, các integron chèn vào có tính chọn lọc cao (high selective) trong các điểm xen đoạn (insertion site) của chúng, thường thì đựơc chèn vào trong plasmit.
Integron chứa gen mã hóa cho prôtêin được gọi là integraz, cần thiết cho sự tái tổ hợp ở điểm chuyên biệt (site-specific recombination). Bộ gen của phagơ λ chèn vào nhiễm sắc thể của E. coli ở một vị trí đặc biệt nhờ họat tính của enzim integraz λ.
Integron cũng chứa trình tự ADN đặc biệt cho phép integraz chèn các nhóm gen gọi là cassette vào nhiễm sắc thể, mà trên đó có promoter cho phép các gen cassette mới được biểu hiện.
5. Các kĩ thuật sinh học phân tử hỗ trợ nghiên cứu di truyền.
5.1. Lai phân tử
Hai mạch ADN có nguồn gốc khác nhau nhưng có trình tự nucleotit bổ trợ có thể liên kết với nhau trong điều kiện phù hợp (về nhiệt độ, độ pH, nồng độ muối trong đệm) để tạo nên một phân tử ADN mới. Hiện tượng liên kết như vậy cũng có thể xảy ra giữa hai mạch ADN với nhau, hoặc giữa hai mạch ARN hoặc giữa ADN và ARN. Phân tử axit nucleic sợi kép mới hình thành được gọi là phân tử lai và quá trình kết cặp giữa các bazơ thuộc hai mạch đơn axit nucleic có nguồn gốc khác nhau theo nguyên tắc bổ sung như vậy được gọi là quá trình lai phân tử.
5.1.1. Phương pháp Southern blot
Phương pháp Southern blot cho phép nghiên cứu ADN của bộ gen, kiểm tra kết quả chuyển gen hoặc kiểm tra sự có mặt của một gen nào đó trong bộ gen của tế bào.
* Quy trình:
- Tách ADN bộ gen của tế bào.
- Sử dụng enzim cắt giới hạn để cắt phân tử ADN thành những đoạn nhỏ, điện di trên gel agarose để tách những đoạn có kích thước khác nhau.
- Gây biến tính ADN trên gel bằng dung dịch NaOH-0,5M, sau đó, chuyển ADN từ gel sang màng lai. Trong quá trình chuyển, vị trí của ADN phải được giữ nguyên.
- ADN cố định trên màng được lai với mẫu dò có đánh dấu phóng xạ, ở nhiệt độ 65°C và thời gian lai khoảng 3- 8 giờ. Sau quá trình lai người ta rửa màng lai để loại bỏ những mẫu dò không bắt cặp chuyên biệt với ADN cố định trên màng lai.
- Dùng kĩ thuật phóng xạ tự ghi để định vị các phân tử lai. Trong kĩ thuật này, người ta đặt một phim nhạy cảm với tia xạ áp sát vào màng lai. Các phân tử lai có đánh dấu phóng xạ sẽ tác động lên phim và kết quả được thể hiện qua các chấm đen trên phim.
* Các ứng dụng quan trọng của Sourthern blot:
- Lập bản đồ giới hạn của một gen.
- Phát hiện các đa dạng trình tự của cùng một gen ở các chủng hay các cá thể khác nhau qua sự so sánh bản đồ giới hạn của chúng.
- Phát hiện các đột biến mất đoạn, đột biến điểm hay tái tổ hợp trên một gen vì chúng làm thay đổi bản đồ giới hạn.
5.1.2. Phương pháp Northern blot
Phương pháp lai Northern blot là phương pháp lai ARN của tế bào với mẫu dò ADN. Về nguyên tắc giống như lai Southern blot, được tiến hành như sau:
ARN (đã được làm biến tính) sẽ được phân tích theo kích thước nhờ điện di trên gel agarose có chứa chất làm biến tính. Các chất làm biến tính có tác dụng ngăn cản sự hình thành cấu trúc bậc hai của ARN sau khi đã biến tính. Và do đó, không cản trở sự di chuyển cũng như sự tách của các ARN trên gel. Sau đó, ARN được chuyển lên màng lai. Những ARN cố định trên màng lai được đem lai với mẫu dò ADN có đánh dấu phóng xạ để tạo phân tử lai ARN-ADN. Các phân tử lai được phát hiện nhờ kĩ thuật phóng xạ tự ghi.
* Ứng dụng:
- Phát hiện sự có mặt mARN trong các mẫu khác nhau.
- Phân tích sự biểu hiện của gen giữa hai mẫu.
- Phát hiện virut ARN.
5.1.3. Lai tại chỗ
Lai tại chỗ là phương pháp lai phân tử, trong đó, trình tự axit nucleic cần tìm nằm ngay trong tế bào hay trong mô.
Lai tại chỗ cho phép nghiên cứu axit nucleic mà không cần qua giai đoạn tách chiết chúng ra khỏi mô, tế bào. Các ứng dụng của kiểu lai này rất đa dạng đi từ kĩ thuật định vị gen trên nhiễm sắc thể, phát hiện dòng vi khuẩn tái tổ hợp trong phương pháp tạo dòng đến việc nghiên cứu một mARN chuyên biệt trong tế bào và mô. Phương pháp này ít tốn kém hơn là lai Southern blot, mẫu dò được đánh dấu bằng huỳnh quang thay cho mẫu dò đánh dấu phóng xạ. Tùy từng loại tế bào và mô có thể thực hiện các phương pháp lai tại chỗ khác nhau.
* Ứng dụng:
- Lai trên tế bào và mô: thường được sử dụng để phát hiện các mARN. Các loại mARN hoạt động ở các giai đoạn khác nhau trong quá trình phát triển tế bào và cơ thể Lai trên tế bào và mô nhằm nghiên cứu giai đoạn hoạt động của mARN, từ đó, tìm hiểu hoạt động của một gen, mối liên quan giữa phiên mã và giải mã, định vị các gen cần nghiên cứu trên nhiễm sắc thể
- Lai trên NST: Phương pháp này cung cấp thông tin chính xác về vị trí và sự phân bố của một trình tự ADN cần tìm trên nhiễm sắc thể nhờ một mẫu dò chuyên biệt.
- Lai trên khuẩn lạc: Phương pháp này được sử dụng để phát hiện dòng vi khuẩn có mang vectơ tái tổ hợp cần tìm trong một ngân hàng gen. Ngân hàng gen được trải trên mặt thạch của hộp petri. Sau khi các khuẩn lạc đạt kích thước nhất định, người ta thu lấy dấu ấn của chúng bằng cách áp một màng lai trên mặt thạch. Mỗi khuẩn lạc sẽ để lại vài tế bào vi khuẩn trên màng lai. Sau đó xử lí màng lai bằng NaOH để làm vỡ tế bào vi khuẩn và làm biến tính ADN. Kết quả phóng xạ tự ghi của dấu ấn cho phép xác định dòng vi khuẩn cần tìm trên hộp pectri ban đầu. Dòng này sẽ được thu nhận và phân tích.
5.2. PCR
5.2.1. Khái niệm
PCR là chữ viết tắt của cụm từ Polymerase Chain Reaction, là Phản ứng chuỗi trùng hợp hoặc "phản ứng khuếch đại gen". PCR là một kỹ thuật phổ biến trong sinh học phân tử nhằm khuyếch đại (tạo ra nhiều bản sao) một đoạn ADN mà không cần sử dụng các sinh vật sống như E. coli hay nấm men.
Trong thực tiễn, PCR được dùng để khuếch đại một đoạn ADN ngắn, đã xác định được một phần; hoặc khuếch đại 1 đoạn ADN ở 1 vùng bất kì trong gennom khi trình tự nucleotit ở 2 đầu đoạn ADN đó đã biết. Đó có thể là một gen đơn, hay một phần của gen. Trái với sinh vật sống, quy trình PCR có thể copy một mảnh ADN ngắn, có thể lên đến 10kb. Một vài phương pháp có thể copy một mảnh kích thuớc lên đến 40kb ít hơn nhiều so với nhiễm sắc thể ADN trong tế bào eukaryote (ví dụ: như tế bào người chứa hơn 3 tỉ cặp base).

PCR cần rất nhiều thành phần, gồm:
- ADN mẫu chứa mảnh ADN cần khuếch đại.
- Cặp mồi (primer), để xác định điểm bắt đầu và kết thúc vùng cần khuếch đại.
- ADN-polymerase xúc tác cho việc nhân lên của ADN.
- Nucleotits (ví dụ dNTP) là nguyên liệu cho ADN-polymerase để xây dựng ADN mới.
- Dung dịch đệm, cung cấp môi trường hóa học cho ADN-polymerase.
Phản ứng PCR được thực hiện trong chu kỳ nhiệt. Đây là máy đun nóng và làm nguội trong ống phản ứng ở nhiệt độ chính xác cho mỗi phản ứng.
5.2.2. Quy trình PCR
Quy trình PCR gồm 20 đến 30 chu kỳ. Mỗi chu kỳ gồm 2-3 giai đoạn biến đổi nhiệt độ (thường là 3). Nhiệt độ và thời gian thực hiện của mỗi chu kỳ phụ thuộc vào
nhiều yếu tố. Những yếu tố này gồm enzim tổng hợp ADN, nồng độ ion hóa trị hai, dNTPs dùng trong phản ứng, và nhiệt độ nóng chảy (Tm) của mồi.
- Giai đoạn 1- Biến tính: Nhiệt độ tăng lên 92-96°C để tách hai sợi ADN ra. Trước chu kỳ 1, ADN thường được biến tính đến thời gian mở chuỗi để đảm bảo mẫu ADN và mồi được phân tách hoàn toàn và chỉ còn dạng sợi đơn.
- Giai đoạn 2- Gắn mồi: Sau khi 2 sợi ADN tách ra, nhiệt độ được hạ thấp xuống để mồi có thể gắn vào sợi ADN đơn. Nhiệt độ giai đoạn này phụ thuộc vào đoạn mồi và thường thấp hơn nhiệt độ biến tính 50°C (45°C - 55°C). Sử dụng sai nhiệt độ trong giai đoạn này dẫn đến việc đoạn mồi không gắn hoàn toàn vào ADN mẫu, hay gắn một cách tùy tiện.
- Giai đoạn 3- Kéo dài: Hỗn hợp ADN và mồi được ủ với enzim Taq polymerase và 4 loại dNTP. Enzim bắt đầu bám vào và hoạt động dọc theo sợi ADN. Nhiệt độ kéo dài phụ thuộc ADN-polymerase. Thời gian của bước này phụ thuộc vào cả ADN-polymerase và chiều dài mạch ADN cần khuếch đại.
Hình 23. Chu kì nhiệt trong kĩ thuật PCR.
Sản phẩm PCR có thể được nhận biết bằng kích thước của chúng qua việc sử dụng điện di trên gel agarose. Kết quả là các đoạn ADN nhỏ hơn sẽ di chuyển nhanh hơn các đoạn lớn qua gel từ cực âm đến cực dương. Kích thước của sản phẩm PCR có thể được xác định bằng việc so sánh với thang ADN, thang này có chứa các ADN đã biết trước kích thước, cũng nằm trong gel.
5.2.3. Tối ưu hoá PCR
Do PCR rất nhạy, nên phải tránh nhiễm các ADN khác có trong phòng thí nghiệm (vi khuẩn, virut, ADN,...). Vì vậy các mẫu ADN, hỗn hợp phản ứng và quy trình ADN, phản ứng phân tích sản phẩm, nên được thực hiện trong một khu vực riêng biệt.
Ở giai đoạn chuẩn bị hỗn hợp phản ứng, phải chuẩn bị phòng riêng biệt với đèn UV. Phải sử dụng găng tay sạch cho mỗi bước PCR cũng như thay thế các pipet.
Thuốc thử dùng trong PCR phải được chuẩn bị riêng biệt và chỉ được dùng cho mục đích này. Các mẫu phải được giữ riêng biệt với các mẫu ADN khác. Phản ứng có kiểm soát (kiểm soát bên trong), ngoại trừ ADN khuôn mẫu, phải luôn được thực hiện, để kiểm tra sự nhiễm bẩn.
5.2.4. Những ứng dụng của PCR
– Xác định các đoạn trình tự cần nghiên cứu.
– Phát hiện đột biến.
– Nghiên cứu quá trình tiến hoá phân tử
– Phục hồi các gen đã tồn tại hàng triệu năm.
– Chọn giống vật nuôi, cây trồng.
– Lựa chọn các cặp cha mẹ thuần chủng trong thời gian ngắn.
– Xác định các loài mới, các loài đặc hữu bằng phưng pháp di truyền phân tử.
– Y học – Khoa học hình sự
– Chuẩn đoán chính xác các bệnh nhiễm trùng từ vi khuẩn, nấm, virut.
– Chuẩn đoán sớm các bệnh gây ra do ung thư, các bệnh di truyền.
– Xác định huyết thống, truy tìm dấu vết tội phạm.
– Là kỹ thuật nền cho các kỹ thuật khác như: RAPD, SSR…
6. Sản xuất vacxin tái tổ hợp dựa trên di truyền vi sinh vật
6.1. Phân loại vacxin
Căn cứ vào nguồn kháng nguyên nhân lên được hay không nhân lên trong cơ thể động vật, người ta chia làm 2 nhóm:
* Nhóm 1: Vacxin có kháng nguyên sống được nhân lên, gồm:
- Vacxin tái tổ hợp có vectơ dẫn truyền
- Vacxin axit nucleic (vacxin ADN)
- Vacxin xóa gen độc
* Nhóm 2: Vacxin có kháng nguyên không nhân lên, gồm: Vacxin chứa kháng nguyên là prôtêin sản xuất bằng kĩ thuật gen
6.2. Vai trò của công nghệ ADN tái tổ hợp trong nhận dạng và sản xuất vacxin
6.2.1. Nhận dạng và tạo dòng các kháng nguyên có tiềm năng vacxin
Nhiều tác nhân gây bệnh gần như không có khả năng nuôi cấy bên ngoài vật chủ tự nhiên của chúng và điều này đã gây cản trở cho các cách tiếp cận truyền thống để phát triển liệu pháp vacxin. Ví dụ: virut viêm gan B (HBV), tác nhân gây bệnh giang mai ở người (Treponema pallidum) và vi khuẩn gây bệnh phong (Mycobacterium leprae) không thể sinh trưởng trong điều kiện in vitro. Bởi vậy, người ta không thể tạo ra các vacxin sống nhược độc hoặc các vacxin bất hoạt bằng cách nuôi cấy các tác nhân này.
Công nghệ ADN tái tổ hợp cho phép chuyển thông tin di truyền từ những sinh vật này vào những vật chủ “dễ nuôi” hơn như là E. coli, nấm men hoặc tế bào động vật có vú.
Không phải tất cả các gen kháng nguyên đều dễ nhận dạng, tạo dòng và biểu hiện như gen kháng nguyên bề mặt của viêm gan. Trình tự nguyên vẹn của genome HBV có kích thước nhỏ hơn 10 kb. Bởi vậy, tương đối đơn giản khi thiết lập khung đọc mở để biểu hiện. Tuy nhiên, ở trường hợp bệnh sốt rét thì lại khác. Genome của ký sinh trùng sốt rét lớn hơn genome của HBV hàng ngàn lần, vì thế người ta đã gặp rất nhiều khó khăn trong việc thu thập các thông tin về trình tự gen thích hợp và sản phẩm của nó để tạo ra bảo vệ miễn dịch, nên người ta cần phải ứng dụng công nghệ ADN tái tổ hợp để sản xuất vacxin.
Điểm khởi đầu của công nghệ ADN tái tổ hợp là xây dựng thư viện ADN của cơ thể được nghiên cứu trong E. coli. Thư viện cADN hoặc genomic ADN có thể cung cấp các phương thức cơ bản để nhận dạng và phân lập các gen quan tâm.
6.2.2. Quy trình sản xuất
* Quy trình sản xuất vacxin bằng công nghệ ADN tái tổ hợp:
- Lựa chọn kháng nguyên đích.
- Phân tích và xác định gen đích.
- Nhân dòng gen.
- Biểu hiện gen trong tế bào chủ.
- Nuôi cấy và thu nhận prôtêin.
- Kiểm tra chất lượng và tinh sạch prôtêin.
- Sản xuất chế phẩm.
- Kiểm tra sản phẩm.
6.2.3. Ưu diểm của vacxin tái tổ hợp và một số thành tựu vacxin
* Ưu điểm:
- Năng suất cao.
- An toàn vì không sử dụng trực tiếp virut gây bệnh.
- Giá thành hạ vì không phải nuôi virut.
* Các vacxin sản xuất bằng công nghệ ADN tái tổ hợp:
- Vacxin chứa kháng nguyên sản xuất bằng ADN tái tổ hợp.
- Vacxin vectơ.
- Vacxin ADN.
- Vacxin mARN.
- Vacxin dựa trên cơ thể chuyển gen (VD: vacxin thực phẩm).
* Vacxin ADN:
Một hướng trong sản xuất vacxin ứng dụng công nghệ ADN tái tổ hợp là các vacxin ADN (miễn dịch di truyền).
Các plasmit mang các gen đặc hiệu cho một hoặc nhiều prôtêin kháng nguyên có thể được dùng để tạo miễn dịch (chủng ngừa). Các plasmit đó được tiêm (thường vào cơ) đưa gen trực tiếp vào trong một số tế bào và được hấp thu bởi các tế bào lân cận nơi kim tiêm được đưa vào.
Ngoài ra, plasmit cũng có thể được đưa vào bằng súng bắn gen (các viên đạn vàng được bọc ADN của plasmit) bằng cách đẩy các plasmit vào trong các tế bào gần bề mặt cơ thể, thường là da hoặc màng nhầy.
Khi vào bên trong tế bào, một vài plasmit tái tổ hợp sẽ đi vào nhân và do gen được điều hòa bởi một promoter mạnh của eukaryote, các tế bào sẽ tổng hợp các kháng nguyên được mã hóa bởi plasmit.
* Vacxin mARN:
Việc tạo ra một sinh khối đủ lớn để bắt đầu nghiên cứu sản xuất một loại vacxin thông thường phải mất từ 4 tới 6 tuần. Trong khi đó với công nghệ mARN, một loại vacxin có thể được tổng hợp và tạo ra chỉ sau vài phút.
Sự khác lớn về tốc độ này là do vacxin chế tạo theo cách thức thông thường phải dựa vào kỹ thuật nuôi cấy tế bào động vật, trong khi hoạt động sản xuất ARN không cần tới tế bào. Đây là một tiến trình sinh hóa, có thể được thực hiện trên các enzim tổng hợp.
Thay vì dùng mầm bệnh giảm độc tính hoặc bị xử lý bất hoạt để "huấn luyện" cơ thể ngăn chặn sự lây nhiễm, như cách thức chế tạo vacxin truyền thống, phương thức mới sử dụng kỹ thuật cao, can thiệp vào cấp độ tế bào và di truyền.
Trước tiên, người ta phải tạo ra các phân tử ADN mạch đôi dạng vòng (plasmit) chuẩn, để dựa vào đó mà chế tạo được mẫu ARN thông tin đã được thay đổi mã di truyền thay vì mầm bệnh. Khi vào cơ thể, các mARN này sẽ "hướng dẫn" nhiều tế bào bên trong chúng ta tạo ra các bản sao của một loại prôtêin (không gây hại) nằm trên bề mặt virut mà cơ thể nh
Khi thấy cơ thể bị tràn ngập nhiều prôtêin lạ, hệ miễn nhiễm sẽ khởi động cơ chế nhận biết, sau đó tấn công tiêu diệt các prôtêin này. Quá trình đó sẽ tạo ra kháng thể, giúp bảo vệ chúng ta khỏi khả năng nhiễm bệnh khỏi virut.
Công nghệ mARN là công cụ tuyệt vời để sản xuất vacxin trong đại dịch COVID-19, với hai sản phẩm nổi tiếng là vacxin Pfizer-BioNTech và ModeARN có hiệu quả ngăn bệnh cao.
Tuy nhiên công nghệ này cũng gặp những khó khăn nhất định, Do mARN – rất mỏng manh và sẽ bị cắt nhỏ bởi các enzim tự nhiên nếu nó được tiêm trực tiếp vào cơ thể người. Để bảo vệ vắc-xin, mARN sẽ được bao bọc trong các bong bóng dầu làm từ các hạt mỡ nano siêu nhỏ.
Do đặc tính mỏng manh, các phân tử mARN sẽ nhanh chóng bị phân rã ở nhiệt độ phòng. Vì vậy vắc-xin mARN cần được bảo quản lạnh và giữ ổn định trong tối đa sáu tháng khi vận chuyển và bảo quản ở nhiệt độ -4° F (-20°C). Điều đó làm cho giá thành của loại vaccin công nghệ mới này cao hơn so với các loại vaccin khác.
II. HỆ THỐNG CÂU HỎI VÀ BÀI TẬP
1. Câu hỏi và bài tập có hướng dẫn.
1.1. Câu hỏi và bài tập hóa sinh tế bào – Sinh học phân tử.
Câu 1.
EF-Tu là một yếu tố kéo dài với GTP tham gia giai đoạn kéo dài chuỗi pôlipeptit ở tế bào nhân sơ. EF-Tu gắn với tất cả các phức hợp aminoaxyl-tARN(aatARN) với ái lực gần như nhau để đưa chúng đến ribôxôm với tần xuất giống nhau. Sau đây là kết quả thí nghiệm xác định sự liên kết của EF-Tu và phức hợp aminoaxyltARN bắt cặp chính xác và không chính xác.
Phức hợp aminoaxyl-tARN Hệ số phân ly(nM)
Ala-tAARNla 6,2
Gln-tAARNla 0,05
Gln –tARNGln 4,4
Ala-tARNGln 260
a. Dựa vào số liệu trên hãy giải thích vì sao hệ thống nhận biết tARN- EF-Tu có thể ngăn ngừa sự ghép sai axit amin trong quá trình dịch mã?
b. Hãy chỉ ra vai trò của EF-Tu trong quá trình dịch mã.
Hướng dẫn trả lời
a. Phức hợp aminoaxyl-tARN bắt cặp chính xác (Ala-tAARNla và Gln-tARNGln) có ái lực gần như nhau với EF-Tu và được chuyển đến vị trí A trên ribôxôm.
- Phức hợp bắt cặp không chính xác Ala-tARNGln gắn với EF-Tu lỏng lẻo hơn nhiều và sẽ phân ly với EF-Tu trước khi tiến đến ribôxôm.
- Phức hợp Gln-tAARNla gắn chặt với EF-Tu làm cho EF-Tu không tách được khỏi chúng tại ribôxôm.
- Do đó, dù ái lực gắn kết cao hay thấp hơn đều ảnh hưởng đến hoạt động của EF-Tu và làm giảm tốc độ gắn vào vị trí A trên ribôxôm của phức hợp aminoaxyl-tARN bắt cặp sai → ngăn ngừa sự ghép cặp sai trong dịch mã..
b. Vai trò của EF-Tu giúp sự bắt cặp chính xác bộ ba đối mã của tARN với bộ ba mã hóa của mARN.
- Sự thủy phân GTP gắn với EF-Tu khi có sự cặp đôi chính xác tạo cấu hình phù hợp cho sự tương tác giữa côđon- anticôđon và đảm bảo cho sự hình thành liên kết peptit xảy ra tiếp theo.
Câu 2.
Enzim A tham gia vào giải độc ammoniac. Ở gà, enzim này được vận chuyển đến ti thể, còn ở người, enzim này định vị ở tế bào chất. Để được vận chuyển tới ti thể, enzim A phải mang đoạn peptit tín hiệu tồn tại ở dạng xoắn alpha lưỡng cực. Trong đó các axit amin tích điện dương nằm ở 1 phía, còn các axit amin kị nước và các axit amin khác nằm ở phía đối diện.
a. Kết quả xây dựng mô hình vòng quay xoắn của 2 trình tự đầu amin (hình dưới đây) cho thấy trình tự 2 tạo ra mô hình vòng quay xoắn lưỡng cực với một phía tích điện dương rõ nét. Trong khi đó trình tự 1 tạo ra vòng quay xoắn với tính lưỡng cực kém hơn, mang quá ít điện tích dương. Do vậy, trình tự 1 là của người và trình tự 2 là của gà .
Dựa vào mô hình vòng quay xoắn (Hình 1) và 2 trình tự đầu amin (-NH2): trình tự 1 và trình tự 2 (Hình 2) của enzim A ở gà và người, hãy cho biết:
a. Trong 2 trình tự này, trình tự nào là của gà và trình tự nào là của người? Giải thích
b. Nếu có 1 lượng lớn enzim A thì có ảnh hưởng đến sự vận chuyển prôtêin khác vào trong ti thể không ? Giải thích.
c. Khi vận chuyển enzim A vào ti thể, có cần prôtêin vận chuyển qua màng ngoài ti thể không? Tại sao?
Hướng dẫn trả lời
Mô hình vòng quay xoắn của 2 trình tự đầu amin
b. Có. Vì enzim A mang peptit tín hiệu sẽ cạnh tranh với các prôtêin khác của ti thể, đồng thời do có số lượng lớn nên sẽ làm giảm hoặc làm ngừng quá trình vận chuyển các prôtêin khác vào trong ti thể
c. Có. Vì enzim A được mã hóa ở nhân và mang peptit tín hiệu nên cần có prôtêin vận chuyển nhận biết được đoạn peptit tín hiệu, chuyển trình tự tín hiệu vào khe gian màng rồi chuyển enzim đi qua màng ngoài ti thể.
Câu 3.
Một nhà khoa học đã tinh sạch được 5 chuỗi peptit khác nhau (từ 1 đến 5), mỗi chuỗi gồm 5 axit amin (được gọi là A, B, C, D, E). Anh ta xác định khối lượng và trình tự axit amin của từng chuỗi peptit. Số liệu thu được được nêu trong bảng dưới đây:

Peptit Trình tự axit amin Khối lượng (Da)
1 BCDACCDEDCB 966
2 ABBCAEEDECB 1099
3 BACDAEAEECA 1357
4 CACADBACAEB 1279

5 EDDCABBCCEE 1014
Khối lượng phân tử của mỗi axit amin được nêu ở bảng sau:
Chú ý: Khối lượng của một phân tử nước là 18 Da.
Hãy xác định C, A, E, B là những axit amin nào trong các axit amin sau: serine, tyrosine, cysteine. glycine?
Hướng dẫn trả lời
Ta sẽ cần phải giải một hệ các phương trình như sau để có thể xác định được các amino acid:
A + 2B + 4C + 3D + E=1146 (1)
2A + 3B + 2C + D+ 3E=1279 (2)
4A + B + 2C +D + 3E =1537 (3)
4A + 2B + 3C + D + E = 1459 (4)
A + 2B + 3C + 2D + 3E = 1194 (5)
Lấy (3) – (2) ↔ 2A - 2B = 258 ↔ A–B =129
A chắc chắn sẽ là tryptophan và B phải là glycine
Sau đó lầy (5) – (4) ↔ D + 2E = 347
2*(2) – (1) ↔ 5E – D = 500
Vậy D là serine, E là cysteine và C là alanine.
(Một điểm cần phải lưu ý ở đây đó là: để hình thành một chuỗi poplypeptit cần n amino acid thì có (n-1) phân tử nước được giải phóng là sản phẩm của phản ứng trùng ngưng, do vậy, cần phải cộng thêm khối lượng này thì ta mới có thể tính
toán.Phần còn lại của câu hỏi là thuần túy toán học, nhưng ta sẽ cần phải tinh ý để nhận ra những phương trình thuận lợi cho tính toán (như (2) và (3)) để có thể trả lời gọn gàng và chính xác nhất.)
Câu 4.
Kết quả nghiên cứu quá trình tái bắt cặp (hồi tính) các đoạn ADN ở người và E. coli thu được trong Hình 1. Trong quá trình này, người ta tăng nhiệt độ để ADN mạch kép tách thành các sợi đơn và hạ nhiệt độ để các sợi tương đồng bắt cặp lại với nhau. Quá trình tái bắt cặp ADN ở E. coli chỉ có một giai đoạn, trong khi ở người lại có hai giai đoạn, gồm giai đoạn đầu diễn ra nhanh (a) và giai đoạn sau diễn ra chậm hơn (b), được phân chia bởi dấu “•” trên Hình 1. Hãy giải thích kết quả thí nghiệm trong Hình 1.
Hướng dẫn trả lời
- Hầu hết các trình tự ADN của E. coli đều tồn tại ở dạng đơn bản.
- Sự hồi tính ADN của E. coli là quá trình đơn giản trong đó mỗi sợi đơn ADN sẽ kết hợp lại chính xác với sợi đơn bổ sung.
- Hệ gen của người có chứa nhiều trình tự ADN lặp lại.
- Nhiều đoạn ADN lặp lại sẽ bắt cặp với nhau tạo nên các vùng sợi kép (hồi tính) nhanh hơn nhiều (hình thành pha nhanh) so với các trình tự ADN có một bản sao duy nhất trong hệ gen (hình thành pha chậm).
Câu 5.
Nuôi cấy tế bào động vật trong môi trường chứa timin được đánh dấu phóng xạ trong 30 phút. Sau đó, tế bào được chuyển sang môi trường chứa timin không đánh dấu phóng xạ để các tế bào tiếp tục phát triển. Tỉ lệ các tế bào có mang ADN đánh dấu phóng xạ bước vào giai đoạn phân chia được xác định liên tục theo thời gian nuôi (Hình 1). Thời điểm 0 giờ là khi tế bào bắt đầu được chuyển sang môi trường không đánh dấu phóng xạ.
a. Sau khi nuôi trong môi trường đánh dấu phóng xạ, có phải tất cả các tế bào sẽ mang ADN có đánh dấu phóng xạ không? Giải thích.
b. Tại sao ở thời gian từ 0 đến 2 giờ (Hình 1) không có tế bào nào chứa ADN đánh dấu phóng xạ được quan sát thấy?
c. Giải thích tại sao lại có sự tăng và giảm và lại tăng lên của đường cong Hình
Hướng dẫn trả lời
a. Không. Vì chỉ các tế bào ở pha S- là pha mà timin phóng xạ có trong môi trường và được gắn vào ADN đang được tổng hợp.
b. Vì không có ADN phóng xạ do các tế bào này không tổng hợp ADN ở giai đoạn có timin phóng xạ trong môi trường.
c. Đường cong đi lên vì có nhiều tế bào đánh dấu phóng xạ bước vào quá trình phân bào (pha S).
Đường cong đi xuống vì các tế bào có đánh dấu phóng xạ kết thúc quá trình phân bào. Các tế bào bước vào phân bào đều là các tế bào không có đánh dấu phóng xạ
Sau 20 giờ đường cong lại tăng vì các tế bào có ADN đánh dấu phóng xạ lại bắt đầu phân chia.
Câu 6.
Hoạt tính của các enzim Wee1 kinase và Cdc25 phosphatase xác định trạng thái phosphoryl hoá của tyrosine 15 trong hợp phần Cdk1 của M-Cdk. Khi tyrosine 15 bị phosphoryl hoá, M-Cdk sẽ bị bất hoạt; khi tyrosine 15 không bị phosphoryl hóa, MCdk ở trạng thái hoạt động (Hình A). Hoạt tính của các enzim Wee1 kinase và Cdc25 phosphatase cũng bị điều khiển bởi quá trình phosphoryl hoá. Sự điều hoà các hoạt tính này có thể được nghiên cứu ở các dịch chiết noãn ếch. Trong các dịch chiết này, Wee1 kinase ở trạng thái hoạt động và Cdc25 phosphatase ở trạng thái bất hoạt. Do vậy, M-Cdk bị bất hoạt vì hợp phần Cdk1 bị phosphoryl hoá ở tyrosine 15. M-Cdk trong các dịch chiết này có thể được hoạt hoá nhanh chóng bằng axit okadaic, là một chất ức chế của enzim serine/threonine phosphatases. Sử dụng các kháng thể đặc hiệu cho Cdk1, Wee1 kinase, và Cdc25 phosphatase, có thể xác định được trạng thái phosphoryl hoá của chúng bằng những thay đổi về sự di chuyển của chúng trên gel điện di (Hình B). Dạng phosphoryl hoá của các prôtêin này thường di chuyển chậm hơn dạng không bị phosphoryl hoá của prôtêin đó.
a. Dựa vào các thông tin trên, hãy cho biết các enzim Wee1 kinase và Cdc25 phosphatase ở trạng thái hoạt động khi nào? Giải thích.
b. Điều gì sẽ xảy ra nếu M-Cdk ở trạng thái hoạt động có thể phosphoryl hoá Wee1 kinase và Cdc25 phosphatase?
Hướng dẫn trả lời
a. – Theo hình A, M-Cdk hoạt động khi Wee1 kinase bất hoạt và Cdc25 phosphatase hoạt động.
- Khi cho axit okadaic vào thì M-Cdk hoạt động → trong môi trường axit okadaic Wee1 kinase bất hoạt và Cdc25 phosphatase hoạt động.
- Theo hình B, trong môi trường axit okadaic thì Wee1 kinase và Cdc25 phosphatase đều bị phosphoryl hóa.
→ Wee1 kinase bị bất hoạt khi bị phosphoryl hóa (Wee1 kinase hoạt động khi không bị phosphoryl hóa) và Cdc25 phosphatase hoạt động khi bị phosphoryl hóa.


b. Nếu M-Cdk ở trạng thái hoạt động có thể phosphoryl hoá Wee1 kinase và Cdc25 phosphatase thì một lượng nhỏ M-Cdk ở trạng thái hoạt động có thể dẫn đến quá trình hoạt hoá nó nhanh chóng và hoàn toàn.
Câu 7.
Trong quá trình giảm phân, prôtêin Rec8 là một loại prôtêin đặc thù của phức hệ kết dính các yếu tố của NST kép được tổng hợp ở kì trung gian và prôtêin này chỉ bị phân rã ở giảm phân II. Các nhà khoa học đã tạo ra tế bào nấm men mang nhiễm sắc thể nhân tạo chứa gen Rec8 và cho biểu hiện đồng thời với tất cả các gen khác trong nguyên phân, từ đó tìm ra prôtêin Shugoshin chính là prôtêin ngăn cản sự phân rã Rec8 ở giảm phân I. Hãy dự đoán:
a. Bằng cách nào người ta biết là prôtêin Shugoshin mà không phải là các prôtêin khác ngăn cản sự phân rã của Rec8?
b. Khuẩn lạc nấm men có tế bào tái tổ hợp biểu hiện đồng thời Rec8 và Shugoshin sẽ như thế nào?
Hướng dẫn trả lời
a. Với các tế bào có Shugoshin được biểu hiện (có prôtêin Shugoshin) thì sự phân li NST trong nguyên phân diễn ra không bình thường.
- Các tế bào có các gen khác được biểu hiện và không có prôtêin Shugoshin thì quá trình nguyên phân diễn ra bình thường.
b. Vì có cả Shugoshin và prôtêin Rec8 cùng được biểu hiện trong nguyên phân nên có NST được phân li, NST khác không phân li do sự ngăn cản của prôtêin Shugoshin. Kết quả tạo ra nhiều loại tế bào con lệch bội khác nhau.
- Trong các tế bào có bộ NST bất thường có thể có kiểu gen gây chết, dẫn đến khuẩn lạc nấm men chậm phát triển.
Câu 8.
Một phòng thì nghiệm đang nghiên cứu một cặp thụ thể GPCR mới được xác định. GPCR – A và GPCR – B. Mỗi loại thụ thể này liên kết với cùng một loại phối tử nhỏ với ái lực như nhau nhưng hoạt hoá các prôtêin G khác nhau tác động đến adenyl cyclase. Khi được hoạt hoá, GPCR – A làm tăng hoạt động của adenyl cyclase, còn GPCR – B làm giảm hoạt động adenyl cyclase. Có 1 dòng tế bào biểu hiện cả GPCR –
A và GPCR – B, các G prôtêin tương ứng và adenyl cyclase. Adenyl cyclase hoạt động
ở mức cơ bản tạo ra nồng độ cAMP ở mức cơ bản. Khi nghiên cứu về một loại vi khuẩn gây bệnh, người ta đã phát hiện vi khuẩn này tiết ra một loại độc tố gây cản trở con đường truyền tín hiệu đã mô tả ở phía trên.
Để xác định hoạt động của độc tố này, người ta xác định mức cAMP nội bào trong các tế bào không được xử lí và các tế bào được xử lí độc tố (phối tử không được thêm vào trong cả 2 lô của thí nghiệm).
Hãy chỉ ra mỗi đột biến sau làm tăng, không thay đổi hay làm giảm lượng cAMP nội bào khi có mặt phối tử trong điều kiện không xử lí độc tố. Lưu ý, cả hai thụ thể GPCR – A và GPCR – B đều liên kết với cùng loại phối tử
Hình 1
Phát biểu nào dưới đây có thể giải thích cơ chế tác động của độc tố được mô tả ở (Hình 1) đến hoạt động của adenyl cyclase?
A. Đột biến ở GPCR – A ngăn chặn sự hoạt hoá G prôtêin.
B. Đột biến ở Gi ngăn chặn sự giải phóng GDP.
C. Đột biến ở Gi ngăn chặn sự thuỷ phân GTP.
D. Độc tố ức chế sự kích hoạt Gi khi hoạt hoá thụ thể.
E. Độc tố ức chế sự thuỷ phân GTP ở Gs.
F. Độc tố giống như GTP và gây ra sự hoạt hoá liên tục tất cả các G prôtêin.
G. Độc tố là chất ức chế của thụ thể GPCR – A.
H. Độc tố là chất hoạt hoá của thụ thể GPCR – B.
Hướng dẫn trả lời
A. Giảm.
B. Giảm. vì ngăn chặn giải phóng GDP làm ngăn chặn hoạt hoá G – prôtêin → không hoạt hoá adenyl cyclase → giảm cAMP.
C. Tăng. Vì không thuỷ phân GTP → G prôtêin giữ ở trạng thái hoạt hoá → liên tục hoạt hoá adenyl cyclase → tăng cAMP.
D. Sai. Vì ức chế kích hoạt Gi → ức chế hoạt hoá G prôtêin → không hoạt hoá adenyl cyclase → giảm cAMP.
E. Đúng. Vì ức chế thuỷ phân GTP → G prôtêin giữ ở trạng thái hoạt hoá → liên tục hoạt hoá adenyl cyclase → tăng cAMP.
F. Đúng. Vì tăng tác dụng của GTP → G prôtêin giữ ở trạng thái hoạt hoá → liên tục hoạt hoá adenyl cyclase → tăng cAMP.
G. Sai. Vì ức chế thụ thể A → giảm hoạt hoá adenyl cyclase → giảm cAMP.
H. Sai. Vì hoạt hoá thụ thể B → giảm hoạt hoá adenyl cyclase → giảm cAMP.
Câu 9.
a. Nuôi cấy cùng một chủng vi khuẩn trong hai môi trường khác nhau, môi trường A có đầy đủ chất dinh dưỡng, môi trường B nghèo chất dinh dưỡng. Sau một thời gian nuôi cấy trong môi trường A, các tế bào vẫn giữ cấu trúc và hình dạng bình thường. Trong môi trường B, ngoài các tế bào bình thường như ở trong môi trường A, người ta còn phát hiện thấy một số tế bào “lạ” có màng tế bào gấp nếp ở nhiều chỗ vào phía trong. Theo thời gian, người ta thấy số lượng các tế bào lạ tăng dần lên. Hãy giải thích kết quả thí nghiệm trên.
b. Nếu cho rằng gen phân mảnh là xu thế tiến hóa có ưu thế ở sinh vật nhân thực, thì màng nhân có vai trò gì về chức năng giúp gen phân mảnh trở nên có ưu thế tiến hóa? Giải thích.
Hướng dẫn trả lời
a. Trong môi trường nuôi cấy B nghèo dinh dưỡng, chọn lọc tự nhiên đã chọn lọc các tế bào có khả năng hấp thu chất dinh dưỡng hiệu quả hơn. Vì thế, những tế bào có màng được gấp nếp nhiều vào phía trong sẽ làm tăng diện tích hấp thu các chất nên được chọn lọc tự nhiên giữ lại, những tế bào bình thường (không có khả năng này) dần dần bị đào thải.
b. - Nhờ có màng nhân ngăn cách ARN sơ cấp với ribôxôm và nhiều phân tử kích thước lớn trong tế bào chất nên ARN sơ cấp phiên mã từ gen phân mảnh có điều kiện để cắt intron ghép exon và hoàn thiện trước khi được dịch mã ở tế bào chất. Sự cắt intron và nối các exon theo các cách khác nhau tạo ra nhiều loại mARN từ cùng một gen qua đó tạo ra nhiều prôtêin ở các loại tế bào khác nhau vì thế tiết kiệm được thông tin di truyền
- Nhờ có màng nhân ngăn cách nên tế bào nhân thực có thêm được cơ chế điều hòa sau phiên mã đối với các gen phân mảnh.
- Nếu màng nhân không được tiến hóa cùng hoặc trước khi có sự tiến hóa của gen phân mảnh thì gen phân mảnh sau khi phiên mã được dịch mã ngay sẽ tạo ra các prôtêin không có chức năng vì cả phần intron cũng được dịch mã tạo ra trình tự axit amin khác thường.
- Gen phân mảnh là cấu trúc cho phép hệ gen sinh vật nhân thực mở rộng hệ gen (tích lũy thêm thông tin di truyền), tăng khả năng tích lũy đột biến trung tính (đột biến câm/giảm thiểu hậu quả của đột biến) làm tăng tính phong phú nguyên liệu cho chọn lọc tự nhiên.
Câu 10.
Hệ prôtêin học bao gồm phân tích biểu hiện của toàn bộ prôtêin. Trong một cách tiếp cận, chiết toàn bộ prôtêin của tế bào đối chứng và tế bào đã xử lý, sau đó phân tách bằng điện di gel hai chiều.
Trong trường hợp chung nhất sẽ tạo hàng trăm hoặc hàng ngàn chấm prôtêin
tách biệt và cho phép so sánh mức biến thiên của mỗi prôtêin trong hai mẫu. đơn giản hóa, ví dụ dưới đây chỉ thể hiện một ít chấm prôtêin. prôtêin phân tách theo chiều thứ nhất dựa vào điện tích bởi tụ tiêu đẳng điện (pH 4-10) và sau đó phân tách theo kích thước bằng điện di gel SDS polyacrylamide. Phát hiện prôtêin bằng cách sử dụng thuốc nhuộm xanh Comassie và đánh số để nhận diện.
a. Xử lý ("+ thuốc") hoặc không xử lý ("Đối chứng") tế bào với thuốc Sau đó chiết và phân tách prôtêin bằng kỹ thuật điện di gel hai chiều. Các gel được nhuộm và thể hiện dưới đây. Có kết luận gì về hiệu quả của thuốc đối với độ biến thiên của các prôtêin 1-7?
b. Bạn nghi ngờ rằng thuốc có thể cảm ứng prôtêin kinase và do đó lặp lại thí nghiệm trong phần (a) sử dụng phosphate vô cơ đánh dấu phóng xạ 32P. Thí nghiệm này sử dụng phim X quang để phát hiện prôtêin đánh dấu 32P trong gel hai chiều. Hình dưới đây thể hiện kết quả trên phim X quang. Có thể kết luận gì về hiệu quả của thuốc trên đối với prôtêin 1-7 từ thí nghiệm này?
c. Để xác định vị trí trong tế bào của cácprôtêin 1-7, dùng kỹ thuật ly tâm vi sai để tách riêng prôtêin ở phần (a) theo phân đoạn nhân và tế bào chất. Hình dưới đây thể hiện kết quả nhuộm sau khi điện di gel hai chiều. Bạn có kết luận gì về vị trí trong tế bào của các prôtêin 1-7?
d. Từ toàn bộ tính chất của prôtêin 1-7 thu được từ dữ liệu của các phần (a), (b) và (c). Mô tả cách xác định nhân dạng của từng prôtêin trong số chúng?
Hướng dẫn trả lời
a. Thuốc không làm thay đổi khối lượng của prôtêin vì vị trí của các prôtêin 1-7 vẫn giữ nguyên ở hai bản 2D-PAGE. Thuốc chỉ làm thay đổi điện tích của prôtêin 1; 2; 4 vì thay đổi đường kính các chấm prôtêin khác nhau trên 2 bản 2D-PAGE. Thuốc không có hiệu quả với prôtêin 3; 5; 6; 7 do điện tích và kích thước không thay đổi trên cả hai bản 2D-PAGE.
b. -prôtêin 2, 6 là prôtêin kinase vì thể hiện kết quả trên phim X quang khi đánh dấu bằng 32P ở bản gel đối chứng. Các prôtêin 1; 3; 4; 5; 7 đều không phải prôtêin kinase vì không thể hiện kết quả ở bản gel đối chứng.
- Thuốc làm prôtêin 3 có hoạt tính kinase vì ở bản gel “+ thuốc” prôtêin 3 đã thể hiện kết quả.
- Vậy thuốc có hiệu quả với prôtêin 1; 2; 3; 4. Thuốc không có hiệu quả với prôtêin 5; 6; 7 do điện tích và kích thước không thay đổi trên cả hai bản 2D-PAGE.
c. - Khi chưa bổ sung thuốc thì prôtêin 1; 5; 6 ở trong nhân tế bào, prôtêin 2; 3; 4; 7 ở tế bào chất. Vì sau khi ly tâm và điện di thì prôtêin 1; 5; 6 chỉ hiển thị trong bản gel của prôtêin nhân tế bào, prôtêin 2; 3; 4; 7 chỉ hiển thị trong bản gel của prôtêin ở tế bào chất.
- Sau khi bổ sung thuốc thì prôtêin 7 được vận chuyển vào trong nhân tế bào,
các prôtêin còn lại vẫn giữ nguyên vị trí.
d. - Sử dụng điện di 2 chiều trên gel để xác định kích thước, khối lượng của prôtêin.
- Sử dụng đánh dấu phóng xạ 32P và chụp ảnh tia X trên bản gel 2 chiều để xác định hoạt tính kinase của prôtêin.
- Sử dụng ly tâm vi sai cùng điện di 2 chiều để xác định vị trí của prôtêin trong tế bào là trong nhân hay trong tế bào chất.
1.2. Câu hỏi và bài tập kết hợp hóa sinh – Vi sinh – Sinh học phân tử.
Câu 1.
X là prôtêin có tác dụng ngăn ngừa sự tăng cân ở người. prôtêin này bất hoạt ở những người béo phì. Các phân tử mARN trưởng thành của X phân lập được từ một số người béo phì của cùng một gia đình cho thấy, chúng thiếu một đoạn trình tự dài 173 nucleotit so với các phân tử mARN trưởng thành phân lập được từ những người bình thường. Khi so sánh trình tự gen mã hóa cho prôtêin X của người bình thường và người béo phì, người ta phát hiện ra rằng không có nucleoti nào bị mất mà chỉ có 1 nucleotit bị thay đổi. Sự thay đổi này xảy ra ở vùng intron của gen.
a. Tại sao việc thay đổi 1 nucleotit lại có thể làm cho mARN trưởng thành của gen đột biến bị mất một đoạn dài 173 nucleotit. Giải thích và minh họa bằng hình vẽ.
b. Giải thích hiện tượng bất hoạt của prôtêin X ở người béo phì.
c. Để xản xuất prôtêin này, người ta sử dụng kỹ thuật di truyền tạo plasmit tái tổ hợp giữa thể truyền với gen mã hóa prôtêin X của người bình thường, sau đó chuyển vào vi khuẩn E. coli để sản xuất sinh khối. Sản phẩm prôtêin tạo ra có bị bất hoạt không? Giải thích.
Hướng dẫn trả lời
a. Đột biến thay thế nucleotit này bằng nucleotit khác xảy ra trong vùng intron của gen nhưng lại làm cho phân tử mARN trưởng thành tổng hợp từ gen đột biến ngắn hơn phân tử mARN trưởng thành của gen bình thường 173 nucleotit, chứng tỏ đột biến trên đã xảy ra ở vùng nhận biết và cắt intron làm cho quá trình cắt intron bị biến đổi
Thay vì chỉ cắt các trình tự intron thì tế bào đã cắt một trình tự gồm 1 đoạn exon xen kẽ 2 đoạn intron và đoạn exon này dài 173 nucleotit nên phân tử mARN trưởng thành của gen đột biến kém phân tử mARN trưởng thành của gen bình thường 173 nucleotit.
b. Đoạn exon bị mất dài 173 nucleotit. Đây là một số không chia hết cho 3, có nghĩa là exon này có nucleotit kết hợp với nucleotit của exon khác để tạo thành một bộ ba hoàn chỉnh.
Khi đoạn exon này bị mất sẽ gây ra hiện tượng dịch khung đọc đối với toàn bộ trình tự nucleotit phía sau exon này, nên phân tử prôtêin tổng hợp từ gen đột biến sẽ mất đoạn axit amin do 173 nucleotit mã hóa đồng thời bị thay đổi toàn bộ trình tự axit amin ở đoạn peptit phía sau, làm cho prôtêin bị mất chức năng (bất hoạt)
c. Sản phẩm prôtêin tạo ra vẫn bị bất hoạt vì plasmit tái tổ hợp tạo ra từ gen mã hóa prôtêin X từ người bình thường mang cả đoạn intron. Đoạn trình tự này vẫn được phiên mã và giải mã bình thường trong cơ thể vi khuẩn. Vì thế, phân tử prôtêin tạo ra có trình tự dài hơn hoặc ngắn hơn trình tự axit amin của prôtêin X. Mặt khác, trình tự axit amin của sản phẩm prôtêin tạo ra sẽ khác với trình tự các axit aminh của prôtêin X.
Câu 2.
Giả sử một gen mã hóa một prôtêin đặc thù ở vi khuẩn có trình tự nuclêôtit của một mạch như sau:
5′ -ATGACAACCCATC……………………..CCCGAGCACAAGTAA- 3′
Để sản xuất ở quy mô lớn loại prôtêin này, người ta tiến hành nhân dòng đoạn gen trên vào một vectơ, tạo plasmit tái tổ hợp và biến nạp vào E. coli. Sau đó người ta tiếp tục cho cảm ứng biểu hiện và sản xuất sinh khối. Để tách prôtêin sản phẩm bằng sắc ký ái lực qua cột ion Ni2+, vectơ biểu hiện thường có một trình tự mã hóa 6 axit amin histadin ở đuôi cacbôxyl hoặc ở đầu amin. Giả sử vị trí phân dòng của vectơ biểu hiện có trình tự là: 5′
CGCTGCAGATGAATATGCGCGGGGATCCGACCACCACCACCACCACCAC- 3′
(Phần in đậm gạch chân là trình tự nhận biết của enzim cắt giới hạn PstI và BamHI, phần in nghiêng gạch chân là trình tự nuclêôtit của 6 bộ ba liên tiếp mã hóa axit amin histadin)
Hãy đề xuất trình tự nuclêôtit của cặp mồi nhân dòng để prôtêin đích được gắn thêm 6 axit amin histadin. Giải thích.
Hướng dẫn trả lời - Để sản phẩm prôtêin có 6 axit amin histadin ở đuôi cacbôxyl thì mã kết thúc phải được phá bỏ để quá trình giải mã kéo dài bao trùm cả 6 bộ ba mã hóa histadin. Đồng thời trình tự đoạn mồi cần phải chứa cả enzim cắt giới hạn để có thể nhân dòng phụ sang vectơ biểu hiện. Mặt khác, để tách được dòng gen, tạo sản phẩm chính xác, trình tự đoạn mồi cần thiết phải bổ sung với mạch gốc.
- Xem xét trình tự của vectơ, phía sau vị trí nhận biết chứa một bộ ba, tiếp đó là 6 bộ ba mã hóa histadin. Vì thế, đột biến phá vỡ mã kết thúc phải làm mất hẳn cả 3 nuclêôtit của mã kết thúc hoặc thêm số nuclêôtit là số chia hết cho 3. Thông thường người ta hạn chế tăng mã bộ ba.
Bởi vậy, trình tự nuclêôtit của cặp mồi tách dòng gen trên có thể là:
Mồi xuôi: 5′ -AGCTGCAGATGACAACCATC- 3′
Mồi ngược: 5′ -ATCGGTACCGAACACGAGCCC- 3′ ’
Ghi chú: Nuclêôtit đầu 5′ có thể không nhất thiết bổ sung với mạch khuôn.

Tuy nhiên, để có đoạn mồi chuẩn, chúng ta cần kiểm tra các đặc điểm khác của mồi trước khi sử dụng. Để đạt các điều kiện tối ưu của mồi, có thể thêm nuclêôtit bất kỳ ở đầu 5′ hoặc kéo dài đoạn bổ sung với mạch khuôn.
Câu 3.
Dưới đây là một đoạn trình tự nucleotitde thuộc vùng mã hóa của một gen quy đinh chuỗi polipeptit có 300 acid amin , mang bộ 3 tương ứng mã mở đầu và chưa xác định được các đầu tận cùng ( 3′ hoặc 5′ ) của đoạn gen này .
A G A T G T A G T A X G G A A T T G A T X X A G T A A G T X A T T
- Phiên mã từ trái sang phải hoặc từ phải sang trái, mARN đều có bộ 3 mở đầu 5′ AUG 3′ . Nhưng nếu theo chiều từ trái sang phải, cách bộ ba 5′ AUG 3′ một bộ ba khác là bộ ba kết thúc 5′ UAA 3′ → Không thỏa mãn
- Do đó, sợi bên trên làm khuôn cho phiên mã, theo chiều từ phải sang trái
- Trình tự và các đầu tận cùng của đoạn mARN, vị trí bộ 3 mở đầu là:
5′ GAAUGAXUUAXUGGAUXAAUUXXGUAXUAXAUXU 3′ +1
Hoặc cũng có thể viết:
3′ UXUAXAUXAUGXXUUAAXUAGGUXAUUXAGUAAG 5′ +1
b. Vẽ mũi tên chỉ chiều phiên mã và viết các đầu tận cùng ( 3′ và 5′ )
5′ A G A T G T A G T A X G G A A T T G A T X X A G T A A G T X A T T X 3′
X
G
T X T A X A T X A T G X X T T A A X T A G G T X A T T X A G T A A
a. Dựa vào trình tự nucleotit của đoạn gen trên , hãy nêu cách xác định sợi làm khuôn cho quá trình phiên mã của gen. Viết trình tự Nu của đoạn mARN được phiên mã từ đoạn gen này với các đầu tận cùng ( 3′ hoặc 5′ ) và viết kí hiệu + 1 để xác đinh bộ ba mở đầu dịch mã.
b. Không thay đỏi các viết thứ tự các Nu , hãy viết lại trình tự Nu của đoạn gen đã cho và bổ sung mũ tên chỉ chiều phiên mã , kí hiệu vị trí tương ứng mã mở đầu ( +1) và các đầu tận cùng ( 3′ hoặc 5′ ) trên 2 sợi của đoạn gen này
Hướng dẫn trả lời
a. Đây là đoạn thuộc vùng mã hóa và mang bộ ba mở đầu dịch mã nên mARN tương ứng được tạo thành phải mang bộ 3 mở đầu 5′ AUG 3′
- Nếu sợi bên dưới làm khuôn cho phiên mã, trình tự mARN là :
5′ AGAUGUAGUAXGGAAUUGAUXXAGUAAGUXAUUX 3′
- Nếu phiên mã theo chiều từ trái sang phải hoặc từ phải sang trái mARN nều xuất hiện bộ 3 mở đầu 5′ AUG 3′ (như gạch chân. Nhưng kế tiếp hoặc gần kế tiếp các bộ ba 5′ AUG 3′ này đều là các bộ ba kết thúc dịch mã ( 5′ UGA 3′ ) → đây không phải là trình tự mARN đúng của gen này
- Sợi bên trên là sợi làm khuôn cho phiên mã. Khi đó mARN có trình tự là:
5′ UXUAXXAUXAUGXXUUAAXUAGGUXAUUXAGUAAG 3′
3′ T X T A X A T X A T G X X T T A A X T A G G T X A T T X A G T A A G 5′ +1
Một nhóm các nhà khoa học thực hiện lại thí nghiệm của Meselson và Stahl (1958) để nghiên cứu mô hình nhân đôi của ADN. Họ đã nuôi vi khuẩn E. coli trong môi trường chỉ có nitơ đồng vị nặng (N15), sau đó chuyển sang môi trường chỉ có nitơ đồng vị nhẹ (N14), tách ADN sau mỗi thế hệ và ly tâm. Kết quả thu được các băng ADN có trọng lượng và tỷ lệ khác nhau như hình 1 (tỷ lệ % thể hiện hàm lượng của các băng ADN trong mỗi thế hệ).
a. Dựa theo kết quả thí nghiệm trên, hãy chứng minh ADN tái bản theo mô hình bán bảo toàn.
b. Tại sao băng ADN trung gian có tỷ lệ giảm dần sau mỗi thế hệ?
c. Giả sử họ nuôi vi khuẩn E. coli trong môi trường chỉ có N14 (thế hệ 0) rồi chuyển sang môi trường chỉ có N15 (từ thế hệ 1), các điều kiện thí nghiệm khác không thay đổi. Hãy xác định tỷ lệ % các băng ADN thu được ở mỗi thế hệ: 0, 1, 2 và 3.
Hướng dẫn trả lời
a. Ba mô hình giải thích cơ chế tái bản ADN: bán bảo toàn, bảo toàn và phân tán. - Thế hệ 1: chỉ có 1 băng ADN trung gian, nên loại bỏ mô hình bảo toàn.
- Thế hệ 2: có 2 băng ADN (nhẹ và trung gian) → loại bỏ mô hình phân tán. Từ
đó kết luận: ADN tái bản theo mô hình bán bảo toàn.
b. Thế hệ 1: Băng ADN trung gian tạo thành từ các phân tử ADN lai (N14-N15)
- Do môi trường nuôi vi khuẩn từ thế hệ 1 trở đi chỉ có N14 làm nguồn vật liệu tổng hợp ADN → sau mỗi thế hệ tỷ lệ này giảm một nửa.
- Thế hệ 2: ADN trung gian giảm còn 1/2,
- Thế hệ 3: ADN trung gian giảm còn 1/4
- Tỷ lệ phân tử ADN nhẹ (N14- N14) tăng lên sau mỗi thế hệ → Tỷ lệ băng ADN trung gian giảm dần sau mỗi thế hệ
c. - Tỷ lệ các băng ADN thu được ở các thế hệ:
+ Thế hệ 0: 100% băng nhẹ
+ Thế hệ 1: 100% băng trung gian.
+ Thế hệ 2: 50% băng trung gian: 50% băng ADN nặng.
+ Thế hệ 3: 25% băng trung gian: 75% băng ADN nặng.
Câu 5.
Trôpônin T là một loại prôtêin liên quan đến hoạt động của cơ. Để thu
được prôtêin này của người trong tế bào vi sinh vật, người ta tiến hành hai thí nghiệm như sau:
Thí nghiệm 1: Tách gen mã hoá trôpônin T từ ADN hệ gen người và chuyển vào vectơ biểu hiện ở nấm men tạo plasmit tái tổ hợp A1; chuyển vào vectơ biểu hiện ở vi khuẩn tạo plasmit tái tổ hợp A2
Thí nghiệm 2: Tổng hợp phân tử cADN bằng cách phiên mã ngược phân tử mARN trưởng thành mã hoá trôpônin T. Sau đó, chuyển phân tử cADN này vào cùng loại vectơ biểu hiện ở nấm men tạo plasmit tái tổ hợp B1; chuyển vào cùng loại vectơ biểu hiện ở vi khuẩn tạo plasmit tái tổ hợp B2.
a. Hãy so sánh kích thước của hai plasmit tái tổ hợp A1 và B1. Giải thích.
b. Khi biến nạp và biểu hiện hai plasmit A1 và B1 trong tế bào nấm men trong cùng điều kiện thí nghiệm, họ thu được hai loại prôtêin có kích thước khác nhau từ plasmit A1 và một loại prôtêin từ plasmit B1. Hãy giải thích kết quả
c. Khi biến nạp và biểu hiện hai plasmit A2 và B2 vào vi khuẩn E. coli trong cùng điều kiện thí nghiệm, prôtêin thu được từ plasmit A2 không có chức năng, còn prôtêin thu được từ plasmit B2 có chức năng bình thường. Hãy giải thích kết quả.
Hướng dẫn trả lời
a. Xét 2 trường hợp:
- TH1: gen Troponin T là gen phân đoạn, thì kích thước plasmit A1 lớn hơn B1, do plasmit A1 mang gen có chứa các đoạn intron, còn plasmit B1 không chứa intron.
-TH2: gen Troponin T là gen không phân đoạn, thì kích thước plasmit A1 bằng B1, do plasmit A1 và B1 đều mang gen không chứa các đoạn intron.
b. Plasmit A1 phiên mã tạo phân tử tiền thân của mARN → trải qua quá trình cắt intron và nối exon → các phân tử mARN trưởng thành khác nhau về số lượng các exon → dịch mã tạo các phân tử prôtêin kích thước khác nhau.
Trong khi đó, plasmit B1 chỉ còn các exon nên không trải qua quá trình cắt nối thay thế và chỉ tạo ra một loại prôtêin.
c. Hệ thống biểu hiện gen ở vi khuẩn không có bộ máy cắt intron và nối exon (spliosome) như ở nấm men.
mARN tạo ra từ plasmit A2 vẫn còn intron nên tạo thành prôtêin không có chức năng. Còn mARN từ plasmit B2 chỉ có các trình tự exon nên tạo prôtêin có chức năng.
Câu 6.
Các thể đột biến ở E. coli trên gen ADNA thường gây chết và được nghiên cứu để phân lập các đột biến điều kiện, mẫn cảm nhiệt độ. Các chủng đột biến này vẫn có thể tái bản ADN của chúng ở nhiệt độ cho phép là 18oC, nhưng không sinh trưởng hay sao chép ADN ở nhiệt độ giới hạn là 37oC. Người ta quan sát thấy như sau: các nghiên cứu sử dụng ADN không xoắn chặt không cần có prôtêin ADNA. Thứ hai, nếu các tế bào nguyên vẹn được nuôi ở 18oC rồi chuyển sang 37oC, ADN tiếp tục tổng hợp ở nhiệt độ này đến khi vòng sao chép được hoan thành và sau đó dừng lại. Những quan sát này cho biết vai trò của sản phẩm gen ADNA là gì?
Hướng dẫn trả lời
Sản phẩm của gen ADNA tạo ra prôtêin ADNA có vai trò là 1 loại enzim tháo xoắn ADN (Helicase) trong quá trình sao chép.
- Các chủng đột biến này có thể có sự mẫn cảm với nhiệt độ của prôtêin ADNA → khi ở nhiệt độ 37oC, prôtêin bị biến tính → ADN không tháo xoắn → tổng hợp ADN không diễn ra.
- Khi tế bào nguyên vẹn được nuôi ở 18oC, prôtêin ADNA có tác dụng tháo xoắn ADNA, khi chuyển sang 37oC, có thể ADN đã tháo xoắn xong → Vẫn tiếp tục nhân đôi.
- Cho đến đợt nhân đôi tiếp theo thì prôtêin ADNA biến tính và không tháo xoắn ADN được → Tổng hợp ADN dừng lại.
Câu 7.

Người ta phân lập được nhiều thể đột biến khuyết dưỡng ở Neurospora (nấm mốc). Phân tích các thể đột biến này, người ta thấy chúng có thể sinh trưởng được trên môi trường tối thiểu có bổ sung các hợp chất khác nhau: A, B, C, D. Bảng dưới đây ghi lại kết quả sinh trưởng của các thể đột biến ở những môi trường có bổ sung các hợp chất tương ứng (+: Sinh trưởng được; -: không sinh trưởng được). Cho biết, các hợp chất A, B, C và D cùng thuộc một con đường hóa sinh.
Thể đột biến Hợp chất A B C D 1 + + - + 2 + + + +
- - - +
+ + + +
Hãy cho biết thứ tự của các hợp chất này trong con đường đó và cho biết các thể đột biến đó thuộc về gen mã hóa các enzim ở bước chuyển hóa nào.
Hướng dẫn trả lời
* Thứ tự: S (cơ chất) → C → A → B → D. Vì:
- Hợp chất nào có tất cả các thể đột biến ( ĐB) đều sinh trưởng ( ST) được sẽ nằm ở cuối cùng trong chuỗi chuyển hóa → Hợp chất D
- Hợp chất nào có ít thể ĐB nhất ST được sẽ nằm ở đầu trong chuỗi chuyển hóa
→ Hợp chất C
- Hợp chất có số thể ĐB ST được nhiều hơn → nằm sau trong chuỗi chuyển hóa (Hợp chất B). Hợp chất có số thể ĐB ST được ít hơn -> nằm trước trong chuỗi chuyển hóa (Hợp chất A).
Thứ tự S → C → A → B → D.
Thể ĐB ST được trên ít hợp chất → ĐB xảy ra ở sau trong chuỗi chuyển hóa.
Thể ĐB ST được trên nhiều hợp chất → ĐB xảy ra ở trước trong chuỗi chuyển hóa.
Câu 8.
Ở sinh vật nhân chuẩn, prôtêin thường bắt đầu tổng họp tại côđôn AUG đầu tiên trên mARN. Tuy nhiên đôi khi điều này không diễn ra đúng như vậy, thay vào đó
ribôxôm quét qua côđôn AUG đầu tiên, và bắt đầu tổng hợp prôtêin ở vị trí AUG nằm bên trong. Để tìm hiểu đặc trưng nào của mARN ảnh hưởng đến hiệu quả khởi đầu tại vị trí AUG, nhiều nghiên cứu đã được thực hiện, sử dụng quá trình tổng hợp chloramphenicol acetyltransferase làm mô hình. Quá trình dịch mã enzim này có thể tạo ra prôtêin gọi là preCAT hoặc một prôtêin nhỏ hơn chút ít, CAT (xem M. Kozak, 2005, Gen 361:13). Hai prôtêin khác nhau ở chỗ một số axit amin tại đầu N của preCAT không nằm trong CAT. Không thể cắt preCAT để thu CAT nhưng khởi đầu dịch mã mARN tại AUG nằm bên trong tạo ra CAT:
preCAT CAT
Khởi đầu Khởi đầu Kết thúc
a. Kết quả từ một số nghiên cứu mang lại giả thiết rằng trình tự 5′ (3)ACCAUGG(+4) 3′ (chữ đỏ thể hiện côđôn khởi đầu AUG) cung cấp điều kiện tối ưu cho khởi đầu tổng họp prôtêin và đảm bảo rằng ribôxôm không quét qua AUG đầu tiên và khởi đầu ở AUG phía sau. Trong một số sơ đồ thể hiện dưới đây, A của AUG khỏi đầu được đánh số (+1); các bazơ phía 5′ của vị trí này được đánh số âm (vì vậy bazơ đầu tiên của trình tự này là (-3)), và các bazơ phía 3′ của vị trí (+1) A được đánh số dương (nên bazơ cuối cùng của trình tự này là (+4)). Để kiểm tra giả thiết rằng trình tự chứa vị trí khởi đầu (-3)ACCAUGG(+4) giúp ngăn chặn quét sót (leaky scanning), nhà nghiên cứu biến đổi trình tự mARN của chloramphenicol acetyhransferase và đánh giá hiệu ứng đối với quá trình dịch mã. Trong hình dưới đây, trình tự (màu đỏ) quanh côđôn AUG đầu tiên (màu đen) của mARN dẫn tới tạo ra preCAT (đường chạy thứ 3). Những biến đổi mARN này được thể hiện phía trên đường chạy của gel (nucleotit bị thay đổi có màu xanh), và toàn bộ prôtêin tạo ra từ mARN đã biến đổi xuất hiện dưới dạng các băng trên gel SDS - polyacrylamide. Cường độ của mỗi băng tương ứng với lượng prôtêin được tổng hợp. Phân tích những thay đổi so với trình tự kiểu dại, và mô tả làm thế nào mà chúng tác động đến quá trình dịch mã. Phải chăng một số vị trí nucleotit có vai trò quan trọng hơn các vị trí khác? Dữ liệu thể hiện trong hình này có hỗ trợ cho giả thiết rằng
trình tự xung quanh AUG đầu tiên tác động đến hiệu quả dịch mã tại vị trí này không? Phải chăng
ACCAUGG là vùng trình tự tối thích để dịch mã khởi đầu tại AUG đầu tiên?
b. Đột biến trong hệ gen có thể cung cấp tri thức về những cơ chế phức tạp của tế bào hoặc về tiến trình phát triển. Phân tích đột biến trong hệ gen sẽ khác nhau thế nào khi đột biến phân tích có tính lặn hoặc trội?
c. Đột biến gây ra một loại bệnh nghiêm trọng về máu được tìm thấy ở một gia đình. Hình dưói đây cho thấy đột biến nằm tại vùng không dịch mã đầu 5′ và làm thay đổi mARN của gen mã hóa hepcidin. Hai phân đoạn trong hình là trình tự mã hóa của gen đột biến và gen bình thường. mARN bị thay đổi không tạo ra hepcidin và
điều này gây ra bệnh. Giải thích vì sao hepcidin không được tổng họp trong các thành viên của gia đình bị đột biến di truyền này? Trong trường họp này có thể suy luận gì về tầm quan trọng của vùng trình tự nơi khởi đầu tổng hợp prôtêin xảy ra?
Hướng dẫn trả lời a. Đường chạy số 3, 4 không có CAT, (trong khi các đường chạy còn lại đều có CAT) và đều có trình tự trước AUG là ACC trong khi các đường chạy khác không có → trình tự ACC quan trọng giúp ngăn chặn quét sót.
- Đường chạy số 1, 2 có CAT trong đó đường chạy 1 có nhiều CAT nhất và chúng đều có trình tự trước AUG là UUU nhưng chỉ có đường chạy 1 bị thay đổi trình tự sau AUG thành U; đường chạy số 3 có nhiều preCAT hơn đường chạy 4 và chỉ đường chạy số 3 có G sau AUG → trình tự G sau AUG quan trọng giúp ngăn chặn quét sót.
- Đường chạy số 5 (kiểu dại) có CAT vì có trình tự trước AUG khác ACC và trình tự sau AUG khác G.
Vậy trình tự số -3 đến -1 có vai trò quan trọng hơn các vị trí khác.
- Dữ liệu thể hiện trong hình có hỗ trợ cho giả thiết rằng trình tự xung quanh
AUG đầu tiên tác động đến hiệu quả dịch mã tại vị trí này.
- Đường chạy số 3 thể hiện khả năng tránh quét sót mạnh nhất → ACCAUGG là vùng trình tự tối thích để dịch mã khởi đầu tại AUG đầu tiên.
b. - Đột biến có tính lặn khi đột biến dẫn tới bất hoạt gen, làm mất chức năng một phần hoặc toàn bộ Đột biến lặn có thể loại bỏ một phần hoặc toàn bộ gen ra khỏi nst, làm gián đoạn biểu hiện gen, hoặc thay đổi cấu trúc dẫn tới thay đổi chức năng prôtêin do gen này mã hoá. Đột biến chỉ biểu hiện ra kiểu hình khi cả hai alen trong cặp tương đồng là dạng đột biến.
- Đột biến có tính trội thường do đột biến thêm chức năng, làm tăng hoạt tính của prôtêin được mã hoá, mang lại kiểu hình mới, hoặc dẫn tới mô hình biểu hiện gen sai lệch về không gian hoặc thời gian. Đột biến biểu hiện ra kiểu hình ngay cả khi một trong hai alen trong cặp tương đồng là dạng đột biến và luôn biểu hiện ở sinh vật đơn bội.
c. Bắt đầu hepcidin

Đột biến
...GCAAUGGGACAGCCAGACAGACGGCACGAUGGCACU.....
- Đột biến thay thế cặp nucleotit C – G bằng T - A làm xuất hiện bộ ba AUG thay thế bộ ba GUG ban đầu.

- Bộ ba AUG là bộ ba mở đầu dịch mã. Khi xuất hiện 1 bộ ba AUG khác trước bộ ba AUG mở đầu dịch mã cho hepcidin thì tARNmet có thể nhận biết và liên kếtbộ ba đối mã vào bộ ba AUG phía trước đó, mở đầu dịch mã từ bộ ba đột biến làm thay đổi thành phần axit amin trong chuỗi polipeptit → hepcidin không được tổng hợp.
- Vùng trình tự nơi khởi đầu tổng hợp prôtêin có vai trò rất quan trọng, giúp tăng ái lực với riboxom và giúp riboxom dịch mã chính xác tại vị trí +1 AUG.
Câu 9.
Gây đột biến chủng nấm men cần uracil để phát triển (ura3-), và phân lập được hai khuẩn lạc đột biến (X và Y). Giao phối các tế bào kiểu a của đột biến X với tế bào kiểu å của đột biến Y để tạo tế bào lưỡng bội. cấy trải bố mẹ (ura3-)t X, Y, và tế bào lưỡng bội trên đĩa thạch chứa uracil và ủ tại 23°c hoặc 32°c. Giám sát sự phát triển của tế bào dựa trên khuẩn lạc hình thành trên đĩa nuôi cấy như thể hiện trong hình dưới đây.
a. Từ dữ liệu trên có thể suy luận gì về các thể đột biến X và Y?
b. Thư viện cADN của nấm men kiểu dại, tạo thành từ plasmit chứa gen URA3+ kiểu dại được sử dụng để biến nạp vào các tế bào X sau đó nuôi cấy như đã trình bày. Mỗi chấm màu đen ở dưới đại diện cho một dòng phát triển trên đĩa petri. Nêu sự khác biệt ở mức phân tử giữa các dòng phát triển trên hai đĩa? Dựa vào kết quả làm thế nào để xác định plasmit chứa gen X kiểu dại? Có thể sử dụng những kết quả trên như thế nào để xác định gen mã hóa X?
...GCAGUGGGACAGCCAGACAGACGGCACGAUGGCACUG... Bình thường 286
c. Tách ADN từ tế bào mẹ, lẫn tế bào X và Y rồi cắt bằng enzim giói
hạn. Mồi PCR được sử dụng để khuếch đại gen mã hóa X ở tế bào mẹ và tế bào X. Mồi bổ sung với các vùng ADN nằm ngay ngoài gen mã hóa X. Gel bên phải thể hiện kết quả PCR. Có thể suy luận gì về đột biến trong gen X từ những dữ liệu này?
- 1 dòng phát triển ở nhiệt độ 320C. Vì gen URA3+ được tái tổ hợp không tương đồng với đoạn gen URA3-, không làm mất gen đột biến nhạy nhiệt nên nấm men phát triển ở điều kiện 320C và không cần uracil.
- Dựa vào kết quả là sự xuất hiện khuẩn lạc hay không có thể xác định plasmit chứa gen X kiểu dại hay không.
- Có thể thu các nấm men ở các khuẩn lạc hai đĩa để giải trình tự và so sánh trình tự, từ đó xác định vị trí gen mã hoá X.
c. - Đột biến trong gen X làm xuất hiện thêm hoặc mất điểm cắt giới hạn nên kết quả Southern của X cho thấy xuất hiện trình tự khác ADN mẹ và Y.
Kết quả Southern PCR d. Tạo gen dung hợp mã hóa prôtêin X kiểu dại và prôtêin phát huỳnh quang xanh (GFP) tại đầu N (GFP-X) hoặc đầu C (X-GFP) của prôtêin X. Biến nạp hai prôtêin dung hợp này (nằm trong plasmit URA3+) vào tế bào X phát triển trong môi trường không chứa uracil. Sau đó quan sát thể biến nạp phát triển tại 32°c và thấy như hình phía dưói bên trái. Bên phải là hình ảnh huỳnh quang của các tế bào X-GFP và GFP-X phát triển tại 230 C ở đó màu xanh lá cây chỉ ra sự có mặt của prôtêin huỳnh quang xanh. Nêu giải thích họp lý cho việc tế bào GFP-X phát triển tại 32°c còn tế bào X-GFP thì không?


- Đột biến nằm giữa gen X chứ không nằm hai đầu điểm cắt giới hạn của X vì kết quả PCR vẫn giống mẹ chứng tỏ hai đầu gen vẫn nguyên vẹn trình tự bổ sung với mồi.
- Vậy đột biến trong gen X là đột biến thêm trình tự cắt giới hạn. d. - Ở 230C là các tế bào có gen X tái tổ hợp tương đồng thay thế gen URA 3nên sống được trong môi trường không có uracil.
- Gắn GFP vào đầu N của X làm tín hiệu cho GFP-X vận chuyển vào trong nhân tế bào và hoạt động trong nhân tế bào, ít chịu ảnh hưởng của nhiệt độ hơn nên GFP-X phát triển tại 320C.
- Gắn GFP vào đầu C của X làm tín hiệu cho X-GFP không được vận chuyển vào nhân và hoạt động ngoài tế bào chất, chịu nhiều ảnh hưởng của nhiệt độ hơn nên tế bào X-GFP không phát triển ở nhiệt độ 320C. (giống X kiểu dại).
Câu 10.
Hướng dẫn trả lời
a. Thể đột biến X và Y không mọc khuẩn lạc ở nhiệt độ 230C, mọc khuẩn lạc ở
320C.
- ura3- và thể lưỡng bội mọc khuẩn lạc ở cả hai điều kiện nhiệt độ.
→ Thể đột biến X, Y bị đột biến nhạy nhiệt, chỉ phát triển trong điều kiện
320C.. thể a của X và thể å của Y là hai đột biến nằm trên hai gen khác nhau vì chúng có thể bổ trợ nhau (mọc khuẩn lạc) ở thể lưỡng bội.
b. - Các dòng phát triển phát triển ở điều kiện 230C có chứa gen URA3+ được tái tổ hợp tương đồng thay thế gen URA3-, làm mất gen đột biến nhạy nhiệt nên nấm men phát triển ở điều kiện 230C và không cần uracil.
ADN vệ tinh là một bộ phận của hệ gen và có mặt ở ADN mã hóa lẫn ADN không mã hóa. Khi nó nằm trong ADN mã hóa, số lượng trình tự lặp có thể làm biến đổi prôtêin. Tuy nhiên tác dụng của các đoạn lặp này ở ADN không mã hóa vẫn chưa được hiểu rõ. Để xác định liệu các đoạn lặp trong vùng promoter có thể thay đổi biểu hiện gen và cô đặc nhiễm sắc thể, Vinces và cộng sự tìm kiếm các trình tự ADN lặp trong promoter trong hệ gen Saccharomyces cerevisiae và xem biến đổi số lượng trình tự lặp ảnh hưởng như thế nào đến biểu hiện gen và đóng gói ADN.
a. Đầu tiên nhóm tìm kiếm ADN vệ tinh ở hệ gen của nhiều chủng s.cerevisiae và thấy 25 phần trăm promoter chứa ít nhất một đoạn lặp. Ngoài ra, cùng một promoter ở các chủng khác nhau thường có số lượng các đoạn lặp ở mỗi vùng ADN vệ tinh khác nhau. Cơ chế nào có thể làm tăng số lượng trình tự lặp ở một vùng ADN vệ tinh?
b. Để xác định xem có mối quan hệ giữa số lượng các trình tự lặp với biểu hiện gen, tiến hành phân tích phiên mã gen SDT1. Promoter SDT1 chứa ADN vệ tinh và số lượng các đoạn lặp trong vùng này được biến đổi từ 0 đến 60 đoạn lặp. Phân tích bằng PCR phiên mã ngược định tính (Q RT-PCR, xem chương 5) cho thấy biểu hiện của SDT1 tăng khi số lượng đoạn lặp tăng từ 0 lên 13. Sau đó biểu hiện của SDT1 giảm
- Phần lớn các dòng không phát triển ở nhiệt độ 32˚C vì gen URA3+ được tái tổ hợp tương đồng thay thế gen URA3-, làm mất gen đột biến nhạy nhiệt. 288
dần khi số lượng trình tự lặp tăng từ 13 lên 60. Có thể kết luận gì về kết quả thí nghiệm này. Giải thích cách dùng Q RT-PCR để phân tích biểu hiện gen SDTl.
c. Kết luận từ phần (b) khiến nhóm nghiên cứu tìm hiểu xem tế bào có thích nghi vói môi trường xung quanh bằng cách biến đổi số lượng trình tự lặp trong ADN vệ tinh trong promoter hay không. Promoter lấy từ gen SDT1 (chứa 48 trình tự lặp) được gắn vào khung đọc mở của URA3, gen chịu trách nhiệm tổng họp uracil. Đặt tế bào mang prôtêin lai này vào môi trường chứa và không chứa uracil. Sau khi sinh trưởng trong mỗi loại môi trường, tiến hành phân tích số lượng các trình tự lặp trong promoter điều khiển biểu hiện URA3. Những tế bào nuôi trong môi trường khuyết ura có số lượng các trình tự lặp trong promoter giảm. Trong khi tế bào nuôi trong môi trường có ura vẫn có 48 trình tự lặp. Có thể kết luận gì về những kết quả này? Dựa vào số liệu ở phần (b), có bao nhiều trình tự lặp ở trong promoter URA3 của tế bào nuôi trong môi trường không có uracil?
d. Vị trí của đoạn lặp ADN trong promoter được so sánh với vị trí nuleosome và thấy rằng mật độ nucleosome trong promoter tỉ lệ thuận với số lượng trình tự lặp tại vị trí ADN đó. Từ đó có thể kết luận gì về trình tự lặp ADN và đóng gói chất nhiễm sắc? Giảm số lượng trình tự lặp ADN có tác dụng gì đến việc histone gắn vào?
Hướng dẫn trả lời
a. - Có thể làm tăng số lượng trình tự lặp bằng cơ chế sao chép trượt trong quá trình tái bản ADN hoặc trao đổi chéo làm xen các trình tự lặp vào vùng ADN vệ tinh.
b. Kết luận: ở số lượng đoạn lặp nhỏ (0 - 13), sự tăng dần số lượng đoạn lặp làm tăng biểu hiện gen. Nhưng ở số lượng đoạn lặp lớn từ 13 trở lên, sự tăng dần số lượng đoạn lặp làm giảm biểu hiện gen.
- Giải thích cách dùng: sử dụng phiên mã ngược để tái tạo ADN từ mARN của gen SDT1 biểu hiện, sau đó dùng PCR để nhân dòng ADN tạo 1 lượng đủ lớn để quan sát, so sánh sự thay đổi trong biểu hiện gen.
c. Kết luận: tế bào có điều chỉnh số lượng trình tự lặp trong ADN vệ tinh trong promoter để thích nghi vói môi trường xung quanh. Trong đó, khi cần tăng biểu hiện gen thì số lượng đoạn lặp trong promoter giảm, khi cần giảm biểu hiện gen thì số lượng đoạn lặp trong promoter tăng.
- Promoter lấy từ gen SDT1 (chứa 48 trình tự lặp) làm biểu hiện của gen URA3 giảm dần vì promoter lấy từ gen SDT1 làm giảm biểu hiện gen khi số lượng đoạn lặp từ 13 trở lên.
- Trong môi trường không có uracil, cần tăng biểu hiện gen nên số lượng đoạn lặp promoter URA3 phải có số đoạn lặp từ 0 – 13.
d. Trình tự lặp ADN tăng làm tăng khả năng đóng gói chất nhiễm sắc. Giảm số lượng trình tự lặp ADN làm ức chế histone gắn vào
1.3. Câu hỏi và bài tập kết hợp vi sinh – Sinh học phân tử - Di truyền
Câu 1.
Ở một vi khuẩn, 3 gen cấu trúc (A, D và E) mã hóa các enzim tương ứng A, D và E. O là operator. Các gen và các trình tự O – A – D – E trên NST. Các enzim này xúc tác con đường sinh tổng hợp Valine. Các đột biến được phân lập trên trình tự/gen này được nghiên cứu để xác định mức tổng hợp các enzim tương ứng khi nồng độ Valine trong tế bào thấp. Các nồng độ enzim được tạo thành ở các thể lưỡng bội một phần mang tổ hợp các đột biến khác nhau được nêu ở bảng dưới đây: Lượng enzim sản xuất Kiểu gen E D A
1. E+D+A+O+/ E+D+A+O+
2. E+D+A+O/
3. E+D-A+O/ E+D+A-O+
4. E+D+A-O/ E+D-A+O+
5. E-D+A+O/ E+D-A+O+
a. prôtêin điều hòa bám vào operator trong operon này là chất ức chế (điều hòa âm tính) hay chất hoạt hóa (điều hòa dương tính)? Giải thích.
b. Operator O có phải là trình tự điều hòa chung của ba gen A, D và E không? Giải thích.
c. Hãy giải thích vì sao ở kiểu gen 3 về nồng độ enzim E đạt thấp?
Hướng dẫn trả lời
a. prôtêin điều hòa là prôtêin ức chế
- Vì dựa vào kiểu gen (KG) 1 khi cả 2 bản sao đều có trình tự O bình thường → lượng enzim A, D, E rất thấp (2,00).
- Mà ở KG 2, có 1 bản sao bị đột biến trình tự O → hàm lượng enzim A, D, E tăng khoảng 20 lần.
b. Trình tự O là trình tự điều hòa chung của cả 3 gen. Vì:
- Ở KG 2: khi có O- ở 1 bản sao có A+, D+, E+ → Nồng độ 3 enzim tăng rất cao.
- Ở KG 4: khi có O- ở 1 bản sao có A-, D+, E+. Bản sao còn lại có O+ và A+
→ Nồng độ chất A bình thường, và chất D, E rất cao.
- KG 5: khi có O- ở 1 bản sao có E-, A+, D+. Bản sao còn lại có O+ và E+ → Nồng độ chất E bình thường , và chất A, D rất cao.
c. Khả năng 1: Sự dịch mã thành công tạo ra sản phẩm đúng chức năng → làm tăng sự dịch mã của gen sau đó trong operon và ngược lại, sự dịch mã tạo ra sản phẩm sai hỏng → giảm sự dịch mã của gen sau đó trong operon.
- VD: gen D bị đột biến → kết thúc dịch mã sớm → tạo ra khoảng cách xa để
Ribôxôm bám vào dịch mã gen E làm giảm sản phẩm enzim E.
- Khả năng 2: ĐB ở trình tự kết thúc dịch mã gen D lại là trình tự để ribôxôm bám vào dịch mã gen E.
Câu 2.
Các trình tự kí hiệu a, b và c có thể là gen mã hóa enzim tổng hợp trp, vùng operator và gen mã hóa chất ức chế. Từ dữ liệu dưới đây, trong đó các kí hiệu (+) hoặc (-) trên các chữ cái biểu diễn tương ứng cho alen kiểu dại hoặc đột biến. Hãy xác định chữ cái nào là kí hiệu cho gen cấu trúc, gen mã hóa chất ức chế và operator (+: enzim tổng hợp tryptophan được biểu hiện; -: không được biểu hiện).
Chủng Kiểu gen Không có Trp Có Trp
1 a-b+c+ + +
2 a+b+c + +
3 a+b-c+ - -
4 a+b+c+/a-b+c - + +
5 a+b+c+/a-bc - + -
6 a+b+c/a-b-c+ + -
7 a-b+c+/a+b-c- + +
Hướng dẫn trả lời
- Ở chủng 3, khi có trình tự b bị ĐB, trình tự a và c bình thường → dù không có trp hay có trp → trp vẫn không được tạo ra → b là gen mã hóa enzim tổng hợp Trp.
- Ở hai chủng 4 và 7 đều có 1 bản sao a+b- và 1 bản sao a-b+ nhưng chúng đều tạo ra Trp ngay cả khi có và không có trp → 2 bản sao ĐB Không bổ sung cho nhau để tạo ra KH bình thường → a là Operator.
- Chủng 6: có 1 bản sao b+c- và 1 bản sao b-c+ → KH Bình thường, 2 bản sao này có thể bổ sung được cho nhau để tạo ra KH bình thường → c là gen ức chế (Mã hóa prôtêin ức chế).
Câu 3.
Thí nghiệm xử lý nấm men tiến hành trên locut tryp4. Đột biến tryp4 làm cho nấm men chỉ sinh trưởng khi được nuôi cấy trên môi trường có bổ sung tryptophan. Alen tryp4-1 là alen đột biến trở lại. Tiến hành thí nghiệm đánh giá tần số đột biến trở lại trong quần thể các tế bào đơn bội. Nuôi cấy tế bào có kiểu giới tính α, kiểu gen tryp4-1 với khoảng 10 triệu tế bào được cấy trên môi trường không chứa tryptophan, người ta thu được 120 khuẩn lạc. Kiểm tra gen của những khuẩn lạc nguyên dưỡng này bằng cách lai mỗi khuẩn lạc với kiểu giới tính a kiểu dại. Từ kết quả của phép lai, các khuẩn lạc nguyên dưỡng trên được chia thành 2 dạng:
Dạng Phép lai Thế hệ con 2/3 dạng 1
α x kiểu dại a Tất cả các thế hệ con đều không phụ thuộc tryptophan
1/3 dạng 2
α x kiểu dại a
721.102 khuẩn lạc thế hệ con không phụ thuộc tryptophan.
138.102 khuẩn lạc thế hệ con phụ thuộc tryptophan.
a. Đột biến nào là đột biến trở lại?
b. tính tần số đột biến trở lại?
Hướng dẫn trả lời
a. Ở dạng 1/3 dạng 2 là dạng đột biến áp chế ( liên quan đến 2 gen- tương tác gen).
Ta thấy tỉ lệ ở đời con là:
138.102 :721.102 =1:3
(Là kết quả của sự tương tác gen khi lai phân tích )
Vì dạng nguyên dưỡng là dạng đột biến nhưng không bị ảnh hưởng bởi đột biến áp chế, còn dạng khuyết dưỡng là dạng có sự áp chế của gen thứ 2
Ta thấy 2/3 dạng 1 cho ra đời con 100% nguyên dưỡng -> Có sự tương tác của 2 gen theo tương tác bổ trợ, đồng thời không có sự áp chế của gen thứ 2
2/3 dạng 1 là đột biến trở lại
b. Tần số đột biến trở lại là:
2 120 3 10000000
× = 8 × 10-6

Câu 4: Oskar, nanos và hunchback là ba gen chính quyết định trục trước sau trong quá trình phát triển phôi ruồi Drosophila. Sơ đồ dưới đây cho thấy mARN và sản phẩm prôtêin của ba gen này phân bố như thế nào trong trứng ruồi (màu càng sẫm thể hiện nồng độ càng cao). Bảng ở dưới liệt kê kết quả phá hỏng một trong các gen này, thông qua phản ánh phân bố prôtêin và phát triển phần trước -sau.
D. Oskar ức chế phiên mã của gen hunchback ở phần đuôi.
E. Prôtêin Hunchback ức chế dịch mã của gen nanos ở phần đầu.
Hướng dẫn trả lời
Nhận xét:
- Đầu bài cho 3 gen Oskar ( qui ước ký hiệu là O), nanos (qui ước kí hiệu là N) và hunchback (qui ước kí hiệu là H) ba gen chính quyết định trục trước sau trong quá trình phát triển phôi ruồi Drosophia.
- Ta thấy 3 gen này phiên mã cho 3 loại mARN và 3 loại prôtêin tương ứng, lượng sản phẩm của 3 gen (cả mARN và prôtêin) được biểu diễn bằng độ đậm nhạt của màu sắc trên các hình ovan, trong đó phía trái thể hiện trục trước và phía tay phải thể hiện trục sau.
→ Khi đột biến N thì sự phân bố của H không bình thường và O phân bố bình thường vậy N ảnh hưởng đến phân bố của H và không ảnh hưởng đến O và thiết lập trục trước sau không bình thường.
Khi đột biến O thì sự phân bố của N và H không bình thường vậy O ảnh hưởng đến phân bố của H và sự thiết lập trục trước sau không bình thường.
Trước Sau
Đột biến ở nanos Đột biến ở hunchback
Đột biến ở oskar
Phân bố của prôtêin Nanos Bình thường Không bình thường
Phân bố của prôtêin Hunchback Không bình thường Không bình thường
Phân bố của prôtêin Oskar Bình thường Bình thường
Thiết lập cực trước –sau bình thường
Không bình thường
Không bình thường Không bình thường
Dựa trên dữ liệu này hãy cho biết khẳng định nào mô tả tương tác giữa ba gen này là đúng?
A. Quá trình dịch mã của gen hunchback bị ức chế bởi prôtêin Nanos
B. Prôtêin Oskar hoạt hóa dịch mã mARN nanos.
C. Hunchback ức chế dịch mã của mARN oskar ở phía đầu.
Khi đột biến H thì sự phân bố của N và O bình thường vậy H không ảnh hưởng đến phân bố của N và O, nhưng thiết lập trục trước sau không bình thường.
A. Sai. Vì khi bình thường prôtêin của gen N ở trục trước ít thì sản phẩm của gen H nhiều và ngược lại prôtêin của N đuôi mà nhiều thì prôtêin H ít. Như vậy có thể khẳng định prôtêin của gen N ức chế dịch mã của H.
Khi đột biến làm hỏng gen, không có prôtêin N thì H phân bố như bình thường nghĩa là gen N không ảnh hưởng đến hoạt động của gen H.
B. Đúng. Khi bình thường prôtêin O ít ở trục trước thì prôtêin N cũng ít ở trục trước và prôtêin O nhiều ở trục sau thì prôtêin N cũng nhiều ở trục sau. Như vậy có thể rút ra giả thuyết prôtêin O hoạt hóa dịch mã mARN của gen N.
Khi đột biến làm hỏng gen, không có prôtêin O thì prôtêin N phân bố không bình thường nghĩa là gen O ảnh hưởng đến hoạt động của gen N theo hướng hoạt hóa mARN của gen N.
C. Sai. Khi bình thường prôtêin H nhiều ở trục trước thì prôtêin O lại ít ở trục trước và prôtêin H ít ở trục sau thì prôtêin O lại nhiều ở trục sau. Như vậy có thể rút ra giả thuyết prôtêin H ức chế dịch mã mARN của gen O ở phần đầu.
Khi đột biến làm hỏng gen, không có prôtêin H thì prôtêin O phân bố bình thường là gen H ảnh hưởng đến hoạt động của gen O.
D. Sai. Khi bình thường gen H luôn được phiên mã kể cả khi nhiều hay ít prôtêin O( độ đậm biểu diễn mARN trên hình vẽ không thay đổi). Như vậy có thể kết luận quá trình phiên mã của gen H không hề phụ thuộc vào gen O.
Khi đột biến làm hỏng gen, không có prôtêin O thì prôtêin H phân bố không bình thường nên prôtêin O chỉ ảnh hưởng tới dịch mã mà không ảnh hưởng tới phiên mã.
E. Sai. Khi bình thường prôtêin H nhiều ở trục trước thì prôtêin N lại ít ở trục trước. Như vậy có thể rút ra giả thuyết prôtêin H ức chế dịch mã mARN của gen N ở phần đầu.

Khi đột biến làm hỏng gen, không có prôtêin H thì prôtêin N phân bố bình thường nghĩa là gen H ảnh hưởng đến hoạt động của gen N.
Câu 5.
Một nhà khoa học có 10 chủng vi khuẩn Escherichia coli mang các đột biến khác nhau nằm trên operon lac của các vi khuẩn này. Khi phân tích ADN của vi khuẩn, nhà khoa học này thấy rằng mỗi chủng mang một trong 5 đột biến: lacZ-, lacY, lacI-, lacIs (lacIs mã hóa LacIs là chất ức chế có thể bám vào operator nhưng không thể bám vào chất cảm ứng), hoặc lacOc (Operator của operon lac đột biến làm chất ức chế không thể bám vào nó). Biết rằng chủng số 6 là một thể đột biến lacZ-. Cấu trúc operon lac được mô tả như hình dưới đây.
Hãy cho biết câu sau đây là Đúng hay Sai? Giải thích
A. Chủng số 7 là thể đột biến lac Z.
B. Chủng số 3 là thể đột biến lac Y.
C. Chủng số 2 và số 4 mang cùng loại đột biến.
D. Nếu chủng số 5 nhận được ADN của chính nó thì thể biến nạp này không thể sinh trưởng được.
Hướng dẫn trả lời
- Các đột biến ở chủng 5, 9 nằm trên cùng 1 gen; 1 và 8 nằm trên cùng 1 gen; 2, 4, 6, 7 nằm cùng 1 gen và 10, 3 nằm cùng 1 gen ( các đột biến không bổ trợ được cho nhau → xảy ra trên cùng 1 gen ). A, B, C đúng:

- Mỗi chủng mang 1 loại đột biến
- Có : LacIs: vi khuẩn không sinh trưởng
- Đột biến LacOc: vi khuẩn luôn sinh trưởng
+) LacI-/LacI-: Sinh trưởng bình thường
+) LacZ-/LacZ-: Không sinh trưởng
Nhà khoa học phân lập được đoạn ADN mang operon lac từ mỗi chủng (gọi là chủng cho) và biến nạp đoạn ADN này vào các chủng khác, tạo ra chủng lưỡng bội từng phần (gọi là chủng nhận). Sau đó, các chủng nhận được nuôi trên môi trường tối thiểu chứa lactose là nguồn carbon duy nhất . Sự sinh trưởng của các chủng được ghi lại trong ảnh dưới đây (+ biểu thị chủng đang sinh trưởng, - biểu thị chủng không sinh trưởng được).
+) lac Y-/LacY-: Không sinh trưởng
- Nếu ở thể dị hợp: LacZ LacZ- và LacY LacY- sinh trưởng bình thường.
- Chủng 6 đột biến lac Z
- Chủng 7 có tác động giống hệt chủng 6, khi biến nạp với chủng 6 → không sinh trưởng → chủng 7 là đột biến LacZ-
- Chủng 5 và chủng 9 có tác động giống hệt nhau, dù biến nạp với chủng nào vi khuẩn vẫn luôn sinh trưởng → chủng 5, 9 đột biến LacOc
- Chủng 1 và chủng 8 có tác động giống hệt nhau biến nạp với các chủng khác thì đều không sinh trưởng ( trừ biến nạp với chủng 5 và 9) → chủng 1 và 8 đột biến LacIs
- Chủng 2 và chủng 4 có tác động giống hệt nhau, biến nạp với các chủng khác đều sinh trưởng được trừ biến nạp với chủng 1 và 8. Và khi biến nạp chủng 2 với chủng 4 đều sinh trưởng được → Chủng 2 và 4 đột biến LacI-
- Chủng 3, 10 có tác động giống nhau, mà khi biến nạp 3 với 10 thì không sinh trưởng → 3 với 10 đột biến LacY-
Câu 6.
Một operon mã hóa enzim 1 và 2 được điều hòa bởi sự chuyển hóa chất X chứa đựng 7 trình tự A, B, C, D, E, F, G chưa biết rõ chức năng. Để làm sáng tỏ chức năng của chúng, sự ảnh hưởng của đột biến ở các trình tự từ A đến G dựa trên sự tổng hợp các enzim được đánh giá thông qua sự có mặt và sự vắng mặt của X.
Có mặt X Vắng mặt X
Đột biến trong trình tự Enzi m1 Enzi m 2 Enzi m1 Enzim2
Không có độ
Hãy xác định vai trò của operon và các trình tự của A, B, C, D, E, F, và G. Giải thích.
Hướng dẫn trả lời
- Operon khi có mặt X thì cả hai enzim đều không được tổng hợp do đó chứng tỏ đây là operon đồng hóa (operon ức chế)
- Thể đột biến B, F biểu hiện giống nhau. Khi có hay không có mặt X thì operon luôn được mở ( luôn tạo ra enzim)
- Bị đột biến vùng O do đó prôtêin ức chế không liên kết được với O ( Lac Oc)
- Thể đột biến D khi có mặt hay không có mặt X thì operon luôn đóng
- Bị đột biến vùng P (ARN pol không liên kết vào P được -> phiên mã không diễn ra, operon luôn đóng)
- Thể đột biến A, E biểu hiện giống nhau khi vắng mặt X chỉ có enzim 2 được tổng hợp.
- Bị đột biến gen tổng hợp enzim 1 → enzim 1 không được tổng hợp
- Thể đột biến C khi vắng mặt X chỉ có enzim 1 được tổng hợp
- Bị đột biến gen tổng hợp enzim 2 → enzim 2 không được tổng hợp
- Thể đột biến G biểu hiện giống kiểu dại → Trình tự không liên quan đến operon mã hóa enzim 1, enzim 2
Câu 7.
Hiện tại, bạn đang nghiên cứu về sự tổng hợp threonin ở E. coli. Để tìm các gen mã hóa các prôtêin quan trọng cho tổng hợp threonin, bạn tiến hành gây đột biến để tìm các đột biến yêu cầu bổ sung threonin cho phát triển. Bạn đã tìm thấy có 9 dòng đột biến và tất cả đều cần bổ sung threonin vào môi trường nuôi cấy để phát triển. Sau đó bạn tiến hành cis-trans test với tất cả các dòng đột biến bạn có. Kết quả thu được như bảng sau (+: phát triển; -: không phát triển):
M t1 M t2 M t3 M t4 M t5 M t6 M t7 M t8 M t9 w t
a. Có bao nhiêu gen mã hóa các enzim tham gia vào quá trình tổng hợp threonin? Giải thích.
b. Khi xem xét quá trình tổng hợp threonin, người ta nhận thấy có 5 cơ chất tham gia vào quá trình đó. Người ta tiến hành nuôi cấy các đột biến ở trên trong các môi trường chứa các cơ chất khác nhau để xác định trình tự quá trình biến đổi. Kết quả thu được trình bày ở bảng sau (+: phát triển; -: không phát triển):
A B C D E Thr eonine
Mt 2 -
Mt 4 -
Mt 7 -
Hãy xác định trật tự chuyển hóa của các chất A, B, C, D, E và các gen mã hóa các enzym xúc tác sự chuyển hóa tương ứng. Giải thích.
Hướng dẫn trả lời
a) Ta thấy:
- Khi cho các dòng đột biến giống nhau lai với nhau
→ Không phát triển, không tạo khuẩn lạc.
→ Đột biến trên cùng một gen nên không bù trừ được cho nhau.
→ Tạo kiểu hình đột biến.
- Khi cho những dòng đột biến khác nhau lai với nhau:
+ Nếu khuẩn lạc phát triển
→ Hai đột biến thuộc hai gen khác nhau bù trừ cho nhau tạo kiểu gen dị hợp
→ Tạo kiểu hình bình thường (kiểu dại).
+ Nếu khuẩn lạc không phát triển
→ Tạo kiểu gen đồng hợp
→ Tạo kiểu hình đột biến.
Từ đó, ta có:
- 1,3,5,6,9 thuộc cùng một gen.
- 2,8 thuộc cùng một gen.
- 4 không bù cho mình, bù cho những dòng đột biến còn lại → gen độc lập
- 7 không bù cho mình, bù cho những dòng đột biến còn lại → gen độc lập
→ Có 4 gen tham gia tổng hợp threonin.
b) sơ đồ chuyển hóa:
A ___Mt1___ D___Mt7__ C___Mt4__ B,E___Mt2___ Threonin
Giải thích
- Các chủng đột biến không sinh trưởng được trong môi trường chỉ chứa A → A là chất đầu dãy chuyển hóa
- Chỉ có chủng đột biến Mt1 sinh trưởng được trong môi trường chỉ chứa D → D là chất thứ 2 của dãy chuyển hóa và chủng Mt1 bị đột biến ở bước chuyển hóa chất A thành chất D
-Trong môi trường chỉ chứa C, có hai chủng Mt1 và Mt7 sống được → chất C là chất thứ 3 của dãy chuyển hóa và chủng Mt7 là chủng bị đột biến ở bước chuyển hóa chất D thành chất C
- Chủng Mt4 sống được trong môi trường chỉ chứa B và E → chất B và E cùng là chất thứ 4 trong dãy chuyển hóa và chủng Mt2 bị đột biến ở bước chuyển hóa chất C thành chất B hoặc E
- Chủng Mt2 chỉ sống được trong môi trường chứa Threonin → chủng này bị đột biến ở bước chuyển hóa chất B hoặc chất E thành Threonin.
Câu 8.
Sắc tố đỏ ở một loài nấm đơn bội được tạo ra bởi một con đường chuyển hóa từ một sắc tố tiền thân qua một số sản phẩm trung gian. Để tìm hiểu con đường chuyển hóa này, người ta thu thập một số chủng đột biến (kí hiệu từ I đến IV). Bảng dưới liệt kê đặc điểm màu sắc của chúng và các con lai đơn bội khi tiến hành lai giữa chúng với nhau.
Chủng hoặc phép lai Màu quan sát thấy
→ Hai đột biến thuộc cùng một gen nên không bù trừ cho nhau 300
Kiểu dại X I X II X III X IV X
I x Kiểu dại X X X
II x III X X X X
II x IV X X
III x IV X X X
Hãy cho biết phát biểu nào dưới đây là đúng hay sai.
A. Ít nhất có 4 gen liên quan đến con đường chuyển hóa này.
B. Chủng I có đột biến ở nhiều hơn một gen liên quan đến con đường chuyển hóanày.
C. Trong con đường chuyển hóa này, enzim chuyển hóa tạo sắc tố hồng ở bước phía trước so với enzim chuyển hóa tạo sắc tố màu be.
D. Cá thể con màu đỏ có thể xuất hiện trong phép lai giữa chủng I vàIV.
Hướng dẫn trả lời
A. Sai B. Đúng C. Sai D. Đúng
Phân tích:
Phép lai I (mau be) x Kiểu dại (đỏ) sinh con đỏ, be, vàng. Phép lai xuất hiện vàng cũng có nghĩa là enzim chuyển hóa tạo sắc tố vàng ở bước phía sau con đường chuyển hóa so với enzim chuyển hóa sắc tố màu be
Phép lai II (be) x (vàng) xuất hiện hồng chứng tỏ là enzim chuyển hóa tạo sắc tố hông ở bước phía sau con đường chuyển hóa so với enzim chuyển hóa tạo sắc tố màu be
A. Sai. Trong con đường này, 3 bước xúc tác bởi tổng cộng 3 enzim là đủ để giải thích kết quả
B. Đúng. I (be) x kiểu tự nhiên, con có be và đỏ (kiểu dại) và con màu vàng mà chỉ có thể giai thích đượng nếu có đột biến cho cả enzim chuyển hóa trung gian màu be và chuyển đổi màu vàng. PL xuất hiện vàng cũng có nghĩa là enzim chuyển hóa tạo sắc tố vàng ở bước phía sau con đường chuyển hóa so với enzim chuyển hóa sắc tố màu be
C. Sai. II (be) x IV (hồng) không tạo ra màu be. Vì chúng ta biết từ B rằng enzim chuyển hóa sắc tố be ở bước phía trước con đường chuyển hóa so với enzim chuyển hóa sắc tố màu vàng, điều nay chỉ có thể được giải thích IV chỉ là đột biến cho gen chuyển hóa trung gian màu hồng, do đó chủng II là đột biến cho cacr hai chuyển hóa enzim mau be và một chuyển hóa màu hồng. Ở PL 3 xuất hiện màu be và không phải màu hồng, màu be ở phía trước con đường chuyển hóa tạo màu hồng
D. Đúng. Con cái có màu đỏ nếu giữa 2 bố mẹ mang ít nhất một bản sao của một loại gen kiểu dại. Chúng ta biết từ B rằng là đột biến duy nhất cho các enzim chuyển đổi màu vàng và màu be và từ C mà IV chỉ là đột biến enzim chuyển đổi do đó màu đỏ (hoang dã)
Câu 9.
Phân lập được nhiều thể đột biến khuyết dưỡng, phân tích cá thể đột biến, thấy sinh trưởng trên môi trường tối thiểu có bổ sung các hợp chất A, B, C, D. Bảng kết quả thể đột biến ở môi trường có bổ sung các chất tương ứng, cho biết A, B, C, D thuộc cùng một con đường hóa sinh.
Thể đột biến A B C
a. Thứ tự các chất này trong con đường chuyển hóa và cho biết các thể đột biến đó bị ảnh hưởng bởi đột biến gen nào?
b. Đột biến nào cùng cặp đột biến nào khác cặp?
Hướng dẫn trả lời
a. - Trong dãy chuyển hóa các bước chuyển hóa càng ở phía sau thì thể đột biến tương ứng càng cần ít chất bổ sung thêm vào môi trường chọn lọc. Ngược lại thì các chất càng ở phía trước của con đường chuyển hóa thì càng có ít chủng đột biến có thể sinh trường. Ở đây chủng đột biến 2 và 4 tương ứng với chất D cần ít chất hơn cả vào môi trường tối thiểu → 2 chủng này bị sai hỏng ở enzim chuyển hóa tiền chất trước các chất A,B,C và D
Tất cả các thể đột biến khi bổ sung chất D đều sinh trường → D là chất cuối cùng trong dãy chuyển hóa
Xét thể đột biến 1: Khi bổ sung chất A,B, D thì sinh trưởng, còn khi bổ sung
chất C thì không sinh trường → do vậy chủng đột biến này bị sai hỏng ở enzim chuyển hóa chất C sang các chất A, B hoặc D → Chất C nằm ở đầu dãy chuyển hóa
Xét thể đột biến 6: Chủng này chỉ sinh trưởng được trên môi trường bổ sung chất B và chất D (trong đó D là chất đứng cuối dãy) → chủng này bị hỏng enzim chuyển hóa từ chất A, C sang B, còn enzim chuyển hóa B sang D vẫn hoạt động bình thường → B nằm ở ngay trước D trong dãy chuyển hóa
Chất A đứng ở giữa C và B trong dãy chuyển hóa
Như vậy ta thu được dãy chuyển hóa sau
Tiền chất -----(2)--- → C -----(1)--- →A----(6)------ → B------(3)----- →D
b, - Các đột biến cùng cặp là các đột biến có các sai hỏng trong cùng 1 enzim chuyển hóa. Từ ý a → các đột biến cùng cặp gồm có: 1 với 7, 2 với 4, 3 với 5 và 6 với 8. Các đột biến khác không phải các cặp trên đều là các đột biến khác cặp.
Câu 10.
Một operon mã hóa 2 enzim 1 và 2 được điều hòa bởi chất chuyển hóa X và chứa 4 đoạn trình tự A, B, C và D với chức năng chưa rõ. Để làm rõ chức năng của những trình tự này, ảnh hưởng của các đột biến trong trình tự các phân đoạn A - D đến sự tổng hợp các enzim được đánh giá khi có mặt và vắng mặt chất X
Thể đột biến Có X Không có X
Đột biến trong trình tự En zim 1 Enz im 2 Enz im 3 Enzi m 4
Không có đột biến + + + + A - -
Phát biểu nào dưới đây là đúng hay sai. Giải thích ?
A. Các enzim 1 và 2 nhiều khả năng tham gia vào con đường tổng hợp X.
B. Đoạn trình tự A mã hóa cho enzim 2.
C. Đoạn trình tự B là vùng promoter.
D. Đoạn trình tự D là một gen điều hòa.
Hướng dẫn trả lời
A. sai. Khi có mặt của X thì gen mã hóa enzume 1,2 đều đượ
1,2 tham gia vào chuyển hóa X giúp tế bào khai thác X làm nguồn dinh dưỡng, không phải tham gia con đường tổng họp chất X
B. sai . khi có đột biến ở A thì cả enzim 1,2 đều không biểu hiện → A ảnh hưởng tới khả năng biểu hiện của cả 1,2 → A có thể là promoter hoặc gen quy định prôtêin hoạt hóa điều khiển hoạt động của operon chứa 1,2
C. sai – B bị đột biến thì cả enzim 1,2 vẫn biểu hiện bình thường → B không phải Promoter ( nếu B promoter thì khi bị đột biến thì 1,2 không biểu hiện)
D. sai. Gen điều hòa quy định pr điều hòa lirn kết với vùng operator khiến AND pol không thể liên kết vào promoter→ ức chế operon biểu hiện → khi gen điều hòa bị đột biến không tạo ra pr điều hòa có chức năng, operon luôn trong trạng thái mở→ Biểu hiện liên tục
Khi D bị đột biến cả hai enzim 1,2đều không biểu hiện → D không phải gen điều hòa
Câu 11.
Các trình tự ADN đa hình được sử dụng rộng rãi trong việc nhận dạng cá thể bằng chỉ thị phân tử. STR (short tandem repeat) bao gồm nhiều trình tự lặp lại của 28 nucleotit và hai đầu đoạn lặp là hai trình tự bảo thủ. Mỗi locut STR thông thường có nhiều hơn hai alen. Đa hình đơn nucleotit (Single nucleotit polymorphism - SNP) là một biến đổi ở một nucleotit trên trình tự ADN giữa các cá thể. Mỗi SNP thường chỉ có 2 alen. Bảy cá thể được xác định kiểu gen ở 2 SNP trên nhiễm sắc thể thường và 2 SNP trên ADN ti thể (mtADN); 2 STR trên nhiễm sắc thể thường và 2 STR liên kết nhiễm sắc thể Y (NRY). Kết quả thu được thể hiện ở bảng sau: Nhiễm sắc thể thường NRY mtADN S NP1 S NP2 S TR1
TR1
nd_6 /C /G 4/14 8/19
I nd_7 C /C G /G 1 4/16 1 9/21 - -
Hãy cho biết câu nào sau đây là đúng hay sai?
A. Nếu sử dụng số lượng các SNP tương đương với số lượng STR, thì phân biệt các cá thể bằng SNP tốt hơn so với STR.
B. Khả năng Ind_6 là con của Ind_2 và Ind_5 là cao hơn so với Ind_3.
C. Ind_4 có thể là anh trai của Ind_6.
D. Ind_7 có thể là cháu gái của ông bà Ind_1 và Ind_6.
Hướng dẫn trả lời
A. Sai: Vì STR có nhiều hơn 2 alen, còn SNP chỉ có 2 alen. Nên khi sử dụng số lượng các SNP tương đương với số lượng STR thì phân biệt các cá thể bằng STR tốt hơn SNP.
B- Sai: Vì Ind_3 có khả năng là con của Ind_2 và Ind_5 cao hơn so với Ind_6 do:
Xét mtADN (ADN ti thể) thì Ind_3 có SNP1 và SNP2 giống hệt Ind_5; còn Ind_6 có SNP1 và SNP2 không giống Ind_2 hoặc Ind_5. Do vậy Ind_3 có khả năng là con của Ind_2 và Ind_5 cao hơn Ind_6.
C-Đúng: Vì mtADN (ADN ti thể) của 2 cá thể Ind_4 và Ind_6 là giống nhau (T, G); các SNP và STR cũng gần giống nhau. Nên khả năng Ind_4 có thể là anh của Ind_6.
D- Đúng:
- Trên mtADN của Ind_7 giống Ind_1 nên Ind_1 có thể là bà ngoại của Ind_7.
- Trên NST thường thì con của Ind_1 giống Ind_6 có:
+ SNP2 là A/G; Ind_7 có SNP2 là G/G.
+ STR1 là 14/15 hoặc 14/13; Ind_7 có STR1 là 14/16.
→ Do vậy Ind_7 có thể là cháu gái của ông bà Ind_1 và Ind_6.
Câu 12.
Một nhà khoa học nhân dòng 1 trình tự mã hóa (coding sequence-CDS)

của một gen vào một vectơ rồi đặt tên plasmit tạo thành là pVN2016. CDS được chèn tại vị trí nhận biết SacII nằm trong vùng MSC (multi cloning site) trong vùng gen lacZ của vectơ (Hình A). CDS được chèn có một vị trí cắt giới hạn Pstl cách bộ ba kết thúc của nó 0,8 kb. Để xác định kích thước và hướng của CDS đã được chèn nhà khoa học đã cắt plasmit này với enzim cắt giới hạn khác nhau, và sản phẩm cắt được trình bày trong hình B.
Dựa vào dữ liệu ở trên, hãy cho biết câu nào sau đây là đúng hay sai:
A. CDS dài 2.6 kb và có vị trí nhận biết EcoRI ở điểm cách một đầu tận cùng của nó khoảng 0,5 kb.
B. SpeI có thể được sử dụng để xác định hướng của CDS.
C. CDS được chèn vào cùng chiều với lacZ.
D. Nếu plasmit pVN2016 bị cắt bởi cả hai enzim SpeI và EcoRI trong đệm Tango (EcoRI và SpeI cắt ở mức tương ứng là 100% và 20%), thì 5 đoạn với kích thước 0,5; 0,8; 1,3; 2,1 và 3,0 kb có thể được xác định bởi điện di trên gel, giả sử không quan sát được các đoạn nhỏ hơn 50 bp.
Hướng dẫn trả lời
A- Đúng:
- pVN2016 (ADN tái tổ hợp) có chiều dài là 56 kb (tổng của các băng 4.3+ 1.3); còn vectơ có chiều dài 3 kb nên CDS có chiều dài là 2,6 kb.
- Khi dùng enzim Sac II trong trong kỹ thuật này thì tạo vectơ vòng hở có 2 vị trí nhận biết của EcoRI ở 2 đầu → cắt bằng EcoRI sẽ tạo ra 1 bản 3kb (của vectơ); mà còn có 2 băng 2,1kb và 0,5kb → Đây là 2 băng tạo ra từ CDS → có vị trí nhận biết EcoRI cách đầu tận cùng của CDS 1 đoạn 0,5kb.
B- Sai:
- Vì SpeI không thể được sử dụng để xác định hướng của CDS do chưa biết được vị trí cắt của SpeI trên CDS.
C- Đúng.
D- Đúng:
- Dựa trên bảng Q93B ta có thể xác định được bản đồ cắt giới hạn.
- Khi cắt bằng EcoRI (100%) và Spe I (20%) có thể thu được các băng:
+ 0,5kb; 2,1kb; 3kb (khi chỉ EcoRI hoạt động).
+ 0,5kb; 0,8kb; 1,3kb; 3kb (khi cả 2 enzim cùng cắt).

Câu 13.
Frederick Sanger (1918-2013) đã phát minh ra kĩ thuật giải trình tự ADN, ARN và prôtêin; Shankar Balasubramanian (1966-đến nay) đã phát minh ra kĩ thuật giải trình tự ADN hiệu năng cao. Trung tâm Chăm sóc Sức khỏe Quốc gia hiện đang giải trình tự 100.000 hệ gen từ các bệnh nhân mắc bệnh hiếm gặp, nhưng các công nghệ giải trình tự khác nhau đều có những giá trị riêng, như được mô tả dưới đây:
Công nghệ Độ dài tối đa của đoạn trình tự có thể đọc được
Tần số đọc lỗi
Tổng số bazo được giải/ mẫu/ngày
Giải trình tự Sanger 900 bp 1/1 000 bp 900 bp (1 phân đoạn)
Máy giải trình tự Illumina 200 bp 1/1 00 bp 3x 1011 bp (>1.5x 109 phân đoạn)
Máy giải trình tự PacificBiosciences 5000 bp 1/1 0 bp 4x 108 bp (>80000 phân đoạn)
Mỗi phát biểu dưới đây về dự án 100.000 hệ gen là đúng hay sai?
A. Công nghệ Illumina là phù hợp nhất để tìm ra những biến đổi đơn nucleotit (các đột biến ở một bazơ đơn lẻ) trong các hệ gen của bệnh nhân.
B. Công nghệ PacificBiosciences là phù hợp nhất để đánh giá những biến đổi trong phiên mã bằng việc giải trình tự ARN.
C. Công nghệ PacificBiosciences là phù hợp nhất để phát hiện sự tái sắp xếp của các đoạn ADN trong các hệ gen bệnh nhân.
D. Giải trình tự Sanger là phù hợp nhất để kiểm định (kiểm tra lại) các kết quả giải trình tự, trước khi dùng thông tin di truyền của bệnh nhân để chỉ định can thiệp y tế
Hướng dẫn trả lời
A- Đúng:
- Vì giải trình tự Sanger tuy tần số đọc lỗi là 1/1/1000 bp nhưng chỉ giải được 1 phân đoạn/1 ngày nên để giải toàn bộ hệ gen và phát hiện biến đổi đơn nucleotit (các
đột biến ở một bazơ đơn lẻ) trong các hệ gen của bệnh nhân sẽ rất tốn thời gian →Không sử dụng.
- Máy giải trình tự PacificBiosciences tuy đọc được 80000 phân đoạn/ ngày nhưng tần số đọc lỗi là 1/10bp nên cũng không phù hợp để phát hiện những biến đổi đơn Nucleotit.
- Giải trình tự Illumina có tần số đọc lỗi là 1/100bp, có thể đọc 1.5x 109 phân đoạn/ngày là phù hợp nhất để phát hiện những biến đổi đơn Nucleotit trong hệ gen của bệnh nhân.
B- Sai:
- Vì công nghệ PacificBiosciences có tần số đọc lỗi là 1/10bp không phù hợp cho việc đánh giá những biến đổi trong quá trình phiên mã bằng dịch mã ARN ( công nghệ phù hợp nhất có thể là Sanger).
C- Đúng:
- Vì công nghệ PacificBiosciences có thể giải trình tự tối đa 1 đoạn dài 5000bp → phù hợp nhất để phát hiện sự tái sắp xếp của ADN trong hệ gen bệnh nhân.
D- Đúng:
- Vì Sanger có tần số lỗi là 1/1000→ thường được dùng để kiểm tra lại một cách chính xác nhất kết quả của việc giải trình tự sau đó mới tiến hành can thiệp y học.
Câu 14.
Một plasmit được biến nạp vào vi khuẩn (1). prôtêin huỳnh quang đỏ (RFP) hoặc prôtêin huỳnh quang xanh (GFP) được biểu hiện từ các gen trên plasmit này khi các nhân tố phiên mã bám vào các promoter. Các trình tự promoter của gen đỏ và xanh là giống nhau. Sự sao chép ADN plasmit bắt đầu ở vị trí khởi đầu sao chép và chạc sao chép di chuyển theo cả hai hướng với tốc độ bằng nhau. Sao chép ADN xảy ra khi các nhân tố sao chép bám vào vị trí khởi đầu sao chép. Cường độ huỳnh quang đỏ và xanh của mỗi tế bào vi khuẩn được đo (2).
Mỗi phát biểu dưới đây là đúng hay sai?
A. Các plasmit đang sao chép có số lượng bản sao gen mã hóa prôtêin huỳnh quang đỏ không bằng với số lượng bản sao gen mã hóa prôtêin huỳnh quang xanh.
B. Tính trung bình, các tế bào ở Vùng A chứa nhiều nhân tố sao chép hơn so với tế bào ở vùng B.
C. Tính trung bình, các tế bào ở vùng A chứa nhiều nhân tố phiên mã hơn các tế bào ở vùng B.
D. Sự tăng hoạt tính của các nhân tố phiên mã được mong đợi làm tăng số lượng các tế bào chứa hàm lượng prôtêin huỳnh quang xanh tương đương (bằng) lượng prôtêin huỳnh quang đỏ.
Hướng dẫn trả lời
A- Sai:
- Vì: Hai gen có chiều dài tương đương nhau, đều cách vị trí khởi đầu sao chép 100bp; chạc sao chép di chuyển về 2 phía với vận tốc như nhau → Các plasmit dạng sao chép có số lượng bản sao GFP và RFP bằng nhau.
B- Sai:
- Không xác định được nồng độ nhân tố sao chép trong các tế bào nếu dựa vào cường độ phát huỳnh quang. Vì nhân tố sao chép không ảnh hưởng lớn đến nồng độ prôtêin tạo ra mà nó phụ thuộc vào nhân tố phiên mã.
C- Sai:
- Vùng A có ít nhân tố phiên mã hơn vùng B vì ở vùng A có cường độ phát huỳnh quang thấp hơn vùng B.
D- Đúng:
- Vì Promoter của 2 gen GFP và RFP giống nhau nên gen được hoạt hóa từ cùng một yếu tố phiên mã → sự tăng hoạt tính của yếu tố phiên mã sẽ làm tăng đồng thời mức độ biểu hiện của gen GFP và RFP → hàm lượng prôtêin xanh và đỏ tạo ra là tương đương nhau.
Câu 15.
Michael Smith (1932-2000) đã phát minh kĩ thuật gây đột biến hướng đích. Gần đây các nhà khoa học đã phát minh công nghệ CRISPR-Cas9 để tiến hành gây đột biến dễ dàng hơn. Prôtêin Cas9 từ Streptococcus pyogens được dẫn bởi một guideARN (ARN chỉ dẫn, gARN) dài 20 bp liên kết với ADN đích. Enzim Cas9 này chỉ có thể cắt sợi kép ADN tại vị trí cách ba nucleotit nằm ngược dòng trình tự 5'-NGG-3' (1). Các phân đoạn ADN sợi kép đã bị cắt bởi các enzim này, và sau đó vẫn có thể được nối trở lại. Gen X cần được làm mất chức năng nhờ enzim Cas9 được mô tả ở đây (2). Một cách khác để knockout gen là sử dụng một cặp gồm Cas9 cải biến có khả năng chỉ tạo ra các vết cắt sợi đơn và ARN chỉ dẫn (guideARN) hướng đích tới các vị trí liền kề.
1. Chọn một trong 5 trình tự guide ARN sẽ chỉ dẫn Cas 9 tới vị trí I?
2. Chọn vị trí NGG có sẵn tốt nhất trên gen đích X để làm mất chức năng của nó?
3. Mỗi phát biểu dưới đây là đúng hay sai?
A. Sử dụng cặp đôi Cas9 và guide ARN bị biến đổi làm tăng các tổn thương sai đích ở các gen khác.

B. Hệ gen Streptococcus pyogens ngẫu nhiên có nhiều vị trí NGG hơn mong đợi.
C. Nếu gen không có các trình tự GG, các prôtêin Cas9 từ các loài khác cần được nghiên cứu.
Hướng dẫn trả lời
1. Trình tự guide ARN sẽ chỉ dẫn Cas 9 tới vị trí I là trình tự 2= 5'AUUU....CCA-3' vì:

- Cas 9 cải biến chỉ có khả năng cắt sợi đơn →ARN chỉ dẫn phải liên kết vào vị trí cắt của Cas 9 để tạo sợi đơn.
- Vị trí cắt cách AGG (I) ở 3 Nucleotit ngược dòng tức sẽ cắt giữa 5'-TAG-3' và 5'GCC-3'.
→ Đoạn ARN chỉ dẫn có trình tự ít nhất là 3'-GAUUU... →có 2 đoạn ARN hợp lý
nhất là 2 và 4 tuy nhiên 4 không hợp lí vì không giúp nhận biết NGG
2. Chọn vị trí NGG có sẵn tốt nhất trên gen đích X để làm mất chức năng của nó là vị trí số II.

3. Chọn
- A- Sai: Vì Cas 9 biến đổi chỉ có khả năng cắt mạch đơn nên chỉ có thể cắt gen đích đã gắn ARN, khác với Cas 9 chưa biến đổi có thể cắt ADN mạch kép → Cas9 và guideARN bị biến đổi làm tăng các tổn thương sai đích.

- B- Sai: Vì nếu có nhiều vị trí NGG thì gen của Streptococcus pyogens sẽ bị tổn thương do hoạt động của Cas 9 và ARN chỉ dẫn.
- C- Đúng: Nếu gen không có trình tự GG → Cas9 giống liên cầu khuẩn không hoạt động để nhận biết cắt nên không cắt được gen → Cần nghiên cứu Cas 9 của loài khác cần được nghiên cứu để tìm ra cơ chế khác.
Câu 16.
Một số enzim giới hạn có các trình tự nhận biết khác nhau nhưng lại tạo ra trình tự đầu dính giống nhau, như SalI và XhoI dưới đây.
Mỗi phát biểu dưới đây là đúng hay sai?
A. Dữ liệu cho thấy đã có 2 bản sao của đoạn chèn được cài vào vectơ biểu hiện.
B. Một nửa số tế bào được nhân dòng được mong đợi là có thể phiên mã mARN của gen x.
C. Một vị trí cắt của XhoI nằm bên ngoài đoạn chèn trong vectơ tái tổ hợp ở thí nghiệm này.
Ảnh điện di dưới đây cho thấy các ADN dạng thẳng nhận được sau khi cắt (mở vòng bằng enzym giới hạn) hoàn toàn các vectơ plasmit biểu hiện không tái tổ hợp (vectơ trống) và tái tổ hợp (vectơ mang gen x). Vectơ biểu hiện chứa một promoter mạnh nằm gần vị trí tách dòng. Đoạn chèn của plasmit tái tổ hợp là sản phẩm cắt của phản ứng với enzim giới hạn SalI. Đoạn chèn được nối vào vectơ đã bị cắt bởi enzim XhoI.
D. Phân đoạn 2 kb được quan sát trên gel được dùng làm mẫu dò để sàng lọc các vectơ tái tổ hợp.
E. Nếu đoạn chèn được tạo thành bởi phản ứng cắt bằng SalI được nối vào một vectơ bị cắt đồng thời bởi cả XhoI và SalI, rồi plasmit tái tổ hợp sau đó bị cắt bởi XhoI, thì sản phẩm thu được có đoạn dài 4 kb.
Hướng dẫn trả lời
A- Sai:
- Vì vectơ biểu hiện chỉ nhận được một bản sao có chiều dài 2 kb.
B- Đúng:
- Vì xác suất gen X cài đúng theo chiều của promoter là 50% nên sẽ có một nửa số tế bào nhân dòng được biểu hiện.
C- Sai:
- Vị trí cắt của XhoI sẽ nằm trong đoạn chèn vì nếu nằm ngoài thì khi cắt plasmit bằng XhoI sẽ làm đứt gãy plasmit và sẽ không có chiều dài 5 kb.
D- Sai:
- Theo bảng ta có thể xác định 1 vị trí cắt của XhoI nằm trong gen X, 1 vị trí cắt của SalI là trong plasmit → khi cắt bằng 2 enzim thì đoạn 2 kb thu được chứa cả gen X và plasmit → không dùng để làm mẫu dò.
E- Sai:
- Plasmit chỉ có vị trí cắt của XhoI →khi thực hiện như vậy vẫn chỉ thu được sản phẩm như ban đầu là 1 băng 7 kb.
Câu 17.
Một trong những vấn đề nảy sinh liên quan đến cơ chế cắt bỏ intron, nối exon là trật tự theo thời gian của việc loại bỏ các intron trong bản mã sao mARN của gen mang nhiều intron có giống với trật tự sắp xếp của các intron hay không.
Để trả lời câu hỏi này, xét gen mã hóa ovomucoid (dài 5,6 kb) có 7 intron (AG), lai Northern (Northern blotting) được tiến hành với ARN được tinh sạch từ nhân của các tế bào biểu hiện ovomucoid. Mẫu dò được sử dụng trong kỹ thuật lai Northern là ADN được đánh dấu có nguồn gốc từ gen mã hóa ovomucoid. Ảnh kết quả Northern blot được nêu ở dưới đây. ARN được tách chiết từ mỗi băng đậm màu trên gel, và sự có mặt hoặc không có mặt của các intron đặc thù trong các phân tử mARN ở nhiều băng khác nhau được kiểm tra sử dụng các mẫu dò đặc hiệu intron. Cuối cùng, các phân tử ARN được phân nhóm dựa trên số lượng các intron bị loại bỏ bằng cắt nối và tính toán cho thấy sự thống nhất về các intron bị mất đi trong các phân tử ARN khác nhau được cắt nối. Các kết quả của các tính toán này được trình bày dưới đây:
Mỗi phát biểu dưới đây là đúng hay sai?
A. Ảnh Northern blot cho thấy thứ tự loại bỏ các intron khỏi bản mã sao sơ cấp ovomucoid không phải là ngẫu nhiên.

B. Dữ liệu ở bảng cho thấy rằng intron E thường nhưng không phải luôn luôn loại bỏ khỏi bản mã sao sơ cấp trước khi intron G bị loại bỏ.
C. Dữ liệu ở bảng cho thấy rằng các intron A, B và C luôn bị loại bỏ khỏi bản sao mã sơ cấp trước các intron D và G.
D. Dữ liệu cho thấy tại thời điểm phân tích, nhìn chung có sự tăng dần về tỉ lệ của các bản sao mã được cắt nối nhiều hơn.
Hướng dẫn trả lời
A- Đúng:
- Nếu thứ tự cắt intron là ngẫu nhiên thì các băng mARN thu được sẽ có độ đậm giống nhau (khác với bản Northern blott).
B- Đúng:
- Vì tần số mất intron E luôn cao hơn tần số mất intron G trong 6 nhóm nên intron E thường bị loại bỏ khỏi bản mã sao sơ cấp trước khi intron G bị loại bỏ.
-Nhưng ở nhóm số 1: Có khả năng 2 intron bị loại bỏ là F và G (không có E) là 60%.
5% = 3%.
→ E thường nhưng không phải luôn luôn bị loại bỏ trước G.
C- Sai:
- Vì qua bảng số liệu ta thấy tần số mất intron A, B, C luôn nhỏ hơn hoặc bằng tần số mất D và G → Do vậy D và G thường dược loại bỏ trước A, B và C.
D- Đúng:
- Các bản sao được cắt nối nhiều hơn, cắt bỏ nhiều intron hơn → kích thước mARN sẽ ngắn hơn.
- Nhìn vào kết quả Northern blott ta có thể thấy băng có kích thước càng nhỏ thì càng đậm (hay mARN bị cắt càng nhiều intron thì số lượng trong nhân càng lớn .
→ Có sự tăng dần các bản sao được cắt nối nhiều hơn.
Câu 18.
Tác nhân gây bệnh ở thực vật (phytopathogen) sử dụng prôtêin tiết "secretomes" của chúng để tấn công cây chủ. Sàng lọc chức năng bẫy tiết men (YST) là phương pháp được sử dụng để phân lập và xác định các prôtêin tiết. Phương pháp này bao gồm việc tạo ra một thư viện vectơ mang các cADN (chứa các trình tự không mã hóa ở đầu 5') được tổng hợp bằng cách sử dụng các ARN phytopathogen dược hợp nhất với một dạng đột biến của gen báo cáo suc2 của Saccharomyces cerevisiae.


Suc2 mã hóa enzim invertase là prôtêin duy nhất được nấm men sử dụng để phân giải sucrose. Sự phân giải xảy ra ở môi trường ngoại bào. Dạng đột biến của gen này là suc2-SP mã hóa một invertase thiếu peptit tín hiệu ở đầu amin (đầu N) cần cho sự tiết của nó. Thư viện dung hợp được sử dụng để chuyển gen vào một chủng nấm men thiếu hụt enzim invertase, và cấy trải các tế bào đã được chuyển gen lên môi trường chọn lọc chứa sucrose. Mỗi tế bào được sống sót dự kiến sẽ chứa một vectơ tái tổ hợp có cADN mã hóa một peptit tín hiệu cho prôtêin dạng dung hợp (chimeric).
Các vectơ được tinh sạch và các cADN của chúng được giải trình tự để xác định các prôtêin được tiết. Hình 1 là sơ đồ của vectơ, và Hình 2 tóm tắt quy trình chuyển gen như được mô tả ở trên. (Em sửa lại hình và chú thích cho cân đối nhé)
Mỗi phát biểu dưới đây là đúng hay sai?
A. Nhằm mục đích sàng lọc, mồi oligo-dT có thể được sử dụng để tổng hợp cADN.
B. Số lượng biến đổi của (các) nucleotit Cytidine sau vị trí Not1 trong vectơ đảm bảo việc có được một khung đọc chính xác.
C. Sự có mặt peptit tín hiệu trong prôtêin dung hợp sẽ không cần thiết đảm bảo cho việc nấm men sinh trưởng được trên môi trường chọn lọc.
Hướng dẫn trả lời
A- Sai:
- cADN có 2 đầu là vị trí cắt của EcoRI (GAATTC) và Not I (GC...C) nên không thể sử dụng đoạn mồi oligo-dT để nhân dòng cADN
B- Đúng:
- Việc có (0-2) C bị biến đổi sau vị trí nhận biết Not I giúp riboxom bám vào và từ đó trượt tiếp để tìm bộ ba mở đầu/ Suc 2-Sp nên bảo toàn khung đọc ( không có trường hợp dịch mã cả c ADN).
C- Đúng:
- Vì trong prôtêin dung hợp có khả năng tự xuất bào mà không cần đến peptit tín hiệu → có thể phân giải saccarozo → có thể sống trong môi trường chọn lọc.
2. Câu hỏi và bài tập tự trả lời
Câu 1.
Hai chủng Hfr 1 và 2 đều là a+b+c+d+e+ được lai với một chủng F– là a-b-cd-e-. Các phép lai bị dừng ở từng giai đoạn cách nhau 5 phút và các kiểu gen của các thể tái tổ hợp được xác định. Kết quả như sau:
Hãy vẽ một bản đồ chứa tất cả các gen và cả hai nhân tố F. Thể hiện F trong các ô nhỏ và hướng truyền gen bằng mũi tên. Trên bản đồ chuẩn, gen b nằm cách điểm khởi đầu truyền gen 20 phút.
Xác định:
a. Chủng nào là chủng đột biến mất đoạn.
b. Vị trí tương ứng của mỗi đột biến mất đoạn với mất đoạn khác ?
c. Vị trí tương ứng của 6 đột biến điểm.
Câu 3.
Bạn phân lập được một số đột biến điểm (ký hiệu từ a đến g) ở vùng rII của phagơ T4. Các phép lai ghép đôi cho tỷ lệ phần trăm các viết tan kiểu dại như sau:
Phép lai % kiểu dại Phép lai % kiểu dại a x b 0,5 a x c 0,3
a x d 0,8 a x f 0,3
b x e 0,4 b x d 1,3


b x f 0,2 c x d 0,5
c x g 0,8 e x g 1,0
b x g 1,6 g x d 0,3
Bạn cũng lai mỗi đột biến điểm với mỗi đột biến mất đoạn và tính các thể tái tổ hợp kiểu dại. Kết quả thu được dưới đây (+ = thể tái tổ hợp kiểu dại, - = thể tái tổ hợp đột biến).
Đột biến điểm
Câu 2.
Có 9 chủng his-, ba trong số đó có những mất đoạn và
6 chủng có những đột biến điểm. Cấy 9 chủng trên môi trường tối thiểu và đếm số khuẩn lạc mọc (+ = mọc được).
1 2 3 4 5 6 7 8 9
+ - + + - + - + +
Phagơ được nhiễm vào một trong 9 chủng và sau đó được sử dụng để tải nạp mỗi chủng mất đoạn. Các thể tải nạp từ mỗi phép lai được cấy trên môi trường tối thiểu và đếm các khuẩn lạc mọc (+ = thể tải nạp mọc được). Dựa trên kết quả sau:
Mất đoạn a b c d e f g
a. Vẽ bản đồ các đột biến điểm, tính khoảng cách.
b. Xác định vị trí của các mất đoạn tương ứng với các đột biến điểm.
c. Các thể đột biến a và e bổ trợ với nhau và e được biết là thuộc rIIB. Các đột biến nào ở rIIA và đột biến nào thuộc rIIB?
d. Những tổ hợp của các mất đoạn nào sẽ tạo ra các thể tái tổ hợp kiểu dại?
Câu 4
Trôpônin T là một loại prôtêin liên quan đến hoạt động của cơ. Để thu được prôtêin này của người trong tế bào vi sinh vật, người ta tiến hành hai thí nghiệm như sau:
Thí nghiệm 1: Tách gen mã hoá trôpônin T từ ADN hệ gen người và chuyển vào vectơ biểu hiện ở nấm men tạo plasmit tái tổ hợp A1; chuyển vào vectơ biểu hiện ở vi khuẩn tạo plasmit tái tổ hợp A2.

Thí nghiệm 2: Tổng hợp phân tử cADN bằng cách phiên mã ngược phân tử mARN trưởng thành mã hoá trôpônin T. Sau đó, chuyển phân tử cADN này vào cùng loại vectơ biểu hiện ở nấm men tạo plasmit tái tổ hợp B1; chuyển vào cùng loại vectơ biểu hiện ở vi khuẩn tạo plasmit tái tổ hợp B2.
a. Hãy so sánh kích thước của hai plasmit tái tổ hợp A1 và B1. Giải thích.
b. Khi biến nạp và biểu hiện hai plasmit A1 và B1 trong tế bào nấm men trong cùng điều kiện thí nghiệm, họ thu được hai loại prôtêin có kích thước khác nhau từ plasmit A1 và một loại prôtêin từ plasmit B1. Hãy giải thích kết quả.
c. Khi biến nạp và biểu hiện hai plasmit A2 và B2 vào vi khuẩn E. coli trong cùng điều kiện thí nghiệm, prôtêin thu được từ plasmit A2 không có chức năng, còn prôtêin thu được từ plasmit B2 có chức năng bình thường. Hãy giải thích kết quả.
Câu 5.
Ở E. coli, khi môi trường không có glucose, galactose được dùng làm nguồn năng lượng. Khi môi trường có đồng thời glucose và galactose, galactose chủ yếu được dùng để tham gia cấu trúc thành tế bào. Operon G gồm các gen cấu trúc mã hóa enzim có vai trò gắn galactose để cấu trúc thành tế bào, hai trình tự khởi đầu phiên mã (một promoter mạnh và một promoter yếu) và trình tự chỉ huy (operator - là vị trí liên kết của prôtêin ức chế). Operon G được điều hòa bởi một gen ức chế. Các nhà nghiên cứu đã phân lập được 6 chủng E. coli tương ứng mang 6 đột biến điểm mất chức năng làm ảnh hưởng tới mức độ biểu hiện của Operon G như ở bảng trên (kí hiệu từ Chủng 1 đến Chủng 6).

Từ các chủng kiểu dại và đột biến, các nhà nghiên cứu tạo được 5 chủng lưỡng bội về 6 đột biến nêu trên ( kí hiệu từ Chủng A đến Chủng E), rồi nuôi các chủng này trên môi trường có hoặc không có glucose ( nhưng luôn có galactose) và xác định mức độ biểu hiện của Operon G. Kết quả nghiên cứu được trình bày ở bảng dưới đây:
Từ các thông tin trên, hãy biện luận để xác định vị trí của mỗi đột biến (từ 1 đến 6) thuộc vùng nào sau đây: vùng mã hóa của các gen cấu trúc, các promoter, operator của Operon G
C. KẾT LUẬN VÀ KIẾN NGHỊ
I. Kết luận
Chuyên đề đã đề cập đến:
- Hệ thống hóa một số kiến thức về vấn đề tích hợp hóa sinh, vi sinh và di truyền.
- Sưu tầm, giới thiệu một số câu hỏi, bài tập vận dụng, một số đề thi thử cho học sinh giỏi.
Với nội dung kiến thức cơ bản về hóa sinh, vi sinh và di truyền cùng với hệ thống câu hỏi, bài tập được đề cập ở trên chúng tôi đã sử dụng trong giảng dạy tại nhà trường và thu được những kết quả nhất định:
+ Học sinh hiểu bài, nắm chắc kiến thức, linh hoạt trong các câu hỏi vận dụng, giải thích tốt nhiều vấn đề có tính thực tiễn cao.
+ Kĩ năng phân tích và khai thác hình vẽ, sơ đồ...của học sinh được nâng cao.
+ Các em hứng thú hơn trong giờ học, tích cực chủ động sáng tạo trong quá trình học để lĩnh hội kiến thức.
+ Nội dung chuyên đề sử dụng tốt trong công tác ôn thi THPT Quốc gia và bồi dưỡng học sinh giỏi các cấp.
II. Kiến nghị
Chuyên đề này đã được tập hợp, chọn lọc từ nhiều nguồn tài liệu liên quan, hy vọng sẽ giúp ích cho các thầy cô giáo và các em học sinh trong quá trình giảng dạy và học tập. Tuy nhiên, nhiều vấn đề còn ít nhiều mang tính chủ quan, kiến thức đưa ra trong phần lý thuyết còn chưa đầy đủ, hệ thống bài tập chưa bao quát hết các nội dung, kính mong các thầy cô giáo và các em học sinh đóng góp bổ sung để chuyên đề được hoàn thiện hơn, hợp lý hơn và có ích hơn trong thực tiễn. Xin trân trọng cảm ơn!
D. TÀI LIỆU THAM KHẢO
Tài liệu tiếng Anh
1. Alberts B. et al. (2008). Molecular Biology of the Cell (Fifth edition). GarlandPublishing. New York
2. Madigan, M.T., Martinko, J.M. and Parker, J., 2012. Biology of Microorganisms. Tenth edition, Prenhall.
3. Stefan H.E. Kaufmann, 2004. Novel Vaccination Strategies. Wiley-VCH VerlagGmb.
4. Uldis N. Streips, Ronald E. Yasbin, 2002. Modern microbial gentics. Secondedition. Wiley-Liss, Inc.
5. Peter J. Russel, 2000. Fundamentals of Gentics. Addison Wesley Longman Inc.
6. Nitesh RAI, Ludwig W, Schleifer KH (2011). Phylogentic identification and in situ detection of individual microbial cells without cultivation. Microbiology Rev.
Tài liệu tiếng Việt
1. Campbell ; “Sinh học”; NXB Giáo dục, 2019.
2. Nguyễn Thành Đạt. Cơ sở sinh học Vi sinh vật. NXB Giáo dục Hà Nội, Tập I(1999), Tập II (2001)
3. Nguyễn Thành Đạt (chủ biên), Mai Thị Hằng. Sinh học vi sinh vật. Nhà xuất bản Giáo dục, 2000.
4. Harvey Lodish, Sinh học phân tử của tế bào.NXB trẻ , 2018
5. Nguyễn Văn Ty ( chủ biên), Tài liệu chuyên sinh học THPT- Vi sinh vật học. NXB Giáo dục, 2009.
6. Đề thi Olympic Quốc tế sinh học quốc tế, đề thi chọn đội tuyển Olympic Quốc tế sinh học, đề thi học sinh giỏi Quốc gia, đề thi HSG các trường Chuyên khu vực DH&ĐBBB, đề thi trại hè Hùng Vương.
Chuyên đề
MỘT SỐ NỘI DUNG TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN
Nguyễn Thị Thu - Trường THPT Chuyên Bắc Giang - Bắc Giang
Chuyên đề đạt giải Ba
Phần thứ nhất: MỞ ĐẦU
1. LÍ DO CHỌN ĐỀ TÀI
Sinh học là môn khoa học nghiên cứu các mối quan hệ của các hệ thống sống ở các cấp độ tổ chức khác nhau từ cấp độ phân tử đến cấp độ sinh quyển. Các mối quan hệ đó rất phức tạp nhưng chặt chẽ và tuân theo những quy luật nhất định.
Trong Sinh học có nhiều phân môn như tế bào-hóa sinh, vi sinh, sinh lý thực vật, sinh lý động vật, di truyền, sinh thái, tiến hóa, việc chia theo phân môn giúp học sinh dễ tiếp cận nhưng lại khó hình dung kiến thức đặc biệt là phần di truyền học tích hợp hóa sinh, vi sinh.
Trong chương trình Sinh học mới năm 2018 phần di truyền học bao gồm di truyền phân tử; Di truyền nhiễm sắc thể; di truyền gen ngoài nhân; mối quan hệ kiểu gen - môi trường - kiểu hình; thành tựu chọn, tạo giống bằng các phương pháp lai hữu tính; di truyền quần thể; di truyền học người. Điểm mới chương trình 2018 so với chương trình 2006 là:
+ Cách tiếp cận trong chương trình 2018 có điểm khác với chương trình 2006: Từ khái quát tới cụ thể, bắt đầu từ khái niệm “Hệ gen”.
+ Khái niệm “Tương tác gen” được cập nhật với những nghiên cứu mới của thế giới
+ Bổ sung, cập nhật những tri thức mới, khai thác cơ sở phân tử của các hiện tượng di truyền và biến dị; làm sáng tỏ bản chất của các hiện tượng này.
+ Bổ sung các thông tin mới liên quan đến các biến đổi ngoại di truyền trong sự biểu hiện của tính trạng.
+ Nhấn mạnh khả năng ứng dụng những tiến bộ của Di truyền học trong khoa học và đời sống hiện nay, liên quan đến y học, nông nghiệp, khoa học hình sự…
+ Phần Quy luật di truyền đi sâu vào việc khai thác bản chất của mối quan hệ gen - protein - tính trạng để giải thích các hiện tượng tương tác gen, tính đa hiệu của gen, gen đa alen, mỗi quy luật đều xuất phát từ bối cảnh/ tình huống có vấn đề khoa học.
+ Phần liên kết gen mở rộng ứng dụng trong lập bản đồ di truyền (dựa vào lai hữu tính) và bản đồ vật lí (dựa trên những thành tựu của các kĩ thuật phân tích ADN, giải trình tự ADN…)
Để học tốt và thi tốt kỳ thi tốt nghiệp THPT, thi học sinh giỏi cấp tỉnh với hình thức trắc nghiệm hoặc trắc nghiệm kết hợp tự luận như hiện nay học sinh cần đổi mới phương pháp học tập và làm quen với hình thức thi cử. Nếu trước đây học và thi môn Sinh học, học sinh cần học thuộc và nhớ từng câu, từng chữ hoặc đối với bài toán học sinh phải giải trọn vẹn các bài toán. Nay học sinh lưu ý trước hết đến sự hiểu bài, hiểu thấu đáo các kiến thức cơ bản đã học vận dụng những hiểu biết đó vào việc phân tích, xác định nhận biết các đáp án đúng sai trong các câu trắc nghiệm. Đặc biệt đối với các câu bài tập làm thế nào để có được kết quả nhanh nhất? Đó là câu hỏi lớn đối với tất cả các giáo viên. Trước thực tế đó đòi hỏi mỗi giáo viên cần xây dựng cách dạy riêng của mình.
Trong những năm gần đây, các bài tập di truyền học tích hợp hóa sinh, vi sinh xuất hiện trong đề thi học sinh giỏi quốc gia môn sinh học ngày càng đa dạng và mức độ khó tăng dần lên. Có thể nói nội dung này đóng một vai trò rất quan trọng trong việc phân hóa học sinh ở bài thi ngày thứ nhất, bài thi ngày thứ hai trong Kì thi chọn học sinh giỏi quốc gia và kì thi vòng 2 chọn đội tuyển Olimpic Quốc tế môn Sinh học. Trái ngược với sự thay đổi của đề thi học sinh giỏi thì nguồn tài liệu để giảng dạy phần này cho giáo viên lại khá hạn chế, không có tính hệ thống.
Từ những cơ sở trên, chúng tôi đã xây dựng hệ cơ sở lý thuyết, cách giải bài tập di truyền tích hợp hóa sinh, vi sinh sưu tầm một số câu hỏi, bài tập chọn lọc nhằm mục tiêu vô cùng thiết thực là giúp cho học sinh thi tốt nghiệp trung học phổ thông, thi học sinh giỏi cấp tỉnh, thi học sinh giỏi quốc gia học tập và nghiên cứu về phần này tốt hơn và đồng thời cũng giúp các đồng nghiệp giảng dạy thuận lợi hơn.
Chính vì vậy chúng tôi chọn đề tài: “Một số nội dung tích hợp hóa sinh, vi sinh và di truyền”
2. MỤC ĐÍCH NGHIÊN CỨU
Nghiên cứu về lí thuyết phần tích hợp hóa sinh, vi sinh và di truyền.
Tổng hợp các câu hỏi và bài tập vận dụng phần tích hợp hóa sinh, vi sinh và di truyền.
Ứng dụng vào việc dạy học sinh chuyên sinh và học sinh khối 12 ôn thi trung học phổ thông quốc gia, thi học sinh giỏi quốc gia môn Sinh học, thi giải toán trên máy tính cầm tay quốc gia nhằm giúp học sinh thu được kết quả cao trong các kì thi này.
3. PHƯƠNG PHÁP NGHIÊN CỨU
Thông qua các bài tập có sẵn hoặc tự đề ra, bài tập trong các đề thi học sinh giỏi quốc gia, bài tập trong các đề thi giải toán trên máy tính khu vực, bài tập trong các đề thi tuyển sinh đại học – cao đẳng, tốt nghiệp THPT, thi THPT quốc gia, sách, tài liệu tham khảo để hướng dẫn học sinh cách làm phần lí thuyết, giải bài tập và phát huy khả năng tích cực, năng động, tư duy sáng tạo trong việc vận dụng kiến thức.
Phần thứ hai: NỘI DUNG CHUYÊN ĐỀ
CHƯƠNG I. CƠ CHẾ DI TRUYỀN Ở MỘT SỐ VIRUT
1.1. Tổng quan về quá trình nhân lên của virus
Quá trình nhân lên của virus trong tế bào bao gồm 7 bước:
Hấp phụ (adsorption)
Xâm nhập và cởi vỏ (penetration và uncoating)
Phiên mã (transcription) tạo mRNA của virus
Dịch mã (translation) mRNA để tạo protein virus
Sao chép (replication) genome
Tự lắp ráp (maturation) protein với genome để tạo virion
Giải phóng (release) ra khỏi tế bào
Nếu các bước 3, 4, 5 nhập vào một bước gọi là bước tổng hợp các thành phần (biosynthesis) thì quá trình nhân lên còn 5 bước.
Ở một số virus, quá trình nhân lên không treo trình tự như trên mà xảy ra đồng thời. Ví dụ cùng lúc đều tiến hành phiên mã, dịch mã, sao chép, lắp ráp và chui ra khỏi tế bào.
1.1.1. Sự hấp phụ
Virus gắn vào thụ thể (receptor) đặc hiệu nằm trên màng sinh chất của tế bào chủ, theo nguyên tắc khoá – chìa. Một số virus còn gắn thêm vào các đồng thụ thể (coreceptor). Sự hấp phụ xảy ra tốt nhất ở 37oC.
Virus gắn được vào các thụ thể là nhờ các liên kêt hoá học như liên kết hydro, ion, Vander Waals, nhưng không có liên kết đồng hoá trị
Vị trí gắn:
+ Virus có các vị trí khác nhau chứa protein gắn vào thụ thể của tế bào.
+ Virus trần có vị trí gắn nằm trên bề mặt capsid, đôi khi nằm sau bên trong (ví dụ virus polio) hoặc đôi khi là đỉnh của khối đa diện (virus lở mồm long móng). Ở virus adeno, vị trí gắn là đầu mút của các sợi mọc ra từ đỉnh capsid khối đa diện. Đối với virus có vỏ ngoài, vị trí gắn là các gai glycoprotein bề mặt.
1.1.2. Xâm nhập và cởi vỏ
Sau khi gắn vào thụ thể, virus phải vượt qua màng sinh chất để xâm nhập vào tế bào theo một trong 2 cơ chế nhập bào hoặc dung hợp.
1.Nhập bào: Cả virus trần và virus có vỏ ngoài đều có thể xâm nhập theo lối nhập bào.
Cơ chế chung bao gồm việc virion gắn vào màng sinh chất nơi mặt trong được bao phủ bởi lớp clathrin (protein dạng sợi tạo thành mạng lưới với các mắt hình đa giác). Virion ấn sâu vào màng tạo hốc rồi khép lại tạo thành túi nội bào hay endosome được lớp clathrin bao phía ngoài. Kênh proton của màng endosome vận chuyển H+ vào trong làm cho pH trong endosome giảm xuống ( pH 4.5 - 5). Endosome sẽ dung hợp với lysosom. Trước khi dung hợp, lớp clathrin bị loại bỏ. pH thấp hoạt hoá enzyme phân giải capsid. Cũng như màng endosome ở một số virus, pH thấp trong endosome dẫn đến việc dung hợp vỏ capsid với màng endosome, làm vỡ vỏ capsid và giải phóng genome.
2. Dung hợp: Khác với virus trần, chỉ có xâm nhập vào tế bào theo lối nhập bào, các virus có vỏ ngoài có thể vào tế bào theo cả 2 cách.
Virus có vỏ ngoài có thể vào tế bào theo cách dung hợp vỏ ngoài virus với màng sinh chất vì chúng có cùng bản chất. Khi 2 màng hoà nhập sẽ đứt ra, nucleocapsid sẽ được chuyển vào tế bào chất. Sự dung hợp này xảy ra với sự tham gia của protein dung hợp (protein F).
Như trên đã nói, virus có vỏ ngoài cũng có thể xâm nhập vào tế bào theo lối nhập bào. Vỏ ngoài virus bám vào thụ thể sau đó ấn lõm tạo endosome. Khi pH trong endosome giảm gai protein F sẽ chồi lên cắm vào màng endosome như chiếc neo, kéo vỏ ngoài virus sát với màng endosome và tiến hành dung hợp.
Hình 1.1. Sự nhập bào theo kiểu thực bào, tạo endosome
Bơm proton làm giảm pH trong endosome, hoạt hoá sự dung hợp giữa vỏ ngoài virus với màng endosome (1b), hoặc hoạt hoá enzyme làm tan màng endosome (2). Dung hợp xảy ra trên bề mặt tế bào giữa vỏ ngoài virus với màng sinh chất.

Sự vận chuyển genome virus vào nhân
Hầu hết virus ARN ở eukaryota tiến hành sao chép trong tế bào chất, vì chúng có thể mã hoá cho tất cả các enzyme cần cho sao chép genome mà không cần đến các enzyme của tế bào nằm trong nhân. Virus cúm A là ngoại lệ, chúng cần bộ máy cắt nối của tế bào nên genome của chúng phải được đưa vào nhân.
Virus retro cũng là virus RNA, nhưng tiến hành sao chép trong nhân. Thoạt đầu chúng nhờ enzyme phiên mã ngược tổng hợp ADN trên khuôn ARN trong tế bào chất, sau đó nằm đợi cho đến khi tế bào bắt đầu phân chia (giai đoạn M), màng nhân tạm thời bị vỡ, ADN cùng với protein liên kết lúc đó mới vào được trong nhân. Do đó virus này chỉ nhân lên được ở giai đoạn tế bào đang phân chia.
Hầu hết virus ADN tiến hành sao chép trong nhân, ngoại trừ virus pox và irido có thể sao chép trong tế bào chất. Đối với virus sao chép trong nhân, protein cấu trúc của chúng có trình tự bám được vào vi ống (microtubule). Vi ống là các ống rỗng, đường kính 25 nm, là thành phần của bộ khung tế bào, có chức năng nâng đỡ và neo giữ nhiều thành phần của tế bào và được ví như đường ray để vận chuyển vật chất hoặc bào quan tới một vị trí nhất định trong tế bào. Một đầu vi ống được ký hiệu là (+) và một đầu ký hiệu là (-). Đầu (+) nằm gần màng sinh chất, còn đầu (-) nằm gần nhân. Một protein gọi là proetin vận chuyển (motor protein) tự nó di chuyển và chở các chất cũng như bào quan dọc theo vi ống từ vùng ngoại vi của tế bào tới vùng gần nhân. Các virus herpes, adeno, parvo, retro sử dụng hệ thống này để chở nucleocapsid vào sát màng nhân.
Màng nhân được cấu tạo từ 2 lớp lipid kép, ở đó có lỗ nhân. Hầu hết nucleocapsid có kích thước quá lớn để có thể lọt qua lỗ nhân. Các phân tử muốn qua lỗ phải tạo phức với các protein chuyên biệt của tế bào có chức năng mang gọi là importin để mang vào hoặc exportin để mang ra khỏi nhân. Kênh có thể mở để cho phép các hạt có kích thước đến 25 nm thậm chí lớn hơn đi qua, ví dụ virus nhỏ như parvo, nhưng với virus lớn hơn thì phải cởi vỏ ở lỗ nhân.
Cởi vỏ cho genome
Cởi toàn bộ hoặc một phần vỏ để giải phóng genome ra khỏi vỏ capsid. Tuỳ loại virus mà quá trình có thể diễn ra tại các vị thí khác nhau:
- Trên bề mặt tế bào, capsid rỗng nằm lại bên ngoài tế bào.
- Cởi vỏ bên trong tế bào chất.
- Cởi vỏ tại lỗ nhân.
- Cởi vỏ bên trong nhân.
Cần nhớ rằng virus xâm nhập thành công vào tế bào không có nghĩa là chúng luôn luôn nhân lên được. Tế bào cũng có cơ chế bảo vệ chống lại virus, ví dụ enzyme từ lysosom có thể làm bất hoạt virus trước và sau cởi vỏ; interferon cảm ứng tạo protein độc ức chế sự nhân lên của virus. Một số virus nhiễm ở dạng tiềm ẩn, chúng không nhân lên, nhưng genome vẫn còn nguyên vẹn và vẫn có tiềm năng nhân lên.
1.1.3. Tổng hợp các thành phần
Sau khi xâm nhận vào tế bào, pha đầu tiên của chu trình nhân lên gọi là pha ẩn. Ở giai đoạn này không phát hiện được bất kỳ virus nào. Đây là đặc điểm chỉ thấy có ở virus. Sự biểu hiện của genome được bắt đầu rất sớm, ngay sau khi virus xâm nhập vào tế bào. Có 4 quá trình xảy ra trước khi hạt virus được lắp ráp:
- Phiên mã tạo mRNA
- Dịch mã sớm tạo protein phi cấu trúc – đó là các enzyme dùng cho sao chép
- Sao chép tạo genome
- Dịch mã muộn tạo protein cấu trúc để cấu tạo capsid và vỏ ngoài
1.1.4. Lắp ráp
Lắp ráp là sự tự kết nối các thành phần virus để tạo ra virion hoàn chỉnh đòi hỏi phải có cấu trúc bền vững, tồn tại được trong môi trường như là một thực thể có khả năng gây nhiễm, tuy nhiên cũng đòi hỏi khi vào trong tế bào cấu trúc này phải không bền vững thì mới có thể giải phóng dễ dàng genome vào tế bào chất. Do vậy virion phải có cơ chế đóng mở kiểu “công-tắc” để có thể biến đổi từ trạng thái bền vững sang
trạng thái không bền vững. Công tắc này liên quan đến việc gắn vào receptor hoặc sự thay đổi pH trong endosome.
Khi số lượng genome và protein cấu trúc được tích luỹ đến ngưỡng thì chúng sẽ tiến hành lắp ráp tạo nucleocapsid.
- Virus có cấu trúc dạng xoắn: Đối với virus ARN đơn có cấu trúc dạng xoắn, lúc đầu một số phân tử protein cấu trúc sẽ bám theo chiều xoắn của genome RNA, sau đó các phân tử khác lần lượt bám theo cho đến khi phủ hết RNA.
- Virus có cấu trúc dạng khối đa diện: Trước hết cần phải lắp ráp một cấu trúc rỗng hình cầu gọi là procapsid. Genome virus chui vào procapsid, sau đó cải biến từ cấu trúc hình cầu sang hình khối đa diện; ví dụ các virus adeno, picorna thực hiện cải biến bằng cách cắt bớt 1 hoặc nhiều protein cấu trúc.
Genome chui vào trong procapsid qua kênh nằm ở vị trí mà sau này sẽ là đỉnh của khối đa diện. Bất kỳ enzyme nào tham gia vào đóng gói genome cũng nằm ở vị trí này. Ở tế bào thực khuẩn cũng như vậy, trước hết cũng phải tạo một “tiền đầu” (prohead) sau đó genome chui qua 1 cái cổng nằm ở một đỉnh. Vị trí này cũng có chức năng nối với đuôi.
a- Đóng gói genome
Trong tế bào có rất nhiều axit nucleic, của cả virus và tế bào. Vậy làm thế nào để genome của virus lại được lựa chon và lắp ráp chứ không phải của tế bào? Sở dĩ như vậy vì virus có một protein chuyên biệt, nhận diện tín hiệu đóng gói, nằm ở vùng có cấu trúc bậc hai của genome. Hầu hết virus có genome sợi đơn có thể đóng gói hoặc sợi dương hoặc sợi âm, nên tín hiệu đóng gói phải có duy nhất ở sợi cần được đóng gói. Genome được nén trong thể tích nhỏ. Virus ADN kép có kích thước lớn như virus herpes, đóng gói genome chặt đến nỗi tạo áp suất lớn gấp 10 lần so với áp suất trong chai rượu sâm – banh.
b- Cơ chế lắp ráp
Trước đây Fraenkel-Conrat đã tách genome ra khỏi capsid của virus đốm thuốc lá, sau đó lại lắp ráp chúng với nhau trong điều kiện pH và sự có mặt của một số ion nhất định, để tạo virus hoàn chỉnh. Với các virus đơn giản, chỉ chứa 1 axit nucleic và một số ít loại protein thì có thể tự lắp ráp một cách đơn giản như trên. Nhưng với các virus phức tạp, như virus herpes và phage có đuôi, thì không thể tái lắp ráp như vậy. Chúng cần phải được lắp ráp trực tiếp trong môi trường của tế bào nhiễm. Khi lắp ráp cần phải có mặt tạm thời 1 protein dùng làm giàn giáo. Các protein cấu trúc theo đó mà lắp vào để tạo capsid. Khi công việc hoàn tất, protein giàn giáo bị loại bỏ khỏi virion hoặc bằng enzyme phân giải hoặc giữ lại để tái sử dụng.
c- Sự tạo màng virion
Vỏ ngoài virus có thể được tạo thành theo 1 trong 2 cơ chế: Cải biến màng sinh chất của tế bào rồi nảy chồi ra ngoài hoặc tự tổng hợp màng mới bao quanh nucleocapsid.
-Cải biến màng sinh chất
Vỏ ngoài của virus thường có nguồn gốc từ màng sinh chất, được cuốn theo khi virus nảy chồi. Vùng màng mà virus sẽ nảy chồi được đính trước 1 hoặc nhiều loại protein đặc hiệu của virus, thường là glycoprotein, các protein này ngâm trong lớp lipid kép. Protein M của virus (các phân tử protein này gắn với nhau tạo thành màng đệm – M) tập trung nhiều ở vùng màng, có ái lực với nhau và đẩy protein tế bào ra khỏi màng. Đôi khi protein tế bào không bị đẩy ra hết nên chúng có thể tham gia vào thành phần vỏ ngoài. Ví dụ vỏ ngoài của virus HIV-1 có chứa protein MHC-II (phức hợp hoà hợp mô chính) của tế bào. Trước khi nảy chồi, protein M tới gắn vào phần đuôi nằm trong tế bào chất của glycoprotein xuyên màng, sau đó nucleocapsid tiến đến bám vào màng M. Nucleocapsid khi nảy chồi sẽ cuốn theo màng tế bào chất và màng M để tạo vỏ ngoài. Không phải tất cả các virus đều có màng M. Ví dụ ở trường hợp virus sốt vàng thì bề mặt nucleocapsid sẽ gắn trực tiếp vào đuôi glycoprotein trong màng.
-Tổng hợp mới màng virus
Chỉ có một số ít virus tạo màng lipid muộn trong quá trình nhân lên. Màng này có thể tham gia hình thành vỏ ngoài (ví dụ virus pox) hoặc nằm trong nhân tạo 1 lớp phía mặt dưới của capsid (ví dụ virus irido).
Khi nhân lên virus baculo tạo ra 2 loại virion có vỏ ngoài: Loại thứ nhất có chức năng lây nhiễm sang tế bào khác trong vật chủ. Loại này có được vỏ ngoài là do nảy chồi qua màng sinh chất. Loại virion thứ hai có chức năng gây nhiễm vào vật chủ mới, vỏ ngoài của nó bao quanh nucleocapsid nằm trong nhân dẫn đến các virion hợp nhất với nhau trong một bọc gọi là thể bọc (occlusion body). Thể bọc giúp virus tránh tác động của môi trường bên ngoài, nên virus có thể duy trì rất lâu ngoài tế bào sống.
1.1.5. Giải phóng virus khỏi tế bào
Đây là giai đoạn cuối cùng của chu trình nhân lên. Nhiều virus được giải phóng khi tế bào bị nổ tung, do thành tế bào bị phân giải, cộng với áp lực lớn trong tế bào. Ví dụ nhiều phage, peptidoglycan bị phân giã dẫn đến làm vỡ tế bào. Số khác lại có khả năng tổng hợp protein ức chế enzyme tham gia vào tổng hợp thành tế bào, làm cho thành tế bào yếu đi nên dễ bị vỡ.
Số lượng virus được tạo thành qua mỗi mẻ là rất lớn. Trong tế bào E.coli có kích thước nhỏ bé, phage T4 có kích thước lớn tạo ra được 200 virion. Virus picorna có kích thước rất nhỏ khi nhiễm vào tế bào động vật có kích thước lớn đã tạo ra đến 100.000 virion.
330
Một số virus không làm tan tế bào mà chỉ chui ra từ từ theo lối nảy chồi, số khác chui ra thông qua việc tạo thành túi hay bọng từ màng lưới nội chất hoặc bộ máy Golgi. Màng bọng dung hợp với màng tế bào chất và đẩy virus ra ngoài. Ở các trường hợp này, tế bào vẫn còn sống thêm một thời gian nữa.
1.2. Các cơ chế di truyền của virus
1.2.1. Sao chép genome của virus
Đây là bước thứ 5 của chu trình nhân lên. Nhìn chung virus ADN và virus ARN sao chép trực tiếp genome của mình thành RNA. Tuy nhiên một số virus ADN khi sao chép cần qua trung gian ARN và một số virus ARN cần qua trung gian DNA.
Các virus ADN tiến hành sao chép trong nhân (trừ virus pox). Các virus có genome nhỏ (virus papilloma) sử dụng ADN polymerase của tế bào, còn virus có genome lớn (ví dụ herpes) thì mã hoá cho enzyme của mình.
Các virus ARN tiến hành sao chép trong tế bào chất (trừ virus cúm và retro), sử dụng enzyme do chúng mã hoá. Trong hầu hết trường hợp, sao chép và phiên mã là một. Enzyme dùng cho sao chép cũng là enzyme dùng cho phiên mã.
Virus ADN kép sao chép trong nhân theo cơ chế bán bảo tồn như ở tế bào, sử dụng enzyme ADN polymerase phụ thuộc ADN của tế bào. Tuy nhiên virus pox là virus ADN kép, sao chép trong tế bào chất sử dụng enzyme ADN polymerase do chúng mã hoá.
Virus ADN đơn (+ hoặc -) tất cả đều sao chép trong nhân, sử dụng ADN polymerase của tế bào và phải qua giai đoạn trung gian tạo sợi ADN kép, gọi là dạng sao chép (RF-replicative form). Tư RF sao chép tạo genome.
Virus ARN kép (ví dụ virus rota) luôn có genome phân đoạn, sao chép trong tế bào chất và sử dụng enzyme ARN polymerase phụ thuộc ARN do chúng mã hoá.
Virus ARN đơn, (+) khi sao chép phải qua bước tạo ARN (-) trung gian làm khuôn để tổng hợp genome ARN (+). Genome ban đầu được dùng làm mRNA để tổng hợp ARN polymerase.
Virus ARN đơn, (-) sao chép trong tế bào chất, sử dụng enzyme ARN polymerase phụ thuộc ARN (RNA-replicase) do chúng mang theo để tổng hợp sợi ARN (+) trung gian làm khuôn tổng hợp genome (-).
Virus ARN (-) phân đoạn (ví dụ virus cúm) sao chép trong nhân, sử dụng ARN polymerase phụ thuộc ARN do chúng mang theo.
Virus ARN (+) phiên mã ngược (ví dụ HIV), trước hết dùng enzyme phiên mã ngược của virus (DNA polymerase phụ thuộc RNA) để tạo ADN kép trong tế bào chất, sau đó vào nhân gắn vào nhiễm sắc thể của tế bào rồi từ đó sao chép tạo genome ARN nhờ enzyme ARN polymerase phụ thuộc ADN của tế bào.
Virus ADN kép phiên mã ngược (ví dụ HBV). muốn sao chép phải qua bước trung gian tạo ARN tiền genome trong nhân, dùng enzyme ARN polymerase phụ thuộc ADN của tế bào. ARN mới sinh ra khỏi nhân làm khuôn để tổng hợp sợi ADN () (sợi L) nhờ enzyme phiên mã ngược của virus. Từ sợi ADN (-) làm khuôn tổng hợp sợi ADN (+) tương bù do enzyme của virus mã hoá.
1.2.1.1. Khởi đầu sao chép
Mỗi genome virus có một trình tự đặc biệt, tại đó bắt đầu sao chép axit nucleic của virus. Khi sao chép cần gắn mồi. Đó là phản ứng đầu tiên của một nucleotide với nhóm –OH tại vị trí khởi đầu sự sao chép genome của nhiều virus (ví dụ rota, rhabdo) bắt đầu khi nucleotide đầu tiên của sợi mới bắt cặp với 1 nucleotide trong ARN của virus. Nucleotide đầu tiên hoạt động có hiệu quả như một mồi để sao chép RNA, khi nhóm 3’-OH của nó gắn với nucleotide thứ 2.
Một số virus ADN đơn (ví dụ virus parvo) sử dụng cách tự tạo mồi. Đầu 3’ của ADN có các trình tự tương bù, nên có thể gập lại, bắt cặp với nhau tạo đầu 3’-OH thay cho mồi.
Để khởi đầu sao chép, nhiều genome ADN và một số genome ARN của virus dùng một phân tử ARN hoặc protein làm mồi.
1.2.1.2. ARN mồi và protein mồi
Sự tổng hợp ADN của tế bào được bắt đầu sau khi một vùng xoắn kép được mở xoắn tạo bong bóng nhờ enzyme helicase và sau khi enzyme primase tổng hợp một đoạn ARN ngắn làm mồi. Cần một mồi cho sợi dẫn đầu và nhiều mồi cho tổng hợp các đoạn Okaseki của sợi muộn (sợi sau). Nucleotide đầu tiên của ADN mới gắn vào 3’OH của ARN mồi.
Một số virus ADN dùng mồi ARN để sao chép genome. Một số (ví dụ virus polymase) dùng primase của tế bào để tổng hợp mồi. Số khác (ví dụ virus herpes và phage T7) lại mã hoá cho primase của riêng mình. Virus retro dùng tRNA của tế bào làm mồi khi ở ngoài tế bào chất, nhưng khi cài xen genome của mình vào nhiễm sắc thể của tế bào nên để sao chép, chúng dùng mồi do primase của tế bào tổng hợp.
Một số virus động vật sử dụng protein làm mồi, trong đó có virus ADN như virus adeno và virus ARN như picorna. Nhóm 3’-OH của serin hoặc tyrosin trong protein sẽ gắn với nucleotide sợi mới.
Virus hepDNAa (ví dụ HBV) là virus ADN kép, dùng mồi là protein để khởi đầu tổng hợp sợi ADN (-) và mồi ARN để khởi đầu tổng hợp ADN (+). Mồi protein và mồi ARN của virus hepDNAa không bị cắt bỏ sau khi vai trò của chúng đã hoàn tất mà vẫn được dính vào đầu 5’ của genome.
1.2.1.3. Sao chép DNA
Mỗi ADN virus có ít nhất một trình tự chuyên biệt để bắt đầu sao chép gọi là trình tự khởi đầu (Ori). Các protein khởi đầu sao chép ADN bám vào vị trí này bao gồm:
- Helicase bám vào vị trí để tháo xoắn.
- Một số protein bám sợi ADN đơn, giữ cho 2 sợi không bắt cặp lại với nhau.
- Một ADN polymerase.
Về cơ bản quá trình sao chép ADN của virus giống như của tế bào. Ở vi khuẩn, số enzyme tham gia ít hơn so với ở eukaryota. Ví dụ helicase-primase của phage T7 (ở E.coli) chỉ là 1 phân tử, trong khi của virus herpes simplex (ở tế bào động vật) là một phức hợp gồm 3 loại protein.
Sự tổng hợp ADN diễn ra gần chạc sao chép. Một trong 2 sợi là sợi dẫn đầu, sợi còn lại là sợi muộn, được tổng hợp thành các đoạn Okaseki sau đó nối lại với nhau nhờ DNA-ligase. Sợi ADN kép mới tạo thành có chứa một mạch của sợi mẹ. Cách sao chép này gọi là bán bảo tồn. Ngược lại với nó là sao chép bảo tồn xảy ra ở một số virus.
Một số genome ADN là phân tử dạng thẳng trong khi một số khác lại là dạng khép vòng. Một số phân tử dạng thẳng khi sao chép lại được khép vòng, cho nên nhiều genome virus được sao chép như là một phân tử vòng tròn. Từ đây có 2 phương thức sao chép.
- Sao chép theo cơ chế theta hay dạng mắt. Từ vị trí Ori tạo ra 2 chạc ba sao chép. Sao chép cùng lúc theo 2 chiều thuận nghịch kim đồng hồ.
- Sao chép theo cơ chế xích ma (δ). Phân tử ADN kép dạng vòng gồm sợi ngoài (+) và sợi trong (-). Sợi ngoài bị cắt đứt ở liên kết photphodiete tạo ra đầu 3’-OH tự do (gọi là điểm sinh trưởng) Sợi trong xoay được dùng làm khuôn. Các nucleotide nối vào đầu 3’-OH để tạo ra 1 sợi ADN mới. Từ sợi ADN mới này lại được gắn mồi tổng hợp mạch bổ sung tạo ADN kép.
Một số virus lúc mới nhiễm sao chép theo cơ chế theta nhưng ở giai đoạn sau lại theo cơ chế xích ma (ví dụ phage ϕx174).
Một số virus ADN như virus herpes và phage T4, kết quả sao chép tạo ra phân tử ADN rất lớn gọi là phân tử trùng lặp (concateme). Mỗi concateme cấu tạo gồm nhiều bản sao genome nối với nhau. Trước khi lắp ráp vào virion concateme sẽ được phân cắt thành các phân tử có kích thước và trình tự của genome.
Thông thường phiên mã là quá trình truyền thông tin di truyền từ ADN sang ARN nhờ enzyme ARN polymerase (chính xác hơn là ARN polymerase phụ thuộc DNA), tuy nhiên genome của virus có thể là RNA, cho nên ở virus phiên mã đơn giản chỉ là quá trình truyền thông tin di truyền từ genome sang mRNA. Có 3 loại ARN polymerase: ARN polymerase I phiên mã rRNA, ARN polymerase II phiên mã các gen mã hoá protein và ARN polymerase III phiên mã rRNA 5S, tRNA và các ARN nhỏ bé khác.
Dựa vào quá trình phiên mã, tạo ra mRNA mà David Baltimore đã phân loại tất cả virus thành 7 nhóm như đã nêu ở phần phân loại: (1) Virus ADN kép, (2) virus ADN đơn (+/-), (3) virus ARN kép, (4) Virus ARN (+), (5) virus ARN (-), (6) virus ARN phiên mã ngược và (7) virus ADN phiên mã ngược.
Axit nucleic sợi đơn (DNA hoặc RNA) có trình tự nucleotide giống với trình tự nucleotide của mRNA quy ước là sợi (+), ngược lại có trình tự tương bù với mRNA được quy ước là sợi (-).
Nhóm I. Virus ADN kép hầu hết tiến hành phiên mã trong nhân, sử dụng ARN polymerase phụ thuộc ADN (tức là ARN polymerase II) của tế bào. Đối với virus ADN kép thì không phân biệt sợi (+) và sợi (-) mà gọi là sợi trái (L) và sợi phải (R), bởi vì genome của hầu hết các virus này có khung đọc mở (ORF) theo cả 2 hướng. Một số virus ADN phiên mã trong tế bào chất (ví dụ virus pox), sử dụng ARN polymerase phụ thuộc ADN do virus mã hoá.
Nhóm II. Virus ADN đơn (+) hoặc (-), tất cả đều phiên mã trong nhân và sử dụng ARN polymerase II của tế bào. Cả virus ADN (+) và ADN (-) khi phiên mã tạo mRNA đều phải qua giai đoạn trung gian ADN kép, dạng sao chép, viết tắt là RF (replicative form).
Nhóm III. Virus ARN kép, tất cả đều phiên mã trong tế bào chất, sử dụng enzyme ARN polymerase phụ thuộc ARN do virus mã hoá để tạo mRNA.
Nhóm IV. Virus ARN (+) có chức năng của mRNA trước khi phiên mã phải dich mã để tạo ARN polymerase phụ thuộc ARN của riêng mình sau đó mới phiên mã tạo mRNA.
Nhóm V. Virus ARN (-) không phân đoạn phiên mã trong tế bào chất, sử dụng ARN polymerase phụ thuộc ARN do chúng mang theo. Virus ARN (-) phân đoạn (ví dụ virus cúm) tuy phiên mã trong nhân nhưng chúng sử dụng ARN polymerase thụ thuộc ARN mang theo, vì tế bào không có enzyme này.
Nhóm VI. Virus retro phiên mã ngược, gồm 2 giai đoạn: Lúc đầu phiên mã genome ARN (+) thành cDNA (-) tiến hành trong tế bào chất nhờ enzyme phiên mã ngược của virus, sau đó ADN vào nhân tích hợp với nhiễm sắc thể của tế bào và tiến hành phiên mã tạo mRNA nhờ enzyme ARN polymerase phụ thuộc ADN của tế bào.
Nhóm VII. Virus ADN kép phiên mã ngược, ví dụ HBV, tiến hành phiên mã

trong nhân dùng enzyme ARN polymerase phụ thuộc ADN của tế bào để tạo ARN (+)
tiền genome, sau đó ra khỏi nhân, dùng enzyme phiên mã ngược của virus để phiên mã
ARN tiền genome thành ADN (-) rồi sau đó tạo genome ADN kép.

Tất cả các virus có genome ARN đều phiên mã trong tế bào chất trừ virus cúm.
Tất cả các virus có genome ARN (-) đều phải mang theo ARN polymerase phụ thuộc ARN để phiên mã.
Năm 1958 Fracis Crick đưa ra lý thuyết Trung tâm (Central Dogma), theo đó dòng thông tin di truyền luôn đi từ ADN mRNA Protein.
Những kiến thức về virus đã đưa đến sự cải biến lý thuyết trung tâm. Nhiều virus có genome ARN được sao thành ARN và một số có genome ARN được phiên mã ngược thành DNA.

Hình 1.2. Phiên mã từ ADN kép của eukaryota
1.2.2.3. Yếu tố phiên mã
1.2.2.2. Promoter và enhancer
Trong promoter của nhiều tế bào eukaryota và virus có trình tự chung (consensus) sau: TATAA/TAA/TA/G. Không phải promoter nào cũng có trình tự giống hệt nhau mà có thể sai lệch, nên gọi là trình tự chung. Trình tự TATAA còn gọi là hộp TATA hay hộp Pridnow nằm ở phía trước điểm khởi đầu phiên mã, khoảng 2530bp. Hộp TATA chịu trách nhiệm cho phép ARN polymerase gắn vào vùng khởi động phiên mã. Ví dụ: Hộp TATA có trong promotor duy nhất của HIV-1, nhưng trong số 4 promoter của HBV thì chỉ có 1 promoter chứa hộp này.
Enhancer (đoạn tăng cường) có trình tự bám vào yếu tố phiên mã. Sự tương tác này làm tăng tốc độ phiên mã bởi ARN polymerase II.
Yếu tố phiên mã là các protein gắn đặc hiệu vào promoter và enhancer để kiểm soát sự biểu hiện gen. Một số virus tạo ra yếu tố phiên mã của riêng mình, ví dụ VP/6 của virus herpes simplex, nó là một thành phần của virion và protein Tax của virus HTLV (virus gây ung thư tế bào T ở người) được tạo ra trong tế bào nhiễm.
Một số yếu tố phiên mã của tế bào cũng tham gia hoạt hoá hoặc kiềm chế sự phiên mã của các gen virus. Các yếu tố phiên mã đặc hiệu mô (tissue – specific transcription factors) cần cho một số virus mang tính đặc hiệu mô nghiêm ngặt, nghĩa là chúng chỉ nhân lên trong một số mô nhất định.
Một số yếu tố phiên mã của tế bào có tên là yếu tố phiên mã chung (general transcription factor) tham gia vào sự kiểm soát biển hiện gen của nhiều loại tế bào và virus. Ví dụ yếu tố phiên mã TFIID bám vào hộp TATA.
Hình 1.3. Sự bám của các yếu tố phiên mã và ARN pol II vào hộp TATA
TFIID là một phức hợp gồm 13 polypeptide, một trong số đó là protein bám hộp TATA. Sau khi TFIID bám vào hộp TATA là đến lượt các yếu tố phiên mã chung khác (TFIIA, TFIIB, TFIIE, TFÌI, TFIIH) và ARN pol II bám vào.
Trong số các yếu tố phiên mã của tế bào bám vào enhancer, có:
- AP1 và AP2 (activator protein 1 và 2) là các protein hoạt hoá.

- SP1 (stimulator protein) là protein kích thích.
- NF-kB (nuclear factor kB) là yếu tố nhân kB.
Hầu hết các yếu tố phiên mã này tham gia vào quá trình phiên mã của HIV-1. Cũng như hoạt hoá sự biểu hiện gen, các yếu tố phiên mã cũng tham gia vào sự kiềm chế biểu hiện gen.
Mọi cơ thể đều có khả năng tự điều hoà các gen của mình. Ví dụ côn trùng có các gen khác nhau và sự biểu hiện các gen này tuỳ thuộc việc chúng đang ở giai đoạn nào: ấu trùng, sâu non, sâu trưởng thành hay bướm. Ở virus cũng vậy, chúng có các gen khác nhau, biểu hiện ở các thời điểm khác nhau. Gen sớm mã hoá cho các protein sớm, đó là các protein không cấu trúc, thường là các enzyme và gen muộn mã hoá cho các protein muộn, đó là các protein cấu trúc, tham gia vào tạo capsid hoặc vỏ ngoài.
1.2.2.4. Biến đổi ARN sau phiên mã
1.2.2.4.1. Gắn mũ vào ARN
Ngay sau khi tổng hợp ARN và trong lúc quá trình phiên mã còn đang tiếp tục thì hầu hết các bản phiên mã (RNA) đã được gắn mũ ở đầu 5’. Mũ là guanosine triphosphate nối với một nucleotide ở đầu 5’ nhờ liên kết 5’-5’, thay vì liên kết 5’-3’ như bình thường.
Hầu hết mRNA ở tế bào eukaryota và ở virus đều có mũ ở đầu 5’.
Mũ có chức năng:
- Giúp mRNA vận chuyển từ nhân ra tế bào chất.
- Bảo vệ mRNA khỏi sự thuỷ phân của enzyme exonuclease.
- Cần cho khởi đầu cho dịch mã.
Enzyme thực hiện gắn mũ của tế bào là guanylyl transferase (thêm guauosin 5’triphosphate) và metyl transferase (thêm nhóm metyl). Các enzyme này nằm trong nhân, nên hầu hết các virus phiên mã trong nhân đều sử dụng. Tuy nhiên các virus phiên mã trong tế bào chất sẽ mã hoá cho enzyme gắn mũ và enzyme metyl hoá của riêng mình.
Virus có genome ARN (-) phân đoạn (cúm A) cần có một cơ chế riêng để đoạt lấy mũ từ mRNA của tế bào. Phức hợp protein của virus giúp ARN polymerase của nó gắn vào đầu 5’ của mRNA đã gắn mũ của tế bào, sau đó nhờ hoạt tính endonuclease của phức hợp sẽ cắt một đoạn khoảng 10-20 nucleotide (chứa cả mũ) ra khỏi đầu 5’ của mRNA. Đoạn này sẽ được dùng làm mồi để khởi đầu phiên mã cho virus.
Không phải tất cả mRNA virus đều có gắn mũ, mRNA của virus picorna (bại liệt) không có mũ Đây là virus phiên mã trong tế bào chất, nên không phải vận chuyển ra khỏi nhân. Chúng có thể tiến hành dịch mã theo cơ chế không phụ thuộc mũ, mà nhờ đoạn IRES.
1.2.2.4.2. Gắn đuôi poly (A)
Các bản phiên mã đầu tiên của eukaryota và virus được ngắn thêm một loạt gốc adenosin ở đầu 3’ để tạo đuôi poly (A). Sự polyadenin hoá ở đầu 3’ làm tăng tính bền của mRNA tránh sự phân giải của exonuclease và có vai trò nhất định trong khởi đầu dịch mã.
Tuy nhiên mRNA của một số virus không gắn đuôi (ví dụ virus reo) nên chức năng kể trên sẽ được thực hiện theo cách khác.
Trước vị trí gắn đuôi có một đoạn tín hiệu gắn đuôi dài 20-30bp. Đoạn này ở SV40 là AATAAA, ở HBV là TATAAA (giống như hộp TATA). Khi phiên mã ARN polymerase tiếp tục chạy dọc khuôn, vượt qua tín hiệu gắn đuôi và vị trí gắn đuôi. Sợi ARN mới tổng hợp được cắt tại vị trí gắn đuôi và các gốc adenosin được lần lượt thêm vào nhờ phức hợp protein, trong đó có enzyme gắn đuôi (poly (A) -polymerase).
1.2.2.4.3. Cắt nối
Một số bản phiên mã có chức năng của mRNA nhưng hầu hết còn cần phải chế biến, cắt bỏ intron để tạo mRNA. Hơn nữa từ mRNA, bằng phương thức cắt nối (ghép đoạn này với đoạn kia), có thể tạo ra nhiều loại mRNA khác nhau. Ví dụ bằng phương pháp cắt nối, HIV-1 có thể tạo ra 30 loại mRNA khác nhau.
1.2.3. Dịch mã của virus
Tham gia vào dịch mã gồm mRNA, tRNA, axit amin và riboxom. Quá trình gồm 3 giai đoạn: Khởi đầu, kéo dài và kết thúc.
Ở eukaryota, mRNA là đơn gen (monocistron) có nghĩa là chỉ có một khung đọc, mã hoá cho 1 protein, còn ở prokaryota, mRNA thường là đa gen (polycistron) có nhiều khung đọc và mã hoá cho nhiều protein.

Có các vùng không mã hoá, còn gọi là vùng không dịch mã, nằm ở phía đầu 5’ (5’-UTR) và đầu 3’ (3’-UTR).
Một khung đọc lớn có thể mã hoá cho một polyprotein lớn sau đó được enzyme phân cắt thành các protein nhỏ với chức năng khác nhau.
Tiểu phần riboxom 40S di chuyển dọc theo ARN theo hướng 5’ 3’ và quét cho đến khi gặp codon khởi đầu AUG ở đầu 5’. Tuy nhiên, một số virus có codon khởi đầu khác. Ví dụ virus Sendai dùng codon khởi đầu là ACG cho một số gen.
Hình 1.4. Dịch mã từ mRNA-monocistron
Một khung đọc mở bắt đầu từ codon khởi đầu AUG ở đầu 5’ của mRNA và kết thúc tại codon kết thúc (UGA, UAA, UAG). Dịch mã theo hướng 5’ 3’. Đầu N của protein được tổng hợp trước.
1.2.3.1. Khởi đầu dịch mã
Như đã nói trên, hầu hết mRNA của eukaryota và virus được gắn mũ và đuôi.
Các cấu trúc này có vai trò quan trọng trong dịch mã.
- Mũ là nơi bám của các yếu tố khởi đầu dịch mã, eIF (e = eukaryota), tRNA gắn với axit amin (Met-tRNAi Met) và tiểu phần riboxom 40S (hình 6.)
- Một protein gọi là protein bám poly (A) bám vào đuôi poly (A). Các protein bám vào đầu sợi ARN có khả năng tương tác và người ta cho rằng sự tương tác này làm cho mRNA truyền tin dẫn đến sự kích thích dịch mã.
Các mRNA không có mũ hoặc đuôi có thể truyền tin theo cách khác.
Hình 1.5. Khởi đầu dịch mã trên m ARN có gắn mũ
a – Các yếu tố khởi đầu eIF, tiểu phần riboxom 40S và t RNA-methionine bám vào đầu 5’ của mRNA, b - Phức hợp quét từ đầu 5’ theo hướng 3’ của mRNA, c – khi tới codon khởi đầu AUG sẽ được anticodon UAC trên tRNA nhận diện, tiểu phần 60S bám vào và các yếu tố eIF sẽ rời ra.
Một số mRNA không có mũ thì sự dịch mã xảy ra theo cơ chế khác. Các eIF thay vì gắn vào mũ, chúng gắn vào vị trí IRES (internal ribosome entry site) gọi là bến đỗ riboxom, đó là đoạn mRNA có cấu trúc bậc 2 ở mức độ cao nằm trước khung đọc, cũng là vị trí gắn của tiểu đơn vị riboxom 40S. Đoạn IRES có ở một số virus RNA, như virus viêm gan C, picorna, đồng thời cũng thấy có ở mRNA của tế bào và mRNA của một số virus khác (ví dụ virus herpes simplex, Sarcoma Rous).

Hình 1.6. Khởi đầu dịch mã trên mRNA không gắn mũ.
Tiểu phần roboxom 40S, các eIF bám vào vị trí IRES (bến đỗ riboxom) nằm phía trước khung đọc.
1.2.3.2. Dịch mã từ m ARN hai khung đọc
Hầu hết mRNA của tế bào và của nhiều virus chỉ có 1 khung đọc mở (ORF), nhưng một số mRNA của virus có 2 hoặc nhiều khung đọc, trong số các mRNA bicistron hoặc polycistron này có một số có chức năng là monocistron, nhưng một số mRNA có cấu trúc là bicistron thì cũng có chức năng là bicistron. Sự khác nhau về tỷ lệ dịch mã của hai khung đọc quy định cơ chế biểu hiện 2 gen ở mức độ khác nhau. Ở nhiều mRNA bicistron có khung đọc chồng lớp. Ở trường hợp khác, khung đọc này nằm gọn trong khung đọc kia.

- Một cơ chế dùng để đọc khung thứ 2 gồm sự quét bỏ sót (leaky): Tiểu đơn vị riboxom 40S có thể quét bỏ qua codon khởi đầu của khung đọc thứ nhất và bắt đầu dịch mã tại điểm khởi đầu của khung đọc thứ 2. Các khung đọc cho hai protein là khác nhau nên hai protein do chúng mã hoá là khác nhau.
Hình 1.7. Sự dịch mã của mRNA bicistron.
– Riboxom có thể dịch mã tại điểm khởi đầu của ORF-1 hoặc có thể quét qua codon khởi đầu của ORF-2.
– Hai codon khởi đầu là của 2 khung đọc khác nhau và protein được tổng hợp là khác nhau.c – Khung đọc 2 được dịch mã là nhờ sự dịch khung riboxom, tạo ra một dạng kéo dài của protein 1.
- Một cơ chế khác để đọc khung đọc thứ hai của mRNA là sự dịch khung (frameshifting) của riboxom: Riboxom nhảy sang khung đọc khác nằm trước điểm kết thúc của khung đọc thứ 1. Do đó nó không bị codon kết thúc của khung đọc thứ nhất nhận ra, mà tiếp tục chạy dọc mRNA để đọc khung đọc thứ nhất kéo dài (hình 19.7c). Sự dịch khung xảy ra khi riboxom di chuyển dọc theo ARN bắt gặp tín hiệu dịch khung (1 trình tự chuyên biệt) có cấu trúc bậc 2, và thường là một pseudoknot (nút giả).
1.2.3.3. Cải biến trong và sau dịch mã
Trong hoặc sau quá trình dịch mã, protein có thể phải qua một hoặc nhiều lần cải biến, bao gồm glycosyl hoá, acyl hoá hoặc phosphoryl hoá.
-Glycosyl hoá

Glycosyl hoá là sự gắn thêm nhóm oligosaccarit vào chuỗi polypeptide. Khi một oligosaccarit được gắn vào nhóm –OH của serin hoặc treonin thì gọi là O-glycosyl hoá, còn nếu gắn vào nhóm -NH2 của asparagin thì gọi là N-glycosyl hoá. Protein
được tổng hợp trong mạng lưới nội chất hạt, nơi bắt đầu N-glycosyl hoá, sau đó chúng
được chuyển tới bộ máy Golgi, nơi hoàn thiện N-glycosuyl hoá nhờ enzyme αmannosidase I và II, và galactosyl transferase. O-glycosyl hoá được thực hiện trong bộ máy Golgi.
Có những virus chỉ cần một loại glycosyl hoá, ví dụ HIV-1 tiến hành Nglycosyl hoá gp120 nhưng cũng có những virus cần cả 2 loại N- và O-glycosyl hoá, ví dụ gC và gD của herpes simplex. Các glycoprotein này được gắn vào vị trí chuyên biệt trên màng sinh chất của tế bào và sẽ là các gai bề mặt sau này của vỏ ngoài. Virus rota không có vỏ ngoài nhưng cũng có glycoprotein. Protein bề mặt VP7 là glycoprotein.
Một số glycoprotein của virus là protein không cấu trúc, ví dụ protein NSP4 của virus rota.
-Acyl hoá
Acyl hoá là sự gắn thêm nhóm acyl (R-CO-) vào phân tử protein. Nhóm acyl thường được gắn là myristyl, trong đó R là CH3-(CH2)12- nhóm myristyl được gắn vào glycin tại đầu N của protein. Hầu như virus không có enzyme N-myristyl transferase để gắn nhóm myristyl, chúng phải sử dụng của tế bào.
Protein myristyl hoá gắn vào màng sinh chất, ví dụ protein Gag của hầu hết virus retro. Nếu không được myristyl hoá thì chúng không gắn được vào màng sinh chất và sự lắp ráp virus sẽ không xảy ra.
- Phosphoryl hoá
Phosphoryl hoá là sự chuyển nhóm phosphate từ một nucleotide, thường là ATP, cho oxy ở nhóm –OH của serin, treonin hoặc tyrosin. Sự chuyển nhóm phosphate nhờ enzyme kinase của tế bào hoặc của virus.
Sự phosphoryl hoá có thể làm thay đổi cấu hình, hoạt tính, vị trí và độ bền của protein. Nhiều quá trình của virus đòi hỏi protein phải được phosphoryl hoá.
- Sự vận chuyển các phân tử trong tế bào eukaryota
Các phân tử của virus được vận chuyển tới các vị trí nhất định trong tế bào.
Genome của virus được đưa vào nhân nhờ trượt theo vi ống và qua lỗ nhân, mRNA virus được vận chuyển từ nhân ra tế bào chất và các protein virus sau khi được tổng hợp có thể được vận chuyển vào các vị trí khác nhau, kể cả vào nhân (Các virus lắp ráp trong nhân).
Nhiều protein có một trình tự axit amin giống như mã số vùng của bưu điện, nó quy định điểm đến của protein. Protein đến gắn vào màng có một trình tự tín hiệu – đó là dãy các gốc axit amin kỵ nước. Sự tổng hợp protein bắt đầu tại các riboxom tự do, nhưng khi trình tự tín hiệu được tổng hợp thì nó sẽ hướng dẫn phức hợp polypeptideriboxom đi vào mạng lưới nội chất và tiếp tục tổng hợp protein tại đó. Vùng mạng lưới nội chất có chứa riboxom vì thế gọi là mạng lưới nội chất sần.
Mỗi một loại protein gắn vào màng có một hoặc nhiều trình tự gắn vào màng, gọi là trình tự neo màng (membrane anchor sequences) giàu gốc axit amin kỵ nước. Một số protein loại này có trình tự tín hiệu hoạt động như cái mỏ neo cắm vào màng. Các protein gắn vào màng khác, ví dụ protein vỏ ngoài của HIV-1 sẽ được chuyển qua màng cho đến khi chạm tới trình tự neo và sau đó trình tự tín hiệu sẽ bị enzyme của tế bào loại bỏ
Nhiều protein được tổng hợp trong mạng lưới nội chất hạt sẽ được tạo bọng để tới bộ máy Golgi. Hầu hết protein gắn màng được glycosyl hoá trong các khoang màng này, sau đó được chuyển tới các màng tế bào chất hoặc màng nhân và có thể nẩy chồi qua các màng này.
Các tế bào biểu mô có mặt đỉnh (ở phía ngoài) và mặt đáy (ở phía trong),
được cấu tạo từ lipid và protein. Khi nhiễm virus có vỏ ngoài vào tế bào biểu mô chúng sẽ nảy chồi qua màng sinh chất một cách giới hạn hoặc qua mặt đỉnh hoặc qua mặt đáy. Ví dụ nếu là VSV (virus chốc mép), chúng sẽ nẩy chồi từ mặt đáy, còn virus cúm A sẽ nảy chồi từ mặt đỉnh. Điều này giải thích vì sao virus cúm nhiễm vào động vật có vú đều khu trú ở đường hô hấp.
Nếu virus nhân lên ở trong nhân thì hầu hết (nhưng không phải tất cả) các protein virus phải được đưa vào nhân. Cơ chế vận chuyển liên quan đến sự nhận diện của tín hiệu định vị nhân (NLS-nuclear localization signal) nằm trên protein của tế bào, gọi là importin, sau đó gắn vào các sợi (fibril) mọc ra từ phức hợp lỗ nhân để vận chuyển qua lỗ nhân.
1.3. Câu hỏi bài tập về cơ chế di truyền ở virut
Câu 1(trích đề thi học sinh giỏi quốc gia năm 2015): Hãy phân biệt quá trình phiên mã và quá trình sao chép trong tế bào chủ của virut chứa hệ gen ARN+ với virut chứa hệ gen AND về: nơi phiên mã, enzim dùng trong phiên mã, nơi sao chép, enzim dùng trong sao chép. Quá trình phiên mã có trùng với quá trình sao chép không?
Virut ARN (+)
Virut ADN
Nơi phiên mã Trong tế bào chất Trong nhân tế bào
Enzim dùng cho phiên
ARN polimeraza phụ thuộc ARN của virut
ARN polimeraza phụ thuộc ADN của tế bào
344
Nơi sao chép Trong tế bào chất Trong nhân tế bào
Enzim dùng cho sao chép
ARN polimeraza phụ thuộc ARN của tế bào
ADN polimeraza phụ thuộc ADN của virut
Ở virut chứa hệ gen ARN(+), quá trình phiên mã trùng với quá trình sao chép còn ở virut chứa hệ gen ADN quá trình phiên mã không trùng với quá trình sao chép.
Câu 2: Hệ gen của các retrovirut (như HIV) và virut ADN kép (như HBV) đều có enzim phiên mã ngược nhưng chúng lại có cơ chế tái bản vật chất di truyền khác nhau. Em hãy chỉ ra những điểm khác biệt đó.
Trả lời Virut retro phiên mã ngược(HIV) Virut ADN kép phiên mã ngược (HBV)
B1: Sử dụng enzim phiên mã ngược của virut để tổng hợp ADN kép trong tế bào chất
B2: ADN kép tích hợp vào NST trong nhân rồi từ đó phiên mã tạo ARN nhờ nzim của tế bào chủ
B1: Sử dụng enzim phiên mã của tế bào để tổng hợp tiền genom virut là ARN (+) trong nhân tế bào
B2: Ra tế bào chất, dùng enzim phiên mã ngược của virut để phiên mã ARN (+) thành ADN (-) rồi sau đó tạo ADN kép.
Câu 3: Một loại virut không có vỏ ngoài, có vật chất di truyền là ARN đơn, sợi (-). Khi xâm nhập vào tế bào, virut này sẽ tái bản hệ gen và tổng hợp prôtein cấu trúc của nó như thế nào? Trả lời
Gồm 3 bước:
- Bước 1: Tống hợp sợi ARN(+) bổ sung với sợi ARN(-) của virut.
- Bước 2: Dùng sợi ARN(+) làm khuôn để tái bản hệ gen ARN(-) của virut.
- Bước 3: Dùng sợi ARN(+) làm mARN để tổng hợp các protein cấu trúc của virut
Câu 4: Mặc dù virut HIV và virut HBV có vật chất di truyền là khác nhau nhưng sau khi xâm nhập vào tế bào người, chúng đều tổng hợp ADN để có thể cài xen vào hệ gen của người. Em hãy nêu những điểm giống nhau trong quá trình tổng hợp ADN của chúng.
Trả lời
- Diễn ra trong tế bào chất.
- Sử dụng enzim phiên mã ngược AND polymeraza phụ thuộc ARN của virut.
- Sử dụng các nucleootit, ATP, các enzim khác của tế bào chủ.
- Sử dụng ARN của virut để tổng hợp AND mạch kép.
Câu 5: Virus chứa hệ gen ARN (+) và virus chứa hệ gen ADN (ký sinh ở tế bào nhân thực) có sự khác biệt như thế nào về: vị trí, enzyme dùng cho hai quá trình phiên mã và tổng hợp vật chất di truyền?
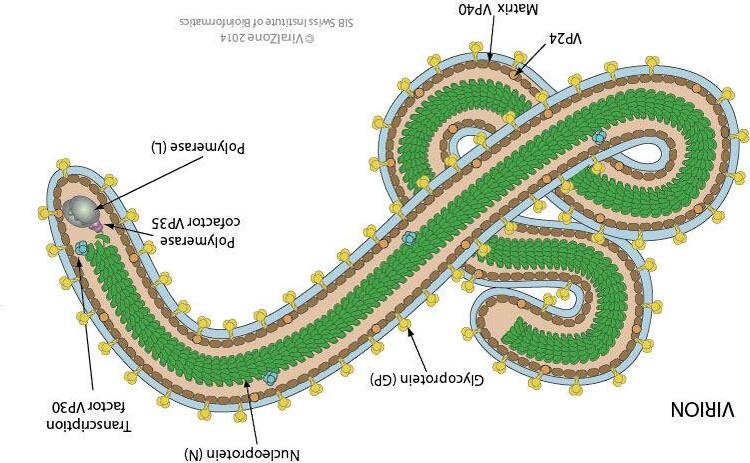
Virus ARN (+) Virus ADN (mạch kép)

Nơi phiên mã Trong tế bào chất Trong nhân tế bào
Enzyme dùng cho phiên mã
Nơi tổng hợp VCDT
Enzyme dùng cho tổng hợp VCDT
ARN polimeraza phụ thuộc ARN của virus
ARN polimeraza phụ thuộc ADN của tế bào
Trong tế bào chất Trong nhân tế bào
ARN polimeraza
phụ thuộc ARN của tế bào
ADN polimeraza phụ thuộc ADN của virus
Câu 6: Cho sơ đồ cấu trúc hai loại virut: virut zika và virut ebola như hình dưới đây:
Sơ đồ cấu trúc virut zika
Sơ đồ cấu trúc virut ebola
Hãy trình bày những điểm khác nhau về hình thái, cấu trúc và quá trình nhân lên của hai loại virut này?
Đặc điểm phân biệt
Virut zika Virut ebolaHình thái Cấu trúc khối: các capsome xếp thành 20 mặt tam giác đều
Cấu trúc Chứa ARN sợi đơn, được dùng như mARN (hay còn gọi là ARN +)
Cấu trúc xoắn: Capsome xoắn theo chiều xoắn của axit nucleic
Chứa ARN sợi đơn, được dùng để tổng hợp mARN (vì trong cấu trúc có enzim polymerase)
- HPV: ADN kép nhân đôi trong nhân theo cơ chế bán bảo tồn, sử dụng ADN polimeraza của tế bào
- HBV: AND nhân đôi theo 2 giai đoạn:
+ ADN ARN diễn ra trong nhân, sử dụng ARN polimeraza của tế bào
+ ARN ADN diễn ra trong tế bào chất, sử dụng enzim phiên mã ngược của virut
Quá trình nhân lên
Sao chép trong tế bào chất: ARN+ đóng vai trò mARN tham gia dịch mã tổng hợp ARN polimerase, ARN polimerase xúc tác tổng hợp ARN-, sau đó từ ARN- tổng hợp mARN để mã hóa vò capsit và cũng từ ARNlàm khuôn để tổng hợp ARN+ (lõi của virut)
Sao chép trong tế bào chất: Sử dụng ARN polimerase do chúng mang theo để tổng hợp ARN+, từ ARN + vừa làm khuôn tổng hợp vỏ capsit vừa làm khuôn tổng hợp ARN- (lõi của virut)
Câu 7: Hệ gen của các retrovirut (như HIV) và virut ADN kép có enzim phiên mã ngược (như HBV) đều có enzim phiên mã ngược nhưng chúng lại có cơ chế tái bản vật chất di truyền khác nhau. Em hãy chỉ ra những điểm khác biệt đó.
Virut retro phiên mã ngược (HIV)
Virut ADN kép phiên mã ngược (HBV)
Hệ gen: 2 sợi ARN (+) giống nhau Hệ gen: ADN kép
B1: Sử dụng enzim phiên mã ngược của virut (ADN-pol phụ thuộc virut) để tổng hợp ADN kép (cADN) trong tế bào chất
B2: ADN kép tích hợp vào NST trong nhân rồi từ đó phiên mã tạo ARN nhờ enzim của tế bào chủ (ARN pol phụ thuộc ADN của tế bào).
B1: Sử dụng enzim phiên mã của tế bào (ARN pol phụ thuộc ADN của tế bào) để tổng hợp tiền genom virut là ARN (+) trong nhân tế bào
B2: Ra tế bào chất, dùng enzim phiên mã ngược của virut (ARN-pol phụ thuộc virut) để phiên mã ARN (+) thành ADN (-) rồi sau đó tạo ADN kép.
Câu 8: HPV và HBV là hai loại virut đều có vật chất di truyền là phân tử ADN kép, tuy nhiên HBV có enzim phiên mã ngược. Quá trình tổng hợp vật chất di truyền của hai virut này trong tế bào chủ có gì khác nhau? Trả lờ
2.1. Khái quát cơ chế điều hòa hoạt động của của Operon đồng hóa, dị hóa ở vi khuẩn
Bộ gen của vi khuẩn được tổ chức theo operon. Một operon có một vùng điều khiển và vùng mã hóa. Vùng điều khiển bao gồm gen điều hoà, promoter và điểm điều hành (operator). Vùng mã hóa bao gồm một số gen cấu trúc nằm kề nhau, mỗi gen cấu trúc mã hóa cho một polypeptide. Các prôtêin được mã hóa trong một operon thường có quan hệ mật thiết với nhau trong một quá trình chuyển hóa sinh hóa nào đó trong tế bào.
Một operon có ít nhất một promoter, tuy nhiên, operon có thể có nhiều hơn một promoter và ái lực của các promoter này đối với ARN-polymerase là khác nhau. Operator là trình tự ADN, nơi mà prôtêin ức chế (repressor prôtêin) gắn vào.
Khái niệm về operon được F. Jacob và các cộng sự của ông nêu ra năm 1961
khi nghiên cứu sự kiểm soát di truyền hấp thụ đường lactose ở E.coli. Với sự khám phá ra cơ chế điều hoà biểu hiện gen, ông đã nhận được giải thưởng Nobel 1965.
Cơ chế điều hòa di truyền của hệ thống lac (lac system) được giải thích bằng
mô hình operon của Francois Jacob và Jacque Monod (1960)
2.1.1. Các khái niệm
2.1.1.1. Gen cơ định (constitutive gene)
Trong hệ gen, cụ thể là trong quá trình chuyển hóa đường glucose, tổng hợp axit amin,… cần có các thành phần tham gia như tARN, rARN, protein cấu thành nên riboxom, ADN polimerase,… và các enzim tham gia vào quá trình trao đổi chất cơ bản thiết yếu cho sự cần thiết của tế bào. Gen đóng vai trò mã hóa cho các thành phần này hoạt động thường xuyên và liên tục ở hầu hết các tế bào được gọi là gen cơ định.
Các cơ thể có ưu thế chọn lọc cao hơn thường có cơ chế điều hòa sao cho các gen tổng hợp những sản phẩm không được sử dụng thường xuyên hoạt động ít hơn và hầu hết chỉ hoạt động trong thời gian cần thiết của tế bào và trong điều kiện môi trường nhất định để nhằm mục đích không tốn năng lượng của tế bào. Đây cũng là một trong những bằng chứng của tiến hóa sau hàng triệu năm tiến hóa của tế bào.
Các ATP là nguồn năng lượng thiết yếu của tế bào, nó có thể mất một cách nhanh chóng và cũng khó để tổng hợp một cách dễ dàng trong điều kiện tự nhiên khắc nghiệt. Vì vậy, mọi dạng sống hiện nay bao gồm cả vi khuẩn và virus đã biết cách “tiết kiệm” chúng bằng các cơ chế biểu hiện gen rất hiệu quả.
2.1.1.2. Sự cảm ứng, gen cảm ứng- sự giải nén, gen bị ức chế:
- Với một chiếc điện thoại thông thường hay smartphone bình thường thì chúng đều ở trạng thái tắt hoặc “ngủ” thực ra thì vẫn xảy ra các quá trình hao tốn pin
nhưng các quá trình này là không đáng kể. Khi xảy ra các hoạt động chạm vào màn hình, hay bấm bàn phím dưới một lực tác động chúng có thể hoạt động một cách mạnh mẽ, hoạt động một cách bất bình thường.
Cũng với hành động như vậy, khi vi khuẩn E.coli được chuyển vào môi trường có lactose làm nguồn hidrat carbon, thì chúng nhanh chóng trong vòng 10 phút tồng hợp các enzym chuyển hóa và sử dụng lactose. Đó là kết quả của việc “bật” sự biểu hiện của gen để đáp ứng lại sự có mặt của tín hiệu nào đó trong môi trường được gọi là sự cảm ứng.
- Các gen được điều hòa theo quá trình “bật”, “tắt’ được gọi là gen cảm ứng (inducible genes). Sản phẩm do gen như vậy mã hóa, nếu là enzym thì gọi là các enzym cảm ứng. Nếu là protein được gọi là protein cảm ứng. Các enzym cảm ứng thường là các enzym tham gia vào quá trình dị hóa.
- Ngược lại, khi sản phẩm tổng hợp một cách dư thừa quá mức hoặc trong môi trường có xảy ra việc dư thừa chất cần thiết cho cơ thể thì tế bào có sự điều hòa ngược lại đó là sự giải nén. Sự giải nén đơn giản là việc “tắt” sự biểu hiện của gen. Các gen được điều hòa theo kiểu này được gọi là các gen bị ức chế (hay gen bị nén). Các enzym tổng hợp từ gen bị ức chế tham gia vào quá trình đồng hóa.
2.1.1.3. Điều hòa dương tính và điều hòa âm tính
- Điều hòa âm tính và dương tính là hai quá trình đi liền với nhau và không thể thiếu trong quá trình ức chế hay kể cả quá trình kích ứng. Cả hai công việc này đều liên quan đến gen điều hòa. Các gen điều hòa này cho sản phẩm trực tiếp điều hòa hoạt động của các gen khác. Trong cơ chế điều hòa dương tính, sản phầm của gen điều hòa có vai trò làm tăng sự biểu hiện của một hay một số gen cấu trúc. Còn trong quá trình điều hòa âm tính, sản phẩm của gen điều hòa thường ức chế hoặc làm tắt sự biểu hiện của gen cấu trúc.
- Các gen điều hòa hoạt động do có các enzym liên kết vào các protein của chúng (protein điều hòa). Vị trí đó được gọi là vị trí liên kết protein điều hòa (RPBS) thường nằm gần promoter của gen mà nó điều hòa.
- Dựa theo hai cơ chế điều hòa thì cũng có hai loại protein gọi là protein hoạt hóa trong hệ thống điều hòa dương tính và protein ức chế trong hệ thống điều hòa âm tính. Không phải protein điều hòa muốn liên kết vào RPBS là được ngay mà cần phải có một phân tử trung gian liên kết cũng giống như muốn các ốc vít được gắn kết chặt hơn thì cần có các goong ở giữa nhằm các ốc đỡ bị chờn ra ngoài. Các phân tử như vậy được gọi là các phân tử tín hiệu. Cũng như các khái niệm khác thì phân tử tín hiệu cũng có hai loại phù hợp với chức năng của chúng, nếu phân tử làm tăng cường sự biểu hiện của gen thì gọi là phân tử kích ứng, còn nếu hạn chế hay kìm hãm sự biểu hiện gen thì gọi là phân tử dồng ức chế.
- Các phân tử này gắn kết vào protein điều hòa và làm thay đổi cấu hình của chúng, dẫn đến sự thay đổi hoạt tính hoặc chức năng của các protein. Việc gắn kết và
350
có kết quả như vậy được gọi là sự biến đổi dị hình. Việc này xảy ra là do sự thay đổi ái lực liên kết của chúng vào các vị trí RPBS ở gần promoter của các gen mà chúng điều khiển.
2.1.2. Các nguyên tắc
2.1.2.1. Biểu hiện gen được điều khiển bởi các protein điều hòa
- Sự biểu hiện gen ở sinh vật nhân sơ hay kể cả sinh vật nhân thực đều phụ thuộc vào các yếu tố môi trường, cụ thể là các tín hiệu từ môi trường. Ở vi khuẩn, điển hình là các phân tử có mặt trong môi trường sống. Và các thông tin từ phân tử tín hiệu thì được chuyển tới gen thông qua các protein điều hòa. Như ta đã biết ở quá trình phiên mã, các ARN polimerase liên kết vào promoter và làm cho phức hệ đang “đóng” chuyển thành ‘mở” và dãn xoắn mạch. Sau đó, ARN polimerase tiếp tục quá trình phiên mã. Điều này chứng minh rằng nó phụ thuộc vào promoter và protein điều hòa.
2.1.2.2. Nhiều promoter được điều khiển bằng ái lực giữa ARN polimerase với
phiên mã đóng chuyển thành mở. Còn trong trường hợp metT, chất hoạt hóa (MerR) tạo ra hiệu ứng tương tự nhưng bằng sự thay đổi cấu hình ADN (promoter).
Qua đây ta thấy được việc điều hòa phiên mã rất đa dạng và phong phú. Ta có thể kết hợp đồng thời nhiều cơ chế khác nhau hoặc sự ức chế phiên mã không nhất thiết phải trải qua sự ngăn cản liên kết giữa ARN pol và promoter.
2.1.2.4. Sự biểu hiện gen từ khoảng cách xa và cấu trúc vòng thắt ADN
ADN
- Bình thường, khi promoter liên kết với ARN polimearase chúng có liên kết yếu làm cho việc khởi đầu phiên mã không thế diễn ra một cách bình thường. Nhưng đôi khi quá trình này vẫn xảy ra theo biểu hiện gen kiểu cơ định, nhưng chỉ ở mức tối thiểu. Còn khi có protein điều hòa gắn giữa ARN polimerase và promoter quá trình phiên mã sẽ diễn ra một cách suôn sẻ bởi liên kết cao hơn do được bổ sung các yếu tố của trình tự liên ứng. Đây được gọi là việc giới hạn tốc độ biểu hiện gene.
- Để điều hòa âm tính một promoter cần có một protein ức chế gắn vào một vị trí bên trong hoặc gối lên promoter. Vị trí này gọi là trình tự điều hành (operator).
Protein này gắn vào operator, ARN polimearase sẽ không gắn được vào promoter và sự phiên mã khoog được thực hiện. Ngược lại, để điều hòa dương tính, một protein điều hòa sẽ giúp ARN pol dễ dàng liên kết với promoter. Hiện tượng này xảy ra bằng cách protein hoạt hóa dùng một phần phân tử của nó để liên kết với ADN tại vị trí gần promoter, còn phần khác tương tác với ARN pol, nhờ vậy đưa enzym đến gần promoter hơn. Cơ chế này gọi là sự liên hợp ADN/protein. Vai trò của protein chỉ như một chất đính kết gắn ADN và ARN pol hay đưa enzym đến gần promoter hơn. Và quá trình mở có thể dễ dàng xảy ra, dẫn đến sự phiên mã diễn ra.
- Operon lac ở vi khuẩn E.coli là một điển hình hoạt động với một protein họat hóa và một protein ức chế
2.1.2.3. Quy tắc dị hình
Quy tắc dị hình là một trong những quy tắc cơ bản nhất trong điều hòa chức năng và hoạt tính protein. Ví dụ: các protein Cyclin hoạt hóa các enzym Cdk tham gia điều khiển chu trình tế bào. Các Cyclin hoạt hóa được enzym này bằng cách liên kết vào enzym và chuyển chúng từ trang thái không hoạt động sang trạng thái hoạt động. Ở glnA, chất hoạt hóa (NtrC) tương tác với ARN pol và thúc đẩy phức hệ khởi đầu
- Các protein điều hòa thường liên kết ở vị trí gần promoter của gen, nhưng điều này không phải lúc nào cũng đúng. Đôi khi protein điều hòa liên kết vào ADN ở vị trí cách gen rất xa. Lúc này, để protein điều hòa có thể tương tác được với gen, thường có sự hình thành các vòng thắt trên phân tử ADN. Trong thực tế, protein hoạt hóa phiên mã NtrC hoạt động theo cách này. Vị trí liên kết ADN của nó nằm ngược dòng promoter (về phía đầu 5’ của mạch mã hóa) 150 bp. Protein này thậm chí có thể hoạt hóa gen ngay cả khi nó liên kết ở vị trí xa hơn (1 kb hoặc hơn). Trong việc điều khiển như vậy, vòng thắt có thể dài tới 3 kb. Một cách khác để đưa các ADN đến gần nhau là nhờ các protein liên kết vào đoạn trình tự ở giữa. Đây là hai cách phổ biến ở sinh vật nhân sơ, còn ở sinh vật nhân thực ta còn thấy có nhiều cách phong phú hơn.
2.1.2.5. Quy tắc dị hình và liên kết ADN – protein
Sự tương tác giữa các đại phân tử nói chung và giữa ADN và ARN pol nói riêng đã hoạt hóa gen từ đó huy động bộ máy phiên mã tới promoter. Trong những trường hợp khác thì gen lại phiên mã theo quy tắc dị hình. Tức là, môt protein liên kết với enzym ARN pol đã liên kết sẵn với ADN và làm thay đổi cấu hình hoặc của enzym và ADN hoặc của promoter, qua đó sự khởi đầu phiên mã có thể diễn ra.
Cả hai quá trình này đều có những vao trò khác nữa trong điều hòa hoạt động của gen và cách tương tác giữa chúng có thể phức tạp hơn. Ví dụ như hai hay nhiều protein điều hòa cùng tương tác với nhau và với ADN. Qua đó chúng hỗ trợ nhau cùng tăng cường liên kết vào vị trí gen được điều khiển. Kiểu tương tác này tạo nên tác dụng hợp lực và có thể nhanh chóng chuyển một gen từ trạng thái không biểu hiện sang trạng thái biểu hiện tối đa. Sự hợp lực của các protein hoạt hóa còn có vai trò tích hợp tín hiệu. Đó là khi các gen chỉ biểu hiện khi đồng thời có mặt nhiều tín hiệu khác nhau. Sự biểu hiện gen ở phago là một ví dụ điển hình cho kiểu điều hòa này.
2.1.2.6. Khởi đầu phiên mã không phải là cơ chế điều hòa duy nhất ở các gen
Sự điều hòa phiên mã không chỉ xảy ra ở bước khởi đầu phiên mã mà còn ở các giai đoạn khác như kéo dài phiên mã, hoàn thiện mARN, dịch mã tổng hợp protein.
2.2. Điều hòa operon tham gia con đường dị hóa ở vi khuẩn
2.2.1. Điều hòa âm tính Operon lactose ở E.coli
Vi khuẩn E.coli có khả năng sử dụng đường lactose như nguồn cacbon chính để phát triển. Lactose là một disacarit, nên khi có mặt lactose trong môi trường, vi khuẩn sản sinh ra enzym β-galactosidase để thuỷ phân lactose, tạo thành các phân tử
monosacarit là glucose và galactose thuận lợi cho quá trình chuyển hóa. Ngoài βgalactosidase ra, tế bào vi khuẩn còn tổng hợp enzym permease làm nhiệm vụ vận chuyển lactose qua màng tế bào. Khi nghiên cứu sự kiểm soát di truyền hấp thụ đường lactose, F. Jacob đã phát hiện cơ chế tổng hợp các enzym hấp thụ đường lactose. Quá trình thí nghiệm có thể mô tả như sau
Khi nuôi vi khuẩn E.coli trên môi trường không có lactose, thì ta thấy nồng độ enzym β-galactosidase và permease do vi khuẩn sinh ra rất thấp: khi đưa đường lactose vào môi trường nuôi thì nồng độ hai enzym này ở tế bào vi khuẩn tăng lên rất nhiều.
Nhưng nếu nuôi vi khuẩn trên môi trường có đường glucose và lactose đồng thời thì nồng độ hai enzym nêu trên trong tế bào thấp hơn so với trường hợp chỉ có đường lactose. Khi phân tích tổng nồng độ mARN có mặt trong tế bào trước và sau khi đưa lactose vào trong môi trường nuôi, thấy rằng: Khi không có mặt lactose trong môi trường thì không xuất hiện mARN mã hóa cho β-galactosidase và permease. Khi bổ sung lactose vào môi trường nuôi thì trong tế bào xuất hiện mARN mã hóa cho hai enzym nêu trên. Từ kết quả thí nghiệm trên, cùng với những kết quả thu được về đột biến gen, F. Jacob và J. Monod đã đưa ra mô hình điều khiển operon lactose như hình bên dưới
Cấu trúc của operon lactose gồm: gen điều hoà (R), promoter (P), operator (O) và ba gen cấu trúc là lacZ mã hóa cho enzym β-galactosidase, lacY mã hóa cho enzym permease và lacA mã hóa cho enzym transacetylase.

Khi không có mặt lactose trong môi trường, gen điều hoà thường xuyên tổng hợp prôtêin ức chế. Prôtêin ức chế có ái lực với điểm điều hành (operator) nên nó gắn vào điểm điều hành, ngăn cản không cho enzym ARN- polymerase thực hiện phiên mã, mARN không thể tổng hợp được, operon đóng.
Khi có mặt lactose trong môi trường, nhờ enzym permease có sẵn ở màng tế bào chuyển một lượng rất ít vào trong tế bào. Khi đã vào trong tế bào, lactose chuyển thành allolactose (có chứa liên kết β-1,6). Allolactose là chất cảm ứng, nó liên kết với prôtêin kìm hãm. Phức hợp này không có ái lực với operator, nên không gắn lên operator được, lúc này, operon mở. ARN- polymerase thực hiện phiên mã các gen cấu trúc.
Hình 2.1.Mô hình hoạt động của operon lactose
Phần lớn sự tổng hợp các enzym hoạt động trong các quá trình phân giải ở tế bào được sự kiểm soát theo cơ chế của operon cảm
2.2.2. Điều hòa dương tính operon lactose
Chức năng của β-galactosidase trong chuyển hóa lactose để tạo ra glucose bằng cách cắt lactose (một sản phẩm cắt khác là galactose cũng có thể được chuyển thành glucose nhờ enzyme của operon galactose). Nếu cả glucose và lactose có mặt trong môi trường sinh trưởng, không cần hoạt động của operon lac. Trong môi trường có glucose, enzyme β- galactosidase không được tạo thành cho đến khi glucose trong môi trường được sử sụng hết. Sự có mặt của glucose làm mRNA không được tổng hợp, do đó không có sự tổng hợp β-galactosidase, vì sự thêm vào nhân tố cảm ứng làm bất hoạt chất kìm hãm. Một yếu tố khác cần cho sự bắt đầu tổng hợp mRNA, hoạt tính của yếu tố này được điều hòa bởi nồng độ glucose.
Glucose có ảnh hưởng ức chế gián tiếp lên sự biểu hiện của operon lac.
Những phân tử nhỏ adenosine monophosphate vòng (cAMP) phân bố rộng rãi trong
mô động vật và trong các cơ thể eukaryote đa bào, có vai trò quan trọng làm chất
trung gian hoạt động hormone. Chất này cũng có trong tế bào E. coli và nhiều tế bào vi khuẩn khác với chức năng khác nhau. cAMP được tổng hợp bởi enzyme adenyl cyclase, và nồng độ của cAMP được điều hòa gián tiếp qua trao đổi chất glucose.
Khi vi khuẩn sinh trưởng ở môi trường chứa glucose, hàm lượng cAMP rất thấp.
Trong môi trường chứa glycerol hoặc các nguồn carbon không thể đi vào con đường hóa sinh được sử dụng để trao đổi chất glucose (con đường glycolytic) hoặc khi vi khuẩn bị đói nguồn năng lượng, nồng độ cAMP cao. Hàm lượng glucose giúp điều hòa nồng độ cAMP trong tế bào và cAMP lại điều hòa hoạt tính của operon lac.
E. coli chứa protein nhận cAMP hay CRP (cyclic AMP receptor protein) còn được gọi là protein hoạt hóa dị hóa CAP (catabolite activator protein), được mã hóa bởi gen crp Đột biến ở gen crp và gen adenyl cyclase làm ngăn cản sự tổng hợp của mRNA lac. Điều này cho thấy chức năng của CRP và cAMP cần thiết cho tổng hợp mRNA lac. CRP và cAMP gắn vào một vị trí khác tạo phức hợp cAMP-CRP được biểu hiện. Phức hợp này là một yếu tố điều hòa hoạt hóa ở hệ thống lac.

ức chế
Khi vắng mặt phức hợp cAMP-CRP, RNA polymerase chỉ bám lỏng lẽo vào promotor. Vì vậy hiếm khi dẫn đến phiên mã vì không có sự tương tác đúng giữa RNA polymerase và promotor. Nhưng RNA polymerase được kích thích gắn vào promotor khi cAMP-CRP được gắn vào DNA.
Những kết quả này giải thích chức năng lactose và glucose cùng nhau tham gia điều hòa phiên mã operon lac như thế nào.
Khi trong môi trường có nhiều nguồn cácbon khác nhau, thông thường vi sinh vật sẽ sử dụng nguồn cácbon quen thuộc trước, sau khi cạn kiệt nguồn đó thì vi sinh vật mới chuyển dần sang sử dụng nguồn cácbon kế tiếp.
Ví dụ: Ở vi khuẩn E.coli nếu trong môi trường nuôi cấy có cả glucozơ và lactozơ thì vi khuẩn sẽ sử dụng glucozơ trước, đến khi cạn dần glucozơ thì mới chuyển sang dùng nguồn cácbon còn lại. Như vậy, có thể khẳng định rằng ở vi sinh vật, gen cơ định mã hóa các enzym chuyển hóa nguồn cácbon nào thì trong môi trường có nhiều nguồn cácbon chúng sẽ sử dụng nguồn cácbon đó trước, vì trong tế bào chúng luôn có sẵn enzym đó.
Vậy bằng cách nào mà vi khuẩn cảm nhận được sự thay đổi nồng độ cácbon đang sử dụng? Đó là nhờ một phân tử AMP vòng (CAMP), phân tử CAMP sẽ được tích lũy dần khi nồng độ glucozơ giảm dần. Mà prôtêin điều hòa hoạt hóa operon Lac là prôtêin khi ở dạng tự do nó không hoạt động. Khi CAMP liên kết với prôtêin hoạt hóa nó trở thành dạng hoạt động (CAP), khi CAP liên kết vào vị trí điều hòa của operon nó sẽ làm tăng ái lực của promotor với ARN polimeraza, dẫn đến gen tăng biểu hiện. Khi tăng nồng độ glucozơ thì lượng CAMP lại giảm đi. Lượng phức hệ CAP giảm, ái lực của promotor với ARN polimeraza giảm, gen phiên mã giảm đi.
Do trình tự liên kết của chất ức chế lacI tại operator phủ lên promotor lac một phần, nên khi lacI liên kết vào operator, nó ngăn cản sự liên kết của ARN polimeraza vào promotor. Còn đối với CAP, khi thiếu vắng prôtêin này, ARN polimeraza chỉ liên kết được rất yếu vào promotor. Bởi vì, promotor lac thiếu một trình tự UP, đồng thời trình tự liên ứng của nó không hoàn chỉnh.
Tổng hợp của mRNA
Hình 2.2. Bốn trạng thái điều hòa của operon lac
Nhu cầu về cAMP-CRP phụ thuộc vào hệ thống ức chế lac, vì đột biến crp và adenyl cyclase không thể tạo mRNA lac ngay cả khi có mặt đột biến ở gen điều hòa (lacI-) và gen chỉ huy (lacO-). Sở dĩ như vậy là vì phức hợp
cAMP-CRP phải gắn vào trình tự base trên DNA ở vùng promotor để xảy ra phiên mã (Hình 7.4). Khác với chất ức chế trong điều hòa âm tính, phức hợp cAMP-CRP là chất điều hòa dương tính. Các thí nghiệm ở điều kiện in vitro
cho thấy: mRNA lac được tổng hợp chỉ khi có cAMP vòng và không có chất
Ngược lại, prôtêin CAP khi liên kết vào operon lac lại có vai trò thúc đẩy operon hoạt động. Tuy vậy, CAP chỉ liên kết mạnh khi môi trường không có glucozơ cAMP là phân tử kích ứng dị hình của CAP. Nghĩa là, chỉ khi cAMP liên kết với CAP, thì prôtêin này mới có cấu hình phù hợp liên kết với ADN.
Khi CAP liên kết vào vị trí này, nó đồng thời huy động và thúc đẩy ARN polimeraza đính kết vào promotor. CAP hoạt hóa operon Lac bằng việc huy động enzym ARN polimeraza đến promotor.
cAMP là phân tử kích ứng dị hình của CAP. Nghĩa là, chỉ khi cAMP liên kết với CAP, thì prôtêin này mới có cấu hình phù hợp liên kết với ADN.
2. 3. Điều hòa Operon đồng hóa ở vi khuẩn
2.3.1. Điều hòa Operon kìm hãm - Operon tryptophan ở E.coli
Operon tryptophan cũng có cấu trúc tương tự như operon lactose, nghĩa là, bao gồm gen điều hoà (R), promoter (P), operator (O) và 5 gen cấu trúc. Mỗi gen cấu trúc mã hóa cho một enzym, xúc tác phản ứng tổng hợp tryptophan. Trong tế bào, tryptophan được tổng hợp bằng một chuỗi 5 phản ứng, mỗi phản ứng được xúc tác bằng một enzym mã hóa trong operon. Năm gen cấu trúc mã hóa cho 5 enzym, lần lượt được ký hiệu là trpE, trpD, trpC, trpB và trpA. Gen trpE nằm ngay sau vùng điều hoà, được phiên mã đầu tiên.

bị ức chế. Khi sự cung cấp tryptophan bị thiếu, sự phiên mã xảy ra. Sự điều hòa của operon lactose tương tự với operon lactose, vì sự tổng hợp mRNA được điều hòa âm tính nhờ chất ức chế. Tuy nhiên, khác với điều hòa ở operon lac, tryptophan hoạt động như chất đồng kìm hãm, kích thích chất ức chế gắn vào operator ngừng sự tổng hợp. Operon tryptophan hoạt động theo kiểu ức chế, điều hòa âm tính.
Tryptophan được tổng hợp qua các giai đoạn khá phức tạp, mỗi giai đoạn có sự xúc tác của một enzyme đặc biệt. Các gen mã hóa cho các enzyme này nằm kề nhau trên nhiễm sắc thể của E.coli. Đó là các gen trpE, trpD, trpC, trpB, trpA. Các enzyme được dịch mã từ một phân tử mRNA đa cistron. Vùng mã hóa gen E được dịch mã trước tiên. Phía trước trpE về đầu 5' có promotor, operator và 2 vùng xếp lần lượt là leader (trpL) và đoạn kìm hãm phiên mã attenuator (trpa, không phải là trpA). Gen ức chế trpR nằm xa operon, tổng hợp protein aporepressor, là chất kìm hãm mà riêng nó không có hoạt tính. Khi tryptophan dư thừa, nó kết hợp với aporepressor tạo chất kìm hãm có hoạt tính, gắn vào operator của operon tryptophan làm dừng phiên mã các gen cấu trúc. Khi nồng độ tryptophan thấp, nó tách khỏi phức hợp kìm hãm và aporepressor mất hoạt tính. Lúc này operator mở ra, RNA polymerase dịch mã 5 gen cấu trúc để tổng hợp 5 enzyme tham gia tổng hợp tryptophan.
2.3.2. Điều hòa Operon phiên mã dở (Attenuation) Operon tryptophan ở E.coli
Kiểu điều hòa thứ hai được phát hiện ở operon tryptophan được gọi là attenuation. Nó dùng sử dịch mã để điều khiển sự phiên mã. Khi có mặt tryptophan nội bào, ngay cả với nồng độ thấp, sự dịch mã một phần vùng leader của mRNA ngay khi vừa được tổng hợp, kết quả làm dừng sự phiên mã trước khi gen cấu trúc đầu tiên của operon được sao chép.
Hình 2.3.Mô hình hoạt động của operon tryptophan
Hoạt động của operon tryptophan khác với operon lactose ở chỗ là prôtêin kìm hãm, bản thân nó không có ái lực với operator nên khi đứng riêng một mình, nó không thể gắn vào điểm điều hành, nên operon mở. Ngược lại, khi prôtêin kìm hãm kết hợp với chất đồng kìm hãm (corepressor), thì sẽ có ái lực với operator nên dễ dàng gắn vào đó, làm operon đóng. Hoạt động của operon có thể mô tả như hình trên. Như vậy, khi lượng tryptophan dư, operon đóng, còn khi thiếu tryptophan thì operon mở
Nhìn chung, cách điều hoà biểu hiện gen ở sinh vật procaryote chủ yếu được thực hiện ở giai đoạn phiên mã.
Operon tryptophan (trp) của E.coli chứa các gen cấu trúc mã hóa cho các enzyme tổng hợp amino acid tryptophan. Operon này được điều hòa theo cách sau: khi tryptophan có mặt đầy đủ trong môi trường sinh trưởng, sự phiên mã của operon
Hình 2.4. Sơ đồ phiên mã của leader trp(A); Chi tiết cấu trúc của 2 codon trp ở vòng 1-2 (B)
Phiên mã dở (attenuation) là kết quả sự tương tác giữa các trình tự DNA trong vùng leader của bản phiên mã trp. Ở tế bào kiểu dại, sự phiên mã operon trp thường được bắt đầu. Tuy nhiên khi có mắt một lượng nhỏ tryptophan, hầu hết phân tử mRNA kết thúc ở vùng 28 base đặc biệt ở trong trình tự leader. Kết quả của sự kết thúc sớm này tạo phân tử mRNA chứa 140 nucleotide chấm dứt một đoạn ngắn của các gen mã hóa cho các enzyme trp. Vùng 28 base xảy ra sự kết thúc phiên mã sớm như thế được gọi là attenuator. Trình tự base của vùng này thường có các tính chất điểm kết thúc, gồm dạng đoạn và vòng (stem-loop) trên mRNA theo sau là trình tự của 8 uridine.
Trình tự leader có các đặc điểm:
- Một vùng có codon AUG và phía sau là codon kết thúc UGA, mã hóa cho một polypeptide chứa 14 amino acid được gọi là leader polypeptide.
- Hai codon tryptophan ở vị trí 10 và 11 trên mRNA của leader polypeptide.

Trình tự lặp lại ngắn này có ý nghĩa trọng điều hòa.
- Bốn đoạn của RNA leader là vùng 1, 2, 3 và 4 tạo thành do khả năng kết cặp của các base với nhau. Các base ở vùng 1 kết cặp với vùng 2, vùng 3 kết cặp với vùng 4.
Khi sự kết cặp xảy ra ở dạng này, sự phiên mã kết thúc ở đoạn đi qua uridine phía trước nucleotide 140. Kiểu kết cặp này xảy ra ở mRNA leader được tinh sạch.
- Một kiểu kết cặp biến đổi có thể xảy ra, trong đó các base vùng 2 kết cặp với vùng 3 nhờ các cặp base ở 2 vùng này gần như bổ sung nhau. Qua mô hình kết cặp base biến đổi này (3-4 hoặc 2-3), sự tổ chức trình tự mRNA có thể điều hòa
phiên mã qua dịch mã của leader polypeptide. Khi vùng leader được phiên mã, sự dịch mã leader polypeptid cũng bắt đầu. Vì có 2 codon của tryptophan trong trình tự mã hóa, nên sự dịch mã nhạy cảm với số lượng tRNAtrp đưa vào.
Nếu môi trường cung cấp đầy đủ tryptophan, ribosome trượt qua codon tryptophan và đi vào vùng 7.1. Sự có mặt của ribosome loại bỏ khả năng kết cặp của vùng khoảng 10 base ở mỗi phía của codon đang dịch mã. Sự có mặt của ribosome ở vùng 2 ngăn cản nó kết cặp với vùng 3. Trong trường hợp này vùng 3 kết cặp với vùng 4, tạo ra điểm kết thúc phiên mã. Sự phiên mã kết thúc khi qua các uridine nằm phía sau vùng 4.
Kết thúc phiên mã 360
Khi số lượng tRNAtrp không đủ, sự dịch mã leader polypeptide bị dừng lại đột ngột ở các codon tryptophan. Sự dùng lại này ngăn cản ribosome tiến vào vùng 2, vì vậy vùng 2 được tự do sẽ kết cặp với vùng 3 làm cản trở sự hình thành cấu trúc kết thúc. Vì vậy phân tử trp mRNA hoàn chỉnh được tạo thành, chứa cả trình tự mã hóa cho gen cấu trúc.
Tóm lại, attenuation là cơ chế điều hòa tinh tế trên cơ sở điều hòa âm tính: Khi tRNAtrp đến đủ cung cấp cho sự dịch mã leader polypeptide, sự phiên mã bị dừng, các trp enzyme không được tổng hợp. Khi nồng độ tRNAtrp quá thấp, sự phiên mã xảy ra cho đến hết, các trp enzyme được tạo nên.
Nhiều operon chịu trách nhiệm tổng hợp các amino acid khác (như các operon của leucine, isoleucine, phenylalanine, histidine) cũng được điều hòa nhờ attenuator với chức năng tạo ra vùng kết cặp biến đổi ở bảnphiên mã. Ở operon histidine vùng mã hóa của leader polypeptide chứa 7 codon histidine kế nhau. Ở operon phenylalanine vùng mã hóa cho leader polypeptide chứa 7 codon phenylalanin chia 3 nhóm.
Điều hòa kiểu attenuation không thể xảy ra ở eukaryote vì ở eukaryote sự phiên mã và dịch mã không xảy ra đồng thời. Sự phiên mã xảy ra trong nhân, còn sự dịch mã xảy ra ở tế bào chất.
Điều hòa ở operon lac và operon trp là ví dụ về một trong số các cơ chế quan trọng điều hòa hoạt động gen ở mức phiên mã của prokaryote.
2.3. Câu hỏi bài tập điều hòa hoạt động Operon đồng hóa, dị hóa ở vi khuẩn
Câu 1. (Đề thi chọn đội tuyển quốc gia vòng 2 dự thi Olympic Sinh học quốc tế năm 2014) : So sánh hoạt động của operon Lac (lactose) và operon Trp (tryptophan) trong điều hoà âm tính ở E. coli?
Gợi ý trả lời:
- Giống nhau:
+ Sự điều hoà của cả hai operon Lac và Trp đều liên quan đến cơ chế điều hoà các gene kiểu âm tính: Nghĩa là, các operon này đều đượ
“tắt” bởi prôtêin điều hoà tương ứng của chúng (đều là các prôtêin ức chế do gene điều hoà tổng hợp).
+ Sự điều hoà của cả hai operon Lac và Tryptophan đều tạo cho tế bào tiết kiệm năng lượng và vật chất trong hoạt động sống của nó.
- Khác nhau:
+ Trong operon Lac, các enzyme tham gia vào con đường chuyển hoá lactose còn gọi là các enzyme cảm ứng do quá trình sinh tổng hợp chúng được gây cảm ứng bởi tín hiệu hoá học (trong trường hợp này là allolactose). Theo nguyên tắc tương tự, trong operon Trp các enzym do operon Trp mã hoá được gọi là các enzyme ức chế
+ Trong operon Tryptophan, khi Tryptophan có sẵn trong môi trường hoặc khi lượng tích luỹ trong tế bào của chúng đã đủ thì chính axit amin này kết hợp với prôtêin điều hoà tạo thành phức hợp đồng ức chế liên kết vào trình tự O (operator) làm dừng quá trình phiên mã. Ngược lại trong operon Lac, allolactose làm bất hoạt protein điều hoà làm cho protein này không liên kết được vào trình tự O, nhờ đó quá trình phiên mã diễn ra.
Câu 2: Một operon mã hóa enzyme 1 và 2 được điều hòa bởi sự chuyển hóa chất X chứa đựng 7 trình tự A, B, C, D, E, F, G chưa biết rõ chức năng. Để làm sáng tỏ chức năng của chúng, sự ảnh hưởng của đột biến ở các trình tự từ A đến G dựa trên sự tổng hợp các enzyme được đánh giá thông qua sự có mặt và sự vắng mặt của X.
Có mặt X Vắng mặt X
Đột biến trong trình tự Enzym 1 Enzym 2 Enzy m1 Enzym2
Không có đột biến - - + +
Hãy xác định vai trò của operon và các trình t
Trả lời
- Operon khi có mặt X thì cả hai enzyme đều không được tổng hợp do đó chứng tỏ đây là operon đồng hóa
- Thể đột biến B, F biểu hiện giống nhau. Khi có hay không có mặt X thì operon luôn được mở ( luôn tạo ra enzyme)
- Bị đột biến vùng O do đó protein ức chế không liên kết được với O ( Lac Oc)
- Thể đột biến D khi có mặt hay không có mặt X thì operon luôn đóng
- Bị đột biến vùng P ( ARN pol không liên kết vào P được -> phiên mã không diễnra, operon luôn đóng)
- Thể đột biến A, E biểu hiện giống nhau khi vắng mặt X chỉ có enzyme 2 được tổng hợp
- Bị đột biến gen tổng hợp enzyme 1 -> enzyme 1 không được tổng hợp
- Thể đột biến C khi vắng mặt X chỉ có enzyme 1 được tổng hợp
- Bị đột biến gen tổng hợp enzyme 2 -> enzyme 2 không được tổng hợp
- Thể đột biến G biểu hiện giống kiểu dại -> Trình tự không liên quan đến operon mã hóa enzyme 1, enzyme 2
Câu 3:Trong 1 thí nghiệm, hoạt tính của enzym β-galactosidase được đánh giá bởi sự có mặt của X-gal và IPTG trong môi trường nuôi cấy. X-gal là một đồng phân của lactose, nó bị chuyển sang màu xanh khi bị phân giải bởi βgalactosidase, nhưng nó không cảm ứng Lac operon. IPTG là chất cảm ứng giống với lactose. Sau khi gây đột biến, để xác định vị trí đột biến, người ta đã nuôi cấy các đột biến ở trong môi trường khác nhau (bảng dưới) và có bổ sung thêm X-gal. Kết quả ở bảngsau:
Điều kiện môi trường nuôi cấy Kiểu hình khuẩn lạc
Có mặt IPTG, không có glucose Màu xanh
Không có mặt IPTG, không có glucose Màu xanh
Có mặt cả IPTG và glucose Màu trắng
Không có mặt IPTG, có mặt glucose Màu trắng
Hãy xác định vị trí đột biến trong operon Lac của chủng đột biến trên. Giải thích.
Trả lời
- Có mặt IPTG, không có glucose -> màu xanh -> không bị đột biến, gen lacZ vẫn được tổng hợp bình thương tạo ra -galatosidase để phân giải Xgal tạo màu xanh
- Không có mặt IPTG, không có glucose -> màu xanh chứng tỏ lacZ vẫn được tổng hợp bình thường -> đột biến xảy ra tại vùng O do đó dù có hay không có cơ chất (IPTG) thì operon Lac vẫn luôn mở
Câu 4: Một số vi khuẩn có cơ chế điều hòa sự tổng hợp enzym liên quan đến quán trình sinh tổng hợp tryptophan (Trp). Trong cấu trúc của operon, phía trước các gen cấu trúc (trpE-A) là một đoạn trình tự leader mã hóa cho đoạn peptit chứa 2 bộ ba mã hóa 2 axit amin Trp liên tiếp. Ở nồng độ cao Trp, ribosome dịch mã mARN của chuỗi peptit leader và rời khỏi bộ ba kết thúc. Hiện tượng này giúp hình thành cấu trúc kẹp tóc giữa đoạn 1-2 và 3-4, dấn đến tín hiệu kết thúc phiên mã sau polyU và ARN polymeraza rời khỏi AND để kết thúc phiên mã (Hình dưới).
Quá trình tổng hợp tryptophan sẽ diễn ra như thế nào trong các trường hợp sau?
Giải thích.
- Đột biến làm mất 1 bộ ba mã hóa trp trong đoạn leader.
- Đột biến làm thay đổi trình tự dẫn đến việc không hình thành được cấu trúc kẹp tóc giữa đoạn 2 và 3.
- Nồng độ enzym aminoacyl-tARN synthetase (xúc tác sự gắn kết trp vào ARNtTrp) bị suy giảm.
Trả lời:
- Khi đột biến làm mất một bộ ba mã hóa Trp, tín hiệu về Trp trong tế bào sẽ thấp hơn khi đang có cả 2 bọ ba mã hóa, vì thế quá trình điều hòa sẽ mở operon chậm hơn so với bình thường.
- Đột biến làm thay đổi trình tự dẫn đến không thể hình thành cấu trúc kẹp tóc 2-3 có thể xảy ra theo 2 trường hợp:
- Đột biến xảy ra ở đoạn 2: Trong trường hợp này, cấu trúc đoạn 3 vẫn bình thường, vì thế cấu trúc kẹp tóc 3-4 luôn được hình thành. Vì thế, operon luôn đóng do ARN polymeraza sẽ luôn rời khỏi tại trình tự polyU và kết thúc phiên mã.
- Đột biến xảy ra ở đoạn 3: Sự xuất hiện đột biến làm mất các nucleotit có thể hình thành cấu trúc kẹp tóc với đoạn 2, thông thường nó sẽ ảnh hưởng tới việc hình thành cấu trúc kẹp tóc giữa đoạn 3 và 4. Trong trường hợp này, cấu trúc kẹp tóc 3-4 không được hình thành, dẫn đến operon luôn mở, phiên mã được tiếp tục ngay cả khi noognf độ axit amin Trp trong tế bào đang ở nồng độ cao.
Tuy nhiên, khi nồng độ tryptophan thấp, ribosome bị dừng tại vị trí 2 mã bộ ba mã hóa tryptophan và cấu trúc kẹp tóc giữa đoạn 2-3 được hình thành. Khi đó trình tự đoạn 4 được tự do và ARN pol tiếp tục phiên mã toàn bộ operon.


* Khi nồng độ enzym aminoacyl-tARN synthetaza bị suy giảm, phức hợp ARNt-Trp sẽ giảm, thiếu nguyên liệu để tổng hợp chuỗi peptit leader, cấu trúc kẹp tóc 2-3 luôn được hình thành, operon luôn mở ngay cả khi nồng độ Trp tăng cao.
Câu 5: Nghiên cứu gen cấu trúc tham gia vào sự chuyển hóa một chất X gồm
A, B, D và E. Một trình tự C liên kết với các trình tự A, B và D, còn E nằm cách xa. Người ta phân lập được các đột biến khác nhau ở trình tự C có ảnh hưởng đến sự biểu hiện của các gen cấu trúc.Họ tạo ra các kiểu gen ở các locus của gen A và trình tự C rồi xác định enzyme do gen A mã hóa có được tạo thành khi có mặt hoặc không có mặt chất X như sau:
Kiểu gen Không có X Có X C+A+ - +
C-A+ - -
C-A+/C+A- - +
CcA-/C-A+ + +
Cc , C: Đột biến.
C+ : Bình thường.
dấu (+) enzyme A được tổng hợp ( được biểu hiện ).
dấu (-) enzyme A không được tổng hợp ( không được biểu hiện ).
a. Hãy cho biết trình tự C đóng vai trò gì trong hoạt động của các gen cấu trúc trên? Giải thích. b. Các kết quả trên cho thấy đây là một operon điều hòa theo cơ chế nào?
c. Đột biến Cc thực ra là đột biến gì? Trả lời
a.Trình tự C là gen điều hòa. Gen điều hòa mang thông tin mã hóa pr chi phối điều hòa hoạt động các gen khác; nên gen C mang thông tin là gen điều hòa chứ không phải là promotor.
b. Khi không có chất X thì gen A không biểu hiện, có chất X thì hoạt động -> X là chất cảm ứng nên đây là một operon điều hòa theo cơ chế cảm ứng.
c. CcA-/C-A+ có hay không có X đều được biểu hiện, nên Cc là một đột biến làm cho kiểu gen luôn luôn kích hoạt gọi là đột biến cơ định -> tạo prôtêin không bám được vào operator.
Câu 6:
a. So sánh cơ chế điều hòa âm tính và điều hòa dương tính ở opêron Lac.
b. Tại sao sự điều hòa hoạt động gen ở sinh vật nhân thực thể hiện khác nhau
ở những giai đoạn phát triển khác nhau của cá thể?
Gợi ý trả lời:
a)* Giống nhau:
Đều để thích ứng với các điều kiện môi trường biến động, đồng thời để tiết
kiệm năng lượng và vật chất của tế bào.
Đều liên quan đến sự tham gia của các gen điều hòa. Các gen
này mã hóa cho các sản phẩm trực tiếp (prôtêin điều hòa) điều
hòa sự biểu hiện của các gen cấu trúc. Đều có hệ thống điều
hòa cảm ứng và ức chế thông qua sự tương tác của các tác
nhân môi trường (vai trò làm tín hiệu điều hòa) với prôtêin
điều hòa.
Khác nhau:
Trong cơ chế điều hòa dương tính, prôtêin điều hòa có vai trò làm
tăng sự biểu hiện của một hoặc một số gene cấu trúc. Còn trong
điều hòa âm tính, prôtêin điều hòa có vai trò ức chế sự biểu hiện của gene cấu trúc.
Trong cơ chế điều hòa dương tính sản sinh protein điều hòa liên
kết với trình tự phần đầu của vùng P (promoter), còn trong điều hòa âm tính, protein điều hòa liên kết với vùng O (operater).
* Sự điều hòa hoạt động gene ở sinh vật nhân thực thể hiện khác nhau ở những giai đoạn phát triển khác nhau của cá thể, vì:
Sinh vật nhân thực thường có cấu tạo cơ thể rất phức tạp, bao gồm các mô và các cơ quan chuyên hóa khác nhau phát sinh từ một tế bào duy nhất (hợp tử). Vì thế, sự điều hòa biểu hiện của nhiều gene vào những giai đoạn khác nhau cần nhiều cơ chế điều hòa tinh tế mới có thể đảm bảo cho cơ thể phát triển và sinh trưởng bình thường.
Trong sự phát sinh cá thể, tùy từng giai đoạn, tùy từng loại mô mà chỉ có một số gene trong tế bào hoạt động. Điều đó được diễn ra nhờ cơ chế điều hòa hoạt động gene.
Câu 7. Trong mô hình điều hòa hoạt động operon Lac, hãy nêu chức năng các trình tự ADN sau: (1) Trình tự chỉ huy (Regulator), (2) Trình tự khởi đầu phiên mã (Promoter), (3) Trình tự điều hoà (Operator), Gene cấu trúc Z (4), Y (5), A(6). Đột biến gene nếu xảy ra ở những trình tự này có thể gây ra những hậu quả gì?
Gợi ý trả lời:
* Chức năng các trình tự ADN trong mô hình điều hòa hoạt động operon Lac:
(1): kiểm soát quá trình phiên mã tổng hợp protein ức chế (repsessor).
(2): chứa các trình tự nu đặc hiệu thực hiện quá trình khởi động phiên mã các gene cấu trúc Z, Y, A của operon Lac.
(3): mang các trình tự đặc hiệu có ái lực cao với protein ức chế (repressor). Khi protein ức chế gắn vào vị trí này thì quá trình phiên mã của operon Lac không diễn ra.
(4): mã hoá cho enzym -galactosidase xúc tác cho phản ứng thủy phân lactôzơ thành glucôzơ và galactose, đồng thời enzim -galactoside còn có tác dụng chuyển hoá một phần lactôzơ thành đồng phân là alolactozo chất kích ứng protein ức chế
(5): mã hoá cho permease (protein vận chuyển) làm nhiệm vụ vận chuyển lactose vào tế bào.
(6): mã hoá cho enzim thiogalactoside transacetylase chuyển nhóm acetyl từ acetyl-coA vào một số hợp chất gây độc với tế bào cũng đồng thời được vận chuyển bằng permease (chức năng giải độc tế bào).
+Đột biến ở gene mã hóa protein ức chế có thể dẫn đến các hậu quả sau:
Có thể xảy ra một đột biến câm, trong các trường hợp:
đột biến nucleotit trong gene này không làm thay đổi trình tự axit amin trong protein ức chế, đột biến thay đổi axit amin trong chuỗi polypeptit của protein ức chế không làm thay đổi khả năng liên kết của protein ức chế với trình tự chỉ huy (O). Hậu quả cuối cùng của các dạng đột biến này là operon Lac hoạt động bình thường không có thay đổi gì liên quan đến sự biểu hiện của các gene cấu trúc Z và Y.
Xảy ra đột biến làm giảm khả năng liên kết của protein ức chế vào
trình tự chỉ huy sự biểu hiện của các gene cấu trúc Z và Y tăng lên.
Làm mất hoàn toàn khả năng liên kết của protein ức chế hoặc protein ức chế không được tạo ra các gene cấu trúc Z và Y được biểu
hiện liên tục.
Xảy ra đột biến làm tăng khả năng liên kết của protein ức chế vào
trình tự chỉ huy sự biểu hiện của các gene cấu trúc Z và Y giảm đi (thực tế trường hợp này hiếm xảy ra).
Câu 8. Trong hoạt động của operon Lac ở vi khuẩn E. coli, nếu đột biến xảy ra
ở gene điều hòa R (còn gọi là lac I) thì có thể dẫn đến những hậu quả gì liên quan đến sự biểu hiện của các gene cấu trúc?
Gợi ý trả lời:
-Operon Lac gồm các phần sau: gene điều hòa R mã hóa cho protein ức chế R, trình
tự khởi động P, trình tự chỉ huy (O), các gen cấu trúc Z, Y và A
-Nếu đột biến xảy ra ở gene R có thể dẫn đến các hậu quả sau:
Xảy ra đột biến câm, trong các trường hợp:
Đột biến nucleotit trong gen này không làm thay đổi trình tự axit amin
trong prôtein ức chế,
Đột biến thay đổi axit amin trong chuỗi polypeptit của prôtein ức chế không làm thay đổi khả năng liên kết của prôtein ức chế với trình tự chỉ huy (O). Hậu quả cuối cùng của các dạng đột biến này là ôperon Lac hoạt động bình thường không có thay đổi gì liên quan đến sự biểu hiện của các gen cấu trúc.
a) Xảy ra đột biến làm giảm khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gen cấu trúc tăng lên.
b) Làm mất hoàn toàn khả năng liên kết của protein ức chế hoặc protein ức chế không được tạo ra các gen cấu trúc biểu hiện liên tục.
c) Xảy ra đột biến làm tăng khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gen cấu trúc giảm đi.
* Kết luận: Đột biến xảy ra ở gene điều hòa R có thể dẫn đến những hậu quả khác nhau trong sự biểu hiện của các gen cấu trúc.
Câu 9. Bình thường người ta thấy loài vi sinh vật nọ không sản xuất ra enzim D, nhưng khi đưa thêm vào môi trường nuôi cấy của chúng một chất dinh dưỡng E thì sau 15 phút người ta thấy enzim D xuất hiện. Hãy giải thích hiện tượng trên về mặt cơ chế di truyền. Trả lời
* Dữ kiện đề bài cho ta nhận định sự điều hoà tổng hợp enzim D ở VSV nọ có thể xảy ra theo cơ chế điều hoà hoạt động gen ở vi khuẩn của F.Jacôp và J.mônô.
- Trong tế bào của VSV, các gen cấu trúc có liên quan về chức năng được phân bố liền nhau thành từng cụm có chung một cơ chế điều hoà được gọi là một opêron.
- 1 Opêrôn gồm các thành phần cơ bản sau:
+1 nhóm gen cấu trúc có liên quan về chức năng.
+ Vùng vận hành (O): là nơi protêin ức chế có thể liên kết làm ngăn cản sự phiên mã, khi không có prôtêin ức chế thì vùng vận hành hoạt động.
+ Vùng khởi động: Nơi ARN polimeraza bám vào và khởi đầu phiên mã.
Sự hoạt động của Opêrôn phụ thuộc váo sự điều khiển của gen điều hoà Opêrôn (R), gen điều hoà không nằm trong thành phần của Opêrôn mà nằm trước Opêrôn.
Bình thường gen điều hoà tổng hợp prôtêin ức chế liên kết với vùng vận hành ngăn cản quá trình phiên mã (không cho Opêrôn hoạt động).
* Trong đề bài thấy bình thường gen điều hoà phiên mã tạo mARN, tổng hợp prôtêin ức chế, chất ức chế bám vào vùng vận hành, do đó các gen cấu trúc không được phiên mã nên enzim D không được tổng hợp.
- Khi đưa thêm chất dinh dưỡng E vào môi trường nuôi cấy VSV nọ thì chính chất này với vai trò là chất cảm ứng gắn với prôtêin ức chế nên nó không gắn vàp vùng vận hành được nữa, vùng vậnh được tự do điều khiển quá trình phiên mã của Opêrôn, mARN của các gen cấu trúc được tổng hợp và quá trình dịch mã xảy ra enzim D được tổng hợp.
Câu 10. Operon là gì? Các thành phần cấu trúc của operon Lac có vai trò gì?
Gợi ý trả lời:
Operon là các gene cấu trúc ở prokaryote có liên quan về chức năng, chịu sự điều khiển chung của một cơ chế điều hoà được phân bố liền nhau thành từng cụm trên ADN. Thông thường, đó là các gene cùng tham gia vào một con đường chuyển hóa.
Một operon là một đơn vị phiên mã đơn bao gồm một chuỗi các gene cấu trúc, một promoter và một operator. Các gene cấu trúc của một operon hoặc là cùng được phiên mã hoặc cùng không được phiên mã vì chúng đều được điều khiển bởi một operator (O) và một promoter (P) duy nhất.
Các thành phần cấu trúc operon Lac có vai trò:
- Một nhóm gen cấu trúc liên quan về chức năng gồm 3 gene cấu trúc (Z, Y, A) kiểm soát tổng hợp 3 protein Z, Y, A, là các enzym tham gia các phản ứng phân giải đường lactozơ:
+ Gene Lac Z quy định tổng hợp enzim β- galactozidase có chức năng phân giải đường lactose thành glucose và galactose, đồng thời còn có vai trò chuyển hóa một phần lactose (liên kết β 1,4D glycoside ) thành một đồng phân allolactose (liên kết β 1,6D glycoside) là phân tử bất hoạt protein ức chế.
+ Gene Lac Y quy định một loại protein permeaza nằm xuyên màng tế bào vi khuẩn làm nhiệm vụ vận chuyển chủ động đường lactose vào trong tế bào vi khuẩn.
+ Gene A quy định tổng hợp enzyme thiogalactoside transacetylase có vai trò giải độc tế bào đối với các hợp chất thiogalactoside cũng được đồng vận chuyển vào tế bào khi permease hoạt động và chuyển nhóm acetyl từ acetyl-coA tới β- galactozidase.
Tất cả 3 gene cấu trúc này đều được phiên mã cùng một lúc tạo thành một phân tử mARN.
- Vùng vận hành (O) là trình tự nuclêôtit đặc biệt để protein ức chế dị hình liên kết ngăn cản phiên mã.
- Vùng khởi động (P) nằm gần đầu 5 của gene Lac Z, là nơi ARN polimezaza bám vào khởi đầu phiên mã đồng thời của ba gene Lac Z, Lac Y, Lac A
Câu 11. So sánh cơ chế điều hòa âm tính và điều hòa dương tính ở opêron Lac.
Trả lời
* Giống nhau:
- Đều để thích ứng với các điều kiện môi trường biến động, đồng thời để tiết kiệm năng lượng và vật chất của tế bào.
- Đều liên quan đến sự tham gia của các gen điều hòa. Các gen này mã hóa cho các sản phẩm trực tiếp (prôtêin điều hòa) điều hòa sự biểu hiện của các
gen cấu trúc. Đều có hệ thống điều hòa cảm ứng và ức chế thông qua sự tương tác của các tác nhân môi trường (vai trò làm tín hiệu điều hòa) với prôtêin điều hòa.
* Khác nhau:
- Trong cơ chế điều hòa dương tính, prôtêin điều hòa có vai trò làm tăng sự biểu hiện của một hoặc một số gen cấu trúc. Còn trong điều hòa âm tính, prôtêin điều hòa có vai trò ức chế sự biểu hiện của gen cấu trúc.
- Trong cơ chế điều hòa dương tính sản sinh prôtêin điều hòa liên kết với trình tự phần đầu của vùng P (promoter), còn trong điều hòa âm tính, prôtêin điều hòa liên kết với vùng O (operater).
Câu 12. So sánh hoạt động của operon lac (lactozơ) và operon trp (tryptophan) trong điều hoà âm tính ở E.coli.
Trả lời
Giống nhau:
- Sự điều hoà của cả hai operon lac và trp đều liên quan đến cơ chế điều hoà các gen kiểu âm tính: Nghĩa là, các operon này đều được “tắt” bởi prôtêin điều hoà tương ứng của chúng (đều là các prôtêin ức chế do gen điều hoà tổng hợp).
- Sự điều hoà của cả hai operon lac và trp đều tạo cho tế bào tiết kiệm năng lượng và vật chất trong hoạt động sống của nó.
Khác nhau: operon Lac operon ức chế
luôn tắt, chỉ mở khi có chất cảm ứng Luôn hoạt động, chỉ tắt khi có chất ức chế
pr ức chế khi mới hình thành đã
ở dạng hoạt động ngay, liên kết vào O và ức chế phiên mã
Enzim cảm ứng thường hoạt động
trong con đường dị hóa
Pr ức chế khi mới tổng hợp ở dạng bất hoạt, chỉ được hoạt hóa khi có chất đồng ức chế.
Enzim ức chế thường hoạt động trong con đường đồng hóa.
- Trong operon lac, các enzim tham gia vào con đường chuyển hoá lactozơ còn gọi là các enzim cảm ứng do quá trình sinh tổng hợp chúng được gây cảm ứng bởi tín hiệu hoá học (trong trường hợp này là allolactozơ). Theo nguyên tắc tương tự, trong operon trp các enzim do operon trp mã hoá được gọi là các enzim ức chế.
- Trong operon trp, khi tryptophan có sẵn trong môi trường hoặc khi
lượng tích luỹ trong tế bào của chúng đã đủ thì chính axit amin này kết hợp với
prôtêin điều hoà tạo thành phức hợp đồng ức chế liên kết vào trình tự O (operator) làm dừng quá trình phiên mã. Ngược lại trong open lac, allolactose
làm bất hoạt prôtêin điều hoà làm cho prôtêin này không liên kết được vào trình
tự O, nhờ đó quá trình phiên mã diễn ra.
Câu 13. Hình thức tổ chức gen theo kiểu Operon đem lại lợi ích gì cho SV?
Cho ví dụ.
Trả lời
Ý nghĩa của việc tổ chức gen theo kiểu Operon:
- Tiết kiệm vật chất di truyền, làm cấu trúc hệ gen gọn nhẹ (cách tổ chức của nhiều gen trong TB nhân sơ).
- Tạo ra sản phẩm các gen nhanh chóng, các sản phẩm gen này thường liên quan đến nhau về chức năng nên cùng lúc tế bào cần lượng tương đương.
VD: Operon Lac ở E.coli tạo ra các protein chuyển hóa lactozo, các gen rARN (ở cả nhân sơ và nhân thực) được tạo ra lượng lớn, đồng thời mới có thể đáp ứng được yêu cầu của TB.
3.1. Cơ sở sinh hóa của tương tác giữa các gen alen
Gen alen là các trạng thái khác nhau của cùng một gen phân bố trong cùng một locus gen. Alen này chỉ khác alen kia ở một hoặc một vài cặp nucleotit và là sản phẩm của quá trình đột biến gen.
Tương tác giữa các gen alen là sự tác động qua lại giữa các alen của cùng một gen để biểu hiện ra kiểu hình theo các dạng:
Trội lặn hoàn toàn.
Trội lặn không hoàn toàn
Đồng trội.
Siêu trội.
Gen gây chết (Alen gây chết).
Gen chịu sự di truyền hiệu quả dòng mẹ (sự DT biểu hiện chậm của Menđen)
Gen biểu hiện chịu ảnh hưởng của giới tính
Gen chịu sự di truyền hiện tượng in vết của gen
3.1.1. Tương tác kiểu trội lặn hoàn toàn
* Khái niệm
Tương tác trội lặn hoàn toàn là hiện tượng trong đó một alen lấn át hoàn toàn sự biểu hiện của một alen khác thuộc cùng một locus. Trong trường hợp này, kiểu hình của thể dị hợp tử hoàn toàn giống kiểu hình của thể đồng hợp trội.
*Cơ sở sinh hóa của tương tác trội lặn hoàn toàn
Ta xét một gen gồm 2 alen A và a, trong đó alen A là trội hoàn toàn so với alen a.
Alen trội A (thường là alen kiểu dại) mã hóa một enzym, alen lặn a (thường là alen đột biến) có thể sinh ra một Protein không có hoặc có rất ít hoạt tính enzym (đột biến nhầm nghĩa) hoặc alen a không sinh ra Protein hoặc chỉ sinh ra một đoạn (đột biến mất nghĩa). Các cá thể dị hợp Aa chỉ sinh ra khoảng ½ số lượng enzim có hoạt tính so với các cá thể đồng hợp trội AA. Nếu số lượng đó là đủ cho tế bào hoặc cơ thể thực hiện các chức năng hóa sinh một cách bình thường thì sẽ xuất hiện kiểu hình bình thường.
Trong trường hợp alen A là alen đột biến trội hơn so với alen a là alen kiểu dại thì
có thể giải thích do enzym của alen đột biến sinh ra ái lực lớn hơn với cơ chất so với enzym của alen kiểu dại. Tuy vậy, enzym đột biến thường không có khả năng xúc tác phản ứng hoặc xúc tác với hiệu quả thấp gây ra hiệu ứng đa số đột biến gen là có hại vì mất cân bằng hài hòa giữa kiểu gen và môi trường trong quá trình chọn lọc tự nhiên
không có sự thay thế hoàn toàn một alen này bằng một alen khác, mà là sự duy trì ưu thế của các thể dị hợp về một hoặc một số cặp alen nào đó.
3.1.4. Gen gây chết (Alen gây chết)
* Khái niệm
Tương tác gen gây chết là hiện tượng khi sự có mặt của alen đó trong kiểu gen sẽ ảnh hưởng không thuận lợi đến sự sinh trưởng và phát triển của sinh vật, do vậy dẫn đến làm giảm sức sống hay gây chết cho cơ thể mang nó.
Có hai dạng tương tác gen gây chết: gây chết trong trạng thái đổng hợp trội và gây chết trong trạng thái đồng hợp lặn.
Các gen theo hiệu quả gây chết thường chia làm 3 nhóm:
Gen gây chết hoàn toàn: là gen làm chết hoàn toàn các cá thể đồng hợp mang nó.
Giả thuyết 1: * Giả thuyết 2: Hình 3.1.Các giả thuyết về cơ sở của tính trội hoàn toàn.
3.1.2. Tương tác trội lặn không hoàn toàn
* Khái niệm
Tương tác trội lặn không hoàn toàn là hiện tượng trong đó một alen trội không hoàn toàn lấn át sự biểu hiện của alen lặn khác thuộc cùng một locus. Kiểu hình của thể dị hợp tử biểu hiện hiểu hình trung gian giữa kiểu hình của hai thể đồng hợp.
* Cơ sở sinh hóa của tương tác trội lặn không hoàn toàn
Có thể giải thích hiện tượng trội không hoàn toàn là do sự có mặt của alen a đã kìm hãm quá trình phiên mã, dịch mã của alen A. Kết quả là sắc tố đỏ hình thành ít hơn mức bình thường khiến cho cây mang kiểu gen Aa cho hoa màu hồng.
Bởi kiểu hình của thể dị hợp là trung gian giữa hai thể đồng hợp, vì vậy ta có thể lý giải trên phương diện sinh hóa rằng hàm lượng sản phẩm tích lũy do một allele trội kiểm soát là không đủ để thể hiện kiểu hình màu đỏ như
trong trường hợp có mặt cả hai alen trội.
3.1.3. Tương tác đồng trội
* Khái niệm
Tương tác đồng trội là hiện tượng cả hai alen trong cơ thể dị hợp cùng hình thành những tính trạng mà nó kiểm soát một cách độc lập với alen cùng cặp.
* Cơ sở sinh hóa của tương tác đồng trội
Hiện tượng đồng trội được giải thích trên cơ sở tính đa hình cân bằng của các sản phẩm Protein được sinh ra từ các alen đồng trội. Trong tự nhiên sự đa hình cân bằng sẽ
Gen nửa gây chết: là gen làm chết nhiều hơn 50% nhưng ít hơn 100% số thể đồng hợp mang nó.
Gen giảm sống: là gen làm chết dưới 50% số thể đồng hợp mang nó. Tuy nhiên, sự phân chia này cũng chỉ có tính chất quy ước vì một gen có thể gây chết hoàn toàn trong điều kiện này nhưng lai lại là gen nửa gây chết trong điều kiện ngoại cảnh khác.
* Cơ sở sinh hóa của tương tác alen gây chết
Hiện tượng tương tác alen gây chết có thể giải thích do alen quy định tổng hợp Protein từ đó quy định tổng hợp những enzyme tham gia quá trình chuyển hóa quyết định sự sống còn của sinh vật.
3.1.5. Di truyền của gen có nhiều alen
Trên thực tế, mỗi một gen không chỉ có hai alen mà có thể có nhiều hơn hai alen, gọi là đa alen. Các alen là những trạng thái cấu trúc khác nhau của cùng một gen phát sinh do đột biến.
Hiện tượng đa alen tồn tại phổ biến trong các quần thể sinh vật khác nhau. Ví dụ, gen kiểm soát màu mắt đỏ-trắng ở ruồi giấm gồm một chuỗi 12 alen, với tính trội giảm dần từ đỏ kiểu dại cho đến trắng đột biến lặn (w) theo thứ tự từ trái sang phải và trên xuống như sau:
Alen
Màu
W+ Wsat Wco Ww Wap3 Wch đỏ dại satsu ma cor al win e aprico t3 cherr y
Alen We Wbl Wap Wi Wt
Màu eossin blood apricot ivory tinged white
Một số gen ở người, chẳng hạn như các gen đối với kháng nguyên bạch cầu người HLA (human leukocyte antigen) xác định các kháng nguyên trên bề
mặt của hầu như tất cả các tế bào có thể có nhiều alen. Ví dụ, gen HLA-B có nhiều hơn 30 alen được xác định khác nhau về mặt kháng nguyên trong một số quần thể. Kết quả của sự đa dạng này là, trong một quần thể có rất nhiều kiểu gen ở gen HLA-B (465 kiểu gen khác nhau, với 30 kiểu đồng hợp và 435 kiểu dị hợp), thực sự tạo nên một dãy biến dị kiểu gen rộng.
3.1.6. Tương tác siêu trội
Hiện tượng siêu trội là hiện tượng thể dị hợp của các alen luôn có sức sống, sức sinh trưởng, sinh sản tốt hơn thể đồng hợp, kể cả thể đồng hợp trội (AA < Aa > aa).
Có thể giải thích là do sự tương tác giữa hai sản phẩm Protein của 2 alen khác nhau về chức phận trong cùng một locus dẫn đến hiệu quả bổ trợ, làm tăng tác động của alen trội.
Tương tác siêu trội gây hiệu ứng ưu thế lai trong lai khác dòng dẫn đến trong quần thể giao phối đa số các kiểu gen ở trạng thái dị hợp.
3.1.7. Gen chịu sự di truyền hiệu quả dòng mẹ (sự DT biểu hiện chậm của Menđen)
Sản phẩm do gen ở trong nhân tạo ra (trước khi thụ tinh) tồn tại ở tế bào chất của tế bào trứng tác động đến sự biểu hiện kiểu hình ở đời sau. Hiện tượng này gọi là "tiền định tế bào chất" hay hiệu ứng dòng mẹ (còn gọi là hiện tượng di truyền Men Đen thể hiện chậm đi 1 thế hệ).
Ví dụ về hiệu quả dòng mẹ: chiều xoắn của vỏ ốc Limnaea peregra. Chiều xoắn này được xác định bởi một cặp alen. D là xoắn phải, d là xoắn trái. Các phép lai cho thấy D là trội so với d và chiều xoắn luôn luôn được xác định bởi kiểu gene của mẹ.
Hình 3.2.Hiệu quả dòng mẹ lên chiều xoắn vỏ ốc
Tiến hành phép lai thuận nghịch giữa đồng hợp tử DD xoắn phải và đồng hợp tử dd xoắn trái:
Cơ sở sinh hóa gen chịu sự di truyền hiệu quả dòng mẹ
Trứng và phôi chịu ảnh hưởng của môi trường cơ thể mẹ nhiều hơn cơ thể bố. Ngay cả khi bị tách khỏi cơ thể mẹ từ một giai đoạn rất sớm, chúng cũng đã nhận được tế bào chất, các chất dinh dưỡng trong trứng từ mẹ và những ảnh hưởng đặc biệt lên hoạt động của gene. Những tiềm năng nhất định của trứng được xác định trước khi thụ tinh và trong một số trường hợp chúng đã chịu ảnh hưởng của môi trường mẹ bao quanh. Sự quyết định trước như vậy do các gene của mẹ hơn là các gene của con, được
gọi là hiệu ứng mẹ. Sự tồn tại của hiệu ứng mẹ nói chung được chứng minh bằng phương pháp lai thuận nghịch, khi đó kết quả phép lai thuận nghịch sẽ khác nhau.
Như vậy hiệu quả dòng mẹ chỉ kéo dài một thế hệ và trường hợp này không thể xem là di truyền qua tế bào chất bởi vì ở đây các đặc tính của tế bào chất đã được xác
định trước bởi tác dụng của các gene trong nhân chứ không phải bởi các gene trong tế bào chất. Nói cách khác ở đây cơ chế di truyền nhiễm sắc thể làm biến đổi tế bào chất của trứng trước khi nó thụ tinh.
3.1.8. Gen biểu hiện chịu ảnh hưởng của giới tính
Trong trường hợp tính trạng trội phụ thuộc vào giới tính thì tỉ lệ kiểu hình của 1 phép lai được tính riêng ở từng giới. Tỉ lệ kiểu hình phân li chung bằng trung bình cộng tỉ lệ kiểu hình ở 2 giới.
3.1.9. Gen chịu sự di truyền hiện tượng in vết của gen
Trong toàn bộ quá trình phân tích về di truyền học Menđen và cơ sở nhiễm sắc thể của sự di truyền, một gen nào đó sẽ cho hiệu quả như nhau bất luận được di truyền từ bố hay từ mẹ. Tuy nhiên, trong một vài trường hợp, ở động vật có vú nhiều tính trạng phụ thuộc vào việc gen biểu hiện được di truyền từ bố hay mẹ Đây chính là khái niệm về di truyền in vết. Những biểu hiện của kiểu hình phụ thuộc vào việc allen quy định đặc điểm được truyền từ bố hay mẹ được gọi là in vết hệ gen (di truyền in vết).
Hiện tượng này khác biệt hoàn toàn với hiện tượng gen liên kết với giới tính, ở đây các gen in vết nằm trên nhiễm sắc thể thường.
* Cơ chế:
In vết hệ gen xảy ra trong quá trình hình thành giao tử và kết quả làm bất hoạt một allen của một gen nhất định. Bởi vì các gen này được in vết khác nhau ở tinh trùng và trứng nên hợp tử chỉ biểu hiện một allen của một gen được in vết (allen đó có thể được di truyền từ bố hoặc di truyền từ mẹ). Sự in vết được truyền và biểu hiện ở tất cả các tế bào cơ thể trong qúa trình phát triển. Trong mỗi thế hệ, sự in vết cũ sẽ được “xóa” đi trong các tế bào sản sinh giao tử và in vết mới được thiết lập trên các nhiễm sắc thể của giao tử đang được hình thành dựa theo giới tính của các cá thể tạo ra giao tử. Trong cùng loài, các gen in vết luôn luôn in vết theo cùng một cách.
Cơ chế liên quan đến in vết gen là sự methyl hóa ADN, gắn nhóm methyl (CH3) vào các nucleotit cytosine của một trong các allen, do đó làm bất hoạt allen, allen này không được biểu hiện trong quá trình phát triển của phôi. Tuy nhiên, đối với một số gen, sự methyl hóa lại làm hoạt hóa sự biểu hiện của allen. Một ví dụ, đó là trường hợp của gen Igf2: Methyl hóa một số cytosine nhất định trên nhiễm sắc thể của bố dẫn đến sự biểu hiện của allen Igf2.
* Một số rối loạn liên quan đến in vết hệ gen, bao gồm: hội chứng Angelman, hội chứng Prader-Willi và hội chứng Beckwith-Wiedemann.
Hội chứng Angelman là một rối loạn của hệ thần kinh, triệu chứng đặc trưng như co giật, khiếm khuyết về ngôn ngữ, hành động kì lạ. Cơ chế gây ra hội chứng
được nghiên cứu trên khoảng 70% số bệnh nhân Angelmen là do một phần của NST số 15 bị xóa (khoảng 4 triệu cặp nucleotit). Trong đó gen đột biến di truyền từ mẹ
không được biểu hiện, gen di truyền từ cha được biểu hiện. Bằng một cách khác, phần bị xóa trên cùng NST số 15, gen đột biến di truyền từ cha không được biểu hiện, gen di truyền từ mẹ được biểu hiện có liên quan đến hội chứng Prader-Willi. Một bệnh khác là hội chứng Beckwith-Wiedemann (BWS), được đặc trưng bởi kích thước sinh lớn bất thường và khuynh hướng dẫn đến khối u. BWS được gây ra bởi sự mất dấu ấn và di truyền tại các vị trí cụ thể trên nhiễm sắc thể 11.
3.2. Cơ sở sinh hóa tương tác giữa các gen không alen
Tương tác gen là sự tác động qua lại của các gen trong quá trình hỉnh thành kiểu hình. Về thực chất, các gen không trực tiếp tương tác với nhau mà chỉ các sản phẩm của chúng như các enzym (bản chất protein) hay giữa các sản phẩm sau hoạt động của enzym phối hợp gây ảnh hưởng qua lại theo những cách khác nhau dẫn đến sự biểu hiện kiểu hình khác nhau.
Sản phẩm enzym của các gen không alen tương tác với nhau:
- Để làm xuất hiện một kiểu hình mới trong trường hợp tương tác bổ trợ
- Enzym do gen này ức chế hoạt động của enzym được tạo ra do gen khác không alen trong trường hợp tương tác át chế
- Cùng làm tăng cường biểu hiện của một tính trạng trong trường hợp tương tác cộng gộp
Dưới dây là các dạng tương tác gen không alen
Tương tác đa gen giữa các gen không alen là sự tác động qua lại giữa 2 hay nhiều gen không alen để cùng quy định một tính trạng theo các dạng:
Tương tác bổ sung (bổ trợ).
Tương tác át chế.
Tương tác cộng gộp.
3.2.1. Tương tác bổ sung
Tương tác bổ sung (tương tác bổ trợ) là kiểu tương tác trong đó các alen của mỗi locus riêng rẽ có biểu hiện kiểu hình riêng. Khi hai hoặc nhiều gen không alen cùng có mặt trong kiểu gen sẽ tạo nên một kiểu hình mới.
Hiện tượng tương tác bổ trợ biểu hiện ra nhiều tỷ lệ kiểu hình F2 khác nhau tùy thuộc vào sự tương tác đặc biệt giữa các gen không alen.
3.2.1.1. Tỷ lệ 9 : 3 : 3 : 1
Ở gà, hình dạng khác nhau của mào là kết quả tương tác giữa 2 gen không alen quy định. Trong đó:
A–B– : gà mào hình quả hồ đào
A–bb : gà mào hình hoa hồng
aaB– : gà mào hình hạt đậu
aabb : gà mào hình lá
Cơ sở sinh hóa
Như vậy, đây là kiểu tương tác giữa hai gen không alen. Trong đó, sự tương tác giữa hai gen trội A và B quy định một kiểu hình riêng. Ở đây, không có sự biến dạng của tỉ lệ phân ly kiểu hình 9 : 3 : 3 : 1.
Cho đến nay, cơ sở phân tử trong việc hình thành 4 hình dạng mào này vẫn chưa được rõ ràng nhưng nhìn một cách tổng quát nhất ta có thể đưa ra giả thuyết như sau.
Kiểu gen aabb sinh ra được sản phẩm gen nhưng sản phẩm gen đó không thể ảnh hưởng tới việc thay đổi hình dạng căn bản của kiểu hình mào gà hình lá.
Alen trội A sinh ra sản phẩm, sản phẩm này sẽ ảnh hưởng tác động qua lại với sản phẩm của gen quy định hình dạng mào gà hình lá để quy định mào hình hoa hồng.
Tương tự, alen trội B sinh ra sản phẩm, sản phẩm này sẽ ảnh hưởng tác động qua lại với sản phẩm của gen quy định hình dạng mào gà hình lá để quy định mào hình hạt đậu.
Khi sản phẩm của cả hai alen trội A và B tương tác với nhau sẽ xuất hiện dạng kiểu hình mới khác với các dạng kiểu hình còn lại là mào hình quả hồ đào.
3.2.1.2. Tỷ lệ 9 : 6 : 1
Ở loài bí Cucubirta peppo, có ba dạng quả đã được nghiên cứu là dạng quả tròn, quả dẹt và quả dài. Đã xác định hình dạng quả là kết quả tương tác giữa hai gen không alen quy định. Ở bí, hình dạng quả là kết quả tương tác giữa 2 gen không alen. Trong
đó:
A–B– : bí quả dẹt A–bb; aaB– : bí quả tròn
aabb : bí quả dài
Cơ sở sinh hóa
Bí quả dài và bí quả tròn là 2 dạng điển hình trong rất nhiều dạng của bí mùa hè (summer squash), hình dạng của quả bí được quy định bởi hai cặp gen không alen.
Điểm đáng chú ý ở đây đó là khi cả hai alen trội không len tương tác với nhau thì ta thu được kết quả vô cùng lý thú: hình dạng mới của quả bí sẽ xuất hiện.
Cơ sở phân tử chính xác về hiện tượng trên cho đến nay vẫn còn là một ẩn số chưa thật sự rõ ràng nhưng giả thuyết sau đây đưa ra đã được phần lớn các nhà khoa học chấp nhận.
Ở bí quả dài có kiểu gen aabb, do sự vắng mặt các sản phẩm của 2 alen A và B nên sản phẩm được tạo ra từ kiểu gen aabb sẽ cho kiểu hình là bí quả dài.
Nếu như thiếu đi một trong hai sản phẩm của hai alen A, B mà không phải cả hai thì hình dạng căn bản của bí quả dài sẽ biến mất, thay vào đó là hình dạng bí quả tròn.
Hình dạng bí quả dẹt có thể được xuất hiện thông qua sự biến đổi của hình dạng bí quả tròn do sự tương tác của cả 2 sản phẩm của 2 alen A và B.
3.2.1.3. Tỷ lệ 9 : 7
- VD1:
Ở ngô, màu sắc hạt là kết quả của sự tương tác giữa hai gen không alen. Trong đó:
A–B–: ngô hạt vàng
A–bb; aaB–; aabb: ngô hạt tím
Ở cây hoa đậu thơm, màu sắc hoa là kết quả của sự tương tác giữa hai gen không alen. Trong đó:
A–B–: cây đậu thơm hoa tía. A–bb; aaB–; aabb: cây đậu thơm hoa trắng.
Cơ sở sinh hóa
Kết quả trên được giải thích bởi sự tương tác của hai gen không alen theo 2 cách:
Cách 1: Tỉ lệ kiểu hình 9/16 cây đậu thơm hoa tía thu được ở F2 nói lên rằng màu sắc ở hoa chỉ xuất hiện khi 2 gen trội không alen tương tác với nhau. Tuy vậy, các quan niệm trước đây chỉ mường tượng rằng sự có mặt của cả hai alen trội trong kiểu gen sẽ quy định hoa tía, còn nếu kiểu gen vắng mặt 1 trong hai alen trội hoặc vắng mặt cả hai thì sẽ quy định hoa màu trắng.
Ngày nay, cơ sở phân tử của hiện tượng trên đã được giải thích rõ ràng theo sơ đồ 1.1.
Theo cách giải thích này thì sắc tố tía được tạo ra là nhờ hai yếu tố là tiền chất do gen A tạo ra và enzim do gen B tạo ra xúc tác phản ứng biến tiền chất A thành sắc tố tía. Theo sơ đồ thì mỗi bước được điều khiển bởi sản phẩm của mỗi gen, có nghĩa gen A điều khiển sự biến đổi từ dạng tiền thân không màu 1 sang dạng tiền thân không màu 2, gen B điều khiển sự biến đổi từ dạng tiền thân không màu 2 sang sản phẩm P cuối cùng của chuỗi phản ứng sinh hóa là sắc tố tía.
Vì vậy, các kiểu gen A–B– đủ hai yếu tố nên sắc tố tía được tổng hợp. Các kiểu gen A–bb; aaB– thiếu một yếu tố và kiểu gen aabb thiếu cả hai yếu tố nên sắc tố tía không được tổng hợp.
Cách 2: Kết quả trên còn có thể được giải thích do hiện tượng đồng át chế lặn xảy ra tức aa > B, b đồng thời bb > A, a. Quan niệm này cũng được rất nhiều người đồng tình. Gen A Gen B
↓ ↓
Enzym A Enzym B
↓ ↓
Dạng tiền thân không màu 1 →Dạng tiền thân không màu 2 → Sản phẩm
P (sắc tố tía)
3.2.2. Tương tác át chế
Tương tác át chế là kiểu tương tác giữa hai hay nhiều gen không alen cùng tham gia quy định một tính trạng. Trong đó, một gen trội (hoặc gen lặn) lấn át sự biểu hiện kiểu hình của gen thuộc locus khác không alen.
Kiểu tương tác này cũng làm biến đổi tỷ lệ phân ly kiểu hình ở F2. Át chế trội xảy ra khi B > C (hoặc ngược lại C > B) và át chế lặn xảy ra khi bb > cc (hoặc cc > bb).
3.2.2.1. Tỷ lệ 12 : 3 : 1 (Át chế trội)
Ở bí mùa hè (Summer squash), sự hình thành màu sắc quả bí là do 2 cặp gen không alen quy định. Trong đó:
B–C–; B–cc: bí trắng bbC–: bí vàng bbcc: bí xanh
Tiến hành phép lai ta thu được kết quả sau:
Pt/c: Bí trắng × Bí xanh
F1: 100% bí trắng. Cho các cơ thể con lai thụ phấn.
F2: 12 bí trắng : 3 bí vàng : 1 bí xanh
Cơ sở sinh hóa
Để giải thích kết quả của phép lai trên ta quan sát vào sơ đồ 1.3
Alen trội C là alen cần thiết cho sự chuyển đổi màu từ bản chất xanh sang vàng của quả bí. Còn alen trội B có khả năng sản sinh ra chất ức chế, chất này sẽ ức chế quá trình chuyển đổi màu sắc từ trắng sang xanh.
Sơ đồ (a) sơ đồ1.3 mô tả con đường hình thành trái bí trắng trên cơ sở kiểu gen B–C–; B–cc. Theo sơ đồ này thì alen B sản sinh ra chất ức chế quá trình chuyển đổi màu từ trắng sang xanh, vì thế bước tiếp theo chuyển đổi màu từ bản chất xanh sang vàng sẽ không thực hiện được nên các cơ thể mang kiểu gen B–C–; B–cc đều có kiểu hình là bí trắng.
Sơ đồ (b) sơ đồ 1.3 mô tả con đường hình thành trái bí vàng trên cơ sở kiểu gen bbC–. Do không có mặt alen B nên chất ức chế không được tạo thành, mặt khác lại có
381
mặt alen C trong kiểu gen nên các cơ thể mang kiểu gen bbC– đều có kiểu hình bí vàng.
Sơ đồ (c) sơ đồ 1.3 mô tả con đường hình thành trái bí xanh trên cơ sở kiểu vắng mặt alen C trong kiểu gen nên các cơ thể mang kiểu gen bbcc đều có kiểu hình là bí xanh.
Con đường hình thành trái bí màu trắng
Gen B Gen C
↓ ↓
Chất ức chế (B) Sản phẩm gen C
↓ ↓
Sắc tố trắng → Sắc tố xanh → Sắc tố vàng
↓
Trái bí trắng (B-C-; B-cc)
Con đường hình thành trái bí màu vàng
Gen C ↓
Không chất ức chế Sản phẩm gen C ↓ ↓
Sắc tố trắng → Sắc tố xanh → Sắc tố vàng → Trái bí vàng
Con đường hình thành trái bí xanh
Không chất ức chế
Sắc tố trắng → Sắc tố xanh → Trái bí xanh (bbcc)
3.2.2.2. Tỷ lệ 13 : 3 (Át chế trội)
Ở gà, sự hình thành màu sắc lông là do hai cặp gen không alen quy định.
Trong đó:
B–C–; bbC–; bbcc: gà lông trắng
B–cc: gà lông đen
Cơ sở sinh hóa
Kết quả của phép lai trên được giải thích do tương tác át chế giữa hai gen không
alen B và C, cụ thể là tương tác át chế trội: CC > B, b.
Alen B là alen quy định việc tạo màu cho bộ lông gà.
382
Sản phẩm của alen C ức chế sự biểu hiện ra kiểu hình của các sản phẩm được sinh ra từ alen B và alen b. Do vậy, các cơ thể có kiểu gen B–C–; bbC– sẽ đều có kiểu hình là gà lông trắng.
Các cơ thể mang kiểu gen bbcc cho kiểu hình gà lông trắng là do gen c tạo màu
ở trạng thái đồng hợp lặn.
Các cơ thể mang kiểu gen B–cc, do sản phẩm tạo màu lông của alen B không bị ức chế nên chúng mang kiểu hình là gà lông đen.
Ví dụ: Màu cánh hoa xanh da trời (blue) của hoa báo xuân di truyền theo mô hình "13: 3" . Màu xanh lơ do sắc tố man-vi-đin (malvidine) tạo ra. Sắc tố này là an-tôxi-a-nin (anthocyanine) đã mê-tyl hóa, được hình thành nhờ xúc tác của một enzym được mã hóa bởi gen trội A, còn alen lặn a không tạo được enzym. Nhưng sự sản xuất sắc tố này bị ngăn cản bởi gen trội B ở lôcut khác. Nên phép lai AaBb × AaBb sinh thế hệ con phân li trung bình là 13/16 trắng (A-B- + aa- -) + 3 xanh (A-bb).

Ngoài tương tác át chế lặn, 2 gen không alen B, C còn tương tác theo kiểubổ trợ do F1 xuất hiện kiểu hình mới là chuột xám nâu (agouti).
Tỷ lệ phân ly 9 : 3 : 4 ta cũng gặp ở ngựa thể hiện qua sơ đồ lai sau:
Pt/c: BBCC (ngựa đen) × bbcc (ngựa trắng)
F1: BbCc (100% ngựa đen)
BbCc (ngựa đen) × BbCc (ngựa đen)
3.2.2.3. Tỷ lệ 9 : 4 : 3 (Át chế lặn)
- Ở chuột, màu sắc lông là do sự tương tác giữa hai gen không alen quy định.
Trong đó:
B–C–: chuột xám nâu (sợi lông có hai đầu mút màu đen, đoạn giữa màu vàng), còn gọi là chuột agouti.
B–cc; bbcc : chuột bạch tạng,
bbC–: chuột đen
Cơ sở sinh hóa
Kết quả của phép lai trên được giải thích thông qua tương tác át chế giữa hai gen không alen B và C, cụ thể là tương tác át chế lặn: cc > B, b.
Alen C quy định việc hình thành sắc tố đen của sợi lông, alen B quy định việc hình thành sắc tố vàng trên sợi lông.
Sản phẩm của alen c ức chế sự biểu hiện ra kiểu hình của các sản phẩm được sinh ra từ alen B và alen b. Do vậy, các cơ thể có kiểu gen B– cc; bbcc sẽ đều có kiểu hình là chuột bạch tạng.
Các cơ thể mang kiểu gen bbC– có alen C quy định việc hình thành sắc tố đen trên sợi lông nên chúng đều có kiểu hình là chuột đen.
F2: 9 ngựa đen (B-C-) : 3 ngựa nâu (bbC-) : 4 ngựa trắng (B-cc; bbcc)
Tuy nhiên kết quả của phép lai trên lại được giải thích theo một cách hoàn toàn khác so với tỷ lệ 9 : 4 : 3 thu được trong ví dụ chuột xám nâu lai với nhau: Tính trạng màu sắc lông ở ngựa do 2 gen không alen B, C tương tác qua lại theo kiểu át chế quy định. Trong đó, gen C quy định việc tổng hợp enzyme C, enzyme này có vai trò xúc tác cho quá trình sinh hóa biến đổi màu sắc lông từ dạng không màu sang màu nâu, gen B quy định việc tổng hợp enzyme B xác tác cho quá trình sinh hóa biến đổi màu sắc lông từ màu nâu sang màu đen theo sơ đồ sau:
Gen C Gen B
↓ ↓
Enzyme C Enzyme B ↓ ↓



Mặt khác, các cơ thể mang kiểu gen B–C– có đầy đủ alen B và C nên chúng có kiểu hình là chuột xám nâu. 384
Lông trắng
→ Lông nâu → Lông đen
Do vậy, các cơ thể có kiểu gen B–C– sẽ cho kiểu hình lông đen. Các cơ thể có kiểu gen bbC– sẽ cho kiểu hình lông màu nâu. Cơ thể mang kiểu gen B–cc không có gen trội C nên thiếu một giai đoạn đầu trong chuỗi phản ứng sinh hóa hình thành màu sắc lông nên kiểu hình là lông trắng. Và như vậy, cơ thể có kiểu gen bbcc thiếu cả 2 alen trội B, C nên hiển nhiên kiểu hình sẽ là lông trắng.
3.2.3. Tương tác cộng gộp
Tương tác cộng gộp là kiểu tương tác giữa hai hay nhiều gen không alen để cùng quy định một tính trạng. Trong đó, mỗi gen trội cùng alen hay khác alen đóng góp một vai trò như nhau vào sự phát triển của cùng một tính trạng.
Có hai kiểu tương tác cộng gộp:
Tương tác cộng gộp có tích lũy
Tương tác cộng gộp không tích lũy
3.2.3.1. Tương tác cộng gộp có tích lũy
Một số tính trạng có liên quan tới năng suất của nhiều vật nuôi, cây trồng như: số lượng hạt trên bắp ngô, sản lượng trứng ở gia cầm … cũng như các tính trạng màu da, chiều cao, màu mắt ở người bị chi phối bởi sự tác động cộng gộp của nhiều gen không alen. Các tính trạng này còn được gọi là tính trạng đa gen hay tính trạng số lượng.
Sự phân ly kiểu hình của các tính trạng đa gen không cho tỷ lệ chất lượng thương phẩm rõ ràng. Các cá thể có biểu hiện kiểu hình dao động khác nhau do nhận nhiều hay ít gen (thường là alen trội) và có thể xếp chúng theo mức độ biểu hiện thành một dãy liên tục.
Dưới đây là một ví dụ điển hình về tương tác cộng gộp.
Màu sắc da ở người là tính trạng đa gen do nhiều gen không alen tương tác quy định.
Phép lai được tiến hành theo sơ đồ sau:
Pt/c: AABBCC (da rất đen) × aabbcc (da rất trắng)
F1: AaBbCc (100% da nâu)
F1 × F1: AaBbCc (da nâu) × AaBbCc (da nâu)
F2: 1 da rất trắng :6 da trắng :15 da trắng vừa : 20 da nâu : 15 da đen vừa : 6 da đen : 1 da rất đen.
Hình 3. 3. Tương tác cộng gộp có tích lũy hình thành màu sắc da ở người
Cơ sở sinh hóa
Tính trạng màu sắc da ở người là do tương tác giữa 3 cặp gen không alen (A và a, B và b, C và c) quy định.
Trong đó mỗi alen trội A, B, C đều có vai trò như nhau trong việc quy định màu sắc da. Kiểu gen mang càng nhiểu alen trội thì kết quả kiểu hình là da càng đen.
Các alen lặn a, b, c không có vai trò trong việc tích lũy sắc tố da. Kiểu gen mang càng nhiều alen lặn thì kiểu hình là da càng trắng.
Có thể giải thích hiện tượng tương tác cộng gộp không tích lũy của các gen không cùng alen bằng cơ chế tiến hóa cấp phân tử. Do đột biến lặp đoạn hoặc chuyển đoạn dẫn đến trên các NST có thể mang các locus gen khác nhau nhưng cùng tạo ra các sản phẩm Protein như nhau. Lượng sản phẩm Protein tạo ra càng nhiều thì có tác động càng mạnh lên sự biểu hiện của tính trạng.
Kết quả ở F2 thông qua phép lai trên ta thu được tỉ lệ một dãy các kiểu hình từ da rất trắng đến da rất đen.
3.2.3.2. Tương tác cộng gộp không tích lũy
Hình dạng quảở cây tề do haicặp gen không alentương tác quy định. Trong đó:
A–B–; aaB–; A–bb: quả hình tam giác
aabb: quả hình bầu dục
Tiến hành phép lai thu được kết quả như sau:
Pt/c: AABB (Quả hình tam giác) × aabb (Quả hình bầu dục)

F1: AaBb (100% quả hình tam giác)
F2: 15 quả hình tam giác (A-B-; A-bb; aaB-) : 1 quả bầu dục (aabb)
Cơ sở sinh hóa
Kết quả của phép lai được giải thích theo 2 cách:
Cách 1: hình dạng quả ở cây tề do tương tác cộng gộp không tích lũy của hai cặp gen không alen (A và a, B và b) quy định. Trong đó, sự có mặt của alen trội (không phụ thuộc vào số alen trội) trong kiểu gen quy định kiểu hình là quả hình tam
giác. Nếu trong kiểu gen không có alen trội thì các cơ thể đó sẽ mang kiểu hình quả bầu dục.
Cách 2: Ngoài ra còn có một giả thuyết khác được đưa ra để giải thích kết quả của phép lai trên. Người ta cho rằng tính trạng hình dạng quả ở cây tề là do tương tác
át chế trội kép giữa 2 cặp gen không alen tức A > B, b và B > A, a.
3.2.4. Tương tác giữa gen trong nhân và gen ngoài nhân
Hình 3.4. Thí nghiệm chứng minh sự di truyền theo hệ mẹ của hiện tượng bất thụ đực.
Tính bất dục do nhiều nguyên nhân, bất dục đực (không tạo phấn hoa hay tạo phấn hoa không có khả năng thụ tinh) ở thực vật có các trường hợp sau:
- Do gene nhân quy định, như gene ms ở cây ngô.
- Do ảnh hưởng của điều kiện môi trường như độ ẩm, quang chiếu, khả năng cung cấp chất dinh dưỡng không đáp ứng đúng nhu cầu sinh lý của cây.
* Ví dụ: gene ms ở cây ngô
- Do lai xa cũng đưa đến các cơ thể lai không có hạt phấn vì NST có nguồn gốc khác nhau không thể tiếp hợp nhau trong giảm phân. Những hiện tượng bất dục này đều có ý nghĩa hạn chế chỉ có bất dục bào chất đực là có vai trò quan trọng. Đó là trường hợp bất dục của hạt phấn bắt nguồn từ tế bào chất, còn nhân thì có thể có điều chỉnh được nhờ đó có thể dùng các cây bất dục bào chất đực để phát huy ưu thế lai ở các đối tượng ngô, cao lương, củ cải đường...
Ví dụ: Xét mối quan hệ giữa kiểu gene, kiểu bào gene và kiểu hình của bắp được sử dụng trong lai một tính mà không cần khử đực ở cây mẹ
STT Kiểu gene Kiểu bào Kiểu hình
gene (hạt phấn)
1 Rfrf S (bất dục) Hỏng
2 Rfrf N (hữu dục) Tốt
3 RfRf hoặc Rfrf N Tốt
4 RfRf hoặc Rfrf S Tốt
Vậy hạt phấn của ngô chỉ bị mất hoạt tính khi có yếu tố bất dục trong tế bào chất mà lại thiếu thiếu gene phục hồi hữu dục (Rf) ở trong nhân, alen của gene này là rf là gene cũng cố tính bất dục.
Cây được phục hồi hữu dục RfrfS cho tự thụ phấn thì ở đời sau sẽ có 1/4 rfrf có hạt phấn hỏng. Nếu lấy bắp của dạng rfrfS thụ phấn của rfrfN, thì phấn hoa của toàn bộ đời sau sẽ bị hỏng, cây này chỉ còn bắp mang nhị cái. Đó là cách dùng phương pháp di truyền để khử cờ ngô. Khi sản xuất hạt giống, những cây này muốn có hạt thì phải thụ phấn hữu dục của những cây bình thường. Nếu muốn dùng những hạt đó để sau này trồng lại thì cây bố phải có kiểu gene RfRf và kiểu bào gene N hoặc S.
Trong sản xuất giống ngô có thể dùng tổ hợp dòng thuần dạng 1 làm cây mẹ và dạng 3 hoặc dạng 4 đồng hợp tử làm cây bố. Như thế sẽ đỡ mất công khử đực ở cây mẹ và hạt lai thu được từ cây mẹ sẽ có kiểu gene Rfrf, kiểu bào gene S. Kiểu gene này đảm bảo được sự thụ phấn bình thường lúc trồng trong sản xuất.
Bất thụ đực tế bào chất ở ngô liên quan đến 2 plasmid dạng thẳng S1 và S2. Chúng ở trong ty thể cùng với mtADN. Một trong những tính chất khó hiểu của plasmid này là chúng có thể thực hiện tái tổ hợp với mtADN.
3.3. Một số câu hỏi, bài tập tương tác gen
3.3.1. Câu hỏi
388
Câu 1: Hiện tại, bạn đang nghiên cứu về sự tổng hợp threonin ở E.coli. Để tìm các gen mã hóa các protein quan trọng cho tổng hợp threonin, bạn tiến hành gây đột biến để tìm các đột biến yêu cầu bổ sung threonin cho phát triển. Bạn đã tìm thấy có 9 dòng đột biến và tất cả đều cần bổ sung threonin vào môi trường nuôi cấy để phát triển. Sau đó bạn tiến hành cis-trans test với tất cả các dòng đột biến bạn có. Kết quả thu được như bảng sau (+: phát triển; -: không phát triển):
=> Tạo kiểu hình bình thường (kiểu dại).
+ Nếu khuẩn lạc không phát triển
=> Hai đột biến thuộc cùng một gen nên không bù trừ cho nhau
=> Tạo kiểu gen đồng hợp
=> Tạo kiểu hình đột biến. Từ đó, ta có:
- 1,3,5,6,9 thuộc cùng một gen.
- 2,8 thuộc cùng một gen.
- 4 không bù cho mình, bù cho những dòng đột biến còn lại => gen độc lập
a. Có bao nhiêu gen mã hóa các enzym tham gia vào quá trình tổng hợp threonin? Giải thích.
b. Khi xem xét quá trình tổng hợp threonin, người ta nhận thấy có
- 7 không bù cho mình, bù cho những dòng đột biến còn lại => gen độc lập => Có 4 gen tham gia tổng hợp threonin.
b) sơ đồ chuyển hóa:
A Mt1 D Mt7 C Mt4 B,E Mt2 Threonin

Giải thích
- Các chủng đột biến không sinh trưởng được trong môi trường chỉ chứa A => A là chất đầu dãy chuyển hóa
5 cơ chất tham gia vào quá trình đó. Người ta tiến hành nuôi cấy các đột biến ở trên trong các môi trường chứa các cơ chất khác nhau để xác định trình tự quá trình biến đổi. Kết quả thu được trình bày ở bảng sau (+: phát triển; -:
không pháttriển):
Hãy xác định trật tự chuyển hóa của các chất A, B, C, D, E và các gen mã hóa các enzym xúc tác sự chuyển hóa tương ứng. Giải thích.
Trả lời
a)Ta thấy:
- Khi cho các dòng đột biến giống nhau lai với nhau
=> Không phát triển, không tạo khuẩn lạc.
=> Đột biến trên cùng một gen nên không bù trừ được cho nhau.
=> Tạo kiểu hình đột biến.
- Khi cho những dòng đột biến khác nhau lai với nhau:
+ Nếu khuẩn lạc phát triển
=> Hai đột biến thuộc hai gen khác nhau bù trừ cho nhau tạo kiểu gen dị hợp
- Chỉ có chủng đột biến Mt1 sinh trưởng được trong môi trường chỉ chứa D => D là chất thứ 2 của dãy chuyển hóa và chủng Mt1 bị đột biến ở bước chuyển hóa chất A thành chất D
-Trong môi trường chỉ chứa C, có hai chủng Mt1 và Mt7 sống được => chất C là chất thứ 3 của dãy chuyển hóa và chủng Mt7 là chủng bị đột biến ở bước chuyển hóa chất D thành chất C
-chủng Mt4 sống được trong môi trường chỉ chứa B và E => chất B và E cùng là chất thứ 4 trong dãy chuyển hóa và chủng Mt2 bị đột biến ở bước chuyển hóa chất C thành chất B hoặc E
-chủng Mt2 chỉ sống được trong môi trường chứa Threonin => chủng này bị đột biến ở bước chuyển hóa chất B hoặc chất E thành Threonin
Câu 2: Trích đề IBO 2012 Có một số các chủng đột biến dinh dưỡng được phân lập từ các chủng nấm mốc bánh mỳ đỏ kiểu hoang dại Neurospora crassa đáp ứng với việc bổ sung thêm các chất nhất định vào môi trường nuôi cấy, nếu phát triển (+) hoặc không phát triển (0). Bảng dưới đây cho biết các số liệu về các thể đột biến gen đơn.
390
a. Hãy chỉ ra thứ tự chuyển hóa (1-5) của 5 sản phẩm chuyển hóa trong quá trình trao đổi trong phiếu trả lời
b. Hãy chỉ ra chủng nào bị ngăn cản ở mỗi một bước trong 4 bước của quá trình chuyển hóa nêu trong phiếu trả lời
Câu 3. A, B, C, D là các chất chuyển hoá trung gian (không theo đúng thứ tự) trong một con đường hoá sinh của tế bào. Người ta tìm thấy 4 thể đột biến khác nhau kí hiệu từ D1- D4. Khi nuôi cấy 4 thể đột biến này lần lượt trong các môi trường được bổ sung chất A, B, C và D, người ta thu được kết quả như sau: D1 chỉ sinh trưởng trong các môi trường có A hoặc D; D2 chỉ sinh trưởng trong các môi trường chứa A hoặc B hoặc D; D3 chỉ sinh trưởng trong môi trường có D; D4 chỉ sinh trưởng trong môi trường có A hoặc B hoặc C hoặc D. Hãy vẽ sơ đồ các bước chuyển hoá của con đường hoá sinh trên và chỉ ra những bước chuyển hoá bị ức chế tương ứng ở các thể đột biến (D1- D4). Giải thích.



Trả lời
Sơ đồ chuyển hoá theo trình tự : C
→ B → A → D
- D1 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá B thành A.
- D2 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá C thành B.
- D3 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá A thành D.
- D4 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá hình thành chất C.
Ta có thể suy ra được trình tự các bước chuyển hoá dựa trên nguyên lý là nếu thể đột biến nào cần phải bổ sung tất cả các chất thì thể đột biến đó bị hỏng gen qui định enzim chuyển hoá tiền chất đầu tiên của con đường chuyển hoá.
Thể đột biến nào chỉ cần bổ sung một chất thì chất đó là sản phẩm cuối cùng của con đường chuyển hoá.
Câu 4. (Trích đề IBO năm 2015) Ở nấm mốc, sự sinh tổng hợp axit amin arginine cần bước chuyển hóa tiền chất thành ornithine; chất này được chuyển hóa tiếp thành citruline là tiền chất trực tiếp được chuyển hóa thành arginine. Mỗi phản ứng trong 3 bước này được xúc tác bởi một enzym riêng rẽ; mỗi enzym được mã hóa bởi một gen (xem Hình)
(tiền chất)
A: Con đường sinh tổng hợp arginine ở nấm mốc. B: Thiết kế thí nghiệm để xác định chuyển hóa: dải màu nâu đen chỉ Nấm mốc sinh trưởng. Chủng kiểu dại mọc trên môi trường tối thiểu, trong khi các chủng đột biến chỉ mọc trên các môi trường được bổ sung (+) chất dinh dưỡng đặc thù. Chọn câu trả lời đúng nhất

A. Các chủng đột biến lớp III có thể mang đột biến ở nhiều hơn 1 gen liên quan đến con đường sinh hóa này
B. Các chủng đột biến lớp I thiếu cả 3 enzym
C. Các thể đột biến thiếu hai enzym A và C có thể mọc nếu môi trường
được bổ sung đồng thời Orthnithine và Citruline
D. Giả sử một chất ức chế enzym B được bổ sung vào môi trường nuôi chủng kiểu dại, gen B sẽ bị bất hoạt
E. Tất cả đều sai
Đáp án : E
3.3.1. Bài tập
Bài tập 1 : Ở phả hệ sau, II-2 là người đầu tiên trong gia đình biểu hiện các triệu chứng liên quan đến đột biến ở gen ti thể (Hội chứng Merff).
a. Hãy đưa ra hai lí do để giải thích hợp lí nhất về việc tại sao người mẹ I-1 không bị bệnh nhưng II-2 bị bệnh này?
b. Bằng cách nào có thể phân biệt hai lí do nêu trong giải thích ở câu a?

Bài tập 2: Gen hiệu ứng mẹ ở ruồi giấm, kí hiệu t là alen lặn, làm ngăn cản sự phát triển của các cấu trúc đầu – đuôi. Ruồi đực kiểu dại được lai với ruồi cái có kiểu gen chưa biết tạo ra ấu trùng không có cấu trúc đầu đuôi và thường chết ở giai đoạn phôi. Hãy xác định kiểu gen và kiểu hình của ruồi cái ở phép lai này. Xác định kiểu gen và kiểu hình của bố mẹ của ruồi cái đó.
Bài giải:
Do ấu trùng tạo ra không có cấu trúc đầu đuôi → kiểu gen của ruồi cái là tt nhưng không bị bệnh do ruồi bố mẹ của ruồi cái là: Tt x Tt hoặc Tt x tt.
Bài tập 3. Ở cây bốn giờ Mirabilus jalapa cùng một cây, một số các nhánh chứa lá màu xanh, một số nhánh chứa lá màu trắng, một số nhánh chứa lá đốm có những mảng màu xanh và trắng xen kẽ. Màu xanh của lá và thân do diệp lục được tổng hợp bởi sự quy định của gen nằm trong lục lạp.
a. Giải thích vì sao cây này có một số nhánh có lá khác nhau như vậy.
b. Khi lấy noãn bào từ nhánh cây lá xanh với hạt phấn từ hoa của bất kì nhánh nào người ta thấy F1 tạo ra tất cả đều có lá màu xanh. Khi thụ phấn noãn của nhánh đốm với hạt phấn từ bất kì nhánh nào, 85% cây con tạo ra có một số nhánh màu xanh, một số có lá màu trắng, một số cá lá đốm; 10% cây con chỉ có lá xanh và 5% cây con có lá trắng nhưng sau đó bị héo và chết sau khi nảy mầm. Hãy giải thích kết quả này. Làm thế nào noãn bào tử nhánh lá đốm tạo ra thế hệ con đều có lá xanh hoặc lá trắng?
Tại sao các cây có lá trắng hoàn toàn nhanh chóng bị chết?
c. Với câu trả lời ở phần b, làm thế nào các cây lá đốm có nhánh trắng vẫn khỏe mạnh bình thường?
Trả lời:
a. Do gen trong lục lạp phân chia độc lập với phân chia nhân. Tế bào hợp tử chứa cả lục lạp bình thường – gen không bị đột biến và cả lục lạp màu trắng – do gen đột biến. Khi tế bào phân chia thì tế bào chất được phân chia không đồng đều về các
tế bào vì thế tế bào nào nhận được chỉ lục lạp bình thường từ đó tạo nhánh toàn lá xanh; tế bào nào nhận được hai loại lục lạp sẽ có lá đốm, tế bào nào nhận được chỉ lục lạp chứa gen đột biến sẽ có màu trắng.
b. Do gen trong tế bào chất của noãn là chủ yếu, còn hạt phấn chứa tinh tử có lượng tế bào chất không đáng kể vì thế gen do tế bào chất quy định di truyền theo dòng mẹ
+ Noãn của cây có lá xanh – toàn lục lạp không bị đột biến dù dùng hạt phấn của cành nào → con đều có lá màu xanh.
+ Noãn của cây có lá đốm → con F1 sẽ chủ yếu là lá đốm, một tỉ lệ nhỏ có cây lá xanh và một tỉ lệ nhỏ cho lá trắng . Vì tế bào mẹ đại bào tử phân chia tế bào chất ngẫu nhiên tạo ra các noãn khác nhau.
- Cây lá trắng chết sau khi mọc mầm vì không có sắc tố quang hợp nên không sống được.
c. Cây đốm vẫn khỏe mạnh vì các lá có lục lạp bình thường quang hợp tạo ra sản phẩm nuôi dưỡng cả cây.
Bài tập 4. Ở ngô có hai dòng tế bào chất: dòng N hữu thụ và dòng S bất thụ ; Gen ở trong nhân: alen Rf – có khả năng phục hồi hữu thụ; alen rf – không có khả năng phục hồi. Chọn P để thu được F1 đồng nhất kiểu gen SRfrf - có ưu thế lai cao
Trả lời:
P. ♀ Srfrf x ♂N(S)RfRf → F1: SRfrf.
Bài tập 5: Cho Pt/c. ♀ lông đen x ♂ lông trắng→ F1: 100% lông đen
a. Cho ♀F1 lai với P♂ lông trắng (trắng do gen lặn quy định). Hãy xác định kết quả phép lai?
b. Cho F1 giao phối với nhau được kết quả KG,KH ở F2 như thế nào?
Cho rằng màu lông do 1 gen quy định nằm trên NST thường và chỉ có 2 KH trên?
Trả lời:
- TH1: Quy luật phân li.
- TH2: Tương tác gen trong nhân và tế bào chất.
P. AA x aa → F1. Aa –đen
Pb. ♀ Aa (F1) x ♂ aa (P) → 1Aa : 1aa (bất thụ)
F1xF1: Aa xAa → F2: 3A-: 1aa (bất thụ)
Bài tập 6. Cho phả hệ sau theo dõi sự di truyền của một bệnh rất hiếm gặp.
a. Hãy cho biết quy luật di truyền chi phối bệnh này?
b. Có cá thể nào trong phả hệ mang đột biến gây bệnh mặc dù không biểu hiện tính trạng bệnh không? Hãy chỉ ra các cá thể đó trong phả hệ (viết chữ số La mã chỉ số thế hệ và số thường chỉ thứ tự từ trái sang phải).
Bài giải:
a. Bệnh hiếm gặp nên tần số alen gây bệnh sẽ thấp, những người kết hôn với người trong gia đình không mang alen bệnh. Chỉ có thể giải thích đây là hiện tượng in vết hệ gen, in vết từ mẹ (III.8 Aa sinh con không bị bệnh) và bệnh do gen trội quy định.
b. Người mẹ II.2 và bố II.1 mang gen bệnh (Aa) nhưng không biểu hiện do được in vết từ thế hệ trước.
Bài tập 7 (Câu 15 đề chọn đội tuyển Olympic sinh học năm 2014 - Ngày 2)
Ở một loài, alen A quy định lông mượt, còn alen a quy định lông xù. Khi lai con cái thuần chủng lông mượt với con đực lông xù, được F1 có tỷ lệ 1 con lông mượt: 1 con lông xù. F1 giao phối với nhau được F2 có tỷ lệ 1 con lông mượt: 1 con lông xù. Giải thích kết quả của phép lai và viết sơ đồ lai từ P đến F2
Hướng dẫn trả lời
- Kết quả ở F1 có tỷ lệ 1 con lông mượt: 1 con lông xù phân bố đều ở hai giới tính, do đó không thể giải thích được bằng di truyền liên kết giới tính và ảnh hưởng của giới tính, Vì cả hai trường hợp này tuy đều cho F1 có tỷ lệ kiểu hình 1:1 nhưng phân hoá giới tính, ví dụ như 1 cái mắt đỏ: 1 đực mắt trắng ở ruồi giấm.
- Kết quả phép lai chỉ có thể giải thích bằng sự di truyền in vết, trong đó alen lặn (a) trong quá trình hình thành giao tử đã được in vết (a*), nghĩa là nó bị methyl hoá xitozin theo hướng hoạt hoá. Do đó với sự hiện diện của a* ở thể dị hợp tử có biểu hiện lông xù.
Sơ đồ lai
P: ♀ Lông mượt x ♂ Lông xù
AA aa
F1: Aa: Aa* → a*- hoạt hoá (in vết)
1 lông mượt : 1 lông xù
F2: 1 AA: 1Aa: 1Aa*: 1aa*
1 lông mượt : 1 lông xù

Bài tập 8 (Câu 15 đề chọn đội tuyển Olympic sinh học năm 2014 - Ngày 2)
Ở cây ngô, alen A quy định lá màu xanh đậm nằm trên nhiễm sắc thể số 1, alen lặn a quy định lá màu xanh nhạt; alen B nằm trên nhiễm sắc thể số 5 quy định bắp ngô to, alen lặn b quy định bắp ngô nhỏ. Cho hai dòng ngô thuần chủng lá màu xanh đậm, bắp to và lá màu xanh nhạt, bắp ngô nhỏ giao phấn với nhau được F1. Các cây F1 đều có khả năng sinh sản bình thường. Khi một cây ngô F1 tự thụ phấn tạo ra một số lượng lớn cá thể ở F2, trong đó 1/4 số cá thể lá màu xanh nhạt, bắp nhỏ. Hãy giải thích kết quả ở F2.
Hướng dẫn trả lời
- Hai dòng thuần giao phấn với nhau cho F1 dị hợp tử về hai cặp gen nằm trên hai nhiễm sắc thể khác nhau. Bình thường một cây F1 khi tự thụ phấn sẽ phải cho ra 1/16 cá thể đời con có lá màu xanh nhạt, bắp nhỏ (aabb). Nhưng trong trường hợp này cây tự thụ phấn lại cho ra tỉ lệ 1/4 aabb nên ta có thể cho rằng đã xảy ra đột biến chuyển đoạn qua lại giữa nhiễm sắc thể số 1 và số 5 dẫn đến alen a nằm trên cùng một nhiễm sắc thể với alen b.
- Hơn thế nữa alen a và alen b phải nằm rất gần điểm đứt gẫy chuyển đoạn khiến cho alen a và b nằm rất gần nhau dẫn đến chúng luôn di truyền cùng nhau. Cây ngô mang đột biến này hoàn toàn hữu thụ nên ta có thể cho rằng đó là cây đột biến chyển đoạn đồng hợp vì nếu là chuyển đoạn dị hợp sẽ bất thụ 50%.
Bài tập 9: Tương tác giữa 1 gen nằm trên NST thường và 1 gen nằm trên NST giới tính (Trích đề thi chọn HSG quốc gia năm học 2015-2016)

Có hai dòng ruồi giấm thuần chủng đều có mắt màu đỏ tươi được kí hiệu là dòng I và dòng II. Để nghiên cứu quy luật di truyền chi phối tính trạng, người ta đã thực hiện hai phép lai dưới đây:
Phép lai 1: Lai các con cái thuộc dòng I với các con đực thuộc dòng II; F1 thu được 100% ruồi con đều có màu mắt kiểu dại.
Phép lai 2: Lai các con cái thuộc dòng II với các con đực thuộc dòng I; F1 thu được 100% các con cái có màu mắt kiểu dại; 100% con đực có màu mắt đỏ tươi.
Từ kết quả của các phép lai trên có thể rút ra được những kết luận gì? Giải thích và viết sơ đồ lai minh họa.
Giải
- Từ kết quả của phép lai 1: Khi lai hai dòng ruồi thuần chủng đều có mắt đỏ tươi với nhau, đời con F1 đều thu được 100% cá thể có màu mắt kiểu dại màu mắt của ruồi giấm do hai gen tương tác kiểu bổ trợ
- Từ kết quả của phép lai 2 ta thấy có sự phân ly không đồng đều ở 2 giới có sự di truyền liên kết giới tính. Khi lai con cái thuộc dòng II với con đực thuộc dòng I
cho ra đời con có tất cả các con cái đều có màu mắt kiểu dại, còn các con đực đều có mắt đỏ tươi di truyền chéo.
- Một trong hai gen quy định tính trạng phải nằm trên nhiễm sắc thể giới tính X tại vùng không tương đồng với Y, gen còn lại nằm trên NST thường (vì nếu cả hai gen cùng nằm trên vùng tương đồng của cặp XY hoặc cùng nằm trên X tại vùng không tương đồng với Y thì sẽ không thu được kết quả như phép lai).
- Từ kết quả của phép lai 1 alen đột biến gây màu mắt đỏ tươi ở dòng I phải nằm trên NST thường. Lý do là nếu alen lặn nằm trên NST giới tính X thì tất cả các con đực sẽ có mắt màu đỏ tươi.
- Từ kết quả của phép lai 2 ta thấy gen lặn quy định màu mắt đỏ tươi phải nằm trên NST X vì tất cả các con đực đều có màu mắt đỏ tươi (có hiện di truyền chéo)
- Tổng hợp kết quả của cả phép lai 1 và 2, ta có thể viết sơ đồ lai chứng minh như sau:
Phép lai 1: P I (đỏ tươi) x II (đỏ tươi)
aaXBXB AAXbY
F1: AaXBXb Mắt kiểu dại
AaXBY Mắt kiểu dại
Phép lai 2: P II (đỏ tươi) x I (đỏ tươi)
AAXbXb aaXBY
F1: AaXBXb Mắt kiểu dại
AaXbY Mắt đỏ tươi
Bài tập 10: Tương tác gen và kiểm định χ2 (khi bình phương) (Trích đề thi chọn HSG quốc gia năm học 2013-2014)
Ở một loài thực vật, người ta thực hiện hai phép lai sau:
Phép lai I: Dòng 1 (hoa trắng) x Dòng 2 (hoa đỏ) được F1 100% hoa trắng.
Cho F1 tự thụ phấn, thu được F2 có 124 cây hoa trắng và 36 cây hoa đỏ.
Phép lai II: Dòng 1 (hoa trắng) x Dòng 3 (hoa đỏ) được F1 100% hoa trắng.
Cho F1 tự thụ phấn, thu được F2 có 122 cây hoa trắng và 38 cây hoa đỏ.
Biết rằng, kiểu gen và cách tác động của gen ở dòng 1 trong hai phép lai giống nhau.
a) Hãy dùng tiêu chuẩn χ2 (khi bình phương) để kiểm định sự phù hợp hay không phù hợp giữa số liệu thực tế và số liệu lí thuyết của 2 phép lai trên. Cho biết, χ2 lí thuyết = 3,84.






b) Giải thích kiểu tác động của gen đối với sự hình thành màu hoa ở kết quả của hai phép lai trên.
c) Cho rằng khi lai dòng 2 với dòng 3 được F1 100% hoa tím. Cho F1 tự thụ phấn, thì kết quả ở F2 sẽ thế nào về kiểu gen và kiểu hình? Giải thích kiểu tác động của gen đối với kết quả của phép lai.
Hướng dẫn
a) Từ kết quả thu được của hai phép lai, tỉ lệ phân li màu hoa ở F2 có thể có hai trường hợp là 13 hoa trắng : 3 hoa đỏ hoặc 3 hoa trắng : 1 hoa đỏ. Nhưng tỉ lệ 3 hoa trắng : 1 hoa đỏ là không phù hợp về mặt di truyền học và các dữ kiện của ý c.
Kiểm định tỉ lệ 13 : 3
- Phép lai I
Hoa trắng Hoa đỏ Tổng số
Số liệu thực tế : 124 36 160
Số liệu lí thuyết : 130 30 160 Độ lệch d : -6 +6
2 36 36
Phép lai II
liệu thực tế : 122 38 160
Số liệu lí thuyết : 130 30 160 Độ lệch d : -8 +8 d
b) - Vì kết quả cả 2 phép lai giống nhau, trong đó dòng 2 và dòng 3 có kiểu gen khác nhau, do đó phải liên quan tới 3 cặp gen.
- P1: AAbbdd (Dòng 1: hoa trắng) x aaBBdd (Dòng 2: hoa đỏ) →F1: AaBbdd (hoa trắng)
F2: (9A-B-dd : 3A-bbdd : 1aabbdd): 13 hoa trắng
3aaB-dd: 3 hoa đỏ
B.
Kiểu tác động của gen: B: hoa đỏ; b: hoa trắng; A át B: hoa trắng; aa không át
- P2: AAbbdd (Dòng 1: hoa trắng) x aabbDD (Dòng 3: hoa đỏ) →F1: AaBbdd (hoa trắng)
F2: (9A-B-dd : 3A-bbdd : 1aabbdd): 13 hoa trắng
3aaB-dd: 3 hoa đỏ
Kiểu tác động của gen: D: hoa đỏ; d: hoa trắng; A át D: hoa trắng; aa không át D.
c) - P3: aabbDD (Dòng 3: hoa đỏ) x aaBBdd (Dòng 2: hoa đỏ) →F1: aaBbDd (hoa tím)
F2: 9aaB-D-: 9 hoa tím
(3aaBbdd : 3aabbD-): 6 hoa đỏ
1aabbdd: 1 hoa trắng
Kiểu tác động của gen: B: hoa đỏ; b: hoa trắng; D: hoa đỏ; d: hoa trắng; B-
D-: hoa tím.
Bài tập 11 (Trích đề thi chọn HSG quốc gia năm học 2012)
Cho rằng ở 1 loài động vật. Lông chỉ có 2 dạng là lông dài và lông ngắn, trong đó KG quy định lông dài là AA, KG quy định lông ngắn là aa. Con đực thuần chủng lông dài cho giao phối với con cái thuần chủng lông ngắn được F1. Cho F1 giao phối ngẫu nhiên với nhau được F2 có số con lông dài chiếm tỉ lệ ¾ ở giới đực và ¼ ở giới cái
a. Giải thích kết quả phép lai
b, Viết sơ đồ lai
Giải: tỉ lệ phân bố không đều ở 2 giới-> Có liên kết giới tính hoặc ảnh hưởng của giới tính đối với sự hình thành tính trạng
- F2 có tỉ lệ phân bố KH 3 lông dài: 1 lông ngắn ở giới đực và ngược lại, nghĩa là sự phân bố các KH không đều ngay ở trong cùng 1 giới tính. Điều này không thể hiện đối với sự DTLK giới tính mà chỉ có với tính trạng ảnh hưởng của giới tính
- Ở F2 giới đực có tỉ lệ 3 lông dài: 1 lông ngắn chứng tỏ thể dị hợp biểu hiện lông dài->-. Thể dị hợp ở giới cái biểu hiện lông ngắn
SĐL Pt/ c ♂ Lông dài (AA) x ♀ lông ngắn aa F1 Aa (♀ lông ngắn, ♂ lông dài)
F1 x F1 ♀ lông ngắn Aa x ♂ lông dài Aa
Giới đực 3 lông dài : 1 lông ngắn
Giới cái 1 lông dài : 3 lông ngắn
Bài tập 12 (Trích đề thi chọn HSG quốc gia năm học 2013-2014)
Ở một loài thực vật, khi cho dòng hoa kép làm mẹ giao phấn với dòng hoa đơn, thu được F1 100% hoa kép. Cho F1 tự thụ phấn, thu được F2 100% hoa kép. Bằng cách nào xác định được tính quy luật sự di truyền của dạng hoa?
Hướng dẫn
- F1 và F2 toàn hoa kép chưa xác định chính xác được tính quy luật của sự di truyền dạng hoa vì kết quả này có thể di truyền qua tế bào chất (di truyền theo dòng mẹ) hoặc di truyền do hiệu ứng dòng mẹ. Vì vậy, muốn xác định được tính quy luật sự di truyền dạng hoa cần phải lai nghịch hay tiếp tục cho F2 tự thụ phấn cho ra F3.
- Cách 1: Tiến hành phép lai nghịch
+ Nếu F1 và F2 toàn hoa đơn giống cây mẹ thì dạng hoa di truyền theo dòng mẹ, nghĩa là di truyền qua tế bào chất.
+ Nếu F1 toàn hoa đơn giống cây mẹ và F2 toàn hoa kép thì dạng hoa di truyền do hiệu ứng dòng mẹ.
- Cách 2: Cho cây F2 tự thụ phấn
+ Nếu F3 toàn hoa đơn giống cây mẹ thì dạng hoa di truyền theo dòng mẹ, nghĩa là di truyền qua tế bào chất.
+ Nếu F3 có tỉ lệ 3 hoa kép : 1 hoa đơn thì dạng hoa di truyền do hiệu ứng dòng mẹ.
Bài tập 13 Tương tác bổ sung của 3 gen
Ở 1 loài thực vật lưỡng bội, tính trạng màu sắc hoa do 3 cặp gen ( A, a; B, b; D, d) phân li độc lập cùng quy định. Kiểu gen có cả 3 alen trội A, B và D cho hoa màu đỏ, kiểu gen có 2 alen trội A và B nhưng không có alen trội D thì cho hoa vàng, các kiểu gen còn lại thì cho hoa trắng. Theo lí thuyết, trong các dự đoán sau có bao nhiêu dự đoán đúng với các dữ liệu đã cho ?
(1). Ở loài này có tối đa 15 kiểu gen quy định hoa trắng.
(2). P: AaBbDd x AabbDd, thu được F1 có số cây hoa trắng chiếm tỉ lệ 62,5%.
(3). P: AABBdd x AabbDD thu được F1, cho F1 giao phấn được F2 có kiểu hình phân li theo tỉ lệ: 12 cây hoa đỏ: 3 cây hoa vàng : 1 cây hoa trắng.
(4). P: AABBDD x aabbDD, thu được F1. F1 tự thụ phấn thu được F2 có kiểu hình phân li theo tỉ lệ: 9 cây hoa đỏ: 7 cây hoa trắng.
A. 4. B. 1. C. 3. D. 7.1.
A+B+D -> đỏ
A+B : Vàng
Số kiểu gen quy định hoa vàng : A-B-dd= 7.1.2=4
=> Tổng số kiểu gen quy định hoa trắng = 33- 8- 4=15 => (1) Đúng
(2) P: AaBbDd x AabbDd
=> F1 Tỉ lệ kiểu gen hoa đỏ : A-B-D- = 3139 42432 xx =
T
ỉ lệ KG hoa vàng: A-B-dd = 3113 42432 xx =
=> Tỉ lệ kiểu gen hoa trắng = 935 1 32328 −−=
(2) => Đúng
(3) P : AABBdd x AabbDD
=> F1 : 1 2 AABbDd ; 1 2 AaBbDD => (3) Sai 935 1 32328 −−=
(4) P: AABBDD x aabbDD => F1 AaBbDD
F1x F1 : AaBbDD x AaBbDD
F2 : -> A-B-D- = 339 4416 x =
=> Tỉ lệ phân ly kiểu hình ở F2 9 đỏ : 7 trắng => Đúng
Đáp án [C]
Bài tập 14 Tương tác át chế của 3 gen
Ở 1 loài động vật, khi cho lai 2 dòng thuần chủng lông màu trắng và lông màu vàng giao phối với nhau được F1 toàn lông màu trắng cho F1 giao phối với nhau được F2 có kiểu hình: 48 con lông màu trắng: 9 con lông đen: 3 con lông xám: 3 con lông nâu; 1 con lông vàng. Hãy giải thích
Giải :
- Kết quả cho thấy màu lông bị chi phối bởi sự tương tác của 3 gen không alen trên NST thường và F1dị hợp về 3 cặp gen
Sơ đồ phân ly ở F2 3/ 4D- = 27 A- B- D- = 27 con lông trắng
3/ 4 B- ¼ dd = 9 A- B- dd = 9 con lông trắng
F2
3/4 A----- 1/ 4 bb
3/ 4D- = 9 A- bb- D- = 9 con lông trắng
(1) Kiểu gen quy định hoa đỏ :A-B-D- : 7.1.2.2=8 402
¼ dd = 3 A- bb- dd = 3 con lông trắng
3/ 4D- = 9 aa- B- D- = 9 con lông đen
3/ 4 B- ¼ dd = 3 aa- B- dd = 3 con lông nâu
1/4 aa----- 1/ 4 bb
3/ 4D- = 9 aa- bb- D- = 3 con lông xám
¼ dd = 1aa- bb- dd = 1 con lông vàng
Nhân xét: alen B: quy định lông nâu, b: lông vàng, alen D; lông xám, d: lông vàng. Các alen trội B và D tác động bổ trợ quy định lông đen: alen A át chế sự hình thành sắc tố màu trắng
Bài tập 15: Tương tác gen có hiện tượng gây chết và độ thâm nhập, độ biểu hiện (Trích đề thi chọn HSG quốc gia năm học 2015-2016)
Một cặp vợ chồng cả hai đều mắc chứng “điếc” do mang một số alen lặn ở 3 gen liên quan đến thính giác: d1 là lặn so với D1, d2 là lặn so với D2, d3 là lặn so với D3 Đồng hợp tử đột biến ở bất cứ 1 trong 3 gen này đều gây “điếc”. Ngoài ra, đồng hợp tử lặn đồng thời ở 2 trong 3 gen này gây chết ở giai đoạn phôi (sảy thai sớm) với độ thâm nhập (độ biểu hiện) là 25%. Đồng hợp tử lặn ở cả 3 gen gây sảy thai sớm với độ thâm nhập là 75%. Với kiểu gen của mẹ là D1d1D2d2d3d3 và của bố là d1d1D2d2D3d3 thì xác suất con của họ được sinh ra (không tính sảy thai) có thính giác bình thường là bao nhiêu? Giải thích.
Giải
Tính tỷ lệ cho từng cặp gen, rồi sử dụng nguyên tắc nhân xác suất (áp dụng cho 3 cặp gen) từ phép lai D1d1D2d2d3d3 x d1d1D2d2D3d thu được tỷ lệ:
3/16 D1- D2-D3- (bình thường) : 3/16 D1- D2-d3d3 (điếc do 1 gen) : 3/16 d1d1-
D2-D3- (điếc do 1 gen) : 1/16 D1- d2d2D3- (điếc do 1 gen) : 1/16 D1- d2d2d3d3 (điếc do 2 gen) : 3/16 d1d1D2-d3d3 (điếc do 2 gen) : 1/16 d1d1d2d2D3- (điếc do 2 gen) : 1/16 d1d1d2d2d3d3 (điếc do 3 gen)
Như vậy, tỉ lệ kiểu hình (chưa tính độ thâm nhập của gen) là 3/16 bình thường :
7/16 điếc do 1 gen : 5/16 điếc do 2 gen : 1/16 điếc do 3 gen
Áp dụng độ thâm nhập của gen do tính trạng “gây chết” với độ thâm nhập không hoàn toàn (dưới 100%) với trường hợp chết do 2 hoặc 3 gen:
- Với 2 gen: 1/4 chết; 3/4 sống và điếc, như vậy ta có 5
/16 điếc do 2 gen x 3/4 sống = 15/64 sống và điếc do 2 gen
- Với 3 gen: 3/4 chết; 1/4 sống và điếc, như vậy ta có 1/16 điếc do 3 gen x 1/4 sống = 1/64 sống và điếc do 3 gen Tổng cộng ta có:
3/16 bình thường : 7/16 điếc do 1 gen : 15/64 điếc do 2 gen : 1/64 điếc do 3 gen = 12/64 bình thường : 28/64 điếc do 1 gen : 15/64 điếc do 2 gen : 1/64 điếc do 3 gen = 56/64 trẻ sống và 8/64 trẻ chết trong giai đoạn phôi (sảy thai).
Như vậy, trong số trẻ sống (56/64), xác suất trẻ bình thường (không bị điếc) là : 12/56 x 100% = 21,4%
Bài tập 16: Tương tác gen và liên kết gen (Trích đề thi chọn HSG quốc gia năm học 2018-2019)
Để tìm hiểu quy luật di truyền chi phối hai tính trạng hình dạng và màu sắc quả bí, một nhà khoa học đã tiến hành lai giữa hai dòng bí thuần chủng quả dẹt, màu xanh với quả dài, màu vàng thu được F1 toàn quả dẹt, màu xanh. Cho F1 tự thụ phấn thu được F2 gồm các kiểu hình có tỷ lệ:
9/16 cây cho quả dẹt, màu xanh
3/16 cây cho quả tròn, màu xanh
2/16 cây cho quả tròn, màu vàng
1/16 cây cho quả tròn, màu trắng
1/16 cây cho quả dài, màu vàng
Biết rằng vị trí các gen trên NST không thay đổi trong quá trình giảm phân.
a) Hãy xác định quy luật di truyền chi phối mỗi tính trạng nêu trên. Viết sơ đồ lai từ P đến F2 về sự di truyền hai tính trạng trên.
b) Xác định tỷ lệ kiểu gen, kiểu hình ở đời con khi cho lai cây F1 với cây có kiểu hình quả dài, màu trắng.
Giải
* Xét riêng sự di truyền của từng cặp tính trạng + Tính trạng hình dạng quả:
Tỷ lệ phân ly ở F2: dẹt : tròn : dài = 9 : 6 : 1 = 16 tổ hợp lai = 4x4.
F1 dị hợp 2 cặp gen không alen cho 4 loại giao tử quy định 1 tính trạng hình dạng quả với tỷ lệ phân ly 9 : 6 : 1 => theo quy luật tương tác gen kiểu bổ trợ. Quy ước: A-B-: quả dẹt; A-bb, và aaB-: quả tròn; aabb: quả dài P: AABB (quả dẹt) x aabb (quả dài)
F1: AaBb (quả dẹt) + Tính trạng màu sắc quả
Tỷ lệ phân ly ở F2: xanh : vàng : trắng = 12 : 3 : 1 = 16 tổ hợp lai = 4×4.
F1 dị hợp 2 cặp gen không alen cho 4 loại giao tử quy định 1 tính trạng màu sắc quả với
tỷ lệ phân ly 12 : 3 : 1 => theo quy luật tương tác gen kiểu át chế do gen trội.
Quy ước: X: quy định màu xanh, gen át chế; x: không át chế; V: quy định màu vàng;
v: quy định màu trắng
404
P: XXvv (quả xanh) x xxVV (quả vàng)
F1: XxVv (quả xanh)

* Xét sự di truyền của cả hai tính trạng
Tỷ lệ phân ly kiểu hình ở F2: 9 : 3 : 2 : 1 : 1 = 16 tổ hợp lai = 4×4
F1 dị hợp về 4 cặp gen, cho 4 loại giao tử khác nhau => 4 cặp gen chi phối 2 tính


trạng tồn tại trên 2 NST liên kết với nhau hoàn toàn.
Sơ đồ lai:
Trường hợp 1: A liên kết với X, B liên kết với V
Tỷ lệ kiểu gen (9): 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1
Tỷ lệ kiểu hình: 9 dẹt, xanh : 3 tròn, xanh : 2 tròn, vàng : 1 dài, vàng : 1 tròn, trắng
Trường hợp 2: A liên kết với V, B liên kết với X
Kết quả tương tự như trường hợp 1
Cây có kiểu hình quả dài màu trắng có kiểu gen đồng hợp tử lặn ở cả 4 cặp gen
Trường hợp 1: A liên kết với X, B liên kết với V

Tỷ lệ phân ly kiểu gen kiểu hình là 1 : 1 : 1 : 1
Trường hợp 2: A liên kết với V, B liên kết với X
Kết quả tương tự như trường hợp 1.
- Khi xét sự di truyền của 2 cặp tính trạng, nếu có 1 tính trạng thường do tương
tác gen, 1 tính trạng liên kết với giới tính, ta dựa vào tỉ lệ phân ly kiểu hình của mỗi tính trạng, suy ra kiểu gen tương ứng. Sau đó kết hợp lại ta có kiểu gen chung về các tính trạng.
Bài tập 17: Tương tác gen và di truyền liên kết giới tính có hoán vị gen (Trích đề thi chọn HSG quốc gia năm học 2016-2017)
Có hai dòng ruồi giấm thuần chủng (A và B) đều có kiểu hình đột biến - mắt xù xì. Lai ruồi cái của dòng A với ruồi đực của dòng B, thu được F1 gồm 100% ruồi cái mắt kiểu dại và 100% ruồi đực mắt xù xì. Cho F1 giao phối ngẫu nhiên thu được F2 gồm 256 ruồi cái mắt kiểu dại, 250 ruồi cái mắt xù xì, 64 ruồi đực mắt kiểu dại và 436 ruồi đực mắt xù xì. Hãy giải thích kết quả và viết sơ đồ lai.
Giải
P thuần chủng về đột biến mắt xù xì → F1: 100% ruồi cái kiểu dại → hai đột biến thuộc về hai gen khác nhau (tương tác bổ sung trong sự quy định kiểu hình mắt hay 2 gen không alen với nhau) → F1 dị hợp tử về 2 cặp gen; đột biến là lặn, kiểu dại là trội.
Qui ước 2 cặp gen tương ứng là A/a và B/b Kiểu hình ở F1 không đồng đều ở 2 giới: 100% ruồi cái mắt kiểu dại; 100% đực mắt xù xì → gen quy định màu mắt nằm trên nhiễm sắc thể giới tính. Từ số lượng cá thể ở đời lai F2 → Tỷ lệ kiểu hình ở F2 là:
+ Con cái: mắt kiểu dại : mắt xù xì = 1:1;
+ Con đực: mắt kiểu dại = 12,8%; mắt xù xì = 87,2%
Nếu 2 gen phân ly độc lập, F1 x F1 không thể cho tỷ lệ phân ly ở F2 như đầu bài đã nêu → 2 gen liên kết không hoàn toàn trên nhiễm sắc thể X; xảy ra hoán vị gen trong giảm phân tạo giao tử ở ruồi cái. P ruồi cái dòng đột biến A (XaBXaB) x ruồi đực dòng đột biến B (XAbY) → F1: ruồi cái (XX) 100% mắt kiểu dại → ruồi cái F1 có kiểu gen dị hợp tử chéo: XaBXAb .
F2: 12,8% số ruồi đực có mắt kiểu dại XABY được tạo thành từ giao tử hoán vị gen XAB của ruồi cái → Tần số hoán vị gen = 12,8 x 2 = 25,6%
F
Con cái: 50% mắt kiểu dại : 50% mắt xù xì
Con đực: 12,8% mắt kiểu dại : 87,2% mắt xù xì
1. KẾT LUẬN
Trên đây chúng tôi đã xây dựng hệ thống cơ sở lý thuyết xây dựng hệ thống cơ sở lý thuyết Một số vấn đề tích hợp hóa sinh, vi sinh và di truyền, đồng thời sưu tầm một số câu hỏi, bài tập chọn lọc nhằm mục tiêu vô cùng thiết thực là giúp cho học sinh thi chuyên Sinh học học tập và nghiên cứu về phần này tốt hơn và đồng thời cũng giúp chính bản thân mình giảng dạy thuận lợi hơn. Phần lí thuyết và các câu hỏi vận dụng mà thực tế chúng tôi đã dạy trên lớp và dùng để bồi dưỡng đội tuyển học sinh giỏi tuỳ từng đối tượng học sinh mà có cách nâng cao khác nhau qua đó đã giúp học sinh nắm được kiến thức, đồng thời các em hiểu sâu hơn về mặt lí thuyết và thấy được ý nghĩa về mặt thực tế Đặc biệt các em giải nhanh dạng trắc nghiệm rất tốt, kỹ năng làm bài thi cũng được nâng lên.
Tuy nhiên đề tài này chắc chắn còn nhiều thiếu sót, chúng tôi rất mong nhận được sự đóng góp của quý thầy cô, đồng nghiệp và bạn bè để đề tài ngày càng hoàn thiện hơn.
1. ĐỀ NGHỊ
- Nội dung đề tài dành cho học sinh ôn tập kiểm tra định kì ở lớp, thi tốt nghiệp, cao đẳng, đại học và bồi dưỡng đội tuyển học sinh giỏi cấp tỉnh và học sinh giỏi quốc gia, bồi dưỡng giáo viên trong các buổi sinh hoạt chuyên môn.
- Đề nghị tổ và trường tạo điều kiện để sang năm tiếp tục phát triển đề tài trên qui mô rộng hơn.
2. Trần Ngọc Danh, Phạm Phương Bình- Tài liệu chuyên Sinh học Trung học phổ thông- Bài tập Di truyền và tiến hoá - Nxb Giáo dục, 2012.
3. Lê Thị Kim Dung – Nguyễn Thị Phương Thảo - Các dạng câu hỏi và bài tập trắc nghiệm Sinh học – NXB Đại học QGHN – 2008
4. Nguyễn Thành Đạt (Tổng chủ biên)- Sách giáo khoa Sinh học 12- NXB Giáo dục, 2008
5. Trần Dũng Hà – Nguyễn Hải Tiến - Phương pháp giải các bài tập sinh học – NXB Đại học QGHN, 2008.
6. Phạm Thành Hổ- Di truyền học- Nxb Giáo dục, 2003.
7. Ngô Văn Hưng, Đỗ Mạnh Hùng, Trần Minh Hương – Giới thiệu đê thi Olympic Quốc tế mônSinh học- Nxb Giáo dục, 2004.
8. Phạm Văn Lập, Trần Ngọc Danh, Đinh Đoàn Long, Tài liệu chuyên Sinh học Trung học phổthông- Di truyền và tiến hoá- Nxb Giáo dục, 2012.
9. Đinh Đòan Long, Đỗ Lê Thăng- Cơ sở di truyền học phân tử và tế bào- Nxb Đại học Quốc Gia Hà Nội, 2013.
10. Vũ Đức Lưu – Sinh học 12 chuyên sâu tập 1- Nxb Đại học Quốc gia Hà Nội, 2014.
11. Vũ Đức Lưu, Nguyễn Thị Hà- Bồi dưỡng học sinh giỏi sinh học Trung học phổ phông - Di truyền và tiến hoá- Nxb Giáo dục, 2013.
12. Vũ Đức Lưu, Nguyễn Thị Hà- Bồi dưỡng học sinh giỏi sinh học Trung học phổ phông, Bài tập di truyền và tiến hoá- Nxb Giáo dục, 2013.
13. Vũ Đức Lưu - Câu hỏi và bài tập trắc nghiệm sinh học 12- Nxb Hà Nội, 2008.
14. Phan Cự Nhân, Nguyễn Minh Công, Đặng Hữu Lanh- Di truyền học- Tập I – Nxb Đại họcSư Phạm, 2004.
15. Phan Cự Nhân, Trần Bá Hoành, Lê Quang Long, Phạm Đình Thái, Hoàng Thị Sản, Mai Đình Yên- Sinh học đại cương – Tập II- Nxb đại học Quốc Gia Hà Nội, 1997.
16. Lê Quang Nghị - Hướng dẫn giải nhanh các dạng bài tập trắc nghiệm và tự luận sinh học -Nxb Đại học Sư Phạm, 2010
17. Phan Khắc Nghệ - Phương pháp giải nhanh các dạng bài tập sinh học –Nxb Đại học Quốc gia Hà Nội, 2012.
TÀI LIỆU THAM KHẢO
t Nam dịch và xuất bản năm2011 408
18. Đỗ Lê Thăng, Đinh Đoàn Long- Chú giải di truyền học -Nxb Giáo dục, 2010
19. Lê Đức Trình- Sinh học phân tử của tế bào- Nxb khoa học kỹ thuật, 2001.
20. Các đề thi học sinh giỏi quốc tế; học sinh giỏi quốc gia; các đề đề xuất học sinh giỏi Duyên hải bắc bộ (2010 -2019), Trại hè Hùng Vương (2009 – 2019) .
409
Chuyên đề
TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN
Trường THPT Chuyên Hoàng Văn Thụ - Hòa Bình
Chuyên đề đạt giải Ba
I. Lí do chọn đề tài
PHẦN I: MỞ ĐẦU
Trong chương trình sinh học cấp trung học phổ thông, phần kiến thức tích hợp hóa sinh, vi sinh và di truyền là một nội dung quan trọng, được đưa nhiều vào đề thi chọn học sinh giỏi quốc gia, quốc tế. Nhiều giáo viên và học sinh còn lúng túng với phần kiến thức này trong khi ôn luyện đội tuyển HSG. Mặc dù hiện nay có rất nhiều tài liệu viết về hóa sinh, vi sinh và di truyền nhưng rất ít tài liệu về tích hợp cả ba nội dung này với nhau và các tài liệu khác nhau với mục đích khác nhau, phần kiến thức này được trình bày theo nhiều cách khác nhau. Vì vậy, chúng tôi biên soạn chuyên đề: “Tích hợp hóa sinh, vi sinh và di truyền” để giúp học sinh có cái nhìn tổng quát hơn, thống nhất hơn. Đồng thời hỗ trợ cho giáo viên kiểm tra kiến thức học sinh thông qua hệ thống các câu hỏi. Đây cũng là nguồn tài liệu ôn tập trong các kì thi trung học phổ thông, học sinh giỏi các cấp, kì thi chọn học sinh giỏi quốc gia và quốc tế.
II. Mục tiêu của chuyên đề
- Hệ thống hóa kiến thức về hóa sinh và di truyền vi sinh vật.
- Giới thiệu một số câu hỏi, bài tập tích hợp phần hóa sinh, di truyền vi sinh vật.
III. Đối tượng áp dụng
- Các đội tuyển ôn thi học sinh giỏi môn Sinh học các cấp.
- Các giáo viên Sinh học.
410
A. LÝ THUYẾT
I. Tích hợp hóa sinh trong cơ sở vật chất di truyền vi sinh vật
1. Axit nucleic

Thực tế thì Axit deoxiribonucleic (ADN) đã được khám phá vào năm 1869 ở trong nhân tế bào bạch cầu của người, nhưng lúc đó người ta vẫn chưa biết rõ được chức năng của nó. Sau đó hợp chất này được tách ra từ nhiều kiểu nhân, nên gọi nó là axit nuclêic (axit nhân).
Năm 1910, bằng phân tích hoá học người ta đã phân biệt được 2 loại axit nuclêic là axit deoxiribonucleic (ADN) và axit ribonucleic (ARN).
Năm 1924, những nghiên cứu hiển vi có sử dụng thuốc nhuộm ADN và prôtêin đã cho thấy cả 2 loại hợp chất này đều có trong nhiễm sắc thể. Một loạt các nghiên cứu khác cũng đã gợi ra mối quan hệ gắn bó giữa ADN và vật chất di truyền. Ví dụ hầu hết các tế bào xôma của một loài có 1 lượng ADN không đổi; trong khi đó hàm lượng của ARN và prôtêin lại thay đổi khá nhiều ở các giai đoạn khác nhau của một tế bào và các tế bào của cùng 1 loài. Nhân của các tế bào giao tử thì lại có hàm lượng ADN bằng một nửa nhân của tế bào xôma cùng loài.
Những dẫn liệu lý thuyết lúc bấy giờ đã đưa ra nhiều tranh cãi về ADN hay prôtêin là vật liệu di truyền bởi lẽ những phân tích hoá học cho thấy ADN kém tính đa dạng hơn so với prôtêin.
Năm 1928, F.Griffith đã bằng thực nghiệm chứng minh được ADN là vật chất di truyền. Thí nghiệm được sơ đồ hóa qui trình như sau:
Hình 1. Thí nghiệm của F.Griffith
Ở vi khuẩn Pneumococus có 2 chủng chủng R và chủng S. Chủng R có khuẩn lạc dạng nhăn nheo, không gây bệnh viêm phổi và chủng S có khuẩn lạc dạng nhẵn bóng gây bẹnh viêm phổi , hai chủng này không phát triển chung với nhau. Khi chuột được tiêm hoặc nhóm tế bào R sống hoặc với nhóm tế bào S chết thì chuột vẫn khoẻ mạnh. Trong trường hợp con chuột bị tiêm một lượng nhỏ chủng R sống và 1 lượng lớn chủng S chết thì chuột lại bị chết vì bị viêm phổi. Phân lập từ các mẫu máu của chuột chết thấy có chủng S sống. Hiện tượng này cho thấy vi khuẩn S không thế tự sống lại được sau khi đã bị đun chết, nhưng các tế bào này đã truyền tính gây bệnh cho tế bào R. Hiện tượng này gọi là biến nạp (transformation).
Năm 1944, T.Avery; Mc Leod và Mc. Carty đã tiến hành thí nghiệm để xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng prôteaza hoặc ARNaze thì hoạt tính biến nạp vẫn còn, chứng tỏ prôtêin và ARN không phải là tác nhân biến nạp. Nhưng nếu tế bào S chết bị xử lý bởi ADNaze thì hoạt tính biến nạp không còn nữa.Chứng tỏ ADN là nhân tố biến nạp. Như vậy hiện tượng biến nạp là một bằng chứng sinh hoá xác nhận rằng ADN mang tín hiệu di truyền. Tuy nhiên tại thời điểm đó vai trò của ADN vẫn chưa được công nhận vì trong thí nghiệm vẫn còn một chút prôtêin.
Năm 1952, A.Hershey và M.Chase đã tiến hành thí nghiệm với bacteriophage T2 đánh dấu S35 và P32 đối với E.coli mới chứng minh được chính xác ADN là vật chất chứa đựng thông tin di truyền và truyền đạt thông tin di truyền.
Hình 2. Thí nghiệm của A.Hershey và M.Chase (1952)
2. Cấu tạo axit nucleic

2.1. Cấu tạo hoá học
2.1.1. Nucleotit – đơn phân cấu tạo axit nucleic
a. Thành phần cấu tạo nucleotit
Cả ADN và ARN đều là các chất trùng phân (polymer) mạch dài gồm nhiều đơn phân (monomer) nối với nhau. Trong đó, mỗi đơn phân của ADN là deoxyribonucleotit, còn của ARN là ribonucleotit.
Mỗi một nucleotit gồm ba thành phần cơ bản:
+ Bazơnitơ: là các dẫn xuất hoặc của purine, gồm adenine (A) và guanine (G); hoặc của pyrimidine, gồm thymine (T), cytosine (C), và uracil (U);

+ Đường pentozơ: Tham gia cấu tạo nucleotit có 2 loại: đường ribose và deoxyribose
+ Axit phot phoric (H3PO4): trong cấu tạo phân tử axit này có 3 gốc axit vì vậy nó có thể kết hợp được với 3 nhóm định chức khác có tính bazơ. Trong cấu trúc của axit nucleic, axit photphoric có 2 nhóm axit liên kết với 2 nhóm OH của 2 phân tử đường trước và sau nó trên mạch polynucleotit và còn lại 1 gốc axit vì vậy chuỗi poly nucleotit mang tính axit
Vị trí các nguyên tử cacbon (C) trên mạch vòng của đường pentose được đánh số từ C-1’ đến C-5’. Các nucleotit mang nhóm phosphate ở vị trí C-5’ có vai trò quan trọng đối với cấu trúc và chức năng của ADN và ARN. Sự có mặt của nhóm phosphate ở vị trí này làm các phân tử ADN và ARN thường mang điện tích âm và có tính axit. Trong điều kiện in vivo, nếu các điện tích của axit nucleic không được trung hòa thì nó không thể đóng gói vào nhiễm sắc thể và nhân tế bào được. Sự trung hòa điện tích diễn ra nhờ sự có mặt của các protein có tính kiềm (còn gọi là tính kiềm) xuất hiện trong tế bào ở cả các sinh vật prokaryote (nhân sơ) và eukaryote (nhân thật). Ở eukaryote, đó là các phân tử protein histon mang tính kiềm, còn ở prokaryote là các polyamin.
b. Sự liên kết giữa các thành phần tham gia cấu tạo 1 nucleotit.
Các bazơ nitơ gắn với đường pentose bằng liên kết cộng hóa trị ở vị trí C-1’ của đường với nitơ ở vị trí số 9 của purine hoặc nitơ ở vị trí số 1 của pyrimidine
Bazơ nitơric + Đường pentozơ
Nucleozit +Axít phốt phoríc
Nucleotit
Hình3.Sơ đồ cấutạo1nucleotit
c. Đồng đẳng của nucleotit
Trong cơ thể thường gặp các nucleotit liên kết với enzim này hay enzim khác vốn đóng vai trò đặc biệt quan trọng. Cấu tạo của các nucleotit deoxiriboza này thường đặc trưng bởi sự hình thành liên kết với 1 phân tử axit phôt phoric ở vị trí C5' của đường pentose, ngoài ra trong tế bào còn có các loại nucleotit liên kết với 2 hoặc 3 gốc photphat. Đại diện quan trọng nhất của các nhóm này là adenosine - 5' - monophotphat (AMP), adenosine -5'- diphotphat (ADP) và adenosine - 5' - triphotphat (ATP). ATP là chất mang năng lượng hoá học hoạt hoá chủ yếu ở trong tế bào.
Loạ i axit nucleic
Purin Pyrimidin Ad enin(A) Gu anin(G) Cyt ozin(C) Ty min (T) Ura xin (U)

AD N A G C T
AR N A G C U
Chú ý:
+ Người ta gọi các đơn phân cấu tạo nên axit nucleic là nucleotit
+ Đơn phân tham gia cấu tạo nên ADN là các nucleotit deoxiriboza
+ Đơn phân tham gia cấu tạo nên ARN là các nucleotit riboza
2.1.2. Các liên kết hoá học chủ yếu trong cấu trúc phân tử axit nucleic
a. Liên kết liên kết cộng hoá trị
- Liên kết phốtpho este: Được hình thành giữa nhóm 3’OH ở phân tử đường với 1 nhóm OH của gốc axit photphoric, liên kết này tạo ra 1 phân tử nước.
- Liên kết N - glycozit: Được hình thành giữa nhóm 1’OH của phân tử đường với 1NH của bazo nitơ loại pirymidin hoặc 9NH của bazơ nitric loại purin, liên kết này tạo ra 1 phân tử nước.
b. Liên kết hidro
- Liên kết hidro là liên kết được hình thành giữa nguyên tử H đã liên kết với 1 nguyên tố hoá học khác với 1 nguyên tố có độ âm điện lớn (O, N) cũng đã liên kết với 1 nguyên tố hoá học khác.
+ Trong cấu trúc của ADN nó được hình thành giữa các bazơnitric trên 2 mạch đơn của ADN: A liên kết với T bởi 2 liên kết hidro, G = X bởi 3 liên kết hidro.
+ Trong cấu trúc phân tử ARN (rARN và tARN được hình thành giữa các nucleotit trên 1 mạch đơn : A=U, G =X.
- Vai trò: Duy trì cấu trúc không gian của axit nucleic.
Hình 4. Sơ đồ cấu trúc AMP, cAMP và ATP
d. Kích thước và khối lượng của 1 nucleotit
- Khối lượng của 1 nucleotit: 300 đvC ()
- Kích thước không gian trung bình của 1 nucleotit là 3,4A0 ()
e. Phân loại nucleotit
2.1.3. Sự liên kết giữa các đơn phân tạo thành chuỗi poly nucleotit
416
- Trên Các nucleotit liên kết với nhau bằng liên kết cộng hoá trị phốtpho dieste
- Chiều liên kết giữa các nucleotit 5’ => 3’,
- Trong cấu trúc phân tử ADN hai mạch đơn có chiều liên kết trái ngược nhau.
trình tự sắp xếp các base chứa nitơ ở trong mạch thứ hai sẽ phụ thuộc một cách nghiêm ngặt vào trình tự sắp xếp các base chứa nitơ trong mạch thứ nhất. Các cặp AdeninThymine và Guanin - Cytosine là những cặp bổ sung.
Hình 5. Sơ đồ cấu tạo 1 đoạn poly nucleotitdeoxyriboza


2.2. Cấu trúc không gian của Axit nucleic
2.2.1. Cấu trúc không gian của ADN
Trong phân tử ADN sợi kép (gọi như vậy vì mỗi phân tử gồm hai mạch đơn polynucleotit), hai mạch polynucleotit liên kết với nhau qua liên kết hydro giữa các nucleotit đối diện trên hai mạch. Liên kết hydro trong ADN hình thành giữa hai mạch thường là A = T và G ≡ C. Trong đó, số gạch nối giữa các nucleotit (A, T, G và C) phản ánh số liên kết hydro giữa chúng; theo đó, giữa A và T có hai liên kết hydro, còn giữa G và C có ba liên kết hydro. Nguyên tắc liên kết này còn gọi là nguyên tắc bổ sung, hay nguyên tắc Chargaff
Cấu trúc không gian của ADN (cấu trúc bậc 2) đầu tiên được J. Watson và
F.Crick đề sướng vào năm 1953 với nội dung cơ bản như sau:

ADN được cấu tạo từ 2 mạch polynucleotit deoxiriboza nằm song song ngược chiều nhau, xoắn lại với nhau theo chu kỳ. Trên mỗi mạch các nucleotit deoxiriboza liên kết lại với nhau nhờ các liên kết phôtphodieste ADN xoắn theo chu kỳ, mỗi chu kỳ xoắn có kích thước là 34A0 gồm có 10 cặp nucleotit. Góc nghiêng giữa các nucleic deoxiriboza cạnh nhau trong từng mạch là 360. Trên mỗi mạch ở mỗi các mononucleotit các base nằm phía trong, còn đường pentose và axit photphoric nằm phía ngoài. Hai mạch đơn polynucleotit deoxiriboza xoắn với nhau xung quanh một trục tưởng tượng và liên kết với nhau thông qua các base chứa nitơ dọc theo phân tử ADN nhờ các liên kết hidro. Các liên kết hidro hướng từ nhóm NH2 của adenin đến nhóm - OH của thymine và từ nhóm - NH2 của guamin đến nhóm - OH của Cytosine. Điều này có nghĩa là trình tự sắp xếp các base chứa nitơ ở một trong hai mạch có thể tuỳ ý, nhưng
Hình 6. J. Watson và F.Crick với mô hình ADN mà 2 ông xây dựng được Sau này người ta còn phát hiện ra nhiều loại ADN nữa đó là dạng A,C,D và Z, các dạng này có sự khác nhau về chiều quay của chuỗi xoắn kép, đường kính vòng xoắn, số cặp bazơ nitơ …(Hình 42).
Trong thực tế mô hình ADN dạng B của Watson và Crick là mô hình cấu trúc phổ biến hơn cả, hai mạch đơn của phân tử ADN sợi kép xoắn xung quanh nhau về phía phải. Cấu trúc xoắn đều đặn như vậy tạo ra hai loại khe trong mỗi vòng xoắn gọi là khe chính (major groove) và khe phụ (minor groove).
Do sự cuộn xoắn của chuỗi ADN sợi kép, khe chính rộng hơn so với khe phụ.
Các nghiên cứu sau này trên các đối tượng sinh vật khác nhau cho thấy các protein
điều hoà hoạt động của gen thường có xu hướng liên kết với trình tự ADN mà nó điều khiển tại vị trí các khe chính nhờ những trình tự axit amin đặc biệt có khả năng hình thành các liên kết hydro với các bazơ nitơ trên phân tử ADN tại khe chính. Mối tương tác này được tìm thấy tương đối phổ biến ở cả sinh vật nhân sơ và nhân thật. Trong khi đó, các khe phụ có xu hướng là vị trí gắn của các protein cấu trúc thường tham gia vào quá trình đóng gói các phân tử ADN ở sinh vật nhân thật. Những protein này, chẳng hạn như histon, chứa một lượng lớn các axit amin mang tính kiềm (tích điện dương) như Arg và Lys, chúng có khuynh hướng tương tác với nhóm phosphate mang điện âm trên phân tử ADN. Vị trí của các cặp bazơ nitơ trong phân tử ADN không nằm dọc trục đường – phosphate giúp giải thích cho đặc tính hấp thụ ánh sáng cực đại của phân
trong môi trường có hàm lượng muối cao hoặc có hiện tượng methyl hóa cytosine, phân tử ADN có thể có cấu hình hiếm gặp hơn là dạng Z (Z-ADN còn gọi là zigzag ADN; hay dạng S-ADN).
Hệ gen của sinh vật nhân sơ nói chung cũng như nhiễm sắc thể của vi khuẩn nói riêng đều nằm trong một vùng tương đối nhỏ. Giống như sinh vật nhân thực, cấu trúc này hình thành nhờ các protein bám ADN đóng gói hệ gen theo một mô hình có tổ chức.
2.2.2 Cấu trúc không gian của ARN
a. mARN
Phân tử mARN có cấu trúc một mạch đơn là bản phiên mã trình tự của gen (thực tế có trình tự giống với mạch mang nghĩa trên phân tử ADN mã hóa cho chuỗi polypeptide tương ứng). Loại ARN này chiếm khoảng 2 - 5 % tổng lượng ARN của tế bào.
Ở sinh vật nhân sơ, phần lớn mARN là bản sao nguyên vẹn trình tự nucleotide được hình thành sau quá trình phiên mã từ phân tử ADN, đồng thời được sử dụng ngay làm khuôn để dịch mã tổng hợp protein.

Ở sinh vật nhân thật, hầu hết các phân tử mARN được hình thành sau phiên mã còn phải trải qua một giai đoạn hoàn thiện (còn gọi là sự chế biến ARN) trong nhân tế bào. Quá trình hoàn thiện mARN gồm 3 sự kiện cơ bản:
+ Lắp “mũ” 7mG (7-methylguanidine triphosphate) vào đầu 5’ tận cùng của phân tử mARN tiền thân (tiền-mARN);
+ Cắt bỏ các đoạn trình tự không mã hoá (intron) và ghép nối các đoạn trình tự mã hoá (exon) với nhau;
+ Lắp ghép vào phần đuôi phân tử mARN bằng trình tự polyA (trình tự chứa từ vài chục đến hàng trăm adenine liên tiếp).
tử ADN ở bước sóng 260 nm (chính chính xác là 257 nm).
Tùy thuộc vào một số yếu tố môi trường xung quanh, như độ pH, trạng thái methyl hóa của các nucleotit, độ ẩm, hàm lượng muối, tính ưa nước, lực ion hoá, sự có mặt của các protein liên kết ADN… mà phân tử ADN sợi kép có thể xuất hiện ở một số dạng cấu hình không gian khác nhau. Trong đó, dạng cấu hình phổ biến nhất là dạng B (viết tắt là B-ADN), đây chính là mô hình được Watson và Crick mô tả. Trong tế bào sống, phần lớn phân tử ADN sợi kép ở dạng cấu hình này, nhưng khi độ ẩm giảm đi (≈75%) thì có thể có sự hình thành của cấu hình dạng A (A-ADN); hoặc nếu
Sản phẩm hình thành sau quá trình hoàn thiện này là phân tử mARN hoàn chỉnh, sẵn sàng cho quá trình dịch mã (tổng hợp protein) diễn ra ở tế bào chất. Kích thước của sợi mARN thường dài từ 900 đến 1200 ribonucleotide, khối lượng trung bình khoảng từ 3x105 đến 4x106 Dal với hệ số lắng dao động trong khoảng từ 6S đến 25S (Hình 7a).
Ở sinh vật nhân thật, phần lớn các phân tử mARN được tổng hợp từ trong nhân tế bào, chỉ trừ một số ít được tổng hợp từ các gen trong ti thể và lạp thể
b. tARN : Hàm lượng ARN vận chuyển (tARN) trong tế bào chiếm khoảng 10 - 15% tổng lượng ARN. Phân tử tARN thường là một mạch polynucleotide ngắn, chứa từ 75 đến 95 ribonucleotide, khối lượng phân tử khoảng 23 - 30 kDa, hệ số lắng là 4S.
Cấu trúc tARN điển hình gồm 1 mạch polynucleotide cuộn xoắn dạng lá phân thùy (Hình 7b), trong đó có một vài đoạn xoắn kép do các ribonuclotide liên kết bổ sung theo nguyên tắc Chargaff. Trong cấu trúc này, tARN có một thuỳ tiếp nhận axit amin có đầu tận cùng luôn là trình tự CCA. Thuỳ đối mã mang bộ ba đối mã phù hợp với mã bộ ba trên phân tử mARN làm mạch khuôn trong quá trình dịch mã. Ngoài ra còn có 1 hoặc 2 thuỳ phụ tùy vào từng loại tARN. Tuy đến nay vai trò của các thuỳ phụ chưa rõ, nhưng dường như chúng có vai trò làm tăng tính ổn định của các phân tử tARN.
Mỗi loại tARN có vai trò vận chuyển một loại axit amin duy nhất. Nhưng một axit amin có thể được vận chuyển bởi nhiều loại tARN khác nhau. Đến nay, đã có trên 300 loại phân tử tARN khác nhau được xác định giữ vai trò vận chuyển 20 loại axit amin cơ bản. Mặc dù các phân tử tARN có thể khác nhau đôi chút về số lượng các ribonucleotide, nhưng chúng có một số đặc điểm cấu hình chung. Theo đó, các phân tử tARN đều là các chuỗi polynucleotide mạch đơn, tự cuộn xoắn bởi các liên kết hydro hình thành giữa các nuleotide bên trong sợi kể từ đầu 5’, đồng thời có một số đặc điểm:
+ Có một nhóm monophosphate ở đầu 5’;
+ Một cấu trúc thùy DHU (còn gọi tắt là thùy DHU) mang bazơ nitơ hiếm gặp dihydrouridin (DHU);
+ Một cấu trúc thùy mang bộ ba đối mã, giúp “dịch” mã bộ ba trên mARN được gọi là “thùy đối mã”;
+ Một cấu trúc thùy bất định, thường có số nucleotide rất khác nhau (từ 3 đến 21 nucleotide) ở các loại phân tử tARN khác nhau;
+ Cấu trúc thùy TΨC mang bazơ nitơ hiếm gặp pseudouridin (kí hiệu Ψ, hoặc ΨU);
+ Phần thân của thùy nhận axit amin thường gồm 7 cặp bazơ nitơ liên kết bổ sung, trong đó có thể có một cặp liên kết bất thường giữa G và U;
+ Một trình tự gồm 3 nucleotide tận cùng đầu 3’ luôn là CCA tìm thấy ở tất cả các loại tARN, trong đó nhóm –OH của A tại đầu 3’ tận cùng chính là vị trí gắn axit amin nhờ hoạt động xúc tác của enzym aminoacyl-tARN synthetase. Enzym này giúp lắp ghép chính xác từng loại axit amin vào phân tử tARN tương ứng vận chuyển nó. Phức hợp tARN sau khi lắp ghép với axit amin được gọi là aminoacyl-tARN (viết tắt là AA-tARN).
c. rARN:
ARN ribosome (Hình 7c) chiếm khoảng 80% tổng lượng ARN trong tế bào. rARN thường có cấu trúc dạng mạch đơn polyribonuleotide với nhiều khúc cuộn, chứa từ dưới 100 đến 1500 ribonucleotide. Các rARN kết hợp với một số phân tử protein đặc biệt tạo thành các ribosome. Ribosome có cấu tạo gồm hai tiểu phần, gọi là tiểu
phần nhỏ và tiểu phần lớn. Trong quá trình dịch mã hai tiểu phần của ribôxôm kết hợp với nhau tạo thành phức hệ ribosome hoàn chỉnh có hoạt tính dịch mã. Khi kết thúc dịch mã, hai tiểu phần lại tách nhau ra và tồn tại riêng rẽ, lúc này chúng không có hoạt tính dịch mã. Ở sinh vật nhân thật, ribosome ở tế bào chất có hệ số lắng là 80S(). Trong đó, tiểu phần nhỏ có hệ số lắng là 40S được cấu tạo từ một phân tử rARN 18S và khoảng 35 phân tử protein cấu trúc; còn tiểu phần lớn có hệ số lắng là 60S được cấu tạo từ 3 phân tử rARN với hệ số lắng là 5S, 5,8S và 28S kết hợp với khoảng 50 phân tử protein cấu trúc. Như vậy, sinh vật nhân thật có 4 loại rARN. ở sinh vật nhân sơ, ribosome có hệ số lắng là 70S, cũng gồm hai tiểu phần. Tiểu phần nhỏ có hệ số lắng là 30S được cấu tạo từ một phân tử rARN 16S và 21 phân tử protein cấu trúc; tiểu phần lớn có hệ số lắng là 50S gồm hai phân tử rARN có hệ số lắng là 5S và 23S kết hợp với 31 phân tử protein cấu trúc. Như vậy, sinh vật nhân sơ có 3 loại rARN.
d) Các loại ARN khác
Trong tế bào sinh vật nhân thật, còn có một số ARN có kích thước nhỏ chỉ chứa khoảng 90 - 300 ribonucleotide được gọi là các snARN (small nuclear RNA).


Các phân tử snARN không tham gia vào quá trình phiên mã, mà thay vào đó, nó kết hợp với một số protein đặc thù để tạo nên phức hợp tham gia vào việc cắt các intron và nối các exon trong quá trình hoàn thiện phân tử mARN, phức hợp này được gọi là phức hệ cắt intron (spliceosome).
Ngoài snARN, một số loại ARN kích thước nhỏ có hoạt tính giống enzym gọi là ribozyme. Ribozyme thực chất là các phân tử ARN kích thước nhỏ (thường chỉ chứa khoảng 40 - 50 ribonucleotide), có trình tự đặc hiệu và có khả năng xúc tác việc cắt các phân tử ARN ở một số vị trí nhất định. Ngoài ra, trong tế bào còn có một số loại ARN khác như SRP ARN (signal recognition particle RNA, còn gọi là ARN nhận biết tín hiệu), snoARN (small nucleolar RNA, còn gọi là ARN hạch nhân), các ARN tham gia điều hòa hoạt động của gen (siARN và miARN).
Hình 7a: mARN Hình 7b: tARN Hình 7c. rARN (riboxom)
Hình 7. Mô hình cấu trúc các loại ARN
3. Tính chất đặc trưng của axit nucleic
3.1. ADN
* Số lượng, thành phần và trình tự nucleotit deoxy riboza trên mỗi mạch đơn.
* Tỷ lệ A+T/ G+X. (đặc trưng cho sự bền vững với nhiệt của ADN, hai phân tử ADN có chiều dài bằng nhau, nếu phân tử nào có tỷ số trên lớn thì phân từ đó có số liên kết hidro ít hơn và kém bền với nhiệt hơn)
* Sự phân bố gen và cách mã hoá thông tin di truyền trên phân tử ADN (lượng
TTDT)
3.2. ARN
* Số lượng, thành phần và trình tự ribônucleotit trong cấu trúc phân tử
* Cấu trúc không gian của nó
4. Chức năng sinh học của axit nucleic
4.1. Chức năng sinh học của ADN
ở phần lớn các loài sinh vật (chỉ trừ một số virut), ADN có chức năng là vật chất mang thông tin di truyền. Để đảm nhiệm chức năng này, ADN có bốn đặc tính cơ bản sau:
+ Có khả năng lưu giữ thông tin ở dạng bền vững cần cho việc cấu tạo, sinh sản và hoạt động của tế bào.
+ Có khả năng sao chép chính xác để thông tin di truyền có thể được truyền từ thế hệ này sang thế hệ kế tiếp thông qua quá trình phân bào hay quá trình sinh sản.
+ Thông tin chứa đựng trong vật chất di truyền phải được dùng để tạo ra các phân tử cần cho cấu tạo và hoạt động của tế bào.
+ Vật liệu di truyền có khả năng biến đổi, nhưng những thay đổi này (đột biến)
chỉ xảy ra ở tần số thấp.
Trong các tế bào sinh vật nhân thật, phần lớn các thông tin di truyền cần thiết của tế bào được lưu giữ trong nhân, chỉ một phần được lưu giữ trong ti thể (cơ quan tử có vai trò hô hấp tế bào, tổng hợp ra phần lớn ATP) hoặc lạp thể (cơ quan quang hợp ở thực vật).
4.2. Chức năng sinh học của ARN
Loại
ARN Chức năng sinh học
mARN Truyền thông tin qui định trình tự axit amin của protein từ
ADN tới ribosome
tARN Dịch các mã bộ ba trên phân tử mARN thành các axit amin
trên phân tử protein
TiềnARN
thành liên kết peptide
Sản phẩm trực tiếp của quá trình phiên mã; là phân tử tiền thân hình thành nên mARN, tARN và rARN hoàn thiện. ở eukaryote, một số phân đoạn ARN intron có vai trò xúc tác (ribozyme) phản ứng cắt chính nó
snARN
Có vai trò xúc tác và cấu trúc trong phức hệ cắt intron (spliceosome) từ các phân tử tiền mARN để tạo thành mARN hoàn thiện.
Là thành phần của phức hệ ARN-protein làm nhiệm vụ nhận biết các peptide tín hiệu trong phân tử protein mới được tổng hợp, giúp "giải phóng" các phân tử protein này khỏi mạng lưới nội chất. sno ARN Tham gia hoàn thiện rARN từ phân tử tiền rARN và đóng gói ribosome tại hạch nhân.
SRP ARN
Telomer ase-ARN Thành phần của enzym telomerase; làm khuôn để tổng hợp trình tự ADN lặp lại tại các đầu mút nhiễm sắc thể ở eukaryote.
gARN Tham gia vào quá trình "biên tập" ADN ti thể ở thực vật và nguyên sinh động vật, và ADN lạp thể ở thực vật.
tmARN ARN tích hợp chức năng của tARN và mARN, giúp giải phóng ribosome khỏi sự "tắc nghẽn" khi dịch mã các phân tử mARN bị mất bộ ba mã kết thúc (stop codon).
M1 ARN Thành phần ARN có vai trò xúc tác của ARNase P, tham gia hoàn thiện các phân tử tARN ở prokaryote.
Các loại ARN can thiệp (siARN và miARN)
Tham gia điều hòa biểu hiện gen ở eukaryote.
Xét về cấu trúc, sở dĩ ARN có thể đảm nhận nhiều chức năng khác nhau trong tế bào (khác với ADN) là do chúng có ba tính chất:
+ ARN có thể tạo liên kết hydro với các phân tử ADN hoặc ARN khác,
+ Mỗi phân tử ARN có cấu hình đặc thù do liên kết hydro hình thành giữa các phần khác nhau của nó,
+ Các ARN có thể mang nhiều nhóm chức khác, nên ARN có thể có hoạt tính xúc tác như các enzym.
Việc ngày càng phát hiện ra nhiều chức năng khác nhau của ARN là cơ sở dẫn đến quan điểm cho rằng: ARN có thể là đại phân tử sinh học đầu tiên xuất hiện trong quá trình tiến hóa (chứ không phải là ADN hay protein như nhiều giả thiết trước đây).
rARN Cấu trúc ribosome và có vai trò xúc tác (ribozyme) hình 424
423
Có thể tưởng tượng về một dạng sống nguyên thủy chỉ có ARN. Ở dạng sống này, ARN vừa có vai trò là vật chất mang thông tin di truyền, vừa biểu hiện chức năng của enzym. Dạng sống đó dần biến đổi thành thế giới sống dựa trên ba loại đại phân tử là ADN, ARN và protein. Một bằng chứng ủng hộ quan điểm protein xuất hiện sau ARN là chức năng của ARN trong ribosome. Trong ribosome, chính thành phần có vai trò xúc tác hình thành liên kết peptide (enzym peptidyl transferase) là ARN. Điểm khác biệt giữa petidyl transferase với các loại ribozyme khác (như ARNase P) là các ribozyme khác đều gây tác động xúc tác lên trung tâm phản ứng là phospho, còn petidyl transferase gây hiệu ứng xúc tác lên trung tâm phản ứng là cacbon, qua đó hình thành liên kết peptide. Có thể ribozyme của ribosome là một “vết tích” sót lại của dạng sống nguyên thủy vốn chỉ có các enzym ARN.
5. Hệ gen của virut
Vật chất di truyền hay hệ gen của virut có các đặc điểm sau đây:
- Có thể là ADN hoặc ARN mà không bao giờ chứa cả hai.
- Có dạng chuỗi đơn hoặc chuỗi kép, dạng thẳng hoặc dạng vòng.
- Hệ gen ADN kép thường có kích thước lớn, hệ gen ARN đơn thường có kích thước nhỏ.
- Tất cả virut ARN kép đều có hệ gen phân đoạn (trong mỗi hạt chứa nhiều đoạn). Virut ARN đơn không phân đoạn, ngoại trừ virut cúm.
- Hệ gen đơn của virut được chia làm 2 loại: dương và âm. Nếu trình tự của hệ gen trùng với trình tự của mARN thì quy ước là hệ gen dương, còn ngược lại (tương bù) gọi là hệ gen âm.
Một số virut chứa enzim phiên mã ngược:
- Virut viêm gan B (HBV) có hệ gen là ADN kép khép vòng, 2 sợi không bằng nhau, sợi dài là (-), sợi ngắn là (+). Khi nhân đôi phải thông qua sự tạo thành ARN trung gian (ADN ARN ADN).
- Virut retro (ví dụ HIV) có hệ gen là 2 sợi ARN giống nhau. Khi nhân đôi phải thông qua sự tạo thành ADN trung gian (ARN ADN ARN).
II. Cơ chế di truyền vi sinh vật
1. Tái bản DNA (DNA replication) ở vi sinh vật.

1.1. Các nguyên tắc và đặc điểm chung của tái bản DNA ở vi sinh vật
- Tái bản theo kiểu bán bảo toàn (semi-conservative).
- Tái bản bắt đầu tại một hoặc nhiều khởi điểm (Ori). Từ khởi điểm, DNA mở xoắn tạo thành hai chạc tái bản (replication fork). Cấu trúc như vậy gọi là đơn vị tái bản (replicon). DNA E. coli trong quá trình tái bản như vậy có cấu trúc giống chữ cái
theta Hy Lạp (θ) nên gọi là tái bản theta. Đối với các DNA mạch vòng, mỗi phân tử chỉ có một Ori; trong khi đó mỗi DNA nhiễm sắc thể eukaryote có nhiều Ori.
- Tham gia vào sự tái bản DNA có nhiều protein và enzyme.
- Tại mỗi chạc tái bản, trước tiên xảy ra sự tổng hợp các đoạn mồi RNA (primer) bởi vì các DNA polymerase tự nó không thể bắt đầu tổng hợp mới được. Mặt khác, do hai sợi đơn của mỗi chạc ngược chiều nhau trong khi các DNA và RNA polymerase chỉ xúc tác theo chiều 3' 5', cho nên sự tái bản DNA trên hai sợi khuôn là không giống nhau: một sợi liên tục gọi là sợi dẫn đầu (leading strand) và một sợi không liên tục gọi là sợi ra chậm (lagging strand). Kiểu tái bản như thế gọi là tái bản nửa gián đoạn (semi-discontinuous), được R. Okazaki phát hiện đầu tiên năm 1969.
Hình 8: Một khởi điểm và hai chạc tái bản sinh trưởng đồng thời theo hai hướng đối lập nhau, mỗi chạc gồm một sợi dẫn đầu và một sợi ra chậm.
Lưu ý: Tất cả các DNA polymerase đều cần có mồi để xúc tác tổng hợp chuỗi theo chiều 5'→3'; và chỉ có một số enzyme này là có hoạt tính đọc sửa (proofreading activity). Khác với E. coli (có ba loại DNA polymerase I, II và III; trong đó Pol II không tham gia tái bản), các tế bào eukaryote có năm loại DNA polymerase (α, β, γ, δ và ε), trong đó ba loại chịu trách nhiệm tái bản DNA nhân là các polymerase α, δ và ε. Polymerase δ chịu trách nhiệm chính cho tổng hợp ở sợi dẫn đầu và polymerase α cho sợi ra chậm. Polymerase γ tái bản DNA của các bào quan ty-lạp thể, còn polymerase β chịu trách nhiệm sửa chửa DNA.
1.2. Cơ chế tái bản ở vi khuẩn
Ở vi khuẩn, thông tin di truyền được truyền một chiều từ thể cho sang thể nhận và tạo ra hợp tử từng phần. Tái tổ hợp ở vi khuẩn có thể thực hiện bằng các đoạn ADN trần trong biến nạp, hay phage trong tải nạp, nhờ giao nạp khi 2 tế bào khác giới tính gắn với nhau.
- Đặc điểm
+ Truyền thông tin một chiều từ tế bào cho (donor) sang tế bào nhận (recipient).
+ Sự tạo thành hợp tử từng phần (merozygote). Thể cho (donor) chỉ chuyển một đoạn của bộ gen sang thể nhận (recipient) nên chỉ lưỡng bội ở một phần, các phần khác đơn bội.
+ Bộ gen thường chỉ là một ADN trần nên chỉ có một nhóm liên kết gen và tái
tổ hợp thực chất là giữa hai phân tử ADN.
- Sinh sản ở vi khuẩn
Tế bào vi khuẩn phân chia theo lối trực phân. Phân tử ADN gắn trực tiếp vào màng sinh chất. Sự sao chép ADN tạo ra hai bản sao gắn chung nhau trên màng sinh chất. Khi tế bào kéo dài ra, các bản sao ADN tách xa nhau do phần màng giữ chúng lớn dần ra. Kiểu sinh sản vô tính này được gọi là ‘phân đôi”hay “ngắt đôi” (“binary fission”) (hình 9). Tế bào vi khuẩn chia nhanh (20 phút trong điều kiện tốt) hơn rất nhiều so với tế bào Eukaryota (24-48 giờ).
E.coli chỉ có 1 sợi NST duy nhất chứa khoảng 4,6 triệu cặp nu, mỗi tế bào
E.coli có thể sao chép toàn bộ phân tử AND và chia đều cho các tế bào con trong vòng
1 giờ.
+ Giai đoạn khởi đầu (initiation) Đối với nhiễm sắc thể E. coli, sự tái bản bắt đầu tại một khởi điểm đặc thù gọi là Ori.
(1) Các protein bám khởi điểm dnaA bám Ori tạo ra cấu trúc nucleoprotein chuyên hoá của khởi điểm;
(2) Cấu trúc này mở xoắn vùng DNA giàu AT để hình thành "phức hợp mở";
(3) Hai phân tử helicase dnaB chui vào phức hợp mở làm mở xoắn khởi điểm theo cả hai hướng, tạo thành hai chạc tái bản (replication fork). Khi cả hai sợi đơn của mỗi chạc được tách ra thì các protein SSB bám vào.
Hình 9: Trực phân ở vi khuẩn Quá trình sao chép ADN được bắt đầu từ điểm xuất phát oriC kéo dài về hai phía song song với quá trình sao chép màng sinh chất, nơi có điểm gắn vào của ADN bộ gen, mọc dài tách 2 phân tử ADN về 2 tế bào con. ADN của E.coli cần 40 phút cho 1 vòng sao chép tương ứng với tốc độ 50.000 cặp bazơ/phút. Phụ thuôc vào tốc độ tăng trưởng, thời gian phân chia tế bào trong khoảng từ 18 đến 60 phút. Như vậy ở các tế bào tăng trưởng nhanh, vòng sao chép mới phải được bắt đầu sớm hơn sự phân bào trước đó như tế bào con đầu tiên.
Hình 10: Hình thành phức hợp mở đầu (primasome) tại khởi điểm với việc tổng hợp đoạn mồi đầu tiên ở một chạc tái bản.
Khởi đầu trong tái bản DNA ở E. coli là sự tổng hợp một đoạn mồi ngắn khoảng 10-12 nucleotide bởi primase (xét chung ở các sinh vật là ~ 5 base). Cả ba loại protein dnaB, dnaC và dnaG hợp thành một phức hợp có tên là primasome (hoặc primosome: thể mở đầu; hình 3).

+ Giai đọan kéo dài (elongation)
Một khi primasome tổng hợp xong một mồi, sự kéo dài chuỗi DNA được bắt đầu bằng một phức hợp giữa primasome và DNA polymerase III hoàn chỉnh, gọi là replisome (thể tái bản). Sự tái bản DNA trên mỗi chạc xảy ra theo kiểu nửa gián đoạn (semi-discontinuous), như (hình 4)
Trên sợi khuôn dẫn đầu (3'→5'): Sau khi primase (primasome) tổng hợp xong đoạn mồi RNA với đầu 3'-OH tự do, enzyme hoàn chỉnh DNA polymerase III (hay replisome) bắt đầu kéo dài chuỗi DNA mới sinh trưởng theo chiều 5'→3' một cách liên tục.

Trên sợi khuôn ra chậm (5'→3'): Sự kéo dài diễn ra không liên tục dưới dạng các đoạn Okazaki. Kích thước trung bình mỗi đoạn Okazaki ở E. coli là 1.000 - 2.000 nucleotide. Quá trình này đòi hỏi sự "mồi hóa" lặp lại, với sự tham gia lần lượt của bốn enzyme sau:
- Cơ chế tái bản ADN (E. coli) 428
(i) primase tổng hợp một đoạn mồi RNA;
(ii) DNA polymerase III kéo dài đoạn Okazaki;
(iii) DNA polymerase I vừa cắt bỏ đoạn mồi vừa lấp khoảng trống
bằng cách kéo dài dần đoạn Okazaki theo sau;
(iv) DNA ligase hàn liền khe hở giữa hai đoạn Okazaki bằng một liên kết phosphodiester.
+ Phức hệ sao chép AND: nhiều protein khác nhau, không di chuyển dọc AND, mà AND sẽ chui qua phức hệ khi sao chép.
+ Đọc và sửa chữa AND: nhờ E. AND Pol. Ở E.coli có khoảng 100 E sửa chữa AND. Sửa chữa dựa trên NTBS.
Sửa chữa bằng cắt bỏ nucleotit nhờ E cắt AND- nuclease sau đó lấp đầy chỗ trống nhờ AND-Pol và AND-ligase.
+ Giai đọan kết thúc (termination)
Do cấu trúc nhiễm sắc thể ở hai nhóm prokaryote và eukaryote là hoàn toàn khác nhau, nên cơ chế kết thúc tái bản của chúng cũng khác nhau.
Cả hai chạc tái bản của DNA E. coli được bắt đầu từ một khởi điểm (ori), và di chuyển hầu như cùng tốc độ theo hai hướng đối lập nhau xung quanh nhiễm sắc thể mạch vòng cho tới khi chúng gặp nhau tại một điểm kết thúc chung đối diện với ori.
Theo Bastia và cộng sự (1997) cũng như nhiều tác giả khác, đây là vùng chứa các trình tự đặc thù gọi là các điểm kết thúc tái bản (replication termini). Tại các trình tự này có các protein kết thúc tái bản (replication terminator protein = RTP) bám vào và các phức hợp protein-DNA này ngăn cản sự di chuyển của các chạc tái bản. Đó là bước đầu tiên của sự hoàn thành một vòng tái bản, và sau đó tách hai nhiễm sắc thể con rời ra một cách có trật tự nhờ xúc tác của topoisomerase IV.
RTP của E. coli có TLPT ~36kD, được mã hóa bởi gene tus (ter) và trong mỗi tế bào có ~80 bản sao của protein này được duy trì hầu như ổn định trong suốt chu kỳ tế bào. Trình tự điều hoà của vùng kết thúc tái bản ở E. coli (R6K), theo Bastia và cộng sự (1997), như sau: 5'NN(A/T)(A/T)(A/T)G(A/T)(A/G)TGTTGTAACTA(A/C)NN3'
Vấn đề kết thúc tái bản ở eukaryote: Mỗi nhiễm sắc thể eukaryote chứa một phân tử DNA sợi kép mạch thẳng kết hợp với nhiều loại protein, có các đầu mút đặc trưng gọi là telomere. Các telomere có cấu trúc đơn giản gồm những trình tự ngắn (6-8 bp) lặp lại nối tiếp cả ngàn lần và đặc thù cho từng loài. Chẳng hạn, ở Tetrahymena là (TTGGGG)n. Một khi đọan mồi đầu tiên trên mỗi sợi được loại bỏ thì nó không có cách nào để bù đắp lại khoảng trống đó, bởi vì DNA không thể nào nới rộng theo chiểu 3'→5'. Các kết quả nghiên cứu đầu tiên của Elizabeth Blackburn và cs đã giải đáp cho vấn đề này. Các đoạn lặp này được gắn thêm vào đầu 3' của các sợi DNA nhờ sự xúc tác của telomerase, một enzyme phiên mã ngược (reverse transcriptase). Chẳng hạn, telomerase của Tetrahymena có một RNA dài 159 nucleotide có chứa trình tự 3'CAACCCCAA5' làm khuôn cho tổng hợp các đoạn lặp 5'-TTGGGG-3'.
Vai trò của các Enzyme(E) tái bản AND ở vi khuẩn E.coli
TT
Enzym Gen mã hóa Chức năng
ADN polymeaz I polA-E; mutD. polA Là E phục hồi chủ yếu. Cắt mồi ARN, lắp chỗ trống
ADN polymeaz II holA-E; mutD. Enzym phục hồi phụ (minor).
ADN polymeaz III polC; ADNE,Q,N,X; Enzym polyme hoá chủ yếu
ADN polymeaz IV polC Phục hồi cấp cứu (SOS repair).
ADN polymeaz V ADNB umuD’2C Phục hồi cấp cứu (SOS repair).
0
Helicase ADNB Tháo xoắn tại chạc ba sao chép
Primase ADNG Tạo mồi mạch ADN mới
Protein gắn
điểm khởi sự (Origin-binding protein).
Protein căng mạch SSB (Singlestrand binding protein)
ADNA Gắn điểm khởi sự sao chép (Ori); tạo thuận lơi cho mở tách mạch.
SSB Ngăn các mạch đơn đã tách không chập lại.
ADN ligaz ligA, ligB Nối các đầu hở trên ADN
2. Cơ chế phiên mã ở vi sinh vật
2.1. Sơ lược về các gene
Một cách tương đối, gene (cistron) là một đọan xác định của bộ gene mã hóa thông tin của một polypeptid hoặc một phân tử RNA chức năng.
Ở các prokaryote và eukaryote bậc thấp, thường có một mối quan hệ đơn giản giữa gene và sản phẩm của nó (một gene - một sản phẩm). Hơn nữa, các gene đồng nghĩa với vùng mã hóa hay khung đọc mở (open reading frame). Nghĩa là, ở các prokaryote các gene liên quan về chức năng thường được tổ chức trong một operon, vì thế có nhiều sản phẩm được dịch mã từ một mRNA polycistron. Trái lại, ở các eukaryote, các gene đồng nghĩa với đơn vị phiên mã (transcription unit) và hầu hết chúng được phiên mã dưới dạng mRNA monocistron.
Ở các bộ gene eukaryote bậc cao, thường có một mối quan hệ phức tạp giữa gene và sản phẩm. Hầu hết các gene đều chứa các intron (các đoạn không mã hóa protein) nằm xen giữa các exon (các đoạn mã hóa protein). Các gene như vậy được gọi là gene phân đoạn (split gene), được Phillip Sharp phát hiện đầu tiên năm 1977.
2.2. Các nguyên tắc và đặc điểm chung của phiên mã
Phiên mã (transcription) là quá trình tổng hợp các RNA khác nhau từ thông tin di truyền chứa đựng trong DNA. Trừ các gene mã hóa protein trong các operon ở vi khuẩn, nói chung, các RNA mới được tổng hợp chỉ là các bản sao sơ cấp (primary transcript) gọi là các pre-RNA. Các pre-RNA này phải trải qua một quá trình sửa đổi để trở thành các RNA trưởng thành (mature) trước khi tham gia vào quá trình sinh tổng hợp protein của tế bào. Quá trình phiên mã các đặc điểm chung sau đây

Hình 11: Phiên mã và dịch mã ở sinh vật nhân sơ.
(i) Diễn ra dưới tác dụng của các enzyme RNA polymerase.

(ii) Chỉ một sợi đơn được dùng làm khuôn cho tổng hợp RNA, gọi là sợi khuôn, sợi mã hoá hay sợi có nghĩa (template/ coding / sense strand); còn sợi bổ sung được gọi là sợi đối khuôn, sợi không mã hoá hay sợi đối nghĩa (antitemplate/ noncoding / antisense strand).
(iii) Phản ứng tổng hợp RNA diễn ra theo nguyên tắc bổ sung và được kéo dài theo chiều 5'→3', ngược với chiều của sợi khuôn.
(iv) Nguyên liệu cho tổng hợp gồm: ATP, UTP, GTP và CTP.
(v) Sản phẩm của phiên mã là các RNA sợi đơn.
(vi) Khởi đầu và kết thúc phiên mã phụ thuộc vào các tín hiệu điều hoà là các trình tự DNA đặc thù nằm trước gen (vùng khởi động) và sau gene.
2.3. RNA polymerase và vùng khởi động (promoter) của VSV nhân sơ
Hình 12: Cấu trúc promoter của prokaryote.
Ở các prokaryote, RNA polymerase hoàn chỉnh (holoenzyme) là một phức hợp gồm nhân tố sigma (σ) và lõi enzyme. Nhân tố σ giúp RNA polymerase nhận biết và bám vào promoter để có thể bắt đầu phiên mã tại vị trí chính xác, và lõi enzyme đóng vai trò xúc tác tổng hợp RNA.
Vùng khởi động (promoter) nói chung nằm kề trước gene và có chứa các trình tự đặc thù cho phép RNA polymerase nhận biết và bám vào. Trình tự quan trọng nhất của promoter là hộp TATA hay hộp Pribnow (Pribnow box) ở vị trí "-10". Ngoài ra, còn có trình tự TTGACA ở vị trí ''-35'', gọi là đoạn nhận biết (recognition sequence).
Các vùng này được bảo tồn cao.
2.4. Các giai đoạn của quá trình phiên mã ở prokaryote
- Khởi đầu: RNA polymerase holoenzyme nhận biết và bám vào promoter, tháo xoắn một đoạn khoảng 12 cặp base. Sau khi tổng hợp ribonucleotide đầu tiên (Appp hoặc Gppp), nhân tố sigma tách ra để đi vào một chu kỳ phiên mã khác, gọi là chu kỳ sigma (sigma cycle).
- Kéo dài: Enzyme lõi tổng hợp sợi RNA dọc theo sợi khuôn.
- Kết thúc: Khi phiên mã xong hai vùng giàu GC và AT nằm sau gene, ở vùng đuôi của sợi RNA hình thành cấu trúc ''kẹp cài tóc'' (hairpin loop) làm dừng sự phiên mã của lõi RNA polymerase. Cuối cùng, dưới tác dụng của nhân tố rho (ρ), sợi RNA và enzyme lõi được giải phóng khỏi DNA khuôn.
3. Dịch mã
Cơ chế của quá trình dịch mã (tổng hợp polypeptide)
- Mở đầu (initiation): Quá trình dịch mã bắt đầu khi một tiểu đơn vị
ribosome bé bám vào mRNA tại vị trí của codon khởi đầu AUG. Lúc này một phân tử tRNA khởi đầu đặc thù mang methionine (ở vi khuẩn là formyl-Met) đi
vào và khớp anticodon của nó với codon mở đầu của mRNA. Kế đó, tiểu đơn vị ribosome lớn bám vào tiểu đơn vị bé tạo ra một ribosome hoạt động hoàn chỉnh.
Lúc này Met-tRNA ở vị trí P và vị trí A để trống; một tRNA thứ hai (ví dụ, tRNAVal) đi vào vị trí A và khớp với codon thứ hai (hình 13A).
- Kéo dài (elongation): Quá trình kéo dài bắt đầu sau khi liên kết peptide đầu tiên được hình thành. Phản ứng này được xúc tác bởi enzyme peptidyl transferase, và kết quả là tạo ra một peptidyl-tRNA ở vị trí A (hình 13.B). Sau đó, ribosome lập tức chuyển dịch sang một codon mới dọc theo mRNA theo chiều 5'→3' (hình 13.C). Phản ứng này đẩy phân tử tRNA tự do vốn ở vị trí P ra ngoài; lúc này peptidyl-tRNA nằm ở vị trí P và vị trí A lại để trống. Một chu kỳ dịch mã mới lại bắt đầu, một aminoacyl-tRNA thứ ba đi vào và khớp anticodon của nó với codon đang để trống ở vị trí A, một liên kết peptide thứ hai được hình thành, và ribosome lại dịch chuyển sang codon kế tiếp. Quá trình nói trên cứ diễn ra một cách tuần tự dọc theo mRNA làm cho chuỗi polypeptide dài dần ra cho đến dịch mã xong codon có nghĩa cuối cùng.
- Kết thúc (termination): Quá trình tổng hợp polypeptide sẽ dừng lại khi codon kết thúc của mRNA đối diện với vị trí A. Lúc này nhân tố giải phóng RF (release factor) đi vào (Hình 13.D); chuỗi polypeptide được tách ra và phóng thích cùng với hai tiểu đơn vị ribosome cũng như tRNA ra khỏi mRNA.
Lưu ý:
(1) Thực ra, trên một mRNA có rất nhiều ribosome cùng hoạt động, gọi là polysome, tạo ra nhiều polypeptide giống nhau.

(2) Trên nguyên tắc, amino acid mở đầu sẽ được cắt bỏ khỏi chuỗi polypeptide. Tuy nhiên, ở các eukaryote không phải lúc nào amino acid mở đầu này cũng bị tách bỏ, mà trong một số protein nó vẫn được giữ lại.
(3) Sau tổng hợp, các chuỗi polypeptide sẽ được sửa đổi và chuyển sang các bậc cấu trúc cao hơn để trở thành các protein chức năng. Thực ra sự biến đổi sau dịch mã còn có các chaperone và nhiều cơ chế tác động phức tạp khác nữa.
(4) Tham gia vào các bước mở đầu, kéo dài và kết thúc còn có các yếu tố protein, với tên gọi tương ứng là các nhân tố mở đầu, kéo dài, và giải phóng cùng với ATP, GTP và các ion Mg2+, K+ và NH4 + .
(5) Trong các tế bào prokaryote, các ribosome và các aminoacyl-tRNA sẽ bám vào đầu 5' của mRNA để bắt đầu quá trình dịch mã trong khi ở đầu 3' của nó quá trình phiên mã đang còn tiếp diễn. Ngược lại, ở các tế bào eukaryote, các pre-mRNA phải trải qua sửa đổi sau phiên mã ở trong nhân, còn dịch mã diễn ra sau đó trong tế bào chất.
(6) Về RNA đối nghĩa (antisense RNA), đây là loại RNA thấy có ở nhiều hệ thống, nhưng rất phổ biến ở các vi khuẩn. Nó được tổng hợp từ sợi đối nghĩa của gene, nên bổ sung với mRNA và có thể tạo thành một sợi kép với nó để gây kìm hãm dịch mã. Vì vậy RNA đối nghĩa còn được gọi là RNA bố sung gây nhiễu mRNA, và được ứng dụng hiệu quả trong điều trị ung thư.
4. Tái tổ hợp di truyền ở vi khuẩn
4.1. Hiện tượng biến nạp ở vi khuẩn Diplococus pneumoniae
Hình 14: Hiện tượng biến nạp
Tiêm vi khuẩn S sống gây bệnh cho chuột - chuột chết.
Tiêm vi khuẩn R sống không gây bệnh - chuột sống.
Tiêm vi khuẩn S bị đun chết cho chuột - chuột sống.
Hỗn hợp vi khuẩn S bị đun chết trộn với vi khuẩn R sống đem tiêm cho chuộtchuột chết. Trong xác chuột chết có vi khuẩn S và R.
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các tế bào chết này đã truyền tính gây bệnh cho tế bào R. Nó được gọi là biến nạp (transformation).
Năm 1944, T.Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng proteaz (enzym phân hủy protein) hoặc ARN-az (enzym phân hủy ARN) hoạt tính biến nạp vẫn còn, chứng tỏ protein và ARN không phải là tác nhân gây biến nạp. Nhưng nếu tế bào S chết bị xử lý bằng ADN-az (enzym chỉ phân hủy đặc hiệu ADN) thì hoạt tính biến nạp không còn nữa, chứng tỏ ADN là nhân tố biến nạp.
Kết quả thí nghiệm có thể tóm tắt như sau:
ADN của S + các tế bào R sống → chuột → chết (có R + S)
Như vậy, biến nạp là hiện tượng truyền thông tin di truyền bằng ADN. Trong biến nạp, ADN trần từ một tế bào vi khuẩn (thể cho) này được truyền sang tế bào vi khuẩn khác (thể nhận). Biến nạp xảy ra khi vi khuẩn nhận ADN ngoại lai và hấp thu vào trong tế bào. Khi tế bào vi khuẩn bị vỡ do bị tan (lysis), ADN vòng tròn của chúng thoát ra môi trường thành các đoạn thẳng với chiều dài khác nhau, có khả năng gây biến nạp cho các tế bào nhận khác.
Cơ chế:
+ Sự phân hủy ADN tế bào cho: ADN tế bào cho có thể là của tế bào tự nhiên bị phân hủy hoặc trong thí nghiệm bị gây chế bằng nhiệt độ cao hay tác nhân phá vỡ tế bào.
+ ADN bám vào bề mặt tế bào: Protein gắn vào ADN.
+ Thâm nhập của ADN: Sợi ADN mạch kép của dòng vi khuẩn S sau khi chui qua màng tế bào của dòng R thì một mạch S sẽ bị nucleaz của tế bào cắt, còn lại một mạch nguyên. + Bắt cặp (Synapsis)và tái tổ hợp: Nhờ sự hỗ trợ của protein RecA ADN của thể nhận R sẽ biến tính tách rời 2 mạch ở 1 đoạn dễ bắt cặp với đoạn ADN thể cho S vừa chui vào.
Hình 15: Sơ đồ diễn biến của quá trình biến nạp ở Vi khuẩn
4.2. Tải nạp ở vi khuẩn + Trong tải nạp (transduction), các virut mang các gen từ tế bào này sang tế bào khác. Ở chu trình tan (lytic cycle), một số bacteriophage gói nhầm ADN vi khuẩn chủ vào capsid. Tế bào bị nhiễm bởi các virut như vậy nhận đoạn ADN của vi khuẩn A khác, chứ không phải ADN của virut. Do vậy, ADN vi khuẩn A tái tổ hợp với ADN nhiễm sắc thể của tế bào chủ B và biến đổi thành phần di truyền (gentic composition).


+ Phage là nhân tố chuyển gen
Thí nghiệm được tiến hành trong ống hình chữ U, ở đáy ống được ngăn cách bằng màng lọc vi khuẩn. Màng có lỗ nhỏ vi khuẩn không qua được, nhưng phage qua được (hình 7). Nhánh A của ống chứa vi khuẩn có khả năng tổng hợp tryptophan (trp+ ), còn nhánh B nuôi các vi khuẩn mất khả năng tổng hợp tryptophan (trp– ). Sau một thời gian nuôi bên nhánh B xuất hiện vi khuẩn có khả năng tổng hợp tryptophan (trp+
). Qua nhiều lần thí nghiệm việc phage tải gen trp+ từ nhánh A sang nhánh B được chứng minh.
Khi ADN của phage xâm nhập tế bào vi khuẩn A, chúng cắt ADN của vi khuẩn A thành nhiều đoạn, đồng thời ADN của phage được sao chép thành nhiều phân tử con và các vỏ capsid của phage cũng được tạo thành. Sau đó các vỏ capsid được lắp ruột ADN vào, phá vỡ tế bào vi khuẩn ra ngoài và tiếp tục xâm nhập các vi khuẩn mới.
Trong quá trình lắp ráp khoảng 1-2% phage vô tình mang đoạn ADN của vi khuẩn có chứa gen. Phage mang gen của vi khuẩn A xâm nhập vi khuẩn B, quá trình tái tổ hợp xảy ra làm gen A gắn vào bộ gen B.
Tải nạp chuyên biệt (Special transduction) Tải nạp chuyên biệt hay hạn chế (restricted transduction) là trường hợp chỉ mang một vài gen nhất định, nó có 4 đặc điểm:

+ Những gen được chuyển nằm sát chỗ prophage gắn vào.
Hình 16: Thí nghiệm chứng minh có tải nạp do virut
Tải nạp chung (Genral transduction) Nhánh B Vi khuẩn Nhánh A Vi khuẩn trp+ Màng lọc vi khuẩn Tải nạp chung xảy ra khi phage mang bất kì gen nào của vi khuẩn A chuyển sang vi khuẩn B. Tải nạp chung (genralized transduction) có các đặc điểm:

+ Thường do phage kiểu P1 thực hiện.
+ Bất kì gen nào của vi khuẩn cũng đều được tải nạp.
+ Tải nạp có được do gói nhầm ADN của tế bào chủ khi phage trưởng thành.
+ Các thể tái tổ hợp đơn bội được tạo ra.
Do không có sự tương đồng giữa trình tự ADN trên các phage này với trình tự ở tế bào chủ, nên không có điểm gắn vào đặc hiệu cho prophage. Bất kì gen nào cũng
được tải nạp vì đầu của phage có thể gói nhầm vào một đoạn ADN của tế bào chủ. Sự
đồng tải nạp (cotransduction) là quá trình tải nạp đồng thời 2 gen. Quá trình phage xâm nhập vào vi khuẩn và sinh sản được mô tả trên hình 4. Đầu tiên phage bám trên
bề mặt của vi khuẩn, sau 4 phút bơm ADN của nó vào tế bào, sau đó chúng sinh sản và độ nửa giờ sau thì làm tan vi khuẩn và giải phóng các phage con mới.
+ Chỉ prophage kiểu λ thực hiện.
+ Do kết quả sự cắt sai của prophage khi tách khỏi nhiễm sắc thể của tế bào chủ.
+ Các vi khuẩn tái tổ hợp có thể lưỡng bội một phần.
Ví dụ: Phage λ chỉ mang gen gal (đồng hóa đường galactoz) từ vi khuẩn này chuyển sang vi khuẩn khác.
Hình 17. Quá trình xâm nhập của phage và làm tan vi khuẩn
ADN phage Nhiễm sắc thể vi khuẩn A Capsid Phage mới phá vỡ tế bào, nhiễm
VK B
Hình 18: Điểm gắn của phage λ vào bộ gen vi khuẩn
Điểm gắn của phage λ vào bộ gen của vi khuẩn nằm giữa 2 gen gal (galactoz) và bio (tổng hợp biotin), tiếp theo xảy ra quá trình tái tổ hợp ở điểm chuyên biệt (sitespecific recombination) chèn ADN của phage vào Nhiễm sắc thể vi khuẩn (hình 9).
438

Đầu của phage chỉ có thể chứa một lượng ADN giới hạn, nó chỉ tải nạp được gen gal hoặc bio. Phage λ tải nạp các gen galactoz được gọi là λgal hay λdg (d = defective: khuyết, g = galactoz). Nếu tế bào gal– được nhiễm bởi λdg (mang gen gal+ ), sự ráp phage biến dạng vào tế bào chủ sẽ tạo lưỡng bội một phần. Sự cắt sai của phage λ rất
hiếm nên tải nạp hạn chế có tần số thấp.
Tuy nhiên, tải nạp tần số cao có thể nhận được trong điều kiện thí nghiệm.
Nếu tế bào vi khuẩn được gây nhiễm kép với phage λ hoang dại và phage λdg, phage hoang dại có thể hỗ trợ chức năng sai sót ở phage biến dạng, và thế hệ con sẽ có cả 2 kiểu với số lượng bằng nhau. Khi dịch tan được dùng tải nạp, quá trình được gọi là tải nạp tần số cao.
Trong nhiều trường hợp, do bộ gen biến dạng λdg không gắn được vào bộ gen của tế bào chủ (nên không sao chép được). Sau mỗi lần phân bào, chỉ một trong 2 tế bào có bộ gen của phage biến dạng; quá trình này được gọi là tải nạp sẩy (abortive transduction).
4.3. Giao nạp (Conjugation)
Vào năm 1946, J. Lederberg và E. Tatum chứng minh có trao đổi vật chất di truyền giữa các vi khuẩn sống. Sự trao đổi này gọi là giao nạp hay tiếp hợp (conjugation). Giao nạp ở vi khuẩn là sự kết hợp nhất thời của hai tế bào có kiểu bắt cặp đối nhau, được tiếp nối bằng sự chuyển một phần vật chất di truyền từ tế bào cho sang tế bào nhận qua cầu tế bào chất, và sau đó các tế bào tách nhau ra (exconjugants).
Giao nạp đòi hỏi sự tiếp xúc trực tiếp giữa 2 loại tế bào được khởi sự bằng ống giao nạp hay tính mao (pilus), một sợi ống nhỏ rất dài do tế bào cho (donor cell) tạo ra (hình 19).
trình, nhất quán với biến nạp, tải nạp ở vi khuẩn và khỏi nhầm lẫn v
ới tiếp h
hoặc (Hfr x F-). Dòng tế bào mang nhân tố F+ được coi là tế bào “đực” và Plasmid có
khả năng tạo protein pilin, từ protein này tạo ống giao nạp là pilus. Sự co lại của pilus
đang nối hai tế bào làm chúng tiếp xúc kề nhau. Tế bào F– được coi là cái (female).
Sau khi giao nạp tế bào F– trở thành F+ . Việc chuyển gen chỉ thực hiện khi plasmid gắn vào bộ gen của vi khuẩn. Trong quá trình chuyển vật chất di truyền sang F– thì ADN của tế bào chủ sao chép và mạch mới có ori đi đầu và F ở cuối (hình 20). Quá trình chuyển ADN từ F+ sang F– có thể bị ngắt quãng.
Hfr X F– Sao chép ADN truyền cho F– Hfr F– tái tổ hợp
Hình 20. Sự truyền ADN từ thể cho sang thể nhận Các gen A, B, C được chuyển một chiều từ Hfr sang F–.

Đoạn gen thể cho bắt cặp với các gen tương đồng của ADN tế bào F– và diễn ra trao đổi chéo tạo tế bào F– tái tổ hợp. Dòng Hfr có tần số lai cao hơn nhiều vì plasmid đã nằm sẵn trong bộ gen. Còn F+ phải qua giai đoạn plasmid gắn vào bộ gen rồi mới chuyển gen.
Trong điều kiện thí nghiệm ở 37oC, nguyên bộ gen của tế bào E. coli được chuyển sang tế bào nhận trong vòng 90 phút. Thường sự giao nạp bị ngắt giữa chừng do các pilus bị gãy, nên ít khi bộ gen được chuyển nguyên vẹn vào tế bào nhận. Lúc đó tế bào F–vẫn là F– . Bằng cách ngắt quãng giao nạp bản đồ di truyền của E.coli được xây dựng và có dạng 1 vòng tròn.
Hình 19. Hai tế bào vi khuẩn giao nạp qua cầu tế bào chất pilus
Hình 21: Đoạn ADN từ thể cho trao đổi chéo với ADN thể nhận tạo F– tái tổ hợp.
Sao chép Plasmid


Plasmid là nhân tố di truyền sao chép độc lập với nhiễm sắc thể (Nhiễm sắc thể) của tế bào chủ. Không giống virut, plasmid không có dạng ngoại bào (extracellular form) và tồn tại trong tế bào đơn giản là ADN tự do và điển hình là dạng vòng tròn. Plasmid và ADN Nhiễm sắc thể của tế bào có vài điểm khác nhau cơ bản: plasmid chỉ mang những gen không thiết yếu (nhưng thường rất có ích), còn những gen thiết yếu nằm trên Nhiễm sắc thể. Plasmid di chuyển giữa các tế bào trong quá trình giao nạp (conjugation), tức sự tiếp xúc giữa hai loại tế bào.
gọi tên loại virut, ví dụ: virut thực vật kí sinh tế bào thực vật, virut động vật kí sinh tế bào động vật. Do đặc điểm này, sự sinh sản của virut khác hẳn với sự sinh sản của tế bào. Điểm nổi bậc là virut tạo ra hàng trăm hay hàng ngàn virion trong mỗi thế hệ.
Do có sự đa dạng như nêu trên, các bộ gen của virut được sao chép theo nhiều con đường khác nhau:
+ Các ADN mạch kép –> ADN mạch kép: kiểu bán bảo tồn.
+ Các ARN mạch kép –> ARN mạch kép: kiểu bán bảo tồn nhờ ARN replicaz do gen tạo enzym sao chép này được mã hóa trong bộ gen của virut ARN.
+ Các ADN hoặc ARN mạch đơn –> chúng làm khuôn (mạch +) để tổng hợp mạch bổ sung (mạch –) thành dạng mạch kép trung gian (+/–), gọi là dạng sao chép (replicative form), mà từ nó lấy mạch – làm khuôn tạo ra mạch đơn + giống bộ gen ban đầu của virut (hình 23).
Hình 22. Hình hiển vi điện tử các plasmid vòng tròn
Enzym liên quan đến việc sao chép plasmid trên thực tế là những enzym bình thường trong tế bào. Vì thế những gen của plasmid chủ yếu liên quan đến sự kiểm soát quá trình khởi sự sao chép và với sự phân bổ những plasmid được sao chép vào các tế bào con.
Tương tự, những plasmid khác nhau hiện diện trong tế bào với số lượng khác nhau, gọi là số bản sao (copy number). Một số plasmid chỉ có 1-3 bản sao trong tế bào, trong khi những plasmid khác lại có đến 100 bản sao. Số lượng bản sao được kiểm soát bởi những gen trên plasmid và bởi sự tương tác gen giữa tế bào chủ và plasmid. Phần lớn các plasmid ở vi khuẩn Gram âm sao chép tương tự như Nhiễm sắc thể vi khuẩn. Việc này thực hiện từ điểm khởi sự sao chép (Ori) và chép theo hai hướng quanh vòng plasmid, tạo ra dạng trung gian theta (θ). Tuy nhiên một số plasmid không sao chép theo hai hướng. Do kích thích nhỏ của ADN plasmid, toàn bộ quá trình sao chép xảy ra rất nhanh, có lẽ bằng 1/10 tổng thời gian diễn ra chu kì phân bào. Hầu hết plasmid của vi khuẩn Gr+ sao chép theo cơ chế vòng xoay tương tự như phage φx. Cơ chế này sẽ tạo ra mạch đơn trung gian, và do đó những plasmid này thỉnh thoảng được xem như plasmid ADN mạch đơn. Phần lớn ADN mạch thẳng sao chép dùng cơ chế liên quan đến một protein gắn với đầu 5’ của mỗi mạch mà được dùng trong tổng hợp mồi ADN. Cơ chế chính của việc chuyển plasmid từ tế bào này sang tế bào khác là giao nạp, mà chức năng thực hiện được mã hóa bởi chính một số plasmid. Giao nạp là một quá trình sao chép ADN và plasmid của tế bào cho và truyền bản sao của Nhiễm sắc thể cùng bản sao plasmid sang tế bào nhận.
5. Cơ chế di truyền của virut
Hình 23. Sao chép của ADN hoặc ARN mạch đơn qua trung gian mạch kép

Ở Retrovirut ARN –> nhờ enzym reverse transcriptaz tổng hợp c-ADN mạch đơn từ khuôn ARN virut –> c-ADN kép –> Chèn vào bộ gen tế bào chủ –> tạo ra bộ gen ARN virut mới và mARN.
Phụ thuộc vào bộ gen ADN hay ARN mạch kép hay đơn, mạch dương (+) hay âm (–), kiểu sao chép bộ gen và tổng hợp mARN, các virut được chia thành 7 lớp (classes).
Một cách sao chép đặc biệt được tế bào vi khuẩn sử dụng trong tiếp hợp để truyền phân tử ADN dạng thẳng sang tế bào khác hoặc được các virut sử dụng để tạo các bộ gen của chúng, mặt khác làm đứt mạch của vòng tròn xoắn kép, tạo đầu hở 3’–OH và 5’–P kết thúc. Helicaz và SSB protein chen vào tạo chẽ 3 sao chép. Sự sao chép được thực hiện không cần mồi (primer) vì mạch 3’–OH sẵn sàng cho việc nối dài như mạch trước (leading strand) nhờ ADN polymeaz I. Đồng thời với sao chép mạch trước, mạch khuôn sau dịch chuyển. Sự dịch chuyển của mạch sau gián đoạn để tổng hợp các đoạn ngắn Okazki như bình thường và đầu 5’ mạch khuôn duỗi thẳng ra (hình 19). Sự sao chép có hình giống chữ sigma (σ) của Hi Lạp, nên được gọi là sao chép sigma (σ replication). Kiểu sao chép σ này còn được gọi là vòng tròn quay (rolling-circle replication), vì mạch khuôn tròn ở giữa không bị đứt và quay tròn làm khuôn cho mạch trước. Sự sao chép kiểu này có thể lặp lại vài lần tạo ra sợi ADN dài, lặp lại nhiều lần bộ gen thẳng của virut, được gọi là concatemer. Enzym endonucleaz cắt ở những điểm khác nhau trên mỗi mạch của ADN tạo ra các đoạn cỡ bộ gen với hai đầu “dính”. Bộ gen thẳng này sẽ tạo thành vòng nhờ bắt cặp bổ sung các đầu “dính”.
Các hạt virut (virut particle) hay virion là những vật kí sinh nội bào bắt buộc (obligate intracellular parasite). Chúng chỉ biểu hiện các gen của chúng và sinh sản bên trong 1 tế bào sống khác. Phụ thuộc vào loại tế bào chủ mà virut kí sinh, người ta 442
441

5.1. Vi rút động vật
Các chu trình sao chép của virut động vật có nhiều điểm tương tự với các virut khác với nhiều biến dạng đáng kể. Ví dụ trường hợp của cúm (influenza virut) hay của Paramyxovirut gồm các virut gây bệnh sởi và quai bị. Các virut cúm và Paramyxovirut có bộ gen là ARN một mạch được gói trong capsid xoắn dẻo. Phía bên ngoài capsid có màng bao là tính chất chung của nhiều nhóm virut động vật và phage (hình 20).
Hình 24. Virut cúm
Bao màng giúp virut xâm nhập tế bào chủ. Khi virut tiếp xúc với tế bào, các glycoprotein thòi ra ở màng bao gắn vào các thụ thể (receptor) trên màng sinh chất (hình 20).

Quá trình này chuyển capsid có chứa ARN vào tế bào chất và ở đó capsid bị mất (bóc vỏ). Các enzym của virut tham gia sao chép ARN của bộ gen và tạo mARN, nhưng bộ máy của tế bào được sử dụng để tổng hợp protein của virut. Các capsid mới được lắp ráp bao các bộ gen của virut và chúng đội màng sinh chất của tế bào mọc chồi rồi thoát ra khỏi tế bào. Bằng cách này các capsid có màng bao chính là màng sinh chất của tế bào chủ cũ và các virion mới có thể sử dụng các bao màng này để hòa nhập với màng của tế bào chủ mới. Toàn bộ chu trình này được gọi là chu trình sinh sản (reproductive cycle), chứ không phải chu trình tan vì các virut có thể thoát ra bằng mọc chồi mà không phải phá vỡ tế bào chủ Đáng chú ý là chu trình sinh sản của các virut loại này diễn ra ở tế bào chất của tế bào và không tác động đến nhân tế bào.
Hình 25. Chu trình sinh sản của virut cúm
Không phải tất cả các màng của virut đều bắt nguồn từ màng sinh chất. Ví dụ, bao màng của herpesvirut từ màng nhân của tế bào chủ. Các bộ gen của herpesvirut là ADN mạch kép , chúng sinh sản bên trong nhân tế bào, sử dụng phối hợp cả enzym của virut và tế bào để sao chép ADN và phiên mã.
Khi ở trong nhân của tế bào, ADN của herpesvirut (tiền virut) tương tự prophage ở vi khuẩn. Provirut ở dạng tiềm ẩn có thể gây một số triệu chứng bệnh ở người. Đôi khi các stress vật lí (như tác động ánh sáng mặt trời mạnh) hay tình cảm có thể làm cho provirut herpes bắt đầu chu trình sinh sản tạo các triệu chứng khó chịu cho con người.
5.2. Các retrovirut
Các virut có bộ gen ARN sinh sản qua trung gian ADN. Retrovirut HTLV-I gây bệnh bạch cầu tế bào T (T-cell leucemia) và retrovirut HIV gây bệnh AIDS làm suy yếu hệ thống miễn dịch (hình 26).Nói chung, retrovirut liên quan đến những bệnh nguy hiểm, khó chữa trị nhất hiện nay của nhân loại, kể cả ung thư.
Tất cả các virut biến đổi tế bào qua việc gắn nucleic axit của virut vào ADN của tế bào chủ. Sự gắn này được ổn định (các provirut không bao giờ bị cắt rời ra như prophage). Đối với các virut gây bệnh ung thư là ADN, sự gắn vào là quá trình trực tiếp. Các retrovirut thì khác, trước hết chúng sử dụng reverse transcriptaz để từ khuôn ARN tạo ra ADN. Sau đó ADN của virut có thể được gắn vào ADN bộ gen của tế bào chủ và chúng cũng sao chép với bộ gen này trong mỗi thế hệ.

Hình 26. Chu trình sinh sản của retrovirut HIV gây bệnh AIDS
5.3. Vi nấm
Hình 27: Nấm men S.cerevisiae dưới kính hiển vi điện tử và kính hiển vi quang học
Nấm men có thể hoàn thành một chu trình sao chép và phân chia khoảng 1,4 giờ. Xuất phát điểm cho sao chép DNA của nấm men gồm nhiều điểm giống với oriC của E. coli. Đó là vùng giàu AT được tách ra khi có một protein khởi động gắn vào điểm kề bên. Các điểm xuất phát sao chép dài hàng ngàn đến hàng chục ngàn nucleotide. Khác với prokaryote, mỗi nhiễm sắc thể eukaryote có nhiều điểm xuất phát sao chép để quá trình sao chép genome của eukaryote có kích thước lớn hơn nhiều xảy ra một cách nhanh chóng. Có khoảng 400 điểm xuất phát sao chép phân bố trên toàn bộ 16 nhiễm sắc thể của nấm men. Sự sao chép xảy ra trực tiếp trên nhiều điểm xuất phát sao chép. Sợi mạch kép được tạo thành ở mỗi điểm xuất phát sao chép kéo dài và nối với sợi kép được tạo thành ở một điểm khác. Khi sao chép trên 2 mạch đơn hoàn thành sẽ tạo ra 2 phân tử DNA con giống hệt nhau.
Sự tổng hợp DNA chỉ xảy ra ở một giai đoạn trong chu trình tế bào, đó là pha S. Ở nấm nấm men có 3 protein cần cho lắp ráp của replisome. Đó là phức hợp DNA polymerase phối hợp hoạt động ở chẽ ba sao chép. Phức hợp nhận biết điểm xuất phát (Origin recognition complex – ORC) sẽ gắn vào trình tự xuất phát của nấm men như DnaA protein của E. coli. Những phức tương tự được tìm thấy ở tất cả các eukaryote. Sự có mặt của ORC làm bổ sung thêm 2 protein khác, Cdc6 và Cdt. Cả 2 protein này và ORC làm kích hoạt helicase và những thành phần khác của replisome. Sự gắn vào của helicase được xem là sự “xác nhận” (license) cho điểm xuất phát, giúp lắp ráp replisome và bắt đầu tổng hợp DNA.
Trong số các gene mã hoá protein, intron chỉ có khoảng 4-5% tất cả các gene của nấm men (276 gene chứa một intron đơn và 7 gene chứa 2 intron riêng rẽ), kích thước trung bình của intron khoảng 2000 nucleotide. Tần số thấp của intron ở nấm men có lẽ liên quan với tần số thấp của các gene giả (pseudogene) giống như intron, phổ biến ở eukaryote bậc cao. Gene giả chứa trình tự mã hoá, nhưng vì thiếu intron và promoter phiên mã nên trình tự mã hoá không được biểu hiện.
Gene mã hoá protein không chỉ là những dạng gene chức năng. Genome của nấm men chứa số lớn gen tạo ra RNA không mã hoá, bao gồm các gene lặp lại mã hoá cho rRNA, 274 gene của tRNA, trong đó có 80 gene chứa nhóm intron đặc biệt, RNA nhân nhỏ (snRNA) tham gia chức năng chế biến rRNA, 5 snRNA tham gia chế biến intron, một vài RNA chưa biết chức năng và 3 RNA như là các tiểu đơn vị chức năng của enzyme Rnase, endoribonuclease và telomere.
Chu trình sống của các loài nấm khác nhau rất đa dạng, nhưng có thể xếp vào 5 kiểu căn bản khác nhau như: 1. Sinh sản vô tính; 2. Thể đơn bội; 3. Thể đơn bội với giai đoạn dị nhân; 4. Đơn bội - Lưỡng bội ; 5. Lưỡng bội.
Ngay trong các nấm sinh sản hữu tính cũng có sự dao động từ hoàn toàn đơn bội như Neurospora đến hoàn toàn lưỡng bội như ở Saccharomyces cerevisiae

Đối với phần lớn nấm sợi, điểm đặc trưng là nhiều nhân cùng hiện diện trong một tế bào. Các nhân này có thể có kiểu gen khác nhau. Trong trường hợp này được gọi là thể dị nhân (heterokaryon) tương tự dị hợp tử.
Ở các vi sinh vật nói chung, các tính trạng hình thái thường không nhiều, nhưng có sự đa dạng lớn về nhiều loại phản ứng hóa học khác nhau. Sự ra đời của giả thuyết nổi tiếng 1 gen - 1 enzym đã dựa trên các đột biến sinh hóa ở N. crassa. Các môi trường chọn lọc được sử dụng để thu thập các dạng lai hiếm hoi giữa các dòng cha mẹ đơn bội mang các đột biến khuyết dưỡng lặn, khi sai hỏng trong chuỗi phản ứng sinh hóa của chúng bổ sungcho nhau. Ngoài ra tính đề kháng với các tác nhân bất lợi được sử dụng rộng rãi làm tính trạng đánh dấu trong các nghiên cứu di truyền ở nấm.
6. Điều hoà biểu hiện gene ở vi khuẩn
6.1.
Mô hình Operon
446

Phần lớn các gene trong bộ gen vi khuẩn được tổ chức thành các đơn vị hoạt động chức năng đặc trưng, gọi là các operon. Các gene cấu trúc trong một operon được điều hoà chung trong quá trình chuyển hoá một hợp chất nhất định của tế bào.
Lần đầu tiên vào năm 1961, Francois Jacob và Jacques Monod (France) đề xuất giả thuyết operon để giải thích sự điều hoà quá trình sinh tổng hợp protein ở vi khuẩncác enzyme tham gia vào con đường hấp thụ và phân giải đường lactose - operon lactose (lac operon). Đây là operon được nghiên cứu kỹ nhất cho đến nay (Hình 24).
Operon là đơn vị tổ chức và hoạt động gene đặc trưng của các bộ gene prokaryote; nó là một phức hợp liên kết chặt chẽ giữa vùng khởi động (promoter) cùng với yếu tố chỉ huy (operator) và nhóm gene cấu trúc do yếu tố này trực tiếp kiểm soát. (Vì vậy nó được gọi là operon).
Tham gia vào điều hòa hoạt động của một operon gồm có bốn yếu tố thuộc hai thành phần chính: (i) các locus cấu trúc (structural loci) và (ii) các locus điều hòa (regulatory loci); trong đó nhóm sau bao gồm yếu tố chỉ huy (operator), vùng khởi động (promoter) và gene điều hòa (regulator gene).
- Một yếu tố chỉ huy (operator = O): trình tự DNA nằm kế trước nhóm gene cấu trúc, là vị trí tương tác với chất ức chế
- Một vùng khởi động (promotor region = P): trình tự DNA nằm trước yếu tố chỉ huy và có thể trùm lên một phần hoặc toàn bộ vùng này, là vị trí bám vào của RNA polymerase để có thể khởi đầu phiên mã tại vị trí chính xác ở sợi khuôn.
- Một gene điều hoà hay còn gọi là gene ức chế (regulatory/ inhibitory gene = R/I): Gene này sinh ra loại protein điều hoà gọi là chất ức chế (repressor) điều hòa hoạt động của nhóm gene cấu trúc thông qua sự tương tác với yếu tố chỉ huy. Mặc dù mỗi gene điều hòa có một vùng khởi động riêng và không có yếu tố chỉ huy, đôi khi người ta vẫn coi chúng là một operon điều hòa; gene này sinh ra chất ức chế một cách ổn định.
6.2. Điều hoà âm tính của các operon cảm ứng: lac operon
Đại diện cho tất cả các operon của các loại đường di- và polysaccharide (mà vi khuẩn sử dụng như một nguồn cung cấp các hợp chất carbon và năng lượng) là operon lactose ở E. coli.
Operon lactose có chức năng sản sinh các enzyme tham gia vào quá trình hấp thụ và phân giải đường lactose (một disacharide) thành galactose và glucose. Nó chỉ hoạt động khi có mặt đường lactose, vì vậy lactose được gọi là chất cảm ứng và lac operon được gọi là operon cảm ứng (inducible) hay operon dị hoá (catabolite). Nói đúng ra, chất cảm ứng là allolactose; lactose (liên kết galactosid dạng β-1,4) bị biến đổi thành chất trung gian trong quá trình thuỷ phân lactose dưới tác dụng của βgalactosidase, gọi là allolactose (liên kết β-1,6).
Cấu trúc của lac operon Các thành phần của lac operon ở E. coli như :

Hình 28. (a) Nhiễm sắc thể E. coli với vị trí tương đối của các operon khác nhau. (b) Cấu trúc phân tử đường lactose; và (c) mô hình operon lactose và chức năng của nó - sản sinh các enzyme hấp thụ và phân giải đường lactose.

- Một nhóm các gene cấu trúc (structural genes) liên quan với nhau về mặt chức năng xếp cạnh nhau, khi phiên mã sẽ tạo ra một phân tử mRNA chung gọi là mRNA đa cistron (polycistronic mRNA). Các enzyme được dịch mã từ một mRNA này sẽ tham gia vào quá trình chuyển hoá (đồng hoá hoặc dị hoá) cụ thể, một chuỗi các phản ứng sinh hoá gồm nhiều khâu nối tiếp hoặc có quan hệ dạng lưới phức tạp. 448
Hình 29: Cấu trúc chi tiết các vùng khác nhau của operon lactose - Nhóm các gene cấu trúc bao gồm ba gene: lacZ, lacY và lacA (nói gọn là Z, Y và A); trong đó lacZ mã hoá cho β galactosidase (thuỷ phân lactose), lacY mã hoá cho permease (vận chuyển lactose qua màng) và lacA mã hoá transacetylase (chức năng không rõ ràng; theo ý nghĩa nó không phải là enzyme liên quan trực tiếp đến sự chuyển hoá lactose).
447
- Yếu tố chỉ huy (lac operator) là trình tự DNA dài ~34 cặp base cách 80 gene



Z chừng 10 cặp base về phía trước, là vị trí tương tác với chất ức chế. Nó chứa trình tự 24 cặp base đối xứng xuôi ngược, giúp chất ức chế (lac repressor) có thể nhận biết và bám vào bằng cách khuếch tán dọc theo DNA từ cả hai phía.
- Vùng khởi động (lac promotor) là đoạn DNA dài chừng 90 cặp base nằm trước và trùm lên lac operator 7 cặp base. Nó chứa hai vị trí tương tác với RNA polymerase và với protein hoạt hoá dị hoá (catabolite activator protein = CAP, hoặc CRP - xem mục V). Điểm khởi đầu phiên mã là vị trí gần cuối của lac promoter.Gene điều hoà (regulatory gene) nằm trước vùng khởi động, mã hoá một protein ức chế gồm bốn polypeptide giống nhau, gọi là tứ phân (tetramer), đều chứa 360 amino acid.
Cơ chế điều hoà âm tính của lac operon
Khi trong môi trường nuôi cấy E. coli vắng mặt lactose (allolactose, chất cảm ứng) thì lac operon không hoạt động, nghĩa là các enzyme tham gia hấp thụ và phân giải lactose không được sinh ra. Nguyên nhân là do chất ức chế của operon (lac repressor) vốn tự thân có hoạt tính, bám chặt vào yếu tố chỉ huy (lac operator) và gây kìm hãm sự phiên mã của các gene cấu trúc Z, Y và A (Hình 26). Do đó các sản phẩm enzyme của lac operon không được tạo ra; tức biểu hiện âm tính.
Hình 31: Chất cảm ứng kết hợp với chất ức chế và làm biến đổi hình dáng của nó; chất ức chế vì vậy không bám được vào lac operator. Kết quả là các gene cấu trúc của lac operon được phiên mã tạo ra phân tử mRNA polycistron và các enzyme tương ứng được tổng hợp.
Phương thức điều hoà như thế được gọi là điều hoà cảm ứng - âm tính, bởi vì chất ức chế lac operon một khi bám vào lac operator sẽ kìm hãm phiên mã, nghĩa là gây hiệu quả âm tính lên sự biểu hiện của các gene; và hoạt động chức năng của protein này lại phụ thuộc vào chất cảm ứng. Nhờ cơ chế điều hoà kiểu liên hệ ngược này mà vi khuẩn có thể thích ứng để tồn tại và phát triển một cách hợp lý.
Hình 30: (a) Chất ức chế bám chặt lac operator gây ức chế phiên mã; (b) Mô hình lac operator (rìa trái) bị bám chặt bởi protein ức chế (rìa phải).
Ngược lại, nếu bổ sung lactose vào môi trường thì một thời gian sau vi khuẩn sẽ bắt đầu hấp thụ và phân giải nó, nghĩa là các enzyme liên quan đã được sinh ra. Sự kiện này được lý giải như sau: Chất cảm ứng (inducer), ở đây là allolactose - dạng biến đổi của lactose - tương tác với chất ức chế (repressor) làm biến đổi cấu hình của chất này. Một phân tử allolactose bám vào một tiểu đơn vị của chất ức chế. Vì vậy chất ức chế mất ái lực và không thể bám vào lac operator; nó tách ra khỏi DNA. Lúc này các gene cấu trúc được phiên mã và các enzyme tương ứng được tổng hợp, nhờ vậy vi khuẩn có thể hấp thụ và phân giải đường lactose (Hình 27). Lactose vì vậy là tác nhân gây cảm ứng (hoạt hoá) lac operon. Ngoài ra, ITPG (isopropyl thiogalactoside) cũng được dùng như một chất cảm ứng nhưng không phải là tác nhân sinh lý.
449
Hình 32: Chất ức chế một khi được bám đầy đủ bởi allolactose thì tách khỏi operator khiến cho sự điều hoà âm tính (sự ức chế) được làm dịu bớt, tuy nhiên RNA polymerase vẫn chưa thể tạo thành một phức hợp bền vững với promoter để có thể khởi đầu phiên mã được.
Về cơ bản, cơ chế "mở" của lac operon được trình bày như trên; nhưng thực ra sự hoạt động của chất cảm ứng mới chỉ làm dịu bớt (alleviation) sự điều hoà âm tính (sự ức chế) của lac operon. Hình 32 cho thấy ngay cả khi chất ức chế đã tách khỏi operator, RNA polymerase vẫn không thể bám ổn định vào promoter và khởi đầu phiên mã - nó không có ái lực đủ cao đối với promoter để bám vào đủ lâu để có thể khởi đầu tạo thành liên kết phosphodiester đầu tiên.
6.3. Cơ chế điều hoà âm tính của trp operon
Khi trong tế bào E. coli dư thừa amino acid tryptophan (sản phẩm cuối cùng của con đường chuyển hoá) thì trp operon ngừng hoạt động và do đó các enzyme tương ứng không được sinh ra. Sự kiện này được giải thích như sau: Chất ức chế bình thường của operon này (trp repressor) tồn tại ở dạng bất hoạt gọi là aporepressor, không có ái lực đối với trp operator, nhưng khi các amio acid dư thừa kết hợp vào sẽ tạo ra phức hợp có hoạt tính, nghĩa là có ái lực với trp operator. Vì thế tryptophan được gọi là chất đồng ức chế (corepressor). Phức hợp này bám vào yếu tố chỉ huy làm kìm hãm phiên mã của trp operon. Ngược lại, khi trong tế bào vắng mặt hay thiếu hụt các amino acid này, tự thân chất ức chế này ở trạng thái bất hoạt nên không bám được vào yếu tố chỉ huy (trp operator). Vì vậy các gene cấu trúc xảy ra sự phiên mã và kết quả là các enzyme tham gia tổng hợp tryptophan được sinh ra. Và một khi hàm lượng amio acid này được tổng hợp ở mức dư thừa sẽ tác động ngược trở lại, kìm hãm hoạt động của trp operon.
NADH dehydrogenase); S-adenosyl methionine (nhường nhóm methyl cho nhiều phân tử, kể cả DNA và "chóp" ở đầu 5' của mRNA eukaryote).
Trong mỗi trường hợp, riboswitch kiểm soát các gene liên quan đến sự chuyển hóa của phân tử đó. Chất chuyển hóa bám vào mRNA đang sinh trưởng và bao gồm cả sự thay đổi biến cấu (allosteric) mà: (i) đối với một số gene nó khiến cho sự tổng hợp mRNA hơn nữa để kết thúc trước khi hình thành một sản phẩm chức năng, và (ii) đối với một số gene khác, nó tăng cường hoàn chỉnh việc tổng hợp mRNA. Trong cả hai trường hợp, kết quả là do sự kiểm soát mức độ của chính chất chuyển hóa ấy. Một số riboswitch kiểm soát sự dịch mã mRNA hơn là phiên mã mRNA. Đã có gợi ý rằng các cơ chế điều hoà này, vốn không liên quan bất kỳ protein nào; nó là một cơ chế còn sót lại từ "giới RNA".
6.4. Sự ức chế dị hoá (Catabolite repression): Điều hoà dương tính của lac operon
Như chúng ta có thể dự đoán, điều hoà dương tính (positive control) là đối lập với điều hoà âm tính; đó là, operon bị đóng, ngưng hoạt động (thực ra là hoạt động của operon bị giảm xuống một mức cơ sở) trừ phi có yếu tố nào đó xen vào bật nó hoạt động trở lại. Trong trường hợp của lac operon, điều này có nghĩa là tách bỏ chất ức chế ra khỏi operator là chưa đủ để kích hoạt operon như đã đề cập trước đây. Nó cần đến một nhân tố dương tính bổ sung thêm.
Hình 33: Operon tryptophan bị kìm hãm (a) và hoạt động (b).

Tóm lại, phương thức điều hòa hoạt động gene theo các cơ chế liên hệ ngược hay phản hồi như thế (feed-back mechanisms) đảm bảo cho bộ gene các vi khuẩn hoạt động một cách hợp lý và nhờ đó các vi khuẩn thích ứng và phát triển trước các điều kiện môi trường luôn thay đổi.
Cần lưu ý rằng, các protein ức chế và đồng ức chế không hẳn là cách duy nhất các vi khuẩn điều hòa phiên mã gene. Ở nhiều vi khuẩn (và một số eukaryote), sự điều hòa mức chuyển hóa nào đó cũng có thể được kiểm soát bởi các riboswitch (tạm dịch là: công tắc ribo). Một riboswitch là một phần của vùng 5'-không được dịch mã (5'-untranslated region = 5'- UTR) trong phân tử mRNA, tại đó có một vị trí bám đặc thù cho chất chuyển hóa (metabolite).
Một số chất chuyển hóa bám vào các riboswitch, như: các purine adenine và guanine; các amino acid glycine và lysine; mononucleotide flavin (nhóm phụ của
Thật vậy, hoạt động của lac operon còn chịu sự kiểm soát của một protein điều hoà dương tính liên quan với sự có mặt của glucose. Cụ thể, khi trong môi trường có mặt đồng thời cả lactose và glucose thì operon lac tạm thời ngưng hoạt động. Hiện tượng này gọi là ức chế dị hoá (catabolite repression). Người ta nhận thấy rằng, khi glucose có mặt ở nồng độ cao thì hàm lượng AMP vòng (3',5'-cyclic adenosine monophosphate = cAMP; Hình 34.a) trong tế bào rất thấp; và ngược lại, khi không có glucose hoặc có không đáng kể thì hàm lượng cAMP trong tế bào được tổng hợp tăng cao. cAMP vì vậy được xem là chất chỉ thị (indicator) của sự vắng mặt glucose và được coi là nhân tố điều hoà dương tính (positive regulator) của các operon dị hoá.
452
Hình 34 (a) Cấu trúc và sự hình thành phân tử cAMP từ ATP. (b) CAP gồm hai monomer giống nhau, mỗi monomer nhận biết một trình tự DNA nhờ vùng xoắn alpha được đánh dấu F; và (c) Trình tự đối xứng của vị trí CAP là các đoạn lặp đảo ngược.

Ngoài ra, còn phát hiện một loại protein điều hoà dương tính có tên là protein hoạt hoá dị hoá (catabolite activator protein = CAP) cũng gọi là protein tiếp nhận / bám cAMP (cyclic AMP receptor / binding protein = CRP) hay protein bám cAMP.
CAP gồm hai tiểu đơn vị giống nhau gọi là homodimer (Hình 34.b); nó chỉ hoạt động khi môi trường nội bào có hàm lượng cAMP cao. Trong trường hợp đó, cAMP kết hợp với CAP tạo thành phức hợp CAP-cAMP và làm tăng ái lực của CAP đối với promoter. Phức hợp này có khả năng nhận biết và bám vào một đoạn 16 bp về phía trước vùng khởi động, với các đoạn lặp đảo ngược (inverted repeats), gọi là vị trí CAP (Hình 34.c). Bằng cách đó RNA polymerase được kích thích bám chặt vào promoter và bắt đầu tổng hơp mRNA ở mức cao.
Như vậy, khác với kiểu điều hoà âm tính do sự tương tác giữa ''chất ức chế và operator'' (tương tác protein-DNA) ở đây sự tương tác xảy ra giữa protein điều hoà thuộc phức hợp CAP-cAMP mà yếu tố chính là CAP với RNA polymerase (tương tác protein-protein) giúp RNA polymersae bám ổn định vào promoter, tăng cường hoạt động phiên mã (điều hoà dương tính) mà chủ yếu là điều chỉnh tốc độ khởi đầu phiên mã. Mặt khác khi CAP/cAMP bám vào, nó làm cho DNA uốn gập đáng kể (khoảng 90 độ). Và như thế, RNA polymerase dễ dàng tách hai sợi của DNA, tạo thành một phức hợp mở
Hình 35: Điều hoà dương tính lac operon
CAP/cAMP cũng có thể kích thích phiên mã các operon cảm ứng khác, bao gồm các ara và gal operon được nghiên cứu kỹ. Ngay khi lac operon tiến hành chuyển hoá lactose, các operon khác này mã hoá cho các enzyme phân cắt các đường biến đổi tương ứng, arabinose và galactose. Như thế, để cho hiệu suất lớn nhất, tất cả ba operon này sẽ vẫn đóng chừng nào tế bào vẫn còn có sẵn glucose. Vì cAMP đáp ứng với nồng độ glucose, nên ta chẳng ngạc nhiên gì khi cả ba operon này cùng chia xẻ một cơ chế điều hoà chung có liên quan cAMP. Phức hợp CAP-cAMP bám ở promoter hoặc gần promoter của mỗi operon và tạo điều kiện thuận lợi cho việc bám vào của RNA polymerase.
6.5. Sự kết thúc phiên mã sớm (Attenuation) ở trp operon

Attenuation (phiên mã dở) là một cơ chế điều hoà gây ra sự kết thúc phiên mã sớm dưới những điều kiện nhất định, bằng cách đó ngăn cản sự biểu hiện của mRNA cần cho sự biểu hiện của các sản phẩm gene tương ứng. Phiên mã dở tạo thành mRNA uốn gập một cách điển hình thành các cấu trúc bậc hai xen kẻ (alternative secondary structures), mà một trong số đó là nhân tố kết thúc độc lập ρ (Rho-independent terminator).
Một cách tiếp cận tin sinh học đã được phát triển để xác định các gene được điều hoà theo kiểu phiên mã dở (Một số bài báo tổng quan hay về phiên mã dở như: Gollnick và Babitzke 2002; Henkin và Yanofsky 2002.)
Operon tryptophan chẳng hạn còn có một kiểu điều hòa phiên mã dở. Nó sử dụng dịch mã để điều khiển sự phiên mã. Khi có mặt tryptophan trong môi trường nội
bào, thậm chí ở nồng độ thấp, sẽ xảy ra sự dịch mã một phần ở vùng leader của mRNA đang được tổng hợp. Kết quả là làm dừng sự phiên mã trước khi gene cấu trúc đầu tiên (trpE) của operon được phiên mã.
vùng 4 và tạo ra điểm kết thúc phiên mã sớm (xảy ra sau khi tổng hợp xong 8 uridine ở ngay sau vùng 4). Khi số lượng tRNATrp đưa vào không đầy đủ, sự dịch mã đoạn dẫn đầu dừng lại đột ngột ở các codon Trp của nó. Điều này ngăn cản ribosome tiến vào vùng 2, do đó vùng này sẽ cặp với vùng 3 gây cản trở việc tạo thành cấu trúc phiên mã dở (trp attenuator). Kết quả là phân tử mRNA đa cistron của operon tryptophan được tạo thành một cách đầy đủ.
Hình 36: Cấu trúc của đoạn dẫn đầu - TrpL (a) và vùng kết thúc phiên mã sớm - trp attenuator với đuôi 3' gồm 8 uridine (b).
Sự kết thúc phiên mã sớm ở operon tryptophan là kết quả của sự tương tác bổ sung nội phân tử giữa các trình tự DNA bên trong vùng leader của bản sao RNA. Kết quả của sự kết thúc phiên mã sớm này tạo ra một mRNA chứa 140 base (hình 36a).
Tại vùng đầu mút 3' của nó xảy ra sự tự bổ sung ở đoạn giàu GC tạo thành một cấu trúc hình vòng trên thân RNA và gây ra sự kết thúc phiên mã sớm. Vùng này được gọi là đoạn phiên mã dở của operon tryptophan (trp attenuator) và ở phần đuôi của mRNA này cũng có 8 base uridine (hình 36.B). Kiểu cấu trúc "kẹp cài tóc" này là tín hiệu kiểm soát kết thúc phiên mã ở prokaryote nói chung.
Với kiểu cấu trúc đặc thù ở đoạn dẫn đầu của trp operon như vậy làm cho nó có
ý nghĩa quan trọng trong điều hoà phiên mã dở, ở chỗ:
(i) tổng hợp một peptide dẫn đầu chứa 14 amino acid;
(ii) trên mRNA của đoạn peptide này chứa hai codon của Trp ở các vị trí 10 và 11;

(iii) ở bốn vùng được đánh số 1-4 xảy ra sự tự bổ sung giữa các vùng 1 và 2 và giữa 3 và 4; và ở một số trường hợp có thể xảy ra sự kết cặp giữa các vùng 2 và 3.
Do trong trình tự mã hóa của trình tự dẫn đầu trpL có hai codon Trp, nên sự dịch mã đoạn này tỏ ra nhạy cảm với số lượng tRNAtrp đưa vào. Nếu môi trường cung
cấp đầy đủ Trp, ribosome trượt qua các codon Trp để đi vào vùng 2. Và sự có mặt của ribosome ở vùng 2 ngăn cản vùng này kết cặp với vùng 3. Khi đó vùng 3 sẽ cặp với
455
B. CÂU HỎI VÀ BÀI TẬP
Câu 1:
Hệ gene của virus HPV-16 (human papilloma virus), một trong những loại virus gây ung thư cổ tử cung phổ biến ở người, chứa gene mã hoá 2 protein ung thư (E6 và E7) làm biến đổi tế bào biểu mô bình thường thành tế bào khối u. Người ta tiến hành các thí nghiệm để phân tích tác động của protein E7 đến các tế bào ở cấp độ phân tử, cụ thể là mối quan hệ của nó với protein retinoblastoma (Rb), một loại protein áp chế khối u liên kết với yếu tố phiên mã E2F điều hoà sự phiên mã của các gene mã hóa protein tham gia pha S của chu kỳ tế bào.
Ở thí nghiệm thứ nhất (Hình 2.1), dòng tế bào ung thư bạch cầu (HL-60) được nuôi cấy trong điều kiện không có (dấu ) hoặc có (dấu +) xử lý với phorbol ester và được đánh dấu bằng 35S-methionine hoặc 32P phosphate. Dịch chiết tế bào được dùng để điện di trên gel SDS-polyacrylamide và xử lý với kháng thể đặc hiệu của protein Rb. Kết quả được ghi lại bằng phóng xạ tự ghi.
Ở thí nghiệm tiếp theo, các tế bào HL-60 được nuôi cấy trong điều kiện không có hoặc có phorbol ester. Dịch chiết tế bào được ủ với các hạt agarose có khả năng liên kết cộng hoá trị với các phân tử E7. Sau khi ủ, phần cặn lắng và dịch nổi được thu lại bằng ly tâm. Kết quả điện di trên gel SDS-polyacrylamide và lai Western sử dụng kháng thể đặc hiệu của Rb với các mẫu dịch chiết tế bào trước khi ủ, dịch nổi và cặn lắng sau khi ủ với hạt agarose được biểu thị trên Hình 2.2 (dấu : không có; dấu +: có các thành phần tương ứng).
c. - Cơ chế tác dụng của protein E7 là liên kết với protein Rb ở dạng khử phosphoryl hoá, giải phóng yếu tố phiên mã E2F dẫn đến sự phiên mã các gen liên quan đến pha S và gây tăng sinh tế bào HL-60.
- Vì khi không xử lý với phorbol ester (mẫu 7), không phát hiện thấy RB ở cặn lắng chứng tỏ E7 liên kết với hạt agarose không tương tác với Rb ở dạng phosphoryl hoá. Khi xử lý với phorbol ester (mẫu 10), Rb xuất hiện ở cặn lắng chứng tỏ có sự liên kết với E7 khi ở dạng khử phosphoryl hoá. Do đó, yếu tố phiên mã E2F được giải phóng và hoạt hóa sự phiên mã các gene liên quan đến pha S.
Câu 2:
Thành tế bào của nhiều vi khuẩn được bao phủ bởi một lớp màng nhầy có thành phần polysaccharide hoặc protein. Người ta tiến hành nghiên cứu sự tổng hợp hyaluronan, một loại polysaccharide có thể tạo màng nhầy, gồm 2 loại đơn phân Nacetylglucosamine (GlcNAc) và acid glucuronic (GlcUA) xếp xen kẽ nhau, dưới tác dụng của enzyme hyaluronan synthase trên màng tế bào.
Hình2.2
Dựa trên các kết quả thí nghiệm, hãy trả lời các câu hỏi sau:
a) Protein Rb hoạt động trong tế bào bình thường ở dạng phosphoryl hoá hay khử phosphoryl hoá? Giải thích.
b) Phorbol ester có tác động gì đến protein Rb? Giải thích.
c) Cơ chế tác dụng của protein E7 đến sự tăng sinh của tế bào HL-60 như
thế nào? Giải thích.
Hướng dẫn
a. Protein Rb hoạt động ở dạng khử phosphoryl hoá. Vì ở thí nghiệm thứ nhất, trong tế bào HL-60 không xử lý phorbol ester, protein Rb ở dạng phosphoryl hoá thể hiện ở băng vạch có khối lượng phân tử cao hơn (Mr cao) khi đánh dấu bằng 35S methionine và 32P phosphate (mẫu 1 và 3). Đây là các tế bào ung thư phân chia nhiều lần có protein Rb ở dạng bất hoạt.

b. Phorbol ester kích thích sự khử phosphoryl hoá protein Rb. Vì khi xử lý với phosbol ester, protein Rb có khối lượng phân tử thấp hơn (Mr thấp) trong thí nghiệm đánh dấu bằng 35S methionine (mẫu 2) và không phát hiện thấy khi đánh dấu bằng 32P phosphate (mẫu 4).
Hình Hình
Hyaluronan synthase được trộn với cơ chất ban đầu - một tetrasaccharide có đánh dấu phóng xạ (Hình 3.1) trong các điều kiện: không bổ sung, có bổ sung riêng lẻ, bổ sung hỗn hợp các phân tử đường ở dạng hoạt hóa cần thiết cho sự tổng hợp (GlcNAc, GlcUA).
Sau thời gian ủ, người ta tiến hành sắc ký bản mỏng sản phẩm thu được ở các điều kiện thí
nghiệm (Hình 3.2, các chữ số bên trái thể
Hình2.1 458
hiện số gốc đường trong phân tử carbohydrate được xác định dựa trên chất chuẩn).
Hình Hình
a) Dựa vào dữ liệu đã cho, hãy cho biết hyaluronan synthase xúc tác
việc gắn thêm các gốc đường ở dạng monosaccharide hay disaccharide? Giải thích.
b) Các gốc đường được gắn vào đầu khử hay đầu không khử của tetrasaccharide ban đầu? Giải thích.
Hyaluronan synthase được trộn với cơ chất ban đầu - một tetrasaccharide có
đánh dấu phóng xạ (Hình 3.1) trong các điều kiện: không bổ sung, có bổ sung riêng lẻ, bổ sung hỗn hợp các phân tử đường ở dạng hoạt hóa cần thiết cho sự tổng hợp (GlcNAc, GlcUA). Sau thời gian ủ, người ta tiến hành sắc ký bản mỏng sản phẩm thu được ở các điều kiện thí nghiệm (Hình 3.2, các chữ số bên trái thể hiện số gốc đường trong phân tử carbohydrate được xác định dựa trên chất chuẩn).
c) Dựa vào dữ liệu đã cho, hãy cho biết hyaluronan synthase xúc tác
việc gắn thêm các gốc đường ở dạng monosaccharide hay disaccharide? Giải thích.
d) Các gốc đường được gắn vào đầu khử hay đầu không khử của tetrasaccharide ban đầu? Giải thích.
e) Dựa vào tốc độ gắn từng loại đường, giải thích tại sao chỉ quan sát
thấy trên bản sắc ký các sản phẩm tổng hợp có số gốc đường là số lẻ
f) Khi nghiên cứu về màng nhầy của vi khuẩn cố định đạm sống tự do trong đất Azotobacter vinelandii ở các điều kiện sống khác nhau, người ta đo
được độ dày trung bình của lớp màng nhầy vi khuẩn trong điều kiện X là 0,01
µm và trong điều kiện Y là 0,05 µm. Khả năng cố định đạm của vi khuẩn A.
vinelandii cao hơn ở điều kiện X hay Y? Giải thích.
Hướng dẫn
a. Enzyme hyaluronan synthase xúc tác gắn các monosaccharide vào tetrasaccharide ban đầu. Vì: khi bổ sung riêng phân tử GlcNAc (làn 3), enzyme đã xúc tác tạo thành phân tử có 5 gốc đường.
b. Các gốc đường được gắn vào đầu không khử của tetrasaccharide ban đầu. Vì: khi bổ sung riêng phân tử GlcUA (làn 4), sự tổng hợp không xảy ra còn khi bổ sung phân tử GlcNAc sự tổng hợp xảy ra. Tetrasaccharide ban đầu có các gốc đường liên kết xen kẽ nhau. Do đó, GlcNAc phải được gắn vào đầu có gốc GlcUA (đầu không khử) của tetrasaccharide ban đầu.
c. Do tốc độ gắn các gốc GlcNAc nhanh hơn nhiều so với gốc GlcUA nên các sản phẩm trung gian có gốc GlcNAc ở đầu không khử (diễn ra sự tổng hợp) sẽ tích lũy do đó các sản phẩm này có số gốc đường là số lẻ
d. Điều kiện Y. Vì vi khuẩn A. vinelandii sống tự do trong đất nên khi chúng cố định đạm bằng phức hệ nitrogenase trong tế bào chất thì O2 phải được giữ một phần bên ngoài tế bào nhờ lớp màng nhày để không ức chế phức hệ nitrogenase.
Câu 3:
459
Một nhà nghiên cứu có 10 chủng vi khuẩn E.coli mang các đột biến khác nhau nằm trên operon lac của các vi khuẩn này. Khi phân tích ADN của vi khuẩn, nhà nghiên cứu đã thấy mỗi chủng mang 1 trong 5 đột biến sau: lacZ, lacY, lacI, lacIS hoặc lacOC. Nhà nghiên cứu này cũng biết chắc chắn rằng chủng 6 là chủng đột biến gen lacZ. Sau đó nhà nghiên cứu này đã phân lập được đoạn ADN mang operon lac từ mỗi chủng (gọi là chủng cho) và biến nạp đoạn ADN này vào các chủng khác, tạo ra chủng lưỡng bội từng phần (gọi là chủng nhận). Các chủng nhận được nuôi trên môi trường tối thiểu chứa lactose là nguồn cung cấp carbon duy nhất. Sự sinh trưởng của các chủng được ghi lại trong hình dưới (+ biểu thị chủng đang sinh trưởng, - biểu thị chủng không sinh trưởng được).
Nếu quy ước sản lượng enzyme β-galactosidase và permase của kiểu hình kiểu dại sản xuất ra trong điều kiện có lactose, thiếu glucose đạt giá trị 100; ở điều kiện có đồng thời cả glucose và lactose đạt giá trị 10.
Hãy xác định giá trị sản lượng tương đương của các enzym ở mỗi dòng lưỡng bội ở bảng trên ở trong 4 điều kiện môi trường của operon Lac:
a) Chỉ có lactose;
b) Chỉ có glucose;
c) Không có cả lactose và glucoe;
d) Có cả glucose và lactose.
460
Hướng dẫn
- Chúng ta nhận thấy, chủng 1 và 8 ức chế tất cả các chủng khác, trừ chủng 5 và 9; chủng 6 và 7 là đột biến xảy ra trên cùng 1 gen; chủng 2 và 4 xảy ra trên cùng 1 gen, nhưng khi kết hợp với nhau thì vẫn biểu hiện kiểu hình kiểu dại; chủng 3 và 10 là các
đột biến xảy tra trên cùng 1 gen.
Vậy, từ dữ kiện của bài ra, ta kết luận: chủng 1 và 8 là đột biến lacIS, chủng 3 và 10 là đột biến lacY; chủng 5 và 9 là đột biến lacOC, chủng 2 và 4 là lacI-, chủng 6 và 7 là lacZ.
- Các dòng lưỡng bội của 5 chủng đột biến trên được tạo thành là: 1-1; 1-2; 1-3;
2-5; 2-6; 3-3; 3-5; 3-6; 5-5; 5-6 và 6-6.
Dòng Chỉ có glucose Chỉ có lactose Có cả lac và glu Không có cả Lac và Glu
4:
Enzim dcm methylase xúc tác phản ứng thêm nhóm methyl vào nucleotit Cytosine thứ 2 trong trình tự CCAGG ở E. coli K-12. Mặt khác, các Cytosine ở vùng trình tự của gen lacI là điểm nóng của đột biến thay thế đồng hoán. Hãy đề xuất 2 phương pháp để có thể khẳng định tần suất đột biến tăng lên ở vị trí này là do methyl hóa cytosine hay không?
Hướng dẫn
- Có 2 nguyên nhân gây đột biến là: do enzym methylase hoặc đột biến thay thế đồng hoán Cytosine thành Timin trong vùng điểm nóng gây đột biến.
- Để xác định nguyên nhân gây đột biến do nguyên nhân nào, ta thực hiện biện pháp knock out các điểm đột biến.
+ Gây đột biến gen dcm tổng hợp dcm methylase. Nếu tần số đột biến giảm, chứng tỏ nguyên nhân đột biến là do dcm methylase. Nếu tần số đột biến vẫn tăng, chứng tỏ nguyên nhân đột biến do đột biến đồng hoán ở vị trí điểm nóng đột biến.
+ Phương pháp thứ 2, loại bỏ nucleotit thứ 2 trong trình tự CCAGG, quan sát tần số đột biến để kết luận nguyên nhân.
Câu 5:
Trong thí nghiệm lai DNA ở 06 loài thực vật thuộc cùng chi Vicia, DNA đã được phân lập từ mỗi loài, gây biến tính bằng nhiệt và phân mảnh thành từ phân đoạn DNA nhỏ rồi tiến hành hồi tính. Ở một thí nghiệm khác, DNA từ vi khuẩn E. coli cũng được phân lập và tiến hành tương tự.
Kết quả của 02 thí nghiệm được thể hiện như đồ thị bên dưới.
461
a. So sánh và giải thích sự khác biệt trong quá trình hồi tính DNA của vi khuẩn E. coli so với các mẫu DNA ở 06 loài thực vật thuộc chi Vicia?
b. Hãy xác định giai
đoạn các trình tự lặp lại và các trình tự duy nhất của DNA ở 06 loài thực vật thuộc chi Vicia.
- Thí nghiệm 1: Nuổi chủng vi khuẩn khuyết dưỡng histidine (his-) trên môi trường dinh dưỡng không bổ sung histidine và có bổ sung B(a)P. Kết quả cho thấy không khác so với mẫu đối chứng, không bổ sung B(a)P.
- Thí nghiệm 2: Tiêm B(a)P vào chuột mỗi tháng một lần. Kết quả cho thấy chuột có xuất hiện khối u ở phổi so với mẫu đối chứng là không tiêm B(a)P.
- Thí nghiệm 3: Ủ B(a)P với dịch chiết của tế bào gan và bổ sung vào môi trường nuôi cấy tương tự ở thí nghiệm 1. Kết quả cho thấy vi khuẩn mọc lên khuẩn lạc nhiều hơn thí nghiệm 1.
a) Dựa vào kết quả thu được của 03 thí nghiệm trên để xác định B(a)P có phải là tác nhân đột biến gen hay không? Vai trò của mẫu đối chứng ở thí nghiệm 1 là gì?
Hướng dẫn a)
- So với DNA của 06 loài thực vật, thì DNA của vi khuẩn E. coli hồi tính nhanh hơn nhiều lần; phần trăm phân đoạn DNA bắt cặp tại từng thời điểm khác nhau gần như tuyến tính.
- Nguyên nhân:
+ DNA của vi khuẩn E. coli có kích thước nhỏ nhìn chung tốc độ hồi tính nhanh; ít vùng trình tự lặp lại quá trình hồi tính diễn ra đơn giản, không chia thành những pha khác nhau (tuyến tính).

+ DNA ở 06 loài thực vật có kích thước lớn nhìn chung tốc độ hồi tính chậm; có nhiều vùng trình tự lặp lại quá trình hồi tính diễn ra phức tạp hơn, có nhiều pha hồi tính nhanh, chậm khác nhau.
b)
- Trong quá trình hồi tính của DNA, các đoạn trình tự lặp lại sẽ được bắt cặp trước do sự tương đồng về trình tự cao hồi tính ở pha sớm; các đoạn trình tự duy nhất thường được bắt cặp chậm hơn hồi tính ở pha muộn.
- Nhìn chung, rõ ràng nhất là giai đoạn 1 giờ đầu tiên ở cả 06 loài đều có sự hồi tính của vùng trình tự lặp lại; các giai đoạn còn lại sẽ có sự hồi tính của vùng trình tự duy nhất.
b) Trong hơn 50 năm qua, tỷ lệ bệnh nhân ung thư phổi ngày càng tăng trong khi tỷ lệ bệnh nhân ung thư dạ dày ngày càng giảm. Hãy đưa ra giải thích cho xu hướng này.
c) Theo thống kê, có hơn 50% trường hợp ung thư vú có thể được trị khỏi nhưng chỉ có khoảng 20% trường hợp ung thư phổi được trị khỏi. Hãy đưa ra giải thích cho sự khác biệt này.
Hướng dẫn
a) - Thí nghiệm 1: Không có sự khác biệt so với mẫu đối chứng B(a)P không phải là tác nhân gây đột biến nên chủng đột biến khuyết dưỡng histidine không phục hồi kiểu hình hoang dại.
- Thí nghiệm 2: Khối u xuất hiện so với mẫu đối chứng B(a)P là tác nhân ung thư (carcinogen) với tế bào chuột.
- Thí nghiệm 3: Khuẩn lạc mọc nhiều hơn ở thí nghiệm 1 chủng đột biến có sự phục hồi đột biến khuyết dưỡng histidine thành kiểu hình hoang dại B(a)P đã được chuyển hóa nhờ dịch chiết của gan tạo thành chất gây đột biến.
Vậy chất B(a)P không tự thân gây đột biến mà cần phải được chuyển hóa thành một chất khác mới có khả năng gây đột biến. Đây được xem là tiền tác nhân gây đột biến (promutagen). Mẫu đối chứng ở thí nghiệm 1 có vai trò chứng tỏ rằng B(a)P không tác động đến sự phục hồi kiểu hình đột biến của vi khuẩn, nếu có xuất hiện khuẩn lạc là do đột biến ngẫu nhiên.
Câu 6: Khói thuốc lá có chứa một chất có tiềm năng gây đột biến là benzo (a) pyrene (viết tắt là B(a)P). Một thí nghiệm đã được tiến hành dựa trên nguyên tắt của Ames như sau: 464
b) Tác động của môi trường ảnh hưởng đến hơn 90% tỷ lệ mắc ung thư ở người. Xu hướng trên thể hiện sự thay đổi mức độ tác động của tác nhân ung thư từ môi trường lên cơ thể người. Mức độ ô nhiễm không khí và xu hướng sử dụng khối thuốc lá liên tục tăng là nguyên nhân chính làm tăng tỷ lệ ung thư phổi; trong khi đó chất lượng thực phẩm ngày càng được
quan tâm, hàm lượng nitrite được loại bỏ khỏi bữa ăn là nguyên nhân chính làm giảm tỷ lệ ung thư dạ dày.
c) Tỷ lệ trị khỏi các loại ung thư khác nhau là khác nhau, điều này ảnh hưởng bởi nhiều yếu tố:
- Ung thư vú dễ phát hiện sớm hơn ung thư phổi nên khả năng xây dựng phát đồ trị liệu sẽ hiệu quả hơn so với ung thư vú. Đa phần, các trường hợp ung thu phổi được phát hiện ở giai đoạn muộn.
- Ung thư vú có thể dễ dàng được loại bỏ dễ dàng và hiệu quả bằng các phương pháp phẫu thuật trong khi ung thư phổi hầu như rất khó để trị bằng phương pháp phẩu thuật mà phải tác động bằng các biện pháp ít đặc hiệu như chiếu xạ, hóa trị,… sẽ để lại nhiều tác dụng phụ.
Câu 7: HNA19-Khi một enzim có mặt ở một loài vi khuẩn, thì con đường chuyển hoá mà enzim đó tham gia thường tồn tại trong loài vi khuẩn này. Bảng 1.1 là tên enzim và phản ứng mà enzim xúc tác được sử dụng làm chỉ thị cho sự xuất hiện của các con đường chuyển hóa mà nó tham gia. Bảng 1.2 thể hiện sự có mặt hay vắng mặt của một số enzim ở bốn loài vi khuẩn khác nhau 1,2, 3 và 4.
Bảng 1.1. Enzim và phản ứng xúc tác tương ứng
Tên enzim Phản ứng xúc tác
Lactat đêhiđrôgenaza (LDH) Axit piruvic + NADH → axit lactic 4NAD+
Alcohol đêhiđrôgenaza (ADH) Axêtanđêhit 4- NADH → Êtanol + NAD+
Xitôcrôm C oxidaza
Vận chuyển electron từ xitôcrôm C tới xitôcrôm a
ATP sintetaza Vận chuyển H+ qua màng tạo ATP từ ADP và Pi
Phức hợp Pyruvate dehydrogenase Xúc tác gắn CoASH với Axit pyruvic để tạo acetyl-CoA
L - + + + +
Hãy cho biết:
a) Loài vi khuẩn nào không thể thực hiện được hô hấp hiếu khí? Giải thích.
b) Các sản phẩm chính mỗi loài vi khuẩn tạo ra sau quá trình chuyển hóa glucozo.
c) Các loài trên sẽ phát triển như nào nếu như bổ sung oxi phân tử vào môi trường.
Hướng dẫn
a).
- Các vi khuẩn không thể thực hiện được hô hấp hiếu khí gồm: loài 1, loài 2, loài 3.
- Loài 1 và loài 2 thiếu xitocrom c oxidaza là enzim chính trong thành phần của quá trình hô hấp hiếu khí (enzym trong chuỗi truyền điện tử).
- Loài 3 thiếu Phức hợp Pyruvate dehydrogenase của giai đoạn trung gian hình thành Acetyl- coA (trước chu trình Crep)
b)
- Loài 1 tạo rượu etanol, CO2 và ATP.
- Loài 2 tạo axit lactic, ATP.
- Loài 3 tạo axit lactic, ATP.
- Loài 4 khi có mặt oxi tạo CO2, H2O và ATP; khi không có mặt oxi tạo rượu etanol, CO2 và ATP.
c)
- Khi bổ sung vào môi trường oxi phân tử thì loài 2 và loài 3 sẽ bị chết nhanh nhất, khả năng chúng là vi khuẩn lactic. Trong điều kiện có oxi, vi khuẩn lactic bị ức chế sinh trưởng vì nó là vi khuẩn kị khí bắt buộc, tế bào thiếu enzim catalaza, SOD giúp chúng tồn tại trong điều kiện có oxi.
- Loài 1 là nấm men rượu – kị khí không bắt buộc. Khi có oxi nấm men chuyển sang hô hấp hiếu khí. Glucôzơ bị phân hủy hoàn toàn; Năng lượng tạo ra nhiều (38 ATP); khi đó chất nhận e là khí oxi.
Tên enzim
Bảng 1.2. Sự có mặt (+) và vắng mặt (-) của mỗi loại enzim trong từng loài vi khuẩn L oài vi khuẩn
LDH AD H Xitôcrô m c oxidaza ATP sinte Phức hợp Pyruvate dehydrogenase
- Loài 4 có khả năng sống được trong cả khi có oxi hoặc không nên sinh trưởng bình thường.
Câu 8: Người ta phân lập được nhiều thể đột biến khuyết dưỡng liên quan đến một con đường chuyển hóa ở một vài vi khuẩn. Khi bổ sung một trong các hợp chất A, B, C, hoặc D vào môi trường nuôi cấy, người ta thấy các thể đột biến sinh trưởng như liệt kê ở bảng 1 dưới đây (dấu + là sinh trưởng được, dấu – là không sinh trưởng
được). Hãy xác định thứ tự của các hợp chất này trong chuỗi chuyển hóa và cho biết các thể đột biến đó mang đột biến ảnh hưởng ở bước nào trong quá trình chuyển hóa đó.
Bảng 1. Sinh trưởng của vi khuẩn trong các môi trường nuôi cấy.
Thể đột biến
p chất
Hướng dẫn Nguyên tắc sắp xếp trật tự các hợp chất trong con đường như sau: Nếu một chất được bổ sung nằm ở phía sau vị trí bị chặn, nó sẽ giúp thể đột biến sinh trưởng được, nếu một hợp chất bổ sung nằm phía trước vị trí bị chặn, nó không có tác dụng khi thêm vào.
Áp dụng nguyên tắc trên ta thấy:
- Nhóm 1 (chủng 2, 4) mọc được khi bổ sung vào môi trường chất A hoặc B hoặc C hoặc D. Do vậy nhóm 1 ảnh hưởng đến 1 bước trước khi tạo thành 4 hợp chất này.
Thứ tự của con đường chuyển hóa là:
→ hợp chất A, B, C, D
Nhóm 1
- Nhóm 2 (chủng 1, 7) vẫn mọc được nếu bổ sung, A hoặc C, hoặc D, nhưng khi bổ sung thêm B thì không mọc được. Do vậy nhóm 2 mang đột biến không
ảnh hưởng đến việc tạo B mà ảnh hưởng đến 1 bước trước khi tạo thành A và C và D.
Và B là chất nằm ở đầu con đường chuyển hóa.
Thứ tự của con đường chuyển hóa là:
→ hợp chất B → hợp chất A, C, D
Nhóm 1 Nhóm 2
- Nhóm 3 (chủng 6, 8) vẫn mọc được nếu bổ sung C hoặc D; nhưng khi bổ sung thêm B hoặc A thì không mọc được. Do vậy nhóm 3 mang đột biến không ảnh hưởng đến việc tạo A và B nhưng ảnh hưởng đến 1 bước trước khi tạo thành C và D. Mà B là chất nằm ở đầu con đường chuyển hóa . Do đó kế tiếp B là A.
Thứ tự của con đường chuyển hóa là:
→ hợp chất B → hợp chất A → hợp chất C, D
Nhóm 1 Nhóm 2 Nhóm 3
- Nhóm 4 (chủng 3, 5) vẫn mọc được nếu bổ sung chất D, nhưng khi
bổ sung thêm vào chất A hoặc B hoặc C thì không mọc được. Do vậy nhóm 4 mang
467
đột biến không ảnh hưởng đến việc tạo A và B và C mà ảnh hưởng đến 1 bước trước khi tạo thành chất D. Và chất D nằm ở cuối con đường chuyển hóa.
Thứ tự của con đường chuyển hóa là:
Hợp chất B → hợp chất A → hợp chất C → hợp chất D
Các thể đột biến mang đột biến ảnh hưởng đến các bước trong quá trình chuyển hóa theo sơ đồ: → hợp chất B → hợp chất A → hợp chất C → hợp chất D
Nhóm 1 Nhóm 2 Nhóm 3 Nhóm 4
Câu 9: Đa số vi khuẩn không chuyển hóa được xylitol để tạo năng lượng. Người ta tiến hành thí nghiệm chuyển một gen mã hóa enzim phân giải xylitol vào E. coli. Một dòng plasmit làm thể truyền được biến đổi để có một gen kháng ampicilin (ampR) và một gen mã hóa β galactozidaza có chứa trình tự nhận biết của enzim cắt giới hạn EcoRI (β galactozidaza có khả năng phân giải X-gal từ không màu thành sản phẩm màu xanh dương). Sau khi đoạn ADN chứa gen mã hóa enzim phân giải xylitol và thể truyền được xử lý riêng rẽ với EcoRI, chúng được trộn với nhau để tái tổ hợp ngẫu nhiên. Hỗn hợp ADN này tiếp tục được cho biến nạp vào một chủng E. coli đã bị đột biến bất hoạt gen Z trong opêron Lac. Sau đó đem cấy trải trên môi trường thạch tối thiểu thu được 3 dòng A, B và C. Tiếp tục nuôi cấy riêng rẽ 3 dòng A, B và C trên môi trường thạch tối thiểu có bổ sung thêm ampicilin và X-gal thu được kết quả như sau:
Dòng A: Xuất hiện khuẩn lạc và không thay đổi màu.
Dòng B: Xuất hiện khuẩn lạc và vùng xung quanh khuẩn lạc chuyển sang màu xanh dương.
Dòng C: Không xuất hiện khuẩn lạc.
a. Giải thích kết quả thí nghiệm.
b. Nếu cả 3 dòng A, B và C được nuôi cấy riêng rẽ trong môi trường chỉ chứa xylitol làm nguồn cacbon duy nhất thì khả năng phát triển của mỗi dòng sẽ như thế nào? Giải thích.
Hướng dẫn
a. Giải thích kết quả thí nghiệm:
- Dòng A phát triển được trên môi trường có ampicilin → có plasmit chứa ampR. Khi nuôi trên X-gal thì màu của môi trường không thay đổi → gen β galactosidaza không được biểu hiện → có hiện tượng biến nạp plasmit tái tổ hợp.
- Dòng B phát triển được trên môi trường có ampicilin và chuyển hóa được Xgal làm môi trường chuyển sang màu xanh → có hiện tượng biến nạp plasmit không tái tổ hợp.
468
- Dòng C không phát triển trên môi trường có ampiclin → không có hiện tượng biến nạp.
b. Khả năng phát triển của 3 dòng khi nuôi cấy trên môi trường có xylitol làm nguồn cacbon duy nhất:
- Dòng A có biến nạp plasmit tái tổ hợp, tuy nhiên không thể chắc chắn plasmit tái tổ hợp này có gen chuyển hóa xylitol. Do đó:
+ Dòng A có thể phát triển được trên môi trường có chứa xylitol nếu plasmit tái tổ hợp có chứa gen chuyển hóa xylitol hoàn chỉnh.
+ Dòng A sẽ không phát triển được trên môi trường chỉ chứa xylitol nếu plasmit tái tổ hợp không chứa gen chuyển hóa xylitol hoàn chỉnh.
- Dòng B có khả năng sử dụng lactozơ (do có gen β galactosidaza trên plasmit hoạt động), nhưng không có khả năng sử dụng xylitol nên không phát triển được trên môi trường chỉ chứa xilytol.
- Dòng C không có biến nạp → không chuyển hóa được xilitol → không phát triển được trên môi trường có chứa xylitol.
Câu 10:
a) Các nhà khoa học thấy rằng phốt phát vô cơ (Pi) thực sự cần thiết cho quá trình lên men glucose bởi nấm men. Khi nguồn cung cấp Pi bị cạn kiệt, quá trình lên men sẽ dừng lại ngay cả khi môi trường vẫn còn glucose. Lúc đó, bên cạnh ethanol và CO2, quá trình lên men còn tích lũy thêm đường fructose 1,6 biphosphate. Giải thích tại sao quá trình lên men bị dừng lại khi nguồn Pi cạn kiệt?
b) Trong một nghiên cứu về chức năng của ti thể, người ta phân lập và chuyển ti thể vào trong một môi trường đệm thích hợp có succinate là nguồn cung cấp điện tử duy nhất cho chuỗi hô hấp. Sau 5 phút ADP được bổ sung vào môi trường. Khoảng 1 phút tiếp theo mỗi chất ức chế (trình bày ở bảng dưới) được bổ sung vào từng ống nghiệm riêng rẽ, sau 10 phút thí nghiệm kết thúc. Nồng độ oxi trong môi trường của từng ống nghiệm thay đổi như thế nào trong suốt thời gian thí nghiệm? giải thích?
Ống nghiệm Chất ức chế Tác dụng
I Atractylosid e Ức chế Protein vận chuyển
ADP/ATP
II Butylmalom ate Ức chế vận chuyển succinate vào ti thể
III Cyanine Ức chế phức hệ cytocrome c oxidase
IV FCCP Làm cho proton thấm được qua màng
V Oligom cin Ứ chế phức hệ ATP synthase
Hướng dẫn a. Quá trình lên men rượu (ethanol) bởi nấm men được chia làm hai giai đoạn: giai đoạn đường phân và giai đoạn tái tạo chất nhận điện tử NAD+ .
+ đường phân cung cấp NADH cho giai đoạn khử acetaldehyde thành ethanol, đồng thời cũng chính giai đoạn khử này giúp tái tạo lại chất nhận điện tử NAD+ cung cấp NAD+ cho giai đoạn đường phân chỉ cần 1 trong hai giai đoạn ngừng hoạt động toàn bộ quá trình ngừng lại.
- Mà trong giai đoạn đường phân, Pi cần thiết cho phản ứng xúc tác bởi enzyme glyceraldehyde 3 phosphate dehydrogenase để chuyển hóa fructose 1,6 biphosphate thành hai phân tử 1,3 biphosphoglycerate, đồng thời giúp chuyển hóa 2 NAD+ thành 2 NADH nhằm cung cấp chất khử cho quá trình hình hình thành ethanol từ acetaldehyde.
Khi nguồn cung cấp Pi bị cạn kiệt NADH không được hình thành quá trình lên men sẽ dừng lại vì không tái tạo lại được chất nhận điện tử NAD+ kết quả cả ethanol, CO2 và đường fructose 1,6 biphosphate được tích lũy.
b. - Trong tất cả các thí nghiệm khi mới bắt đầu nồng độ oxi trong môi trường giảm dần do hô hấp tế bào sử dụng succinate, khi cho thêm ADP thì sự tổng hợp ATP tăng chuỗi truyền điện tử hoạt động tăng.
- Khi bổ sung chất ức chế Atractyloside làm cho quá trình vận chuyển ADP đi vào và ATP đi ra ti thể bị ức chế làm giảm quá trình tổng hợp ATP do đó mức tiêu thụ oxi chậm dần.
- Khi bổ sung chất ức chế Butylmalomate và Cyanine đều ức chế chuỗi truyền điện tử làm ngừng quá trình tiêu thụ oxi nên nồng độ oxi ngừng giảm.
- Khi bổ sung chất ức chế FCCP làm nồng độ oxi giảm nhanh do làm tăng tính thấm của màng với proton làm tăng nồng độ H+ ở chất nền và làm tăng chuỗi truyền điện tử nên tăng lượng tiêu thụ oxi làm nồng độ oxi giảm nhanh.
- Khi bổ sung chất ức chế Oligomycin giống trường hợp 1.
Câu 11: Cho các chủng E.coli thuộc các thể lưỡng bội một phần có kiểu gen sau đây:
(I) I+ O- Z+ / I+ O+ Z-
(II) I+ O+ Z- / I- O+ Z+
(III) I+ O+ Z- / IS O+ Z+
Biết rằng trong Operon Lac, I+, O+, Z+ tương ứng là các trình tự kiểu dại
470
của gen mã hóa protein ức chế (I), vùng vận hành (O) và gen lacZ. O-, Z- là các trình tự đột biến mất chức năng so với trình tự kiểu dại tương ứng. I- là đột biến làm protein ức chế mất khả năng gắn vùng vận hành. IS là đột biến làm protein ức chế mất khả năng gắn vào đồng phân của lactose, nhưng vẫn bám vào Operator.
a. Trong môi trường không có glucose, hãy đánh giá sự biểu hiện của gen Lac
Z của các thể lưỡng bội một phần nói trên khi môi trường có nuôi cấy có hoặc không có lactose?
b. Trong các thể lưỡng bội 1 phần nói trên, hãy xác định tương quan trội lặn giữa các alen: I+, I-, IS. Giải thích?
Hướng dẫn
a. – Chủng I: Lac Z biểu hiện cơ định - luôn biểu hiện hay biểu hiện liên tục cho dù môi trường có hay không có lactozơ
– Chủng II: Lac Z cảm ứng được. Có Lactozơ thì biểu hiện, không lactozơ thì không biểu hiện.
– Chủng III: Lac Z không biểu hiện– luôn không biểu hiện dù môi trường có hay không có lactozơ
b. - Các thể đột biến nói trên thì kiểu gen II và III đều là thể dị hợp về LacI. Ở thể dị hợp, alen trội được biểu hiện ra kiểu hình
- Chúng II: Cảm ứng được tức là có Lactozơ thì biểu hiện, không lactozơ thì
không biểu hiện.Do I+ tạo protein ức chế, Chủng II biểu hiện kiểu hình của I+ → I+ >
I-
- Chủng III: biểu hiện kiểu hình của IS. Nên IS > I+
Tương quan trội lặn IS > I+ > I-
Câu 12. Trình bày điểm giống nhau và khác nhau giữa operon cảm ứng và operon ức chế?
Hướng dẫn - Giống nhau: Đều được tắt bởi dạng hoạt hóa của protein điều hòa (pr ức chế)
- Khác nhau: operon cảm ứng operon ức chế
Luôn tắt, chỉ mở khi có chất cảm ứng.
Protein ức chế khi mới hình thành đã ở dạng hoạt động ngay, liên kết vào O và ức chế phiên mã.
Luôn hoạt động, chỉ tắt khi có chất ức chế.
Prorein ức chế khi mới tổng
hợp ở dạng bất hoạt, chỉ được hoạt hóa khi có chất đồng ức chế
Enzim cảm ứng thường hoạt động trong con đường dị hóa
Enzim ức chế thường hoạt động trong con đường đồng hóa.
Câu 13. Cho biết A, B, C, D là các chất chuyển hóa trung gian (không đúng thứ tự) trong một con đường hóa sinh của tế bào. Người ta tìm thấy 4 thể đột biến khác nhau kí hiệu từ D1 – D4. Khi nuôi cấy 4 thể đột biến này lần lượt trong các môi trường bổ sung chất A, B, C, D người ta thu được kết quả như sau: D1 chỉ sinh trưởng trong môi trường có A và có D; D2 chỉ sinh trưởng trong môi trường chứa A hoặc B hoặc D; D3 chỉ sinh trưởng trong môi trường có D; D4 chỉ sinh trưởng trong môi trường có A hoặc B hoặc C hoặc D. hãy vẽ sơ đồ các bước chuyển hóa của con đường hóa sinh trên và chỉ ra các bước chuyển hóa bị ức chế tương ứng ở các thể đột biến (D1 – D4). Giải thích?
Hướng dẫn
- Sơ đồ chuyển hóa theo trình tự: C B A D.
- D1 bị đột biến làm mất chức năng của gen mã hóa enzim chuyển hóa B thành A.
- D2 bị đột biến làm mất chức năng của gen mã hóa enzim chuyển hóa C thành B.
- D3 bị đột biến làm mất chức năng của gen mã hóa enzim chuyển hóa A thành D.
- D4 bị đột biến làm mất chức năng của gen mã hóa enzim chuyển hóa hình thành chất C.
Ta có thể suy ra được trình tự các bước chuyển hóa trên dựa theo nguyên lí là nếu đột biến nào cần bổ sung tất cả các chất thì thể đột biến đó bị hỏng gen quy định enzim chuyển hóa tiền chất đầu tiên của con đường chuyển hóa. Đột biến nào cần bổ sung một chất thì chất đó là sản phẩm cuối cùng của con đường chuyển hóa.
Câu 14. Khi xử lí tế bào E.coli kiểu dại riêng rẽ với hóa chất acridin và 5-BU, người ta thu được tương ứng 2 dòng đột biến LacZ – 1 và LacZ – 2 mang đột biến điểm trong gen LacZ. Hai dòng đột biến này nhiều khả năng mang loại đột biến gen nào? Giải thích.
Hướng dẫn
- LacZ-1 là đột biến dịch khung vì acridin thường cài xen vào giữa các bazo nito và nếu tái bản xảy ra thường dẫn đến việc xen vào hay mất đi một hoặc 1 số cặp nucleotit, không phải bội số của 3 (tương ứng codon), dẫn đến dịch mã lệch khung đọc.
- LacZ-2 là đột biết thay thế bazo (đồng hoán) vì 5-BU thường cài xen vào vị trí bazo loại Timin trong quá trình tái bản, và lần tái bản sau đó thay thế bằng Xitozin. Dẫn đến thay thế cặp A-T bằng G-X.
Câu 15. Hiện tại, bạn đang nghiên cứu về sự tổng hợp threonine ở E.coli. Để tìm các gen mã hóa các protein quan trọng cho tổng hợp threonine, bạn tiến hành gây đột biến để tìm các đột biến yêu cầu cần bổ sung threonine cho sự phát triển. Bạn đã tìm thấy có 9 dòng đột biến và tất cả đều cần bổ sung threonine vào môi trường nuôi cấy để phát triển. Sau đó, bạn tiến hành cis – trans test với tất cả các dòng đột biến bạn có. Kết quả thu được như bảng sau. (+: phát triển; - : không phát triển)
- Tạo kiểu hình đột biến.
Khi ta cho các dòng đột biến khác nhau lai với nhau:
- Nếu khuẩn lạc phát triển: Hai đột biến thuộc 2 gen khác nhau, bù trừ cho nhau tạo kiểu gen dị hợp tạo kiểu hình bình thường (kiểu dại).
- Nếu khuẩn lạc không phát triển: Hai đột biến thuộc cùng 1 gen nên không bù trừ cho nhau, tạo kiểu gen đồng hợp Tạo kiểu hình đột biến.
m
t1 m t2 m t3 m t4 m t5 m t6 m t7 m t8 m t9 w t
Từ đó, ta có:
- 1, 3, 5, 6, 9 thuộc cùng 1 gen.
- 2, 8 thuộc cùng 1 gen.
- 4 không bù cho mình, bù cho những dòng đột biến còn lại gen độc lập.
- 7 không bù cho mình, bù cho những dòng đột biến còn lại gen
độc lập.
Có 4 gen tham gia tổng hợp threonine.
Câu 16. Một Operon mã hóa enzim 1 và 2 được điều hòa bởi sự chuyển hóa chất X chứa đựng 7 trình tự A, B, C, D, E, F, G, H chưa rõ chức năng. Để làm sáng tỏ chức năng của chúng, sự ảnh hưởng của đột biến ở các trình tự từ A đến G dựa trên dự tổng hợp các enzyme được đánh giá thông qua sự có mặt và sự vắng mặt của X. Có mặt X Không có mặt X
Đột biến trong trình tự Ez 1 Ez 2 Ez 1 Ez 2
Không có đột biến - - + +
A - - - +
B + + +
C - - + -
D - - - -
E - - - +
F + + + +
Có bao nhiêu gen mã hóa các enzim tham gia vào quá trình tổng hợp threonine?
Giải thích.
Hướng dẫn
Ta thấy:
Khi cho các dòng đột biến giống nhau lai với nhau:
- Không phát triển không tạo khuẩn lạc.
- Đột biến trên cùng 1 gen không bù trừ được cho nhau.
G - - + +
Hãy xác định vai trò của Operon và các trình tự của A, B, C, D, E, F, G. Giải thích
Hướng dẫn - Operon khi có mặt X thì cả hai ez đều không được tổng hợp do đó đây không phải operon đồng hóa.
474
- Thể đột biến B, F biểu hiện giống nhau. Khi có hay không có X
thì operon luôn mở (luôn tạo enzim).

Bị ĐB vùng O (LacOc)
- Thể đột biến D khi có mặt hay không có mặt X cũng luôn đóng.
Bị đột biến vùng P
- Thể đột biến A, E có biểu hiện giống nhau. Khi vắng mặt X chỉ có
ez 2 được tổng hợp.
ĐB gen tổng hợp ez 1
- Thể đột biến C khi vắng mặt X chỉ tổng hợp ez 1.
ĐB gen tổng hợp ez 2
- Thể đột biến G biểu hiện giống kiểu dại trình tự đb không liên
quan đến operon.
Các bài tập tự giải
Câu 17: Artemisinin là một loại thuốc chống bệnh rốt rét được chiết rút từ cây ngải hoa vàng (Artemisia annua), hiện nay được sản xuất nhờ tổng hợp hữu cơ qua axit artemisinic thu từ sinh khối vi sinh vật chuyển gen.
a) Quá trình tạo plasmit tái tổ hợp từ gen mã hoá enzim tổng hợp axit artemisinic (của A. annua) có gì khác nhau nếu người ta muốn biểu hiện gen ở (1) nấm men rượu và (2) vi khuẩn E. coli? Giải thích.
b) Để tạo plasmit tái tổ hợp (biểu hiện ở E. coli) từ thể truyền gồm một gen kháng kháng sinh tetracylin (tetR) và một gen mã hoá β galactozidaza (có khả năng phân giải X-gal từ không màu thành sản phẩm màu xanh dương) có chứa trình tự nhận biết của enzim cắt giới hạn EcoRI, người ta tiến hành các bước sau:
B1: Xử lí gen mã hoá enzim tổng hợp artemisinic (đã cải biến phù hợp) và thể truyền với enzim EcoRI.
B2: Trộn các đoạn gen thu được ở trên với nhau và bổ sung enzim ADN ligaza.
B3: Cho các plamsit thu được vào hỗn hợp tế bào E. coli mang đột biến ở gen lacZ trên nhiễm sắc thể.
B4: Cấy rải tất cả vi khuẩn lên bề mặt môi trường thạch argar có bổ sung tetracylin và X-gal.
Hãy cho biết các sản phẩm có thể thu được qua bước (2), (3) và giải thích mục đích của bước (4).
Câu 18: Virus cúm A
Dịch cúm theo mùa cướp đi sinh mạng của hơn 500.000 người mỗi năm trên toàn thế giới. Nhiều hóa chất đã được thử nghiệm để ngăn chặn sự nhân lên của virus cúm A trong cơ thể. Trong một thí nghiệm, trước khi tiếp xúc virus cúm A, tế bào chủ được xử lí lần lượt với mỗi loại hóa chất sau: zanamivir (chất ức chế neuraminidase),
NH4Cl (duy trì pH cao của lysosome), actinomycin D (ức chế sự phiên mã). Hãy dự đoán tác động của các hóa chất trên đối với quá trình nhân lên của virus cúm A.
Câu 19: Các virut cúm A thuộc loại Orthomyxovirus lớp V, được chia nhóm dựa vào hai protein có trên bề mặt virut: hemagglutinin (H) và neuraminidaza (N). Có 18 loại hemagglutinin khác nhau (H1-H18) và có 11 loại neuraminidaza khác nhau (N1-N11).
a. Hãy cho biết đặc điểm xâm nhập và tổng hợp hệ gen của virut cúm A.
b. Tại sao virut cúm A có tốc độ biến đổi rất nhanh?
c. Tamiflu là thuốc được sử dụng để điều trị bệnh cúm bằng cách ức chế protein N của virut. Tamiflu ức chế quá trình nào trong chu trình nhân lên của virut?
d. Nếu đột biến làm thay đổi protein H trên bề mặt virut thì sẽ ảnh hưởng gì đến khả năng gây bệnh của virut đó?
Câu 20: Khi tiếp nhận tín hiệu từ yếu tố sinh trưởng có nguồn gốc tiểu cầu (PDGF), thụ thể PDGFR – một loại thụ thể tyrosine kinase gồm 2 tiểu phần – được hoạt hóa và tự phosphoryl hóa ở nhiều vị trí tyrosine trên vùng nội bào của nó. Những gốc tyrosine được phosphoryl hóa là các vị trí liên kết với các protein tương tác với thụ thể. Một trong những đáp ứng tế bào đối với tín hiệu PDGF là tăng cường tổng hợp ADN, được xác định thông qua việc đo mật độ tín hiệu thymidine phóng xạ kết hợp vào ADN. Để xác định vai trò của các protein (kí hiệu A, B, C và D ở hình dưới) trong đáp ứng tế bào, người ta tạo ra các dạng đột biến (kí hiệu từ 2 đến 9) của thụ thể PDGFR chứa một hoặc một số gốc tyrosine sẽ được phosphoryl hóa. Mỗi loại thụ thể PDGFR đột biến được biểu hiện ở các tế bào không tự tổng hợp thụ thể PDGFR và những vị trí tyrosine có mặt trên thụ thể đều được phosphoryl hóa dưới tác động của tín hiệu PDGF. Số liệu về mức độ tăng cường tổng hợp ADN trong các tế bào có thụ thể kiểu dại (kí hiệu 1) và thụ thể đột biến (từ 2 đến 9) được biểu thị ở trên hình. Biết rằng, mức tăng cường tổng hợp ADN ở tế bào có thụ thể kiểu dại được coi là 100%.
Hãy cho biết:
476
a. Có protein nào trong số 4 protein (từ A đến D) ức chế sự tăng cường tổng hợp ADN không? Giải thích.
b. Protein nào trong số 4 protein (từ A đến D) tăng cường tổng hợp ADN? Giải thích.

c. Nếu đột biến 5 chứa serine thay thế cho tyrosine ở vị trí 1021 thì mức độ tăng cường tổng hợp ADN có thay đổi không? Giải thích.
d. Ở các tế bào có thụ thể PDGFR kiểu dại, nếu có thêm thụ thể PDGFR đột biến được biểu hiện ở mức rất cao thì đột biến xảy ra ở vùng ngoại bào hay ở vùng nội bào có ảnh hưởng lớn hơn đến sự tăng cường tổng hợp ADN? Giải thích.
a. Hãy nhận xét về sự thay đổi tần số các nhóm alen của gen mã hóa protease của HIV khi bệnh nhân dùng thuốc ức chế protease và giải thích nguyên nhân.
b. Hãy dự đoán sự thay đổi của đồ thị tần số các nhóm alen của gen mã hóa protease của quần thể 2 sau khoảng 150 ngày dùng thuốc.
c. Hãy nhận định ba yếu tố quan trọng có thể tác động làm thay đổi nhanh tần số các nhóm alen của gen mã hóa protease của HIV như dữ liệu mô tả.
d. Hãy dự đoán xu hướng thay đổi tần số các nhóm alen nếu bệnh nhân ngừng sử dụng thuốc ức chế protease sau đợt điều trị trên. Giải thích.
Câu 22: a) Để nghiên cứu ảnh hưởng của môi trường đến quá trình lên men và chất lượng dưa cải muối, người ta tiến hành muối dưa ở nồng độ 2,5% và 210C với 4 nhóm thí nghiệm gồm: lên men tự phát (A), bổ sung vi khuẩn Leuconostoc mesenteroides (B), bổ sung vi khuẩn Lactococcus lactis (C) và bổ sung nước dưa cũ (D). Kết quả thu được sau 28 ngày lên men thể hiện ở bảng và hình dưới:
(1) Sắp xếp các nhóm thí nghiệm theo hiệu quả lên men tăng dần. Giải thích.
Câu 21: Virus gây hội chứng suy giảm miễn dịch ở người (HIV) thuộc nhóm retrovirus có hệ gen gồm hai bản sao ARN với kích thước khoảng 10 kb. Sau khi xâm nhập vào tế bào chủ, HIV sử dụng enzym phiên mã ngược để tạo nên ADN mạch kép từ chính hệ gen của nó.
Phân tử ADN này sẽ chèn vào ADN của tế bào chủ và tiến hành phiên mã tạo thành hệ gen ARN virus. Hệ gen ARN này sẽ được đóng gói vào trong hạt virus rồi giải phóng ra khỏi tế bào chủ bằng protease của virus. Một trong những liệu pháp nhằm ức chế sự nhân lên của HIV là sử dụng các nucleotide giả (đã loại bỏ gốc phosphat) để ức chế enzym phiên mã ngược nhưng không ảnh hưởng đến ADN polymerase của tế bào chủ. Một liệu pháp khác là sử dụng các chất ức chế protease nhằm ngăn chặn sự phóng thích các hạt virus mới.
Doukhan và Delwart (2001) đã nghiên cứu tần số các nhóm alen của gen mã hóa protease của hai quần thể virus HIV (kí hiệu quần thể 1 và 2 tương ứng ở hai bệnh nhân 1 và 2) sau khi họ được dùng thuốc ức chế protease. Kết quả của nghiên cứu được biểu thị trong hai đồ thị dưới đây.
Trên quan điểm tiến hóa quần thể, hãy trả lời các câu hỏi sau:
477
(2) Nhóm nào có chất lượng sản phẩm tốt nhất? Giải thích.
Chỉ tiêu A B C D
Mùi hương Hă ng Nh ẹ Dễ chịu H ăng
Màu sắc Và ng Và ng rơm Vàng rơm V àng nhạt
Mùi vị Ch ua Ch ua nhẹ Chua đặc trưng Đ ắng
Kết cấu dưa Tr ung bình Mề m Giòn M ềm
Bảo quản được 4 tháng 2 –3 tháng
tháng
– 8 tuần
b) Phân lập vi khuẩn L. lactis từ nước dưa và Clostridium botulinum từ đất rồi
478
nuôi trong môi trường kị khí thích hợp, sau đó nhỏ vài giọt H2O2 vào ống nghiệm chứa mỗi loại vi khuẩn, người ta không thấy hiện tượng xảy ra. Giải thích kết quả thu được. Từ kết quả có thể phân loại mỗi loài trên dựa vào nhu cầu ôxi cho sinh trưởng được không? Tại sao?
c) Khi chuyển L. lactis ra môi trường thoáng khí và nuôi trên đĩa thạch dinh dưỡng, vi khuẩn sinh trưởng rất chậm mặc dù có nguồn cacbon dồi dào. Nếu bổ sung sắt (thành phần của protein Hem) vào môi trường, vi khuẩn sinh trưởng nhanh hơn và bắt đầu tiêu thụ O2. Hãy giải thích hiện tượng trên. Sắt có phải nhân tố sinh trưởng đối với L. lactis không? Giải thích.
dạy các lớp chuyên và bồi dưỡng học sinh giỏi tại trường chúng tôi và thu được những kết quả nhất định:
- Học sinh hiểu bài, nắm chắc kiến thức, linh hoạt trong các câu hỏi vận dụng, giải thích tốt nhiều vấn đề có tính thực tiễn cao.
- Các em hứng thú hơn trong giờ học, tích cực chủ động sáng tạo trong quá trình học để lĩnh hội kiến thức.
- Nội dung chuyên đề sử dụng tốt trong công tác bồi dưỡng học sinh giỏi các cấp.
Chuyên đề này đã được tìm hiểu, tập hợp, chọn lọc từ nhiều nguồn tài liệu liên quan để trước hết là giúp các tác giả trong quá trình giảng dạy, đồng thời hỗ trợ các em học sinh cùng tìm hiểu nội dung này. Hy vọng rằng chuyên đề sẽ giúp ích cho các thầy cô giáo và các em học sinh trong quá trình giảng dạy và học tập. Tuy nhiên, nhiều vấn đề còn ít nhiều mang tính chủ quan, kiến thức đưa ra trong phần lý thuyết còn chưa đầy đủ, có thể còn chưa chính xác, hệ thống bài tập chưa bao quát hết các nội dung, đề nghị các thầy cô giáo và các em học sinh đóng góp bổ sung để chuyên đề được hoàn thiện hơn, hợp lý hơn và có ích hơn trong thực tiễn.
TÀI LIỆU THAM KHẢO
1. Sinh học, Campbell . Reece, 2011.
2. Sinh học 12, Nguyễn Thành Đạt, Phạm Văn Lập, Đặng Hữu Lanh, Mai Sỹ Tuấn, 2012.
3. Sinh học 10, Nguyễn Thành Đạt, Phạm Văn Lập, Trần Dụ Chi, Trịnh Nguyên Giao, Phạm Văn Tỵ, 2021.
3. Tài liệu giáo khoa chuyên Sinh học THPT Vi sinh vật học, Phạm Văn Tỵ, Nguyễn Vĩnh Hà, 2009.
PHẦN III: KẾT LUẬN VÀ KIẾN NGHỊ
Nội dung chuyên đề đã trình bày được:
- Hệ thống kiến thức về hóa sinh và di truyền vi sinh vật.
- Sưu tầm, giới thiệu một số câu hỏi, bài tập vận dụng tích hợp hóa sinh và di truyền vi sinh vật.
Với nội dung kiến thức cơ bản về tích hợp hóa sinh và di truyền vi sinh vật cùng với hệ thống câu hỏi, bài tập được đề cập ở trên chúng tôi đã sử dụng trong giảng
479
4. Tài liệu giáo khoa chuyên Sinh học THPT Di truyền và Tiến hóa, Phạm Văn Lập, Trần Ngọc Danh, Đinh Đoàn Long,2015.
5. Bồi dưỡng học sinh giỏi Sinh học THPT phần Di truyền và Tiến hóa, Vũ Đức Lưu, 2011.
6. Bồi dưỡng Học sinh giỏi Sinh học 12, Phan Khắc Nghệ, 2013.
7. Các website trên internet về Hóa sinh, Vi sinh và Di truyền
8. Các đề thi học sinh giỏi quốc gia, quốc tế; các đề thi đề xuất
Chuyên đề
HỆ THỐNG LÝ THUYẾT VÀ BÀI TẬP TÍCH HỢP
HOÁ SINH, VI SINH, DI TRUYỀN
Trường THPT Chuyên Lê Khiết - Quảng Ngãi
Chuyên đề đạt giải Ba
PHẦN 1: MỞ ĐẦU
1. Lý do chọn đề tài
Trong khoảng 150 năm trở lại đây, vi sinh vật học được xem là lĩnh vực có những bước phát triển vượt bậc so với các lĩnh vực khác của Sinh học. Là một ngành khoa học ứng dụng, vi sinh vật học đi đầu trong nhiều nghiên cứu đột phá về con người, thuốc thú y, nông nghiệp, công nghiệp, năng lượng và môi trường. Phải khẳng định rằng, cùng với sự phát triển của mình, vi sinh vật học đã có rất nhiều tác động tích cực đến sự phát triển của các ngành khoa học khác và mọi mặt trong cuộc sống của con người.
Di truyền học là một trong những ngành khoa học đã tạo được rất nhiều bước ngoặt quan trọng nhờ vi sinh vật học. Bắt đầu từ năm 1944 với nghiên cứu của D. T.
Avery, C. M. Macleod và M. Mc. Carty thực nghiệm trên vi khuẩn đã chứng minh quá trình biến nạp được thực hiện thông qua ADN. Năm 1957, H. Fraenkel – Conrat và B. Singer đã chứng minh thêm vai trò của axit nuclêic trong truyền đạt thông tin di truyền qua thí nghiệm trên virut khảm thuốc lá. Cùng với phát hiện vĩ đại về cấu trúc ADN của J. D. Watson và F. H. C. Crick năm 1952, phát hiện về vai trò của Operon trong việc “đóng, mở gen” của F. Jacob và J. Monod, việc xác định được mã di truyền của M. Nirenberg và G. Khorana (1961 – 1965),… đã giúp con người có được bức tranh toàn cảnh về cấu trúc, chức năng, quy luật vận động của vật liệu di truyền ở cả sinh vật nhân sơ và nhân thực. Từ đây, hình thành các lĩnh vực di truyền học hoá sinh và di truyền học vi sinh vật, hai nền tảng chính cho sự ra đời của di truyền học phân tử (1953) và công nghệ ADN tái tổ hợp (1978).[2]
Bên cạnh đó, thành tựu của các ngành khoa học khác, đặc biệt là hóa sinh học đã giúp cho các nhà vi sinh vật học hiểu rõ hơn về thành phần cấu tạo, hoạt động chức năng và các quá trình chuyển hóa xảy ra trong tế bào vi sinh vật, làm nền tảng cho các nghiên cứu liên quan đến vi sinh vật, giúp cho vi sinh vật học không ngừng phát triển. Hóa sinh vi sinh vật không chỉ là nội dung giảng dạy trong chương trình Sinh học THPT mà còn là học phần quan trọng trong chương trình đào tạo của các ngành: công nghệ thực phẩm, y dược, công nghệ sinh học, nông lâm,….
nâng cao chất lượng bồi dưỡng học sinh giỏi, đóng góp thêm vào nguồn tài liệu tham khảo cho học sinh và quý đồng nghiệp, quan trọng hơn là hoàn thiện kiến thức của bản thân, chúng tôi tiến hành nghiên cứu đề tài “Hệ thống lý thuyết và bài tập tích hợp hóa sinh – vi sinh – di truyền”.
2. Mục đích của đề tài
- Cung cấp thêm tài liệu tham khảo về kiến thức tích hợp vi sinh – hóa sinh và vi sinh – di truyền cho học sinh và giáo viên.
- Xây dựng hệ thống bài tập tích hợp vi sinh – hóa sinh – di truyền, từ đó rèn luyện kĩ năng vận dụng kiến thức để giải quyết vấn đề cho học sinh.
- Xây dựng được mạch kiến thức cơ bản, có thể áp dụng để dạy nền cho mọi đối tượng học sinh.
Do đó, hóa sinh vi sinh vật và di truyền vi sinh vật là hai nội dung thường xuyên xuất hiện trong các đề thi Học sinh giỏi và Olimpic. Đây là hai nội dung không mới nhưng tương đối khó đối với học sinh và cả giáo viên dạy bồi dưỡng vì phạm vi kiến thức rất rộng, bài tập đa dạng, thế giới của các vi sinh vật “bé nhỏ” lại không thể quan sát bằng mắt thường nên kiến thức mang tính trừu tượng cao. Với mong muốn 482
PHẦN 2: NỘI DUNG
1. HỆ THỐNG LÝ THUYẾT TÍCH HỢP VI SINH – HÓA SINH
Cơ thể của tất cả sinh vật đều được cấu tạo từ các nguyên tố hóa học. Các nguyên tố đa lượng như C, H, O, N, Ca, P, S,… đóng vai trò quyết định xây dựng nên
481
các chất hữu cơ, các cấu trúc của tế bào và cơ thể. Các nguyên tố vi lượng như Mo, Fe, Cu, Zn,… có vai trò quan trọng trong các hoạt động sống như quá trình hóa thẩm thấu, hoạt hóa enzim,…[4]. Vì vậy, việc nghiên cứu thành phần cấu tạo và các quá trình chuyển hóa vật chất trong hệ thống sống (Hóa sinh học) có vai trò rất quan trọng, liên quan chặt chẽ đến tất cả các lĩnh vực khác của Sinh học, trong đó có Vi sinh vật học.
Vi sinh vật tuy có kích thước nhỏ bé, cấu trúc cơ thể tương đối đơn giản nhưng chúng có tốc độ sinh trưởng, sinh sản rất nhanh và hoạt động trao đổi chất vô cùng mạnh mẽ. Vi sinh vật có thể phân giải hầu hết tất cả các loại chất có trên thế giới, kể cả những chất rất khó phân giải, hoặc những chất gây hại đến nhóm sinh vật khác. Bên cạnh khả năng phân giải, vi sinh vật còn có khả năng tổng hợp nhiều hợp chất hữu cơ phức tạp, trong điều kiện nhiệt độ, áp suất bình thường. Do đó, nghiên cứu các hoạt động trao đổi vật chất ở vi sinh vật có ý nghĩa to lớn, được ứng dụng trong thực tiễn đời sống, trong sản xuất công – nông nghiệp và công nghệ sinh học.
1.1. Dinh dưỡng của vi sinh vật
1.1.1. Nhu cầu dinh dưỡng của vi sinh vật
Vi sinh vật cần ở môi trường tự nhiên hay môi trường nuôi cấy các chất dinh dưỡng để xây dựng nên các hợp chất của tế bào và những hợp chất dùng cho trao đổi năng lượng. Các chất dinh dưỡng mà vi sinh vật cần đều được cấu thành từ các nguyên tố hóa học. Tuy nhiên, chỉ có một vài nguyên tố là thành phần thiết yếu, có vai trò chi phối hệ thống sống: hydro (H), oxy (O), cacbon (C), nitơ (N), phốt pho (P), lưu huỳnh (S) và selen (Se). Ngoài ra, có ít nhất 50 nguyên tố khác, mặc dù không thiết yếu nhưng có tham gia vào các quá trình chuyển hóa trong tế bào của vi sinh vật [6].
Ngoài nước (chiếm từ 70 - 80% khối lượng của tế bào vi sinh vật) thì trong tế bào chủ yếu là các đại phân tử protein, axit nucleic, lipit và polysaccarit. Trong đó, protein chiếm 55% khối lượng chất khô của tế bào [6].
1.1.2. Cacbon, nitơ và các nguyên tố đa lượng
Tất cả các tế bào đều cần carbon và hầu hết các sinh vật nhân sơ cần các hợp chất hữu cơ để cung cấp nguồn cacbon cho chúng. Các vi sinh vật tự dưỡng xây dựng cấu trúc tế bào của chúng từ nguồn cacbon là CO2. Cacbon chiếm khoảng 50% khối lượng khô của tế bào vi khuẩn [6].
Tế bào vi khuẩn có khoảng 13% nitơ chứa trong protein, axit nucleic và một số thành phần cấu tạo khác của tế bào. Lượng lớn nitơ có sẵn trong tự nhiên là amoniac (NH3), nitrat (NO3 -), hoặc khí nitơ (N2). Hầu như tất cả các sinh vật nhân sơ đều có thể sử dụng NH3 làm nguồn nitơ, nhiều vi sinh vật cũng có thể sử dụng NO3 - và một số có thể sử dụng chất hữu cơ nguồn nitơ, chẳng hạn như axit amin. N2 chỉ được sử dụng bởi các vi sinh vật cố định nitơ [6].
Nguồn nitơ là nguồn cung cấp N cho vi sinh vật để tổng hợp nên các hợp chất chứa N trong tế bào. Chỉ một số ít vi sinh vật tự dưỡng (thuộc nhóm ammon hoá, nhóm nitrate hoá) dùng muối ammone, muối nitrate làm nguồn năng lượng. Trong điều kiện thiếu nguồn C và không có oxy, một số vi sinh vật kỵ khí có thể sử dụng một số axit amin làm nguồn năng lượng [2].

Năng lực hấp thu muối ammone và nitrate ở vi sinh vật là khá mạnh. Ion NH4 + sau khi được tế bào hấp thu có thể được trực tiếp sử dụng. Còn nitrate sau khi được hấp thụ cần khử thành NH4 + rồi mới được vi sinh vật sử dụng [6].
Ngoài C, N, O và H (từ H2O), nhiều nguyên tố đa lượng khác cần thiết cho tế bào nhưng thường với lượng nhỏ hơn [6]:
+ Phôtpho là thành phần cấu tạo axit nucleic, phôtpholipit,… và thường được cung cấp cho tế bào dưới dạng phốt phát (PO4 2-).
+ Lưu huỳnh là thành phần cấu tạo axit amin cysteine, methionine và một số loại vitamin như thiamine, biotin,… Lưu huỳnh thường được cung cấp cho các tế bào dưới dạng muối sulfat (SO4 2-).
+ Kali (K) cần thiết cho hoạt động của một số enzym.
+ Magiê (Mg) cần thiết để ổn định ribosome, màng, axit nucleic và cũng cần cho hoạt động của nhiều enzym.
+ Canxi (Ca) và natri (Na) là những chất dinh dưỡng quan trọng cho một số ít vi sinh vật, đặc biệt vi sinh vật biển rất cần natri.
1.1.3. Nguyên tố vi lượng và các nhân tố sinh trưởng
Có những nguyên tố hóa học mà vi sinh vật chỉ cần một lượng rất nhỏ cho sự sinh trưởng của chúng và chỉ chiếm ít hơn 0,01% khối lượng của tế bào gọi là các nguyên tố vi lượng: B, Mo, Se, Cu,…. Tuy chỉ chiếm tỉ lệ rất nhỏ nhưng vi sinh vật không thể sống nếu thiếu các nguyên tố này. Đứng đầu trong số các nguyên tố vi lượng mà vi sinh vật cần là sắt (Fe). Sắt là thành phần chính của các cytochromes và
483
các protein sắt - lưu huỳnh (có vai trò quan trọng trong chuỗi vận chuyển điện tử của quá trình hô hấp tế bào). Nhiều nguyên tố vi lượng là thành phần không thể thiếu của các enzim [6].

Nhân tố sinh trưởng là những hợp chất hữu cơ mà một số vi sinh vật cần cho sự sinh trưởng, phát triển của chúng nhưng lại không thể tự tổng hợp được. Các nhân tố sinh trưởng bao gồm 3 loại hợp chất: các axit amin, các vitamin, bazơ purine và pyrimidine [3].
Mỗi loài vi sinh vật cần nhân tố sinh trưởng với liều lượng nhất định và rất thấp. Nhu cầu đối với nhân tố sinh trưởng của một loài vi sinh vật đôi khi được đáp ứng khi có mặt một loài vi sinh vật khác có khả năng tổng hợp nhân tố sinh trưởng đó, gọi là hiện tượng đồng dưỡng.
Dựa vào nhu cầu của vi sinh vật về nhân tố sinh trưởng, người ta chia chúng thành hai nhóm [3]:
+ Vi sinh vật nguyên dưỡng: là những vi sinh vật không nhất thiết cần các nhân tố sinh trưởng, có thể nuôi cấy chúng trên môi trường tối thiểu.
+ Vi sinh vật khuyết dưỡng: là những vi sinh vật đòi hỏi các chất hữu cơ nhất định cần cho sự sinh trưởng của chúng.
Cùng một loài vi sinh vật nhưng nhu cầu đối với nhân tố sinh trưởng

cũng thay đổi tuỳ theo điều kiện môi trường. Ví dụ Mucor rouxii khi sinh trưởng trong điều kiện kỵ khí thì cần thiamin (B1) và biotin (H), nhưng trong điều kiện hiếu khí thì lại tự tổng hợp được các vitamin này [2].
1.2 Các kiểu dinh dưỡng của vi sinh vật
Căn cứ vào nguồn cacbon, nguồn năng lượng và nguồn điện tử, người ta phân chia vi sinh vật thành các kiểu dinh dưỡng (Hình 1.2) [6].
Một nhóm lớn các vi khuẩn hóa dưỡng hữu cơ là những vi sinh vật gây bệnh hoặc rất được chú ý trong y học, các vi khuẩn tạp nhiễm vào thức ăn, các vi khuẩn được sử dụng trong công nghiệp để tổng hợp các chất kháng sinh, các loại vitamin, các axit amin,…[3]
Những vi khuẩn dinh dưỡng ký sinh là những vi sinh vật sống trong tế bào chủ như các loài Rickettsia, Chlamydia,… chúng lấy năng lượng từ cơ thể chủ nhờ ký sinh bắt buộc [3].
Hình 1.2. Các kiểu dinh dưỡng của vi sinh vật.
1.3. Chuyển hóa vật chất và năng lượng ở vi sinh vật Sự trao đổi chất ở một tế bào hay một cơ thể là một quá trình gồm toàn bộ các phản ứng hóa học xảy ra trong tế bào hoặc cơ thể. Các phản ứng này có thể là những phản ứng phân giải hay phản ứng tổng hợp các hợp chất khác nhau, mà qua đó tế bào có thể thu được hoặc tiêu tốn năng lượng.
Quá trình trao đổi năng lượng là quá trình phân giải có kèm theo giải phóng năng lượng. Ở sinh vật bậc cao quá trình dị hoá và trao đổi năng lượng chỉ là một, đó là sự oxi hoá các chất hữu cơ trong cơ thể để giải phóng năng lượng.

Ở vi sinh vật quá trình trao đổi năng lượng không chỉ là quá trình dị hoá mà chủ yếu còn là quá trình phân giải các chất được hấp thụ từ bên ngoài [2].
1.3.1 Sinh tổng hợp các chất ở vi sinh vật
Hoạt động sinh tổng hợp các chất ở vi sinh vật được chia thành hai loại như sau [3]:
- Sinh tổng hợp các sản phẩm sơ cấp: gồm nhiều giai đoạn, thông qua hàng loạt các phản ứng trung gian để tổng hợp nên các đại phân tử hữu cơ tham gia cấu tạo nên các thành phần cấu trúc của tế bào như: màng nhày, thành tế bào, màng sinh chất,….
485
- Sinh tổng hợp các sản phẩm thứ cấp có các đặc điểm sau:
+ Các chất trao đổi thứ cấp là những chất không nhất thiết cần cho sự sinh trưởng, phát triển của cơ thể vi sinh vật như: chất kháng sinh,…
+ Có rất nhiều dạng cấu trúc và thay đổi lớn về hoạt tính sinh học.
+ Được sinh ra bằng con đường tổng hợp riêng, con đường tổng hợp trung gian, con đường trao đổi chất sơ cấp.
+ Không diễn ra suốt trong tất cả các giai đọan của quá trình sống mà chủ yếu ở pha cân bằng động (sau pha lũy thừa) và rất phức tạp. Các phản ứng được xúc tác bởi các enzim đặc trưng, khác với các enzim tham gia quá trình tổng hợp sơ cấp.
+ Sự tổng hợp các chất trao đổi thứ cấp được điều khiển bởi tập hợp các gen thường nằm liên kết với các gen điều chỉnh và ức chế ở các plasmid. Cơ chế kiểm soát của quá trình tổng hợp các chất trao đổi thứ cấp được cảm ứng tăng cường do trạng thái sinh lý của cơ thể và điều kiện nuôi cấy.
+ Một chất trao đổi thứ cấp thường chỉ được tổng hợp nhờ một số cơ thể vi sinh vật nhất định. Ví dụ: Penicilline được tổng hợp bởi Penicillium chrysogenum, Erythromycine được tổng hợp bởi Streptomyces erythreus,….
1.3.2 Trao đổi năng lượng
Năng lượng chính là động lực trao đổi chất của tế bào, các phản ứng cung cấp năng lượng là yếu tố sống còn của tế bào.
Trong thế giới vi sinh vật có sự khác biệt rất lớn về khả năng sử dụng các nguồn năng lượng [3]:
+ Vi sinh vật hiếu khí có khả năng tổng hợp ATP cho mình từ đường bị phân giải theo con đường đường phân và chu trình Krebs nhờ chuỗi hô hấp trên màng sinh chất (đối với vi sinh vật nhân sơ) hoặc trên màng trong của ty thể ở vi sinh vật nhân thực.
+ Vi sinh vật kỵ khí thu năng lượng nhờ quá trình lên men các loại đường hoặc nhờ chuỗi vận chuyển electron trên màng sinh chất với chất nhận electron cuối cùng là một chất vô cơ không phải O2.
Tóm lại, hô hấp kỵ khí, hô hấp hiếu khí và lên men đều là quá trình phân giải chất hữu cơ và chuyển hóa năng lượng trong chất hữu cơ thành năng lượng ATP để sử dụng cho các hoạt động sống của tế bào vi sinh vật. Hô hấp kị khí và hô hấp hiếu khí trải qua 3 giai đoạn: phân giải đường tạo ra axit pyruvic, chu trình Crep và chuỗi truyền electron hô hấp. Lên men gồm 2 giai đoạn là phân giải đường tạo axit pyruvic và biến đổi axit pyruvic thành sản phẩm lên men.
1.3.2.1 Sự phân giải glucozơ thành pyruvat
487
Vi sinh vật phân giải đường thành Pyruvate và các chất trung gian tương tự theo ba con đường: đường phân, con đường pentose-phosphate và con đường Entner – Doudoroff [3].
* Con đường đường phân hay Embden-Meyerhof-Parnas (EMP)
Đây là con đường được biết sớm nhất và phổ biến nhất dùng phân giải glucose thành pyruvate. Đường phân gặp ở tất cả các nhóm chủ yếu của vi sinh vật và hoạt động trong sự có mặt cũng như vắng mặt của oxy. Quá trình này diễn ra trong tế bào chất của cả vi sinh vật nhân sơ và nhân thực.
Glucose chuyển thành Pyruvat qua 10 phản ứng từ 1 đến 10 (Hình 1.3) tạo ra các chất trung gian đều ở dạng photphoryl hóa [6].
Phản ứng đặc trưng của con đường EMP là sự phân cắt Fructose-1, 6bisphosphate có 6C thành 2 chất Triozophosphate có 3C. Vì vậy, người ta còn gọi nó với cái tên là con đường Hecxozodiphosphat (HDP).
488
Hình 1.3. 10 phản ứng chi tiết của đường phân.
* Con đường PP (Pentozo-phosphat hay hexo-monophosphate)
Con đường này có thể được dùng đồng thời với con đường đường phân và con đường Entner - Doudoroff, diễn ra trong điều kiện hiếu khí cũng như kỵ khí và có vai trò quan trọng trong sinh tổng hợp cũng như trong phân giải.
Hai enzyme đặc trưng của con đường chuyển hóa này là: Transketolase xúc tác chuyển nhóm ketol 2C và Transaldolase xúc tác chuyển nhóm 3C từ sedoheptulose - 7 - phosphate với glyceraldehyde-3-phosphate. Phương trình tổng quát: 3gluco-6phosphate + 6NADP+ + 3H2O → 2 fructo-6-phosphate + glyceraldehyde-3-phosphate

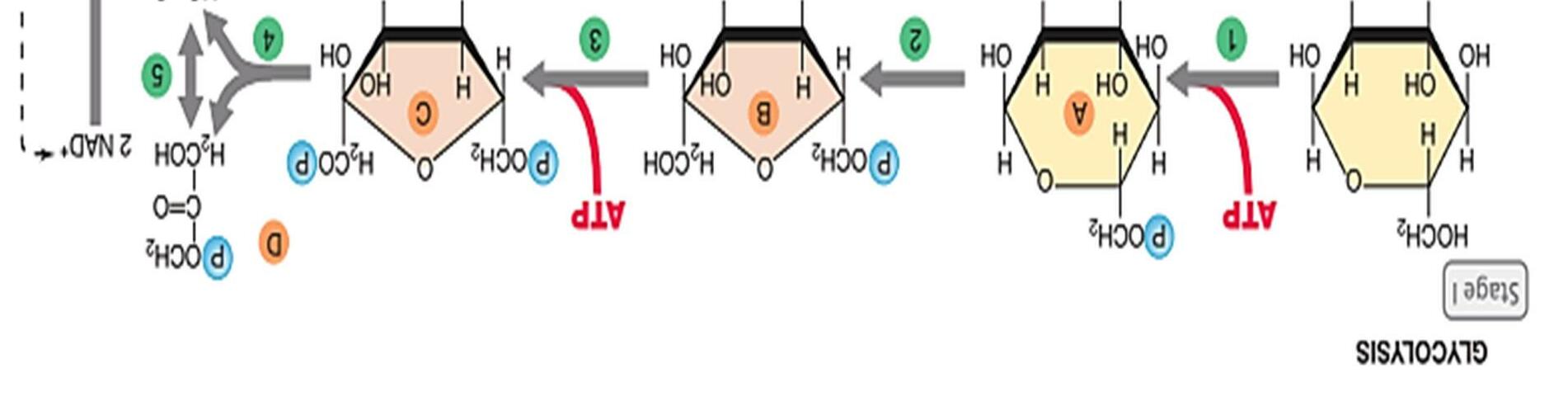
+ 3CO2 + 6 NADPH + 6H+
Các chất trung gian nói trên đượ
sử dụng trong hai con đường:
+ Fructose-6- phosphate có thể được chuyển trở lại thành glucose-6-phosphate, còn glyceraldehyde-3- phosphate được chuyển thành Pyruvate bởi các enzyme của đường phân.
+ Glyceraldehyde-3-phosphate cũng có thể trở lại con đường PP qua việc tạo thành glucose-6-phosphate. Điều này dẫn đến sự phân giải hoàn toàn glucose-6phosphate thành CO2 và tạo thành một lượng lớn NADPH.


Con đường PP có một số chức năng trong quá trình chuyển hóa vật chất:
- NADPH là nguồn cung cấp electron cho việc khử các phân tử trong sinh tổng hợp.
- Con đường tổng hợp các đường 4 C và 5 C dùng vào một số mục đích:
+ Đường erytrose-4-phosphate được dùng để tổng hợp các acid amin thơm và vitamin B6 (piridoxal).
+ Ribose-5-phosphate là thành phần chủ yếu của các acid nucleic và ribulose1,5-diphosphate (chất nhận CO2 đầu tiên trong quang hợp).
Khi một vi sinh vật đang sinh trưởng trên một nguồn carbon là pentose, con đường này cũng có thể cung cấp carbon cho việc tổng hợp hexose.
- Các chất trung gian trong con đường PP có thể được dùng để tạo thành ATP.
Vì các đường 5C là những chất trung gian, do đó con đường PP có thể được dùng để chuyển hoá pentose cũng như hexose.
Ở nhiều loài vi sinh vật, hai con đường EMP và PP là những con đường phổ biến và bổ sung cho nhau, cùng phát huy tác dụng trong tế bào. Con đường PP thường có vai trò quan trọng trong sinh tổng hợp. Mức độ hoạt động của mỗi con đường tùy thuộc vào trạng thái sinh trưởng của tế bào. Trong giai đoạn sinh trưởng mạnh mẽ nhất 2 con đường được sử dụng với tỉ lệ 2:1 (2 EMP: 1PP). Tỉ lệ sử dụng hai con đường này là không giống nhau ở các loài vi sinh vật khác nhau.
490
Hình 1.4. Con đường Pentozo phosphate.
* Con đường Enter-Doudoroff (ED hay KDPG)
Hầu hết vi khuẩn sử dụng các con đường đường phân và pentozo-phosphat nhưng một số lại sử dụng con đường Entner-Doudoroff thay cho đường phân. Con đường Entner-Doudoroff thường gặp ở các chi Pseudomonas, Rhizobium, Azotobacter, Agrobacterium và một vài chi vi khuẩn gram âm khác.
Con đường Entner-Doudoroff phân giải glucose thành pyruvate, 1 ATP, 1 NADH và 1 NADPH.


Hình 1.5. Con đường KDPG.
Do con đường Entner-Doudoroff không tạo thành các phosphate đường 5C và 4C nên tế bào vẫn cần sự hoạt động đồng thời của cả con đường PP.
Mặc dù một phần năng lượng có thể thu được từ sự phân giải glucose thành pyruvate qua các con đường nói trên, tuy nhiên phần lớn năng lượng nằm trong các NADH và nhất là pyruvate. Số phận của Pyruvate như thế nào là tùy thuộc vào hệ enzim của vi sinh vật. Nếu ở các vi sinh vật hiếu khí thì pyruvate sẽ tiếp tục tham gia vào chu trình Krebs và bị phân giải hoàn toàn.
1.3.2.2. Chu trình Krebs (Tricacboxylic)
Muốn tham gia vào chu trình Krebs, axit pyruvic phải được oxi hóa thành
Acetyl CoA nhờ phức hợp đa enzyme pyruvate dehydrogenase theo PTTQ như sau: CH3COCOOH + CoA-SH + NAD+ → CH3CO-SCoA + NADH + CO2



CoA là chất có nguồn gốc từ vitamin B. Acetyl - CoA là một phân tử cao năng
bao gồm coenzyme A và acid acetic nối với nhau qua liên kết cao năng tiol este.
Acetyl CoA trực tiếp tham gia vào chu trình Krebs (Hình 1.6).

Chuỗi vận chuyển electron được nghiên cứu kỹ nhất là chuỗi ở ti thể (Hình 1.7)
[1]. Chuỗi bao gồm một dãy các chất mang electron hoạt động phối hợp với nhau để vận chuyển electron từ các chất cho như NADH và FADH2 tới O2.
1. Oxaloacetate có thể đượctạoratừ 1 hợp chất 3 cacbon kếthợpvớiCO2



2. Acetyl CoA chuyển nhóm acetyl cho oxaloacetate tạo ra citrate (hợp chất có 6 cacbon.
3. Qua một loạt các phản ứng oxy hóa khử và biến đổi, hợp chất sáu cacbon cuối cùng được chuyển đổi trở lại bốn carbon (oxaloacetate). Bắt đầu một chu trìnhkhácvớisự bổ sung phân tử acetyl-CoA tiếp theo.
Hình 1.6. Các phản ứng cơ bản của chu trình Crep.
Chu trình Krebs gắn bó chặt chẽ với các quá trình đồng hóa, nó cung cấp những chất ban đầu để xây dựng protein, lipit và gluxxit. Ngược lại, khi cần hoặc nếu dư thừa thì các chất này lại có thể tham gia vào chu trình Krebs.
1.3.2.3. Chuỗi vận chuyển electron hô hấp và quá trình phosphoryl hóa oxi hóa


Khi một phân tử glucose trải qua con đường đường phân và chu trình Krebs chỉ khoảng 4 phân tử ATP được tạo thành, còn hầu hết ATP thu được là từ sự oxy hoá NADH và FADH2 trong chuỗi vận chuyển electron.
* Chuỗi vận chuyển electron hô hấp
493
Các electron di chuyển từ các chất mang có thế khử âm hơn tới các chất mang có thế khử dương hơn, cuối cùng kết hợp với O2 và H+ tạo thành nước. Các electron vận chuyển xuôi theo gradien thế năng tương tự như nước chảy xuôi qua một dãy thác ghềnh. Sự khác nhau trong thế khử giữa O2 và NADH là khoảng 1,14V thuận lợi cho
Hình 1.7. Chuỗi vận chuyển electron ở ty thể.
việc giải phóng một lượng lớn năng lượng. Sự thay đổi thế năng ở một số điểm trong chuỗi đủ lớn để cung cấp năng lượng cho việc tạo thành ATP tương tự như năng lượng từ thác nước chảy xuống các bánh xe dùng để sản xuất điện. Chuỗi vận chuyển electron đã phân tách toàn bộ năng lượng thoát ra thành các bước nhỏ [2].
Các chất mang trong chuỗi vận chuyển electron nằm trên màng trong của ti thể hoặc màng sinh chất ở vi khuẩn vì:
+ Các chất này cần ở gần nhau để thuận tiện cho việc chuyền các electron cho nhau.
+ Mục đích của chuỗi là bơm H+ qua màng để tạo thành gradient H+, do đó nếu không có màng cũng sẽ không có gradient. Chuỗi vận chuyển ở ti thể được sắp xếp thành 4 phức hợp các chất mang, mỗi phức hợp có thể vận chuyển một phần của các electron trên con đường tới O2. CoQ và Cytochrome c liên kết các phức hợp với nhau.
Mặc dù chuỗi hô hấp ở một số vi khuẩn tương tự như ở ti thể nhưng nói chung chúng rất khác nhau [2]:
+ Các chất mang electron thường không giống nhau và các chuỗi vận chuyển electron ở vi khuẩn có thể phân nhánh mạnh mẽ. Các electron thường có thể đi vào chuỗi ở một số điểm và rời khỏi chuỗi qua một số oxydase tận cùng.
+ Số Cytochrome có thể hoạt động, bản chất của các Cytochrome và đặc biệt là sự có mặt hay vắng mặt của 1 loại Cytochrome C.
+ Bản chất của Cytochrome cuối cùng, loại Cytochrome đảm bảo vận chuyển electron đến O2: Cytochrome O ở E. Coli, Cytochrome a3 ở Paracoccus denitrificans,...

+ Các chuỗi ở vi khuẩn cũng có thể ngắn hơn và có tỉ lệ P/O (thể hiện số ATP có thể được tạo thành từ ADP và Pi mỗi khi một cặp electron chuyển từ NADH hoặc FADH2 tới O2) thấp hơn các chuỗi ở ti thể. Như vậy, các chuỗi vận chuyển electron ở sinh vật nhân sơ và sinh vật nhân thật khác nhau về cấu trúc chi tiết mặc dù chúng đều hoạt động theo các nguyên tắc cơ bản như nhau.
Ở E. coli, các electron cũng được vận chuyển từ NADH tới các chất nhận và vận chuyển các proton qua màng sinh chất nhưng chuỗi vận chuyển ở E. coli hoàn toàn khác với ở ti thể [2].
+ Chuỗi ở E. coli phân nhánh và chứa các Cytochrome rất khác nhau. CoQ hoặc ubiquinol cung cấp các electron cho cả hai nhánh nhưng chúng hoạt động dưới các điều kiện sinh trưởng khác nhau.
+ Nhánh Cytochrome d có ái lực rất cao đối với oxy và hoạt động ở nồng độ oxy thấp. Nhánh này hoạt động kém hiệu quả hơn nhánh Cytochrome o vì không chủ động bơm proton. Nhánh Cytochrome o có ái lực cao trung bình đối với oxy, là một bơm proton và hoạt động ở nồng độ oxy cao hơn.
Paracoccus denitrificans là một vi khuẩn đất, gram âm, kị khí không bắt buộc, có thể sinh trưởng dị dưỡng với hàng loạt chất dinh dưỡng hoặc tự dưỡng với H2 và CO2 nhờ NO3 là chất nhận electron. Chuỗi vận chuyển electron hiếu khí gồm 4 phức hợp như ở ti thể (Hình 1.8). Ngoài các chất cho như NADH, succinat vi khuẩn nói trên còn oxy hoá metanol, metylamin như nguồn carbon duy nhất cho sinh trưởng. Các electron đi vào chuỗi truyền electron ở vị trí Cytochrome c. Metanol bị oxy hoá thành formaldehit, chất này được chuyển thành CO2 và đi vào chu trình Calvin. Khi vi khuẩn sinh trưởng kỵ khí với NO3− là chất nhận electron chuỗi sẽ được sắp xếp hoàn toàn khác [6].
Hình 1.8. Chuỗi truyền electron ở Paracoccus denitrificans, vi sinh vật được chọn là hình mẫu cho các nghiên cứu về hô hấp tế bào.
* Quá trình phosphoryl hóa oxi hóa
Cơ chế của phosphoryl hoá oxy hoá được chấp nhận rộng rãi theo giả thuyết hoá thẩm thấu (chemiosmosis) do nhà sinh hoá người Anh Peter Mitchell đề xuất đầu tiên vào năm 1951. Theo giả thuyết này, chuỗi vận chuyển electron được sắp xếp sao cho các proton được đẩy từ chất nền ti thể ra phía ngoài, còn các electron thì được vận chuyển bên trong chuỗi (hình 1.9). Kết quả là xuất hiện một động lực proton (proton motive force, PMF) bao gồm một gradient proton và một thế hiệu màng do sự phân bố không đều của điện tích. Khi các proton di chuyển trở lại chất nền ti thể nhờ PMF, ATP sẽ được tổng hợp [2].
Ở vi khuẩn cũng diễn ra quá trình tương tự: dòng electron tạo điều kiện cho các proton di chuyển ra phía ngoài qua màng sinh chất. Sau đó, ATP được tổng hợp khi các proton này khuếch tán trở lại tế bào. PMF cũng có thể hướng dẫn vận chuyển các phân tử qua màng cũng như sự quay của tiên mao nghĩa là đóng vai trò trung tâm trong sinh lý học của vi khuẩn. Giả thuyết hoá thẩm thấu được chấp nhận bởi hầu hết các nhà vi sinh vật học. Tuy nhiên, ở một số vi khuẩn biển ưa mặn các ion natri có thể được dùng để hướng dẫn tổng hợp ATP [2].
495
Hình 1.9. Hóa thẩm gắn kết chuỗi chuyền electron với sự tổng hợp ATP.
Ở nồng độ đủ cao, nhiều hoá chất kìm hãm tổng hợp ATP trong điều kiện hiếu khí và thậm chí có thể giết chết tế bào. Các chất này có thể được phân thành hai loại [2]:
+ Một số ngăn cản trực tiếp việc vận chuyển electron như: chất kháng sinh pierixidin cạnh tranh với CoQ; chất kháng sinh antimixin A cản trở việc vận chuyển electron giữa các Cytochrome b và c; xianit và azit làm ngừng việc vận chuyển electron giữa Cytochrome a và O2 do chúng có cấu trúc tương tự với O2.
Khi chuyển từ điều kiện kỵ khí sang hiếu khí, nhiều vi sinh vật giảm mạnh tốc độ phân giải đường và chuyển sang hô hấp hiếu khí. Hiện tượng này được gọi là hiệu
ứng Pasteur. Hiệu ứng này rất có lợi cho vi sinh vật vì chỉ cần phân giải một lượng đường ít hơn mà vẫn thu được sản lượng ATP như nhau.
1.3.2.4. Hô hấp kị khí
Nhiều vi khuẩn có các chuỗi vận chuyển hoạt động với các chất nhận electron là các phân tử vô cơ từ bên ngoài khác O2. Quá trình này được gọi là hô hấp kỵ khí. Các chất nhận electron chủ yếu là nitrate, sulfate và CO2, ngoài ra cũng có thể có các kim loại và một số phân tử hữu cơ [3].
Một số vi khuẩn có thể sử dụng nitrate làm chất nhận electron và tổng hợp ATP. Nitrate có thể bị khử thành nitrite bởi nitrate – reductase (enzyme thay thế cho Cytochrome oxydase) theo phương trình: NO3- + 2e + 2H+ → NO2- + H2O

Tuy nhiên, sự khử nitrate thành nitrite không phải là con đường thu nhận ATP có hiệu quả, nitrite tạo thành lại rất độc. Vì vậy nitrate thường bị khử tiếp thành N2 (sản phẩm không độc) trong quá trình gọi là phản nitrate hóa theo phương trình tổng quát:
2NO3 - + 10e- + 12H+ → N2 + 6H2O
Phản nitrate hoá là một quá trình nhiều bước bao gồm 4 enzyme tham gia: nitrate - reductase, nitrite- reductase, oxyt nitric- reductase và oxyt nitrơ – reductase: NO3→ NO2 - → NO → N2O →N2
Ở động vật có vú, NO tác dụng như một chất dẫn truyền thần kinh đóng vai trò điều chỉnh huyết áp và được các đại thực bào sử dụng để tiêu diệt vi khuẩn cũng như các tế bào u.
Quá trình phản nitrate hóa được nghiên cứu kỹ ở vi khuẩn đất, gram âm Paracoccus denitrificans. Chuỗi vận chuyển electron ở chúng chứa nitrate - reductase và oxyt nitric - reductase gắn vào màng còn nitrite reductase và oxyt nitrơ - reductase thì tồn tại trong chu chất. 4 enzyme sử dụng các electron từ CoQ và các Cytochrome C để khử nitrate và sản ra PMF.
+ Một nhóm chất kìm hãm khác gọi là chất cách li (uncoupler) có tác dụng đình chỉ tổng hợp ATP nhưng không ức chế việc vận chuyển electron. Thông thường, sự vận chuyển electron liên kết chặt chẽ với phosphoryl hoá oxy hoá sao cho tốc độ tổng hợp ATP điều hoà tốc độ vận chuyển electron. Tổng hợp ATP diễn ra càng nhanh thì chuỗi vận chuyển electron hoạt động cũng càng nhanh. Các chất cách li tách riêng phosphoryl hoá oxy hoá khỏi vận chuyển electron. Vì vậy, năng lượng do chuỗi thải ra sẽ ở dạng nhiệt chứ không phải ATP. Nhiều chất cách li như dinitrophenol và valinomixin có thể cho phép các ion H+, K+ và các ion khác qua màng mà không hoạt hoá ATPsynthase. Kết quả là các gradient pH và ion bị phá huỷ. Valinomixin cũng có thể liên kết trực tiếp với ATPsynthase và kìm hãm hoạt tính của enzyme này. 498
Phản nitrate hoá gặp ở một số loài thuộc các chi Pseudomonas, Paracoccus và Bacillus. Chúng sử dụng con đường này thay cho hô hấp hiếu khí bình thường và có thể được xem là các vi khuẩn kỵ khí tuỳ tiện. Khi O2 có mặt chúng thực hiện hô hấp hiếu khí (tổng hợp nitrate reductase bị kiềm chế bởi O2). Trong đất kỵ khí, phản nitrate hoá làm tổn thất nitơ của đất và ảnh hưởng xấu đến độ phì của đất.
Hai nhóm vi khuẩn chủ yếu khác sử dụng hô hấp kỵ khí là các vi khuẩn kỵ khí bắt buộc:
+ Nhóm vi khuẩn sử dụng CO2 hoặc carbonat làm chất nhận electron cuối cùng
được gọi là các vi khuẩn sinh metan vì chúng khử CO2 thành metan (CH4).
+ Vi khuẩn Desulfovibrio sử dụng Sulfate làm chất nhận electron cuối cùng.
Sulfate bị khử thành sulfua (S2- hoặc H2S) và 8 electron được tiếp nhận.
SO4 2- + 8e- + 8H+ → S2- + 4H2O
So với hô hấp hiếu khí thì hô hấp kỵ khí có hiệu quả tổng hợp ATP thấp hơn. Vì
các chất nhận electron có sức hút với electron thấp hơn O
2. Tuy nhiên, hô hấp kỵ khí là có ích vì có hiệu quả hơn lên men và cho phép tổng hợp ATP nhờ vận chuyển electron và phosphoryl hoá oxy hoá trong điều kiện vắng mặt O2. Hô hấp kỵ khí chiếm ưu thế trong đất và các bùn kỵ khí.
Nếu trong môi trường đặc biệt tồn tại O2, nitrate, ion mangan, ion ferric (Fe3+), sulfate và CO2 ta sẽ thấy diễn ra thứ tự sử dụng chất nhận electron như sau [2]:
+ Ở các vi sinh vật có khả năng hô hấp với O2 và nitrate, oxy được sử dụng trước tiên vì nó kìm hãm việc sử dụng nitrate.
+ Các vi khuẩn khử sulfate và vi khuẩn sinh metan bị kìm hãm khi có mặt O2 vì chúng là bọn kỵ khí bắt buộc.
+ Khi O2, nitrate bị cạn kiệt và các sản phẩm lên men, kể cả hydro được tích luỹ thì sẽ bắt đầu diễn ra sự cạnh tranh sử dụng các chất nhận electron khác: Mangan và sắt sẽ được sử dụng đầu tiên. Tiếp theo, là sự cạnh tranh của vi khuẩn sử dụng sulfate và vi khuẩn sinh metan. Hiệu quả năng lượng sẽ cao hơn khi sulfate là chất nhận electron. Vi khuẩn khử sulfate (Desulfovibrio) sẽ sinh trưởng nhanh và sử dụng hydro sẵn có với tốc độ nhanh hơn vi khuẩn sinh metan (Methanobacterium)
+ Khi sulfate cạn kiện, Desulfovibrio không oxy hoá hydro nữa và nồng độ hydro tăng lên. Cuối cùng, vi khuẩn sinh metan chiếm ưu thế và khử CO2 thành CH4
Bên cạnh quá trình phosphoryl hóa oxy hóa cơ chất, còn tồn tại những con đường khác, đặc biệt là con đường oxy hóa trực tiếp nhờ các enzim vận chuyển các electron từ cơ chất đến O2 với sự tạo thành ion superoxide.


FADH2 + O2 → FAD + H2O2 (với sự xúc tác của FAD-oxydase)
Những con đường này cho rất ít hoặc không cho năng lượng hữu ích. Bên cạnh đó, H2O2 rất độc và có thể làm chết tế bào, do đó nó cần được phân giải ngay bởi enzim catalase, peroxidase hoặc superoxide dismutase.
Ngoài ra, trong quá trình khử O2 tạo thành H2O cũng sinh ra các sản phẩm phụ có hại cho tế bào và cần có enzim phân giải (Hình 1.10) [6].
Hình 1.10. Các sản phẩm gây hại cho tế bào sinh ra từ quá trình khử O2 và các enzyme phân giải chúng.
Vì vậy, những vi khuẩn không có các enzim như trên thì bắt buộc phải sống kỵ khí, vì O2 là chất gây độc đối với chúng. Có một số vi khuẩn chịu được hiếu khí như Streptococcus nhờ có các enzim flavin (NAD-oxydase và NAD-peroxydase). Người ta cũng dựa vào số và lượng các enzim có thể phân giải H2O2 để quyết định tính hiếu khí hay kị khí ở các vi khuẩn [3].
1.3.2.5. Lên men
Khi vắng mặt hô hấp hiếu khí hoặc kỵ khí, NADH không bị oxy hoá bởi chuỗi vận chuyển electron do thiếu chất nhận electron từ bên ngoài. Tuy nhiên, NADH tạo thành trong con đường đường phân vẫn cần phải được oxy hoá trở lại thành NAD+ . Nếu NAD+ không được tái tạo việc oxy hoá glyceraldehyde-3-phosphate sẽ không diễn ra và đường phân sẽ ngừng hoạt động. Vì vậy, chức năng chủ yếu của lên men là tái tạo NAD+ cho đường phân [1]. Để khắc phục tình trạng trên nhiều vi sinh vật đã giảm hoặc làm ngừng hoạt tính của enzyme pyruvate dehydrogenase và sử dụng pyruvate hay một trong các chất dẫn xuất của pyruvate như chất nhận electron và hydro nhằm oxy hoá NADH trong một quá trình gọi là lên men [2].
Trong lên men vi sinh vật cần chú ý hai điểm:
+ NADH được oxy hoá thành NAD+
+ Chất nhận electron thường là pyruvate hoặc một chất dẫn xuất từ pyruvate như acetaldehyde.
Trong quá trình lên men, cơ chất bị oxy hoá một phần, ATP chỉ được tạo thành bởi phosphoryl hoá ở mức độ cơ chất và O2 là không cần thiết. Vi sinh vật thực hiện
499
nhiều quá trình lên men khác nhau nhưng trong phạm vi của chuyên đề này, chúng tôi chỉ giới thiệu hai quá trình lên men thường gặp nhất là lên men etylic và lên men lactic.
* Lên men etylic
Rượu etylic là một trong số các sản phẩm lên men phổ biến nhất gặp ở vi sinh vật. Gặp ở cả nấm, vi khuẩn, tảo, động vật nguyên sinh. Vi sinh vật sản sinh rượu etylic chủ yếu là nấm men, đặc biệt là các chủng thuộc Saccharomyces cerevisiae [2].
Quá trình lên men etylic được mô tả tóm tắt ở Hình 1.11 sau đây [1]:
chứng minh sữa chua là kết quả hoạt động của một nhóm vi sinh vậ
sinh vật lên men lactic.
Lên men lactic là quá trình chuyển hóa kỵ khí các hợp chất đường thành acid lactic và một số sản phẩm khác (Hình 1.12) [1].
Hình 1.11. Sơ đồ tóm tắt quá trình lên men ethanol ở vi sinh vật.
Nếu trong môi trường lên men có chất dihydrosulfit, sẽ làm giảm lượng ethanol tạo ra. Carl Neuberg đã nhận thấy chất này tuy không độc đối với nấm men nhưng khi thêm vào môi trường chứa glucose đang được nấm men lên men thì acetaldehyde sẽ bị kết tủa do tạo thành phức chất:
CH3-CHO + NaHSO3 → CH3-CHOH-SO3Na
Lúc này, glyceryl sẽ xuất hiện như một sản phẩm lên men mới, đồng thời hiệu suất ethanol và CO2 sẽ giảm xuống. Trên thực tế, quá trình lên men với sự có mặt của dihydrosulfit (hay còn gọi là lên men Neuberg) đã được sử dụng trong công nghiệp để sản xuất glyceryl.
Một nguyên nhân khác làm sụt giảm lượng ethanol tạo thành là do sự cạnh tranh của NADH giữa quá trình hô hấp hiếu khí với quá trình lên men (hiệu ứng Pasteur) [3].
* Lên men lactic
Con người từ lâu đã sử dụng vi sinh vật lên men lactic để bảo quản và chế biến các loại thức ăn, đặc biệt là thức ăn có nguồn gốc từ sữa. Năm 1857, L. Pasteur đã
Hình 1.12. Sơ đồ tóm tắt quá trình lên men lactic đồng hình.
Những vi khuẩn lên men lactic rất đa dạng, có thể thuộc nhóm Streptococcus, Leuconostoc, Pedicoccus, Lactobacillus, Bifidobacterium… Có 2 kiểu lên men lactic ở vi sinh vật là: lên men lactic đồng hình và lên men latic dị hình [2].
- Lên men lactic đồng hình là quá trình lên men trong đó sản phẩm acid lactic chiếm đa số (trên 90%). Trong quá trình lên men lactic đồng hình, glucose được chuyển hóa theo chu trình Embden-Mayerhoff tạo thành acid pyruvic. Vì trong vi khuẩn lên men lactic đồng hình không có enzyme carboxylase nên acid pyruvic không phân hủy nữa mà tiếp tục khử thành acid lactic.
- Lên men lactic dị hình là quá trình lên men trong đó sản phẩm tạo ra ngoài acid lactic (40%) thì còn một lượng đáng kể các sản phẩm phụ như acid acetic (10%), ethanol (10%), acid succinic (20%), CO2… đôi khi không có các khí mà thay vào đó là sự tích lũy một lượng acid formic.
Vi khuẩn lên men lactic dị hình không có các enzyme cơ bản của chu trình Embden-Mayerhoff là aldolase triozophosphatizomeras, vì vậy, chúng đi theo con đường Pentose phosphate để tạo thành 2 phân tử: 3-phospho glyceraldehyde và phospho acetyl. 3-phospho glyceraldehyde sẽ qua một số phản ứng lên men đồng hình để chuyển thành acid lactic, còn phospho acetyl sẽ chuyển thành ethanol và có thể chuyển thành acid acetic. Do đó, trong lên men lactic dị hình ngoài acid lactic còn có thêm các sản phẩm phụ khác.


Lên men lactic được ứng dụng để muối chua rau củ, làm sữa chua, ủ chua các loại cây cỏ dùng trong chăn nuôi để phân giải một số polysaccharide và protein thành các đoạn ngắn hơn, đồng thời bổ sung nhiều loại vitamin do vi khuẩn tổng hợp nên.
Bộ gene của E. coli là một phân tử DNA sợi kép vòng có kích thước khoảng 4.639.221 bp, với 4290 gene mã hóa protein cộng với 53 gene RNA. Thường tập trung ở "vùng nhân", không có màng nhân bao bọc, và ở trạng thái siêu xoắn dưới sự kiểm soát của các topoisomerase
Mỗi bộ gene có khoảng 4,6 triệu cặp bazơ với khoảng 100 vòng siêu xoắn; mỗi vòng chứa khoảng 40 ngàn cặp
Hình 2.1. Bộ gen của E. coli K-12.
bazơ. Ngoài ra, còn có nhiều phân tử DNA sợi kép vòng khác có kích thước bé phân bố rải rác trong tế bào chất, gọi là các plasmid [7].
Hình 1.13. Một số quá trình lên men cơ bản và thực phẩm lên men tương
ứng.
2. HỆ THỐNG LÝ THUYẾT TÍCH HỢP VI SINH – DI TRUYỀN
Đầu thập niên 1940, các vi sinh vật trở thành đối tượng nghiên cứu chính yếu của di truyền học. Từ đây, hình thành các lĩnh vực di truyền học hoá sinh và di truyền học vi sinh vật, hai nền tản chính cho sự ra đời của di truyền học phân tử (1953) và công nghệ ADN tái tổ hợp (1978) [2].
Có thể nói, từ khi ra đời cho đến nay, sự phát triển vô cùng nhanh chóng của di truyền học vi sinh vật, đặc biệt là sự tiến bộ của công nghệ sinh học đã có những tác động mạnh mẽ lên nhiều ngành khoa học và trên mọi mặt của đời sống, kinh tế, chính trị và xã hội toàn cầu.
Di truyền vi sinh vật là lĩnh vực rất rộng, vì vậy trong phạm vi của chuyên đề này, chúng tôi chỉ tập trung vào hai phần là cơ sở phân tử của di truyền và điều hòa biểu hiện gen của vi khuẩn. Đây là hai nội dung thường xuyên xuất hiện trong đề thi học sinh giỏi các cấp và Olimpic, đồng thời cũng đóng góp thêm tài liệu tham khảo vào mảng di truyền mà các chuyên đề tham gia Hội thảo năm 2019 đã viết.
2.1 Tổ chức phân tử của các nhiễm sắc thể vi khuẩn
2.1.1. Tổ chức bộ gene của các vi khuẩn Nhóm prokaryote bao gồm các vi khuẩn (bacteria) và vi khuẩn cổ (archaeobacteria) là các sinh vật có tổ chức bộ gen đơn giản.
Các plasmid là những mẫu ADN nhỏ, ngắn, dạng vòng (khép kín), sợi đôi nằm ngoài nhiễm sắc thể, được tìm thấy đầu tiên trong tế bào một số vi khuẩn. Chúng sao chép được là nhờ một số enzyme có mặt trong tế bào vi khuẩn và không phụ thuộc vào sự sao chép nhiễm sắc thể vi khuẩn. Tùy các kiểu của plasmid mà số bản sao plasmid bởi vi khuẩn sẽ khác nhau. Một số plasmid chỉ có một bản sao duy nhất vì chúng tự tái bản chỉ một lần trong mổi lần phân bào.



Một số khác có số bản sao lớn vì chúng tái bản được nhiều lần trong mỗi chu kỳ phân bào. Những plasmid có từ 10 đến 100 bản sao trong tế bào chủ được xem như plasmid có bản sao cao. Plasmid khác có từ 1 đến 4 bản sao trong tế bào chủ, được xếp vào nhóm có bản sao thấp. Trong sinh vật eucaryote, plasmid chỉ có trong tế bào nấm men.
Mỗi plasmid đều có một chuỗi mã di truyền (sequence) mang chức măng tự tái bản ADN. Nếu không có vị trí khởi đầu phiên mã (ori) này, ADN không thể tự tái lập trong tế bào chủ. Với tính chất tự tái bản, plasmid là một vector chuyển gen để nhân dòng ADN cần thiết [7].
Do kích thước nhỏ nên plasmid chỉ chứa rất ít gen chọn lọc, thường đặc tính chọn lọc là kháng kháng sinh. Từ khi được phát hiện đến nay, các plasmid không ngừng được cải tiến và ngày càng được có thêm nhiều đặc tính quí cho việc tạo dòng. Trong phòng thí nghiệm, người ta sử dụng các plasmid nhân tạo mà được tạo ra từ các plasmid tự nhiên và cài thêm một số chuỗi ADN.
pBR322 có kích thước 4.363bp và hai gen kháng thuốc, một chống chịu được ampicilline (ampR) và một chống chịu được tetracycline (tetR).
Có nhiều điểm nhận biết bởi enzyme cắt hạn chế và trong số đó có nhiều điểm nhận biết nằm trong gen kháng kháng sinh. Ví dụ, gen kháng ampicilline có ba trình tự nhận biết bởi ba enzyme cắt hạn chế là PstI, PvuI, ScaI. Còn gen kháng
Hình 2.2. Cấu tạo plasmid pBR322.

tetracycline có sáu điểm nhận biết là: EcoRV, BamHI, SphI, SlaI, XmaIII, NnuI.
Việc cài ADN lạ được tiến hành một trong hai gen kháng thuốc. Nếu chỉ còn một gen kháng với kháng sinh là do một trong hai gen đã không nhận đoạn cài. Điều này cho phép để chọn lựa các plasmid tái tổ hợp. Một ứng dụng nữa của pBR322 là chúng có số bản sao lớn. Thực nghiệm cho thấy cứ 15 plasmid tái tổ hợp được biến nạp trong E. Coli, số lượng này có thể tăng lên từ 1.000 đến 3.000 bản sao trong điều kiện nuôi cấy tốt [6].
2.1.2. Kích thước bộ gene các prokaryote
Trước đây, người ta cho rằng tất cả các bộ gene prokaryote là bé hơn nhiều các bộ gene eukaryote. Tuy nhiên, việc áp dụng các kỹ thuật mới để xây dựng các bản đồ vật lý và xác định trình tự đầy đủ bộ gene đã chỉ ra rằng có một sự đa dạng rất lớn trong kích thước và tổ chức của các bộ gene prokaryote. Kích thước nhiễm sắc thể của các vi khuẩn biến thiên từ 0,6 Mbp đến 10 Mbp, còn kích thước nhiễm sắc thể của các Archae biến thiên từ 0,5 Mbp đến 5,8 Mbp [7].
Bộ gene của vi khuẩn cổ bé nhất được xác định cho đến nay là từ Nanoarchaeum equtans, một thể cộng sinh bắt buộc nhỏ nhất có kích thước bộ gene là 491 Kbp. Sinh vật này không có các gene cần thiết cho tổng hợp các lipid, các amino acid, các nucleotide và các vitamin, và như thế chúng phải sinh trưởng bằng cách kết hợp chặt chẽ với sinh vật khác để được cung cấp các chất dinh dưỡng này.
Bộ gene vi khuẩn ngày nay bé nhất được xác định là từ Mycoplasma genitalium, một vi khuẩn gây bệnh ký sinh nội bào bắt buộc có kích thước bộ gene bằng 580 Kbp.
các khối kiến trúc cơ sở. Vì vậy, các vi sinh vật sống tự do bé nhất có kích thước bộ gene trên 1 Mbp.
Một đặc điểm thường được dùng để phân biệt các bộ gene của các prokaryote and eukaryote là số lượng DNA "rác" ("junk" DNA). Trên nguyên tắc chung, các prokaryote có xu hướng rất ít DNA dạng này (điển hình là chưa đến 15% của bộ gene) và các eukaryote thì có số lượng đáng kể DNA này [7].
2.1.3. Tái bản DNA (DNA replication)
2.1.3.1. Các nguyên tắc và đặc điểm chung của tái bản DNA
Trong khi khám phá ra cấu trúc DNA, Watson và Crick đã đưa ra dự đoán chính xác rằng sự tái bản DNA phải diễn ra theo kiểu bán bảo toàn (semiconservative). Các thí nghiệm sau đó ở E. coli của Meselson và Stahl (1958) và John Cairn (1961) đã nhanh chóng khẳng định điều dự đoán này. Dưới đây là các nguyên tắc chung của tái bản DNA [7].
- Tái bản theo kiểu bán bảo toàn (semi-conservative).
- Tái bản bắt đầu tại một hoặc nhiều khởi điểm (Ori). Từ khởi điểm, DNA mở xoắn tạo thành hai chạc tái bản (replication fork). Cấu trúc như vậy gọi là đơn vị tái bản (replicon). DNA E. coli trong quá trình tái bản như vậy có cấu trúc giống chữ cái theta Hy Lạp (θ) nên gọi là tái bản theta (Hình 2.3) [6]. Đối với các DNA mạch vòng, mỗi phân tử chỉ có một Ori; trong khi đó mỗi DNA nhiễm sắc thể eukaryote có nhiều Ori
- Tham gia vào sự tái bản DNA có nhiều protein và enzyme.
- Tại mỗi chạc tái bản, trước tiên xảy ra sự tổng hợp các đoạn mồi RNA (primer) bởi vì các DNA polymerase tự nó không thể bắt đầu tổng hợp mới được. Mặt khác, do hai sợi đơn của mỗi chạc ngược chiều nhau trong khi các DNA và RNA polymerase chỉ xúc tác theo chiều 3’ → 5', cho nên sự tái bản DNA trên hai sợi khuôn là không giống nhau: một sợi liên tục gọi là sợi dẫn đầu (leading strand) và một sợi không liên tục gọi là sợi ra chậm (lagging strand). Kiểu tái bản như vậy gọi là tái bản nửa gián đoạn (semi-discontinuous), được R. Okazaki phát hiện đầu tiên năm 1969.
Khác với Nanoarchaeum và Mycoplasma, các vi khuẩn sống tự do phải dành ra nhiều gene thực hiện các con đường sinh tổng hợp và vận chuyển các dưỡng chất và 506
Hình 2.3. Tái bản DNA và cấu trúc theta ở E. coli.
Tất cả các DNA polymerase đều cần có mồi để xúc tác tổng hợp chuỗi theo chiều 5'→3' và chỉ có một số enzyme này là có hoạt tính đọc sửa (proofreading activity). Khác với E. coli (có ba loại DNA polymerase I, II và III; trong đó Pol II không tham gia tái bản), các tế bào eukaryote có năm loại DNA polymerase (α, β, γ, δ và ε), trong đó ba loại chịu trách nhiệm tái bản DNA nhân là các polymerase α, δ và ε
Polymerase δ chịu trách nhiệm chính cho tổng hợp ở sợi dẫn đầu và polymerase α cho sợi ra chậm. Polymerase γ tái bản DNA của các bào quan ty thể, lục lạp; polymerase β chịu trách nhiệm sửa chữa DNA.


2.1.3.2. Các thành phần của bộ máy tái bản DNA ở E. coli.
Các enzim và protein tham gia tái bản DNA ở E. coli được trình bày ở bảng sau: [7], [6]
Thành phần Chức năng
Protein dnaA Bám trình tự DNA của điểm khởi đầu
protein dnaB (Helicase) Tháo xoắn DNA tại điểm khởi đầu
Protein Rep Helicase (mở xoắn DNA ở chạc tái bản)
Protein SSB Bám DNA sợi đơn
DNA polymerase III
DNA polymerase I
DNA ligase
Polymerase tái bản chính yếu: sử dụng mạch DNA “mẹ” làm khuôn, tổng hợp mạch DNA mới bằng cách bổ sung các nucleotit vào đầu 3’ của mạch DNA sẵn có hoặc đoạn mồi qua liên kết cộng hóa trị.
Tách bỏ mồi và thay thế bằng các nucleotit của DNA.
Nối đầu 3’ của đoạn DNA đã thay thế đoạn mồi với phần còn lại của mạch dẫn đầu hoặc nối các đoạn Okazaki của mạch ra chậm.
2.1.3.3. Cơ chế tái bản DNA (E. coli)
* Giai đoạn khởi đầu
Đối với nhiễm sắc thể E. coli, sự tái bản bắt đầu tại một khởi điểm đặc thù gọi là Ori. Quá trình diễn biến ở khởi điểm E. coli cho đến lúc hình thành hai chạc có thể tóm tắt như sau:
- Các protein bám khởi điểm dnaA bám Ori tạo ra cấu trúc nucleoprotein chuyên hoá của khởi điểm. Cấu trúc này mở xoắn vùng DNA giàu AT để hình thành "phức hợp mở".
- Hai phân tử helicase dnaB chui vào phức hợp mở làm mở xoắn khởi điểm theo cả hai hướng, tạo thành hai chạc tái bản (replication fork). Khi cả hai sợi đơn của mỗi chạc được tách ra thì các protein SSB bám vào.
Khởi đầu trong tái bản DNA ở E. coli là sự tổng hợp một đoạn mồi ngắn khoảng 10-12 nucleotide bởi primase (xét chung ở các sinh vật là ~ 5 base). Cả ba loại protein dnaB, dnaC và dnaG hợp thành một phức hợp có tên là primasome.
* Giai đọan kéo dài (elongation)
Khi primasome tổng hợp xong một mồi, sự kéo dài chuỗi DNA được bắt đầu bằng một phức hợp giữa primasome và DNA polymerase III hoàn chỉnh, gọi là replisome (thể tái bản). Sự tái bản DNA trên mỗi chạc xảy ra theo kiểu nửa gián đoạn
Primas ome
protein dnaC Bám protein dnaB
protein dnaG (Primase)
Tổng hợp RNA mồi
DNA gyrase Mở siêu xoắn ngược đằng trước mỗi chạc
Trên mạch dẫn đầu (3'→5'): Sau khi primase (primasome) tổng hợp xong đoạn mồi RNA với đầu 3'-OH tự do, enzyme DNA polymerase III bắt đầu kéo dài chuỗi DNA mới sinh trưởng theo chiều 5'→3' một cách liên tục.
Trên mạch ra chậm (5'→3'): Sự kéo dài mạch diễn ra không liên tục dưới dạng các đoạn Okazaki. Kích thước trung bình mỗi đoạn Okazaki ở E. coli là 1.000 - 2.000
nucleotide. Quá trình này đòi hỏi sự "mồi hóa" lặp lại, với sự tham gia lần lượt của bốn enzyme:
- Primase tổng hợp một đoạn mồi RNA.
- DNA polymerase III kéo dài đoạn Okazaki.
- DNA polymerase I vừa cắt bỏ đoạn mồi vừa lấp khoảng trống bằng cách kéo dài dần đoạn Okazaki theo sau.

- DNA ligase hàn liền khe hở giữa hai đoạn Okazaki bằng một liên kết phosphodiester.

* Giai đọan kết thúc (termination)
Cả hai chạc tái bản của DNA E. coli được bắt đầu từ một khởi điểm (ori), và di chuyển hầu như cùng tốc độ theo hai hướng đối lập nhau xung quanh nhiễm sắc thể mạch vòng cho tới khi chúng gặp nhau tại một điểm kết thúc chung đối diện với ori Đây là vùng chứa các trình tự đặc thù gọi là các điểm kết thúc tái bản. Tại các trình tự này, có các protein kết thúc tái bản (replication terminator protein = RTP) bám vào và các phức hợp protein-DNA này ngăn cản sự di chuyển của các chạc tái bản. Đó là bước đầu tiên của sự hoàn thành một vòng tái bản, và sau đó tách hai nhiễm sắc thể con rời ra một cách có trật tự nhờ xúc tác của topoisomerase IV
RTP của E. coli được mã hóa bởi gene tus và trong mỗi tế bào có khoảng 80 bản sao của protein này được duy trì hầu như ổn định trong suốt chu kỳ tế bào.
Diễn biến chính trong quá trình tái bản DNA ở prokaryote được trình bày ở Hình 2.4 [1].

Hình 2.4. Tóm tắt quá trình sao chép DNA ở vi khuẩn.
2.1.4. Phiên mã (Transcription) ở prokaryote

2.1.4.1. Polycistronic mRNA và Operon
Ở các prokaryote, các gene mã hóa cho các enzyme của một con đường trao đổi chất cụ thể thường tập hợp với nhau thành một cụm trên nhiễm sắc thể của vi khuẩn. Ví dụ: E. coli tổng hợp tryptophan từ một phân tử tiền chất qua một con đường gồm nhiều bước. Mỗi phản ứng của con đường này đều được xúc tác bởi một enzyme đặc hiệu; và 5 gene mã hóa tương ứng cho các tiểu đơn vị của những enzyme này tập hợp với nhau thành một cụm trên nhiễm sắc thể của vi khuẩn E. coli. Một promoter duy nhất được dùng chung cho cả 5 gene; nghĩa là, các gene này tập hợp lại thành một đơn vị phiên mã duy nhất. Như vậy, sản phẩm của phiên mã sẽ là một phân tử mRNA dài, chứa trình tự mã hóa cho cả 5 chuỗi polypeptide cấu tạo nên các enzyme tham gia vào con đường sinh tổng hợp tryptophan. Một mRNA như vậy được gọi là một polycistronic mRNA [1]
509
Một nhóm các gene liên quan về chức năng được ghép vào cùng một đơn vị phiên mã, dùng chung một “công tắc bật-tắt” như vậy gọi là một operon [1].
Trái lại, ở các eukaryote, các gene đồng nghĩa với đơn vị phiên mã
Hình 2.5. Cấu trúc của operon và polycistronic mRNA ở prokaryote.


(transcription unit) và hầu hết chúng được phiên mã dưới dạng monocistronic mRNA Ở các bộ gene eukaryote bậc cao, thường có một mối quan hệ phức tạp giữa gene và sản phẩm. Hầu hết các gene đều chứa các intron (các đoạn không mã hóa protein) nằm xen giữa các exon (các đoạn mã hóa protein). Các gene như vậy được gọi là gene phân đoạn, được Phillip Sharp phát hiện đầu tiên năm 1977 [7].
2.1.4.2. Các nguyên tắc và đặc điểm chung của phiên mã
Phiên mã là quá trình tổng hợp các RNA khác nhau từ thông tin di truyền chứa đựng trong DNA. Trừ các gene mã hóa protein trong các operon ở vi khuẩn, nói chung, các RNA mới được tổng hợp chỉ là các bản sao sơ cấp (primary transcript) gọi là các pre-RNA. Các pre-RNA này phải trải qua một quá trình sửa đổi để trở thành các RNA trưởng thành (mature) trước khi tham gia vào quá trình sinh tổng hợp protein của tế bào.
Quá trình phiên mã DNA các đặc điểm chung sau đây [7]:
(1) Diễn ra dưới tác dụng của các enzyme RNA polymerase.
(2) Chỉ một sợi đơn được dùng làm khuôn cho tổng hợp RNA, gọi là sợi khuôn, sợi mã hoá hay sợi có nghĩa (template/ coding / sense strand); còn sợi bổ sung được gọi là sợi đối khuôn, sợi không mã hoá hay sợi đối nghĩa (antitemplate/ non-coding / antisense strand).
(3) Phản ứng tổng hợp RNA diễn ra theo nguyên tắc bổ sung và được kéo dài theo chiều 5'→3', ngược với chiều của sợi khuôn.
(4) Nguyên liệu cho tổng hợp gồm: ATP, UTP, GTP và CTP.
(5) Sản phẩm của phiên mã là các RNA sợi đơn.
2.1.4.3. RNA polymerase và vùng khởi động (promoter) của prokaryote
Hình 2.6. Sự tương tác của RNA polymerase với một promoter của vi khuẩn.

Ở các prokaryote, RNA polymerase hoàn chỉnh là một phức hợp gồm nhân tố sigma (σ) và lõi enzyme nên gọi là RNA polymerase holoenzyme [6].
+ Nhân tố σ giúp RNA polymerase nhận biết và bám vào promoter để có thể bắt đầu phiên mã tại vị trí chính xác.
+ Lõi enzyme đóng vai trò xúc tác tổng hợp RNA.
Vùng khởi động (promoter) nói chung nằm kề trước gene và có chứa các trình tự đặc thù cho phép RNA polymerase nhận biết và bám vào. Trình tự quan trọng nhất của promoter là hộp TATA hay hộp Pribnow (Pribnow box) vị trí "-10" (Hình 2.5) [6]. Ngoài ra, còn có trình tự TTGACA ở vị trí ''-35'', gọi là đoạn nhận biết. Các vùng này được bảo tồn cao.
Khởi đầu và kết thúc phiên mã phụ thuộc vào các tín hiệu điều hoà là các trình tự DNA đặc thù nằm trước gen (vùng khởi động) và sau gene. 512
2.1.4.4. Các giai đoạn của quá trình phiên mã ở prokaryote
- Khởi đầu: RNA polymerase holoenzyme nhận biết và bám vào promoter, tháo xoắn một đoạn khoảng 12 cặp base. Sau khi tổng hợp ribonucleotide đầu tiên, nhân tố sigma tách ra để đi vào một chu kỳ phiên mã khác, gọi là chu kỳ sigma (sigma cycle) (Hình 2.6).
- Kéo dài: Enzyme lõi tổng hợp sợi RNA dọc theo sợi khuôn.
- Kết thúc: Khi phiên mã xong hai vùng giàu GC và AT nằm sau gene, ở vùng đuôi của sợi RNA hình thành cấu trúc ''kẹp tóc'' (stem-loop) (Hình 2.7) làm dừng sự phiên mã của lõi RNA polymerase. Cuối cùng,
Hình
dưới tác dụng của nhân tố rho (ρ), sợi RNA và enzyme lõi được giải phóng khỏi DNA khuôn.

2.1.5. Dịch mã (translation)
Trong quá trình dịch mã, tế bào tiến hành “phiên dịch” thông điệp di truyền trên phân tử mRNA thành chuỗi polypeptide tương ứng. Thông điệp di truyền là chuỗi các bộ ba nucleotide trên mRNA, “phiên dịch viên” là các phân tử tRNA, nguyên liệu là các amino acid và nhà máy tổng hợp protein là các ribosome [6].
2.1.5.1. “Phiên dịch viên” RNA vận chuyển (tRNA)
Giống với các loại RNA khác trong tế bào, các phân tử RNA vận chuyển được phiên mã từ mạch khuôn DNA. Các tRNA có hai chức năng chính là mang amino acid và đọc mã trên mRNA. Có 86 tRNA ở E. coli. Hầu hết các tRNA có khoảng 75-80 nucleotide và có cấu trúc bậc hai mở rộng do các tương tác cặp base (A-U và G-C) ở một số đoạn của chúng cũng như cấu trúc bậc ba (không phải dạng siêu xoắn, mà nó có kiểu uốn gập thêm lần nữa trong không gian ba chiều) [7].
Hình 2.9. Cấu trúc của một tRNA.
Trong thành phần nucleotide của các tRNA có khá nhiều base hiếm tập trung ở các vòng thân như: 5',6'-dihydrouridine (DHU), inosine (I), ribothymidine (T), pseudouridine (Ψ),...

Các phân tử tRNA thường rất giống nhau ở nhiều đoạn và khác nhau chủ yếu ở bộ ba đối mã (anticodon). Mỗi tRNA thường có 3-4 vòng trên thân (tính từ đầu 5') với chức năng khác nhau như sau [6]:
- Vòng D (D loop) nhận biết aminoacyl-tRNA synthetase.


- Vòng anticodon đọc mã mRNA bằng sự kết cặp anticodon-codon.
- Vòng "phụ" (extra loop) có thể không có ở một số tRNA.
- Vòng TΨC nhận biết ribosome để đi vào đúng "vị trí A" .
- Và cuối cùng là đoạn mạch thẳng -CCA ở đầu 3': vị trí gắn vào của amino acid đã được hoạt hoá để tạo thành aminoacyl-tRNA.
Sự dịch mã chính xác từ mRNA đến protein được quyết định bởi hai quá trình đều dựa trên cơ chế nhận biết phân tử [1].
513
514
2.7. Phiên mã ở Prokaryote. Hình 2.8. Hình thành cấu trúc kẹp tóc và kết thúc phiên mã ở prokaryote.- Đầu tiên là tRNA liên kết với codon trên mRNA nhất định phải vận chuyển tới ribosome đúng loại amino acid mà codon đó mã hóa. Sự bắt cặp chính xác giữa tRNA và amino acid được quyết định bởi enzyme aminoacyl-tRNA synthetase. Trung
tâm hoạt động của mỗi loại aminoacyl-tRNA synthetase chỉ phù hợp cho sự bắt cặp
đặc thù của một loại amino acid và tRNA. Có 20 loại aminoacyl-tRNA synthetase
khác nhau, mỗi loại dành cho một amino acid; mỗi aminoacyl-tRNA synthetase có thể
liên kết với nhiều tRNA cùng mã hóa cho một loại amino acid.
- Thứ hai là sự bắt cặp giữa anticodon trên tRNA với codon trên mRNA. Có 61 codon trên mRNA
nhưng chỉ có khoảng 45 loại tRNA. Điều này cho thấy rằng một số tRNA có thể liên kết với nhiều hơn một bộ ba mã hóa. Sự bắt cặp “linh hoạt” như vậy có thể do nguyên tắc bắt cặp bổ sung giữa base thứ ba của anticodon trên tRNA với base tương ứng trên
Hình 2.10. Sự bắt cặp linh động giữa anticodon và codon tại vị trí wobble.
mRNA là “lỏng lẻo” (wobble position) hơn so với các base ở hai vị trí còn lại (Hình 2.9). Sự bắt cặp “linh động” này giải thích vì sao các bộ ba đồng nghĩa (cùng mã hóa cho một amino acid) thường chỉ khác nhau ở base thứ ba.
2.1.5.2. Ribosome – nhà máy tổng hợp protein của tế bào
Các ribosome tạo điều kiện cho sự bắt cặp giữa các anticodon trên tRNA với các codon tương ứng trên mRNA trong quá trình tổng hợp protein. Mỗi ribosome đều gồm hai tiểu phần, lần lượt được gọi là tiểu phần lớn và tiểu phần bé. Ở cả vi khuẩn và các sinh vật nhân thực, tiểu phần lớn và tiểu phần bé chỉ lắp ráp với nhau để hình thành nên ribosome khi chúng đã đính kết vào một phân tử mRNA. Khoảng 2/3 khối lượng ribosome là các rRNA, bao gồm 3 phân tử ở vi khuẩn và 4 phân tử ở các sinh vật nhân thực. Do phần lớn mỗi tế bào luôn chứa hàng ngàn ribosome nên rRNA thường là loại RNA phổ biến nhất trong tế bào [1].
Cấu trúc của ribosome phản ánh chức năng của nó là đưa mRNA tiếp xúc với tRNA đã nạp amino acid. Ngoài vị trí liên kết với mRNA, mỗi ribosome đều có 3 vị trí liên kết tRNA [1].
- Vị trí P (vị trí petidyl-tRNA): giữ tRNA mang chuỗi polypeptide đang được kéo dài.
- Vị trí A (vị trí aminoacyl-tRNA): giữ tRNA mang amino acid tiếp theo được bổ sung vào chuỗi polypeptide.

- Vị trí E (exit): nơi giải phóng tRNA tự do (không liên kết với amino acid) khỏi ribosome.
Ribosome giữ mRNA và tRNA ở những vị trí áp sát vào nhau, đồng thời đưa các amino acid mới tới sát cạnh đầu C (đầu cacboxyl) của chuỗi polypeptide đang kéo dài. Sau đó, nó xúc tác cho sự hình thành liên kết peptide. Khi chuỗi polypeptide đã đủ dài, nó chui qua một kênh thoát thuộc tiểu phần lớn của ribosome. Khi chuỗi polypeptide đã tổng hợp xong, nó được giải phóng vào bào tương của tế bào qua kênh thoát.
Nhiều nghiên cứu chỉ ra rằng, chính rRNA giữ vai trò quan trọng trong cấu trúc và chức năng của ribosome. Các phân tử protein giúp thay đổi cấu hình không gian của các rRNA khi các phân tử này thực hiện vai trò xúc tác trong quá trình dịch mã. Các rRNA là thành phần chính tạo nên giao diện tiếp xúc giữa hai tiểu phần ribosome tại các vị trí A và P, đồng thời cũng là trung tâm xúc tác hình thành liên kết peptide. Vì vây, ribosome có thể được xem là một ribozyme khổng lồ! [1]
2.1.5.3. Quá trình dịch mã
Dịch mã là quá trình sinh tổng hợp protein diễn ra trong tế bào chất, được chia làm hai giai đoạn là hoạt hoá amino acid và tổng hợp chuỗi polipeptide [7].
* Hoạt hoá amino acid
Đây là quá trình tạo nguồn các aminoacyl-tRNA cho dịch mã. Mỗi amino acid được đính vào tRNA thích hợp thông qua hai phản ứng nhờ enzyme aminoacyl-tRNA synthetase đặc thù cùng với ATP và Mg2+ .
R−CH(NH2)−COOH + ATP → R−CH(NH2)−CO~AMP + PP
R−CH(NH2)−CO~AMP + tRNA → R−CH(NH2)−CO~tRNA + AMP
* Tổng hợp chuỗi polypeptide (Hình 2.10 và 2.11)
Mở đầu (initiation): Quá trình dịch mã bắt đầu khi một tiểu đơn vị ribosome bé bám vào mRNA tại vị trí của codon khởi đầu AUG. Lúc này một phân tử tRNA khởi đầu đặc thù mang methionine (ở vi khuẩn là formyl-Met) đi vào và khớp anticodon của nó với codon mở đầu của mRNA. Kế đó, tiểu đơn vị ribosome lớn bám vào tiểu đơn vị bé tạo ra một ribosome hoạt động hoàn chỉnh. Lúc này Met-tRNA ở vị trí P và vị trí A để trống; một tRNA thứ hai (ví dụ, tRNAVal) đi vào vị trí A và khớp với codon thứ hai.
Hình 2.11. Giai đoạn mở đầu của quá trình dịch mã ở vi khuẩn.
Kéo dài (elongation): Quá trình kéo dài bắt đầu sau khi liên kết peptide đầu tiên được hình thành. Phản ứng này được xúc tác bởi enzyme peptidyl transferase, và kết quả là tạo ra một peptidyl-tRNA ở vị trí 1. Sau đó, ribosome lập tức chuyển dịch sang một codon mới dọc theo mRNA theo chiều 5'→3'. Phản ứng này đẩy phân tử tRNA tự do vốn ở vị trí P ra ngoài; lúc này peptidyl-tRNA nằm ở vị trí P và vị trí A lại để trống. Một chu kỳ dịch mã mới lại bắt đầu, một aminoacyl-tRNA thứ ba đi vào và khớp anticodon của nó với codon đang để trống ở vị trí A, một liên kết peptide thứ hai được hình thành, và ribosome lại dịch chuyển sang codon kế tiếp. Quá trình nói trên diễn ra một cách tuần tự dọc theo mRNA làm cho chuỗi polypeptide dài dần ra đến khi dịch mã xong codon có nghĩa cuối cùng.

Kết thúc (termination): Quá trình tổng hợp chuỗi polypeptide sẽ dừng lại khi codon kết thúc của mRNA đối diện với vị trí A. Lúc này, nhân tố giải phóng RF (release factor) đi vào; chuỗi polypeptide được tách ra và phóng thích cùng với hai tiểu đơn vị ribosome, tRNA cũng ra khỏi mRNA.



Hình 2.12. Giai đoạn kéo dài chuỗi polypeptide.
Lưu ý:
(1) Thực ra, trên một mRNA có rất nhiều ribosome cùng hoạt động, gọi là polysome, tạo ra nhiều polypeptide giống nhau.
(2) Trên nguyên tắc, amino acid mở đầu sẽ được cắt bỏ khỏi chuỗi polypeptide. Tuy nhiên, ở các eukaryote không phải lúc nào amino acid mở đầu này cũng bị tách bỏ, mà trong một số protein nó vẫn được giữ lại.
(3) Sau tổng hợp, các chuỗi polypeptide sẽ được sửa đổi và chuyển sang các bậc cấu trúc cao hơn để trở thành các protein chức năng. Thực ra sự biến đổi sau dịch mã còn có các chaperone và nhiều cơ chế tác động phức tạp khác nữa.
(4) Tham gia vào các bước mở đầu, kéo dài và kết thúc còn có các yếu tố protein, với tên gọi tương ứng là các nhân tố mở đầu, kéo dài, và giải phóng cùng với ATP, GTP và các ion Mg2+, K+ và NH4 +
(5) Trong các tế bào prokaryote, các ribosome và các aminoacyl-tRNA sẽ bám vào đầu 5' của mRNA để bắt đầu quá trình dịch mã trong khi ở đầu 3' của nó quá trình phiên mã đang còn tiếp diễn. Ngược lại, ở các tế bào eukaryote, các pre-mRNA phải trải qua sửa đổi sau phiên mã ở trong nhân, còn dịch mã diễn ra sau đó trong tế bào chất.
(6) Về RNA đối nghĩa (antisense RNA), đây là loại RNA thấy có ở nhiều hệ thống, nhưng rất phổ biến ở các vi khuẩn. Nó được tổng hợp từ sợi đối nghĩa của gene, nên bổ sung với mRNA và có thể tạo thành một sợi kép với nó để gây kìm hãm dịch
mã. Vì vậy RNA đối nghĩa còn được gọi là RNA bố sung gây nhiễu mRNA, và được ứng dụng hiệu quả trong điều trị ung thư
2.1.5.4. tmRNA – ARN cứu hộ ribosome của tế bào vi khuẩn
Một mRNA bị thiếu codon kết thúc là nguyên nhân gây trở ngại cho quá trình phiên mã. Một mRNA có thể bị thiếu codon kết thúc do nhiều nguyên nhân như: một đột biến đã loại bỏ codon kết thúc, sự tổng hợp mRNA bị lỗi, hoặc mRNA bị suy thoái một phần trước khi nó được dịch mã,…. Nếu một ribosome đi đến cuối phân tử mRNA và không có codon kết thúc, yếu tố giải phóng không thể liên kết và ribosome không thể giải phóng khỏi mRNA. Nói cách khác, ribosome này đã bị “kẹt” lại trên mRNA [6].

Hình 2.13. Giải cứu ribosome bởi tmRNA.
Tế bào vi khuẩn có chứa một phân tử RNA nhỏ, gọi là tmRNA, làm nhiệm vụ giải phóng ribosome bị “kẹt”. tmRNA có cấu trúc giống với cả tRNA vận chuyển amino acid alanine và mRNA có chứa một đoạn ngắn có thể được dịch mã.
Khi tmRNA va chạm với một ribosome đang bị “kẹt”, nó sẽ liên kết với mRNA bị lỗi. Quá trình tổng hợp protein có thể được tiến hành sau đó, đầu tiên là thêm alanin vào tmRNA và sau đó là dịch mã đoạn thông tin ngắn trên tmRNA. tmRNA có chứa một codon kết thúc và do đó, yếu tố giải phóng có thể liên kết để giải phóng ribosome. Chuỗi polypeptide được tạo ra từ quá trình “giải cứu ribosome” này là một sản phẩm lỗi và sẽ bị phân hủy sau đó. Điều này được thực hiện nhờ một chuỗi ngắn các axit amin được mã hóa bởi tmRNA được thêm vào phần cuối của protein bị lỗi; như tín hiệu cho một protease phân hủy protein lỗi. Do đó, nhờ hoạt động của tmRNA, các ribosome bị “kẹt” không bị bất hoạt mà được giải phóng để tham gia vào quá trình tổng hợp protein tiếp theo [6].
2.2. Điều hòa biểu hiện gen ở vi khuẩn
2.2.1. Các nguyên lý điều hòa Cho đến nay, các cơ chế điều hoà được hiểu rõ nhất là các cơ chế ở vi khuẩn và phage. Trong các hệ thống này, hoạt động điều hoà mở - đóng (on-off regulatory activity) xảy ra thông qua kiểm soát sự phiên mã ở chỗ: sự tổng hợp một mRNA (mở)
519
cụ thể chỉ xảy ra khi sản phẩm của gene được cần đến và bị kìm hãm (đóng) khi sản phẩm này không thực sự cần đến. Đó chính là cơ chế liên hệ ngược (feed-back). Trong trường hợp sau, nói cho đúng là sự phiên mã diễn ra rất thấp, với một ít sản phẩm gene có mặt [7].
Ở các vi khuẩn, khi một vài enzyme hoạt động theo một trình tự trong một con đường chuyển hoá thì thường hoặc là tất cả các enzyme này được sinh ra hoặc không. Hiện tượng này gọi là điều hoà phối hợp (coordinate regulation), do mRNA của các prokaryote thuộc kiểu polycistron.
Một số cơ chế điều hoà phiên mã có tính phổ biến; một cơ chế cụ thể được sử dụng thường phụ thuộc vào các enzyme được điều hoà hoạt động trong các con đường chuyển hóa thuộc kiểu phân giải (dị hoá) hay tổng hợp (đồng hoá). Chẳng hạn, trong một hệ thống phân giải nhiều bước thì sự có sẵn các phân tử để phân giải thường xác định việc tổng hợp các enzyme trong con đường chuyển hoá đó. Trái lại, trong con đường sinh tổng hợp, sản phẩm cuối của con đường chuyển hoá này thường đóng vai trò là phân tử điều hoà. Ngay cả trong một hệ thống mà trong đó một phân tử protein (không nhất thiết phải là một enzyme) được dịch mã từ một mRNA monocistron, protein đó có thể tự điều hoà (autoregulated) - nghĩa là, bản thân nó có thể kìm hãm sự khởi đầu phiên mã và nồng độ cao của protein này sẽ khiến cho tổng hợp mRNA của nó ít lại [7].
(a) (b) (c)
A B C X
Enzyme đầu tiên (a) là một enzyme dị lập thể. Nó có đặc điểm thay đổi cấu hình không gian khi có mặt sản phẩm cuối cùng nhằm giảm bớt hoạt tính xúc tác của mình. Ở enzyme này, ngoài vị trí gắn với cơ chất A (trung tâm phản ứng), nó còn có một hay nhiều vị trí gắn với sản phẩm cuối cùng x gọi là trung tâm dị lập thể. Trung tâm phản ứng và trung tâm dị lập thể tách biệt nhau về không gian và khác nhau về cấu trúc. Trạng thái hoạt động của enzyme này được đặc trưng ở chỗ: nó có khả năng liên kết với cơ chất A và nếu bên cạnh cơ chất A còn có sự hiện diện của X ở mức độ dư thừa so với nhu cầu của cơ thể vi sinh vật thì sẽ xảy ra sự bao vay của trung tâm dị lập thể, làm cho trung tâm phản ứng bị biến đổi cấu hình không gian đến mức khiến cho enzyme (a) không thể gắn được với cơ chất A mà chỉ gắn với X. Như vậy enzyme (a) sẽ không có hiệu lực chuyển hóa A thành B. Chuỗi sinh tổng hợp X sẽ bị gián đoạn. Do đó, X sẽ bị giảm số lượng.
Sự ức chế quá trình sinh tổng hợp này xảy ra nhanh và rất có hiệu quả. Người ta nhận thấy enzyme (a) tuy vẫn được cơ thể vi sinh vật tổng hợp nên nhưng bất hoạt. Sự điều hòa này ở mức độ enzyme.

Các cơ chế phân tử đối với mỗi một kiểu điều hoà sai khác nhau rất lớn nhưng thường rơi vào một trong hai tiêu chí chính yếu sau: điều hoà âm tính (negative regulation) và điều hoà dương tính (positive regulation). Trong hệ thống được điều hoà theo kiểu âm tính, chất ức chế (inhibitor/ repressor) có mặt trong tế bào và gây cản trở phiên mã. Một chất đối lập của chất ức chế, gọi chung là chất cảm ứng (inducer), cần thiết cho sự khởi đầu phiên mã. Trong hệ thống được điều hoà theo kiểu dương tính, phân tử hiệu ứng (có thể là một protein) kích hoạt vùng khởi động (promoter) làm tăng cường hiệu quả phiên mã. Cần lưu ý rằng, sự điều hoà âm tính và dương tính không phải loại trừ lẫn nhau, và một số hệ thống được điều hoà theo cả hai cách này bằng cách sử dụng hai chất điều hoà để đáp ứng với các điều kiện khác nhau trong tế bào.
Một hệ thống phân giải có thể được điều hoà hoặc dương tính hoặc âm tính.
Trong con đường sinh tổng hợp, sản phẩm cuối cùng thường điều hoà âm tính sự tổng hợp của riêng nó; ở kiểu điều hoà âm tính đơn giản nhất, sự vắng mặt của sản phẩm này làm tăng cường sự tổng hợp ra nó và sự có mặt của sản phẩm lại làm giảm mức tổng hợp của nó.
Các phương thức điều hoà ở các prokaryote rõ ràng là đơn giản hơn và được hiểu rõ hơn so với ở các eukaryote, mặc dù lượng thông tin có được về các hệ thống eukaryote đang được tích luỹ với một tốc độ phi thường.
2.2.2. Mô hình Operon
Phần lớn các gene trong bộ gen vi khuẩn được tổ chức thành các đơn vị hoạt động chức năng đặc trưng, gọi là các operon. Năm 1961, Francois Jacob và Jacques Monod (France) đề xuất giả thuyết operon để giải thích sự điều hoà quá trình sinh tổng hợp protein ở vi khuẩn - các enzyme tham gia vào con đường hấp thụ và phân giải đường lactose - operon lactose. Cho tới nay, mô hình
trí của các maltose operon và lactose operon.
và cơ chế hoạt động của nhiều operon khác như operon arginine, operon maltose, operon tryptophan,…. cũng đã được nghiên cứu rõ [6]. Tuy nhiên, trong phạm vi của chuyên đề này chúng tôi chỉ xin đề cập đến operon lactose và operon tryptophan – hai operon được nghiên cứu sớm nhất và thường xuyên xuất hiện trong các đề thi học sinh giỏi trong nước, IBO.
Tham gia vào điều hòa hoạt động của một operon gồm có bốn yếu tố thuộc hai thành phần chính: (i) các locus cấu trúc (structural loci) và (ii) các locus điều hòa (regulatory loci); trong đó nhóm sau bao gồm yếu tố chỉ huy (operator), vùng khởi động (promoter) và gene điều hòa (regulator gene) [7].
2.2.2.1. Nhóm gen cấu trúc
Nhóm gen này đảm nhận việc mã hóa cấu trúc của các phân tử protein – enzyme. Các gen này thường xếp liền nhau. Trong Operon Lac của E. coli, các gen cấu trúc gồm 3 gen kí hiệu là Z, Y và A [6].
- Gen Z: mã hóa thứ tự amino acid của β-galactosidase.
Hình 2.15. Mô hình cấu trúc và cơ chế hoạt động của lactose operon.
- Gen Y: mã hóa cấu trúc của enzyme thẩm thấu galactosidepermease.
- Gen A: mã hóa cấu trúc của enzyme thiogalactoseacetyltransferase.

Cả 3 gen cấu trúc nói trên tạo thành một đơn vị phiên mã.
2.2.2.2. Gen chỉ huy (Operator)
Là đoạn DNA nằm kề bên nhóm gen cấu trúc (Hình 2.14). Nhờ tác dụng gắn với chất ức chế, Operator làm việc như một công tắc phụ trách việc đóng mở hoạt động của nhóm gen cấu trúc [6].
2.2.2.3. Gen khởi động (Promoter)
Là đoạn DNA nằm kề phía trước Operator, là nơi gắn enzyme RNA polymerase. Enzyme này xúc tác cho quá trình tổng hợp mRNA của nhóm gen cấu trúc. Khi chất ức chế gắn vào Operator thì phân tử RNA polymerase bị cản trở, không thể di chuyển dọc theo mạch khuôn DNA dẫn đến các gen cấu trúc bị kiềm chế và không tạo được protein cũng như enzyme tương ứng. Do vị trí và chức năng như vậy nên người ta gọi là gen Promoter [7].
521
Hình 2.14. Nhiễm sắc thể của E. coli với vị2.2.2.4. Gen điều hòa
Gen này chịu trách nhiệm mã hóa việc tổng hợp nên một protein đặc biệt đóng
vai trò chất ức chế. Thường nó chỉ được tổng hợp với một lượng không đáng kể trong tế bào (khoảng 10-20 phân tử/tế bào). Đặc điểm của chất ức chế là một protein biến cấu oligomer, có 2 tâm đặc thù, 2 tâm này làm cho chất ức chế có khả năng hoặc gắn được với chất cảm ứng hoặc gắn với Operator. Chất ức chế có ái lực lớn đối với Operator nên thông thường nó hay gắn vào Operator [7].
Nhóm gen cấu trúc, gen Operator và gen Promoter được tổ hợp lại thành một đơn vị gọi là Operon. Mỗi một Operon tương ứng với một gen điều hòa và chịu sự kiểm soát nghiêm ngặt của gen này.
Khi trong môi trường xuất hiện chất cảm ứng. Chất này có ái lực với chất ức chế, nó tác dụng với chất ức chế Điều đáng lưu ý ở đây là chất cảm ứng tác dụng với chất ức chế làm cho chất ức chế bị bất hoạt. Cụ thể: Khi chất cảm ứng gắn với chất ức chế, nó làm thay đổi cấu hình không gian của chất ức chế, do đó làm giảm ái lực của tâm gắn chất ức chế với Operator, từ đó giúp Operator tự giải thoát khỏi chất ức chế. Lúc này Operon sẽ được giải phóng. Liên đới gen Promoter sẽ được giải phóng. Enzyme RNA polymerase không bị cản trở và nó được di chuyển dọc theo mạch khuôn DNA. Quá trình tổng hợp mRNA được bắt đầu, các gen cấu trúc sẽ làm việc để tiến hành tổng hợp protein, enzyme sẽ được hình thành. Chính enzyme (enzyme cảm ứng ) được tạo thành này sẽ có trách nhiệm phân hủy cơ chất khó đồng hóa (chất cảm ứng).
Theo sơ đồ của F.Jacob và J.Monod, để hình thành một enzyme cảm ứng phải hội tụ đủ 4 điều kiện: gen, nguyên liệu xây dựng, năng lượng và chất cảm ứng.
2.2.3. Điều hòa âm tính của các Operon cảm ứng
Điều hòa âm tính của các Operon cảm ứng còn gọi là điều hòa tổng hợp enzyme nhờ sự ức chế dị hóa: Phức hợp Protein ức chế - chất cảm ứng không gắn được vào Operator, quá trình phiên mã được khởi động.
Đại diện cho tất cả các operon của các loại đường disaccharide và polysaccharide (mà vi khuẩn sử dụng như một nguồn cung cấp các hợp chất carbon và năng lượng) là operon lactose ở E. coli.
2.2.3.1. Cơ chế điều hòa âm cảm ứng
Operon lactose có chức năng sản sinh các enzyme tham gia vào quá trình hấp thụ và phân giải đường lactose thành galactose và glucose. Nó chỉ hoạt động khi có mặt đường lactose, vì vậy lactose được gọi là chất cảm ứng và lac operon được gọi là operon cảm ứng (inducible) hay operon dị hoá (catabolite) [7]. Nói đúng ra, chất cảm ứng là allolactose; lactose (liên kết galactosid dạng β-1,4) bị biến đổi thành chất trung gian trong quá trình thuỷ phân lactose dưới tác dụng của β-galactosidase, gọi là allolactose (liên kết β-1,6) [6].
523
Khi trong môi trường nuôi cấy E. coli vắng mặt lactose (allolactose, chất cảm ứng) thì lac operon không hoạt động, nghĩa là các enzyme tham gia hấp thụ và phân giải lactose không được sinh ra. Nguyên nhân là do chất ức chế của operon (lac repressor) vốn tự thân có hoạt tính, bám chặt vào yếu tố chỉ huy (lac operator) và gây kìm hãm sự phiên mã của các gene cấu trúc Z, Y và A. Do đó các sản phẩm enzyme của lac operon không được tạo ra (biểu hiện âm tính).
Ngược lại, nếu bổ sung lactose vào môi trường thì một thời gian sau vi khuẩn sẽ bắt đầu hấp thụ và phân giải nó, nghĩa là các enzyme liên quan đã được sinh ra. Điều này được lý giải như sau: Chất cảm ứng (inducer), ở đây là allolactose - dạng biến đổi của lactose - tương tác với chất ức chế (repressor) làm biến đổi cấu hình của chất này. Một phân tử allolactose bám vào một tiểu đơn vị của chất ức chế. Vì vậy chất ức chế mất ái lực và không thể bám vào lac operator và nó tách ra khỏi DNA. Lúc này các gene cấu trúc được phiên mã và các enzyme tương ứng đượ c tổng hợp, nhờ vậy vi khuẩn có thể hấp thụ và phân gi ải đường lactose. Lactose vì vậy là tác nhân gây cảm ứng (hoạt hoá) lac operon. Ngoài ra, ITPG (isopropyl thiogalactoside) cũng được dùng như một chất cảm ứng nhưng không phải là tác nhân sinh lý.
Phương thức điều hoà như thế được gọi là điều hoà cảm ứng - âm tính, bởi vì chất ức chế lac operon một khi bám vào lac operator sẽ kìm hãm phiên mã, nghĩa là gây hiệu quả âm tính lên sự biểu hiện của các gene và hoạt động chức năng của protein này lại phụ thuộc vào chất cảm ứng. Nhờ cơ chế điều hoà kiểu liên hệ ngược này mà vi khuẩn có thể thích ứng để tồn tại và phát triển một cách hợp lý [7].
2.2.3.2. Điều hòa tổng hợp enzyme nhờ sự ức chế dị hóa (điều hòa dương tính của Lac Operon) Người ta nhận thấy rằng, nếu trong môi trường nuôi cấy vi sinh vật có nhiều nguồn cơ chất, trước hết xảy ra sự tổng hợp các enzyme xúc tác cho sự phân giải cơ chất dễ sử dụng nhất. Sự tổng hợp các enzyme xúc tác phân giải các cơ chất khác bị ức chế bởi sự ức chế dị hóa. Ví dụ: Trong môi trường nuôi cấy có 2 nguồn carbohydrate là glucose và lactose. Trước tiên vi sinh vật sẽ hình thành các enzyme phân giải glucose. Sự cảm ứng để tổng hợp enzyme phân giải lactose là β-galactosidase bị ức chế bởi sự ức chế dị hóa.
Cơ chế ức chế dị hóa đã được nghiên cứu khá chi tiết ở vi khuẩn E.coli với việc điều hòa tổng hợp enzyme β-galactosidase.
Trong số các hợp chất dạng carbohydrate, glucose là nguồn cơ chất được sử dụng thích hợp nhất. Vì vậy, khi có mặt glucose thì nhiều enzyme khác của quá trình dị hóa cũng như trao đổi chất trung gian không được tổng hợp. Người ta gọi hiện tượng này là hiệu ứng glucose.
Thật ra không phải bản thân glucose mà là một chất trao đổi khác được hình thành trong quá trình dị hóa glucose sẽ có nhiệm vụ đối với sự ức chế này. Nhiều thực nghiệm cho thấy chất đó chính là cyclic adenosine monophosphate (cAMP). cAMP là một phân tử quan trọng trong nhiều hệ thống kiểm soát trao đổi
Hình 2.16. Phản ứng tạo cAMP từ ATP.
chất ở cả sinh vật nhân sơ và nhân thực [6].
Các cơ chất được đồng hóa nhanh như glucose ức chế sự tạo thành cAMP và kích thích sự phân hủy nó. Khi có mặt glucose trong môi trường thì hàm lượng cAMP phải ở mức độ thấp. Chỉ sau khi glucose được tiêu thụ hết thì nồng độ cAMP ở trong môi trường mới tăng lên. Hoạt động của Operon lac được mô tả ở Hình 2.16 và chúng ta sẽ xem xét cụ thể trong 3 trường hợp sau.
a. Khi môi trường không có glucose, có lactose
Khi nồng độ glucose trong tế bào xuống thấp, một lượng lớn cAMP sẽ được tích lũy. Lúc này, cAMP sẽ liên kết với protein thụ thể kí hiệu là CRP (cyclic AMP receptor protein) tạo thành phức hệ cAMP – (CRP). Chỉ khi liên kết với cAMP thì
CRP mới bám được vào Promoter của Operon Lac và RNA polymerase mới gắn vào

Promoter để phiên mã các enzim phân giải lactose. Đồng thời, lactose sẽ liên kết với
protein ức chế, làm thay đổi cấu hình không gian của protein ức chế, khiến cho nó không gắn được với Operator để ngăn cản phiên mã các gen cấu trúc.
RNA polymerase được 2 yếu tố kích thích hoạt động, dẫn đến tổng hợp mRNA và tổng hợp nên β-galactosidase, dó đó người ta gọi là Lac Operon được điều khiển kép.
b. Khi môi trường có lactose và có glucose

Trong môi trường, nếu ngoài lactose còn được bổ sung glucose thì lúc ấy hàm lượng cAMP sẽ bị giảm mạnh, không tạo được phức hệ với cAMP- CRP, do đó RNA polymerase không hoạt động hoặc hoạt động rất yếu. Vì vậy, ngay cả khi có mặt cơ chất cảm ứng lactose thì gen
Hình 2.17. Sơ đồ hoạt động của operon Lac.
cấu trúc cũng không được phiên mã và không tạo thành enzyme hoặc hoạt động này xảy ra rất yếu.
c. Khi môi trường không có glucose, không có lactose
Khi trong môi trường không có glucose như đã phân tích ở trên, RNA polymerase sẽ được hoạt hóa. Tuy nhiên, nếu vắng mặt cả lactose thì protein ức chế sẽ gắn vào Operator nên Operon Lac vẫn bị ức chế. Do đó, mRNA không được tổng hợp, không có sự tổng hợp enzyme.
2.2.4. Điều hòa âm tính của các Operon ức chế
Đây là cơ chế kiểm soát âm ức chế: phức hợp protein ức chế - chất cảm ứng có khả năng gắn vào Operator ức chế quá trình phiên mã.
Đại diện cho tất cả các operon của các loại amino acid và các vitamine là Operon tryptophan (trp operon) ở E. coli [6]. Operon tryptophan có chức năng sản sinh các enzyme đồng hoá tham gia vào quá trình sinh tổng hợp amino acid tryptophan (tryp) cho tế bào; thiếu amino acid này vi khuẩn không thể tổng hợp protein được và
nó sẽ chết. Trp Operon cũng chịu sự điều hoà âm tính như lac Operon, chỉ hoạt động khi môi trường nội bào thiếu hụt và không hoạt động (bị ức chế) khi dư thừa tryp (sản phẩm cuối của con đường sinh tổng hợp). Vì vậy trp Operon được gọi là Operon ức chế hay Operon đồng hoá.
Trp Operon của E. coli có chứa năm gene cấu trúc chính ( trpE, trpD, trpC, trpB và trpA) mã hoá cho các enzyme tham gia vào các con đường sinh tổng hợp tryp theo một cơ chế rất phức tạp. trp Operon có một số đặc điểm khác như: trình tự Operator nằm lọt trong Promoter; gene điều hoà nằm cách xa Operon về phía trước (Hình 2.17) [6].

2) Đo độ hấp phụ ánh sáng của môi trường nuôi cấy bằng máy đo quang phổ (với bước sóng A600 = 1 tương ứng 8x108 tế bào vi khuẩn/ml).
3) Pha loãng môi trường nuôi cấy vài lần sau đó cấy trải trên đĩa thạch và đếm số khuẩn lạc (xem hình dưới) để tính số lượng khuẩn lạc hình thành trên một ml (cfu/ml).
Hình 2.18. Mô hình cấu trúc operon tryptophan.

Khi trong môi trường có hiện tượng dư thừa tryp so với nhu cầu của tế bào, tryp sẽ gắn với protein ức chế làm thay đổi cấu hình không gian của protein khiến cho protein ức chế có khả năng gắn với Operator. Chất ức chế bình thường của Operon này tồn tại ở dạng bất hoạt, không có ái lực đối với trp Operator, nhưng khi các amio acid dư thừa kết hợp vào sẽ tạo ra phức hợp có hoạt tính, nghĩa là có ái lực với trp Operator. Vì thế tryp được gọi là chất đồng ức chế. Phức hợp này bám vào yếu tố chỉ huy làm kìm hãm phiên mã của trp operon.
Khi protein ức chế gắn với Operator sẽ làm ngưng trệ quá trình phiên mã, ức chế Operon dẫn đến các enzyme không được tổng hợp, việc sản xuất tryp cũng bị gián đoạn. Trong khi đó, tế bào vẫn tiếp tục sử dụng tryptophan khiến cho số lượng acid amin này giảm đến mức không đủ để đáp ứng nhu cầu. Dẫn đến, protein ức chế bị thiếu yếu tố hoạt hóa do đó bị bất hoạt và không còn khả năng gắn với Operator. Operon được giải phóng và các enzyme được tổng hợp, việc sản xuất tryp được tiến hành trở lại, gọi là sự giải ức chế [7].
3. BÀI TẬP TÍCH HỢP HÓA SINH – VI SINH – DI TRUYỀN
Các câu hỏi được trích từ các đề thi IBO, Chọn học sinh giỏi cấp Quốc gia,…..
Câu 1. Đếm vi khuẩn trong môi trường lỏng có thể thực hiện bằng các phương pháp khác nhau:
Hãy chỉ ra các phát biểu dưới đây là đúng hay sai? Giải thích vì sao.
A. Mẫu nuôi cấy có giá trị A600 = 0,1 và thời gian nhân đôi là 30 phút sẽ đạt tới 4x108 tế bào/ml trong thời gian chưa đầy 1 giờ 30 phút.
B. Phương pháp đếm khuẩn lạc trên đĩa nuôi cấy sẽ cho số tế bào vi khuẩn có xu hướng it hơn phương pháp đếm dưới kính hiển vi.
C. Sử dụng phương pháp cấy gạt trên đĩa cho số tế bào sống chính xác nhất (từ hình vẽ), dịch nuôi cấy môi trường X ước tính chứa khoảng 1.6x105 cfu/ml.
D. Khi lặp lại cấy gạt đĩa nhiều lần trên đĩa với độ pha loãng f (từ hình vẽ), một vài đĩa sẽ xuất hiện khuẩn lạc.
Hướng dẫn trả lời
A. Đúng
Mẫu nuôi cấy có giá trị OD600= 0,1 và 30 phút nhân đôi 1 lần thì sau 30 phút OD600 = 0,2; sau 1 giờ là 0,4 và sau 1 giờ 30 phút là 0,8.
1) Các tế bào có thể được đếm dưới kính hiển vi bằng cách sử dụng buồng đếm. 528
Mà OD600 = 1 tương ứng với 8 x108 tế bào => mẫu nuôi cấy sẽ đạ
4x108 tế bào/ml trong thời gian chưa đầy 1 giờ 30 phút và OD600 = 0,5.
B. Đúng Phương pháp đếm khuẩn lạc chỉ đếm được các tế bào sống, đếm dưới kính hiển vi sẽ đếm được cả tế bào sống và tế bào chết.
C. Đúng Có 160 khuẩn lạc, độ pha loãng 1000 lần => dịch nuôi cấy môi trường X ước tính chứa khoảng 1.6x105 cfu/ml.
D. Đúng Độ pha loãng f chứa 0,16 tế bào/ml . Theo thống kê thì cấy khoảng 6 lần với độ pha loãng này sẽ có đĩa xuất hiện khuẩn lạc.
Câu 2. Ba chủng nấm men (Saccharomyces cerevisiae) đã được biến đổi gen sao cho mỗi chủng chứa 1 gen mã hóa một trong ba enzyme của quá trình đường phân (Tpi, Eno hoặc Pyk); các gen này được điều khiển bởi một trình tự khởi động (promoter) bị bất hoạt bởi doxycycline, nên sự bổ sung doxycycline làm giảm quá trình tổng hợp enzyme tương ứng. Nồng độ của một số chất chuyển hóa được đo ở mỗi chủng nấm men được nuôi trong môi trường glucose với các nồng độ doxycycline khác nhau, đối chiếu với các nồng độ tương ứng nhưng không chứa doxycycline. Các bước liên quan của quá trình đường phân được thể hiện trên kết quả các phép đo với các chất chuyển hóa và enzyme được viết tắt như sau:

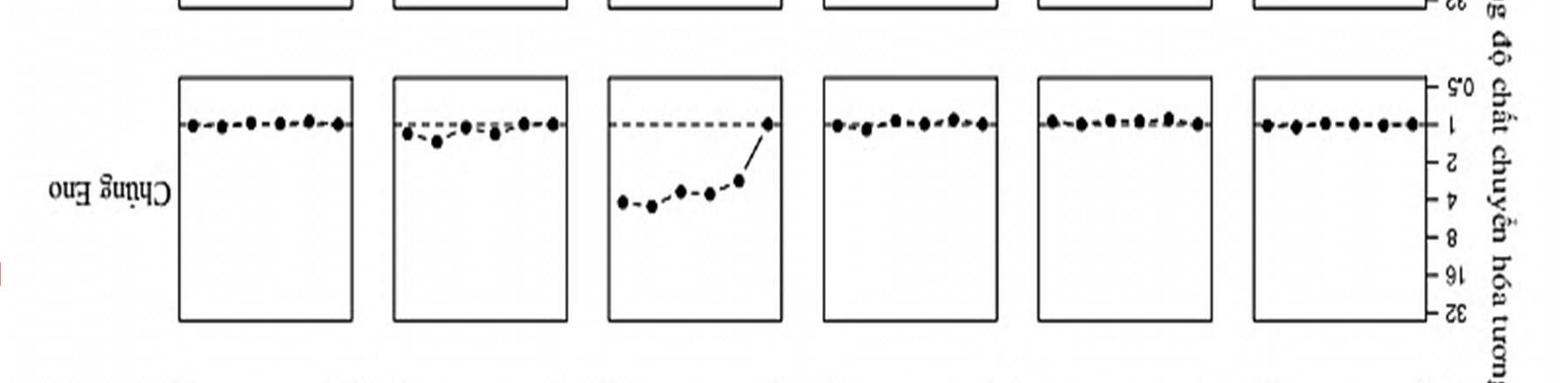


Hãy cho biết các phát biểu sau là đúng hay sai? Giải thích vì sao.
A. Điều hòa làm giảm (âm tính) một trong các enzyme này sẽ làm thay đổi nồng độ các chất chuyển hóa trong toàn bộ con đường.
B. Điều hòa làm giảm (âm tính) một trong các enzyme sẽ ảnh hưởng đến nồng độ cơ chất hơn là nồng độ của sản phẩm tạo thành.
C. Sự cân bằng của phản ứng từ 2+3-PG → PEP nghiêng về phía sản phẩm hơn là sự cân bằng của phản ứng từ FBP → DHAP.
D. Nồng độ F6P được cho là không bị ảnh hưởng bởi điều hòa làm giảm Aldolase.
Hướng dẫn trả lời:
A . Sai
B. Đúng
C . Sai
529
Nếu 1 enzim nào đó bị giảm nồng độ và nồng độ cơ chất của enzim đó tăng lên thì cũng không làm thay đổi nồng độ của các chất chuyển hóa khác trong toàn bộ con đường.
Theo đồ thị ta thấy, điều hòa làm giảm (âm tính) một trong các enzyme sẽ ảnh hưởng đến nồng độ cơ chất hơn là nồng độ của sản phẩm tạo thành.
Theo đồ thị ta thấy, nồng độ enzim Tpi giảm làm ảnh hưởng mạnh mẽ (hơn 10 lần) đến nồng độ DHAP, nhưng nồng độ FBP không đổi. Do đó, cân bằng của phản ứng FBP → DHAP là nghiêng về phía của DHAP.
D
Đúng
.
Ngược lại, việc điều chỉnh giảm Pyk không chỉ ảnh hưởng đến nồng độ của PEP, mà còn ảnh hưởng tới nồng độ của 2 + 3PG. Điều này cho thấy, nồng độ PEP tăng lên sẽ thay đổi trạng thái cân bằng của phản ứng 2 + 3PG → PEP theo hướng trở lại 2 + 3PG, do đó cân bằng về phía
sản phẩm không mạnh bằng FBP → DHAP.
Trong phản ứng F6P → FBP, ATP bị thủy phân để phosphoryl hóa

F6P. Do đó, trạng thái cân bằng nghiêng hẳn về phía FBP.
Khi giảm nồng độ enzim Aldolase sẽ làm tăng nồng độ FBP, nhưng
vì trạng thái cân bằng của phản ứng F6P → FBP nghiêng mạnh về phía
FBP (thậm chí còn mạnh hơn phản ứng FBP → DHAP) nên nồng độ F6P
sẽ không bị ảnh hưởng.
Câu 3. Hình dưới đây minh họa một phần con đường shikimate - một con đường tổng hợp các axit amin vòng thơm ở vi khuẩn.
Đúng vậy, đây là 1 phản ứng khử
C. Sai Trong phản ứng c, cơ chất bị phosphoryl hóa. Vì vậy, Y là ATP, GDP và ADP không thể phosphoryl hóa cơ chất.

D. Sai Trong phản ứng d, một nhóm phosphate bị loại khỏi cơ chất.
Câu 4. Trong vùng sáng của môi trường nước ngọt và nước mặn, nơi ánh sáng xuyên xuống, các vi khuẩn lam được tìm thấy ở phần trên của vùng nước, vi khuẩn tía và xanh lục được tìm thấy ở phần dưới của vùng nước. Giải thích nào sau đây là đúng nhất về sự phân bố theo chiều thẳng đứng của các vi khuẩn quang hợp.
A. Vi khuẩn tía và xanh lục là các vi khuẩn kỵ khí, vi khuẩn lam là vi khuẩn hiếu khí.
B. Vi khuẩn tía và xanh lục có khả năng sử dụng độ dài sóng của ánh sáng tốt hơn, trong khi đó vi khuẩn lam không sử dụng hiệu quả.
C. Sự cách biệt về môi trường sống tăng lên do cạnh tranh dinh dưỡng và ôxy. D. Vi khuẩn lam có khả năng sử dụng oxy và truyền electron quang hợp tốt hơn. E. Đó là kết quả thích nghi với nhiệt độ thấp của vi khuẩn tía và xanh lục.
Đáp án: B.
Câu 5. Một phòng thí nghiệm vi sinh học gần đây đã phân lập vi sinh vật Thermus szegediensis từ một suối nước nóng ở Szeged. Trong một chuỗi các thí nghiệm, đầu tiên, họ nuôi cấy T. szegediensis trong các điều kiện khác nhau để kiểm tra nhu cầu dinh dưỡng của nó. Các điều kiện và kết quả được thể hiện trong bảng dưới đây.

Hãy chỉ ra các phát biểu sau là đúng hay sai. Giải thích.
A. Trong phản ứng a, X là H2O.
B. Trong phản ứng b, cơ chất bị khử
C. Trong phản ứng c, Y là ADP hoặc GDP.
D. Phản ứng d là phản ứng photphoryl hóa cơ chất.
A. Đúng Phản ứng a là phản ứng tách nước, cơ chất bị loại nước. Vì vậy, X là H2O.
B. NADPH + H+ là một chất khử. Số oxi hóa của cacbon trong nhóm cacboxyl ở cơ chất cũng thay đổi từ + 2 thành +1 ở sản phẩm tạo thành. Vì 532
Phòng thí nghiệm đã tìm kiếm các gen của T. szegediensis đóng vai trò quan trọng đối với sự tồn tại của vi sinh vật này ở nhiệt độ cao. Các nhà khoa học đã phân lập được 6 chủng đột biến (M1 - M6). Sau đó, tiến hành phép thử bổ sung như ở bảng sau: (”+”: có sinh trưởng, ”-”: không sinh trưởng, WT: wild type – tế bào kiểu dại) để xem chúng sinh trưởng như thế nào ở nhiệt độ cao (T = 56°C)
5.1. Kiểu dinh dưỡng của T. szegediensis là?
A. Quang tự dưỡng B. Hóa tự dưỡng
C. Quang dị dưỡng D. Hóa dị dưỡng
Đáp án: B
5.2. T. szegediensis nhiều khả năng sử dụng nguồn năng lượng nào sau đây để tổng hợp các hợp chất hữu cơ
A. SO4 2- B. Ánh sáng C. H2S D. NO3-
E. H2O F. Diệp lục G. Axit lactic

Đáp án: C.
5.3. Hãy ghi dấu X để chỉ ra các đột biến là trội hay lặn (A. Trội; B. Lặn)
A B C D E
Hình 1. Đĩa A = Môi trường giàu dinh dưỡng. Đĩa B = Môi trường tối thiểu. Đĩa C = Môi trường tối thiểu + Phenylalanine. Đĩa D = Môi trường tối thiểu + Leucine. Đĩa E = Môi trường tối thiểu + Arginine.



Dựa vào Hình 1, hãy ghi dấu X để chỉ ra những nhận định đúng cho các khuẩn lạc (1-8). Đánh dấu tất cả các ô còn lại bằng dấu O.
6.1 Những đột biến nào có cùng nhu cầu dinh dưỡng giống như tế bào kiểu dại (WT)?
6.2 Em sẽ sử dụng những khuẩn lạc nào để phát hiện các gen tham gia vào con đường tổng hợp arginine?
Trong một thí nghiệm khác, em lấy một nhóm các thể đột biến khác và cấy chúng lên đĩa như trong Hình 2.
Hình 2. Đĩa 1 = Môi trường giàu dinh dưỡng. Đĩa 2 = Môi trường giàu dinh dưỡng không có Arginine.
6.3 Những khuẩn lạc nào (1-8) trong Hình 2 không thể sản xuất arginine?
Đáp án:
5.4. Từ dữ liệu trong bảng trên, em có thể xác định ít nhất bao nhiêu gen khác nhau đóng vai trò quan trọng cho phép T. szegediensis tồn tại ở nhiệt độ cao?
Đáp án: có 2 hoặc 3 gen.
Câu 6. Beadle và Tatum đề xuất rằng một gen tạo ra một enzyme. Họ đã gây đột biến nấm mốc và quan sát thấy các khuẩn lạc khác nhau cần chất bổ sung khác nhau trong môi trường bình thường để tồn tại. 534
Câu 7. A, B, C là các loại thuốc phổ biến được dùng để điều trị bệnh nhiễm trùng do vi khuẩn hoặc virut. Khả năng của các loại thuốc được thể hiện ở bảng dưới đây: Th uốc Đặc điểm
ãy cho biết :
A Liên kết vào phía trước vị trí A trên tiểu đơn vị 50S của
ribôxôm
B Một chất tương tự guanin
C Ức chế ADN gyraza
a. Bước nào trong quá trình tổng hợp protein bị ức chế bởi mỗi loại thuốc? Giải thích.
b. Thuốc nào có hiệu quả hơn để chống lại vi khuẩn? Giải thích.
Đáp án
a. - Thuốc C ức chế ADN gyraza, enzim giúp giải toả lực căng ADN trước chạc sao chép. Do đó, quá trình tổng hợp ARN từ ADN (phiên mã) và bắt đầu dịch mã (tổng hợp protein) bị ức chế
- Thuốc A ức chế chuyển vị aminoacyl bằng cách can thiệp vào việc chuyển tARN từ vị trí A sang vị trí P. Do đó, kéo dài chuỗi polypeptide bị ức chế bởi thuốc B.
- Thuốc B: một chất tương tự guanin, có khả năng liên kết với polymeraza của virut và ức chế tổng hợp ADN của virut.
b. Thuốc A chống vi khuẩn hiệu quả hơn vì nó hoạt động bằng cách liên kết với vị trí A trên tiểu đơn vị 50S của ribôxôm vi khuẩn và ức chế sự kéo dài của chuỗi polypeptid. Do đó, nó sẽ không gây ra bất kỳ ảnh hưởng nào đến cơ thể người vì ribôxôm của sinh vật nhân thực là loại 80S bao gồm hai tiểu phần 60S và 40S trong đó ribôxôm của vi khuẩn là loại 70S gồm hai tiểu phần là 50S và 30S.
Câu 8. Operon fox chứa trình tự các đoạn A, B, C và D (có thể là đoạn trình tự gen cấu trúc hoặc gen điều hòa), để mã hóa cho enzyme 1 và enzyme 2. Đột biến ở các đoạn trình tự A, B, C và D có thể gây nên sự tổng hợp (+) hoặc không tổng hợp (-) các enzyme (thể hiện ở bảng dưới). Hãy cho biết các đoạn trình tự A, B, C và D tương ứng với thành phần nào của operon và gen điều hòa.
Không có mặt fox Có mặt fox
Đột biển Enzy me 1 Enzym e 2 Enzym e 1 Enzym e 2
Không đột biến
- Ở thể đột biến A, khi có mặt Fox, sản phẩm enzyme 2 vẫn được biểu hiện, nhưng enzyme 1 thì không kể cả khi có hay không Fox. Do đó A phải xảy ra đột biến cấu trúc của gen mã hóa cho enzyme 1.
- Đối với thể đột biến B, cả hai enzyme 1 và 2 đều không được biểu hiện bất kể trong điều kiện có hay không có Fox. Do đó, trình tự đột biến có khả năng cao là vùng khởi động promoter, ngăn chặn RNA polymerase bám và trượt qua operon Fox để tiến hành phiên mã.
- Tương tự với A, đột biến ở C chỉ ảnh hưởng làm enzyme 2 luôn không biểu hiện trong khi enzyme 1 vẫn biểu hiện kiểu cảm ứng. Vì vậy thể đột biến C xảy ra đột biến cấu trúc gen mã hóa cho enzyme 2.
- Thể đột biến D luôn tạo ra enzyme 1 và 2 cho dù môi trường có mặt Fox hay không. Đột biến này có khả năng cao xảy ra tại vùng gen điều hòa, sản phẩm của gen điều hòa không có khả năng bám vào operator trong bất kì điều kiện nào.
Câu 9. Các nhà khoa học phân lập 6 dòng vi khuẩn kỵ khí sống trong đất ký hiệu (A-F) để nghiên cứu vai trò của chúng đối với chu trình nitơ. Mỗi dòng được nuôi cấy trong 4 loại môi trường chứa các chất khác nhau: (1) Peptone, (2) Ammonium, (3) Nitrate, và (4) Nitrite. Trong đó, chỉ có môi trường chứa nitrat mới có carbohydrate dưới dạng nguồn cacbon cho vi khuẩn. Sau 7 ngày, các vi khuẩn được kiểm tra sinh hóa để phân tích những thay đổi trong môi trường và kết quả thu được trình bày ở bảng dưới đây:
(Chú thích: “+”: vi khuẩn có sinh trưởng, “-”: vi khuẩn không sinh trưởng; pH+ = pH của môi trường tăng; NO2 - = dương tính với kiểm tra nitrit; NO3 - = dương tính với kiểm tra nitrat; gas = có khí gas trong môi trường nuôi cấy).
Môi trường nuôi
cấy
Chủng vi khuẩn
A B C D E F
Peptone +, pH+ +, pH++ , pH++ , pH+
Ammoni um -+ , NO2 - - -
Nitrate +, + - + - +
Em hãy cho biết các phát biểu sau đây là đúng hay sai:
A. C thuộc nhóm vi khuẩn nitrat hóa.
B. B và D thuộc nhóm vi khuẩn phản nitrat hóa.
C. C và E là những vi khuẩn hóa dị dưỡng.
D. A và F có thể sử dụng nitrat như một chất oxy hóa để tạo ra năng lượng.
Đáp án: A Đúng
C là vi khuẩn nitrat hóa: oxy hóa amoni → nitrit → nitrat. C đã
được quan sát thấy trên môi trường amoni, trong đó nó chuyển hóa NH3
thành NO2B . Sai
Vi khuẩn phản nitrat sẽ thực hiện quá trình khử nitrat thành N2. Sự tăng trưởng của các chủng B và D được quan sát thấy trên môi trường peptone cho thấy cả hai chủng đều phân giải peptone để tăng trưởng tạo ra amôn, quá trình amôn hóa này làm tăng độ pH của môi trường
C . Sai
D Đúng
- Các chủng A, B, D, và F thuộc nhóm hóa dị dưỡng vì chúng sử dụng hợp chất nitơ hữu cơ để sinh năng lượng cho hoạt động sống.
- C và E: hóa tự dưỡng.
Sự tăng trưởng của chủng A và F được quan sát thấy trên môi trường nitrat, môi trường này cũng chứa carbohydrate như một nguồn carbon. Sản xuất khí trên môi trường cho thấy cả hai chủng đều có thể tiến hành hô hấp kị khí chuyển hóa nitrat (NO3) thành nitơ tự do (N2) (Vì vi khuẩn là kị khí, nên không thể sản xuất ra khí CO2 từ hô hấp hiếu khí).
Câu 10. Operon arabinose ở vi khuẩn Escherichia coli không biểu hiện khi môi trường không có arabinose. Điều này liên quan đến protein Ara C. Protein này liên kết với vùng vận hành (operator) của operon arabinose và có vai trò ức chế phiên mã. Bình thường operon arabinose được biểu hiện khi có mặt arabinose. Tuy nhiên, trong các thể đột biến mất gen AraC, operon arabinose không biểu hiện thậm chí khi môi trường có arabinose. Dựa vào những thông tin trên, phát biểu nào dưới đây là hợp lý nhất khi đề cập tới AraC.
A. Sự phiên mã gen AraC được cảm ứng bởi arabinose.
B. Sự phiên mã của gen AraC bị ngăn cản bởi arabinose.
C. Protein AraC được chuyển thành một chất hoạt hóa khi có arabinose.
D. Protein AraC bị phân hủy khi có mặt arabinose.


Đáp án: C
Câu 11. Một số vi khuẩn có cơ chế bổ sung để điều hòa tổng hợp các enzym tham gia vào quá trình sinh tổng hợp trytophan (Trp). Ở phía trước của các gen cấu trúc (trpA-E) trong operon trytophan (Trp) có một đoạn trình tự dẫn đầu (trpL), mã hóa cho đoạn peptide dẫn đầu. trpL chứa hai bộ ba mã hóa tryptophan liên tiếp.
Ở nồng độ tryptophan cao, ribôxôm dịch mã mARN đoạn trình tự peptit dẫn đầu và dừng lại ở bộ ba kết thúc, do đó che khuất đoạn 2 của mARN và cho phép hai đoạn 3 và 4 tạo cấu trúc kẹp tóc. Cấu trúc kẹp tóc được theo sau bởi chuỗi poly-U là tín hiệu kết thúc phiên mã làm cho enzyme ARN polymerase (RNA pol) tách khỏi ADN.
Tuy nhiên, ở nồng độ tryptophan thấp, ribôxôm bị nghẽn lại tại vị trí các bộ ba mã hóa triptophan cho phép hai đoạn 2 và 3 hình thành cấu trúc kẹp tóc. Nhờ vậy, toàn bộ operon tryptophan có thể được phiên mã bởi ARN polymerase.
Hãy cho biết các phát biểu sau đây là đúng hay sai. Giải thích.
A. Cơ chế điều hòa giống hệt như vậy cũng xuất hiện ở các gen trong nhân của các sinh vật nhân thực.
538
B. Khi giảm nồng độ aminoacyl-tARN synthetase (enzyme gắn tryptophan vào tARN Trp), thì sự phiên mã của các gen trpA-E sẽ bị bất hoạt ở một nồng độ tryptophan thấp hơn.


C. Sau khi loại bỏ một trong hai bộ ba mã hóa tryptophan trong đoạn trình tự dẫn đầu, thì sự phiên mã của các gen trpA-E sẽ bị bất hoạt ở nồng độ tryptophan thấp hơn.
D. Trong trường hợp một đột biến làm mất ổn định cấu trúc kẹp tóc 2-3, thì sự phiên mã các gen trpA-E sẽ bị bất hoạt ở nồng độ tryptophan thấp hơn.
Đáp án:
A. Sai
Cơ chế điều hòa như trên chỉ xảy ra khi quá trình dịch mã diễn ra lúc quá trình phiên mã chưa kết thúc. Vì vậy, nó không thể xuất hiện ở các gen trong nhân của các sinh vật nhân thực.
prôtêin bằng cách thêm phân tử nước (ma = 0.018kDa) vào cạnh lysin (K) và arginin (R), Trypsin được bổ sung và khối lượng của các đoạn peptide sau đó được xác định bằng phổ khối (màu đỏ). Một phản ứng đối chứng mà ở đó Pup không được bổ sung, được tiến hành tương tự (màu đen). Khối lượng được làm tròn về 2 chữ số thập phân. Lưu ý rằng có các đoạn Pup nằm ngoài vùng đo được.

B. Sai
C. Đúng
D. Đúng
Khi giảm nồng độ aminoacyl-tARN synthetase (enzyme gắn tryptophan vào tARN Trp), thì sự phiên mã của các gen trpA-E sẽ bị bất hoạt ở một nồng độ tryptophan cao hơn. Vì phức hợp tARN-Trp được hình thành chậm hơn so với bình thường.
Nếu chỉ có 1 bộ ba mã hóa Trp thì ở nồng độ tryptophan thấp hơn vẫn diễn ra quá trình dịch mã đoạn trình tự peptit dẫn đầu, vì vậy sự phiên mã của các gen trpA-E sẽ bị bất hoạt.
Một đột biến làm mất ổn định cấu trúc kẹp tóc 2-3 sẽ thúc đẩy hình thành cấu trúc kẹp tóc đoạn 3-4 ở nồng độ Trp thấp, vì vậy sẽ ức chế sự phiên mã của các gen trpA-E.
Câu 12. Gắn thêm Pupyl (Pupylation) là sự cải biến prôtêin sau dịch mã ở một số xạ khuẩn Actinobacteriat, trong đó một đoạn prôtêin ngắn Pup được nối vào chuỗi bên Lysin của phân tử protein đích bởi enzym Pup ligase.
Các trình tự protein X và Pup cũng như khối lượng phân tử (ma) của các đoạn peptide được nêu dưới đây.
Để xác định liệu protein X có được gắn thêm Pup không, người ta cho prôtêin X tinh sạch được ủ với Pup và enzym Pup ligase qua đêm. Enzym Trypsin thủy phân
Em hãy cho biết các phát biểu sau đây là đúng hay sai. Giải thích.
A. Hiệu quả Trypsin như trên dẫn đến một vài đoạn peptit không được cắt hoàn toàn.
B. Phản ứng thủy phân bởi trypsin bị ức chế cạnh vị trí lysin được pupyl hóa.
C. Dưới những điều kiện của thí nghiệm trên, sự pupyl hóa chỉ xảy ra ở các lysin đơn lẻ (không xảy ra đồng thời trên các lysin liền kề).
D. 90% prôtêin đích bị pupyl hóa.
Đáp án:
540
Lưu ý: Khối lượng cho trong hình thứ hai tương ứng với các monopeptit thử nghiệm. Trong quá trình thủy phân trypsin, nước (ma 0.018 kDa) được thêm vào, do đó, khi tính khối lượng của một đipeptit, khối lượng nước cần trừ đi.
A . Đúng
B . Đúng
C . Đúng
Khối lượng tương ứng với các đipeptit có thể thấy trong phổ khối đen, ví dụ: khối lượng 3,05 kDa tương ứng với một đipeptit được tạo thành bởi hai peptit thử nghiệm đầu tiên.
Peptit được pupyl hóa lớn nhất là 2,78 kDa tương ứng với đipeptit
được tạo thành bởi 1 monopeptit pupyl hóa và 1 monopeptit không pupyl hóa không quan sát thấy (cao nhất sẽ là 1,46 kDa của lysine).
Có hai đỉnh (2,78 và 3,81 kDa) có mặt trong quang phổ màu đỏ (gắn Pup) nhưng không có trong phổ màu đen. Đỉnh 2,78 kDa tương ứng với một
đipeptit được tạo thành bởi hai peptit cuối cùng của protein, đỉnh 3,81 tương
ứng với một tripeptit được tạo thành bởi ba peptit cuối cùng của protein.
Hai đỉnh (1,22 và 1,33 kDa) giảm sau khi bổ sung Pup. Đỉnh 1,33kDa
có mật độ tương đối từ 0,6 xuống 0,25. Do đó, protein được pupyl hóa
D . Sai
khoảng 1- (0,25/0,6) = 58%, không phải 90%. Thêm vào đó, đỉnh tương ứng
với một peptit pupyl hóa (2,78 kDa) có mật độ tương đối chỉ 0,4 và đỉnh
tương ứng với tripeptit được pupyl hóa (3,81 kDa) có mật độ tương đối khoảng 0,15, mức độ pupyl hóa sẽ chỉ khoảng 55%.
Câu 13. Enzym giới hạn EcoRI là một ADN endonucleaza có trình tự nhận biết là GAATTC. Nó được tìm thấy lần đầu ở Escherichia coli, vì vậy nó được đặt tên là EcoRI. Để tạo lượng lớn endonucleaza này, đoạn ADN mã hóa gen được cài vào vectơ biểu hiện plasmid, rồi plasmid tái tổ hợp được chuyển vào tế bào E. coli để sản xuất enzym tái tổ hợp phục vụ cho nghiên cứu. Tại sao ADN tế bào chủ không bị cắt bởi EcoRI tái tổ hợp?
A. ADN của tế bào chủ không có các vị trí nhận biết của EcoRI.
B. EcoRI bị các tế bào chủ tiết đẩy ra ngoài.
C. Các nhân tố môi trường như nhiệt độ hay pH ức chế hoạt động của EcoRI.
D. Tế bào chủ E. coli sản sinh các chất ức chế ngăn cản hoạt động của EcoRI.
E. Các trình tự nhận biết của EcoRI trong ADN tế bào chủ được biến đổi hóa học.
Đáp án: E
Câu 14. Khi E. coli được nuôi trên môi trường chứa hỗn hợp glucôzơ và lactôzơ, động học về sự
tăng trưởng của vi khuẩn thu được như đồ thị dưới đây.
Chú thích hình: Bacterial cells/mL = Số tế bào vi khuẩn / mL
Time (min) = Thời gian (phút).
a. Cặp đồ thị nào dưới đây phản ánh đúng sự thay đổi nồng độ glucôzơ trong môi trường và sự thay đổi hoạt tính của β-galactosidaza trong tế bào?
Chú thích hình: Glucose Concentration = Nồng độ glucôzơ;
β-galactosidase activity = Hoạt tính β-galactosidaza; (arbitrary units) = (đơn vị tương đối); Time (min) = phút



Đáp án: Cặp đồ thị hình D
b. Đồ thị dưới đây cho thấy kiểu biểu hiện mRNA lac ở các tế bào E. coli kiểu dại và kiểu đột biến sau khi lactôzơ được bổ sung vào môi trường đã cạn kiệt glucôzơ.
Hãy chỉ ra, trong các thể đột biến dưới đây, các thể đột biến nào có thể và các thể đột biến nào không thể có kiểu biểu hiện giống thể đột biến ở đồ thị trên bằng cách đánh dấu (√) vào các ô tương ứng trong Phiếu trả lời.
I. Thể đột biến ở E. coli mất khả năng biểu hiện protein ức chế (repressor).

II. Thể đột biến ở E. coli mà ở đó protein ức chế có khả năng liên kết vào trình tự chỉ huy (operator) nhưng không có khả năng liên kết với lactôzơ.
III. Thể đột biến ở E. coli mà ở đó promoter bị đột biến dẫn đến protein ức chế không còn khả năng liên kết vào nó.
IV. Thể đột biến ở E. coli mà ở đó ARN polymerase không có khả năng liên kết vào trình tự khởi đầu phiên mã (promoter) của operôn lac.
Thể đột biến Có thể biểu hiện Không thể biểu hiện
Câu 15. Dưa cải muối chua là món ăn quen thuộc của chúng ta. Vi sinh vật thường thấy trong dịch lên men gồm vi khuẩn lactic, nấm men và nấm sợi. Hình dưới đây thể hiện số lượng tế bào sống (log CFU/ml) của 3 nhóm vi sinh vật khác nhau và giá trị pH trong quá trình lên men lactic dưa cải. Ôxi hòa tan trong dịch lên men giảm theo thời gian và được sử dụng hết sau ngày thứ 22.
a. Nguyên nhân nào làm giá trị pH giảm từ ngày thứ nhất đến ngày thứ ba?
Chú thích hình:
Level of lac mRNA = Mức biểu hiện của mARN lac;



Wild-type = Kiểu dại;
Mutant = Thể đột biến;
Time = Thời gian;
Lactose = Lactôzơ
b. Tại sao nấm men sinh trưởng nhanh từ ngày thứ 10 đến ngày thứ 26 và giảm mạnh sau ngày thứ 26?
c. Vì sao nấm sợi vẫn duy trì được khả năng sinh trưởng vào giai đoạn cuối của quá trình lên men?
544
Đáp án:
a. - pH giảm do lượng axit được vi sinh vật tạo ra nhiều và giải phóng vào môi trường, trong đó chủ yếu là axit lactic do vi khuẩn lactic sinh ra, ngoài ra còn có các axit hữu cơ khác như axit pyruvic, các axit trung gian trong chu trình crep…
- Tương ứng với đồ thị: vi khuẩn lactic tăng nhanh số lượng từ ngày 1 đến ngày 3 (đạt số lượng cao nhất vào khoảng ngày 5, 6).
b. - Môi trường có độ pH từ 3,5 – 4,0 thuận lợi cho sự phát triển của nấm men
→ Nấm men sinh trưởng nhanh từ ngày thứ 10, và đạt số lượng lớn nhất vào khoảng ngày 22 – 26.

- Ôxi hòa tan trong dịch lên men giảm theo thời gian và được sử dụng hết sau ngày thứ 22.
- Nấm men chuyển từ hô hấp hiếu khí sang hô hấp kị khí nên quá trình sinh trưởng giảm. Độ pH thấp duy trì lâu (dưới 3,5) ức chế sự sinh trưởng của nấm men → Số lượng nấm men giảm mạnh sau ngày 26.
c. Nấm sợi vẫn duy trì được khả năng sinh trưởng vào giai đoạn cuối của quá trình lên men vì:
- Nấm sợi có khả năng chịu đựng cao với môi trường có độ pH thấp (pH dưới 3,5).
- Càng về cuối quá trình lên men, sự hoạt động mạnh của vi khuẩn lactic và nấm men làm môi trường lên men càng trở nên ưu trương. Nấm sợi có khả năng chịu áp suất thấm thấu cao.
Câu 16. Khả năng hấp thụ saccharose của một chủng vi khuẩn sống ở biển được xác định bằng việc nuôi các tế bào này trong môi trường có saccharose (là nguồn carbon duy nhất) được đánh dấu phóng xạ 14 C trong thời gian ngắn. Sau đó, các tế bào được thu, rửa và đo sự có mặt của saccharose đã được đánh dấu phóng xạ 14 C. Sự hấp thụ saccharose theo thời gian được đo ở các môi trường có bổ sung Na+, K+, Li+, Na+ và chất X (chất ức chế tạo gradient H+). Kết quả nghiên cứu khả năng hấp thụ saccharose của các tế bào vi khuẩn này được thể hiện ở bảng dưới đây:
a. Sự hấp thu saccharose được thực hiện theo cơ chế đồng vận chuyển (Na+/saccharose).
- Sự hấp thu saccharose là tích cực, tiêu tốn năng lượng (ATP)
- Sự hấp thu đạt hiệu quả cao khi có mặt Na+ và bị ức chế khi có mặt của X, một chất ức chế tạo gradient proton, ức chế tạo ATP.
b. Sự có mặt của Li+ và K+ → sự hấp thụ saccharose không tăng cao như sự có mặt của Na+. Vì các ion này được vận chuyển chậm qua hệ thống đồng vận chuyển Na+/saccharose dẫn đến saccharose được hấp thụ chậm.
Câu 17. Khi một enzyme có mặt ở một loài vi khuẩn thì con đường chuyển hóa mà enzyme đó tham gia thường tồn tại trong loài vi khuẩn này.
- Bảng 17.1 là tên enzyme và phản ứng mà enzyme xúc tác được sử dụng làm chỉ thị cho sự xuất hiện của các con đường chuyển hóa mà nó tham gia.
Bảng 17.1 Enzyme và phản ứng xúc tác tương ứng
- Bảng 17.2 thể hiện sự có mặt hay vắng mặt của các enzyme ở bốn loài vi khuẩn khác nhau 1, 2, 3 và 4.


Bảng 17.2 Sự có mặt (+) và vắng mặt (-) của mỗi loại enzyme trong từng loài vi khuẩn.
a. Hãy cho biết sự hấp thu saccharose ở vi khuẩn trên được thực hiện theo cơ chế nào? Giải thích.
b. Giải thích tác động của K+, Li+ lên sự hấp thu saccharose.
545
Hãy cho biết:
a. Loài vi khuẩn nào KHÔNG thể thực hiện được hô hấp hiếu khí? Giải thích.
b. Các sản phẩm chính mỗi loài vi khuẩn tạo ra sau quá trình chuyển hóa glucose.
Đáp án:
a. - Các vi khuẩn không thể thực hiện được hô hấp hiếu khí gồm: loài 1, loài 2 và loài 3.
- Giải thích: do loài 1 và loài 2 thiếu cytochrome c oxidase là enzyme chính trong thành phần của quá trình hô hấp hiếu khí (chuỗi truyền điện tử), loài 3 thiếu enzyme citrate synthase của chu trình krebs.
b.
- Loài 1: tạo rượu ethanol, CO2 và ATP.
- Loài 2: tạo acid lactic và ATP.
- Loài 3: tạo acid lactic và ATP.
- Loài 4: khi có mặt oxi tạo CO2 và H2O còn khi không có mặt oxi tạo ethanol, CO2 và ATP .
Câu 18. a. Hoạt động của coenzim NADH trong hô hấp tế bào và quá trình lên men có gì khác nhau?
b. Vi khuẩn có những đặc tính cơ bản nào mà người ta dùng chúng trong các nghiên cứu di truyền học hiện đại?
c. Trong kỹ thuật di truyền, việc lựa chọn plasmit cần quan tâm đến những đặc điểm nào?
Đáp án:
a. - Thông thường hệ gen vi khuẩn là đơn bội (có 1 nst) nên gen khi bị đột biến sẽ biểu hiện ngay ra kiểu hình, sinh sản nhanh dễ nuôi cấy trong phòng thí nghiệm. - Có các plasmit có thể dùng làm thể truyền và có khả năng tạo biến dị tổ hợp như: biến nạp, tải nạp, tiếp hợp.
b. - Trong hô hấp tế bào, NADH đi vào chuỗi truyền điện tử để tổng hợp ATP, chất nhận H+ và e- cuối cùng là oxi.
- Trong quá trình lên men, NADH không đi vào chuỗi truyền e- mà nhường H+ và e- tới sản phẩm trung gian để hình thành axit lactic hoặc rượu, chất nhận H+ và ecuối cùng tạo nên axit lactic hoặc ethanol.
c. - Plasmit có kích thước ngắn, có gen chuẩn (gen đánh dấu), có điểm cắt của enzim giới hạn, có thể nhân lên nhiều bản sao trong tế bào nhận.
Câu 19. Có một ống hình chữ U, ở hai nhánh bổ sung hai chủng vi khuẩn kháng kháng sinh:
+ Một nhánh bổ sung chủng S. aureus kháng penixili- Chủng A
+ Một nhánh bổ sung chủng S. aureus kháng Chloramphenicol- Chủng B
547
Sau một thời gian nuôi cấy, người ta thấy ngoài hai chủng trên còn xuất hiện một chủng mới kháng cả penixilin và chloramphenicol (Chủng C).
a. Làm thế nào để chứng minh có sự tồn tại của chủng C?
b. Chủng vi khuẩn C có thể được hình thành bằng các cơ chế nào?
c. Nếu bổ sung enzim ADN - aza vào hai nhánh của bình chữ U thì chủng C có được tạo ra không? Tại sao? Đáp án:
a. Chứng minh có sự tồn tại của chủng C:
- Sau một thời gian nuôi cấy cả hai chủng A và B trong ống hình chữ U, lấy các tế bào trong ống hình chữ U.
- Sử dụng môi trường có cả hai loại kháng sinh penixilin và chloramphenicol để nuôi cấy các tế bào. Nếu có tế bào sống sót thì chứng tỏ có sự tồn tại của chủng C.
b. Chủng vi khuẩn C có thể được hình thành bằng các cơ chế tiếp hợp, tải nạp, biến nạp.
c. Nếu bổ sung enzim ADN-aza vào hai nhánh của bình chữ U thì chủng C vẫn có thể được tạo ra bằng tiếp hợp hoặc bằng tải nạp nhưng không thể bằng biến nạp vì các ADN-aza sẽ phân giải các plasmit khi ở ngoài tế bào.
Câu 20. Dòng nước chảy ra từ các mỏ khai thác sắt ở Thái Nguyên chứa nhiều ion sắt, sulphate và một số ion kim loại khác. Dòng nước này chảy vào sông, suối, ao, hồ sẽ gây ô nhiễm, làm cho sinh vật thủy sinh chết hàng loạt. Người ta xử lý loại bỏ sắt của nước thải này bằng cách sử dụng vi khuẩn khử lưu huỳnh. Cho dòng nước thải chảy qua tháp phản ứng (là một hệ thống kín) có nhồi chất hữu cơ như rơm, rạ đã được trộn vi khuẩn khử lưu huỳnh thì nước thu được từ tháp phản ứng không còn một số ion, đáy tháp có kết tủa màu đen. Hãy giải thích:
a. Vi khuẩn khử lưu huỳnh có kiểu dinh dưỡng nào?
b. Chất hữu cơ (rơm, rạ) và sulfate có tác dụng gì?
c. Kết tủa có màu đen ở đáy tháp là gì?
Đáp án:
a. Vi khuẩn khử lưu huỳnh là vi khuẩn dị dưỡng. Chúng tiến hành hô hấp kị khí tạo ra năng lượng cho các hoạt động sống.
b. Chất hữu cơ là chất cho điện tử; sulfate là chất nhận điện tử trong hô hấp kị khí.
c. Sản phẩm của quá trình khử lưu huỳnh là khí sulfua. Sulfua kết hợp với kim loại tạo thành hợp chất sunfua-kim loại (trong trường hợp này là FeS). FeS có màu đen và được tạo thành kết tủa ở đáy của tháp phản ứng.
Câu 21. Có 5 chất kháng sinh (A, B, C, D và E) được kiểm tra về hiệu lực chống vi khuẩn gây bệnh Staphylococcus aureus. Với từng chất kháng sinh, người ta tẩm ướt một khoanh giấy thấm tròn với dịch chứa 2mg chất kháng sinh tương ứng rồi lần lượt đặt chúng lên môi trường thạch nuôi cấy vi khuẩn Staphylococcus aureus, kết quả thu được như hình 1 dưới đây. Được biết 5 chất kháng sinh này gây độc với người ở các liều lượng khác nhau như số liệu trình bày trên hình 2.

+ Kháng sinh A và D sử dụng liều cao mới gây độc đối với người, nhưng không có hiệu lực hoặc có hiệu lực thấp đối với Staphylococcus aureus.
+ Kháng sinh C sử dụng liều thấp (< 2mg) đã gây độc đối với người và có hiệu lực thấp.
+ Kháng sinh E có hiệu lực diệt vi khuẩn Staphylococcus aureus rất cao, nhưng sử dụng liều thấp (< 1mg) đã gây độc đối với người.
Câu 22. Để đo nồng độ vi khuẩn có trong dịch X, các nhà nghiên cứu có thể dùng 2 phương pháp:
(1) Pha loãng nhiều bước rồi đo nồng độ dịch cuối cùng Theo đó, lấy 1ml dịch vi khuẩn ban đầu cho vào 8ml nước ở ống 1, rồi tiếp tục cho đến ống 3. Sau đó, lấy 1 ml từ ống 3 quét lên đĩa petri rồi ủ trong 5h, sau đó quan sát sự hình thành khuẩn lạc.
a. Hãy sắp xếp hiệu lực diệt vi khuẩn Staphylococcus aureus của 5 loại thuốc kháng sinh (A → E) theo thứ tự giảm dần? Giải thích.
b. Ở liều dùng 2mg, kháng sinh nào (A → E) vừa an toàn cho người sử dụng vừa có hiệu lực diệt vi khuẩn Staphylococcus aureus cao? Giải thích.
Đáp án:
a. Theo hình 1:
+ Thuốc kháng sinh E có vùng vi khuẩn không mọc rộng nhất → có hiệu lực cao nhất.
+ Thuốc kháng sinh D và C có vùng vi khuẩn không mọc gần bằng nhau, nhưng nhỏ hơn thuốc kháng sinh E → D và C có hiệu lực bằng nhau, nhưng thấp hơn E.
+ Thuốc kháng sinh B có vùng vi khuẩn không mọc nhỏ hơn E nhưng > D và C.
+ Thuốc kháng sinh A không có vùng vi khuẩn không mọc → không có hiệu lực đối với vi khuẩn Staphylococcus aureus → hiệu lực diệt vi khuẩn Staphylococcus aureus của 5 loại thuốc kháng sinh (A → E) theo thứ tự giảm dần là:
E > B > D = C > A.
b. Ở liều dùng 2mg, chỉ có kháng sinh B là vừa an toàn cho người sử dụng vừa có hiệu lực diệt vi khuẩn Staphylococcus aureus cao.
Vì:
549
(2) Đo độ hấp thụ ánh sáng bằng máy quang phổ Chiếu ánh sáng có bước sóng 600nm và đo độ hấp thụ ánh sáng của dung dịch (A600). Đo được A600 của dịch X là 3,75 x 10-7 (A600 = 1 tương đương với 8x108 tế bào/ml)
Cho 2 phage lây nhiễm vào 1ml dung dịch X. Biết phage này không làm tan tế bào khi xuất bào, tế bào mẹ nhiễm phage chỉ truyền phage cho 1 tế bào con. Thời gian thế hệ của vi khuẩn là 20 phút. Thời gian từ khi phage xâm nhập đến khi chuẩn bị lắp ráp là 10 phút, thời gian phage lắp ráp xuất bào và xâm nhập vào tế bào khác là 5 phút. Trong một tế bào, một phage gây sinh tổng hợp nên 32 phage mới. Để xác định thời điểm mà số lượng phage bằng số lượng vi khuẩn, An đã lập phương trình sau, với
550

N là số lượng vi khuẩn ban đầu, t là thời gian cần thiết để số lượng phage và vi khuẩn đạt bằng nhau.
N x 2t/20 = 2 x 32t/15
a. Tính nồng độ vi khuẩn trong dịch X theo phương pháp (1)
b. Tính nồng độ vi khuẩn trong dịch X theo phương pháp (2)
c. Vì sao lại có sự khác nhau giữa kết quả của 2 phương pháp?
d. Tính t theo phương trình của An.
e. Kết quả t theo phương trình của An có đúng không? Nếu không, khả năng là do đâu? Thật sự có tồn tại thời điểm mà số phage bằng số vi khuẩn không?
Đáp án:
a. - Hệ số pha loãng là: (1 x 2 x 2)/(9 x 10 x 3) = 2/135
- Nồng độ dịch X = số khuẩn lạc : (hệ số pha loãng) = 4 : 2/135 = 270 (vk/l)
b. Nồng độ dịch X: 8 x 108 x 3,75 x 10-7 = 300 (vk/l)
c. Khác nhau là do việc đo độ hấp thụ của dịch không phân biệt được đâu là vi khuẩn sống, đâu là xác vi khuẩn đã chết.
d. Thay N = 270 → 300 x 2t/20 = 2x 32t/15 → t = 25p
e. - Sau 25 phút, cả vi khuẩn lẫn phage mới chỉ nhân lên 1 lần, theo đó, có 540 vi khuẩn và 64 phage, vậy phương trình trên sai.
- Lí do là vì sự tăng số lượng của một thể sinh học là không đều theo thời gian, theo đó, trong 20 phút thì chỉ có thời điểm cuối cùng vi khuẩn mới nhân đôi, phage nhân số lượng chỉ trong 5 phút cuối của 15 phút trong một chu trình. Kết quả 25 phút trên ám chỉ rằng tại mọi thời điểm trong 25 phút, sự tăng số lượng luôn diễn ra.
- Sau 30 phút, số lượng phage tăng lên 2048, còn sau 40 phút thì số lượng vi khuẩn mới đạt 1080 con, lúc này, phage đã đạt 2048. Như vậy, không thời điểm nào số lượng hai thể sinh học này bằng nhau vì phage tăng số lượng nhanh hơn vi khuẩn.
Câu 23: Giả sử em xác định được 1 gen ở nấm men rượu, tương đồng với gen mã hóa tiểu đơn vị của enzyme telomerase từ động vật nguyên sinh. Sau đó em tạo môt đột biến đích làm mất một bản sao của gen ở chủng lưỡng bội của nấm men và sau đó cảm ứng để tạo bào tử và sinh ra các tế bào đơn bội. Tất cả bốn bào tử nảy mầm tốt, và em có thể tạo khuẩn lạc trên đĩa thạch. Cứ sau 3 ngày, em cấy ria lại các khuẩn lạc lên môi trường thạch đĩa mới. Sau 4 lần cấy chuyển liên tục như vậy, các thế hệ sau của 2 trong số 4 bào tử ban đầu sinh trưởng rất kém, thậm chí không sinh trưởng. Em lấy các tế bào từ các đĩa gốc nuôi 3, 6 và 9 ngày để tách chiết ADN, sau đó cắt mẫu ADN ở vị trí cách điểm khởi đầu của vùng lặp lại ở đầu mút của nhiễm sắc thể khoảng 35 nucleotit. Phân tách các phân đoạn bằng điện di, và lai chúng với mẫu dò đặc hiệu đầu mút đánh dấu phóng xạ (các băng màu tối) (Hình Q.52). Giả thiết là thời gian thế hệ là 6 giờ
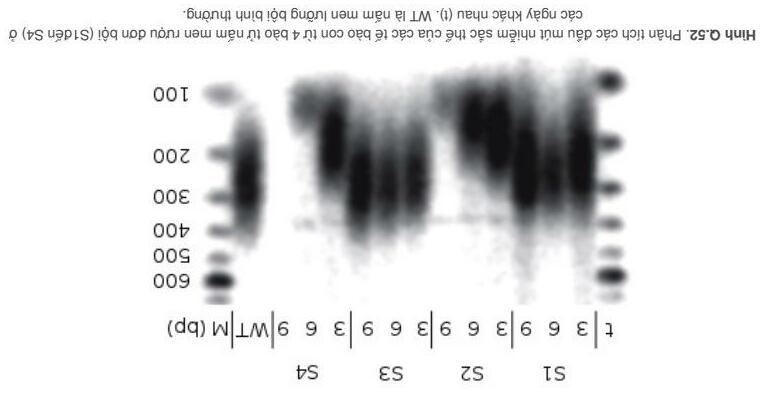
551
Hãy chỉ ra các câu sau là Đúng hay Sai
A. Độ dài trung bình của đầu mút của nấm men rượu là 300 nucleotit.
B. Bào tử 2 và 4 có vẻ thiếu enzyme tổng hợp đầu mút telomerase.
C. Đầu mút nhiễm sắc thể nấm men rượu mất không quá 20 nucleotit sau mỗi lần nhân đôi nhiễm sắc thể.
D. Các nấm men rượu mất các đầu mút sẽ có kích thước tế bào bình thường.
Đáp án:
- Phát biểu sai: A, B
- Phát biểu đúng: C, D
Câu 24. Mr. Trung nhân dòng 1 trình tự mã hóa (coding sequence-CDS) của một gen vào một vector rồi đặt tên plasmid tạo thành là pVN2016. CDS được chèn tại vị trí nhận biết SacII nằm trong vùng MSC (multi cloning site) trong vùng gen lacZ của vector (Hình Q.93A). CDS được chèn có một vị trí cắt giới hạn Pstl cách bộ ba kết thúc của nó 0,8 kb. Để xác định kích thước và hướng của CDS đã được chèn, Mr. Trung đã cắt plasmid này với enzyme cắt giới hạn khác nhau, và sản phẩm cắt được trình bày trong hình Q.93B.
Dựa vào dữ liệu ở trên, hãy cho biết câu nào sau đây là đúng hay sai
A. CDS dài 2.6 kb và có vị trí nhận biết EcoRI ở điểm cách một đầu tận cùng của nó khoảng 0,5 kb.

B. SpeI có thể được sử dụng để xác định hướng của CDS.
C. CDS được chèn vào cùng chiều với lacZ.
D. Nếu plasmid pVN2016 bị cắt bởi cả hai enzyme SpeI và EcoRI trong đệm Tango (EcoRI và SpeI cắt ở mức tương ứng là 100% và 20%), thì 5 đoạn với kích


thước 0,5; 0,8; 1,3; 2,1 và 3,0 kb có thể được xác định bởi điện di trên gel, giả sử
khôngquan sát được các đoạn nhỏ hơn 50 bp.
Đáp án
- Phát biểu đúng: A, D
- Phát biểu sai: B, C
Câu 24. Michael Smith (1932-2000) đã phát minh kĩ thuật gây đột biến hướng đích. Gần đây các nhà khoa học đã phát minh công nghệ CRISPR-Cas9 để tiến hành gây đột biến dễ dàng hơn. Protein Cas9 từ Streptococcus pyogenes được dẫn bởi một guideARN (ARN chỉ dẫn, gARN) dài 20 bp liên kết với ADN đích. Enzyme Cas9 này chỉ có thể cắt sợi kép ADN tại vị trí cách ba nucleotide nằm ngược dòng trình tự 5'NGG-3' (1). Các phân đoạn ADN sợi kép đã bị cắt bởi các enzyme này, và sau đó vẫn có thể được nối trở lại. Gen X cần được làm mất chức năng nhờ enzyme Cas9 được mô tả ở đây (2). Một cách khác để knockout gen là sử dụng một cặp gồm Cas9 cải biến có khả năng chỉ tạo ra các vết cắt sợi đơn và ARN chỉ dẫn (guideRNA) hướng đích tới các vị trí liền kề
Chọn trình tự guideARN sẽ chỉ dẫn Cas9 tới vị trí I.


1 = 5'-ATTTTTCGACGTTTAGGCCA-3'
2 = 5'-AUUUUUCGACGUUUAGGCCA-3'
3 = 5'-AUAAAACGUGCAAAUCCGGU-3'
4 = 5'-UUUGCACGUUUAGGCCAAGG-3'
5 = 5'-UUUUUGGGGGCCCAGGAUCU-3'
Mỗi phát biểu dưới đây là đúng hay sai?
A. Sử dụng cặp đôi Cas9 và guideARN bị biến đổi làm tăng các tổn thương sai đích ở các gen khác.
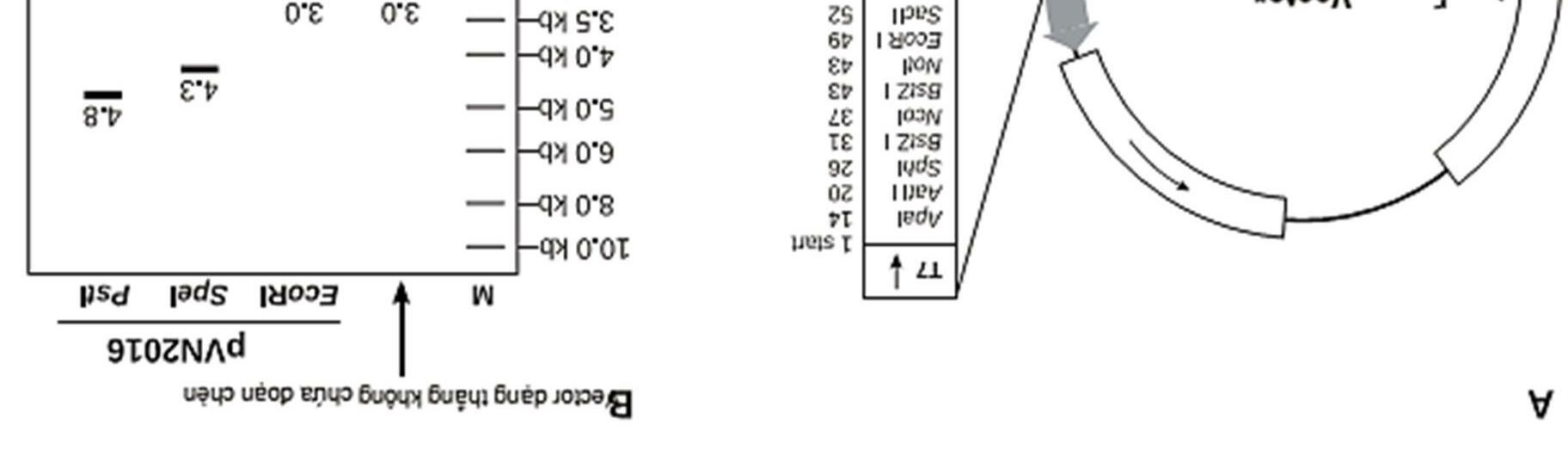
B. Hệ gen Streptococcus pyogenes ngẫu nhiên có nhiều vị trí NGG hơn mong đợi.
C. Nếu gen không có các trình tự GG, các protein Cas9 từ các loài khác cần được nghiên cứu.
Đáp án:
- Trình tự đúng: 2 = 5 – AUUUUC …………………..
- Phát biểu đúng: C
- Phát biểu sai: A và B
Câu 25.
a. Khi E.Coli được nuôi trên môi trường chứa hỗn hợp glucozơ và lactozơ, sự tăng trưởng của vi khuẩn được ghi lại theo đồ thị bên. Căn cứ vào đồ thị, hãy cho biết:
- Nồng độ glucôzơ trong môi trường nuôi cấy cao nhất và thấp nhất ở thời điểm nào? Giải thích.
- Vào thời điểm nào của quá trình nuôi cấy thì vi khuẩn tiết ra enzym Galactosidaza. Giải thích.
b. Cho vi khuẩn phản nitrat hóa vào bình nuôi cấy đựng dung dịch KNO3 , glucôzơ và các nhân tố cần thiết cho sự sinh trưởng, sau đó đậy kín bình lại. Sau một thời gian hãy nhận xét về sự biến đổi của hàm lượng ôxi, N2 và CO2. Giải thích.
Đáp án:
a. - Nồng độ glucose cao nhất ở thời điểm bắt đầu nuôi cấy (0phút) : khi đó vi khuẩn chưa sử dụng glucose.
- Nồng độ glucose thấp nhất ở khoảng thời gian 100 phút : khi đó vi khuẩn sử dụng hết glucose.
- Theo đồ thị thì ở khoảng sau phút thứ 100, nguồn dinh dưỡng glucozo đã cạn kiệt vi khuẩn sử dụng lactozo, lúc này enzym Glactosidaza được tiết ra.
b. - Nhận xét: Hàm lượng O2 giảm, hàm lượng N2 và CO2 tăng.
- Ban đầu vi khuẩn hô hấp hiếu khí nên sử dụng hết ôxi có trong bình.
- Vi khuẩn chuyển sang hô hấp khị khí, sử dụng NO3 - làm chất nhận điện tử và giải phóng N2.
- Hô hấp hiếu khí và kị khí đều thải CO2.
Câu 26. a. Người ta cho 80 ml nước chiết thịt (thịt bò hay thịt lợn nạc) vô trùng vào hai bình tam giác cỡ 100 ml (kí hiệu là bình A và B), sau đó cho vào mỗi bình 0,50 gam đất vườn được lấy ở cùng vị trí và thời điểm. Cả hai bình đều được bịt kín bằng nút cao su, đun sôi (100oC) trong 5 phút và đưa vào phòng nuôi cấy có nhiệt độ từ 30-35oC. Sau 1 ngày người ta lấy bình thí nghiệm B ra và đun sôi (100oC) trong 5 phút, sau đó lại đưa vào phòng nuôi cấy. Sau 3 ngày cả hai bình thí nghiệm được mở ra thì thấy bình thí nghiệm A có mùi thối, còn bình thí nghiệm B gần như không có mùi thối. Giải thích.
b. Trong sản xuất các chế phẩm vi sinh vật, người ta có thể dùng phương pháp nuôi cấy liên tục và không liên tục. Giả sử có 2 chủng xạ khuẩn, một chủng có khả năng sinh enzim A, một chủng khác có khả năng sinh kháng sinh B. Hãy chọn phương pháp nuôi cấy cho mỗi chủng xạ khuẩn để thu được lượng enzim A, kháng sinh B cao nhất và giải thích lí do chọn?
Đáp án:
a. - Ở phương pháp nuôi cấy liên tục, người ta thường xuyên bổ sung chất dinh dưỡng và lấy đi một lượng dịch nuôi tương đương, tạo được môi trường ổn định, do vậy VSV sinh trưởng ổn định ở pha lũy thừa. Enzim là sản phẩm bậc I được hình thành ở pha tiềm phát và pha lũy thừa, vì vậy chọn phương pháp nuôi cấy liên tục là thích hợp nhất, thu được lượng enzim A cao nhất.
- Ở phương pháp nuôi cấy không liên tục (từng mẻ), sự sinh trưởng của VSV diễn ra theo đường cong gồm 4 pha: tiềm phát, lũy thừa, cân bằng và suy vong.
Chất kháng sinh là sản phẩm bậc II được hình thành ở pha cân bằng, pha này cho lượng kháng sinh nhiều nhất (nuôi cấy liên tục không có pha cân bằng), vì vậy chọn phương pháp nuôi cấy không liên tục là thích hợp nhất, thu được lượng kháng sinh B cao nhất.
b. Trong 0,5 g đất chứa nhiều mầm vi sinh vật, ở nhiệt độ sôi 100oC các tế bào dinh dưỡng đều chết, chỉ còn lại nội bào tử (endospore) của vi khuẩn.
- Trong bình thí nghiệm A, các nội bào tử vi khuẩn sẽ nảy mầm và phân giải protein của nước thịt trong điều kiện kị khí. Nước thịt là môi trường dư thừa hợp chất nitơ và thiếu hợp chất cacbon, nên những vi khuẩn kị khí sẽ khử amin giải phóng NH3, H2S để sử dụng cacbohydrat làm nguồn năng lượng trong lên men.
- Vì vậy, khi mở nắp ống nghiệm các loại khí NH3, H2S bay lên gây thối rất khó chịu, còn gọi là quá trình amôn hoá kị khí là lên men thối.
- Trong bình thí nghiệm B, các nội bào tử này mầm hình thành tế bào dinh dưỡng chúng bị tiêu diệt sau 1 ngày bị đun sôi lần thứ hai, do đó protein không bị phân giải, kết quả không có mùi.
Câu 27. Nghiên cứu sự điều hòa biểu hiện gen của Operon lac ở một chủng E. coli đột biến, người ta phát hiện thấy có bất thường. Để xác định nguyên nhân của sự bất thường đó xảy ra ở vị trí nào trong Operon Lac, người ta đánh giá mức độ biểu hiện của gen LacZ ở chủng E. coli đột biến này trong các điều kiện môi trường nuôi cấy khác nhau.
Bảng 1. Mức độ biểu hiện của gen LacZ trong các điều kiện nuôi cấy.
Lactose Glucose

Điều kiện môi trường nuôi cấy Mức độ biểu hiện gen
Có Không 100
555
556
a. Từ thông tin ở bảng 2, hãy cho biết nguyên nhân bất thường xảy ra ở vị trí nào trên operon Lac ở chủng E. coli đột biến này? Giải thích.
b. Tại sao từ một phân tử mARN được phiên mã từ Operon Lac nhưng các gen khác nhau lại được dịch mã với tốc độ khác nhau.
Đáp án:
a. Bất thường xảy ra ở vùng vận hành Operator.
Vì: Môi trường không có lactose và glucose nhưng gen LacZ vẫn biểu hiện
=> protein ức chế không gắn được vào vùng vận hành Operator
Khi môi trường có glucose thì hàm lượng AMPv giảm, CAP không tạo phức hợp với AMPv, RNA polymerase không hoạt động, gen LacZ không biểu hiện.

b. Giải thích:
- Tốc độ dịch mã được xác định bởi sự có mặt của các loại tRNA khác nhau, tương ứng với các codon đặc hiệu trong mRNA.
- Hàm lượng tương đối của các tRNA khác nhau trong tế bào về cơ bản là khác nhau.
- Do đó, các gen khác nhau sẽ có thành phần các codon khác nhau, đoạn mRNA nào có chứa nhiều codon "hiếm" thì sự dịch mã diễn ra chậm hơn các mRNA có chứa nhiều codon thông dụng.
Câu 28. Để tổng hợp một loại protein đơn giản của người nhờ vi khuẩn qua kỹ thuật DNA tái tổ hợp, người ta có 2 cách: 1) cách thứ nhất: tách gen mã hóa protein trực tiếp từ hệ gen trong nhân tế bào, rồi cài đoạn gen đó vào plasmid của vi khuẩn nhờ enzyme ligase; 2) Cách thứ hai: tách RNAm trưởng thành của gen mã hóa protein đó, sau đó dùng enzyme phiên mã ngược tổng hợp lại gen (cDNA), rồi cài đoạn cDNA này vào plasmid nhờ enzyme ligase. Trong thực tế, người ta thường chọn cách nào? Tại sao?
Đáp án:
Trong thực tế, người ta chọn cách thứ 2. Vì:
- DNA tách trực tiếp từ hệ gen người thường mang intron, còn cDNA được tổng hợp từ RNAm không mang intron.
- Các tế bào vi khuẩn không có khả năng cắt bỏ các intron của các gen Eukaryote, nên đoạn DNA cài tách trực tiếp từ nhân không tạo ra protein bình thường.
- Đoạn DNA phiên mã ngược chính là bản sao tương ứng của RNAm dùng để dịch mã protein, có kích thước ngắn hơn nên dễ tách dòng và biểu hiện gen trong điều kiện in vitro.
Câu 29. Hình 5 mô tả cơ chế điều hòa tổng hợp enzyme tổng hợp trytophan ở vi khuẩn E. coli.
a. Khi môi trường có hiện tượng dư thừa tryptophan so với nhu cầu của tế bào thì cơ chế điều hòa sẽ hoạt động như thế nào?
b. Người ta sử dụng một loại thuốc gây đột biến để làm sai hỏng cơ chế điều hòa tổng hợp enzyme để tạo nên chủng E. coli đột biến có khả năng tổng hợp liên tục trytophan. Hãy cho biết, thuốc gây đột biến có thể đã tác động đến vị trí nào trong cơ chế điều hòa tổng hợp enzyme trên? Giải thích.
Đáp án:
a. Khi môi trường dư thừa tryptophan:
- Tryptophan sẽ gắn vào protein ức chế, làm thay đổi cấu hình protein ức chế.
- Lúc này protein ức chế có khả năng gắn với Operator làm ngừng phiên mã.
- Các enzyme tổng hợp tryptophan không được tổng hợp, tế bào vẫn sử dụng tryptophan làm cho hàm lượng tryptophan giảm dần.
- Khi hàm lượng tryptophan xuống mức thấp, không gắn với protein ức chế, do đó protein này không gắn được với Operator, các enzyme được tổng hợp và hàm lượng tryptophan tăng dần.
b. - Thuốc gây đột biến có thể đã tác động đến vùng gen điều hòa (Regulator) vì sai hỏng trong việc tổng hợp protein ức chế làm cho protein này không được tổng hợp hoặc tổng hợp được nhưng không gắn được với tryptophan để ức chế Operon. Do đó, tryptophan được tổng hợp liên tục.
- Thuốc gây đột biến có thể đã tác động đến vùng gen vận hành Operator làm mất khả năng gắn kết với protein ức chế. Do đó, tryptophan được tổng hợp liên tục.
Câu 30. Cho sơ đồ chuyển gen nhờ plasmid vào vi khuẩn đường ruột E. coli như hình 7.
558
a. Điền các cấu trúc thích hợp tương ứng với các vị trí đánh số trên sơ đồ hình 7.
b. Để phục vụ cho quá trình phân tích, xử lí các chế phẩm DNA, người ta sử dụng 2 nhóm enzyme phục vụ hai hướng chính là enzyme phân cắt và enzyme nối. Enzyme phân cắt DNA được gọi là nuclease, gồm 2 nhóm là endonuclease và exonuclease. Enzyme nối DNA thường dùng là E. coli DNA ligase và T4 DNA ligase. Dựa vào chức năng, em hãy cho biết các enzyme này khác nhau như thế nào?
Đáp án:
a. 1. Đoạn gen (DNA) cần chuyển; 2. Plasmid làm thể truyền; 3. DNA tái tổ hợp;
4. DNA tái tổ hợp trong tế bào vi khuẩn; 5. DNA của vi khuẩn; 6. DNA tái tổ hợp của thế hệ tiếp theo.
b. - Endonuclease: cắt DNA ở giữa phân tử
- Exonuclease: cắt DNA từ 2 đầu mút của phân tử DNA.
- E. coli DNA ligase: nối hai trình tự DNA có đầu so le.
- T4 DNA ligase: nối hai trình tự DNA có đầu so le hoặc đầu bằng.
Câu 31. Trong hoạt động của operon Lac ở vi khuẩn E. coli, nếu đột biến xảy ra ở gen điều hòa R (còn gọi là lac I) thì có thể dẫn đến những hậu quả gì liên quan đến sự biểu hiện của các gen cấu trúc?
Đáp án:
Nếu đột biến xảy ra ở gen R có thể dẫn đến các hậu quả sau:
- Xảy ra đột biến câm, Operon Lac hoạt động bình thường, sự biểu hiện của các gen cấu trúc bình thường. Gồm các trường hợp sau:
+ Đột biến nucleotit trong gen này không làm thay đổi trình tự axit amin trong protein ức chế
+ Đột biến thay đổi axit amin trong chuỗi polypeptit của protein ức chế không làm thay đổi khả năng liên kết của protein ức chế với trình tự chỉ huy (O).
- Xảy ra đột biến làm giảm khả năng liên kết của protein ức chế vào trình tự chỉ huy → sự biểu hiện của các gen cấu trúc tăng lên.
- Đột biến làm mất hoàn toàn khả năng liên kết của protein ức chế hoặc protein ức chế không được tạo ra → các gen cấu trúc biểu hiện liên tục.
- Xảy ra đột biến làm tăng khả năng liên kết của protein ức chế vào trình tự chỉ huy → sự biểu hiện của các gen cấu trúc giảm đi.
Câu 32. a. Các locut gen dưới đây cùng tham gia vào Operon Lac:

z = gen cấu trúc mã hoá enzim β galactosidase
i = gen ức chế
o = operator


Hãy xét xem các chủng vi khuẩn có các kiểu gen dưới đây có thể tổng hợp được enzim β galactosidase không, và nếu được thì trong điều kiện nào?
Kiểu gen các chủng β galactosidase (+ = có, - = không)
Không có lactose Có lactose
1. io+z+
2. io+z -
3. i+o+z -/io+z+
4. i+o+z+/i+oz -
b. Trong các Operon dưới đây, r = gen ức chế, p = promoter và a, b, c là các gen cấu trúc. Các locus gen bị đột biến và bất hoạt được kí hiệu bằng dấu trừ trên đầu các
560
chữ cái. Hãy xác định xem Operon nào trong các Operon dưới đây hoạt động liên tục, không hoạt động hoặc hoạt động điều hoà? Giải thích.
Chủng 1. rp+o+a+b+c+
Chủng 2. r+po+a+b+c+/ rp+o+a+b+c+
c. Gen a, b, c đều cần để tổng hợp một loại prôtêin. Những chủng nào dưới đây
có khả năng tổng hợp prôtêin đó và trong điều kiện nào?
Chủng 1. p+o+a+B-c+
Chủng 2. p+o+a -b+c+/ p+o+a+B-c+
Chủng 3. po+a+b+c+/ p+o+a+b+c -
a.
Đáp án:
Kiểu gen các chủng
β galactosidase (+ = có, - = không)
Không có lactose Có lactose
1. io+z+ + +
2. io+z- -
3. i+o+z -/io+z+ - +
4. i+o+z+/i+oz - - +
b. Chủng 1: Operon hoạt động liên tục, vì gen điều hòa bị bất hoạt, các gen khác bình thường.
Chủng 2: Operon hoạt động bình thường vì protein ức chế được tổng hợp ở dòng 1 cung cấp cho Operon dòng 2.
c. Chủng 2 tổng hợp được protein trên trong điều kiện có chất cảm ứng vì cả 3 gen a, b, c đều hoạt động bình thường.
Chủng 1 không tổng hợp được enzyme b, chủng 3 không tổng hợp được enzyme c nên cả 2 chủng không tổng hợp được protein trên.
Câu 33. Cho biết một Operon chịu cảm ứng có các thành phần nằm trong một vùng kề nhau theo trật tự PQRS. Operon này sinh ra hai loại enzym 1 và 2. Đột biến ở mỗi vùng sẽ sinh ra các enzym sau đây (+ = chỉ được sinh ra khi có chất cảm ứng, 0 = không có enzym, C = enzym được tạo ra một cách cơ định).
a. P, Q, R, S là kí kiệu của những Đ EE
561
vùng gen nào?
b. Dựa vào câu a hãy tiên đoán sự có
mặt của enzym 1 và 2 (+, 0 hoặc C) ở các
thể lưỡng bội một phần dưới đây.
Trường hợp 1: P+Q-R+S+/ P-Q+R+S+
Trường hợp 2. P-Q+R+S+/ P+Q-R+S-
Đáp án:
a. - Vùng R bị đột biến, các enzyme không được hình thành => R là vùng gen khởi động.
- Vùng S bị đột biến, các enzyme đều được tổng hợp cơ định => S là vùng gen chỉ huy.
- Đột biến ở P, enzyme 2 không được tổng hợp => P là vùng gen cấu trúc quy định enzyme 2.
- Đột biến ở Q, enzyme 1không được tổng hợp => Q là vùng gen cấu trúc quy định enzyme 1.
b. - Trường hợp 1: P+Q-R+S+/ P-Q+R+S+ :
+ Khi có chất cảm ứng, enzyme 1 và 2 đều được tổng hợp.
+ Khi không có chất cảm ứng, enzyme 1 và 2 không được tổng hợp.
- Trường hợp 2. P-Q+R+S+/ P+Q-R+S:
+ Khi có chất cảm ứng, enzyme 1 và 2 đều được tổng hợp.
+ Khi không có chất cảm ứng, enzyme 1 không được tổng hợp, enzyme 2 được tổng hợp cơ định vì vùng gen chỉ huy S không gắn được với protein ức chế
Câu 34. Phân tích ADN theo phương pháp Sanger cho kết quả điện di trên agarose của 2 mẫu kiểu dại (wild type) và mẫu đột biến (mutant) như hình vẽ bên dưới.

a. Hãy cho biết trình tự 5’ – 3’ của mẫu kiểu dại.
b. Hãy cho biết trình tự 5’ – 3’ của mẫu đột biến. Loại đột biến nào đã xảy ra?
Đáp án:
a. 5’TGGCGTAAAGTCTGGCATCC3’
b. 5’TGGCGTAAGTCTGGCATCC3’. Đột biến điểm dạng mất cặp nucleotit
A-T đã xảy ra làm khung đọc thay đổi từ bộ ba thứ 3 trở đi.
Câu 35. Để xác định chính xác điểm bắt đầu phiên mã (TSS) của một promoter của một gen mới được phát hiện ở vi khuẩn, một mồi được đánh dấu phóng xạ có trình tự bổ sung với đầu 3' của gen được dùng để giải trình tự theo phương pháp Sanger và
Kéo dài mồi của mARN. Sự kéo dài mồi của mARN (tương tự như tổng hợp cADN)
được lặp đi, lặp lại trên mARN được phiên mã khi bổ sung thêm yếu tố phiên mã α
Phát biểu nào là đúng hay sai?
A. Các loại polymerase khác nhau được dùng trong giải trình tự Sanger và để kéo dài mồi.
B. mARN của gen này có CUCAGUAC là 8 bazơ đầu tiên ngay sau TSS được tìm thấy trong các tế bào này.
C. Nhiều TSS tồn tại trong gen này.
D. Phiên mã được điều hòa chính bởi yếu tố α
Đáp án:
Các phân đoạn thu được sau đó được phân tách bởi điện di trên gel; hình ảnh phóng xạ tự chụp như sau:
A . Đúng
B . Đúng
Trong giải trình tự Sanger, người ta sử dụng enzyme DNA polymerase. Ngược lại, để kéo dài đoạn mồi, người ta sử dụng enzyme phiên mã ngược.


Làn PE trên kết quả điện di cho biết chiều dài của mRNA. Bazơ đầu tiên (đầu 5') của mRNA chính tương ứng với đoạn trình tự sắp xếp có cùng độ dài (tại G). Bazơ thứ hai của mRNAc tương ứng với trình tự sắp xếp thấp hơn một dải trên bản điện di (tại A).
C . Đúng Đối với gen này, có 1 TSS chính và 1 TSS phụ tồn tại.
D . Sai Phiên mã khi vắng mặt hoặc có mặt yếu tố α thì vẫn tổng hợp cùng một tỉ lệ mRNA bắt đầu từ TSS chính và phụ
563
1. Kết luận Như vậy, qua chuyên đề này tôi đã giải quyết những vấn đề sau đây:
- Giúp giáo viên và học sinh có cái nhìn khái quát về lý thuyết tích hợp hóa sinh – vi sinh – di truyền. Hệ thống hóa toàn bộ kiến thức cơ bản nhất cùng với nhiều hình ảnh minh họa, giúp giáo viên và học sinh thuận lợi hơn trong nghiên cứu, giảng dạy, ôn tập.
- Có nhiều câu hỏi, bài tập vận dụng dùng trong quá trình hướng dẫn học sinh nghiên cứu chuyên đề để giúp học sinh vận dụng, giải quyết những vấn đề thực tiễn và ôn tập, củng cố lại kiến thức.
- Bên cạnh đó nhờ nghiên cứu chuyên đề này tôi cũng rút ra kinh nghiệm cho việc giảng dạy kiến thức tích hợp hóa sinh – vi sinh – di truyền nói riêng và các chuyên đề khác của sinh học nói chung.
2. Kiến nghị
Việc tìm hiểu, nghiên cứu kiến thức tích hợp hóa sinh – vi sinh – di truyền là rất cần thiết để tạo ra những nghiên cứu đột phá về con người, thuốc thú y, nông nghiệp, công nghiệp, năng lượng và môi trường. Từ đó, có những tác động tích cực đến sự phát triển của các ngành khoa học khác và mọi mặt trong cuộc sống của con người. Vì vậy, cần được tạo điều kiện để học sinh học tập và nghiên cứu ngay ở bậc trung học.
Với thời gian hạn chế, kiến thức bản thân có hạn nên đề tài không tránh khỏi những sai sót, tôi rất mong nhận được sự góp ý của quý đồng nghiệp để đề tài được hoàn thiện hơn. Chân thành cảm ơn quý đồng nghiệp!
TÀI LIỆU THAM KHẢO
[1]. Campbell & Reece, Biology, NXB Giáo dục Việt Nam dịch và xuất bản năm 2011.
[2]. Nguyễn Lân Dũng, Nguyễn Đình Quyến, Phạm Văn Ty, Vi sinh vật học, NXB Giáo dục, 2007.
[3]. Nguyễn Thành Đạt, Cơ sở sinh học vi sinh vật, tập 1+2, NXB Giáo dục, 2000.
[4]. Nguyễn Thành Đạt (tổng chủ biên), Sách giáo khoa Sinh học 10, NXB Giáo dục, 2008.
[5]. Nguyễn Thành Đạt (tổng chủ biên), Sách giáo khoa Sinh học 12, NXB Giáo dục, 2008.
[6]. Madigan & Martinko, Brock biology of microorganisms, 14th edition, 2015.
[7]. Hoàng Trọng Phán (chủ biên), Giáo trình di truyền học vi sinh vật và ứng dụng, NXB Đại học Huế, 2008.
[8]. Đề thi IBO và các đề thi học sinh giỏi cấp quốc gia môn Sinh học.
566
MỤCLỤC
Chuyên đề
TÍCH HỢP DI TRUYỀN HOÁ SINH VI SINH VẬT
Đỗ Thuỳ Dung, Trần Thị Loan - Trường THPT Chuyên Lào Cai .......................................
Chuyên đề:
MỘT SỐ VẤN ĐỀ TÍCH HỢP
HÓA SINH – VI SINH – DI TRUYỀN HỌC VI KHUẨN
Nguyễn Thị Cúc - Trường THPT Chuyên Biên Hòa – Hà Nam ........................................ 131
Chuyên đề
TÍCH HỢP DI TRUYỀN HOÁ SINH VI SINH VẬT
Nguyễn Mạnh Quỳnh, Nguyễn Thị Thu Trang - THPT Chuyên Thái Nguyên .................
Chuyên đề
MỘT SỐ NỘI DUNG TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN
Nguyễn Thị Thu - Trường THPT Chuyên Bắc Giang - Bắc Giang ...................................
Chuyên đề
TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN Trường THPT Chuyên Hoàng Văn Thụ - Hòa Bình .........................................................
Chuyên đề
HỆ THỐNG LÝ THUYẾT VÀ BÀI TẬP TÍCH HỢP HOÁ SINH, VI SINH, DI TRUYỀN Trường THPT Chuyên Lê Khiết - Quảng Ngãi.................................................................
233
567