CHUYÊN ĐỀ MÔN SINH HỌC DUYÊN HẢI BẮC BỘ Ths Nguyễn Thanh Tú eBook Collection CÁC CHUYÊN ĐỀ MÔN SINH HỌC ĐẠT GIẢI TẠI HỘI THẢO CÁC TRƯỜNG CHUYÊN KHU VỰC DUYÊN HẢI BẮC BỘ NĂM 2022 Chuyên đề Một số nội dung tích hợp hóa sinh, vi sinh và di truyền WORD VERSION | 2023 EDITION ORDER NOW / CHUYỂN GIAO QUA EMAIL TAILIEUCHUANTHAMKHAO@GMAIL.COM Hỗ trợ trực tuyến Fb www.facebook.com/DayKemQuyNhon Mobi/Zalo 0905779594 Tài liệu chuẩn tham khảo Phát triển kênh bởi Ths Nguyễn Thanh Tú Đơn vị tài trợ / phát hành / chia sẻ học thuật : Nguyen Thanh Tu Group vectorstock com/28062415
Chuyên đề MỘT SỐ NỘI DUNG TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN Nguyễn Thị Thu - Trường THPT Chuyên Bắc Giang - Bắc Giang Chuyên đề đạt giải Ba Phần thứ nhất: MỞ ĐẦU
1. LÍ DO CHỌN ĐỀ TÀI
Sinh học là môn khoa học nghiên cứu các mối quan hệ của các hệ thống sống ở các cấp độ tổ chức khác nhau từ cấp độ phân tử đến cấp độ sinh quyển. Các mối quan hệ đó rất phức tạp nhưng chặt chẽ và tuân theo những quy luật nhất định.
Trong Sinh học có nhiều phân môn như tế bào-hóa sinh, vi sinh, sinh lý thực vật, sinh lý động vật, di truyền, sinh thái, tiến hóa, việc chia theo phân môn giúp học sinh dễ tiếp cận nhưng lại khó hình dung kiến thức đặc biệt là phần di truyền học tích hợp hóa sinh, vi sinh.
Trong chương trình Sinh học mới năm 2018 phần di truyền học bao gồm di truyền phân tử; Di truyền nhiễm sắc thể; di truyền gen ngoài nhân; mối quan hệ kiểu gen - môi trường - kiểu hình; thành tựu chọn, tạo giống bằng các phương pháp lai hữu tính; di truyền quần thể; di truyền học người. Điểm mới chương trình 2018 so với chương trình 2006 là:
+ Cách tiếp cận trong chương trình 2018 có điểm khác với chương trình 2006: Từ khái quát tới cụ thể, bắt đầu từ khái niệm “Hệ gen”.
+ Khái niệm “Tương tác gen” được cập nhật với những nghiên cứu mới của thế giới
+ Bổ sung, cập nhật những tri thức mới, khai thác cơ sở phân tử của các hiện tượng di truyền và biến dị; làm sáng tỏ bản chất của các hiện tượng này.
+ Bổ sung các thông tin mới liên quan đến các biến đổi ngoại di truyền trong sự biểu hiện của tính trạng.
+ Nhấn mạnh khả năng ứng dụng những tiến bộ của Di truyền học trong khoa học và đời sống hiện nay, liên quan đến y học, nông nghiệp, khoa học hình sự…
+ Phần Quy luật di truyền đi sâu vào việc khai thác bản chất của mối quan hệ gen - protein - tính trạng để giải thích các hiện tượng tương tác gen, tính đa hiệu của gen, gen đa alen, mỗi quy luật đều xuất phát từ bối cảnh/ tình huống có vấn đề khoa học.
+ Phần liên kết gen mở rộng ứng dụng trong lập bản đồ di truyền (dựa vào lai hữu tính) và bản đồ vật lí (dựa trên những thành tựu của các kĩ thuật phân tích ADN, giải trình tự ADN…)
DẠYKÈMQUYNHƠN
325
OFFICIAL
Để học tốt và thi tốt kỳ thi tốt nghiệp THPT, thi học sinh giỏi cấp tỉnh với hình thức trắc nghiệm hoặc trắc nghiệm kết hợp tự luận như hiện nay học sinh cần đổi mới phương pháp học tập và làm quen với hình thức thi cử. Nếu trước đây học và thi môn Sinh học, học sinh cần học thuộc và nhớ từng câu, từng chữ hoặc đối với bài toán học sinh phải giải trọn vẹn các bài toán. Nay học sinh lưu ý trước hết đến sự hiểu bài, hiểu thấu đáo các kiến thức cơ bản đã học vận dụng những hiểu biết đó vào việc phân tích, xác định nhận biết các đáp án đúng sai trong các câu trắc nghiệm. Đặc biệt đối với các câu bài tập làm thế nào để có được kết quả nhanh nhất? Đó là câu hỏi lớn đối với tất cả các giáo viên. Trước thực tế đó đòi hỏi mỗi giáo viên cần xây dựng cách dạy riêng của mình.
Trong những năm gần đây, các bài tập di truyền học tích hợp hóa sinh, vi sinh xuất hiện trong đề thi học sinh giỏi quốc gia môn sinh học ngày càng đa dạng và mức độ khó tăng dần lên. Có thể nói nội dung này đóng một vai trò rất quan trọng trong việc phân hóa học sinh ở bài thi ngày thứ nhất, bài thi ngày thứ hai trong Kì thi chọn học sinh giỏi quốc gia và kì thi vòng 2 chọn đội tuyển Olimpic Quốc tế môn Sinh học. Trái ngược với sự thay đổi của đề thi học sinh giỏi thì nguồn tài liệu để giảng dạy phần này cho giáo viên lại khá hạn chế, không có tính hệ thống. Từ những cơ sở trên, chúng tôi đã xây dựng hệ cơ sở lý thuyết, cách giải bài tập di truyền tích hợp hóa sinh, vi sinh sưu tầm một số câu hỏi, bài tập chọn lọc nhằm mục tiêu vô cùng thiết thực là giúp cho học sinh thi tốt nghiệp trung học phổ thông, thi học sinh giỏi cấp tỉnh, thi học sinh giỏi quốc gia học tập và nghiên cứu về phần này tốt hơn và đồng thời cũng giúp các đồng nghiệp giảng dạy thuận lợi hơn. Chính vì vậy chúng tôi chọn đề tài: “Một số nội dung tích hợp hóa sinh, vi sinh và di truyền”
OFFICIAL
2. MỤC ĐÍCH NGHIÊN CỨU Nghiên cứu về lí thuyết phần tích hợp hóa sinh, vi sinh và di truyền. Tổng hợp các câu hỏi và bài tập vận dụng phần tích hợp hóa sinh, vi sinh và di truyền.
Ứng dụng vào việc dạy học sinh chuyên sinh và học sinh khối 12 ôn thi trung học phổ thông quốc gia, thi học sinh giỏi quốc gia môn Sinh học, thi giải toán trên máy tính cầm tay quốc gia nhằm giúp học sinh thu được kết quả cao trong các kì thi này.
3. PHƯƠNG PHÁP NGHIÊN CỨU
Thông qua các bài tập có sẵn hoặc tự đề ra, bài tập trong các đề thi học sinh giỏi quốc gia, bài tập trong các đề thi giải toán trên máy tính khu vực, bài tập trong các đề thi tuyển sinh đại học – cao đẳng, tốt nghiệp THPT, thi THPT quốc gia, sách, tài liệu tham khảo để hướng dẫn học sinh cách làm phần lí thuyết, giải bài tập và phát huy khả năng tích cực, năng động, tư duy sáng tạo trong việc vận dụng kiến thức.
DẠYKÈMQUYNHƠN
326
Phần thứ hai: NỘI DUNG CHUYÊN ĐỀ CHƯƠNG I. CƠ CHẾ DI TRUYỀN Ở MỘT SỐ VIRUT
1.1. Tổng quan về quá trình nhân lên của virus Quá trình nhân lên của virus trong tế bào bao gồm 7 bước:
➢ Hấp phụ (adsorption)
➢ Xâm nhập và cởi vỏ (penetration và uncoating)
➢ Phiên mã (transcription) tạo mRNA của virus
➢ Dịch mã (translation) mRNA để tạo protein virus
➢ Sao chép (replication) genome
➢ Tự lắp ráp (maturation) protein với genome để tạo virion
➢ Giải phóng (release) ra khỏi tế bào
Nếu các bước 3, 4, 5 nhập vào một bước gọi là bước tổng hợp các thành phần (biosynthesis) thì quá trình nhân lên còn 5 bước.
Ở một số virus, quá trình nhân lên không treo trình tự như trên mà xảy ra đồng thời. Ví dụ cùng lúc đều tiến hành phiên mã, dịch mã, sao chép, lắp ráp và chui ra khỏi tế bào.
1.1.1. Sự hấp phụ
Virus gắn vào thụ thể (receptor) đặc hiệu nằm trên màng sinh chất của tế bào chủ, theo nguyên tắc khoá – chìa. Một số virus còn gắn thêm vào các đồng thụ thể (coreceptor). Sự hấp phụ xảy ra tốt nhất ở 37oC.
DẠYKÈMQUYNHƠN
327
OFFICIAL
Virus gắn được vào các thụ thể là nhờ các liên kêt hoá học như liên kết hydro, ion, Vander Waals, nhưng không có liên kết đồng hoá trị.
Vị trí gắn:
+ Virus có các vị trí khác nhau chứa protein gắn vào thụ thể của tế bào.
+ Virus trần có vị trí gắn nằm trên bề mặt capsid, đôi khi nằm sau bên trong (ví dụ virus polio) hoặc đôi khi là đỉnh của khối đa diện (virus lở mồm long móng). Ở virus adeno, vị trí gắn là đầu mút của các sợi mọc ra từ đỉnh capsid khối đa diện. Đối với virus có vỏ ngoài, vị trí gắn là các gai glycoprotein bề mặt.
1.1.2. Xâm nhập và cởi vỏ
Sau khi gắn vào thụ thể, virus phải vượt qua màng sinh chất để xâm nhập vào tế bào theo một trong 2 cơ chế nhập bào hoặc dung hợp.
1.Nhập bào: Cả virus trần và virus có vỏ ngoài đều có thể xâm nhập theo lối nhập bào.
Cơ chế chung bao gồm việc virion gắn vào màng sinh chất nơi mặt trong được bao phủ bởi lớp clathrin (protein dạng sợi tạo thành mạng lưới với các mắt hình đa giác). Virion ấn sâu vào màng tạo hốc rồi khép lại tạo thành túi nội bào hay endosome được lớp clathrin bao phía ngoài. Kênh proton của màng endosome vận chuyển H+ vào trong làm cho pH trong endosome giảm xuống ( pH 4.5 - 5). Endosome sẽ dung hợp với lysosom. Trước khi dung hợp, lớp clathrin bị loại bỏ. pH thấp hoạt hoá enzyme phân giải capsid. Cũng như màng endosome ở một số virus, pH thấp trong endosome dẫn đến việc dung hợp vỏ capsid với màng endosome, làm vỡ vỏ capsid và giải phóng genome.
2. Dung hợp: Khác với virus trần, chỉ có xâm nhập vào tế bào theo lối nhập bào, các virus có vỏ ngoài có thể vào tế bào theo cả 2 cách.
Virus có vỏ ngoài có thể vào tế bào theo cách dung hợp vỏ ngoài virus với màng sinh chất vì chúng có cùng bản chất. Khi 2 màng hoà nhập sẽ đứt ra, nucleocapsid sẽ được chuyển vào tế bào chất. Sự dung hợp này xảy ra với sự tham gia của protein dung hợp (protein F).
Như trên đã nói, virus có vỏ ngoài cũng có thể xâm nhập vào tế bào theo lối nhập bào. Vỏ ngoài virus bám vào thụ thể sau đó ấn lõm tạo endosome. Khi pH trong endosome giảm gai protein F sẽ chồi lên cắm vào màng endosome như chiếc neo, kéo vỏ ngoài virus sát với màng endosome và tiến hành dung hợp.
DẠYKÈMQUYNHƠN
328
OFFICIAL
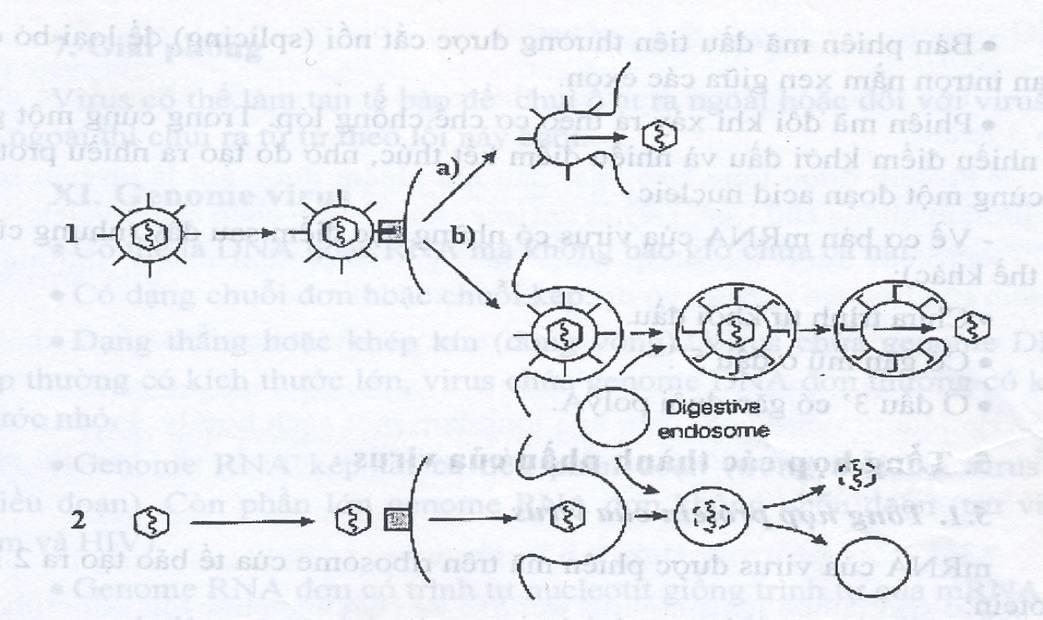
Hình 1.1. Sự nhập bào theo kiểu thực bào, tạo endosome
Bơm proton làm giảm pH trong endosome, hoạt hoá sự dung hợp giữa vỏ ngoài virus với màng endosome (1b), hoặc hoạt hoá enzyme làm tan màng endosome (2). Dung hợp xảy ra trên bề mặt tế bào giữa vỏ ngoài virus với màng sinh chất. Sự vận chuyển genome virus vào nhân
Hầu hết virus ARN ở eukaryota tiến hành sao chép trong tế bào chất, vì chúng có thể mã hoá cho tất cả các enzyme cần cho sao chép genome mà không cần đến các enzyme của tế bào nằm trong nhân. Virus cúm A là ngoại lệ, chúng cần bộ máy cắt nối của tế bào nên genome của chúng phải được đưa vào nhân. Virus retro cũng là virus RNA, nhưng tiến hành sao chép trong nhân. Thoạt đầu chúng nhờ enzyme phiên mã ngược tổng hợp ADN trên khuôn ARN trong tế bào chất, sau đó nằm đợi cho đến khi tế bào bắt đầu phân chia (giai đoạn M), màng nhân tạm thời bị vỡ, ADN cùng với protein liên kết lúc đó mới vào được trong nhân. Do đó virus này chỉ nhân lên được ở giai đoạn tế bào đang phân chia. Hầu hết virus ADN tiến hành sao chép trong nhân, ngoại trừ virus pox và irido có thể sao chép trong tế bào chất. Đối với virus sao chép trong nhân, protein cấu trúc của chúng có trình tự bám được vào vi ống (microtubule). Vi ống là các ống rỗng, đường kính 25 nm, là thành phần của bộ khung tế bào, có chức năng nâng đỡ và neo giữ nhiều thành phần của tế bào và được ví như đường ray để vận chuyển vật chất hoặc bào quan tới một vị trí nhất định trong tế bào. Một đầu vi ống được ký hiệu là (+) và một đầu ký hiệu là (-). Đầu (+) nằm gần màng sinh chất, còn đầu (-) nằm gần nhân. Một protein gọi là proetin vận chuyển (motor protein) tự nó di chuyển và chở các chất cũng như bào quan dọc theo vi ống từ vùng ngoại vi của tế bào tới vùng gần nhân. Các virus herpes, adeno, parvo, retro sử dụng hệ thống này để chở nucleocapsid vào sát màng nhân.
DẠYKÈMQUYNHƠN
329
OFFICIAL
Màng nhân được cấu tạo từ 2 lớp lipid kép, ở đó có lỗ nhân. Hầu hết nucleocapsid có kích thước quá lớn để có thể lọt qua lỗ nhân. Các phân tử muốn qua lỗ phải tạo phức với các protein chuyên biệt của tế bào có chức năng mang gọi là importin để mang vào hoặc exportin để mang ra khỏi nhân. Kênh có thể mở để cho phép các hạt có kích thước đến 25 nm thậm chí lớn hơn đi qua, ví dụ virus nhỏ như parvo, nhưng với virus lớn hơn thì phải cởi vỏ ở lỗ nhân. Cởi vỏ cho genome Cởi toàn bộ hoặc một phần vỏ để giải phóng genome ra khỏi vỏ capsid. Tuỳ loại virus mà quá trình có thể diễn ra tại các vị thí khác nhau:
- Trên bề mặt tế bào, capsid rỗng nằm lại bên ngoài tế bào.
- Cởi vỏ bên trong tế bào chất.
- Cởi vỏ tại lỗ nhân.
- Cởi vỏ bên trong nhân.
Cần nhớ rằng virus xâm nhập thành công vào tế bào không có nghĩa là chúng luôn luôn nhân lên được. Tế bào cũng có cơ chế bảo vệ chống lại virus, ví dụ enzyme từ lysosom có thể làm bất hoạt virus trước và sau cởi vỏ; interferon cảm ứng tạo protein độc ức chế sự nhân lên của virus. Một số virus nhiễm ở dạng tiềm ẩn, chúng không nhân lên, nhưng genome vẫn còn nguyên vẹn và vẫn có tiềm năng nhân lên.
1.1.3. Tổng hợp các thành phần
Sau khi xâm nhận vào tế bào, pha đầu tiên của chu trình nhân lên gọi là pha ẩn. Ở giai đoạn này không phát hiện được bất kỳ virus nào. Đây là đặc điểm chỉ thấy có ở virus. Sự biểu hiện của genome được bắt đầu rất sớm, ngay sau khi virus xâm nhập vào tế bào. Có 4 quá trình xảy ra trước khi hạt virus được lắp ráp:
- Phiên mã tạo mRNA
- Dịch mã sớm tạo protein phi cấu trúc – đó là các enzyme dùng cho sao chép
- Sao chép tạo genome
- Dịch mã muộn tạo protein cấu trúc để cấu tạo capsid và vỏ ngoài
1.1.4. Lắp ráp
Lắp ráp là sự tự kết nối các thành phần virus để tạo ra virion hoàn chỉnh đòi hỏi phải có cấu trúc bền vững, tồn tại được trong môi trường như là một thực thể có khả năng gây nhiễm, tuy nhiên cũng đòi hỏi khi vào trong tế bào cấu trúc này phải không bền vững thì mới có thể giải phóng dễ dàng genome vào tế bào chất. Do vậy virion phải có cơ chế đóng mở kiểu “công-tắc” để có thể biến đổi từ trạng thái bền vững sang
DẠYKÈMQUYNHƠN
OFFICIAL 330
trạng thái không bền vững. Công tắc này liên quan đến việc gắn vào receptor hoặc sự thay đổi pH trong endosome. Khi số lượng genome và protein cấu trúc được tích luỹ đến ngưỡng thì chúng sẽ tiến hành lắp ráp tạo nucleocapsid.
- Virus có cấu trúc dạng xoắn: Đối với virus ARN đơn có cấu trúc dạng xoắn, lúc đầu một số phân tử protein cấu trúc sẽ bám theo chiều xoắn của genome RNA, sau đó các phân tử khác lần lượt bám theo cho đến khi phủ hết RNA.
- Virus có cấu trúc dạng khối đa diện: Trước hết cần phải lắp ráp một cấu trúc rỗng hình cầu gọi là procapsid. Genome virus chui vào procapsid, sau đó cải biến từ cấu trúc hình cầu sang hình khối đa diện; ví dụ các virus adeno, picorna thực hiện cải biến bằng cách cắt bớt 1 hoặc nhiều protein cấu trúc. Genome chui vào trong procapsid qua kênh nằm ở vị trí mà sau này sẽ là đỉnh của khối đa diện. Bất kỳ enzyme nào tham gia vào đóng gói genome cũng nằm ở vị trí này. Ở tế bào thực khuẩn cũng như vậy, trước hết cũng phải tạo một “tiền đầu” (prohead) sau đó genome chui qua 1 cái cổng nằm ở một đỉnh. Vị trí này cũng có chức năng nối với đuôi.
a- Đóng gói genome
Trong tế bào có rất nhiều axit nucleic, của cả virus và tế bào. Vậy làm thế nào để genome của virus lại được lựa chon và lắp ráp chứ không phải của tế bào? Sở dĩ như vậy vì virus có một protein chuyên biệt, nhận diện tín hiệu đóng gói, nằm ở vùng có cấu trúc bậc hai của genome. Hầu hết virus có genome sợi đơn có thể đóng gói hoặc sợi dương hoặc sợi âm, nên tín hiệu đóng gói phải có duy nhất ở sợi cần được đóng gói. Genome được nén trong thể tích nhỏ. Virus ADN kép có kích thước lớn như virus herpes, đóng gói genome chặt đến nỗi tạo áp suất lớn gấp 10 lần so với áp suất trong chai rượu sâm – banh. b- Cơ chế lắp ráp
Trước đây Fraenkel-Conrat đã tách genome ra khỏi capsid của virus đốm thuốc lá, sau đó lại lắp ráp chúng với nhau trong điều kiện pH và sự có mặt của một số ion nhất định, để tạo virus hoàn chỉnh. Với các virus đơn giản, chỉ chứa 1 axit nucleic và một số ít loại protein thì có thể tự lắp ráp một cách đơn giản như trên. Nhưng với các virus phức tạp, như virus herpes và phage có đuôi, thì không thể tái lắp ráp như vậy. Chúng cần phải được lắp ráp trực tiếp trong môi trường của tế bào nhiễm. Khi lắp ráp cần phải có mặt tạm thời 1 protein dùng làm giàn giáo. Các protein cấu trúc theo đó mà lắp vào để tạo capsid. Khi công việc hoàn tất, protein giàn giáo bị loại bỏ khỏi virion hoặc bằng enzyme phân giải hoặc giữ lại để tái sử dụng. c- Sự tạo màng virion
DẠYKÈMQUYNHƠN
331
OFFICIAL
Vỏ ngoài virus có thể được tạo thành theo 1 trong 2 cơ chế: Cải biến màng sinh chất của tế bào rồi nảy chồi ra ngoài hoặc tự tổng hợp màng mới bao quanh nucleocapsid. -Cải biến màng sinh chất
Vỏ ngoài của virus thường có nguồn gốc từ màng sinh chất, được cuốn theo khi virus nảy chồi. Vùng màng mà virus sẽ nảy chồi được đính trước 1 hoặc nhiều loại protein đặc hiệu của virus, thường là glycoprotein, các protein này ngâm trong lớp lipid kép. Protein M của virus (các phân tử protein này gắn với nhau tạo thành màng đệm – M) tập trung nhiều ở vùng màng, có ái lực với nhau và đẩy protein tế bào ra khỏi màng. Đôi khi protein tế bào không bị đẩy ra hết nên chúng có thể tham gia vào thành phần vỏ ngoài. Ví dụ vỏ ngoài của virus HIV-1 có chứa protein MHC-II (phức hợp hoà hợp mô chính) của tế bào. Trước khi nảy chồi, protein M tới gắn vào phần đuôi nằm trong tế bào chất của glycoprotein xuyên màng, sau đó nucleocapsid tiến đến bám vào màng M. Nucleocapsid khi nảy chồi sẽ cuốn theo màng tế bào chất và màng M để tạo vỏ ngoài. Không phải tất cả các virus đều có màng M. Ví dụ ở trường hợp virus sốt vàng thì bề mặt nucleocapsid sẽ gắn trực tiếp vào đuôi glycoprotein trong màng.
-Tổng hợp mới màng virus
Chỉ có một số ít virus tạo màng lipid muộn trong quá trình nhân lên. Màng này có thể tham gia hình thành vỏ ngoài (ví dụ virus pox) hoặc nằm trong nhân tạo 1 lớp phía mặt dưới của capsid (ví dụ virus irido).
Khi nhân lên virus baculo tạo ra 2 loại virion có vỏ ngoài: Loại thứ nhất có chức năng lây nhiễm sang tế bào khác trong vật chủ. Loại này có được vỏ ngoài là do nảy chồi qua màng sinh chất. Loại virion thứ hai có chức năng gây nhiễm vào vật chủ mới, vỏ ngoài của nó bao quanh nucleocapsid nằm trong nhân dẫn đến các virion hợp nhất với nhau trong một bọc gọi là thể bọc (occlusion body). Thể bọc giúp virus tránh tác động của môi trường bên ngoài, nên virus có thể duy trì rất lâu ngoài tế bào sống.
1.1.5. Giải phóng virus khỏi tế bào Đây là giai đoạn cuối cùng của chu trình nhân lên. Nhiều virus được giải phóng khi tế bào bị nổ tung, do thành tế bào bị phân giải, cộng với áp lực lớn trong tế bào. Ví dụ nhiều phage, peptidoglycan bị phân giã dẫn đến làm vỡ tế bào. Số khác lại có khả năng tổng hợp protein ức chế enzyme tham gia vào tổng hợp thành tế bào, làm cho thành tế bào yếu đi nên dễ bị vỡ. Số lượng virus được tạo thành qua mỗi mẻ là rất lớn. Trong tế bào E.coli có kích thước nhỏ bé, phage T4 có kích thước lớn tạo ra được 200 virion. Virus picorna có kích thước rất nhỏ khi nhiễm vào tế bào động vật có kích thước lớn đã tạo ra đến 100.000 virion.
DẠYKÈMQUYNHƠN
332
OFFICIAL
Một số virus không làm tan tế bào mà chỉ chui ra từ từ theo lối nảy chồi, số khác chui ra thông qua việc tạo thành túi hay bọng từ màng lưới nội chất hoặc bộ máy Golgi. Màng bọng dung hợp với màng tế bào chất và đẩy virus ra ngoài. Ở các trường hợp này, tế bào vẫn còn sống thêm một thời gian nữa.
1.2. Các cơ chế di truyền của virus
1.2.1. Sao chép genome của virus
Đây là bước thứ 5 của chu trình nhân lên. Nhìn chung virus ADN và virus ARN sao chép trực tiếp genome của mình thành RNA. Tuy nhiên một số virus ADN khi sao chép cần qua trung gian ARN và một số virus ARN cần qua trung gian DNA.
Các virus ADN tiến hành sao chép trong nhân (trừ virus pox). Các virus có genome nhỏ (virus papilloma) sử dụng ADN polymerase của tế bào, còn virus có genome lớn (ví dụ herpes) thì mã hoá cho enzyme của mình.
Các virus ARN tiến hành sao chép trong tế bào chất (trừ virus cúm và retro), sử dụng enzyme do chúng mã hoá. Trong hầu hết trường hợp, sao chép và phiên mã là một. Enzyme dùng cho sao chép cũng là enzyme dùng cho phiên mã.
➢ Virus ADN kép sao chép trong nhân theo cơ chế bán bảo tồn như ở tế bào, sử dụng enzyme ADN polymerase phụ thuộc ADN của tế bào. Tuy nhiên virus pox là virus ADN kép, sao chép trong tế bào chất sử dụng enzyme ADN polymerase do chúng mã hoá.
➢ Virus ADN đơn (+ hoặc -) tất cả đều sao chép trong nhân, sử dụng ADN polymerase của tế bào và phải qua giai đoạn trung gian tạo sợi ADN kép, gọi là dạng sao chép (RF-replicative form). Tư RF sao chép tạo genome.
➢ Virus ARN kép (ví dụ virus rota) luôn có genome phân đoạn, sao chép trong tế bào chất và sử dụng enzyme ARN polymerase phụ thuộc ARN do chúng mã hoá.
➢ Virus ARN đơn, (+) khi sao chép phải qua bước tạo ARN (-) trung gian làm khuôn để tổng hợp genome ARN (+). Genome ban đầu được dùng làm mRNA để tổng hợp ARN polymerase.
➢ Virus ARN đơn, (-) sao chép trong tế bào chất, sử dụng enzyme ARN polymerase phụ thuộc ARN (RNA-replicase) do chúng mang theo để tổng hợp sợi ARN (+) trung gian làm khuôn tổng hợp genome (-).
Virus ARN (-) phân đoạn (ví dụ virus cúm) sao chép trong nhân, sử dụng ARN polymerase phụ thuộc ARN do chúng mang theo.
❖ Virus ARN (+) phiên mã ngược (ví dụ HIV), trước hết dùng enzyme phiên mã ngược của virus (DNA polymerase phụ thuộc RNA) để tạo ADN kép trong tế bào chất, sau đó vào nhân gắn vào nhiễm sắc thể của tế bào rồi từ đó sao chép tạo genome ARN nhờ enzyme ARN polymerase phụ thuộc ADN của tế bào.
DẠYKÈMQUYNHƠN
333
OFFICIAL
❖ Virus ADN kép phiên mã ngược (ví dụ HBV). muốn sao chép phải qua bước trung gian tạo ARN tiền genome trong nhân, dùng enzyme ARN polymerase phụ thuộc ADN của tế bào. ARN mới sinh ra khỏi nhân làm khuôn để tổng hợp sợi ADN () (sợi L) nhờ enzyme phiên mã ngược của virus. Từ sợi ADN (-) làm khuôn tổng hợp sợi ADN (+) tương bù do enzyme của virus mã hoá.
1.2.1.1. Khởi đầu sao chép Mỗi genome virus có một trình tự đặc biệt, tại đó bắt đầu sao chép axit nucleic của virus. Khi sao chép cần gắn mồi. Đó là phản ứng đầu tiên của một nucleotide với nhóm –OH tại vị trí khởi đầu sự sao chép genome của nhiều virus (ví dụ rota, rhabdo) bắt đầu khi nucleotide đầu tiên của sợi mới bắt cặp với 1 nucleotide trong ARN của virus. Nucleotide đầu tiên hoạt động có hiệu quả như một mồi để sao chép RNA, khi nhóm 3’-OH của nó gắn với nucleotide thứ 2.
Một số virus ADN đơn (ví dụ virus parvo) sử dụng cách tự tạo mồi. Đầu 3’ của ADN có các trình tự tương bù, nên có thể gập lại, bắt cặp với nhau tạo đầu 3’-OH thay cho mồi.
Để khởi đầu sao chép, nhiều genome ADN và một số genome ARN của virus dùng một phân tử ARN hoặc protein làm mồi.
1.2.1.2. ARN mồi và protein mồi
Sự tổng hợp ADN của tế bào được bắt đầu sau khi một vùng xoắn kép được mở xoắn tạo bong bóng nhờ enzyme helicase và sau khi enzyme primase tổng hợp một đoạn ARN ngắn làm mồi. Cần một mồi cho sợi dẫn đầu và nhiều mồi cho tổng hợp các đoạn Okaseki của sợi muộn (sợi sau). Nucleotide đầu tiên của ADN mới gắn vào 3’OH của ARN mồi.
Một số virus ADN dùng mồi ARN để sao chép genome. Một số (ví dụ virus polymase) dùng primase của tế bào để tổng hợp mồi. Số khác (ví dụ virus herpes và phage T7) lại mã hoá cho primase của riêng mình. Virus retro dùng tRNA của tế bào làm mồi khi ở ngoài tế bào chất, nhưng khi cài xen genome của mình vào nhiễm sắc thể của tế bào nên để sao chép, chúng dùng mồi do primase của tế bào tổng hợp. Một số virus động vật sử dụng protein làm mồi, trong đó có virus ADN như virus adeno và virus ARN như picorna. Nhóm 3’-OH của serin hoặc tyrosin trong protein sẽ gắn với nucleotide sợi mới.
Virus hepDNAa (ví dụ HBV) là virus ADN kép, dùng mồi là protein để khởi đầu tổng hợp sợi ADN (-) và mồi ARN để khởi đầu tổng hợp ADN (+). Mồi protein và mồi ARN của virus hepDNAa không bị cắt bỏ sau khi vai trò của chúng đã hoàn tất mà vẫn được dính vào đầu 5’ của genome.
DẠYKÈMQUYNHƠN
OFFICIAL 334
1.2.1.3. Sao chép DNA
Mỗi ADN virus có ít nhất một trình tự chuyên biệt để bắt đầu sao chép gọi là trình tự khởi đầu (Ori). Các protein khởi đầu sao chép ADN bám vào vị trí này bao gồm:
- Helicase bám vào vị trí để tháo xoắn.
- Một số protein bám sợi ADN đơn, giữ cho 2 sợi không bắt cặp lại với nhau.
- Một ADN polymerase.
Về cơ bản quá trình sao chép ADN của virus giống như của tế bào. Ở vi khuẩn, số enzyme tham gia ít hơn so với ở eukaryota. Ví dụ helicase-primase của phage T7 (ở E.coli) chỉ là 1 phân tử, trong khi của virus herpes simplex (ở tế bào động vật) là một phức hợp gồm 3 loại protein.
Sự tổng hợp ADN diễn ra gần chạc sao chép. Một trong 2 sợi là sợi dẫn đầu, sợi còn lại là sợi muộn, được tổng hợp thành các đoạn Okaseki sau đó nối lại với nhau nhờ DNA-ligase. Sợi ADN kép mới tạo thành có chứa một mạch của sợi mẹ. Cách sao chép này gọi là bán bảo tồn. Ngược lại với nó là sao chép bảo tồn xảy ra ở một số virus.
Một số genome ADN là phân tử dạng thẳng trong khi một số khác lại là dạng khép vòng. Một số phân tử dạng thẳng khi sao chép lại được khép vòng, cho nên nhiều genome virus được sao chép như là một phân tử vòng tròn. Từ đây có 2 phương thức sao chép.
- Sao chép theo cơ chế theta hay dạng mắt. Từ vị trí Ori tạo ra 2 chạc ba sao chép. Sao chép cùng lúc theo 2 chiều thuận nghịch kim đồng hồ.
- Sao chép theo cơ chế xích ma (δ). Phân tử ADN kép dạng vòng gồm sợi ngoài (+) và sợi trong (-). Sợi ngoài bị cắt đứt ở liên kết photphodiete tạo ra đầu 3’-OH tự do (gọi là điểm sinh trưởng) Sợi trong xoay được dùng làm khuôn. Các nucleotide nối vào đầu 3’-OH để tạo ra 1 sợi ADN mới. Từ sợi ADN mới này lại được gắn mồi tổng hợp mạch bổ sung tạo ADN kép.
Một số virus lúc mới nhiễm sao chép theo cơ chế theta nhưng ở giai đoạn sau lại theo cơ chế xích ma (ví dụ phage ϕx174).
Một số virus ADN như virus herpes và phage T4, kết quả sao chép tạo ra phân tử ADN rất lớn gọi là phân tử trùng lặp (concateme). Mỗi concateme cấu tạo gồm nhiều bản sao genome nối với nhau. Trước khi lắp ráp vào virion concateme sẽ được phân cắt thành các phân tử có kích thước và trình tự của genome.
1.2.2. Phiên mã của virus
1.2.2.1. Phiên mã genome virus
DẠYKÈMQUYNHƠN
335
OFFICIAL
Thông thường phiên mã là quá trình truyền thông tin di truyền từ ADN sang ARN nhờ enzyme ARN polymerase (chính xác hơn là ARN polymerase phụ thuộc DNA), tuy nhiên genome của virus có thể là RNA, cho nên ở virus phiên mã đơn giản chỉ là quá trình truyền thông tin di truyền từ genome sang mRNA. Có 3 loại ARN polymerase: ARN polymerase I phiên mã rRNA, ARN polymerase II phiên mã các gen mã hoá protein và ARN polymerase III phiên mã rRNA 5S, tRNA và các ARN nhỏ bé khác.
Dựa vào quá trình phiên mã, tạo ra mRNA mà David Baltimore đã phân loại tất cả virus thành 7 nhóm như đã nêu ở phần phân loại: (1) Virus ADN kép, (2) virus ADN đơn (+/-), (3) virus ARN kép, (4) Virus ARN (+), (5) virus ARN (-), (6) virus ARN phiên mã ngược và (7) virus ADN phiên mã ngược.
Axit nucleic sợi đơn (DNA hoặc RNA) có trình tự nucleotide giống với trình tự nucleotide của mRNA quy ước là sợi (+), ngược lại có trình tự tương bù với mRNA được quy ước là sợi (-).
Nhóm I. Virus ADN kép hầu hết tiến hành phiên mã trong nhân, sử dụng ARN polymerase phụ thuộc ADN (tức là ARN polymerase II) của tế bào. Đối với virus ADN kép thì không phân biệt sợi (+) và sợi (-) mà gọi là sợi trái (L) và sợi phải (R), bởi vì genome của hầu hết các virus này có khung đọc mở (ORF) theo cả 2 hướng. Một số virus ADN phiên mã trong tế bào chất (ví dụ virus pox), sử dụng ARN polymerase phụ thuộc ADN do virus mã hoá.
Nhóm II. Virus ADN đơn (+) hoặc (-), tất cả đều phiên mã trong nhân và sử dụng ARN polymerase II của tế bào. Cả virus ADN (+) và ADN (-) khi phiên mã tạo mRNA đều phải qua giai đoạn trung gian ADN kép, dạng sao chép, viết tắt là RF (replicative form).
Nhóm III. Virus ARN kép, tất cả đều phiên mã trong tế bào chất, sử dụng enzyme ARN polymerase phụ thuộc ARN do virus mã hoá để tạo mRNA.
Nhóm IV. Virus ARN (+) có chức năng của mRNA trước khi phiên mã phải dich mã để tạo ARN polymerase phụ thuộc ARN của riêng mình sau đó mới phiên mã tạo mRNA.
Nhóm V. Virus ARN (-) không phân đoạn phiên mã trong tế bào chất, sử dụng ARN polymerase phụ thuộc ARN do chúng mang theo. Virus ARN (-) phân đoạn (ví dụ virus cúm) tuy phiên mã trong nhân nhưng chúng sử dụng ARN polymerase thụ thuộc ARN mang theo, vì tế bào không có enzyme này.
Nhóm VI. Virus retro phiên mã ngược, gồm 2 giai đoạn: Lúc đầu phiên mã genome ARN (+) thành cDNA (-) tiến hành trong tế bào chất nhờ enzyme phiên mã ngược của virus, sau đó ADN vào nhân tích hợp với nhiễm sắc thể của tế bào và tiến hành phiên mã tạo mRNA nhờ enzyme ARN polymerase phụ thuộc ADN của tế bào.
DẠYKÈMQUYNHƠN
336
OFFICIAL
Nhóm VII. Virus ADN kép phiên mã ngược, ví dụ HBV, tiến hành phiên mã trong nhân dùng enzyme ARN polymerase phụ thuộc ADN của tế bào để tạo ARN (+) tiền genome, sau đó ra khỏi nhân, dùng enzyme phiên mã ngược của virus để phiên mã ARN tiền genome thành ADN (-) rồi sau đó tạo genome ADN kép.
Tất cả các virus có genome ARN đều phiên mã trong tế bào chất trừ virus cúm. Tất cả các virus có genome ARN (-) đều phải mang theo ARN polymerase phụ thuộc ARN để phiên mã.
Năm 1958 Fracis Crick đưa ra lý thuyết Trung tâm (Central Dogma), theo đó dòng thông tin di truyền luôn đi từ ADN mRNA Protein.
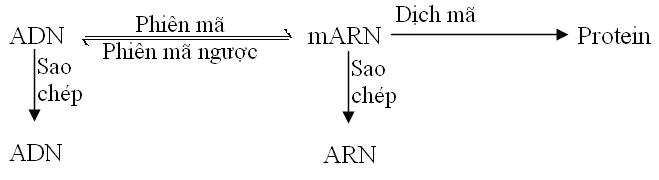
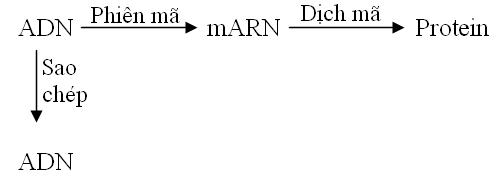
Những kiến thức về virus đã đưa đến sự cải biến lý thuyết trung tâm. Nhiều virus có genome ARN được sao thành ARN và một số có genome ARN được phiên mã ngược thành DNA.
1.2.2.2. Promoter và enhancer
Trong promoter của nhiều tế bào eukaryota và virus có trình tự chung (consensus) sau: TATAA/TAA/TA/G. Không phải promoter nào cũng có trình tự giống hệt nhau mà có thể sai lệch, nên gọi là trình tự chung. Trình tự TATAA còn gọi là hộp TATA hay hộp Pridnow nằm ở phía trước điểm khởi đầu phiên mã, khoảng 2530bp. Hộp TATA chịu trách nhiệm cho phép ARN polymerase gắn vào vùng khởi động phiên mã. Ví dụ: Hộp TATA có trong promotor duy nhất của HIV-1, nhưng trong số 4 promoter của HBV thì chỉ có 1 promoter chứa hộp này. Enhancer (đoạn tăng cường) có trình tự bám vào yếu tố phiên mã. Sự tương tác này làm tăng tốc độ phiên mã bởi ARN polymerase II.
DẠYKÈMQUYNHƠN
OFFICIAL 337
Hình 1.2. Phiên mã từ ADN kép của eukaryota 1.2.2.3. Yếu tố phiên mã
Yếu tố phiên mã là các protein gắn đặc hiệu vào promoter và enhancer để kiểm soát sự biểu hiện gen. Một số virus tạo ra yếu tố phiên mã của riêng mình, ví dụ VP/6 của virus herpes simplex, nó là một thành phần của virion và protein Tax của virus HTLV (virus gây ung thư tế bào T ở người) được tạo ra trong tế bào nhiễm. Một số yếu tố phiên mã của tế bào cũng tham gia hoạt hoá hoặc kiềm chế sự phiên mã của các gen virus. Các yếu tố phiên mã đặc hiệu mô (tissue – specific transcription factors) cần cho một số virus mang tính đặc hiệu mô nghiêm ngặt, nghĩa là chúng chỉ nhân lên trong một số mô nhất định.
Một số yếu tố phiên mã của tế bào có tên là yếu tố phiên mã chung (general transcription factor) tham gia vào sự kiểm soát biển hiện gen của nhiều loại tế bào và virus. Ví dụ yếu tố phiên mã TFIID bám vào hộp TATA.
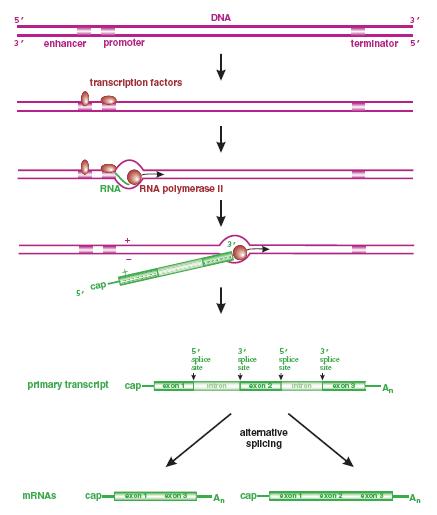
DẠYKÈMQUYNHƠN OFFICIAL 338
Hình 1.3. Sự bám của các yếu tố phiên mã và ARN pol II vào hộp TATA TFIID là một phức hợp gồm 13 polypeptide, một trong số đó là protein bám hộp TATA. Sau khi TFIID bám vào hộp TATA là đến lượt các yếu tố phiên mã chung khác (TFIIA, TFIIB, TFIIE, TFÌI, TFIIH) và ARN pol II bám vào.
Trong số các yếu tố phiên mã của tế bào bám vào enhancer, có:
- AP1 và AP2 (activator protein 1 và 2) là các protein hoạt hoá.
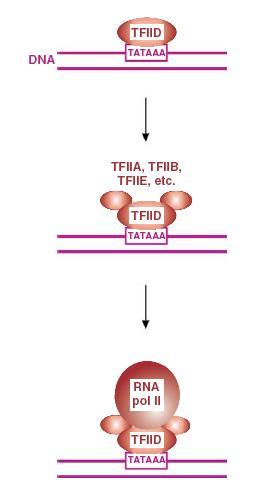
- SP1 (stimulator protein) là protein kích thích.
- NF-kB (nuclear factor kB) là yếu tố nhân kB.
Hầu hết các yếu tố phiên mã này tham gia vào quá trình phiên mã của HIV-1. Cũng như hoạt hoá sự biểu hiện gen, các yếu tố phiên mã cũng tham gia vào sự kiềm chế biểu hiện gen.
Mọi cơ thể đều có khả năng tự điều hoà các gen của mình. Ví dụ côn trùng có các gen khác nhau và sự biểu hiện các gen này tuỳ thuộc việc chúng đang ở giai đoạn nào: ấu trùng, sâu non, sâu trưởng thành hay bướm. Ở virus cũng vậy, chúng có các gen khác nhau, biểu hiện ở các thời điểm khác nhau. Gen sớm mã hoá cho các protein sớm, đó là các protein không cấu trúc, thường là các enzyme và gen muộn mã hoá cho các protein muộn, đó là các protein cấu trúc, tham gia vào tạo capsid hoặc vỏ ngoài.
1.2.2.4. Biến đổi ARN sau phiên mã
1.2.2.4.1. Gắn mũ vào ARN
DẠYKÈMQUYNHƠN
339
OFFICIAL
Ngay sau khi tổng hợp ARN và trong lúc quá trình phiên mã còn đang tiếp tục thì hầu hết các bản phiên mã (RNA) đã được gắn mũ ở đầu 5’. Mũ là guanosine triphosphate nối với một nucleotide ở đầu 5’ nhờ liên kết 5’-5’, thay vì liên kết 5’-3’ như bình thường.
Hầu hết mRNA ở tế bào eukaryota và ở virus đều có mũ ở đầu 5’.
Mũ có chức năng:
- Giúp mRNA vận chuyển từ nhân ra tế bào chất. - Bảo vệ mRNA khỏi sự thuỷ phân của enzyme exonuclease.
- Cần cho khởi đầu cho dịch mã.
Enzyme thực hiện gắn mũ của tế bào là guanylyl transferase (thêm guauosin 5’triphosphate) và metyl transferase (thêm nhóm metyl). Các enzyme này nằm trong nhân, nên hầu hết các virus phiên mã trong nhân đều sử dụng. Tuy nhiên các virus phiên mã trong tế bào chất sẽ mã hoá cho enzyme gắn mũ và enzyme metyl hoá của riêng mình.
Virus có genome ARN (-) phân đoạn (cúm A) cần có một cơ chế riêng để đoạt lấy mũ từ mRNA của tế bào. Phức hợp protein của virus giúp ARN polymerase của nó gắn vào đầu 5’ của mRNA đã gắn mũ của tế bào, sau đó nhờ hoạt tính endonuclease của phức hợp sẽ cắt một đoạn khoảng 10-20 nucleotide (chứa cả mũ) ra khỏi đầu 5’ của mRNA. Đoạn này sẽ được dùng làm mồi để khởi đầu phiên mã cho virus.
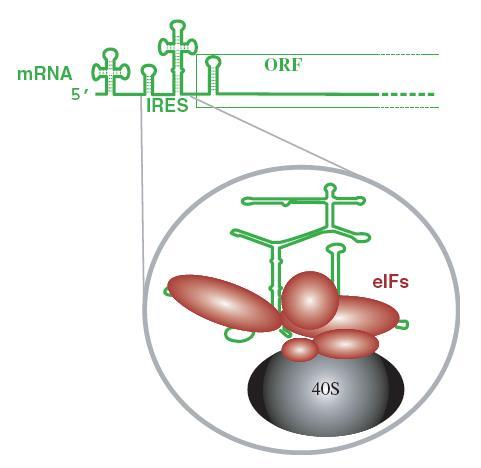
Không phải tất cả mRNA virus đều có gắn mũ, mRNA của virus picorna (bại liệt) không có mũ. Đây là virus phiên mã trong tế bào chất, nên không phải vận chuyển ra khỏi nhân. Chúng có thể tiến hành dịch mã theo cơ chế không phụ thuộc mũ, mà nhờ đoạn IRES.
1.2.2.4.2. Gắn đuôi poly (A)
Các bản phiên mã đầu tiên của eukaryota và virus được ngắn thêm một loạt gốc adenosin ở đầu 3’ để tạo đuôi poly (A). Sự polyadenin hoá ở đầu 3’ làm tăng tính bền của mRNA tránh sự phân giải của exonuclease và có vai trò nhất định trong khởi đầu dịch mã.
Tuy nhiên mRNA của một số virus không gắn đuôi (ví dụ virus reo) nên chức năng kể trên sẽ được thực hiện theo cách khác.
Trước vị trí gắn đuôi có một đoạn tín hiệu gắn đuôi dài 20-30bp. Đoạn này ở SV40 là AATAAA, ở HBV là TATAAA (giống như hộp TATA). Khi phiên mã ARN polymerase tiếp tục chạy dọc khuôn, vượt qua tín hiệu gắn đuôi và vị trí gắn đuôi. Sợi ARN mới tổng hợp được cắt tại vị trí gắn đuôi và các gốc adenosin được lần lượt thêm vào nhờ phức hợp protein, trong đó có enzyme gắn đuôi (poly (A) -polymerase).
1.2.2.4.3. Cắt nối
DẠYKÈMQUYNHƠN
340
OFFICIAL
Một số bản phiên mã có chức năng của mRNA nhưng hầu hết còn cần phải chế biến, cắt bỏ intron để tạo mRNA. Hơn nữa từ mRNA, bằng phương thức cắt nối (ghép đoạn này với đoạn kia), có thể tạo ra nhiều loại mRNA khác nhau. Ví dụ bằng phương pháp cắt nối, HIV-1 có thể tạo ra 30 loại mRNA khác nhau.
1.2.3. Dịch mã của virus
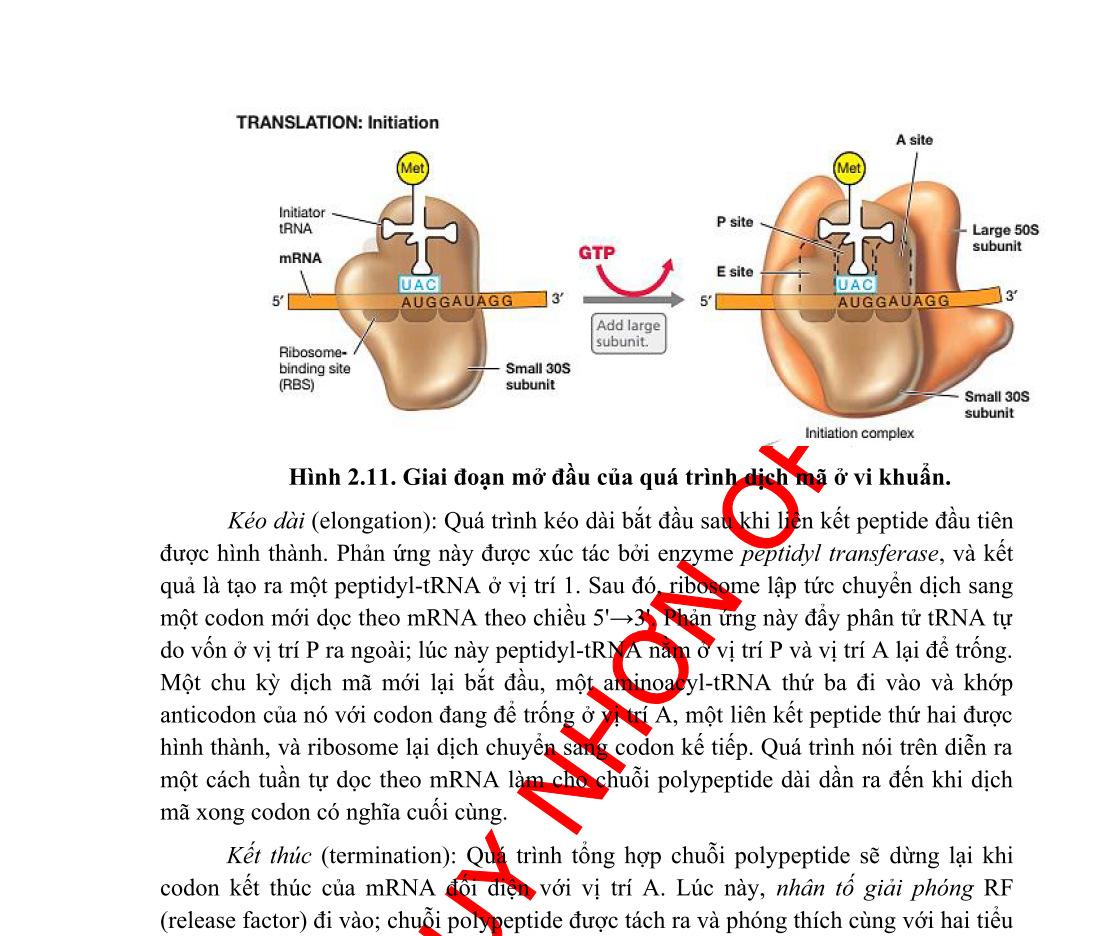
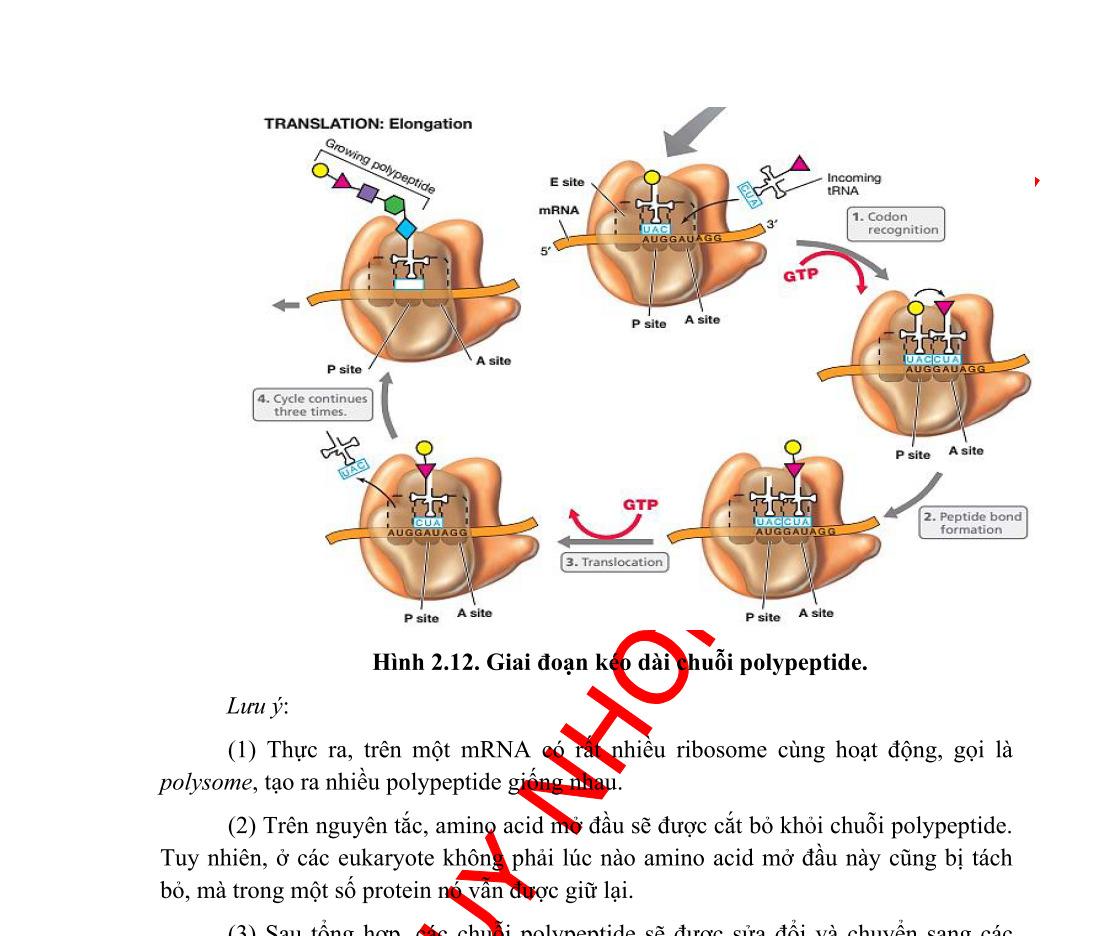
Tham gia vào dịch mã gồm mRNA, tRNA, axit amin và riboxom. Quá trình gồm 3 giai đoạn: Khởi đầu, kéo dài và kết thúc.
Ở eukaryota, mRNA là đơn gen (monocistron) có nghĩa là chỉ có một khung đọc, mã hoá cho 1 protein, còn ở prokaryota, mRNA thường là đa gen (polycistron) có nhiều khung đọc và mã hoá cho nhiều protein.
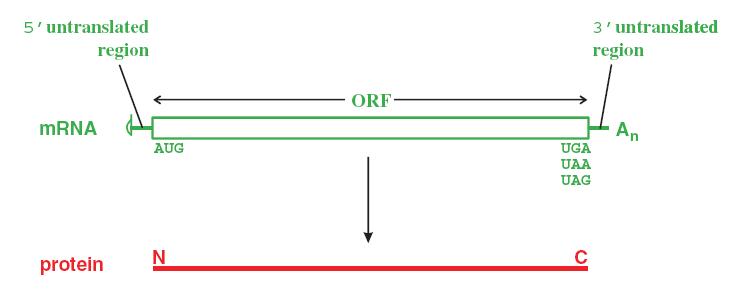
Có các vùng không mã hoá, còn gọi là vùng không dịch mã, nằm ở phía đầu 5’ (5’-UTR) và đầu 3’ (3’-UTR).
Một khung đọc lớn có thể mã hoá cho một polyprotein lớn sau đó được enzyme phân cắt thành các protein nhỏ với chức năng khác nhau.
Hình 1.4. Dịch mã từ mRNA-monocistron
Một khung đọc mở bắt đầu từ codon khởi đầu AUG ở đầu 5’ của mRNA và kết thúc tại codon kết thúc (UGA, UAA, UAG). Dịch mã theo hướng 5’ 3’. Đầu N của protein được tổng hợp trước.
1.2.3.1. Khởi đầu dịch mã
Như đã nói trên, hầu hết mRNA của eukaryota và virus được gắn mũ và đuôi. Các cấu trúc này có vai trò quan trọng trong dịch mã.
- Mũ là nơi bám của các yếu tố khởi đầu dịch mã, eIF (e = eukaryota), tRNA gắn với axit amin (Met-tRNAi Met) và tiểu phần riboxom 40S (hình 6.)
- Một protein gọi là protein bám poly (A) bám vào đuôi poly (A). Các protein bám vào đầu sợi ARN có khả năng tương tác và người ta cho rằng sự tương tác này làm cho mRNA truyền tin dẫn đến sự kích thích dịch mã.
Các mRNA không có mũ hoặc đuôi có thể truyền tin theo cách khác.
DẠYKÈMQUYNHƠN
341
OFFICIAL
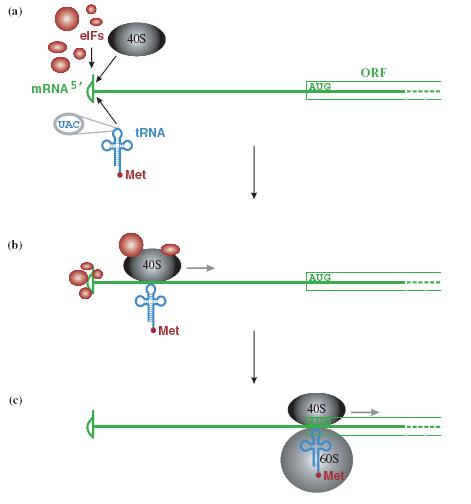
Tiểu phần riboxom 40S di chuyển dọc theo ARN theo hướng 5’ 3’ và quét cho đến khi gặp codon khởi đầu AUG ở đầu 5’. Tuy nhiên, một số virus có codon khởi đầu khác. Ví dụ virus Sendai dùng codon khởi đầu là ACG cho một số gen.
Hình 1.5. Khởi đầu dịch mã trên m ARN có gắn mũ a – Các yếu tố khởi đầu eIF, tiểu phần riboxom 40S và t RNA-methionine bám vào đầu 5’ của mRNA, b - Phức hợp quét từ đầu 5’ theo hướng 3’ của mRNA, c – khi tới codon khởi đầu AUG sẽ được anticodon UAC trên tRNA nhận diện, tiểu phần 60S bám vào và các yếu tố eIF sẽ rời ra.
Một số mRNA không có mũ thì sự dịch mã xảy ra theo cơ chế khác. Các eIF thay vì gắn vào mũ, chúng gắn vào vị trí IRES (internal ribosome entry site) gọi là bến đỗ riboxom, đó là đoạn mRNA có cấu trúc bậc 2 ở mức độ cao nằm trước khung đọc, cũng là vị trí gắn của tiểu đơn vị riboxom 40S. Đoạn IRES có ở một số virus RNA, như virus viêm gan C, picorna, đồng thời cũng thấy có ở mRNA của tế bào và mRNA của một số virus khác (ví dụ virus herpes simplex, Sarcoma Rous).
DẠYKÈMQUYNHƠN
342
OFFICIAL
Hình 1.6. Khởi đầu dịch mã trên mRNA không gắn mũ.
Tiểu phần roboxom 40S, các eIF bám vào vị trí IRES (bến đỗ riboxom) nằm phía trước khung đọc.
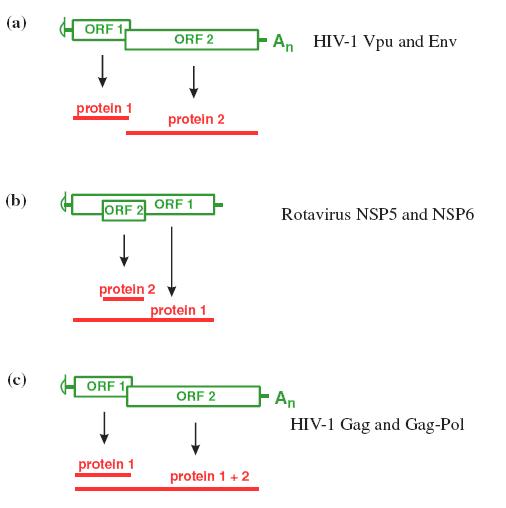
1.2.3.2. Dịch mã từ m ARN hai khung đọc Hầu hết mRNA của tế bào và của nhiều virus chỉ có 1 khung đọc mở (ORF), nhưng một số mRNA của virus có 2 hoặc nhiều khung đọc, trong số các mRNA bicistron hoặc polycistron này có một số có chức năng là monocistron, nhưng một số mRNA có cấu trúc là bicistron thì cũng có chức năng là bicistron. Sự khác nhau về tỷ lệ dịch mã của hai khung đọc quy định cơ chế biểu hiện 2 gen ở mức độ khác nhau. Ở nhiều mRNA bicistron có khung đọc chồng lớp. Ở trường hợp khác, khung đọc này nằm gọn trong khung đọc kia.
- Một cơ chế dùng để đọc khung thứ 2 gồm sự quét bỏ sót (leaky): Tiểu đơn vị riboxom 40S có thể quét bỏ qua codon khởi đầu của khung đọc thứ nhất và bắt đầu dịch mã tại điểm khởi đầu của khung đọc thứ 2. Các khung đọc cho hai protein là khác nhau nên hai protein do chúng mã hoá là khác nhau.
DẠYKÈMQUYNHƠN
343
OFFICIAL
Hình 1.7. Sự dịch mã của mRNA bicistron.
– Riboxom có thể dịch mã tại điểm khởi đầu của ORF-1 hoặc có thể quét qua codon khởi đầu của ORF-2.
– Hai codon khởi đầu là của 2 khung đọc khác nhau và protein được tổng hợp là khác nhau.c – Khung đọc 2 được dịch mã là nhờ sự dịch khung riboxom, tạo ra một dạng kéo dài của protein 1.
- Một cơ chế khác để đọc khung đọc thứ hai của mRNA là sự dịch khung (frameshifting) của riboxom: Riboxom nhảy sang khung đọc khác nằm trước điểm kết thúc của khung đọc thứ 1. Do đó nó không bị codon kết thúc của khung đọc thứ nhất nhận ra, mà tiếp tục chạy dọc mRNA để đọc khung đọc thứ nhất kéo dài (hình 19.7c). Sự dịch khung xảy ra khi riboxom di chuyển dọc theo ARN bắt gặp tín hiệu dịch khung (1 trình tự chuyên biệt) có cấu trúc bậc 2, và thường là một pseudoknot (nút giả)
1.2.3.3. Cải biến trong và sau dịch mã
Trong hoặc sau quá trình dịch mã, protein có thể phải qua một hoặc nhiều lần cải biến, bao gồm glycosyl hoá, acyl hoá hoặc phosphoryl hoá.
-Glycosyl hoá
DẠYKÈMQUYNHƠN OFFICIAL 344
Glycosyl hoá là sự gắn thêm nhóm oligosaccarit vào chuỗi polypeptide. Khi một oligosaccarit được gắn vào nhóm –OH của serin hoặc treonin thì gọi là O-glycosyl hoá, còn nếu gắn vào nhóm -NH2 của asparagin thì gọi là N-glycosyl hoá. Protein được tổng hợp trong mạng lưới nội chất hạt, nơi bắt đầu N-glycosyl hoá, sau đó chúng được chuyển tới bộ máy Golgi, nơi hoàn thiện N-glycosuyl hoá nhờ enzyme αmannosidase I và II, và galactosyl transferase. O-glycosyl hoá được thực hiện trong bộ máy Golgi.
Có những virus chỉ cần một loại glycosyl hoá, ví dụ HIV-1 tiến hành Nglycosyl hoá gp120 nhưng cũng có những virus cần cả 2 loại N- và O-glycosyl hoá, ví dụ gC và gD của herpes simplex. Các glycoprotein này được gắn vào vị trí chuyên biệt trên màng sinh chất của tế bào và sẽ là các gai bề mặt sau này của vỏ ngoài. Virus rota không có vỏ ngoài nhưng cũng có glycoprotein. Protein bề mặt VP7 là glycoprotein. Một số glycoprotein của virus là protein không cấu trúc, ví dụ protein NSP4 của virus rota.
-Acyl hoá
Acyl hoá là sự gắn thêm nhóm acyl (R-CO-) vào phân tử protein. Nhóm acyl thường được gắn là myristyl, trong đó R là CH3-(CH2)12- nhóm myristyl được gắn vào glycin tại đầu N của protein. Hầu như virus không có enzyme N-myristyl transferase để gắn nhóm myristyl, chúng phải sử dụng của tế bào.
Protein myristyl hoá gắn vào màng sinh chất, ví dụ protein Gag của hầu hết virus retro. Nếu không được myristyl hoá thì chúng không gắn được vào màng sinh chất và sự lắp ráp virus sẽ không xảy ra.
- Phosphoryl hoá
Phosphoryl hoá là sự chuyển nhóm phosphate từ một nucleotide, thường là ATP, cho oxy ở nhóm –OH của serin, treonin hoặc tyrosin. Sự chuyển nhóm phosphate nhờ enzyme kinase của tế bào hoặc của virus.
Sự phosphoryl hoá có thể làm thay đổi cấu hình, hoạt tính, vị trí và độ bền của protein. Nhiều quá trình của virus đòi hỏi protein phải được phosphoryl hoá.
- Sự vận chuyển các phân tử trong tế bào eukaryota
Các phân tử của virus được vận chuyển tới các vị trí nhất định trong tế bào. Genome của virus được đưa vào nhân nhờ trượt theo vi ống và qua lỗ nhân, mRNA virus được vận chuyển từ nhân ra tế bào chất và các protein virus sau khi được tổng hợp có thể được vận chuyển vào các vị trí khác nhau, kể cả vào nhân (Các virus lắp ráp trong nhân).
DẠYKÈMQUYNHƠN
345
OFFICIAL
Nhiều protein có một trình tự axit amin giống như mã số vùng của bưu điện, nó quy định điểm đến của protein. Protein đến gắn vào màng có một trình tự tín hiệu – đó là dãy các gốc axit amin kỵ nước. Sự tổng hợp protein bắt đầu tại các riboxom tự do, nhưng khi trình tự tín hiệu được tổng hợp thì nó sẽ hướng dẫn phức hợp polypeptideriboxom đi vào mạng lưới nội chất và tiếp tục tổng hợp protein tại đó. Vùng mạng lưới nội chất có chứa riboxom vì thế gọi là mạng lưới nội chất sần.
Mỗi một loại protein gắn vào màng có một hoặc nhiều trình tự gắn vào màng, gọi là trình tự neo màng (membrane anchor sequences) giàu gốc axit amin kỵ nước. Một số protein loại này có trình tự tín hiệu hoạt động như cái mỏ neo cắm vào màng. Các protein gắn vào màng khác, ví dụ protein vỏ ngoài của HIV-1 sẽ được chuyển qua màng cho đến khi chạm tới trình tự neo và sau đó trình tự tín hiệu sẽ bị enzyme của tế bào loại bỏ.
Nhiều protein được tổng hợp trong mạng lưới nội chất hạt sẽ được tạo bọng để tới bộ máy Golgi. Hầu hết protein gắn màng được glycosyl hoá trong các khoang màng này, sau đó được chuyển tới các màng tế bào chất hoặc màng nhân và có thể nẩy chồi qua các màng này. Các tế bào biểu mô có mặt đỉnh (ở phía ngoài) và mặt đáy (ở phía trong), được cấu tạo từ lipid và protein. Khi nhiễm virus có vỏ ngoài vào tế bào biểu mô chúng sẽ nảy chồi qua màng sinh chất một cách giới hạn hoặc qua mặt đỉnh hoặc qua mặt đáy. Ví dụ nếu là VSV (virus chốc mép), chúng sẽ nẩy chồi từ mặt đáy, còn virus cúm A sẽ nảy chồi từ mặt đỉnh. Điều này giải thích vì sao virus cúm nhiễm vào động vật có vú đều khu trú ở đường hô hấp.
Nếu virus nhân lên ở trong nhân thì hầu hết (nhưng không phải tất cả) các protein virus phải được đưa vào nhân. Cơ chế vận chuyển liên quan đến sự nhận diện của tín hiệu định vị nhân (NLS-nuclear localization signal) nằm trên protein của tế bào, gọi là importin, sau đó gắn vào các sợi (fibril) mọc ra từ phức hợp lỗ nhân để vận chuyển qua lỗ nhân.
1.3. Câu hỏi bài tập về cơ chế di truyền ở virut
Câu 1(trích đề thi học sinh giỏi quốc gia năm 2015): Hãy phân biệt quá trình phiên mã và quá trình sao chép trong tế bào chủ của virut chứa hệ gen ARN+ với virut chứa hệ gen AND về: nơi phiên mã, enzim dùng trong phiên mã, nơi sao chép, enzim dùng trong sao chép. Quá trình phiên mã có trùng với quá trình sao chép không?
Virut ADN Nơi phiên mã Trong tế bào chất Trong nhân tế bào
Enzim dùng cho phiên
Virut ARN (+)
ARN polimeraza phụ thuộc ARN của virut
ARN polimeraza phụ thuộc ADN của tế bào
DẠYKÈMQUYNHƠN
346
OFFICIAL
mã
Nơi sao chép Trong tế bào chất Trong nhân tế bào
Enzim dùng cho sao chép
ARN polimeraza phụ thuộc ARN của tế bào
ADN polimeraza phụ thuộc ADN của virut
Ở virut chứa hệ gen ARN(+), quá trình phiên mã trùng với quá trình sao chép còn ở virut chứa hệ gen ADN quá trình phiên mã không trùng với quá trình sao chép.
Câu 2: Hệ gen của các retrovirut (như HIV) và virut ADN kép (như HBV) đều có enzim phiên mã ngược nhưng chúng lại có cơ chế tái bản vật chất di truyền khác nhau. Em hãy chỉ ra những điểm khác biệt đó.
Trả lời
OFFICIAL
Virut retro phiên mã ngược(HIV) Virut ADN kép phiên mã ngược (HBV)
B1: Sử dụng enzim phiên mã ngược của virut để tổng hợp ADN kép trong tế bào chất
B2: ADN kép tích hợp vào NST trong nhân rồi từ đó phiên mã tạo ARN nhờ nzim của tế bào chủ
B1: Sử dụng enzim phiên mã của tế bào để tổng hợp tiền genom virut là ARN (+) trong nhân tế bào
B2: Ra tế bào chất, dùng enzim phiên mã ngược của virut để phiên mã ARN (+) thành ADN (-) rồi sau đó tạo ADN kép.
Câu 3: Một loại virut không có vỏ ngoài, có vật chất di truyền là ARN đơn, sợi (-). Khi xâm nhập vào tế bào, virut này sẽ tái bản hệ gen và tổng hợp prôtein cấu trúc của nó như thế nào?
Trả lời
Gồm 3 bước:
- Bước 1: Tống hợp sợi ARN(+) bổ sung với sợi ARN(-) của virut. - Bước 2: Dùng sợi ARN(+) làm khuôn để tái bản hệ gen ARN(-) của virut. - Bước 3: Dùng sợi ARN(+) làm mARN để tổng hợp các protein cấu trúc của virut
Câu 4: Mặc dù virut HIV và virut HBV có vật chất di truyền là khác nhau nhưng sau khi xâm nhập vào tế bào người, chúng đều tổng hợp ADN để có thể cài xen vào hệ gen của người. Em hãy nêu những điểm giống nhau trong quá trình tổng hợp ADN của chúng.
Trả lời
DẠYKÈMQUYNHƠN
347
- Diễn ra trong tế bào chất.
- Sử dụng enzim phiên mã ngược AND polymeraza phụ thuộc ARN của virut.
- Sử dụng các nucleootit, ATP, các enzim khác của tế bào chủ
- Sử dụng ARN của virut để tổng hợp AND mạch kép.
Câu 5: Virus chứa hệ gen ARN (+) và virus chứa hệ gen ADN (ký sinh ở tế bào nhân thực) có sự khác biệt như thế nào về: vị trí, enzyme dùng cho hai quá trình phiên mã và tổng hợp vật chất di truyền?
Virus ARN (+) Virus ADN (mạch kép) Nơi phiên mã Trong tế bào chất
Enzyme dùng cho phiên mã
ARN polimeraza phụ thuộc ARN của virus
Trong nhân tế bào
ARN polimeraza phụ thuộc ADN của tế bào
OFFICIAL
Nơi tổng hợp VCDT Trong tế bào chất Trong nhân tế bào
Enzyme dùng cho tổng hợp VCDT
ARN polimeraza phụ thuộc ARN của tế bào
ADN polimeraza phụ thuộc ADN của virus
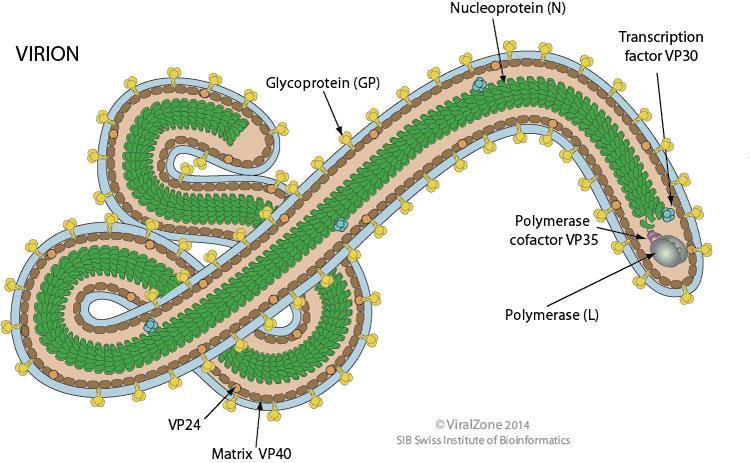
Câu 6: Cho sơ đồ cấu trúc hai loại virut: virut zika và virut ebola như hình dưới đây:
Sơ đồ cấu trúc virut zika
Sơ đồ cấu trúc virut ebola Hãy trình bày những điểm khác nhau về hình thái, cấu trúc và quá trình nhân lên của hai loại virut này?
DẠYKÈMQUYNHƠN
348
Đặc điểm phân biệt
Virut zika
Virut ebola Hình thái
Cấu trúc khối: các capsome xếp thành 20 mặt tam giác đều
Cấu trúc xoắn: Capsome xoắn theo chiều xoắn của axit nucleic
Cấu trúc
Chứa ARN sợi đơn, được dùng như mARN (hay còn gọi là ARN +)
Quá trình nhân lên
Sao chép trong tế bào chất: ARN+ đóng vai trò mARN tham gia dịch mã tổng hợp ARN polimerase, ARN polimerase xúc tác tổng hợp ARN-, sau đó từ ARN- tổng hợp mARN để mã hóa vò capsit và cũng từ ARNlàm khuôn để tổng hợp ARN+ (lõi của virut)
Chứa ARN sợi đơn, được dùng để tổng hợp mARN (vì trong cấu trúc có enzim polymerase)
Sao chép trong tế bào chất: Sử dụng ARN polimerase do chúng mang theo để tổng hợp ARN+, từ ARN + vừa làm khuôn tổng hợp vỏ capsit vừa làm khuôn tổng hợp ARN- (lõi của virut)
Câu 7: Hệ gen của các retrovirut (như HIV) và virut ADN kép có enzim phiên mã ngược (như HBV) đều có enzim phiên mã ngược nhưng chúng lại có cơ chế tái bản vật chất di truyền khác nhau. Em hãy chỉ ra những điểm khác biệt đó.
Virut retro phiên mã ngược (HIV)
Hệ gen: 2 sợi ARN (+) giống nhau
B1: Sử dụng enzim phiên mã ngược của virut (ADN-pol phụ thuộc virut) để tổng hợp ADN kép (cADN) trong tế bào chất
B2: ADN kép tích hợp vào NST trong nhân rồi từ đó phiên mã tạo ARN nhờ enzim của tế bào chủ (ARN pol phụ thuộc ADN của tế bào).
Virut ADN kép phiên mã ngược (HBV)
Hệ gen: ADN kép
B1: Sử dụng enzim phiên mã của tế bào (ARN pol phụ thuộc ADN của tế bào) để tổng hợp tiền genom virut là ARN (+) trong nhân tế bào
B2: Ra tế bào chất, dùng enzim phiên mã ngược của virut (ARN-pol phụ thuộc virut) để phiên mã ARN (+) thành ADN (-) rồi sau đó tạo ADN kép.
Câu 8: HPV và HBV là hai loại virut đều có vật chất di truyền là phân tử ADN kép, tuy nhiên HBV có enzim phiên mã ngược. Quá trình tổng hợp vật chất di truyền của hai virut này trong tế bào chủ có gì khác nhau?
Trả lời:
DẠYKÈMQUYNHƠN
349 Trả lờ
OFFICIAL
i
- HPV: ADN kép nhân đôi trong nhân theo cơ chế bán bảo tồn, sử dụng ADN polimeraza của tế bào
- HBV: AND nhân đôi theo 2 giai đoạn:
+ ADN → ARN diễn ra trong nhân, sử dụng ARN polimeraza của tế bào
+ ARN → ADN diễn ra trong tế bào chất, sử dụng enzim phiên mã ngược của virut
OFFICIAL
DẠYKÈMQUYNHƠN
350
2.1. Khái quát cơ chế điều hòa hoạt động của của Operon đồng hóa, dị hóa ở vi khuẩn
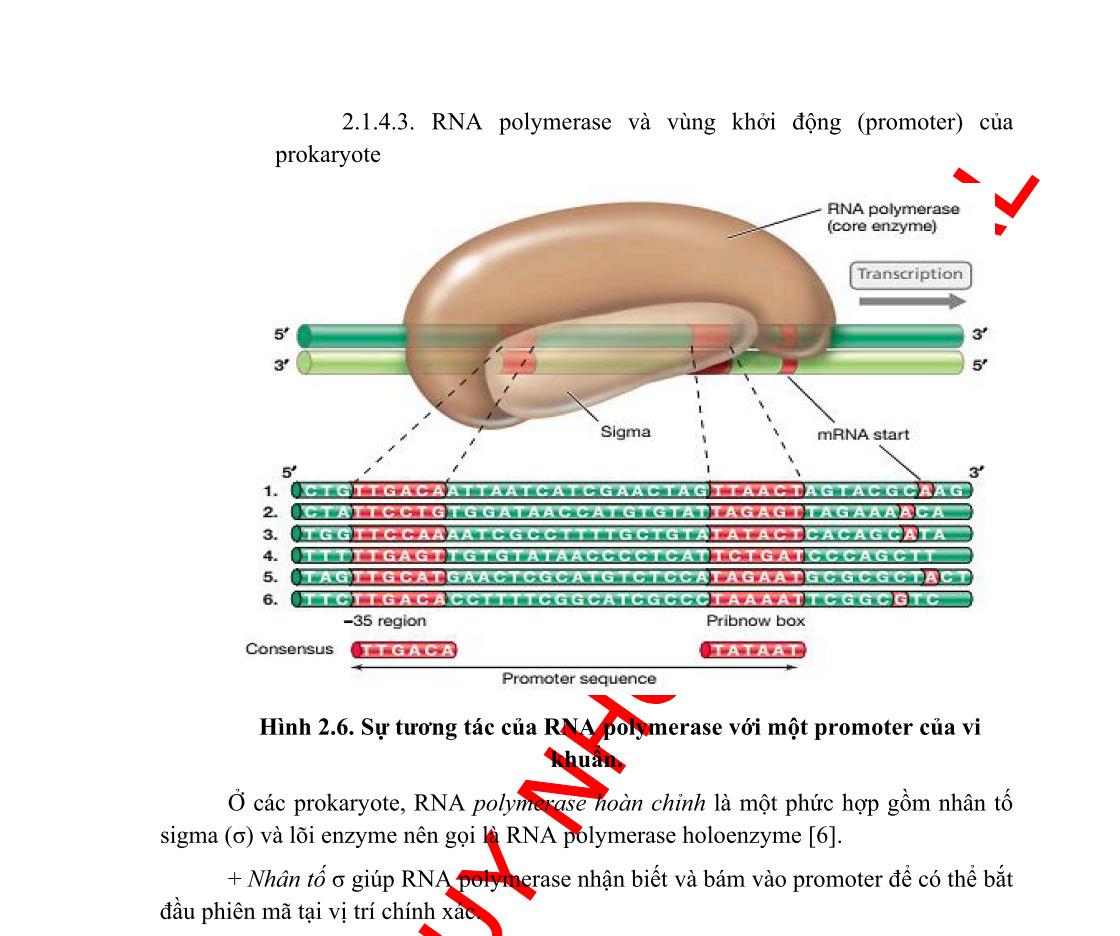
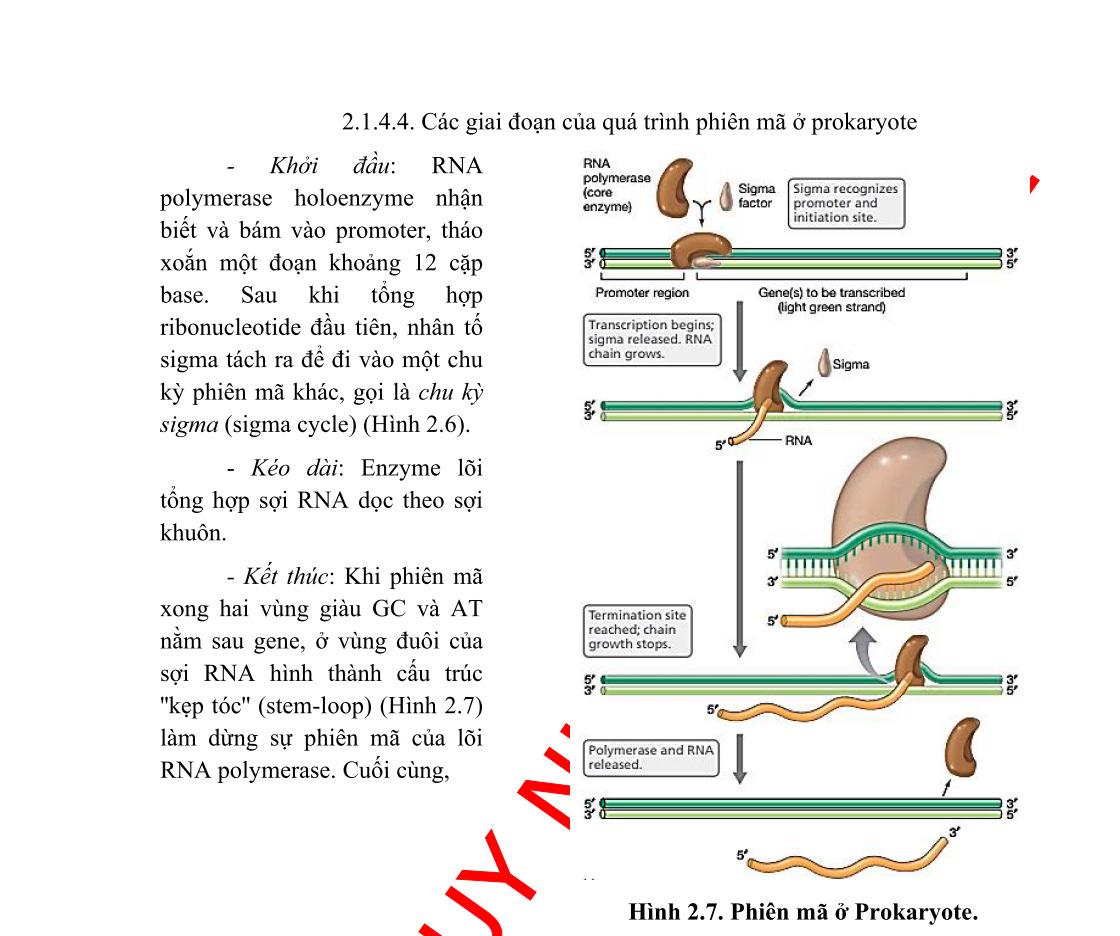
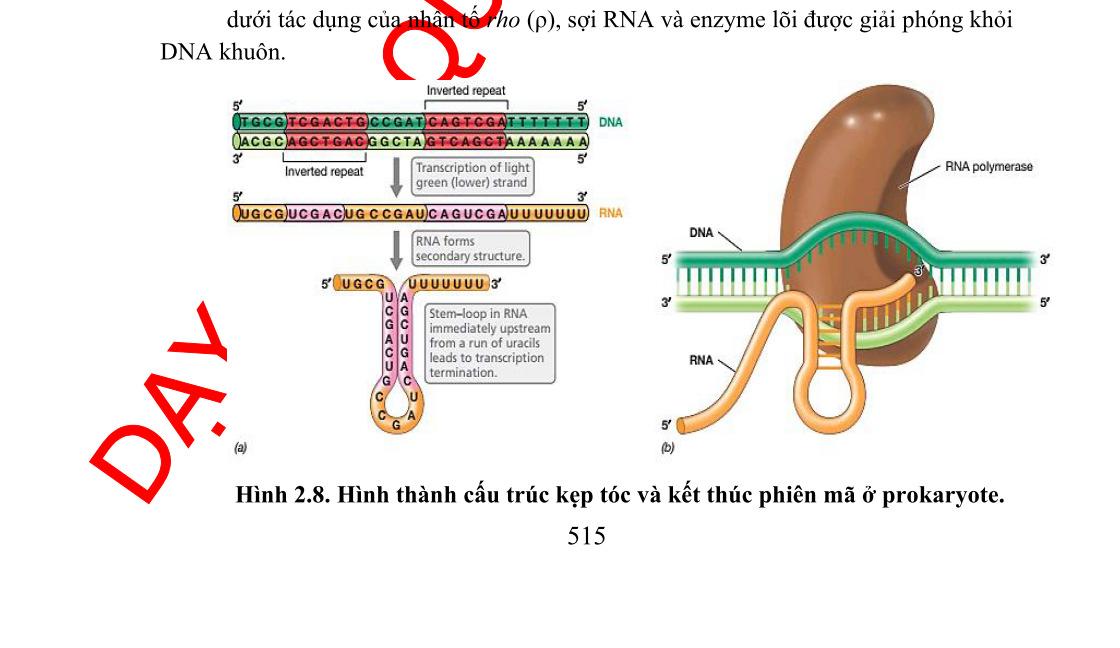
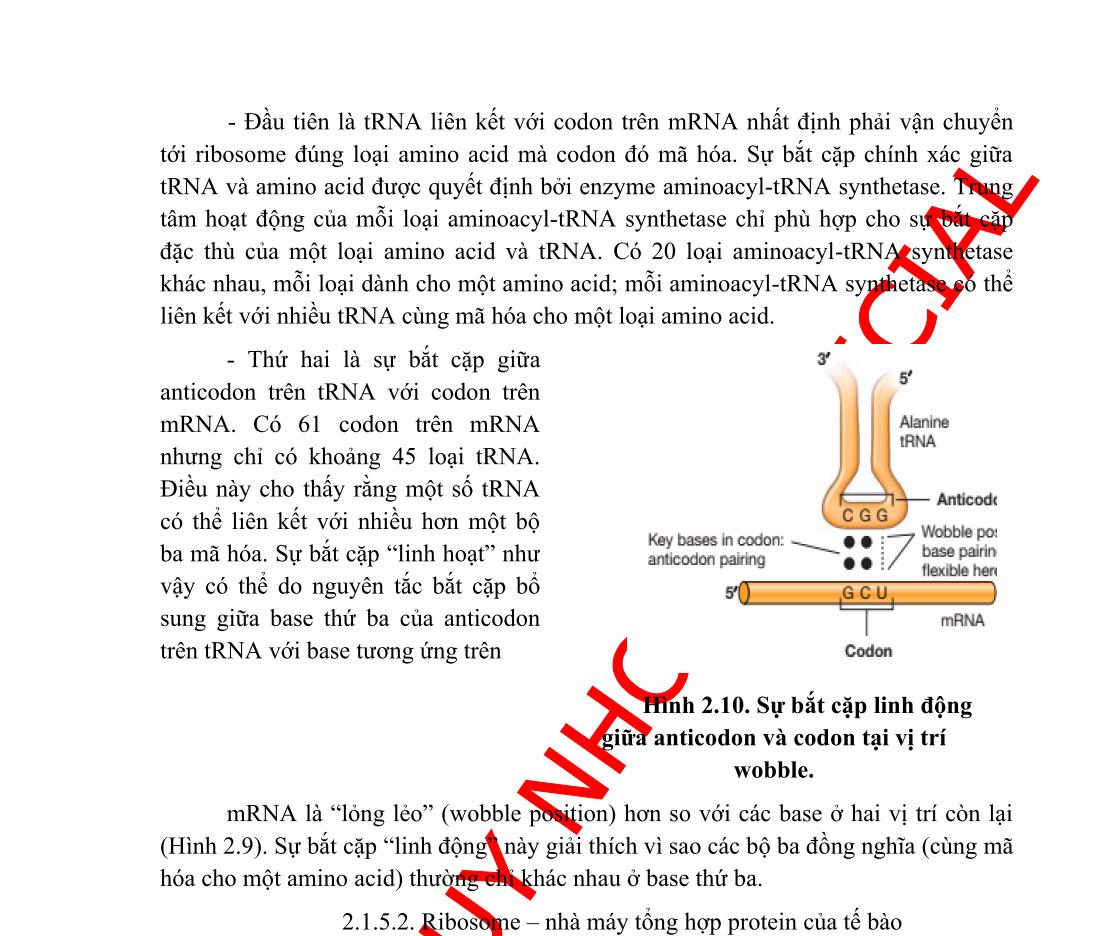
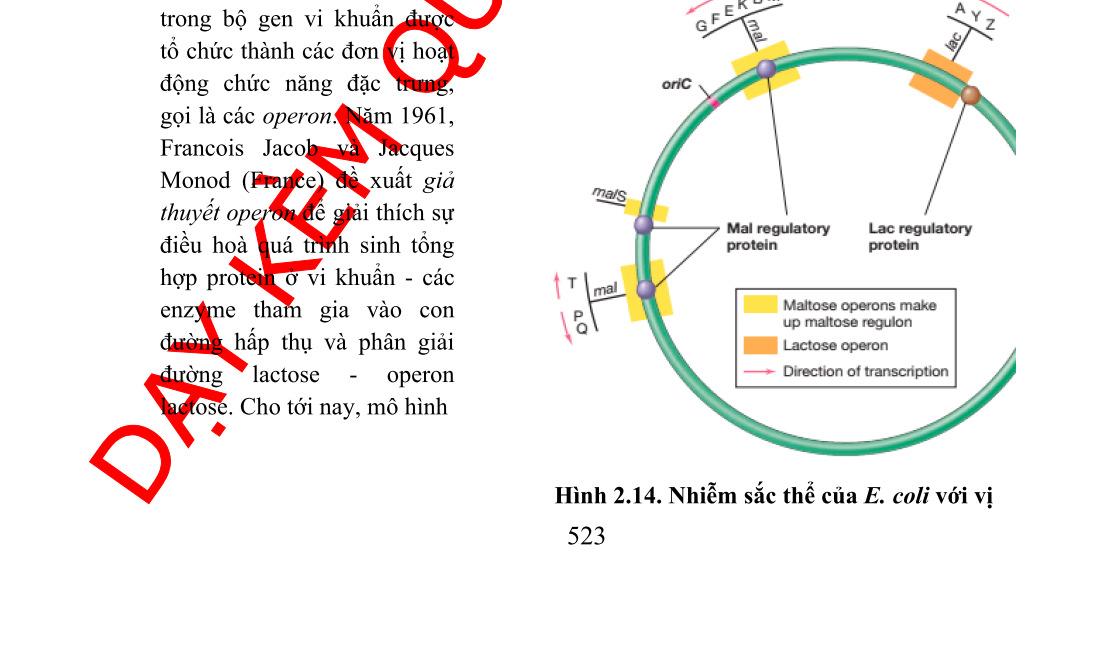
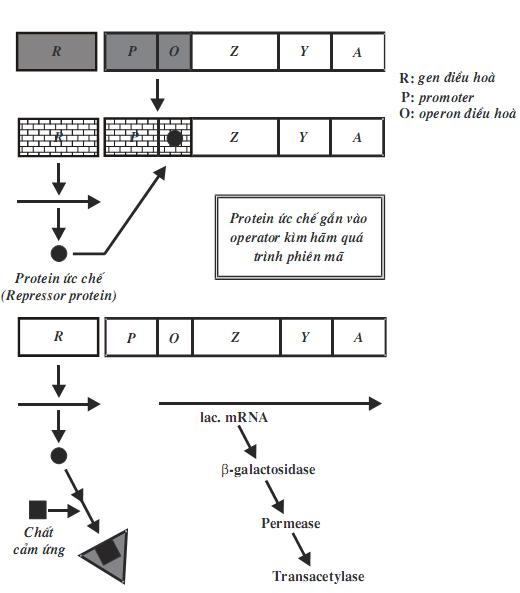
Bộ gen của vi khuẩn được tổ chức theo operon. Một operon có một vùng điều khiển và vùng mã hóa. Vùng điều khiển bao gồm gen điều hoà, promoter và điểm điều hành (operator). Vùng mã hóa bao gồm một số gen cấu trúc nằm kề nhau, mỗi gen cấu trúc mã hóa cho một polypeptide. Các prôtêin được mã hóa trong một operon thường có quan hệ mật thiết với nhau trong một quá trình chuyển hóa sinh hóa nào đó trong tế bào.
Một operon có ít nhất một promoter, tuy nhiên, operon có thể có nhiều hơn một promoter và ái lực của các promoter này đối với ARN-polymerase là khác nhau. Operator là trình tự ADN, nơi mà prôtêin ức chế (repressor prôtêin) gắn vào.
Khái niệm về operon được F. Jacob và các cộng sự của ông nêu ra năm 1961 khi nghiên cứu sự kiểm soát di truyền hấp thụ đường lactose ở E.coli. Với sự khám phá ra cơ chế điều hoà biểu hiện gen, ông đã nhận được giải thưởng Nobel 1965. Cơ chế điều hòa di truyền của hệ thống lac (lac system) được giải thích bằng mô hình operon của Francois Jacob và Jacque Monod (1960)
2.1.1. Các khái niệm
2.1.1.1. Gen cơ định (constitutive gene)
Trong hệ gen, cụ thể là trong quá trình chuyển hóa đường glucose, tổng hợp axit amin,… cần có các thành phần tham gia như tARN, rARN, protein cấu thành nên riboxom, ADN polimerase,… và các enzim tham gia vào quá trình trao đổi chất cơ bản thiết yếu cho sự cần thiết của tế bào. Gen đóng vai trò mã hóa cho các thành phần này hoạt động thường xuyên và liên tục ở hầu hết các tế bào được gọi là gen cơ định. Các cơ thể có ưu thế chọn lọc cao hơn thường có cơ chế điều hòa sao cho các gen tổng hợp những sản phẩm không được sử dụng thường xuyên hoạt động ít hơn và hầu hết chỉ hoạt động trong thời gian cần thiết của tế bào và trong điều kiện môi trường nhất định để nhằm mục đích không tốn năng lượng của tế bào. Đây cũng là một trong những bằng chứng của tiến hóa sau hàng triệu năm tiến hóa của tế bào. Các ATP là nguồn năng lượng thiết yếu của tế bào, nó có thể mất một cách nhanh chóng và cũng khó để tổng hợp một cách dễ dàng trong điều kiện tự nhiên khắc nghiệt. Vì vậy, mọi dạng sống hiện nay bao gồm cả vi khuẩn và virus đã biết cách “tiết kiệm” chúng bằng các cơ chế biểu hiện gen rất hiệu quả. 2.1.1.2. Sự cảm ứng, gen cảm ứng- sự giải nén, gen bị ức chế: - Với một chiếc điện thoại thông thường hay smartphone bình thường thì chúng đều ở trạng thái tắt hoặc “ngủ” thực ra thì vẫn xảy ra các quá trình hao tốn pin
DẠYKÈMQUYNHƠN
351
ẨN
OFFICIAL
CHƯƠNG II: ĐIỀU HÒA HOẠT ĐỘNG GEN Ở VI KHU
nhưng các quá trình này là không đáng kể. Khi xảy ra các hoạt động chạm vào màn hình, hay bấm bàn phím dưới một lực tác động chúng có thể hoạt động một cách mạnh mẽ, hoạt động một cách bất bình thường.
Cũng với hành động như vậy, khi vi khuẩn E.coli được chuyển vào môi trường có lactose làm nguồn hidrat carbon, thì chúng nhanh chóng trong vòng 10 phút tồng hợp các enzym chuyển hóa và sử dụng lactose. Đó là kết quả của việc “bật” sự biểu hiện của gen để đáp ứng lại sự có mặt của tín hiệu nào đó trong môi trường được gọi là sự cảm ứng.
OFFICIAL
- Các gen được điều hòa theo quá trình “bật”, “tắt’ được gọi là gen cảm ứng (inducible genes). Sản phẩm do gen như vậy mã hóa, nếu là enzym thì gọi là các enzym cảm ứng. Nếu là protein được gọi là protein cảm ứng. Các enzym cảm ứng thường là các enzym tham gia vào quá trình dị hóa - Ngược lại, khi sản phẩm tổng hợp một cách dư thừa quá mức hoặc trong môi trường có xảy ra việc dư thừa chất cần thiết cho cơ thể thì tế bào có sự điều hòa ngược lại đó là sự giải nén. Sự giải nén đơn giản là việc “tắt” sự biểu hiện của gen. Các gen được điều hòa theo kiểu này được gọi là các gen bị ức chế (hay gen bị nén). Các enzym tổng hợp từ gen bị ức chế tham gia vào quá trình đồng hóa. 2.1.1.3. Điều hòa dương tính và điều hòa âm tính - Điều hòa âm tính và dương tính là hai quá trình đi liền với nhau và không thể thiếu trong quá trình ức chế hay kể cả quá trình kích ứng. Cả hai công việc này đều liên quan đến gen điều hòa. Các gen điều hòa này cho sản phẩm trực tiếp điều hòa hoạt động của các gen khác. Trong cơ chế điều hòa dương tính, sản phầm của gen điều hòa có vai trò làm tăng sự biểu hiện của một hay một số gen cấu trúc. Còn trong quá trình điều hòa âm tính, sản phẩm của gen điều hòa thường ức chế hoặc làm tắt sự biểu hiện của gen cấu trúc.
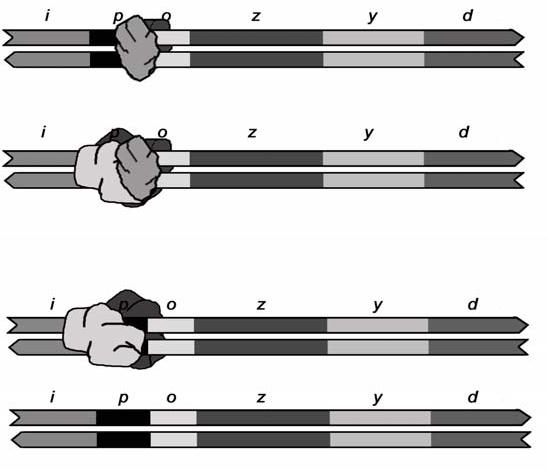
- Các gen điều hòa hoạt động do có các enzym liên kết vào các protein của chúng (protein điều hòa). Vị trí đó được gọi là vị trí liên kết protein điều hòa (RPBS) thường nằm gần promoter của gen mà nó điều hòa.
- Dựa theo hai cơ chế điều hòa thì cũng có hai loại protein gọi là protein hoạt hóa trong hệ thống điều hòa dương tính và protein ức chế trong hệ thống điều hòa âm tính. Không phải protein điều hòa muốn liên kết vào RPBS là được ngay mà cần phải có một phân tử trung gian liên kết cũng giống như muốn các ốc vít được gắn kết chặt hơn thì cần có các goong ở giữa nhằm các ốc đỡ bị chờn ra ngoài. Các phân tử như vậy được gọi là các phân tử tín hiệu. Cũng như các khái niệm khác thì phân tử tín hiệu cũng có hai loại phù hợp với chức năng của chúng, nếu phân tử làm tăng cường sự biểu hiện của gen thì gọi là phân tử kích ứng, còn nếu hạn chế hay kìm hãm sự biểu hiện gen thì gọi là phân tử dồng ức chế - Các phân tử này gắn kết vào protein điều hòa và làm thay đổi cấu hình của chúng, dẫn đến sự thay đổi hoạt tính hoặc chức năng của các protein. Việc gắn kết và
DẠYKÈMQUYNHƠN
352
có kết quả như vậy được gọi là sự biến đổi dị hình. Việc này xảy ra là do sự thay đổi ái lực liên kết của chúng vào các vị trí RPBS ở gần promoter của các gen mà chúng điều khiển.
2.1.2. Các nguyên tắc
2.1.2.1. Biểu hiện gen được điều khiển bởi các protein điều hòa
OFFICIAL
- Sự biểu hiện gen ở sinh vật nhân sơ hay kể cả sinh vật nhân thực đều phụ thuộc vào các yếu tố môi trường, cụ thể là các tín hiệu từ môi trường. Ở vi khuẩn, điển hình là các phân tử có mặt trong môi trường sống. Và các thông tin từ phân tử tín hiệu thì được chuyển tới gen thông qua các protein điều hòa. Như ta đã biết ở quá trình phiên mã, các ARN polimerase liên kết vào promoter và làm cho phức hệ đang “đóng” chuyển thành ‘mở” và dãn xoắn mạch. Sau đó, ARN polimerase tiếp tục quá trình phiên mã. Điều này chứng minh rằng nó phụ thuộc vào promoter và protein điều hòa. 2.1.2.2. Nhiều promoter được điều khiển bằng ái lực giữa ARN polimerase với ADN - Bình thường, khi promoter liên kết với ARN polimearase chúng có liên kết yếu làm cho việc khởi đầu phiên mã không thế diễn ra một cách bình thường. Nhưng đôi khi quá trình này vẫn xảy ra theo biểu hiện gen kiểu cơ định, nhưng chỉ ở mức tối thiểu. Còn khi có protein điều hòa gắn giữa ARN polimerase và promoter quá trình phiên mã sẽ diễn ra một cách suôn sẻ bởi liên kết cao hơn do được bổ sung các yếu tố của trình tự liên ứng. Đây được gọi là việc giới hạn tốc độ biểu hiện gene. - Để điều hòa âm tính một promoter cần có một protein ức chế gắn vào một vị trí bên trong hoặc gối lên promoter. Vị trí này gọi là trình tự điều hành (operator). Protein này gắn vào operator, ARN polimearase sẽ không gắn được vào promoter và sự phiên mã khoog được thực hiện. Ngược lại, để điều hòa dương tính, một protein điều hòa sẽ giúp ARN pol dễ dàng liên kết với promoter. Hiện tượng này xảy ra bằng cách protein hoạt hóa dùng một phần phân tử của nó để liên kết với ADN tại vị trí gần promoter, còn phần khác tương tác với ARN pol, nhờ vậy đưa enzym đến gần promoter hơn. Cơ chế này gọi là sự liên hợp ADN/protein. Vai trò của protein chỉ như một chất đính kết gắn ADN và ARN pol hay đưa enzym đến gần promoter hơn. Và quá trình mở có thể dễ dàng xảy ra, dẫn đến sự phiên mã diễn ra. - Operon lac ở vi khuẩn E.coli là một điển hình hoạt động với một protein họat hóa và một protein ức chế
2.1.2.3. Quy tắc dị hình Quy tắc dị hình là một trong những quy tắc cơ bản nhất trong điều hòa chức năng và hoạt tính protein. Ví dụ: các protein Cyclin hoạt hóa các enzym Cdk tham gia điều khiển chu trình tế bào. Các Cyclin hoạt hóa được enzym này bằng cách liên kết vào enzym và chuyển chúng từ trang thái không hoạt động sang trạng thái hoạt động. Ở glnA, chất hoạt hóa (NtrC) tương tác với ARN pol và thúc đẩy phức hệ khởi đầu
DẠYKÈMQUYNHƠN
353
phiên mã đóng chuyển thành mở. Còn trong trường hợp metT, chất hoạt hóa (MerR) tạo ra hiệu ứng tương tự nhưng bằng sự thay đổi cấu hình ADN (promoter).
Qua đây ta thấy được việc điều hòa phiên mã rất đa dạng và phong phú. Ta có thể kết hợp đồng thời nhiều cơ chế khác nhau hoặc sự ức chế phiên mã không nhất thiết phải trải qua sự ngăn cản liên kết giữa ARN pol và promoter.
2.1.2.4. Sự biểu hiện gen từ khoảng cách xa và cấu trúc vòng thắt ADN
OFFICIAL
- Các protein điều hòa thường liên kết ở vị trí gần promoter của gen, nhưng điều này không phải lúc nào cũng đúng. Đôi khi protein điều hòa liên kết vào ADN ở vị trí cách gen rất xa. Lúc này, để protein điều hòa có thể tương tác được với gen, thường có sự hình thành các vòng thắt trên phân tử ADN. Trong thực tế, protein hoạt hóa phiên mã NtrC hoạt động theo cách này. Vị trí liên kết ADN của nó nằm ngược dòng promoter (về phía đầu 5’ của mạch mã hóa) 150 bp. Protein này thậm chí có thể hoạt hóa gen ngay cả khi nó liên kết ở vị trí xa hơn (1 kb hoặc hơn). Trong việc điều khiển như vậy, vòng thắt có thể dài tới 3 kb. Một cách khác để đưa các ADN đến gần nhau là nhờ các protein liên kết vào đoạn trình tự ở giữa. Đây là hai cách phổ biến ở sinh vật nhân sơ, còn ở sinh vật nhân thực ta còn thấy có nhiều cách phong phú hơn. 2.1.2.5. Quy tắc dị hình và liên kết ADN – protein Sự tương tác giữa các đại phân tử nói chung và giữa ADN và ARN pol nói riêng đã hoạt hóa gen từ đó huy động bộ máy phiên mã tới promoter. Trong những trường hợp khác thì gen lại phiên mã theo quy tắc dị hình. Tức là, môt protein liên kết với enzym ARN pol đã liên kết sẵn với ADN và làm thay đổi cấu hình hoặc của enzym và ADN hoặc của promoter, qua đó sự khởi đầu phiên mã có thể diễn ra. Cả hai quá trình này đều có những vao trò khác nữa trong điều hòa hoạt động của gen và cách tương tác giữa chúng có thể phức tạp hơn. Ví dụ như hai hay nhiều protein điều hòa cùng tương tác với nhau và với ADN. Qua đó chúng hỗ trợ nhau cùng tăng cường liên kết vào vị trí gen được điều khiển. Kiểu tương tác này tạo nên tác dụng hợp lực và có thể nhanh chóng chuyển một gen từ trạng thái không biểu hiện sang trạng thái biểu hiện tối đa. Sự hợp lực của các protein hoạt hóa còn có vai trò tích hợp tín hiệu. Đó là khi các gen chỉ biểu hiện khi đồng thời có mặt nhiều tín hiệu khác nhau. Sự biểu hiện gen ở phago là một ví dụ điển hình cho kiểu điều hòa này.
2.1.2.6. Khởi đầu phiên mã không phải là cơ chế điều hòa duy nhất ở các gen Sự điều hòa phiên mã không chỉ xảy ra ở bước khởi đầu phiên mã mà còn ở các giai đoạn khác như kéo dài phiên mã, hoàn thiện mARN, dịch mã tổng hợp protein.
2.2. Điều hòa operon tham gia con đường dị hóa ở vi khuẩn
2.2.1. Điều hòa âm tính Operon lactose ở E.coli
Vi khuẩn E.coli có khả năng sử dụng đường lactose như nguồn cacbon chính để phát triển. Lactose là một disacarit, nên khi có mặt lactose trong môi trường, vi khuẩn sản sinh ra enzym β-galactosidase để thuỷ phân lactose, tạo thành các phân tử
DẠYKÈMQUYNHƠN
354
monosacarit là glucose và galactose thuận lợi cho quá trình chuyển hóa. Ngoài βgalactosidase ra, tế bào vi khuẩn còn tổng hợp enzym permease làm nhiệm vụ vận chuyển lactose qua màng tế bào. Khi nghiên cứu sự kiểm soát di truyền hấp thụ đường lactose, F. Jacob đã phát hiện cơ chế tổng hợp các enzym hấp thụ đường lactose. Quá trình thí nghiệm có thể mô tả như sau
Khi nuôi vi khuẩn E.coli trên môi trường không có lactose, thì ta thấy nồng độ enzym β-galactosidase và permease do vi khuẩn sinh ra rất thấp: khi đưa đường lactose vào môi trường nuôi thì nồng độ hai enzym này ở tế bào vi khuẩn tăng lên rất nhiều.
Nhưng nếu nuôi vi khuẩn trên môi trường có đường glucose và lactose đồng thời thì nồng độ hai enzym nêu trên trong tế bào thấp hơn so với trường hợp chỉ có đường lactose. Khi phân tích tổng nồng độ mARN có mặt trong tế bào trước và sau khi đưa lactose vào trong môi trường nuôi, thấy rằng: Khi không có mặt lactose trong môi trường thì không xuất hiện mARN mã hóa cho β-galactosidase và permease. Khi bổ sung lactose vào môi trường nuôi thì trong tế bào xuất hiện mARN mã hóa cho hai enzym nêu trên. Từ kết quả thí nghiệm trên, cùng với những kết quả thu được về đột biến gen, F. Jacob và J. Monod đã đưa ra mô hình điều khiển operon lactose như hình bên dưới
Cấu trúc của operon lactose gồm: gen điều hoà (R), promoter (P), operator (O) và ba gen cấu trúc là lacZ mã hóa cho enzym β-galactosidase, lacY mã hóa cho enzym permease và lacA mã hóa cho enzym transacetylase.
Khi không có mặt lactose trong môi trường, gen điều hoà thường xuyên tổng hợp prôtêin ức chế. Prôtêin ức chế có ái lực với điểm điều hành (operator) nên nó gắn vào điểm điều hành, ngăn cản không cho enzym ARN- polymerase thực hiện phiên mã, mARN không thể tổng hợp được, operon đóng.
Khi có mặt lactose trong môi trường, nhờ enzym permease có sẵn ở màng tế bào chuyển một lượng rất ít vào trong tế bào. Khi đã vào trong tế bào, lactose chuyển thành allolactose (có chứa liên kết β-1,6). Allolactose là chất cảm ứng, nó liên kết với prôtêin kìm hãm. Phức hợp này không có ái lực với operator, nên không gắn lên operator được, lúc này, operon mở. ARN- polymerase thực hiện phiên mã các gen cấu trúc.
DẠYKÈMQUYNHƠN
OFFICIAL 355
Hình 2.1.Mô hình hoạt động của operon lactose Phần lớn sự tổng hợp các enzym hoạt động trong các quá trình phân giải ở tế bào được sự kiểm soát theo cơ chế của operon cảm 2.2.2. Điều hòa dương tính operon lactose Chức năng của β-galactosidase trong chuyển hóa lactose để tạo ra glucose bằng cách cắt lactose (một sản phẩm cắt khác là galactose cũng có thể được chuyển thành glucose nhờ enzyme của operon galactose). Nếu cả glucose và lactose có mặt trong môi trường sinh trưởng, không cần hoạt động của operon lac. Trong môi trường có glucose, enzyme β- galactosidase không được tạo thành cho đến khi glucose trong môi trường được sử sụng hết. Sự có mặt của glucose làm mRNA không được tổng hợp, do đó không có sự tổng hợp β-galactosidase, vì sự thêm vào nhân tố cảm ứng làm bất hoạt chất kìm hãm. Một yếu tố khác cần cho sự bắt đầu tổng hợp mRNA, hoạt tính của yếu tố này được điều hòa bởi nồng độ glucose. Glucose có ảnh hưởng ức chế gián tiếp lên sự biểu hiện của operon lac.
DẠYKÈMQUYNHƠN
356
OFFICIAL
Những phân tử nhỏ adenosine monophosphate vòng (cAMP) phân bố rộng rãi trong mô động vật và trong các cơ thể eukaryote đa bào, có vai trò quan trọng làm chất trung gian hoạt động hormone. Chất này cũng có trong tế bào E. coli và nhiều tế bào vi khuẩn khác với chức năng khác nhau. cAMP được tổng hợp bởi enzyme adenyl cyclase, và nồng độ của cAMP được điều hòa gián tiếp qua trao đổi chất glucose. Khi vi khuẩn sinh trưởng ở môi trường chứa glucose, hàm lượng cAMP rất thấp. Trong môi trường chứa glycerol hoặc các nguồn carbon không thể đi vào con đường hóa sinh được sử dụng để trao đổi chất glucose (con đường glycolytic) hoặc khi vi khuẩn bị đói nguồn năng lượng, nồng độ cAMP cao. Hàm lượng glucose giúp điều hòa nồng độ cAMP trong tế bào và cAMP lại điều hòa hoạt tính của operon lac.
E. coli chứa protein nhận cAMP hay CRP (cyclic AMP receptor protein) còn được gọi là protein hoạt hóa dị hóa CAP (catabolite activator protein), được mã hóa bởi gen crp. Đột biến ở gen crp và gen adenyl cyclase làm ngăn cản sự tổng hợp của mRNA lac.Điều này cho thấy chức năng của CRP và cAMP cần thiết cho tổng hợp mRNA lac. CRP và cAMP gắn vào một vị trí khác tạo phức hợp cAMP-CRP được biểu hiện. Phức hợp này là một yếu tố điều hòa hoạt hóa ở hệ thống lac
OFFICIAL
không
Repressor không
Phức hợp cAMP-CRP Có Phiênmã không
Tổng hợp của mRNA
Hình 2.2. Bốn trạng thái điều hòa của operon lac Nhu cầu về cAMP-CRP phụ thuộc vào hệ thống ức chế lac, vì đột biến crp và adenyl cyclase không thể tạo mRNA lac ngay cả khi có mặt đột biến ở gen điều hòa (lacI-) và gen chỉ huy (lacO-). Sở dĩ như vậy là vì phức hợp cAMP-CRP phải gắn vào trình tự base trên DNA ở vùng promotor để xảy ra phiên mã (Hình 7.4). Khác với chất ức chế trong điều hòa âm tính, phức hợp cAMP-CRP là chất điều hòa dương tính. Các thí nghiệm ở điều kiện in vitro cho thấy: mRNA lac được tổng hợp chỉ khi có cAMP vòng và không có chất
DẠYKÈMQUYNHƠN
357
ức chế.
Khi vắng mặt phức hợp cAMP-CRP, RNA polymerase chỉ bám lỏng lẽo vào promotor. Vì vậy hiếm khi dẫn đến phiên mã vì không có sự tương tác đúng giữa RNA polymerase và promotor. Nhưng RNA polymerase được kích thích gắn vào promotor khi cAMP-CRP được gắn vào DNA.
Những kết quả này giải thích chức năng lactose và glucose cùng nhau tham gia điều hòa phiên mã operon lac như thế nào.
Khi trong môi trường có nhiều nguồn cácbon khác nhau, thông thường vi sinh vật sẽ sử dụng nguồn cácbon quen thuộc trước, sau khi cạn kiệt nguồn đó thì vi sinh vật mới chuyển dần sang sử dụng nguồn cácbon kế tiếp.
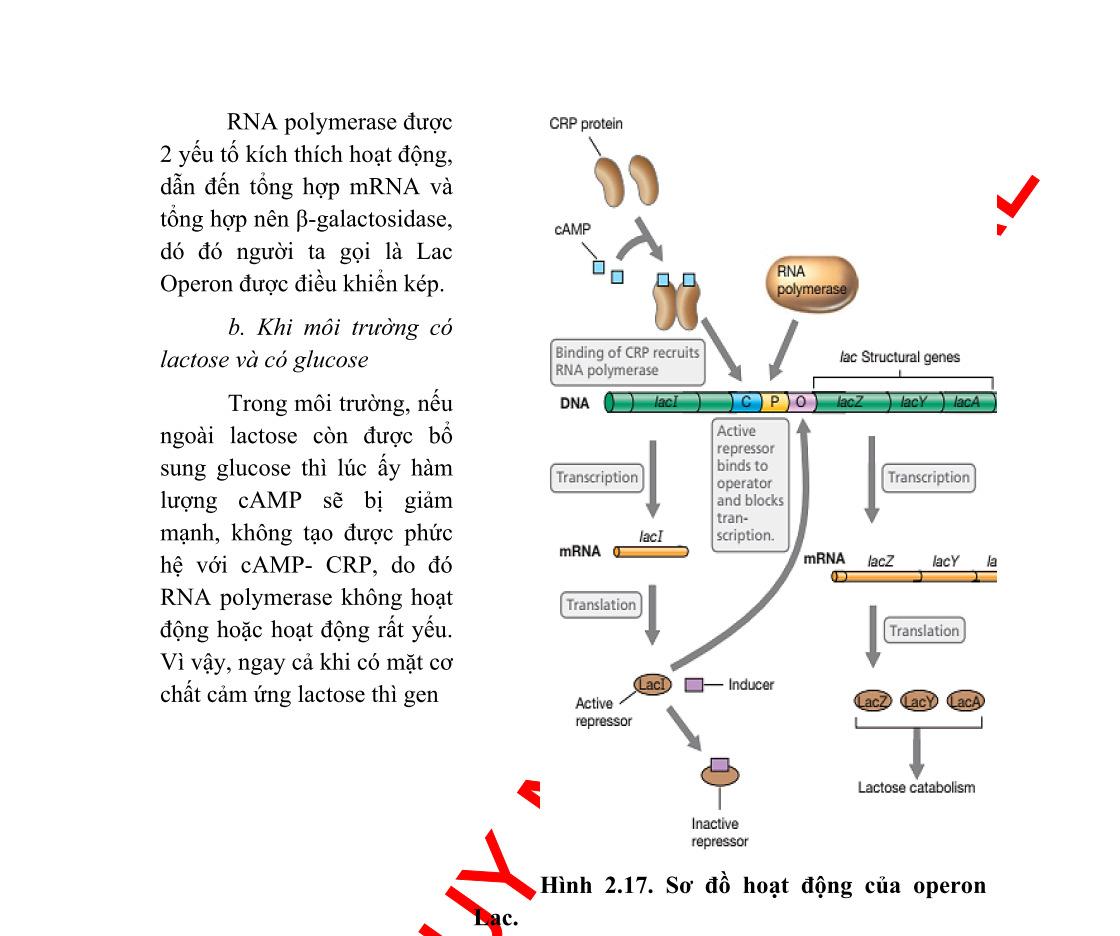
Ví dụ: Ở vi khuẩn E.coli nếu trong môi trường nuôi cấy có cả glucozơ và lactozơ thì vi khuẩn sẽ sử dụng glucozơ trước, đến khi cạn dần glucozơ thì mới chuyển sang dùng nguồn cácbon còn lại. Như vậy, có thể khẳng định rằng ở vi sinh vật, gen cơ định mã hóa các enzym chuyển hóa nguồn cácbon nào thì trong môi trường có nhiều nguồn cácbon chúng sẽ sử dụng nguồn cácbon đó trước, vì trong tế bào chúng luôn có sẵn enzym đó. Vậy bằng cách nào mà vi khuẩn cảm nhận được sự thay đổi nồng độ cácbon đang sử dụng? Đó là nhờ một phân tử AMP vòng (CAMP), phân tử CAMP sẽ được tích lũy dần khi nồng độ glucozơ giảm dần. Mà prôtêin điều hòa hoạt hóa operon Lac là prôtêin khi ở dạng tự do nó không hoạt động. Khi CAMP liên kết với prôtêin hoạt hóa nó trở thành dạng hoạt động (CAP), khi CAP liên kết vào vị trí điều hòa của operon nó sẽ làm tăng ái lực của promotor với ARN polimeraza, dẫn đến gen tăng biểu hiện. Khi tăng nồng độ glucozơ thì lượng CAMP lại giảm đi. Lượng phức hệ CAP giảm, ái lực của promotor với ARN polimeraza giảm, gen phiên mã giảm đi.
Do trình tự liên kết của chất ức chế lacI tại operator phủ lên promotor lac một phần, nên khi lacI liên kết vào operator, nó ngăn cản sự liên kết của ARN polimeraza vào promotor. Còn đối với CAP, khi thiếu vắng prôtêin này, ARN polimeraza chỉ liên kết được rất yếu vào promotor. Bởi vì, promotor lac thiếu một trình tự UP, đồng thời trình tự liên ứng của nó không hoàn chỉnh.
Ngược lại, prôtêin CAP khi liên kết vào operon lac lại có vai trò thúc đẩy operon hoạt động. Tuy vậy, CAP chỉ liên kết mạnh khi môi trường không có glucozơ. cAMP là phân tử kích ứng dị hình của CAP. Nghĩa là, chỉ khi cAMP liên kết với CAP, thì prôtêin này mới có cấu hình phù hợp liên kết với ADN.
Khi CAP liên kết vào vị trí này, nó đồng thời huy động và thúc đẩy ARN polimeraza đính kết vào promotor. CAP hoạt hóa operon Lac bằng việc huy động enzym ARN polimeraza đến promotor. cAMP là phân tử kích ứng dị hình của CAP. Nghĩa là, chỉ khi cAMP liên kết với CAP, thì prôtêin này mới có cấu hình phù hợp liên kết với ADN.
DẠYKÈMQUYNHƠN
OFFICIAL 358
2. 3. Điều hòa Operon đồng hóa ở vi khuẩn
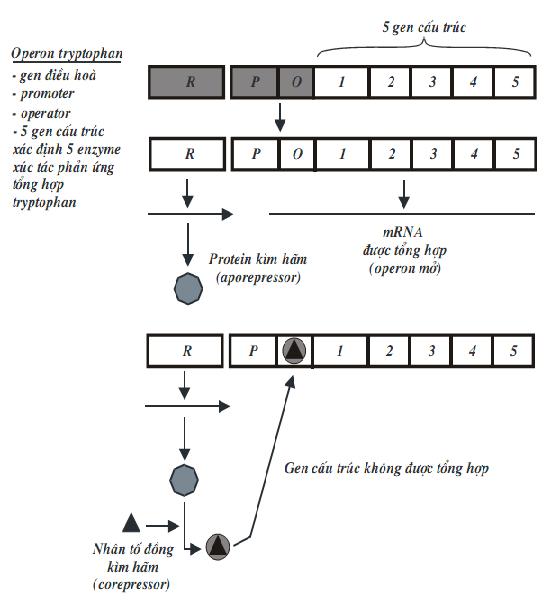
2.3.1. Điều hòa Operon kìm hãm - Operon tryptophan ở E.coli Operon tryptophan cũng có cấu trúc tương tự như operon lactose, nghĩa là, bao gồm gen điều hoà (R), promoter (P), operator (O) và 5 gen cấu trúc. Mỗi gen cấu trúc mã hóa cho một enzym, xúc tác phản ứng tổng hợp tryptophan. Trong tế bào, tryptophan được tổng hợp bằng một chuỗi 5 phản ứng, mỗi phản ứng được xúc tác bằng một enzym mã hóa trong operon. Năm gen cấu trúc mã hóa cho 5 enzym, lần lượt được ký hiệu là trpE, trpD, trpC, trpB và trpA. Gen trpE nằm ngay sau vùng điều hoà, được phiên mã đầu tiên.
Hình 2.3.Mô hình hoạt động của operon tryptophan
Hoạt động của operon tryptophan khác với operon lactose ở chỗ là prôtêin kìm hãm, bản thân nó không có ái lực với operator nên khi đứng riêng một mình, nó không thể gắn vào điểm điều hành, nên operon mở. Ngược lại, khi prôtêin kìm hãm kết hợp với chất đồng kìm hãm (corepressor), thì sẽ có ái lực với operator nên dễ dàng gắn vào đó, làm operon đóng. Hoạt động của operon có thể mô tả như hình trên. Như vậy, khi lượng tryptophan dư, operon đóng, còn khi thiếu tryptophan thì operon mở. Nhìn chung, cách điều hoà biểu hiện gen ở sinh vật procaryote chủ yếu được thực hiện ở giai đoạn phiên mã. Operon tryptophan (trp) của E.coli chứa các gen cấu trúc mã hóa cho các enzyme tổng hợp amino acid tryptophan. Operon này được điều hòa theo cách sau: khi tryptophan có mặt đầy đủ trong môi trường sinh trưởng, sự phiên mã của operon
DẠYKÈMQUYNHƠN
359
OFFICIAL
bị ức chế. Khi sự cung cấp tryptophan bị thiếu, sự phiên mã xảy ra. Sự điều hòa của operon lactose tương tự với operon lactose, vì sự tổng hợp mRNA được điều hòa âm tính nhờ chất ức chế. Tuy nhiên, khác với điều hòa ở operon lac, tryptophan hoạt động như chất đồng kìm hãm, kích thích chất ức chế gắn vào operator ngừng sự tổng hợp. Operon tryptophan hoạt động theo kiểu ức chế, điều hòa âm tính.
Tryptophan được tổng hợp qua các giai đoạn khá phức tạp, mỗi giai đoạn có sự xúc tác của một enzyme đặc biệt. Các gen mã hóa cho các enzyme này nằm kề nhau trên nhiễm sắc thể của E.coli. Đó là các gen trpE, trpD, trpC, trpB, trpA. Các enzyme được dịch mã từ một phân tử mRNA đa cistron. Vùng mã hóa gen E được dịch mã trước tiên. Phía trước trpE về đầu 5' có promotor, operator và 2 vùng xếp lần lượt là leader (trpL) và đoạn kìm hãm phiên mã attenuator (trpa, không phải là trpA). Gen ức chế trpR nằm xa operon, tổng hợp protein aporepressor, là chất kìm hãm mà riêng nó không có hoạt tính. Khi tryptophan dư thừa, nó kết hợp với aporepressor tạo chất kìm hãm có hoạt tính, gắn vào operator của operon tryptophan làm dừng phiên mã các gen cấu trúc. Khi nồng độ tryptophan thấp, nó tách khỏi phức hợp kìm hãm và aporepressor mất hoạt tính. Lúc này operator mở ra, RNA polymerase dịch mã 5 gen cấu trúc để tổng hợp 5 enzyme tham gia tổng hợp tryptophan.
2.3.2. Điều hòa Operon phiên mã dở (Attenuation) Operon tryptophan ở E.coli Kiểu điều hòa thứ hai được phát hiện ở operon tryptophan được gọi là attenuation. Nó dùng sử dịch mã để điều khiển sự phiên mã. Khi có mặt tryptophan nội bào, ngay cả với nồng độ thấp, sự dịch mã một phần vùng leader của mRNA ngay khi vừa được tổng hợp, kết quả làm dừng sự phiên mã trước khi gen cấu trúc đầu tiên của operon được sao chép.
DẠYKÈMQUYNHƠN
360
OFFICIAL
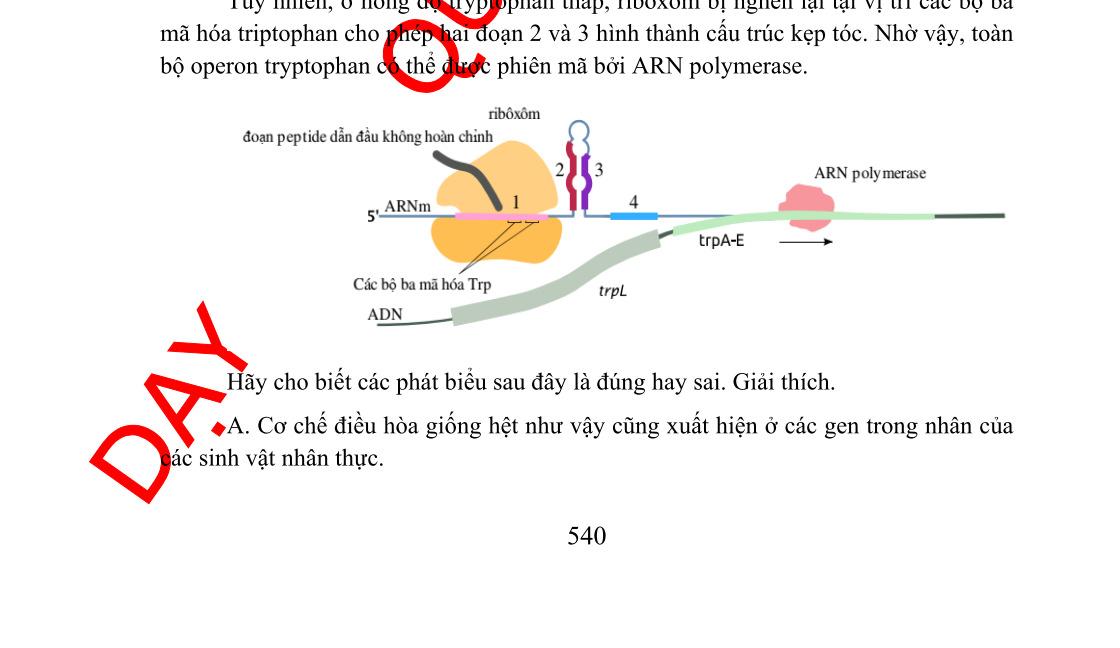
Hình 2.4. Sơ đồ phiên mã của leader trp(A); Chi tiết cấu trúc của 2 codon trp ở vòng 1-2 (B)
Phiên mã dở (attenuation) là kết quả sự tương tác giữa các trình tự DNA trong vùng leader của bản phiên mã trp. Ở tế bào kiểu dại, sự phiên mã operon trp thường được bắt đầu. Tuy nhiên khi có mắt một lượng nhỏ tryptophan, hầu hết phân tử mRNA kết thúc ở vùng 28 base đặc biệt ở trong trình tự leader. Kết quả của sự kết thúc sớm này tạo phân tử mRNA chứa 140 nucleotide chấm dứt một đoạn ngắn của các gen mã hóa cho các enzyme trp. Vùng 28 base xảy ra sự kết thúc phiên mã sớm như thế được gọi là attenuator. Trình tự base của vùng này thường có các tính chất điểm kết thúc, gồm dạng đoạn và vòng (stem-loop) trên mRNA theo sau là trình tự của 8 uridine.
Trình tự leader có các đặc điểm:
- Một vùng có codon AUG và phía sau là codon kết thúc UGA, mã hóa cho một polypeptide chứa 14 amino acid được gọi là leader polypeptide.
- Hai codon tryptophan ở vị trí 10 và 11 trên mRNA của leader polypeptide. Trình tự lặp lại ngắn này có ý nghĩa trọng điều hòa.
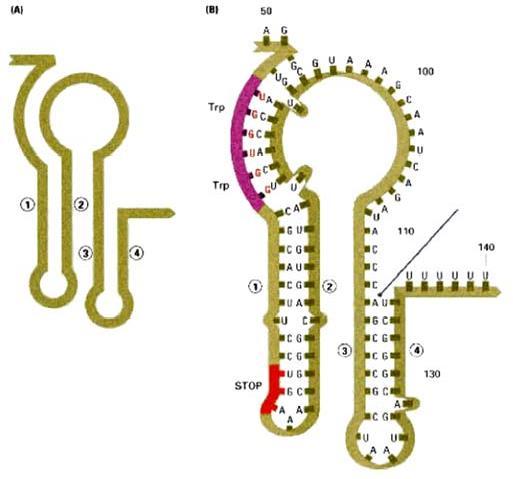
- Bốn đoạn của RNA leader là vùng 1, 2, 3 và 4 tạo thành do khả năng kết cặp của các base với nhau. Các base ở vùng 1 kết cặp với vùng 2, vùng 3 kết cặp với vùng 4.
Khi sự kết cặp xảy ra ở dạng này, sự phiên mã kết thúc ở đoạn đi qua uridine phía trước nucleotide 140. Kiểu kết cặp này xảy ra ở mRNA leader được tinh sạch. - Một kiểu kết cặp biến đổi có thể xảy ra, trong đó các base vùng 2 kết cặp với vùng 3 nhờ các cặp base ở 2 vùng này gần như bổ sung nhau. Qua mô hình kết cặp base biến đổi này (3-4 hoặc 2-3), sự tổ chức trình tự mRNA có thể điều hòa
DẠYKÈMQUYNHƠN
361
OFFICIAL
[A [B
Kết thúc phiên mã
phiên mã qua dịch mã của leader polypeptide. Khi vùng leader được phiên mã, sự dịch mã leader polypeptid cũng bắt đầu. Vì có 2 codon của tryptophan trong trình tự mã hóa, nên sự dịch mã nhạy cảm với số lượng tRNAtrp đưa vào. Nếu môi trường cung cấp đầy đủ tryptophan, ribosome trượt qua codon tryptophan và đi vào vùng 7.1. Sự có mặt của ribosome loại bỏ khả năng kết cặp của vùng khoảng 10 base ở mỗi phía của codon đang dịch mã. Sự có mặt của ribosome ở vùng 2 ngăn cản nó kết cặp với vùng 3. Trong trường hợp này vùng 3 kết cặp với vùng 4, tạo ra điểm kết thúc phiên mã. Sự phiên mã kết thúc khi qua các uridine nằm phía sau vùng 4. Khi số lượng tRNAtrp không đủ, sự dịch mã leader polypeptide bị dừng lại đột ngột ở các codon tryptophan. Sự dùng lại này ngăn cản ribosome tiến vào vùng 2, vì vậy vùng 2 được tự do sẽ kết cặp với vùng 3 làm cản trở sự hình thành cấu trúc kết thúc. Vì vậy phân tử trp mRNA hoàn chỉnh được tạo thành, chứa cả trình tự mã hóa cho gen cấu trúc.
Tóm lại, attenuation là cơ chế điều hòa tinh tế trên cơ sở điều hòa âm tính: Khi tRNAtrp đến đủ cung cấp cho sự dịch mã leader polypeptide, sự phiên mã bị dừng, các trp enzyme không được tổng hợp. Khi nồng độ tRNAtrp quá thấp, sự phiên mã xảy ra cho đến hết, các trp enzyme được tạo nên. Nhiều operon chịu trách nhiệm tổng hợp các amino acid khác (như các operon của leucine, isoleucine, phenylalanine, histidine) cũng được điều hòa nhờ attenuator với chức năng tạo ra vùng kết cặp biến đổi ở bảnphiên mã. Ở operon histidine vùng mã hóa của leader polypeptide chứa 7 codon histidine kế nhau. Ở operon phenylalanine vùng mã hóa cho leader polypeptide chứa 7 codon phenylalanin chia 3 nhóm.
Điều hòa kiểu attenuation không thể xảy ra ở eukaryote vì ở eukaryote sự phiên mã và dịch mã không xảy ra đồng thời. Sự phiên mã xảy ra trong nhân, còn sự dịch mã xảy ra ở tế bào chất.
Điều hòa ở operon lac và operon trp là ví dụ về một trong số các cơ chế quan trọng điều hòa hoạt động gen ở mức phiên mã của prokaryote.
2.3. Câu hỏi bài tập điều hòa hoạt động Operon đồng hóa, dị hóa ở
DẠYKÈMQUYNHƠN
OFFICIAL 362
vi
Câu 1. (Đề thi
quốc tế năm 2014) : So
động của operon Lac (lactose) và operon Trp (tryptophan) trong điều hoà âm tính ở E. coli? Gợi ý trả lời: - Giống nhau: + Sự điều hoà của cả hai operon Lac và Trp đều liên quan đến cơ chế điều hoà các gene kiểu âm tính: Nghĩa là, các operon này đều được
khuẩn
chọn đội tuyển quốc gia vòng 2 dự thi Olympic Sinh học
sánh hoạt
“tắt” bởi prôtêin điều hoà tương ứng của chúng (đều là các prôtêin ức chế do gene điều hoà tổng hợp).
+ Sự điều hoà của cả hai operon Lac và Tryptophan đều tạo cho tế bào tiết kiệm năng lượng và vật chất trong hoạt động sống của nó.
- Khác nhau:
+ Trong operon Lac, các enzyme tham gia vào con đường chuyển hoá lactose còn gọi là các enzyme cảm ứng do quá trình sinh tổng hợp chúng được gây cảm ứng bởi tín hiệu hoá học (trong trường hợp này là allolactose). Theo nguyên tắc tương tự, trong operon Trp các enzym do operon Trp mã hoá được gọi là các enzyme ức chế.
OFFICIAL
+ Trong operon Tryptophan, khi Tryptophan có sẵn trong môi trường hoặc khi lượng tích luỹ trong tế bào của chúng đã đủ thì chính axit amin này kết hợp với prôtêin điều hoà tạo thành phức hợp đồng ức chế liên kết vào trình tự O (operator) làm dừng quá trình phiên mã. Ngược lại trong operon Lac, allolactose làm bất hoạt protein điều hoà làm cho protein này không liên kết được vào trình tự O, nhờ đó quá trình phiên mã diễn ra.
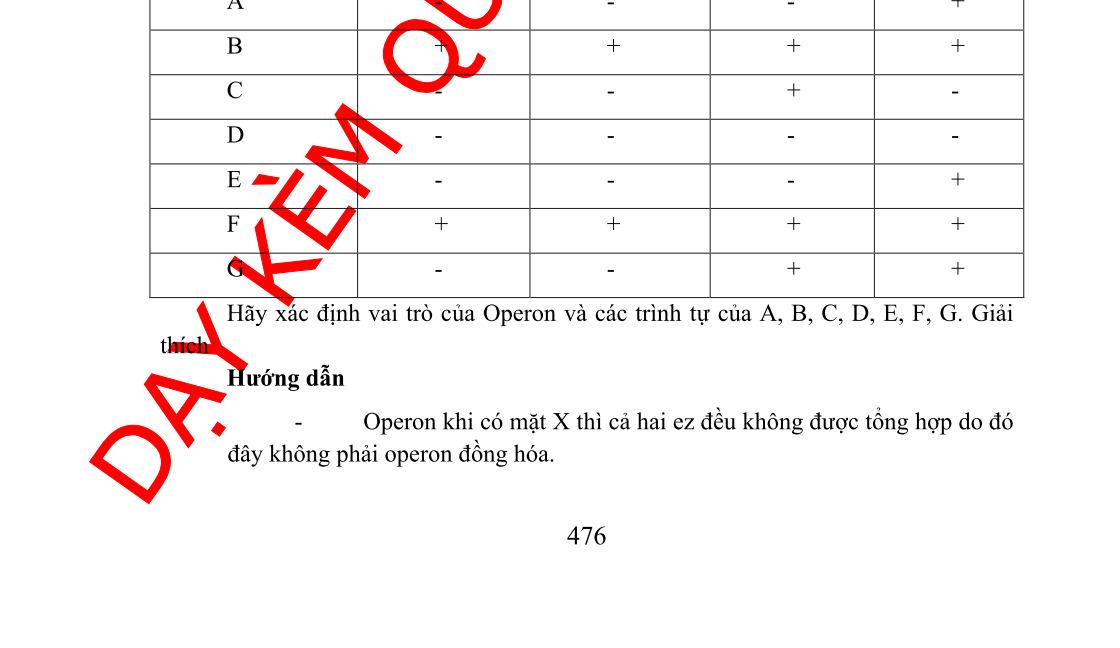
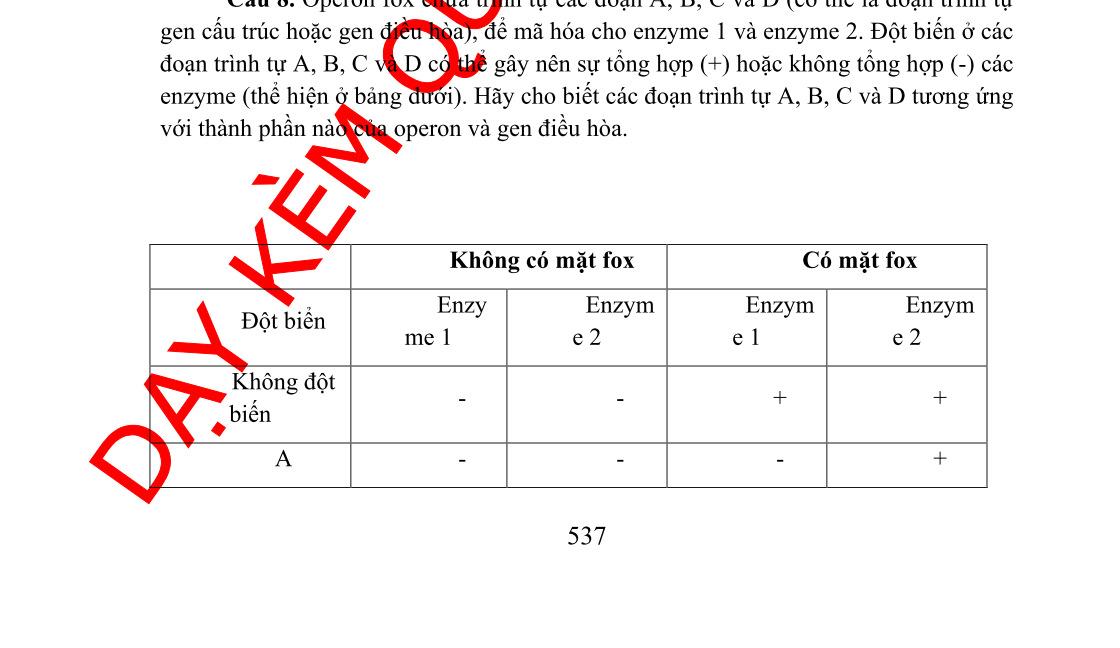
Câu 2: Một operon mã hóa enzyme 1 và 2 được điều hòa bởi sự chuyển hóa chất X chứa đựng 7 trình tự A, B, C, D, E, F, G chưa biết rõ chức năng. Để làm sáng tỏ chức năng của chúng, sự ảnh hưởng của đột biến ở các trình tự từ A đến G dựa trên sự tổng hợp các enzyme được đánh giá thông qua sự có mặt và sự vắng mặt của X. Có mặt X Vắng mặt X Đột biến trong trình tự Enzym 1 Enzym 2 Enzy m1 Enzym2 Không có đột biến - - + + A - - - + B + + + + C - - +D - - -E - - - + F + + + + G - - + + Hãy xác định vai trò của operon và các trình tự của A, B, C, D, E, F, và G. Giải thích.
DẠYKÈMQUYNHƠN
363
Trả lời
- Operon khi có mặt X thì cả hai enzyme đều không được tổng hợp do đó chứng tỏ đây là operon đồng hóa
- Thể đột biến B, F biểu hiện giống nhau. Khi có hay không có mặt X thì operon luôn được mở ( luôn tạo ra enzyme)
- Bị đột biến vùng O do đó protein ức chế không liên kết được với O ( Lac Oc)
- Thể đột biến D khi có mặt hay không có mặt X thì operon luôn đóng
- Bị đột biến vùng P ( ARN pol không liên kết vào P được -> phiên mã không diễnra, operon luôn đóng)
- Thể đột biến A, E biểu hiện giống nhau khi vắng mặt X chỉ có enzyme 2 được tổng hợp
OFFICIAL
- Bị đột biến gen tổng hợp enzyme 1 -> enzyme 1 không được tổng hợp
- Thể đột biến C khi vắng mặt X chỉ có enzyme 1 được tổng hợp
- Bị đột biến gen tổng hợp enzyme 2 -> enzyme 2 không được tổng hợp
- Thể đột biến G biểu hiện giống kiểu dại -> Trình tự không liên quan đến operon mã hóa enzyme 1, enzyme 2
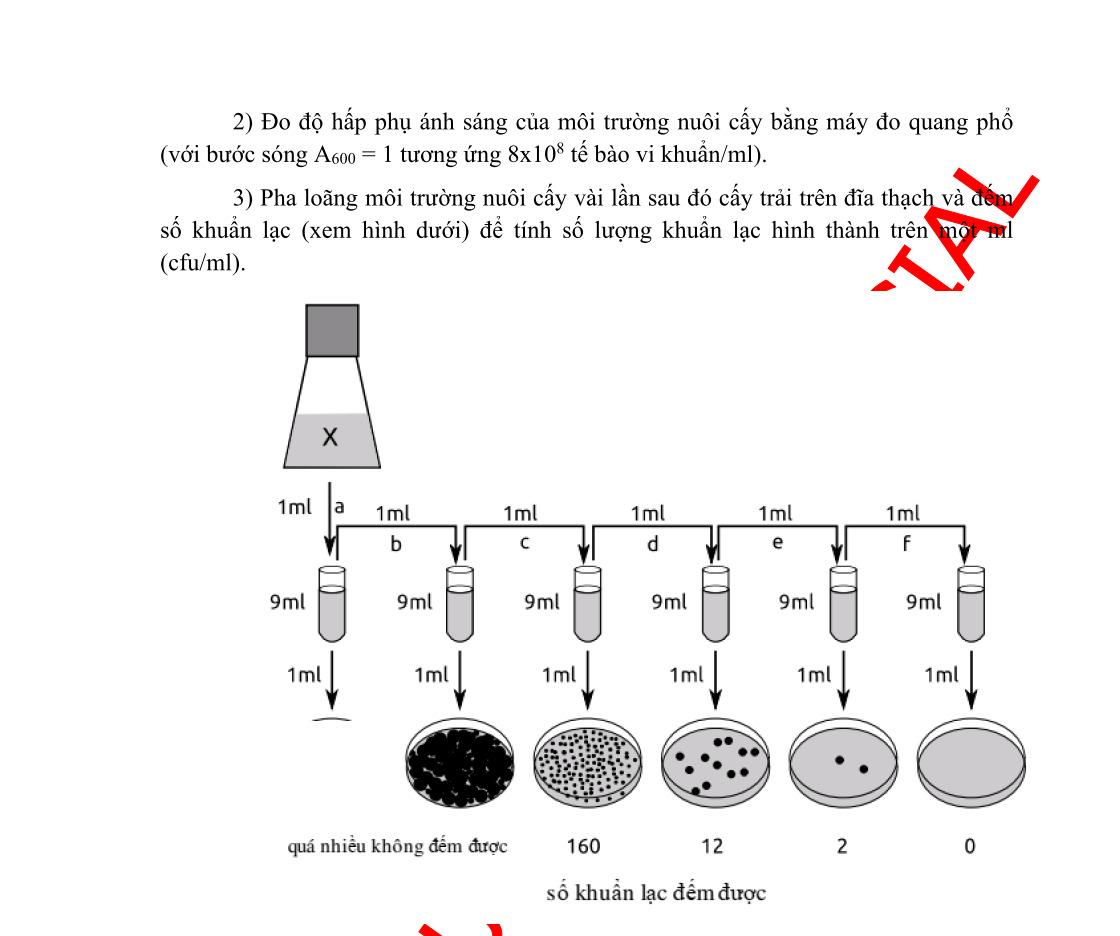
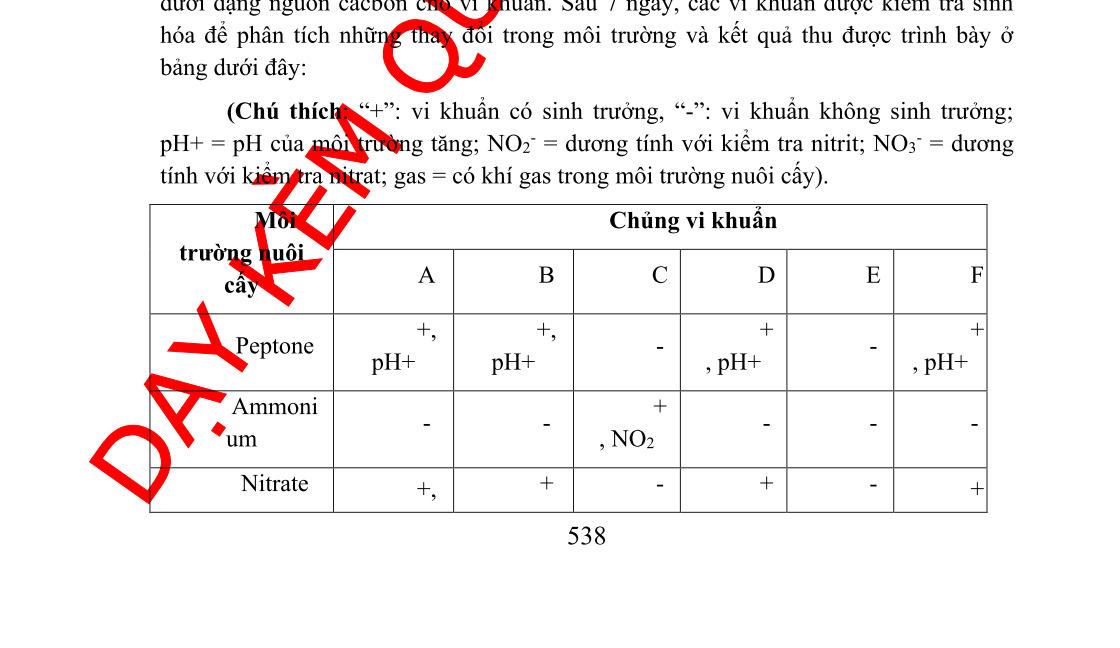
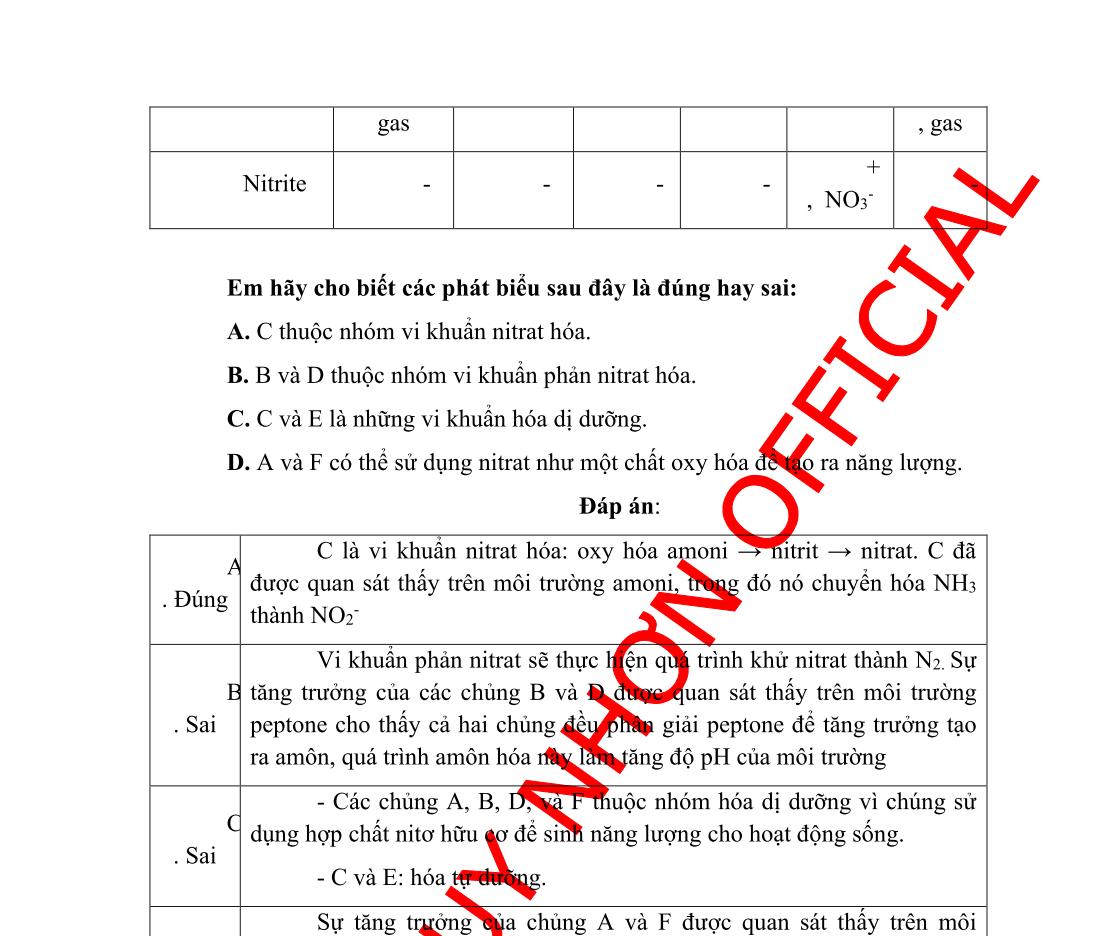
Câu 3:Trong 1 thí nghiệm, hoạt tính của enzym β-galactosidase được đánh giá bởi sự có mặt của X-gal và IPTG trong môi trường nuôi cấy. X-gal là một đồng phân của lactose, nó bị chuyển sang màu xanh khi bị phân giải bởi βgalactosidase, nhưng nó không cảm ứng Lac operon. IPTG là chất cảm ứng giống với lactose. Sau khi gây đột biến, để xác định vị trí đột biến, người ta đã nuôi cấy các đột biến ở trong môi trường khác nhau (bảng dưới) và có bổ sung thêm X-gal. Kết quả ở bảngsau:
Điều kiện môi trường nuôi cấy
Có mặt IPTG, không có glucose
Không có mặt IPTG, không có glucose
Có mặt cả IPTG và glucose
Không có mặt IPTG, có mặt glucose
Kiểu hình khuẩn lạc
Màu xanh
Màu xanh
Màu trắng
Màu trắng
Hãy xác định vị trí đột biến trong operon Lac của chủng đột biến trên. Giải thích. Trả lời
DẠYKÈMQUYNHƠN
364
- Có mặt IPTG, không có glucose -> màu xanh -> không bị đột biến, gen lacZ vẫn được tổng hợp bình thương tạo ra -galatosidase để phân giải Xgal tạo màu xanh
- Không có mặt IPTG, không có glucose -> màu xanh chứng tỏ lacZ vẫn được tổng hợp bình thường -> đột biến xảy ra tại vùng O do đó dù có hay không có cơ chất (IPTG) thì operon Lac vẫn luôn mở
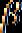
Câu 4: Một số vi khuẩn có cơ chế điều hòa sự tổng hợp enzym liên quan đến quán trình sinh tổng hợp tryptophan (Trp). Trong cấu trúc của operon, phía trước các gen cấu trúc (trpE-A) là một đoạn trình tự leader mã hóa cho đoạn peptit chứa 2 bộ ba mã hóa 2 axit amin Trp liên tiếp. Ở nồng độ cao Trp, ribosome dịch mã mARN của chuỗi peptit leader và rời khỏi bộ ba kết thúc. Hiện tượng này giúp hình thành cấu trúc kẹp tóc giữa đoạn 1-2 và 3-4, dấn đến tín hiệu kết thúc phiên mã sau polyU và ARN polymeraza rời khỏi AND để kết thúc phiên mã (Hình dưới).
OFFICIAL
Tuy nhiên, khi nồng độ tryptophan thấp, ribosome bị dừng tại vị trí 2 mã bộ ba mã hóa tryptophan và cấu trúc kẹp tóc giữa đoạn 2-3 được hình thành. Khi đó trình tự đoạn 4 được tự do và ARN pol tiếp tục phiên mã toàn bộ operon.
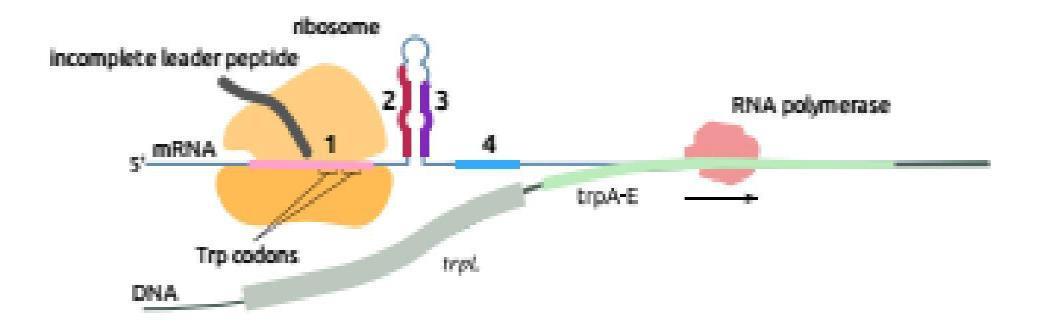
DẠYKÈMQUYNHƠN
365
Quá trình tổng hợp tryptophan sẽ diễn ra như thế nào trong các trường hợp sau? Giải thích.
- Đột biến làm mất 1 bộ ba mã hóa trp trong đoạn leader.
- Đột biến làm thay đổi trình tự dẫn đến việc không hình thành được cấu trúc kẹp tóc giữa đoạn 2 và 3.
- Nồng độ enzym aminoacyl-tARN synthetase (xúc tác sự gắn kết trp vào ARNtTrp) bị suy giảm.
Trả lời:
- Khi đột biến làm mất một bộ ba mã hóa Trp, tín hiệu về Trp trong tế bào sẽ thấp hơn khi đang có cả 2 bọ ba mã hóa, vì thế quá trình điều hòa sẽ mở operon chậm hơn so với bình thường.
OFFICIAL
- Đột biến làm thay đổi trình tự dẫn đến không thể hình thành cấu trúc kẹp tóc 2-3 có thể xảy ra theo 2 trường hợp: - Đột biến xảy ra ở đoạn 2: Trong trường hợp này, cấu trúc đoạn 3 vẫn bình thường, vì thế cấu trúc kẹp tóc 3-4 luôn được hình thành. Vì thế, operon luôn đóng do ARN polymeraza sẽ luôn rời khỏi tại trình tự polyU và kết thúc phiên mã. - Đột biến xảy ra ở đoạn 3: Sự xuất hiện đột biến làm mất các nucleotit có thể hình thành cấu trúc kẹp tóc với đoạn 2, thông thường nó sẽ ảnh hưởng tới việc hình thành cấu trúc kẹp tóc giữa đoạn 3 và 4. Trong trường hợp này, cấu trúc kẹp tóc 3-4 không được hình thành, dẫn đến operon luôn mở, phiên mã được tiếp tục ngay cả khi noognf độ axit amin Trp trong tế bào đang ở nồng độ cao.
* Khi nồng độ enzym aminoacyl-tARN synthetaza bị suy giảm, phức hợp ARNt-Trp sẽ giảm, thiếu nguyên liệu để tổng hợp chuỗi peptit leader, cấu trúc kẹp tóc 2-3 luôn được hình thành, operon luôn mở ngay cả khi nồng độ Trp tăng cao.
Câu 5: Nghiên cứu gen cấu trúc tham gia vào sự chuyển hóa một chất X gồm A, B, D và E. Một trình tự C liên kết với các trình tự A, B và D, còn E nằm cách xa. Người ta phân lập được các đột biến khác nhau ở trình tự C có ảnh hưởng đến sự biểu hiện của các gen cấu trúc.Họ tạo ra các kiểu gen ở các locus của gen A và trình tự C rồi xác định enzyme do gen A mã hóa có được tạo thành khi có mặt hoặc không có mặt chất X như sau: Kiểu gen Không có X Có X C+A+ - +
DẠYKÈMQUYNHƠN
366
C-A+ - -
C-A+/C+A- +
CcA-/C-A+ + +
Cc , C- : Đột biến.
C+ : Bình thường.
dấu (+) enzyme A được tổng hợp ( được biểu hiện ).
dấu (-) enzyme A không được tổng hợp ( không được biểu hiện ).
a. Hãy cho biết trình tự C đóng vai trò gì trong hoạt động của các gen cấu trúc trên? Giải thích. b. Các kết quả trên cho thấy đây là một operon điều hòa theo cơ chế nào?
c. Đột biến Cc thực ra là đột biến gì?
Trả lời
a.Trình tự C là gen điều hòa. Gen điều hòa mang thông tin mã hóa pr chi phối điều hòa hoạt động các gen khác; nên gen C mang thông tin là gen điều hòa chứ không phải là promotor.
b. Khi không có chất X thì gen A không biểu hiện, có chất X thì hoạt động -> X là chất cảm ứng nên đây là một operon điều hòa theo cơ chế cảm ứng.
c. CcA-/C-A+ có hay không có X đều được biểu hiện, nên Cc là một đột biến làm cho kiểu gen luôn luôn kích hoạt gọi là đột biến cơ định -> tạo prôtêin không bám được vào operator.
Câu 6:
a. So sánh cơ chế điều hòa âm tính và điều hòa dương tính ở opêron Lac.
b. Tại sao sự điều hòa hoạt động gen ở sinh vật nhân thực thể hiện khác nhau ở những giai đoạn phát triển khác nhau của cá thể? Gợi ý trả lời:
a)* Giống nhau:
Đều để thích ứng với các điều kiện môi trường biến động, đồng thời để tiết kiệm năng lượng và vật chất của tế bào.
Đều liên quan đến sự tham gia của các gen điều hòa. Các gen này mã hóa cho các sản phẩm trực tiếp (prôtêin điều hòa) điều hòa sự biểu hiện của các gen cấu trúc. Đều có hệ thống điều hòa cảm ứng và ức chế thông qua sự tương tác của các tác nhân môi trường (vai trò làm tín hiệu điều hòa) với prôtêin điều hòa.
DẠYKÈMQUYNHƠN
367
OFFICIAL
- Khác nhau:
Trong cơ chế điều hòa dương tính, prôtêin điều hòa có vai trò làm tăng sự biểu hiện của một hoặc một số gene cấu trúc. Còn trong điều hòa âm tính, prôtêin điều hòa có vai trò ức chế sự biểu hiện của gene cấu trúc
Trong cơ chế điều hòa dương tính sản sinh protein điều hòa liên kết với trình tự phần đầu của vùng P (promoter), còn trong điều hòa âm tính, protein điều hòa liên kết với vùng O (operater).
OFFICIAL
* Sự điều hòa hoạt động gene ở sinh vật nhân thực thể hiện khác nhau ở những giai đoạn phát triển khác nhau của cá thể, vì: Sinh vật nhân thực thường có cấu tạo cơ thể rất phức tạp, bao gồm các mô và các cơ quan chuyên hóa khác nhau phát sinh từ một tế bào duy nhất (hợp tử). Vì thế, sự điều hòa biểu hiện của nhiều gene vào những giai đoạn khác nhau cần nhiều cơ chế điều hòa tinh tế mới có thể đảm bảo cho cơ thể phát triển và sinh trưởng bình thường.
Trong sự phát sinh cá thể, tùy từng giai đoạn, tùy từng loại mô mà chỉ có một số gene trong tế bào hoạt động. Điều đó được diễn ra nhờ cơ chế điều hòa hoạt động gene.
Câu 7. Trong mô hình điều hòa hoạt động operon Lac, hãy nêu chức năng các trình tự ADN sau: (1) Trình tự chỉ huy (Regulator), (2) Trình tự khởi đầu phiên mã (Promoter), (3) Trình tự điều hoà (Operator), Gene cấu trúc Z (4), Y (5), A(6). Đột biến gene nếu xảy ra ở những trình tự này có thể gây ra những hậu quả gì?
Gợi ý trả lời:
* Chức năng các trình tự ADN trong mô hình điều hòa hoạt động operon Lac:
(1): kiểm soát quá trình phiên mã tổng hợp protein ức chế (repsessor).
(2): chứa các trình tự nu đặc hiệu thực hiện quá trình khởi động phiên mã các gene cấu trúc Z, Y, A của operon Lac.
(3): mang các trình tự đặc hiệu có ái lực cao với protein ức chế (repressor). Khi protein ức chế gắn vào vị trí này thì quá trình phiên mã của operon Lac không diễn ra.
(4): mã hoá cho enzym -galactosidase xúc tác cho phản ứng thủy phân lactôzơ thành glucôzơ và galactose, đồng thời enzim -galactoside còn có tác dụng chuyển hoá một phần lactôzơ thành đồng phân là alolactozo chất kích ứng protein ức chế
(5): mã hoá cho permease (protein vận chuyển) làm nhiệm vụ vận chuyển lactose vào tế bào.
DẠYKÈMQUYNHƠN
368
(6): mã hoá cho enzim thiogalactoside transacetylase chuyển nhóm acetyl từ acetyl-coA vào một số hợp chất gây độc với tế bào cũng đồng thời được vận chuyển bằng permease (chức năng giải độc tế bào). +Đột biến ở gene mã hóa protein ức chế có thể dẫn đến các hậu quả sau: Có thể xảy ra một đột biến câm, trong các trường hợp: đột biến nucleotit trong gene này không làm thay đổi trình tự axit amin trong protein ức chế, đột biến thay đổi axit amin trong chuỗi polypeptit của protein ức chế không làm thay đổi khả năng liên kết của protein ức chế với trình tự chỉ huy (O). Hậu quả cuối cùng của các dạng đột biến này là operon Lac hoạt động bình thường không có thay đổi gì liên quan đến sự biểu hiện của các gene cấu trúc Z và Y.
OFFICIAL
Xảy ra đột biến làm giảm khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gene cấu trúc Z và Y tăng lên. Làm mất hoàn toàn khả năng liên kết của protein ức chế hoặc protein ức chế không được tạo ra các gene cấu trúc Z và Y được biểu hiện liên tục. Xảy ra đột biến làm tăng khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gene cấu trúc Z và Y giảm đi (thực tế trường hợp này hiếm xảy ra).
Câu 8. Trong hoạt động của operon Lac ở vi khuẩn E. coli, nếu đột biến xảy ra ở gene điều hòa R (còn gọi là lac I) thì có thể dẫn đến những hậu quả gì liên quan đến sự biểu hiện của các gene cấu trúc? Gợi ý trả lời: -Operon Lac gồm các phần sau: gene điều hòa R mã hóa cho protein ức chế R, trình tự khởi động P, trình tự chỉ huy (O), các gen cấu trúc Z, Y và A -Nếu đột biến xảy ra ở gene R có thể dẫn đến các hậu quả sau: Xảy ra đột biến câm, trong các trường hợp: Đột biến nucleotit trong gen này không làm thay đổi trình tự axit amin trong prôtein ức chế, Đột biến thay đổi axit amin trong chuỗi polypeptit của prôtein ức chế không làm thay đổi khả năng liên kết của prôtein ức chế với trình tự chỉ huy (O). Hậu quả cuối cùng của các dạng đột biến này là ôperon Lac hoạt động bình thường không có thay đổi gì liên quan đến sự biểu hiện của các gen cấu trúc.
a) Xảy ra đột biến làm giảm khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gen cấu trúc tăng lên.
b) Làm mất hoàn toàn khả năng liên kết của protein ức chế hoặc protein ức chế không được tạo ra các gen cấu trúc biểu hiện liên tục.
DẠYKÈMQUYNHƠN
369
c) Xảy ra đột biến làm tăng khả năng liên kết của protein ức chế vào trình tự chỉ huy sự biểu hiện của các gen cấu trúc giảm đi.
* Kết luận: Đột biến xảy ra ở gene điều hòa R có thể dẫn đến những hậu quả khác nhau trong sự biểu hiện của các gen cấu trúc.
Câu 9. Bình thường người ta thấy loài vi sinh vật nọ không sản xuất ra enzim D, nhưng khi đưa thêm vào môi trường nuôi cấy của chúng một chất dinh dưỡng E thì sau 15 phút người ta thấy enzim D xuất hiện. Hãy giải thích hiện tượng trên về mặt cơ chế di truyền. Trả lời
* Dữ kiện đề bài cho ta nhận định sự điều hoà tổng hợp enzim D ở VSV nọ có thể xảy ra theo cơ chế điều hoà hoạt động gen ở vi khuẩn của F.Jacôp và J.mônô.
- Trong tế bào của VSV, các gen cấu trúc có liên quan về chức năng được phân bố liền nhau thành từng cụm có chung một cơ chế điều hoà được gọi là một opêron.
- 1 Opêrôn gồm các thành phần cơ bản sau:
+1 nhóm gen cấu trúc có liên quan về chức năng.
+ Vùng vận hành (O): là nơi protêin ức chế có thể liên kết làm ngăn cản sự phiên mã, khi không có prôtêin ức chế thì vùng vận hành hoạt động.
+ Vùng khởi động: Nơi ARN polimeraza bám vào và khởi đầu phiên mã.
Sự hoạt động của Opêrôn phụ thuộc váo sự điều khiển của gen điều hoà Opêrôn (R), gen điều hoà không nằm trong thành phần của Opêrôn mà nằm trước Opêrôn.
Bình thường gen điều hoà tổng hợp prôtêin ức chế liên kết với vùng vận hành ngăn cản quá trình phiên mã (không cho Opêrôn hoạt động).
* Trong đề bài thấy bình thường gen điều hoà phiên mã tạo mARN, tổng hợp prôtêin ức chế, chất ức chế bám vào vùng vận hành, do đó các gen cấu trúc không được phiên mã nên enzim D không được tổng hợp.
- Khi đưa thêm chất dinh dưỡng E vào môi trường nuôi cấy VSV nọ thì chính chất này với vai trò là chất cảm ứng gắn với prôtêin ức chế nên nó không gắn vàp vùng vận hành được nữa, vùng vậnh được tự do điều khiển quá trình phiên mã của Opêrôn, mARN của các gen cấu trúc được tổng hợp và quá trình dịch mã xảy ra enzim D được tổng hợp.
Câu 10. Operon là gì? Các thành phần cấu trúc của operon Lac có vai trò gì? Gợi ý trả lời:
DẠYKÈMQUYNHƠN
OFFICIAL 370
Operon là các gene cấu trúc ở prokaryote có liên quan về chức năng, chịu sự điều khiển chung của một cơ chế điều hoà được phân bố liền nhau thành từng cụm trên ADN. Thông thường, đó là các gene cùng tham gia vào một con đường chuyển hóa.
Một operon là một đơn vị phiên mã đơn bao gồm một chuỗi các gene cấu trúc, một promoter và một operator. Các gene cấu trúc của một operon hoặc là cùng được phiên mã hoặc cùng không được phiên mã vì chúng đều được điều khiển bởi một operator (O) và một promoter (P) duy nhất.
Các thành phần cấu trúc operon Lac có vai trò:
- Một nhóm gen cấu trúc liên quan về chức năng gồm 3 gene cấu trúc (Z, Y, A) kiểm soát tổng hợp 3 protein Z, Y, A, là các enzym tham gia các phản ứng phân giải đường lactozơ:
OFFICIAL
+ Gene Lac Z quy định tổng hợp enzim β- galactozidase có chức năng phân giải đường lactose thành glucose và galactose, đồng thời còn có vai trò chuyển hóa một phần lactose (liên kết β 1,4D glycoside ) thành một đồng phân allolactose (liên kết β 1,6D glycoside) là phân tử bất hoạt protein ức chế.
+ Gene Lac Y quy định một loại protein permeaza nằm xuyên màng tế bào vi khuẩn làm nhiệm vụ vận chuyển chủ động đường lactose vào trong tế bào vi khuẩn.
+ Gene A quy định tổng hợp enzyme thiogalactoside transacetylase có vai trò giải độc tế bào đối với các hợp chất thiogalactoside cũng được đồng vận chuyển vào tế bào khi permease hoạt động và chuyển nhóm acetyl từ acetyl-coA tới β- galactozidase.
Tất cả 3 gene cấu trúc này đều được phiên mã cùng một lúc tạo thành một phân tử mARN.
- Vùng vận hành (O) là trình tự nuclêôtit đặc biệt để protein ức chế dị hình liên kết ngăn cản phiên mã.
- Vùng khởi động (P) nằm gần đầu 5, của gene Lac Z, là nơi ARN polimezaza bám vào khởi đầu phiên mã đồng thời của ba gene Lac Z, Lac Y, Lac A
Câu 11. So sánh cơ chế điều hòa âm tính và điều hòa dương tính ở opêron Lac.
Trả lời
* Giống nhau: - Đều để thích ứng với các điều kiện môi trường biến động, đồng thời để tiết kiệm năng lượng và vật chất của tế bào. - Đều liên quan đến sự tham gia của các gen điều hòa. Các gen này mã hóa cho các sản phẩm trực tiếp (prôtêin điều hòa) điều hòa sự biểu hiện của các
DẠYKÈMQUYNHƠN
371
gen cấu trúc. Đều có hệ thống điều hòa cảm ứng và ức chế thông qua sự tương tác của các tác nhân môi trường (vai trò làm tín hiệu điều hòa) với prôtêin điều hòa.
* Khác nhau:
- Trong cơ chế điều hòa dương tính, prôtêin điều hòa có vai trò làm tăng sự biểu hiện của một hoặc một số gen cấu trúc. Còn trong điều hòa âm tính, prôtêin điều hòa có vai trò ức chế sự biểu hiện của gen cấu trúc.
- Trong cơ chế điều hòa dương tính sản sinh prôtêin điều hòa liên kết với trình tự phần đầu của vùng P (promoter), còn trong điều hòa âm tính, prôtêin điều hòa liênkết vớivùngO (operater).
Câu 12. So sánh hoạt động của operon lac (lactozơ) và operon trp (tryptophan) trong điều hoà âm tính ở E.coli.
Trả lời Giống nhau:
OFFICIAL
- Sự điều hoà của cả hai operon lac và trp đều liên quan đến cơ chế điều hoà các gen kiểu âm tính: Nghĩa là, các operon này đều được “tắt” bởi prôtêin điều hoà tương ứng của chúng (đều là các prôtêin ức chế do gen điều hoà tổng hợp).
- Sự điều hoà của cả hai operon lac và trp đều tạo cho tế bào tiết kiệm năng lượng và vật chất trong hoạt động sống của nó.
Khác nhau: operon Lac operon ức chế luôn tắt, chỉ mở khi có chất cảm ứng Luôn hoạt động, chỉ tắt khi có chất ức chế.
pr ức chế khi mới hình thành đã ở dạng hoạt động ngay, liên kết vào O và ức chế phiên mã
Enzim cảm ứng thường hoạt động trong con đường dị hóa
Pr ức chế khi mới tổng hợp ở dạng bất hoạt, chỉ được hoạt hóa khi có chất đồng ức chế.
Enzim ức chế thường hoạt động trong con đường đồng hóa.
- Trong operon lac, các enzim tham gia vào con đường chuyển hoá lactozơ còn gọi là các enzim cảm ứng do quá trình sinh tổng hợp chúng được gây cảm ứng bởi tín hiệu hoá học (trong trường hợp này là allolactozơ). Theo nguyên tắc tương tự, trong operon trp các enzim do operon trp mã hoá được gọi là các enzim ức chế
DẠYKÈMQUYNHƠN
372
- Trong operon trp, khi tryptophan có sẵn trong môi trường hoặc khi lượng tích luỹ trong tế bào của chúng đã đủ thì chính axit amin này kết hợp với prôtêin điều hoà tạo thành phức hợp đồng ức chế liên kết vào trình tự O (operator) làm dừng quá trình phiên mã. Ngược lại trong open lac, allolactose làm bất hoạt prôtêin điều hoà làm cho prôtêin này không liên kết được vào trình tự O, nhờ đó quá trình phiên mã diễn ra.
Câu 13. Hình thức tổ chức gen theo kiểu Operon đem lại lợi ích gì cho SV? Cho ví dụ.
Trả lời
Ý nghĩa của việc tổ chức gen theo kiểu Operon:
- Tiết kiệm vật chất di truyền, làm cấu trúc hệ gen gọn nhẹ (cách tổ chức của nhiều gen trong TB nhân sơ).
- Tạo ra sản phẩm các gen nhanh chóng, các sản phẩm gen này thường liên quan đến nhau về chức năng nên cùng lúc tế bào cần lượng tương đương. VD: Operon Lac ở E.coli tạo ra các protein chuyển hóa lactozo, các gen rARN (ở cả nhân sơ và nhân thực) được tạo ra lượng lớn, đồng thời mới có thể đáp ứng được yêu cầu củaTB.
OFFICIAL
DẠYKÈMQUYNHƠN
373
CHƯƠNG III. CƠ SỞ SINH HÓA CỦA TƯƠNG TÁC GEN
3.1. Cơ sở sinh hóa của tương tác giữa các gen alen
Gen alen là các trạng thái khác nhau của cùng một gen phân bố trong cùng một locus gen. Alen này chỉ khác alen kia ở một hoặc một vài cặp nucleotit và là sản phẩm của quá trình đột biến gen.
Tương tác giữa các gen alen là sự tác động qua lại giữa các alen của cùng một gen để biểu hiện ra kiểu hình theo các dạng:
➢ Trội lặn hoàn toàn.
➢ Trội lặn không hoàn toàn
➢ Đồng trội.
➢ Siêu trội.
➢ Gen gây chết (Alen gây chết).
➢ Gen chịu sự di truyền hiệu quả dòng mẹ (sự DT biểu hiện chậm của Menđen)
➢ Gen biểu hiện chịu ảnh hưởng của giới tính
➢ Gen chịu sự di truyền hiện tượng in vết của gen
3.1.1. Tương tác kiểu trội lặn hoàn toàn
* Khái niệm
Tương tác trội lặn hoàn toàn là hiện tượng trong đó một alen lấn át hoàn toàn sự biểu hiện của một alen khác thuộc cùng một locus. Trong trường hợp này, kiểu hình của thể dị hợp tử hoàn toàn giống kiểu hình của thể đồng hợp trội.
*Cơ sở sinh hóa của tương tác trội lặn hoàn toàn
Ta xét một gen gồm 2 alen A và a, trong đó alen A là trội hoàn toàn so với alen a.
Alen trội A (thường là alen kiểu dại) mã hóa một enzym, alen lặn a (thường là alen đột biến) có thể sinh ra một Protein không có hoặc có rất ít hoạt tính enzym (đột biến nhầm nghĩa) hoặc alen a không sinh ra Protein hoặc chỉ sinh ra một đoạn (đột biến mất nghĩa). Các cá thể dị hợp Aa chỉ sinh ra khoảng ½ số lượng enzim có hoạt tính so với các cá thể đồng hợp trội AA. Nếu số lượng đó là đủ cho tế bào hoặc cơ thể thực hiện các chức năng hóa sinh một cách bình thường thì sẽ xuất hiện kiểu hình bình thường.
Trong trường hợp alen A là alen đột biến trội hơn so với alen a là alen kiểu dại thì
DẠYKÈMQUYNHƠN
374
OFFICIAL
có thể giải thích do enzym của alen đột biến sinh ra ái lực lớn hơn với cơ chất so với enzym của alen kiểu dại. Tuy vậy, enzym đột biến thường không có khả năng xúc tác phản ứng hoặc xúc tác với hiệu quả thấp gây ra hiệu ứng đa số đột biến gen là có hại vì mất cân bằng hài hòa giữa kiểu gen và môi trường trong quá trình chọn lọc tự nhiên *
Giả thuyết 1:
* Giả thuyết 2: Hình 3.1.Các giả thuyết về cơ sở của tính trội hoàn toàn. 3.1.2. Tương tác trội lặn không hoàn toàn
* Khái niệm Tương tác trội lặn không hoàn toàn là hiện tượng trong đó một alen trội không hoàn toàn lấn át sự biểu hiện của alen lặn khác thuộc cùng một locus. Kiểu hình của thể dị hợp tử biểu hiện hiểu hình trung gian giữa kiểu hình của hai thể đồng hợp.
* Cơ sở sinh hóa của tương tác trội lặn không hoàn toàn
Có thể giải thích hiện tượng trội không hoàn toàn là do sự có mặt của alen a đã kìm hãm quá trình phiên mã, dịch mã của alen A. Kết quả là sắc tố đỏ hình thành ít hơn mức bình thường khiến cho cây mang kiểu gen Aa cho hoa màu hồng.
Bởi kiểu hình của thể dị hợp là trung gian giữa hai thể đồng hợp, vì vậy ta có thể lý giải trên phương diện sinh hóa rằng hàm lượng sản phẩm tích lũy do một allele trội kiểm soát là không đủ để thể hiện kiểu hình màu đỏ như trong
DẠYKÈMQUYNHƠN
375
OFFICIAL
3.1.3. Tương tác đồng trội * Khái niệm Tương tác đồng trội là hiện tượng cả hai alen trong cơ thể dị hợp cùng hình thành những tính trạng mà nó kiểm soát một cách độc lập với alen cùng cặp. * Cơ sở sinh hóa của tương tác đồng trội Hiện tượng đồng trội được giải thích trên cơ sở tính đa hình cân bằng của các sản phẩm Protein được sinh ra từ các alen đồng trội. Trong tự nhiên sự đa hình cân bằng sẽ
trường hợp có mặt cả hai alen trội.
không có sự thay thế hoàn toàn một alen này bằng một alen khác, mà là sự duy trì ưu thế của các thể dị hợp về một hoặc một số cặp alen nào đó.
3.1.4. Gen gây chết (Alen gây chết)
* Khái niệm Tương tác gen gây chết là hiện tượng khi sự có mặt của alen đó trong kiểu gen sẽ ảnh hưởng không thuận lợi đến sự sinh trưởng và phát triển của sinh vật, do vậy dẫn đến làm giảm sức sống hay gây chết cho cơ thể mang nó.
Có hai dạng tương tác gen gây chết: gây chết trong trạng thái đổng hợp trội và gây chết trong trạng thái đồng hợp lặn.
Các gen theo hiệu quả gây chết thường chia làm 3 nhóm:
➢ Gen gây chết hoàn toàn: là gen làm chết hoàn toàn các cá thể đồng hợp mang nó.
OFFICIAL
➢ Gen nửa gây chết: là gen làm chết nhiều hơn 50% nhưng ít hơn 100% số thể đồng hợp mang nó.
➢ Gen giảm sống: là gen làm chết dưới 50% số thể đồng hợp mang nó. Tuy nhiên, sự phân chia này cũng chỉ có tính chất quy ước vì một gen có thể gây chết hoàn toàn trong điều kiện này nhưng lai lại là gen nửa gây chết trong điều kiện ngoại cảnh khác.
* Cơ sở sinh hóa của tương tác alen gây chết
Hiện tượng tương tác alen gây chết có thể giải thích do alen quy định tổng hợp Protein từ đó quy định tổng hợp những enzyme tham gia quá trình chuyển hóa quyết định sự sống còn của sinh vật.
3.1.5. Di truyền của gen có nhiều alen
Trên thực tế, mỗi một gen không chỉ có hai alen mà có thể có nhiều hơn hai alen, gọi là đa alen. Các alen là những trạng thái cấu trúc khác nhau của cùng một gen phát sinh do đột biến.
Hiện tượng đa alen tồn tại phổ biến trong các quần thể sinh vật khác nhau. Ví dụ, gen kiểm soát màu mắt đỏ-trắng ở ruồi giấm gồm một chuỗi 12 alen, với tính trội giảm dần từ đỏ kiểu dại cho đến trắng đột biến lặn (w) theo thứ tự từ trái sang phải và trên xuống như sau:
DẠYKÈMQUYNHƠN
376
Màu Alen We Wbl Wap W
Wt Màu
W+ Wsat Wco Ww Wap3 Wch đỏ
Alen
i
eossin blood apricot ivory tinged white Một số gen ở người, chẳng hạn như các gen đối với kháng nguyên bạch cầu người HLA (human leukocyte antigen) xác định các kháng nguyên trên bề
dại satsu ma cor al win e aprico t3 cherr y
mặt của hầu như tất cả các tế bào có thể có nhiều alen. Ví dụ, gen HLA-B có nhiều hơn 30 alen được xác định khác nhau về mặt kháng nguyên trong một số quần thể. Kết quả của sự đa dạng này là, trong một quần thể có rất nhiều kiểu gen ở gen HLA-B (465 kiểu gen khác nhau, với 30 kiểu đồng hợp và 435 kiểu dị hợp), thực sự tạo nên một dãy biến dị kiểu gen rộng.
DẠYKÈMQUYNHƠN OFFICIAL
377
3.1.6. Tương tác siêu trội Hiện tượng siêu trội là hiện tượng thể dị hợp của các alen luôn có sức sống, sức sinh trưởng, sinh sản tốt hơn thể đồng hợp, kể cả thể đồng hợp trội (AA < Aa > aa).
Có thể giải thích là do sự tương tác giữa hai sản phẩm Protein của 2 alen khác nhau về chức phận trong cùng một locus dẫn đến hiệu quả bổ trợ, làm tăng tác động của alen trội.
Tương tác siêu trội gây hiệu ứng ưu thế lai trong lai khác dòng dẫn đến trong quần thể giao phối đa số các kiểu gen ở trạng thái dị hợp.
3.1.7. Gen chịu sự di truyền hiệu quả dòng mẹ (sự DT biểu hiện chậm của
Menđen)
Sản phẩm do gen ở trong nhân tạo ra (trước khi thụ tinh) tồn tại ở tế bào chất của tế bào trứng tác động đến sự biểu hiện kiểu hình ở đời sau. Hiện tượng này gọi là "tiền định tế bào chất" hay hiệu ứng dòng mẹ (còn gọi là hiện tượng di truyền Men Đen thể hiện chậm đi 1 thế hệ). Ví dụ về hiệu quả dòng mẹ: chiều xoắn của vỏ ốc Limnaea peregra. Chiều xoắn này được xác định bởi một cặp alen. D là xoắn phải, d là xoắn trái. Các phép lai cho thấy D là trội so với d và chiều xoắn
OFFICIAL
n rất sớm, chúng cũng đã nhận được tế bào chất, các chất dinh dưỡng trong trứng từ mẹ và những ảnh hưởng đặc biệt lên hoạt động của gene. Những tiềm năng nhất định của trứng được xác định trước khi thụ tinh và trong một số trường hợp chúng đã chịu ảnh hưởng của môi trường mẹ bao quanh. Sự quyết định trước như vậy do các gene của mẹ hơn là các gene của con, được
DẠYKÈMQUYNHƠN
378
Hình 3.2.Hiệu quả dòng mẹ lên chiều xoắn vỏ ốc Tiến hành phép lai thuận nghịch giữa đồng hợp tử DD xoắn phải và đồng hợp tử dd xoắn trái: Cơ sở sinh hóa gen chịu sự di truyền hiệu quả dòng mẹ Trứng và phôi chịu ảnh hưởng của môi trường cơ thể mẹ nhiều hơn cơ thể bố. Ngay cả khi bị tách khỏi cơ thể mẹ từ một giai đoạ
luôn luôn được xác định bởi kiểu gene của mẹ.
gọi là hiệu ứng mẹ. Sự tồn tại của hiệu ứng mẹ nói chung được chứng minh bằng phương pháp lai thuận nghịch, khi đó kết quả phép lai thuận nghịch sẽ khác nhau. Như vậy hiệu quả dòng mẹ chỉ kéo dài một thế hệ và trường hợp này không thể xem là di truyền qua tế bào chất bởi vì ở đây các đặc tính của tế bào chất đã được xác định trước bởi tác dụng của các gene trong nhân chứ không phải bởi các gene trong tế bào chất. Nói cách khác ở đây cơ chế di truyền nhiễm sắc thể làm biến đổi tế bào chất của trứng trước khi nó thụ tinh.
3.1.8. Gen biểu hiện chịu ảnh hưởng của giới tính
Trong trường hợp tính trạng trội phụ thuộc vào giới tính thì tỉ lệ kiểu hình của 1 phép lai được tính riêng ở từng giới. Tỉ lệ kiểu hình phân li chung bằng trung bình cộng tỉ lệ kiểu hình ở 2 giới.
3.1.9. Gen chịu sự di truyền hiện tượng in vết của gen
Trong toàn bộ quá trình phân tích về di truyền học Menđen và cơ sở nhiễm sắc thể của sự di truyền, một gen nào đó sẽ cho hiệu quả như nhau bất luận được di truyền từ bố hay từ mẹ. Tuy nhiên, trong một vài trường hợp, ở động vật có vú nhiều tính trạng phụ thuộc vào việc gen biểu hiện được di truyền từ bố hay mẹ. Đây chính là khái niệm về di truyền in vết. Những biểu hiện của kiểu hình phụ thuộc vào việc allen quy định đặc điểm được truyền từ bố hay mẹ được gọi là in vết hệ gen (di truyền in vết). Hiện tượng này khác biệt hoàn toàn với hiện tượng gen liên kết với giới tính, ở đây các gen in vết nằm trên nhiễm sắc thể thường. * Cơ chế:
In vết hệ gen xảy ra trong quá trình hình thành giao tử và kết quả làm bất hoạt một allen của một gen nhất định. Bởi vì các gen này được in vết khác nhau ở tinh trùng và trứng nên hợp tử chỉ biểu hiện một allen của một gen được in vết (allen đó có thể được di truyền từ bố hoặc di truyền từ mẹ). Sự in vết được truyền và biểu hiện ở tất cả các tế bào cơ thể trong qúa trình phát triển. Trong mỗi thế hệ, sự in vết cũ sẽ được “xóa” đi trong các tế bào sản sinh giao tử và in vết mới được thiết lập trên các nhiễm sắc thể của giao tử đang được hình thành dựa theo giới tính của các cá thể tạo ra giao tử. Trong cùng loài, các gen in vết luôn luôn in vết theo cùng một cách.
Cơ chế liên quan đến in vết gen là sự methyl hóa ADN, gắn nhóm methyl (CH3) vào các nucleotit cytosine của một trong các allen, do đó làm bất hoạt allen, allen này không được biểu hiện trong quá trình phát triển của phôi. Tuy nhiên, đối với một số gen, sự methyl hóa lại làm hoạt hóa sự biểu hiện của allen. Một ví dụ, đó là trường hợp của gen Igf2: Methyl hóa một số cytosine nhất định trên nhiễm sắc thể của bố dẫn đến sự biểu hiện của allen Igf2. * Một số rối loạn liên quan đến in vết hệ gen, bao gồm: hội chứng Angelman, hội chứng Prader-Willi và hội chứng Beckwith-Wiedemann.
DẠYKÈMQUYNHƠN
OFFICIAL 379
Hội chứng Angelman là một rối loạn của hệ thần kinh, triệu chứng đặc trưng như co giật, khiếm khuyết về ngôn ngữ, hành động kì lạ. Cơ chế gây ra hội chứng được nghiên cứu trên khoảng 70% số bệnh nhân Angelmen là do một phần của NST số 15 bị xóa (khoảng 4 triệu cặp nucleotit). Trong đó gen đột biến di truyền từ mẹ không được biểu hiện, gen di truyền từ cha được biểu hiện. Bằng một cách khác, phần bị xóa trên cùng NST số 15, gen đột biến di truyền từ cha không được biểu hiện, gen di truyền từ mẹ được biểu hiện có liên quan đến hội chứng Prader-Willi. Một bệnh khác là hội chứng Beckwith-Wiedemann (BWS), được đặc trưng bởi kích thước sinh lớn bất thường và khuynh hướng dẫn đến khối u. BWS được gây ra bởi sự mất dấu ấn và di truyền tại các vị trí cụ thể trên nhiễm sắc thể 11.
3.2. Cơ sở sinh hóa tương tác giữa các gen không alen
OFFICIAL
Tương tác gen là sự tác động qua lại của các gen trong quá trình hỉnh thành kiểu hình. Về thực chất, các gen không trực tiếp tương tác với nhau mà chỉ các sản phẩm của chúng như các enzym (bản chất protein) hay giữa các sản phẩm sau hoạt động của enzym phối hợp gây ảnh hưởng qua lại theo những cách khác nhau dẫn đến sự biểu hiện kiểu hình khác nhau.
Sản phẩm enzym của các gen không alen tương tác với nhau:
- Để làm xuất hiện một kiểu hình mới trong trường hợp tương tác bổ trợ.
- Enzym do gen này ức chế hoạt động của enzym được tạo ra do gen khác không alen trong trường hợp tương tác át chế.
- Cùng làm tăng cường biểu hiện của một tính trạng trong trường hợp tương tác cộng gộp
Dưới dây là các dạng tương tác gen không alen
Tương tác đa gen giữa các gen không alen là sự tác động qua lại giữa 2 hay nhiều gen không alen để cùng quy định một tính trạng theo các dạng:
Tương tác bổ sung (bổ trợ).
Tương tác át chế.
Tương tác cộng gộp.
3.2.1. Tương tác bổ sung
Tương tác bổ sung (tương tác bổ trợ) là kiểu tương tác trong đó các alen của mỗi locus riêng rẽ có biểu hiện kiểu hình riêng. Khi hai hoặc nhiều gen không alen cùng có mặt trong kiểu gen sẽ tạo nên một kiểu hình mới.
Hiện tượng tương tác bổ trợ biểu hiện ra nhiều tỷ lệ kiểu hình F2 khác nhau tùy thuộc vào sự tương tác đặc biệt giữa các gen không alen.
DẠYKÈMQUYNHƠN
380
3.2.1.1. Tỷ lệ 9 : 3 : 3 : 1
➢ Ở gà, hình dạng khác nhau của mào là kết quả tương tác giữa 2 gen không alen quy định. Trong đó: A–B– : gà mào hình quả hồ đào A–bb : gà mào hình hoa hồng aaB– : gà mào hình hạt đậu aabb : gà mào hình lá
Cơ sở sinh hóa Như vậy, đây là kiểu tương tác giữa hai gen không alen. Trong đó, sự tương tác giữa hai gen trội A và B quy định một kiểu hình riêng. Ở đây, không có sự biến dạng của tỉ lệ phân ly kiểu hình 9 : 3 : 3 : 1.
Cho đến nay, cơ sở phân tử trong việc hình thành 4 hình dạng mào này vẫn chưa được rõ ràng nhưng nhìn một cách tổng quát nhất ta có thể đưa ra giả thuyết như sau.
Kiểu gen aabb sinh ra được sản phẩm gen nhưng sản phẩm gen đó không thể ảnh hưởng tới việc thay đổi hình dạng căn bản của kiểu hình mào gà hình lá.
Alen trội A sinh ra sản phẩm, sản phẩm này sẽ ảnh hưởng tác động qua lại với sản phẩm của gen quy định hình dạng mào gà hình lá để quy định mào hình hoa hồng.
Tương tự, alen trội B sinh ra sản phẩm, sản phẩm này sẽ ảnh hưởng tác động qua lại với sản phẩm của gen quy định hình dạng mào gà hình lá để quy định mào hình hạt đậu.
Khi sản phẩm của cả hai alen trội A và B tương tác với nhau sẽ xuất hiện dạng kiểu hình mới khác với các dạng kiểu hình còn lại là mào hình quả hồ đào.
3.2.1.2. Tỷ lệ 9 : 6 : 1
Ở loài bí Cucubirta peppo, có ba dạng quả đã được nghiên cứu là dạng quả tròn, quả dẹt và quả dài Đã xác định hình dạng quả là kết quả tương tác giữa hai gen không alen quy định. Ở bí, hình dạng quả là kết quả tương tác giữa 2 gen không alen. Trong đó:
A–B– : bí quả dẹt A–bb; aaB– : bí quả tròn aabb : bí quả dài ➢ Cơ sở sinh hóa Bí quả dài và bí quả tròn là 2 dạng điển hình trong rất nhiều dạng của bí mùa hè (summer squash), hình dạng của quả bí được quy định bởi hai cặp gen không alen. Điểm đáng chú ý ở đây đó là khi cả hai alen trội không len tương tác với nhau thì ta thu được kết quả vô cùng lý thú: hình dạng mới của quả bí sẽ xuất hiện.
DẠYKÈMQUYNHƠN
381
OFFICIAL
Cơ sở phân tử chính xác về hiện tượng trên cho đến nay vẫn còn là một ẩn số chưa thật sự rõ ràng nhưng giả thuyết sau đây đưa ra đã được phần lớn các nhà khoa học chấp nhận. Ở bí quả dài có kiểu gen aabb, do sự vắng mặt các sản phẩm của 2 alen A và B nên sản phẩm được tạo ra từ kiểu gen aabb sẽ cho kiểu hình là bí quả dài. Nếu như thiếu đi một trong hai sản phẩm của hai alen A, B mà không phải cả hai thì hình dạng căn bản của bí quả dài sẽ biến mất, thay vào đó là hình dạng bí quả tròn.
Hình dạng bí quả dẹt có thể được xuất hiện thông qua sự biến đổi của hình dạng bí quả tròn do sự tương tác của cả 2 sản phẩm của 2 alen A và B. 3.2.1.3. Tỷ lệ 9 : 7 - VD1: Ở ngô, màu sắc hạt là kết quả của sự tương tác giữa hai gen không alen. Trong đó:
A–B–: ngô hạt vàng
A–bb; aaB–; aabb: ngô hạt tím
➢ Ở cây hoa đậu thơm, màu sắc hoa là kết quả của sự tương tác giữa hai gen không alen. Trong đó:
A–B–: cây đậu thơm hoa tía. A–bb; aaB–; aabb: cây đậu thơm hoa trắng.
➢ Cơ sở sinh hóa
Kết quả trên được giải thích bởi sự tương tác của hai gen không alen theo 2 cách: Cách 1: Tỉ lệ kiểu hình 9/16 cây đậu thơm hoa tía thu được ở F2 nói lên rằng màu sắc ở hoa chỉ xuất hiện khi 2 gen trội không alen tương tác với nhau. Tuy vậy, các quan niệm trước đây chỉ mường tượng rằng sự có mặt của cả hai alen trội trong kiểu gen sẽ quy định hoa tía, còn nếu kiểu gen vắng mặt 1 trong hai alen trội hoặc vắng mặt cả hai thì sẽ quy định hoa màu trắng.
Ngày nay, cơ sở phân tử của hiện tượng trên đã được giải thích rõ ràng theo sơ đồ 1.1.
Theo cách giải thích này thì sắc tố tía được tạo ra là nhờ hai yếu tố là tiền chất do gen A tạo ra và enzim do gen B tạo ra xúc tác phản ứng biến tiền chất A thành sắc tố tía. Theo sơ đồ thì mỗi bước được điều khiển bởi sản phẩm của mỗi gen, có nghĩa gen A điều khiển sự biến đổi từ dạng tiền thân không màu 1 sang dạng tiền thân không màu 2, gen B điều khiển sự biến đổi từ dạng tiền thân không màu 2 sang sản phẩm P cuối cùng của chuỗi phản ứng sinh hóa là sắc tố tía. Vì vậy, các kiểu gen A–B– đủ hai yếu tố nên sắc tố tía được tổng hợp. Các kiểu gen A–bb; aaB– thiếu một yếu tố và kiểu gen aabb thiếu cả hai yếu tố nên sắc tố tía không được tổng hợp.
DẠYKÈMQUYNHƠN
OFFICIAL 382
Cách 2: Kết quả trên còn có thể được giải thích do hiện tượng đồng át chế lặn xảy ra tức aa > B, b đồng thời bb > A, a. Quan niệm này cũng được rất nhiều người đồng tình. Gen A Gen B ↓ ↓ Enzym A Enzym B ↓ ↓
Dạng tiền thân không màu 1 →Dạng tiền thân không màu 2 → Sản phẩm P (sắc tố tía)
3.2.2. Tương tác át chế Tương tác át chế là kiểu tương tác giữa hai hay nhiều gen không alen cùng tham gia quy định một tính trạng. Trong đó, một gen trội (hoặc gen lặn) lấn át sự biểu hiện kiểu hình của gen thuộc locus khác không alen.
Kiểu tương tác này cũng làm biến đổi tỷ lệ phân ly kiểu hình ở F2. Át chế trội xảy ra khi B > C (hoặc ngược lại C > B) và át chế lặn xảy ra khi bb > cc (hoặc cc > bb).
3.2.2.1. Tỷ lệ 12 : 3 : 1 (Át chế trội)
➢ Ở bí mùa hè (Summer squash), sự hình thành màu sắc quả bí là do 2 cặp gen không alen quy định. Trong đó:
B–C–; B–cc: bí trắng bbC–: bí vàng bbcc: bí xanh
➢ Tiến hành phép lai ta thu được kết quả sau:
Pt/c: Bí trắng × Bí xanh
F1: 100% bí trắng. Cho các cơ thể con lai thụ phấn.
F2: 12 bí trắng : 3 bí vàng : 1 bí xanh
➢ Cơ sở sinh hóa Để giải thích kết quả của phép lai trên ta quan sát vào sơ đồ 1.3. Alen trội C là alen cần thiết cho sự chuyển đổi màu từ bản chất xanh sang vàng của quả bí. Còn alen trội B có khả năng sản sinh ra chất ức chế, chất này sẽ ức chế quá trình chuyển đổi màu sắc từ trắng sang xanh.
Sơ đồ (a) sơ đồ1.3 mô tả con đường hình thành trái bí trắng trên cơ sở kiểu gen B–C–; B–cc. Theo sơ đồ này thì alen B sản sinh ra chất ức chế quá trình chuyển đổi màu từ trắng sang xanh, vì thế bước tiếp theo chuyển đổi màu từ bản chất xanh sang vàng sẽ không thực hiện được nên các cơ thể mang kiểu gen B–C–; B–cc đều có kiểu hình là bí trắng. Sơ đồ (b) sơ đồ 1.3 mô tả con đường hình thành trái bí vàng trên cơ sở kiểu gen bbC–. Do không có mặt alen B nên chất ức chế không được tạo thành, mặt khác lại có
DẠYKÈMQUYNHƠN
383
OFFICIAL
mặt alen C trong kiểu gen nên các cơ thể mang kiểu gen bbC– đều có kiểu hình bí vàng.
Sơ đồ (c) sơ đồ 1.3 mô tả con đường hình thành trái bí xanh trên cơ sở kiểu vắng mặt alen C trong kiểu gen nên các cơ thể mang kiểu gen bbcc đều có kiểu hình là bí xanh.
Con đường hình thành trái bí màu trắng
Gen B Gen C ↓ ↓ Chất ức chế (B) Sản phẩm gen C ↓ ↓
Sắc tố trắng → Sắc tố xanh → Sắc tố vàng ↓
Trái bí trắng (B-C-; B-cc) Con đường hình thành trái bí màu vàng Gen C ↓ Không chất ức chế Sản phẩm gen C ↓ ↓ Sắc tố trắng → Sắc tố xanh → Sắc tố vàng → Trái bí vàng Con đường hình thành trái bí xanh Không chất ức chế
DẠYKÈMQUYNHƠN
384
OFFICIAL
3 (Át chế trội) ➢ Ở gà, sự hình thành màu sắc lông là do hai cặp gen không
Trong đó: B–C–; bbC–; bbcc: gà lông trắng B–cc: gà lông đen ➢ Cơ sở sinh hóa Kết quả của phép lai trên được giải thích do
tác át chế giữ
alen B và C, cụ thể là tương
át chế trộ
CC >
. Alen B là alen quy định việc tạo
ộ
↓ Sắc tố trắng → Sắc tố xanh → Trái bí xanh (bbcc) 3.2.2.2. Tỷ lệ 13 :
alen quy định.
tương
a hai gen không
tác
i:
B, b
màu cho b
lông gà.
Sản phẩm của alen C ức chế sự biểu hiện ra kiểu hình của các sản phẩm được sinh ra từ alen B và alen b. Do vậy, các cơ thể có kiểu gen B–C–; bbC– sẽ đều có kiểu hình là gà lông trắng.
Các cơ thể mang kiểu gen bbcc cho kiểu hình gà lông trắng là do gen c tạo màu ở trạng thái đồng hợp lặn.
Các cơ thể mang kiểu gen B–cc, do sản phẩm tạo màu lông của alen B không bị ức chế nên chúng mang kiểu hình là gà lông đen.
Ví dụ: Màu cánh hoa xanh da trời (blue) của hoa báo xuân di truyền theo mô hình "13: 3" . Màu xanh lơ do sắc tố man-vi-đin (malvidine) tạo ra. Sắc tố này là an-tôxi-a-nin (anthocyanine) đã mê-tyl hóa, được hình thành nhờ xúc tác của một enzym được mã hóa bởi gen trội A, còn alen lặn a không tạo được enzym. Nhưng sự sản xuất sắc tố này bị ngăn cản bởi gen trội B ở lôcut khác. Nên phép lai AaBb × AaBb sinh thế hệ con phân li trung bình là 13/16 trắng (A-B- + aa- -) + 3 xanh (A-bb).
OFFICIAL
3.2.2.3. Tỷ lệ 9 : 4 : 3 (Át chế lặn)

- Ở chuột, màu sắc lông là do sự tương tác giữa hai gen không alen quy định. Trong đó:
B–C–: chuột xám nâu (sợi lông có hai đầu mút màu đen, đoạn giữa màu vàng), còn gọi là chuột agouti.
B–cc; bbcc : chuột bạch tạng, bbC–: chuột đen ➢ Cơ sở sinh hóa
Kết quả của phép lai trên được giải thích thông qua tương tác át chế giữa hai gen không alen B và C, cụ thể là tương tác át chế lặn: cc > B, b.
Alen C quy định việc hình thành sắc tố đen của sợi lông, alen B quy định việc hình thành sắc tố vàng trên sợi lông.
Sản phẩm của alen c ức chế sự biểu hiện ra kiểu hình của các sản phẩm được sinh ra từ alen B và alen b. Do vậy, các cơ thể có kiểu gen B– cc; bbcc sẽ đều có kiểu hình là chuột bạch tạng. Các cơ thể mang kiểu gen bbC– có alen C quy định việc hình thành sắc tố đen trên sợi lông nên chúng đều có kiểu hình là chuột đen. Mặt khác, các cơ thể mang kiểu gen B–C– có đầy đủ alen B và C nên chúng có kiểu hình là chuột xám nâu.
DẠYKÈMQUYNHƠN
385
Ngoài tương tác át chế lặn, 2 gen không alen B, C còn tương tác theo kiểubổ trợ do F1 xuất hiện kiểu hình mới là chuột xám nâu (agouti).


Tỷ lệ phân ly 9 : 3 : 4 ta cũng gặp ở ngựa thể hiện qua sơ đồ lai sau:
OFFICIAL
Pt/c: BBCC (ngựa đen) × bbcc (ngựa trắng) F1: BbCc (100% ngựa đen) BbCc (ngựa đen) × BbCc (ngựa đen)

F2: 9 ngựa đen (B-C-) : 3 ngựa nâu (bbC-) : 4 ngựa trắng (B-cc; bbcc)
Tuy nhiên kết quả của phép lai trên lại được giải thích theo một cách hoàn toàn khác so với tỷ lệ 9 : 4 : 3 thu được trong ví dụ chuột xám nâu lai với nhau: Tính trạng màu sắc lông ở ngựa do 2 gen không alen B, C tương tác qua lại theo kiểu át chế quy định. Trong đó, gen C quy định việc tổng hợp enzyme C, enzyme này có vai trò xúc tác cho quá trình sinh hóa biến đổi màu sắc lông từ dạng không màu sang màu nâu, gen B quy định việc tổng hợp enzyme B xác tác cho quá trình sinh hóa biến đổi màu sắc lông từ màu nâu sang màu đen theo sơ đồ
DẠYKÈMQUYNHƠN
386
Gen C Gen B ↓ ↓ Enzyme C Enzyme B ↓ ↓ Lông trắng → Lông nâu → Lông đen
sau:
Do vậy, các cơ thể có kiểu gen B–C– sẽ cho kiểu hình lông đen. Các cơ thể có kiểu gen bbC– sẽ cho kiểu hình lông màu nâu. Cơ thể mang kiểu gen B–cc không có gen trội C nên thiếu một giai đoạn đầu trong chuỗi phản ứng sinh hóa hình thành màu sắc lông nên kiểu hình là lông trắng. Và như vậy, cơ thể có kiểu gen bbcc thiếu cả 2 alen trội B, C nên hiển nhiên kiểu hình sẽ là lông trắng.
3.2.3. Tương tác cộng gộp
Tương tác cộng gộp là kiểu tương tác giữa hai hay nhiều gen không alen để cùng quy định một tính trạng. Trong đó, mỗi gen trội cùng alen hay khác alen đóng góp một vai trò như nhau vào sự phát triển của cùng một tính trạng.
Có hai kiểu tương tác cộng gộp:
➢ Tương tác cộng gộp có tích lũy ➢ Tương tác cộng gộp không tích lũy
OFFICIAL
3.2.3.1. Tương tác cộng gộp có tích lũy Một số tính trạng có liên quan tới năng suất của nhiều vật nuôi, cây trồng như: số lượng hạt trên bắp ngô, sản lượng trứng ở gia cầm … cũng như các tính trạng màu da, chiều cao, màu mắt ở người bị chi phối bởi sự tác động cộng gộp của nhiều gen không alen. Các tính trạng này còn được gọi là tính trạng đa gen hay tính trạng số lượng.
Sự phân ly kiểu hình của các tính trạng đa gen không cho tỷ lệ chất lượng thương phẩm rõ ràng. Các cá thể có biểu hiện kiểu hình dao động khác nhau do nhận nhiều hay ít gen (thường là alen trội) và có thể xếp chúng theo mức độ biểu hiện thành một dãy liên tục.
Dưới đây là một ví dụ điển hình về tương tác cộng gộp.
➢ Màu sắc da ở người là tính trạng đa gen do nhiều gen không alen tương tác quy định.
➢ Phép lai được tiến hành theo sơ đồ sau:
Pt/c: AABBCC (da rất đen) × aabbcc (da rất trắng)
F1: AaBbCc (100% da nâu)
F1 × F1: AaBbCc (da nâu) × AaBbCc (da nâu)
F2: 1 da rất trắng :6 da trắng :15 da trắng vừa : 20 da nâu : 15 da đen vừa : 6 da đen : 1 da rất đen.
DẠYKÈMQUYNHƠN
387
Hình 3. 3. Tương tác cộng gộp có tích lũy hình thành màu sắc da ở người
➢ Cơ sở sinh hóa
Tính trạng màu sắc da ở người là do tương tác giữa 3 cặp gen không alen (A và a, B và b, C và c) quy định.
Trong đó mỗi alen trội A, B, C đều có vai trò như nhau trong việc quy định màu sắc da. Kiểu gen mang càng nhiểu alen trội thì kết quả kiểu hình là da càng đen.
Các alen lặn a, b, c không có vai trò trong việc tích lũy sắc tố da. Kiểu gen mang càng nhiều alen lặn thì kiểu hình là da càng trắng.
Có thể giải thích hiện tượng tương tác cộng gộp không tích lũy của các gen không cùng alen bằng cơ chế tiến hóa cấp phân tử Do đột biến lặp đoạn hoặc chuyển đoạn dẫn đến trên các NST có thể mang các locus gen khác nhau nhưng cùng tạo ra các sản phẩm Protein như nhau. Lượng sản phẩm Protein tạo ra càng nhiều thì có tác động càng mạnh lên sự biểu hiện của tính trạng. Kết quả ở F2 thông qua phép lai trên ta thu được tỉ lệ một dãy các kiểu hình từ da rất trắng đến da rất đen. 3.2.3.2. Tương tác cộng gộp không tích lũy ➢ Hìnhdạngquảởcâytềdohaicặpgenkhôngalentươngtácquyđịnh.Trongđó: A–B–; aaB–; A–bb: quả hình tam giác aabb: quả hình bầu dục ➢ Tiến hành phép lai thu được kết quả như sau: Pt/c: AABB (Quả hình tam giác) × aabb (Quả hình bầu dục)
F1: AaBb (100% quả hình tam giác) F2: 15 quả hình tam giác (A-B-; A-bb; aaB-) : 1 quả bầu dục (aabb) ➢ Cơ sở sinh hóa Kết quả của phép lai được giải thích theo 2 cách: Cách 1: hình dạng quả ở cây tề do tương tác cộng gộp không tích lũy của hai cặp gen không alen (A và a, B và b) quy định. Trong đó, sự có mặt của alen trội (không phụ thuộc vào số alen trội) trong kiểu gen quy định kiểu hình là quả hình tam
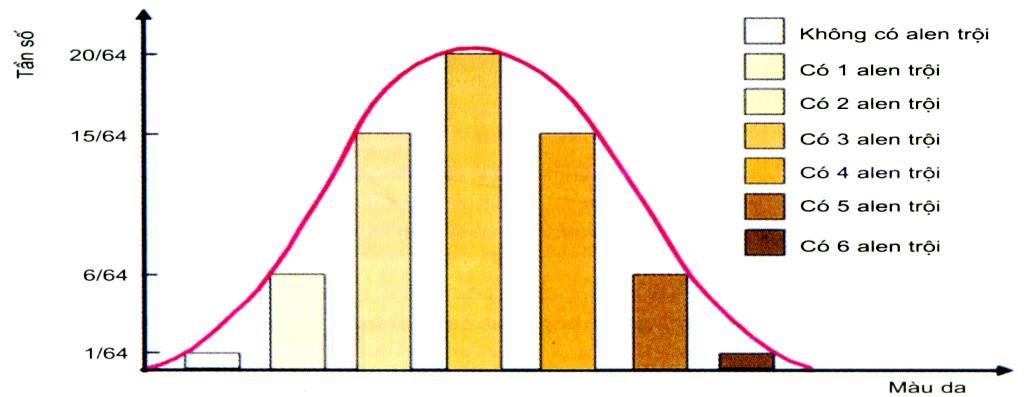
DẠYKÈMQUYNHƠN
388
OFFICIAL
giác. Nếu trong kiểu gen không có alen trội thì các cơ thể đó sẽ mang kiểu hình quả bầu dục.
Cách 2: Ngoài ra còn có một giả thuyết khác được đưa ra để giải thích kết quả của phép lai trên. Người ta cho rằng tính trạng hình dạng quả ở cây tề là do tương tác át chế trội kép giữa 2 cặp gen không alen tức A > B, b và B > A, a.
3.2.4. Tương tác giữa gen trong nhân và gen ngoài nhân
OFFICIAL
Hình 3.4. Thí nghiệm chứng minh sự di truyền theo hệ mẹ của hiện tượng bất thụ đực.
Tính bất dục do nhiều nguyên nhân, bất dục đực (không tạo phấn hoa hay tạo phấn hoa không có khả năng thụ tinh) ở thực vật có các trường hợp sau:
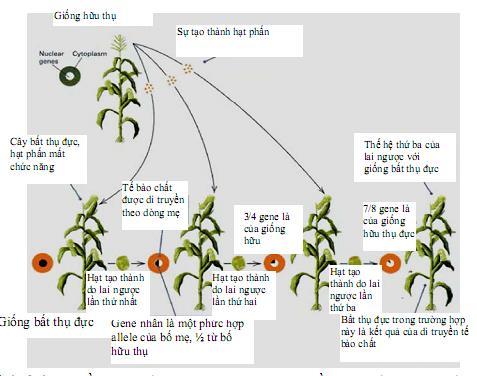
- Do gene nhân quy định, như gene ms ở cây ngô.
- Do ảnh hưởng của điều kiện môi trường như độ ẩm, quang chiếu, khả năng cung cấp chất dinh dưỡng không đáp ứng đúng nhu cầu sinh lý của cây.
* Ví dụ: gene ms ở cây ngô
- Do lai xa cũng đưa đến các cơ thể lai không có hạt phấn vì NST có nguồn gốc khác nhau không thể tiếp hợp nhau trong giảm phân. Những hiện tượng bất dục này đều có ý nghĩa hạn chế chỉ có bất dục bào chất đực là có vai trò quan trọng. Đó là trường hợp bất dục của hạt phấn bắt nguồn từ tế bào chất, còn nhân thì có thể có điều chỉnh được nhờ đó có thể dùng các cây bất dục bào chất đực để phát huy ưu thế lai ở các đối tượng ngô, cao lương, củ cải đường... Ví dụ: Xét mối quan hệ giữa kiểu gene, kiểu bào gene và kiểu hình của bắp được sử dụng trong lai một tính mà không cần khử đực ở cây mẹ STT Kiểu gene Kiểu bào Kiểu hình
DẠYKÈMQUYNHƠN
389
gene (hạt phấn)
Rfrf S (bất dục) Hỏng 2
1
Rfrf N (hữu dục) Tốt 3
RfRf hoặc Rfrf N Tốt 4
RfRf hoặc Rfrf S Tốt
Vậy hạt phấn của ngô chỉ bị mất hoạt tính khi có yếu tố bất dục trong tế bào chất mà lại thiếu thiếu gene phục hồi hữu dục (Rf) ở trong nhân, alen của gene này là rf là gene cũng cố tính bất dục. Cây được phục hồi hữu dục RfrfS cho tự thụ phấn thì ở đời sau sẽ có 1/4 rfrf có hạt phấn hỏng. Nếu lấy bắp của dạng rfrfS thụ phấn của rfrfN, thì phấn hoa của toàn bộ đời sau sẽ bị hỏng, cây này chỉ còn bắp mang nhị cái. Đó là cách dùng phương pháp di truyền để khử cờ ngô. Khi sản xuất hạt giống, những cây này muốn có hạt thì phải thụ phấn hữu dục của những cây bình thường. Nếu muốn dùng những hạt đó để sau này trồng lại thì cây bố phải có kiểu gene RfRf và kiểu bào gene N hoặc S. Trong sản xuất giống ngô có thể dùng tổ hợp dòng thuần dạng 1 làm cây mẹ và dạng 3 hoặc dạng 4 đồng hợp tử làm cây bố. Như thế sẽ đỡ mất công khử đực ở cây mẹ và hạt lai thu được từ cây mẹ sẽ có kiểu gene Rfrf, kiểu bào gene S. Kiểu gene này đảm bảo được sự thụ phấn bình thường lúc trồng trong sản xuất.
Bất thụ đực tế bào chất ở ngô liên quan đến 2 plasmid dạng thẳng S1 và S2. Chúng ở trong ty thể cùng với mtADN. Một trong những tính chất khó hiểu của plasmid này là chúng có thể thực hiện tái tổ hợp với mtADN.
3.3. Một số câu hỏi, bài tập tương tác gen
3.3.1. Câu hỏi
DẠYKÈMQUYNHƠN
390
OFFICIAL
Câu 1: Hiện tại, bạn đang nghiên cứu về sự tổng hợp threonin ở E.coli. Để tìm các gen mã hóa các protein quan trọng cho tổng hợp threonin, bạn tiến hành gây đột biến để tìm các đột biến yêu cầu bổ sung threonin cho phát triển. Bạn đã tìm thấy có 9 dòng đột biến và tất cả đều cần bổ sung threonin vào môi trường nuôi cấy để phát triển. Sau đó bạn tiến hành cis-trans test với tất cả các dòng đột biến bạn có. Kết quả thu được như bảng sau (+: phát triển; -: không phát triển):
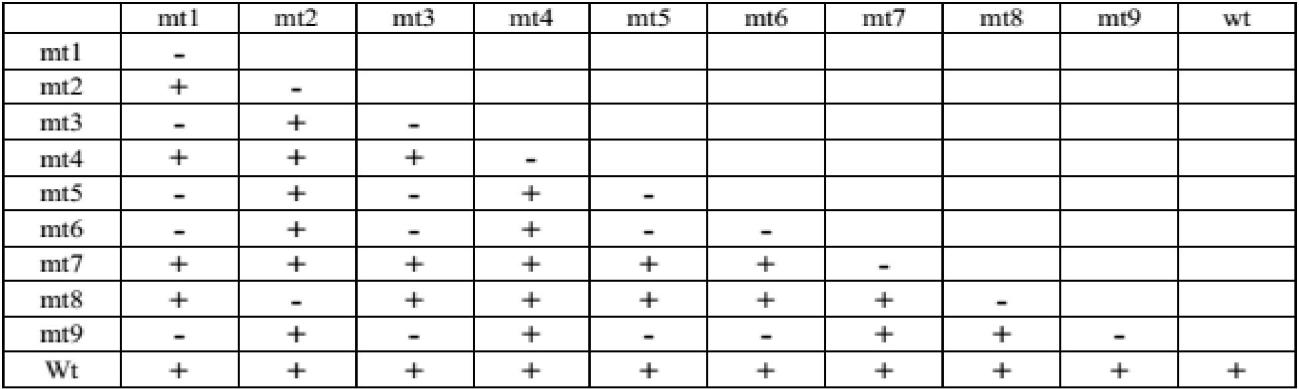
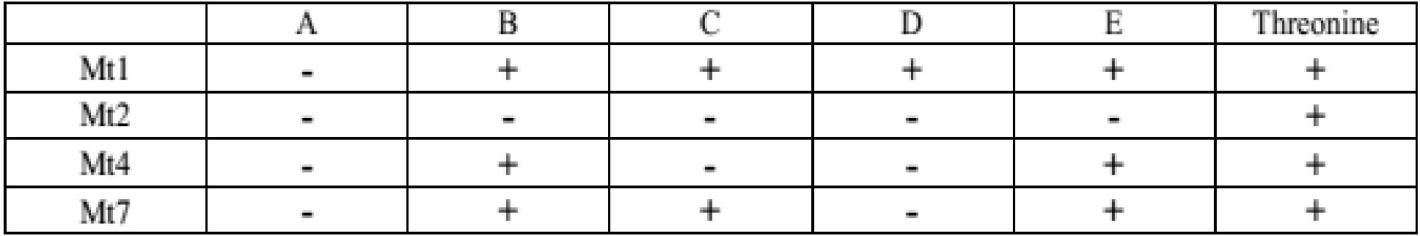
OFFICIAL
a. Có bao nhiêu gen mã hóa các enzym tham gia vào quá trình tổng hợp threonin? Giải thích.
b. Khi xem xét quá trình tổng hợp threonin, người ta nhận thấy có
5 cơ chất tham gia vào quá trình đó. Người ta tiến hành nuôi cấy các đột biến ở trên trong các môi trường chứa các cơ chất khác nhau để xác định trình tự quá trình biến đổi. Kết quả thu được trình bày ở bảng sau (+: phát triển; -: không pháttriển):
Hãy xác định trật tự chuyển hóa của các chất A, B, C, D, E và các gen mã hóa các enzym xúc tác sự chuyển hóa tương ứng. Giải thích. Trả lời
a)Ta thấy: - Khi cho các dòng đột biến giống nhau lai với nhau
=> Không phát triển, không tạo khuẩn lạc.
=> Đột biến trên cùng một gen nên không bù trừ được cho nhau.
=> Tạo kiểu hình đột biến.
- Khi cho những dòng đột biến khác nhau lai với nhau:
+ Nếu khuẩn lạc phát triển
=> Hai đột biến thuộc hai gen khác nhau bù trừ cho nhau tạo kiểu gen dị hợp
DẠYKÈMQUYNHƠN
391
=> Tạo kiểu hình bình thường (kiểu dại).
+ Nếu khuẩn lạc không phát triển
=> Hai đột biến thuộc cùng một gen nên không bù trừ cho nhau
=> Tạo kiểu gen đồng hợp
=> Tạo kiểu hình đột biến. Từ đó, ta có: - 1,3,5,6,9 thuộc cùng một gen. - 2,8 thuộc cùng một gen.
- 4 không bù cho mình, bù cho những dòng đột biến còn lại => gen độclập - 7 không bù cho mình, bù cho những dòng đột biến còn lại => gen độclập
=> Có 4 gen tham gia tổng hợp threonin.
b) sơ đồ chuyển hóa:
A Mt1 D Mt7 C Mt4 B,E Mt2 Threonin
Giải thích
- Các chủng đột biến không sinh trưởng được trong môi trường chỉ chứa A => A là chất đầu dãy chuyển hóa - Chỉ có chủng đột biến Mt1 sinh trưởng được trong môi trường chỉ chứa D => D là chất thứ 2 của dãy chuyển hóa và chủng Mt1 bị đột biến ở bước chuyển hóa chất A thành chất D
-Trong môi trường chỉ chứa C, có hai chủng Mt1 và Mt7 sống được => chất C là chất thứ 3 của dãy chuyển hóa và chủng Mt7 là chủng bị đột biến ở bước chuyển hóa chất D thành chất C
-chủng Mt4 sống được trong môi trường chỉ chứa B và E => chất B và E cùng là chất thứ 4 trong dãy chuyển hóa và chủng Mt2 bị đột biến ở bước chuyển hóa chất C thành chất B hoặc E -chủng Mt2 chỉ sống được trong môi trường chứa Threonin => chủng này bị đột biến ở bước chuyển hóa chất B hoặc chất E thành Threonin
DẠYKÈMQUYNHƠN
OFFICIAL 392
Câu
lập từ các chủng
ấ
ố
việc bổ
2: Trích đề IBO 2012 Có một số các chủng đột biến dinh dưỡng được phân
n
m m
c bánh mỳ đỏ kiểu hoang dại Neurospora crassa đáp ứng với
sung thêm các chất nhất định vào môi trường nuôi cấy, nếu phát triển (+) hoặc không phát triển (0). Bảng dưới đây cho biết các số liệu về các thể đột biến gen đơn.
a. Hãy chỉ ra thứ tự chuyển hóa (1-5) của 5 sản phẩm chuyển hóa trong quá trình trao đổi trong phiếu trả lời
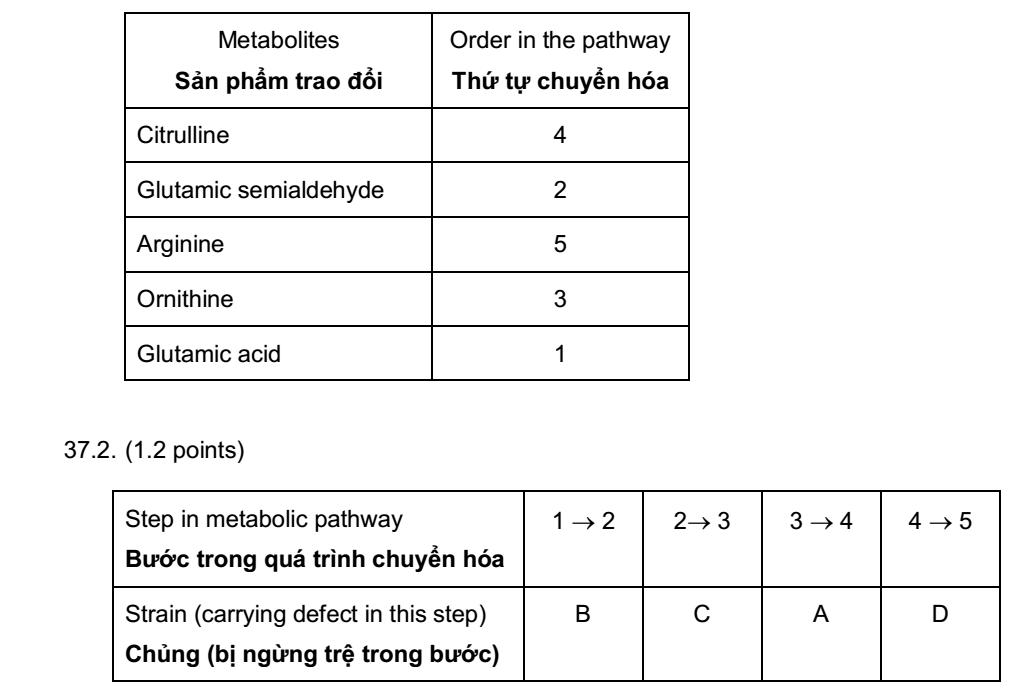
b. Hãy chỉ ra chủng nào bị ngăn cản ở mỗi một bước trong 4 bước của quá trình chuyển hóa nêu trong phiếu trả lời
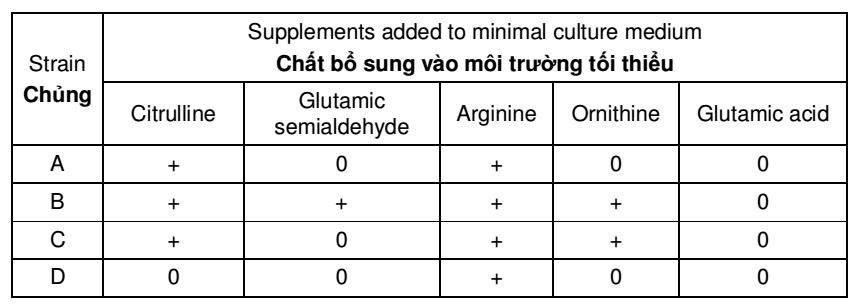
DẠYKÈMQUYNHƠN
393
OFFICIAL
Câu 3. A, B, C, D là các chất chuyển hoá trung gian (không theo đúng thứ tự) trong một con đường hoá sinh của tế bào. Người ta tìm thấy 4 thể đột biến khác nhau kí hiệu từ D1- D4. Khi nuôi cấy 4 thể đột biến này lần lượt trong các môi trường được bổ sung chất A, B, C và D, người ta thu được kết quả như sau: D1 chỉ sinh trưởng trong các môi trường có A hoặc D; D2 chỉ sinh trưởng trong các môi trường chứa A hoặc B hoặc D; D3 chỉ sinh trưởng trong môi trường có D; D4 chỉ sinh trưởng trong môi trường có A hoặc B hoặc C hoặc D. Hãy vẽ sơ đồ các bước chuyển hoá của con đường hoá sinh trên và chỉ ra những bước chuyển hoá bị ức chế tương ứng ở các thể đột biến (D1- D4). Giải thích.
Trả lời
Sơ đồ chuyển hoá theo trình tự : C
→ B → A → D
- D1 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá B thành A. - D2 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá C thànhB.
OFFICIAL
- D3 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá A thành D.
- D4 bị đột biến làm mất chức năng của gen mã hoá enzim chuyển hoá hình thành chất C.
Ta có thể suy ra được trình tự các bước chuyển hoá dựa trên nguyên lý là nếu thể đột biến nào cần phải bổ sung tất cả các chất thì thể đột biến đó bị hỏng gen qui định enzim chuyển hoá tiền chất đầu tiên của con đường chuyển hoá. Thể đột biến nào chỉ cần bổ sung một chất thì chất đó là sản phẩm cuối cùng của con đường chuyển hoá.
Câu 4. (Trích đề IBO năm 2015) Ở nấm mốc, sự sinh tổng hợp axit amin arginine cần bước chuyển hóa tiền chất thành ornithine; chất này được chuyển hóa tiếp thành citruline là tiền chất trực tiếp được chuyển hóa thành arginine. Mỗi phản ứng trong 3 bước này được xúc tác bởi một enzym riêng rẽ; mỗi enzym được mã hóa bởi một gen (xem Hình) (tiền chất)
DẠYKÈMQUYNHƠN
394
A: Con đường sinh tổng hợp arginine ở nấm mốc. B: Thiết kế thí nghiệm để xác định chuyển hóa: dải màu nâu đen chỉ Nấm mốc sinh trưởng. Chủng kiểu dại mọc trên môi trường tối thiểu, trong khi các chủng đột biến chỉ mọc trên các môi trường được bổ sung (+) chất dinh dưỡng đặc thù. Chọn câu trả lời đúng nhất
A. Các chủng đột biến lớp III có thể mang đột biến ở nhiều hơn 1 gen liên quan đến con đường sinh hóa này
B. Các chủng đột biến lớp I thiếu cả 3 enzym
C. Các thể đột biến thiếu hai enzym A và C có thể mọc nếu môi trường được bổ sung đồng thời Orthnithine và Citruline
D. Giả sử một chất ức chế enzym B được bổ sung vào môi trường nuôi chủng kiểu dại, gen B sẽ bị bất hoạt E. Tất cả đều sai Đáp án :
DẠYKÈMQUYNHƠN
395
OFFICIAL
E 3.3.1. Bài tập Bài tập 1 : Ở phả hệ sau, II-2 là người đầu tiên trong gia đình biểu hiện các triệu chứng liên quan đến đột biến ở gen ti thể (Hội chứng Merff).
a. Hãy đưa ra hai lí do để giải thích hợp lí nhất về việc tại sao người mẹ I-1 không bị bệnh nhưng II-2 bị bệnh này?
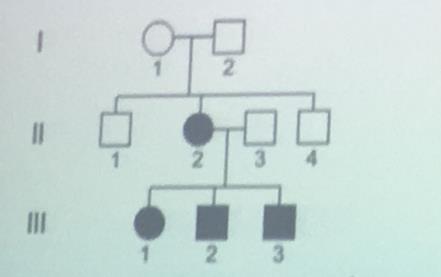
b. Bằng cách nào có thể phân biệt hai lí do nêu trong giải thích ở câu a?
Bài tập 2: Gen hiệu ứng mẹ ở ruồi giấm, kí hiệu t là alen lặn, làm ngăn cản sự phát triển của các cấu trúc đầu – đuôi. Ruồi đực kiểu dại được lai với ruồi cái có kiểu gen chưa biết tạo ra ấu trùng không có cấu trúc đầu đuôi và thường chết ở giai đoạn phôi. Hãy xác định kiểu gen và kiểu hình của ruồi cái ở phép lai này. Xác định kiểu gen và kiểu hình của bố mẹ của ruồi cái đó.
Bài giải:
Do ấu trùng tạo ra không có cấu trúc đầu đuôi → kiểu gen của ruồi cái là tt nhưng không bị bệnh do ruồi bố mẹ của ruồi cái là: Tt x Tt hoặc Tt x tt.
Bài tập 3. Ở cây bốn giờ Mirabilus jalapa cùng một cây, một số các nhánh chứa lá màu xanh, một số nhánh chứa lá màu trắng, một số nhánh chứa lá đốm có những mảng màu xanh và trắng xen kẽ. Màu xanh của lá và thân do diệp lục được tổng hợp bởi sự quy định của gen nằm trong lục lạp.
a. Giải thích vì sao cây này có một số nhánh có lá khác nhau như vậy.
b. Khi lấy noãn bào từ nhánh cây lá xanh với hạt phấn từ hoa của bất kì nhánh nào người ta thấy F1 tạo ra tất cả đều có lá màu xanh. Khi thụ phấn noãn của nhánh đốm với hạt phấn từ bất kì nhánh nào, 85% cây con tạo ra có một số nhánh màu xanh, một số có lá màu trắng, một số cá lá đốm; 10% cây con chỉ có lá xanh và 5% cây con có lá trắng nhưng sau đó bị héo và chết sau khi nảy mầm. Hãy giải thích kết quả này. Làm thế nào noãn bào tử nhánh lá đốm tạo ra thế hệ con đều có lá xanh hoặc lá trắng? Tại sao các cây có lá trắng hoàn toàn nhanh chóng bị chết?
c. Với câu trả lời ở phần b, làm thế nào các cây lá đốm có nhánh trắng vẫn khỏe mạnh bình thường?
Trả lời:
a. Do gen trong lục lạp phân chia độc lập với phân chia nhân. Tế bào hợp tử chứa cả lục lạp bình thường – gen không bị đột biến và cả lục lạp màu trắng – do gen đột biến. Khi tế bào phân chia thì tế bào chất được phân chia không đồng đều về các
DẠYKÈMQUYNHƠN
396
OFFICIAL
tế bào vì thế tế bào nào nhận được chỉ lục lạp bình thường từ đó tạo nhánh toàn lá xanh; tế bào nào nhận được hai loại lục lạp sẽ có lá đốm, tế bào nào nhận được chỉ lục lạp chứa gen đột biến sẽ có màu trắng.
b. Do gen trong tế bào chất của noãn là chủ yếu, còn hạt phấn chứa tinh tử có lượng tế bào chất không đáng kể vì thế gen do tế bào chất quy định di truyền theo dòng mẹ.
+ Noãn của cây có lá xanh – toàn lục lạp không bị đột biến dù dùng hạt phấn của cành nào → con đều có lá màu xanh.
+ Noãn của cây có lá đốm → con F1 sẽ chủ yếu là lá đốm, một tỉ lệ nhỏ có cây lá xanh và một tỉ lệ nhỏ cho lá trắng . Vì tế bào mẹ đại bào tử phân chia tế bào chất ngẫu nhiên tạo ra các noãn khác nhau.
OFFICIAL
- Cây lá trắng chết sau khi mọc mầm vì không có sắc tố quang hợp nên không sống được. c. Cây đốm vẫn khỏe mạnh vì các lá có lục lạp bình thường quang hợp tạo ra sản phẩm nuôi dưỡng cả cây.
Bài tập 4. Ở ngô có hai dòng tế bào chất: dòng N hữu thụ và dòng S bất thụ ; Gen ở trong nhân: alen Rf – có khả năng phục hồi hữu thụ; alen rf – không có khả năng phục hồi. Chọn P để thu được F1 đồng nhất kiểu gen SRfrf - có ưu thế lai cao
Trả lời:
P. ♀ Srfrf x ♂N(S)RfRf → F1: SRfrf.
Bài tập 5: Cho Pt/c. ♀ lông đen x ♂ lông trắng→ F1: 100% lông đen
a. Cho ♀F1 lai với P♂ lông trắng (trắng do gen lặn quy định). Hãy xác định kết quả phép lai?
b. Cho F1 giao phối với nhau được kết quả KG,KH ở F2 như thế nào?
Cho rằng màu lông do 1 gen quy định nằm trên NST thường và chỉ có 2 KH trên?
Trả lời:
- TH1: Quy luật phân
DẠYKÈMQUYNHƠN
397
li. - TH2: Tương tác gen
nhân và tế bào chất. P. AA x aa → F1. Aa –đen Pb. ♀ Aa (F1) x ♂ aa (P) → 1Aa : 1aa (bất thụ) F1xF1: Aa xAa → F2: 3A-: 1aa
ất thụ) Bài tập 6. Cho phả hệ sau
ự di
ền của một bệnh rất hiế
ặ
trong
(b
theo dõi s
truy
m g
p.
a. Hãy cho biết quy luật di truyền chi phối bệnh này?
b. Có cá thể nào trong phả hệ mang đột biến gây bệnh mặc dù không biểu hiện tính trạng bệnh không? Hãy chỉ ra các cá thể đó trong phả hệ (viết chữ số La mã chỉ số thế hệ và số thường chỉ thứ tự từ trái sang phải).
Bài giải:
a. Bệnh hiếm gặp nên tần số alen gây bệnh sẽ thấp, những người kết hôn với người trong gia đình không mang alen bệnh. Chỉ có thể giải thích đây là hiện tượng in vết hệ gen, in vết từ mẹ (III.8 Aa sinh con không bị bệnh) và bệnh do gen trội quy định.
OFFICIAL
b. Người mẹ II.2 và bố II.1 mang gen bệnh (Aa) nhưng không biểu hiện do được in vết từ thế hệ trước.
Bài tập 7 (Câu 15 đề chọn đội tuyển Olympic sinh học năm 2014 - Ngày 2) Ở một loài, alen A quy định lông mượt, còn alen a quy định lông xù. Khi lai con cái thuần chủng lông mượt với con đực lông xù, được F1 có tỷ lệ 1 con lông mượt: 1 con lông xù. F1 giao phối với nhau được F2 có tỷ lệ 1 con lông mượt: 1 con lông xù. Giải thích kết quả của phép lai và viết sơ đồ lai từ P đến F2.
Hướng dẫn trả lời
- Kết quả ở F1 có tỷ lệ 1 con lông mượt: 1 con lông xù phân bố đều ở hai giới tính, do đó không thể giải thích được bằng di truyền liên kết giới tính và ảnh hưởng của giới tính, Vì cả hai trường hợp này tuy đều cho F1 có tỷ lệ kiểu hình 1:1 nhưng phân hoá giới tính, ví dụ như 1 cái mắt đỏ: 1 đực mắt trắng ở ruồi giấm.
- Kết quả phép lai chỉ có thể giải thích bằng sự di truyền in vết, trong đó alen lặn (a) trong quá trình hình thành giao tử đã được in vết (a*), nghĩa là nó bị methyl hoá xitozin theo hướng hoạt hoá. Do đó với sự hiện diện của a* ở thể dị hợp tử có biểu hiện lông xù.
Sơ đồ lai
P: ♀ Lông mượt x ♂ Lông xù AA aa
F1: Aa: Aa* → a*- hoạt hoá (in vết)
1 lông mượt : 1 lông xù
F2: 1 AA: 1Aa: 1Aa*: 1aa* 1 lông mượt : 1 lông xù
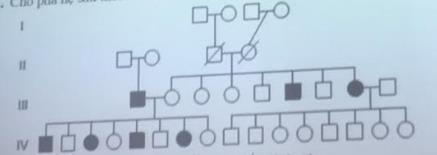
DẠYKÈMQUYNHƠN
398
Bài tập 8 (Câu 15 đề chọn đội tuyển Olympic sinh học năm 2014 - Ngày 2) Ở cây ngô, alen A quy định lá màu xanh đậm nằm trên nhiễm sắc thể số 1, alen lặn a quy định lá màu xanh nhạt; alen B nằm trên nhiễm sắc thể số 5 quy định bắp ngô to, alen lặn b quy định bắp ngô nhỏ. Cho hai dòng ngô thuần chủng lá màu xanh đậm, bắp to và lá màu xanh nhạt, bắp ngô nhỏ giao phấn với nhau được F1. Các cây F1 đều có khả năng sinh sản bình thường. Khi một cây ngô F1 tự thụ phấn tạo ra một số lượng lớn cá thể ở F2, trong đó 1/4 số cá thể lá màu xanh nhạt, bắp nhỏ. Hãy giải thích kết quả ở F2. Hướng dẫn trả lời - Hai dòng thuần giao phấn với nhau cho F1 dị hợp tử về hai cặp gen nằm trên hai nhiễm sắc thể khác nhau. Bình thường một cây F1 khi tự thụ phấn sẽ phải cho ra 1/16 cá thể đời con có lá màu xanh nhạt, bắp nhỏ (aabb). Nhưng trong trường hợp này cây tự thụ phấn lại cho ra tỉ lệ 1/4 aabb nên ta có thể cho rằng đã xảy ra đột biến chuyển đoạn qua lại giữa nhiễm sắc thể số 1 và số 5 dẫn đến alen a nằm trên cùng một nhiễm sắc thể với alen b. - Hơn thế nữa alen a và alen b phải nằm rất gần điểm đứt gẫy chuyển đoạn khiến cho alen a và b nằm rất gần nhau dẫn đến chúng luôn di truyền cùng nhau. Cây ngô mang đột biến này hoàn toàn hữu thụ nên ta có thể cho rằng đó là cây đột biến chyển đoạn đồng hợp vì nếu là chuyển đoạn dị hợp sẽ bất thụ 50%.
OFFICIAL
Bài tập 9: Tương tác giữa 1 gen nằm trên NST thường và 1 gen nằm trên NST giới tính (Trích đề thi chọn HSG quốc gia năm học 2015-2016)
Có hai dòng ruồi giấm thuần chủng đều có mắt màu đỏ tươi được kí hiệu là dòng I và dòng II. Để nghiên cứu quy luật di truyền chi phối tính trạng, người ta đã thực hiện hai phép lai dưới đây:
Phép lai 1: Lai các con cái thuộc dòng I với các con đực thuộc dòng II; F1 thu được 100% ruồi con đều có màu mắt kiểu dại.
Phép lai 2: Lai các con cái thuộc dòng II với các con đực thuộc dòng I; F1 thu được 100% các con cái có màu mắt kiểu dại; 100% con đực có màu mắt đỏ tươi.
Từ kết quả của các phép lai trên có thể rút ra được những kết luận gì? Giải thích và viết sơ đồ lai minh họa.
Giải
- Từ kết quả của phép lai 1: Khi lai hai dòng ruồi thuần chủng đều có mắt đỏ tươi với nhau, đời con F1 đều thu được 100% cá thể có màu mắt kiểu dại→ màu mắt của ruồi giấm do hai gen tương tác kiểu bổ trợ - Từ kết quả của phép lai 2 ta thấy có sự phân ly không đồng đều ở 2 giới→ có sự di truyền liên kết giới tính. Khi lai con cái thuộc dòng II với con đực thuộc dòng I
DẠYKÈMQUYNHƠN
399
cho ra đời con có tất cả các con cái đều có màu mắt kiểu dại, còn các con đực đều có mắt đỏ tươi→ di truyền chéo.
- Một trong hai gen quy định tính trạng phải nằm trên nhiễm sắc thể giới tính X tại vùng không tương đồng với Y, gen còn lại nằm trên NST thường (vì nếu cả hai gen cùng nằm trên vùng tương đồng của cặp XY hoặc cùng nằm trên X tại vùng không tương đồng với Y thì sẽ không thu được kết quả như phép lai).
- Từ kết quả của phép lai 1→ alen đột biến gây màu mắt đỏ tươi ở dòng I phải nằm trên NST thường. Lý do là nếu alen lặn nằm trên NST giới tính X thì tất cả các con đực sẽ có mắt màu đỏ tươi.
- Từ kết quả của phép lai 2 ta thấy gen lặn quy định màu mắt đỏ tươi phải nằm trên NST X vì tất cả các con đực đều có màu mắt đỏ tươi (có hiện di truyền chéo)
- Tổng hợp kết quả của cả phép lai 1 và 2, ta có thể viết sơ đồ lai chứng minh như sau:
Phép lai 1: P I (đỏ tươi) x II (đỏ tươi) aaXBXB AAXbY


F1: AaXBXb Mắt kiểu dại AaXBY Mắt kiểu dại
Phép lai 2: P II (đỏ tươi) x I (đỏ tươi) AAXbXb aaXBY
F1: AaXBXb Mắt kiểu dại AaXbY Mắt đỏ tươi
Bài tập 10: Tương tác gen và kiểm định χ2 (khi bình phương) (Trích đề thi chọn HSG quốc gia năm học 2013-2014) Ở một loài thực vật, người ta thực hiện hai phép lai sau: Phép lai I: Dòng 1 (hoa trắng) x Dòng 2 (hoa đỏ) được F1 100% hoa trắng. Cho F1 tự thụ phấn, thu được F2 có 124 cây hoa trắng và 36 cây hoa đỏ.
Phép lai II: Dòng 1 (hoa trắng) x Dòng 3 (hoa đỏ) được F1 100% hoa trắng. Cho F1 tự thụ phấn, thu được F2 có 122 cây hoa trắng và 38 cây hoa đỏ. Biết rằng, kiểu gen và cách tác động của gen ở dòng 1 trong hai phép lai giống nhau.
a) Hãy dùng tiêu chuẩn χ2 (khi bình phương) để kiểm định sự phù hợp hay không phù hợp giữa số liệu thực tế và số liệu lí thuyết của 2 phép lai trên. Cho biết, χ2 lí thuyết = 3,84.
DẠYKÈMQUYNHƠN
OFFICIAL 400
b) Giải thích kiểu tác động của gen đối với sự hình thành màu hoa ở kết quả của hai phép lai trên.
c) Cho rằng khi lai dòng 2 với dòng 3 được F1 100% hoa tím. Cho F1 tự thụ phấn, thì kết quả ở F2 sẽ thế nào về kiểu gen và kiểu hình? Giải thích kiểu tác động của gen đối với kết quả của phép lai.
Hướng dẫn
a) Từ kết quả thu được của hai phép lai, tỉ lệ phân li màu hoa ở F2 có thể có hai trường hợp là 13 hoa trắng : 3 hoa đỏ hoặc 3 hoa trắng : 1 hoa đỏ. Nhưng tỉ lệ 3 hoa trắng : 1 hoa đỏ là không phù hợp về mặt di truyền học và các dữ kiện của ý c. Kiểm định tỉ lệ 13 : 3 - Phép lai I
Hoa trắng Hoa đỏ Tổng số Số liệu thực tế : 124 36 160 Số liệu lí thuyết : 130 30 160 Độ lệch d : -6 +6 d2 36 36 χ2 36/130 + 36/30 = 1,47 < 3,84 tỉ lệ 13 : 3 chấp nhận được. - Phép lai II
Hoa trắng Hoa đỏ Tổng số Số liệu thực tế : 122 38 160 Số liệu lí thuyết : 130 30 160 Độ lệch d : -8 +8 d2 64 64 χ2 64/130 + 64/30 = 2,64 < 3,84 tỉ lệ 13 : 3 chấp nhận được. b) - Vì kết quả cả 2 phép lai giống nhau, trong đó dòng 2 và dòng 3 có kiểu gen khác nhau, do đó phải liên quan tới 3 cặp gen. - P1: AAbbdd (Dòng 1: hoa trắng) x aaBBdd (Dòng 2: hoa đỏ) →F1: AaBbdd (hoa trắng)
F2: (9A-B-dd : 3A-bbdd : 1aabbdd): 13 hoa trắng 3aaB-dd: 3 hoa đỏ
DẠYKÈMQUYNHƠN
OFFICIAL 401
Kiểu tác động của gen: B: hoa đỏ; b: hoa trắng; A át B: hoa trắng; aa không át B.
- P2: AAbbdd (Dòng 1: hoa trắng) x aabbDD (Dòng 3: hoa đỏ) →F1: AaBbdd (hoa trắng)
F2: (9A-B-dd : 3A-bbdd : 1aabbdd): 13 hoa trắng 3aaB-dd: 3 hoa đỏ
Kiểu tác động của gen: D: hoa đỏ; d: hoa trắng; A át D: hoa trắng; aa không át D.
c) - P3: aabbDD (Dòng 3: hoa đỏ) x aaBBdd (Dòng 2: hoa đỏ) →F1: aaBbDd (hoa tím)
F2: 9aaB-D-: 9 hoa tím (3aaBbdd : 3aabbD-): 6 hoa đỏ 1aabbdd: 1 hoa trắng
Kiểu tác động của gen: B: hoa đỏ; b: hoa trắng; D: hoa đỏ; d: hoa trắng; BD-: hoa tím.
Bài tập 11 (Trích đề thi chọn HSG quốc gia năm học 2012)
Cho rằng ở 1 loài động vật. Lông chỉ có 2 dạng là lông dài và lông ngắn, trong đó KG quy định lông dài là AA, KG quy định lông ngắn là aa. Con đực thuần chủng lông dài cho giao phối với con cái thuần chủng lông ngắn được F1. Cho F1 giao phối ngẫu nhiên với nhau được F2 có số con lông dài chiếm tỉ lệ ¾ ở giới đực và ¼ ở giới cái
a. Giải thích kết quả phép lai b, Viết sơ đồ lai Giải: tỉ lệ phân bố không đều ở 2 giới-> Có liên kết giới tính hoặc ảnh hưởng của giới tính đối với sự hình thành tính trạng - F2 có tỉ lệ phân bố KH 3 lông dài: 1 lông ngắn ở giới đực và ngược lại, nghĩa là sự phân bố các KH không đều ngay ở trong cùng 1 giới tính. Điều này không thể hiện đối với sự DTLK giới tính mà chỉ có với tính trạng ảnh hưởng của giới tính
- Ở F2 giới đực có tỉ lệ 3 lông dài: 1 lông ngắn chứng tỏ thể dị hợp biểu hiện lông dài->-. Thể dị hợp ở giới cái biểu hiện lông ngắn
SĐL Pt/ c ♂ Lông dài (AA) x ♀ lông ngắn aa → F1 Aa (♀ lông ngắn, ♂ lông dài)
F1 x F1 ♀ lông ngắn Aa x ♂ lông dài Aa
Giới đực 3 lông dài : 1 lông ngắn Giới cái 1 lông dài : 3 lông ngắn
DẠYKÈMQUYNHƠN
OFFICIAL 402
Bài tập 12 (Trích đề thi chọn HSG quốc gia năm học 2013-2014)
Ở một loài thực vật, khi cho dòng hoa kép làm mẹ giao phấn với dòng hoa đơn, thu được F1 100% hoa kép. Cho F1 tự thụ phấn, thu được F2 100% hoa kép. Bằng cách nào xác định được tính quy luật sự di truyền của dạng hoa?
Hướng dẫn
- F1 và F2 toàn hoa kép chưa xác định chính xác được tính quy luật của sự di truyền dạng hoa vì kết quả này có thể di truyền qua tế bào chất (di truyền theo dòng mẹ) hoặc di truyền do hiệu ứng dòng mẹ. Vì vậy, muốn xác định được tính quy luật sự di truyền dạng hoa cần phải lai nghịch hay tiếp tục cho F2 tự thụ phấn cho ra F3
- Cách 1: Tiến hành phép lai nghịch
+ Nếu F1 và F2 toàn hoa đơn giống cây mẹ thì dạng hoa di truyền theo dòng mẹ, nghĩa là di truyền qua tế bào chất.
OFFICIAL
+ Nếu F1 toàn hoa đơn giống cây mẹ và F2 toàn hoa kép thì dạng hoa di truyền do hiệu ứng dòng mẹ.
- Cách 2: Cho cây F2 tự thụ phấn
+ Nếu F3 toàn hoa đơn giống cây mẹ thì dạng hoa di truyền theo dòng mẹ, nghĩa là di truyền qua tế bào chất. + Nếu F3 có tỉ lệ 3 hoa kép : 1 hoa đơn thì dạng hoa di truyền do hiệu ứng dòng mẹ.
Bài tập 13 Tương tác bổ sung của 3 gen Ở 1 loài thực vật lưỡng bội, tính trạng màu sắc hoa do 3 cặp gen ( A, a; B, b; D, d) phân li độc lập cùng quy định. Kiểu gen có cả 3 alen trội A, B và D cho hoa màu đỏ, kiểu gen có 2 alen trội A và B nhưng không có alen trội D thì cho hoa vàng, các kiểu gen còn lại thì cho hoa trắng. Theo lí thuyết, trong các dự đoán sau có bao nhiêu dự đoán đúng với các dữ liệu đã cho ?
(1). Ở loài này có tối đa 15 kiểu gen quy định hoa trắng.
(2). P: AaBbDd x AabbDd, thu được F1 có số cây hoa trắng chiếm tỉ lệ 62,5%.
(3). P: AABBdd x AabbDD thu được F1, cho F1 giao phấn được F2 có kiểu hình phân li theo tỉ lệ: 12 cây hoa đỏ: 3 cây hoa vàng : 1 cây hoa trắng.
(4). P: AABBDD x aabbDD, thu được F1. F1 tự thụ phấn thu được F2 có kiểu hình phân li theo tỉ lệ: 9 cây hoa đỏ: 7 cây hoa trắng.
A. 4. B. 1. C. 3. D. 7.1.
A+B+D -> đỏ
A+B : Vàng
(1) Kiểu gen quy định hoa đỏ :A-B-D- : 7.1.2.2=8
DẠYKÈMQUYNHƠN
403
Số kiểu gen quy định hoa vàng : A-B-dd= 7.1.2=4
=> Tổng số kiểu gen quy định hoa trắng = 33 - 8- 4=15 => (1) Đúng
(2) P: AaBbDd x AabbDd
=> F1 Tỉ lệ kiểu gen hoa đỏ : A-B-D- = 3139 42432 xx =
Tỉ lệ KG hoa vàng: A-B-dd = 3113 42432 xx =
=> Tỉ lệ kiểu gen hoa trắng = 935 1 32328 −−=
(2) => Đúng
(3) P : AABBdd x AabbDD
=> F1 : 1 2 AABbDd ; 1 2 AaBbDD => (3) Sai 935 1 32328 −−=
(4) P: AABBDD x aabbDD => F1 AaBbDD
F1x F1 : AaBbDD x AaBbDD
F2 : -> A-B-D- = 339 4416 x = => Tỉ lệ phân ly kiểu hình ở F2 9 đỏ : 7 trắng => Đúng Đáp án [C]
Bài tập 14 Tương tác át chế của 3 gen Ở 1 loài động vật, khi cho lai 2 dòng thuần chủng lông màu trắng và lông màu vàng giao phối với nhau được F1 toàn lông màu trắng cho F1 giao phối với nhau được F2 có kiểu hình: 48 con lông màu trắng: 9 con lông đen: 3 con lông xám: 3 con lông nâu; 1 con lông vàng. Hãy giải thích Giải : - Kết quả cho thấy màu lông bị chi phối bởi sự tương tác của 3 gen không alen trên NST thường và F1dị hợp về 3 cặp gen Sơ đồ phân ly ở F2 3/ 4D- = 27 A- B- D- = 27 con lông trắng
3/ 4 B- ¼ dd = 9 A- B- dd = 9 con lông trắng F2
3/ 4D- = 9 A- bb- D- = 9 con lông trắng 3/4 A 1/ 4 bb
DẠYKÈMQUYNHƠN
404
OFFICIAL
¼ dd = 3 A- bb- dd = 3 con lông trắng
3/ 4D- = 9 aa- B- D- = 9 con lông đen
3/ 4 B- ¼ dd = 3 aa- B- dd = 3 con lông nâu
3/ 4D- = 9 aa- bb- D- = 3 con lông xám
1/4 aa 1/ 4 bb
¼ dd = 1aa- bb- dd = 1 con lông vàng
Nhân xét: alen B: quy định lông nâu, b: lông vàng, alen D; lông xám, d: lông vàng. Các alen trội B và D tác động bổ trợ quy định lông đen: alen A át chế sự hình thành sắc tố→ màu trắng Bài tập 15: Tương tác gen có hiện tượng gây chết và độ thâm nhập, độ biểu hiện (Trích đề thi chọn HSG quốc gia năm học 2015-2016) Một cặp vợ chồng cả hai đều mắc chứng “điếc” do mang một số alen lặn ở 3 gen liên quan đến thính giác: d1 là lặn so với D1, d2 là lặn so với D2, d3 là lặn so với D3. Đồng hợp tử đột biến ở bất cứ 1 trong 3 gen này đều gây “điếc”. Ngoài ra, đồng hợp tử lặn đồng thời ở 2 trong 3 gen này gây chết ở giai đoạn phôi (sảy thai sớm) với độ thâm nhập (độ biểu hiện) là 25%. Đồng hợp tử lặn ở cả 3 gen gây sảy thai sớm với độ thâm nhập là 75%. Với kiểu gen của mẹ là D1d1D2d2d3d3 và của bố là d1d1D2d2D3d3 thì xác suất con của họ được sinh ra (không tính sảy thai) có thính giác bình thường là bao nhiêu? Giải thích.
Giải
Tính tỷ lệ cho từng cặp gen, rồi sử dụng nguyên tắc nhân xác suất (áp dụng cho 3 cặp gen) từ phép lai D1d1D2d2d3d3 x d1d1D2d2D3d thu được tỷ lệ: 3/16 D1- D2-D3- (bình thường) : 3/16 D1- D2-d3d3 (điếc do 1 gen) : 3/16 d1d1D2-D3- (điếc do 1 gen) : 1/16 D1- d2d2D3- (điếc do 1 gen) : 1/16 D1- d2d2d3d3 (điếc do 2 gen) : 3/16 d1d1D2-d3d3 (điếc do 2 gen) : 1/16 d1d1d2d2D3- (điếc do 2 gen) : 1/16 d1d1d2d2d3d3 (điếc do 3 gen) Như vậy, tỉ lệ kiểu hình (chưa tính độ thâm nhập của gen) là 3/16 bình thường : 7/16 điếc do 1 gen : 5/16 điếc do 2 gen : 1/16 điếc do 3 gen Áp dụng độ thâm nhập của gen do tính trạng “gây chết” với độ thâm nhập không hoàn toàn (dưới 100%) với trường hợp chết do 2 hoặc 3 gen: - Với 2 gen: 1/4 chết; 3/4 sống và điếc, như vậy ta có 5 /16 điếc do 2 gen x 3/4 sống = 15/64 sống và điếc do 2 gen
DẠYKÈMQUYNHƠN
405
OFFICIAL
- Với 3 gen: 3/4 chết; 1/4 sống và điếc, như vậy ta có 1/16 điếc do 3 gen x 1/4 sống = 1/64 sống và điếc do 3 gen Tổng cộng ta có: 3/16 bình thường : 7/16 điếc do 1 gen : 15/64 điếc do 2 gen : 1/64 điếc do 3 gen = 12/64 bình thường : 28/64 điếc do 1 gen : 15/64 điếc do 2 gen : 1/64 điếc do 3 gen = 56/64 trẻ sống và 8/64 trẻ chết trong giai đoạn phôi (sảy thai).
Như vậy, trong số trẻ sống (56/64), xác suất trẻ bình thường (không bị điếc) là : 12/56 x 100% = 21,4%
Bài tập 16: Tương tác gen và liên kết gen (Trích đề thi chọn HSG quốc gia năm học 2018-2019)
OFFICIAL
Để tìm hiểu quy luật di truyền chi phối hai tính trạng hình dạng và màu sắc quả bí, một nhà khoa học đã tiến hành lai giữa hai dòng bí thuần chủng quả dẹt, màu xanh với quả dài, màu vàng thu được F1 toàn quả dẹt, màu xanh. Cho F1 tự thụ phấn thu được F2 gồm các kiểu hình có tỷ lệ: 9/16 cây cho quả dẹt, màu xanh 3/16 cây cho quả tròn, màu xanh 2/16 cây cho quả tròn, màu vàng 1/16 cây cho quả tròn, màu trắng 1/16 cây cho quả dài, màu vàng Biết rằng vị trí các gen trên NST không thay đổi trong quá trình giảm phân. a) Hãy xác định quy luật di truyền chi phối mỗi tính trạng nêu trên. Viết sơ đồ lai từ P đến F2 về sự di truyền hai tính trạng trên. b) Xác định tỷ lệ kiểu gen, kiểu hình ở đời con khi cho lai cây F1 với cây có kiểu hình quả dài, màu trắng. Giải
* Xét riêng sự di truyền của từng cặp tính trạng + Tính trạng hình dạng quả: Tỷ lệ phân ly ở F2: dẹt : tròn : dài = 9 : 6 : 1 = 16 tổ hợp lai = 4x4. F1 dị hợp 2 cặp gen không alen cho 4 loại giao tử quy định 1 tính trạng hình dạng quả với tỷ lệ phân ly 9 : 6 : 1 => theo quy luật tương tác gen kiểu bổ trợ. Quy ước: A-B-: quả dẹt; A-bb, và aaB-: quả tròn; aabb: quả dài P: AABB (quả dẹt) x aabb (quả dài) F1: AaBb (quả dẹt) + Tính trạng màu sắc quả Tỷ lệ phân ly ở
406
DẠYKÈMQUYNHƠN
F2:
F1 dị hợp 2 cặp gen
với tỷ lệ phân
Quy ước:
v:
xanh : vàng : trắng = 12 : 3 : 1 = 16 tổ hợp lai = 4×4.
không alen cho 4 loại giao tử quy định 1 tính trạng màu sắc quả
ly 12 : 3 : 1 => theo quy luật tương tác gen kiểu át chế do gen trội
X: quy định màu xanh, gen át chế; x: không át chế; V: quy định màu vàng;
quy định màu trắng
P: XXvv (quả xanh) x xxVV (quả vàng) F1: XxVv (quả xanh)
* Xét sự di truyền của cả hai tính trạng Tỷ lệ phân ly kiểu hình ở F2: 9 : 3 : 2 : 1 : 1 = 16 tổ hợp lai = 4×4 F1 dị hợp về 4 cặp gen, cho 4 loại giao tử khác nhau => 4 cặp gen chi phối 2 tính trạng tồn tại trên 2 NST liên kết với nhau hoàn toàn. Sơ đồ lai: Trường hợp 1: A liên kết với X, B liên kết với V
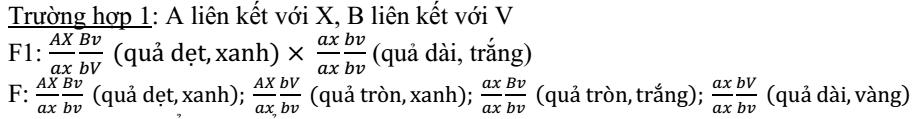
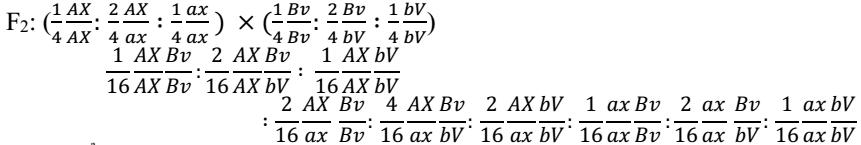
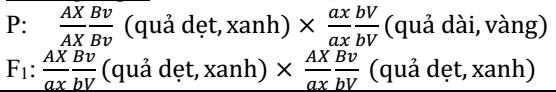
OFFICIAL
Tỷ lệ kiểu gen (9): 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1 Tỷ lệ kiểu hình: 9 dẹt, xanh : 3 tròn, xanh : 2 tròn, vàng : 1 dài, vàng : 1 tròn, trắng Trường hợp 2: A liên kết với V, B liên kết với X Kết quả tương tự như trường hợp 1
Cây có kiểu hình quả dài màu trắng có kiểu gen đồng hợp tử lặn ở cả 4 cặp gen Trường hợp 1: A liên kết với X, B liên kết với V
Tỷ lệ phân ly kiểu gen kiểu hình là 1 : 1 : 1 : 1 Trường hợp 2: A liên kết với V, B liên kết với X Kết quả tương tự như trường hợp 1.
- Khi xét sự di truyền của 2 cặp tính trạng, nếu có 1 tính trạng thường do tương tác gen, 1 tính trạng liên kết với giới tính, ta dựa vào tỉ lệ phân ly kiểu hình của mỗi tính trạng, suy ra kiểu gen tương ứng. Sau đó kết hợp lại ta có kiểu gen chung về các tính trạng.
Bài tập 17: Tương tác gen và di truyền liên kết giới tính có hoán vị gen (Trích đề thi chọn HSG quốc gia năm học 2016-2017)
DẠYKÈMQUYNHƠN
407
Có hai dòng ruồi giấm thuần chủng (A và B) đều có kiểu hình đột biến - mắt xù xì. Lai ruồi cái của dòng A với ruồi đực của dòng B, thu được F1 gồm 100% ruồi cái mắt kiểu dại và 100% ruồi đực mắt xù xì. Cho F1 giao phối ngẫu nhiên thu được F2 gồm 256 ruồi cái mắt kiểu dại, 250 ruồi cái mắt xù xì, 64 ruồi đực mắt kiểu dại và 436 ruồi đực mắt xù xì. Hãy giải thích kết quả và viết sơ đồ lai. Giải
P thuần chủng về đột biến mắt xù xì → F1: 100% ruồi cái kiểu dại → hai đột biến thuộc về hai gen khác nhau (tương tác bổ sung trong sự quy định kiểu hình mắt hay 2 gen không alen với nhau) → F1 dị hợp tử về 2 cặp gen; đột biến là lặn, kiểu dại là trội. Qui ước 2 cặp gen tương ứng là A/a và B/b Kiểu hình ở F1 không đồng đều ở 2 giới: 100% ruồi cái mắt kiểu dại; 100% đực mắt xù xì → gen quy định màu mắt nằm trên nhiễm sắc thể giới tính. Từ số lượng cá thể ở đời lai F2 → Tỷ lệ kiểu hình ở F2 là: + Con cái: mắt kiểu dại : mắt xù xì = 1:1; + Con đực: mắt kiểu dại = 12,8%; mắt xù xì = 87,2%

Nếu 2 gen phân ly độc lập, F1 x F1 không thể cho tỷ lệ phân ly ở F2 như đầu bài đã nêu → 2 gen liên kết không hoàn toàn trên nhiễm sắc thể X; xảy ra hoán vị gen trong giảm phân tạo giao tử ở ruồi cái. P ruồi cái dòng đột biến A (XaBXaB) x ruồi đực dòng đột biến B (XAbY) → F1: ruồi cái (XX) 100% mắt kiểu dại → ruồi cái F1 có kiểu gen dị hợp tử chéo: XaBXAb . F2: 12,8% số ruồi đực có mắt kiểu dại XABY được tạo thành từ giao tử hoán vị gen XAB của ruồi cái → Tần số hoán vị gen = 12,8 x 2 = 25,6%
F 2 Con cái: 50% mắt kiểu dại : 50% mắt xù xì Con đực: 12,8% mắt kiểu dại : 87,2% mắt xù xì
DẠYKÈMQUYNHƠN
OFFICIAL 408
1. KẾT LUẬN
Trên đây chúng tôi đã xây dựng hệ thống cơ sở lý thuyết xây dựng hệ thống cơ sở lý thuyết Một số vấn đề tích hợp hóa sinh, vi sinh và di truyền, đồng thời sưu tầm một số câu hỏi, bài tập chọn lọc nhằm mục tiêu vô cùng thiết thực là giúp cho học sinh thi chuyên Sinh học học tập và nghiên cứu về phần này tốt hơn và đồng thời cũng giúp chính bản thân mình giảng dạy thuận lợi hơn. Phần lí thuyết và các câu hỏi vận dụng mà thực tế chúng tôi đã dạy trên lớp và dùng để bồi dưỡng đội tuyển học sinh giỏi tuỳ từng đối tượng học sinh mà có cách nâng cao khác nhau qua đó đã giúp học sinh nắm được kiến thức, đồng thời các em hiểu sâu hơn về mặt lí thuyết và thấy được ý nghĩa về mặt thực tế. Đặc biệt các em giải nhanh dạng trắc nghiệm rất tốt, kỹ năng làm bài thi cũng được nâng lên.
Tuy nhiên đề tài này chắc chắn còn nhiều thiếu sót, chúng tôi rất mong nhận được sự đóng góp của quý thầy cô, đồng nghiệp và bạn bè để đề tài ngày càng hoàn thiện hơn.
1. ĐỀ NGHỊ
- Nội dung đề tài dành cho học sinh ôn tập kiểm tra định kì ở lớp, thi tốt nghiệp, cao đẳng, đại học và bồi dưỡng đội tuyển học sinh giỏi cấp tỉnh và học sinh giỏi quốc gia, bồi dưỡng giáo viên trong các buổi sinh hoạt chuyên môn. - Đề nghị tổ và trường tạo điều kiện để sang năm tiếp tục phát triển đề tài trên qui mô rộng hơn.
TÀI LIỆU THAM KHẢO
1. Campbell & Reece, Sinh học 8th- Nhà xuất bản giáo dục Việt Nam dịch và xuất bản năm2011
DẠYKÈMQUYNHƠN
409 Phần thứ 3: KẾT LUẬN VÀ KIẾN
Ị
OFFICIAL
NGH
2. Trần Ngọc Danh, Phạm Phương Bình- Tài liệu chuyên Sinh học Trung học phổ thông- Bài tập Di truyền và tiến hoá - Nxb Giáo dục, 2012.
3. Lê Thị Kim Dung – Nguyễn Thị Phương Thảo - Các dạng câu hỏi và bài tập trắc nghiệm Sinh học – NXB Đại học QGHN – 2008
4. Nguyễn Thành Đạt (Tổng chủ biên)- Sách giáo khoa Sinh học 12- NXB Giáo dục, 2008
5. Trần Dũng Hà – Nguyễn Hải Tiến - Phương pháp giải các bài tập sinh học – NXB Đại học QGHN, 2008.
6. Phạm Thành Hổ- Di truyền học- Nxb Giáo dục, 2003.
7. Ngô Văn Hưng, Đỗ Mạnh Hùng, Trần Minh Hương – Giới thiệu đê thi Olympic Quốc tế mônSinh học- Nxb Giáo dục, 2004.
8. Phạm Văn Lập, Trần Ngọc Danh, Đinh Đoàn Long, Tài liệu chuyên Sinh học Trung học phổthông- Di truyền và tiến hoá- Nxb Giáo dục, 2012.
9. Đinh Đòan Long, Đỗ Lê Thăng- Cơ sở di truyền học phân tử và tế bào- Nxb Đại học Quốc Gia Hà Nội, 2013.
10. Vũ Đức Lưu – Sinh học 12 chuyên sâu tập 1- Nxb Đại học Quốc gia Hà Nội, 2014.
11. Vũ Đức Lưu, Nguyễn Thị Hà- Bồi dưỡng học sinh giỏi sinh học Trung học phổ phông - Di truyền và tiến hoá- Nxb Giáo dục, 2013.
12. Vũ Đức Lưu, Nguyễn Thị Hà- Bồi dưỡng học sinh giỏi sinh học Trung học phổ phông, Bài tập di truyền và tiến hoá- Nxb Giáo dục, 2013.
13. Vũ Đức Lưu - Câu hỏi và bài tập trắc nghiệm sinh học 12- Nxb Hà Nội, 2008.
14. Phan Cự Nhân, Nguyễn Minh Công, Đặng Hữu Lanh- Di truyền học- Tập I – Nxb Đại họcSư Phạm, 2004.
15. Phan Cự Nhân, Trần Bá Hoành, Lê Quang Long, Phạm Đình Thái, Hoàng Thị Sản, Mai Đình Yên- Sinh học đại cương – Tập II- Nxb đại học Quốc Gia Hà Nội, 1997.
16. Lê Quang Nghị - Hướng dẫn giải nhanh các dạng bài tập trắc nghiệm và tự luận sinh học -Nxb Đại học Sư Phạm, 2010
17. Phan Khắc Nghệ - Phương pháp giải nhanh các dạng bài tập sinh học –Nxb Đại học Quốc gia Hà Nội, 2012.
18. Đỗ Lê Thăng, Đinh Đoàn Long- Chú giải di truyền học -Nxb Giáo dục, 2010
19. Lê Đức Trình- Sinh học phân tử của tế bào- Nxb khoa học kỹ thuật, 2001.
DẠYKÈMQUYNHƠN
OFFICIAL 410
20. Các đề thi học sinh giỏi quốc tế; học sinh giỏi quốc gia; các đề đề xuất học sinh giỏi Duyên hải bắc bộ (2010 -2019), Trại hè Hùng Vương (2009 – 2019) .
411
DẠYKÈMQUYNHƠN OFFICIAL
Chuyên đề
TÍCH HỢP HOÁ SINH, VI SINH VÀ DI TRUYỀN
Trường THPT Chuyên Hoàng Văn Thụ - Hòa Bình
Chuyên đề đạt giải Ba
I. Lí do chọn đề tài
PHẦN I: MỞ ĐẦU
Trong chương trình sinh học cấp trung học phổ thông, phần kiến thức tích hợp hóa sinh, vi sinh và di truyền là một nội dung quan trọng, được đưa nhiều vào đề thi chọn học sinh giỏi quốc gia, quốc tế. Nhiều giáo viên và học sinh còn lúng túng với phần kiến thức này trong khi ôn luyện đội tuyển HSG. Mặc dù hiện nay có rất nhiều tài liệu viết về hóa sinh, vi sinh và di truyền nhưng rất ít tài liệu về tích hợp cả ba nội dung này với nhau và các tài liệu khác nhau với mục đích khác nhau, phần kiến thức này được trình bày theo nhiều cách khác nhau. Vì vậy, chúng tôi biên soạn chuyên đề: “Tích hợp hóa sinh, vi sinh và di truyền” để giúp học sinh có cái nhìn tổng quát hơn, thống nhất hơn. Đồng thời hỗ trợ cho giáo viên kiểm tra kiến thức học sinh thông qua hệ thống các câu hỏi. Đây cũng là nguồn tài liệu ôn tập trong các kì thi trung học phổ thông, học sinh giỏi các cấp, kì thi chọn học sinh giỏi quốc gia và quốc tế.
II. Mục tiêu của chuyên đề
- Hệ thống hóa kiến thức về hóa sinh và di truyền vi sinh vật.
- Giới thiệu một số câu hỏi, bài tập tích hợp phần hóa sinh, di truyền vi sinh vật.
III. Đối tượng áp dụng
- Các đội tuyển ôn thi học sinh giỏi môn Sinh học các cấp.
- Các giáo viên Sinh học.
DẠYKÈMQUYNHƠN
412
OFFICIAL
A. LÝ THUYẾT
I. Tích hợp hóa sinh trong cơ sở vật chất di truyền vi sinh vật
1. Axit nucleic
Thực tế thì Axit deoxiribonucleic (ADN) đã được khám phá vào năm 1869 ở trong nhân tế bào bạch cầu của người, nhưng lúc đó người ta vẫn chưa biết rõ được chức năng của nó. Sau đó hợp chất này được tách ra từ nhiều kiểu nhân, nên gọi nó là axit nuclêic (axit nhân).
Năm 1910, bằng phân tích hoá học người ta đã phân biệt được 2 loại axit nuclêic là axit deoxiribonucleic (ADN) và axit ribonucleic (ARN).
Năm 1924, những nghiên cứu hiển vi có sử dụng thuốc nhuộm ADN và prôtêin đã cho thấy cả 2 loại hợp chất này đều có trong nhiễm sắc thể. Một loạt các nghiên cứu khác cũng đã gợi ra mối quan hệ gắn bó giữa ADN và vật chất di truyền. Ví dụ hầu hết các tế bào xôma của một loài có 1 lượng ADN không đổi; trong khi đó hàm lượng của ARN và prôtêin lại thay đổi khá nhiều ở các giai đoạn khác nhau của một tế bào và các tế bào của cùng 1 loài. Nhân của các tế bào giao tử thì lại có hàm lượng ADN bằng một nửa nhân của tế bào xôma cùng loài.
Những dẫn liệu lý thuyết lúc bấy giờ đã đưa ra nhiều tranh cãi về ADN hay prôtêin là vật liệu di truyền bởi lẽ những phân tích hoá học cho thấy ADN kém tính đa dạng hơn so với prôtêin.
Năm 1928, F.Griffith đã bằng thực nghiệm chứng minh được ADN là vật chất di truyền. Thí nghiệm được sơ đồ hóa qui trình như sau:
DẠYKÈMQUYNHƠN
413 PHẦ
ỘI
OFFICIAL
N II. N
DUNG CHUYÊN ĐỀ
Hình 1. Thí nghiệm của F.Griffith
Ở vi khuẩn Pneumococus có 2 chủng chủng R và chủng S. Chủng R có khuẩn lạc dạng nhăn nheo, không gây bệnh viêm phổi và chủng S có khuẩn lạc dạng nhẵn bóng gây bẹnh viêm phổi , hai chủng này không phát triển chung với nhau. Khi chuột được tiêm hoặc nhóm tế bào R sống hoặc với nhóm tế bào S chết thì chuột vẫn khoẻ mạnh. Trong trường hợp con chuột bị tiêm một lượng nhỏ chủng R sống và 1 lượng lớn chủng S chết thì chuột lại bị chết vì bị viêm phổi. Phân lập từ các mẫu máu của chuột chết thấy có chủng S sống. Hiện tượng này cho thấy vi khuẩn S không thế tự sống lại được sau khi đã bị đun chết, nhưng các tế bào này đã truyền tính gây bệnh cho tế bào R. Hiện tượng này gọi là biến nạp (transformation).
Năm 1944, T.Avery; Mc Leod và Mc. Carty đã tiến hành thí nghiệm để xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng prôteaza hoặc ARNaze thì hoạt tính biến nạp vẫn còn, chứng tỏ prôtêin và ARN không phải là tác nhân biến nạp. Nhưng nếu tế bào S chết bị xử lý bởi ADNaze thì hoạt tính biến nạp không còn nữa.Chứng tỏ ADN là nhân tố biến nạp. Như vậy hiện tượng biến nạp là một bằng chứng sinh hoá xác nhận rằng ADN mang tín hiệu di truyền. Tuy nhiên tại thời điểm đó vai trò của ADN vẫn chưa được công nhận vì trong thí nghiệm vẫn còn một chút prôtêin.
Năm 1952, A.Hershey và M.Chase đã tiến hành thí nghiệm với bacteriophage T2 đánh dấu S35 và P32 đối với E.coli mới chứng minh được chính xác ADN là vật chất chứa đựng thông tin di truyền và truyền đạt thông tin di truyền.
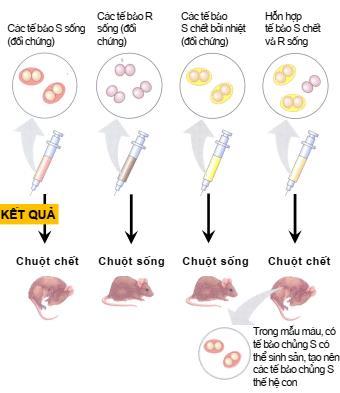
DẠYKÈMQUYNHƠN
414
OFFICIAL
Hình 2. Thí nghiệm của A.Hershey và M.Chase (1952)
2. Cấu tạo axit nucleic
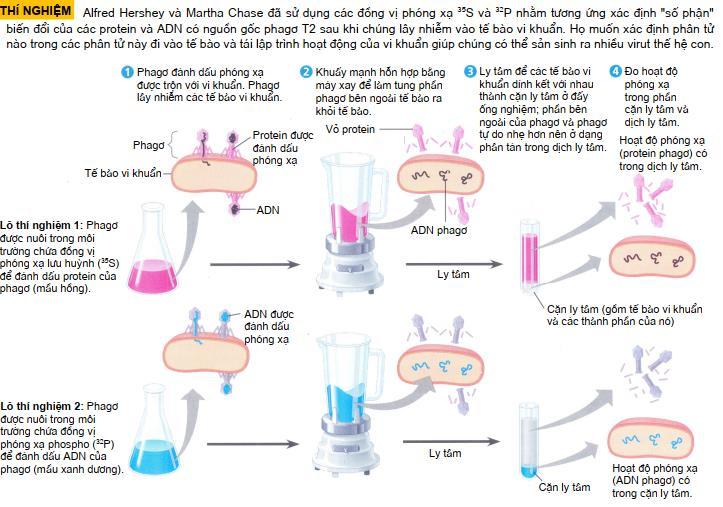
2.1. Cấu tạo hoá học
2.1.1. Nucleotit – đơn phân cấu tạo axit nucleic
a. Thành phần cấu tạo nucleotit
Cả ADN và ARN đều là các chất trùng phân (polymer) mạch dài gồm nhiều đơn phân (monomer) nối với nhau. Trong đó, mỗi đơn phân của ADN là deoxyribonucleotit, còn của ARN là ribonucleotit.
415
DẠYKÈMQUYNHƠN OFFICIAL
Mỗi một nucleotit gồm ba thành phần cơ bản:
+ Bazơnitơ: là các dẫn xuất hoặc của purine, gồm adenine (A) và guanine (G); hoặc của pyrimidine, gồm thymine (T), cytosine (C), và uracil (U);
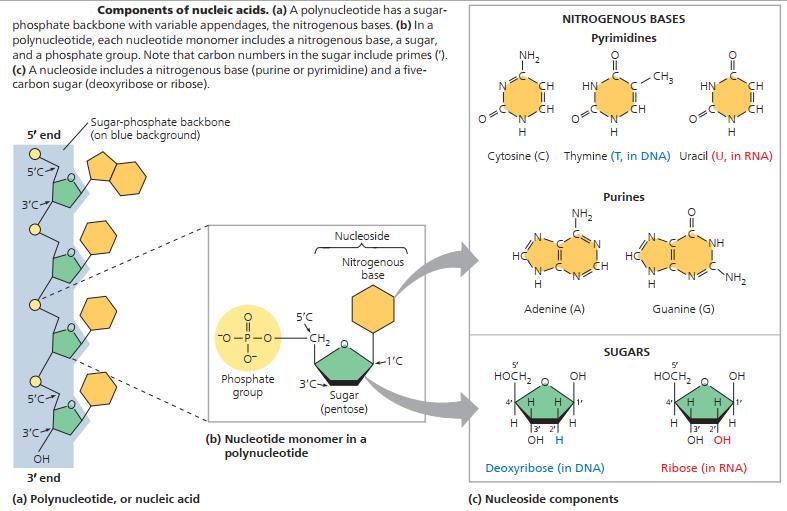
+ Đường pentozơ: Tham gia cấu tạo nucleotit có 2 loại: đường ribose và deoxyribose
+ Axit phot phoric (H3PO4): trong cấu tạo phân tử axit này có 3 gốc axit vì vậy nó có thể kết hợp được với 3 nhóm định chức khác có tính bazơ. Trong cấu trúc của axit nucleic, axit photphoric có 2 nhóm axit liên kết với 2 nhóm OH của 2 phân tử đường trước và sau nó trên mạch polynucleotit và còn lại 1 gốc axit vì vậy chuỗi poly nucleotit mang tính axit Vị trí các nguyên tử cacbon (C) trên mạch vòng của đường pentose được đánh số từ C-1’ đến C-5’. Các nucleotit mang nhóm phosphate ở vị trí C-5’ có vai trò quan trọng đối với cấu trúc và chức năng của ADN và ARN. Sự có mặt của nhóm phosphate ở vị trí này làm các phân tử ADN và ARN thường mang điện tích âm và có tính axit. Trong điều kiện in vivo, nếu các điện tích của axit nucleic không được trung hòa thì nó không thể đóng gói vào nhiễm sắc thể và nhân tế bào được. Sự trung hòa điện tích diễn ra nhờ sự có mặt của các protein có tính kiềm (còn gọi là tính kiềm) xuất hiện trong tế bào ở cả các sinh vật prokaryote (nhân sơ) và eukaryote (nhân thật). Ở eukaryote, đó là các phân tử protein histon mang tính kiềm, còn ở prokaryote là các polyamin. b. Sự liên kết giữa các thành phần tham gia cấu tạo 1 nucleotit.
DẠYKÈMQUYNHƠN
416
OFFICIAL
Các bazơ nitơ gắn với đường pentose bằng liên kết cộng hóa trị ở vị trí C-1’ của đường với nitơ ở vị trí số 9 của purine hoặc nitơ ở vị trí số 1 của pyrimidine
c. Đồng đẳng của nucleotit
Trong cơ thể thường gặp các nucleotit liên kết với enzim này hay enzim khác vốn đóng vai trò đặc biệt quan trọng. Cấu tạo của các nucleotit deoxiriboza này thường đặc trưng bởi sự hình thành liên kết với 1 phân tử axit phôt phoric ở vị trí C5' của đường pentose, ngoài ra trong tế bào còn có các loại nucleotit liên kết với 2 hoặc 3 gốc photphat. Đại diện quan trọng nhất của các nhóm này là adenosine - 5' - monophotphat (AMP), adenosine -5'- diphotphat (ADP) và adenosine - 5' - triphotphat (ATP). ATP là chất mang năng lượng hoá học hoạt hoá chủ yếu ở trong tế bào.
OFFICIAL
tphoríc

DẠYKÈMQUYNHƠN
417
Sơ đồ cấu trúc AMP,
và ATP d. Kích thước và khối lượng của 1 nucleotit - Khối lượng của 1 nucleotit:
() - Kích thước không gian
của 1 nucleotit là
0 () e. Phân loại nucleotit
ấ
Hình 4.
cAMP
300 đvC
trung bình
3,4A
Bazơ nitơric + Đường pentozơ Nucleozit +Axítphố
Nucleotit Hình 3. Sơ đồ c
u tạo 1 nucleotit
Loạ i axit nucleic
Purin Pyrimidin Ad enin(A) Gu anin(G) Cyt ozin(C) Ty min (T) Ura xin (U)
AD N A G C T
AR N A G C U
Chú ý: + Người ta gọi các đơn phân cấu tạo nên axit nucleic là nucleotit + Đơn phân tham gia cấu tạo nên ADN là các nucleotit deoxiriboza + Đơn phân tham gia cấu tạo nên ARN là các nucleotit riboza 2.1.2. Các liên kết hoá học chủ yếu trong cấu trúc phân tử axit nucleic a. Liên kết liên kết cộng hoá trị - Liên kết phốtpho este: Được hình thành giữa nhóm 3’OH ở phân tử đường với 1 nhóm OH của gốc axit photphoric, liên kết này tạo ra 1 phân tử nước. - Liên kết N - glycozit: Được hình thành giữa nhóm 1’OH của phân tử đường với 1NH của bazo nitơ loại pirymidin hoặc 9NH của bazơ nitric loại purin, liên kết này tạo ra 1 phân tử nước.
b. Liên kết hidro
- Liên kết hidro là liên kết được hình thành giữa nguyên tử H đã liên kết với 1 nguyên tố hoá học khác với 1 nguyên tố có độ âm điện lớn (O, N) cũng đã liên kết với 1 nguyên tố hoá học khác.
+ Trong cấu trúc của ADN nó được hình thành giữa các bazơnitric trên 2 mạch đơn của ADN: A liên kết với T bởi 2 liên kết hidro, G = X bởi 3 liên kết hidro.
+ Trong cấu trúc phân tử ARN (rARN và tARN được hình thành giữa các nucleotit trên 1 mạch đơn : A=U, G =X. - Vai trò: Duy trì cấu trúc không gian của axit nucleic. 2.1.3. Sự liên kết giữa các đơn phân tạo thành chuỗi poly nucleotit
DẠYKÈMQUYNHƠN
418
OFFICIAL
- Trên Các nucleotit liên kết với nhau bằng liên kết cộng hoá trị phốtpho dieste
- Chiều liên kết giữa các nucleotit 5’ => 3’,
- Trong cấu trúc phân tử ADN hai mạch đơn có chiều liên kết trái ngược nhau.
OFFICIAL
Hình 5. Sơ đồ cấu tạo 1 đoạn poly nucleotitdeoxyriboza
2.2. Cấu trúc không gian của Axit nucleic
2.2.1. Cấu trúc không gian của ADN
Trong phân tử ADN sợi kép (gọi như vậy vì mỗi phân tử gồm hai mạch đơn polynucleotit), hai mạch polynucleotit liên kết với nhau qua liên kết hydro giữa các nucleotit đối diện trên hai mạch. Liên kết hydro trong ADN hình thành giữa hai mạch thường là A = T và G ≡ C. Trong đó, số gạch nối giữa các nucleotit (A, T, G và C) phản ánh số liên kết hydro giữa chúng; theo đó, giữa A và T có hai liên kết hydro, còn giữa G và C có ba liên kết hydro. Nguyên tắc liên kết này còn gọi là nguyên tắc bổ sung, hay nguyên tắc Chargaff Cấu trúc không gian của ADN (cấu trúc bậc 2) đầu tiên được J. Watson và F.Crick đề sướng vào năm 1953 với nội dung cơ bản như sau: ADN được cấu tạo từ 2 mạch polynucleotit deoxiriboza nằm song song ngược chiều nhau, xoắn lại với nhau theo chu kỳ. Trên mỗi mạch các nucleotit deoxiriboza liên kết lại với nhau nhờ các liên kết phôtphodieste ADN xoắn theo chu kỳ, mỗi chu kỳ xoắn có kích thước là 34A0 gồm có 10 cặp nucleotit. Góc nghiêng giữa các nucleic deoxiriboza cạnh nhau trong từng mạch là 360. Trên mỗi mạch ở mỗi các mononucleotit các base nằm phía trong, còn đường pentose và axit photphoric nằm phía ngoài. Hai mạch đơn polynucleotit deoxiriboza xoắn với nhau xung quanh một trục tưởng tượng và liên kết với nhau thông qua các base chứa nitơ dọc theo phân tử ADN nhờ các liên kết hidro. Các liên kết hidro hướng từ nhóm NH2 của adenin đến nhóm - OH của thymine và từ nhóm - NH2 của guamin đến nhóm - OH của Cytosine. Điều này có nghĩa là trình tự sắp xếp các base chứa nitơ ở một trong hai mạch có thể tuỳ ý, nhưng
DẠYKÈMQUYNHƠN
419
5’ 5’ 3’ 3’
trình tự sắp xếp các base chứa nitơ ở trong mạch thứ hai sẽ phụ thuộc một cách nghiêm ngặt vào trình tự sắp xếp các base chứa nitơ trong mạch thứ nhất. Các cặp AdeninThymine và Guanin - Cytosine là những cặpbổ sung
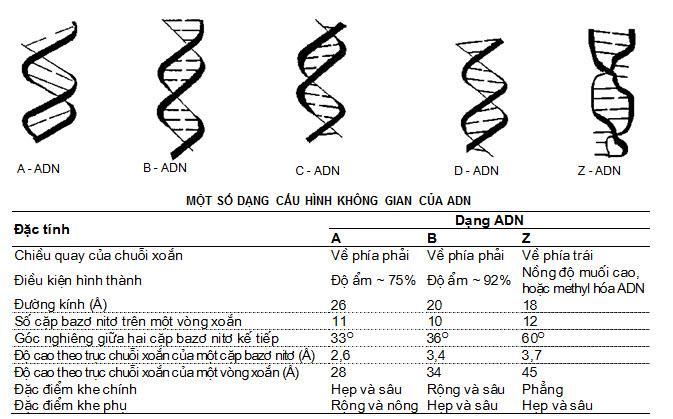
Hình 6. J. Watson và F.Crick với mô hình ADN mà 2 ông xây dựng được Sau này người ta còn phát hiện ra nhiều loại ADN nữa đó là dạng A,C,D và Z, các dạng này có sự khác nhau về chiều quay của chuỗi xoắn kép, đường kính vòng xoắn, số cặp bazơ nitơ …(Hình 42).

DẠYKÈMQUYNHƠN OFFICIAL 420
Trong thực tế mô hình ADN dạng B của Watson và Crick là mô hình cấu trúc phổ biến hơn cả, hai mạch đơn của phân tử ADN sợi kép xoắn xung quanh nhau về phía phải. Cấu trúc xoắn đều đặn như vậy tạo ra hai loại khe trong mỗi vòng xoắn gọi là khe chính (major groove) và khe phụ (minor groove).
Do sự cuộn xoắn của chuỗi ADN sợi kép, khe chính rộng hơn so với khe phụ. Các nghiên cứu sau này trên các đối tượng sinh vật khác nhau cho thấy các protein điều hoà hoạt động của gen thường có xu hướng liên kết với trình tự ADN mà nó điều khiển tại vị trí các khe chính nhờ những trình tự axit amin đặc biệt có khả năng hình thành các liên kết hydro với các bazơ nitơ trên phân tử ADN tại khe chính. Mối tương tác này được tìm thấy tương đối phổ biến ở cả sinh vật nhân sơ và nhân thật. Trong khi đó, các khe phụ có xu hướng là vị trí gắn của các protein cấu trúc thường tham gia vào quá trình đóng gói các phân tử ADN ở sinh vật nhân thật. Những protein này, chẳng hạn như histon, chứa một lượng lớn các axit amin mang tính kiềm (tích điện dương) như Arg và Lys, chúng có khuynh hướng tương tác với nhóm phosphate mang điện âm trên phân tử ADN. Vị trí của các cặp bazơ nitơ trong phân tử ADN không nằm dọc trục đường – phosphate giúp giải thích cho đặc tính hấp thụ ánh sáng cực đại của phân tử ADN ở bước sóng 260 nm (chính chính xác là 257 nm). Tùy thuộc vào một số yếu tố môi trường xung quanh, như độ pH, trạng thái methyl hóa của các nucleotit, độ ẩm, hàm lượng muối, tính ưa nước, lực ion hoá, sự có mặt của các protein liên kết ADN… mà phân tử ADN sợi kép có thể xuất hiện ở một số dạng cấu hình không gian khác nhau. Trong đó, dạng cấu hình phổ biến nhất là dạng B (viết tắt là B-ADN), đây chính là mô hình được Watson và Crick mô tả. Trong tế bào sống, phần lớn phân tử ADN sợi kép ở dạng cấu hình này, nhưng khi độ ẩm giảm đi (≈75%) thì có thể có sự hình thành của cấu hình dạng A (A-ADN); hoặc nếu
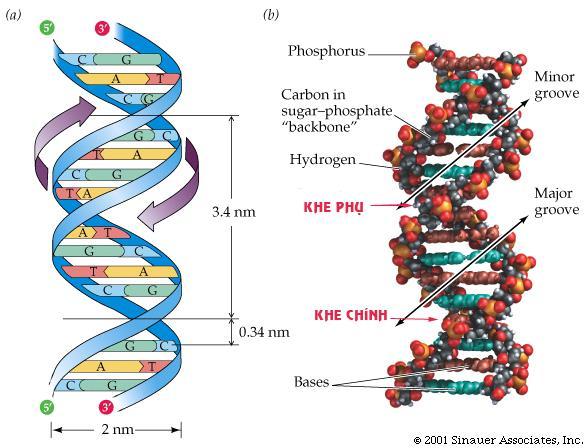
OFFICIAL
DẠYKÈMQUYNHƠN
421
trong môi trường có hàm lượng muối cao hoặc có hiện tượng methyl hóa cytosine, phân tử ADN có thể có cấu hình hiếm gặp hơn là dạng Z (Z-ADN còn gọi là zigzag ADN; hay dạng S-ADN).
Hệ gen của sinh vật nhân sơ nói chung cũng như nhiễm sắc thể của vi khuẩn nói riêng đều nằm trong một vùng tương đối nhỏ. Giống như sinh vật nhân thực, cấu trúc này hình thành nhờ các protein bám ADN đóng gói hệ gen theo một mô hình có tổ chức.
2.2.2 Cấu trúc không gian của ARN
a. mARN
Phân tử mARN có cấu trúc một mạch đơn là bản phiên mã trình tự của gen (thực tế có trình tự giống với mạch mang nghĩa trên phân tử ADN mã hóa cho chuỗi polypeptide tương ứng). Loại ARN này chiếm khoảng 2 - 5 % tổng lượng ARN của tế bào.
Ở sinh vật nhân sơ, phần lớn mARN là bản sao nguyên vẹn trình tự nucleotide được hình thành sau quá trình phiên mã từ phân tử ADN, đồng thời được sử dụng ngay làm khuôn để dịch mã tổng hợp protein.
Ở sinh vật nhân thật, hầu hết các phân tử mARN được hình thành sau phiên mã còn phải trải qua một giai đoạn hoàn thiện (còn gọi là sự chế biến ARN) trong nhân tế bào. Quá trình hoàn thiện mARN gồm 3 sự kiện cơ bản:
+ Lắp “mũ” 7mG (7-methylguanidine triphosphate) vào đầu 5’ tận cùng của phân tử mARN tiền thân (tiền-mARN);
+ Cắt bỏ các đoạn trình tự không mã hoá (intron) và ghép nối các đoạn trình tự mã hoá (exon) với nhau;
+ Lắp ghép vào phần đuôi phân tử mARN bằng trình tự polyA (trình tự chứa từ vài chục đến hàng trăm adenine liên tiếp).
Sản phẩm hình thành sau quá trình hoàn thiện này là phân tử mARN hoàn chỉnh, sẵn sàng cho quá trình dịch mã (tổng hợp protein) diễn ra ở tế bào chất. Kích thước của sợi mARN thường dài từ 900 đến 1200 ribonucleotide, khối lượng trung bình khoảng từ 3x105 đến 4x106 Dal với hệ số lắng dao động trong khoảng từ 6S đến 25S (Hình 7a).
Ở sinh vật nhân thật, phần lớn các phân tử mARN được tổng hợp từ trong nhân tế bào, chỉ trừ một số ít được tổng hợp từ các gen trong ti thể và lạp thể.
b. tARN : Hàm lượng ARN vận chuyển (tARN) trong tế bào chiếm khoảng 10 - 15% tổng lượng ARN. Phân tử tARN thường là một mạch polynucleotide ngắn, chứa từ 75 đến 95 ribonucleotide, khối lượng phân tử khoảng 23 - 30 kDa, hệ số lắng là 4S.
DẠYKÈMQUYNHƠN
OFFICIAL 422
Cấu trúc tARN điển hình gồm 1 mạch polynucleotide cuộn xoắn dạng lá phân thùy (Hình 7b), trong đó có một vài đoạn xoắn kép do các ribonuclotide liên kết bổ sung theo nguyên tắc Chargaff. Trong cấu trúc này, tARN có một thuỳ tiếp nhận axit amin có đầu tận cùng luôn là trình tự CCA. Thuỳ đối mã mang bộ ba đối mã phù hợp với mã bộ ba trên phân tử mARN làm mạch khuôn trong quá trình dịch mã. Ngoài ra còn có 1 hoặc 2 thuỳ phụ tùy vào từng loại tARN. Tuy đến nay vai trò của các thuỳ phụ chưa rõ, nhưng dường như chúng có vai trò làm tăng tính ổn định của các phân tử tARN.
OFFICIAL
Mỗi loại tARN có vai trò vận chuyển một loại axit amin duy nhất. Nhưng một axit amin có thể được vận chuyển bởi nhiều loại tARN khác nhau. Đến nay, đã có trên 300 loại phân tử tARN khác nhau được xác định giữ vai trò vận chuyển 20 loại axit amin cơ bản. Mặc dù các phân tử tARN có thể khác nhau đôi chút về số lượng các ribonucleotide, nhưng chúng có một số đặc điểm cấu hình chung. Theo đó, các phân tử tARN đều là các chuỗi polynucleotide mạch đơn, tự cuộn xoắn bởi các liên kết hydro hình thành giữa các nuleotide bên trong sợi kể từ đầu 5’, đồng thời có một số đặc điểm:
+ Có một nhóm monophosphate ở đầu 5’;
+ Một cấu trúc thùy DHU (còn gọi tắt là thùy DHU) mang bazơ nitơ hiếm gặp dihydrouridin (DHU);
+ Một cấu trúc thùy mang bộ ba đối mã, giúp “dịch” mã bộ ba trên mARN được gọi là “thùy đối mã”;
+ Một cấu trúc thùy bất định, thường có số nucleotide rất khác nhau (từ 3 đến 21 nucleotide) ở các loại phân tử tARN khác nhau;
+ Cấu trúc thùy TΨC mang bazơ nitơ hiếm gặp pseudouridin (kí hiệu Ψ, hoặc ΨU);
+ Phần thân của thùy nhận axit amin thường gồm 7 cặp bazơ nitơ liên kết bổ sung, trong đó có thể có một cặp liên kết bất thường giữa G và U;
+ Một trình tự gồm 3 nucleotide tận cùng đầu 3’ luôn là CCA tìm thấy ở tất cả các loại tARN, trong đó nhóm –OH của A tại đầu 3’ tận cùng chính là vị trí gắn axit amin nhờ hoạt động xúc tác của enzym aminoacyl-tARN synthetase. Enzym này giúp lắp ghép chính xác từng loại axit amin vào phân tử tARN tương ứng vận chuyển nó. Phức hợp tARN sau khi lắp ghép với axit amin được gọi là aminoacyl-tARN (viết tắt là AA-tARN).
c. rARN:
ARN ribosome (Hình 7c) chiếm khoảng 80% tổng lượng ARN trong tế bào. rARN thường có cấu trúc dạng mạch đơn polyribonuleotide với nhiều khúc cuộn, chứa từ dưới 100 đến 1500 ribonucleotide. Các rARN kết hợp với một số phân tử protein đặc biệt tạo thành các ribosome. Ribosome có cấu tạo gồm hai tiểu phần, gọi là tiểu
DẠYKÈMQUYNHƠN
423
phần nhỏ và tiểu phần lớn. Trong quá trình dịch mã hai tiểu phần của ribôxôm kết hợp với nhau tạo thành phức hệ ribosome hoàn chỉnh có hoạt tính dịch mã. Khi kết thúc dịch mã, hai tiểu phần lại tách nhau ra và tồn tại riêng rẽ, lúc này chúng không có hoạt tính dịch mã. Ở sinh vật nhân thật, ribosome ở tế bào chất có hệ số lắng là 80S(). Trong đó, tiểu phần nhỏ có hệ số lắng là 40S được cấu tạo từ một phân tử rARN 18S và khoảng 35 phân tử protein cấu trúc; còn tiểu phần lớn có hệ số lắng là 60S được cấu tạo từ 3 phân tử rARN với hệ số lắng là 5S, 5,8S và 28S kết hợp với khoảng 50 phân tử protein cấu trúc. Như vậy, sinh vật nhân thật có 4 loại rARN. ở sinh vật nhân sơ, ribosome có hệ số lắng là 70S, cũng gồm hai tiểu phần. Tiểu phần nhỏ có hệ số lắng là 30S được cấu tạo từ một phân tử rARN 16S và 21 phân tử protein cấu trúc; tiểu phần lớn có hệ số lắng là 50S gồm hai phân tử rARN có hệ số lắng là 5S và 23S kết hợp với 31 phân tử protein cấu trúc. Như vậy, sinh vật nhân sơ có 3 loại rARN.
OFFICIAL
d) Các loại ARN khác Trong tế bào sinh vật nhân thật, còn có một số ARN có kích thước nhỏ chỉ chứa khoảng 90 - 300 ribonucleotide được gọi là các snARN (small nuclear RNA). Các phân tử snARN không tham gia vào quá trình phiên mã, mà thay vào đó, nó kết hợp với một số protein đặc thù để tạo nên phức hợp tham gia vào việc cắt các intron và nối các exon trong quá trình hoàn thiện phân tử mARN, phức hợp này được gọi là phức hệ cắt intron (spliceosome).
Ngoài snARN, một số loại ARN kích thước nhỏ có hoạt tính giống enzym gọi là ribozyme. Ribozyme thực chất là các phân tử ARN kích thước nhỏ (thường chỉ chứa khoảng 40 - 50 ribonucleotide), có trình tự đặc hiệu và có khả năng xúc tác việc cắt các phân tử ARN ở một số vị trí nhất định. Ngoài ra, trong tế bào còn có một số loại ARN khác như SRP ARN (signal recognition particle RNA, còn gọi là ARN nhận biết tín hiệu), snoARN (small nucleolar RNA, còn gọi là ARN hạch nhân), các ARN tham gia điều hòa hoạt động của gen (siARN và miARN).
DẠYKÈMQUYNHƠN
424
Hình 7a: mARN Hình 7b: tARN Hình 7c. rARN (riboxom) Hình 7. Mô hình cấu trúc
loại ARN 3. Tính chất
trưng của axit nucleic 3.1. ADN
các
đặc
* Số lượng, thành phần và trình tự nucleotit deoxy riboza trên mỗi mạch đơn.
* Tỷ lệ A+T/ G+X. (đặc trưng cho sự bền vững với nhiệt của ADN, hai phân tử ADN có chiều dài bằng nhau, nếu phân tử nào có tỷ số trên lớn thì phân từ đó có số liên kết hidro ít hơn và kém bền với nhiệt hơn)
* Sự phân bố gen và cách mã hoá thông tin di truyền trên phân tử ADN (lượng TTDT)
3.2. ARN
* Số lượng, thành phần và trình tự ribônucleotit trong cấu trúc phân tử
* Cấu trúc không gian của nó
4. Chức năng sinh học của axit nucleic
4.1. Chức năng sinh học của ADN
ở phần lớn các loài sinh vật (chỉ trừ một số virut), ADN có chức năng là vật chất mang thông tin di truyền. Để đảm nhiệm chức năng này, ADN có bốn đặc tính cơ bản sau:
+ Có khả năng lưu giữ thông tin ở dạng bền vững cần cho việc cấu tạo, sinh sản và hoạt động của tế bào.
+ Có khả năng sao chép chính xác để thông tin di truyền có thể được truyền từ thế hệ này sang thế hệ kế tiếp thông qua quá trình phân bào hay quá trình sinh sản.
+ Thông tin chứa đựng trong vật chất di truyền phải được dùng để tạo ra các phân tử cần cho cấu tạo và hoạt động của tế bào.
+ Vật liệu di truyền có khả năng biến đổi, nhưng những thay đổi này (đột biến) chỉ xảy ra ở tần số thấp.
Trong các tế bào sinh vật nhân thật, phần lớn các thông tin di truyền cần thiết của tế bào được lưu giữ trong nhân, chỉ một phần được lưu giữ trong ti thể (cơ quan tử có vai trò hô hấp tế bào, tổng hợp ra phần lớn ATP) hoặc lạp thể (cơ quan quang hợp ở thực vật).
4.2. Chức năng sinh học của ARN
Loại ARN
mARN
tARN
Chức năng sinh học
Truyền thông tin qui định trình tự axit amin của protein từ ADN tới ribosome
Dịch các mã bộ ba trên phân tử mARN thành các axit amin trên phân tử protein
rARN Cấu trúc ribosome và có vai trò xúc tác (ribozyme) hình
DẠYKÈMQUYNHƠN
OFFICIAL 425
TiềnARN
thành liên kết peptide
Sản phẩm trực tiếp của quá trình phiên mã; là phân tử tiền thân hình thành nên mARN, tARN và rARN hoàn thiện. ở eukaryote, một số phân đoạn ARN intron có vai trò xúc tác (ribozyme) phản ứng cắt chính nó
snARN
SRP ARN
sno
ARN
Telomer ase-ARN
Có vai trò xúc tác và cấu trúc trong phức hệ cắt intron (spliceosome) từ các phân tử tiền mARN để tạo thành mARN hoàn thiện.
Là thành phần của phức hệ ARN-protein làm nhiệm vụ nhận biết các peptide tín hiệu trong phân tử protein mới được tổng hợp, giúp "giải phóng" các phân tử protein này khỏi mạng lưới nội chất.
Tham gia hoàn thiện rARN từ phân tử tiền rARN và đóng gói ribosome tại hạch nhân.
Thành phần của enzym telomerase; làm khuôn để tổng hợp trình tự ADN lặp lại tại các đầu mút nhiễm sắc thể ở eukaryote.
gARN Tham gia vào quá trình "biên tập" ADN ti thể ở thực vật và nguyên sinh động vật, và ADN lạp thể ở thực vật.
tmARN ARN tích hợp chức năng của tARN và mARN, giúp giải phóng ribosome khỏi sự "tắc nghẽn" khi dịch mã các phân tử mARN bị mất bộ ba mã kết thúc (stop codon).
M1 ARN
Các loại ARN can thiệp (siARN và miARN)
Thành phần ARN có vai trò xúc tác của ARNase P, tham gia hoàn thiện các phân tử tARN ở prokaryote.
Tham gia điều hòa biểu hiện gen ở eukaryote.
Xét về cấu trúc, sở dĩ ARN có thể đảm nhận nhiều chức năng khác nhau trong tế bào (khác với ADN) là do chúng có ba tính chất:
+ ARN có thể tạo liên kết hydro với các phân tử ADN hoặc ARN khác,
+ Mỗi phân tử ARN có cấu hình đặc thù do liên kết hydro hình thành giữa các phần khác nhau của nó,
+ Các ARN có thể mang nhiều nhóm chức khác, nên ARN có thể có hoạt tính xúc tác như các enzym.
Việc ngày càng phát hiện ra nhiều chức năng khác nhau của ARN là cơ sở dẫn đến quan điểm cho rằng: ARN có thể là đại phân tử sinh học đầu tiên xuất hiện trong quá trình tiến hóa (chứ không phải là ADN hay protein như nhiều giả thiết trước đây).
426
DẠYKÈMQUYNHƠN
OFFICIAL
Có thể tưởng tượng về một dạng sống nguyên thủy chỉ có ARN. Ở dạng sống này, ARN vừa có vai trò là vật chất mang thông tin di truyền, vừa biểu hiện chức năng của enzym. Dạng sống đó dần biến đổi thành thế giới sống dựa trên ba loại đại phân tử là ADN, ARN và protein. Một bằng chứng ủng hộ quan điểm protein xuất hiện sau ARN là chức năng của ARN trong ribosome. Trong ribosome, chính thành phần có vai trò xúc tác hình thành liên kết peptide (enzym peptidyl transferase) là ARN. Điểm khác biệt giữa petidyl transferase với các loại ribozyme khác (như ARNase P) là các ribozyme khác đều gây tác động xúc tác lên trung tâm phản ứng là phospho, còn petidyl transferase gây hiệu ứng xúc tác lên trung tâm phản ứng là cacbon, qua đó hình thành liên kết peptide. Có thể ribozyme của ribosome là một “vết tích” sót lại của dạng sống nguyên thủy vốn chỉ có các enzym ARN.
5. Hệ gen của virut
Vật chất di truyền hay hệ gen của virut có các đặc điểm sau đây:
- Có thể là ADN hoặc ARN mà không bao giờ chứa cả hai.
OFFICIAL
- Có dạng chuỗi đơn hoặc chuỗi kép, dạng thẳng hoặc dạng vòng.
- Hệ gen ADN kép thường có kích thước lớn, hệ gen ARN đơn thường có kích thước nhỏ.
- Tất cả virut ARN kép đều có hệ gen phân đoạn (trong mỗi hạt chứa nhiều đoạn). Virut ARN đơn không phân đoạn, ngoại trừ virut cúm.
- Hệ gen đơn của virut được chia làm 2 loại: dương và âm. Nếu trình tự của hệ gen trùng với trình tự của mARN thì quy ước là hệ gen dương, còn ngược lại (tương bù) gọi là hệ gen âm.
Một số virut chứa enzim phiên mã ngược:
- Virut viêm gan B (HBV) có hệ gen là ADN kép khép vòng, 2 sợi không bằng nhau, sợi dài là (-), sợi ngắn là (+). Khi nhân đôi phải thông qua sự tạo thành ARN trung gian (ADN → ARN → ADN).
- Virut retro (ví dụ HIV) có hệ gen là 2 sợi ARN giống nhau. Khi nhân đôi phải thông qua sự tạo thành ADN trung gian (ARN → ADN → ARN).
DẠYKÈMQUYNHƠN
427
II. Cơ chế di truyền vi sinh vật 1. Tái bản DNA (DNA replication) ở vi sinh vật. 1.1. Các nguyên tắc và đặc điểm chung của tái bản DNA ở vi sinh vật - Tái bản theo kiểu bán bảo toàn (semi-conservative). - Tái bản bắt đầu tại một hoặc nhiều khởi điểm
Từ khởi điểm, DNA
ở xoắn tạo thành hai chạc tái bản (replication fork). Cấu trúc như vậy gọi
bản (replicon). DNA E. coli
y có cấu
ống
ữ
(Ori).
m
là đơn vị tái
trong quá trình tái bản như vậ
trúc gi
ch
cái
theta Hy Lạp (θ) nên gọi là tái bản theta. Đối với các DNA mạch vòng, mỗi phân tử chỉ có một Ori; trong khi đó mỗi DNA nhiễm sắc thể eukaryote có nhiều Ori.
- Tham gia vào sự tái bản DNA có nhiều protein và enzyme.
- Tại mỗi chạc tái bản, trước tiên xảy ra sự tổng hợp các đoạn mồi RNA (primer) bởi vì các DNA polymerase tự nó không thể bắt đầu tổng hợp mới được. Mặt khác, do hai sợi đơn của mỗi chạc ngược chiều nhau trong khi các DNA và RNA polymerase chỉ xúc tác theo chiều 3'→ 5', cho nên sự tái bản DNA trên hai sợi khuôn là không giống nhau: một sợi liên tục gọi là sợi dẫn đầu (leading strand) và một sợi không liên tục gọi là sợi ra chậm (lagging strand). Kiểu tái bản như thế gọi là tái bản nửa gián đoạn (semi-discontinuous), được R. Okazaki phát hiện đầu tiên năm 1969.
OFFICIAL
Hình 8: Một khởi điểm và hai chạc tái bản sinh trưởng đồng thời theo hai hướng đối lập nhau, mỗi chạc gồm một sợi dẫn đầu và một sợi ra chậm.
Lưu ý: Tất cả các DNA polymerase đều cần có mồi để xúc tác tổng hợp chuỗi theo chiều 5'→3'; và chỉ có một số enzyme này là có hoạt tính đọc sửa (proofreading activity). Khác với E. coli (có ba loại DNA polymerase I, II và III; trong đó Pol II không tham gia tái bản), các tế bào eukaryote có năm loại DNA polymerase (α, β, γ, δ và ε), trong đó ba loại chịu trách nhiệm tái bản DNA nhân là các polymerase α, δ và ε. Polymerase δ chịu trách nhiệm chính cho tổng hợp ở sợi dẫn đầu và polymerase α cho sợi ra chậm. Polymerase γ tái bản DNA của các bào quan ty-lạp thể, còn polymerase β chịu trách nhiệm sửa chửa DNA.
1.2. Cơ chế tái bản ở vi khuẩn Ở vi khuẩn, thông tin di truyền được truyền một chiều từ thể cho sang thể nhận và tạo ra hợp tử từng phần. Tái tổ hợp ở vi khuẩn có thể thực hiện bằng các đoạn ADN trần trong biến nạp, hay phage trong tải nạp, nhờ giao nạp khi 2 tế bào khác giới tính gắn với nhau. - Đặc điểm + Truyền thông tin một chiều từ tế bào cho (donor) sang tế bào nhận (recipient).
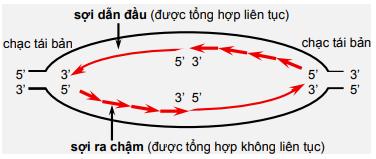
DẠYKÈMQUYNHƠN
428
+ Sự tạo thành hợp tử từng phần (merozygote). Thể cho (donor) chỉ chuyển một đoạn của bộ gen sang thể nhận (recipient) nên chỉ lưỡng bội ở một phần, các phần khác đơn bội.
+ Bộ gen thường chỉ là một ADN trần nên chỉ có một nhóm liên kết gen và tái tổ hợp thực chất là giữa hai phân tử ADN.
- Sinh sản ở vi khuẩn
Tế bào vi khuẩn phân chia theo lối trực phân. Phân tử ADN gắn trực tiếp vào màng sinh chất. Sự sao chép ADN tạo ra hai bản sao gắn chung nhau trên màng sinh chất. Khi tế bào kéo dài ra, các bản sao ADN tách xa nhau do phần màng giữ chúng lớn dần ra. Kiểu sinh sản vô tính này được gọi là ‘phân đôi”hay “ngắt đôi” (“binary fission”) (hình 9). Tế bào vi khuẩn chia nhanh (20 phút trong điều kiện tốt) hơn rất nhiều so với tế bào Eukaryota (24-48 giờ).
OFFICIAL
Hình 9: Trực phân ở vi khuẩn Quá trình sao chép ADN được bắt đầu từ điểm xuất phát oriC kéo dài về hai phía song song với quá trình sao chép màng sinh chất, nơi có điểm gắn vào của ADN bộ gen, mọc dài tách 2 phân tử ADN về 2 tế bào con. ADN của E.coli cần 40 phút cho 1 vòng sao chép tương ứng với tốc độ 50.000 cặp bazơ/phút. Phụ thuôc vào tốc độ tăng trưởng, thời gian phân chia tế bào trong khoảng từ 18 đến 60 phút. Như vậy ở các tế bào tăng trưởng nhanh, vòng sao chép mới phải được bắt đầu sớm hơn sự phân bào trước đó như tế bào con đầu tiên.
- Cơ chế tái bản ADN (E. coli)
DẠYKÈMQUYNHƠN
429
E.coli chỉ có 1 sợi NST duy nhất chứa khoảng 4,6 triệu cặp nu, mỗi tế bào E.coli có thể sao chép toàn bộ phân tử AND và chia đều cho các tế bào con trong vòng 1 giờ + Giai đoạn khởi đầu (initiation) Đối với nhiễm sắc thể E. coli, sự tái bản bắt đầu tại một khởi điểm đặc thù gọi là Ori.
(1) Các protein bám khởi điểm dnaA bám Ori tạo ra cấu trúc nucleoprotein chuyên hoá của khởi điểm;
(2) Cấu trúc này mở xoắn vùng DNA giàu AT để hình thành "phức hợp mở";
(3) Hai phân tử helicase dnaB chui vào phức hợp mở làm mở xoắn khởi điểm theo cả hai hướng, tạo thành hai chạc tái bản (replication fork). Khi cả hai sợi đơn của mỗi chạc được tách ra thì các protein SSB bám vào.
Hình 10: Hình thành phức hợp mở đầu (primasome) tại khởi điểm với việc tổng hợp đoạn mồi đầu tiên ở một chạc tái bản. Khởi đầu trong tái bản DNA ở E. coli là sự tổng hợp một đoạn mồi ngắn khoảng 10-12 nucleotide bởi primase (xét chung ở các sinh vật là ~ 5 base). Cả ba loại protein dnaB, dnaC và dnaG hợp thành một phức hợp có tên là primasome (hoặc primosome: thể mở đầu; hình 3).
+ Giai đọan kéo dài (elongation)
Một khi primasome tổng hợp xong một mồi, sự kéo dài chuỗi DNA được bắt đầu bằng một phức hợp giữa primasome và DNA polymerase III hoàn chỉnh, gọi là replisome (thể tái bản). Sự tái bản DNA trên mỗi chạc xảy ra theo kiểu nửa gián đoạn (semi-discontinuous), như (hình 4)
Trên sợi khuôn dẫn đầu (3'→5'): Sau khi primase (primasome) tổng hợp xong đoạn mồi RNA với đầu 3'-OH tự do, enzyme hoàn chỉnh DNA polymerase III (hay replisome) bắt đầu kéo dài chuỗi DNA mới sinh trưởng theo chiều 5'→3' một cách liên tục.
Trên sợi khuôn ra chậm (5'→3'): Sự kéo dài diễn ra không liên tục dưới dạng các đoạn Okazaki. Kích thước trung bình mỗi đoạn Okazaki ở E. coli là 1.000 - 2.000 nucleotide. Quá trình này đòi hỏi sự "mồi hóa" lặp lại, với sự tham gia lần lượt của bốn enzyme sau: (i) primase tổng hợp một đoạn mồi RNA; (ii) DNA polymerase III kéo dài đoạn Okazaki;
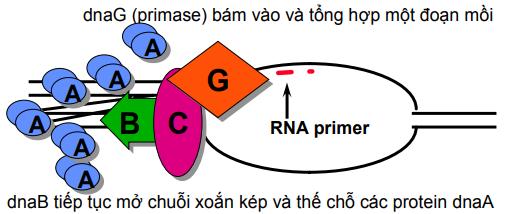
DẠYKÈMQUYNHƠN
OFFICIAL 430
(iii) DNA polymerase I vừa cắt bỏ đoạn mồi vừa lấp khoảng trống bằng cách kéo dài dần đoạn Okazaki theo sau;
(iv) DNA ligase hàn liền khe hở giữa hai đoạn Okazaki bằng một liên kết phosphodiester.
+ Phức hệ sao chép AND: nhiều protein khác nhau, không di chuyển dọc AND, mà AND sẽ chui qua phức hệ khi sao chép.
+ Đọc và sửa chữa AND: nhờ E. AND Pol. Ở E.coli có khoảng 100 E sửa chữa AND. Sửa chữa dựa trên NTBS.
Sửa chữa bằng cắt bỏ nucleotit nhờ E cắt AND- nuclease sau đó lấp đầy chỗ trống nhờ AND-Pol và AND-ligase.
+ Giai đọan kết thúc (termination)
Do cấu trúc nhiễm sắc thể ở hai nhóm prokaryote và eukaryote là hoàn toàn khác nhau, nên cơ chế kết thúc tái bản của chúng cũng khác nhau.
OFFICIAL
Cả hai chạc tái bản của DNA E. coli được bắt đầu từ một khởi điểm (ori), và di chuyển hầu như cùng tốc độ theo hai hướng đối lập nhau xung quanh nhiễm sắc thể mạch vòng cho tới khi chúng gặp nhau tại một điểm kết thúc chung đối diện với ori. Theo Bastia và cộng sự (1997) cũng như nhiều tác giả khác, đây là vùng chứa các trình tự đặc thù gọi là các điểm kết thúc tái bản (replication termini). Tại các trình tự này có các protein kết thúc tái bản (replication terminator protein = RTP) bám vào và các phức hợp protein-DNA này ngăn cản sự di chuyển của các chạc tái bản. Đó là bước đầu tiên của sự hoàn thành một vòng tái bản, và sau đó tách hai nhiễm sắc thể con rời ra một cách có trật tự nhờ xúc tác của topoisomerase IV.
RTP của E. coli có TLPT ~36kD, được mã hóa bởi gene tus (ter) và trong mỗi tế bào có ~80 bản sao của protein này được duy trì hầu như ổn định trong suốt chu kỳ tế bào. Trình tự điều hoà của vùng kết thúc tái bản ở E. coli (R6K), theo Bastia và cộng sự (1997), như sau: 5'NN(A/T)(A/T)(A/T)G(A/T)(A/G)TGTTGTAACTA(A/C)NN3'
Vấn đề kết thúc tái bản ở eukaryote: Mỗi nhiễm sắc thể eukaryote chứa một phân tử DNA sợi kép mạch thẳng kết hợp với nhiều loại protein, có các đầu mút đặc trưng gọi là telomere. Các telomere có cấu trúc đơn giản gồm những trình tự ngắn (6-8 bp) lặp lại nối tiếp cả ngàn lần và đặc thù cho từng loài. Chẳng hạn, ở Tetrahymena là (TTGGGG)n. Một khi đọan mồi đầu tiên trên mỗi sợi được loại bỏ thì nó không có cách nào để bù đắp lại khoảng trống đó, bởi vì DNA không thể nào nới rộng theo chiểu 3'→5'. Các kết quả nghiên cứu đầu tiên của Elizabeth Blackburn và cs đã giải đáp cho vấn đề này. Các đoạn lặp này được gắn thêm vào đầu 3' của các sợi DNA nhờ sự xúc tác của telomerase, một enzyme phiên mã ngược (reverse transcriptase). Chẳng hạn, telomerase của Tetrahymena có một RNA dài 159 nucleotide có chứa trình tự 3'CAACCCCAA5' làm khuôn cho tổng hợp các đoạn lặp 5'-TTGGGG-3'. Vai trò của các Enzyme(E) tái bản AND ở vi khuẩn E.coli
DẠYKÈMQUYNHƠN
431
S TT Enzym Gen mã hóa Chức năng
1 ADN polymeaz I polA-E; mutD. polA
Là E phục hồi chủ yếu. Cắt mồi ARN, lắp chỗ trống
2 ADN polymeaz II holA-E; mutD. Enzym phục hồi phụ (minor).
3 ADN polymeaz III polC; ADNE,Q,N,X; Enzym polyme hoá chủ yếu
4 ADN polymeaz IV polC Phục hồi cấp cứu (SOS repair).
5 ADN polymeaz V ADNB umuD’2C Phục hồi cấp cứu (SOS repair).
432
DẠYKÈMQUYNHƠN OFFICIAL
6 Helicase ADNB Tháo xoắn tại chạc ba sao chép
7
Primase ADNG
8 Protein gắn điểm khởi sự (Origin-binding protein).
9 Protein căng mạch SSB (Singlestrand binding protein)
1 0
ADNA
Tạo mồi mạch ADN mới
Gắn điểm khởi sự sao chép (Ori); tạo thuận lơi cho mở tách mạch.
SSB
Ngăn các mạch đơn đã tách không chập lại.
ADN ligaz ligA, ligB
Nối các đầu hở trên ADN
2. Cơ chế phiên mã ở vi sinh vật 2.1. Sơ lược về các gene Một cách tương đối, gene (cistron) là một đọan xác định của bộ gene mã hóa thông tin của một polypeptid hoặc một phân tử RNA chức năng. Ở các prokaryote và eukaryote bậc thấp, thường có một mối quan hệ đơn giản giữa gene và sản phẩm của nó (một gene - một sản phẩm). Hơn nữa, các gene đồng nghĩa với vùng mã hóa hay khung đọc mở (open reading frame). Nghĩa là, ở các prokaryote các gene liên quan về chức năng thường được tổ chức trong một operon, vì thế có nhiều sản phẩm được dịch mã từ một mRNA polycistron. Trái lại, ở các eukaryote, các gene đồng nghĩa với đơn vị phiên mã (transcription unit) và hầu hết chúng được phiên mã dưới dạng mRNA monocistron. Ở các bộ gene eukaryote bậc cao, thường có một mối quan hệ phức tạp giữa gene và sản phẩm. Hầu hết các gene đều chứa các intron (các đoạn không mã hóa protein) nằm xen giữa các exon (các đoạn mã hóa protein). Các gene như vậy được gọi là gene phân đoạn (split gene), được Phillip Sharp phát hiện đầu tiên năm 1977.
2.2. Các nguyên tắc và đặc điểm chung của phiên mã
Phiên mã (transcription) là quá trình tổng hợp các RNA khác nhau từ thông tin di truyền chứa đựng trong DNA. Trừ các gene mã hóa protein trong các operon ở vi khuẩn, nói chung, các RNA mới được tổng hợp chỉ là các bản sao sơ cấp (primary transcript) gọi là các pre-RNA. Các pre-RNA này phải trải qua một quá trình sửa đổi để trở thành các RNA trưởng thành (mature) trước khi tham gia vào quá trình sinh tổng hợp protein của tế bào. Quá trình phiên mã các đặc điểm chung sau đây
DẠYKÈMQUYNHƠN
433
OFFICIAL
Hình 11: Phiên mã và dịch mã ở sinh vật nhân sơ.
(i) Diễn ra dưới tác dụng của các enzyme RNA polymerase.
(ii) Chỉ một sợi đơn được dùng làm khuôn cho tổng hợp RNA, gọi là sợi khuôn, sợi mã hoá hay sợi có nghĩa (template/ coding / sense strand); còn sợi bổ sung được gọi là sợi đối khuôn, sợi không mã hoá hay sợi đối nghĩa (antitemplate/ noncoding / antisense strand).
(iii) Phản ứng tổng hợp RNA diễn ra theo nguyên tắc bổ sung và được kéo dài theo chiều 5'→3', ngược với chiều của sợi khuôn.
(iv) Nguyên liệu cho tổng hợp gồm: ATP, UTP, GTP và CTP.
(v) Sản phẩm của phiên mã là các RNA sợi đơn.
(vi) Khởi đầu và kết thúc phiên mã phụ thuộc vào các tín hiệu điều hoà là các trình tự DNA đặc thù nằm trước gen (vùng khởi động) và sau gene.
2.3. RNA polymerase và vùng khởi động (promoter) của VSV nhân sơ
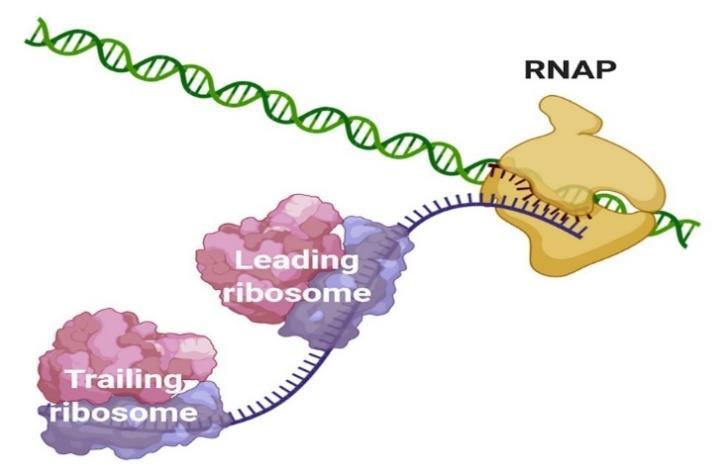
Hình 12: Cấu trúc promoter của prokaryote. Ở các prokaryote, RNA polymerase hoàn chỉnh (holoenzyme) là một phức hợp gồm nhân tố sigma (σ) và lõi enzyme. Nhân tố σ giúp RNA polymerase nhận biết và bám vào promoter để có thể bắt đầu phiên mã tại vị trí chính xác, và lõi enzyme đóng vai trò xúc tác tổng hợp RNA.
Vùng khởi động (promoter) nói chung nằm kề trước gene và có chứa các trình tự đặc thù cho phép RNA polymerase nhận biết và bám vào. Trình tự quan trọng nhất của promoter là hộp TATA hay hộp Pribnow (Pribnow box) ở vị trí "-10". Ngoài ra, còn có trình tự TTGACA ở vị trí ''-35'', gọi là đoạn nhận biết (recognition sequence). Các vùng này được bảo tồn cao.
2.4. Các giai đoạn của quá trình phiên mã ở prokaryote
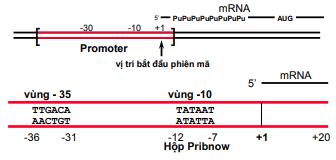
DẠYKÈMQUYNHƠN
434
OFFICIAL
- Khởi đầu: RNA polymerase holoenzyme nhận biết và bám vào promoter, tháo xoắn một đoạn khoảng 12 cặp base. Sau khi tổng hợp ribonucleotide đầu tiên (Appp hoặc Gppp), nhân tố sigma tách ra để đi vào một chu kỳ phiên mã khác, gọi là chu kỳ sigma (sigma cycle).
- Kéo dài: Enzyme lõi tổng hợp sợi RNA dọc theo sợi khuôn.
- Kết thúc: Khi phiên mã xong hai vùng giàu GC và AT nằm sau gene, ở vùng đuôi của sợi RNA hình thành cấu trúc ''kẹp cài tóc'' (hairpin loop) làm dừng sự phiên mã của lõi RNA polymerase. Cuối cùng, dưới tác dụng của nhân tố rho (ρ), sợi RNA và enzyme lõi được giải phóng khỏi DNA khuôn.
3. Dịch mã Cơ chế của quá trình dịch mã (tổng hợp polypeptide) - Mở đầu (initiation): Quá trình dịch mã bắt đầu khi một tiểu đơn vị ribosome bé bám vào mRNA tại vị trí của codon khởi đầu AUG. Lúc này một phân tử tRNA khởi đầu đặc thù mang methionine (ở vi khuẩn là formyl-Met) đi vào và khớp anticodon của nó với codon mở đầu của mRNA. Kế đó, tiểu đơn vị ribosome lớn bám vào tiểu đơn vị bé tạo ra một ribosome hoạt động hoàn chỉnh. Lúc này Met-tRNA ở vị trí P và vị trí A để trống; một tRNA thứ hai (ví dụ, tRNAVal) đi vào vị trí A và khớp với codon thứ hai (hình 13A).
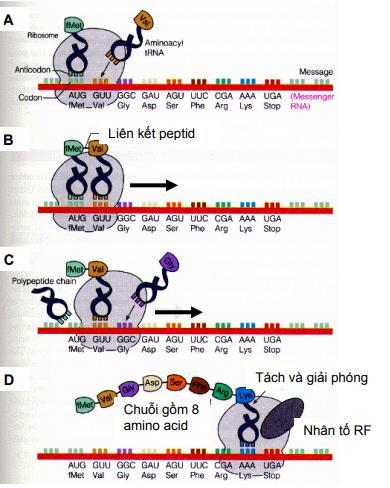
OFFICIAL
DẠYKÈMQUYNHƠN
435
Hình
13: Cơ chế dịch mã
- Kéo dài (elongation): Quá trình kéo dài bắt đầu sau khi liên kết peptide đầu tiên được hình thành. Phản ứng này được xúc tác bởi enzyme peptidyl transferase, và kết quả là tạo ra một peptidyl-tRNA ở vị trí A (hình 13.B). Sau đó, ribosome lập tức chuyển dịch sang một codon mới dọc theo mRNA theo chiều 5'→3' (hình 13.C). Phản ứng này đẩy phân tử tRNA tự do vốn ở vị trí P ra ngoài; lúc này peptidyl-tRNA nằm ở vị trí P và vị trí A lại để trống. Một chu kỳ dịch mã mới lại bắt đầu, một aminoacyl-tRNA thứ ba đi vào và khớp anticodon của nó với codon đang để trống ở vị trí A, một liên kết peptide thứ hai được hình thành, và ribosome lại dịch chuyển sang codon kế tiếp. Quá trình nói trên cứ diễn ra một cách tuần tự dọc theo mRNA làm cho chuỗi polypeptide dài dần ra cho đến dịch mã xong codon có nghĩa cuối cùng.
- Kết thúc (termination): Quá trình tổng hợp polypeptide sẽ dừng lại khi codon kết thúc của mRNA đối diện với vị trí A. Lúc này nhân tố giải phóng RF (release factor) đi vào (Hình 13.D); chuỗi polypeptide được tách ra và phóng thích cùng với hai tiểu đơn vị ribosome cũng như tRNA ra khỏi mRNA. ➢ Lưu ý:
OFFICIAL
(1) Thực ra, trên một mRNA có rất nhiều ribosome cùng hoạt động, gọi là polysome, tạo ra nhiều polypeptide giống nhau.
(2) Trên nguyên tắc, amino acid mở đầu sẽ được cắt bỏ khỏi chuỗi polypeptide. Tuy nhiên, ở các eukaryote không phải lúc nào amino acid mở đầu này cũng bị tách bỏ, mà trong một số protein nó vẫn được giữ lại.
(3) Sau tổng hợp, các chuỗi polypeptide sẽ được sửa đổi và chuyển sang các bậc cấu trúc cao hơn để trở thành các protein chức năng. Thực ra sự biến đổi sau dịch mã còn có các chaperone và nhiều cơ chế tác động phức tạp khác nữa.
(4) Tham gia vào các bước mở đầu, kéo dài và kết thúc còn có các yếu tố protein, với tên gọi tương ứng là các nhân tố mở đầu, kéo dài, và giải phóng cùng với ATP, GTP và các ion Mg2+, K+ và NH4 +
(5) Trong các tế bào prokaryote, các ribosome và các aminoacyl-tRNA sẽ bám vào đầu 5' của mRNA để bắt đầu quá trình dịch mã trong khi ở đầu 3' của nó quá trình phiên mã đang còn tiếp diễn. Ngược lại, ở các tế bào eukaryote, các pre-mRNA phải trải qua sửa đổi sau phiên mã ở trong nhân, còn dịch mã diễn ra sau đó trong tế bào chất.
(6) Về RNA đối nghĩa (antisense RNA), đây là loại RNA thấy có ở nhiều hệ thống, nhưng rất phổ biến ở các vi khuẩn. Nó được tổng hợp từ sợi đối nghĩa của gene, nên bổ sung với mRNA và có thể tạo thành một sợi kép với nó để gây kìm hãm dịch mã. Vì vậy RNA đối nghĩa còn được gọi là RNA bố sung gây nhiễu mRNA, và được ứng dụng hiệu quả trong điều trị ung thư.
DẠYKÈMQUYNHƠN
436
4. Tái tổ hợp di truyền ở vi khuẩn 4.1. Hiện tượng biến nạp ở vi khuẩn Diplococus pneumoniae
Hình 14: Hiện tượng biến nạp
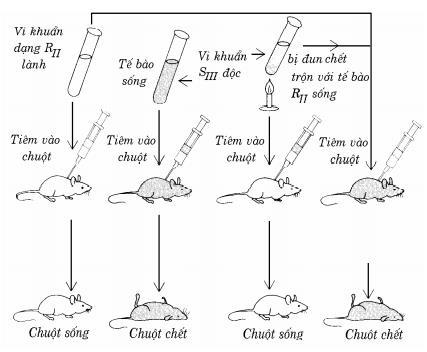
Tiêm vi khuẩn S sống gây bệnh cho chuột - chuột chết.
Tiêm vi khuẩn R sống không gây bệnh - chuột sống.
Tiêm vi khuẩn S bị đun chết cho chuột - chuột sống.
Hỗn hợp vi khuẩn S bị đun chết trộn với vi khuẩn R sống đem tiêm cho chuộtchuột chết. Trong xác chuột chết có vi khuẩn S và R.
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các tế bào chết này đã truyền tính gây bệnh cho tế bào R. Nó được gọi là biến nạp (transformation).
Năm 1944, T.Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu các tế bào S bị xử lý bằng proteaz (enzym phân hủy protein) hoặc ARN-az (enzym phân hủy ARN) hoạt tính biến nạp vẫn còn, chứng tỏ protein và ARN không phải là tác nhân gây biến nạp. Nhưng nếu tế bào S chết bị xử lý bằng ADN-az (enzym chỉ phân hủy đặc hiệu ADN) thì hoạt tính biến nạp không còn nữa, chứng tỏ ADN là nhân tố biến nạp. Kết quả thí nghiệm có thể tóm tắt như sau: ADN của S + các tế bào R sống → chuột → chết (có R + S) Như vậy, biến nạp là hiện tượng truyền thông tin di truyền bằng ADN. Trong biến nạp, ADN trần từ một tế bào vi khuẩn (thể cho) này được truyền sang tế bào vi khuẩn khác (thể nhận). Biến nạp xảy ra khi vi khuẩn nhận ADN ngoại lai và hấp thu vào trong tế bào. Khi tế bào vi khuẩn bị vỡ do bị tan (lysis), ADN vòng tròn của chúng thoát ra môi trường thành các đoạn thẳng với chiều dài khác nhau, có khả năng gây biến nạp cho các tế bào nhận khác. Cơ chế:
DẠYKÈMQUYNHƠN
437
OFFICIAL
+ Sự phân hủy ADN tế bào cho: ADN tế bào cho có thể là của tế bào tự nhiên bị phân hủy hoặc trong thí nghiệm bị gây chế bằng nhiệt độ cao hay tác nhân phá vỡ tế bào.
+ ADN bám vào bề mặt tế bào: Protein gắn vào ADN.
OFFICIAL
+ Thâm nhập của ADN: Sợi ADN mạch kép của dòng vi khuẩn S sau khi chui qua màng tế bào của dòng R thì một mạch S sẽ bị nucleaz của tế bào cắt, còn lại một mạch nguyên. + Bắt cặp (Synapsis)và tái tổ hợp: Nhờ sự hỗ trợ của protein RecA ADN của thể nhận R sẽ biến tính tách rời 2 mạch ở 1 đoạn dễ bắt cặp với đoạn ADN thể cho S vừa chui vào. Hình 15: Sơ đồ diễn biến của quá trình biến nạp ở Vi khuẩn 4.2. Tải nạp ở vi khuẩn
+ Trong tải nạp (transduction), các virut mang các gen từ tế bào này sang tế bào khác. Ở chu trình tan (lytic cycle), một số bacteriophage gói nhầm ADN vi khuẩn chủ vào capsid. Tế bào bị nhiễm bởi các virut như vậy nhận đoạn ADN của vi khuẩn A khác, chứ không phải ADN của virut. Do vậy, ADN vi khuẩn A tái tổ hợp với ADN nhiễm sắc thể của tế bào chủ B và biến đổi thành phần di truyền (gentic composition).
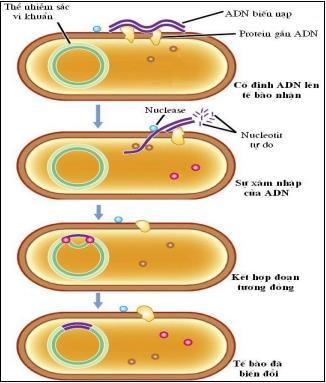
+ Phage là nhân tố chuyển gen
Thí nghiệm được tiến hành trong ống hình chữ U, ở đáy ống được ngăn cách bằng màng lọc vi khuẩn. Màng có lỗ nhỏ vi khuẩn không qua được, nhưng phage qua được (hình 7). Nhánh A của ống chứa vi khuẩn có khả năng tổng hợp tryptophan (trp+ ), còn nhánh B nuôi các vi khuẩn mất khả năng tổng hợp tryptophan (trp– ). Sau một thời gian nuôi bên nhánh B xuất hiện vi khuẩn có khả năng tổng hợp tryptophan (trp+
438
DẠYKÈMQUYNHƠN
). Qua nhiều lần thí nghiệm việc phage tải gen trp+ từ nhánh A sang nhánh B được chứng minh.
Hình 16: Thí nghiệm chứng minh có tải nạp do virut
OFFICIAL
Tải nạp chung (Genral transduction) Nhánh B Vi khuẩn Nhánh A Vi khuẩn trp+ Màng lọc vi khuẩn Tải nạp chung xảy ra khi phage mang bất kì gen nào của vi khuẩn A chuyển sang vi khuẩn B. Tải nạp chung (genralized transduction) có các đặc điểm:
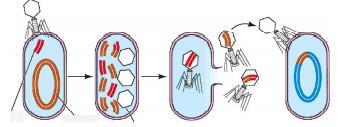
+ Thường do phage kiểu P1 thực hiện.
+ Bất kì gen nào của vi khuẩn cũng đều được tải nạp.
+ Tải nạp có được do gói nhầm ADN của tế bào chủ khi phage trưởng thành.
+ Các thể tái tổ hợp đơn bội được tạo ra.
Do không có sự tương đồng giữa trình tự ADN trên các phage này với trình tự ở tế bào chủ, nên không có điểm gắn vào đặc hiệu cho prophage. Bất kì gen nào cũng được tải nạp vì đầu của phage có thể gói nhầm vào một đoạn ADN của tế bào chủ. Sự đồng tải nạp (cotransduction) là quá trình tải nạp đồng thời 2 gen. Quá trình phage xâm nhập vào vi khuẩn và sinh sản được mô tả trên hình 4. Đầu tiên phage bám trên bề mặt của vi khuẩn, sau 4 phút bơm ADN của nó vào tế bào, sau đó chúng sinh sản và độ nửa giờ sau thì làm tan vi khuẩn và giải phóng các phage con mới.
Hình 17. Quá trình xâm nhập của phage và làm tan vi khuẩn ADN phage Nhiễm sắc thể vi khuẩn A Capsid Phage mới phá vỡ tế bào, nhiễm VK B
DẠYKÈMQUYNHƠN
439
Khi ADN của phage xâm nhập tế bào vi khuẩn A, chúng cắt ADN của vi khuẩn A thành nhiều đoạn, đồng thời ADN của phage được sao chép thành nhiều phân tử con và các vỏ capsid của phage cũng được tạo thành. Sau đó các vỏ capsid được lắp ruột ADN vào, phá vỡ tế bào vi khuẩn ra ngoài và tiếp tục xâm nhập các vi khuẩn mới. Trong quá trình lắp ráp khoảng 1-2% phage vô tình mang đoạn ADN của vi khuẩn có chứa gen. Phage mang gen của vi khuẩn A xâm nhập vi khuẩn B, quá trình tái tổ hợp xảy ra làm gen A gắn vào bộ gen B.
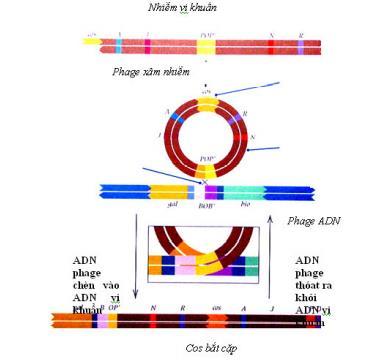
Tải nạp chuyên biệt (Special transduction) Tải nạp chuyên biệt hay hạn chế (restricted transduction) là trường hợp chỉ mang một vài gen nhất định, nó có 4 đặc điểm:
+ Những gen được chuyển nằm sát chỗ prophage gắn vào.
+ Chỉ prophage kiểu λ thực hiện.
+ Do kết quả sự cắt sai của prophage khi tách khỏi nhiễm sắc thể của tế bào chủ
OFFICIAL
+ Các vi khuẩn tái tổ hợp có thể lưỡng bội một phần.
Ví dụ: Phage λ chỉ mang gen gal (đồng hóa đường galactoz) từ vi khuẩn này chuyển sang vi khuẩn khác.
Hình 18: Điểm gắn của phage λ vào bộ gen vi khuẩn Điểm gắn của phage λ vào bộ gen của vi khuẩn nằm giữa 2 gen gal (galactoz) và bio (tổng hợp biotin), tiếp theo xảy ra quá trình tái tổ hợp ở điểm chuyên biệt (sitespecific recombination) chèn ADN của phage vào Nhiễm sắc thể vi khuẩn (hình 9).
DẠYKÈMQUYNHƠN
440
Đầ
u của phage chỉ có thể chứa một lượng ADN giới hạn, nó chỉ tải nạp được gen gal hoặc bio. Phage λ tải nạp các gen galactoz được gọi là λgal hay λdg (d = defective: khuyết, g = galactoz). Nếu tế bào gal– được nhiễm bởi λdg (mang gen gal+ ), sự ráp phage biến dạng vào tế bào chủ sẽ tạo lưỡng bội một phần. Sự cắt sai của phage λ rất hiếm nên tải nạp hạn chế có tần số thấp.
Tuy nhiên, tải nạp tần số cao có thể nhận được trong điều kiện thí nghiệm. Nếu tế bào vi khuẩn được gây nhiễm kép với phage λ hoang dại và phage λdg, phage hoang dại có thể hỗ trợ chức năng sai sót ở phage biến dạng, và thế hệ con sẽ có cả 2 kiểu với số lượng bằng nhau. Khi dịch tan được dùng tải nạp, quá trình được gọi là tải nạp tần số cao.
Trong nhiều trường hợp, do bộ gen biến dạng λdg không gắn được vào bộ gen của tế bào chủ (nên không sao chép được). Sau mỗi lần phân bào, chỉ một trong 2 tế bào có bộ gen của phage biến dạng; quá trình này được gọi là tải nạp sẩy (abortive transduction).
4.3. Giao nạp (Conjugation)
Vào năm 1946, J. Lederberg và E. Tatum chứng minh có trao đổi vật chất di truyền giữa các vi khuẩn sống. Sự trao đổi này gọi là giao nạp hay tiếp hợp (conjugation). Giao nạp ở vi khuẩn là sự kết hợp nhất thời của hai tế bào có kiểu bắt cặp đối nhau, được tiếp nối bằng sự chuyển một phần vật chất di truyền từ tế bào cho sang tế bào nhận qua cầu tế bào chất, và sau đó các tế bào tách nhau ra (exconjugants). Giao nạp đòi hỏi sự tiếp xúc trực tiếp giữa 2 loại tế bào được khởi sự bằng ống giao nạp hay tính mao (pilus), một sợi ống nhỏ rất dài do tế bào cho (donor cell) tạo ra (hình 19).
Hình 19. Hai tế bào vi khuẩn giao nạp qua cầu tế bào chất pilus ADN được truyền qua một cầu tế bào chất mỏng (a thin cytoplasmic bridge) gọi là ống giao nạp (conjugation tube). Kiểu sao chép sigma (σ) được tế bào vi khuẩn sử dụng trong giao nạp để truyền phân tử ADN dạng thẳng sang tế bào khác. Khi đọan ADN vào bên trong tế bào nhận (recipient cell), nó thực hiện tái tổ hợp với các gen tương đồng (homologous gens). Về thuật ngữ, nhiều tác giả dùng “tiếp hợp” để chỉ quá trình này, theo chúng tôi, dùng “giao nạp” tốt hơn, vì phản ánh được bản chất của quá
DẠYKÈMQUYNHƠN
OFFICIAL 441
trình, nhất quán với biến nạp, tải nạp ở vi khuẩn và khỏi nhầm lẫn với tiếp hợp của nhiễm sắc thể. Muốn xảy ra tái tổ hợp thì 2 dòng vi khuẩn phải tiếp xúc với nhau (F+ x F -) hoặc (Hfr x F-). Dòng tế bào mang nhân tố F+ được coi là tế bào “đực” và Plasmid có khả năng tạo protein pilin, từ protein này tạo ống giao nạp là pilus. Sự co lại của pilus đang nối hai tế bào làm chúng tiếp xúc kề nhau. Tế bào F– được coi là cái (female). Sau khi giao nạp tế bào F– trở thành F+ . Việc chuyển gen chỉ thực hiện khi plasmid gắn vào bộ gen của vi khuẩn. Trong quá trình chuyển vật chất di truyền sang F– thì ADN của tế bào chủ sao chép và mạch mới có ori đi đầu và F ở cuối (hình 20). Quá trình chuyển ADN từ F+ sang F– có thể bị ngắt quãng. Hfr X F– Sao chép ADN truyền cho F– Hfr F– tái tổ hợp Hình 20. Sự truyền ADN từ thể cho sang thể nhận Các gen A, B, C được chuyển một chiều từ Hfr sang F–.
Đoạn gen thể cho bắt cặp với các gen tương đồng của ADN tế bào F– và diễn ra trao đổi chéo tạo tế bào F– tái tổ hợp. Dòng Hfr có tần số lai cao hơn nhiều vì plasmid đã nằm sẵn trong bộ gen. Còn F+ phải qua giai đoạn plasmid gắn vào bộ gen rồi mới chuyển gen.
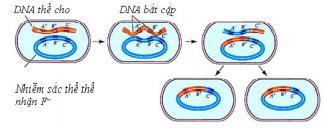
Trong điều kiện thí nghiệm ở 37oC, nguyên bộ gen của tế bào E. coli được chuyển sang tế bào nhận trong vòng 90 phút. Thường sự giao nạp bị ngắt giữa chừng do các pilus bị gãy, nên ít khi bộ gen được chuyển nguyên vẹn vào tế bào nhận. Lúc đó tế bào F–vẫn là F– . Bằng cách ngắt quãng giao nạp bản đồ di truyền của E.coli được xây dựng và có dạng 1 vòng tròn.
Hình 21: Đoạn ADN từ thể cho trao đổi chéo với ADN thể nhận tạo F– tái tổ hợp.
Sao chép Plasmid
DẠYKÈMQUYNHƠN
OFFICIAL 442
Plasmid là nhân tố di truyền sao chép độc lập với nhiễm sắc thể (Nhiễm sắc thể) của tế bào chủ. Không giống virut, plasmid không có dạng ngoại bào (extracellular form) và tồn tại trong tế bào đơn giản là ADN tự do và điển hình là dạng vòng tròn. Plasmid và ADN Nhiễm sắc thể của tế bào có vài điểm khác nhau cơ bản: plasmid chỉ mang những gen không thiết yếu (nhưng thường rất có ích), còn những gen thiết yếu nằm trên Nhiễm sắc thể. Plasmid di chuyển giữa các tế bào trong quá trình giao nạp (conjugation), tức sự tiếp xúc giữa hai loại tế bào.
OFFICIAL
Hình 22. Hình hiển vi điện tử các plasmid vòng tròn Enzym liên quan đến việc sao chép plasmid trên thực tế là những enzym bình thường trong tế bào. Vì thế những gen của plasmid chủ yếu liên quan đến sự kiểm soát quá trình khởi sự sao chép và với sự phân bổ những plasmid được sao chép vào các tế bào con. Tương tự, những plasmid khác nhau hiện diện trong tế bào với số lượng khác nhau, gọi là số bản sao (copy number). Một số plasmid chỉ có 1-3 bản sao trong tế bào, trong khi những plasmid khác lại có đến 100 bản sao. Số lượng bản sao được kiểm soát bởi những gen trên plasmid và bởi sự tương tác gen giữa tế bào chủ và plasmid. Phần lớn các plasmid ở vi khuẩn Gram âm sao chép tương tự như Nhiễm sắc thể vi khuẩn. Việc này thực hiện từ điểm khởi sự sao chép (Ori) và chép theo hai hướng quanh vòng plasmid, tạo ra dạng trung gian theta (θ). Tuy nhiên một số plasmid không sao chép theo hai hướng. Do kích thích nhỏ của ADN plasmid, toàn bộ quá trình sao chép xảy ra rất nhanh, có lẽ bằng 1/10 tổng thời gian diễn ra chu kì phân bào. Hầu hết plasmid của vi khuẩn Gr+ sao chép theo cơ chế vòng xoay tương tự như phage φx. Cơ chế này sẽ tạo ra mạch đơn trung gian, và do đó những plasmid này thỉnh thoảng được xem như plasmid ADN mạch đơn. Phần lớn ADN mạch thẳng sao chép dùng cơ chế liên quan đến một protein gắn với đầu 5’ của mỗi mạch mà được dùng trong tổng hợp mồi ADN. Cơ chế chính của việc chuyển plasmid từ tế bào này sang tế bào khác là giao nạp, mà chức năng thực hiện được mã hóa bởi chính một số plasmid. Giao nạp là một quá trình sao chép ADN và plasmid của tế bào cho và truyền bản sao của Nhiễm sắc thể cùng bản sao plasmid sang tế bào nhận.
5. Cơ chế di truyền của virut Các hạt virut (virut particle) hay virion là những vật kí sinh nội bào bắt buộc (obligate intracellular parasite). Chúng chỉ biểu hiện các gen của chúng và sinh sản bên trong 1 tế bào sống khác. Phụ thuộc vào loại tế bào chủ mà virut kí sinh, người ta
DẠYKÈMQUYNHƠN
443
gọi tên loại virut, ví dụ: virut thực vật kí sinh tế bào thực vật, virut động vật kí sinh tế bào động vật. Do đặc điểm này, sự sinh sản của virut khác hẳn với sự sinh sản của tế bào. Điểm nổi bậc là virut tạo ra hàng trăm hay hàng ngàn virion trong mỗi thế hệ
Do có sự đa dạng như nêu trên, các bộ gen của virut được sao chép theo nhiều con đường khác nhau:
+ Các ADN mạch kép –> ADN mạch kép: kiểu bán bảo tồn. + Các ARN mạch kép –> ARN mạch kép: kiểu bán bảo tồn nhờ ARN replicaz do gen tạo enzym sao chép này được mã hóa trong bộ gen của virut ARN.
+ Các ADN hoặc ARN mạch đơn –> chúng làm khuôn (mạch +) để tổng hợp mạch bổ sung (mạch –) thành dạng mạch kép trung gian (+/–), gọi là dạng sao chép (replicative form), mà từ nó lấy mạch – làm khuôn tạo ra mạch đơn + giống bộ gen ban đầu của virut (hình 23).
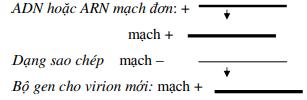
OFFICIAL
Hình 23. Sao chép của ADN hoặc ARN mạch đơn qua trung gian mạch kép Ở Retrovirut ARN –> nhờ enzym reverse transcriptaz tổng hợp c-ADN mạch đơn từ khuôn ARN virut –> c-ADN kép –> Chèn vào bộ gen tế bào chủ –> tạo ra bộ gen ARN virut mới và mARN. Phụ thuộc vào bộ gen ADN hay ARN mạch kép hay đơn, mạch dương (+) hay âm (–), kiểu sao chép bộ gen và tổng hợp mARN, các virut được chia thành 7 lớp (classes). Một cách sao chép đặc biệt được tế bào vi khuẩn sử dụng trong tiếp hợp để truyền phân tử ADN dạng thẳng sang tế bào khác hoặc được các virut sử dụng để tạo các bộ gen của chúng, mặt khác làm đứt mạch của vòng tròn xoắn kép, tạo đầu hở 3’–OH và 5’–P kết thúc. Helicaz và SSB protein chen vào tạo chẽ 3 sao chép. Sự sao chép được thực hiện không cần mồi (primer) vì mạch 3’–OH sẵn sàng cho việc nối dài như mạch trước (leading strand) nhờ ADN polymeaz I. Đồng thời với sao chép mạch trước, mạch khuôn sau dịch chuyển. Sự dịch chuyển của mạch sau gián đoạn để tổng hợp các đoạn ngắn Okazki như bình thường và đầu 5’ mạch khuôn duỗi thẳng ra (hình 19). Sự sao chép có hình giống chữ sigma (σ) của Hi Lạp, nên được gọi là sao chép sigma (σ replication). Kiểu sao chép σ này còn được gọi là vòng tròn quay (rolling-circle replication), vì mạch khuôn tròn ở giữa không bị đứt và quay tròn làm khuôn cho mạch trước. Sự sao chép kiểu này có thể lặp lại vài lần tạo ra sợi ADN dài, lặp lại nhiều lần bộ gen thẳng của virut, được gọi là concatemer. Enzym endonucleaz cắt ở những điểm khác nhau trên mỗi mạch của ADN tạo ra các đoạn cỡ bộ gen với hai đầu “dính”. Bộ gen thẳng này sẽ tạo thành vòng nhờ bắt cặp bổ sung các đầu “dính”.
DẠYKÈMQUYNHƠN
444
5.1. Vi rút động vật
Các chu trình sao chép của virut động vật có nhiều điểm tương tự với các virut khác với nhiều biến dạng đáng kể. Ví dụ trường hợp của cúm (influenza virut) hay của Paramyxovirut gồm các virut gây bệnh sởi và quai bị. Các virut cúm và Paramyxovirut có bộ gen là ARN một mạch được gói trong capsid xoắn dẻo. Phía bên ngoài capsid có màng bao là tính chất chung của nhiều nhóm virut động vật và phage (hình 20).
OFFICIAL
Hình
24. Virut cúm
Bao màng giúp virut xâm nhập tế bào chủ. Khi virut tiếp xúc với tế bào, các glycoprotein thòi ra ở màng bao gắn vào các thụ thể (receptor) trên màng sinh chất (hình 20).
Quá trình này chuyển capsid có chứa ARN vào tế bào chất và ở đó capsid bị mất (bóc vỏ). Các enzym của virut tham gia sao chép ARN của bộ gen và tạo mARN, nhưng bộ máy của tế bào được sử dụng để tổng hợp protein của virut. Các capsid mới được lắp ráp bao các bộ gen của virut và chúng đội màng sinh chất của tế bào mọc chồi rồi thoát ra khỏi tế bào. Bằng cách này các capsid có màng bao chính là màng sinh chất của tế bào chủ cũ và các virion mới có thể sử dụng các bao màng này để hòa nhập với màng của tế bào chủ mới. Toàn bộ chu trình này được gọi là chu trình sinh sản (reproductive cycle), chứ không phải chu trình tan vì các virut có thể thoát ra bằng mọc chồi mà không phải phá vỡ tế bào chủ. Đáng chú ý là chu trình sinh sản của các virut loại này diễn ra ở tế bào chất của tế bào và không tác động đến nhân tế bào.
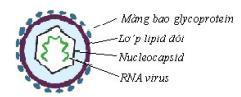
DẠYKÈMQUYNHƠN
445
Hình 25. Chu trình sinh sản của virut cúm Không phải tất cả các màng của virut đều bắt nguồn từ màng sinh chất. Ví dụ, bao màng của herpesvirut từ màng nhân của tế bào chủ. Các bộ gen của herpesvirut là ADN mạch kép , chúng sinh sản bên trong nhân tế bào, sử dụng phối hợp cả enzym của virut và tế bào để sao chép ADN và phiên mã.
Khi ở trong nhân của tế bào, ADN của herpesvirut (tiền virut) tương tự prophage ở vi khuẩn. Provirut ở dạng tiềm ẩn có thể gây một số triệu chứng bệnh ở người. Đôi khi các stress vật lí (như tác động ánh sáng mặt trời mạnh) hay tình cảm có thể làm cho provirut herpes bắt đầu chu trình sinh sản tạo các triệu chứng khó chịu cho con người.
5.2. Các retrovirut
Các virut có bộ gen ARN sinh sản qua trung gian ADN. Retrovirut HTLV-I gây bệnh bạch cầu tế bào T (T-cell leucemia) và retrovirut HIV gây bệnh AIDS làm suy yếu hệ thống miễn dịch (hình 26).Nói chung, retrovirut liên quan đến những bệnh nguy hiểm, khó chữa trị nhất hiện nay của nhân loại, kể cả ung thư. Tất cả các virut biến đổi tế bào qua việc gắn nucleic axit của virut vào ADN của tế bào chủ. Sự gắn này được ổn định (các provirut không bao giờ bị cắt rời ra như prophage). Đối với các virut gây bệnh ung thư là ADN, sự gắn vào là quá trình trực tiếp. Các retrovirut thì khác, trước hết chúng sử dụng reverse transcriptaz để từ khuôn ARN tạo ra ADN. Sau đó ADN của virut có thể được gắn vào ADN bộ gen của tế bào chủ và chúng cũng sao chép với bộ gen này trong mỗi thế hệ.
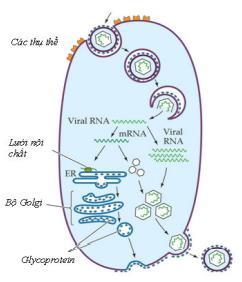
DẠYKÈMQUYNHƠN
446
OFFICIAL
Hình 26. Chu trình sinh sản của retrovirut HIV gây bệnh AIDS 5.3. Vi nấm
Hình 27: Nấm men S.cerevisiae dưới kính hiển vi điện tử và kính hiển vi quang học Nấm men có thể hoàn thành một chu trình sao chép và phân chia khoảng 1,4 giờ. Xuất phát điểm cho sao chép DNA của nấm men gồm nhiều điểm giống với oriC của E. coli. Đó là vùng giàu AT được tách ra khi có một protein khởi động gắn vào điểm kề bên. Các điểm xuất phát sao chép dài hàng ngàn đến hàng chục ngàn nucleotide. Khác với prokaryote, mỗi nhiễm sắc thể eukaryote có nhiều điểm xuất phát sao chép để quá trình sao chép genome của eukaryote có kích thước lớn hơn nhiều xảy ra một cách nhanh chóng. Có khoảng 400 điểm xuất phát sao chép phân bố trên toàn bộ 16 nhiễm sắc thể của nấm men. Sự sao chép xảy ra trực tiếp trên nhiều điểm xuất phát sao chép. Sợi mạch kép được tạo thành ở mỗi điểm xuất phát sao chép kéo dài và nối với sợi kép được tạo thành ở một điểm khác. Khi sao chép trên 2 mạch đơn hoàn thành sẽ tạo ra 2 phân tử DNA con giống hệt nhau.
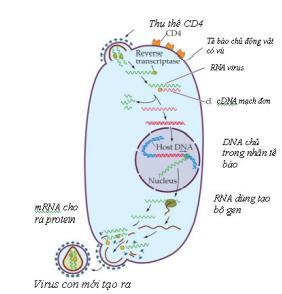
DẠYKÈMQUYNHƠN
447
OFFICIAL
Sự tổng hợp DNA chỉ xảy ra ở một giai đoạn trong chu trình tế bào, đó là pha S. Ở nấm nấm men có 3 protein cần cho lắp ráp của replisome. Đó là phức hợp DNA polymerase phối hợp hoạt động ở chẽ ba sao chép. Phức hợp nhận biết điểm xuất phát (Origin recognition complex – ORC) sẽ gắn vào trình tự xuất phát của nấm men như DnaA protein của E. coli. Những phức tương tự được tìm thấy ở tất cả các eukaryote. Sự có mặt của ORC làm bổ sung thêm 2 protein khác, Cdc6 và Cdt. Cả 2 protein này và ORC làm kích hoạt helicase và những thành phần khác của replisome. Sự gắn vào của helicase được xem là sự “xác nhận” (license) cho điểm xuất phát, giúp lắp ráp replisome và bắt đầu tổng hợp DNA. Trong số các gene mã hoá protein, intron chỉ có khoảng 4-5% tất cả các gene của nấm men (276 gene chứa một intron đơn và 7 gene chứa 2 intron riêng rẽ), kích thước trung bình của intron khoảng 2000 nucleotide. Tần số thấp của intron ở nấm men có lẽ liên quan với tần số thấp của các gene giả (pseudogene) giống như intron, phổ biến ở eukaryote bậc cao. Gene giả chứa trình tự mã hoá, nhưng vì thiếu intron và promoter phiên mã nên trình tự mã hoá không được biểu hiện.
OFFICIAL
Gene mã hoá protein không chỉ là những dạng gene chức năng. Genome của nấm men chứa số lớn gen tạo ra RNA không mã hoá, bao gồm các gene lặp lại mã hoá cho rRNA, 274 gene của tRNA, trong đó có 80 gene chứa nhóm intron đặc biệt, RNA nhân nhỏ (snRNA) tham gia chức năng chế biến rRNA, 5 snRNA tham gia chế biến intron, một vài RNA chưa biết chức năng và 3 RNA như là các tiểu đơn vị chức năng của enzyme Rnase, endoribonuclease và telomere.
Chu trình sống của các loài nấm khác nhau rất đa dạng, nhưng có thể xếp vào 5 kiểu căn bản khác nhau như: 1. Sinh sản vô tính; 2. Thể đơn bội; 3. Thể đơn bội với giai đoạn dị nhân; 4. Đơn bội - Lưỡng bội ; 5. Lưỡng bội.
Ngay trong các nấm sinh sản hữu tính cũng có sự dao động từ hoàn toàn đơn bội như Neurospora đến hoàn toàn lưỡng bội như ở Saccharomyces cerevisiae Đối với phần lớn nấm sợi, điểm đặc trưng là nhiều nhân cùng hiện diện trong một tế bào. Các nhân này có thể có kiểu gen khác nhau. Trong trường hợp này được gọi là thể dị nhân (heterokaryon) tương tự dị hợp tử.
Ở các vi sinh vật nói chung, các tính trạng hình thái thường không nhiều, nhưng có sự đa dạng lớn về nhiều loại phản ứng hóa học khác nhau. Sự ra đời của giả thuyết nổi tiếng 1 gen - 1 enzym đã dựa trên các đột biến sinh hóa ở N. crassa. Các môi trường chọn lọc được sử dụng để thu thập các dạng lai hiếm hoi giữa các dòng cha mẹ đơn bội mang các đột biến khuyết dưỡng lặn, khi sai hỏng trong chuỗi phản ứng sinh hóa của chúng bổ sungcho nhau. Ngoài ra tính đề kháng với các tác nhân bất lợi được sử d
ng rộng rãi làm tính trạng đánh dấu trong các nghiên cứu di truyền ở nấm.
DẠYKÈMQUYNHƠN
448
6.
u
ể
6.1.
ụ
Điề
hoà bi
u hiện gene ở vi khuẩn
Mô hình Operon
Phần lớn các gene trong bộ gen vi khuẩn được tổ chức thành các đơn vị hoạt động chức năng đặc trưng, gọi là các operon. Các gene cấu trúc trong một operon được điều hoà chung trong quá trình chuyển hoá một hợp chất nhất định của tế bào. Lần đầu tiên vào năm 1961, Francois Jacob và Jacques Monod (France) đề xuất giả thuyết operon để giải thích sự điều hoà quá trình sinh tổng hợp protein ở vi khuẩncác enzyme tham gia vào con đường hấp thụ và phân giải đường lactose - operon lactose (lac operon). Đây là operon được nghiên cứu kỹ nhất cho đến nay (Hình 24).
Operon là đơn vị tổ chức và hoạt động gene đặc trưng của các bộ gene prokaryote; nó là một phức hợp liên kết chặt chẽ giữa vùng khởi động (promoter) cùng với yếu tố chỉ huy (operator) và nhóm gene cấu trúc do yếu tố này trực tiếp kiểm soát. (Vì vậy nó được gọi là operon).
Tham gia vào điều hòa hoạt động của một operon gồm có bốn yếu tố thuộc hai thành phần chính: (i) các locus cấu trúc (structural loci) và (ii) các locus điều hòa (regulatory loci); trong đó nhóm sau bao gồm yếu tố chỉ huy (operator), vùng khởi động (promoter) và gene điều hòa (regulator gene).
OFFICIAL
Hình 28. (a) Nhiễm sắc thể E. coli với vị trí tương đối của các operon khác nhau. (b) Cấu trúc phân tử đường lactose; và (c) mô hình operon lactose và chức năng của nó - sản sinh các enzyme hấp thụ và phân giải đường lactose. - Một nhóm các gene cấu trúc (structural genes) liên quan với nhau về mặt chức năng xếp cạnh nhau, khi phiên mã sẽ tạo ra một phân tử mRNA chung gọi là mRNA đa cistron (polycistronic mRNA). Các enzyme được dịch mã từ một mRNA này sẽ tham gia vào quá trình chuyển hoá (đồng hoá hoặc dị hoá) cụ thể, một chuỗi các phản ứng sinh hoá gồm nhiều khâu nối tiếp hoặc có quan hệ dạng lưới phức tạp.
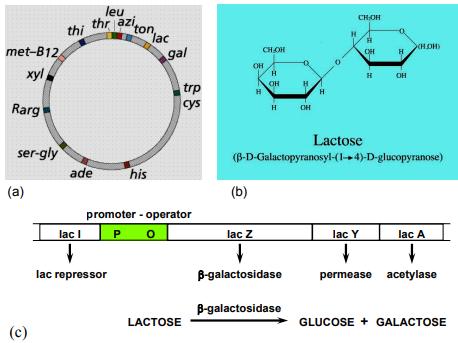
DẠYKÈMQUYNHƠN
449
- Một yếu tố chỉ huy (operator = O): trình tự DNA nằm kế trước nhóm gene cấu trúc, là vị trí tương tác với chất ức chế.
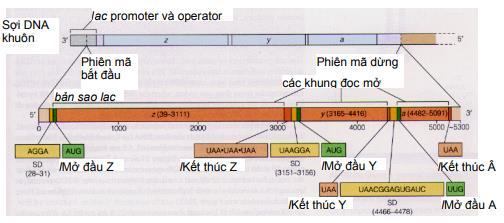
- Một vùng khởi động (promotor region = P): trình tự DNA nằm trước yếu tố chỉ huy và có thể trùm lên một phần hoặc toàn bộ vùng này, là vị trí bám vào của RNA polymerase để có thể khởi đầu phiên mã tại vị trí chính xác ở sợi khuôn.
- Một gene điều hoà hay còn gọi là gene ức chế (regulatory/ inhibitory gene = R/I): Gene này sinh ra loại protein điều hoà gọi là chất ức chế (repressor) điều hòa hoạt động của nhóm gene cấu trúc thông qua sự tương tác với yếu tố chỉ huy. Mặc dù mỗi gene điều hòa có một vùng khởi động riêng và không có yếu tố chỉ huy, đôi khi người ta vẫn coi chúng là một operon điều hòa; gene này sinh ra chất ức chế một cách ổn định.
OFFICIAL
6.2. Điều hoà âm tính của các operon cảm ứng: lac operon Đại diện cho tất cả các operon của các loại đường di- và polysaccharide (mà vi khuẩn sử dụng như một nguồn cung cấp các hợp chất carbon và năng lượng) là operon lactose ở E. coli.
Operon lactose có chức năng sản sinh các enzyme tham gia vào quá trình hấp thụ và phân giải đường lactose (một disacharide) thành galactose và glucose. Nó chỉ hoạt động khi có mặt đường lactose, vì vậy lactose được gọi là chất cảm ứng và lac operon được gọi là operon cảm ứng (inducible) hay operon dị hoá (catabolite). Nói đúng ra, chất cảm ứng là allolactose; lactose (liên kết galactosid dạng β-1,4) bị biến đổi thành chất trung gian trong quá trình thuỷ phân lactose dưới tác dụng của βgalactosidase, gọi là allolactose (liên kết β-1,6). Cấu trúc của lac operon Các thành phần của lac operon ở E. coli như :
Hình 29: Cấu trúc chi tiết các vùng khác nhau của operon lactose - Nhóm các gene cấu trúc bao gồm ba gene: lacZ, lacY và lacA (nói gọn là Z, Y và A); trong đó lacZ mã hoá cho β galactosidase (thuỷ phân lactose), lacY mã hoá cho permease (vận chuyển lactose qua màng) và lacA mã hoá transacetylase (chức năng không rõ ràng; theo ý nghĩa nó không phải là enzyme liên quan trực tiếp đến sự chuyển hoá lactose).
DẠYKÈMQUYNHƠN
450
- Yếu tố chỉ huy (lac operator) là trình tự DNA dài ~34 cặp base cách 80 gene Z chừng 10 cặp base về phía trước, là vị trí tương tác với chất ức chế. Nó chứa trình tự 24 cặp base đối xứng xuôi ngược, giúp chất ức chế (lac repressor) có thể nhận biết và bám vào bằng cách khuếch tán dọc theo DNA từ cả hai phía. - Vùng khởi động (lac promotor) là đoạn DNA dài chừng 90 cặp base nằm trước và trùm lên lac operator 7 cặp base. Nó chứa hai vị trí tương tác với RNA polymerase và với protein hoạt hoá dị hoá (catabolite activator protein = CAP, hoặc CRP - xem mục V). Điểm khởi đầu phiên mã là vị trí gần cuối của lac promoter.Gene điều hoà (regulatory gene) nằm trước vùng khởi động, mã hoá một protein ức chế gồm bốn polypeptide giống nhau, gọi là tứ phân (tetramer), đều chứa 360 amino acid.
Cơ chế điều hoà âm tính của lac operon

Khi trong môi trường nuôi cấy E. coli vắng mặt lactose (allolactose, chất cảm ứng) thì lac operon không hoạt động, nghĩa là các enzyme tham gia hấp thụ và phân giải lactose không được sinh ra. Nguyên nhân là do chất ức chế của operon (lac repressor) vốn tự thân có hoạt tính, bám chặt vào yếu tố chỉ huy (lac operator) và gây kìm hãm sự phiên mã của các gene cấu trúc Z, Y và A (Hình 26). Do đó các sản phẩm enzyme của lac operon không được tạo ra; tức biểu hiện âm tính.
OFFICIAL
Hình 30: (a) Chất ức chế bám chặt lac operator gây ức chế phiên mã; (b) Mô hình lac operator (rìa trái) bị bám chặt bởi protein ức chế (rìa phải). Ngược lại, nếu bổ sung lactose vào môi trường thì một thời gian sau vi khuẩn sẽ bắt đầu hấp thụ và phân giải nó, nghĩa là các enzyme liên quan đã được sinh ra. Sự kiện này được lý giải như sau: Chất cảm ứng (inducer), ở đây là allolactose - dạng biến đổi của lactose - tương tác với chất ức chế (repressor) làm biến đổi cấu hình của chất này. Một phân tử allolactose bám vào một tiểu đơn vị của chất ức chế. Vì vậy chất ức chế mất ái lực và không thể bám vào lac operator; nó tách ra khỏi DNA. Lúc này các gene cấu trúc được phiên mã và các enzyme tương ứng được tổng hợp, nhờ vậy vi khuẩn có thể hấp thụ và phân giải đường lactose (Hình 27). Lactose vì vậy là tác nhân gây cảm ứng (hoạt hoá) lac operon. Ngoài ra, ITPG (isopropyl thiogalactoside) cũng được dùng như một chất cảm ứng nhưng không phải là tác nhân sinh lý.
DẠYKÈMQUYNHƠN
451
Hình 31: Chất cảm ứng kết hợp với chất ức chế và làm biến đổi hình dáng của nó; chất ức chế vì vậy không bám được vào lac operator. Kết quả là các gene cấu trúc của lac operon được phiên mã tạo ra phân tử mRNA polycistron và các enzyme tương ứng được tổng hợp.
Phương thức điều hoà như thế được gọi là điều hoà cảm ứng - âm tính, bởi vì chất ức chế lac operon một khi bám vào lac operator sẽ kìm hãm phiên mã, nghĩa là gây hiệu quả âm tính lên sự biểu hiện của các gene; và hoạt động chức năng của protein này lại phụ thuộc vào chất cảm ứng. Nhờ cơ chế điều hoà kiểu liên hệ ngược này mà vi khuẩn có thể thích ứng để tồn tại và phát triển một cách hợp lý. Hình
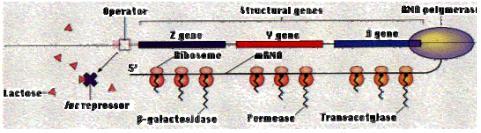
cơ bản, cơ chế "mở" của lac operon được trình bày như trên; nhưng thực ra sự hoạt động của chất cảm ứng mới chỉ làm dịu bớt (alleviation) sự điều hoà âm tính (sự ức chế) của lac operon. Hình 32 cho thấy ngay cả khi chất ức chế đã tách khỏi operator, RNA polymerase vẫn không thể bám ổn định vào promoter và khởi đầu phiên mã - nó không có ái lực đủ cao đối với promoter để bám vào đủ lâu để có thể khởi đầu tạo thành liên kết phosphodiester đầu tiên.
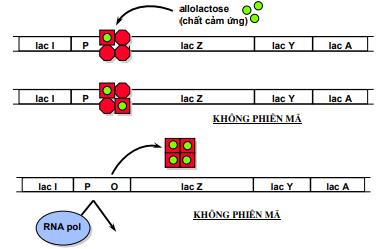
DẠYKÈMQUYNHƠN
452
OFFICIAL
được làm dịu bớt, tuy nhiên RNA polymerase vẫn chưa thể tạo thành một phức hợp bền vững với promoter để có thể khởi đầu phiên mã được. Về
32: Chất ức chế một khi được bám đầy đủ bởi allolactose thì tách khỏi operator khiến cho sự điều hoà âm tính (sự ức chế)
6.3. Cơ chế điều hoà âm tính của trp operon
Khi trong tế bào E. coli dư thừa amino acid tryptophan (sản phẩm cuối cùng của con đường chuyển hoá) thì trp operon ngừng hoạt động và do đó các enzyme tương ứng không được sinh ra. Sự kiện này được giải thích như sau: Chất ức chế bình thường của operon này (trp repressor) tồn tại ở dạng bất hoạt gọi là aporepressor, không có ái lực đối với trp operator, nhưng khi các amio acid dư thừa kết hợp vào sẽ tạo ra phức hợp có hoạt tính, nghĩa là có ái lực với trp operator. Vì thế tryptophan được gọi là chất đồng ức chế (corepressor). Phức hợp này bám vào yếu tố chỉ huy làm kìm hãm phiên mã của trp operon. Ngược lại, khi trong tế bào vắng mặt hay thiếu hụt các amino acid này, tự thân chất ức chế này ở trạng thái bất hoạt nên không bám được vào yếu tố chỉ huy (trp operator). Vì vậy các gene cấu trúc xảy ra sự phiên mã và kết quả là các enzyme tham gia tổng hợp tryptophan được sinh ra. Và một khi hàm lượng amio acid này được tổng hợp ở mức dư thừa sẽ tác động ngược trở lại, kìm hãm hoạt động của trp operon.
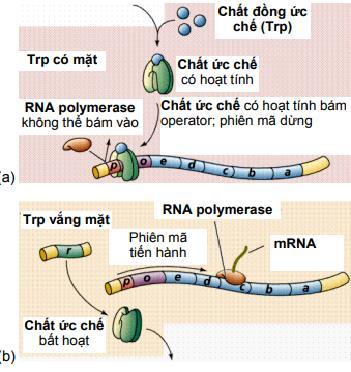
OFFICIAL
Hình 33: Operon tryptophan bị kìm hãm (a) và hoạt động (b). Tóm lại, phương thức điều hòa hoạt động gene theo các cơ chế liên hệ ngược hay phản hồi như thế (feed-back mechanisms) đảm bảo cho bộ gene các vi khuẩn hoạt động một cách hợp lý và nhờ đó các vi khuẩn thích ứng và phát triển trước các điều kiện môi trường luôn thay đổi. ➢ Cần lưu ý rằng, các protein ức chế và đồng ức chế không hẳn là cách duy nhất các vi khuẩn điều hòa phiên mã gene. Ở nhiều vi khuẩn (và một số eukaryote), sự điều hòa mức chuyển hóa nào đó cũng có thể được kiểm soát bởi các riboswitch (tạm dịch là: công tắc ribo). Một riboswitch là một phần của vùng 5'-không được dịch mã (5'-untranslated region = 5'- UTR) trong phân tử mRNA, tại đó có một vị trí bám đặc thù cho chất chuyển hóa (metabolite). Một số chất chuyển hóa bám vào các riboswitch, như: các purine adenine và guanine; các amino acid glycine và lysine; mononucleotide flavin (nhóm phụ của
DẠYKÈMQUYNHƠN
453
NADH dehydrogenase); S-adenosyl methionine (nhường nhóm methyl cho nhiều phân tử, kể cả DNA và "chóp" ở đầu 5' của mRNA eukaryote).
Trong mỗi trường hợp, riboswitch kiểm soát các gene liên quan đến sự chuyển hóa của phân tử đó. Chất chuyển hóa bám vào mRNA đang sinh trưởng và bao gồm cả sự thay đổi biến cấu (allosteric) mà: (i) đối với một số gene nó khiến cho sự tổng hợp mRNA hơn nữa để kết thúc trước khi hình thành một sản phẩm chức năng, và (ii) đối với một số gene khác, nó tăng cường hoàn chỉnh việc tổng hợp mRNA. Trong cả hai trường hợp, kết quả là do sự kiểm soát mức độ của chính chất chuyển hóa ấy. Một số riboswitch kiểm soát sự dịch mã mRNA hơn là phiên mã mRNA. Đã có gợi ý rằng các cơ chế điều hoà này, vốn không liên quan bất kỳ protein nào; nó là một cơ chế còn sót lại từ "giới RNA".
6.4. Sự ức chế dị hoá (Catabolite repression): Điều hoà dương tính của lac operon
Như chúng ta có thể dự đoán, điều hoà dương tính (positive control) là đối lập với điều hoà âm tính; đó là, operon bị đóng, ngưng hoạt động (thực ra là hoạt động của operon bị giảm xuống một mức cơ sở) trừ phi có yếu tố nào đó xen vào bật nó hoạt động trở lại. Trong trường hợp của lac operon, điều này có nghĩa là tách bỏ chất ức chế ra khỏi operator là chưa đủ để kích hoạt operon như đã đề cập trước đây. Nó cần đến một nhân tố dương tính bổ sung thêm. Thật vậy, hoạt động của lac operon còn chịu sự kiểm soát của một protein điều hoà dương tính liên quan với sự có mặt của glucose. Cụ thể, khi trong môi trường có mặt đồng thời cả lactose và glucose thì operon lac tạm thời ngưng hoạt động. Hiện tượng này gọi là ức chế dị hoá (catabolite repression). Người ta nhận thấy rằng, khi glucose có mặt ở nồng độ cao thì hàm lượng AMP vòng (3',5'-cyclic adenosine monophosphate = cAMP; Hình 34.a) trong tế bào rất thấp; và ngược lại, khi không có glucose hoặc có không đáng kể thì hàm lượng cAMP trong tế bào được tổng hợp tăng cao. cAMP vì vậy được xem là chất chỉ thị (indicator) của sự vắng mặt glucose và được coi là nhân tố điều hoà dương tính (positive regulator) của các operon dị hoá.
DẠYKÈMQUYNHƠN
OFFICIAL 454
Hình 34 (a) Cấu trúc và sự hình thành phân tử cAMP từ ATP. (b) CAP gồm hai monomer giống nhau, mỗi monomer nhận biết một trình tự DNA nhờ vùng xoắn alpha được đánh dấu F; và (c) Trình tự đối xứng của vị trí CAP là các đoạn lặp đảo ngược.
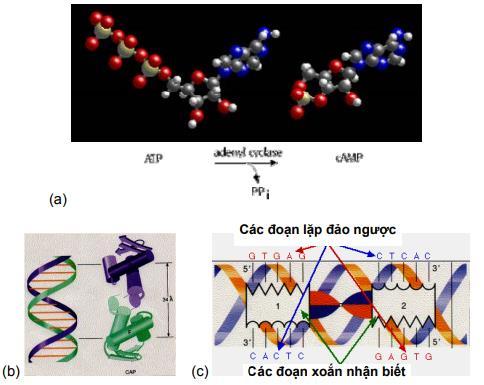
Ngoài ra, còn phát hiện một loại protein điều hoà dương tính có tên là protein hoạt hoá dị hoá (catabolite activator protein = CAP) cũng gọi là protein tiếp nhận / bám cAMP (cyclic AMP receptor / binding protein = CRP) hay protein bám cAMP. CAP gồm hai tiểu đơn vị giống nhau gọi là homodimer (Hình 34.b); nó chỉ hoạt động khi môi trường nội bào có hàm lượng cAMP cao. Trong trường hợp đó, cAMP kết hợp với CAP tạo thành phức hợp CAP-cAMP và làm tăng ái lực của CAP đối với promoter. Phức hợp này có khả năng nhận biết và bám vào một đoạn 16 bp về phía trước vùng khởi động, với các đoạn lặp đảo ngược (inverted repeats), gọi là vị trí CAP (Hình 34.c). Bằng cách đó RNA polymerase được kích thích bám chặt vào promoter và bắt đầu tổng hơp mRNA ở mức cao.
Như vậy, khác với kiểu điều hoà âm tính do sự tương tác giữa ''chất ức chế và operator'' (tương tác protein-DNA) ở đây sự tương tác xảy ra giữa protein điều hoà thuộc phức hợp CAP-cAMP mà yếu tố chính là CAP với RNA polymerase (tương tác protein-protein) giúp RNA polymersae bám ổn định vào promoter, tăng cường hoạt động phiên mã (điều hoà dương tính) mà chủ yếu là điều chỉnh tốc độ khởi đầu phiên mã. Mặt khác khi CAP/cAMP bám vào, nó làm cho DNA uốn gập đáng kể (khoảng 90 độ). Và như thế, RNA polymerase dễ dàng tách hai sợi của DNA, tạo thành một phức hợp mở.
DẠYKÈMQUYNHƠN
455
OFFICIAL
Hình 35: Điều hoà dương tính lac operon CAP/cAMP cũng có thể kích thích phiên mã các operon cảm ứng khác, bao gồm các ara và gal operon được nghiên cứu kỹ. Ngay khi lac operon tiến hành chuyển hoá lactose, các operon khác này mã hoá cho các enzyme phân cắt các đường biến đổi tương ứng, arabinose và galactose. Như thế, để cho hiệu suất lớn nhất, tất cả ba operon này sẽ vẫn đóng chừng nào tế bào vẫn còn có sẵn glucose. Vì cAMP đáp ứng với nồng độ glucose, nên ta chẳng ngạc nhiên gì khi cả ba operon này cùng chia xẻ một cơ chế điều hoà chung có liên quan cAMP. Phức hợp CAP-cAMP bám ở promoter hoặc gần promoter của mỗi operon và tạo điều kiện thuận lợi cho việc bám vào của RNA polymerase.
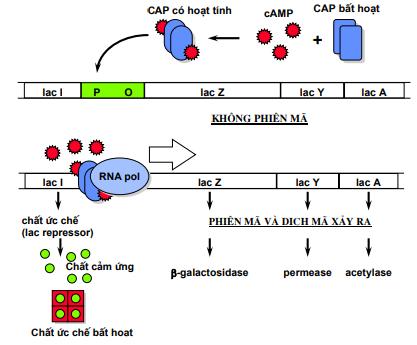
6.5. Sự kết thúc phiên mã sớm (Attenuation) ở trp operon Attenuation (phiên mã dở) là một cơ chế điều hoà gây ra sự kết thúc phiên mã sớm dưới những điều kiện nhất định, bằng cách đó ngăn cản sự biểu hiện của mRNA cần cho sự biểu hiện của các sản phẩm gene tương ứng. Phiên mã dở tạo thành mRNA uốn gập một cách điển hình thành các cấu trúc bậc hai xen kẻ (alternative secondary structures), mà một trong số đó là nhân tố kết thúc độc lập ρ (Rho-independent terminator).
Một cách tiếp cận tin sinh học đã được phát triển để xác định các gene được điều hoà theo kiểu phiên mã dở (Một số bài báo tổng quan hay về phiên mã dở như: Gollnick và Babitzke 2002; Henkin và Yanofsky 2002.)
Operon tryptophan chẳng hạn còn có một kiểu điều hòa phiên mã dở. Nó sử dụng dịch mã để điều khiển sự phiên mã. Khi có mặt tryptophan trong môi trường nội
DẠYKÈMQUYNHƠN
456
OFFICIAL
bào, thậm chí ở nồng độ thấp, sẽ xảy ra sự dịch mã một phần ở vùng leader của mRNA đang được tổng hợp. Kết quả là làm dừng sự phiên mã trước khi gene cấu trúc đầu tiên (trpE) của operon được phiên mã.
OFFICIAL
Hình 36: Cấu trúc của đoạn dẫn đầu - TrpL (a) và vùng kết thúc phiên mã sớm - trp attenuator với đuôi 3' gồm 8 uridine (b). Sự kết thúc phiên mã sớm ở operon tryptophan là kết quả của sự tương tác bổ sung nội phân tử giữa các trình tự DNA bên trong vùng leader của bản sao RNA. Kết quả của sự kết thúc phiên mã sớm này tạo ra một mRNA chứa 140 base (hình 36a) Tại vùng đầu mút 3' của nó xảy ra sự tự bổ sung ở đoạn giàu GC tạo thành một cấu trúc hình vòng trên thân RNA và gây ra sự kết thúc phiên mã sớm. Vùng này được gọi là đoạn phiên mã dở của operon tryptophan (trp attenuator) và ở phần đuôi của mRNA này cũng có 8 base uridine (hình 36.B). Kiểu cấu trúc "kẹp cài tóc" này là tín hiệu kiểm soát kết thúc phiên mã ở prokaryote nói chung. Với kiểu cấu trúc đặc thù ở đoạn dẫn đầu của trp operon như vậy làm cho nó có ý nghĩa quan trọng trong điều hoà phiên mã dở, ở chỗ:
(i) tổng hợp một peptide dẫn đầu chứa 14 amino acid; (ii) trên mRNA của đoạn peptide này chứa hai codon của Trp ở các vị trí 10 và 11; (iii) ở bốn vùng được đánh số 1-4 xảy ra sự tự bổ sung giữa các vùng 1 và 2 và giữa 3 và 4; và ở một số trường hợp có thể xảy ra sự kết cặp giữa các vùng 2 và 3. Do trong trình tự mã hóa của trình tự dẫn đầu trpL có hai codon Trp, nên sự dịch mã đoạn này tỏ ra nhạy cảm với số lượng tRNAtrp đưa vào. Nếu môi trường cung cấp đầy đủ Trp, ribosome trượt qua các codon Trp để đi vào vùng 2. Và sự có mặt của ribosome ở vùng 2 ngăn cản vùng này kết cặp với vùng 3. Khi đó vùng 3 sẽ cặp với
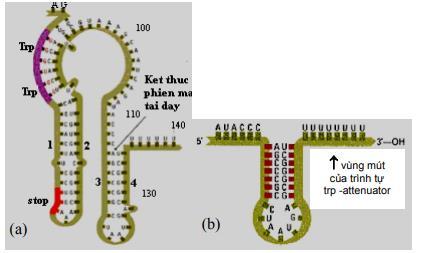
DẠYKÈMQUYNHƠN
457
vùng 4 và tạo ra điểm kết thúc phiên mã sớm (xảy ra sau khi tổng hợp xong 8 uridine ở ngay sau vùng 4). Khi số lượng tRNATrp đưa vào không đầy đủ, sự dịch mã đoạn dẫn đầu dừng lại đột ngột ở các codon Trp của nó. Điều này ngăn cản ribosome tiến vào vùng 2, do đó vùng này sẽ cặp với vùng 3 gây cản trở việc tạo thành cấu trúc phiên mã dở (trp attenuator). Kết quả là phân tử mRNA đa cistron của operon tryptophan được tạo thành một cách đầy đủ.
OFFICIAL
B. CÂU HỎI VÀ BÀI TẬP
Câu 1:
Hệ gene của virus HPV-16 (human papilloma virus), một trong những loại virus gây ung thư cổ tử cung phổ biến ở người, chứa gene mã hoá 2 protein ung thư (E6 và E7) làm biến đổi tế bào biểu mô bình thường thành tế bào khối u. Người ta tiến hành các thí nghiệm để phân tích tác động của protein E7 đến các tế bào ở cấp độ phân tử, cụ thể là mối quan hệ của nó với protein retinoblastoma (Rb), một loại protein áp chế khối u liên kết với yếu tố phiên mã E2F điều hoà sự phiên mã của các gene mã hóa protein tham gia pha S của chu kỳ tế bào. Ở thí nghiệm thứ nhất (Hình 2.1), dòng tế bào ung thư bạch cầu (HL-60) được nuôi cấy trong điều kiện không có (dấu ) hoặc có (dấu +) xử lý với phorbol ester và được đánh dấu bằng 35S-methionine hoặc 32P phosphate. Dịch chiết tế bào được dùng để điện di trên gel SDS-polyacrylamide và xử lý với kháng thể đặc hiệu của protein Rb. Kết quả được ghi lại bằng phóng xạ tự ghi.
DẠYKÈMQUYNHƠN
458
Ở thí nghiệm tiếp theo, các tế bào HL-60 được nuôi cấy trong điều kiện không có hoặc có phorbol ester. Dịch chiết tế bào được ủ với các hạt agarose có khả năng liên kết cộng hoá trị với các phân tử E7. Sau khi ủ, phần cặn lắng và dịch nổi được thu lại bằng ly tâm. Kết quả điện di trên gel SDS-polyacrylamide và lai Western sử dụng kháng thể đặc hiệu của Rb với các mẫu dịch chiết tế bào trước khi ủ, dịch nổi và cặn lắng sau khi ủ với hạt agarose được biểu thị trên Hình 2.2 (dấu : không có; dấu +: có các thành phần tương ứng).
OFFICIAL
Hình 2.1
Hình 2.2
Dựa trên các kết quả thí nghiệm, hãy trả lời các câu hỏi sau:
a) Protein Rb hoạt động trong tế bào bình thường ở dạng phosphoryl hoá hay khử phosphoryl hoá? Giải thích.
b) Phorbol ester có tác động gì đến protein Rb? Giải thích.
c) Cơ chế tác dụng của protein E7 đến sự tăng sinh của tế bào HL-60 như thế nào? Giải thích. Hướng dẫn
a. Protein Rb hoạt động ở dạng khử phosphoryl hoá. Vì ở thí nghiệm thứ nhất, trong tế bào HL-60 không xử lý phorbol ester, protein Rb ở dạng phosphoryl hoá thể hiện ở băng vạch có khối lượng phân tử cao hơn (Mr cao) khi đánh dấu bằng 35S methionine và 32P phosphate (mẫu 1 và 3). Đây là các tế bào ung thư phân chia nhiều lần có protein Rb ở dạng bất hoạt.
b. Phorbol ester kích thích sự khử phosphoryl hoá protein Rb. Vì khi xử lý với phosbol ester, protein Rb có khối lượng phân tử thấp hơn (Mr thấp) trong thí nghiệm đánh dấu bằng 35S methionine (mẫu 2) và không phát hiện thấy khi đánh dấu bằng 32P phosphate (mẫu 4).

DẠYKÈMQUYNHƠN
459
c. - Cơ chế tác dụng của protein E7 là liên kết với protein Rb ở dạng khử phosphoryl hoá, giải phóng yếu tố phiên mã E2F dẫn đến sự phiên mã các gen liên quan đến pha S và gây tăng sinh tế bào HL-60.
- Vì khi không xử lý với phorbol ester (mẫu 7), không phát hiện thấy RB ở cặn lắng chứng tỏ E7 liên kết với hạt agarose không tương tác với Rb ở dạng phosphoryl hoá. Khi xử lý với phorbol ester (mẫu 10), Rb xuất hiện ở cặn lắng chứng tỏ có sự liên kết với E7 khi ở dạng khử phosphoryl hoá. Do đó, yếu tố phiên mã E2F được giải phóng và hoạt hóa sự phiên mã các gene liên quan đến pha S.
Câu 2:
Thành tế bào của nhiều vi khuẩn được bao phủ bởi một lớp màng nhầy có thành phần polysaccharide hoặc protein. Người ta tiến hành nghiên cứu sự tổng hợp hyaluronan, một loại polysaccharide có thể tạo màng nhầy, gồm 2 loại đơn phân Nacetylglucosamine (GlcNAc) và acid glucuronic (GlcUA) xếp xen kẽ nhau, dưới tác dụng của enzyme hyaluronan synthase trên màng tế bào.
OFFICIAL
Hyaluronan synthase được trộn với cơ chất ban đầu - một tetrasaccharide có đánh dấu phóng xạ (Hình 3.1) trong các điều kiện: không bổ sung, có bổ sung riêng lẻ, bổ sung hỗn hợp các phân tử đường ở dạng hoạt hóa cần thiết cho sự tổng hợp (GlcNAc, GlcUA). Sau thời gian ủ, người ta tiến hành sắc ký bản mỏng sản phẩm thu được ở các điều kiện thí nghiệm (Hình 3.2, các chữ số bên trái thể hiện số gốc đường trong phân tử carbohydrate được xác định dựa trên chất chuẩn).
Hình 3.1 Hình 3.2
DẠYKÈMQUYNHƠN
460
a) Dựa vào dữ liệu đã cho, hãy cho biết hyaluronan synthase xúc tác việc gắn thêm các gốc đường ở dạng monosaccharide hay disaccharide? Giải thích.
b) Các gốc đường được gắn vào đầu khử hay đầu không khử của tetrasaccharide ban đầu? Giải thích. Hyaluronan synthase được trộn với cơ chất ban đầu - một tetrasaccharide có đánh dấu phóng xạ (Hình 3.1) trong các điều kiện: không bổ sung, có bổ sung riêng lẻ, bổ sung hỗn hợp các phân tử đường ở dạng hoạt hóa cần thiết cho sự tổng hợp (GlcNAc, GlcUA). Sau thời gian ủ, người ta tiến hành sắc ký bản mỏng sản phẩm thu được ở các điều kiện thí nghiệm (Hình 3.2, các chữ số bên trái thể hiện số gốc đường trong phân tử carbohydrate được xác định dựa trên chất chuẩn).
c) Dựa vào dữ liệu đã cho, hãy cho biết hyaluronan synthase xúc tác việc gắn thêm các gốc đường ở dạng monosaccharide hay disaccharide? Giải thích.
OFFICIAL
d) Các gốc đường được gắn vào đầu khử hay đầu không khử của tetrasaccharide ban đầu? Giải thích.
e) Dựa vào tốc độ gắn từng loại đường, giải thích tại sao chỉ quan sát thấy trên bản sắc ký các sản phẩm tổng hợp có số gốc đường là số lẻ.
f) Khi nghiên cứu về màng nhầy của vi khuẩn cố định đạm sống tự do trong đất Azotobacter vinelandii ở các điều kiện sống khác nhau, người ta đo được độ dày trung bình của lớp màng nhầy vi khuẩn trong điều kiện X là 0,01 µm và trong điều kiện Y là 0,05 µm. Khả năng cố định đạm của vi khuẩn A. vinelandii cao hơn ở điều kiện X hay Y? Giải thích. Hướng dẫn
a. Enzyme hyaluronan synthase xúc tác gắn các monosaccharide vào tetrasaccharide ban đầu. Vì: khi bổ sung riêng phân tử GlcNAc (làn 3), enzyme đã xúc tác tạo thành phân tử có 5 gốc đường.
b. Các gốc đường được gắn vào đầu không khử của tetrasaccharide ban đầu. Vì: khi bổ sung riêng phân tử GlcUA (làn 4), sự tổng hợp không xảy ra còn khi bổ sung phân tử GlcNAc sự tổng hợp xảy ra. Tetrasaccharide ban đầu có các gốc đường liên kết xen kẽ nhau. Do đó, GlcNAc phải được gắn vào đầu có gốc GlcUA (đầu không khử) của tetrasaccharide ban đầu. c. Do tốc độ gắn các gốc GlcNAc nhanh hơn nhiều so với gốc GlcUA nên các sản phẩm trung gian có gốc GlcNAc ở đầu không khử (diễn ra sự tổng hợp) sẽ tích lũy do đó các sản phẩm này có số gốc đường là số lẻ d. Điều kiện Y. Vì vi khuẩn A. vinelandii sống tự do trong đất nên khi chúng cố định đạm bằng phức hệ nitrogenase trong tế bào chất thì O2 phải được giữ một phần bên ngoài tế bào nhờ lớp màng nhày để không ức chế phức hệ nitrogenase. Câu 3:
DẠYKÈMQUYNHƠN
461
Một nhà nghiên cứu có 10 chủng vi khuẩn E.coli mang các đột biến khác nhau nằm trên operon lac của các vi khuẩn này. Khi phân tích ADN của vi khuẩn, nhà nghiên cứu đã thấy mỗi chủng mang 1 trong 5 đột biến sau: lacZ, lacY, lacI, lacIS hoặc lacOC. Nhà nghiên cứu này cũng biết chắc chắn rằng chủng 6 là chủng đột biến gen lacZ. Sau đó nhà nghiên cứu này đã phân lập được đoạn ADN mang operon lac từ mỗi chủng (gọi là chủng cho) và biến nạp đoạn ADN này vào các chủng khác, tạo ra chủng lưỡng bội từng phần (gọi là chủng nhận). Các chủng nhận được nuôi trên môi trường tối thiểu chứa lactose là nguồn cung cấp carbon duy nhất. Sự sinh trưởng của các chủng được ghi lại trong hình dưới (+ biểu thị chủng đang sinh trưởng, - biểu thị chủng không sinh trưởng được). 23 - + 4 - + + 5 + + + + 6 - + + + + 7 - + + + +8 - - - - + -9 + + + + + + + + 1 0 - + - + + + + - + 1 2 3 4 5 6 7 8 9 Nếu quy ước sản lượng enzyme β-galactosidase và permase của kiểu hình kiểu dại sản xuất ra trong điều kiện có lactose, thiếu glucose đạt giá trị 100; ở
OFFICIAL
DẠYKÈMQUYNHƠN
462
ng thời cả glucose
Hãy xác định giá
ị
ản
ỗi
bội ở bảng trên ở trong
a) Chỉ có lactose; b) Chỉ có glucose; c) Không
cả lactose
d) Có cả glucose
điều kiện có đồ
và lactose đạt giá trị 10.
tr
s
lượng tương đương của các enzym ở m
dòng lưỡng
4 điều kiện môi trường của operon Lac:
có
và glucoe;
và lactose.
Hướng dẫn
- Chúng ta nhận thấy, chủng 1 và 8 ức chế tất cả các chủng khác, trừ chủng 5 và 9; chủng 6 và 7 là đột biến xảy ra trên cùng 1 gen; chủng 2 và 4 xảy ra trên cùng 1 gen, nhưng khi kết hợp với nhau thì vẫn biểu hiện kiểu hình kiểu dại; chủng 3 và 10 là các đột biến xảy tra trên cùng 1 gen.
Vậy, từ dữ kiện của bài ra, ta kết luận: chủng 1 và 8 là đột biến lacIS, chủng 3 và 10 là đột biến lacY; chủng 5 và 9 là đột biến lacOC, chủng 2 và 4 là lacI-, chủng 6 và 7 là lacZ. - Các dòng lưỡng bội của 5 chủng đột biến trên được tạo thành là: 1-1; 1-2; 1-3; 1-5; 1-6; 2-2; 2-3; 2-5; 2-6; 3-3; 3-5; 3-6; 5-5; 5-6 và 6-6.
OFFICIAL
(1-2)
(1-3)
(1-5)
(1-6)
DẠYKÈMQUYNHƠN
463
L
L
L
L
L
Dòng Chỉ có glucose Chỉ có lactose Có cả lac và glu Không có cả Lac và Glu
acZ
acY
acZ
acY
acZ L acY L acZ L acY lacIS/lacIS (1-1) 0 0 0 0 0 0 0 0 lacIS/lacI-
0 0 0 0 0 0 0 0 lacIS/lacY-
0 0 0 0 0 0 0 0 lacIS/lacO C
1 0 1 0 1 00 1 00 1 0 1 0 1 00 1 00 lacIS/lacZ-
0 0 0 0 0 0 0 0 lacI-/ lacI(2-2) 2 0 2 0 2 00 2 00 2 0 2 0 2 00 2 00 lacI-/ lacY- (2-3) 0 0 2 00 1 00 2 0 1 0 0 0 lacI-/ lacOC (2-5) 2 0 2 0 2 00 2 00 2 0 2 0 2 00 2 00 lacI-/ lacZ(2-6) 0 0 1 00 2 00 1 0 2 0 0 0 lacY-/ 0 0 2 0 2 0 0 0
lacY- (3-3) 00 0 lacY-/ lacOC(3-5)
1 0 1 0 2 00 1 00 2 0 1 0 1 0 1 0 lacY-/ lacZ- (3-6)
0 0 1 00 1 00 1 0 1 0 0 0 lacOC/ lacOC (5-5) 2 0 2 0 2 00 2 00 2 0 2 0 2 00 2 00 lacOC/ lacZ- (5-6) 1 0 1 0 1 00 2 00 1 0 2 0 1 0 1 0 lacZ-/ lacZ- (6-6) 0 0 0 2 00 0 2 0 0 0
Câu 4: Enzim dcm methylase xúc tác phản ứng thêm nhóm methyl vào nucleotit Cytosine thứ 2 trong trình tự CCAGG ở E. coli K-12. Mặt khác, các Cytosine ở vùng trình tự của gen lacI là điểm nóng của đột biến thay thế đồng hoán. Hãy đề xuất 2 phương pháp để có thể khẳng định tần suất đột biến tăng lên ở vị trí này là do methyl hóa cytosine hay không?
Hướng dẫn
- Có 2 nguyên nhân gây đột biến là: do enzym methylase hoặc đột biến thay thế đồng hoán Cytosine thành Timin trong vùng điểm nóng gây đột biến.
- Để xác định nguyên nhân gây đột biến do nguyên nhân nào, ta thực hiện biện pháp knock out các điểm đột biến.
+ Gây đột biến gen dcm tổng hợp dcm methylase. Nếu tần số đột biến giảm, chứng tỏ nguyên nhân đột biến là do dcm methylase. Nếu tần số đột biến vẫn tăng, chứng tỏ nguyên nhân đột biến do đột biến đồng hoán ở vị trí điểm nóng đột biến.
+ Phương pháp thứ 2, loại bỏ nucleotit thứ 2 trong trình tự CCAGG, quan sát tần số đột biến để kết luận nguyên nhân.
Câu 5:
Trong
DẠYKÈMQUYNHƠN
464
OFFICIAL
loài thực vật thuộ
ằng nhiệ
thí nghiệm
Kết quả củ
thí nghiệm lai DNA ở 06
c cùng chi Vicia, DNA đã được phân lập từ mỗi loài, gây biến tính b
t và phân mảnh thành từ phân đoạn DNA nhỏ rồi tiến hành hồi tính. Ở một
khác, DNA từ vi khuẩn E. coli cũng được phân lập và tiến hành tương tự.
a 02 thí nghiệm được thể hiện như đồ thị bên dưới.
Hướng dẫn a)
a. So sánh và giải thích sự khác biệt trong quá trình hồi tính DNA của vi khuẩn E. coli so với các mẫu DNA ở 06 loài thực vật thuộc chi Vicia?
b. Hãy xác định giai đoạn các trình tự lặp lại và các trình tự duy nhất của DNA ở 06 loài thực vật thuộc chi Vicia
OFFICIAL
- So với DNA của 06 loài thực vật, thì DNA của vi khuẩn E. coli hồi tính nhanh hơn nhiều lần; phần trăm phân đoạn DNA bắt cặp tại từng thời điểm khác nhau gần như tuyến tính.
- Nguyên nhân:
+ DNA của vi khuẩn E. coli có kích thước nhỏ → nhìn chung tốc độ hồi tính nhanh; ít vùng trình tự lặp lại → quá trình hồi tính diễn ra đơn giản, không chia thành những pha khác nhau (tuyến tính).
+ DNA ở 06 loài thực vật có kích thước lớn → nhìn chung tốc độ hồi tính chậm; có nhiều vùng trình tự lặp lại → quá trình hồi tính diễn ra phức tạp hơn, có nhiều pha hồi tính nhanh, chậm khác nhau.
b)
- Trong quá trình hồi tính của DNA, các đoạn trình tự lặp lại sẽ được bắt cặp trước do sự tương đồng về trình tự cao → hồi tính ở pha sớm; các đoạn trình tự duy nhất thường được bắt cặp chậm hơn → hồi tính ở pha muộn.
- Nhìn chung, rõ ràng nhất là giai đoạn 1 giờ đầu tiên ở cả 06 loài đều có sự hồi tính của vùng trình tự lặp lại; các giai đoạn còn lại sẽ có sự hồi tính của vùng trình tự duy nhất.
Câu 6: Khói thuốc lá có chứa một chất có tiềm năng gây đột biến là benzo (a) pyrene (viết tắt là B(a)P). Một thí nghiệm đã được tiến hành dựa trên nguyên tắt của Ames như sau:
DẠYKÈMQUYNHƠN
465
- Thí nghiệm 1: Nuổi chủng vi khuẩn khuyết dưỡng histidine (his-) trên môi trường dinh dưỡng không bổ sung histidine và có bổ sung B(a)P. Kết quả cho thấy không khác so với mẫu đối chứng, không bổ sung B(a)P.
- Thí nghiệm 2: Tiêm B(a)P vào chuột mỗi tháng một lần. Kết quả cho thấy chuột có xuất hiện khối u ở phổi so với mẫu đối chứng là không tiêm B(a)P.
- Thí nghiệm 3: Ủ B(a)P với dịch chiết của tế bào gan và bổ sung vào môi trường nuôi cấy tương tự ở thí nghiệm 1. Kết quả cho thấy vi khuẩn mọc lên khuẩn lạc nhiều hơn thí nghiệm 1.
a) Dựa vào kết quả thu được của 03 thí nghiệm trên để xác định B(a)P có phải là tác nhân đột biến gen hay không? Vai trò của mẫu đối chứng ở thí nghiệm 1 là gì?
OFFICIAL
b) Trong hơn 50 năm qua, tỷ lệ bệnh nhân ung thư phổi ngày càng tăng trong khi tỷ lệ bệnh nhân ung thư dạ dày ngày càng giảm. Hãy đưa ra giải thích cho xu hướng này.
c) Theo thống kê, có hơn 50% trường hợp ung thư vú có thể được trị khỏi nhưng chỉ có khoảng 20% trường hợp ung thư phổi được trị khỏi. Hãy đưa ra giải thích cho sự khác biệt này. Hướng dẫn
a) - Thí nghiệm 1: Không có sự khác biệt so với mẫu đối chứng → B(a)P không phải là tác nhân gây đột biến nên chủng đột biến khuyết dưỡng histidine không phục hồi kiểu hình hoang dại.
- Thí nghiệm 2: Khối u xuất hiện so với mẫu đối chứng → B(a)P là tác nhân ung thư (carcinogen) với tế bào chuột.
- Thí nghiệm 3: Khuẩn lạc mọc nhiều hơn ở thí nghiệm 1 → chủng đột biến có sự phục hồi đột biến khuyết dưỡng histidine thành kiểu hình hoang dại → B(a)P đã được chuyển hóa nhờ dịch chiết của gan tạo thành chất gây đột biến.
→ Vậy chất B(a)P không tự thân gây đột biến mà cần phải được chuyển hóa thành một chất khác mới có khả năng gây đột biến. Đây được xem là tiền tác nhân gây đột biến (promutagen). Mẫu đối chứng ở thí nghiệm 1 có vai trò chứng tỏ rằng B(a)P không tác động đến sự phục hồi kiểu hình đột biến của vi khuẩn, nếu có xuất hiện khuẩn lạc là do đột biến ngẫu nhiên.
b) Tác động của môi trường ảnh hưởng đến hơn 90% tỷ lệ mắc ung thư ở người. Xu hướng trên thể hiện sự thay đổi mức độ tác động của tác nhân ung thư từ môi trường lên cơ thể người. Mức độ ô nhiễm không khí và xu hướng sử dụng khối thuốc lá liên tục tăng là nguyên nhân chính làm tăng tỷ lệ ung thư phổi; trong khi đó chất lượng thực phẩm ngày càng được
DẠYKÈMQUYNHƠN
466
quan tâm, hàm lượng nitrite được loại bỏ khỏi bữa ăn là nguyên nhân chính làm giảm tỷ lệ ung thư dạ dày.
c) Tỷ lệ trị khỏi các loại ung thư khác nhau là khác nhau, điều này ảnh hưởng bởi nhiều yếu tố:
- Ung thư vú dễ phát hiện sớm hơn ung thư phổi nên khả năng xây dựng phát đồ trị liệu sẽ hiệu quả hơn so với ung thư vú. Đa phần, các trường hợp ung thu phổi được phát hiện ở giai đoạn muộn.
- Ung thư vú có thể dễ dàng được loại bỏ dễ dàng và hiệu quả bằng các phương pháp phẫu thuật trong khi ung thư phổi hầu như rất khó để trị bằng phương pháp phẩu thuật mà phải tác động bằng các biện pháp ít đặc hiệu như chiếu xạ, hóa trị,… sẽ để lại nhiều tác dụng phụ.
Câu 7: HNA19-Khi một enzim có mặt ở một loài vi khuẩn, thì con đường chuyển hoá mà enzim đó tham gia thường tồn tại trong loài vi khuẩn này. Bảng 1.1 là tên enzim và phản ứng mà enzim xúc tác được sử dụng làm chỉ thị cho sự xuất hiện của các con đường chuyển hóa mà nó tham gia. Bảng 1.2 thể hiện sự có mặt hay vắng mặt của một số enzim ở bốn loài vi khuẩn khác nhau 1,2, 3 và 4. Bảng 1.1. Enzim và phản ứng xúc tác tương ứng Tên enzim Phản ứng xúc tác
Lactat đêhiđrôgenaza (LDH)
Axit piruvic + NADH → axit lactic 4NAD+ Alcohol đêhiđrôgenaza (ADH)
Axêtanđêhit 4- NADH → Êtanol + NAD+ Xitôcrôm C oxidaza
Vận chuyển electron từ xitôcrôm C tới xitôcrôm a
ATP sintetaza
Vận chuyển H+ qua màng tạo ATP từ ADP và Pi
Xúc tác gắn CoASH với Axit pyruvic để tạo acetyl-CoA Bảng 1.2. Sự có mặt (+) và vắng mặt (-) của mỗi loại enzim trong từng loài vi khuẩn
Phức hợp Pyruvate dehydrogenase
L oài vi khuẩn
Tên enzim LDH AD H Xitôcrô m c oxidaza ATP sinte taza
Phức hợp Pyruvate dehydrogenase L oài 1 - + - +L oài 2 + - - +L oài 3 + - + + -
DẠYKÈMQUYNHƠN
OFFICIAL 467
L
oài 4 - + + + + Hãy cho biết:
a) Loài vi khuẩn nào không thể thực hiện được hô hấp hiếu khí? Giải thích.
b) Các sản phẩm chính mỗi loài vi khuẩn tạo ra sau quá trình chuyển hóa glucozo.
c) Các loài trên sẽ phát triển như nào nếu như bổ sung oxi phân tử vào môi trường.
Hướng dẫn
a).
- Các vi khuẩn không thể thực hiện được hô hấp hiếu khí gồm: loài 1, loài 2, loài 3.
- Loài 1 và loài 2 thiếu xitocrom c oxidaza là enzim chính trong thành phần của quá trình hô hấp hiếu khí (enzym trong chuỗi truyền điện tử).
- Loài 3 thiếu Phức hợp Pyruvate dehydrogenase của giai đoạn trung gian hình thành Acetyl- coA (trước chu trình Crep)
b)
- Loài 1 tạo rượu etanol, CO2 và ATP.
- Loài 2 tạo axit lactic, ATP.
- Loài 3 tạo axit lactic, ATP.
- Loài 4 khi có mặt oxi tạo CO2, H2O và ATP; khi không có mặt oxi tạo rượu etanol, CO2 và ATP.
c)
- Khi bổ sung vào môi trường oxi phân tử thì loài 2 và loài 3 sẽ bị chết nhanh nhất, khả năng chúng là vi khuẩn lactic. Trong điều kiện có oxi, vi khuẩn lactic bị ức chế sinh trưởng vì nó là vi khuẩn kị khí bắt buộc, tế bào thiếu enzim catalaza, SOD giúp chúng tồn tại trong điều kiện có oxi.
- Loài 1 là nấm men rượu – kị khí không bắt buộc. Khi có oxi nấm men chuyển sang hô hấp hiếu khí. Glucôzơ bị phân hủy hoàn toàn; Năng lượng tạo ra nhiều (38 ATP); khi đó chất nhận e là khí oxi.
- Loài 4 có khả năng sống được trong cả khi có oxi hoặc không nên sinh trưởng bình thường.
Câu 8: Người ta phân lập được nhiều thể đột biến khuyết dưỡng liên quan đến một con đường chuyển hóa ở một vài vi khuẩn. Khi bổ sung một trong các hợp chất A, B, C, hoặc D vào môi trường nuôi cấy, người ta thấy các thể đột biến sinh trưởng như liệt kê ở bảng 1 dưới đây (dấu + là sinh trưởng được, dấu – là không sinh trưởng
DẠYKÈMQUYNHƠN
468
OFFICIAL