
THE PHENOLOGY OF HIBERNATING BATS IN CHALK MINES IN WEST SUFFOLK OVER 76 YEARS WITH COMMENTS ON SURVIVAL OF THE DIFFERENT SPECIES
ROBERT
E STEBBINGS
Background
In the period following the Second World War, a group of undergraduates, mostly studying at Cambridge University, decided to initiate a study of bats hibernating in two series of derelict chalk mines at Bury St. Edmunds. At that time, there were five chalk pits in Bury with tunnels radiating from the cliffs created in the pits. Their study involved occasional winter visits where the locations of animals were marked on hand drawn plans and soon involved putting numbered aluminium bird rings on the bats’ wings. The first bats were ringed in the 1947/48 winter and continued each following winter. By 1951 the students had dispersed but a local resident of Bury was Owen Gilbert, first a student at Manchester and then Bangor Universities. Because Owen lived away from Bury and became a soil scientist at the then newly formed Nature Conservancy based in Grange-over-Sands, Cumbria, he looked for someone to continue the study. My first visit was on 2 January 1952.
At those early times there were few reliable publications detailing knowledge about bats. Those which helped focussed thoughts on what was known and what was unknown, and the evident lack of knowledge helped direct initial studies. Michael Blackmore (1948) and Harrison Matthews (1952) helped by giving advice in person and through their books.
At that time there were four known species traditionally visiting the mines for hibernation, Daubenton’s, Myotis daubentonii, Natterer’s, M. nattereri, Whiskered, M. mystacinus and Brown long-eared, Plecotus auritus. I was taught how to distinguish between these. In addition, I was shown how to find, catch and handle the animals and then to apply rings safely. Owen had produced new rings manufactured by Hughes, also bird-style rings with the inscription UCNW Bangor with number (University College of North Wales).
East Anglia: Origin of mines in
chalk
East Anglia is not generally known for its mines. But looking around Suffolk and Norfolk it is obvious many buildings were constructed up to the early twentieth century largely from flint and lime mortar with some brick on corners and around windows and doors. Many of the mediaeval churches, and a few castles, were constructed in this way often with expensive stone sourced from far away, which was used for facing but with a core of flint and mortar in-fill. Walking around the ruins of the Abbey at Bury St Edmunds immediately impresses the fact a large amount of mortar was used in its construction. Thus, there had been local sources for lime which would have been made locally in kilns and transported by horse and cart.
Amongst the main areas in Suffolk where chalk out-crops is the north-west of the county including around Bury St Edmunds. It was deposited in warm seas of the early Cretaceous era, roughly around 90–100 million years ago. Traditionally, the chalk was subdivided into three broad units: Lower, Middle and Upper Chalk, with the boundaries marked by hard beds that formed distinctive features in the landscape.
The ‘white chalk’ types are amalgamated into one group with that at Bury St Edmunds being the lower layer of that type (Woods et al., 2012).
In fact, all the towns, including the city of Norwich, which sat on shallow soils covering the chalk, once had mines of varying sizes. In Norwich, the combined length of tunnels in the several mines were measured in many kilometres. At Bury St Edmunds, there were at least five mines within the town, all within a kilometre of the Abbey but the Horringer mine was over two kilometres away and may not have contributed to supplying much lime to the town.
Most of the chalk quarrying originally seems to have started as open cast with pick and shovel with the processing kilns situated on the edge or even within the developing quarry. The overburden of soil first needed to be removed and at some stage it was seen to be more efficient just to create adits and removing the best chalk for processing. This was especially relevant where the overburden was particularly thick. The kilns needed a supply of timber, and it is assumed nearby woodland coppicing provided the fuel. Around London in Essex, Hertfordshire and Surrey, chalk was covered by a thick layer of soils and shafts were dug to extract chalk. These features are termed ‘dean’ holes. Some are up to 20m deep.
It has been suggested the mining in some areas may date back as early as the 12th century for lime used in construction, especially in the larger towns. Smaller settlements probably relied more on open pits rather than mining which introduced hazards not present in open cast quarrying. Although there are few records it seems chalk mining ceased as a consequence of the WW1.
At that time, digging machinery and lorries became widespread so the hand winning of chalk became obsolete and small-scale chalk winning was uneconomic. Some of the old mines were seen as presenting a hazard to people wandering into the labyrinth of tunnels so had their entrances bulldozed. Others were abandoned with entrances remaining open.
In all these mines, bats would have found the tunnels as soon as they had been created. Current observations in Wiltshire limestone mines which are under active mining, shows they still attract bats which usually choose the darker corners for hibernation. The often-loud noise of machinery and fan-forced ventilation fails to deter the animals seeking underground refuge.
’
Description of Horringer ‘Caves
There are two separate pits within the original quarried area surrounded by woodland. The adjacent house within the woodland replaced an earlier religious building in the early 19th Century. It is not known whether tunnelling began simultaneously in both pits, but it seems likely the west pit was exploited first with the largest length of tunnels, and a second set of tunnels in the east apparently started with the aim of joining both systems together. The joining tunnel is narrow and barely suitable for extraction of the chalk. It is possible the two pits were exploited by different owners. The effect of the resulting pattern has served to benefit hibernating bats because of the air currents generated by the different levels - the east pit entrances are lower than those in the west.

The dimensions of tunnels are variable throughout the system
A theodolite survey of the tunnels was prepared by Brian Francis (1952). These plans have been modified since and form the maps used throughout this paper.
The dimensions of tunnels are variable throughout the system with some as narrow as 1.4m but most are around 1.8m wide. Heights vary from 1.6m to mostly about 3.0m and the highest up to 5.6m. Generally, the width of passages at roof level is narrower because of the need to maintain coherence of the tunnel in the soft chalk. Several roof falls have occurred where three or more tunnels meet or where the excavated area became too wide. Domes have been created trapping warm air which bats have utilised to digest meals especially in autumn when they first start moving in for hibernation.
Protection for bats
For almost the first 30 years of this study bats were often regarded as ‘vermin’ and local Councils would visit residents where the presence of bats was causing anxiety to kill the bats. Often the Council ‘rat catcher’ would be dispatched to get rid of bats by any method, even poisoning using cyanide. This continued into the early 1970s. In 1975, the Protection of Wild Creatures and Wild Plants Act was passed and that gave some protection to two rare bat species the Greater Horseshoe, Rhinolophus ferrumequinum, and Greater Mouse-eared, Myotis myotis, but all ringing of bats became controlled by licencing. This was both because of the significant disturbance

caused by capture and handling, and the fact that some bats became injured by the rings which are the principal form of individual identification marking. As a result of the Wildlife and Countryside Act 1981, all bats became protected.
At Horringer, unauthorised access was made periodically, sometimes by people burning various materials and clearly causing great disturbance. As a result, steel grilles were installed across the six entrances in 1970. These were broken on several occasions so much more substantial grilles were constructed in 1979.
Aims of this study
During the 76 winters of this study the questions being addressed varied over time as answers were discovered. For convenience, this paper addresses these differing aims in turn for each section. Similarly, the methods changed as the studies progressed and these are detailed in each section. Throughout the study, with the exception of the first ten years, a principal concern has been the well-being of the animals which, to some extent, limited the quantity of results which could be achieved.
These observations were amongst the first of any being made on wild-living bats although some work had been started in the Netherlands in the 1930s, which was not published until 1952 (Bels). After WW2, new studies started, also predominately in the Netherlands and their preliminary results were similarly reflected in the results found at Horringer.
SECTION 1
1.1. The initial aim was to establish whether the same bats returned to the same hibernaculum each winter.
1.2. Secondly, whether the activities of the research workers were affecting the behaviour of the bats.
1.3. Eventually, the important concern was added as to how injuries caused by the bird-style rings could be prevented.
Methods
On each survey visit a search was made for bats using hand torches. When a bat was located it was secured by hand and the location recorded on a plan of the mine. A ring was placed on the wing and the species, sex and number recorded on the plan. If a bat already had a ring the same information was recorded. The first rings had the inscription Saffron Walden MUS with a number while later this changed to ZOO UCNW BANGOR with a number. Later still all rings had LOND ZOO with a number. A total of 1367 bats were ringed in this project covering three periods. The first was from the winter of 1947/48 to 1960/61, (991 ringed) then one winter each for 1970/71 (150 ringed) and 1977/78 (226 ringed). No other kind of individual marking of bats was undertaken.
Results
1.1. Of the first 153 bats ringed in 1947/48 to 1949/50 only 14 were recovered at least once in the same location. These included Daubenton’s, Natterer’s, whiskered s.l. and long eared (subsequently all the latter have been confirmed as Brown long-eared. Whiskered bats were later found to include both Myotis mystacinus and M. brandti). Later periods of this project where ringing was undertaken again, had considerably higher recovery rates.
1.2. It was concluded that bats either left the tunnels when disturbed or became more furtive following first capture and more difficult to find showing the behaviour of the bats did change as a result of capture and examination (Gilbert & Stebbings, 1958). It was found bats are active through the winter apparently not just as a result of disturbance
1.3. Overall, the ratio of males to females for all the bats in hibernation showed a predominance of males from a 1:1 sex ratio at the 2.5 per cent probability level (χ 2=5.08).
1.4. One worrying aspect of ringing was the injuries caused at the point where the often-sharp ring ends rubbed against the wing membrane. It was impossible to judge the true incidence of injury because of the low numbers of recoveries. Of those recovered, the proportion with injury was often at least one third. The rings were designed for the scaly legs of birds. They were made of plain, soft aluminium and stamped out of thin tape. This resulted in sharp ends which rubbed against the wing membrane. Also, being soft metal, some bats chewed the ring making more sharp edges and squashing the ring onto the membrane. It is unknown whether bats died from infections, but that was possible as some injuries became substantial.
Conclusion
1.1. Because the capture, handling, arousing from torpor and release of bats appeared to have an effect on their behaviour, all future studies were to consider their welfare as central to the work.
1.2. The Mammal Society was established in 1954 and Michael Blackmore soon formed a Bat Group. One of the first considerations was the problem of injuries to bats caused by bat rings. As a result, with the considerable assistance of Joe Pickvance of the Birmingham University Extra-Mural department, the quest began to find a ring design to minimise or prevent any injury. Thoughts were gathered from all those who had been ringing bats up to the mid-1950s and new designs were produced. These were trialled at Horringer and elsewhere and eventually a design was produced with a hard magnesium alloy which had all the rough edges removed in the manufacture by Lambourns of Birmingham. Placed correctly and carefully, these caused no injury to bats.
SECTION 2
2.1. Similar observations continued through the 1950s to the winter of 1960/61 but minimising the amount of disturbance caused to individual bats. At the end of that period all ringing ceased and survey visits were to record only the species and location of the animals. If there was a ringed bat the number was read only if it could be done without touching or disturbing the individual.
2.2. The principal aim was to observe how numbers varied each year after the apparent decline from 1947/48 to 1953/54 which had been considered to be caused by over-disturbance by the surveyors (see Figure 1).

Figure 1. Horringer: Maximum number of bats each winter from 1947/48 – 2022/23.
2.3. Consideration was given to the exceptional winter of 1962/63 where extremely low temperatures were recorded across southern Britain lasting continuously for almost three months into March. The question was ‘did this winter affect survival of bat populations in the area?’
Results
2.1. In the 14 winters from 1947/48 to 1960/61 there were a total of 73 complete surveys for bats. There were 2 in August, 10 in September, 9 in October, 8 in November, 16 in December, 6 in January, 8 in February, 7 in March, 6 in April and one in May. Bats were found in each month with just one in August 1949 and six in May 1950.
2.2. The largest number of bats in hibernation each year was generally at the end of December (10 of 14 winters).
2.3. Generally female Daubenton’s arrive in the hibernaculum before the males although the latter become more numerous and dominant by the end of the year, (Gilbert & Stebbings 1958). In September, 27 females were recorded but with only 7 males (79% females) and in October 71 females arrived compared to 32 males (69% females). It was noted the females arriving early appeared to be adults as judged by their dark brown colouration while juveniles of the year had greyish brown fur and they tended to arrive from November onwards.
2.4. After the decline in abundance of bats in the first seven years, there was a steady increase averaging about five bats per year, (Figure

Figure 2. Horringer: Maximum number of bats each winter from 1953/54 – 1963/64.
2.5. From Figure 1 it was apparent there had been a halving of the bats arriving for hibernation in the year following the severe 1962/63 winter. However, thereafter there was a steady increase at the average rate of 6.4 bats per year for 16 years, (Figure 3).

Figure 3. Horringer: Maximum number of bats each winter from 1965 – 1980.
SECTION 3
In order to secure an improved picture of how the bats utilise the Horringer mine, two winters were set aside for detailed surveys. Searches were conducted in the same way as previously. Also, consideration was given to how numbers in hibernation varied over several winters.
3.1. In these two winters, 1971/72 and 1978/79, visits for surveys were undertaken at 15-day intervals from August to April inclusive for the first, and September to April with monthly surveys for the second. In both winters, every bat located was caught and ringed. However, on repeat surveys if a bat was already ringed it would not be disturbed. If the ring was in a position where the number could be read, then it was recorded.
3.2. In addition, the pattern of changing numbers of bats within the tunnels throughout a winter could be determined.
3.3. Also, an aim was to discover whether there was a pattern of visiting of the hibernaculum by bats which was consistent in different years and whether the different species showed similar patterns. Four winters were selected for comparison as they had sufficient survey visits for meaningful comparison. Those winters were 1971/72, 1974/75, 1977/78 and 1978/79.
3.4. From the two years of detailed observations involving ringing, some knowledge could be gained about the longevity and survival rates of the two most abundant species.
Results
3.1. The pattern of occupation of the tunnels by bats in the 1971/72 winter is shown in Figure 4. The accumulative number of counts for the 17 two-weekly surveys was 465 bats. At this time, each bat was disturbed once in order to ring and sex individuals. This disturbance could be expected to affect the number of bats counted on each visit so the counts would be lower than if no disturbance was

3.2. When the pattern of occupation for the four separate winters was plotted as a proportion of each year’s maximum, it showed considerable variation (Figure 5). In 1977/78 (total count 411, n=7) the bats steadily entered hibernation reaching a peak by the first of December. In contrast, the year later, 1978/79 (total count 498, n=7) the ingress by bats was much slower reaching only 40% Januarysix weeks later than previous year. For each year, the pattern of leaving after

Figure 5. Horringer: Movement of bats into and out of the mine in four winters, 1971/72 (⚫), 1974/75 (◆), 1977/78 () & 1978/79 (◼).
3.3. The ingress of bats entering the tunnels was examined for the two winters when ringing occurred, 1971/72 and 1977/78 as a proportion of the maximum total reached. It was found that there was as much as a two month variation between the 50% point of entering the mine for combined results for Daubenton’s and Natterer’s (Figure 6). The results did show the peak number was recorded by 1 March in 1971/72, but only achieved by 1 May in 1977/78. This shows bats continue to visit throughout the winter and roost in places where they can be counted as individuals.

Figure 6. Horringer: Movement of bats into the mine 1977/78 (⚫) % of final total/15 days intervals (Daubenton’s + Natterer’s)
s, it is seen

Figure 7. Horringer: Movement of bats into the mine 1971/72 . % of final total/15 days intervals Daubenton’s (⚫) & Natterer’s (◼)
SECTION 4
4.1. As a result of ringing the bats in two winters, 1971/72 and 1977/78, some detailed observations were made on precise roost selection by a few individual bats. It had long been noted that individual holes and crevices were regularly occupied each winter by the same species of bat but without ringing it was not possible to know whether it was the same bat.
4.2. In addition, in the winter 1978/79 many hundreds of temperature measurements were made using a rapid response temperature recorder (typically recording within 5 seconds). Temperatures were recorded at roof level as well as at ground level throughout the whole mine and at the exact position where each animal was in hibernation. This was done on each month through the winter. As a result, it was possible to relate the position where bats roosted to the temperatures of their selection. Care was required to ensure heat from the recorder’s bodies did not influence the recorded temperature.
Results
4.1. In the later periods of ringing bats, the individual animals were disturbed by handling only once on initial capture. However, for individual animals which roosted at lower heights on the walls of the adits and happened to present their ring in a position where the number could be read without touching the bat, it allowed the assembly at a roost selection history for a few individuals.
4.2. Figure 8 shows the roosting behaviour of a single adult female Daubenton’s bat through a span of four winters, 1971/72, not recorded in 1972/73, but was recorded again in 1973/74 and 1974/75. The bat had been caught and ringed in September 1971 and it never used the same precise roost location again. However, it was identified on ten other occasions and all within about 20m in the same area of the mine. The bat was in the same location in October and November 1971, noted as absent in December 1971 but it reappeared in the same site by January 1972 where it was recorded in February and March. There was no knowledge whether the bat remained hidden in the tunnels for December or whether it left and was elsewhere.

4.3. Similar results were noted for an adult female Natterer’s ringed in December 1971 but relocated in January 1972 roosting in a cold, partially light area near an entrance where it occurred also a year later (Figure 9). The same bat was in the same crevice as originally captured three years later in 1974 and again two years after that. Over the six winters it was only not recorded in 1975/76. It could have been present but roosting somewhere where the ring could not be read.

Figure 9. Movements of individual hibernating bats as winter progresses. Natterer’s female Ring 1881.

Figure 10. Seasonal changes through the winter of 1971/72 in percentage of bats occupying the warmer, stable parts of the mine (all species). Bats gradually concentrate in the cooler, but more variable temperature zones near entrances. (Fitted line y=89.4-5.6x r2 0.87 n=466)
4.4. The detailed temperature recordings made throughout the winter of 1978/79 are plotted in Figures 11–17. These illustrate how the temperatures in the tunnels change through the winter and as expected, gradually cool throughout the mine with the maximum length of tunnels being coldest from late January to February (See Figure 10). However, even the places furthest from entrances gradually become colder although there is no noticeable air circulation in most areas. On windy days, air currents can be detected in passages which connect areas between entrances. In such weather, bats either vacate the long thin passage connecting the two pits or if remaining, then hide away deeper in crevices out of the air flow.






Figures 11–17. Detailed temperature recordings made throughout the winter of 1978/79 showing how temperatures gradually cool with the maximum length of tunnels being coldest from late January to February.

4.5. The numbers of bats found on each survey day throughout 1978/79 were plotted on the temperature plans and the number found in each temperature zone was expressed as a percentage of each total for that month. The number of Natterer’s bats observed were too few to be of significance on their own (Table 1) so they were combined with Daubenton’s for comparison with ambient temperatures found throughout Horringer. Therefore, only the abundant species, the Daubenton’s and Natterer’s were counted. The results are shown in Table 2. In the table the figure beneath the date is the total number of bats found on that survey date. The bold percentage figures highlight where the largest proportion of bats were located. Overall, it was clear how the bats gradually moved towards the colder entrance areas as winter progressed.
Table 1. Numbers of Natterer’s bats found per temperature regime found throughout the winter of 1978/79.
Table 2. Percentage of bats in each temperature range as winter progresses. Figures beneath months are the number of bats counted for that date. (Blank sections indicate no choice was available) Figures in bold highlight the principal numbers of bats.
SECTION 5
This section concerns various characteristics of the different species found in Horringer over the full 76 winters. Daubenton’s have always been most abundant with fewer Natterer’s more-or-less following proportionally. The other species have been Brown long-eared, Plecotus auritus, whiskered, which after 1971 were found to include Myotis mystacinus and M. brandti, and Barbastelle, Barbastella barbastellus.
5.1. Were the numbers of Daubenton’s and Natterer’s proportionally similar throughout the study or was one more likely to be increasing or declining differentially?
5.2. From the two periods of ringing bats in 1971/72 and 1977/78 with subsequent surveys each winter, what were the survival characteristics for the combined results for Daubenton’s and Natterer’s bats? Was the survival similar for the two periods?
5.3. Some Brown long-eared bats were found almost every winter throughout the study. Was the number found similar throughout or have numbers showed any trends?
5.4. At the beginning of the study, whiskered (s.l.) bats were found in most winters but none has been seen since the winter of 1999/2000. Was this loss driven by any sudden event or was it a gradual attrition?
5.5. A few Barbastelle bats have been found intermittently through the study. Is there any pattern for the occurrence of this uncommon species?
Results
5.1. The maximum number of Daubenton’s and Natterer’s bats found each winter in the Horringer hibernaculum is shown in Figure 18. When plotting the ratio of the two species related to Daubenton’s for the period 1967/68 to 2022/23 it shows the ratio has remained more-or-less equal throughout the 56 winters at 73% Daubenton’s (Figure 19). Prior to 1967/68, the number of bats found was very variable and low with an average total of 32 bats per winter for all species.
5.2. During the two winters when ringing took place in the 1970s, each bat was disturbed once only for identification, sexing and ringing. Subsequent winter surveys recorded the number of individuals which were already ringed to those un-ringed. In order to be able to distinguish in which winter the bats were ringed, during the first winter, 1971/72 all bats were ringed on the right forearm and in the second winter they were ringed on the left. In subsequent years, usually a few bats were too tightly squeezed into crevices which did not allow determination whether they were ringed or not. Therefore, the resultant survival calculations are underestimates (Figure 20). Nevertheless, the rate of survival was similar for the two periods with the 1977/78 ringed bats having a higher level. From Figure 1 it is evident the population from 1971/72 for the following eight years showed numbers each winter increased at the rate of about 6.4 bats per year (Figure 3). Shortly after 1977/78 the rate of increase rose rapidly.

17. Horringer: Maximum number of bats each winter from 1947—2023. Daubenton’s (blue) & Natterer’s (red)

18. Horringer: Ratio of Daubenton’s bats to Natterer’s bats each winter from 1967/68 2022/23

Figure 20. Horringer: Survival of ringed bats from 1971/72 & 1977/78 (combined results of Daubenton’s + Natterer’s bats)
5.3. Brown long-eared bats have been found usually in small numbers and if several visits were made through a winter, they tended to be found early in October but may then be absent later in the winter. The maxima counted per winter

Figure 21. Horringer: Maximum number of Brown long-eared bats each winter from 1947/48 2022/23.
Suffolk Nat. Soc. 59 (2023)
5.4. At the start of this project in the 1940s and 1950s, whiskered bats were encountered regularly but not in large numbers. Following the papers by Hanák, (1970, 1971) in which the whiskered bats were separated into two distinct species, it was possible to determine which species we had in the West Suffolk chalk mines. However, most whiskered bats were found before 1971. Checking all records, there were 80 of whiskered s.l. but after separation there were only two Myotis mystacinus critically identified (both males) and eight Myotis brandti (five males and three females). A further nine Brandt’s were seen in the same winters as ones critically identified and as they were not
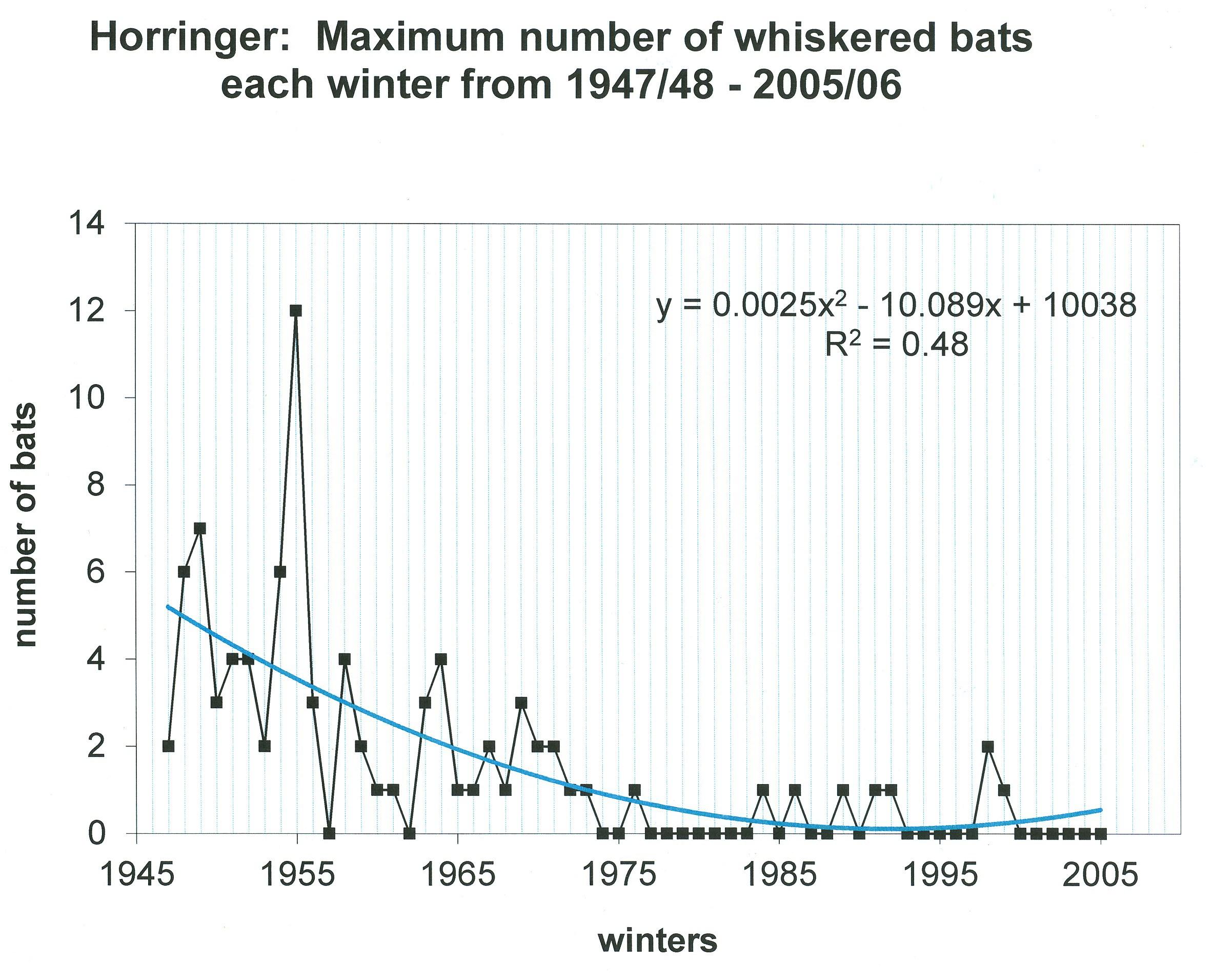
22. Horringer: Maximum number of whiskered bats each winter from 1947/48 2005/06.
5.5. Only 25 records of the Barbastelle have been made in the 76 winters (Figure

Figure 23. Horringer: Maximum number of Barbastelle bats each winter from 1947/48 2005/06.
5.6. Miscellaneous observations: Three species records were quite unexpected. Firstly, a Lesser Horseshoe bat, (Rhinolophus hipposideros, was found on 18 December 1958 which was in the same tunnel on 9 January 1959 and also on 31 January, 14 March and 20 April 1959, (Stebbings, 1960). At the time only two other Lesser Horseshoes had been recorded from East Anglia. Since then, one other Lesser Horseshoe has been found and that was at Elveden (Suffolk) some 18km north of Bury St. Edmunds. It was first found on 10 February 1996 and every subsequent year to 6 February 2016. The only year it was not present was 1997. (Arthur Rivett, pers. comm.).
A second unexpected bat, not usually found underground, was a pipistrelle, s.l. found within chalk tunnels on 11 November 1958 and recorded in the same place on 18 December 1958. It was located again on 10 October 1969 at Buxton, Norfolk, some 74km to the northeast and eleven winters later. It had just been killed by a cat.
The third bat which was exceptional was a Serotine, Eptesicus serotinus, found and ringed on 1 January 1997. The previous month had been unusually cold with nearby temperatures down to -10oC and most nights were frosty.
Serotines have been recorded hibernating in derelict limestone mines (author’s records) over several years, but this behaviour is quite exceptional.
From around 1960 for ten years there were several Daubenton’s bats showing varying degrees of flavism. In 1968 a fully flavistic specimen was found at Horringer while one had been found across the town in February 1961

Discussion
Pivotal to all the studies on bats carried out by this author over a long career working on bats and other animals, was the discovery by the mid-1950s that bats were sensitive to disturbance. It appeared, once disturbed, the bats tended to avoid being caught again (Gilbert & Stebbings, 1958; Rybar, 1973). This, of course could be expected because unscheduled arousals cause bats to utilise energy which they had stored in the autumn. Several such awakenings with subsequent loss of body mass could lead to death before food becomes available again in spring (Stebbings, 1969). Several observations were made on bats in other situations which showed how quickly bats can lose mass on disturbance (Stebbings, 1966). A 9.47g Brown longeared bat lost mass when handled to take measurements at the rate of 0.022g/ minute a total of 0.45g in 20 minutes.
Studies on bats in mainland Europe began in late 1920s and 1930s which often incorporated field and laboratory investigations (e.g. in Germany, Eisentraut, 1934, 1935, 1936). In Britain, amongst the first field bat studies involving bat ringing began in the late 1940s (Hesketh, 1951; Hooper & Hooper, 1956). However, by the mid1950s, research in the Netherlands showed similar results to those being found in Horringer with reductions in numbers of bats being found in hibernacula seemingly caused by observers (Sluiter, & Heerdt, 1957, 1964; Heerdt & Sluiter, 1957). Comparative findings were reported in Poland by Krzanowski (1959). It was noteworthy that when catching and ringing ceased at Horringer, the number of bats being recorded gradually increased again.
Concern about the disturbance being caused to bats in hibernation and damage caused to them have been expressed many times (e.g., Bárta, et al., 1981; Bezem, et al., 1960; Rybar, 1973; Stebbings, 1978; Topal, 1962). Topal considered bats could not be ringed without causing injuries and was concerned as to how ringing could be reducing survival. Stebbings (1965) estimated about 20% of the bats ringed at Horringer suffered some injury from the rings. However, all those, like most bat research workers across the world at that time, used soft aluminium rings which bats chewed and distorted squeezing them onto the forearm and soft membrane. From all the studies, none established any connection between injuries caused by rings and mortality. Perhaps this is not surprising as bats are likely to die away from the hibernaculum where they were ringed and remain undiscovered.
The movement of bats into and out of underground mines through each winter has been well documented (Baagøe, et al., 1988; Bezem, et al., 1964; BagrowskaUrbańczyk & Urbańczyk, 1983; Daan, 1973; Egsbaek, & Jensen, 1963; Punt, & Parma. 1964.) However, the reasons for the movements were not clearly understood until the thermal preferences for each species had been found, particularly by laboratory experimentation as well as under field conditions (Harmata, 1969; & 1973). Unfortunately, Harmata observed only Brown long-eared, Natterer’s and Barbastelle bats in his studies of those found at Horringer. In winter, the Brown long-eared were found hibernating in temperatures from -3 to +11oC with mean of 5.4oC and
Natterer’s, 6 to 10oC and Barbastelle at -3 to +9oC with mean of 3.6oC. His conclusion was the Brown long-eared and Barbastelle preferred cold temperatures which was reflected in those bats arriving in underground places intermittently in the coldest weather.
At Horringer we were able to show how the two most abundant species, Daubenton’s and Natterer’s responded to the changing ambient temperatures occurring through the tunnels. These results were similar to those recorded on mainland Europe, (Daan, 1967, 1970; Daan et al., 1980: Daan & Wichers, 1968; Dorgelo & Punt, 1969; Gaisler, 1970; Kuipers & Daan, 1970; Wijngaarden, 1968). This internal ‘migration’ of bats throughout the hibernation period is now accepted in addition to bats which appear to move in and out of any given hibernaculum through all winters. What is known is that every winter is different in the pattern of movements apparently depending on the weather. What has not been established is how weather affects these movements. Much more research is needed to analyse the weather with regard to the response by bats. Some attempt has been made in regard to the results at Horringer but it became obvious the analysis is complex requiring considerable resources to effect some useful result.
Throughout the study at Horringer the relative proportion of Daubenton’s and Natterer’s remained remarkably consistent despite total numbers varying considerably. However, when the population was increasing rapidly in the late 1970s and early 1980s this had an effect on increasing survival rates which might have been predicted. The overall pattern of population change as shown in Figure 1 will be discussed further, below. Actual survival and longevity of individual bats was observed with only a few achieving a maximum predicted life span. The longest records were for male Daubenton’s with four individuals exceeding 10 years from ringing to last recovery. Generally, the age of each was not known at the time of ringing. However, one bat ringed on 28 December 1949 was last seen on 30 December 1967 and hence at least 18.5 years old from birth, assumed to be in June (Stebbings, 1965, 1968). It had been seen on six other winters through its life. A female Natterer’s bat ringed on 27 December 1951 had been recorded only twice but refound on 25 January 1964, an age of at least 12 years. Another female Natterer’s bat was ringed on 2 December 1971 and refound on seven winters with the last sighting on 29 January 1989, therefore at least 17.5 years old. One male whiskered s.l. bat was recorded seven years after ringing and it had been recaptured five times. No Brown long-eared bats were seen again in any winter after that of its ringing. Similar maxima results for the Netherlands were recorded with Daubenton’s, 15.5, Natterer’s, 17.5, whiskered, 18.5 and Brown long-eared with 17.5 years respectively (Heerdt & Sluiter, 1955; Heerdt, 1961).
The number of Brown long-eared bats has never been large with a maximum of twelve, but the variation was always unpredictable. Nevertheless, the apparent increase from 1947 to mid-1980s with subsequent decline again to the 2020s, suggests environmental factors outside the hibernaculum are the cause.
Similarly, the apparent decline in whiskered bats s.l. is not surprising as it was never an abundant species in East Anglia (Barrett-Hamilton, 1910: Sergeant & Harrison, 1947). After whiskered bats were separated into the two species (M. mystacinus & M. brandti) it was found both occurred in Horringer but most records pre-dated that discovery (Stebbings, 1991). The confirmed records for Suffolk show a total of 13 Brandt’s for Bury St Edmunds sites and only two Whiskered. Elsewhere in Suffolk there was only another single specimen for both species and eleven specimens which were not critically identified. Ten of those also were in Horringer. Generally, Whiskered bats are more abundant in the west and south of Britain (authors observations). Within mainland Europe they are widespread and abundant (Daan, 1973; Sluiter et al.,1956) but further north in Jutland, Denmark, they are less common (Egsbaek & Jensen, 1963). The last whiskered bats s.l. bats seen at Horringer were two in 1998/99 and a single specimen the following winter. None has been found for the last 23 winters. A small number of records (eight) have been made using ultrasound detectors but there are some concerns as to how accurate these are.
The species with the fewest records in Horringer is the Barbastelle with a total of 25, from the first in 1954/55 to the most recent in 2005/06. Generally, this species has always been considered rare across Britain but recent observations using ultrasonic detectors have demonstrated it is more widespread (Harris & Yalden, 2008). It has been found to be mostly dependant on trees for roosts, often behind loose bark or in splits. Also, it has been found in the roots of trees growing on banks where the soil has eroded (Tony Vine, pers. comm.). In eastern and northern Europe, it is found hibernating in large numbers in old buildings and mines, e.g. Bogdanowicz, & Urbańczyk (1983).
Throughout the study when surveys are usually made between 09.00 - 12.00 a few bats often were found awake, sometimes grooming but others were seen flying along the passages. Squeaking was heard and on investigation one bat had landed on another with the awake bat clearly trying to awaken the other for mating (Gilbert & Stebbings, 1958). This behaviour has been observed with both Daubenton’s and Natterer’s and in all months of the winter. This has been recorded in other hibernacula (Roer & Egsbaek, 1969).
Another aspect of behaviour which became known early in the project was that it was necessary to walk carefully through the tunnels and avoid walking on any loose, soft scree. Often squeaking was heard under-ground and on one occasion a hole was carefully excavated by hand in an effort to find the squeaking bat. In the course of removing chalk lumps, six torpid bats (all Daubenton’s) were found before finding the squeaking individual. Again, similar situations have been recorded (Egsbaek et al., 1971; Roer & Egsbaek, 1966). Earlier it had been reported (Stebbings, 1965) that as a result of a roof fall in Horringer in 1956 in a section of tunnel which had been surveyed for bats previously that morning, two Daubenton’s bats were found in the rubble which could not have been seen earlier.
So, this raises the question, ‘how many bats are present in the mines when surveys are undertaken?’ Comparing visual counts within hibernacula were related to the
number recorded leaving the roost caught by traps and nets and some projects used automatic recorders (Baagøe et al., 1988; Degn et al., 1995). Typically, single visual counts in these studies recorded small numbers, e.g., 50 bats, but as a result of trapping and automatic recorders, the estimated population size was 3500–5000 Daubenton’s bats. Krzanowski (1959) counted bats in a cave on 602 occasions and recorded a maximum of 15 bats. His average was 2.5 bats per visit. However, from detailed mark and recapture studies he recorded an average of 247 bats present, representing just 6% for the highest count of 15.
By accident we found some observations which showed results directly comparable to Horringer. At Grimes Graves neolithic flint mines near Brandon, several of us had been entering during winters from the late 1960s in order to count bats (Stebbings, 1992; 1993). The total was always less than ten, comprising Daubenton’s and Natterer’s. The concrete cap to the 10m deep shaft had a 600 x 700mm man-hole cover which was raised about 20mm to allow some ventilation to the flint mine. Bats landed on the ground level concrete cap and crawled through the narrow gap to gain entry. On 11 November 1980 seven Daubenton’s were counted in the tunnels which ramify all around the base of the shaft. Early in 1981 English Heritage placed a small hut over the manhole which had no ventilation holes large enough for bats to pass through. Over the course of several weeks the resident custodian found bats within the hut each morning and released them outside. Over 120 were caught and released. Clearly these bats had been present within the mine and most had been hidden when a bat survey was undertaken. From this it appears no more than 8% of bats present could be seen by three experienced bat surveyors.
Knowing that on occasions while surveying Horringer it became apparent through accidents that hidden bats were found, if there was a similar proportion to that found at Grimes Graves it suggests the number present could be 4375 bats when visual counts were around 350. Additionally, because it has been found there is movement into and out again of bats through the winter, the total number of bats using the mines could be several thousand annually.
Figure 1 shows the great variation in maximum numbers of bats counted each winter. After the low numbers recorded in the 1950s and 1960s, the number rose gradually at first then suddenly increased through the 1970s and 1980s. No direct correlation was obvious either with weather or landscape changes. However, around 1970, it was estimated around 30 million elm trees died and were felled. These would include many containing bat roosts (Potter, et al., 2011). Was this assumed loss of roosts a factor in numbers of bats in Horringer?
Since then, the number fell sharply and suddenly in the last winter 2022/23 rose again. The 1950s were characterised by the loss of almost half the hedgerows with their trees across lowland Britain (Pollard, et al., 1974). In addition, the Wildlife Trusts report half a million ponds have been lost in the last 100 years along with their rich wildlife so together this huge loss of habitat will have greatly reduced the number of insects available for bats. Williams (1938) working at the Rothamsted Experimental Station investigated the behaviour of flying insects at night in particular. Following on
from that pioneering work, L. R. Taylor (pers. comm.) stated that the density of flying insects at night from 1947 to 1953 declined by 50%. The implication being that bats had to fly twice as long to catch food at the end of that period compared with the start. There does not appear to be comparable data to suggest how insect densities have changed since 1953, but with loss of ‘wild’ habitat and the use of fertilisers and pesticides they are likely to have reduced insect diversity and abundance considerably.
Also, the use of pesticides in the environment was widespread after WW2 certainly causing mortality and possible sub-lethal effects on bat populations (Jefferies, 1972). Other landscape changes such as ploughing flood plain pastures up to the riverbanks would further reduce insect populations on which the bats depend (Robinson & Stebbings, 1994). In addition to landscape degredation, there has been treatment of timbers in buildings, often using persistent pesticides (Shore, et al., 1990; 1991; Stebbings, 1988). These kinds of problems have been reported across all of Europe but usually it was not known which were the most significant causes of observed declines in bat populations (Stebbings, 1988).
Finally, what is the meaning of the apparent increase in numbers of bats in Horringer from 1950 to 1990s? Clearly, over the 76 years of this study there has been enormous loss of wildlife habitat including loss of roosts, declines in insect food and use of pesticides all of which are known to impact bats. It is possible with the number of roosts being reduced bats seek to use remaining sites. It does seem likely the sudden large rise last winter could be attributed to the unusually long, severe cold spell in December which encouraged more bats to seek hibernation sites away from freezing conditions.
Acknowledgements
This project was begun by students at Cambridge in the late 1940s. As they finished their degrees they dispersed and Owen Gilbert, who’s family lived at Bury St. Edmunds, became involved, but he too was studying and working far away. In late 1951 I was invited to become involved as I lived only a kilometre away from Horringer. My first training was on 2 January 1952 and only a second one a month later before being left to continue by myself. I did receive practical advice from Michael Blackmore and later from L. Harrison Matthews when preparing reports. Further support came from the SNS former President Lord (Jock) Cranbrook. He supported my bat studies from the 1950s and was himself interested in bats and their welfare and additionally, was a leader in achieving legal protection for bats in Britain.
Over the years, many people assisted with the field surveys and especially from around 1990 Arthur Rivett (Suffolk Bat Group) helped and, following the property becoming a care home and now a health centre, he has generously been the focus for access and ongoing site management, especially repairs to grilles. Although first grilled around 1970, substantial grilles were installed in 1990 by Maurice Webber. I am grateful for the Suffolk whiskered records which Martin Sanford has provided. Finally, I wish to thank Antony Mould for helping to guide me through the analysis of results and preparation of the figures. Critical comment on this paper was kindly given by Russell Stebbings, University Museum of Zoology, Cambridge.
References
Barrett-Hamilton, G. E. H. (1910). A history of British Mammals: Vol. 1 - Bats. Gurney & Jackson, London.
Baagøe, H. J., Degn, H. J. & Nielsen, P. (1988). Departure dynamics of Myotis daubentoni (Chiroptera) leaving a large hibernaculum. Vidensk. Meddr. Dansk naturh. Foren. 147, 7–24.
Bagrowska-Urbańczyk, E. & Urbańczyk, Z. (1983). Structure and dynamics of a winter colony of bats. Acta theriologica. 28, 183–196.
Bárta, Z., Červený, J., Gaisler, J., Hanák, P., Hanák, V., Horáček, I., Hůrka, L., Miles, P., Nevrlý, M., Rumler, Z., Sklenář, J. & Žalman, J. (1981). [Results of the winter census of bats inCzechoslovakia; 1969–1979]. (inCzech). Okresniho muzea v Moste 3, 71–116 Bels, L. (1952). Fifteen years of bat banding in the Netherlands. Publ. Natuurh. Genoots. Limburg 5, 1–99.
Bezem, J. J., Sluiter, J. W. & Heerdt, P. F. van. (1960). Population statistics of five species of the bat genus Myotis and one of the Rhinolophus, hibernating in the caves of S. Limburg. Arch. Néerl Zool. 13, (4) 511–539.
Bezem, J. J., Sluiter, J. W. & Heerdt, P. F. van. (1964). Some characteristics of the hibernating locations of various species of bats in South Limburg. I & II. Proc. Kon. Ned. Akad. Wet. (C) 67, 325–350.
Blackmore, M. (1948). Mammals in Britain. London, Collins.
Bogdanowicz, W. & Urbańczyk, Z. (1983). Some ecological aspects of bats hibernating in City of Poznań. Acta Theriologica. 28, 371–385.
Daan, S. (1967). De Geulhemergroeve: gevolgen van de afsluiting vaneen mergelgroeve voor het vleermuizenbestand. Natuurh. Maandbl. 56, 154–160.
Daan, S. (1970). Photographic recording of natural activity in hibernating bats. Bijdragen tot de Dierkunde 40, (1) 13–16.
Daan, S. (1973). Activity during natural hibernation in three species of Vespertillionid bats. Arch. Neérl. Zool. 23, (1) 1–71.
Daan, S. (1980). Long term changes in bat populations in the Netherlands: a summary. Lutra. 22, 95–105.
Daan, S., Glas, G. H. & Voute, A. M. (1980). De Nederlandse vleermuizen: Bestandsontwikkelingen in winter - en zomerkwartieren. Lutra 22, 1–118.
Daan, S., & Wichers, H. J. (1968). Habitat selection of bats hibernating in a limestone cave. Z. Saugetier., 33, 262–287.
Degn, H. J., Andersen, B. B. & Baagøe, H. (1995). Automatic registration of bat activity through the year at Mønsted Limestone mine, Denmark. Z. Saugetier., 60, 129–135.
Dorgelo, J. & Punt, A. (1969). Abundance and internal migration of hibernating bats in an artificial limestone cave. (Sibbergroeve). Lynx. n.s. 10, 134–143.
Egsbaek, W. & Jensen, B. (1963). Results of bat banding in Denmark. Vidensk. Meddr. dansk Naturh. Foren. 125, 269–296.
Egsbaek, W., Kirk, K. & Roer, H. (1971). Beringungsergebnisse an der Wasserfledermaus (Myotis daubentoni) und Teichfledermaus (Myotis dasycneme) in Jütland. Decheniana-Beihefte, 18, 51–55.
Eisentraut, M. (1934). Markierungsversuche bie Fledermaüse. Z. Morph. Oekol. 28, 553–560.
Eisentraut, M. (1935). Ergebnisse der Fledermausberingung nach dreijähriger Versuchzeit. Z. wiss. Biol. 31, 1–26.
Eisentraut, M. (1936). Zur Fortpflanzungsbiologie der Fledermaüse. Z. Morph. Oekol. 31, 27–63.
Francis, B. (1952). Cave exploration of Bury St. Edmunds. East Anglian Mag. 11, 335–340.
Gaisler, J. (1970). Remarks on the thermopreferendum of Palearctic bats in their natural habitats. Bijdragen tot de Dierkunde. 40, (1) 33–35.
Gilbert, O. & Stebbings, R. E. (1958). Winter roosts of bats in West Suffolk. Proc. zool. Soc. Lond., 131, 329–333.
Hanák, V. (1970). Notes on the distribution and systematics of Myotis mystacinus Kuhl, 1819. Bijdr. Dierk, 40, 40–44.
Hanák, V. (1971). Myotis brandtii (Eversmann, 1845) (Vespertillionidae Chiroptera) in der Tschechoslowakei. Vĕst. čsl. Spol. Zool. 35, 175–185.
Harmata, W. (1969). The thermopreferendum of some species of bats (Chiroptera). Acta theriol., 14, 49–62.
Harmata, W. (1973). The thermopreferendum of some species of bats (Chiroptera) in natural conditions. Zesz. Nauk. UJ Prace zool. 19, 127–141.
Harris, S. & Yalden, D. W. Eds. (2008). Mammals of the British Isles: Handbook, 4th Edition. The Mammal Society, Southampton.
Heerdt, P. F. van. (1961). New data on longevity in bats. Natuurh. Maandbl 50, 36.
Heerdt, P. F. van, & Sluiter, J. W. (1955). Longevity in bats. Nat. Hist. Maandblad. 44, 35–36.
Heerdt, P. F. van, & Sluiter, J. W. (1957). The results of bat banding in the Netherlands in 1956. Nat. Hist. Maandblad. 46, 13–16.
Hesketh, G. E. (1951). Ringing bats in Denbighshire. Naturalist, Lond. 1951, 177–181. Hooper, J. H. & Hooper, W. M. (1956). Habits and movements of cave-dwelling bats in Devonshire. Proc. zool. Soc. Lond. 127, 1–26.
Jefferies, D. J. (1972). Organochlorine insecticide residues in British bats and their significance. J. Zool., Lond 166, 245–263.
Krzanowski, A. (1959). Some major aspects of population turnover in wintering bats in a cave at Puławy (Poland). Acta Theriologica, 3, 27–42.
Kuipers, B. & Daan, S. (1970). Internal migration of hibernating bats: Response to seasonal variation in cave microclimate. Bijdragen tot de Dierkunde. 40, (1) 51–55.
Matthews, L. H. (1952). British Mammals. London, Collins.
Mitchell-Jones, A. J., Cooke, A. S., Boyd, I. & Stebbings, R. E. (1989). Bats and remedial timber treatment chemicals - a review. Mammal Review, 19, 93–110.
Nagel, A. & Nagel, R. (1991). How do bats choose optimal temperatures for hibernation? Comp. Biochem. Physiol. 99A (3) 323–326.
Nagel, A. & Nagel, R. (1991). Remarks on the problem of optimal ambient temperatures in hibernating bats. Myotis, 29, 109–114.
(2023)
Pollard, E., Hooper, M. D. & Moore, N. W. (1974). Hedges. New Naturalist. Collins, London.
Potter, C., Harwood, T., Knight, J. & Tomlinson, I. (2011). Learning from history, predicting the future: the UK Dutch elm disease outbreak in relation to contemporary tree disease threats. Philos Trans R Soc Lond B Biol Sci. 366, (1573) 1966–74.
Punt, A. & Parma, S. (1964). On the hibernation of bats in a marl cave. Publ. Natuurh. Gen. Limburg, 13, 45–59.
Robinson, M. F. & Stebbings, R. E. (1994). Changing land-use in south Cambridgeshire: its effect on Serotine Bats. Nature in Cambridgeshire, 36, 62–68.
Roer, H. & Egsbaek, W. (1966). Zur Biologie einer skandinavischen Population der Wasserfledermaus (Myotis daubentoni) (Chiroptera) Z. Säugetierk. 31, 440–453.
Roer, H. & Egsbaek, W. (1969). Uber die Balz der Wasserfledermaus (Myotis daubentoni) (Chiroptera) im Winterquartier. Lynx. n.s.10, 85–91.
Rybar, P. (1973). Remarks on banding and protection of bats. Period. biol. 75, 177–179.
Sergeant, D. E. & Harrison, D. L. (1947). An occurrence of the whiskered bat (Myotis mystacinus Kuhl.) Naturalist, Lond. 1947, 152.
Shore, R. F., Boyd, I. L., Leach, D. V., Stebbings, R. E. & Myhill, D. G. (1990). Organochlorine residues in roof timbers and possible implications for bats. Environ. Pollut. 64, 179–188.
Shore, R. F., Myhill, D. G., French, M. C., Leach, D. V. & Stebbings, R. E. (1991). Toxicity and Tissue Distribution of Pentachlorophenol and Permethrin in Pipistrelle Bats Experimentally Exposed to Treated Timber. Environ. Pollut. 73, 101–118.
Sluiter, J. W. & Heerdt, P. F. van. (1957). Distribution and decline of bat populations in S. Limburg. Natuurh. Maandbl 46, 134–143.
Sluiter, J. W. & Heerdt, P. F. van. (1964). Distribution and abundance of Bats in S. Limburg from 1958 till 1962. Natuurh. Maandbl. 53, 164–173.
Sluiter, J. W., Heerdt, P. F. van & Bezem, J. J. (1956). Population statistics of the bat Myotis mystacinus, based on the mark-recapture method. Arch. Néerl Zool. 12, (1) 64–88.
Stebbings, R. E. (1960). Lesser horseshoe bat in West Suffolk. Proc. zool. Soc. Lond., 133, 483.
Stebbings, R. E. (1965). Observations during sixteen years on winter roosts of bats in West Suffolk. Proc. zool. Soc. Lond., 144, 137–134.
Stebbings, R. E. (1966). Bats under stress. Stud. Speleol., 1, 168–173.
Stebbings, R. E. (1968). Longevity of Vespertilionid bats in Britain. J. Zool., 156, 530–531.
Stebbings, R. E. (1969). Observer influence on bat behaviour. Lynx, 10, 93–100.
Stebbings, R. E. (1978). Marking bats. In: Animal marking: recognition marking of animals in research, Ed. B. Stonehouse, 81–94. London: Macmillan.
Stebbings, R. E. (1988). Conservation of European Bats. Christopher Helm, London. 246pp.
Stebbings, R. E. (1988). Bats and buildings. Proceedings 8th British Pest Control Conference. 8pp.
Stebbings, R. E. (1991). Bats: Order Chiroptera. In: Handbook of British Mammals. 3rd Edn. Eds. G. B. Corbet & S. Harris. Blackwell Scientific, London. 81–87, 99–108, and 123–130.
Stebbings, R. E. (1992). Bats in Greywell Tunnel, Basingstoke Canal. Rep. English Nature, Newbury. 89pp.
Stebbings, R. E. (1993). The Greywell Tunnel. An internationally important haven for bats. English Nature (South Region). Newbury. 32pp.
Topal, G. A. (1962). Some experience and results of bat banding in Hungary.Symposium Theriologicum (proceedings of the International Symposium on Methods of Mammalogical Investigation held in Brno, Czechoslovakia, 1960) 339–344.
Wijngaarden, A. van. (1968). De natuurlijke luchtcirculatie in ondergrondse kalksteengroeven in Zuid-Limburg. Publ. Natuurh. Gen. Limburg 17, 5–14.
Williams, C. B. (1939). An analysis of four years captures of insects in a light trap. Part I. General survey; sex proportion; phenology; and time of flight. Trans. R. Ent. Soc. Lond. 89, 79–131.
Woods, M. A., Mortimore, R. N. & Wood, C. J. (2012). The Chalk of Suffolk. In: Dixon, R., (ed.) A celebration of Suffolk geology: GeoSuffolk 10th Anniversary Volume. Ipswich, UK, 105–131.
Robert Stebbings
Willowbrook House
Kings Cliffe
Peterborough
PE8 6XN
