Download full Single-molecule fluorescence spectroscopy of the folding of a repeat protein 1st editi
Single-Molecule
Fluorescence Spectroscopy of the Folding of a Repeat Protein 1st Edition Sharona Cohen (Auth.)
Visit to download the full and correct content document: https://textbookfull.com/product/single-molecule-fluorescence-spectroscopy-of-the-fol ding-of-a-repeat-protein-1st-edition-sharona-cohen-auth/
More products digital (pdf, epub, mobi) instant download maybe you interests ...
Practical Fluorescence Spectroscopy 1st Edition Zygmunt (Karol) Gryczynski
Angle Resolved Photoemission Spectroscopy on High Temperature Superconductors Studies of Bi2212 and Single Layer FeSe Film Grown on SrTiO3 Substrate 1st Edition Junfeng He (Auth.)
Single-Molecule Fluorescence Spectroscopy of the Folding of a Repeat Protein
Springer Theses
Recognizing Outstanding Ph.D. Research
Aims and Scope
The series “Springer Theses” brings together a selection of the very best Ph.D. theses from around the world and across the physical sciences. Nominated and endorsed by two recognized specialists, each published volume has been selected for its scientific excellence and the high impact of its contents for the pertinent field of research. For greater accessibility to non-specialists, the published versions include an extended introduction, as well as a foreword by the student’s supervisor explaining the special relevance of the work for the field. As a whole, the series will provide a valuable resource both for newcomers to the research fields described, and for other scientists seeking detailed background information on special questions. Finally, it provides an accredited documentation of the valuable contributions made by today’s younger generation of scientists.
Theses are accepted into the series by invited nomination only and must fulfill all of the following criteria
• They must be written in good English.
• The topic should fall within the confines of Chemistry, Physics, Earth Sciences, Engineering and related interdisciplinary fields such as Materials, Nanoscience, Chemical Engineering, Complex Systems and Biophysics.
• The work reported in the thesis must represent a significant scientific advance.
• If the thesis includes previously published material, permission to reproduce this must be gained from the respective copyright holder.
• They must have been examined and passed during the 12 months prior to nomination.
• Each thesis should include a foreword by the supervisor outlining the significance of its content.
• The theses should have a clearly defined structure including an introduction accessible to scientists not expert in that particular field.
More information about this series at http://www.springer.com/series/8790
Sharona Cohen
Single-Molecule
Fluorescence Spectroscopy
of the Folding of a Repeat Protein
Doctoral Thesis accepted by Weizmann Institute of Science, Rehovot, Israel
Author Dr. Sharona Cohen
Department
of Chemical Physics
Weizmann Institute of
Science
Rehovot
Israel
Supervisor
Prof. Gilad Haran
Department
of Chemical Physics
Weizmann Institute of Science
Rehovot
Israel
ISSN 2190-5053
ISSN 2190-5061 (electronic)
ISBN 978-3-319-09557-8 ISBN 978-3-319-09558-5 (eBook)
DOI 10.1007/978-3-319-09558-5
Library of Congress Control Number: 2014946189
Springer Cham Heidelberg New York Dordrecht London
This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. Exempted from this legal reservation are brief excerpts in connection with reviews or scholarly analysis or material supplied specifically for the purpose of being entered and executed on a computer system, for exclusive use by the purchaser of the work. Duplication of this publication or parts thereof is permitted only under the provisions of the Copyright Law of the Publisher’s location, in its current version, and permission for use must always be obtained from Springer. Permissions for use may be obtained through RightsLink at the Copyright Clearance Center. Violations are liable to prosecution under the respective Copyright Law. The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use.
While the advice and information in this book are believed to be true and accurate at the date of publication, neither the authors nor the editors nor the publisher can accept any legal responsibility for any errors or omissions that may be made. The publisher makes no warranty, express or implied, with respect to the material contained herein.
Printed on acid-free paper
Springer is part of Springer Science+Business Media (www.springer.com)
Parts of this thesis have been published in the following documents:
Sharona S. Cohen, Inbal Riven, Aitziber L. Cortajarena, Lucia De Rosa, Luca D. D’Andrea, Lynne Regan, Gilad Haran, Probing the molecular origin of nativestate flexibility in repeat proteins. J Am Chem Soc, Volume 137, 2015, Pages 10367–10373.
Supervisor’s Foreword
The protein folding field has undergone enormous transformation in recent years, led mainly by the introduction of new techniques that facilitate the study of folding on the single-molecule level, based on either sensitive fluorescence methodology or force spectroscopy. The progress in the field has also been driven by the realization that, in addition to the more familiar globular proteins, there are novel groups of proteins whose folding behavior requires the development of new paradigms. Among these proteins are two important groups. The first is the group of intrinsically unfolded proteins, many of which fold only upon interaction with their binding partners. The second group includes the repeat proteins, a diverse set of proteins made of short structural units that repeat themselves and form nonglobular, elongated structures resembling solenoids.
The Ph.D. thesis of Dr. Sharona Cohen marries the two themes discussed above. Dr. Cohen employed single-molecule fluorescence spectroscopy to probe the folding and dynamics of tetratricopeptide repeat proteins, whose repeat units consist of 34 amino acids. In particular, she studied a designed protein made of three repeats, with fluorescent probes inserted in several positions. Fluorescence resonance energy transfer (FRET) was used to learn how the conformation of the protein changes in the presence of chemical denaturants.
The big surprise in Dr. Cohen’s results is that in addition to the unfolding transition of the protein, clearly observed in single-molecule FRET histograms, there is interesting behavior related to changes in the structure of the folded state. As it turns out, the folded protein becomes more and more loose as the concentration of denaturant is increased, like a solenoid whose spring constant is weakened gradually. This finding immediately suggests that the source of the spring behavior of this protein is to be found in the interfaces between repeat units, which are mostly hydrophobic in nature, and can be loosened by the denaturant. This explanation is commensurate with molecular dynamics simulations conducted by others on similar proteins.
Multiple questions are raised by this study such as: What are the timescales of the dynamics related to the spring behavior of these repeat proteins? What is
the functional role of this behavior? Single-molecule techniques, in combination with computer simulations, will no doubt answer some of these questions in the not-so-far future. The beautiful set of experiments documented in this thesis lays the foundation for these future studies.
Rehovot, June 2014 Prof. Gilad Haran
Acknowledgments
I express sincere gratitude to my mentor, Prof. Gilad Haran, for his support and generous assistance throughout my research program. I also thank my fellow lab members, Menahem Pirchi, Inbal Riven, Haim Aviram, Mila Goomanovsky, Nir Zohar, Yunmin Jung, and Rita August for their help. I thank Ellen Merlo for financial support. I am indebted to the Weizmann Institute of Science for giving me the opportunity and for providing me a superb environment to research my field of interest. For their never ending support and dedication, I thank my dear parents, Mehrdad Sedghani and Diana Soleimani. Last but not least, I am grateful to my husband Shlomo Cohen for love, patience, understanding, and encouragement.
Abbreviations
ADP Avalanche Photo Diodes
ANK Ankyrin
CD Circular Dichroism
CTPR Consensus TPR
CTPR3 CTPR with three repeats
DIC Dichroic mirror
FCS Fluorescent correlation spectroscopy
FRET Förster Resonance Energy Transfer
GuHCl Guanidine hydrochloride
Heat Huntington, Elongation factor 3, protein phosphatase 2A, and Yeast Kinase TOR1
HSQC Heteronuclear Single Quantum Coherence
HX Hydrogen Exchange
MS Motorized stage
NMR Nuclear magnetic resonance
PDB Protein Data Bank
PH Pinhole
PMT Photomultiplier tube
SM-FRET Single-Molecule FRET
TPR Tetratrico Peptide Repeat
Chapter 1 Introduction
Repeat proteins account for more than 5 % of the proteins in multi-cellular organisms. In contrast to globular proteins that have extensive long-range contacts, the structures of repeat proteins show mainly intra- and inter-repeat local contacts. These simple topologies facilitate modeling of the folding mechanism. Tetratricopeptide repeat (TPR) is a 34-amino acid helix-turn-helix motif found in tandem arrays in many natural proteins [1].
The design and characterization of a series of consensus TPR (CTPR) proteins, built as arrays of multiple tandem copies of a consensus sequence, enable further simplification of the folding model. In a series of experiments we used Single Molecule Förster Resonance Energy Transfer (SM-FRET) to study the folding of a CTPR protein with three repeats.
In our experiment, single molecules of double-labeled CTPR were allowed to diffuse through the focus of a confocal microscope. These measurements, which were carried out for two FRET pairs, map the folding behavior of two and three repeats. Measured photon bursts were used to obtain FRET efficiency histograms in different chemical denaturant concentrations. For both FRET pairs, a continuous shift of FRET efficiency histograms is found at mild denaturant concentrations, followed by an all-or-none transition to the unfolded state. Global fitting of histograms showed that the continuous shift is not a result of an interchange between two stable populations with similar FRET efficiencies. Statistical analysis of single-molecule trajectories obtained from measurements of immobilized molecules prove the absence of a slow transition between possible FRET populations. Correlation analysis was used to observe the dynamics of FRET efficiency. These measurements indicate that the conformational dynamics is faster than a microsecond timescale. We conclude that the initial continuous shift of FRET efficiency is caused by a continuous expansion of the structure.
Combining the result of nuclear magnetic resonance (NMR) and SM-FRET measurements we determine that during global expansion the secondary structure is preserved and the tertiary structure slightly expanded. This expansion is followed by a barrier crossing event which involves unfolding of the secondary structure. Modification of a previously proposed Ising model for the folding of CTPR proteins is suggested.
S. Cohen, Single-Molecule Fluorescence Spectroscopy of the Folding of a Repeat Protein, Springer Theses, DOI 10.1007/978-3-319-09558-5_1
Our model is in agreement with previous studies of the flexibility of tertiary structure in repeat proteins, suggesting that this flexibility plays a role in binding to other proteins or ligands [2].
1.1 Repeat Proteins
Repeat proteins, whether as independent folds or as segments within larger proteins, are composed of tandem arrays of a small structural unit. The non-globular folds of repeat proteins are formed from stacks of repeated motifs of 20–40 amino acids, often producing extended structures. Repeat proteins are widely distributed in various biological species [3], and are involved in a myriad of essential biological processes. They account for more than 5 % of the proteins in the multi-cellular organism category of the Swiss-Prot database. Ankyrin (ANK) repeats, tetratricopeptide (TPR), leucine-rich repeats, Heat (Huntington, Elongation factor 3, protein phosphatase 2A, and Yeast Kinase TOR1), beta helical repeats, and armadillo repeats are common examples of repeat proteins.
In contrast to globular proteins with complicated and irregular tertiary structures, the tertiary structures of repeat proteins are simple and modular. The degree of the simplicity of this topology is usually determined by the contact order, which is the average sequence distance between all pairs of contacting residues normalized by the total sequence length. Typical contact order values for repeat proteins range from 0.05 to 0.07 (equal to sequence separations of about 6–7 residues), whereas the contact order values of globular proteins range from 0.1 to 0.2 (equal to sequence separations of about 10–30 residues) [4]. Owing to the modularity and simplicity of their structure, repeat proteins have received increasing attention by scientists in various fields such as protein engineering, protein design, protein folding, and molecular dynamics simulations.
The non-globular, elongated shapes of repeat proteins result in larger surfaces for molecular recognition and interactions. Recognition of different binding proteins, based on their sequence and structure, is the most common function of repeat proteins.
In repeat proteins, amino acid residues that are important for binding are distinct and different from those that are important for maintaining the stability of the fold; therefore, consensus designed proteins with fixed framework positions and variable binding positions can be designed to construct novel binding molecules. Additionally, the linearity in the structure of repeat proteins suggests that repeat protein modules, based on the consensus sequence, may be added, allowing extended structures without altering the overall topology [5]. Design of repeats in the laboratory, based on the consensus sequences of individual repeats [6], is therefore possible. Consensus TPR (CTPR) [7], ANK [8, 9], leucine-rich [10], beta-propeller [11], and armadillo repeats [12] with the same structure as the natural repeat have been successfully designed.
TPRs [1, 13] are one of the common structural motifs, and are found in tandem arrays of 2–16 motifs of 34 amino acid residues. The three-dimensional structure
of a single TPR is a helix–turn–helix. Adjacent TPR units are stacked in parallel and form a right-handed superhelix. More than 5,000 TPR-containing proteins have been identified in different organisms. For more than 100 of them, their structures have been solved and deposited in the Protein Data Bank (PDB). Similar to other repeat proteins, the large surface area presented by their super helix elongated structures [1] makes them particularly well suited to mediate interactions with other proteins [14–16].
TPRs participate in a diverse set of processes: they are involved in peroxisomal targeting [17], in co-chaperones interacting with heat shock proteins 70 and 90 [18], and in synaptic vesicle fusion [19]. In addition, TPRs interact with transcriptional repressor proteins [20] and are associated with bacterial pathogenesis [21, 22], protein sorting, and translocation to the outer membrane of chloroplasts and mitochondria [23, 24], and more. They are also found in magnetosome-associated proteins in magnetotactic bacteria [25] and in the outer membrane assembly of alphaproteobacteria [26]. Mutations in TPR-containing proteins have been associated with a variety of human diseases [13].
The engineered structure of CTPRs [27, 28], with high stability and desired binding specificity, make them unique and useful for diverse applications. Recent studies have used CTPRs for generating anti-cancer agents [29] and functional biofilms [30].
1.2 Folding Stability of Repeat Proteins
The effect of insertion of repeat modules on the stability and folding kinetics for two series of repeat proteins, namely, the CTPR and ANK series [31, 32], was previously studied. In both series the insertion of repeat modules stabilizes the protein and reduces the unfolding rate.
In general, individual repeats are intrinsically unstable; the addition of a repeat with favorable interface interactions between repeats stabilizes the fold. Since there is no long-range interaction in repeat proteins, assuming that nonadjacent repeats are independent of one another, to a good approximation, the stabilities of a construct can be described as the sum of energy terms associated with each repeat [5]. One-dimensional Ising models have been used with great success to explain the unfolding behavior of repeat proteins at equilibrium.
The Ising model is a statistical model, conceived in 1920, to describe the magnetization of a series of spins with only nearest neighbor interactions between them. In this model each spin is an independent unit that can be either up or down. The free energy of the system is a linear summation of energy associated with each unit [33]. The one-dimensional Ising model has been used for more than half a century to describe the helix-coil transition in the formation of α helices. In this model each residue is considered to be ordered in a helical state, or be in a coil state [34]. Local contact between adjacent residues results in cooperative folding behavior.
structures fluctuate until they collide and pack tightly together to form the native structure; and (2) the nucleation-condensation model, in which the secondary and tertiary structures of the protein are formed at the same time [43]. Recent studies in the field show that there are many proteins that exhibit the characteristics of both the diffusion-collision and nucleation-condensation models [44]. The Ising model is one model that describes such a behavior. According to this model, folding can be initiated from any of the helices, a la the diffusion collision model, but propagation of folding occurs simultaneously for secondary and tertiary structures, as in the nucleation condensation model.
Modern theories and recent experimental results propose that the folding process can be explained by a funnel-shaped energy landscape [45, 46]. According to this view, which is opposed to a historical view, various parallel pathways are involved in the folding process. These pathways encompass numerous structures, although all lead to the final state, which is the most stable thermodynamic state under native conditions. At any stage during its folding, the protein exists as an ensemble of conformations and can be trapped transiently in many local energy minima. Detailed structural characterization of those conformations that are sampled by the protein during folding is not yet technically possible. However, new single-molecule techniques, combined with statistical analysis [47–50], enable us to obtain some insight about the folding pathways. Owing to the simplicity of the fold in repeat proteins, structural characterization of intermediate states and folding pathways is feasible.
Studying the thermodynamics of folding, in particular, is a powerful method to get insight into the mechanism involved in the process. This method allows us to map the local minima on the energy landscape and calculate the propensity for the protein to be trapped in these minima. The rate of transition between any two local minima and the order of transitions during the process can also be obtained.
The thermodynamic behavior of small proteins is typically characterized by a two-state model [51–55]. A one-dimensional free energy surface is used to explain the folding of such proteins, and the crossing of the energy barrier between folded and unfolded conformations is the rate-limiting step [56]. According to this model, proteins rarely exist in the transition state region, and the transition over the barrier is an all-or-none event.
During the folding of certain proteins, stable intermediate states have been reported to exist under mild denaturation conditions [57–59]. Circular dichroism and NMR measurements indicate that considerable amounts of secondary structure exist in such intermediate states. However, as indicated by near-UV circular dichroism, as well as by the NMR and fluorescent spectroscopic behavior of aromatic residues [60–62], significant mobility seems to exist in the hydrophobic core, which is not present in the native state of these proteins. This kind of intermediate state was termed the molten globule, which was defined as a structure with loosened tertiary structure that conserves its native secondary structure.
Single-molecule techniques provide new insights into the protein folding field. Using these techniques, partially unfolded stable states have been found in large proteins [48, 63, 64]. In these reports the diversity of the folding pathways
of proteins was observed experimentally. In a few other proteins the continuous changes in an order parameter during folding were observed, suggesting that even for small proteins, folding may be more complicated than was previously thought [65–69]. Clear structural characterization of such processes, however, remains a challenge.
1.4 Single-Molecule Fluorescence Spectroscopy
Single-molecule spectroscopy provides a fundamental advantage in resolving and quantifying the properties of individual molecules or subpopulations by direct examination of the molecular processes. This information is usually inaccessible in classical ensemble experiments due to the averaging. Generally, two different single-molecule methods are used to investigate biomolecules at a single-molecule level [49]. The first method is force spectroscopy, where either the atomic force microscope [70, 71] or the optical trap [72] is used to obtain a detailed structural understanding of molecules under mechanical force. The second method is fluorescence spectroscopy [50, 73–76], which in combination with Förster Resonance Energy Transfer (FRET), is widely used as an accurate technique for distance measurements, and which was termed “a spectroscopic ruler”. FRET was discovered in 1948 by Forster [77], who demonstrated that dipole-dipole interactions between donor to acceptor fluorophores can lead to efficient energy transfer between them. This energy transfer strongly depends on the distance between the two probes. Single-Molecule FRET (SM-FRET) has been successfully and extensively used to investigate a variety of processes such as protein folding [50, 63, 78] and misfolding [79], the conformational dynamics of proteins [80], protein-protein interactions [81, 82], intrinsically unstructured proteins [83–85], protein function in vitro [86] and in vivo [87]. It has also been used to study structures involving DNA [88] and RNA [89].
In particular, in the field of protein folding, the modern view proposing the existence of a diversity of folding pathways has attracted much attention to single-molecule spectroscopy techniques. Single-molecule fluorescence spectroscopy was used to obtain detailed information about the folding pathways of single-molecule proteins in equilibrium for fast [64] and for slow [63] folding processes. This was achieved by visualizing the change in FRET efficiency as a function of time at the single-molecule level in different denaturant concentrations and by using statistical analysis to analyze them. SM-FRET was also used to study the protein folding transition path time [90]. In order to investigate the behavior of single molecules under conditions far from equilibrium, time-resolved measurements of FRET efficiency were carried out by abrupt change of the denaturant concentration in micro-fluidic devices [91–93]. SM-FRET was also used to obtain information about unfolded states. Expansion of the unfolded state by increasing the denaturant concentration was observed by single-molecule FRET equilibrium and non-equilibrium measurements [94, 95]. FRET, in combination with fluorescent
correlation spectroscopy, was used to quantify the internal conformational friction of the unfolded states and the intrinsic denatured states [96].
Single-molecule measurements show that the protein folding process for some proteins is very complicated. Diversity in the folding of different proteins was experimentally observed [63, 66, 69].
1.5 The Main Findings of This Work
A CTPR protein with three repeats (CTPR3) was labeled at different positions with two pairs of fluorophores suitable for FRET. Single-molecule measurements were carried out on freely diffusing molecules. Histograms of FRET efficiency were measured in different denaturant concentrations. These histograms revealed two populations: one at a low FRET value, and another at a high FRET value. The population with a low FRET value was found to represent the unfolded state. The population with a high FRET value was found to represent the folded state under denaturant-free conditions. This population was observed to shift continuously to lower FRET values with increasing denaturant concentration.
We demonstrated that continuous loosening of the tertiary structure is responsible for this shift and that crossing the barrier to the unfolded state occurs when the secondary structure melts. This folding model is in opposition to the nucleation condensation model, which proposes coincident propagation of folding of secondary and tertiary structures. The loosening of the tertiary structure is believed to be important in TPR functionality because of its direct relationship to its affinity for binding to other proteins.
References
1. Blatch GL, Lassle M (1999) The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays 21(11):932–939
2. Forwood JK et al (2010) Quantitative structural analysis of importin-beta flexibility: paradigm for solenoid protein structures. Structure 18(9):1171–1183
3. Andrade MA, Perez-Iratxeta C, Ponting CP (2001) Protein repeats: structures, functions, and evolution. J Struct Biol 134(2–3):117–131
4. Javadi Y, Itzhaki LS (2013) Tandem-repeat proteins: regularity plus modularity equals design-ability. Curr Opin Struct Biol 23(4):622–631
5. Mello CC, Barrick D (2004) An experimentally determined protein folding energy landscape. Proc Natl Acad Sci USA 101(39):14102–14107
6. Forrer P et al (2004) Consensus design of repeat proteins. ChemBioChem 5(2):183–189
7. Main ER et al (2003) Design of stable alpha-helical arrays from an idealized TPR motif. Structure 11(5):497–508
8. Mosavi LK, Minor DL Jr, Peng ZY (2002) Consensus-derived structural determinants of the ankyrin repeat motif. Proc Natl Acad Sci USA 99(25):16029–16034
9. Binz HK et al (2003) Designing repeat proteins: well-expressed, soluble and stable proteins from combinatorial libraries of consensus ankyrin repeat proteins. J Mol Biol 332(2):489–503
10. Stumpp MT et al (2003) Designing repeat proteins: modular leucine-rich repeat protein libraries based on the mammalian ribonuclease inhibitor family. J Mol Biol 332(2):471–487
11. Nikkhah M et al (2006) Engineering of beta-propeller protein scaffolds by multiple gene duplication and fusion of an idealized WD repeat. Biomol Eng 23(4):185–194
12. Parmeggiani F et al (2008) Designed armadillo repeat proteins as general peptide-binding scaffolds: Consensus design and computational optimization of the hydrophobic core. J Mol Biol 376(5):1282–1304
13. D’Andrea LD, Regan L (2003) TPR proteins: the versatile helix. Trends Biochem Sci 28(12):655–662
14. Tang KS et al (1999) Stability and folding of the tumour suppressor protein p16. J Mol Biol 285(4):1869–1886
15. Das AK, Cohen PW, Barford D (1998) The structure of the tetratricopeptide repeats of protein phosphatase 5: implications for TPR-mediated protein-protein interactions. EMBO J 17(5):1192–1199
16. Zeytuni N, Zarivach R (2012) Structural and functional discussion of the tetra-trico-peptide repeat, a protein interaction module. Structure 20(3):397–405
17. Fransen M et al (2008) Comparison of the PTS1- and Rab8b-binding properties of Pex5p and Pex5Rp/TRIP8b. Biochim Et Biophys Acta-Mol Cell Res 1783(5):864–873
18. Ramsey AJ, Russell LC, Chinkers M (2009) C-terminal sequences of hsp70 and hsp90 as non-specific anchors for tetratricopeptide repeat (TPR) proteins. Biochem J 423:411–419
19. Young JC, Barral JM, Hartl FU (2003) More than folding: localized functions of cytosolic chaperones. Trends Biochem Sci 28(10):541–547
20. Smith RL, Redd MJ, Johnson AD (1995) The tetratricopeptide repeats of ssn6 interact with the homeo domain of alpha-2. Genes Dev 9(23):2903–2910
21. Edqvist PJ et al (2006) Tetratricopeptide repeats in the type III secretion chaperone, LcrH: their role in substrate binding and secretion. Mol Microbiol 59(1):31–44
22. Tiwari D et al (2009) Key residues in mycobacterium tuberculosis protein kinase G play a role in regulating kinase activity and survival in the host. J Biol Chem 284(40):27467–27479
23. Mirus O et al (2009) Evolutionarily evolved discriminators in the 3-TPR domain of the Toc64 family involved in protein translocation at the outer membrane of chloroplasts and mitochondria. J Mol Model 15(8):971–982
24. Baker MJ et al (2007) Mitochondrial protein-import machinery: correlating structure with function. Trends Cell Biol 17(9):456–464
25. Zeytuni N et al (2011) Self-recognition mechanism of MamA, a magnetosome-associated TPR-containing protein, promotes complex assembly. Proc Natl Acad Sci USA 108(33):E480–E487
26. Gatsos X et al (2008) Protein secretion and outer membrane assembly in Alphaproteobacteria. FEMS Microbiol Rev 32(6):995–1009
27. Cortajarena AL et al (2010) Designed proteins to modulate cellular networks. ACS Chem Biol 5(6):545–552
28. Cortajarena AL et al (2004) Protein design to understand peptide ligand recognition by tetratricopeptide repeat proteins. Protein Eng Des Sel 17(4):399–409
29. Cortajarena AL, Yi F, Regan L (2008) Designed TPR modules as novel anticancer agents. ACS Chem Biol 3(3):161–166
30. Grove TZ, Regan L, Cortajarena AL (2013) Nanostructured functional films from engineered repeat proteins. J Roy Soc Interface 10(83):20130051
31. Wetzel SK et al (2008) Folding and unfolding mechanism of highly stable full-consensus ankyrin repeat proteins. J Mol Biol 376(1):241–257
32. Javadi Y, Main ER (2009) Exploring the folding energy landscape of a series of designed consensus tetratricopeptide repeat proteins. Proc Natl Acad Sci USA 106(41):17383–17388
References
33. Gallavotti G (1999) Statistical mechanics : a short treatise. Texts and monographs in physics, Springer, Berlin, xiv, 339 p
34. Scott RA 3rd (1967) Statistical mechanical studies of polypeptides. 1. Theory of the helixcoil transition including right- and left-handed helical states. Biopolymers 5(10):931–951
35. Kajander T et al (2005) A new folding paradigm for repeat proteins. J Am Chem Soc 127(29):10188–10190
36. Cortajarena AL, Mochrie SGJ, Regan L (2011) Modulating repeat protein stability: the effect of individual helix stability on the collective behavior of the ensemble. Protein Sci 20(6):1042–1047
37. Phillips JJ et al (2012) Modulation of the multistate folding of designed TPR proteins through intrinsic and extrinsic factors. Protein Sci 21(3):327–338
38. Bradley CM, Barrick D (2002) Limits of cooperativity in a structurally modular protein: response of the Notch ankyrin domain to analogous alanine substitutions in each repeat. J Mol Biol 324(2):373–386
39. Zweifel ME, Barrick D (2001) Studies of the ankyrin repeats of the Drosophila melanogaster Notch receptor. 1. Solution conformational and hydrodynamic properties. Biochemistry 40(48):14344–14356
40. Ferreiro DU et al (2008) The energy landscapes of repeat-containing proteins: topology, cooperativity, and the folding funnels of one-dimensional architectures. PLoS Comput Biol 4(5):e1000070
41. Hagai T et al (2012) Modulation of folding kinetics of repeat proteins: interplay between intra- and interdomain interactions. Biophys J 103(7):1555–1565
42. Serquera D et al (2010) Mechanical unfolding of an ankyrin repeat protein. Biophys J 98(7):1294–1301
43. Karplus M, Weaver DL (1976) Protein-folding dynamics. Nature 260(5550):404–406
44. Travaglini-Allocatelli C et al (2009) Folding and stability of globular proteins and implications for function. Curr Opin Struct Biol 19(1):3–7
45. Sali A, Shakhnovich E, Karplus M (1994) How does a protein fold? Nature 369(6477):248–251
46. Karplus M (1997) The Levinthal paradox: yesterday and today. Fold Des 2(4):S69–S75
47. Schuler B (2005) Single-molecule fluorescence spectroscopy of protein folding. ChemPhysChem 6(7):1206–1220
48. Stigler J et al (2011) The complex folding network of single calmodulin molecules. Science 334(6055):512–516
49. Borgia A, Williams PM, Clarke J (2008) Single-molecule studies of protein folding. Annu Rev Biochem 77:101–125
50. Haran G (2003) Single-molecule fluorescence spectroscopy of biomolecular folding. J Phys Condens Matter 15:R1291–R1317
51. Privalov PL (1979) Stability of proteins: small globular proteins. Adv Protein Chem 33:167–241
52. Weikl TR, Palassini M, Dill KA (2004) Cooperativity in two-state protein folding kinetics. Protein Sci 13(3):822–829
53. Segawa SI, Sugihara M (1984) Characterization of the transition-state of Lysozyme unfolding. 1. effect of protein solvent interactions on the transition-state. Biopolymers 23(11):2473–2488
54. Jackson SE, Fersht AR (1991) Folding of chymotrypsin inhibitor 2. 1. Evidence for a twostate transition. Biochemistry 30(43):10428–10435
55. Baker D (2000) A surprising simplicity to protein folding. Nature 405(6782):39–42
56. Kubelka J, Hofrichter J, Eaton WA (2004) The protein folding ‘speed limit’. Curr Opin Struct Biol 14(1):76–88
57. Ptitsyn OB et al (1990) Evidence for a molten globule state as a general intermediate in protein folding. FEBS Lett 262(1):20–24
58. Christensen H, Pain RH (1991) Molten globule intermediates and protein folding. Eur Biophys J 19(5):221–229
59. Ohgushi M, Wada A (1983) Molten-globule state—a compact form of globular-proteins with mobile side-chains. FEBS Lett 164(1):21–24
60. Carrey EA, Pain RH (1978) Conformation of a stable intermediate on the folding pathway of Staphylococcus aureus penicillinase. Biochim Biophys Acta 533(1):12–22
61. Baum J et al (1989) Characterization of a partly folded protein by NMR methods: studies on the molten globule state of guinea pig alpha-lactalbumin. Biochemistry 28(1):7–13
62. Roder H, Elove GA, Englander SW (1988) Structural characterization of folding intermediates in cytochrome c by H-exchange labelling and proton NMR. Nature 335(6192):700–704
63. Pirchi M et al (2011) Single-molecule fluorescence spectroscopy maps the folding landscape of a large protein. Nat Commun 2:493
64. Chung HS et al (2011) Extracting rate coefficients from single-molecule photon trajectories and fret efficiency histograms for a fast-folding protein. J Phys Chem A 115(16):3642–3656
65. Knott M, Chan HS (2006) Criteria for downhill protein folding: calorimetry, chevron plot, kinetic relaxation, and single-molecule radius of gyration in chain models with subdued degrees of cooperativity. Proteins 65(2):373–391
66. Liu J et al (2012) Exploring one-state downhill protein folding in single molecules. Proc Natl Acad Sci USA 109(1):179–184
67. Favrin G et al (2003) Two-state folding over a weak free-energy barrier. Biophys J 85(3):1457–1465
68. Naganathan AN, Orozco M (2011) The native ensemble and folding of a protein moltenglobule: functional consequence of downhill folding. J Am Chem Soc 133(31):12154–12161
69. Lindhoud S (2012) Visualization and charachterisation of apoflavodoxin folding, wageningen university. p 140
70. Engel A, Gaub HE, Muller DJ (1999) Atomic force microscopy: a forceful way with single molecules. Curr Biol 9(4):R133–R136
71. Rief M et al (1997) Reversible unfolding of individual titin immunoglobulin domains by AFM. Science 276(5315):1109–1112
72. Kellermayer MS et al (1997) Folding-unfolding transitions in single titin molecules characterized with laser tweezers. Science 276(5315):1112–1116
73. Moerner WE (2002) A dozen years of single-molecule spectroscopy in physics, chemistry, and biophysics. J Phys Chem B 106(5):910–927
74. Haustein E, Schwille P (2004) Single-molecule spectroscopic methods. Curr Opin Struct Biol 14(5):531–540
75. Michalet X et al (2003) The power and prospects of fluorescence microscopies and spectroscopies. Annu Rev Biophys Biomol Struct 32:161–182
76. Roy R, Hohng S, Ha T (2008) A practical guide to single-molecule FRET. Nat Methods 5(6):507–516
77. Forster T (1948) Zwischenmolecular engergiewanderung and floureszenz. Ann Phys Ny 2:55
78. Schuler B, Lipman EA, Eaton WA (2002) Probing the free-energy surface for protein folding with single-molecule fluorescence spectroscopy. Nature 419(6908):743–747
79. Borgia MB et al (2011) Single-molecule fluorescence reveals sequence-specific misfolding in multidomain proteins. Nature 474(7353):662–665
80. Hanson JA et al (2007) Illuminating the mechanistic roles of enzyme conformational dynamics. Proc Natl Acad Sci USA 104(46):18055–18060
81. Bae W, Choi M-G, Hyeon C, Shin Y-K, Yoon T-Y (2013) Real-time observation of multipleprotein complex formation with single-molecule FRET. J Am Chem Soc 135:10254–10257
82. Truong K, Ikura M (2001) The use of FRET imaging microscopy to detect proteinprotein interactions and protein conformational changes in vivo. Curr Opin Struct Biol 11(5):573–578
83. Ohashi T et al (2007) An experimental study of GFP-based FRET, with application to intrinsically unstructured proteins. Protein Sci 16(7):1429–1438
84. Schuler B et al (2012) Application of confocal single-molecule FRET to intrinsically disordered proteins. Methods Mol Biol 896:21–45
85. Nath A et al (2012) The conformational ensembles of alpha-synuclein and tau: combining single-molecule FRET and simulations. Biophys J 103(9):1940–1949
90. Chung HS et al (2012) Single-molecule fluorescence experiments determine protein folding transition path times. Science 335(6071):981–984
91. Lipman EA et al (2003) Single-molecule measurement of protein folding kinetics. Science 301(5637):1233–1235
92. Wunderlich B et al (2013) Microfluidic mixer designed for performing single-molecule kinetics with confocal detection on timescales from milliseconds to minutes. Nat Protoc 8(8):1459–1474
93. Kinoshita M et al (2007) Development of a technique for the investigation of folding dynamics of single proteins for extended time periods. Proc Natl Acad Sci USA 104(25):10453–10458
94. Schuler B, Eaton WA (2008) Protein folding studied by single-molecule FRET. Curr Opin Struct Biol 18(1):16–26
95. Sherman E, Haran G (2006) Coil-globule transition in the denatured state of a small protein. Proc Natl Acad Sci USA 103(31):11539–11543
96. Soranno A et al (2012) Quantifying internal friction in unfolded and intrinsically disordered proteins with single-molecule spectroscopy. Proc Natl Acad Sci USA 109(44):17800–17806
Chapter 2 Methods
2.1 Protein Purification and Labeling
This protein purification and labeling was done in collaboration with Luca Domenico D’Andrea, Istituto di Biostrutture e Bioimmagini, CNR, Napoli, Italy. The protein cloning, expression and purification was carried out according to the protocol published previously [1].
The amino acid sequence of CTPR3 is as follows:
. The crystal structure of CTPR3 with the same color coding is shown in Fig. 2.1
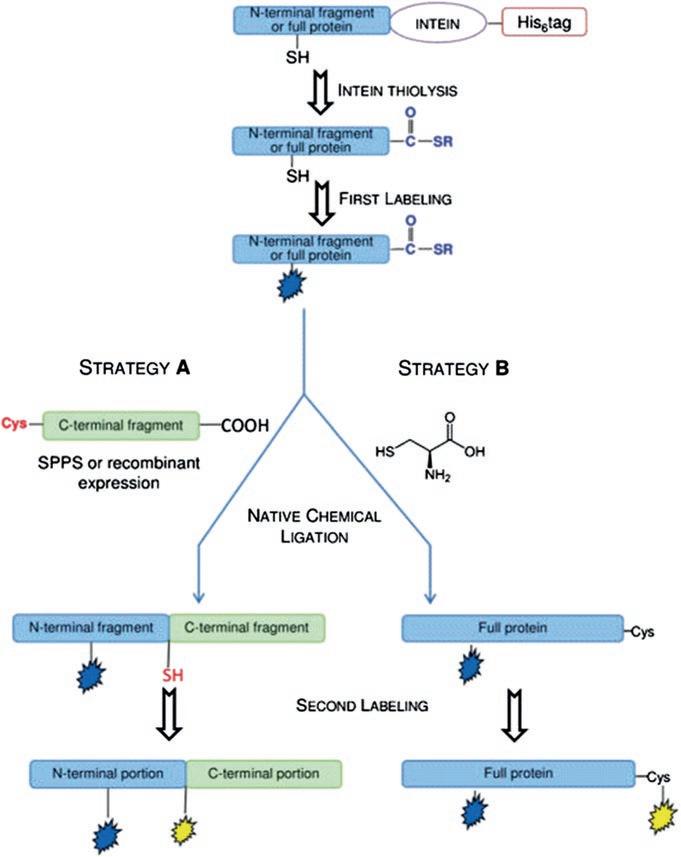
A novel strategy was developed for site specific labeling of CTPR3 [2]. This method is based on the use of the expressed protein ligation. Schematic representation of this process is shown in Fig. 2.2. This technique requires the splitting of the protein of interest into two fragments, each bearing a single cysteine. The N-terminal fragment is expressed in a bacterial host as a fusion protein with the Mycobacterium xenopi gyrase A intein (Mxe GyrA intein). The expressed chimeric protein is also endowed with a C-terminal His6-tag, allowing its purification by affinity chromatography. After purification, the fusion protein is incubated with a thiol, which induces the splicing of Mxe GyrA intein and the release of the protein fragment as a C-terminal thioester. The thioester protein is then isolated from the intein and selectively labeled on its single Cys residue upon reaction with a probe containing a maleimide group as sulfhydryl-reactive moiety. The C-terminal fragment, carrying an N-terminal Cys residue, can be chemically synthesized or expressed in bacteria depending on the fragment size. The native chemical ligation reaction between the mono-labeled thioester fragment and the C-terminal fragment affords the full-length mono-labeled protein.
The Cys residue involved in NCL reaction is then exploited to introduce the second probe into the protein (Fig. 2.2, strategy A). A simpler approach is used when the second probe is located at the C-terminus. In this case, the full-length protein containing a single Cys at the required position is expressed as an intein fusion protein. After purification and first labeling, the full-length thioester protein is reacted with L-Cys in a NCL reaction. The newly introduced Cys residue is then labeled with the second probe (Fig. 2.2, strategy B). A final re-folding step is required before their utilization.
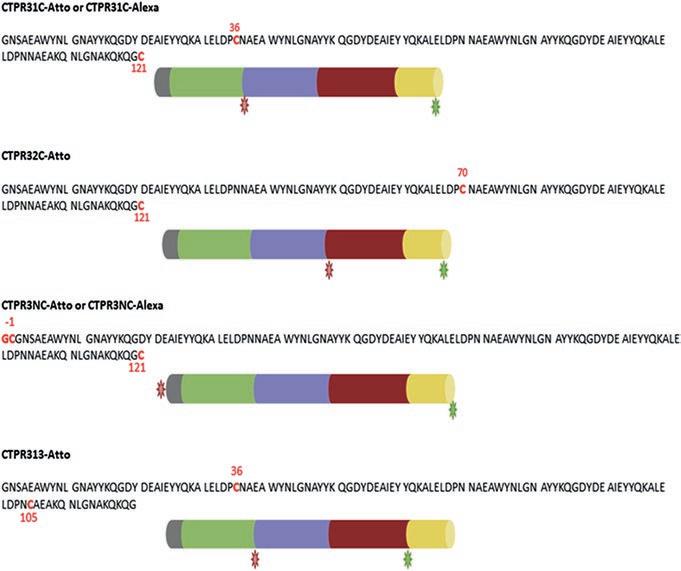
The details of the labeling process was reported for CTPR with Atto dyes [2]. This method was also used for the labeling of CTPR3 with Alexa-maleimide dyes (Life technologies Invitrogen). Amino acid sequences of the CTPR3 variants and a schematic representation of double labeled CTPR are shown in Fig. 2.3.
CTPR31C Atto is the CTPR3 double labeled with Atto 488 on site 121, and Atto 647 N on site 36.
CTPR32C Atto is the CTPR3 double labeled with Atto 488 on site 121, and Atto 647 N on site 70.
CTPR3NC Atto is the CTPR3 double labeled with Atto 488 on site 121, and Atto 647 N on site 1.
CTPR313 Atto is the CTPR3 double labeled with Atto 488 on site 105, and Atto 647 N on site 12.
CTPR31C Alexa is the CTPR3 double labeled with Alexa 488 on site 121, and Alexa 594 on site 36.
CTPR3NC Alexa is the CTPR3 double labeled with Alexa 488 on site 121, and Alexa 594 on site 1.
Finally, for control measurements, CTPR3 is labeled only with Alexa 488 in position 121(CTPR3SL-Alexa488).
Fig. 2.2 A schematic representation of the semi-synthetic strategy adopted for the preparation of doubly labeled proteins. The N-terminal portion of the protein was prepared as a C-terminal thioester through expression with the Mxe GyrA intein as fusion partner. After splicing, the N-terminal fragment of the protein, carrying a C-terminal thioester group, was labeled with the first probe and then reacted by native chemical ligation with the C-terminal remaining portion of the protein containing an N-terminal Cys residue (strategy A) or a L-Cys (strategy B). Finally, the full protein is labeled with the second probe on the Cys reactive in the native chemical ligation reaction
Fig. 2.3 Amino acid sequences and schematic representation of the CTPR3 variants and double labeled proteins. Red colored amino acids represent insertion and/or mutation. When double labeled with Atto dyes, green stars represent Atto 488 and red stars represent Atto 647 N. When double labeled with Alexa dyes, green stars represent Alexa 488 and red stars represent Alexa 594
All measurements were performed in 50 mM phosphate buffer, 150 mM NaCl, at pH 6.8. Index of refraction of stock solution was determined using a Fisher Abbe refractometer (Fisher Scientific Co.). Concentration of guanidine hydrochloride (GuHCl) was obtained using the equation below where, Δn is the difference between index of refraction of buffer with and without GuHCl.
2.3 Circular Dichroism Spectroscopy
Circular Dichroism (CD) spectroscopy was performed on a AVIV Model 215 CD spectrophotometer (AVIV Instruments, Lakewood, NJ). CD experiments were performed at protein concentration of 6 μM, 25 °C, in 222 nm wavelength.
Ensemble/Bulk Steady-State
2.4 Ensemble/Bulk Steady-State Measurements
Ensemble/bulk fluorescence studies were conducted with a Fluorolog spectrofluorometer (Jobin-Yvon) equipped with Glan-Thompson polarizers. CTPR3 labeled with Alexa 488 and the double-labeled CTPR31C were used for control experiments, including evaluation of fluorescence quantum yield and anisotropy values as a function of GuHCl concentration (see Sect. 3.12). CTPR31C was also used for measuring the ensemble/bulk denaturation curve.
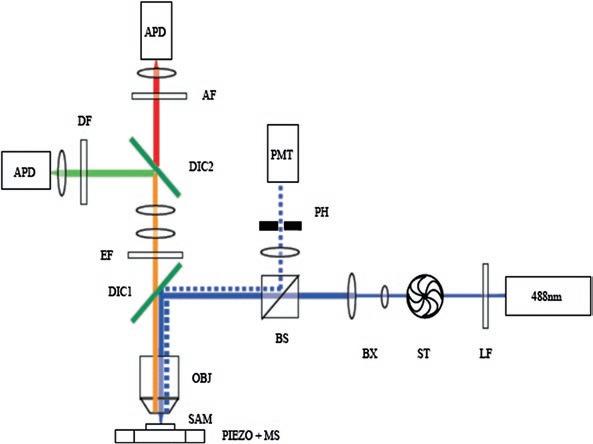
2.5 Single-Molecule Setup and Measurement Methodology
Single-molecule setup
A custom-made single-molecule microscope for automated data collection was constructed, as described in Fig. 2.4 . The beam of an argon ion laser (SpectraPhysics model 163, 488 nm) was passed through a laser-line cleanup filter (LP, Chroma Z488/10x) and expanded five times (BX) to fill the back aperture of a 100x oil-immersion objective (OBJ, Zeiss FLUAR 100x/1.3NA). A sample cell (SAM) was mounted on a custom-designed sample holder on top of a capacitance-feedback 100 × 100 × 20 μ m piezo stage (PIEZO, PI P-517) controlled by a dedicated DSP (PI E-710). The sample was excited by the focused
Fig. 2.4 Schematic representation of the confocal set up
laser beam and the fluorescent emission from the sample was collected by the objective, passed through the first dichroic mirror (DIC1, Chroma 505DCLP), filtered from remaining excitation light (EF, Semrock LP02-488RU), and split into two detection channels by a second dichroic mirror (DIC2, Chroma 565DCLP). The donor dye emission and the acceptor dye emission were filtered by respective emission filters (DF, Semrock FF01-536/40 and AF, Semrock FF01-624/40) and focused onto two single-photon avalanche photodiodes (APD, Perkin-Elmer SPCM-AQR-15). Each APD delivered outputs a TTL pulse output for each incoming photon. The arrival times of each of these photons was recorded by a counter/timer card (National Instruments NI6602) with time resolution of 12.5 ns. A standalone TCSPC module (PicoHarp 300, PicoQuant) was used instead of the counter/timer card in free diffusion experiments. In order to generate an auto-focus mechanism, back-reflected laser light from the sample surface was focused on a pinhole (PH) and measured by a photomultiplier tube (PMT, Hamamatsu H8249). When exactly focused, the PMT position was adjusted to register the maximum photon flux. The piezo stage was mounted on top of a coarse motorized stage (MS) to allow largerscale motion of the sample. When measurements were not in progress, to prevent excess illumination, a mechanical shutter (ST, Uniblitz LS6T2) blocked the laser.
The encapsulation of proteins
Liposomes made of egg phosphatidylcholine and a fraction of 1:500 of 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(cap biotinyl) (both from Avanti Lipids) were prepared by extrusion [3] in the appropriate buffer, using a disposable 0.1 μm syringe connected to Anopore filter (Whatman Anotop-10). To prepare protein-loaded liposomes, we added labeled protein to a final concentration of ~0.2 μM, chosen so that one out of ~10 liposomes would contain a single molecule [4]. Liposomes were separated from non-encapsulated proteins by size exclusion column (GE Healthcare MicroSpin S-400 HR).
Flow cells
Sample cells were made of two glass #1.5 coverslips (Thomas Scientific), glued together with two Teflon strips. After incubation, to prevent evaporation, the cell was sealed with silicon grease.
2.5.1 Measurements on Freely-Diffusing Molecules
Sample preparation
Flow cells filled with a dilute solution of protein molecules (20–45 pM), and 0.01 % Tween to minimize protein adsorption, at different denaturant concentrations were prepared.
Another random document with no related content on Scribd:
tradition the dance used at one time to take place in the churchyard on a Sunday. The horns are said to be those of the reindeer, and from this it may possibly be inferred that they were brought to Abbots Bromley by Scandinavian settlers. The remaining performers represent a hobby-horse, a clown, a woman, and an archer, who makes believe to shoot the horned men[550] .
The motifs of the dances and their chansons must also at first have been determined by the nature of the festivals at which they took place. There were dances, no doubt, at such domestic festivals as weddings and funerals[551] . In Flanders it is still the custom to dance at the funeral of a young girl, and a very charming chanson is used[552] . The development of epic poetry from the cantilenae of the war-festival has been noted in a former chapter. At the agricultural festivals, the primary motif is, of course, the desire for the fertility of the crops and herds. The song becomes, as in the Anglo-Saxon charm, so often referred to, practically a prayer[553] . With this, and with the use of ‘Thread the Needle’ at Châtre ‘to make the hemp grow,’ may be compared the games known to modern children, as to Gargantua, in which the operations of the farmer’s year, and in particular his prayer for his crops, are mimicked in a ronde[554] . Allusions to the process of the seasons, above all to the delight of the renouveau in spring, would naturally also find a place in the festival songs. The words of the famous thirteenth-century lyric were perhaps written to be sung to the twinkling feet of English girls in a round. It has the necessary refrain:
‘Sumer is icumen in, Lhude sing cuccu!
Groweth sed and bloweth med And springth the wdë nu, Sing cuccu!
‘Awë bleteth after lomb, Lhouth after calvë cu. Bulloc sterteth, buckë verteth, Murie sing cuccu!
‘Cuccu, cuccu, wel singës thu, cuccu; Ne swik thu naver nu. Sing cuccu nu. Sing cuccu. Sing cuccu. Sing cuccu nu!’[555]
The savour of the spring is still in the English May songs, the French maierolles or calendes de mai and the Italian calen di maggio. But for the rest they have either become little but mere quête songs, or else, under the influence of the priests, have taken on a Christian colouring[556] . At Oxford the ‘merry ketches’ sung by choristers on the top of Magdalen tower on May morning were replaced in the seventeenth century by the hymn now used[557] . Another very popular Mayers’ song would seem to show that the Puritans, in despair of abolishing the festival, tried to reform it.
‘Remember
us poor Mayers all, And thus we do begin To lead our lives in righteousness, Or else we die in sin.
‘We have been rambling all this night, And almost all this day: And now returned back again, We have brought you a branch of May.
‘A branch of May we have brought you, And at your door it stands; It is but a sprout, but it’s well budded out, By the work of our Lord’s hands,’ &c.[558]
Another religious element, besides prayer, may have entered into the pre-Christian festival songs; and that is myth. A stage in the evolution of drama from the Dionysiac dithyramb was the introduction of mythical narratives about the wanderings and victories of the god, to be chanted or recited by the choragus. The relation of the choragus to the chorus bears a close analogy to that
between the leader of the mediaeval carole and his companions who sang the refrain. This leader probably represents the Keltic or Teutonic priest at the head of his band of worshippers; and one may suspect that in the north and west of Europe, as in Greece, the pauses of the festival dance provided the occasion on which the earliest strata of stories about the gods, the hieratic as distinguished from the literary myths, took shape. If so the development of divine myth was very closely parallel to that of heroic myth[559] .
After religion, the commonest motif of dance and song at the village festivals must have been love. This is quite in keeping with the amorous licence which was one of their characteristics. The goddess of the fertility of earth was also the goddess of the fertility of women. The ecclesiastical prohibitions lay particular stress upon the orationes amatoriae and the cantica turpia et luxuriosa which the women sang at the church doors, and only as love-songs can be interpreted the winileodi forbidden to the inmates of convents by a capitulary of 789[560] . The love-interest continues to be prominent in the folk-song, or the minstrel song still in close relation to folk-song, of mediaeval and modern France. The beautiful wooing chanson of Transformations, which savants have found it difficult to believe not to be a supercherie, is sung by harvesters and by lace-makers at the pillow[561] . That of Marion, an ironic expression of wifely submission, belongs to Shrove Tuesday[562] . These are modern, but the following, from the Chansonnier de St. Germain, may be a genuine mediaeval folk-song of Limousin provenance:
‘A l’entrada dal tems clar, eya, Per joja recomençar, eya, Et per jelos irritar, eya, Vol la regina mostrar Qu’el’ es si amoroza. Alavi’, alavia jelos, Laissaz nos, laissaz nos Ballar entre nos, entre nos[563].’
The ‘queen’ here is, of course, the festival queen or lady of the May, the regina avrillosa of the Latin writers, la reine, la mariée, l’épousée, la trimousette of popular custom[564] . The defiance of the jelos, and the desire of the queen and her maidens to dance alone, recall the conventional freedom of women from restraint in May, the month of their ancient sex-festival, and the month in which the mediaeval wifebeater still ran notable danger of a chevauchée.
The amorous note recurs in those types of minstrel song which are most directly founded upon folk models. Such are the chansons à danser with their refrains, the chansons de mal mariées, in which the ‘jalous’ is often introduced, the aubes and the pastourelles[565] . Common in all of these is the spring setting proper to the chansons of our festivals, and of the ‘queen’ or ‘king’ there is from time to time mention. The leading theme of the pastourelles is the wooing, successful or the reverse, of a shepherdess by a knight. But the shepherdess has generally also a lover of her own degree, and for this pair the names of Robin and Marion seem to have been conventionally appropriated. Robin was perhaps borrowed by the pastourelles from the widely spread refrain
‘Robins m’aime, Robins m’a: Robins m’a demandée: si m’ara[566].’
The borrowing may, of course, have been the other way round, but the close relation of the chanson à danser with its refrain to the dance suggests that this was the earliest type of lyric minstrelsy to be evolved, as well as the closest to the folk-song pattern. The pastourelle forms a link between folk-song and drama, for towards the end of the thirteenth century Adan de la Hale, known as ‘le Bossu,’ a minstrel of Arras, wrote a Jeu de Robin et Marion, which is practically a pastourelle par personnages. The familiar theme is preserved. A knight woos Marion, who is faithful to her Robin. Repulsed, he rides away, but returns and beats Robin. All, however, ends happily with dances and jeux amongst the peasants. Adan de la Hale was one of the train of Count Robert of Artois in Italy. The
play may originally have been written about 1283 for the delectation of the court of Robert’s kinsman, Charles, king of Naples, but the extant version was probably produced about 1290 at Arras, when the poet was already dead. Another hand has prefixed a dramatic prologue, the Jeu du Pèlerin, glorifying Adan, and has also made some interpolations in the text designed to localize the action near Arras. The performers are not likely to have been villagers: they may have been the members of some puy or literary society, which had taken over the celebration of the summer festival. In any case the Jeu de Robin et Marion is the earliest and not the least charming of pastoral comedies[567] .
It is impossible exactly to parallel from the history of English literature this interaction of folk-song and minstrelsy at the French fête du mai. For unfortunately no body of English mediaeval lyric exists. Even ‘Sumer is icumen in’ only owes its preservation to the happy accident which led some priest to fit sacred words to the secular tune; while the few pieces recovered from a Harleian manuscript of the reign of Edward I, beautiful as they are, read like adaptations less of English folk-song, than of French lyric itself[568] . Nevertheless, the village summer festival of England seems to have closely resembled that of France, and to have likewise taken in the long run a dramatic turn. A short sketch of it will not be without interest.
I have quoted at the beginning of this discussion of folk-customs the thirteenth-century condemnations of the Inductio Maii by Bishop Grosseteste of Lincoln and of the ludi de Rege et Regina by Bishop Chanteloup of Worcester. The ludus de Rege et Regina is not indeed necessarily to be identified with the Inductio Maii, for the harvest feast or Inductio Autumni of Bishop Grosseteste had also its ‘king’ and ‘queen,’ and so too had some of the feasts in the winter cycle, notably Twelfth night[569] . It is, however, in the summer feast held usually on the first of May or at Whitsuntide[570] , that these rustic dignitaries are more particularly prominent. Before the middle of the fifteenth century I have not come across many notices of them. That a summer king was familiar in Scotland is implied by the jest of
Robert Bruce’s wife after his coronation at Scone in 1306[571] . In 1412 a ‘somerkyng’ received a reward from the bursar of Winchester College[572] . But from about 1450 onwards they begin to appear frequently in local records. The whole ludus is generally known as a ‘May-play’ or ‘May-game,’ or as a ‘king-play[573],’ ‘king’s revel[574],’ or ‘king-game[575].’ The leading personages are indifferently the ‘king’ and ‘queen,’ or ‘lord’ and ‘lady.’ But sometimes the king is more specifically the ‘somerkyng’ or rex aestivalis. At other times he is the ‘lord of misrule[576],’ or takes a local title, such as that of the ‘Abbot of Marham,’ ‘Mardall,’ ‘Marrall,’ ‘Marram,’ ‘Mayvole’ or ‘Mayvoll’ at Shrewsbury[577] , and the ‘Abbot of Bon-Accord’ at Aberdeen[578] . The use of an ecclesiastical term will be explained in a later chapter[579] . The queen appears to have been sometimes known as a ‘whitepot’ queen[580] . And finally the king and queen receive, in many widely separated places, the names of Robin Hood and Maid Marian, and are accompanied in their revels by Little John, Friar Tuck, and the whole joyous fellowship of Sherwood Forest[581] . This affiliation of the ludus de Rege et Regina to the Robin Hood legend is so curious as to deserve a moment’s examination[582] .
The earliest recorded mention of Robin Hood is in Langland’s Piers Plowman, written about 1377. Here he is coupled with another great popular hero of the north as a subject of current songs:
‘But I can rymes of Robyn hood, and Randolf erle of Chestre[583].’
In the following century his fame as a great outlaw spread far and wide, especially in the north and the midlands[584] . The Scottish chronicler Bower tells us in 1447 that whether for comedy or tragedy no other subject of romance and minstrelsy had such a hold upon the common folk[585] . The first of the extant ballads of the cycle, A Gest of Robyn Hode, was probably printed before 1500, and in composition may be at least a century earlier. A recent investigator of the legend, and a very able one, denies to Robin Hood any
traceable historic origin. He is, says Dr Child, ‘absolutely a creation of the ballad muse.’ However this may be, the version of the Elizabethan playwright Anthony Munday, who made him an earl of Huntingdon and the lover of Matilda the daughter of Lord Fitzwater, may be taken as merely a fabrication. And whether he is historical or not, it is difficult to see how he got, as by the sixteenth century he did get, into the May-game. One theory is that he was there from the beginning, and that he is in fact a mythological figure, whose name but faintly disguises either Woden in the aspect of a vegetation deity[586] , or a minor wood-spirit Hode, who also survives in the Hodeken of German legend[587] . Against this it may be pointed out, firstly that Hood is not an uncommon English name, probably meaning nothing but ‘à-Wood’ or ‘of the wood[588],’ and secondly that we have seen no reason to suppose that the mock king, which is the part assigned to Robin Hood in the May-game, was ever regarded as an incarnation of the fertilization spirit at all. He is the priest of that spirit, slain at its festival, but nothing more. I venture to offer a more plausible explanation. It is noticeable that whereas in the May-game Robin Hood and Maid Marian are inseparable, in the early ballads Maid Marian has no part. She is barely mentioned in one or two of the latest ones[589] . Moreover Marian is not an English but a French name, and we have already seen that Robin and Marion are the typical shepherd and shepherdess of the French pastourelles and of Adan de la Hale’s dramatic jeu founded upon these. I suggest then, that the names were introduced by the minstrels into English and transferred from the French fêtes du mai to the ‘lord’ and ‘lady’ of the corresponding English May-game. Robin Hood grew up independently from heroic cantilenae, but owing to the similarity of name he was identified with the other Robin, and brought Little John, Friar Tuck and the rest with him into the Maygame. On the other hand Maid Marian, who does not properly belong to the heroic legend, was in turn, naturally enough, adopted into the later ballads. This is an hypothesis, but not, I think, an unlikely hypothesis.
Of what, then, did the May-game, as it took shape in the fifteenth and sixteenth centuries, consist? Primarily, no doubt, of a quête or
‘gaderyng.’ In many places this became a parochial, or even a municipal, affair. In 1498 the corporation of Wells possessed moneys ‘provenientes ante hoc tempus de Robynhode[590].’ Elsewhere the churchwardens paid the expenses of the feast and accounted for the receipts in the annual parish budget[591] . There are many entries concerning the May-game in the accounts of Kingston-on-Thames during some half a century. In 1506 it is recorded that ‘Wylm. Kempe’ was ‘kenge’ and ‘Joan Whytebrede’ was ‘quen.’ In 1513 and again in 1536 the game went to Croydon[592] . Similarly the accounts of New Romney note that in 1422 or thereabouts the men of Lydd ‘came with their may and ours[593],’ and those of Reading St. Lawrence that in 1505 came ‘Robyn Hod of Handley and his company’ and in 1507 ‘Robyn Hod and his company from ffynchamsted[594].’ In contemporary Scotland James IV gave a present at midsummer in 1503 ‘to Robin Hude of Perth[595].’ It would hardly have been worth while, however, to carry the May-game from one village or town to another, had it been nothing but a procession with a garland and a ‘gaderyng’; and as a matter of fact we find that in England as in France dramatic performances came to be associated with the summer folk-festivals. The London ‘Maying’ included stage plays[596] . At Shrewsbury lusores under the Abbot of Marham acted interludes ‘for the glee of the town’ at Pentecost[597] . The guild of St. Luke at Norwich performed secular as well as miracle plays, and the guild of Holy Cross at Abingdon held its feast on May 3 with ‘pageants, plays and May-games,’ as early as 1445[598] . Some of these plays were doubtless miracles, but so far as they were secular, the subjects of them were naturally drawn, in the absence of pastourelles, from the ballads of the Robin Hood cycle[599] . Amongst the Paston letters is preserved one written in 1473, in which the writer laments the loss of a servant, whom he has kept ‘thys iij yer to pleye Seynt Jorge and Robyn Hod and the Shryff off Nottyngham[600].’ Moreover, some specimens of the plays themselves are still extant. One of them, unfortunately only a fragment, must be the very play referred to in the letter just quoted, for its subject is ‘Robin Hood and the Sheriff of Nottingham,’ and it is
found on a scrap of paper formerly in the possession of Sir John Fenn, the first editor of the Paston Letters[601] . A second play on ‘Robin Hood and the Friar’ and a fragment of a third on ‘Robin Hood and the Potter’ were printed by Copland in the edition of the Gest of Robyn Hode published by him about 1550[602] . The Robin Hood plays are, of course, subsequent to the development of religious drama which will be discussed in the next volume. They are of the nature of interludes, and were doubtless written, like the plays of Adan de la Hale, by some clerk or minstrel for the delectation of the villagers. They are, therefore, in a less degree folk-drama, than the examples which we shall have to consider in the next chapter. But it is worthy of notice, that even in the hey-day of the stage under Elizabeth and James I, the summer festival continued to supply motives to the dramatists. Anthony Munday’s Downfall and Death of Robert Earl of Huntingdon[603] , Chapman’s May-Day, and Jonson’s delightful fragment The Sad Shepherd form an interesting group of pastoral comedies, affinities to which may be traced in the As You Like It and Winter’s Tale of Shakespeare himself.
As has been said, it is impossible to establish any direct affiliation between the Robin Hood plays and earlier caroles on the same theme, in the way in which this can be done for the jeu of Adan de la Hale, and the Robin and Marion of the pastourelles. The extant Robin Hood ballads are certainly not caroles; they are probably not folk-song at all, but minstrelsy of a somewhat debased type. The only actual trace of such caroles that has been come across is the mention of ‘Robene hude’ as the name of a dance in the Complaynt of Scotland about 1548[604] . Dances, however, of one kind or another, there undoubtedly were at the May-games. The Wells corporation accounts mention puellae tripudiantes in close relation with Robynhode[605] And particularly there was the morris-dance, which was so universally in use on May-day, that it borrowed, almost in permanence, for its leading character the name of Maid Marian. The morris-dance, however, is common to nearly all the village feasts, and its origin and nature will be matter for discussion in the next chapter.
In many places, even during the Middle Ages, and still more afterwards, the summer feast dropped out or degenerated. It became a mere beer-swilling, an ‘ale[606].’ And so we find in the sixteenth century a ‘king-ale[607]’ or a ‘Robin Hood’s ale[608],’ and in modern times a ‘Whitsun-ale[609],’ a ‘lamb-ale[610]’ or a ‘gyst-ale[611]’ beside the ‘church-ales’ and ‘scot-ales’ which the thirteenth-century bishops had already condemned[612] . On the other hand, the village festival found its way to court, and became a sumptuous pageant under the splendour-loving Tudors. For this, indeed, there was Arthurian precedent in the romance of Malory, who records how Guenever was taken by Sir Meliagraunce, when ‘as the queen had mayed and all her knights, all were bedashed with herbs, mosses, and flowers, in the best manner and freshest[613].’ The chronicler Hall tells of the Mayings of Henry VIII in 1510, 1511, and 1515. In the last of these some hundred and thirty persons took part. Henry was entertained by Robin Hood and the rest with shooting-matches and a collation of venison in a bower; and returning was met by a chariot in which rode the Lady May and the Lady Flora, while on the five horses sat the Ladies Humidity, Vert, Vegetave, Pleasaunce and Sweet Odour[614] . Obviously the pastime has here degenerated in another direction. It has become learned, allegorical, and pseudoclassic. At the Reformation the May-game and the May-pole were marks for Puritan onslaught. Latimer, in one of his sermons before Edward VI, complains how, when he had intended to preach in a certain country town on his way to London, he was told that he could not be heard, for ‘it is Robyn hoodes daye. The parishe are gone a brode to gather for Robyn hoode[615].’ Machyn’s Diary mentions the breaking of a May-pole in Fenchurch by the lord mayor of 1552[616] , and the revival of elaborate and heterogeneous May-games throughout London during the brief span of Queen Mary[617] . The Elizabethan Puritans renewed the attack, but though something may have been done by reforming municipalities here and there to put down the festivals[618] , the ecclesiastical authorities could not be induced to go much beyond forbidding them to take place in churchyards[619] . William Stafford, indeed, declared in 1581 that
‘May-games, wakes, and revels’ were ‘now laid down[620],’ but the violent abuse directed against them only two years later by Philip Stubbes, which may be taken as a fair sample of the Puritan polemic as a whole, shows that this was far from being really the case[621]. In Scotland the Parliament ordered, as early as 1555, that no one ‘be chosen Robert Hude, nor Lytill Johne, Abbot of vnressoun, Quenis of Maij, nor vtherwyse, nouther in Burgh nor to landwart in ony tyme to cum[622].’ But the prohibition was not very effective, for in 1577 and 1578 the General Assembly is found petitioning for its renewal[623] . And in England no similar action was taken until 1644 when the Long Parliament decreed the destruction of such May-poles as the municipalities had spared. Naturally this policy was reversed at the Restoration, and a new London pole was erected in the Strand, hard by Somerset House, which endured until 1717[624] .
CHAPTER IX
THE SWORD-DANCE
[Bibliographical Note.—The books mentioned in the bibliographical note to the last chapter should be consulted on the general tendency to μίμησις in festival dance and song. The symbolical dramatic ceremonies of the renouveau are collected by Dr. J. G. Frazer in The Golden Bough. The sword-dance has been the subject of two elaborate studies: K. Müllenhoff, Ueber den Schwerttanz, in Festgaben für Gustav Homeyer (1871), iii, with additions in Zeitschrift für deutsches Alterthum, xviii. 9, xx. 10; and F. A. Mayer, Ein deutsches Schwerttanzspiel aus Ungarn (with full bibliography), in Zeitschrift für Völkerpsychologie (1889), 204, 416. The best accounts of the morris-dance are in F. Douce, Illustrations of Shakespeare (1807, new ed. 1839), and A. Burton, Rushbearing (1891), 95.]
The last two chapters have afforded more than one example of village festival customs ultimately taking shape as drama. But neither the English Robin Hood plays, nor the French Jeu de Robin et Marion, can be regarded as folk-drama in the proper sense of the word. They were written not by the folk themselves, but by trouvères or minstrels for the folk; and at a period when the independent evolution of the religious play had already set a model of dramatic composition. Probably the same is true of the Hox Tuesday play in the form in which we may conjecture it to have been presented before Elizabeth late in the sixteenth century. Nevertheless it is possible to trace, apart from minstrel intervention and apart from imitation of miracles, the existence of certain embryonic dramatic tendencies in the village ceremonies themselves. Too much must not be made of these. Jacob Grimm was inclined to find in them the first
vague beginnings of the whole of modern drama[625] . This is demonstrably wrong. Modern drama arose, by a fairly well defined line of evolution, from a threefold source, the ecclesiastical liturgy, the farce of the mimes, the classical revivals of humanism. Folkdrama contributed but the tiniest rill to the mighty stream. Such as it was, however, a couple of further chapters may be not unprofitably spent in its analysis.
The festival customs include a number of dramatic rites which appear to have been originally symbolical expressions of the facts of seasonal recurrence lying at the root of the festivals themselves. The antithesis of winter and summer, the renouveau of spring, are mimed in three or four distinct fashions. The first and the most important, as well as the most widespread of these, is the mock representation of a death or burial. Dr. Frazer has collected many instances of the ceremony known as the ‘expulsion of Death[626].’ This takes place at various dates in spring and early summer, but most often on the fourth Sunday in Lent, one of the many names of which is consequently Todten-Sonntag. An effigy is made, generally of straw, but in some cases of birch twigs, a beechen bough, or other such material. This is called Death, is treated with marks of fear, hatred or contempt, and is finally carried in procession, and thrust over the boundary of the village. Or it is torn in pieces, buried, burnt, or thrown into a river or pool. Sometimes the health or other welfare of the folk during the year is held to depend on the rite being duly performed. The fragments of Death have fertilizing efficacy for women and cattle; they are put in the fields, the mangers, the hens’ nests. Here and there women alone take part in the ceremony, but more often it is common to the whole village. The expulsion of Death is found in various parts of Teutonic Germany, but especially in districts such as Thuringia, Bohemia, Silesia, where the population is wholly or mainly Slavonic. A similar custom, known both in Slavonic districts and in Italy, France, and Spain, had the name of ‘sawing the old woman.’ At Florence, for instance, the effigy of an old woman was placed on a ladder. At Mid Lent it was sawn through, and the nuts and dried fruits with which it was stuffed scrambled for by the
crowd. At Palermo there was a still more realistic representation with a real old woman, to whose neck a bladder of blood was fitted[627] .
The ‘Death’ of the German and Slavonic form of the custom has clearly come to be regarded as the personification of the forces of evil within the village; and the ceremony of expulsion may be compared with other periodical rites, European and non-European, in which evil spirits are similarly expelled[628] . The effigy may even be regarded in the light of a scapegoat, bearing away the sins of the community[629] . But it is doubtful how far the notion of evil spirits warring against the good spirits which protect man and his crops is a European, or at any rate a primitive European one[630]; and it may perhaps be taken for granted that what was originally thought to be expelled in the rite was not so much either ‘Death’ or ‘Sin’ as winter This view is confirmed by the evidence of an eighth-century homily, which speaks of the expulsion of winter in February as a relic of pagan belief[631] . Moreover, the expulsion of Death is often found in the closest relation to the more widespread custom of bringing summer, in the shape of green tree or bough, into the village. The procession which carries away the dead effigy brings back the summer tree; and the rhymes used treat the two events as connected[632] .
The homily just quoted suggests that the mock funeral or expulsion of winter was no new thing in the eighth century. On the other hand, it can hardly be supposed that customs which imply such abstract ideas as death, or even as summer and winter, belong to the earliest stages of the village festival. What has happened is what happens in other forms of festival play. The instinct of play, in this case finding vent in a dramatic representation of the succession of summer to winter, has taken hold of and adapted to its own purposes elements in the celebrations which, once significant, have gradually come to be mere traditional survivals. Such are the ceremonial burial in the ground, the ceremonial burning, the ceremonial plunging into water, of the representative of the fertilization spirit. In particular, the southern term ‘the old woman’ suggests that the effigy expelled or destroyed is none other than the ‘corn mother’ or ‘harvest-May,’
fashioned to represent the fertilization spirit out of the last sheaf at harvest, and preserved until its place is taken by a new and green representative in the spring.
There are, however, other versions of the mock death in which the central figure of the little drama is not the representative of the fertilization spirit itself, but one of the worshippers. In Bavaria the Whitsuntide Pfingstl is dressed in leaves and water-plants with a cap of peonies. He is soused with water, and then, in mimicry, has his head cut off. Similar customs prevail in the Erzgebirge and elsewhere[633] . We have seen this Pfingstl before. He is the Jack in the green, the worshipper clad in the god under whose protection he desires to put himself[634] . But how can the killing of him symbolize the spring, for obviously it is the coming summer, not the dying winter, that the leaf-clad figure must represent? The fact is that the Bavarian drama is not complete. The full ceremony is found in other parts of Germany. Thus in Saxony and Thuringia a ‘wild man’ covered with leaves and moss is hunted in a wood, caught, and executed. Then comes forward a lad dressed as a doctor, who brings the victim to life again by bleeding[635] . Even so annually the summer dies and has its resurrection. In Swabia, again, on Shrove Tuesday, ‘Dr. Eisenbart’ bleeds a man to death, and afterwards revives him. This same Dr. Eisenbart appears also in the Swabian Whitsuntide execution, although here too the actual resurrection seems to have dropped out of the ceremony[636] . It is interesting to note that the green man of the peasantry, who dies and lives again, reappears as the Green Knight in one of the most famous divisions of Arthurian romance[637] .
The mock death or burial type of folk-drama resolves itself, then, into two varieties. In one, it is winter whose passing is represented, and for this the discarded harvest-May serves as a nucleus. In the other, which is not really complete without a resurrection, it is summer, whose death is mimed merely as a preliminary to its joyful renewal; and this too is built up around a fragment of ancient cult in the person of the leaf-clad worshipper, who is, indeed, none other than the priest-king, once actually, and still in some sort and show,
slain at the festival[638] . In the instances so far dealt with, the original significance of the rite is still fairly traceable. But there are others into which new meanings, due to the influence of Christian custom, have been read. In many parts of Germany customs closely analogous to those of the expulsion of winter or Death take place on Shrove Tuesday, and have suffered metamorphosis into ‘burial of the Carnival[639].’ England affords the ‘Jack o’ Lent’ effigy which is taken to represent Judas Iscariot[640] , the Lincoln ‘funeral of Alleluia[641],’ the Tenby ‘making Christ’s bed[642],’ the Monkton ‘risin’ and buryin’ Peter[643].’ The truth that the vitality of a folk custom is far greater than that of any single interpretation of it is admirably illustrated.
Two other symbolical representations of the phenomena of the renouveau must be very briefly treated. At Briançon in Dauphiné, instead of a death and resurrection, is used a pretty little May-day drama, in which the leaf-clad man falls into sleep upon the ground and is awakened by the kiss of a maiden[644] . Russia has a similar custom; and such a magic kiss, bringing summer with it, lies at the heart of the story of the Sleeping Beauty. Indeed, the marriage of heaven and earth seems to have been a myth very early invented by the Aryan mind to explain the fertility of crops beneath the rain, and it probably received dramatic form in religious ceremonies both in Greece and Italy[645] . Finally, there is a fairly widespread spring custom of holding a dramatic fight between two parties, one clad in green to represent summer, the other in straw or fur to represent winter. Waldron describes this in the Isle of Man[646]; Olaus Magnus in Sweden[647] . Grimm says that it is found in various districts on both sides of the middle Rhine[648] . Perhaps both this dramatic battle and that of the Coventry Hox Tuesday owe their origin to the struggle for the fertilizing head of a sacrificial animal, which also issued in football and similar games. Dr Frazer quotes several instances from all parts of the world in which a mock fight, or an interchange of abuse and raillery taking the place of an actual fight, serves as a crop-charm[649] . The summer and winter battle gave to literature a famous type of neo-Latin and Romance débat[650] . In one of the most interesting forms of this, the eighth-or ninth-century Conflictus
Veris et Hiemis, the subject of dispute is the cuckoo, which spring praises and winter chides, while the shepherds declare that he must be drowned or stolen away, because summer cometh not. The cuckoo is everywhere a characteristic bird of spring, and his coming was probably a primitive signal for the high summer festival[651] .
The symbolical dramas of the seasons stand alone and independent, but it may safely be asserted that drama first arose at the village feasts in close relation to the dance. That dancing, like all the arts, tends to be mimetic is a fact which did not escape the attention of Aristotle[652] . The pantomimes of the decadent Roman stage are a case in point. Greek tragedy itself had grown out of the Dionysiac dithyramb, and travellers describe how readily the dances of the modern savage take shape as primitive dramas of war, hunting, love, religion, labour, or domestic life[653] Doubtless this was the case also with the caroles of the European festivals. The types of chanson most immediately derived from these are full of dialogue, and already on the point of bursting into drama. That they did do this, with the aid of the minstrels, in the Jeu de Robin et de Marion we have seen[654] . A curious passage in the Itinerarium Cambriae of Giraldus Cambrensis ( † 1188) describes a dance of peasants in and about the church of St. Elined, near Brecknock on the Gwyl Awst, in which the ordinary operations of the village life, such as ploughing, sewing, spinning were mimetically represented[655] . Such dances seem to survive in some of the rondes or ‘singing-games,’ so frequently dramatic, of children[656] . On the whole, perhaps, these connect themselves rather with the domestic than with the strictly agricultural element in village cult. A large proportion of them are concerned with marriage. But the domestic and the agricultural cannot be altogether dissociated. The game of ‘Nuts in May,’ for instance, seems to have as its kernel a reminiscence of marriage by capture; but the ‘nuts’ or rather ‘knots’ or ‘posies’ ‘in May’ certainly suggest a setting at a seasonal festival. So too, with ‘Round the Mulberry Bush.’ The mimicry here is of domestic operations, but the ‘bush’ recalls the sacred tree, the natural centre of the seasonal dances. The closest parallels to the dance described by Giraldus Cambrensis are to be found in the
rondes of ‘Oats and Beans and Barley’ and ‘Would you know how doth the Peasant?’, in which the chief, though not always the only, subjects of mimicry are ploughing, sowing and the like, and which frequently contain a prayer or aspiration for the welfare of the crops[657] .
I have treated the mimetic element of budding drama in the agricultural festivals as being primarily a manifestation of the activities of play determined in its direction by the dominant interests of the occasion, and finding its material in the débris of ritual custom left over from forgotten stages of religious thought. It is possible also to hold that the mimesis is more closely interwoven with the religious and practical side of the festivals, and is in fact yet another example of that primitive magical notion of causation by the production of the similar, which is at the root of the rain-and sun-charms. Certainly the village dramas, like the other ceremonies which they accompany, are often regarded as influencing the luck of the farmer’s year; just as the hunting-and war-dances of savages are often regarded not merely as amusement or as practice for actual war and hunting, but as charms to secure success in these pursuits[658] . But it does not seem clear to me that in this case the magical efficacy belongs to the drama from the beginning, and I incline to look upon it as merely part of the sanctity of the feast as a whole, which has attached itself in the course of time even to that side of it which began as play.
The evolution of folk-drama out of folk-dance may be most completely studied through a comparison of the various types of European sword-dance with the so-called ‘mummers’,’ ‘guisers’,’ or ‘Pace-eggers’’ play of Saint George. The history of the sword-dance has received a good deal of attention from German archaeologists, who, however, perhaps from imperfect acquaintance with the English data, have stopped short of the affiliation to it of the play[659] . The dance itself can boast a hoar antiquity. Tacitus describes it as the one form of spectaculum to be seen at the gatherings of the Germans with whom he was conversant. The dancers were young men who leapt with much agility amongst menacing spear-points and sword-blades[660] . Some centuries later the use of sweorda-
gelac as a metaphor for battle in Beowulf shows that the term was known to the continental ancestors of the Anglo-Saxons[661] . Then follows a long gap in the record, bridged only by a doubtful reference in an eighth-century Frankish homily[662] , and a possible representation in a ninth-century Latin and Anglo-Saxon manuscript[663] . The minstrels seem to have adopted the sworddance into their repertory[664] , but the earliest mediaeval notice of it as a popular ludus is at Nuremberg in 1350. From that date onwards until quite recent years it crops up frequently, alike at Shrovetide, Christmas and other folk festivals, and as an element in the revels at weddings, royal entries, and the like[665] . It is fairly widespread throughout Germany. It is found in Italy, where it is called the mattaccino[666] , and in Spain (matachin), and under this name or that of the danse des bouffons it was known both in France and England at the Renaissance[667] . It is given by Paradin in his Le Blason des Danses and, with the music and cuts of the performers, by Tabourot in his Orchésographie (1588)[668] . These are the sophisticated versions of courtly halls. But about the same date Olaus Magnus describes it as a folk-dance, to the accompaniment of pipes or cantilenae, in Sweden[669] . In England, the main area of the acknowledged sword-dance is in the north. It is found, according to Mr. Henderson, from the Humber to the Cheviots; and it extends as far south as Cheshire and Nottinghamshire[670] . Outlying examples are recorded from Winchester[671] and from Devonshire[672] . In Scotland Sir Walter Scott found it among the farthest Hebrides, and it has also been traced in Fifeshire[673]
The name of danse des bouffons sometimes given to the sworddance may be explained by a very constant feature of the English examples, in which the dancers generally include or are accompanied by one or more comic or grotesque personages. The types of these grotesques are not kept very distinct in the descriptions, or, probably, in fact. But they appear to be fundamentally two. There is the ‘Tommy’ or ‘fool,’ who wears the skin and tail of a fox or some other animal, and there is the ‘Bessy,’ who is a man dressed in a woman’s clothes. And they can be paralleled
from outside England. A Narr or Fasching (carnival fool) is a figure in several German sword-dances, and in one from Bohemia he has his female counterpart in a Mehlweib[674] .
With the cantilenae noticed by Olaus Magnus may be compared the sets of verses with which several modern sword-dances, both in these islands and in Germany, are provided. They are sung before or during part of the dances, and as a rule are little more than an introduction of the performers, to whom they give distinctive names. If they contain any incident, it is generally of the nature of a quarrel, in which one of the dancers or one of the grotesques is killed. To this point it will be necessary to return. The names given to the characters are sometimes extremely nondescript; sometimes, under a more or less literary influence, of an heroic order. Here and there a touch of something more primitive may be detected. Five sets of verses from the north of England are available in print. Two of these are of Durham provenance One, from Houghton-le-Spring, has, besides the skin-clad ‘Tommy’ and the ‘Bessy,’ five dancers. These are King George, a Squire’s Son also called Alick or Alex, a King of Sicily, Little Foxey, and a Pitman[675] . The other Durham version has a captain called True Blue, a Squire’s Son, Mr. Snip a tailor, a Prodigal Son (replaced in later years by a Sailor), a Skipper, a Jolly Dog. There is only one clown, who calls himself a ‘fool,’ and acts as treasurer. He is named Bessy, but wears a hairy cap with a fox’s brush pendent[676] . Two other versions come from Yorkshire. At Wharfdale there are seven dancers, Thomas the clown, his son Tom, Captain Brown, Obadiah Trim a tailor, a Foppish Knight, Love-ale a vintner, and Bridget the clown’s wife[677] . At Linton in Craven there are five, the clown, Nelson, Jack Tar, Tosspot, and Miser a woman[678] . The fifth version is of unnamed locality. It has two clowns, Tommy in skin and tail, and Bessy, and amongst the dancers are a Squire’s Son and a Tailor[679] . Such a nomenclature will not repay much analysis. The ‘Squire,’ whose son figures amongst the dancers, is identical with the ‘Tommy,’ although why he should have a son I do not know. Similarly, the ‘Bridget’ at Wharfdale and the ‘Miser’ at Linton correspond to the ‘Bessy’ who appears elsewhere.