Primates of the past
If, via some Jurassic Park-like resurrection, the earliest primates appeared in a zoo somewhere, they wouldn’t attract much attention. Few people would adulate our ancestors absent the knowledge that their issue includes humankind. Instead, these small, arboreal mammals would have no more claim to fame than flying lemurs or tree shrews have. If a time machine enabled us to visit the first primates in their day, they’d probably be disappointing: just another species living the high life in trees. We would witness little more than ordinary mammals eating a few bugs, berries, and blossoms at night and dozing through the day, and they did little else for millions of years.
Then one day, about 50 million years ago, a hot-house world teeming with trees saw the first glimmer of a gradual cooling trend that would continue, off and on, for 16 million years. In time, it caused rainforests to contract: bad news for our primate ancestors, which depended on forests for life and limbs. More hardship followed, beginning 34 million years ago when global cooling accelerated sharply. Forests contracted yet more, nutrients began to vary markedly from season-to-season, and large predators appeared on the scene. It wasn’t pleasant to be a primate back then, but somehow our ancestors survived. Many species did not. The primates that made it through those tough times gave birth to all that followed.
It’s only natural for modern primates to pop into the mind’s eye at the mention of extinct ones, but primates of the past differed from those alive today. For one thing, when the earth cooled 34 million years ago, most primates were small animals. Only later did some become the larger animals we know today. Likewise, today’s primates live long lives, but that hasn’t always been the case, either. Our ancestors had short lifespans, like most small mammals. The transition from a brief life to a lengthy one was a big part of primate evolution, which means that it was also a big part of human evolution.
Today, nearly everyone understands that human evolution is part of primate evolution. As obvious as this fact seems, we have only known that we are primates for about 0.1% of our existence as a species, maybe less. It was Linnaeus who first classified people as primates ~300 years ago, and it caused quite a commotion at the time. Even to this day, some people renounce their primate heritage, as if unsavory behavior by monkeys at the zoo reflects badly on all of us. On the contrary, I’m proud to be a primate, despite the pompous name of our order.
Primates, prelates, and priority
Why pompous? Unlike other orders, which have labels that emphasize their physical features or behaviors, ours coveys a deepseated conceit. Dictionaries and etymologies reveal the selfcongratulatory nature of the word “primate.” Merriam-Webster (https://www.merriam-webster.com/dictionary/primate) provides a typical example, defining “primate” as:
. . . a bishop who has precedence in a province, a group of provinces, or a nation. archaic: one first in authority or rank: leader.
. . . any of an order (Primates) of mammals that are characterized especially by advanced development of binocular vision resulting in stereoscopic depth perception, specialization of the hands and feet for grasping, and enlargement of the cerebral hemispheres and that include humans, apes, monkeys, and related forms (such as lemurs and tarsiers).
The first usage is uncommon, but it does turn up now and then. The Primate of All England is the Archbishop of Canterbury, and the Pope is also the Primate of Italy. What could these prelates possibly have in common with an order of mammals? The answer is that all the meanings of “primate” derive from the Latin primas, meaning first. Thus, primates were once thought to be an order that ranked first in an ordered sequence of orders.
In contrast to the narcissism inherent in the name “primate,” other orders have more modest monikers. Bats, for example, compose the order Chiroptera, derived from the Greek kheirfor hand and pteron for wing. In any language, the name hand-wing captures the essence of being a bat. Charmingly, Eulipotyphla—a group that includes hedgehogs along with various shrews and moles—means fat and blind: not very flattering and a bit of an exaggeration, but reasonably descriptive. Rodentia comes from the Latin for gnaw, a specialization of the order; Scandentia derives from the Latin for climb, which tree shrews do quite well; and Carnivora means flesh eater. These names say something about each order’s distinctive characteristics, but ours implies that we outrank ordinary orders.
At long last, the practice of ranking animals based on their similarity to humans, called the scala naturae or the evolutionary scale, has begun to disappear from the neuroscience literature. According to that outmoded idea, the more closely an animal resembles us, the higher they perch on the “ladder of life.”
References to a phylogenetic scale persisted in our field for far too long, but evolutionary biologists have known better for generations. If you hear or read anything about higher primates or lower vertebrates, you are witnessing the dying embers of the scala naturae. Contemporary biology rejects the idea that orders have an order.
More importantly, no amount of similarity to humans alters evolutionary relationships. Even if dolphins could squeak sonnets in iambic pentameter, they would still be more distantly related to humans than rabbits are; elephants might mourn their dearly departed friends, but rats remain more closely related to us without giving a rat’s ass about the demise of their fellow vermin; and polar bears might make magnificent mothers, but they are far more distant relatives than the worst mothers in the mammalian world: tree shrews, which only visit their newborns every two days or so— and then only briefly. To understand cortical evolution in primates, phylogenetic relationships matter; the place of primates on an imaginary “ladder of life” does not. Primates are the animals most
closely related to us, but they don’t rank higher on a phylogenetic scale.
Primates in their proper place
Regardless, primates don’t belong on ladders; the proper place of primates is in trees. Not only did early species live in trees, but primates are one branch of the tree of life. As our ancestors raced for survival along both kinds of branches, they evolved a large cerebral cortex. In primates of the present day, most of the brain consists of cortex, but that wasn’t true of early species. Like the Dodo’s running party, every living primate has won a race, and a colossal cortex is one of the prizes.
The inspiration for Lewis Carroll’s Dodo might provide a clue about why primates won that particular prize. Dodos were large, flightless pigeons that once lived in dense forests on Mauritius, an island in the Indian Ocean. They thrived for millennia; then one fine day in the 1600s human colonists arrived, and the dodo’s days were numbered. Several anthropogenic factors contributed to their extinction, including the introduction of pigs, rats, and monkeys that plundered the dodo’s nests. The fact that islanders hunted these 20pound birds didn’t do them any good either, but habitat reduction from deforestation was probably the main reason for their demise. Likewise, deforestation has caused many primate species to go the way of the dodo. The earth’s climate has cooled several times over the past 50 million years, which decreased atmospheric moisture and caused rainforests to dry and contract. Global cooling threatened species that relied on the nutrients and protection that rainforests provide, as primates did back then and many do today. The effects of global climate change might provide some hints about why primates evolved such a large cerebral cortex.
As its subtitle says, this book explores what primates are, what primates were, and why the cortex changed. Today, all primates are remarkably brainy; 50 million years ago, primates were run-of-the-
mill placental mammals with a cortex to match. The life and times of primates in-between are why the cortex changed.
Acknowledgments
I thank the following colleagues for their comments on individual chapters: Mary Baldwin, Betsy Murray, Daniel Pine, Todd Preuss, and Georg Striedter. Samantha White, Caleb Darden, and Jensen Palmer read the entire book and discussed the chapters with me at length, and I thank Mark Laubach for arranging those meetings. My commissioning editor at Oxford University Press, Martin Baum, encouraged me to undertake this project and provided crucial support along the way, as did Phoebe Aldridge-Turner, who guided the manuscript from submission to publication. Anya Hastwell edited the manuscript with skill and dedication.
I owe a different kind of debt to Jon Kaas, which the dedication of this book acknowledges. The book you are holding in your hands would not exist if he and his students hadn’t performed the decades of research that they did. One of them, Leah Krubitzer, deserves special recognition. When I was a graduate student in the 1970s, my research focused on the somatosensory cortex of rats, cats, squirrels, squirrel monkeys, and both rhesus and crab-eating macaques. I also did some work on the auditory cortex of mustache bats. Naturally, I tried to understand how the cortical maps of these diverse mammals related to each other. But it was all a muddle until one day at a Society for Neuroscience annual meeting. Leah presented a poster that summarized her cortical mapping studies of the duckbilled platypus and some ideas about cortical evolution in mammals. Within half an hour, my knowledge of comparative cortical anatomy became much better organized. This book is one among many consequences of that 30-minute discussion. Todd Preuss also deserves special recognition for his contributions to understanding cortical evolution in primates, and more than a few of the ideas in this book stem from my coauthors on previous books, in
chronological order: Reza Shadmehr, Dick Passingham, Betsy Murray, Kim Graham, and Mary Baldwin, whose inspired artwork enlivens many of the illustrations in this book.
Listoffiguresandtables
Referencefigure
Listofabbreviations:text
Listofabbreviations:figures
Epigraph
PART I. WHAT PRIMATES ARE
1.
Topics tackled
Overview
Why?
What?
When?
Why now?
Why not?
Why not now?
Why try?
2. Contents
Chapter summary
References
Compact cladistics
Overview
Introduction
Taxing terminology
Homology, homoplasy, and analogy
Trees and scales
Old and new areas
Chapter summary
References
Present primates
Overview
Introduction
Taxing taxonomy
Principal primate clades
Strepsirrhines
Haplorhines
Catarrhines
Chapter summary
To “the” or not to “the”
References
PART II. WHAT PRIMATES WERE
Prolog to paleontology
Overview
Introduction
The dating scene
Bodies from bones
Focus on forests
Chapter summary
References
5.
Arboreal adaptations
Overview
Introduction
Primates true and stem
Into the trees
Chapter summary
Cortical considerations
References
Primate paleoecology
Overview
Introduction
Paleocene plesiadapiforms
Eocene Euprimates
Oligocene openings and Miocene monkeys
Miocene modifications and Plio-Pleistocene primates
Chapter summary
References
PART III. WHAT PRIMATE CORTEX WAS
Great grades of gray
Overview
Introduction
Measures and misconceptions
Grades and clades
Eocene expansions
Chapter summary
References
Greater grades of gray
Overview
9.
Introduction
Miocene monkeys and apes
Plio-Pleistocene hominins
Body or brain?
Chapter summary
References
Tempo and temperature
Overview
Introduction
Time travel
Cooling and crisis
Corticalization and speciation
Cortex and corpus
Chapter summary
References
10.
Other orders
Overview
Introduction
Pride of place
The origin of mammals
Brain expansion
Chapter summary
Parietofrontia
References
PART IV. WHAT PRIMATE CORTEX IS
11.
Cortical comparisons
Overview
Introduction
A Declaration of Independence
Flying primates, feathered apes
Crucial comparisons
Misconceptions: minor and massive
Chapter summary
References
Suites of specializations
Overview
Introduction
From tip to toe
Suite success
Chapter summary
References
Anthropoid adaptations
Overview
Introduction
The big chill
Frontal-field phylogeny
Changes at the top
Sights and sounds on the side
Guilt by association
Chapter summary
References
Human hemispheres
Overview
Introduction
Whole hemispheres
Area analysis
Sensational size
Temporal tracts
Allocortical alterations
Cortex and chromosomes
Chapter summary
References
PART V. WHY THE CORTEX CHANGED
Eocene expansions
Overview
Introduction
What’s new is old
The cortex complete
Eocene enlargements
Chapter summary
References
Anthropoid augmentations
Overview
Introduction
Groups, grub, and gray matter
Statistics and significance
Climate and cortex
Principal proposal
Chapter summary
References
17. 18. Pleistocene prizes
Overview
Introduction
Suggestions for selection
Self and social systems
Chapter summary
References
Corticalization and composition
Overview
Introduction
Cortical chauvinism
Parts and primates
Questions and conclusions
Instead of intelligence: representations
Chapter summary
References
Epilogue
Crucialglossary Extendedglossary
Index
List of figures and tables
Figures
Reference figure
The brain of an extinct primate
Graphical definitions of cladistic terms
Phyletic dwarfism in callitrichid primates
Differing views of primate relations
Mammalian evolutionary tree before molecular phylogenies
An evolutionary tree of eutherian mammals
An evolutionary tree of Euarchontoglires
An evolutionary tree of Euarchontoglires, emphasizing anthropoids
An evolutionary tree of catarrhines
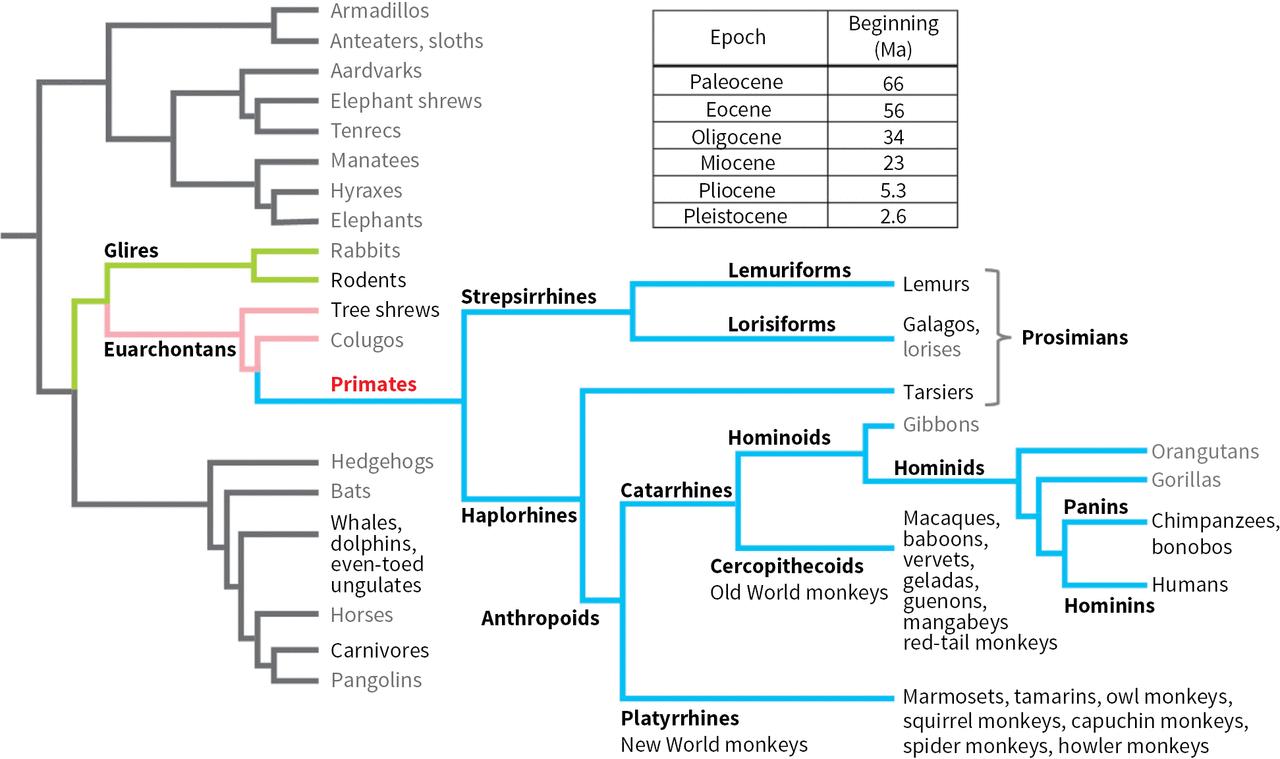
Geological epochs and eras
Chronograms of Euarchontoglires and primates
Tooth morphology and diet
Global surface temperature during the Cenozoic
An evolutionary tree of primates
Dentition in plesiadapiforms
Eocene geography
Convergent evolution in distantly related primates
Chronogram of Euarchontans
Anthropoid extinctions during the Oligocene bottleneck in North Africa
The evolution of body size in anthropoids
Selective pressures and anthropoid adaptations
Evolutionary trajectories of femur morphology in anthropoids
Climate change and adaptive radiations
Brain–body relationships in primates and other mammals
Virtual cranial endocasts of fossil primates
Eocene grade-shifts in encephalization
Eocene grade-shifts in corticalization
Longevity and encephalization
Summary of Eocene grade-shifts
Encephalization quotients in mammals
Upward grade-shifts during anthropoid evolution
The emergence and loss of sulci in anthropoids
Olfactory bulb contraction
Cortical expansion in hominins
Encephalization in hominins
Encephalization quotients and estimated divergence times in hominins and panins
Changes in human brains and brain shape in hominids
Skull shape in Homospecies
Anterior temporal bulging in four human species and chimpanzees
Grade-shifts in brain size–body size allometry in primates
Summary of Eocene and Miocene grade-shifts
Upward grade-shifts in cortex size and periods of global cooling
Brain size, body size, land productivity, and rainfall
EQ values in fossil and modern mammals
Cortical grade-shifts in cetaceans
Cortical grade-shifts in artiodactyls
Cortical grade-shifts in carnivores
Cortical grade-shifts in rodents
Diversity of Euarchontoglires cortex
Tree shrew phylogeny
Grade difference in posterior parietal cortex
Ideas about homologies among frontal areas in rodents and primates
Types of cerebral cortex
Cortical organization in mammals
Cortical maps in selected Euarchontoglires
Action maps in selected Euarchontoglires
Transcortical networks
Relative size of the frontal lobe in Euarchontoglires
Cortical maps of an anthropoid and a strepsirrhine
Types of frontal cortex in selected Euarchontoglires
Phylogeny of traits involved in grasping and manipulation
Fingers, “foveas,” and frugivory
Action maps in galagos, squirrel monkeys, and rhesus macaques
Prefrontal predominance in humans
Expansion of the prefrontal cortex in humans
Preferential expansion of prefrontal cortex
Myelin density in the cortex of four anthropoids
Relative expansion of cortical regions
Evolutionary changes in the temporal lobe of anthropoids and frontotemporal pathways
Hippocampal contraction in anthropoids, followed by expansion in humans
Neuronal density and counts for the cerebral cortex
Genetics of cortical expansion in hominins
Conjunctive representations in the cortex of primates
The ventral visual stream in anthropoids
Primate social and mating systems
Relation between clique size and corticalization
Anthropoid synapomorphies
The sources of visual feature conjunctions in frontal cortex: dorsal and ventral visual streams
Transcortical networks in humans
Technological knowledge and cortical expansion
Cortical evolution in primates
Cortical evolution in primates: simplified
Table
17.1
Selective factors that might have contributed to cortical expansion in hominins
Reference figure. Evolutionary relationships among the mammals mentioned in this book. The table at the top gives onset dates for the geological epochs discussed most frequently in the text. Abbreviation: Ma, million years ago
List of abbreviations: text
A1 Primary auditory area
AF Arcuate fascicle
AGm Medial agranular frontal area (in rodents)
AIP Anterior intraparietal cortex
ARHGAP11B Rho GTPase-activating protein 11B
BOLD Blood oxygen-level decrease
C3 and C4 Two metabolic pathways for photosynthesis
CA3 Third area of Ammon’s horn (Cornu Ammonis), the hippocampus
DM Dorsomedial visual area
DS–DC Diagonal sequence, diagonally coupled [gait]
EECO Early Eocene climatic optimum
EOCT Eocene–Oligocene climatic transition
EQ Encephalization quotient
ES Extrastriate cortex
FEF Frontal eye field
fMRI Functional magnetic resonance imaging
FOXP2 A gene involved in orofacial coordination
Fr2 Second frontal area (in rodents), also known as AGm
FST Fundus superior temporal area
HARE5 Human-accelerated regulatory enhancer 5
HSD Hominini-specific deletion
IQ Intelligence quotient
ICZN International Commission on Zoological Nomenclature
IFOF Inferior frontal–occipital fascicle
ILF Inferior longitudinal fascicle
L Long wavelength [cone photoreceptor]
LCA Last common ancestor
LGN Lateral genicular nucleus
LIP Lateral intraparietal cortex
LT-ICMS Long-train intracortical microstimulation
LUCA Last universal common ancestor
M Middle wavelength [cone photoreceptor]
M1 Primary motor cortex
M1c Caudal primary motor cortex
M2 In primates, the supplementary motor area; in tree shrews, a motor area
Ma Million years ago
MECO Middle Eocene climatic optimum
MIP Medial intraparietal area
MMCO Middle Miocene climatic optimum
MMCT Middle Miocene climatic transition
MRI Magnetic resonance imaging
MST Middle superior temporal area
MT Middle temporal area
MTc Middle temporal crescent area
NOTCH2NL Notch homolog 2 N-terminal-like protein
PC Principal component
PCA Principal components analysis
PETM Paleocene–Eocene thermal maximum
PFdl Dorsolateral prefrontal cortex
PFdm Dorsomedial prefrontal cortex
PFo Granular orbital prefrontal cortex
PFp Polar prefrontal cortex; frontal-pole cortex
PFvl Ventrolateral prefrontal cortex; also known as ventral prefrontal cortex
PIm Middle inferior pulvinar nucleus of the thalamus
PM 1., Premotor cortex (in primates); 2., posteromedial area (in rodents)
PMd Dorsal premotor cortex
PMv Ventral premotor cortex
Pom Posterior medial nucleus of the thalamus
PP, PPC Posterior parietal cortex
PPr Rostral posterior parietal cortex
PR Public relations
PrCm Medial precentral area (in rodents), also known as AGm
PV Ventral parietal somatosensory area
Rex Tyrannosaurus rex
RMA Rostral motor area (in rodents), also known as AGm
S Short wavelength [cone photoreceptor]
S1 Primary (or first) somatosensory area
S2
Second (or secondary) somatosensory area
SC Caudal somatosensory area
SMA Supplementary motor area, also known as M2
SRGAP2 Slit protein, roundabout-receptor GTPase-activating protein 2
ST-ICMS Short-train intracortical microstimulation
T . rex Tyrannosaurus rex
TA Anterior temporal area
TD Dorsal temporal area
TP Posterior temporal area
TPJ Temporal–parietal junction
V1 Primary (or first) visual area; striate cortex
V2 Second (or secondary) visual area
V3 Third visual area
VIP Ventral intraparietal cortex
List of abbreviations: figures
A. Australopithecus
a, A Anterior
A1 Primary auditory cortex
AB Auditory belt cortex
AC Anterior cingulate cortex
AF Arcuate fascicle
AGm Medial agranular area, also known as Fr2, PrCm, and RMA
AIP Anterior intraparietal area
Anc Ancestral
AP Anterior-posterior
APB Posterior auditory belt cortex
AS Arcuate sulcus
Aud Auditory cortex
c Caudal
CA3 An area within the hippocampus
CBLN2 Cerebellin 2precursor
cc Corpus callosum
CG Cingulate gyrus
CgS Cingulate sulcus
CLI Claustral isocortex
CMc Caudal cingulate motor area
CMr Rostral cingulate motor area
CMv Ventral cingulate motor area
CoA Cortical nucleus of the amygdala
CS Central sulcus
d, D Dorsal
DF Dorsal frontal area
DL Dorsolateral visual area
DLPFC Dorsolateral prefrontal cortex, also abbreviated PFdl
DM Dorsomedial visual area
DMPFC Dorsomedial prefrontal cortex, also abbreviated PFdm
Dys Dysgranular cortex
EECO Early Eocene climatic optimum
EmC Extreme capsule
EOCT Eocene–Oligocene climatic transition
EQ Encephalization quotient
ERh Entorhinal cortex
ES Extrastriate visual areas
FEF Frontal eye field
FPC Frontopolar cortex, also abbreviated PFp
Fr2 Second frontal area, also known as AGm
FST Fundus of the superior temporal cortex
G Gustatory cortex, also known as the primary gustatory cortex
G1 Primary gustatory cortex
Gr Granular
H. Homo
Hippos Hippopotamuses
hl Hindlimb representation of the primary motor cortex
HSD Human-specific deletion (of a gene sequence)
i Inferior
Ia Agranular insular cortex
Id Dysgranular insular cortex
Ig Granular insular cortex
IFOF Inferior fronto-occipital fascicle
IL Infralimbic cortex
ILF Inferior longitudinal fascicle
Ins Insular cortex
IPS Intraparietal sulcus
IT Inferior temporal visual cortex
ITS Inferior temporal sulcus
l, L Lateral
LatS Lateral (Sylvian) sulcus (or fissure)
LCA Last common ancestor
LIP Lateral intraparietal area
LO Lateral orbitofrontal cortex
LunS Lunate sulcus
m, M Medial
M1 Primary motor cortex
M1c Caudal primary motor cortex