Priručnik sa smjernicama i mjerama prilagodbe



Fakultet šumarstva i drvne tehnologije Sveučilišta u Zagrebu
Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu
Agronomski fakultet Sveučilišta u Zagrebu
Zagreb, siječanj 2023. godine

O projektu
Projektom Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima - "MEMORIE" se želi doprinijeti rješavanju problema nedostatka podataka i radova o utjecaju, ranjivosti, otpornosti i mogućnosti prilagodbe na klimatske promjene u 3 ranjiva sektora (šumarstvo, prirodni ekosustavi i bioraznolikost, i poljoprivreda).
Dobivene spoznaje i baze podataka će omogućiti analize ranjivosti i otpornosti šuma, prirode i biološke raznolikosti i poljoprivrede u odnosu na predviđene modele klimatskih promjena. Na taj način će se unaprijediti znanja koja će omogućiti razvoj mjera prilagodbe uzajamnom suradnjom multisektorskog istraživačkog tima i dionika.
Cilj projekta jest suradnjom istraživačkih institucija putem primijenjenih istraživanja u ranjivim sektorima predložiti 10 mjera prilagodbe klimatskim promjenama za dionike.
Istraživačkim aktivnostima u projektu “MEMORIE” će se definirati konkretne mjere koje dionici mogu praktično implementirati u svakodnevni rad. Korištenjem rezultata primijenjenih istraživanja u okviru ovoga projekta razvit će se metode prilagodbe na klimatske promjene kojima će se značajno poboljšati već postojeći postupci upravljanja i gospodarenja prirodnim resursima u ranjivim sektorima. Rezultati istraživanja su naročito važni za gospodarenje i upravljanje prirodnim resursima čiji je razvoj usko povezan te snažno ovisan o klimi, a prilagodba u većini situacija spora.
Vrijednost projekta: 3.150.324,95 HRK
Ukupno prihvatljivi troškovi: 3.077.080,77 kn
Intenzitet EU potpore: 85%
Intenzitet nacionalne potpore: 15% financirao Fond za zaštitu okoliša i energetsku
učinkovitost
Bespovratna sredstva: 2.615.518,14 HRK
Razdoblje trajanje projekta: 36 mjeseci - u razdoblju od siječnja 2020. do siječnja 2023.
Naručitelj projekta: Ministarstvo gospodarstva i održivog razvoja
www.sumfak.unizg.hr/memorie
• Korisnik: Sveučilište u Zagrebu, Fakultet šumarstva i drvne tehnologije
• Partner 1: Sveučilište u Zagrebu, Agronomski fakultet
• Partner 2: Sveučilište u Zagrebu, Prirodoslovno-matematički fakultet
Više informacija na : QR kod
Predgovor
Ovaj priručnik nastao je kao rezultat projekta Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima - “MEMORIE”. Projektom se doprinijelo rješavanju problema nedostatka podataka i radova o utjecaju, ranjivosti, otpornosti i mogućnosti prilagodbe na klimatske promjene u 3 ranjiva sektora koja se temelje na prirodnim resursima (šumarstvo, prirodni ekosustavi i bioraznolikost te poljoprivreda), sukladno Sedmom nacionalnom izvješću Republike Hrvatske prema Okvirnoj konvenciji Ujedinjenih Naroda o promjeni klime
Provedene istraživačke aktivnosti u projektu “MEMORIE” predlažu konkretne mjere koje dionici mogu praktično implementirati u svakodnevni rad. Korištenjem rezultata primijenjenih istraživanja u okviru ovoga projekta razvijeni su prijedlozi konkretnih mjera prilagodbe na klimatske promjene kojima će se značajno poboljšati već postojeći postupci upravljanja i gospodarenja prirodnim resursima u ranjivim sektorima.
 Autori
Autori
Bioraznolikost Poljoprivreda Šumarstvo 6 54 66
Klimatske promjene i ŠUMARSTVO
Stjepan Mikac1*
Marko Orešković1
Domagoj Trlin1
Anja Žmegač3
Stjepan Dekanić2
Vinko Paulić1
Damir Ugarković1
Milan Oršanić1
Igor Anić1
1 / Sveučilište u Zagrebu, Fakultet šumarstva i drvne tehnologije, Zavod za ekologiju i uzgajanje šuma, Svetošimunska cesta 23, 10000 Zagreb, Hrvatska
* / email: smikac@sumfak.hr
2 / PERCEPTIVES j.d.o.o., A.T. Mimare 4, 10000 Zagreb, Hrvatska
3 / Technical University of Munich, Hans-Carl-v.-Carlowitz-Platz 2 85354 Freising, Germany
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 6
1. UVOD
Klimatske promjene su jedan od gorućih problema u svijetu s dalekosežnim posljedicama na socioekonomski, ekološki i održivi razvoj svih zemalja. Iako smo svjesni promjena, naše poznavanje njihovog porijekla ali još važnije mogućih posljedica je još uvijek ograničeno.
Stope i brzina kojom se klima mijenja znatno premašuju vrijednosti kojima se šumski ekosustavi mogu brzo prilagoditi. Za dugovječne organizme kao što su stabla odnosno šume, brza progresija klimatskih promjena znači da će na kraju svog života doživjeti izrazito drugačiju klimu u odnosu na uvjete pod kojima su se razvijali. Takav scenarij sve veće neprilagođenosti šuma na promjene vjerojatno će negativno utjecati na osiguravanje širokog spektra usluga koje ovi vrijedni ekosustavi pružaju te će u konačnici dovesti do brojnih nepovoljnih ekoloških i ekonomskih posljedica za cijelo gospodarstvo i društvo u cjelini. Utjecaji klimatskih promjena na šumske ekosustave mogu biti neposredni i posredni (FAO i CIFOR 2019). Neposredni utjecaji uključuju posljedice postupnih promjena klimatskih parametara i/ili ekstremnih vremenskih događaja kao što su na primjer: promjene u temperaturnim obrascima, toplinski valovi, suše, mraz, olujni vjetrovi i dr. Posredno klimatske promjene utječu na proizvodnost i funkcioniranje šumskih ekosustava putem promjena do kojih dovode kod drugih živih sastavnica ekosustava, kao što su: oprašivači, štetnici, prijenosnici bolesti i invazivne vrste. Također u obzir treba uzeti i utjecaje ljudskih aktivnosti, kao što je prenamjena zemljišta i neodrživa praksa korištenja zemljišta, koji u međudjelovanju s neposrednim i posrednim utjecajima klimatskih promjena zajednički određuju ranjivost i otpornost neke vrste ili staništa na klimatske promjene.
Prema Lindner i dr. (2010) utjecaji klimatskih promjena na šumske ekosustave mogu biti i pozitivni i negativni. U sjeverozapadnoj Europi očekuje se pozitivni učinak na rast stabala kao posljedica veće koncentracije CO2, većih temperatura i produljenja vegetacijske sezone (Myneni i dr. 1997). S druge strane, povećanje suše i učestalosti vremenskih ekstrema donijet će brojne negativne posljedice, pri čemu je vjerojatno da bi negativni učinci mogli nadmašiti pozitivne. U svakom slučaju pretpostavka je da će se utjecaj klimatskih promjena značajno reflektirati na geografski marginalnim populacijama (Linares & Camarero 2012), u ekosustavima koji su oslabljeni i destabilizirani antropogenim djelovanjem (Markham 1996) ili u ekosustavima koji imaju vrlo usku ekološku nišu. Genetski i ekološki mehanizmi tijekom povijesti su olakšali prilagodbu na promjene te su prirodnom selekcijom rezultirali razvojem optimalnih ekotipova prilagođenih lokalnim uvjetima što se najbolje može potvrditi kod vrsta iz roda (Quercus sp.) (Kremer 2020). No istodobno u uvjetima naglih klimatskih promjena, ekološki optimum se mijenja, ali postoji i određeni vremenski odmak prije nego što se razvije optimalni ekotip koji je prilagođen na novonastale uvjete (Bürger and Krall, 2004).
Utjecaj neposrednih klimatskih promjena na šumske ekosustave radi pojednostavljenog shvaćanja možemo podijeliti u dvije skupine. Prva skupina obuhvaća spore promjene uzrokovane konstantnim porastom temperature zraka, povećanjem suše te povećanom koncentracijom CO2 u odnosu na predindustrijsko razdoblje. Druga skupina obuhvaća neočekivane i nepredvidive ekstremne klimatske događaje (elementarne nepogode) kao što su: poplave, suše, orkanski vjetrovi, oluje, ledolomi, snjegolomi, požari i sl. koji nanose direktne ekološke (promjene biološke raznolikosti, usluga ekosustava i dr.) i materijalne štete. U posljednjih 20 godina samo na šumskim ekosustavima u Republici Hrvatskoj štete koje su nastale kao izravna ili neizravna posljedica klimatskih promjena višestruko nadmašuju sve zabilježene štete u posljednjem stoljeću.
ŠUMARSTVO 7
Ukupna površina šuma i šumskih zemljišta u RH iznosi 2.759.039 ha, što je oko 47% ukupne kopnene površine Republike Hrvatske. Od toga je 2.097.318,16 ha u vlasništvu RH, dok je ostatak od 661.720,89 ha u vlasništvu privatnih posjednika. Glavninom šuma i šumskih zemljišta u državnom vlasništvu (96%) temeljem Zakona o šumama, gospodari trgovačko društvo „Hrvatske šume d.o.o.“. Prema namjeni šume mogu biti gospodarske (temeljna je namjena proizvodnja šumskih proizvoda uz očuvanje i unapređenje usluga ekosustava) i zaštitne (prvenstveno služe za zaštitu zemljišta, voda, naselja, objekata i druge imovine), a iznimno šume i šumska zemljišta prema namjeni mogu biti šume posebne namjene (zaštićeni dijelovi prirode, sjemenske sastojine, šume za znanstvena istraživanja, šume za potrebe obrane RH, urbane šume i šume prema posebnim propisima). Bez obzira na veliki udio šuma i šumskog zemljišta u ukupnoj površini RH, tek je 55.3% visokog uzgojnog oblika (nastale generativno iz sjemena). Relativno veliki ostatak od 44.7% su šume niskog uzgojnog oblika te razni oblici degradiranih šuma. Degradirani oblici šumskih sastojina su posljedica neprimjerenog upravljanja šumama u prošlosti, šumskih požara i negativnog utjecaja ostalih abiotičkih ili biotičkih čimbenika. Gospodarenje šumama s ekonomskog gledišta isključivo se odvija u šumama sjemenjača kojih ima svega 0,9 milijuna hektara (55.3%) i koje osiguravaju najvrjednije drvne sortimente.
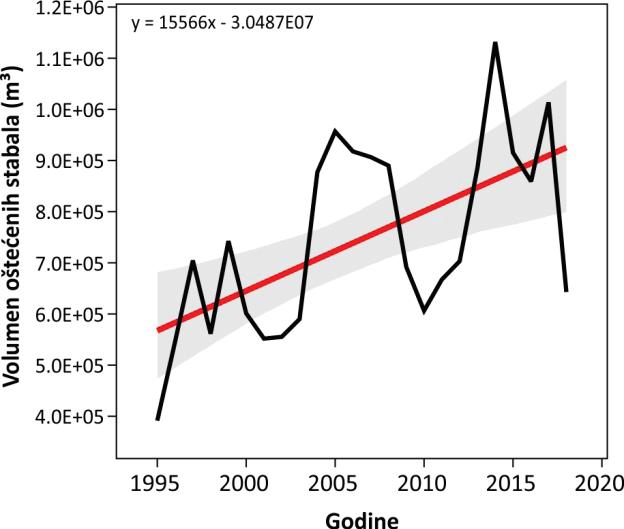
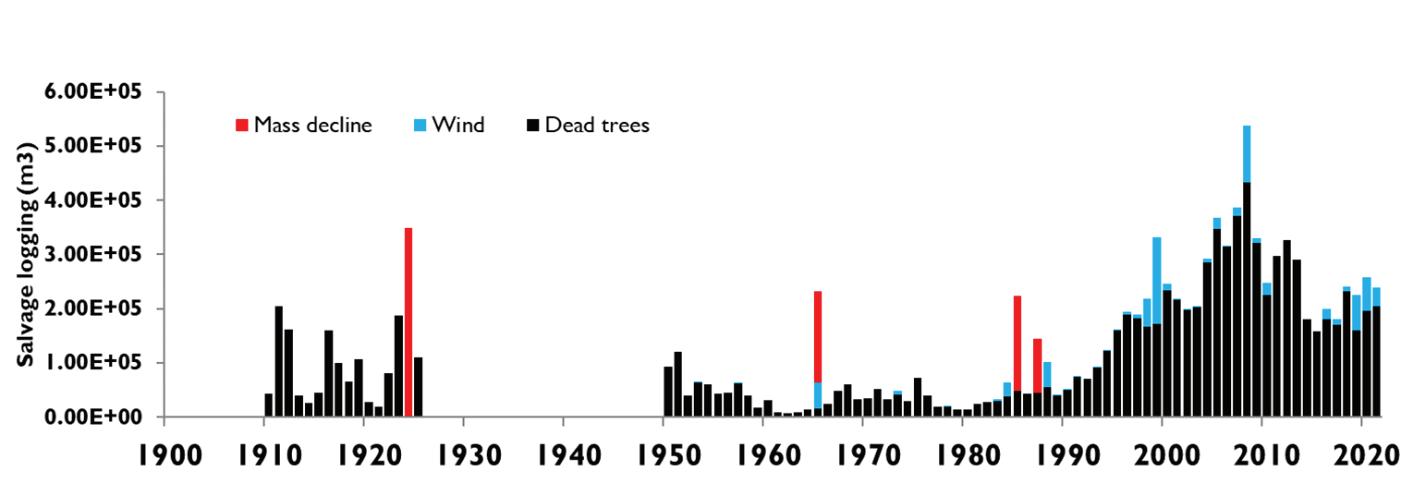
Šumarstvo kao gospodarska djelatnost ima značajan utjecaj na razvoj ruralnih područja, iako njegov udio u nacionalnom BDP-u iznosi svega 1,2 %. Održivo upravljanje šumama u RH ima višestoljetnu tradiciju očuvanja i poboljšanja zaštite bioraznolikosti i brojnih usluga ekosustava. Ukupni broj zaposlenih u šumarstvu iznosi oko 9.400 osoba. Klimatske promjene praćene šumskim požarima, ledolomima, vjetroizvalama, sušenjem i propadanjem vrsta drveća i napadima štetnika (potkornjaci i patogeni organizmi) direktno i indirektno uzrokuju štete u šumama na cijelom području Republike Hrvatske. Ukupan broj evidentiranih šteta prema posječenoj drvnoj masi (volumen drva u m3) u Republici Hrvatskoj od 1995. do 2021. godine iznosi gotovo 20 miljuna m3 drveta. Od toga je 63.8 % suhih stabala (odumiranje uzrokovano kompleksnim čimbenicima), 18 % stabala uklonjenih kao sanitar (zaražena, insektima napadnutih, oštećena stabla) 15.6 % stabala oštećenih vjetrom (vjetroizvale + vjetrolomi) te oko 2 % stabala uklonjenih zbog šteta od snijega i leda.
Slika 1. Trend povećanja šteta (u m3 drvne mase) nastao kao posljedica prirodnih nepogoda u razdoblju od 1995 do 2020. godine (izvor: Hrvatske šume d.o.o.)

Prema količini posječene mase oštećenih stabala od prirodnih nepogoda najoštećenije vrste su hrast lužnjak (Quercus robur L.) sa 6.960.066 m3, obična jela (Abies alba Mill.) sa 3.987.371 m3, obična bukva (Fagus sylvatica L.) sa 1 830 106 m3 te obična smreka (Picea abies Karst.) sa 1.340.131 m3 drvne mase. Najveći udio šteta od stabala koja su odumrla od kompleksnih čimbenika utvrđen je za hrast lužnjak (86%), poljski jasen (83%), smreku (70%) i jelu (48%). Kod obične bukve najveći dio šteta prouzrokovan je vjetrom i to u iznosu od 54%. Značajan udio šteta od vjetra utvrđen je i kod hrasta kitnjaka u iznosu od 25% te kod crnog bora u iznosu od 19%. Kod jele i smreke ovako veliki postotak sušenja vjerojatno se
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 8
najvećim dijelom odnosi na sušenje prouzrokovano patogenim organizmima (potkornjacima), dok je kod jasena gotovo sav udio sušaca nastao zbog patogene gljive Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz & Hosoya.
Dinamika šteta od prirodnih nepogoda tijekom promatranog razdoblja ukazuje na značajan porast šteta na šumskom drveću kod svih vrsta. Analizom prosječnih iznosa štete, uzimajući u obzir individualne događaje, utvrđeno je da su najveće štete prouzrokovane vjetrom (vjetroizvale, vjetrolomi) zatim sušom, ledom, potkornjacima i snijegom. Vjetar kao jedna od najznačajnijih nepogoda naročito je izražen 1999., 2008., 2014. te 2017. godine sa pojedinačnim iznosom i preko 300 000 m3 drvne mase. Dugoročno planiranje kao i provedba aktivnih mjera prilagodbe u šumarstvu zahtjeva poznavanje svih mogućih scenarija utjecaja klime na rast i razvoj pojedinih vrsta drveća. Način na koji će se vrste prilagoditi budućim trendovima kretanja klime jedno je od osnovnih ekoloških pitanja koje zaokuplja brojne znanstvenike, praktičare i donositelje odluka. Neadekvatna prilagodba gospodarenja šumama ima potencijalno štetne učinke na funkcioniranje ekosustava, biološku raznolikost, zdravlje i turizam. Štoviše, s obzirom na to da su mnoge vrste koje žive u šumama vrlo ovisne o prevladavajućim vrstama drveća, odgođeni odgovor na promjenjivu klimu može pridonijeti izumiranju i prikriti klimatski uzrokovan gubitak biološke raznolikosti. Međutim, klimatske promjene vjerojatno će pojačati elementarne nepogode koje direktno i relativno brzo mijenjaju stanište za razne vrste biljaka i životinja.
Kako bismo unaprijedili kvalitetu i raspoloživost podataka iz područja klime te poboljšali spoznaje o utjecaju, ranjivosti vrsta drveća i razvili konkretne mjere prilagodbe klimatskim promjenama neophodno je u što kraćem vremenu prikupiti kvalitetne podatke i popuniti praznine u podacima i znanju te provesti analize koje će rezultirati konkretnim mjerama prilagodbe. Stoga je ovaj projekt usmjeren na prikupljanje podataka o klimi, analize utjecaja klime na šume, biološku raznolikost koristeći interdisciplinarni istraživački pristup.
ŠUMARSTVO 9
2. MATERIJAL I METODE RADA
2.1. Istraživanja budućih rasprostranjenosti šumskih vrsta drveća
Pod pojmom rasprostranjenosti određene vrsta drveća možemo smatrati prikladnost staništa za određenu vrstu drveća prema trenutnim ali i budućim okolišnim čimbenicima kao što su klima, tlo, hidrografske i orografske prilike i dr. Modeliranje prikladnosti staništa (ponekad zvano modeliranje distribucije vrsta ili ekološke niše vrste) je metoda za predviđanje prikladnosti određenog područja za vrstu ili grupu vrsta, na temelju njihovog trenutnog (povijesno poznatog) odnosa prema okolišnim čimbenicima. Kakva će biti buduća dinamika i prostorna distribucija pogodnosti staništa za glavne vrste drveća u šumskim ekosustavima Republike Hrvatske ključno je pitanje za predlaganje nekih od konkretnih mjera prilagodbe posebice u promijenjenim uvjetima uzrokovanim klimatskim promjenama. Takve mjere zahtijevaju znanje o trenutačnoj prostornoj rasprostranjenosti pogodnosti staništa za vrste drveća, utjecaju ekoloških čimbenika (npr. klima, tlo) kako bi otkrili ugrožena područja ali i moguća nova područja ekološke prikladnosti uzimajući u obzir modele projekcija klimatskih promjena. Korištenje bolje prilagođenih vrsta šumskog drveća (genotipovi) i selekcija podrijetla (genotipizacija) imaju potencijal poboljšati otpornost šumskih sustava i omogućiti potpomognute migracijske strategije (Hanewinkel et al., 2014.; Marchi i Ducci, 2018.), čime se pomažu adaptivni procesi šumskih ekosustava (Ferrarini i sur., 2016). No osim toga izbor vrsta za pošumljavanje na neobraslim proizvodnim površinama također nalazi uporište u analizama buduće prikladnosti staništa za određenu vrstu drveća. Kako bismo prilagodili svakodnevne stručne aktivnosti poput kvalitetnog odabira vrsta drveća za pošumljavanja i uzgajanje ali i bolje usmjerili monitoring i praćenje promjena u šumskim ekosustavima potrebno je odrediti područja koja više neće biti pogodna za određenu vrstu ali i eventualno nova (pogodna) područja obzirom na povijesni raspon ekoloških čimbenika.
2.1.1. Metode rada
Za modeliranje trenutne te potencijalne buduće rasprostranjenosti pogodnosti staništa za glavne vrste drveća korištena je platforma za modeliranje skupnih distribucija vrsta (BIOMOD2). BIOMOD2 se učestalo koristi za istraživanje utjecaja klimatskih promjena na šume i vrste drveća ali i druge taksonomske skupine. U analizama su korištena četiri modela distribucije i to: generalizirani linearni modeli (GLM), generalizirani aditivni modeli (GAM), generalizirani modeli pojačanja (GBM) i slučajna šuma (RF). Za svaku pojedinu vrstu drveća model je kalibriran na trenutnim podacima o pojavnosti vrste te je predviđena buduća potencijalna distribucija kao karta buduće vjerojatnosti pogodnosti staništa, čije su vrijednosti pretvorene radi uštede memorije u cijele brojeve u rasponu od nula do tisuću. Konačno, za svaku vrstu drveća izračunali smo konsenzusni model uprosječivanjem (medijan vrijednost) projekcija pojedinačnih modela. Prisutnosti i odsutnosti jednako su ponderirani postavljanjem parametra prevalencije BIOMOD2 na 0,5. Projicirali smo modele (kalibrirane za trenutne uvjete) u budućnost koristeći dva scenarija emisija (RCP 4.5 i RCP 8.5) i dva 40-godišnja vremenska razdoblja. S popisa potencijalnih prediktorskih varijabli (klime i tla) odabrane su samo one koje objašnjavaju većinu varijacija u promatranoj prisutnosti i odsutnosti svake vrste rekurzivnim pristupom eliminacije (RFE) implementiranim unutar algoritma slučajne šume (Breiman 2001). Unutar RFE pristupa, varijable su eliminirane iterativno, počevši od punog skupa potencijalnih prediktora i zadržavajući samo one varijable koje smanjuju srednju kvadratnu pogrešku u odnosu na slučajne permutacije iste varijable. Identificirane su varijable koje su bile u linearnoj korelaciji s drugim varijablama i imale faktore inflacije varijance VIF > 5, prag koji se često koristi za otkrivanje mulikolinearnosti (Craney i Surles 2002; Thompson et al. 2017). Identificirane kolinearne varijable s nižom vrijednošću prema Akaike informacijskim kriterijima (AIC) (Akaike 1974) zadržane su za daljnji razvoj modela. Ovaj podskup nekoreliranih klimatskih varijabli i
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 10
varijabli tla korištene su kao prediktorske varijable za razvoj skupnih modela distribucije vrsta. Podaci o trenutnoj rasprostranjenosti vrsta drveća preuzeti su iz baze prve Nacionalne inventure šuma CRONFI (Čavlović 2010). CRONFI plohe ravnomjerno prekrivaju različite tipove šuma i šumske vegetacije u RH a sadržavaju podatke o geografskom položaju ploha (Latitude, Longitude), nadmorskoj visini te brojnim biometrijskom podacima kao što su promjeri i visine stabala te vrste drveća. Iz navedene baze preuzeti su podaci na razini svake pojedine plohe i to: prostorni položaj plohe (Latitude, Longitude) te prisustvo vrste (0 - nema, 1 – vrsta je zabilježena). Na ovaj način je kreirana baza od preko 7000 ploha.
prema odabranim vrstama drveća Vrsta
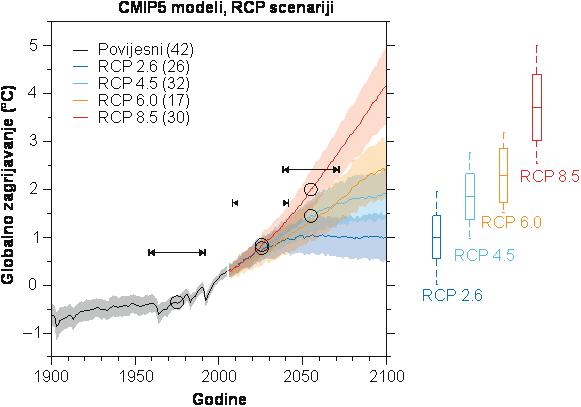
Klimatski podaci korišteni u ovom istraživanju preuzeti su iz baze ClimateEU (Marchi i dr. 2020). Podaci predstavljaju homogenizirane nizove osnovnih i izvedenih varijabli temperature zraka i oborina. Inicijalno baza je kreirana u prostornoj rezoluciji od 1 km2 te je kao takva korištena za modeliranje utjecaja klimatskih promjena na prikladnost staništa za rast i razvoj odabranih vrsta drveća. Softver ClimateEU se može se koristiti za procjenu više od 50 mjesečnih, sezonskih i godišnjih varijabli, uključujući mnoge ekonomski ili biološki relevantne varijable. Za prostorno modeliranje staništa i pojavnosti vrsta, koristili smo prostorno interpolirane klimatske podatke rezolucije 1 × 1 km u Albersovoj projekciji. Za izradu predviđanja za normalno razdoblje 1961. – 1990. i za dva predviđena vremenska odsječka korištene su skupne projekcije 2020-ih (2011. – 2040.), 2050. (2041. – 2070.). Navedene skupne projekcije obuhvaćaju prosjek za 15 modela opće cirkulacije atmosfere i oceana koji odgovaraju IPCC izvješću o procjeni 5 (CanESM2, ACCESS1.0, IPSL-CM5A-MR, MIROC5, MPI-ESM-LR, CCSM4, HadGEM2-ES, CNRM-CM5, CSIRO Mk 3.6, GFDL-CM3, INM-CM4, MRI-CGCM3, MIROC-ESM, CESM1-CAM5 i GISS-E2R), koji predstavljaju sve klastere sličnih AOGCM-ova i imaju visoku statistiku validacije u svojim CMIP3 ekvivalentima (Marchi et al. , 2020). Svako vremensko razdoblje predviđeno je za reprezentativni put koncentracije (RCP) s vrijednošću zračenja od +8,5 W/m2. Ova forsirana vrijednost rezultira projiciranim globalnim zagrijavanjem od približno +2,0°C (±0,6) do 2050-ih i +3,7°C (±0,9) do 2080-ih (Marchi et al., 2020).
Slika 2. Projekcije globalnog zagrijavanja prema modelima emisija. Strelice na grafu predstavljaju raspon vremenskih razdoblja za koje su napravljene projekcije pogodnosti staništa.
ŠUMARSTVO 11
(latinski naziv) Broj ploha s vrstom Broj ploha
vrste Obična jela (Abies alba Mill.) 533 6789 Obična smreka (Picea abies Karst) 262 7060 Obična bukva (Fagus sylvatica L.) 2321 5001 Hrast lužnjak (Quercus robur L.) 846 6476 Poljski jasen (Fraxinus angustifolia Vahl) 353 6969 Hrast crnika (Quercus ilex L.) 201 7121 Alespki bor (Pinus halepensis) 162 7160
Tablica 1. Broj ploha
bez
Podaci o fizičkim i kemijskim svojstvima tala preuzeti su iz baze SOILGRIDS v2.0 (Poggio i dr. 2021). Preuzeti podaci predstavljaju prosjeke vrijednosti za sve slojeve tala do dubine od 60 cm (zona dubine korijenskog sustava većine naših vrsta drveća). Podaci su izvorno generirani u prostornoj rezoluciji od 250 m koja je naknadno pomoću bilinearne interpolacije uprosječena na prostornu rezoluciju od 1 km2 identičnu rezoluciji klimatskih podataka. U analizama prikladnosti staništa su korišteni podaci o fizičkim i hidrološkim svojstvima tala i to: BDOD – gustoća tla (cg/cm3), CEC – kapacitet izmjenjivih kationa (mmol(c)/kg), Clay – gravimetrijski sadržaj čestica gline u tlu (g/kg), FC – poljski kapacitet tla za vodu (mm), Nitro - sadržaj ukupnog dušika (amonijak, organski i reducirani dušik) izmjeren Kjeldahlovom digestijom plus nitratnitrit (cg/kg), PAW – biljci pristupačna voda u tlu FC – WP) (mm), PH – reakcija tla (sadržaj vodikovih iona H+ u vodi) (x10), Sand - gravimetrijski sadržaj čestica pijeska u tlu (g/kg), Siltgravimetrijski sadržaj praškastih čestica u tlu (g/kg), THS – sadržaj vode u kod potpunog zasićenja tla (mm), WP – sadržaj vode u točki uvenuća (mm).
2.2. Istraživanja osjetljivosti i otpornosti šumskih ekosustava na klimatske promjene u prošlosti
2.2.1. Postavljanje ploha ne terenu
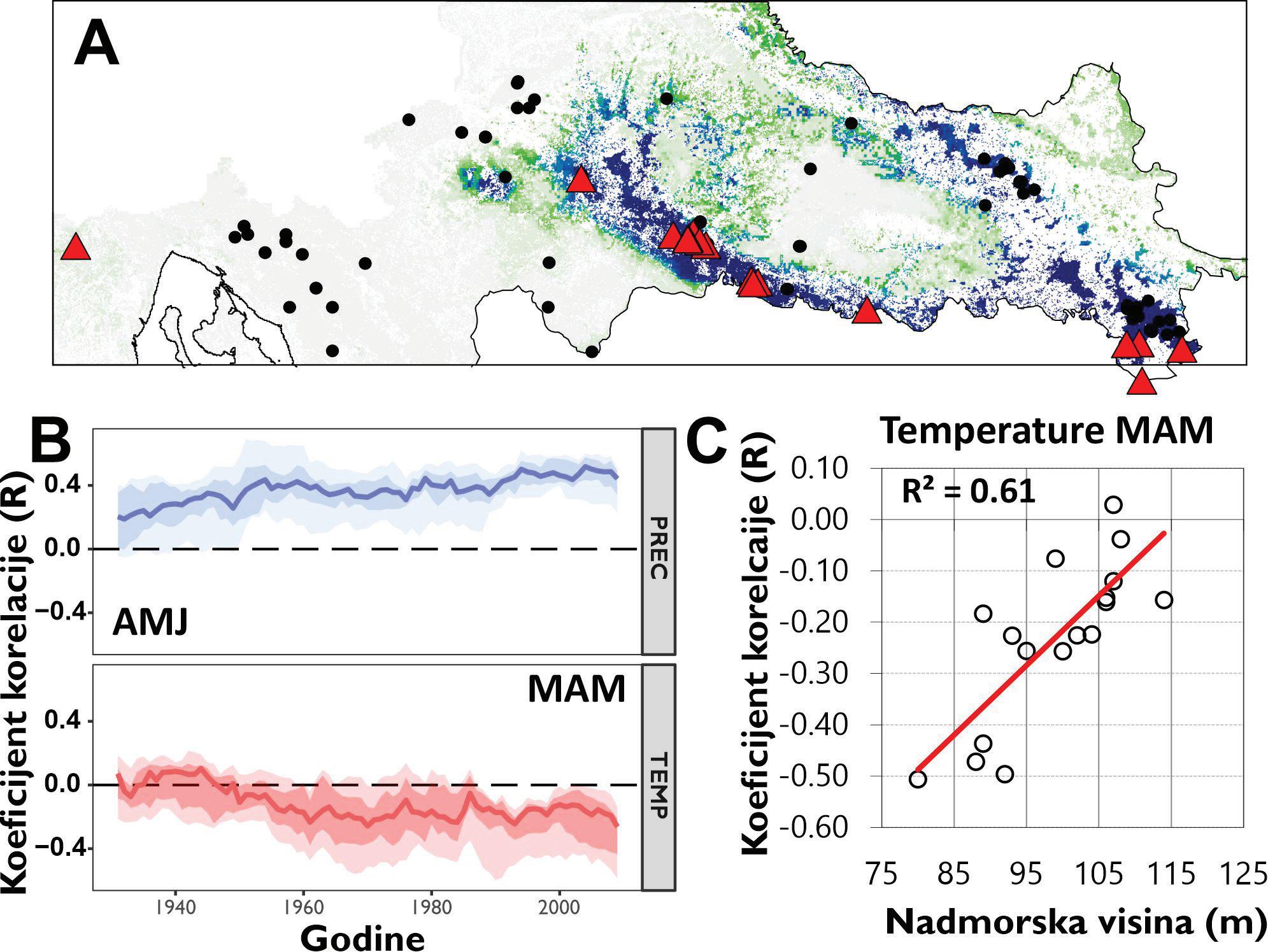
Odabir lokacije za postavljanje pokusne plohe ovisi o puno različitih čimbenika kao što su: struktura sastojine, starost, oblik sastojine, ekspozicija i inklinacija terena, utjecaj čovjeka kroz povijest i dr. Prilikom postavljanja pokusnih ploha za potrebe projekta MEMORIE, obraćena je posebna pažnja da pokusne plohe budu osnovane u što starijim šumskim kompleksima (zbog zahvaćanja što duljeg vremenskog razdoblja interakcije klime i rasta stabala), te da reprezentativno predstavljaju šumske sastojine i glavne vrste drveća Republike Hrvatske.
2.2.2. Izmjera podataka na plohama
U tako odabranim sastojinama odabrano je minimalno 15 dominantnih stabala za uzorkovanje kako bi što bolje predstavljali odabrani lokalitet. Na svakom lokalitetu, tj. svakoj pokusnoj plohi je utvrđena nadmorska visina (m), prevladavajući nagib i ekspozicija. Na svakom pojedinom stablu su izmjerene sljedeće vrijednosti: prsni promjer (cm), visina (m), vrsta drveta, morfološka oštećenja, stanje krošnje i opći izgled vitaliteta stabla, živo/odumrlo stablo, dubeće mrtvo stablo/ležeće mrtvo drvo
2.2.3. Prikupljanje uzoraka sa stabala
Na svim odabranim stablima pomoću Pressler-ovog svrdla promjera 4.9 mm prikupljena su po dva uzorka. Uzorci su uzimani na prsnoj visini (~ 1,30 m od razine tla) nasuprotno jedan od drugoga te okomito na nagib terena kako bi se izbjeglo uzorkovanje reakcijskog drva. Nakon vađenja uzorci su označeni rednim brojem plohe i identifikacijskim oznakama (lokalitet, ploha, vrsta drveća, broj uzorka) prema uputama ITRDB (The International Tree-Ring Data Bank) te stavljeni u kartonske spremnike za transport.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 12
2.2.4. Sušenje i lijepljenje uzoraka
Nakon povratka s terena prikupljeni uzorci su stavljeni na sušenje na sobnoj temperaturi od 24 - 48 h. Prvi korak u procesu obrade, nakon sušenja izvrtaka je lijepljenje uzoraka na za to izrađene drvene podloške. Prilikom lijepljena uzoraka bitno je postaviti uzorak tako da su provodni elementi (traheje odnosno traheide) postavljeni vertikalno kako se nalaze u deblu stabla, kako bi godovi bili jasno uočljivi nakon obrade.

2.2.5. Mehanička i kemijska obrada uzoraka
Nakon sušenja i lijepljenja obavljena je gruba i fina (poliranje) mehanička obrada uzoraka i preparacija pomoću uređaja Core-Microtome. Mikrotom je uređaj dizajniran za površinsku pripremu cijelih izvrtaka (duljine do 70cm), omogućuje mikroskopski precizno rezanje ravnih površina (debljine 0.1 mm) kako bi se jasno vidjele granice godova. Kod nekih uzoraka kod kojih nakon mehaničke obrade i dalje nisu jasno vidljive granice godova (obična bukva) potrebno je kemijski tretirati uzorak kako bi se bolje vidjele granice ranog i kasnog drva. Uzorke smreke i jele (četinjača) ponekad je potrebno dodatno očistiti od smole (Soxhletov aparat) u kombiniranoj otopini toluena i etanola 2:1 u trajanju od 48 h.
ŠUMARSTVO 13
Slika 3. Prikupljanje podataka i uzoraka na terenskim plohama. Izgled svježe izvađenog uzorka s jasno vidljivim širinama godova (gore desno).
2.2.6. Digitalizacija uzoraka
Pripremljeni uzorci se skeniraju pomoću sustava ATRICS (Advanced Tree Ring Image Capturing System) koji se sastoji od automatiziranog mjernog stola i digitalne kamere (Olympus) povezanih sa računalom. Fotografije nastale prilikom pomicanja mjernog stola spajaju se u jednu snimku visoke rezolucije koja predstavlja polazište za izmjeru širine godova. Vrlo bitan korak prilikom skeniranja je postavljanje kalibracijskog stakalca na površinu uzorka kako bi se prilikom očitavanja vrijednosti širine godova iz piksela mogle pretvoriti u stotinke milimetra.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima

14
Slika 4. Laboratorijska obrada prikupljenih uzoraka. Lijepljenje uzoraka u drvene predloške A. Priprema uzoraka pomoću Core Microtoma B. Skeniranje uzoraka na sustavu ATRICS C.
2.2.7. Izmjera širina godova
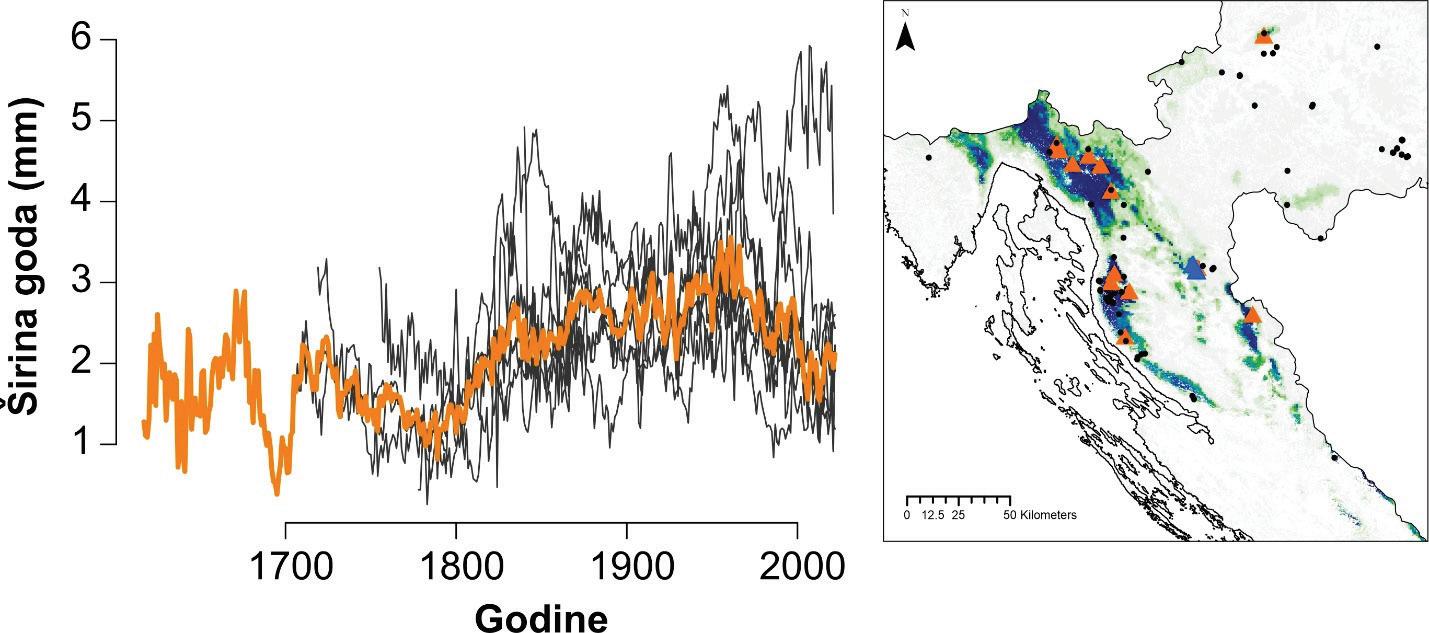
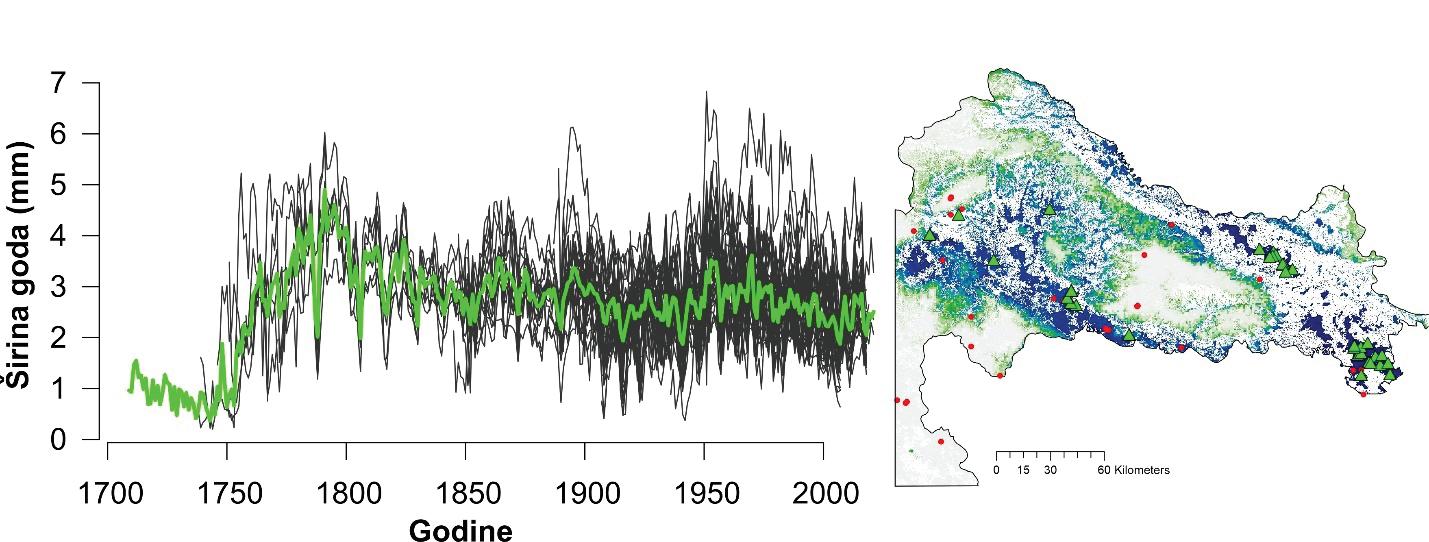
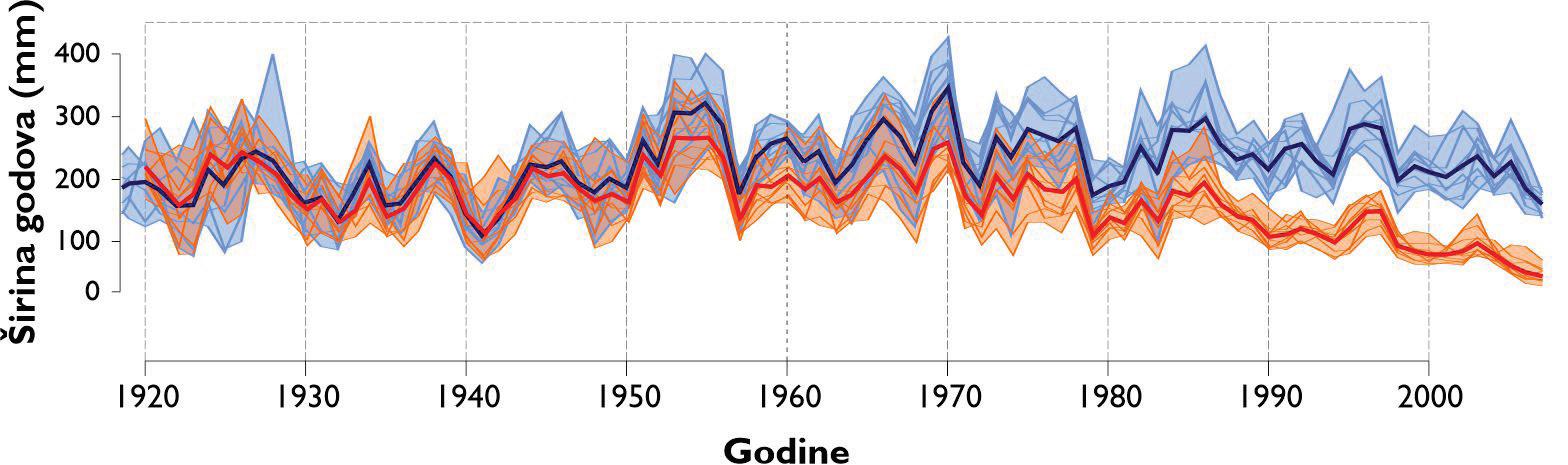
Izmjera širina godova se provodi u programu CooRecorder na prethodno snimljenim uzorcima. Izmjera se provodi digitalno a podaci o širinama godova se spremaju u tabelarni format s vrijednostima širine goda u 1000/mm i godinom u kojoj je nastao god počevši od godine uzorkovanja ili prethodne ukoliko god nije cijeli, odnosno ako stablo još nije završilo rast za tekuću godinu. Takvi podaci se spremaju u standardni format ekstenzije .rwl te kao takvi služe za daljnju statističku obradu i analizu. Očitane širine godova se u naknadnoj analizi uprosječuju na razini jednog stabla, pokusne plohe i vrste drveta te se na taj način stvara krivulja radijalnog prirasta odnosno širina godova (slika 9.)
Slika 5. Izgled digitaliziranih uzoraka obične bukve (gore) i obične jele (dolje) s izmjerenim vrijednostima širina godova
2.2.8. Analiza i obrada uzoraka
Unakrsno datiranje izmjerenih serija širina godova napravljeno je vizualno uz statističku provjeru kvalitete datacije pomoću programa COFECHA (Holmes, 1983). Prije datiranja serije širina godova s istih stabala su uprosječene u jednu individualnu seriju. Kvalitetno datirane serije širina godova su detrednirane kako bi se uklonio efekt starosti stabala te drugih šumova koji nisu direktno povezani s klimom (npr. growth releases). Standardizacija je provedena pomoću programa ARSTAN (Cook 1985) koristeći funkciju spline (50% prekidom frekvencije) na 32 godine na prethodno stabiliziranim serijama koristeći power transformaciju kako bi se producirao homoskedastički indeks (Cook & Peters, 1997). Reziduali izmjerenih vrijednosti i detrendiranih funkcija su izračunati kao razlika (Helama, Lindholm, Timonen, & Eronen, 2004). Za svaki lokalitet rezidualne vrijednosti serija širina godova su uprosječene u individualnu kronologiju koristeći biweight robust mean (Cook, 1985). U naknadnim analizama klimatske osjetljivosti korištena je ARSTAN kronologija. Analize klimatske osjetljivosti napravljena je pomoću jednostavnih (Persons) korelacija između indeksnih kronologija (RWI) i mjesečnih vrijednosti klimatskih čimbenika za razdoblje od 19 mjeseci (od lipnja prethodne godine do listopada tekuće godine) za razdoblje od 1950 – 2010. godine. Sezonske korelacije (prosjek temperatura zraka i suma oborina za razdoblje od nekoliko uzastopnih mjeseci: 2, 3 i 4 mjeseca) napravljena je pomoću seascorr funkcije u paketu treeclim (Zang 2015).

ŠUMARSTVO 15
3. REZULTATI
3.1. Obična jela (Abies alba Mill.)
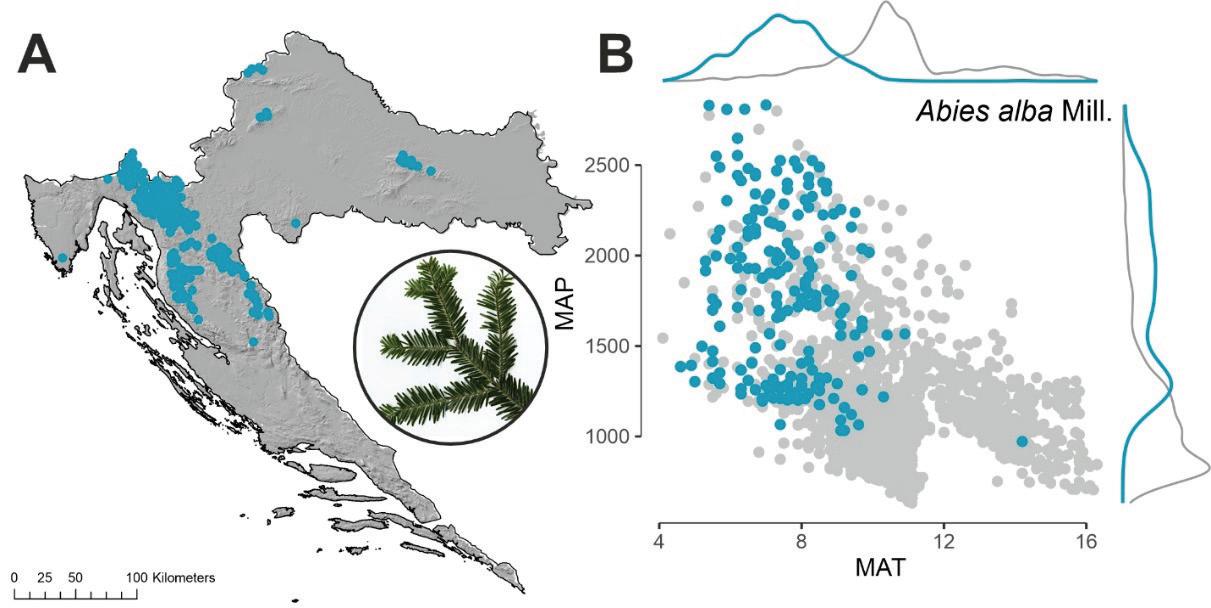
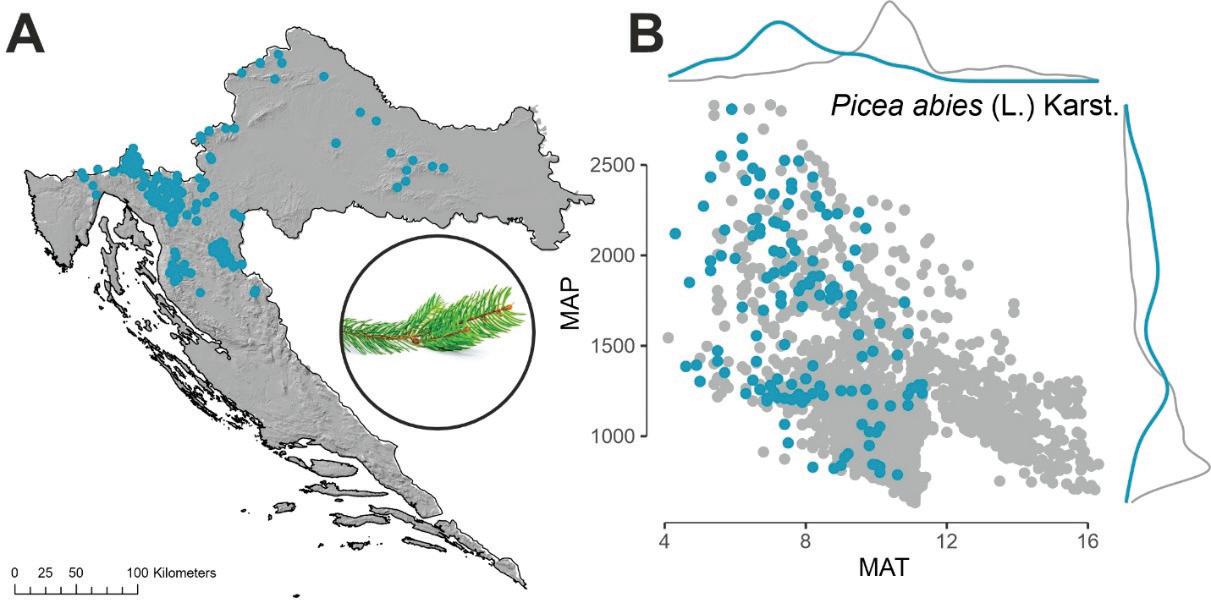
Obična jela (Abies alba Mill.) pridolazi u području koje u klimatskom smislu karakteriziraju prosječne godišnje temperature zraka niže od 12ºC (optimum oko 8 ºC). Što se tiče oborina, jela pridolazi u širokom rasponu od 1000 pa sve do preko 2500 mm godišnje (gorski vegetacijski pojasevi dinarskoga gorja te manjim djelom na najvišim predjelima kontinentalnih brda).
Slika 6. Prostorni položaj nalazišta obične jele u RH (Abies alba Mill.) A. Odnos između prosječne temperature zraka MAT (ºC) i ukupne količine oborina MAP (mm) za plohe na kojima se pojavljuje jela (plave točke) u odnosu na plohe gdje nema jele (sive točke) B.
Tijekom posljednjih nekoliko desetaka godina sastojine u čijem sastavu pridolazi obična jela pretrpjele su značajna i brojna oštećenja kao posljedice klimatskih promjena. U veljači 2014. godine na širem području Gorskog kotara zabilježena je ekstremna ledena kiša koja je prouzročila štetu od oko 200.000 m3 obične jele i smreke od čega je polovina stabala prelomljena a oko 15% izvaljeno. Nakon ledene kiše uslijedio je napad potkornjaka 2017. godine, a 2018. godine i snažno orkansko nevrijeme koje je oštetilo oko 1.200.000 m3 obične jele.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 16
Rezultati analize prikladnosti staništa za običnu jelu ukazuju na dobru do izvrsnu predikciju prema vrijednostima TOC-a te odličnu prema vrijednostima ROC-a. Čimbenici koji najviše pridonose modelu su: referentna evaporacija (Eref) >40%, suma oborina u ljeto (PPT_sm) oko 22%, oborine u zimu (PPT_wt) i kontinetalnost klime (TD) oko 16% dok je postotni sadržaj pijeska u tlu oko 3%.
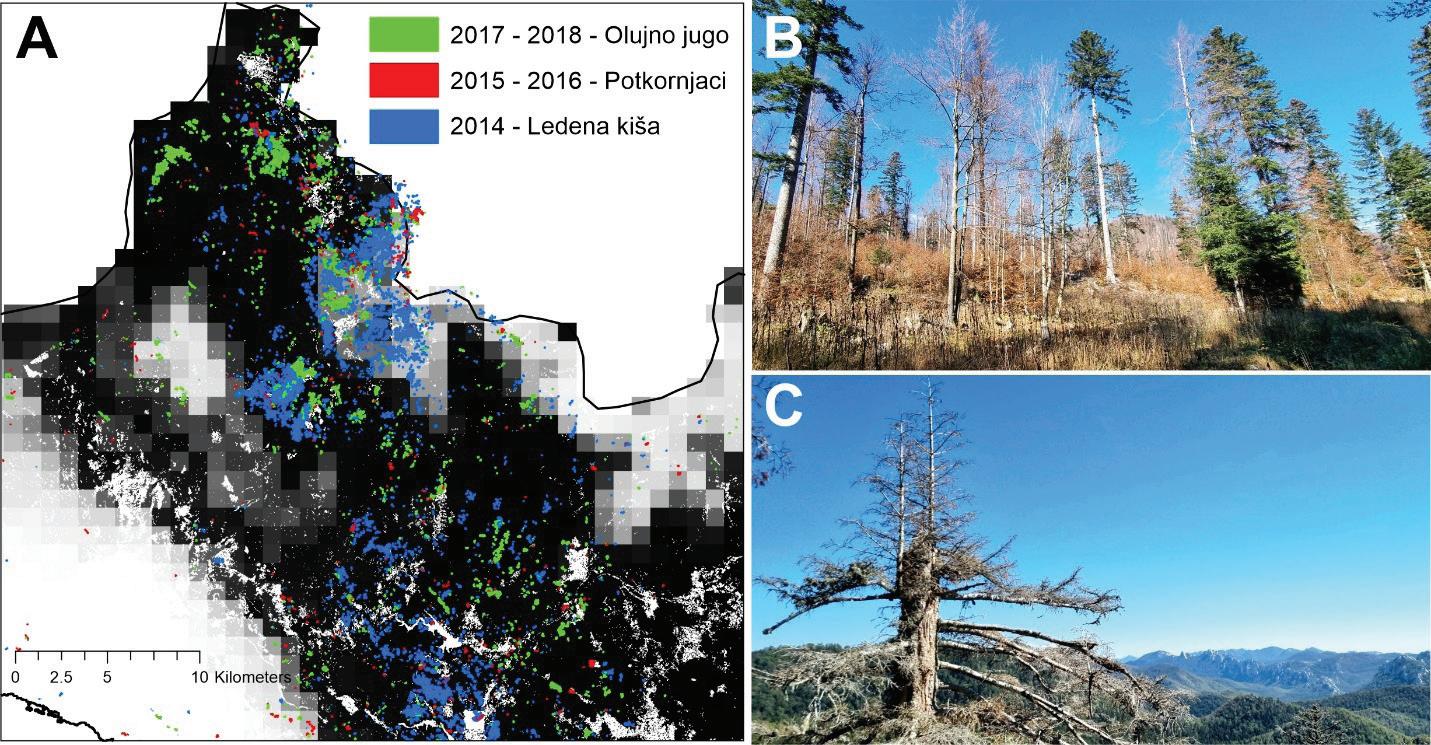
ŠUMARSTVO 17
Slika 7. Prostorni položaj šteta u posljednjih 10-ak godina na području Gorskog kotara prema tipovima prirodnih i biotskih nepogoda A. Sastojina koja je značajno oštećena ledenom kišom (stanje 2022. godine) B. Pojedinačna sušenja stabala jele na području srednjeg Velebita C.
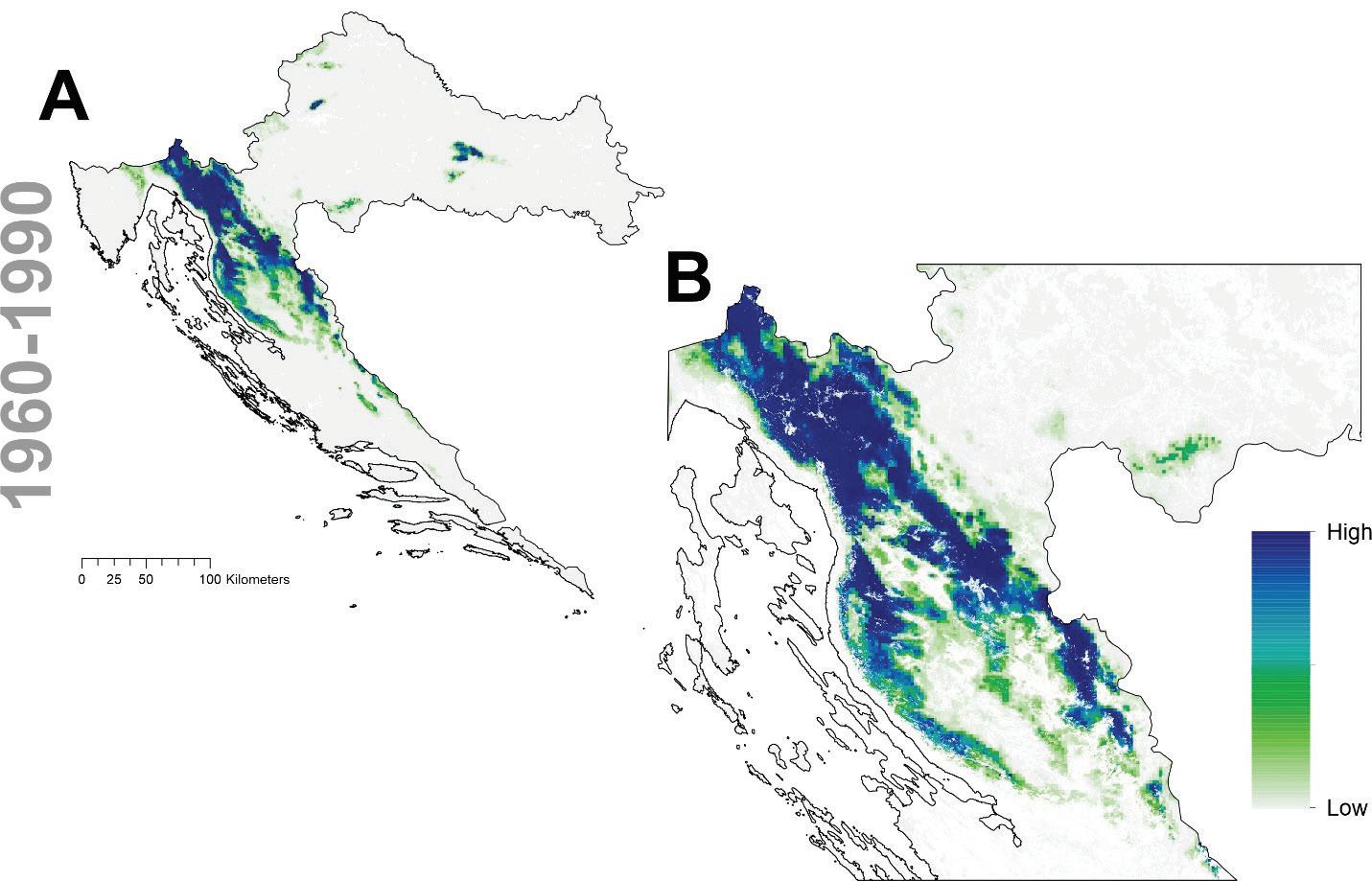
Slika 8. Projekcija trenutne pogodnosti staništa obične jelu u Hrvatskoj A te uvećano za područje dinarske Hrvatske B
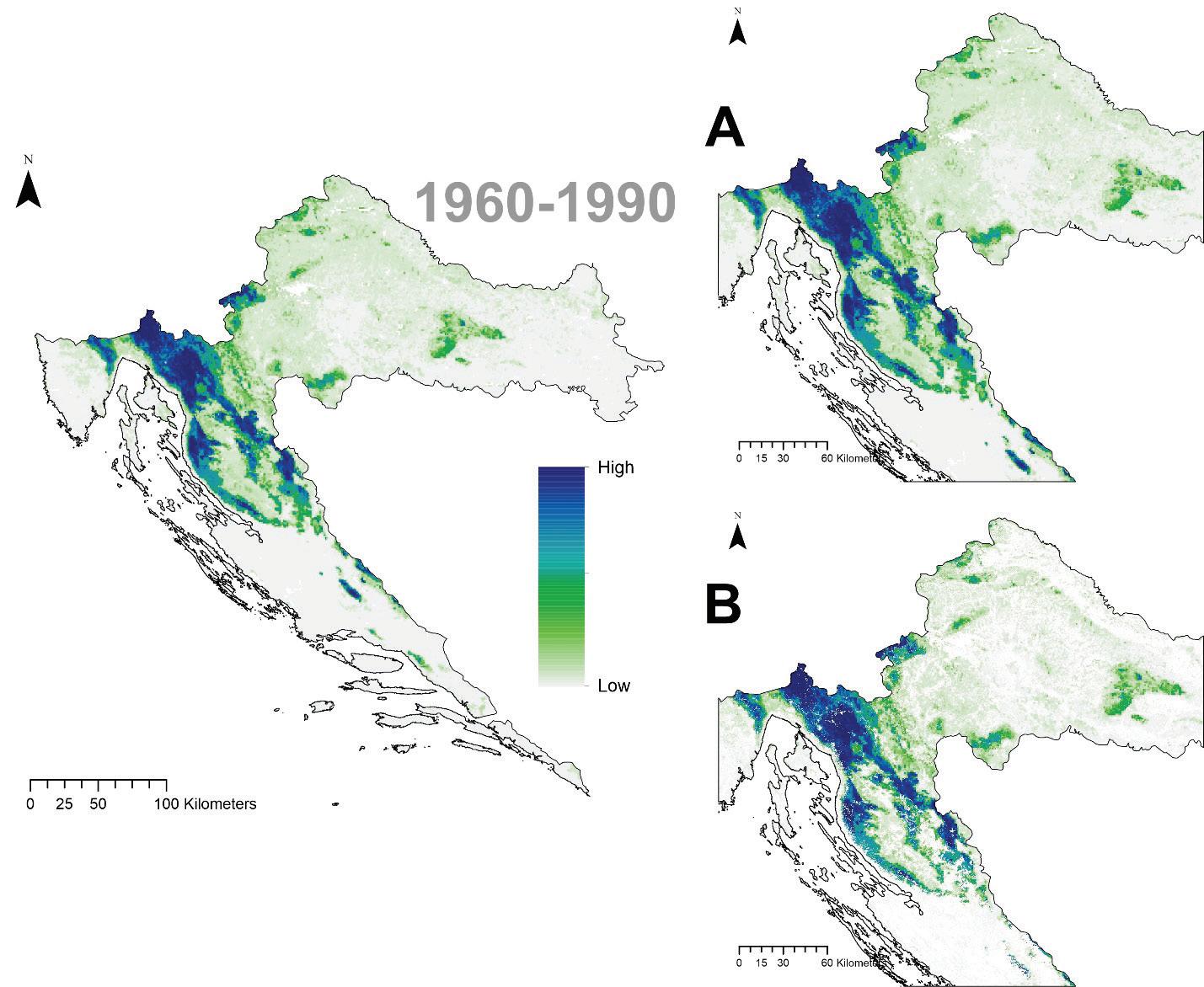
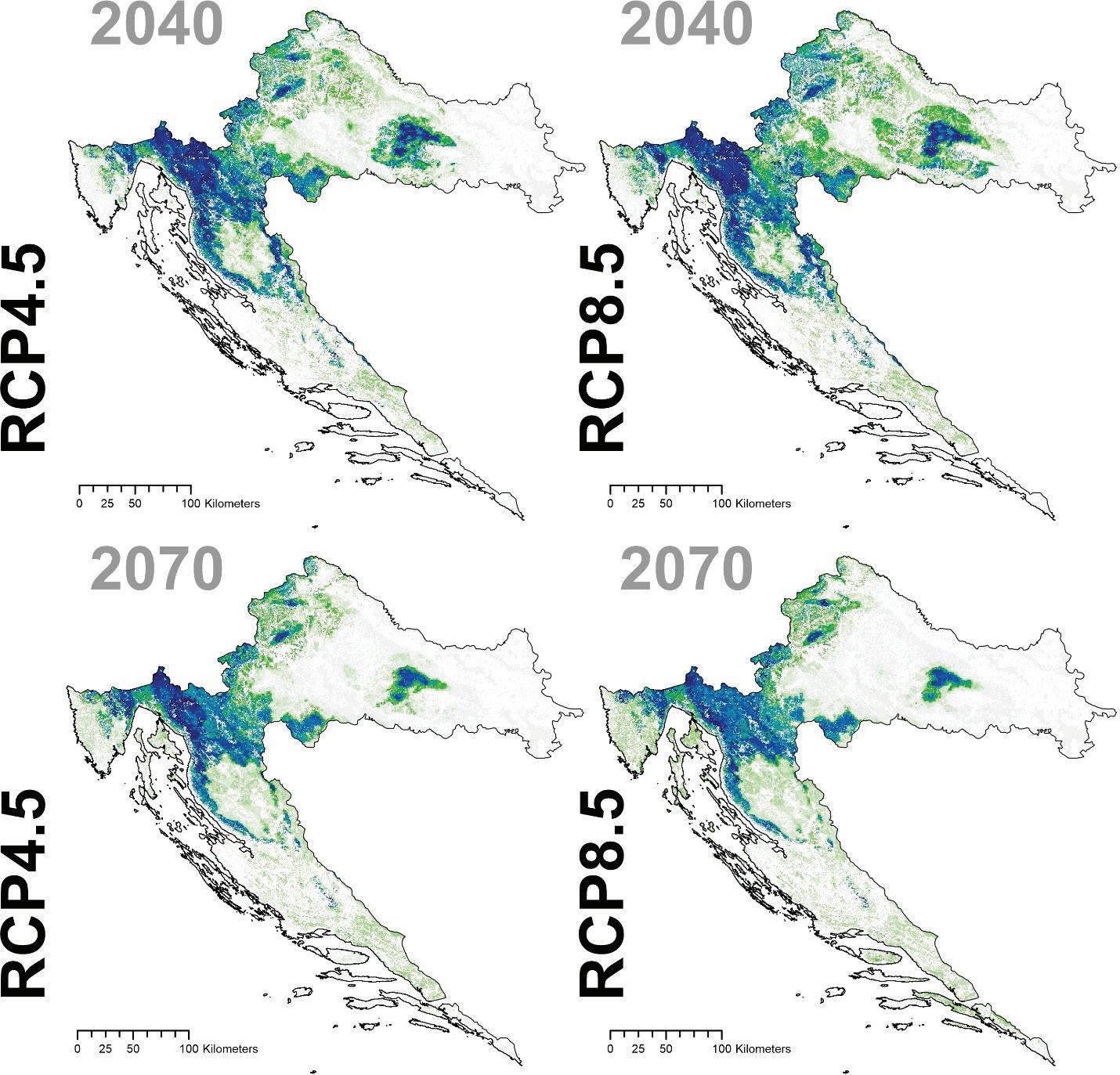
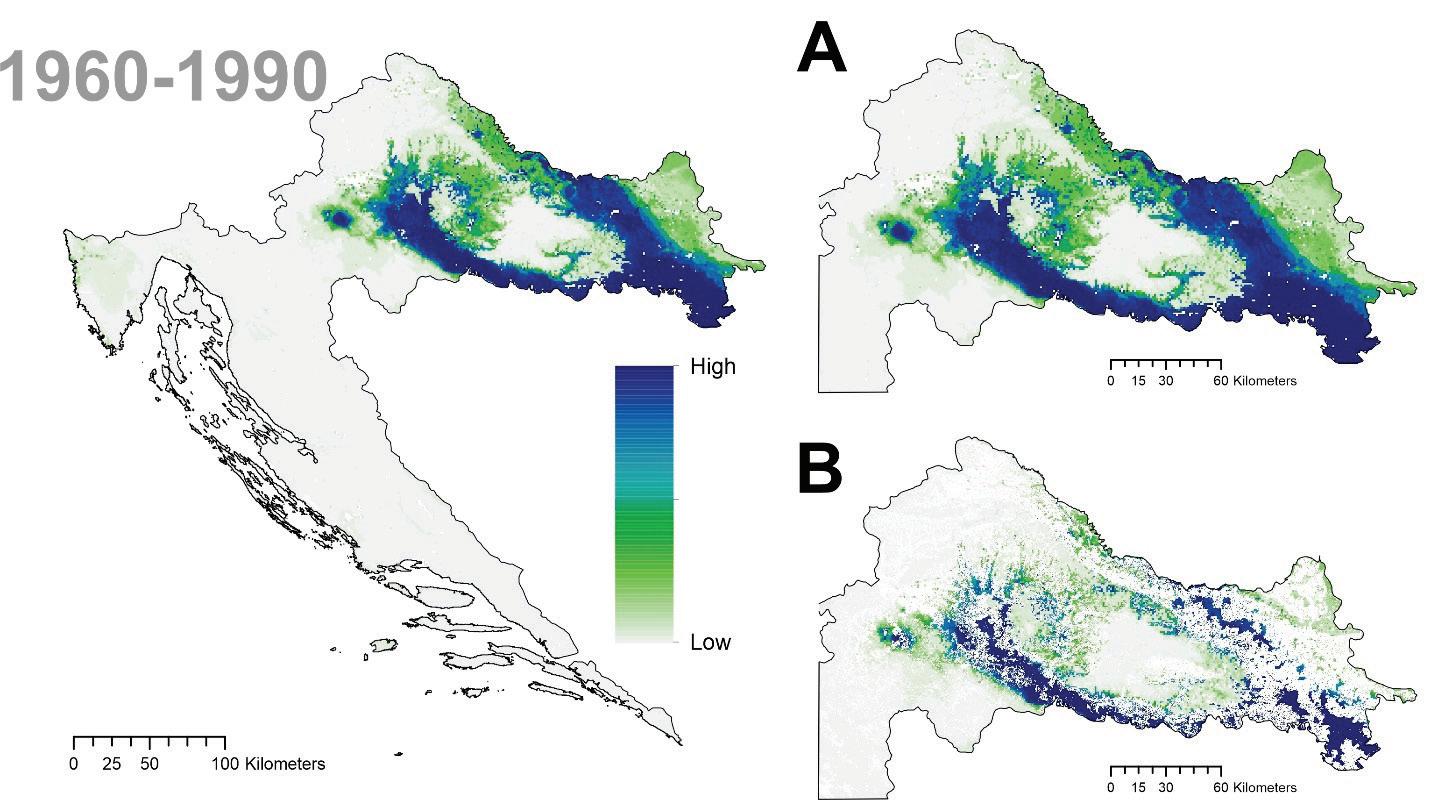
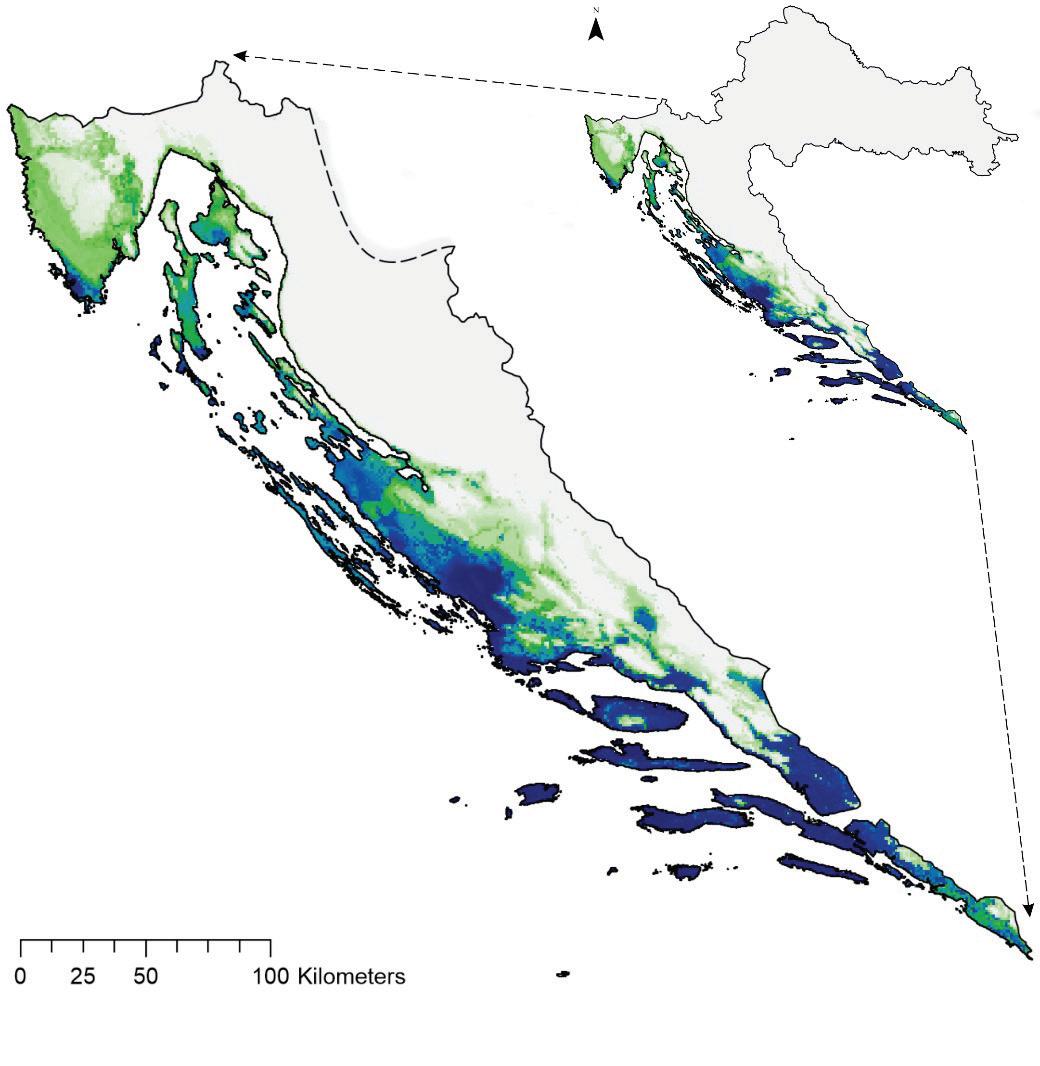
Prema očekivanim modelima klimatskih promjena združeni model predviđa brzu promjenu pogodnog staništa za običnu jelu i to prvenstveno na istočnom dijelu njenog trenutnog areala. Drugim riječima područja Male i Velike Kapele ali i dio Gorskog kotara na nižim nadmorskim visinama neće biti pogodni za rast i razvoj šuma u kojima danas pridolazi obična jela. Trenutačno nije jasno koliko dugo jela još može dobro rasti na tim područjima unatoč tome što će uskoro postati neprikladna. Modelirana prostorna distribucija obične jele ima tendenciju podcjenjivanja široke ekološke amplitude vrste, iako se vrlo dobro uklapa u srž njezine trenutne distribucije. Također treba imati na umu da su predviđanja za sva klimatska razdoblja ograničena na trenutni raspon vrste. Ako je apsolutna vjerojatnost prisutnosti predviđena ispod 400 (0,4), stanište je označeno kao nepogodno u odnosu na osnovnu projekciju za razdoblje od 1961. do 1990.
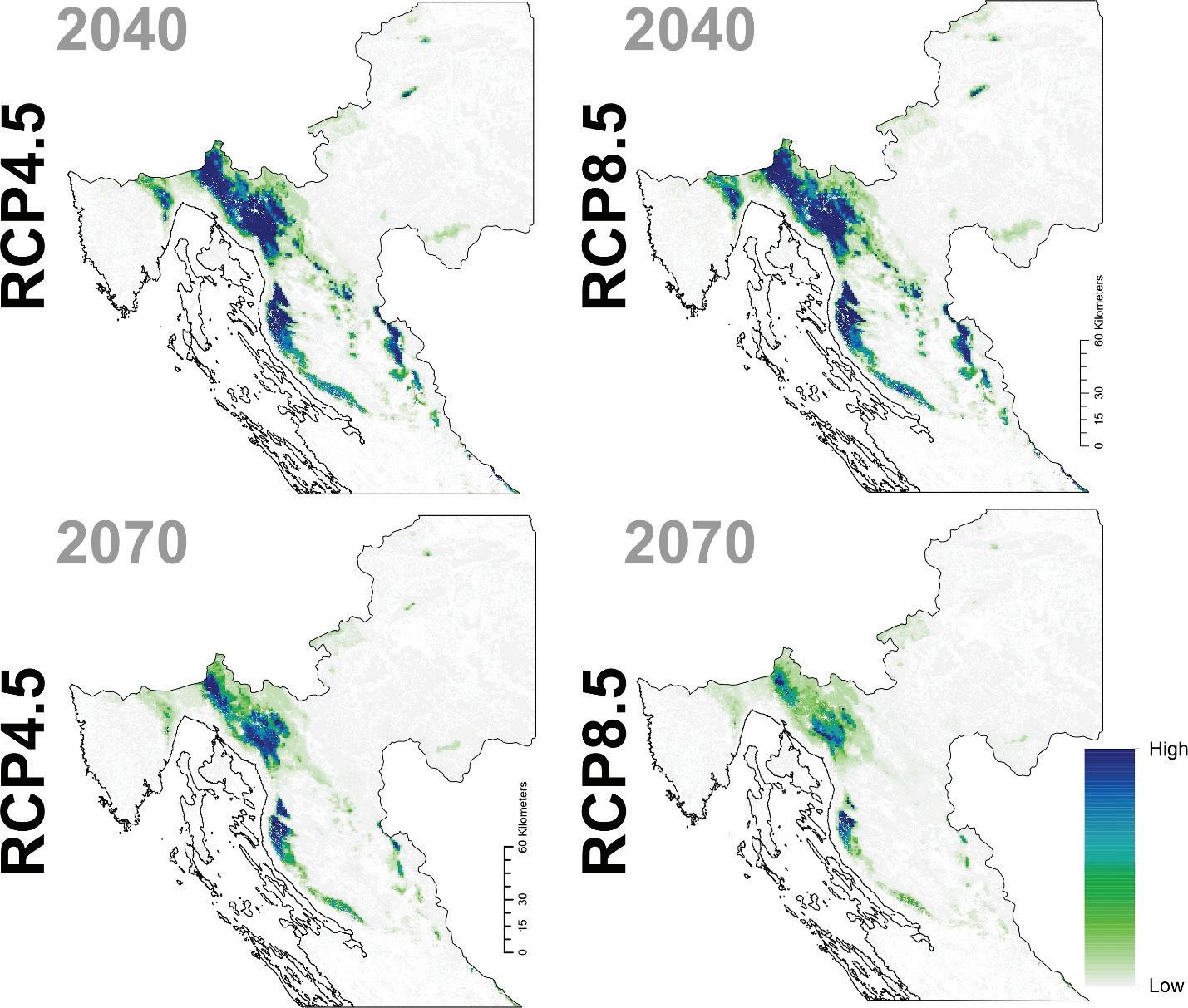
za običnu jelu u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (prvo od 2011. – 2040. godine te drugo od 2041. – 2070. godine. Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
Općenito je poznato da obična jela može tolerirati topliju klimu, pa čak i nešto sušu klimu. Analizom klimatske osjetljivosti pomoću uzoraka iz stabala za razdoblje od 1950. do 2009. godine utvrđena je značajna (P<0.05) pozitivna osjetljivost radijalnog prirasta na oborine u ljeto tekuće godine (godina u kojoj je nastao god) i to na ukupnu količinu oborina od mjeseca svibnja do srpnja (MJJ). Korelacije se kreću u rasponu od 0.06 do 0.43. Također je utvrđena i značajna pozitivna osjetljivost (P<0.05) na prosječnu temperaturu zraka u zimi (prosjek za mjesece siječanj, veljaču i ožujak, JFM).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 18
Slika 9. Buduće projekcije pogodnosti staništa
Slika
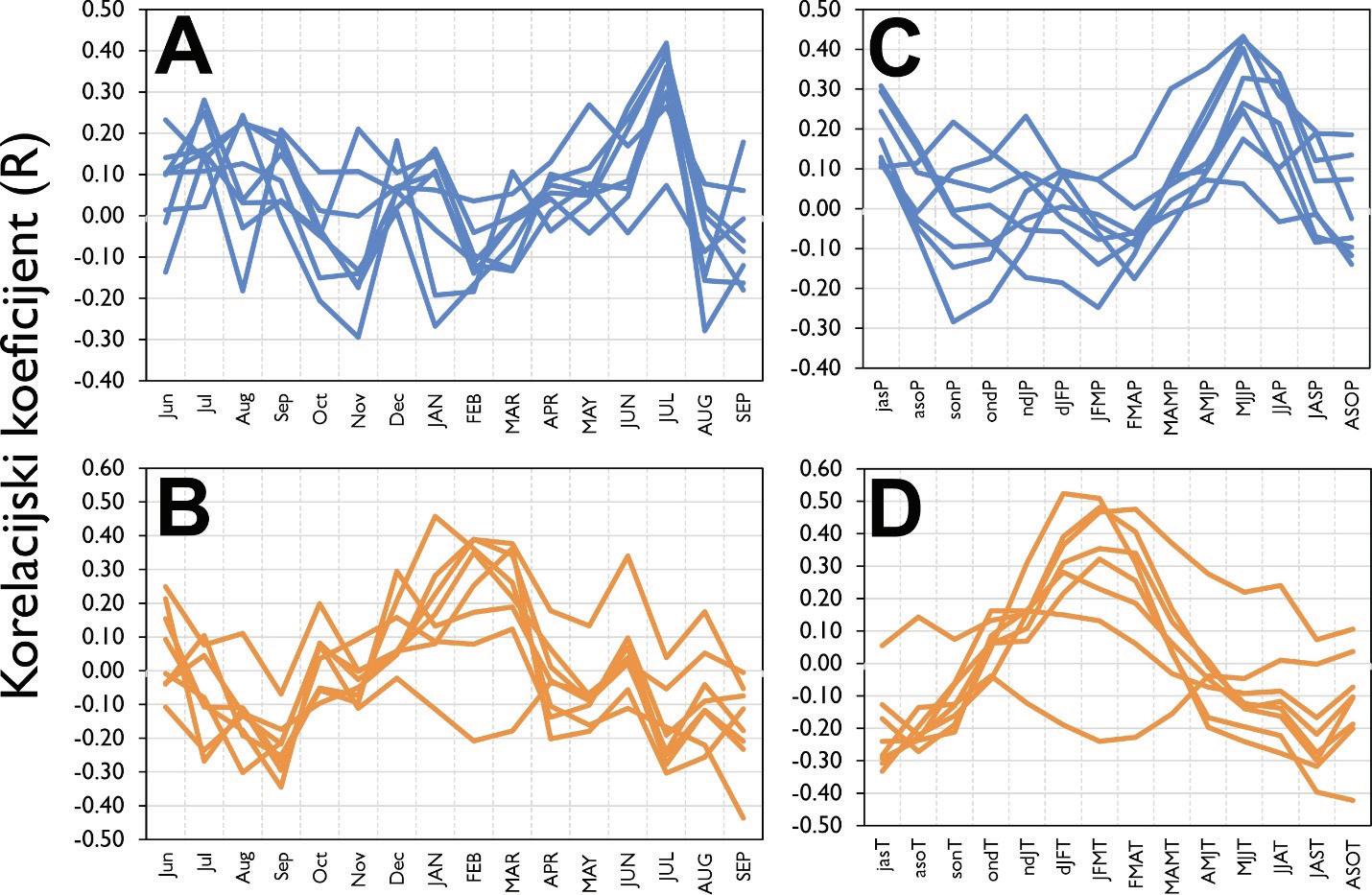
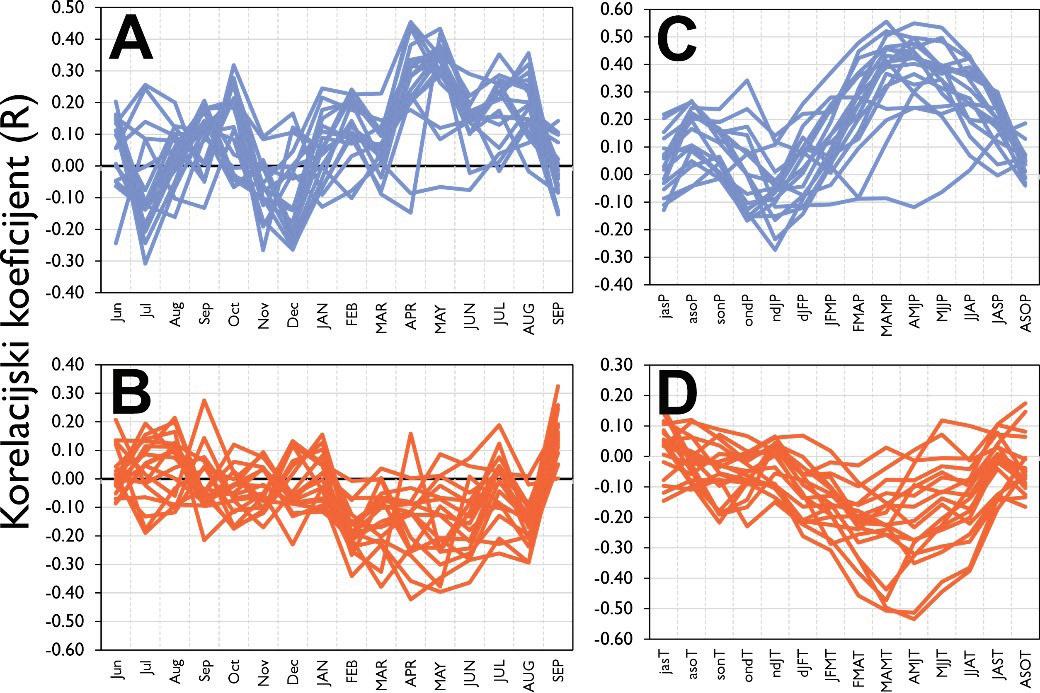
kronologija širina godova obične jele na istraživanim plohama. Prostorni položaj pokusnih ploha (narančasti trokuti). Vrijednosti korelacijskih koeficijenata između mjesečnih vrijednosti klimatskih čimbenika i to: A – oborine (mm) po mjesecima, C - suma oborina za 3 uzastopna mjeseca, B – temperature zraka i D -prosjek temperatura zraka za tri uzastopna mjeseca.
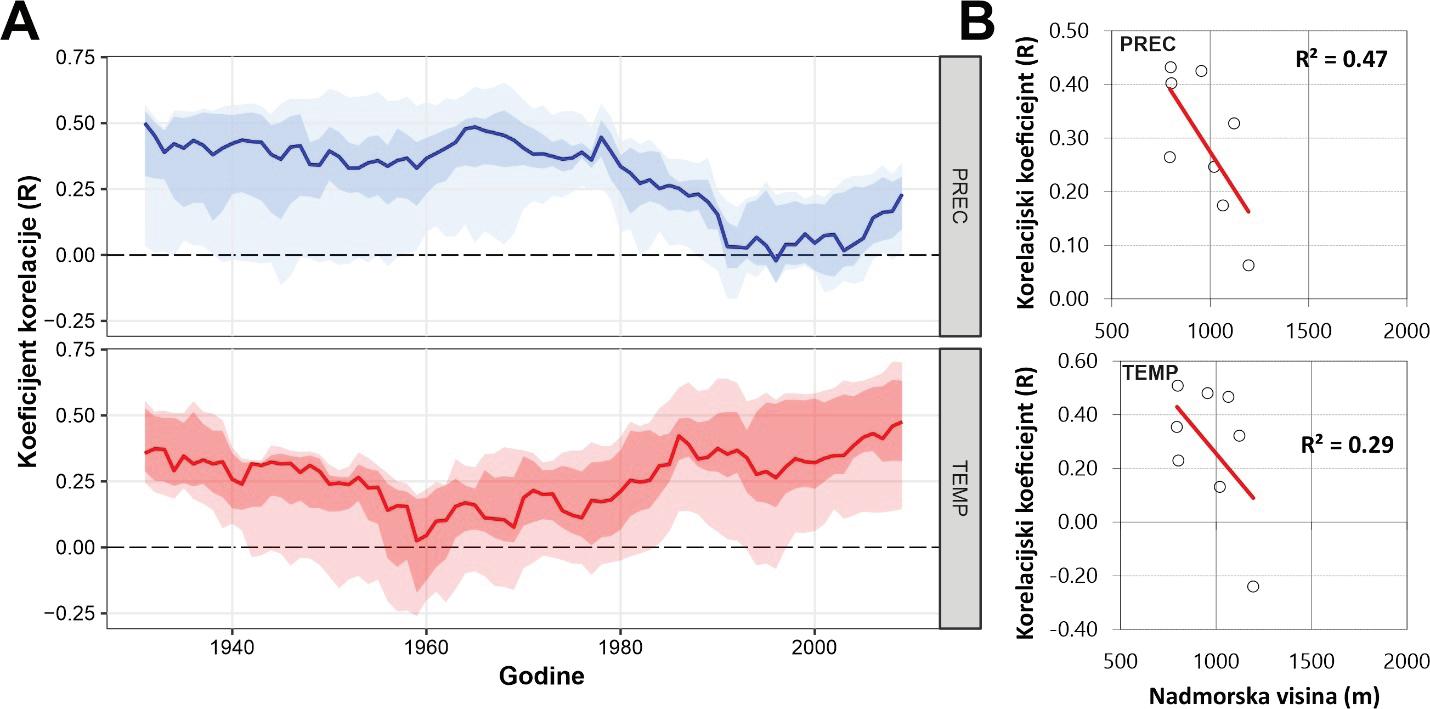
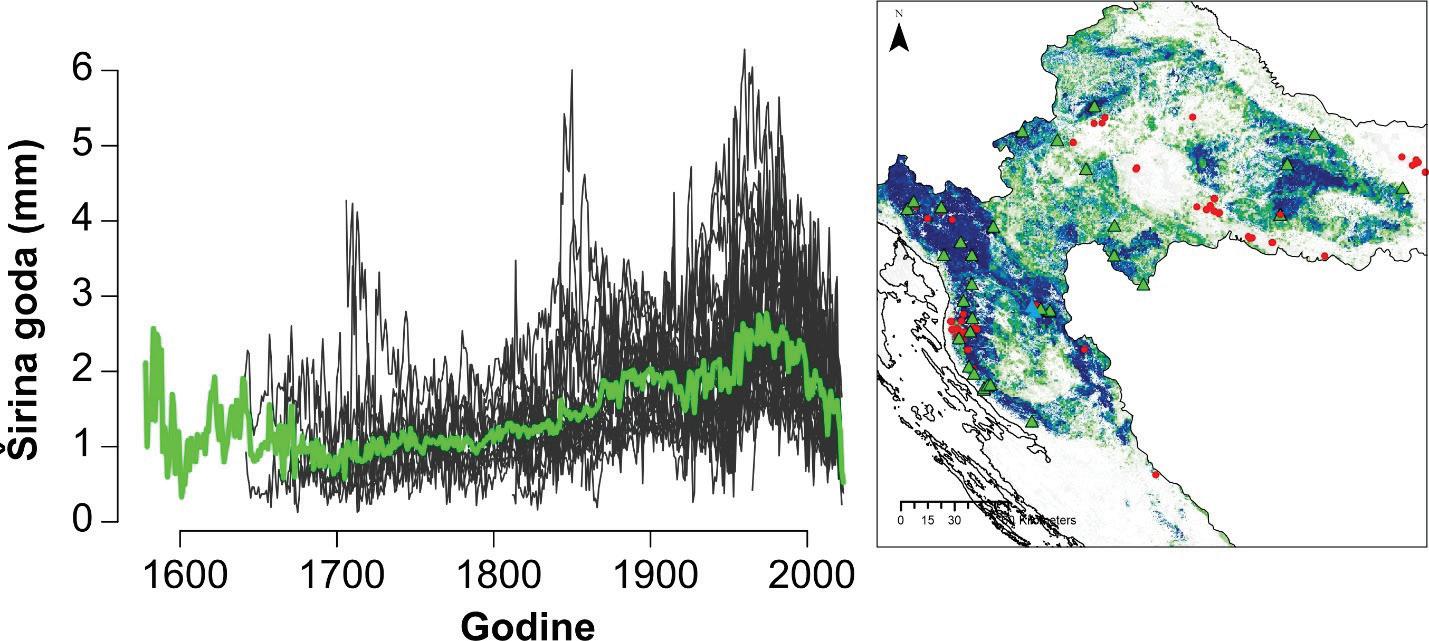
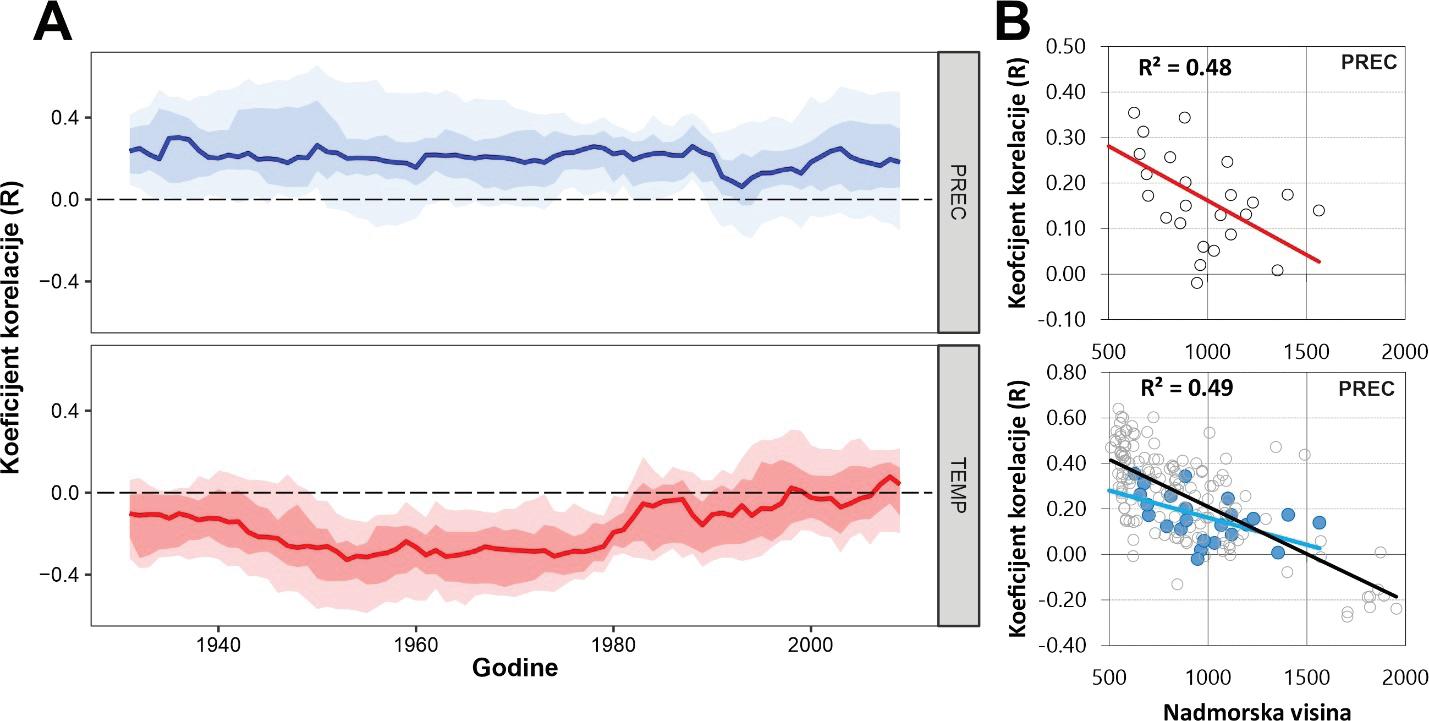
Analiza pomičnih korelacija za period od 30. godina i razdoblje od 1900 – 2010. godine ukazuje da se klimatska osjetljivost jele značajno mijenja tijekom promatranog razdoblja. Odnosno oborine u ljeto tekuće godine nakon 1980. godine nisu više značajne za rast stabala jele gotovo na svim lokalitetima. Za razliku od oborina, jela postaje sve osjetljivija na prosječnu temperaturu zraka od siječnja do ožujka tekuće godine. Usporedbom klimatske osjetljivosti prema nadmorskim visinama utvrđeno je da su populacije jele koje rastu na nižim nadmorskim visinama značajno ovisne o ljetnim oborinama u odnosu na populacije koje rastu na višim nadmorskim visinama kao i na temperaturu zraka zimi (Slika 11).
ŠUMARSTVO 19
10. Prosječne vrijednosti
Slika 11. Pomične korelacije za duljinu perioda od 30 godina te razdoblje od 1900. – 2010. godine za oborine u ljeto –od svibnja do srpnja (MJJ) i temperaturu zraka zimi – prosjek od siječnja do ožujka (JFM).
3.2. Obična smreka (Picea abies Karst.)
U Hrvatskoj obična smreka zauzima površinu od 16,670 ha odnosno 1% površine šuma. Ukupna drvna zaliha iznosi oko 3 milijuna m3 (Oršanić i dr. 2008). Smreka prirodno pridolazi u subalpskom vegetacijskom pojasu na nadmorskim visinama od 1200 – 1600 m gdje tvori čiste sastojine. Osim toga pojavljuje se u manjem udjelu do 5% u pojasu bukovo-jelovih šuma kao prateća vrsta (Matić 2010). Na području sjevernoga Velebita areal smreke je disjunktan na površini od 2200 ha unutar klimata bukve (Vukelić i Rukavina 2005). Njen pridolazak je uvjetovan specifičnim orografskim prilikama krškog reljefa koji modificira klimatske prilike. Smreka uglavnom zauzima hladnije i vlažnije vrtače, udoline i mrazišta. U takvim reljefno uvjetovanim klimatskim inverzijama smreka nadoknađuje nedostatak vlage i oborina deficitarnih na dinarskome kršu. U ekološkom smislu smreka dobro podnosi niske temperature zraka i mraz. Ne podnosi visoke temperature zraka i niske količine oborina. Što se tiče makroklimatskih prilika za područje Hrvatske obuhvaća identično područje kao i obična jela s rasponom temperatura zraka od 4 do 12 ºC te širokim rasponom oborina od 1000 mm pa sve do preko 2500 mm godišnje (Slika 12).
nalazišta obične smreke u RH (Picea abies Karst.) A. Odnos između prosječne temperature zraka MAT (ºC) i ukupne količine oborina MAP (mm) za plohe na kojima se pojavljuje smreka (plave točke) u odnosu na plohe gdje nema smreke (sive točke) B.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 20
Slika 12. Prostorni položaj
Pogoršano zdravstveno stanje i odumiranje smreke je prisutno unazad nekoliko godina u cijeloj Europi (Kucbel i dr. 2009). Sastojine obične smreke su na području sjevernoga Velebita posljednjih nekoliko godina pretrpjela značajnija sušenja. Odumiranje šuma u planinskim područjima zabilježeno je u čitavoj Europi. Mnogo je potencijalnih i uzajamno povezanih razloga za to. Kumulativni utjecaj nekoliko čimbenika kao što su klimatske promjene, zagađenje zraka te dugovječno preferiranje smreke na nekom staništu rezultiralo je promjenama u kruženju hranjiva, akumulaciji listinca te acidifikaciji tla. Te procese opisali su podrobno Fanta (1997) i Emmer i dr. (1998, 2000) te ih pri tome nazvali antropogenom borelizacijom. Prema istraživanjima Robertsa i dr. (1989) jedan od razloga odumiranja smreke u Srednjoj Europi je folijarni deficit magnezija (Mg). On rezultira klorozom starijih iglica te tako djeluje na redukciju fotosinteze i translokaciju ugljikohidrata. Prema Maueru i dr. (2008) visoke temperature u kombinaciji s nedostatkom oborina su predisponirajući čimbenici sušenju smreke. Oslabljena stabla tada su predisponirana napadu mednjače (Armillara mellea). Temperatura zraka i oborine važan su čimbenik koji utječe na rast i razvoj stabala. U ekstremnim uvjetima, kao što su planinska područja klima je glavni čimbenik koji limitira rast stabala (Fritts 1976). Prema Savva i dr. (2006) temperatura zraka je glavni ekološki čimbenik koji utječe na rast stabala u planinskim predjelima, dok oborine imaju značajniji utjecaj na rast na nižim nadmorskim visinama. Utjecaj temperature na rast stabala je najznačajniji na početku vegetacijske sezone kada niske temperature mogu produljiti početak stvaranja kambija (Fritts 1976).

ŠUMARSTVO 21
Slika 13. Izgled sastojina obuhvaćenih sušenjem i propadanjem tijekom posljednjih 15-ak godina.
Visoke temperature zraka tijekom vegetacijskog razdoblja uglavnom utječu pozitivno na rast stabla. Ipak ako su previsoke mogu utjecati na smanjenje bilance ugljika što za posljedicu ima opadanje prirasta stabla (Čermák 2007). Prema Rybníčeku i dr. (2009) proljetne i zimske oborine prethodne godine, te oborine u proljeće i ljeto tekuće godine su od izuzetne važnosti za rast stabala. Kao i jela obična smreka je značajno ugrožena recentnim promjenama klime i jedina vrsta koja je u relativnom udjelu u posljednjih nekoliko godina pretrpjela enormne štete. Od 2001. do 2021. godine ukupno je evidentirano oko 1.600.000 m3 štete kao izravna posljedica klimatskih promjena diljem Hrvatske što iznosi oko 30% ukupne drvne zalihe ove vrste u nas.
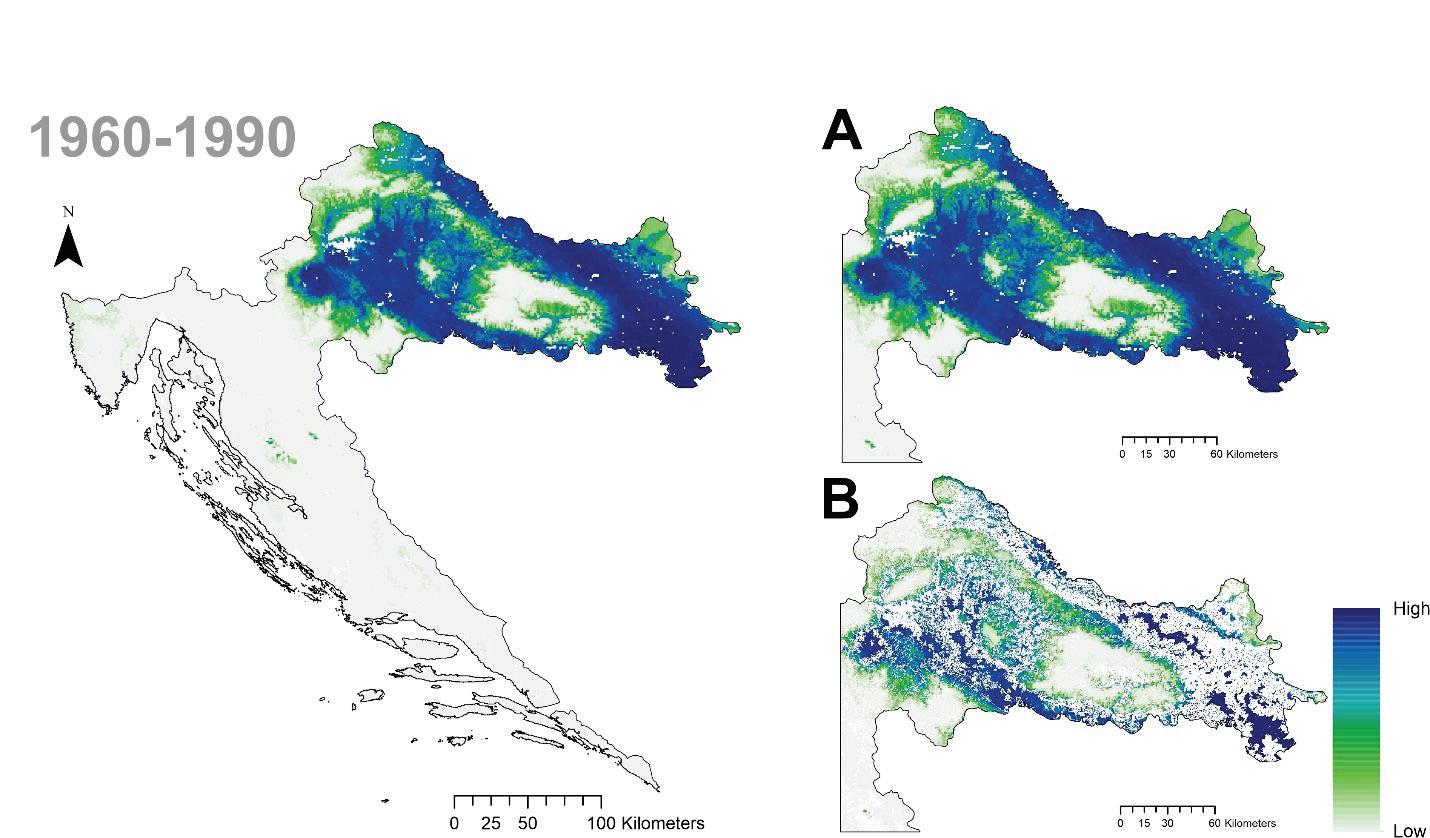
Model trenutne pogodnosti staništa za običnu smreku se vrlo dobro poklapa s njenom stvarnom rasprostranjenosti. Čimbenici koji najviše pridonose modelu pogodnosti staništa su: ukupna količina oborina u ljeto (PPT_sm), evaporacija (Eref) i kapacitet tla za vodu (PAW).
Slika 14. Projekcija prostorne razdiobe trenutne pogodnosti staništa obične smreke u Hrvatskoj. Plavom bojom (High) označena su područja visoke pogodnosti staništa obzirom na promatrane okolišne čimbenike. A Potencijalna karta pogodnosti staništa jele te B stvarna karta pogodnosti staništa (bez nešumskih površina).
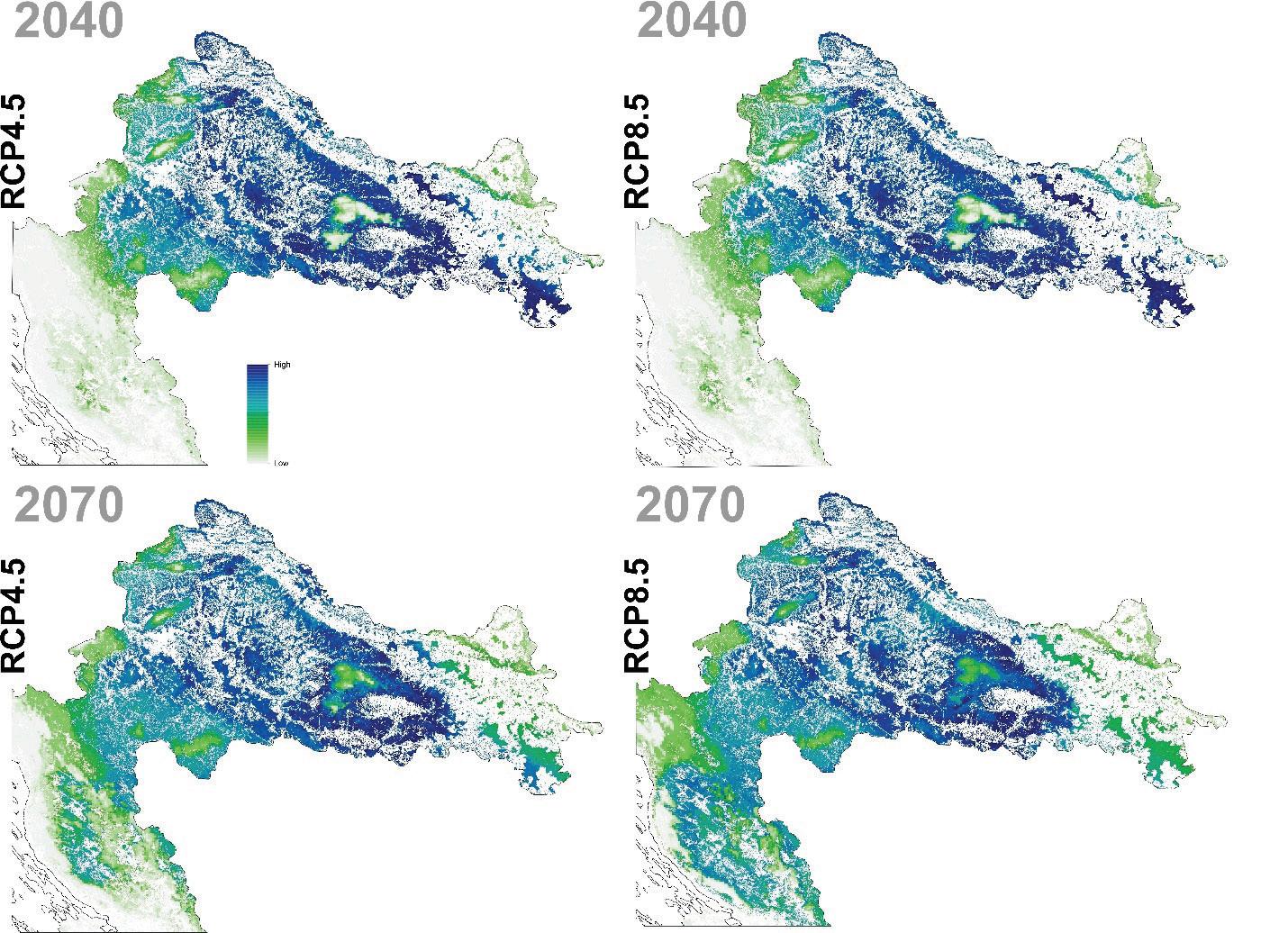
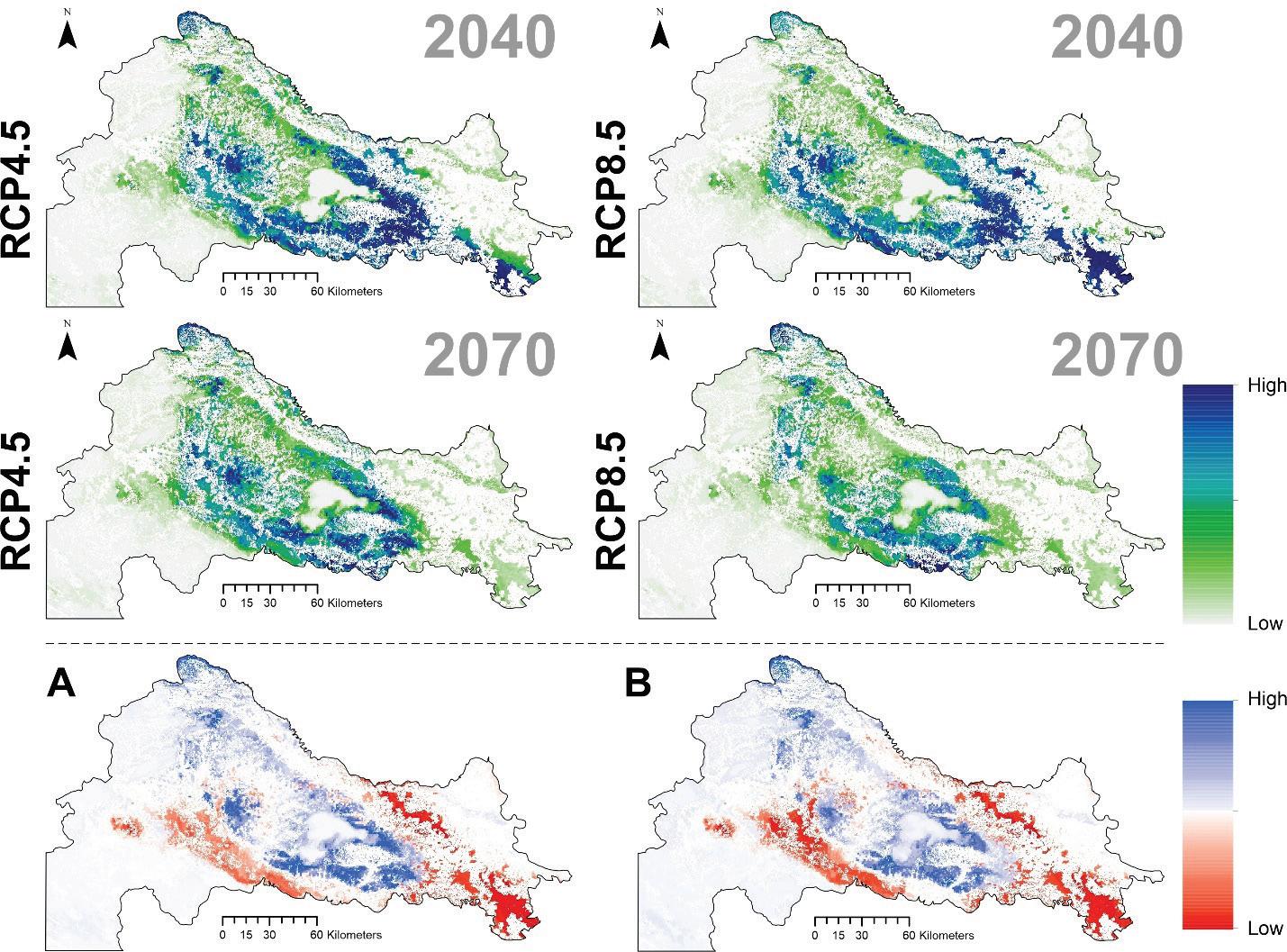
Buduće projekcije prikladnosti staništa ukazuju kao i kod jele na značajan gubitak staništa poglavito u istočnom dijelu Dinarida (Slika 15.) Područja na današnjem istočnom dijelu geografske rasprostranjenosti neće više biti pogodna za rast obične smreke dok područja Gorskog kotara i dijelovi Sjevernog Velebita i dalje ostaju potencijalno pogodni za rast obične smreke.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 22
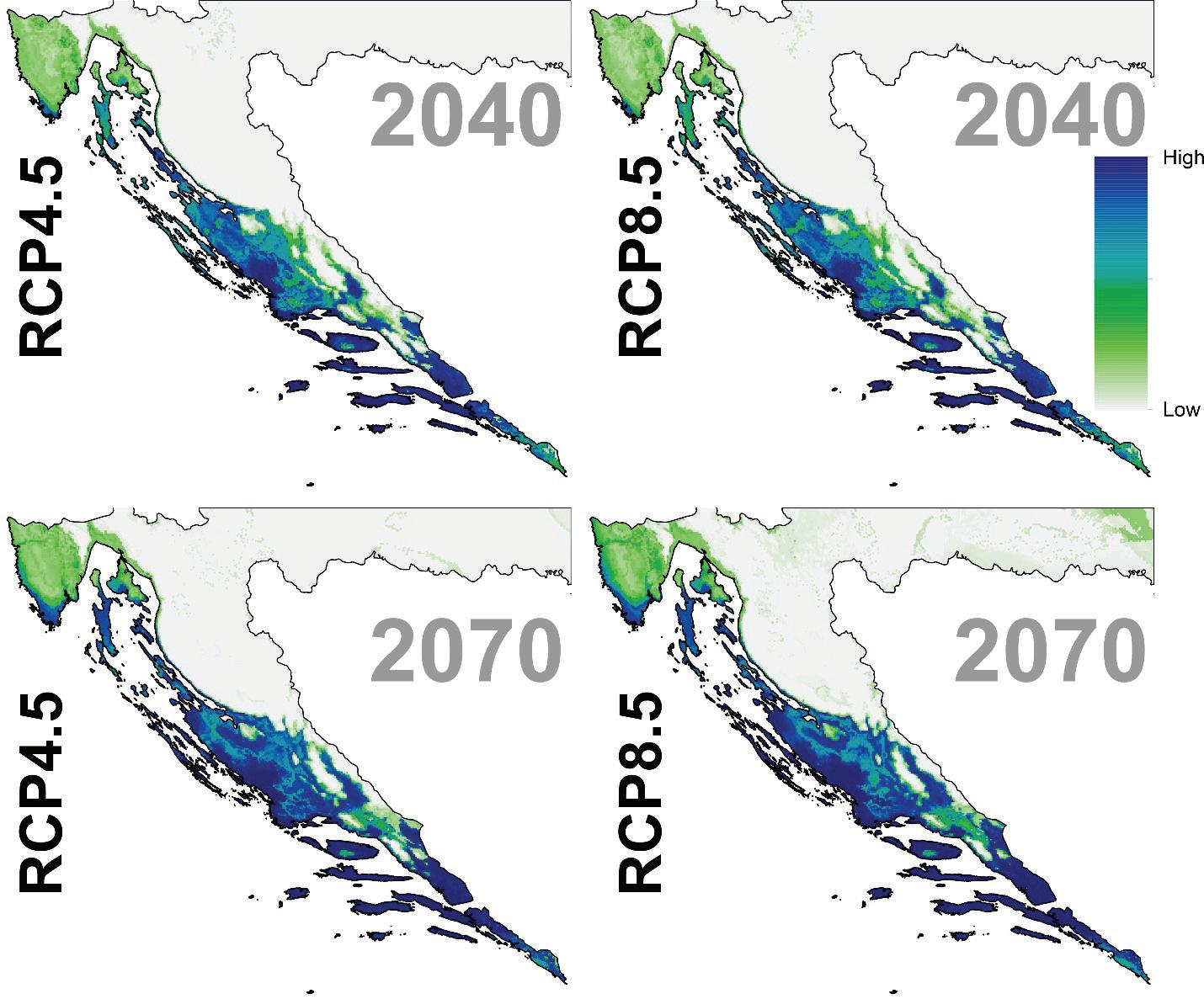
Slika 15. Buduće projekcije pogodnosti staništa za običnu smreku u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (prvo od 2011. – 2040. godine i drugo od 2041. – 2070. godine. Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
Analizom klimatske osjetljivosti kronologija smreke utvrđena je značajna pozitivna ovisnost radijalnog prirast o oborinama u srpnju tekuće godine dok je s druge strane radijalni prirast značajno limitiran visokim temperaturama zraka u ljeto prethodne godine (prosjek od srpnja do rujna).
ŠUMARSTVO 23
Slika 16. Prosječne vrijednosti kronologija širina godova obične smreke na istraživanim plohama. Prostorni položaj pokusnih ploha (narančasti trokuti). Vrijednosti korelacijskih koeficijenata između mjesečnih vrijednosti klimatskih čimbenika i to: A – oborine (mm) po mjesecima, C - suma oborina za 3 uzastopna mjeseca, B – temperature zraka i D -prosjek temperatura zraka za tri uzastopna mjeseca.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 24
3.3. Obična bukva (Fagus sylvatica L.)
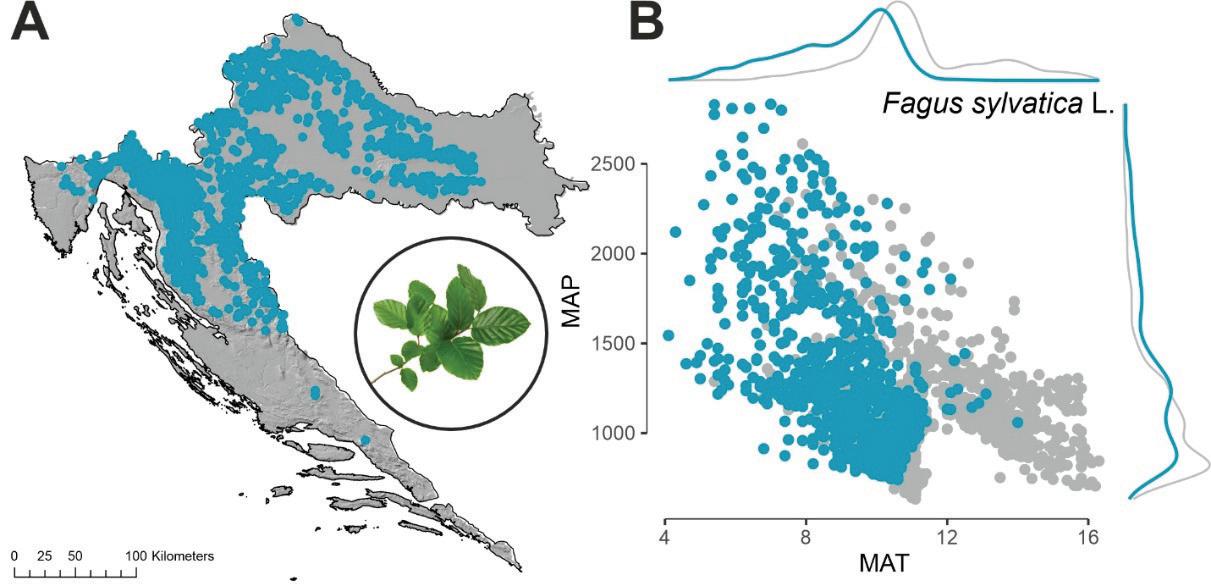
Obična bukva (Fagus sylvatica L.) je najzastupljenija vrsta drveća u Hrvatskoj. Ona zauzima oko 28% sveukupne površine šuma, a po drvnoj zalihi gotovo 40% od ukupne drvne zalihe svih vrsta drveća u Hrvatskoj. Bukva je vrsta široke ekološke valencije, pridolazi u gotovo svim vegetacijskim pojasevima od nizine (u primjesama sa hrastom lužnjakom) pa sve do subalpinskog vegetacijskog pojasa gdje ujedno tvori i gornju granicu šumske vegetacije. Obzirom na prosječnu temperaturu zraka i oborine bukva pokazuje širok raspon pridolaska (Slika 17).
Slika 17. Prostorni položaj nalazišta obične bukve u RH (Fagus sylvatica L.) A. Odnos između prosječne temperature zraka MAT (ºC) i ukupne količine oborina MAP (mm) za plohe na kojima se pojavljuje bukva (plave točke) u odnosu na plohe gdje nema bukve (sive točke) B

Obična bukva vrlo teško podnosi ekstremne klimatske uvjete jer ponik strada od mraza i suše što je česta pojava u prirodnom arealu bukovih šuma ili na rubu njihova areala, osobito na južnom dijelu (Harapin, 2003). Dosadašnje spoznaje o sušenju šuma obične bukve u Hrvatskoj i Europi govore u prilog negativnim klimatskim čimbenicima koji se smatraju uzročnikom velikog sušenja šuma na
ŠUMARSTVO 25
početku 20. stoljeća.
Slika 18. Odumiranje bukovih sastojina na Velebitu prouzrokovano sušom
U tom periodu bilo je više godina koje se navode kao ekstremno sušne, a obična bukva pokazuje izrazitu osjetljivost ako je njezina kora izravno izložena suncu gdje dolazi do pregrijavanja (suncožara) što uzrokuje odumiranje stabala (Slika 19B). Recentna istraživanja ukazuju na prevladavajući pad rasta stabala obične bukve u razdoblju od 1955. do 2016. godine diljem Europe. Ovo smanjenje je široko rasprostranjeno s iznimkom sjevernih područja u Danskoj, Norveškoj i Švedskoj kao i na višim nadmorskim visinama u planinskim područjima. Smanjenje prirasta kod obične bukve izraženije je pri istim stanišnim uvjetima nego kod ostalih vrsta drveća koje pridolaze u zajednici s običnom bukvom. Od sredine 20. stoljeća temperature su počele značajno rasti, a sušna razdoblja se pojavljivati već od proljeća (travanj-svibanj) kada je ujedno i najveća fiziološka aktivnost u biljkama (početak vegetacijskog razdoblja). Pad prirasta na području zapadne, središnje i jugoistočne Europe prvenstveno je povezan s negativnim učinkom visokih temperatura i dugotrajnih suša u prošlogodišnjem vegetacijskom razdoblju (Dulamsuren i dr., 2017). Također je potvrđena i smanjena kambijalna aktivnost obične bukve u razdobljima velike vlažnosti zraka uzrokovane visokim ljetnim temperaturama, a to znači smanjenje prirasta (Kocher i dr., 2012). U zadnjih šest desetljeća prirast obične bukve u Europi je u značajnom padu i većina stabala ove vrste danas raste sporije nego prije 60 godina. Najizraženiji pad bilježi se na području južne Europe gdje se stopa pada prirasta procjenjuje na oko 20 %. Negativni učinci povezani s klimom u budućnosti imaju tendenciju rasta što će se u narednih tridesetak godina još značajnije odraziti na smanjenje prirasta obične bukve s naglaskom na područje južne Europe gdje se očekuje pad prirasta i preko 50% (Martinez del Castillo i dr., 2022). Obična bukva, iako vrlo konkurentna i superiorna u odnosu na druge vrste drveća s kojima pridolazi u zajednicama, vrlo je osjetljiva na klimatske ekstreme, a konstantan pad prirasta promatran desetljećima unazad najbolji je pokazatelj njezine ranjivosti.

Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 26
Slika 19. Subalpinske sastojine obične bukve na gornjoj granici vegetacije A. Sušenje bukve kao posljedica ekstremno visokih temperatura i suše 2022.godine B i sastojine obične bukve u enklavama Biokova C
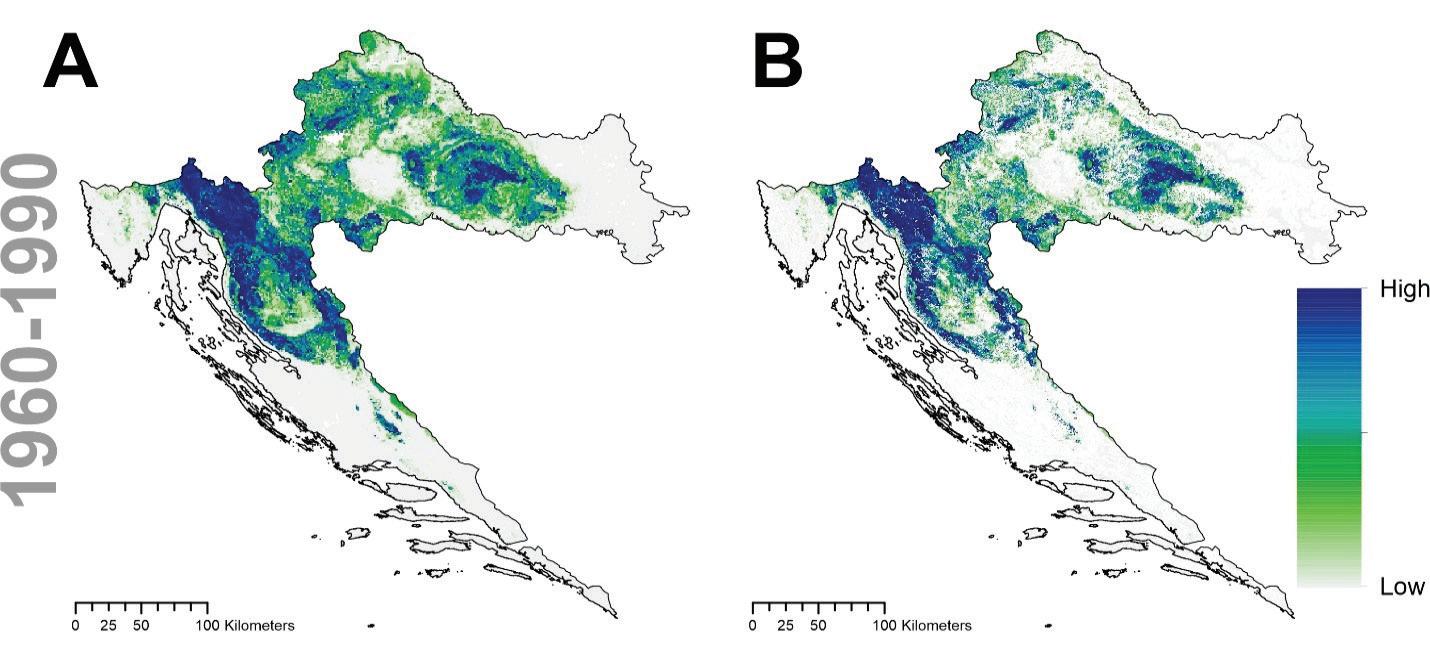
Model pogodnosti staništa za trenutne uvjete klime prikazan je na slici 20. Najznačajnije varijable koje pridonose modelu pogodnosti staništa za običnu bukvu su količina oborina u ljeto (PPT_sm), kontinentalnost (TD), postotni sadržaj gline u tlu (Clay) i oborine zimi (PPT_wt).
Slika 20. Projekcija trenutne pogodnosti staništa obične bukve u Hrvatskoj i to: A potencijalna (uključujući nešumske površine) i B stvarna (bez nešumskih površina).
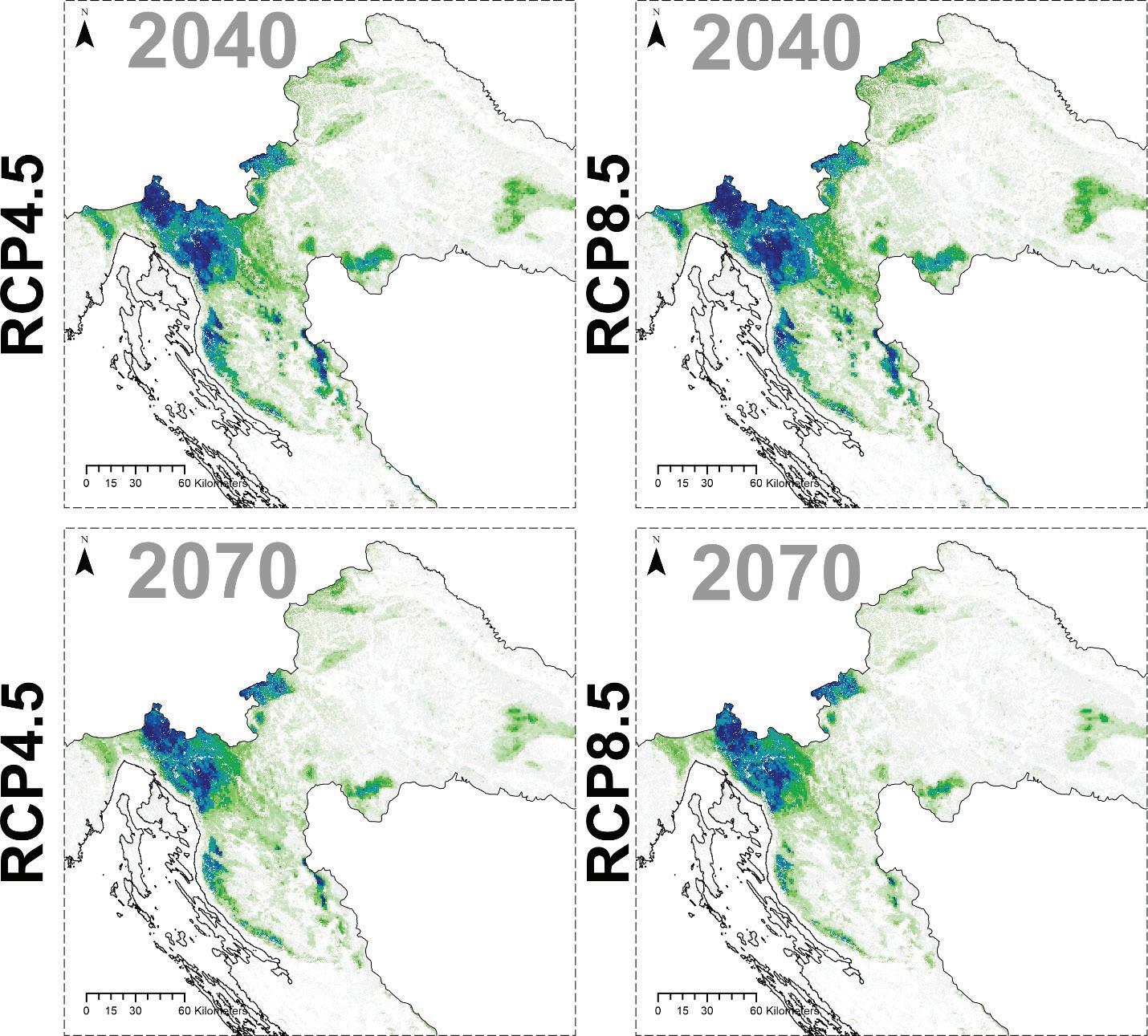
Prema očekivanom modelu klimatskih promjena predviđa se brza promjena ekološke pogodnosti obične bukve i to naročito u kontinentalnom dijelu Hrvatske ali i u nižim predjelima Like. Pogodno stanište za običnu bukvu u budućnosti prema oba scenarija emisija RCP4.5 i RCP8.5 predviđaju opstanak samo na većim nadmorskim visinama Gorskog kotara i Velebita te na kontinentalnim gorama (Slika 21).
ŠUMARSTVO 27
Slika 21. Buduće projekcije pogodnosti staništa za običnu bukvu u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (od 2011. – 2040. godine, od 2041. – 2070. godine). Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
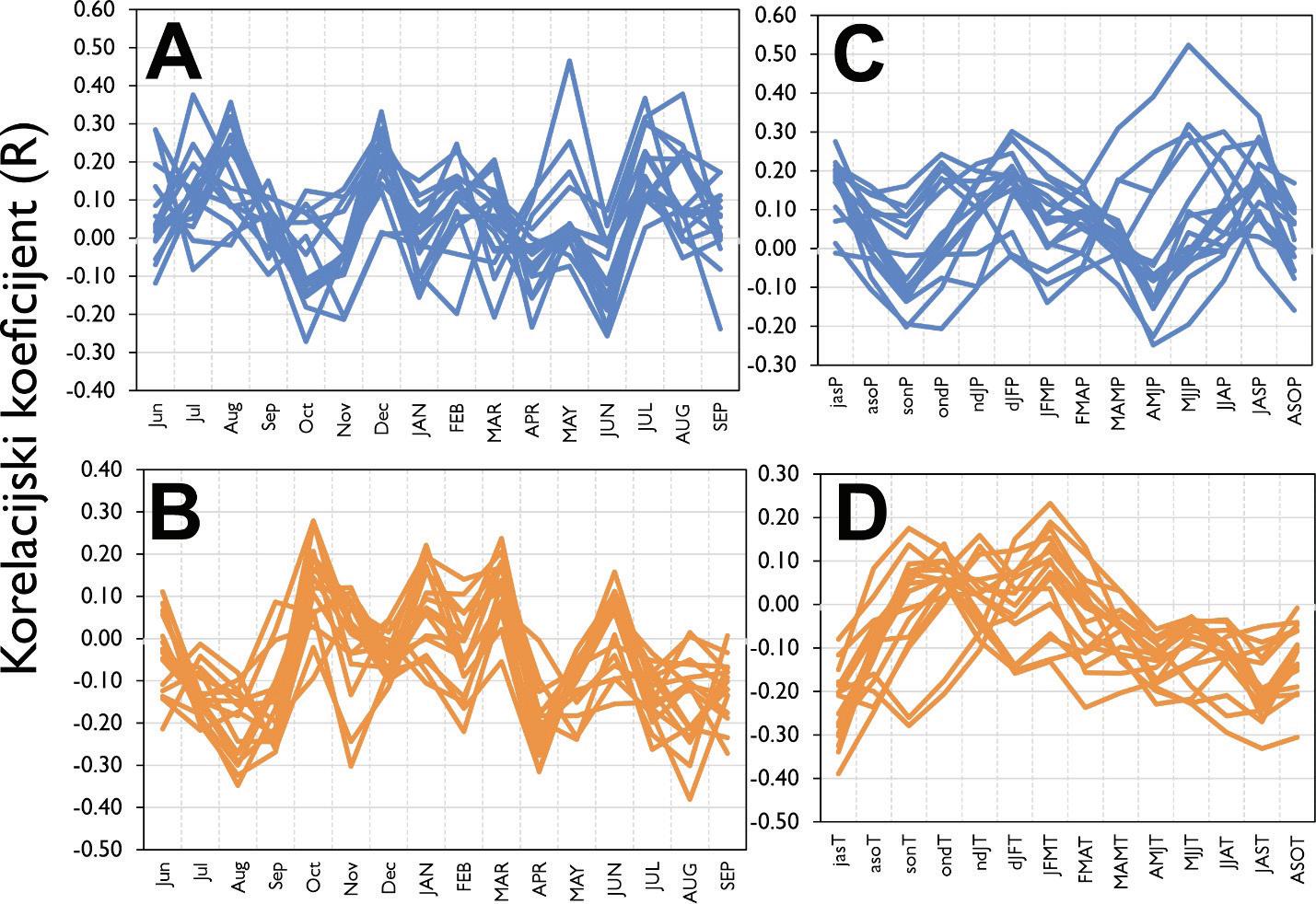
Rezultati analize klimatske osjetljivosti bukve ukazuju da je bukva značajno pozitivno osjetljiva na oborine u lipnju, odnosno na ukupne oborine u razdoblju od mjeseca svibnja do srpnja (MJJ). Veće količine oborina u ljeto (od svibnja do srpnja) utječu pozitivno na rast stabala bukve na svim istraživanim lokalitetima. Vrijednost ove zakonitosti značajno ovisi o nadmorskoj visini. Stabla koja rastu na nižim nadmorskim visinama osjetljivija su na manjak oborina od onih na višim nadmorskim visinama u ljetnim mjesecima. Analizom pomičnih korelacija utvrđeno je da je ova zakonitost stabilna tijekom posljednjih 100 godina i ne pokazuje promjene.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 28
Slika 22. Prosječne vrijednosti kronologija širina godova obične bukve na istraživanim plohama. Prostorni položaj pokusnih ploha (zeleni trokuti). A - Vrijednosti pomičnih korelacijskih koeficijenata između kronologija bukve i sezonskih vrijednosti sume oborina od svibnja do srpnja (plavo) te s temperaturom zraka od svibnja do srpnja (crveno). B – odnos korelacijskih koeficijenata sa sezonskim vrijednostima oborina prema nadmorskoj visini (gore) te u odnosu na poznate europske populacije (dolje).
ŠUMARSTVO 29
3.4. Hrast lužnjak (Quercus robur L.)
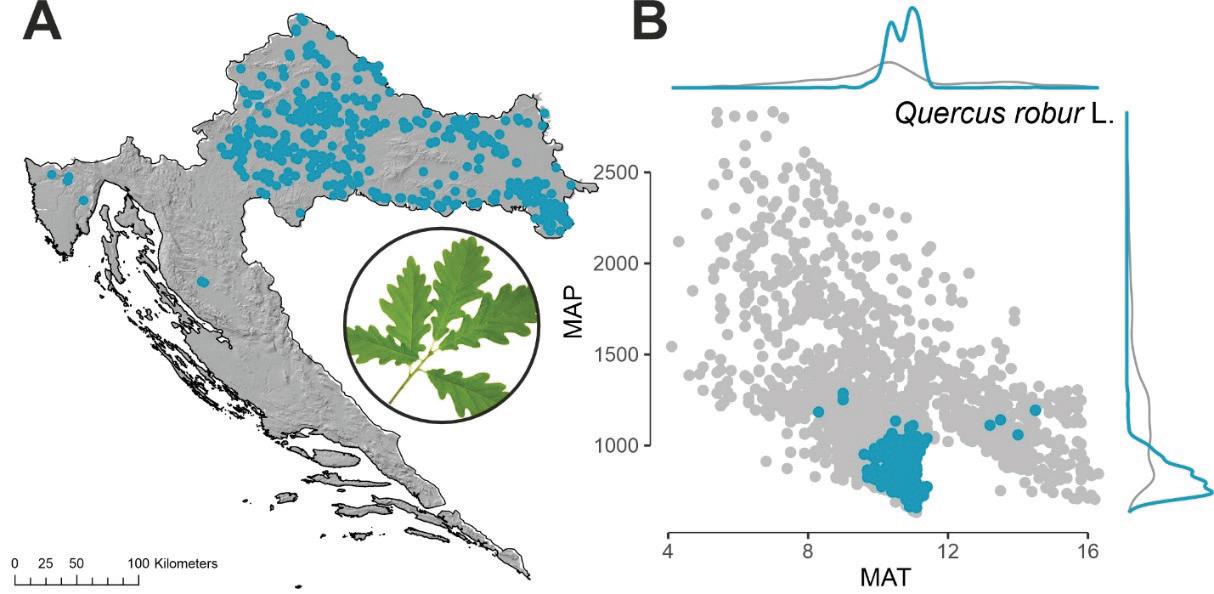
Hrast lužnjak jedna je od ekološki i gospodarski najvažnijih vrsta drveća u Hrvatskoj. Rasprostire se uglavnom u kontinentalnoj regiji u nizinskom vegetacijskom pojasu na površini od oko 200.000 ha. No osim toga pridolazi sporadično i u dinarskim krškim poljima u Lici na nadmorskim visinama između 700 i 900 m. Obzirom na klimatske prilike (oborine i temperaturu zraka) hrast lužnjak je vrsta uske ekološke valencije u odnosu na većinu prethodno spomenutih vrsta drveća. Što se tiče oborina njegova pojavnost je uglavnom u područjima s manjom količinom oborina (ispod 1400 mm) te područja s prosječnom godišnjom temperaturom zraka između 10 i 12 °C.
Slika 23. Prostorni položaj nalazišta hrasta lužnjaka u RH (Quercus robur L.) korišten za modeliranje pogodnosti staništa A. Odnos između prosječne temperature zraka MAT (ºC) i ukupne količine oborina MAP (mm) za plohe na kojima se pojavljuje hrast (plave točke) u odnosu na plohe gdje nema hrasta (sive točke) B.
Hrast lužnjak (Quercus robur L.) široko je rasprostranjena vrsta drveća u Europi. Unatoč velikoj geografskoj rasprostranjenosti i širokoj ekološkoj niši često se navodi da je varijabilnost rasta hrasta lužnjaka ograničena strogo definiranim lokalnim prilikama naročito hidromorfnim uvjetima tala (Thomas i sur., 2002). Prema dosadašnjim spoznajama hrast lužnjak je vrsta otporna na ekstremnu sušu i visoke temperature (Kuster i sur., 2014) a osjetljiv na kasne proljetne mrazove i nisku temperaturu zraka (Vitasse i sur., 2019). Tijekom suše hrast lužnjak sposoban je brzo raspodijeliti resurse u rast korijena kako bi iskoristio vlagu i hranjiva, istovremeno smanjujući gubitak vode transpiracijom (Thomas i Gausling, 2000). Međutim, niz regionalnih epizoda odumiranja hrasta tijekom 20-og stoljeća se pripisuje kombinaciji predisponirajući ekstremnih klimatskih (suše, mraz) i štetnih biotskih čimbenika (Andersson et al. 2011., Denman i sur., 2014.; Delatour 1983, Führer 1998., Levanič i sur., 2011). Tijekom posljednjih 20-ak godina zabilježena su brojna sušenja, odumiranja i propadanja hrasta lužnjaka diljem Hrvatske. Ukupna količina posječenih suhih i/ili odumrlih stabala iznosi oko 6.000.000 m3 što je oko 17% drvne zalihe hrasta lužnjaka u Hrvatskoj. Sušenja zahvaćaju gotovo sve dobne razrede hrastovih šuma a u relativnom iznosu su najveća u mlađim sastojinama.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 30
Slika 25. Prikaz posječenih stabala hrasta lužnjaka u Hrvatskoj tijekom posljednjih 120. godina. Crveni histogrami označavaju pojedinačna masovna odumiranja
Karta trenutne pogodnosti staništa hrasta lužnjaka u Hrvatskoj se dobro poklapa s njegovom stvarnom rasprostranjenosti. Najznačajniji čimbenici koji pridonose predikciji su broj dana s temperaturom zraka većom od 18°C (DDabove18), suma oborina u ljeto (PPT_sm) i jesen (PPT_at) te manje značajni (<5%) kapacitet tla za vodu (PAW) i postotno učešće pijeska u tlu (Sand).
Slika 26. Projekcija trenutne pogodnosti staništa hrasta lužnjaka u Hrvatskoj. Potencijalna pogodnost uključujući i nešumske površine A te stvarna (isključene nešumske površine) karta pogodnosti staništa B
Buduće projekcije pogodnosti staništa ukazuju na značajne promjene pogodnog staništa za rast i razvoj sastojina hrasta lužnjaka i to naročito na krajnjem istoku Hrvatske gdje se ujedno nalaze i naši najvrjedniji kompleksi ove vrijedne vrste (šumski bazeni Spačva i Koška). Kao pogodna staništa u budućnosti ostaju ona na zapadu trenutnog areala ali i na području ličkih polja (Slika 27).
ŠUMARSTVO 31
lužnjak u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (prvo od 2011. – 2040. godine i drugo od 2041. – 2070. godine). Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
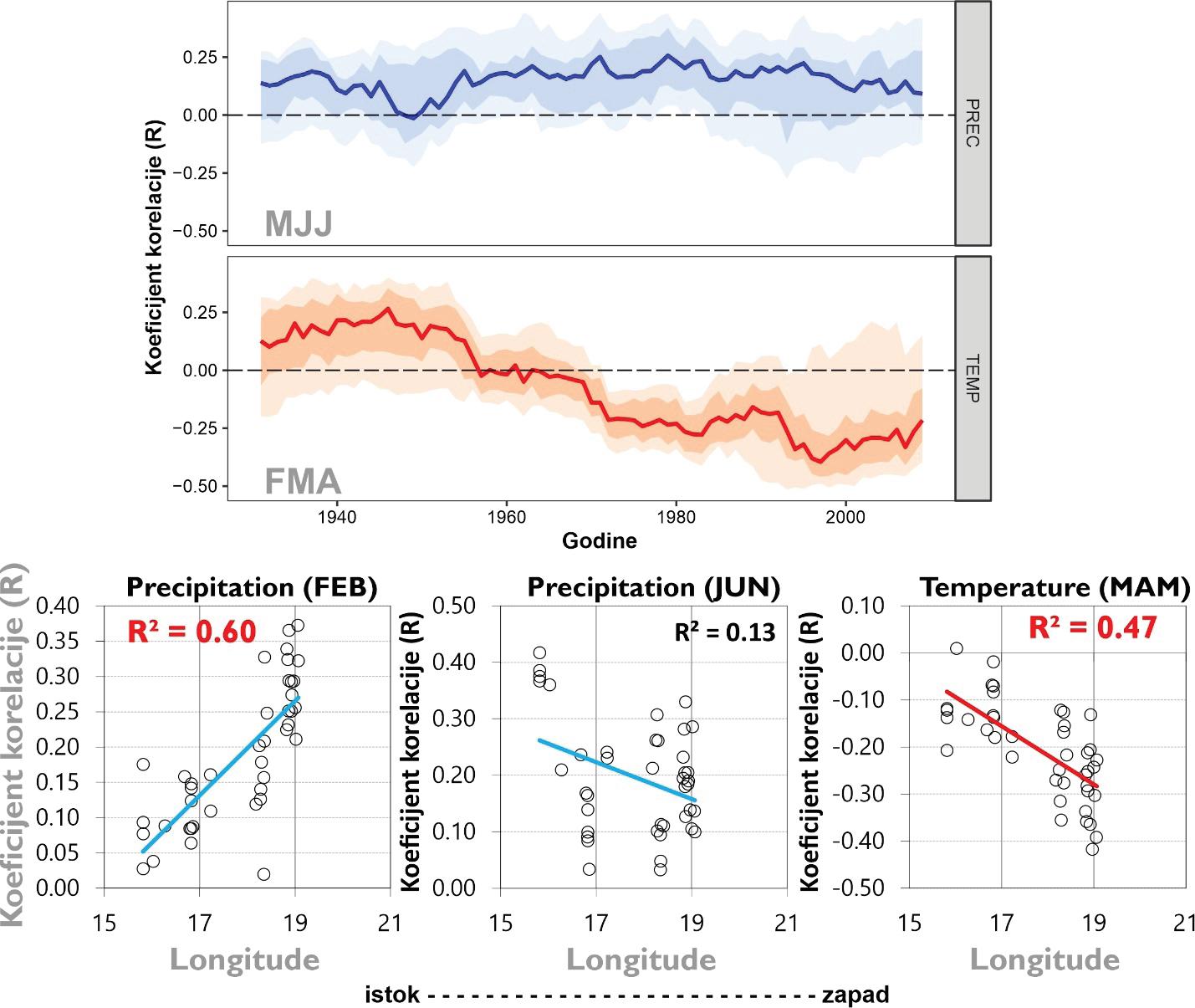
Analizama klimatske osjetljivosti kronologija rasta hrasta lužnjaka i klime (oborine i temperatura zraka) utvrđena je značajna klimatska osjetljivost na oborine u veljači i lipnju tekuće godine te negativna osjetljivost na temperature zraka u rano proljetnim mjesecima (prosjek temperatura od veljače do travnja) tekuće godine.
Tijekom promatranog razdoblja (posljednjih 100 godina) hrast nije mijenjao osjetljivost na oborine (Slika 28). Za razliku od oborina utvrđena je značajna promjena u klimatskoj osjetljivosti hrasta lužnjaka s temperaturom zraka u proljeće. Tijekom posljednjih 100 godina naročito nakon 1960. godine hrast postaje sve osjetljiviji na visoke proljetne temperature. Drugim riječima visoke temperature zraka (od veljače to travnja) sve više limitiraju rast hrasta lužnjaka.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 32
Slika 27. Buduće projekcije pogodnosti staništa za hrast
Slika 28. Prosječne vrijednosti kronologija širina godova hrasta lužnjaka na istraživanim plohama. Prostorni položaj pokusnih ploha (zeleni trokuti).
Usporedbom klimatske osjetljivosti prema geografskom položaju utvrđeno je da su populacije hrasta lužnjaka na krajnjem istoku areala (šumski bazeni Spačva i Koška) značajnije osjetljivije na deficit oborina u veljači u odnosu na zapadne populacije (Slika 29). Također populacije na istoku pokazuju i veće negativne korelacije s temperaturama zraka u odnosu na zapadne populacije (Slika 29).
Na području šumskog bazena Spačva (Slika 30) utvrđene su razlike u radijalnom prirastu vitalnih i odumrlih stabala hrasta lužnjaka pri čemu je utvrđeno da prosječni radijalni prirast odumrlih stabala zaostaje za prosječnim radijalnim prirastom vitalnih stabala dulje vremensko razdoblje (Dekanić 2014, Mikac i dr. 2020). Odumiranju hrasta lužnjaka i povećanom broju sušaca uvijek prethodi značajan pad radijalnog prirasta koji može trajati i nekoliko desetljeća prije konačnog odumiranja stabla.
Slika 29. Pomične korelacije s oborinama u ljeto (MJJ) te temperaturama zraka u proljeće (FMA) tekuće godine tijekom posljednjih 100 godina. Odnos između koeficijenata korelacije između kronologija rasta stabala hrasta i glavnih klimatskih čimbenika prema geografskoj dužini.
ŠUMARSTVO 33
Slika 30. Usporedba kronologija širina godova na području Spačve između živih (vitalnih) i (odumirućih i/ili odumrlih) stabala hrasta lužnjaka (crvena boja).
3.5. Poljski jasen (Fraxinus angustifolia Vahl)
Poljski jasen je rasprostranjen uglavnom u nizinskom dijelu Hrvatske u poplavnim područjima uz velike nizinske rijeke kao što su: Sava, Drava, Kupa i Dunav. Predikcijski model pogodnosti staništa pokazuje dobru podudarnost sa stvarnom distribucijom poljskog jasena u Republici Hrvatskoj. TSS vrijednost za združeni model iznosi 0.857. Najznačajnije varijable koje pridonose modelu pogodnosti staništa su oborine u jesen (PPT_at) s više od 40%, prosječna temperatura zraka u ljeto (Tave_sm) i oborine u ljeto (PPT_sm) te sadržaj pijeska u tlu (Sand).
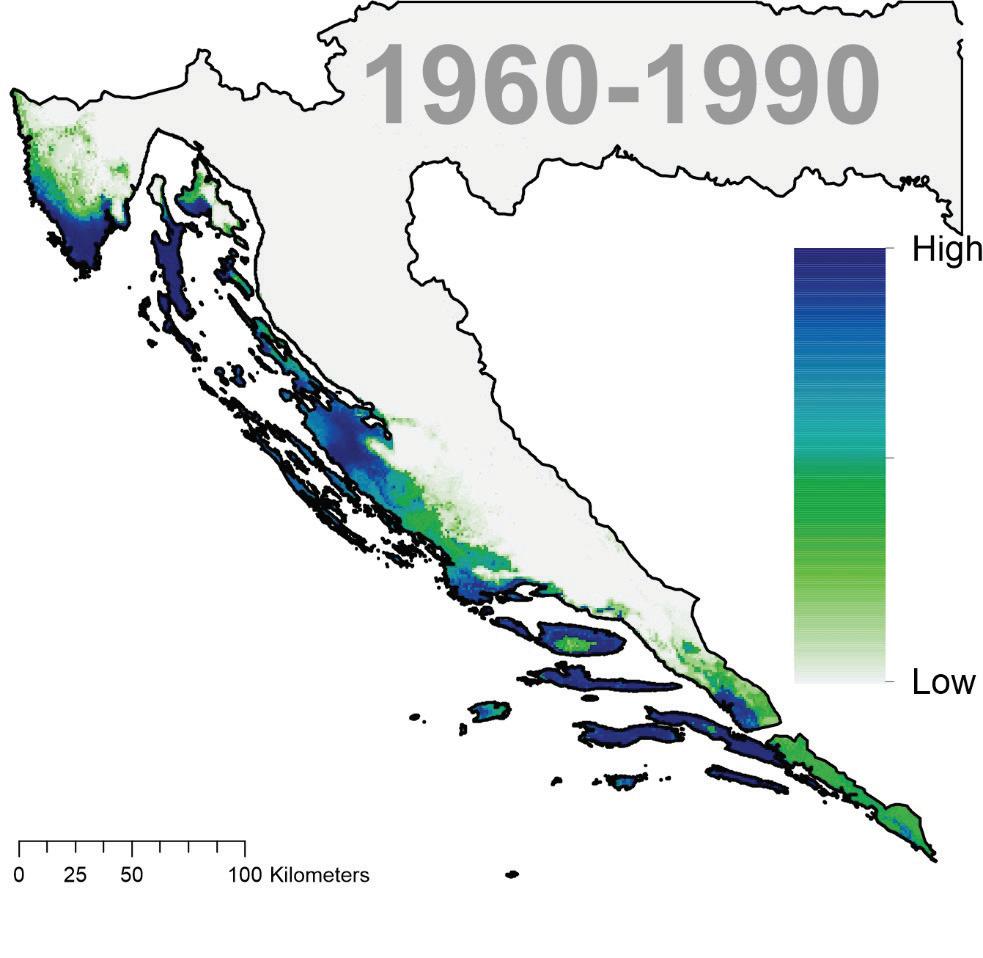
trenutne pogodnosti staništa poljskog jasena u Hrvatskoj za razdoblje klimatskih podataka od 1960 – 1990. godine. Trenutna potencijalna pogodnost A i stvarna pogodnost stanišnih prilika s obzirom na trenutnu rasprostranjenost šuma B
Sušenje i propadanje sastojina poljskog jasena (Fraxinus angustifolia Vahl) u posljednjih nekoliko godina poprimilo je katastrofalne razmjere na čitavom području pridolaska ove vrste u Republici Hrvatskoj. Prema procijeni, trenutno je sušenjem i propadanjem zahvaćeno i potencijalno ugroženo više do 30.000 ha sastojina poljskog jasena s ukupno 12 milijuna m3 drvne mase. Osim velike ekonomske i gospodarske štete nesagledivo su veće ekološke štete, naročito u područjima ekološke mreže Natura 2000. Uzroci sušenja i katastrofalnog propadanja jasenovih šuma dijelom se pripisuju gradaciji invazivne gljive Hymenoscyphus fraxineus, ali i nepovoljnom djelovanju klimatskih čimbenika poglavito suša unazad nekoliko godina. Sušenja i propadanja stabala poljskog jasena su u pojedinim šumskim sastojinama katastrofalna, često puta je sušenjem zahvaćeno i više od 90 % stabala u sastojini.
Mjere
promjenama za održivo upravljanje prirodnim resursima 34
prilagodbe klimatskim
Slika 31. Projekcija
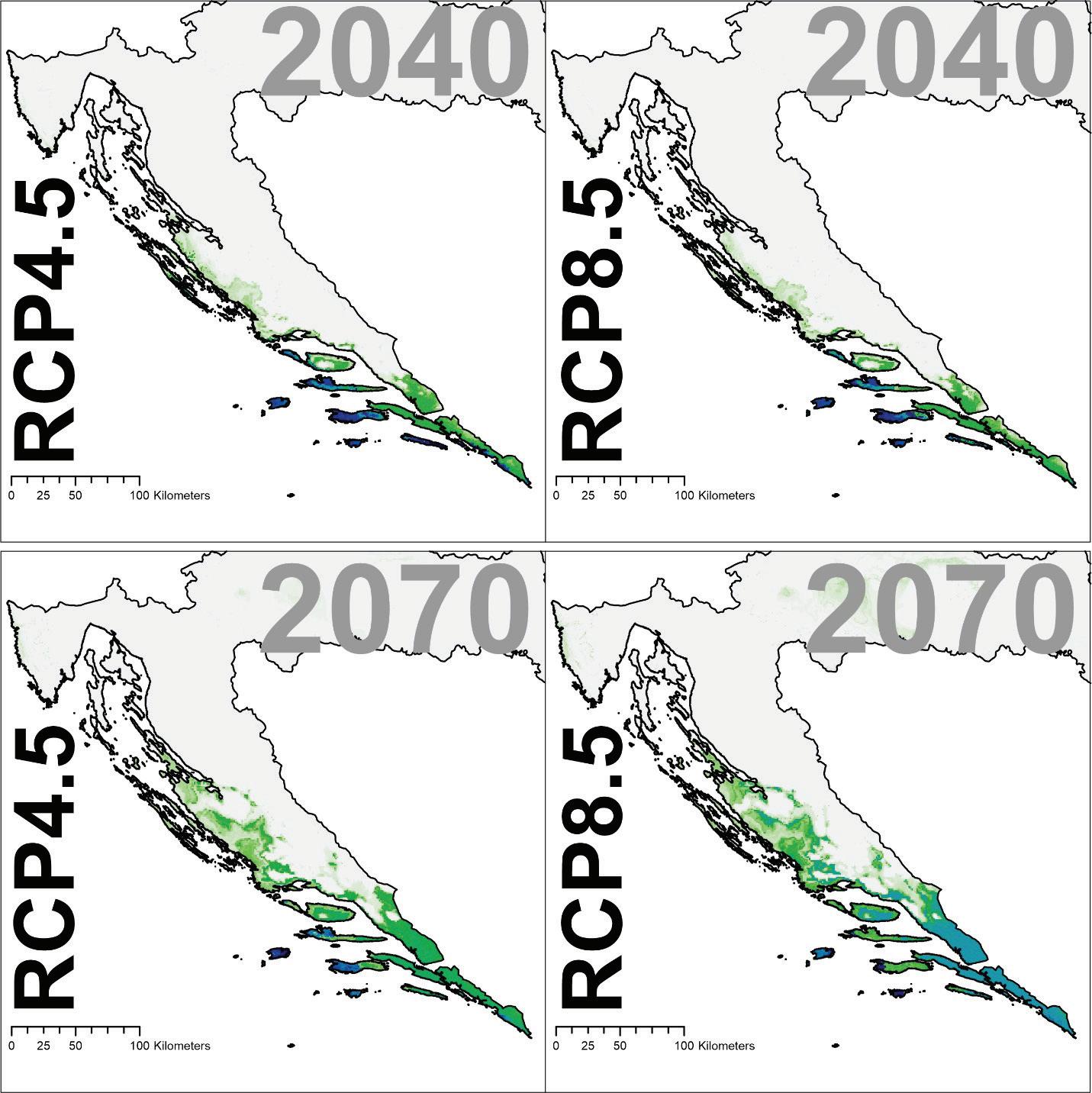
Modeli buduće prikladnosti staništa poljskog jasena ukazuju na značajni gubitak prikladnosti poglavito na istoku Hrvatske ali i na zapadnom dijelu njegove trenutne geografske distribucije i to posebice u gornjoj i srednjoj Posavini. Pogodnija staništa za poljski jasen predviđena su samo na nešto većim visinama unutar brežuljkastog vegetacijskog pojasa.
za poljski jasen u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (od 2011. – 2040. godine, od 2041. – 2070. godine). Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).

ŠUMARSTVO 35
Slika 32. Izgled sastojine poljskog jasena zahvaćen intenzivnim sušenjem i propadanjem
Slika 33. Buduće projekcije pogodnosti staništa
Analizom indeksnih kronologija širina godova poljskog jasena utvrđena je značajna pozitivna korelacija između dinamike rasta stabala jasena i oborina u rano ljeto tekuće godine (suma oborina od travnja do lipnja). Drugim riječima veća količina oborina u periodu od travnja do lipnja pogoduje boljem rastu stabala poljskog jasena. Istovremeno visoke temperature zraka limitiraju rast stabala poljskog jasena odnosno djeluju negativno i to posebice u rano proljeće (prosjek temperatura od ožujka do svibnja) (Slika 34). Najviše korelacije, odnosno najveću klimatsku osjetljivost pokazuju populacije poljskog jasena koje rastu na najnižim reljefnim depresijama (barama) i to na području srednje i donje Posavine. Stoga možemo konstatirati da će ove sastojine biti najugroženije od recentnih promjena klime.
Slika 34. Vrijednosti korelacijskih koeficijenata između kronologija poljskog jasena i mjesečnih vrijednosti klimatskih čimbenika i to: A – oborine (mm) po mjesecima, C - suma oborina za 3 uzastopna mjeseca, B – temperature zraka i D -prosjek temperatura zraka za tri uzastopna mjeseca.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 36
položaj pokusnih ploha poljskog jasena A. Pomične korelacije između indeksnih kronologija poljskog jasena i oborina od travnja do lipnja i temperature zraka od ožujka do svibnja B. Odnos koeficijenta korelacije o nadmorskoj visini istraživanih ploha C
ŠUMARSTVO 37
Slika 35. Prostorni
3.6. Alepski bor (Pinus halepensis Mill.)
Alepski bor je tipična mediteranska vrsta. Prirodno je rasprostranjen u srednjoj i južnoj Dalmaciji. Karta pogodnosti staništa pokazuje dobru podudarnost sa stvarnom geografskom rasprostranjenošću vrste. Najznačajnije varijable koje pridonose modelu pogodnosti staništa su MCMT – prosječna temperatura najhladnijeg mjeseca sa 51%, količina oborina u ljeto PPT_sm sa 35% i količina oborina u jesen PPT_at sa 14%.
U odnosu na ostale vrste drveća modeli budućih pogodnosti staništa ukazuju na proširenje pogodnog staništa u budućnosti (Slika 37.).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 38
Slika 36. Projekcija trenutne pogodnosti staništa alepskog bora u Hrvatskoj za razdoblje klimatskih podataka od 1960 – 1990. godine.
projekcije pogodnosti staništa za alepski bor u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (od 2011 – 2040. godine, od 2041. – 2070. godine). Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
ŠUMARSTVO 39
Slika 37. Buduće
3.7. Hrast crnika (Quercus ilex L.)
Hrast crnika obuhvaća slično područje kao i alepski bor. Za razliku od alepskog bora ipak zahtjeva nešto vlažnija tla. Karta pogodnosti staništa pokazuje dobru podudarnost sa stvarnom geografskom rasprostranjenošću vrste. Najznačajnije varijable koje pridonose modelu pogodnosti staništa su TD – kontinentalnost klime sa 52%, količina oborina u ljeto PPT_sm sa 30% i količina oborina u proljeće PPT_sp sa 18%.
Slika 38. Projekcija trenutne pogodnosti staništa hrasta crnike u Hrvatskoj za razdoblje klimatskih podataka od 1960 – 1990. godine.
U odnosu na alepski bor, hrast crnika prema svim modelima pokazuje značajno smanjenje pogodnosti staništa te redistribuciju povoljnog staništa prema južnim predjelima Hrvatske (Slika 39.).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 40
ŠUMARSTVO 41
Slika 39. Buduće projekcije pogodnosti staništa za hrast crniku u Republici Hrvatskoj za dva scenarija emisija (RCP4.5 i RCP8.5) za dva razdoblja (od 2011 – 2040. godine, od 2041. – 2070. godine). Plavom bojom su označena područja visoke pogodnosti staništa u rasponu od 0 do 1000 (odnosno od 0 – 100%).
4. ZAKLJUČCI S MJERAMA PRILAGODBE
Klimatske promjene na području Republike Hrvatske već su uvelike prisutne, a u narednom srednjoročnom i dugoročnom razdoblju donose njihov još i intenzivniji direktni utjecaji na uvjete za rast stabala i funkcioniranje šumskih ekosustava i to u dva osnovna smjera. Postupnim povećanjem temperature mijenjaju se okvirni uvjeti za rast, posebice s obzirom na hidrološki ciklus. Povećanjem temperature povećava se i potencijalna evapotranspiracija te isparavanje s tla i vegetacije. Procjena promjena režima oborina povezana je s puno većom razinom nesigurnosti, ali vjerojatno je da se količine oborina na godišnjoj razini neće mijenjati u većoj mjeri, no poprilično je sigurno da će doći do promjena u rasporedu oborina tijekom godine.
Osim postupnih promjena okvirnih uvjeta za rast, povećat će se i učestalost i intenzitet ekstremnih vremenskih događaja. Vrlo je vjerojatna smjena vegetacijskih sezona koje će po svom karakteru biti ili vrlo sušne (veća učestalost pojavljivanja) ili ekstremno vlažne sa suviškom vode u ciklusu. Također se očekuje veća učestalost intenzivnih oluja sa snažnim vjetrovima i posljedično velikim i naglim poremećajima u strukturi i funkcioniranju šumskih ekosustava putem vjetroloma i vjetroizvala velikoga broja stabala.
S predviđenim budućim promjenama klime, geografske distribucije vrsta drveća odnosno pogodnosti staništa će se proširiti ili smanjiti, geografski položaj ekoloških zona će se prostorno redistribuirati, produktivnost šumskih ekosustava će se promijeniti i sami ekosustavi bi se mogli reorganizirati nakon intenzivnih poremećaja (prirodnih nepogoda). Prilagodba gospodarenja na klimatske promjene uključuje praćenje i predviđanje promjena te poduzimanje radnji kako bi se izbjegle negativne posljedice ili iskoristile potencijalne dobrobiti tih promjena (Levina i Tirpak 2006). Usvajanje i provedba načela i prakse održivog gospodarenja šumama može pružiti čvrstu osnovu za suočavanje s izazovima klimatskih promjena. Međutim, Innes i sur. (2009.) istaknuli su da će naš neuspjeh u implementaciji dosadašnjih komponenti održivog gospodarenja šumama u mnogim šumama vjerojatno ograničiti sposobnost prilagodbe klimatskim promjenama. Upravitelji šuma trebat će planirati na više prostornih i vremenskih razina i usvojiti prilagodljivije i suradničke pristupe gospodarenju kako bi odgovorili na buduće izazove.
Provedena istraživanja u području šumarstva u sklopu projekta MEMORIE omogućila su bolje razumijevanje povijesnog ali i mogućeg budućeg direktnog utjecaja klimatskih promjena na rasta i razvoj glavnih vrsta drveća u Republici Hrvatskoj. Pri tome su korištene najmodernije metode znanstvenoistraživačkog rada trenutno dostupne. Novostvorena znanja i baze podataka omogućavaju bolje razumijevanje ekologije glavnih vrsta drveća te njihov odnos prema klimi i klimatskim promjenama.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 42
Uzimajući u obzir prostorne distribucije glavnih vrsta drveća te projekcije prema dva scenarija emisija za dva razdoblja možemo zaključiti da će kod većine vrsta (osim alepskog bora) doći do smanjenja prikladnosti staništa u budućnosti. To nužno ne znači da vrste više neće moći rasti na tim staništima nego da staništa neće biti u potpunosti pogodna, odnosno da će vrste biti ranjivije na promjene.
Naročito izražene posljedice po rast stabala istraživanih vrsta drveća mogu se očekivati u slučaju promjena nekog ili svih čimbenika koji su u ovome istraživanju utvrđeni kao ključni za definiranje ekološke niše pojedine vrste i/ili za radijalni prirast pojedinih stabala (Tablica 2).
U skladu s intenzitetom i smjerom promjena navedenih čimbenika potrebno je koncipirati i mjere prilagodbe šumskih ekosustava. Iako se generalni smjerovi promjena osnovnih klimatskih čimbenika za područje RH mogu odrediti s popriličnom sigurnošću (naročito za temperature zraka), procjena budućih klimatskih uvjeta za svaku pojedinu šumsku sastojinu povezana je s velikom nesigurnosti. Neki od najvažnijih izvora nesigurnosti procjene obuhvaćaju:
• osnovna nesigurnost svakog korištenog modela klimatskih promjena koja je neizostavno sadržana u svakoj procjeni koja se obavlja metodologijom matematičkog modeliranja,
• nesigurnosti povezane s „usitnjavanjem“ (downscalling) rezultata globalnih i regionalnih klimatskih modela na lokalnu razinu zadovoljavajuće razlučivosti, naročito uzimajući u obzir iznimno veliku orografsku i klimatsku razvedenost RH,
• s obzirom na kaotičnu prirodu globalnih geopolitičkih odnosa trenutačno je nemoguće odrediti po kojem će se scenariju klimatske promjene u konačnici odvijati (RCP4.5 ili RCP8.5, ili možda neki hibridni put kao kombinacija ova dva ili još i nekih drugih scenarija).
U okviru projekta MEMORIE definirane su i konkretne mjere za prilagodbu gospodarenja šumskim ekosustavima Republike Hrvatske. Mjere su oblikovane s ciljem uspostave temelja i ishodišta za budući sveobuhvatni sustav prilagodbe u okviru kojega će svi zainteresirani dionici u gospodarenju šumskim ekosustavima RH imati na raspolaganju alate i podatke pomoću kojih mogu na odgovarajući način rješavati konkretne i lokalne probleme vezane uz negativne utjecaje klimatskih promjena. Shodno tome, mjere koje se navode u ovome priručniku predstavljaju početak razvoja kataloga konkretnih mjera za prilagodbu upravljanja šumskim ekosustavima u RH.
Mjere su generalno koncipirane i funkcionalno grupirane tako da se njihovim provođenjem mogu ostvariti sljedeći osnovni ciljevi prilagodbe na području RH:
CILJEVI KOJE SE ODNOSE NA ŠUME, VRSTE I RADOVE U ŠUMAMA:
1. povećanje otpornosti šumskih vrsta i ekosustava na postupne promjene klimatskih čimbenika kao i na učestalije pojave naglih ekstremnih vremenskih događaja,
2. unaprjeđenje postupaka sanacije šumskih površina nakon pojave ekstremnih događaja,
3. povećanje raznolikosti vrsta šumskih drveća, povećanje unutarvrsne genetske raznolikosti i povećanje strukturne raznolikosti šumskih sastojina.
CILJEVI KOJI SE ODNOSE NA UNAPRJEĐENJE SUSTAVA PRILAGODBE U RH
1. uspostava sustava za kontinuirano i detaljno motrenje klimatskih i stanišnih promjena u šumskim ekosustavima, modeliranje i prognozu,
2. uspostava sustava za provođenje ciljanih istraživanja novih metoda i alata prilagođenih konkretnim lokacijama, staništima i vrstama drveća,
3. unaprjeđenje institucionalne potpore, zakonskih okvira i potpore krajnjim korisnicima.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 44
A. MJERE KOJE SE ODNOSE NA ŠUME, VRSTE I RADOVE U ŠUMAMA
Mjera A.1. Povećanje otpornosti vrsta drveća i šumskih sastojina na klimatske promjene
Podmjera: Intenziviranje radova njege šumskih sastojina
U mlađim razvojnim stadijima intenzivirati provođenje šumskouzgojnih zahvata njege šuma na način da se poveća vitalnosti i stabilnost sastojina.
Razviti i testirati nove metode proreda s ciljem povećanja kapaciteta produkcije sastojina, povećanja vitaliteta i smanjenja mortaliteta. Novi modeli proreda sastojina moraju obuhvatiti koncenzus između dosadašnjih kriterija (npr. kvaliteta drvne mase) i novih kriterija kao što su vitalitet i otpornost sastojina na buduće uvjete klime. Također modeli moraju biti prilagođeni svakoj pojedinoj vrsti drveća i budućim ekološkim uvjetima uslijed promjena klime.
U proredama forsirati ostavljanje, kvalitetnih, vitalnih i zdravih stabala kojima se mora osigurati prostorni raspored koji im omogućava pravilan razvoj krošanja odgovarajuće veličine.
Podmjera: Poboljšanje stanja degradiranih sastojina
U šumama niskog uzgojnog oblika (panjače) provoditi intenzivne šumskouzgojne radove trijebljenja te posredno stvoriti uvjete za konverziju prema visokom uzgojnom obliku.
Degradirane stadije visokih šuma (šikare, šibljaci, makije i garizi) zaštititi od dalje degradacije i šumskouzgojnim radovima (popunjavanja) postepeno prevoditi u visoke uzgojne oblike gdje je to moguće.
Podmjera: Prilagodba postupaka obnove sastojina
Obnove šumskih sastojina u regularnom gospodarenju provoditi na malim površinama (dijelovima sastojina). Povećati broj sjekoreda kroz dulje pomladno razdoblje.
U slučajevima dobrog trenutnog stanja starih sastojina gdje uvjeti staništa dozvoljavaju produljiti ophodnje.
Kod obnove pionirskih vrsta drveća (mediteranski borovi, npr. crni i alepski bor) na pomladnoj površini ostavljati manji broj pričuvaka starih stabala. Ova stara stabla ne treba nikako uklanjati s pomladne površine jer mogu biti izvor sjemena uslijed opetovanih prirodnih požara.
U prebornim sastojinama (šume s običnom jelom) obavezno provoditi normalno preborno gospodarenje. Posebnu pažnju posvetiti uspješnoj obnovi obične jele. Na kvalitetnijem tlu obnove prebornih sastojina umjesto stablimičnog načina provoditi obnovu grupimičnom ili skupinastom metodom.
Podmjera: Prilagoditi radove pošumljavanja, popunjavanja i umjetne bonove Potrebno je gdje je god moguće povećati površine pod šumama pošumljavanjem neobrasloga šumskog zemljišta. Prioritet treba dati površinama koje se naslanjaju na postojeće šumske površine i time povećavati komplekse jedinstvenih šumskih površina ili povezivati razdvojene šumske cjeline čime se povećava njihova sposobnost buferiranja eksternih negativnih utjecaja.
Prilikom pošumljavanja voditi računa o izboru vrsta i načinima sadnje. Vrste kojima se obavljaju radovi trebaju biti prilagođene sadašnjim ali i budućim klimatskim uvjetima konkretnog staništa. Svakom po-
ŠUMARSTVO 45
šumljavanju treba prethoditi detaljna analiza stanišnih uvjeta, konzultacija dostupnih projekcija klimatskih promjena barem na regionalnoj razini i tek u skladu s tim obaviti izbor vrsta drveća za pošumljavanje, popunjavanje ili umjetnu obnovu.
Kod radova popunjavanja, umjetne obnove ili pošumljavanja forsirati kvalitetni sadni materijal u pravilu obloženog korijena te istovremeno starije sadnice koje imaju veću mogućnost preživljavanja.
Mjera A.2. Unaprjeđenje postupaka sanacije šumskih sastojina nakon prirodnih nepogoda
Sanacije sastojina ili dijelova sastojina koje su zahvaćene prirodnim nepogodama kao što su ledolomi, vjetroizvale i vjetrolomovi dio odumrle mase (mrtvog drveta) treba ostavljati na površini, kao supstrat za regeneraciju osjetljivih vrsta npr. obične smreke.
Nakon sanacije obavezno provoditi popunjavanje sastojina. Popunjavanju treba prethoditi analiza intenziteta stanišnih promjena uslijed nastale nepogode i određivanje mogućih smjerova klimatskih promjena na konkretnoj lokaciji. U skladu s tim analizama treba odabrati vrste drveća za popunjavanje (sadnjom ili sjetvom sjemena ili sadnjom sadnica) pri čemu treba nastojati postići prirodni omjer smjese jednak onome prije nastupa nepogode. Ukoliko to nije u cijelosti moguće zbog prevelikog rizika uslijed zbog budućih klimatskih promjena, onda je potrebno odabrati one vrste koje će biti bolje prilagođene projicirane buduće klimatske uvjete.
Na površinama (u pravilu smrekovim sastojinama) koje su zahvaćene napadom potkornjaka i drugim prirodnim nepogodama koja nisu više pogodna za običnu smreku treba saditi jelu i/ili druge otporne vrste. Posebice se odnosi na površine gdje smreka nije autohtona vrsta. Nakon elementarnih nepogoda provoditi brze mjere sanacije staništa i sastojine s onim vrstama koje imaju najveći kapacitet adsorpcije na buduću klimu.
Mjera A.3. Povećati raznolikost vrsta drveća, genetske raznolikosti
Potrebno je u značajnoj mjeri u postojećim i budućim šumskim sastojinama raditi na aktivnom povećanju broja vrsta drveća koje ih tvore. S povećanjem brojnosti vrsta drveća smanjuje se rizik od pojave potencijalno velikih šteta do kojih dolazi kada se u sastojinama u kojima dominira jedna ili dvije vrste drveća pojave novi štetnici, ili ukoliko klimatski uvjeti pogoduju gradaciji i neuobičajeno velikoj brojnosti nekih štetnika koji do sada nisu izazivali veće štete.
Kod obnove sastojina potrebno je koristiti genetski materijal sa značajno većom genetskom raznolikošću. Drugim riječima, neće biti dovoljno samo povećati broj različitih vrsta drveća u šumskim sastojinama, nego će i svaka od tih vrsta zasebno trebati veću genetsku raznolikost. Ovim se načinom povećava širina moguće reakcije na promijenjene uvjete koji će na ovome području vladati za 30, 50 ili 100 godina. Naime, veća genetska raznolikost šumskog reprodukcijskog materijala osigurava veći raspon genetskih konstitucija čime se povećava adaptivna sposobnost stabala na buduće promjene.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 46
B. „SOFT“ MJERE ZA USPOSTAVU SUSTAVA PRILAGODBE U RH
Mjera B.1. Uspostaviti sustav za kontinuirano i detaljno motrenje klimatskih, stanišnih i sastojinskih promjena u šumama RH
Uspostaviti jedinstveni program nacionalnog praćenja utjecaja klimatskih promjena na šumske ekosustave kojim će se obuhvatiti šume u svim oblicima vlasništva (državno vlasništvo kojim gospodare Hrvatske šume ali i ostali dionici (nacionalni parkovi i dr.), i šume u privatnom vlasništvu kako pojedinaca tako i pravnih osoba).
Programom praćenja treba osigurati sustavno prikupljanje podataka o promjenama u sljedećim sastavnicama:
• Zdravstveno stanje i vitalnost šuma
• Produktivnost šuma i šumskih ekosustava
• Biološka raznolikost šuma
• Praćenje otpornosti i oporavka šumskih ekosustava nakon prirodnih nepogoda
• Praćenje dinamike šumskih ekosustava (gospodarskih i prašumskih sastojina)
Potrebno je uspostaviti sustav senzora u šumama Hrvatske za kontinuirano i detaljno motrenje klimatskih parametara na subregionalnoj razini, mikroklimatskih parametara unutar sastojina, parametara za procjenu promjena u hidrološkim ciklusima, kao i parametara kojima će se pratiti promjene u šumskom tlu. Motrenje klimatskih parametara na subregionalnoj razini omogućiti će korekciju globalno dostupnih dugoročnih klimatskih podataka i poboljšati njihovu razlučivost i primjenjivost za potrebe adaptacije na klimatske promjene.
Uspostavljeni sustav motrenja mora osigurati povezivost podataka o klimatskim i stanišnim promjenama sa podacima o reakciji stabala i sastojina na utvrđene promjene. Jedino se na taj način mogu točno kvantificirati konkretni utjecaji klimatskih promjena na vitalitet stabala i proizvodnost i stabilnost šumskih sastojina. Također se na taj način može obavljati motrenje uspješnosti provedenih adaptacijskih mjera i po potrebi u budućnosti vršiti korekcije kako se bude odvijao tijek promjena klimatskih uvjeta.
Mjera B.2. Sustavno provoditi ciljana istraživanja sa svrhom izrade novih metoda, alata i tehničkih rješenja prilagodbe za konkretne lokacije, staništa i vrste drveća
Utvrditi listu potreba za novim adaptivnim metodama u šumama Hrvatske, uzimajući u obzir vrstu drveća, stanišni tip, biogeografsku regiju i trenutačni i neposredno budući intenzitet negativnih posljedica i šteta uslijed utjecaja klimatskih primjena. Listu je potrebno složiti prema prioritetima i uspostaviti sustavno istraživanje i projektne okvire unutar kojih će se pristupiti istraživanju čija svrha i konkretni rezultat mora biti konkretna, primjenjiva i detaljno razrađena adaptivna mjera za rješavanje konkretnog adaptacijskog problema/slučaja.
U skladu s utvrđenim utjecajima klimatskih promjena, potrebno je preispitati postojeće i razviti nove modele proreda u regularnom kao i modele doznaka u prebornom gospodarenju šumama.
U poslovima uređivanja šuma, konkretno prilikom izrade Programa gospodarenja, svakako je potrebno detaljno preispitati sve podloge koje se koriste u određivanju i propisivanju etata i šumskih radova u pojedinim odsjecima. Primjerice, prirasno-prihodne tablice nikako više ne mogu biti vjerodostojan sustav prema kojem se određuje potencijalna proizvodnost šuma u budućem razdoblju za koje se propis
ŠUMARSTVO 47
izrađuje (uzimajući u obzir polu razdoblja i razdoblja, propisi se određuju za sljedećih 40 godina). Stoga je potrebno žurno uspostaviti istraživanja i projekte u okviru kojih se čim prije mora prilagoditi postojeće metode uređivanja šuma na način da se u njih uključe procjene utjecaja klimatskih promjena.
Potrebno je intenzivirati istraživačke aktivnosti na razvoju novih tehničkih rješenja koja će pospješiti primanje i preživljavanje mladih biljaka na terenu. To se prije svega odnosi na nove supstrate i materijale koji povećavaju kapacitete vode u tlu čime će se sadnicama prilikom presadnje na teren povećati otpornost na sve češće pojave ekstremnih sušnih događaja.
Uspostaviti pilot projekte kojima će se u šumskim ekosustavima ispitati mogućnosti primjene sustava prirodnih mjera zadržavanja vode (Natural Water Retention Measures – NWRM). Alati u okviru ovih mjera (male retencije, jezera i slično) mogu poslužiti kako bi se ublažile negativne posljedice unutargodišnje promjene režima oborina, primjerice, zadržavanjem prekomjernih količina vode izvan vegetacijske sezone na području šumske sastojine za upotrebu stabala u vegetacijskoj sezoni u slučaju pojave suše.
Mjera B.3. Prilagodba zakonskih okvira i unaprjeđenje potpore krajnjim korisnicima
Potrebno je žurno prilagoditi zakonodavne propise gospodarenja šumama u Hrvatskoj kako bi se omogućila primjena adaptacijskih mjera bez sputavanja trenutačnim rigidnim zakonodavnim okvirom unutar čijih dokumenata se adaptacija na klimatske promjene trenutačno uopće ne spominje. Adaptivne aktivnosti koje će trebati provoditi čim prije u šumama Hrvatske u mnogim će slučajevima biti u suprotnosti ili koliziji sa trenutačno važećim propisima Programa gospodarenja koji su određeni bez uzimanja u obzir potencijalnih utjecaja klimatskih promjena. U takvim slučajevima, adaptivne aktivnosti kojima se spašava šumski ekosustav su pravno gledajući ilegalne. Izmjeni zakonskih propisa svakako treba pristupiti sustavno, pri čemu su ključne promjene u Zakonu o šumama, Pravilniku o uređivanju šuma, Zakonu o šumskom reprodukcijskom materijalu i dr.
Potpora krajnjim korisnicima
Potrebno je u sljedećem kratkoročnom razdoblju uvelike poboljšati institucionalnu potporu svim šumoposjednicima i dionicima u procesima upravljanja šumskim resursima RH u njihovim naporima da prilagode svoje gospodarenje klimatskim promjenama. To se prije svega odnosi na edukaciju dionika o negativnim posljedicama klimatskih promjena na šumske ekosustave koji se očekuju u budućnosti, te adaptivnim mjerama kojima se ti negativni utjecaji mogu spriječiti ili ublažiti.
Također je potrebno poboljšati dostupnosti i primjenjivosti podataka i alata krajnjim korisnicima koji su im potrebni za prilagodbu na klimatske promjene za konkretna staništa, šumske sastojine i vrste šumskog drveća. Ovo je moguće ostvariti primjerice izradom regionalnih projekcija klimatskih promjena s katalozima potencijalnih negativnih utjecaja na vrste drveća koje u toj regiji dolaze koristeći u početnoj fazi podatke prikupljene projektom MEMORIE.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 48
5. LITERATURA
Auer, I.; Böhm, R.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Matulla, C.; Briffa, K.; Jones, P.; Efthymiadis, D.; Brunetti, M.; Nanni, T.; Maugeri, M.; Mercalli, L.; Mestre, O.; Moisselin, J.-M.; Begert, M.; Müller-Westermeier, G.; Kveton, V.; Bochnicek, O.; Stastny, P.; Lapin, M.; Szalai, S.; Szentimrey, T.; Cegnar, T.; Dolinar, M.; Gajic-Capka, M.; Zaninovic, K.; Majstorovic, Z.; Nieplova, E. HISTALP—Historical Instrumental Climatological Surface Time Series of the Greater Alpine Region. Int. J. Climatol. 2007, 27 (1), 17–46. https://doi.org/10.1002/joc.1377.
Giorgi, F.; Lionello, P. Climate Change Projections for the Mediterranean Region. Global and Planetary Change 2008, 63 (2–3), 90–104. https://doi.org/10.1016/j.gloplacha.2007.09.005.
Climate Change 2014: Mitigation of Climate Change ; Working Group III Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., IPCC, IPCC, Eds.; Climate change 2014; Cambridge Univ. Press: New York, NY, 2014.
Floodplain Management: Reducing Flood Risks and Restoring Healthy Ecosystems; European Environment Agency, 2016
Gaume, E.; Bain, V.; Bernardara, P.; Newinger, O.; Barbuc, M.; Bateman, A.; Blaškovičová, L.; Blöschl, G.; Borga, M.; Dumitrescu, A.; Daliakopoulos, I.; Garcia, J.; Irimescu, A.; Kohnova, S.; Koutroulis, A.; Marchi, L.; Matreata, S.; Medina, V.; Preciso, E.; Sempere-Torres, D.; Stancalie, G.; Szolgay, J.; Tsanis, I.; Velasco, D.; Viglione, A. A Compilation of Data on European Flash Floods. Journal of Hydrology 2009, 367 (1–2), 70–78. https://doi.org/10.1016/j.jhydrol.2008.12.028.
Jaun, S.; Ahrens, B.; Walser, A.; Ewen, T.; Schär, C. A Probabilistic View on the August 2005 Floods in the Upper Rhine Catchment. Nat. Hazards Earth Syst. Sci. 2008, 8 (2), 281–291. https://doi. org/10.5194/nhess-8-281-2008.
Kundzewicz, Z. W.; Pińskwar, I.; Brakenridge, G. R. Large Floods in Europe, 1985–2009. Hydrological Sciences Journal 2013, 58 (1), 1–7. https://doi.org/10.1080/02626667.2012.745082.
Floods in the Balkans: Bosnia and Herzegovina, Croatia and Serbia Situation Report; Situation report; WHO, 2014.
Communication on Flood Risk Management; Flood Prevention, Protection and Mitigation; European commission, 2004
Alfieri, L.; Burek, P.; Feyen, L.; Forzieri, G. Global Warming Increases the Frequency of River Floods in Europe. Hydrol. Earth Syst. Sci. Discuss. 2015, 12 (1), 1119–1152. https://doi.org/10.5194/hessd-12-1119-2015.
Winsemius, H. C.; Aerts, J. C. J. H.; van Beek, L. P. H.; Bierkens, M. F. P.; Bouwman, A.; Jongman, B.; Kwadijk, J. C. J.; Ligtvoet, W.; Lucas, P. L.; van Vuuren, D. P.; Ward, P. J. Global Drivers of Future River Flood Risk. Nature Clim Change 2016, 6 (4), 381–385. https://doi.org/10.1038/nclimate2893.
Fink, A. H.; Brücher, T.; Krüger, A.; Leckebusch, G. C.; Pinto, J. G.; Ulbrich, U. The 2003 European Summer Heatwaves and Drought -Synoptic Diagnosis and Impacts: European Heatwave - Impacts. Weather 2004, 59 (8), 209–216. https://doi.org/10.1256/wea.73.04.
ŠUMARSTVO 49
Hanel, M.; Rakovec, O.; Markonis, Y.; Máca, P.; Samaniego, L.; Kyselý, J.; Kumar, R. Revisiting the Recent European Droughts from a Long-Term Perspective. Sci Rep 2018, 8 (1), 9499. https://doi. org/10.1038/s41598-018-27464-4.
Cook, E. R.; Jacoby, G. C. Potomac River Streamflow Since 1730 as Reconstructed by Tree Rings. Journal of Climate and Applied Meteorology 1983, 22 (10), 1659–1672.
Kames, S. Sensitivity of Vessels in Black Ash (Fraxinus Nigra Marsh.) Tree Rings to Fire and Hydro-Climatic Variables. 2009, 174.
Perez-Valdivia, C.; Sauchyn, D. Tree-Ring Reconstruction of Groundwater Levels in Alberta, Canada: Long Term Hydroclimatic Variability. Dendrochronologia 2011, 29 (1), 41–47. https://doi.org/10.1016/j.dendro.2010.09.001.
Shah, S. K.; Bhattacharyya, A.; Chaudhary, V. Streamflow Reconstruction of Eastern Himalaya River, Lachen ‘Chhu’, North Sikkim, Based on Tree-Ring Data of Larix Griffithiana from Zemu Glacier Basin. Dendrochronologia 2014, 32 (2), 97–106. https://doi.org/10.1016/j.dendro.2014.01.005.
Speer, J. H.; Shah, S. K.; Truettner, C.; Pacheco, A.; Bekker, M. F.; Dukpa, D.; Cook, E. R.; Tenzin, K. Flood History and River Flow Variability Recorded in Tree Rings on the Dhur River, Bhutan. Dendrochronologia 2019, 56, 125605. https://doi.org/10.1016/j.dendro.2019.125605.
Tockner, K.; Stanford, J. A. Riverine Flood Plains: Present State and Future Trends. Envir. Conserv. 2002, 29 (3), 308–330. https://doi.org/10.1017/S037689290200022X.
Robinson, M.; Cognard-Plancq, A.-L.; Cosandey, C.; David, J.; Durand, P.; Führer, H.-W.; Hall, R.; Hendriques, M. O.; Marc, V.; McCarthy, R.; McDonnell, M.; Martin, C.; Nisbet, T.; O’Dea, P.; Rodgers, M.; Zollner, A. Studies of the Impact of Forests on Peak Flows and Baseflows: A European Perspective. Forest Ecology and Management 2003, 186 (1–3), 85–97. https://doi.org/10.1016/S03781127(03)00238-X.
Bradshaw, C. J. A.; Sodhi, N. S.; Peh, K. S.-H.; Brook, B. W. Global Evidence That Deforestation Amplifies Flood Risk and Severity in the Developing World. Global Change Biol 2007, 13 (11), 2379–2395.
https://doi.org/10.1111/j.1365-2486.2007.01446.x.
Hughes, F. M. R.; del Tánago, M. G.; Mountford, J. O. Restoring Floodplain Forests in Europe. In A Goal-Oriented Approach to Forest Landscape Restoration; Stanturf, J., Madsen, P., Lamb, D., Eds.; World Forests; Springer Netherlands: Dordrecht, 2012; Vol. 16, pp 393–422. https://doi. org/10.1007/978-94-007-5338-9_15.
Schneider, E. Floodplain Restoration of Large European Rivers, with Examples from the Rhine and the Danube. In Restoration of Lakes, Streams, Floodplains, and Bogs in Europe; Eiseltová, M., Ed.; Wetlands: Ecology, Conservation and Management; Springer Netherlands: Dordrecht, 2010; Vol. 3, pp 185–223. https://doi.org/10.1007/978-90-481-9265-6_11.
Horner, G. J.; Baker, P. J.; Nally, R. M.; Cunningham, S. C.; Thomson, J. R.; Hamilton, F. Mortality of Developing Floodplain Forests Subjected to a Drying Climate and Water Extraction. Global Change Biology 2009, 15 (9), 2176–2186. https://doi.org/10.1111/j.1365-2486.2009.01915.x.
Pittock, J.; Hansen, L. J.; Abell, R. Running Dry: Freshwater Biodiversity, Protected Areas and Climate Change. Biodiversity 2008, 9 (3–4), 30–38. https://doi.org/10.1080/14888386.2008.9712905.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 50
Boshier, D.; Oxford Forestry Institute. Ash Species in Europe: Biological Characteristics and Practical Guidelines for Sustainable Use; Oxford Forestry Institute: Oxford, 2005.
Marques, I. C. G. Fraxinus Angustifolia Vahl as a Valuable Species in Riparian Rehabilitation Projects. From Annual Growth to Habitat Preference of Narrow-Leaved Ash in Southern Portugal. masterThesis, ISA-UL, 2016
Mikac, S.; Žmegač, A.; Trlin, D.; Paulić, V.; Oršanić, M.; Anić, I. Drought-Induced Shift in Tree Response to Climate in Floodplain Forests of Southeastern Europe. Sci Rep 2018, 8 (1), 16495. https://doi. org/10.1038/s41598-018-34875-w.
Muñoz, N. G.; Linares, J. C.; Castro-Díez, P.; Sass-Klaassen, U. Contrasting Secondary Growth and Water Use Efficiency Patterns in Native and Exotic Trees Co-Occurring in Inner Spain Riparian Forests. Forest Systems 2015, 24 (1), 017. https://doi.org/10.5424/fs/2015241-06586.
Ratna, S. B.; Ratnam, J. V.; Behera, S. K.; Cherchi, A.; Wang, W.; Yamagata, T. The Unusual Wet Summer (July) of 2014 in Southern Europe. Atmospheric Research 2017, 189, 61–68. https://doi.org/10.1016/j.atmosres.2017.01.017.
Levanič, T. Atrics – A New System for Image Acquisition in Dendrochronology. trre 2007, 63 (2), 117–122. https://doi.org/10.3959/1536-1098-63.2.117.
Baillie, M. G. L.; Pilcher, J. R. A Simple Crossdating Program for Tree-Ring Research. 1973
Schweingruber, F. H. Tree Rings: Basics and Applications of Dendrochronology; 1988.
Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree Ring Bulletin 1983, 44, 69–78.
Program ARSTAN, a Tree-Ring Standardization Program Based on Detrending and Autoregressive Time Series Modeling with Interactive Graphics; ree-Ring Laboratory Lamont Doherty Earth Observatory of Columbia University: New York, NY, 2005.
Cook, E. R.; Peters, K. Calculating Unbiased Tree-Ring Indices for the Study of Climatic and Environmental Change. The Holocene 1997, 7 (3), 361–370. https://doi.org/10.1177/095968369700700314.
Mosteller, F.; Tukey, J. W. Data Analysis and Regression : A Second Course in Statistics; Reading, Mass. : Addison-Wesley Pub. Co, 1977
Harris, I.; Jones, P. D.; Osborn, T. J.; Lister, D. H. Updated High-Resolution Grids of Monthly Climatic Observations - the CRU TS3.10 Dataset: UPDATED HIGH-RESOLUTION GRIDS OF MONTHLY CLIMATIC OBSERVATIONS. Int. J. Climatol. 2014, 34 (3), 623–642. https://doi.org/10.1002/joc.3711.
Zang, C.; Biondi, F. Dendroclimatic Calibration in R: The BootRes Package for Response and Correlation Function Analysis. Dendrochronologia 2013, 31 (1), 68–74. https://doi.org/10.1016/j.dendro.2012.08.001.
Salas, J. D.; Tarawneh, Z.; Biondi, F. A Hydrological Record Extension Model for Reconstructing Streamflows from Tree-Ring Chronologies: DENDROHYDROLOGICAL RECORD EXTENSION MODEL. Hydrol. Process. 2015, 29 (4), 544–556. https://doi.org/10.1002/hyp.10160.
ŠUMARSTVO 51
Meko, D.; Stockton, C. W.; Boggess, W. R. THE TREE-RING RECORD OF SEVERE SUSTAINED DROUGHT. J Am Water Resources Assoc 1995, 31 (5), 789–801. https://doi.org/10.1111/j.1752-1688.1995. tb03401.x.
Fritts, H. C. Tree Rings and Climate; Academic Press: London ; New York, 1976.
Koprowski, M.; Okoński, B.; Gričar, J.; Puchałka, R. Streamflow as an Ecological Factor Influencing Radial Growth of European Ash (Fraxinus Excelsior (L.)). Ecological Indicators 2018, 85, 390–399. https://doi.org/10.1016/j.ecolind.2017.09.051.
Alley, W. M. The Palmer Drought Severity Index: Limitations and Assumptions. Journal of climate and applied meteorology 1984, 23, 1100–1109. https://doi.org/10.1175/1520-0450(1984)023<1100:TPDSIL>2.0.CO;2.
Pušpure, I.; Gerra-Inohosa, L.; Matisons, R.; Laiviņš, M. Tree-Ring Width of European Ash Differing by Crown Condition and Its Relationship with Climatic Factors in Latvia. Baltic Forestry 2017, 23 (1), 244–252.
Okonski, B. RADIAL GROWTH OF PEDUNCULATE OAK AND EUROPEAN ASH ON ACTIVE RIVER TERRACES: HYDROLOGIC AND CLIMATIC CONTROLS. Infrastruktura i Ekologia Terenów Wiejskich / Infrastucture and Ecology of Rural Areas 2017, No. III/1, 1075–1091. https://doi.org/10.14597/infraeco.2017.3.1.083.
Buntgen, U.; Tegel, W.; Nicolussi, K.; McCormick, M.; Frank, D.; Trouet, V.; Kaplan, J. O.; Herzig, F.; Heussner, K.-U.; Wanner, H.; Luterbacher, J.; Esper, J. 2500 Years of European Climate Variability and Human Susceptibility. Science 2011, 331 (6017), 578–582. https://doi.org/10.1126/science.1197175.
Cooper, R. J.; Melvin, T. M.; Tyers, I.; Wilson, R. J. S.; Briffa, K. R. A Tree-Ring Reconstruction of East Anglian (UK) Hydroclimate Variability over the Last Millennium. Clim Dyn 2013, 40 (3–4), 1019–1039. https://doi.org/10.1007/s00382-012-1328-x.
Raffalli-Delerce, G.; Masson-Delmotte, V.; Dupouey, J. L.; Stievenard, M.; Breda, N.; Moisselin, J. M. Reconstruction of Summer Droughts Using Tree-Ring Cellulose Isotopes: A Calibration Study with Living Oaks from Brittany (Western France). Tellus B: Chemical and Physical Meteorology 2004, 56 (2), 160–174. https://doi.org/10.3402/tellusb.v56i2.16405.
Čufar, K.; De Luis, M.; Eckstein, D.; Kajfež-Bogataj, L. Reconstructing Dry and Wet Summers in SE Slovenia from Oak Tree-Ring Series. Int J Biometeorol 2008, 52 (7), 607–615. https://doi.org/10.1007/ s00484-008-0153-8.
Zang, C. & Biondi, F. Treeclim: an R package for the numerical calibration of proxy-climate relationships. Ecography 38, 431-436 (2014).
Tessier, A., Carignan, R. & Belzile, N. Processes Occurring at the Sediment-Water Interface: Emphasis on Trace Elements. In: Buffle, J. and DeVitre, R.R., Eds., Chemical and Biological Regulation of Aquatic System 137-173 (1994).
Tumajer, J., Treml, V. Response of floodplain pedunculate oak (Quercus robur L.) tree-ring width and vessel anatomy to climatic trends and extreme hydroclimatic events. Forest Ecology and Management 379, 185–194 (2016).
Rozas, V. Dendrochronology of pedunculate oak (Quercus robur L.) in an old-growth pollarded woodland
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 52
in northern Spain: tree-ring growth responses to climate. Ann. For. Sci. 62, 209-218 (2005).
Schneider-Jacoby, M., Schwaderer, G., & Fremuth, W. The South-Eastern European Green Belt. In Ullrich, K. & Riecken, U. (Eds.), The green belt of Europe - From vision to reality. IUCN, Gland, (2006).
Friedrichs, D. A., Neuwirth, B., Winiger, M. & Löffler, J. Methodologically induced differences in oak site classifications in a homogeneous tree-ring network. Dendrochronologia 27, 21–30 (2009).
Führer, E. Oak Decline in Central Europe: A Synopsis of Hypotheses. Proceedings: Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects. USDA Forest Service General Technical Report NE-247, 7-24 (1998).
Hughes, F. & Richards, K. The flooded forest. The FLOBAR2 Project (2003).
Kern, Z. et al. Multiple tree-ring proxies (earlywood width, latewood width and δ13C) from pedunculate oak (Quercus robur L.), Hungary. Quaternary International 293, 257-267 (2013).
Klimo, E. & Hager, H. Floodplain forests of the temperate zone of Europe. Lesnická práce (2008).
FAO i CIFOR, 2019: FAO Framework Methodology for Climate Change Vulnerability Assessments of Forests and Forest Dependent People. Rome, str: 1-71.
Dekanić, S. 2014: Morfološka i dendrokronološka analiza oštećenih stabala hrasta hrasta lužnjaka (Quercus robur L.) u starim sastojinama spačvanskog bazena. Doktorska disertacija, Šumarski fakultet Sveučilišta u Zagrebu.
Mikac, S., Trlin, D., Žmegač, A., Dekanić, S., Miklić, B., Anić, I., 2020: Utjecaj recentnih promjena klime na nizinske šumske ekosustave hrasta lužnjaka (Quercus robur L.) u Hrvatskoj. Zbornik radova sa znanstvenog skupa Gospodarenje šumama u uvjetima klimatskih promjena i prirodnih nepogoda, ur. Anić, I., str. 1-15.
ŠUMARSTVO 53
Klimatske promjene i POLJOPRIVREDA
1 / Agronomski fakultet Sveučilišta u Zagrebu, Odsjek za agroekologiju Zavod za ishranu bilja. Svetošimunska cesta 25, 10000 Zagreb
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 54
prof. dr. sc. Milan Poljak1 zv. prof. dr. sc. Boris Lazarević1
1. Uvod
Krumpir (Solanum tuberosum L.) je uz pšenicu (Triticum aestivum L.), kukuruz (Zea mays L.) i rižu (Oryza sativa L.) jedna od najznačajnijih poljoprivrednih kultura u svijetu. Uzgaja se diljem svijeta te ostvaruje relativno visoke prinose, uz to visoke je nutritivne vrijednosti što ga čini i izrazito važnom kulturom sa stanovišta sigurnosti hrane i borbe protiv pothranjenosti, dva glavna cilja FAO Agende za održivi razvoj do 2030.
Godišnja svjetska proizvodnja krumpira iznosi 376,875,686 tona. Najveći pojedinačni svjetski proizvođač krumpira je Kina s 99,122,420 tona godišnje, a slijedi je Indija s 43,770,000 tona godišnje. Najveće prinose ostvaruju proizvođači iz SAD-a, prosječno 49 t/ha. Krumpir je u Hrvatskoj u 2021 godini uzgajan na 8786 ha, a proizvedeno je 127 826 t s prosječnim prinosom 14.5 t/ha (Tablica 1).
Tablica 1. Godišnja proizvodnja krumpira u Hrvatskoj u posljednjih pet godina (površine, ukupni i prosječni prinosi).
Globalni klimatski podaci pokazuju da je posljednjih sedam godina najtoplijih sedam godina od kad postoje klimatološka mjerenja, a 2020 je bila za 1.02 °C toplija u odnosu na prosjek 1951-1980. Globalne klimatske promjene povećavaju učestalost pojavljivanja i trajanje nepovoljnih klimatskih uvjeta za uzgoj poljoprivrednih kultura. Kako bi se mogli oduprijeti negativnom utjecaju visokih temperatura na krumpir i pronašli metode za sprječavanje šteta od visokih temperatura važno je utvrditi poremećaje (morfološke i fiziološke) koje visoke temperature izazivaju kod krumpira. Klimatske promjene se povezuju s povećanom koncentracijom CO2 u atmosferi. Povećanje koncentracije CO2 u atmosferi će pogodovati većini poljoprivrednih kultura, pa tako i krumpiru. No, krumpir je izrazito osjetljiv na visoke temperature te se procjenjuje da će klimatske promjene izazvati pad prinosa krumpira za 18-32%.
U sklopu projekta „MEMORIE – Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima“ provedena su istraživanja utjecaja visokih temperatura na morfološka i fiziološka svojstva krumpira. Pokus je proveden na tri sorte krumpira (Agria, Bellarosa i Desiree) koje su uzgajane u kontroliranim uvjetima komora rasta, pri različitim temperaturnim dan/noć režimima: 20/15 °C, 25/20 °C, 30/25 °C, 35/30 °C, te u poljskim uvjetima tijekom vegetacijskih sezona 2021. i 2022. Poljski pokusi su obavljani na lokaciji Lovinac (44° 23´ 24˝ N, 15° 40´ 48˝ E) u dva roka sadnje (rani i kasni rok) te uz navodnjavanje i bez navodnjavanja. Sve ostale agrotehničke mjere su bile su sukladne dobroj poljoprivrednoj praksi i istovjetne za sve članove pokusa. Mjerenja su obuhvaćala morfološke analize upotrebom 3D multispektralnog skenera (PlantEye F500), analize spektralne refleksije i vegetacijskih indeksa multispektralnim kamerama (CropReporter), analize svjetlosnih reakcija fotosinteze odnosno klorofilne fluorescencije (CropReporter), analize izmjene plinova (tamnih reakcija fotosinteze i transpiracije) (LI6800), mjerenje prinosa i dr.
55 POLJOPRIVREDA
2017 9833 156 089 15.9 2018 9272 182 261 19.7 2019 9387 173 149 18.4 2020 9325 174 279 18.7 2021 8786 127 826 14.5
Godina Ukupna površina (ha) Ukupna proizvodnja (t) Prosječni prinos (t/ha)
2. Utjecaj temperature na razvoj krumpira
Provedena istraživanja ukazuju da visoke temperature negativno utječu na rast i razvoj krumpira, na fiziološke procese te u konačnici smanjuju prinos i kvalitetu gomolja. Morfološka svojstva krumpira prikazana su slikom 1.
U nastavku iznosimo glavne zaključke istraživanja utjecaja visokih temperatura na krumpir.
Utjecaj temperature na morfološka svojstva krumpira:
Visoke temperature različito utječu na rast i razvoj pojedinih organa te mijenjaju morfološka svojstva biljke krumpira.
• Temperature više od 25 °C značajno remete razvoj stolona.
• Optimalna temperatura za rast korijena je 20 °C, dok kod viših temperatura dolazi do slabijeg transporta asimilata (šećera) iz lista u korijen zbog čega korijen zaostaje u rastu.
• Optimalna temperatura za zametanje listova je 28 °C, dok je optimalna temperatura za rast lista i za trajnost lisne površine 25 °C.
• Visoke temperature potiču izduživanje stabljike (dužinski rast) i rast postranih grana (grananje biljaka krumpira).
• Stoga su biljke krumpira pri višim temperaturama (iznad 25 °C) više, uz veći broj postranih grana, ali s manjim lišćem i manjom lisnom površinom (Slika 2, Grafikon 1).
• Takav habitus nadzemnih organa lošije pokriva tlo i smanjuje primanje svjetlosne energije.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 56
Slika 1. Morfologija biljke krumpira
u
uzgajanih pri različitim temperaturnim tretmanima (20/15 °C (T1), 25/20 °C (T2), 30/25 °C (T3), 35/30 °C (T4)). Različita mala podebljana slova označavaju statistički značajne razlike između srednjih vrijednosti temperaturnih tretmana.

Utjecaj temperature na fiziološka svojstva krumpira:
Optimalna temperatura usjeva krumpira za proces fotosinteze je 24 °C. Visoke temperature ubrzavaju starenje lista, smanjuj sadržaj fotosintetskih pigmenata, ubrzavaju disanje i fotorespiraciju, oštećuju fotosustave i smanjuju intenzitet fotosinteze. Međutim biljka ima mogućnost aklimatizacije na visoke temperature te je naše istraživanje pokazalo da:
• Povišenjem temperature s 20 °C raste refleksija listova u zelenom, crvenom i plavom dijelu spektra, što ukazuje na lošiju apsorpciju fotosintetski aktivne radijacije.
• Temperature iznad 25 °C uzrokuju smanjenje sadržaja antocijana, a temperature iznad 30 °C smanjenje sadržaja klorofila (Slika 4, Grafikon 2).
• Temperature iznad 25 °C uzrokuju smanjenje broja fostosustava (PSII), no fotokemijske reakcije su narušene tek kod temperatura od 35 °C (Slika 5, Grafikon 3).
• Suprotno literaturnim navodima, s porastom temperature od 20 do 30 °C raste intenzitet fotosinteze što ukazuje na sposobnost aklimatizacije biljaka na visoke temperature (Grafikon 4).
• No osim porasta fotosinteze još je izraženiji porast provodljivosti puči i transpiracije zbog čega se smanjuje efikasnost korištenja vode (Grafikon 4).
• Naprednim statističkim analizama odabrana su svojstva koja mogu poslužiti kao markeri temperaturnog stresa i omogućuju razlikovanje biljka uzgajanih pod različitim temperaturnim uvjetima.
Ta svojstva su: lisna površina, sadržaj antocijana u listu, minimalna klorofilna fluorescencija i efikasnost korištenja vode.
57 POLJOPRIVREDA
Slika 2. Utjecaj temperature na morfologiju krumpira. Krumpir uzgajan pri 20/15 °C (T1), 25/20 °C (T2), 30/25 °C (T3), 35/30 °C (T4). Biljke su naklijane pri 20/15 °C te su nakon razvijena prva dva lista izložene navedenim temperaturnim tretmanima (T1-T4). Slike su dobivene skeniranjem biljaka izloženih temperaturnim tretmanima u trajanju od 10 dana, skeniranje je obavljeno 3D multispektralnim skenerom PlantEye F500.
Grafikon 1. Rezultati analize visine biljaka i lisne površine sorata korištenih
pokusu,
Grafikon 2. Rezultati analize vegetacijskih indeksa (sadržaj antocijana (ARI) i sadržaj klorofila (CHI)) sorata korištenih u pokusu, uzgajanih pri različitim temperaturnim tretmanima (20/15 °C (T1), 25/20 °C (T2), 30/25 °C (T3), 35/30 °C (T4). Različita mala podebljana slova označavaju statistički značajne razlike između srednjih vrijednosti temperaturnih tretmana.
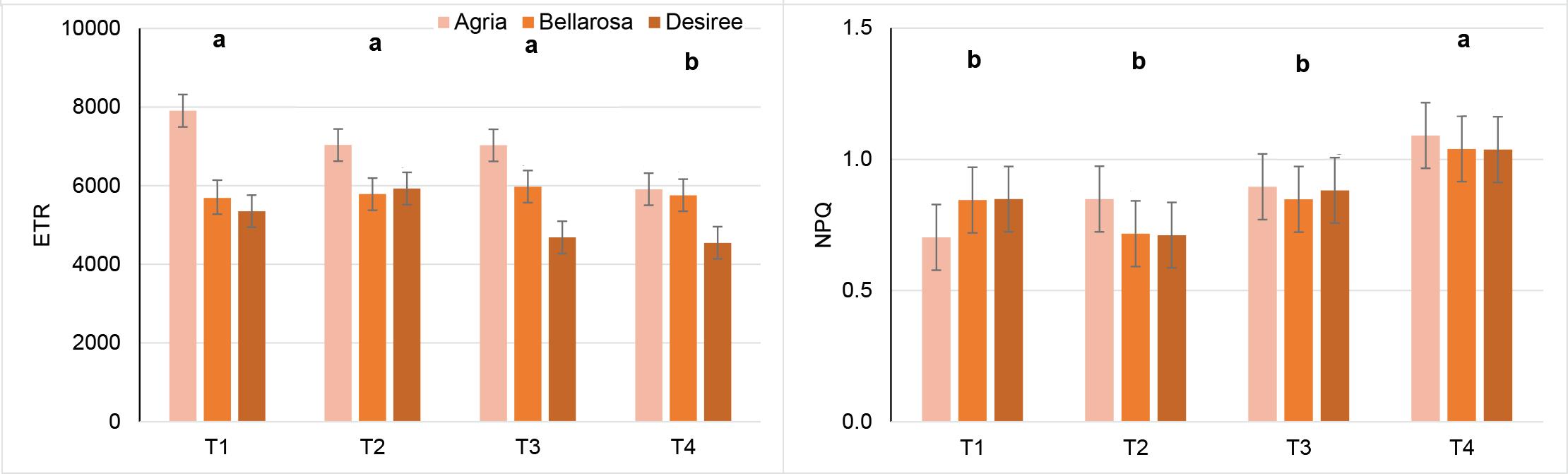

Slika 3. Primjeri analiziranih vegetacijskih indeksa (sadržaja antocijana (ARI) i sadržaja klorofila (CHI)) te slika u boji (Color) biljaka krumpira uzgajanih pri 20/15 °C (T1) i 35/30 °C (T4). Biljke su snimljene CropReporter uređajem.
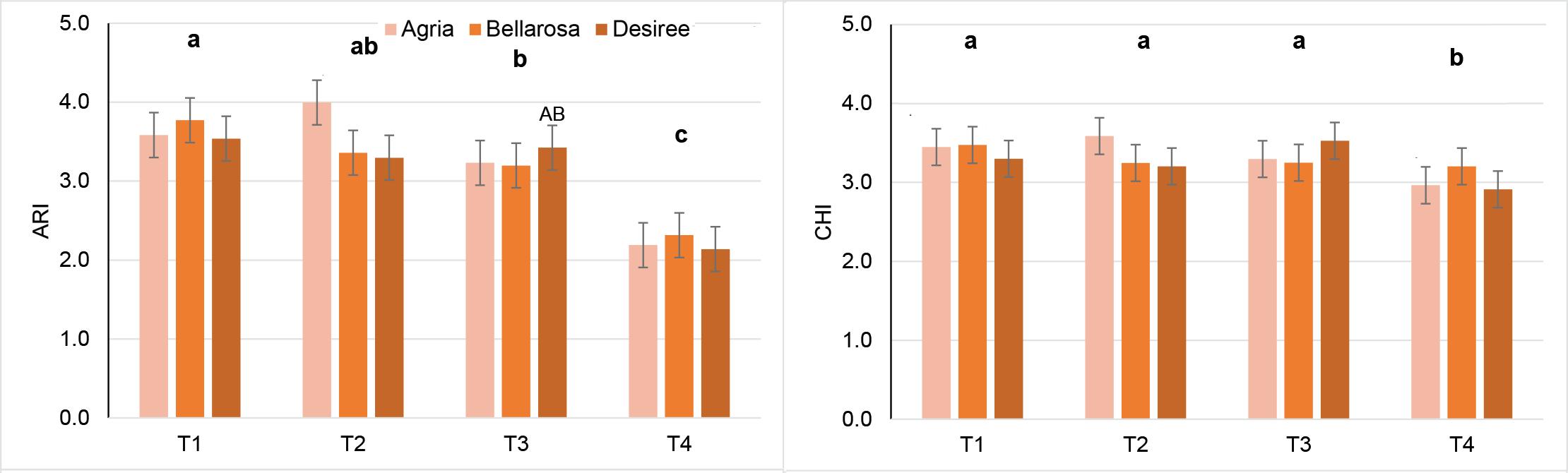
(20/15
(NPQ))
(T1), 25/20
pri
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 58
Grafikon 3. Rezultati analize klorofilne fluorescencije (parametara svjetlosnih reakcija fotosinteze: brzina transporta elektrona (ETR) i nefotokemijskog čišćenja
kod sorata krumpira korištenih u pokusu, uzgajanih
različitim temperaturnim tretmanima
°C
°C (T2), 30/25 °C (T3), 35/30 °C (T4). Različita mala podebljana slova označavaju statistički značajne razlike između srednjih vrijednosti temperaturnih tretmana.
analiziranih parametara klorofilne fluorescencije (efektivna efikasnost fotosustava (Fq’/Fm’), brzina transporta elektrona (ETR) nefotokemijsko čišćenje (NPQ) biljaka krumpira uzgajanih pri 20/15 °C (T1) i 35/30 °C (T4). Biljke su snimljene CropReporter uređajem.
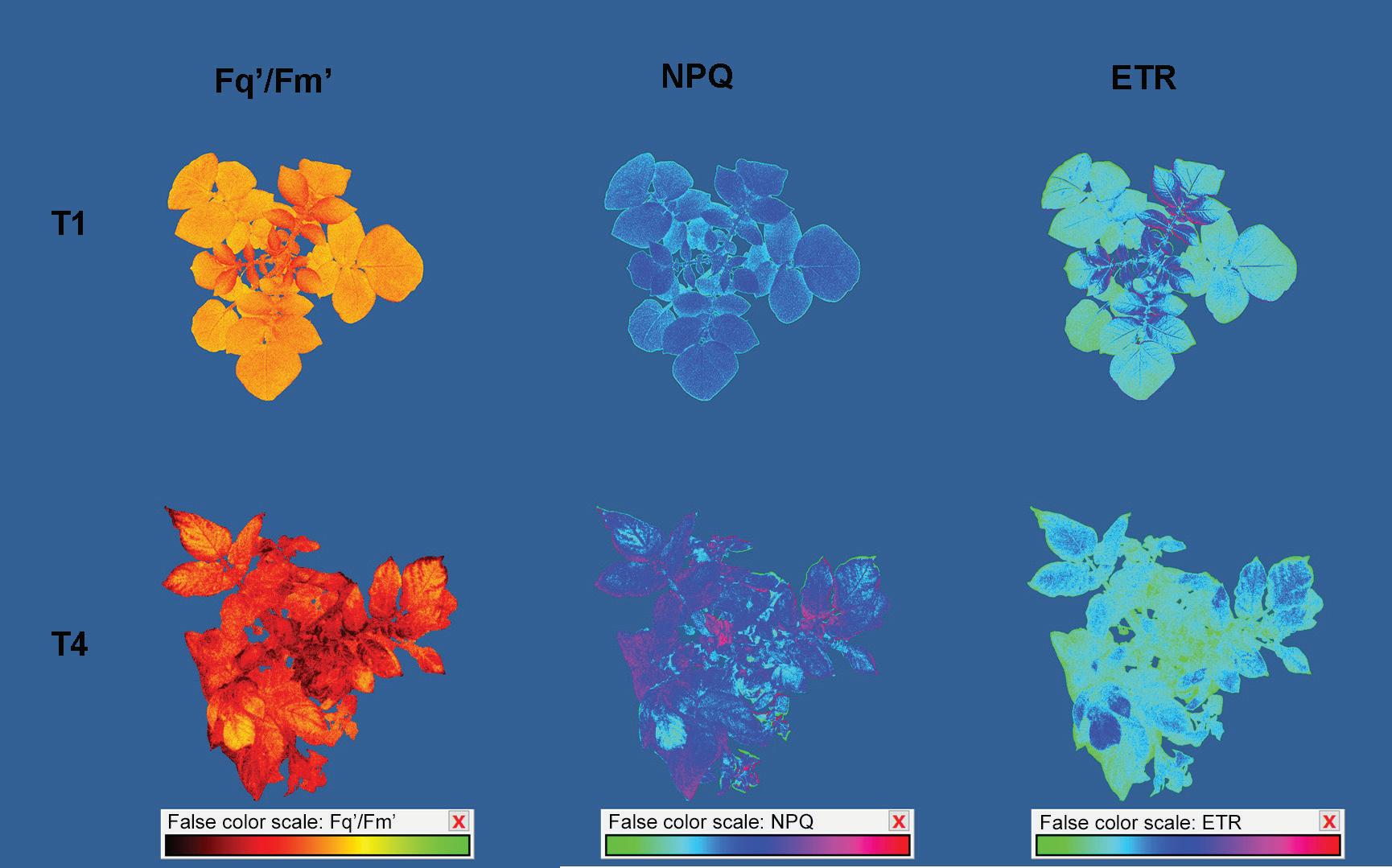
Grafikon
Rezultati analize izmjene plinova (intenzitet fotosinteze (A) i intenzitet transpiracije (E) kod sorata krumpira korištenih u pokusu, uzgajanih pri različitim temperaturnim tretmanima (20/15 °C (T1), 25/20 °C (T2), 30/25 °C (T3), 35/30 °C (T4). Različita mala podebljana slova označavaju statistički značajne razlike između srednjih vrijednosti temperaturnih tretmana.
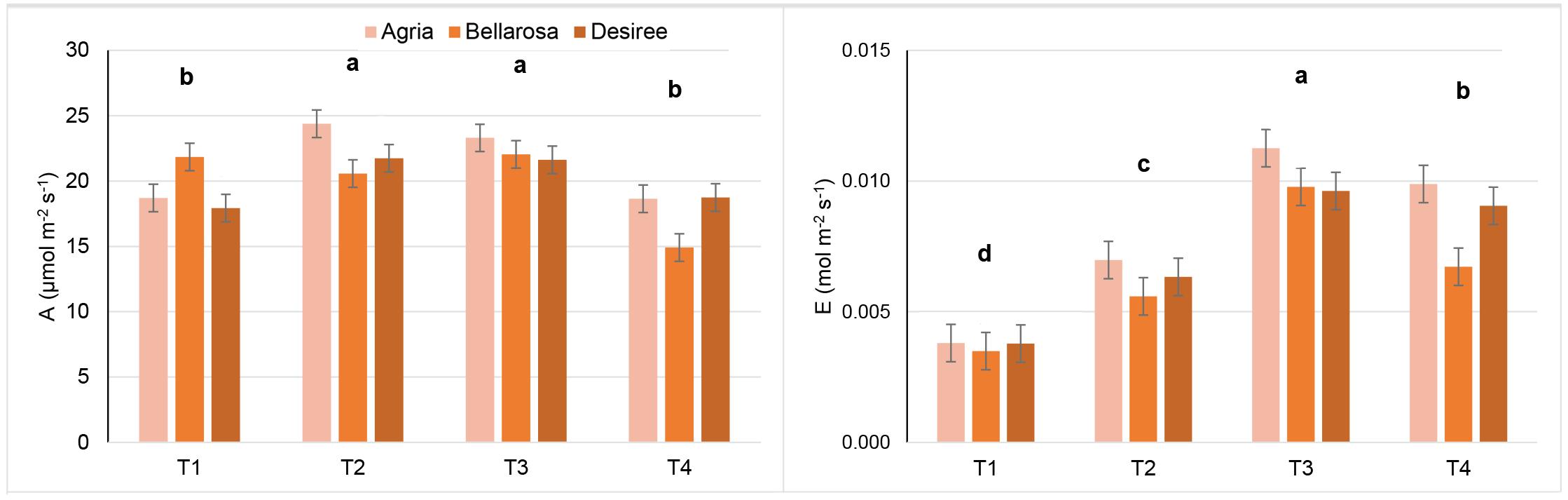
59 POLJOPRIVREDA
Slika 4. Primjeri
4.
Utjecaj vremena sadnje i navodnjavanja na prinos krumpira:
• Optimalna temperatura tla za razvoj gomolja iznosi 15 °C, a s porastom temperature tla s 15 na 23 °C usporava se inicijacija i zametanje gomolja te nalijevanje gomolja.
• Temperature iznad 30 °C uzrokuju prekid nalijevanja gomolja (prijenosa šećera u gomolj), izazivaju pojavu smeđih mrlja u gomolju te skraćuju period mirovanja gomolja (dormantnost) što izaziva preuranjeno proklijavanje gomolja u skladištu.
Podaci naših istraživanja koja su uključivala dva roka sadnje (rani i kasni), tri sorte krumpira (Agria, Bellarosa i Desiree) te navodnjavanje i izostavljanje navodnjavanja (Slika 5) ukazuju:
• Ranija sadnja u odnosu na kasniju, neovisno o sorti ostvaruje značajno veće prosječne prinose (29.4 t/ha rana u usporedi s 25.4 t/ha u kasnoj sadnji).
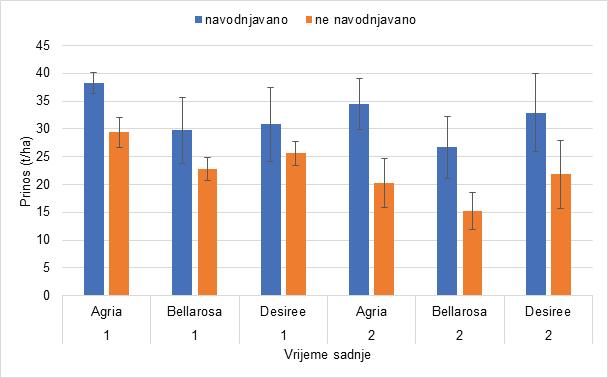
• Navodnjavanje kod svih sorti značajno povećava prosječne prinose (32.1 t/ha kod navodnjavanog u odnosu na 22.5 t/ha kod nenavodnjavanog) (Grafikon 5).
Grafikon 5.Prosječni prinosi krumpira i standardne devijacije prinosa, s poljskih pokusa za tri istraživane sorte (Agria, Bellarosa i Desiree) sađene u dva roka (rani i kasni) te uz primjenu navodnjavanja ili bez navodnjavanja.

Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 60
Slika 5. Poljski pokus s dva roka sadnje (rani i kasni), tri sorte krumpira (Agria, Bellarosa i Desiree) te navodnjavanje i izostavljanje navodnjavanja postavljen u Lovincu 2022.
3. Mjere prilagodbe
Svakako temelj svake uspješne proizvodnje krumpira jest korištenje zdravog, certificiranog sadnog materijala (gomolja), no ovdje ćemo se osvrnuti na moguće mjere ublažavanja temperaturnog stresa u uzgoju krumpira. Osim što se mora boriti protiv klimatskih promjena te smanjiti emisije stakleničkih plinova, poljoprivredna proizvodnja se mora prilagoditi novonastalim, često nepovoljnim, klimatskim uvjetima koji se javljaju zbog klimatskih promjena. Metode prilagodbe na visoke temperature uključuju selekciju novih sorata krumpira otpornih na visoke temperature, promjene proizvodnih područja, primjenu navodnjavanja i povećanje efikasnosti korištenja vode, i dr.
U nastavku ćemo izložiti neka područja u kojima se može ostvariti značajan napredak, a time i povećati otpornost na klimatskim promjenama izazvane temperaturne stresove.
1. Oplemenjivanje - tolerantne sorte
Krumpir je visoko prihodovna kultura i postoji nekoliko vrlo razvijenih oplemenjivačkih centara za krumpir. Selekcijski i oplemnjivački posao je dugotrajan i skup no s razvojem novih molekularnih metoda oplemenjivački proces se ubrzava i stvaraju se nove sorte prilagođene specifičnim agroklimatskim uvjetima, pa tako i sorte s povećanom otpornošću na visoke temperature.
2. Promjena proizvodnih područja
Ova praksa još nije zaživjela na području Republike Hrvatske i za nju još uvijek nema potrebe, no s porastom temperatura, neka od tradicionalno pogodnih područja za uzgoj krumpira morat će preći na druge kulture, dok će se uzgoj krumpira premještati sjevernije i na više nadmorske visine. Osim samih dnevnih temperatura, u uzgoju krumpira štetni utjecaj imaju i visoke noćne temperature. Stoga su pogodna područja za uzgoj krumpira u Hrvatskoj područja na višim nadmorskim visinama, gdje hladne noći omogućuju hlađenje tla što pogoduje razvoju korijena, stolona, zametanju i nalijevanju gomolja. Naša područja Like i Gorskog kotara predstavljaju pogodna područja za uzgoj krumpira, gdje se samo u ljetnim mjesecima mogu očekivati visoke dnevne temperature koje mogu negativno djelovati na produktivnost krumpira. U tim područjima relativno hladne noći omogućuju hlađenje tla što pozitivno utječe na rast i razvoj gomolja.
3. Jesenska priprema tla i veći humci
U nekim državama prisutna je praksa jesenske potpune pripreme tla za sadnju krumpira, a krumpir se sadi na proljeće. Ova priprema tla uključuje stvaranje humaka (koji odgovaraju širini od 3-5 budućih redova krumpira) (Slika 6.). Tako pripremljeno tlo se brže prosušuje u proljeće i može se obaviti ranija sadnja krumpira. No ovakav pristup mijenja tradicionalan način pripreme tla za sadnju krumpira koji se primjenjuje u Hrvatskoj.
61 POLJOPRIVREDA
Nadalje, modifikacija veličine humka u koji se sade krumpiri odnosno debljine sloja tla koji se koristi za ogrtanje može ublažiti učinak visokih temperatura. Ovo se može postići stvaranjem većih humaka ili višekratnim ogrtanjem krumpira. Veća količina tla kojom se ogrće krumpir djeluje kao izolacijski materijala i sniziti temperaturu te na taj način zaštiti stolone i gomolje od visokih temperatura.
4. Ranija sadnja ili sadnja naklijalih gomolja
Kao što je navedeno, najveći problem u zametanju gomolja te kasnijem rastu i nalijevanju gomolja su visoke temperature (osobito iznad 25 °C). Takve temperature kod nas nastupaju krajem proljeća i početkom ljeta. U takvim uvjetima ranija sadnja ima prednost pred kasnijim rokovima sadnje, jer omogućuje biljkama da razviju veću biomasu do vremena pojave visokih temperatura. Veći korijenov sustav u takvim uvjetima omogućuje bolje primanje vode, a veća nadzemna biomasa omogućuje bolje zasjenjivanje i manje gubitke vode evaporacijom.

Krumpir se može saditi kad temperatura tla na dubini 10 cm postigne 6-8 °C. Rana sadnja nosi rizik sadnje u hladno tlo, te ako nakon sadnje nastupi period niskih temperatura može doći do usporavanja klijanja i nicanja te pojačanog napada bolesti (npr. rizoktonija).
Jedna od mogućnosti kojom se postiže ubrzan početni razvoj usjeva i postizanje veće biomase do nastupa visokih temperatura jest naklijavanje gomolja prije sadnje. Može se provoditi samo djelomično naklijavanje (tzv. mini-naklijali gomolji) kada gomolji puste male žuto-zelene klice (tek što su krenule) ili se mogu koristiti stvarno naklijali gomolji kod kojih su klice zelene i čvrste, dugačke 1 do 2 cm.

5. Malčiranje
Primjena malčiranja (pokrivanje tla organskim materijalima) može umanjiti stres visokih temperatura. Zabilježeni su pozitivni učinci malča od slame, rižine slame, trave, lišća i sl. materijala (Slika 7). Istraživanja ukazuju da krumpir uzgajan pod malčem u usporedbi s ne malčiranim površinama ima značajno veći indeks lisne površine, učinkovitost korištenja vode, primanje fotosintetski aktivnog zračenja te u konačnici prinos gomolja. Temperatura tla ispod malča za je za 4– 6 °C niža u odnosu na ne malčirano tlo, a malč pomaže i zadržavanju vlage tla.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 62
Slika 6. Jesenska priprema humaka za sadnju krumpira u proljeće.
Slika 7. Upotreba malča od slame u uzgoju krumpira
6. Navodnjavanje i povećanje efikasnosti korištenja vode
Klimatske promjene uz povećanje temperatura često donose i problem duljih sušnih perioda. Usjevi dobro opskrbljeni vodom imaju veće mogućnosti hlađenja transpiracijom i na taj način bolje podnose visoke temperature. Većina sorata krumpira ima relativno slabo razvijen, plitak korijenov sustav. Zbog toga je važno uskladištiti vlagu u tlu tijekom zime. Dobrom pripremom tla i gnojidbom omogućiti rani razvoj velikog korijenovog sustava koji će omogućiti bolje usvajanje vode, ali i razvoj nadzemne biljne mase koja brzo zatvara redove i na taj način zasjenjuje tlo i smanjuje gubitke vode evaporacijom.
Navodnjavanje daje vrlo dobre rezultate u smanjenju negativnog utjecaja suše i temperaturnog stresa na prinos krumpira. Navodnjavanjem se može utjecati i na raspored gomolja. Naime, u uvjetima veće vlažnosti gomolji se zameću na većoj udaljenosti od matične biljke, dok se u sušnijim uvjetima gomolji nalaze bliže „majčinskom“ gomolju. Obzirom na sve veću nestašicu vode te potrebe uštede energije, trebalo bi težiti pametnim metodama navodnjavanja koje uključuju senzore za mjerenja vlage tla te meteorološke stanice za izračun evapotranspiraicje, kako bi se optimizirao učinak navodnjavanja, a smanjili troškovi.
Nadalje vrlo je bitna modernizacija u smislu precizne poljoprivrede koja integrira veliki broj klimatoloških i pedoloških podataka kako bi se optimizirala primjena gnojiva, sredstava za zaštitu bilja i navodnjavanje vrlo je. Moderni poljoprivredni proizvođači se više ne zadovoljavaju informacijama s najbliže meto postaje, već na svojim poljima prate mikroklimatske podatke u stvarnom vremenu. Sve je veći broj i onih koji dronovima ili satelitskim snimkama mapiraju svoja polja i na taj način upravljaju agrotehničkim mjerama poput obrade, zaštite, gnojidbe i navodnjavanja. U Kanadi, primjerice, koriste variable rate irrigation (VRI) sustav koji kontrolira navodnjavanje i aplicira vodu tamo gdje je potrebno i u potrebnim količinama, dok se vlažna mjesta ne navodnjavaju.
7. Praćenje pojave bolesti i štetnika
Klimatske promjene, osobito porast temperature pogoduje povećanju brojnosti i ubrzavanju razvoja bolesti i štetnika poljoprivrednih kultura. S toplijim zimama sve je veća brojnost jedinki koje prezimljuju, a s ranim porastom temperature u proljeće ranije počinju životni ciklusi bolesti i štetnika. Nadalje više temperature tijekom vegetacije ubrzavaju njihov razvoj, zbog šega dolazi do povećanog broja generacija i većih populacija te samim time većih šteta na poljoprivrednim kulturama.
Ovi problemi se ne odnose samo na izravne štetnike i bolesti krumpira, već i na pojavu i brojnost lisnih uši koje su važni prijenosnici virusa krumpira. Isto tako promjene klime utjecati će na promjenu prevladavajućih sojeva bolesti zbog čega će se mjere zaštite morati prilagoditi novim zahtjevima.
U tom smislu morat će se uvesti metode praćenja i predviđanja pojave bolesti i štetnika kao temelj za provođenje integrirane zaštite bilja.
8. Hlađenje skladišta
U fazi uzgoja krumpira u polju cilj nam je ostvariti maksimalne prinose. Nakon berbe krumpira u fazi skladištenja cilj nam je smanjiti gubitke i zadržati kvalitetu. S porastom temperatura, osobito jesenskih, kada se obavlja berba krumpira, povećavaju se gubici u uskladištenom krumpiru. Kod nas su vrlo rijetka skladišta s mogućnošću kontrole temperature i atmosfere u skladištu. Investiranje u skladišta s mogućnošću hlađenja krumpira i kontrolom atmosfere biti će važna mjera prilagodbe klimatskim promjenama.
63 POLJOPRIVREDA
Literatura
FAO. 2020. FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/SCL (accessed on 23 November 2022).
Hijmans, R.J. The Effect of Climate Change on Global Potato Production. Am. J. Potato Res. 2003, 80, 271–279. https://doi.org/10.1007/BF02855363.
Birch, P.R.J.; Bryan, G.; Fenton, B.; Gilroy, E.M.; Hein, I.; Jones, J.T.; Prashar, A.; Taylor, M.A.; Torrance, L.; Toth, I.K. Crops That Feed the World 8: Potato: Are the Trends of Increased Global Production Sustainable? Food Secur. 2012, 4, 477–508. https://doi.org/10.1007/s12571-012-0220-1.
FAO. 2020. Available online: http://www.fao.org/3/ca9692en/online/ca9692en.html#chapter-1_1 (accessed on 20 September 2022).
https://podaci.dzs.hr/hr/podaci/poljoprivreda/ - pristupljeno 15/12/2022
2021 Tied for 6th Warmest Year in Continued Trend, NASA Analysis Shows|NASA. Available online: https://www.nasa.gov/press-release/2021-tied-for-6th-warmest-year-in-continued-trendnasa-analysis-shows (accessed on 23 November 2022).
IPCC. Summary for Policymakers. In Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1–30.
Levy, D.; Veilleux, R.E. Adaptation of Potato to High Temperatures and Salinity—A Review. Am. J. Potato Res. 2007, 84, 487–506. https://doi.org/10.1007/BF02987885.
Struik, P.C. Responses of the Potato Plant to the Temperature. In Potato Biology and Biotechnology; Vreugdenhil, D., Bradshaw, J., Gebhardt, C., Govers, F., Taylor, M., MacKerron, D., Ross, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 367–393.
Fleisher, D.H.; Timlin, D. Modeling Expansion of Individual Leaves in the Potato Canopy. Agric. Meteorol. 2006, 139, 84–93. https://doi.org/10.1016/j.agrformet.2006.06.002.
Lee, Y.H.; Sang, W.G.; Baek, J.K.; Kim, J.H.; Shin, P.; Seo, M.C.; Cho, J.I. The Effect of Concurrent Elevation in CO2 and Temperature on the Growth, Photosynthesis, and Yield of Potato Crops. PLoS ONE 2020, 15, e0241081. https://doi.org/10.1371/journal.pone.0241081.
Yamori, W.; Hikosaka, K.; Way, D.A. Temperature Response of Photosynthesis in C3, C4, and CAM Plants: Temperature Acclimation and Temperature Adaptation. Photosynth. Res. 2014, 119, 101–117. https://doi.org/10.1007/s11120-013-9874-6.
Prange, R.K.; Mcrae, K.B.; Midmore, D.J.; Deng, R. Reduction in Potato Growth at High Temperature: Role of Photosynthesis and Dark Respiration. Am. Potato J. 2008, 67, 357–369.
Midmore, D.J. Potato (Solanum spp.) in the Hot Tropics I. Soil Temperature Effects on Emergence, Plant Development and Yield. Field Crops Res. 1984, 8, 255–271. https://doi.org/10.1016/0378-4290(84)90073-X.
64
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima
Benoit, G.R.; Stanley, C.D.; Grant, W.J.; Torrey, D.B. Potato Top Growth as Influenced by Temperatures. Am. Potato J. 1983, 60, 489–501. https://doi.org/10.1007/BF02877320.
Ewing, E.E.; Struik, P.C. Tuber Formation in Potato: Induction, Initiation, and Growth. Hortic. Rev. (Am. Soc. Hortic. Sci.) 2010, 14, 89–198.
Hancock, R.D.; Morris, W.L.; Ducreux, L.J.M.; Morris, J.A.; Usman, M.; Verrall, S.R.; Fuller, J.; Simpson, C.G.; Zhang, R.; Hedley, P.E.; et al. Physiological, Biochemical and Molecular Responses of the Potato (Solanum Tuberosum L.) Plant to Moderately Elevated Temperature. Plant Cell Environ. 2014, 37, 439–450. https://doi.org/10.1111/pce.12168
Dahal, K.; Li, X.Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving Potato Stress Tolerance and Tuber Yield under a Climate Change Scenario—A Current Overview. Front. Plant Sci. 2019, 10, 563
65 POLJOPRIVREDA
Klimatske promjene i BIORAZNOLIKOST


Izvođač: Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu
Istraživač, voditelj projektnog modula: doc. dr. sc. Andreja Brigić, Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu

Istraživač 2: prof. dr. sc. Renata Matoničkin Kepčija, Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu
Suradnik na projektu: dr. sc. Fran Rebrina, Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu
Suradnici: dr. sc. Ana Ješovnik, Zavod za zaštitu okoliša i prirode, Ministarstvo gospodarstva i održivog razvoja
dr. sc. Jana Bedek, Institut Ruđer Bošković
dr. sc. Vedran Šegota, Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 66
1. UVOD
Pojam biološka raznolikost podrazumijeva varijabilnost živog svijeta na svim razinama – od molekula (gena) do ekosustava, uključujući sve evolucijske, ekološke i ostale procese u kojima sudjeluju živa bića (Naeem i sur., 2016). Biološka raznolikost je od izuzetne važnosti za dobrobit ljudskog društva jer se nalazi u izravnoj vezi s funkcijama ekosustava, o kojima ovise brojne usluge koje pružaju zdravi ekosustavi, poput pročišćavanja zraka i vode, oprašivanja, obrane od prirodnih katastrofa i recikliranja hranjivih tvari (Balvanera i sur., 2017). Klimatske promjene, ubrzane povećanim otpuštanjem stakleničkih plinova uslijed ljudskih aktivnosti, danas predstavljaju jednu od najvećih prijetnji biološkoj raznolikosti na svim razinama organizacije (Bellard i sur., 2012). Procjenjuje se da je od sredine 19. st. do 2005. godine prosječna globalna temperatura porasla za oko 0,8°C, a predviđa se i daljnji porast između 1,4°C i 5,8°C do 2100. godine (SCBD: https://www.cbd.int/climate/intro.shtml).
S jačanjem klimatskih promjena dolazi do sve većeg pritiska na šumske ekosustave zbog povećane učestalosti ekstremnih i teško predvidivih događaja poput jakih suša, požara i razornih oluja, što zahtijeva brzu prilagodbu u načinima gospodarenja šumskim ekosustavima, s ciljem ublažavanja negativnih posljedica (FAO, 2013). Iako šumski požari manjih razmjera mogu povećati heterogenost staništa te za posljedicu imati povećanje lokalne biološke raznolikosti (Bhaskar i sur., 2019; Moretti i sur., 2004), požari većih razmjera, koji postaju sve češći i snažniji u sušnijoj i toplijoj klimi (Mansoor i sur., 2022), mogu uzrokovati značajne promjene unutar šumskog matriksa. Takvi požari predstavljaju osobitu ugrozu za brojne vrste šumskih specijalista s ograničenom sposobnošću širenja areala (Moretti i sur., 2004). Slične negativne posljedice imaju sve učestaliji ledolomi i vjetroizvale u šumskim staništima; prema nekim procjenama, od ledoloma je samo u 2014. godini oštećeno ili uništeno čak 16,24% listopadnih, 24,32% crnogoričnih te 54,83% mješovitih šuma u Republici Hrvatskoj (Šimić Milas i sur., 2015).
Cretovi, kao specifičan tip močvarnih staništa uključenih u Dodatak I Europske direktive o staništima (Eionet, 2018), na prostoru Republike Hrvatske kritično su ugroženi sukcesijom vegetacije i hidrološkim promjenama uslijed napuštanja tradicionalnog načina gospodarenja i nepovoljnih klimatskih uvjeta (Alegro i Topić, 2017; Brigić i sur., 2017). S obzirom da izravno ovise o visini podzemnih voda i količini oborina, promjene hidrološkog režima cretnih staništa omogućuju efikasno praćenje utjecaja klimatskih promjena (Joosten i sur., 2012). Dosadašnja istraživanja unutar sektora Prirodni ekosustavi i bioraznolikost u Hrvatskoj ukazuju na ubrzano smanjenje cretnih površina (Topić i Stančić, 2006), pri čemu su primjerice na cretu Dubravica u 55 godina potpuno nestale dvije manje cretne površine, a najveći cret je sveden na 37% izvorne površine (Hršak, 1996). Rezultat toga su lokalna izumiranja rijetkih i ugroženih cretnih vrsta (Bujan i sur., 2015; Topić i Stančić, 2006).
Iako već odgovori pojedinih vrsta mogu ukazati na promjene u okolišu, ne smijemo zanemariti činjenicu da su u prirodi vrste međusobno povezane nizom izravnih i neizravnih interakcija, primjerice trofičkim odnosima ili natjecanjem za resurse, te tvore biološke zajednice (Walther, 2010). Prema tome, svaka promjena na razini vrste može dovesti do promjene u ekološkim interakcijama, što će u konačnici imati posljedice na funkcioniranje zajednica i ekosustava u cjelini (Caddy-Retalic i sur., 2019). Stoga su primijenjena istraživanja na razini zajednica pojedinih indikatorskih skupina nužna za potpunije razumijevanje posljedica klimatskih promjena na biološku raznolikost šumskih odnosno cretnih ekosustava (Walther, 2010). Međutim, sustavna istraživanja utjecaja klimatskih promjena na biološku raznolikost ovih ekosustava u Republici Hrvatskoj općenito su malobrojna, a na razini zajednica gotovo u potpunosti izostaju.
U današnje vrijeme, klimatske promjene su jedan od vodećih uzroka smanjenja brojnosti i raznolikosti terestričkih člankonožaca (Deutsch i sur., 2008). Trčci (Coleoptera: Carabidae), mravi (Hymenoptera: Formicidae), ravnokrilci (Orthoptera) i kopneni jednakonožni rakovi (Isopoda: Oniscidea) važne su sa-
67 BIORAZNOLIKOST
stavnice terestričkih ekosustava zbog velike brojnosti i biomase te sudjelovanja u mnogim važnim ekološkim procesima i interakcijama (Hochkirch i sur., 2016; Hölldobler i Wilson, 1990; Lövei i Sunderland, 1996; Magrini i sur., 2011). Zbog visoke osjetljivosti na promjene u okolišu, ove se skupine člankonožaca često koriste kao bioindikatori, kako na razini vrste, tako i na razini zajednice (Angulo i sur., 2016; Antonović i sur., 2012; Brigić i sur., 2017; Fartmann i sur., 2012; Rainio i Niemela, 2003). Posljedice klimatskih promjena na nekom staništu moguće je detektirati i na mikroskopskoj razini; mikroorganizmi tla poput fagotrofnih praživotinja (Protozoa) također se često koriste kao bioindikatori zbog velike raširenosti, brojnosti te osjetljivosti na okolišne promjene (Payne, 2013).
U sklopu ovog projekta, unutar sektora Prirodni ekosustavi i bioraznolikost, provedeno je primijenjeno istraživanje s dva glavna cilja:
(1) Procijeniti ranjivost šumskih i cretnih ekosustava na klimatske promjene, na temelju odgovora odabranih indikatorskih skupina terestričkih organizama (Carabidae, Formicidae, Orthoptera, Isopoda, Protozoa), uslijed izravnih i neizravnih posljedica klimatskih promjena (požari, ledolomi i vjetroizvale u šumskim staništima, sukcesijske i hidrološke promjene na cretnom staništu).
(2) Temeljem rezultata pod (1), predložiti sustav za praćenje stanja (monitoring) biološke raznolikosti šumskih i cretnih ekosustava te definirati konkretne mjere prilagodbe klimatskim promjenama, s konačnim ciljem povećanja otpornosti ovih ekosustava na klimatske promjene te očuvanja usluga koje ti ekosustavi pružaju.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 68
2. MATERIJALI I METODE
2.1. Terensko istraživanje 1 - Utjecaj požara na biološku raznolikost šumskih ekosustava
2.1.1. Područje istraživanja
Istraživanje je provedeno na području Nacionalnog Parka Sjeverni Velebit, koji s površinom od 10.900 ha čini 5,35% područja Velebita (Javna ustanova Park prirode Velebit, 2017). Klimatska obilježja Velebita definirana su njegovim graničnim položajem između kontinentalnog i mediteranskog dijela Hrvatske (Perica i Orešić, 1999), pri čemu su izražene klimatološke razlike između kontinentalnijih SI obronaka te JZ (primorskih) obronaka pod većim utjecajem mediteranske klime. Prosječna godišnja temperatura s JZ strane masiva iznosi oko 10°C na 600-700 m n.v., dok je prosječna godišnja količina oborina 1.8002.000 mm (mjerna postaja Zavižan) (Zaninović i sur., 2008). Za istraživanje utjecaja požara na šumski ekosustav odabrana je lokacija u blizini prijevoja Veliki Alan duž ceste Krasno-Jablanac na primorskim obroncima Velebita, unutar primorske šume bukve (Seslerio autumnali-Fagetum M. Wraber ex Borhidi 1963) (Bardi i sur., 2016) na oko 1.100-1.200 m n.v. (Slika 1a). Lokacija obuhvaća dio požarišta Sjeverni Velebit iz 2017. godine ukupne površine 726,40 ha (Javna ustanova Park prirode Velebit, 2017)



2.1.2. Dizajn istraživanja i uzorkovanje
Istraživanje je postavljeno u dva glavna tipa staništa: 1) opožarenim staništima, gdje je uslijed požara došlo do nastanka šumskih čistina koje su danas djelomično zahvaćene sukcesijom vegetacije (Slika 1b) te 2) kontrolnim šumskim staništima, nezahvaćenima požarom (Slika 1c). Unutar svakog tipa staništa odabrano je devet postaja (replikata). Postaje su međusobno bile udaljene najmanje 150 m kako bi se osigurala neovisnost replikata (Slika 1a). Na svakoj postaji beskralježnjaci tla prikupljani su pomoću pet lovnih posuda koje su bile postavljene u linijskom transektu s međusobnom udaljenošću > 5 m. Ukupno je postavljeno 90 lovnih posuda (2 staništa x 9 postaja x 5 lovnih posuda), a svaka se sastojala od plastične čaše (vol. 500 ml) ukopane u tlo i djelomično ispunjene mješavinom antifriza i vode (omjer 3:2), u koju je dodana kap deterdženta za razbijanje površinske napetosti. Svaka lovna posuda bila je natkrivena bitumenskim krovićem u svrhu zaštite uzoraka od kiše i listinca. Na svakoj postaji također je proveden po jedan linijski transekt kečerom (promjer = 40 cm, dubina = 60 cm) duljine 30 m (1 m = 1 zamah), kojim je uzorkovana zajednica ravnokrilaca (ukupno 18 transekata po terenskom izlasku; 2 staništa x 9 postaja). Uzorci iz lovnih posuda odnosno kečera konzervirani su u 75%-tnom etanolu. Praživotinje su uzorkovane na način da su na svakoj postaji prikupljena dva uzorka tla (svaki vol. 100 ml) iz površinskog
69 BIORAZNOLIKOST
(Slika 1a)
(Slika 1c)
(Slika 1b)
sloja (7 cm dubine) s dvije točke međusobno udaljene > 5 m. Ukupno je prikupljeno 36 uzoraka po terenskom izlasku (2 staništa x 9 postaja x 2 uzorka).
Sva uzorkovanja su provođena jednom mjesečno tijekom 2021. godine, pri čemu su lovne posude pražnjene od srpnja do studenog, istovremeno s uzorkovanjem praživotinja, dok su transekti kečerom provođeni od srpnja do rujna (u vrijeme najveće aktivnosti ravnokrilaca). Pri svakom terenskom izlasku mjereni su i sljedeći okolišni čimbenici uz svaku lovnu posudu (= 5 mjerenja po postaji, iz kojih je izračunata srednja vrijednost): temperatura (ºC) i vlažnost zraka (%) (digitalnim termo-higrometrom TROTEC T200), temperatura tla (ºC) na dubini od 7 cm (digitalnim termometrom sa sondom TFA-Dostmann) te vlažnost tla (%) (higrometrom sa sondom FieldScout TDR100). Prilikom terenskog izlaska u srpnju izmjerena je i pH vrijednost tla (pH-metrom FieldScout SoilStikTM) na svakoj postaji.
Pri terenskom izlasku u kolovozu provedeno je i istraživanje vegetacije na svakoj postaji. Površina uzorkovanja je bila konstantna i iznosila je 100 m2. Većina uzorkovanih ploha imala je oblik kvadrata (10 x 10 m), dok je u iznimnim slučajevima oblik bio drugačiji, uz istu površinu. Ovako opisani pristup odabiru postaja i uzorkovanju osigurava jednak istraživački napor u svim tipovima vegetacije na sistematičan i standardiziran način. Vegetacija je na plohama uzorkovana metodom fitocenoloških snimki, što znači da su popisivane sve prisutne svojte te procijenjena njihova pokrovnost uz korištenje Braun-Blanquetove skale, proširene prema Barkman i sur. (1964) (Tablica 1).
Tablica 1. Skala po Braun-Blanquetu (proširena prema Barkmanu i sur.) za kombiniranu procjenu abundancije i pokrovnosti svojti u vegetacijskoj snimci.
r jedna jedinka u snimci
+ 2 – 5 jedinki u snimci, pokrovnost < 5%
1 6 – 50 jedinki u snimci, pokrovnost < 5%
2m >50 jedinki u snimci, pokrovnost <5%
2a pokrovnost 5 -15%, bez obzira na broj jedinki
2b pokrovnost 16 – 25%, bez obzira na broj jedinki
3 pokrovnost 26 -50%, bez obzira na broj jedinki
4 pokrovnost 51 – 75%, bez obzira na broj jedinki
5 pokrovnost 76 -100%, bez obzira na broj jedinki
Prilikom izrade snimki procijenjene su i vrijednosti sljedećih parametara na svakoj postaji: ukupna pokrovnost pojedinih slojeva vegetacije (sloj drveća, sloj grmlja, sloj prizemnog rašća i sloj mahovina) (%), otvorenost krošnji (%), površina tla pod listincem (%) te površina tla pod skeletom (skeletnost) (%). Sav biljni materijal koji nije bilo moguće identificirati na terenu prikupljen je u terenski herbarij za naknadnu identifikaciju u laboratoriju.
Također, kako bi se detaljnije procijenila otvorenost sklopa krošanja, u kolovozu 2022. godine na svakoj je postaji pomoću digitalnog fotoaparata Nikon Coolpix 8400 s FC-E9 hemisfernim objektivom (tzv. riblje oko, engl. fisheye) fotografiran sklop krošanja na pet različitih točaka međusobno udaljenih najmanje 5 m, na visini od oko 1,4 m iznad tla. Fotografiranje je provođeno u uvjetima optimalne razine prirodne svjetlosti, od oko 17:30 do 20:30 h. Otvorenost sklopa, odnosno postotak otvorenog neba (predstavljenog bijelim pikselima) vidljivog kroz krošnje stabala (crni pikseli), izračunata je u programu WinSCANOPY Pro 2006 (Regent, 2006), pri čemu je refleksija na pojedinim fotografijama uklonjena koristeći ručnu klasifikaciju piksela, kako bi se umanjila greška.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 70
2.2.
Terensko istraživanje 2 - Utjecaj ledoloma i vjetroizvala na biološku raznolikost šumskih ekosustava
2.2.1. Područje istraživanja
Ovo istraživanje je provedeno u Nacionalnom parku Risnjak i u neposrednoj okolici Parka, na području Šumarije Gerovo (UŠP Delnice), u Gorskom kotaru. NP Risnjak se prostire na ukupno 6.350 ha (Pešić, 2019), dok Šumarija Gerovo raspolaže s četiri gospodarske jedinice ukupne površine 7.168,98 ha, od čega čak 98% otpada na obraslo šumsko zemljište (Šoštarić, 2016). Istraživano područje ima umjereno toplu vlažnu klimu s toplim ljetom (Šegota i Filipčić, 2003), s prosječnom godišnjom temperaturom oko 7°C i prosječnom godišnjom količinom oborina 1.840-1.850 mm (mjerna postaja Parg) (Zaninović i sur., 2008). Istraživanje je postavljeno unutar šume bukve i jele (Omphalodo-Fagetum (Tregubov 1957) Marinček i sur. 1993) na oko 800-1.000 m n.v., na relaciji Lazac-Šegine-Btov laz unutar NP Risnjak te na relaciji Šegine-Lividraga u okviru Šumarije (Slika 2a). Na ovom je području u zimu 2013./2014. došlo do opsežnog ledoloma u šumskim staništima zbog velike količine ledene kiše i mokrog snijega u kratkom vremenskom razdoblju (Sertić, 2016; Šoštarić, 2016). Na istim je lokacijama u zimu 2017./2018. orkansko jugo uzrokovalo intenzivne vjetroizvale i djelomično vjetrolome (Pešić, 2019).
2.2.2. Dizajn istraživanja i uzorkovanje
Kako bismo ispitali utjecaj ekstremnih klimatskih događaja, ali i načina gospodarenja zahvaćenim šumskim površinama na njihovu biološku raznolikost, istraživanje je postavljeno unutar tri glavna tipa staništa: 1) staništa zahvaćena ledolomom i vjetroizvalama unutar gospodarene šume (Šumarija Gerovo; Slika 2b), 2) staništa zahvaćena istim poremećajima unutar negospodarene šume (NP Risnjak; Slika 2c) te 3) kontrolna šumska staništa, nezahvaćena ledolomom i vjetroizvalama, unutar negospodarene šume (Slika 2d). U svakom tipu staništa postavljeno je osam postaja, koje su bile međusobno udaljene barem 120 m kako bi se osigurala neovisnost replikata (Slika 2a)


Na svakoj postaji, beskralježnjaci tla su prikupljani pomoću pet lovnih posuda u linijskom transektu. Ukupno je postavljeno 120 lovnih posuda (3 staništa x 8 postaja x 5 lovnih posuda). Međutim, uzorkovanje ravnokrilaca kečerom nije provođeno jer je nakon probnog uzorkovanja u srpnju uočen gotovo potpuni izostanak vrsta koje obitavaju na vegetaciji. Praživotinje su uzorkovane na pet od osam postaja u svakom tipu staništa, s po dva uzorka po postaji (ukupno 30 uzoraka po izlasku; 3 staništa x 5 postaja x 2 uzorka). Uzorkovanje je provođeno jednom mjesečno od lipnja do listopada 2021. godine, prema metodologiji koja je detaljnije opisana u odjeljku 2.1.2. Pri svakom terenskom izlasku mjerena je temperatura (ºC) i vlažnost zraka (%), temperatura (ºC) i vlažnost tla (%), a u lipnju i srpnju izmjerena je i pH vrijednost tla te je početkom srpnja provedeno istraživanje vegetacije i procjena stanišnih parametara na svakoj postaji, prema ranije opisanoj metodologiji (vidi 2.1.2).

71 BIORAZNOLIKOST
(Slika 2b)
(Slika 2c)
(Slika 2d)
ekosustava
2.3.1. Područje istraživanja
Istraživanje utjecaja posljedica klimatskih promjena na cretni ekosustav provedeno je na cretu Trstenik u Gorskom kotaru, koji se nalazi u krškoj depresiji sjeveroistočno od mjesta Klana u zaleđu grada Rijeke, na oko 960 m n.v. (Modrić Surina, 2011; Šišić i sur., 2009) (Slika 3a). Površina predložena za zaštitu u vidu posebnog rezervata iznosi 12,5 ha (Šišić i sur., 2009), no sami cret se prostire na otprilike 5 ha površine (Modrić Surina, 2011). Područje ima umjereno toplu vlažnu klimu s toplim ljetom (Šegota i Filipčić, 2003), s prosječnom godišnjom temperaturom 6-7°C i prosječnom godišnjom količinom oborina od oko 2.700 mm (Modrić Surina, 2011). Cret Trstenik 2007. godine je uvršten u Nacionalnu ekološku mrežu Natura 2000 kao kritično ugroženi stanični tip - prijelazni acidofilni cret s elementima nadignutog creta (Randić, 2018). Središnji dio creta obrastao je sastojinama vrsta Sphagnum magellanicum Brid.-Polytrichum strictum Menzies ex Brid., dok na marginama nalazimo fragmente bazofilne cretne vegetacije sveze Caricion davallianae Klika 1934 (Alegro i Topić, 2017). Iako se nalazi u zoni šuma bukve i jele (Omphalodo-Fagetum (Tregubov 1957) Marinček i sur. 1993), zbog temperaturne inverzije cret okružuje šuma smreke (Haquetio-Piceetum Zupančič (1976) 1999) (Modrić Surina, 2011).



2.3.2. Dizajn istraživanja i uzorkovanje
Istraživanje je postavljeno unutar tri glavna tipa staništa: 1) središnji (vlažni) dio creta (Slika 3b), 2) (suhi) dio creta zahvaćen uznapredovalom sukcesijom prema sastojinama modre beskoljenke (Molinia caerulea (L.) Moench) (Slika 3c) te 3) šumski rub (Slika 3d). U svakom tipu staništa postavljeno je šest postaja (Slika 3a) s maksimalnom mogućom međusobnom udaljenošću, kako bi se osigurala neovisnost replikata.
Beskralježnjaci tla su prikupljani pomoću četiri lovne posude (vol. 350 ml) u linijskom transektu na svakoj postaji. Ukupno su postavljene 72 lovne posude (3 staništa x 6 postaja x 4 lovne posude). Na svakoj postaji, za uzorkovanje ravnokrilaca na vegetaciji proveden je po jedan linijski transekt kečerom (ukupno 18 transekata po terenskom izlasku; 3 staništa x 6 postaja) te su prikupljena po dva uzorka tla za analizu zajednice praživotinja (ukupno 36 uzoraka po izlasku; 3 staništa x 6 postaja x 2 uzorka). Uzorkovanje beskralježnjaka tla i praživotinja je provođeno jednom mjesečno od lipnja do listopada 2021. godine, dok su uzorci kečerom prikupljani jednom mjesečno od srpnja do rujna (u vrijeme najveće aktivnosti ravnokrilaca), prema metodologiji koja je detaljno opisana u odjeljku 2.1.2. Pri svakom terenskom izlasku također je mjerena temperatura (ºC) i vlažnost zraka (%), temperatura (ºC) i vlažnost tla (%), pH vrijednost tla (u lipnju i srpnju) te je početkom srpnja provedeno istraživanje vegetacije na svakoj postaji, prema ranije opisanoj metodologiji (vidi 2.1.2). Od stanišnih parametara prilikom izrade vegetacijskih snimki procijenjena je pokrovnost pojedinih slojeva vegetacije (sloj drveća, sloj grmlja i sloj prizemnog rašća) (%).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 72
2.3. Terensko istraživanje 3 - Utjecaj sukcesijskih i hidroloških promjena na biološku raznolikost cretnog
(Slika 3c) (Slika 3b)
(Slika 3d)
2.4. Laboratorijski rad i statistička obrada podataka
2.4.1. Identifikacija
Nakon svakog terenskog izlaska, uzorci iz lovnih posuda i kečera su sortirani po indikatorskim skupinama (Carabidae, Formicidae, Orthoptera, Isopoda), označeni te pohranjeni u 75%-tnom etanolu na Biološkom odsjeku Prirodoslovno-matematičkog fakulteta za daljnju obradu. Prikupljeni materijal određen je do najniže moguće taksonomske razine uz upotrebu odgovarajućih identifikacijskih ključeva za svaku skupinu: Carabidae (Freude i sur., 2006; Hůrka, 1996; Mlynář, 1977; Trautner i Geigenmüller, 1987; Turin i sur., 2003), Formicidae (Radchenko i Elmes, 2010; Seifert, 2018; AntWeb: https://www.antweb.org), Orthoptera (Bellmann i sur., 2019; Harz, 1975, 1969; Sardet i sur., 2015), Isopoda (REF). Vrste koje nije bilo moguće odrediti morfološki zbog problema u taksonomiji identificirane su DNA sekvenciranjem (Macrogen Europe B.V.).
U laboratoriju uzorci tla s praživotinjama prebačeni su u Petrijeve zdjelice unutar 24 sata od uzorkovanja, prema metodi non-flooded Petri dish (Foissner, 1992). Nakon inkubacije od 6-7 dana, izdvojeno je po 3 ml svakog uzorka te konzervirano u jednakom volumenu 4%-tne otopine formalina. U svakom uzorku praživotinje su zatim određene do najniže moguće taksonomske razine uz upotrebu odgovarajućih identifikacijskih ključeva (REF).
2.4.2. Statistička obrada podataka
Iz podataka prikupljenih svakim od tri terenska istraživanja (vidi 2.1., 2.2. i 2.3.), u programu Primer 6.0 (Clarke i Gorley, 2006) izračunati su sljedeći biocenološki parametri za svaku indikatorsku skupinu na svakoj postaji za svaki mjesec uzorkovanja: brojnost jedinki (N), bogatstvo vrsta (S), Pielou indeks ujednačenosti (J), Shannonov (H’) i Simpsonov indeks raznolikosti (1-λ). Iz dobivenih vrijednosti indeksa raznolikosti izračunata je raznolikost 1. reda (Shannon) odnosno 2. reda (Simpson) izražena u tzv. efektivnom broju vrsta (Jost, 2006). Prilikom analize zajednica ravnokrilaca, odvojeno su analizirani podaci prikupljeni lovnim posudama odnosno kečerom. S obzirom na proceduralna ograničenja (različiti volumeni uzoraka prilikom pregledavanja materijala zbog različitog isparavanja konzervansa), za praživotinje je umjesto brojnosti korišten relativni udio svake vrste u uzorku (%) kao pouzdaniji pokazatelj zastupljenosti.
Promjene u prostornoj dinamici zajednica indikatorskih organizama i okolišnih čimbenika na postajama utjecanim posljedicama klimatskih promjena u odnosu na kontrolu testirane su generaliziranim linearnim mješovitim modelima (GLMM) u programu SPSS Statistics (IBM Corp., 2013). Raspodjela podataka za svaki biocenološki odnosno okolišni parametar prethodno je testirana Shapiro-Wilk W testom. Ovisno o raspodjeli, odabran je linearni model (u slučaju normalne raspodjele), model s Poissonovom, negativnom binomnom ili gama raspodjelom (za podatke koji nisu imali normalnu raspodjelu).
Struktura podataka bila je sljedeća: stanište (npr. O = opožareno, K = kontrola) s replikatima (npr. O1, O2, O3…) je tretirano kao subjekt, a mjeseci uzorkovanja kao ponovljena mjerenja (osim za parametre koji su mjereni samo jednom tokom istraživanja). Utjecaj staništa na pojedini parametar (npr. brojnost jedinki) je bio čimbenik od interesa (fixed effect), dok su replikati i mjeseci uključeni u model kao nasumični čimbenici (random effects). Ako je model bio statistički značajan, razlike u pojedinim parametrima između staništa ispitane su pomoću testa najmanje značajne razlike (least significant difference, LSD). U svim modelima korištena je autoregresijska struktura kovarijance 1. reda (first-order autoregressive, AR1) za nasumične čimbenike te kriterij apsolutne konvergencije parametara. Rezultati su grafički prikazani u programu RStudio (RStudio Team, 2020) koristeći R paket ggplot2
73 BIORAZNOLIKOST
Utjecaj okolišnih čimbenika na prostornu raspodjelu vrsta pojedine indikatorske skupine na istraživanom području ispitan je kanoničkom analizom korespondencije (CCA) ili analizom redundancije (RDA), ovisno o duljini SD jedinica, u programu Canoco ver. 5.11 (ter Braak i Šmilauer, 2012). U analizu su uključeni okolišni parametri za koje su prethodno zabilježene statistički značajne razlike između istraživanih staništa (GLMM, p<0.05), a iz analize su izbačeni parametri koji su međusobno korelirani. Za zajednice trčaka i ravnokrilaca (bez obzira na metodu uzorkovanja), vrste zastupljene s manje od pet jedinki u ukupnom ulovu su isključene iz analize (Lepš i Šmilauer, 2014). Za zajednice mrava, nije uzeta u obzir zastupljenost nego prisutnost/odsutnost vrste na pojedinoj postaji, pri čemu su iz analize isključene vrste s jednom ili dvije jedinke u ukupnom ulovu (Longino, 2000). Za zajednice jednakonožaca iz analize su isključene vrste zastupljene s manje od deset jedinki (Risnjak) odnosno pet jedinki (Velebit) u ukupnom ulovu ili su sve vrste uključene u analizu (Trstenik). Brojnosti vrsta su transformirane po drugom korijenu (Carabidae, Formicidae), četvrtom korijenu (Orthoptera, Isopoda) ili su korištene vrijednosti bez transformacije (Protozoa) te je njihov odnos s okolišnim parametrima ispitan Monte Carlo testom s 499 (Orthoptera, Isopoda, Protozoa) ili 999 permutacija (Carabidae, Formicidae) (ter Braak i Šmilauer, 2012).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 74
3. REZULTATI
3.1. Utjecaj požara na biološku raznolikost šumskih ekosustava
3.1.1. Okolišni čimbenici
Generaliziranim linearnim mješovitim modelima utvrđene su statistički značajne razlike između opožarenih i kontrolnih postaja u sedam okolišnih čimbenika (Slika 4, 5). Opožarene postaje imale su statistički značajno višu temperaturu zraka (GLMM, F=10,839, d.f.1=1, d.f.2=88, p=0,001) i tla (F=17,784, d.f.1=1, d.f.2=88, p<0,001) te veću otvorenost sklopa krošanja (F=316,395, d.f.1=1, d.f.2=88, p<0,001) i zastupljenost prizemnog sloja vegetacije (F=4,739, d.f.1=1, d.f.2=16, p=0,045) od kontrolnih postaja (Slika 4ac, 5ac). S druge strane, neutjecane šumske površine odlikovala je statistički značajno viša vlažnost zraka (F=22,558, d.f.1=1, d.f.2=88, p<0,001), veća zastupljenost sloja drveća (F=155,102, d.f.1=1, d.f.2=16, p<0,001) i, posljedično, veći udio listinca (F=17,892, d.f.1=1, d.f.2=16, p=0,001) u odnosu na opožarene površine (Slika 4b, 5bd). Vlažnost tla (F=0,009, d.f.1=1, d.f.2=88, p=0,925), pH tla (F=4,114, d.f.1=1, d.f.2=16, p=0,060), zastupljenost sloja grmlja (F=0,002, d.f.1=1, d.f.2=16, p=0,966), skeletnost tla (F=0,046, d.f.1=1, d.f.2=16, p=0,833) i zastupljenost sloja mahovina (F=2,904, d.f.1=1, d.f.2=16, p=0,108) nisu se statistički značajno razlikovale između postaja.
Slika 4. Razlike u okolišnim čimbenicima između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit: a) temperatura zraka, b) vlažnost zraka, c) temperatura tla. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
75 BIORAZNOLIKOST
Slika 5. Razlike u okolišnim čimbenicima između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit: a) otvorenost sklopa krošanja, zastupljenost b) sloja drveća, c) prizemnog sloja vegetacije, d) udio listinca. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
3.1.2. Prostorna dinamika zajednica indikatorskih organizama
Brojnost ravnokrilaca uzorkovanih kečerom (GLMM, F=32,330, d.f.1=1, d.f.2=52, p<0,001) i jednakonožaca (F=4,487, d.f.1=1, d.f.2=88, p=0,037) razlikovala se između staništa, uz statistički značajno više vrijednosti na opožarenim nego na kontrolnim postajama u obje skupine (Slika 6). Brojnost ravnokrilaca uzorkovanih lovnim posudama nije se statistički značajno razlikovala između staništa (F=2,125, d.f.1=1, d.f.2=88, p=0,148).
Slika 6. Razlike u brojnosti jedinki: a) ravnokrilaca (Orthoptera) uzorkovanih kečerom, b) jednakonožaca (Isopoda), između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 76
Bogatstvo vrsta (F=17,912-33,200, d.f.1=1, d.f.2=52, 88, p<0,001), raznolikost 1. (F=33,236-82,100, d.f.1=1, d.f.2=52, 88, p<0,001) i 2. reda (F=63,477-120,856, d.f.1=1, d.f.2=52, 88, p<0,001) te ujednačenost (F=51,802-71,517, d.f.1=1, d.f.2=52, 88, p<0,001) zajednica ravnokrilaca također su imali statistički značajno više vrijednosti na opožarenim nego na kontrolnim postajama, bez obzira na metodu uzorkovanja (Slika 7, 8, 9bc, 10ab). Isti je statistički značajan obrazac pratila i raznolikost 2. reda (F=4,948, d.f.1=1, d.f.2=88, p=0,029) u zajednicama mrava (Slika 9a), dok je ujednačenost zajednica jednakonožaca pokazala suprotan odgovor (F=4,557, d.f.1=1, d.f.2=88, p=0,036), uz statistički značajno više vrijednosti na kontrolnim nego na opožarenim postajama (Slika 10c)
Slika 7. Razlike u bogatstvu vrsta ravnokrilaca (Orthoptera) uzorkovanih a) lovnim posudama, b) kečerom, između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
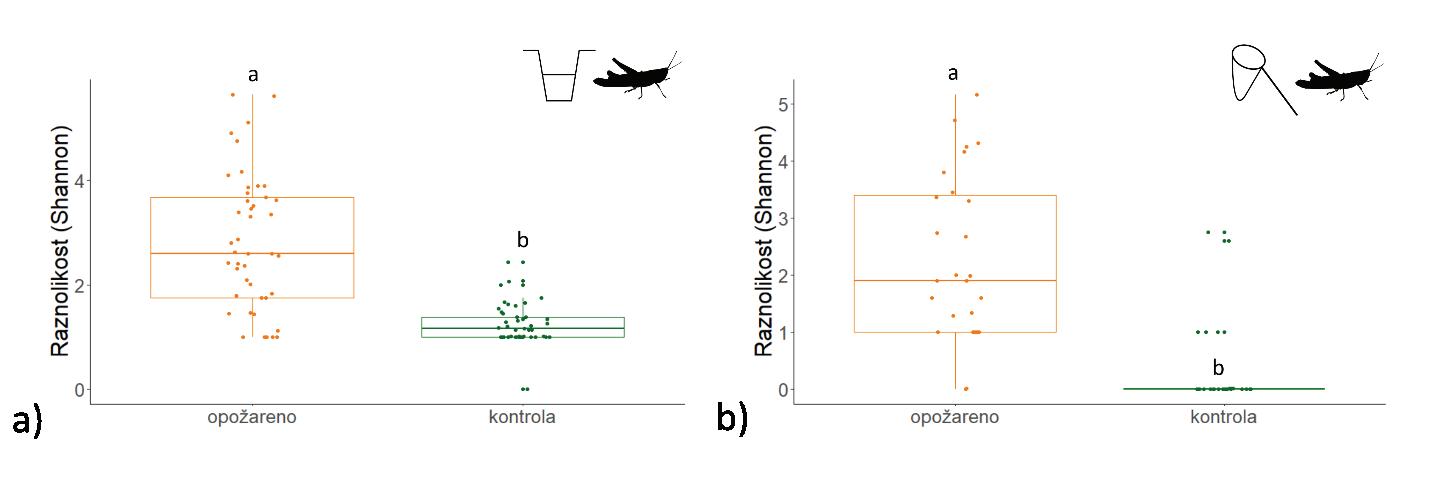
Slika 8. Razlike u raznolikosti 1. reda (iz Shannonovog indeksa) za ravnokrilce (Orthoptera) uzorkovane a) lovnim posudama, b) kečerom, između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
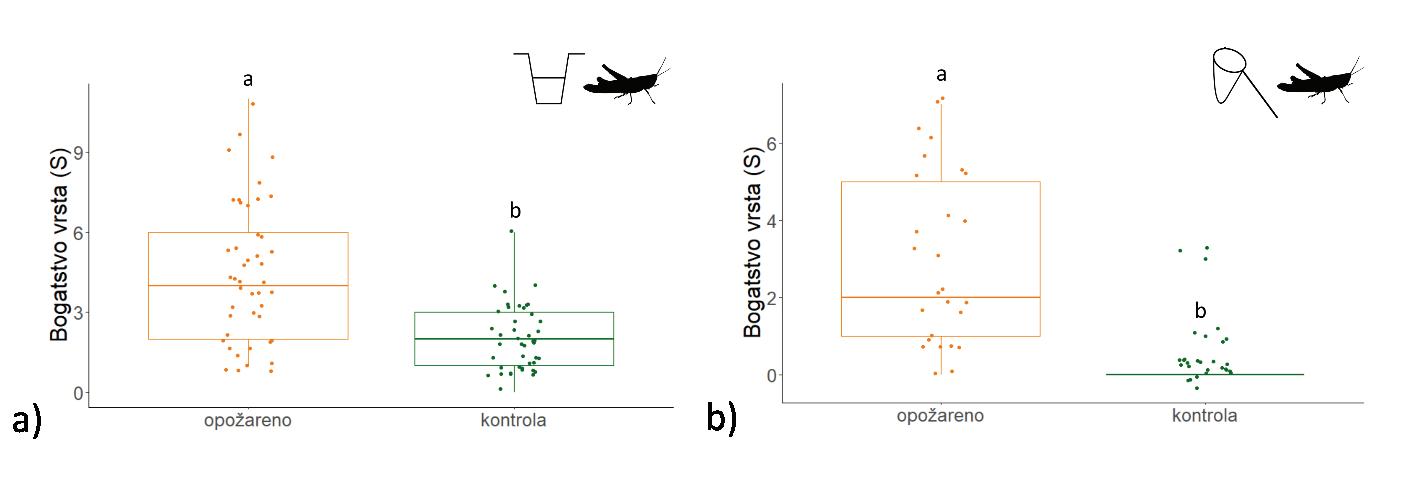
77 BIORAZNOLIKOST
Slika 8. Razlike u raznolikosti 2. reda (iz Simpsonovog indeksa): a) mrava (Formicidae) te ravnokrilaca (Orthoptera) uzorkovanih b) lovnim posudama, c) kečerom, između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
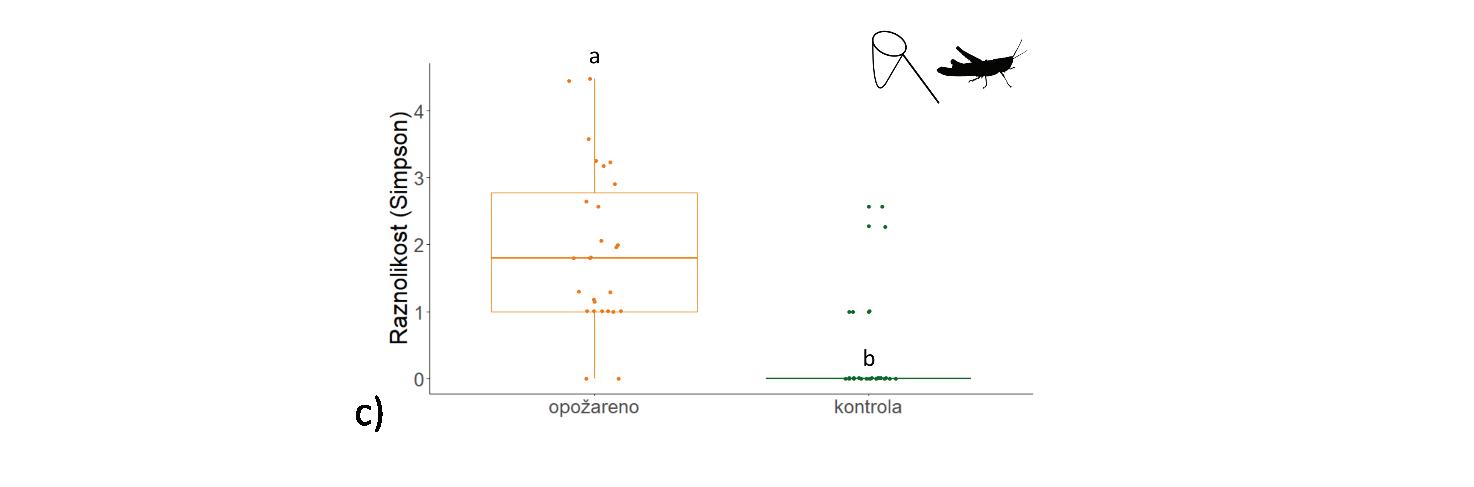
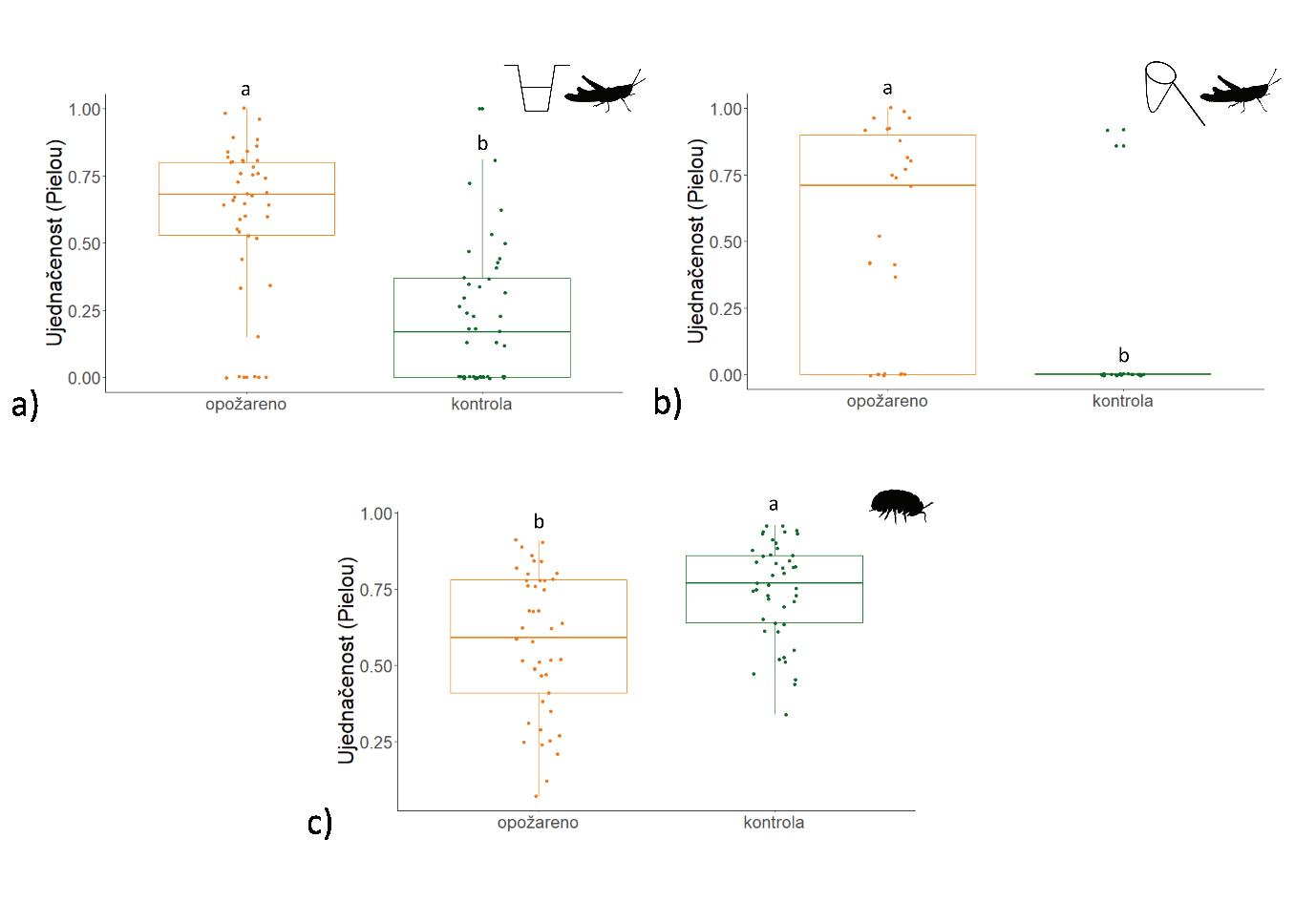
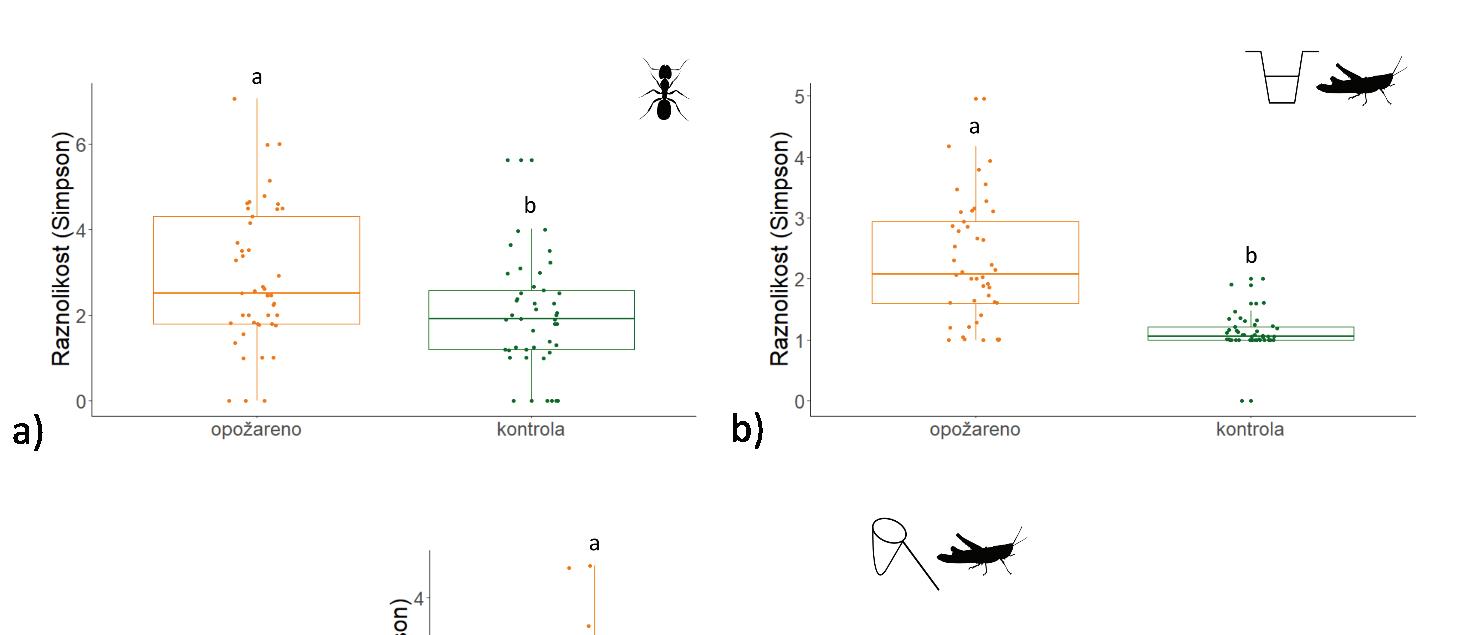
Slika 9. Razlike u ujednačenosti zajednica (Pielou indeks): ravnokrilaca (Orthoptera) uzorkovanih a) lovnim posudama, b) kečerom, c) jednakonožaca (Isopoda), između opožarenih (narančasto) i kontrolnih (tamnozeleno) šumskih staništa u NP Sjeverni Velebit. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Nisu zabilježene statistički značajne razlike između staništa u preostalim biocenološkim parametrima za zajednice mrava (F=2,259-3,786, d.f.1=1, d.f.2=88, p=0,136-0,055) i jednakonožaca (F=0,241-2,198, d.f.1=1, d.f.2=88, p=0,625-0,142) te ni u jednom parametru za zajednice trčaka (F=0,153-1,490, d.f.1=1, d.f.2=88, p=0,697-0,225) i praživotinja (F=0,327-3,337, d.f.1=1, d.f.2=178, p=0,568-0,069).
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 78
3.1.3. Utjecaj okolišnih čimbenika na prostornu raspodjelu vrsta indikatorskih organizama
Utjecaj razlika u okolišnim čimbenicima između opožarenih i kontrolnih šumskih staništa na prostornu raspodjelu vrsta indikatorskih organizama ispitan je kanoničkom analizom korespondencije (CCA) odnosno analizom redundancije (RDA), koje su pokazale statistički značajnu ordinaciju za sve indikatorske skupine (pseudo-F=1,4-4,5, p=0,031-0,002) osim praživotinja (pseudo-F=1,1, p=0,290) (Slika 10). Prve dvije osi ordinacije zajedno objašnjavaju 22,33% (u zajednicama mrava) do čak 67% varijacije u sastavu vrsta (u zajednicama ravnokrilaca uzorkovanih kečerom). U većine skupina uočljivo je jasno odvajanje opožarenih i kontrolnih postaja po 1. osi ordinacije , uz nešto veća preklapanja u zajednicama mrava (Slika 10b)
Slika 10. Ordinacijski dijagrami kanoničke analize korespondencije (a,b) odnosno analize redundancije (c-e) s prikazom odnosa vrsta (plave strelice/trokutići): a) trčaka (Carabidae), b) mrava (Formicidae), ravnokrilaca (Orthoptera) uzorkovanih c) lovnim posudama, d) kečerom te e) jednakonožaca (Isopoda) i mjerenih okolišnih čimbenika (crvene strelice). Postaje u opožarenim šumskim staništima u NP Sjeverni Velebit prikazane su narančastim, a kontrolne postaje tamnozelenim kvadratićima. Prikazane su samo statistički značajne ordinacije (p<0,050). Puna imena vrsta uključenih u analizu navedena su u prilogu.
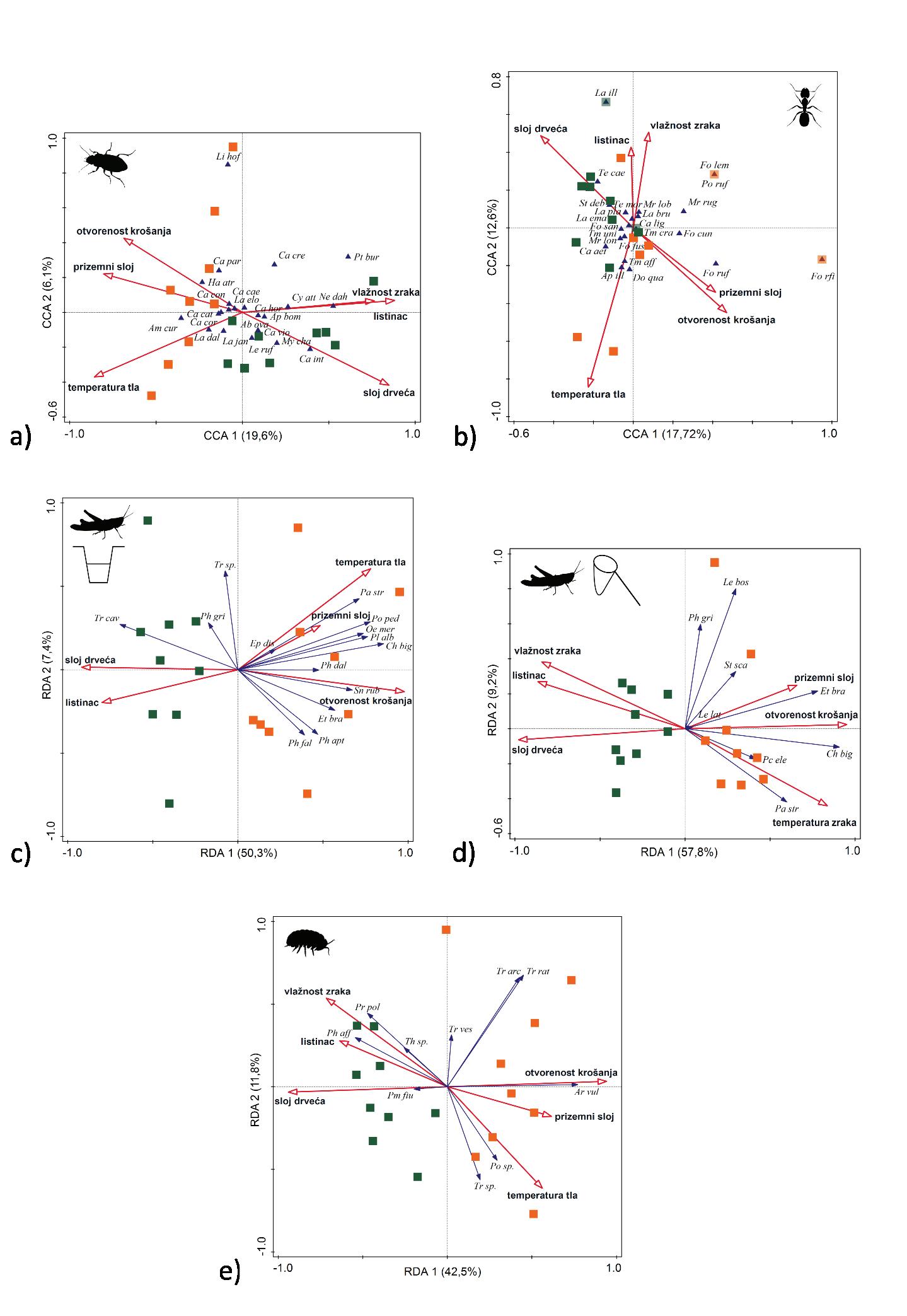
79 BIORAZNOLIKOST
Najveći utjecaj na sastav vrsta u opožarenim staništima imala je otvorenost sklopa krošanja te posljedično viša temperatura tla (odnosno zraka za ravnokrilce uzorkovane kečerom; Slika 10d) te veća zastupljenost prizemnog sloja vegetacije (Slika 10). S druge strane, u neutjecanoj šumi sastav vrsta bio je prvenstveno određen većom zastupljenošću sloja drveća te stoga višom vlažnošću zraka (potonji čimbenik nije imao statistički značajan utjecaj jedino na sastav vrsta ravnokrilaca uzorkovanih lovnim posudama; Slika 10c) i većim udjelom listinca (Slika 10)
3.2. Utjecaj ledoloma i vjetroizvala na biološku raznolikost šumskih ekosustava
3.2.1. Okolišni čimbenici
Ukupno deset okolišnih čimbenika pokazalo je statistički značajne razlike između utjecanih gospodarenih, utjecanih negospodarenih i/ili kontrolnih postaja (Slika 11, 12). Na gospodarenim šumskim površinama zabilježena je statistički značajno veća zastupljenost prizemnog sloja vegetacije (GLMM, F=6,074, d.f.1=2, d.f.2=21, p=0,008) te manja zastupljenost sloja mahovina (F=8,125, d.f.1=2, d.f.2=21, p=0,002) i listinca (F=27,212, d.f.1=2, d.f.2=21, p<0,001) u odnosu na negospodarene i kontrolne postaje (Slika 12cde). Također, temperatura zraka (F=22,691, d.f.1=2, d.f.2=117, p<0,001) i tla (F=38,352, d.f.1=2, d.f.2=117, p<0,001) postupno se smanjivala, a zastupljenost sloja drveća povećavala (F=44,879, d.f.1=2, d.f.2=21, p<0,001) od gospodarenih prema kontrolnim postajama (Slika 11ac, 12b). S druge strane, kontrolne postaje imale su statistički značajno višu vlažnost zraka (F=6,136, d.f.1=2, d.f.2=117, p=0,003) i pH tla (F=7,239, d.f.1=2, d.f.2=44, p=0,002) te manju otvorenost sklopa krošanja (F=61,551, d.f.1=2, d.f.2=21, p<0,001) u odnosu na utjecane postaje, bez obzira na tip gospodarenja (Slika 11bd, 12a). Skeletnost tla je jedini čimbenik čije su najviše vrijednosti zabilježene na negospodarenim postajama te su se postupno smanjivale preko kontrolnih prema gospodarenim postajama (F=20,219, d.f.1=2, d.f.2=21, p<0,001; Slika 11e). Vlažnost tla (F=2,221, d.f.1=2, d.f.2=117, p=0,113) i zastupljenost sloja grmlja (F=2,567, d.f.1=2, d.f.2=21, p=0,101) nisu se statistički značajno razlikovale između postaja.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 80
Slika 11. Razlike u okolišnim čimbenicima između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka: a) temperatura zraka, b) vlažnost zraka, c) temperatura tla, d) pH tla, e) udio skeletnog tla. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
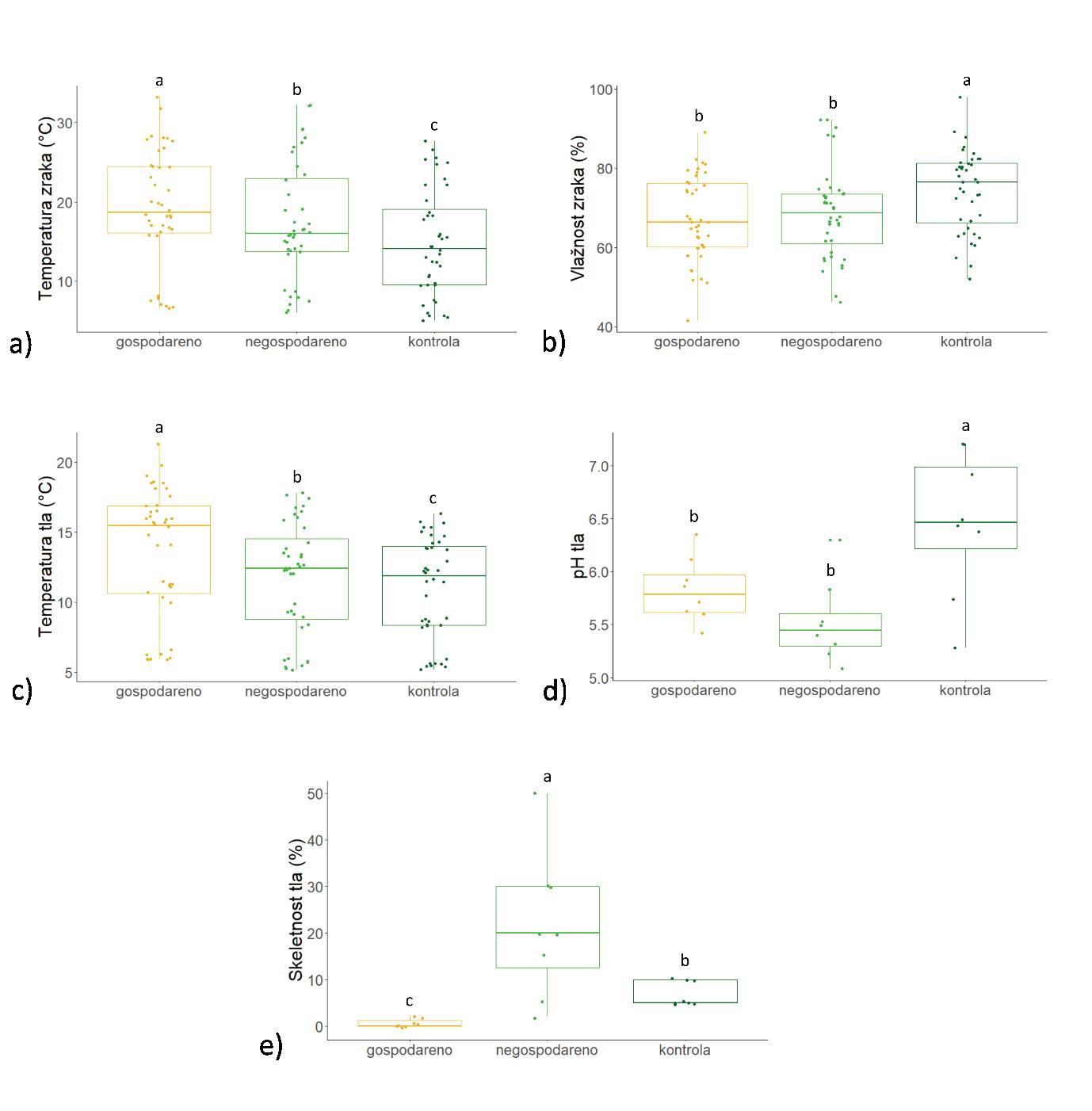
81 BIORAZNOLIKOST
Slika 12. Razlike u okolišnim čimbenicima između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka: a) otvorenost sklopa krošanja, zastupljenost b) sloja drveća, c) prizemnog sloja vegetacije, d) sloja mahovina, e) udio listinca. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
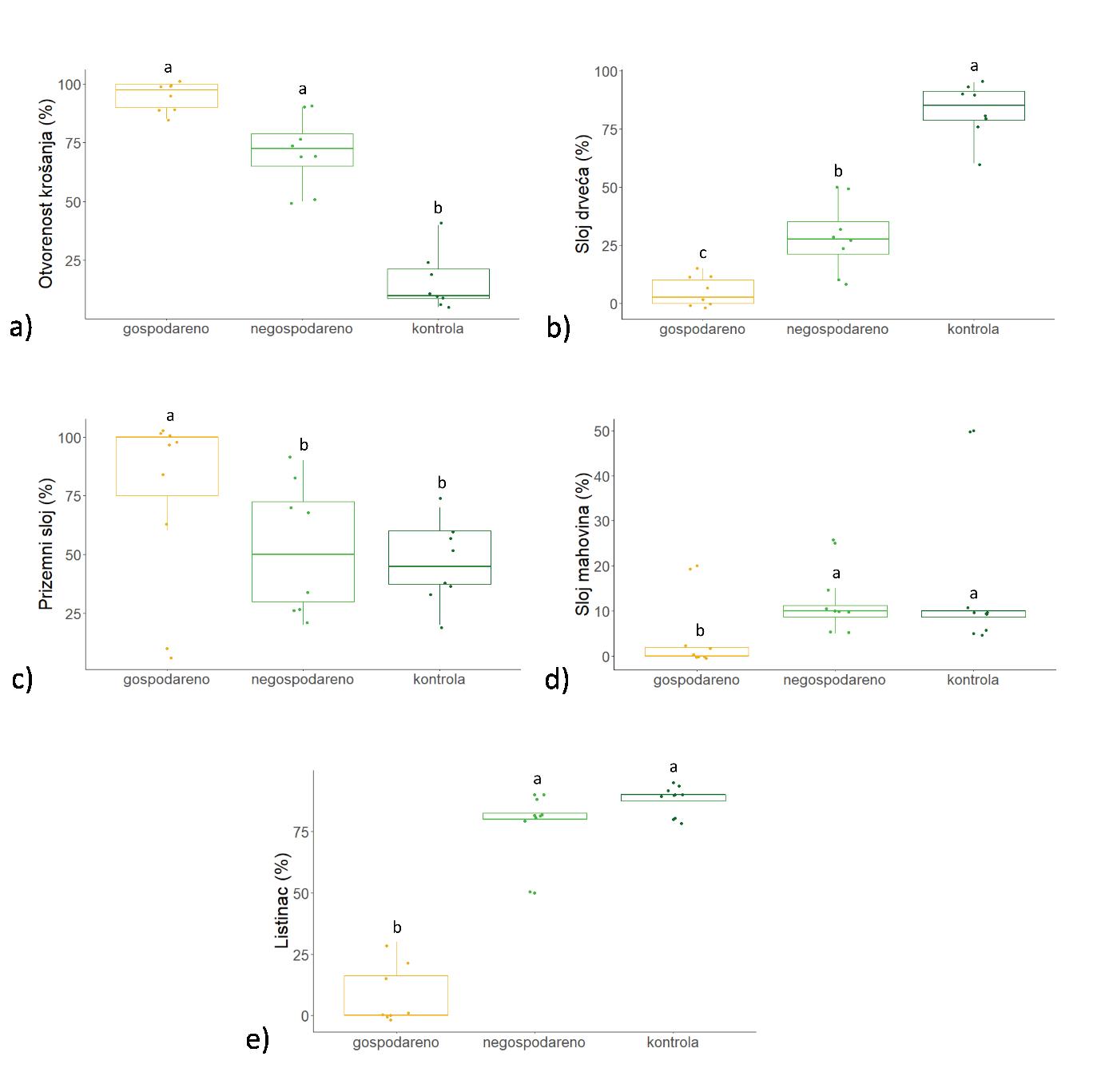
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 82
3.2.2. Prostorna dinamika zajednica indikatorskih organizama
Generaliziranim linearnim mješovitim modelima ustanovljen je statistički značajan pad brojnosti trčaka (F=6,521, d.f.1=2, d.f.2=117, p=0,002) i ravnokrilaca (F=19,505, d.f.1=2, d.f.2=117, p<0,001) na utjecanim gospodarenim u odnosu na negospodarene i kontrolne postaje (Slika 13). Bogatstvo svojti ravnokrilaca imalo je isti statistički značajan obrazac (F=10,800, d.f.1=2, d.f.2=117, p<0,001; Slika 14b), dok je suprotan obrazac zabilježen za bogatstvo vrsta mrava (F=7,955, d.f.1=2, d.f.2=117, p=0,001), uz statistički značajno više vrijednosti na gospodarenim u odnosu na negospodarene i kontrolne postaje (Slika 14a).
Slika 13.
u
jedinki: a) trčaka (Carabidae), b) ravnokrilaca (Orthoptera), između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
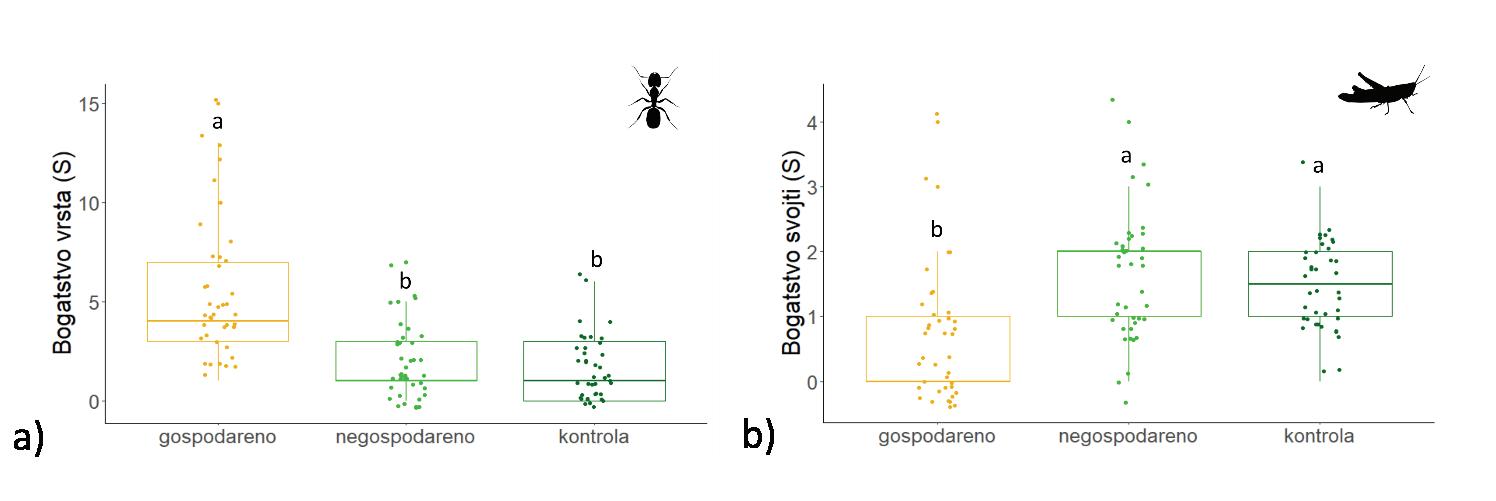
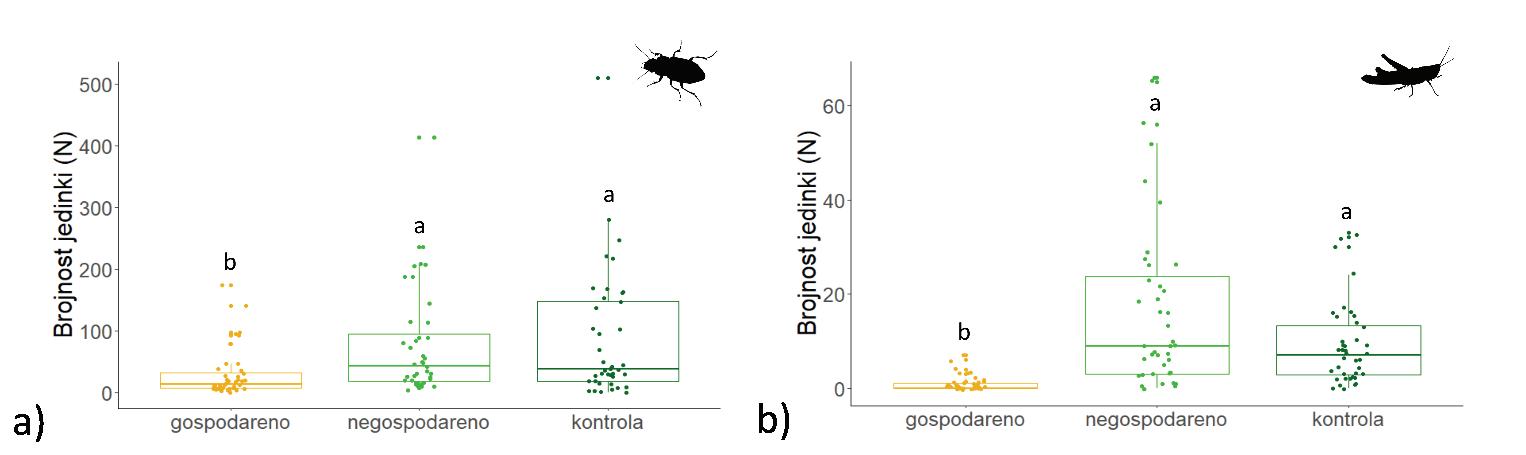
Slika 14. Razlike u bogatstvu vrsta/svojti: a) mrava (Formicidae), b) ravnokrilaca (Orthoptera), između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Raznolikost 1. i 2. reda statistički se značajno razlikovala između istraživanih postaja u tri indikatorske skupine, pokazujući asimetrične odgovore (Slika 15, 16). Utjecane gospodarene postaje podržavale su statistički značajno veću raznolikost trčaka (F=4,965, d.f.1=2, d.f.2=117, p=0,009) i mrava (F=13,23115,086, d.f.1=2, d.f.2=117, p<0,001) u odnosu na negospodarene i kontrolne postaje (Slika 15b, 16ab), s izuzetkom raznolikosti 1. reda u zajednicama trčaka (F=4,135, d.f.1=2, d.f.2=117, p=0,018), koja je u negospodarenoj šumi imala intermedijarne vrijednosti u odnosu na druga dva tipa staništa (Slika 15a). Nasuprot tome, raznolikost ravnokrilaca imala je statistički značajno niže vrijednosti na gospodarenim nego na negospodarenim i kontrolnim postajama (F=9,594-9,951, d.f.1=2, d.f.2=117, p<0,001; Slika 15c, 16c).
83 BIORAZNOLIKOST
Razlike
brojnosti
Slika 15. Razlike u raznolikosti 1. reda (iz Shannonovog indeksa): a) trčaka (Carabidae), b) mrava (Formicidae), c) ravnokrilaca (Orthoptera), između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
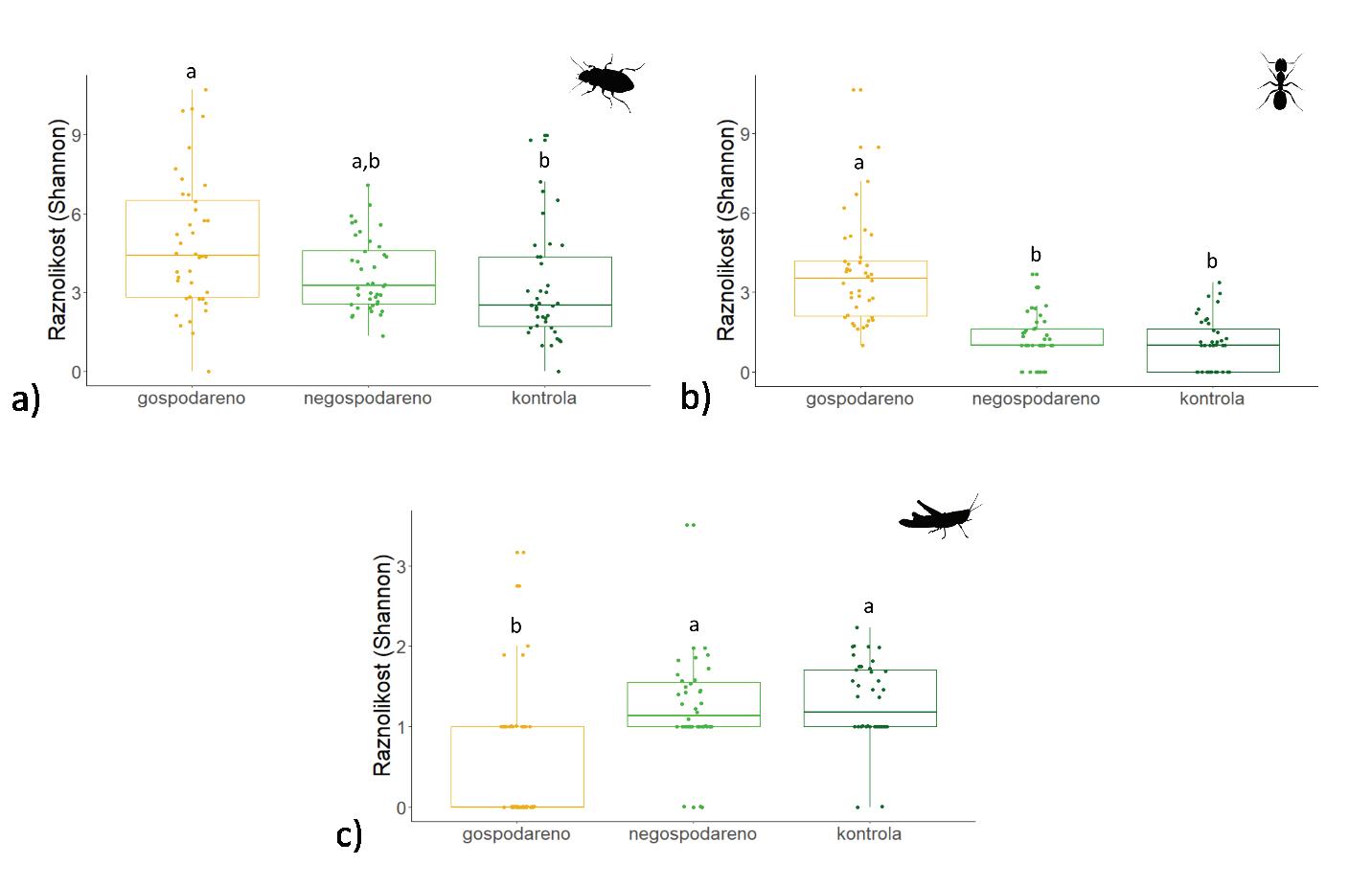
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 84
Slika 16. Razlike u raznolikosti 2. reda (iz Simpsonovog indeksa): a) trčaka (Carabidae), b) mrava (Formicidae), c) ravnokrilaca (Orthoptera), između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Ujednačenost zajednica mrava bila je također u prvom redu određena tipom gospodarenja, pokazujući statistički značajno više vrijednosti na gospodarenim u odnosu na druga dva tipa staništa (F=26,220, d.f.1=2, d.f.2=117, p<0,001; Slika 17b), dok je poremećaj na staništu uzrokovan ledolomom odnosno vjetroizvalama imao veći utjecaj na ujednačenost zajednica trčaka nego tip gospodarenja, uz statistički značajno niže vrijednosti na kontrolnim u odnosu na utjecane postaje (F=8,807, d.f.1=2, d.f.2=117, p<0,001; Slika 17a).
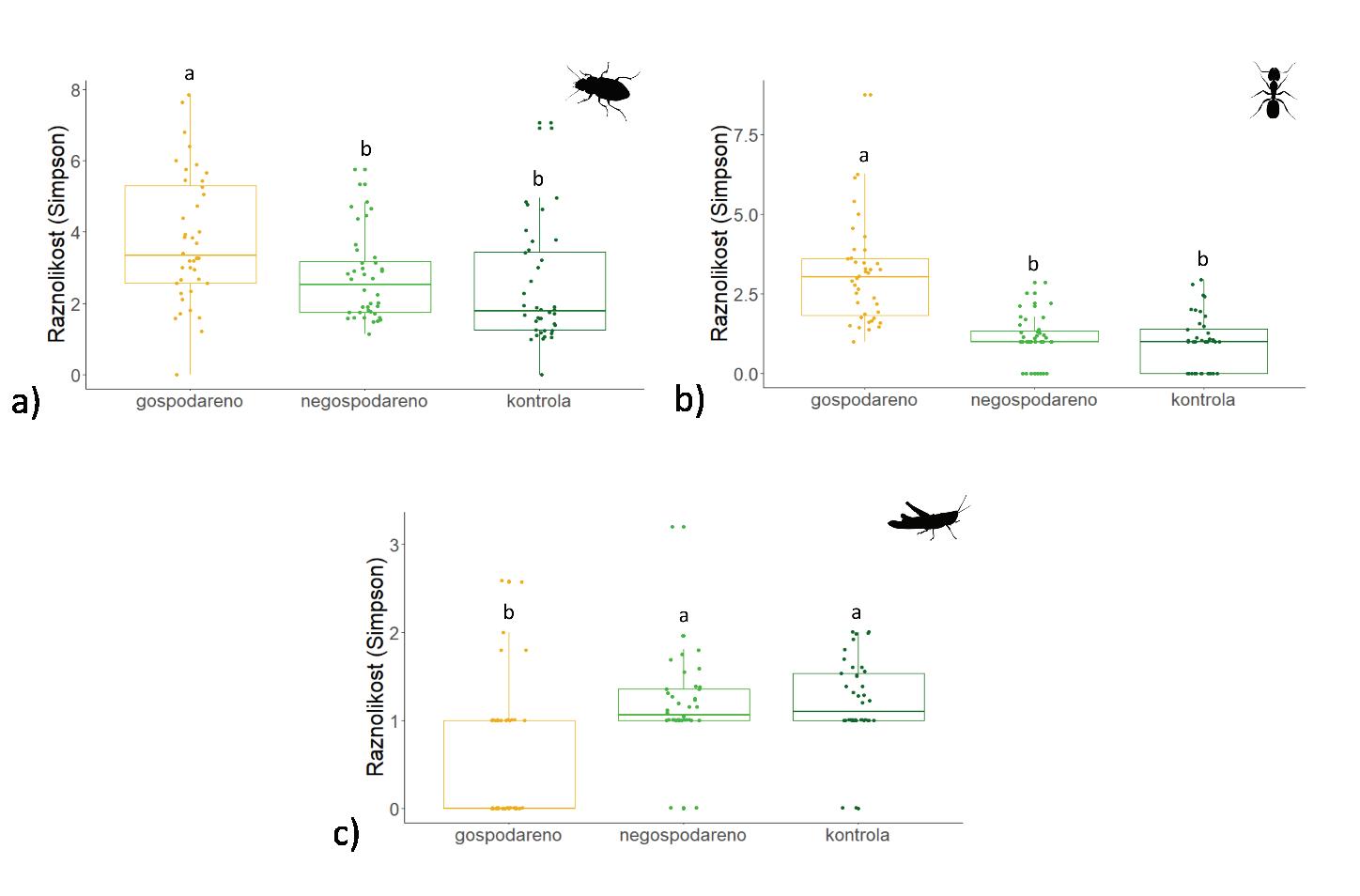
85 BIORAZNOLIKOST
Slika 17. Razlike u ujednačenosti zajednica (Pielou indeks): a) trčaka (Carabidae), b) mrava (Formicidae), između gospodarenih (žuto) i negospodarenih (svijetlozeleno) šumskih staništa utjecanih ledolomom i vjetroizvalama te neutjecanih (kontrolnih) staništa (tamnozeleno) na području Risnjaka. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
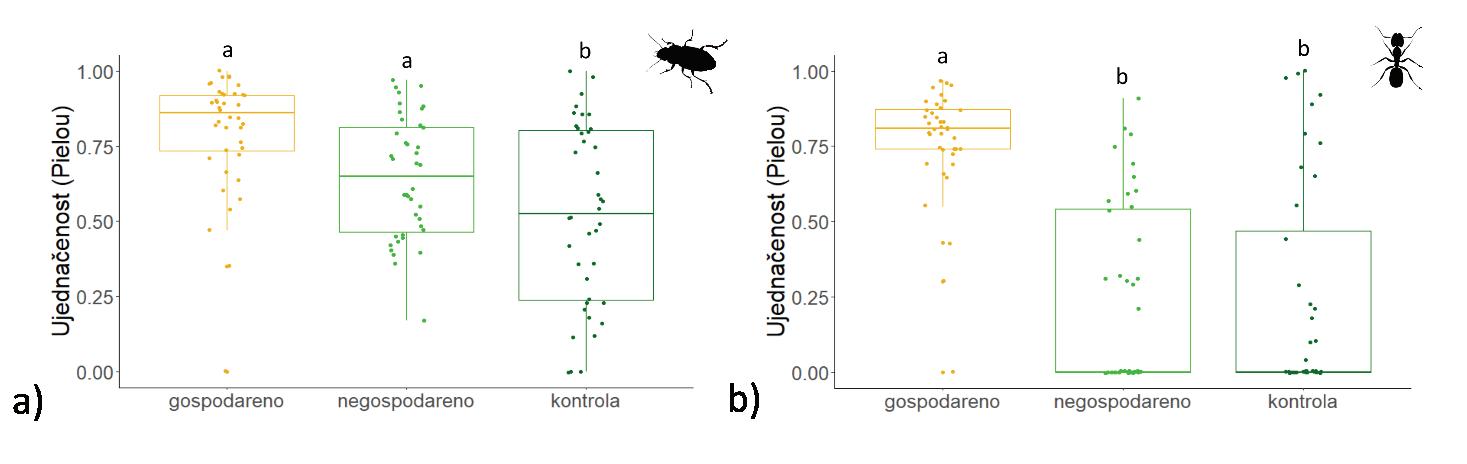
Nisu zabilježene statistički značajne razlike između staništa u bogatstvu vrsta trčaka (F=0,372, d.f.1=2, d.f.2=117, p=0,690), brojnosti mrava (F=1,925, d.f.1=2, d.f.2=117, p=0,150) i ujednačenosti zajednica ravnokrilaca (F=0,183, d.f.1=2, d.f.2=117, p=0,833) te ni u jednom parametru za zajednice jednakonožaca (F=0,024-1,130, d.f.1=2, d.f.2=117, p=0,977-0,327) i praživotinja (F=0,395-1,409, d.f.1=2, d.f.2=117, p=0,675-0,248).
3.2.3. Utjecaj okolišnih čimbenika na prostornu raspodjelu vrsta indikatorskih organizama
Analizom redundancije (RDA) ispitan je utjecaj razlika u okolišnim čimbenicima između utjecanih gospodarenih, negospodarenih i kontrolnih šumskih staništa na prostornu raspodjelu vrsta indikatorskih organizama; ordinacija je bila statistički značajna za sve skupine (pseudo-F=2,3-4,5, p=0,008-0,002) osim praživotinja (pseudo-F=0,9, p=0,742) (Slika 18). Prve dvije osi ordinacije zajedno objašnjavaju 34,45% (u zajednicama mrava) do 60,37% varijacije u sastavu vrsta (u zajednicama ravnokrilaca). Dok je u svih skupina osim jednakonožaca (Slika 18d) vidljivo uglavnom jasno odvajanje utjecanih gospodarenih postaja od postaja u preostala dva tipa staništa po 1. osi (Slika 18abc), odvajanje utjecanih negospodarenih od kontrolnih postaja po 2. osi uočljivo je samo u trčaka i, s nešto većim preklapanjem, u ravnokrilaca (Slika 18ac)
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 86
Slika 18. Ordinacijski dijagrami analize redundancije s prikazom odnosa vrsta (plave strelice): a) trčaka (Carabidae), b) mrava (Formicidae), c) ravnokrilaca (Orthoptera) te d) jednakonožaca (Isopoda) i mjerenih okolišnih čimbenika (crvene strelice). Postaje u gospodarenim odnosno negospodarenim šumskim staništima utjecanima ledolomom/ vjetroizvalama na području Risnjaka prikazane su žutim odnosno svijetlozelenim kvadratićima, a kontrolne postaje tamnozelenim kvadratićima. Prikazane su samo statistički značajne ordinacije (p<0,050). Puna imena vrsta uključenih u analizu navedena su u prilogu.
Sastav vrsta na utjecanim gospodarenim postajama bio je u prvom redu određen višim vrijednostima temperature tla, koja je bila korelirana s otvorenošću sklopa krošanja i, posljedično, većom zastupljenošću prizemnog sloja vegetacije (Slika 18). Nasuprot tome, najveći utjecaj na prostornu raspodjelu vrsta u druga dva tipa staništa imao je povišen udio listinca te, osobito u neutjecanim šumskim staništima, više vrijednosti pH tla i veća zastupljenost sloja drveća (Slika 18), s kojom je u vezi i povećana vlažnost zraka (potonji čimbenik nije imao statistički značajan utjecaj na zajednice trčaka i mrava; Slika 18ab). Za razliku od ostalih skupina, sastav vrsta jednakonožaca bio je u velikoj mjeri određen i većom zastupljenošću sloja mahovina na utjecanim negospodarenim i kontrolnim postajama (Slika 18d)
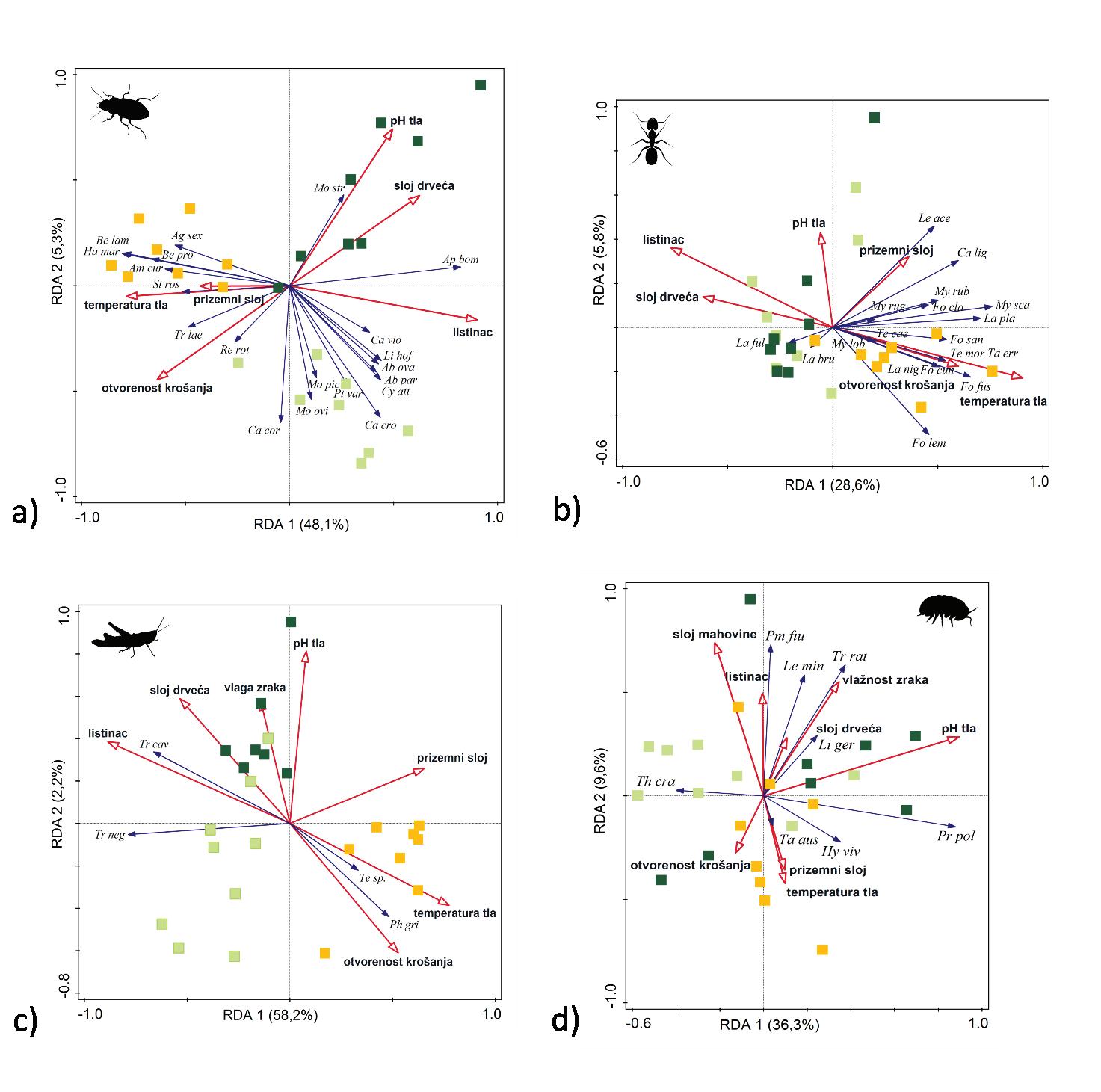
87 BIORAZNOLIKOST
3.3. Utjecaj sukcesijskih i hidroloških promjena na biološku raznolikost cretnog ekosustava
3.3.1. Okolišni čimbenici
Svi mjereni okolišni čimbenici statistički su se značajno razlikovali između postaja u središnjem (vlažnom) dijelu creta, postaja u sukcesijskom (suhom) dijelu creta te postaja na šumskom rubu (Slika 19, 20). Dok su statistički značajno više vrijednosti temperature zraka (GLMM, F=9,523, d.f.1=2, d.f.2=87, p<0,001) i temperature tla (F=11,086, d.f.1=2, d.f.2=87, p<0,001) zabilježene u središnjem i sukcesijskom dijelu creta u odnosu na šumski rub (Slika 19ac), vlažnost zraka (F=19,933, d.f.1=2, d.f.2=87, p<0,001) i vlažnost tla (F=27,303, d.f.1=2, d.f.2=87, p<0,001) pokazivale su asimetričan odgovor; dok se prva povećavala, potonja se smanjivala od središnjeg dijela creta prema šumskom rubu (Slika 19bd). Središnji dio creta se izdvajao po statistički značajno višim vrijednostima pH tla (F=5,762, d.f.1=2, d.f.2=33, p=0,007), dok se šumski rub izdvajao po većoj zastupljenosti sloja drveća (F=3.955,076, d.f.1=2, d.f.2=15, p<0,001) i manjoj zastupljenosti prizemnog sloja vegetacije (F=83,874, d.f.1=2, d.f.2=15, p<0,001) u odnosu na druga dva tipa staništa (Slika 19e, 20ac). Nadalje, zastupljenost grmlja se postupno smanjivala od šumskog ruba prema sukcesijskom dijelu creta (F=32,490, d.f.1=2, d.f.2=15, p<0,001; Slika 20b).
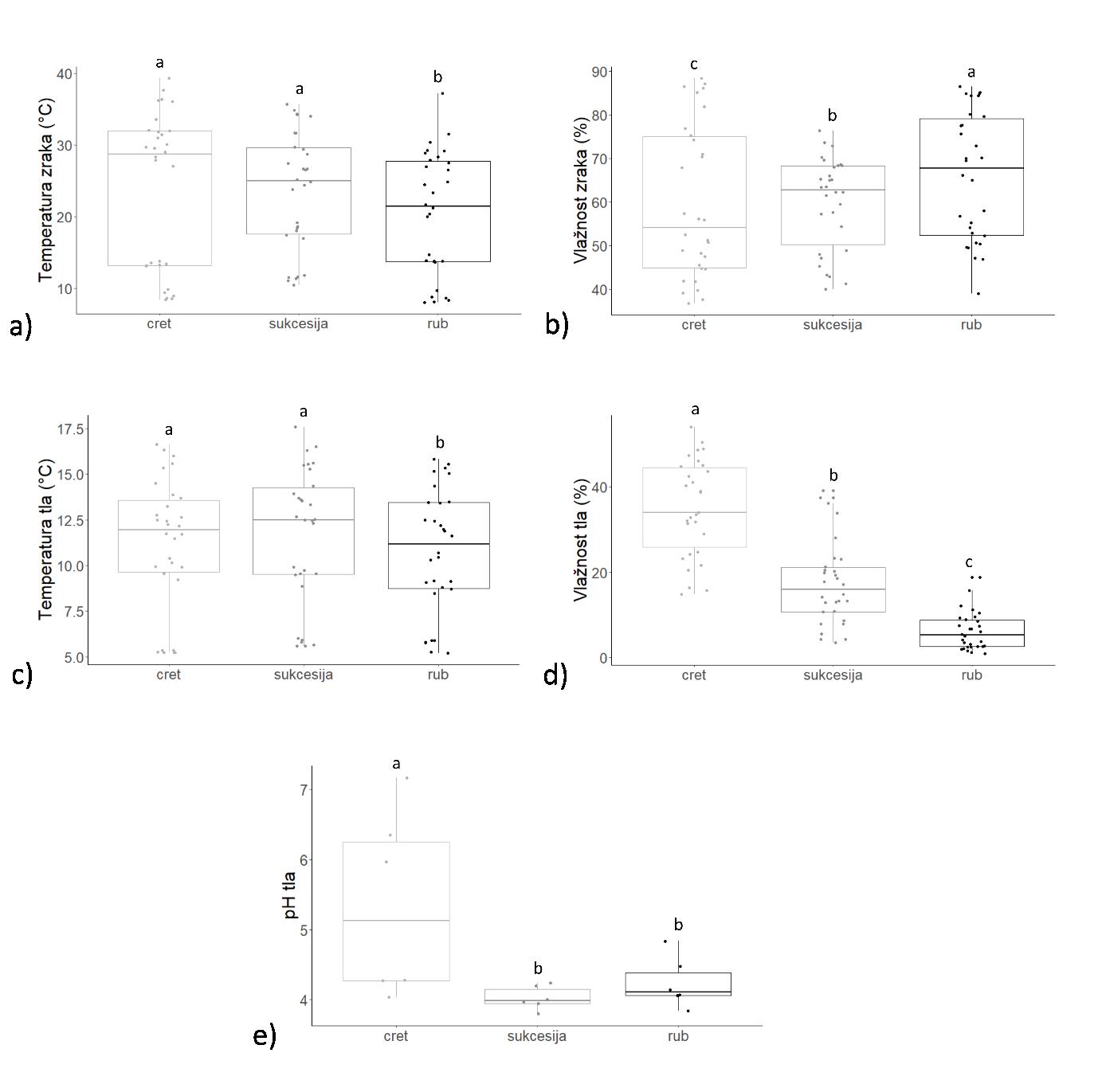
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 88
Slika 19. Razlike u okolišnim čimbenicima između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku: a) temperatura zraka, b) vlažnost zraka, c) temperatura tla, d) vlažnost tla, e) pH tla. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Slika 20. Razlike u okolišnim čimbenicima između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku: zastupljenost a) sloja drveća, b) sloja grmlja, c) prizemnog sloja vegetacije. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za čimbenike koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
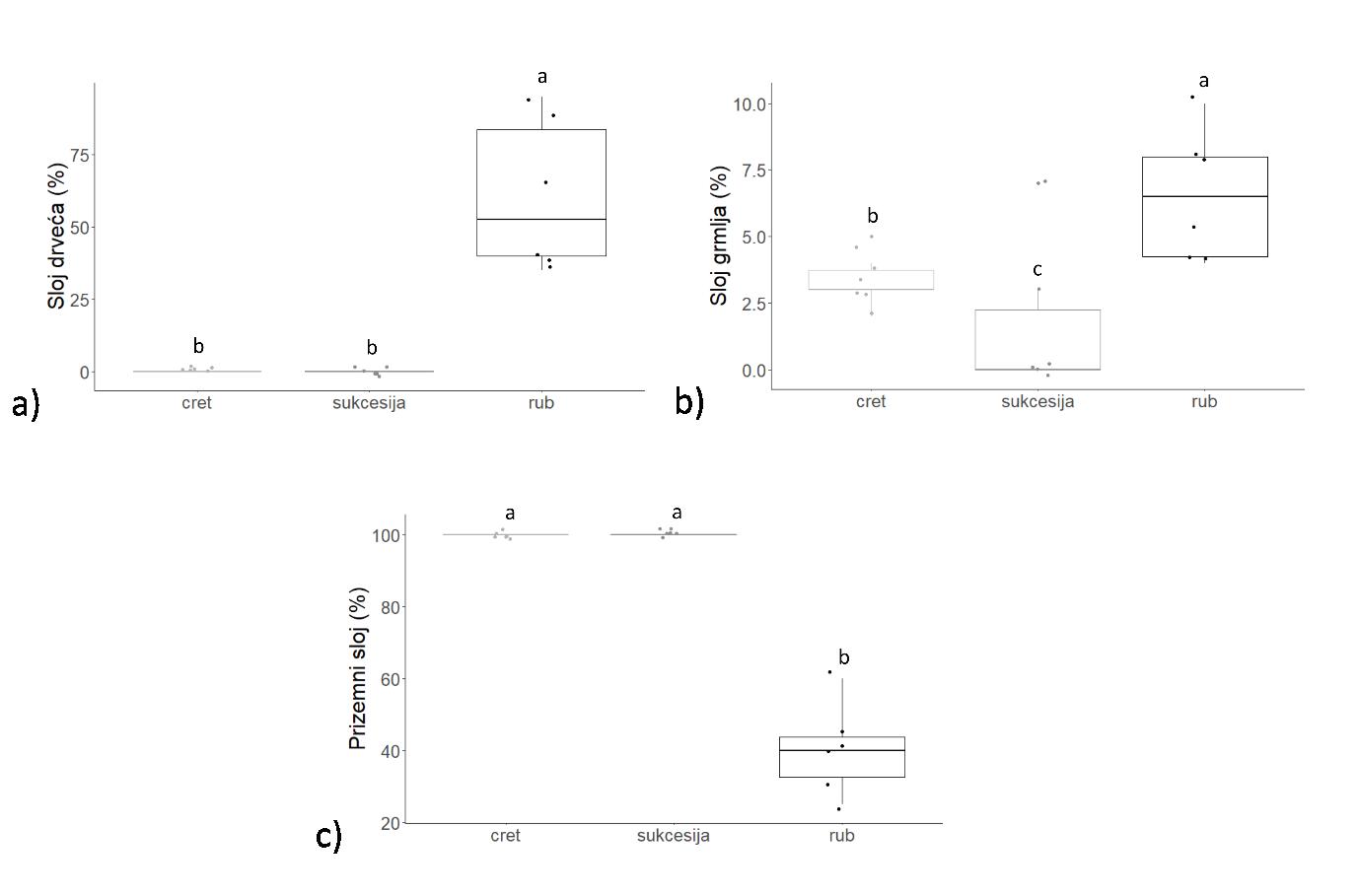
89 BIORAZNOLIKOST
3.3.2. Prostorna dinamika zajednica indikatorskih organizama
Brojnost jedinki statistički se značajno razlikovala između istraživanih postaja u dvije indikatorske skupine, uz varijabilne obrasce (Slika 21). Dok je brojnost trčaka bila najviša na rubu šume, a najniža u središnjem dijelu creta, s intermedijarnim vrijednostima u sukcesijskom dijelu (GLMM, F=4,057, d.f.1=2, d.f.2=87, p=0,021; Slika 21a), brojnost ravnokrilaca uzorkovanih lovnim posudama bila je najviša u sukcesijskom dijelu creta, a najniža na rubu, pokazujući intermedijarne vrijednosti u središnjem dijelu creta (F=4,836, d.f.1=2, d.f.2=87, p=0,010, Slika 21b). S druge strane, zabilježen je statistički značajan pad brojnosti ravnokrilaca uzorkovanih kečerom na šumskom rubu u odnosu na druga dva tipa staništa (F=15,926, d.f.1=2, d.f.2=51, p<0,001; Slika 21c).
Slika 21. Razlike u brojnosti jedinki: a) trčaka (Carabidae) te ravnokrilaca (Orthoptera) uzorkovanih b) lovnim posudama, c) kečerom, između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
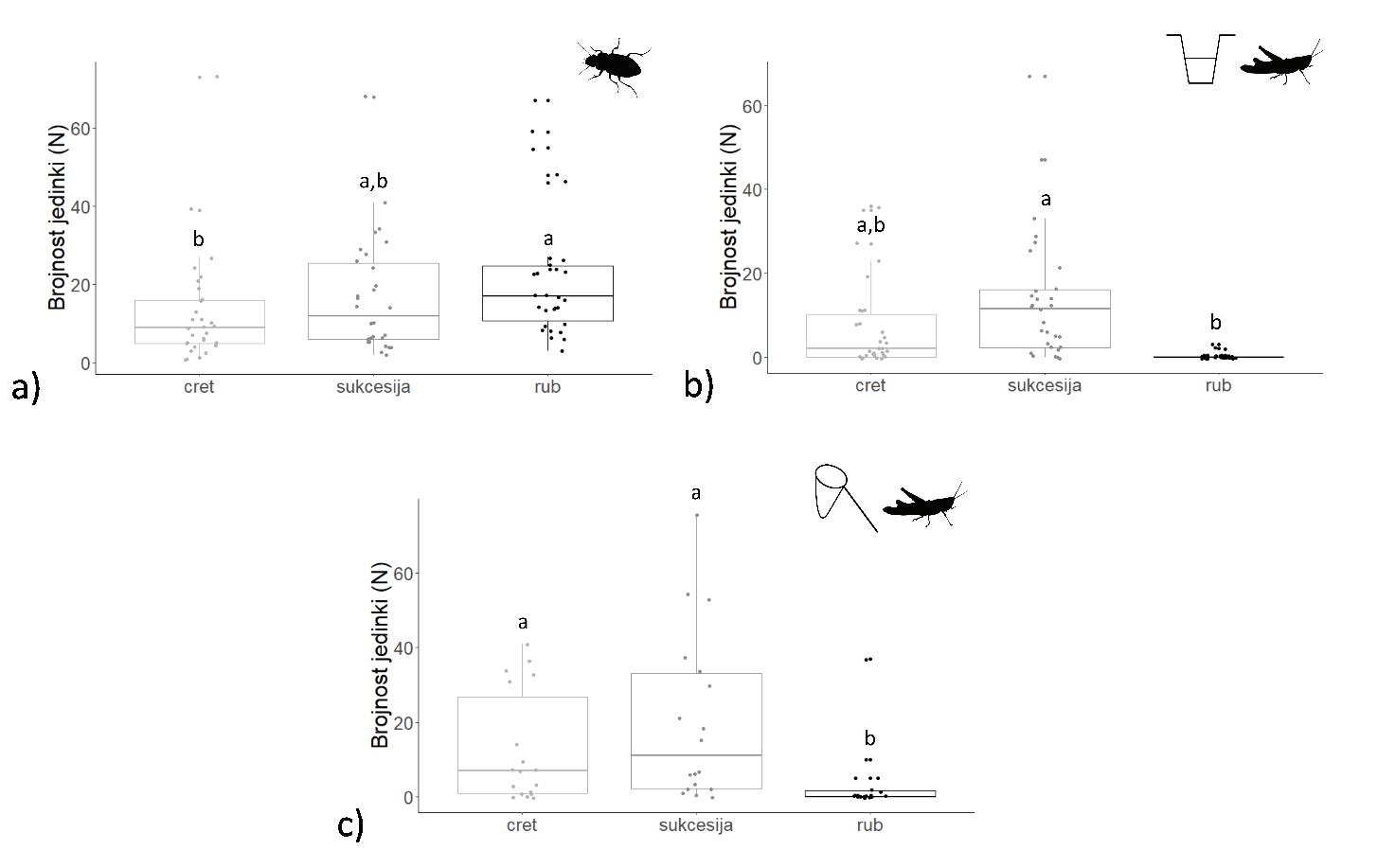
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 90
Isti statistički značajan obrazac pokazalo je bogatstvo vrsta ravnokrilaca (F=10,348-37,166, d.f.1=2, d.f.2=51, 87, p<0,001), bez obzira na metodu uzorkovanja (Slika 22bc), kao i bogatstvo svojti praživotinja (F=16,263, d.f.1=2, d.f.2=177, p<0,001; Slika 22d), dok je u trčaka zabilježen suprotan obrazac (F=11,192, d.f.1=2, d.f.2=87, p<0,001), s postupnim povećanjem bogatstva vrsta od središnjeg dijela creta prema šumskom rubu (Slika 22a)
Slika 22. Razlike u bogatstvu vrsta/svojti: a) trčaka (Carabidae), ravnokrilaca (Orthoptera) uzorkovanih b) lovnim posudama, c) kečerom te d) praživotinja (Protozoa), između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
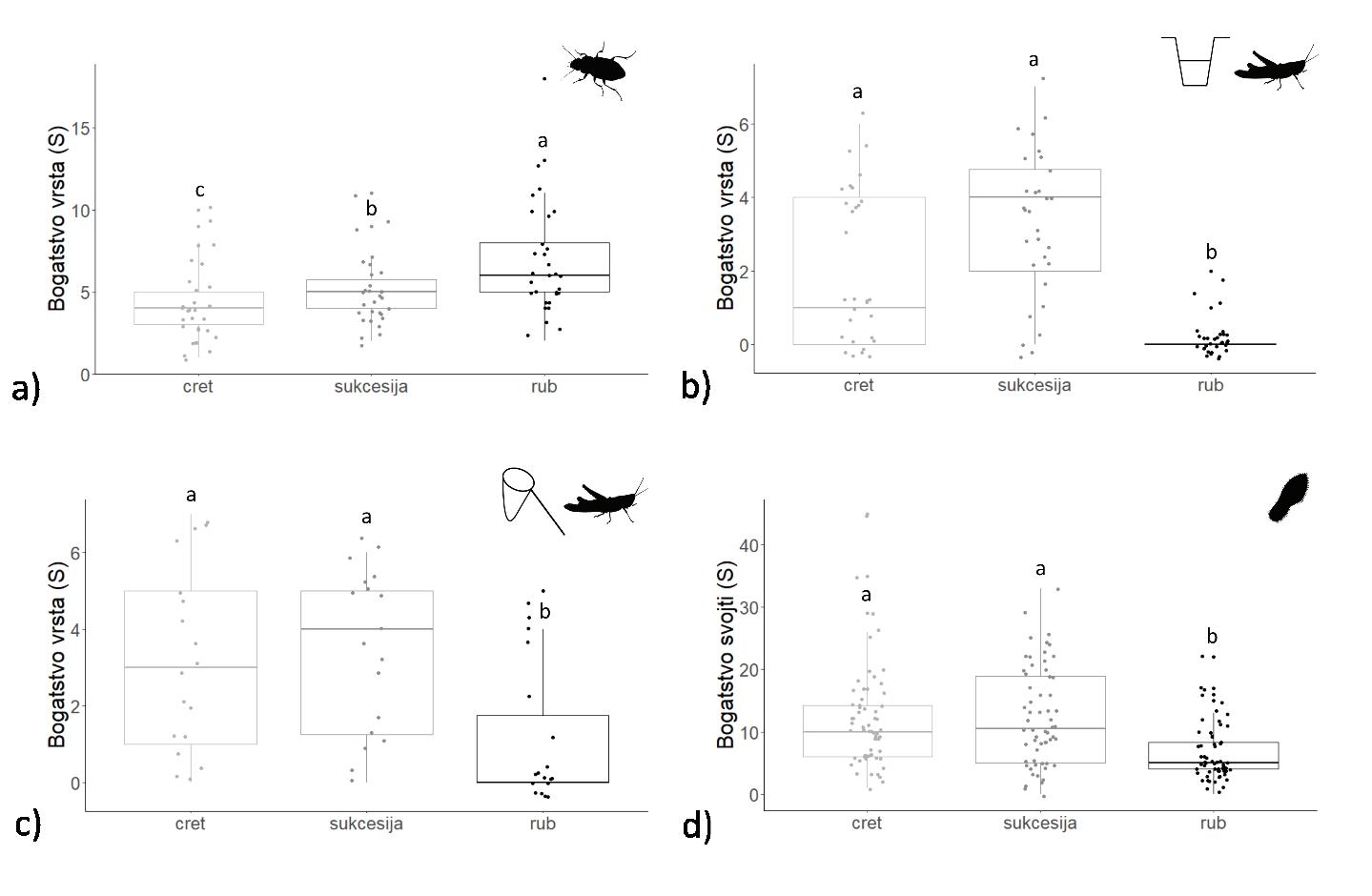
91 BIORAZNOLIKOST
Raznolikost 1. reda pokazala je statistički značajan prostorni obrazac u četiri, a raznolikost 2. reda u tri indikatorske skupine (Slika 23, 24). U zajednicama trčaka, raznolikost 1. (F=11,588, d.f.1=2, d.f.2=87, p<0,001) odnosno 2. reda (F=10,499, d.f.1=2, d.f.2=87, p<0,001) bila je statistički značajno veća na šumskom rubu u odnosu na druga dva tipa staništa (Slika 23a, 24a), dok je u zajednicama jednakonožaca zabilježen statistički značajan pad raznolikosti 1. reda u sukcesijskom dijelu creta (F=3,399, d.f.1=2, d.f.2=87, p=0,038; Slika 23d). Nasuprot tome, u zajednicama ravnokrilaca (F=10,334-34,061, d.f.1=2, d.f.2=51, 87, p<0,001) i praživotinja (F=26,676-32,537, d.f.1=2, d.f.2=177, p<0,001) zabilježen je statistički značajan pad raznolikosti na šumskom rubu, bez razlika između središnjeg i sukcesijskog dijela creta (Slika 23bce, 24cd)
Slika 23. Razlike u raznolikosti 1. reda (iz Shannonovog indeksa): a) trčaka (Carabidae), ravnokrilaca (Orthoptera) uzorkovanih b) lovnim posudama, c) kečerom, d) jednakonožaca (Isopoda) te e) praživotinja (Protozoa), između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
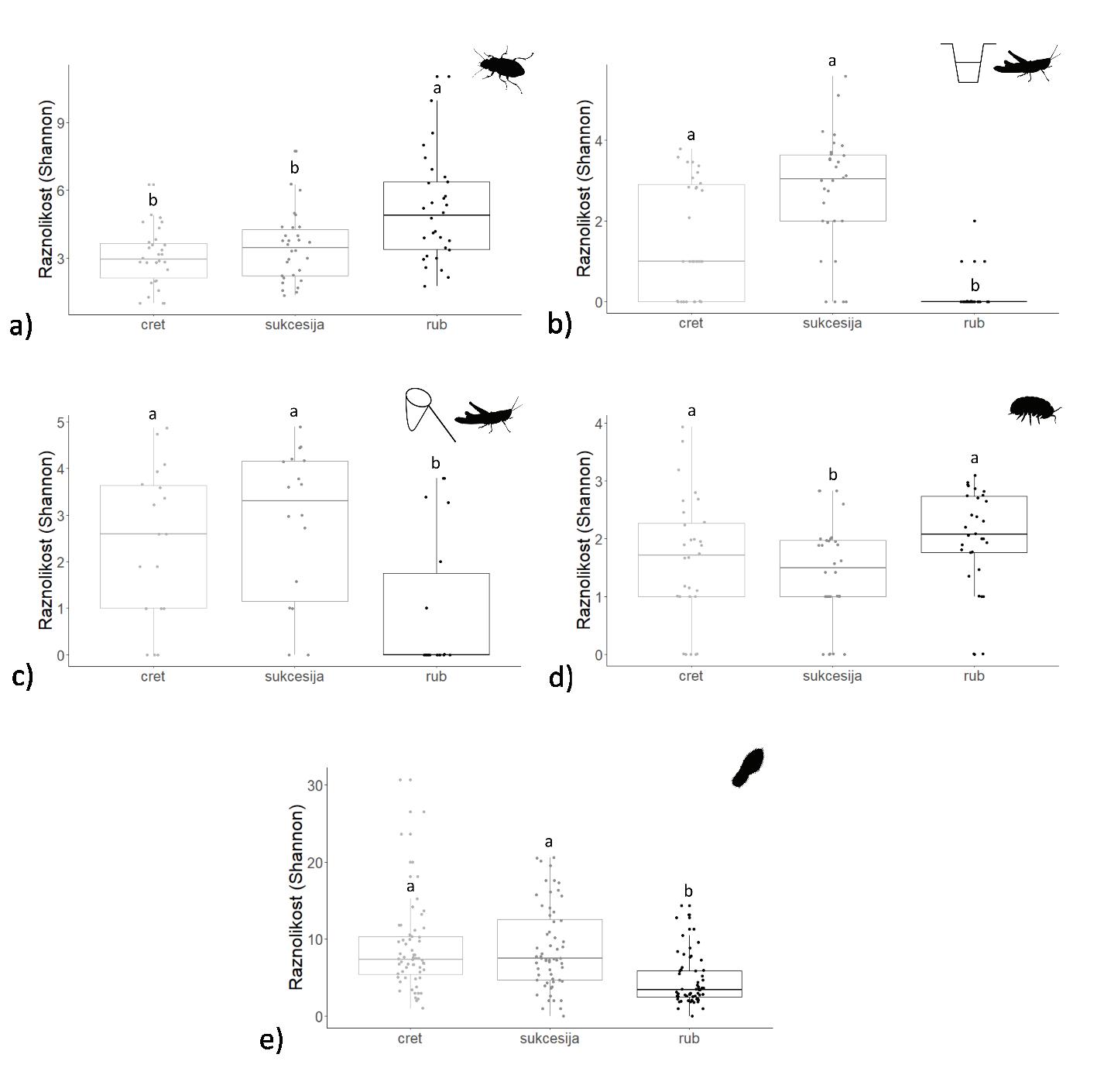
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 92
Slika 24. Razlike u raznolikosti 2. reda (iz Simpsonovog indeksa): a) trčaka (Carabidae), ravnokrilaca (Orthoptera) uzorkovanih b) lovnim posudama, c) kečerom te d) praživotinja (Protozoa), između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
Ujednačenost zajednica ravnokrilaca (F=5,127-31,659, d.f.1=2, d.f.2=51, 87, p=0,009-<0,001) odnosno praživotinja (F=11,339, d.f.1=2, d.f.2=177, p<0,001) pokazivala je isti statistički značajan obrazac (Slika 25). Izuzetak je bila raznolikost 2. reda u zajednicama ravnokrilaca uzorkovanih lovnim posudama (F=466,983, d.f.1=2, d.f.2=87, p<0,001), koja je u središnjem dijelu creta imala intermedijarne vrijednosti između sukcesijskog dijela s najvišom te šumskog ruba s najnižom raznolikošću (Slika 24b).
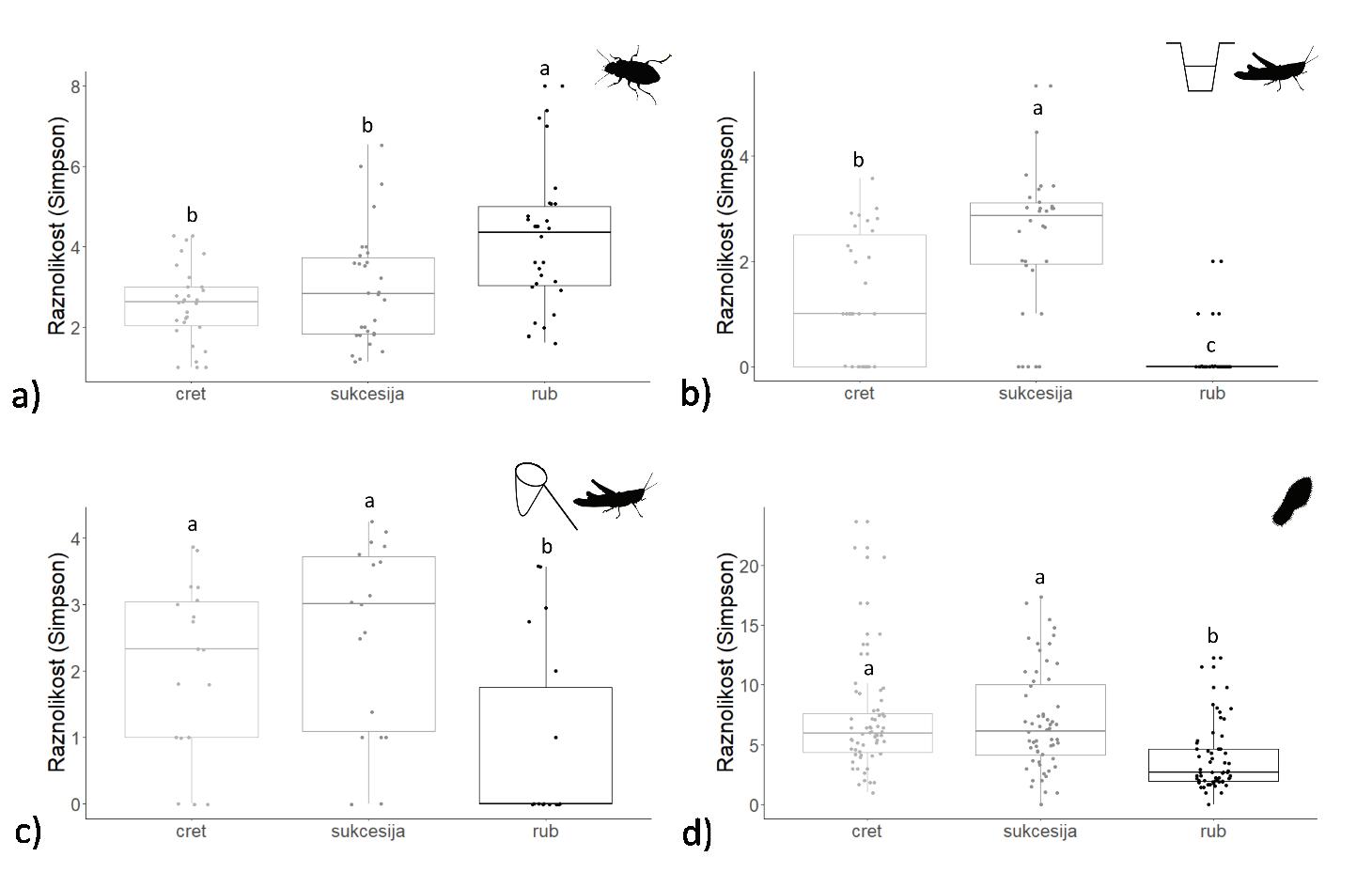
93 BIORAZNOLIKOST
Slika 25. Razlike u ujednačenosti zajednica (Pielou indeks): ravnokrilaca (Orthoptera) uzorkovanih a) lovnim posudama, b) kečerom te c) praživotinja (Protozoa), između središnjeg (vlažnog) dijela creta (svijetlosivo), sukcesijskog (suhog) dijela creta (tamnosivo) i šumskog ruba (crno) na Trsteniku. Prikazani su 1. kvartil, srednja vrijednost, 3. kvartil, standardna devijacija i raspršenje podataka za parametre koji pokazuju statistički značajne razlike između staništa, naznačene različitim slovima (GLMM, post hoc test najmanje značajne razlike, p<0,050).
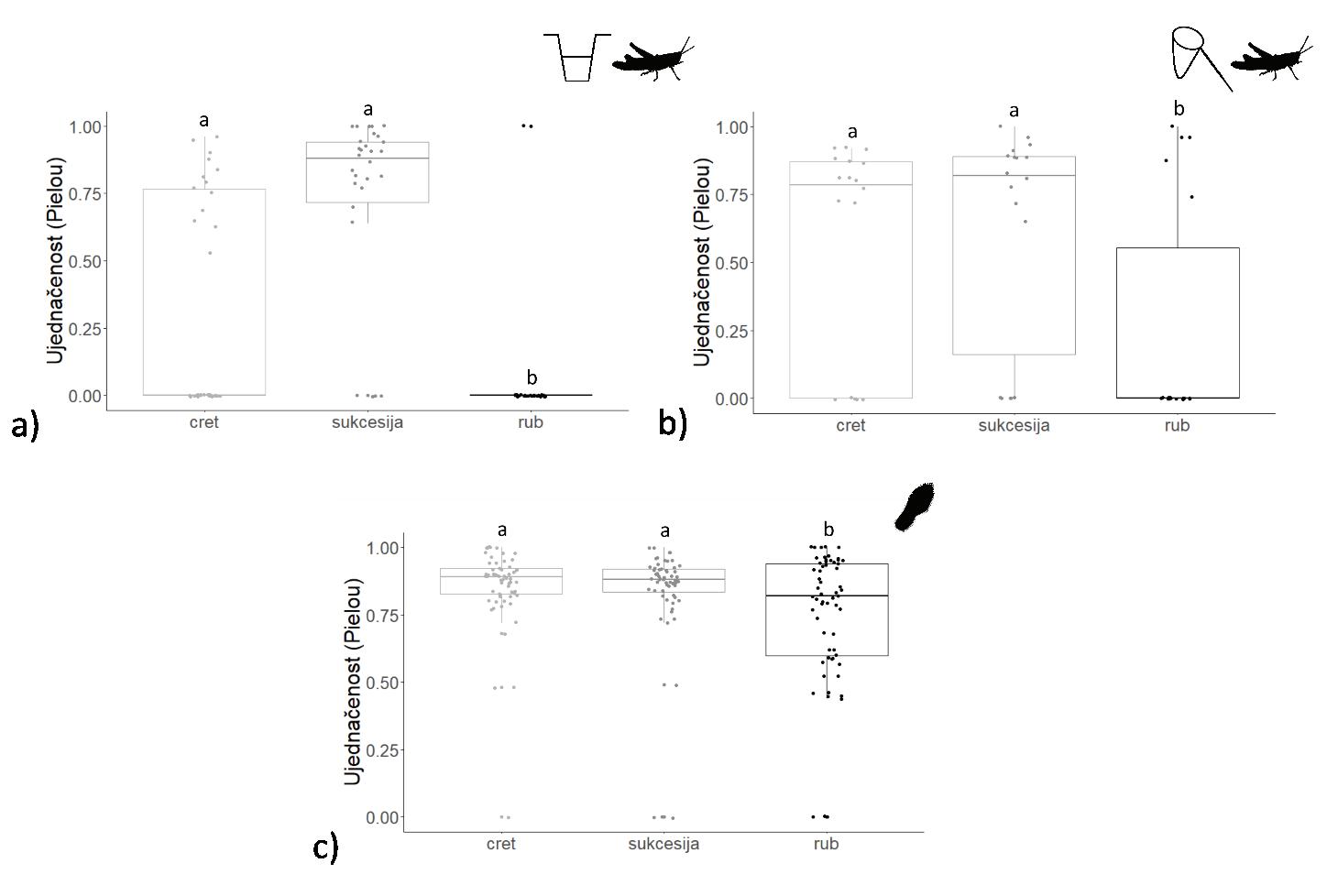
Nisu zabilježene statistički značajne razlike između staništa u ujednačenosti zajednica trčaka (F=2,021, d.f.1=2, d.f.2=87, p=0,139), u preostalim biocenološkim parametrima za zajednice jednakonožaca (F=0,418-1,340, d.f.1=2, d.f.2=87, p=0,660-0,267) te ni u jednom parametru za zajednice mrava (F=0,1901,639, d.f.1=2, d.f.2=87, p=0,827-0,200).
3.3.3. Utjecaj okolišnih čimbenika na prostornu raspodjelu vrsta indikatorskih organizama
Utjecaj promjena okolišnih čimbenika od središnjeg dijela creta prema šumskom rubu na prostornu raspodjelu vrsta ispitan je analizom redundancije (RDA), koja je imala statistički značajnu ordinaciju za sve indikatorske skupine (pseudo-F=1,9-6,7, p=0,018-0,001) (Slika 26). Prve dvije osi ordinacije zajedno objašnjavaju 29,44% (u zajednicama mrava) do čak 71,34% varijacije u sastavu vrsta (u zajednicama ravnokrilaca uzorkovanih lovnim posudama). U većine skupina vidljivo je jasno odvajanje postaja na šumskom rubu od postaja u preostala dva tipa staništa po 1. osi ordinacije (Slika 26), s izuzetkom jednakonožaca i mrava, kod kojih je prisutno znatno preklapanje postaja u sva tri tipa staništa (Slika 26be)
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 94
Slika 26. Ordinacijski dijagrami analize redundancije s prikazom odnosa vrsta (plave strelice): a) trčaka (Carabidae), b) mrava (Formicidae), ravnokrilaca (Orthoptera) uzorkovanih c) lovnim posudama, d) kečerom, e) jednakonožaca (Isopoda) te f) praživotinja (Protozoa) i mjerenih okolišnih čimbenika (crvene strelice). Postaje u središnjem (vlažnom) dijelu creta Trstenik prikazane su svijetlosivim, u sukcesijskom (suhom) dijelu creta tamnosivim te na šumskom rubu crnim kvadratićima. Puna imena vrsta uključenih u analizu navedena su u prilogu.
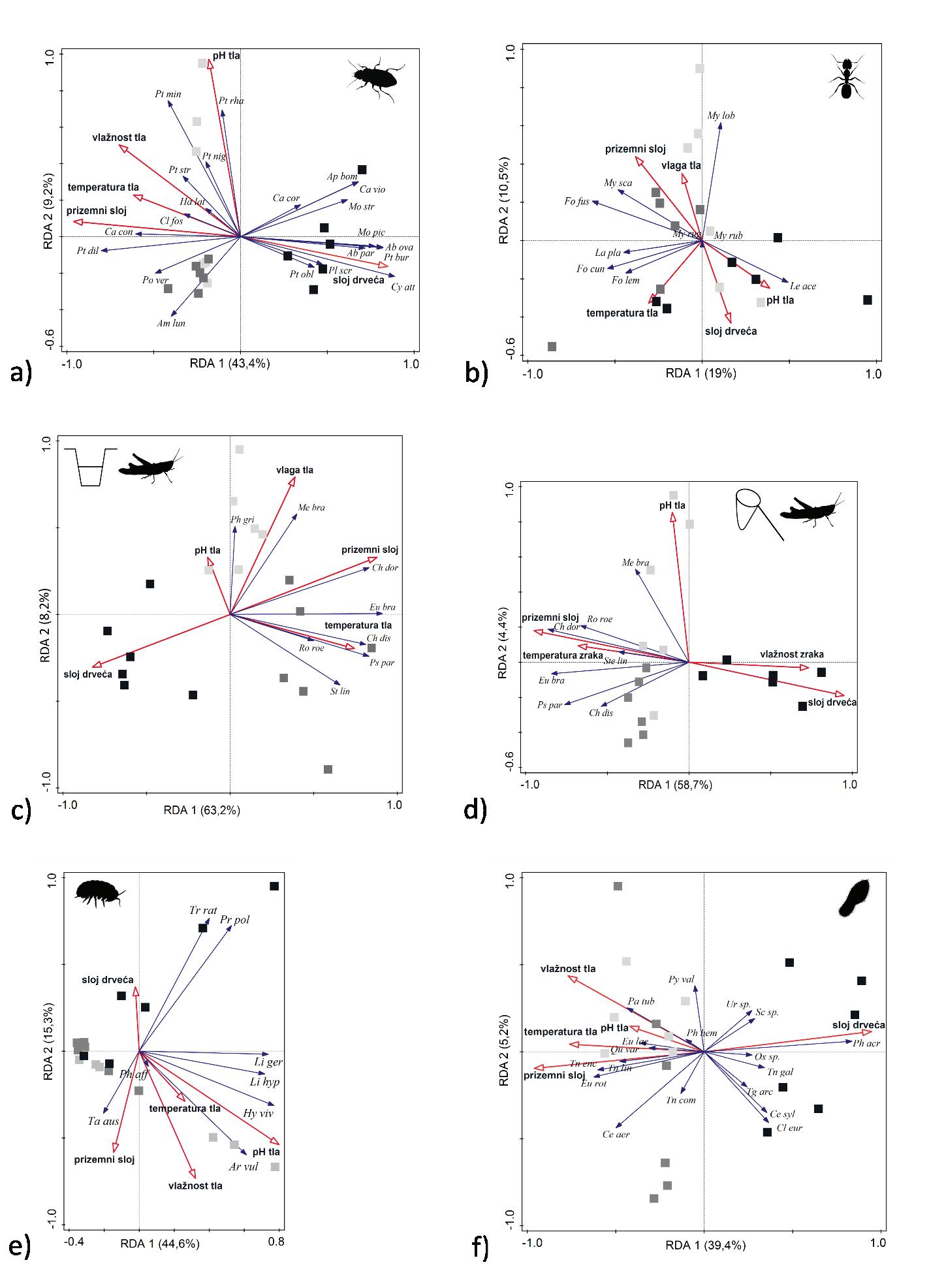
Najvažniji prediktor sastava zajednica na šumskom rubu bila je povećana zastupljenost sloja drveća (Slika 26), no prostorna raspodjela ravnokrilaca uzorkovanih kečerom bila je određena i povišenim vrijednostima vlažnosti zraka u ovom tipu staništa (Slika 26d). S druge strane, na cretnom staništu najveći je utjecaj na sastav vrsta gotovo svih indikatorskih skupina imala visoka zastupljenost prizemnog sloja
vegetacije uz povišene vrijednosti temperature tla odnosno zraka te, osobito u središnjem dijelu creta, povišena vlažnost i pH vrijednost tla (Slika 26)
95 BIORAZNOLIKOST
4. PRIJEDLOG MJERA PRILAGODBE KLIMATSKIM PROMJENAMA
4.1. Mjere prilagodbe šumskih ekosustava na direktne posljedice klimatskih promjena – šumski požari, ledolomi i vjetroizvale
Istraživanjem utjecaja požara na biološku raznolikost šumskih ekosustava u NP Sjeverni Velebit, u većine indikatorskih skupina terestričkih organizama nisu zabilježene razlike u biocenološkim parametrima zajednice na opožarenim površinama u odnosu na neutjecani šumski matriks. S jedne strane, tome vjerojatno doprinosi mala površina opožarenih staništa u kombinaciji sa značajkama istraživanih šumskih sastojina; primorske bukove šume su svijetle, termofilne, s velikom zastupljenošću malih prirodnih čistina, zbog čega je kontrast između šumskog matriksa i čistina nastalih uslijed požara razmjerno slab. Upravo su prirodne šumske čistine vjerojatno poslužile kao koridori u obliku „otoka“ (engl. stepping stones) za naseljavanje opožarenih površina vrstama otvorenih staništa, koje je doprinijelo povećanju raznolikosti pojedinih skupina (npr. ravnokrilaca, mrava) na čistinama nastalim uslijed požara. S druge strane, kako se opožarene površine uglavnom nalaze na nedostupnim lokacijama, na njima nije provođen nikakav oblik gospodarenja (npr. sječa, uklanjanje srušenih debala itd.) te su prepuštene prirodnoj sukcesiji vegetacije, uslijed koje je kroz četiri godine došlo do djelomične obnove šumskog matriksa. Posljedično, došlo je i do obnove većine zajednica koje naseljavaju ova staništa, o čemu svjedoči izostanak odgovora na poremećaj uzrokovan požarom.
Na temelju svega navedenog, kao mjere prilagodbe šumskih ekosustava na negativne posljedice požara, sve učestalije i raširenije pojave uslijed klimatskih promjena, iz perspektive biološke raznolikosti predlaže se: 1) u slučaju požara manjih razmjera (zahvaćene manje površine šumskog staništa), ne uklanjati srušena i oštećena stabla kako bi se u što većoj mjeri očuvala zatvorenost sklopa i omogućila prirodna obnova šumskog staništa sukcesijom vegetacije te 2) u slučaju požara većih razmjera (zahvaćene veće površine šumskog staništa), kada nije moguća dovoljno brza prirodna obnova sukcesijom vegetacije, pošumljavati zahvaćene površine vrstama drveća koje grade okolni šumski matriks.
4.2. Mjere prilagodbe šumskih ekosustava na direktne posljedice klimatskih promjena
ledolomi i vjetroizvale
Istraživanjem utjecaja ledoloma i vjetroizvala na biološku raznolikost šumskih ekosustava na području Risnjaka ustanovljeno je da tip gospodarenja zahvaćenim šumskim površinama ima daleko veći utjecaj na njihovu biološku raznolikost nego sam poremećaj. Sječom i uklanjanjem srušenih odnosno oštećenih stabala, na gospodarenim površinama došlo je do gotovo potpunog otvaranja sklopa krošanja, čime su nastale razmjerno velike površine otvorenog staništa okružene nezahvaćenim šumskim matriksom.
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 96
–
Izrazit kontrast u abiotičkim i vegetacijskim značajkama ovih dvaju tipova staništa rezultirao je promjenama u zajednicama većine indikatorskih skupina terestričkih organizama. Pritom, na gospodarenim površinama, lokalno povećanje raznolikosti (npr. trčaka, mrava) uslijed kolonizacije vrstama otvorenih staništa prati smanjenje brojnosti jedinki (npr. trčaka, ravnokrilaca) zbog smanjene pogodnosti staništa za inače vrlo brojne šumske vrste (npr. Aptinus bombarda, Troglophilus neglectus), čime su znatno narušeni sastav i struktura šumskih zajednica. Nasuprot tome, negospodarene površine zahvaćene ledolomom i vjetroizvalama, koje su bile prepuštene prirodnoj obnovi šumskog matriksa sukcesijom vegetacije, sedam godina nakon poremećaja podržavaju jednaku brojnost jedinki, bogatstvo vrsta i raznolikost gotovo svih indikatorskih skupina kao nezahvaćeni šumski matriks.
Na temelju svega navedenog, kao mjere prilagodbe šumskih ekosustava na negativne posljedice ledoloma i vjetroizvala, uslijed sve veće učestalosti ekstremnih klimatskih događaja, iz perspektive biološke raznolikosti predlaže se: 1) ne uklanjati srušena i oštećena stabla, uz eventualno selektivno uklanjanje većih trupaca u svrhu omogućavanja prohodnosti, kako bi se u što većoj mjeri očuvala zatvorenost sklopa i osigurala dostupnost mikrostaništa te 2) posebnu pažnju posvetiti održavanju prebornosti odnosno prirodne slojevitosti šumske vegetacije, čime se povećava potencijal za prirodnu obnovu šumskog staništa nakon daljnjih poremećaja.
4.3. Mjere prilagodbe cretnog ekosustava na sukcesijske i hidrološke promjene uvjetovane klimatskim promjenama
Istraživanjem utjecaja sukcesijskih i hidroloških promjena na biološku raznolikost cretnog ekosustava na Trsteniku, ustanovljeno je da se raznolikost većine indikatorskih skupina ne razlikuje između središnjeg (vlažnog) i sukcesijskog (suhog) dijela creta, što upućuje na narušeno hidrološko stanje i uznapredovalu sukcesiju modrom beskoljenkom (Molinia caerulea) u oba dijela creta. Iako su na cretu zabilježene tirfofilne indikatorske vrste, poput trčaka Pterostichus rhaeticus i P. diligens, s obzirom na uznapredovalu sukcesiju njihov je dugoročni opstanak, kao i opstanak mahova tresetara (rod Sphagnum), više nego upitan. S druge strane, u zajednicama većine indikatorskih skupina terestričkih organizama zabilježene su promjene na šumskom rubu u odnosu na sam cret. S obzirom na ekološke preferencije pojedinih skupina, odgovori su bili pozitivni (povećanje raznolikosti na rubu zbog prisustva i vrsta otvorenih staništa i šumskih vrsta, npr. trčci, mravi) ili negativni (skupine koje preferiraju otvorene tipove staništa poput ravnokrilaca, odnosno povišenu vlažnost tla poput praživotinja).
Temeljem gore navedenih rezultata, predlažu se sljedeće mjere prilagodbe cretnog ekosustava na sukcesijske i hidrološke promjene uvjetovane promjenama klime, iz perspektive biološke raznolikosti: 1) unaprjeđenje hidrološkog stanja creta, u prvom redu povišenjem vodnog lica na cretu te sprječavanjem otjecanja vode s creta, što se postiže kompleksnim restauracijskim zahvatima (npr. ugradnja vodonepropusnih podloga), 2) uklanjanje drvenastih i zeljastih biljaka koje zauzimaju cret, pri čemu je nužno ukloniti sve dijelove biljke, uključujući korijen, 3) redovita košnja modre beskoljenke s ciljem ograničavanja i usporavanja daljnje sukcesije te 4) presađivanje mahova tresetara (autohtone vrste koje naseljavaju cret) s ciljem obnove cretnog staništa odnosno osiguravanja povoljnih uvjeta za opstanak cretnih vrsta i zajednica.
97 BIORAZNOLIKOST
Literatura
Alegro, A., Topić, J., 2017. Croatia, in: Joosten, H., Tanneberger, F., Moen, A. (Eds.), Mires and Peatlands of Europe: Status, Distribution and Conservation. Schweizerbart Science, Stuttgart, pp. 329–335.
Angulo, E., Boulay, R., Ruano, F., Tinaut, A., Cerdá, X., 2016. Anthropogenic impacts in protected areas: Assessing the efficiency of conservation efforts using Mediterranean ant communities. PeerJ 2016. https://doi.org/10.7717/peerj.2773
Antonović, I., Brigić, A., Sedlar, Z., Bedek, J., Šoštarić, R., 2012. Terrestrial isopod community as indicator of succession in a peat bog. Zookeys 176, 171–188. https://doi.org/10.3897/zookeys.176.2379
Balvanera, P., Quijas, S., Karp, D.S., Ash, N., Bennett, E.M., Boumans, R., Brown, C., Chan, K.M.A., Chaplin-Kramer, R., Halpern, B.S., Honey-Rosés, J., Kim, C.-K., Cramer, W., Martínez-Harms, M.J., Mooney, H., Mwampamba, T., Nel, J., Polasky, S., Reyers, B., Roman, J., Turner, W., Scholes, R.J., Tallis, H., Thonicke, K., Villa, F., Walpole, M., Walz, A., 2017. Ecosystem Services, in: Walters, M., Scholes, R. (Eds.), The GEO Handbook on Biodiversity Observation Networks. Springer International Publishing, Cham, pp. 39–78. https://doi.org/10.1007/978-3-319-27288-7_3
Bardi, A., Papini, P., Quaglino, E., Biondi, E., Topić, J., Milović, M., Pandža, M., Kaligarič, M., Oriolo, G., Roland, V., Batina, A., Kirin, T., 2016. Karta prirodnih i poluprirodnih ne-šumskih kopnenih i slatkovodnih staništa Republike Hrvatske. AGRISTUDIO s.r.l., TEMI S.r.l., TIMESIS S.r.l., HAOP.
Barkman, J.J., Doing, H., Segal, S., 1964. Kritische Bemerkungen und Vorschläge zur Quantitativen Vegetationsanalyse. Acta Botanica Neerlandica 13, 394–419.
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., Courchamp, F., 2012. Impacts of climate change on the future of biodiversity. Ecol Lett 15, 365–377. https://doi.org/10.1111/j.1461-0248.2011.01736.x
Bellmann, H., Rutschmann, F., Roesti, C., Hochkirch, A., 2019. Der Kosmos Heuschreckenführer: Die Heuschrecken Mitteleuropas und die Wichtigsten Arten Südeuropas, Second Edition. ed. Kosmos Verlag, Stuttgart, Germany.
Bhaskar, D., Easa, P.S., Sreejith, K.A., Skejo, J., Hochkirch, A., 2019. Large scale burning for a threatened ungulate in a biodiversity hotspot is detrimental for grasshoppers (Orthoptera: Caelifera). Biodivers Conserv 28, 3221–3237. https://doi.org/10.1007/s10531-019-01816-6
Brigić, A., Bujan, J., Alegro, A., Šegota, V., Ternjej, I., 2017. Spatial distribution of insect indicator taxa as a basis for peat bog conservation planning. Ecol Indic 80, 344–353. https://doi.org/10.1016/j. ecolind.2017.05.007
Bujan, J., Brigić, A., Sedlar, Z., Šoštarić, R., 2015. Progressive vegetation succession of fen habitats promotes the lack of habitat specialist ants. Insectes Soc 62, 415–422. https://doi.org/10.1007/ s00040-015-0420-8
Caddy-Retalic, S., Hoffmann, B.D., Guerin, G.R., Andersen, A.N., Wardle, G.M., McInerney, F.A., Lowe, A.J., 2019. Plant and ant assemblages predicted to decouple under climate change. Divers Distrib 25, 551–567. https://doi.org/10.1111/ddi.12858
Clarke, K.R., Gorley, R.N., 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth, U.K.
Deutsch, C.A., Tewksbury, J.J., Huey, R.B., Sheldon, K.S., Ghalambor, C.K., Haak, D.C., Martin, P.R., 2008. Impacts of climate warming on terrestrial ectotherms across latitude. PNAS 105, 6668–6672
Eionet, 2018. Reference portal for Natura, 2000 [WWW Document]. https://bd. eionet.europa.eu/activities/Natura_2000/reference_portal. [accessed 18 September 2022].
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 98
FAO, 2013. Climate change guidelines for forest managers, FAO Forestry No. 172. ed. Food and Agriculture Organization of the United Nations, Rome, Italy.
Fartmann, T., Krämer, B., Stelzner, F., Poniatowski, D., 2012. Orthoptera as ecological indicators for succession in steppe grassland. Ecol Indic 20, 337–344. https://doi.org/10.1016/j.ecolind.2012.03.002
Foissner, W., 1992. Estimating the Species Richness of Soil Protozoa Using the “non-flooded Petri dish method,” in: Lee, J.J., Soldo, A.T. (Eds.), Protocols in Protozoology. Allen Press, Lawrence, Kansas, pp. B101–B102.
Freude, H., Harde, K.-W., Lohse, G.A., Klausnitzer, B., 2006. Die Käfer Mitteleuropas. II Band. Adephaga
1. Elsevier GmbH.
Harz, K., 1975. The Orthoptera of Europe II. Dr. W. Junk N.V. - Publishers. https://doi.org/10.1007/97894-017-2513-2_1
Harz, K., 1969. The Orthoptera of Europe I. Dr. W. Junk N.V. - Publishers. Hochkirch, A., Nieto, A., Criado, M.G., Cálix, M., Braud, Yoan., Buzzetti, F.M., Chobanov, Dragan., Willemse, Luc., Zuna-Kratky, Thomas., Vega, Pablo.B., Bushell, Mark., Clemente, M.Eulalia., Correas, J.R., Dusoulier, R., Ferreira, Sónia., Fontana, Paolo., García, M.Dolores., Heller, K.-G., Iorgu, I.S., Ivković, Slobodan., Kati, Vassiliki., Kleukers, Roy., Krištín, Anton., Lemonnier-Darcemont, Michèle., Lemos, Paulo., Massa, Bruno., Monnerat, Christian., Papapavlou, K.P., Prunier, Florent., Pushkar, Taras., Roesti, Christian., Rutschmann, Florin., Şirin, Deniz., Skejo, Josip., Szövényi, Gergely., Tzirkalli, Elli., Vedenina, Varvara., Domenech, Joan.B., Barros, Francisco., Tapia, P.J.C., Defaut, Bernard., Fartmann, Thomas., Gomboc, Stanislav., Gutiérrez-Rodríguez, Jorge., Holuša, Jaroslav., Illich, Inge., Karjalainen, Sami., Kočárek, Petr., Korsunovskaya, Olga., Liana, Anna., López, Heriberto., Morin, Didier., Olmo-Vidal, J.M., Puskás, Gellért., Savitsky, Vladimir., Stalling, Thomas., Tumbrinck, Josef., 2016. European Red List of grasshoppers, crickets and bush-crickets. Publications Office of the European Union. https://doi.org/10.2779/60944
Hölldobler, B., Wilson, E.O., 1990. The Ants. Springer Berlin Heidelberg, Berlin, Heidelberg. https://doi. org/10.1007/978-3-662-10306-7
Hršak, V., 1996. Vegetation succession at acidic fen near Dubravica in the Hrvatsko Zagorje region. Natura Croatica 5, 1–10.
Hůrka, K., 1996. Carabidae of the Czech and Slovak Republics. Ing. Vit Kabourek. IBM Corp., 2013. IBM SPSS Statistics for Windows, Version 22.0.
Javna ustanova Park prirode Velebit, 2017. Izvještaj o opožarenim površinama na području Parka prirode Velebit. Gospić.
Joosten, H., Tapio-Biström, M.-L., Tol, S., 2012. Peatlands - guidance for climate change mitigation through conservation, rehabilitation and sustainable use, Second Edition. ed. Food and Agriculture Organization of the United Nations and Wetlands International, Rome, Italy.
Jost, L., 2006. Entropy and diversity. Oikos 113, 363–375. https://doi.org/10.1111/j.2006.00301299.14714.x
Lepš, J., Šmilauer, P., 2014. Multivariate Analysis of Ecological Data using CANOCO 5, 2nd edition. Cambridge University Press. https://doi.org/https://doi.org/10.1017/CBO9781139627061
99 BIORAZNOLIKOST
Longino, J.T., 2000. What to do with the data, in: Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R. (Eds.), Measuring and Monitoring Biological Diversity: Standard Methods for Ground-Living Ants. Smithsonian Institution Press, Washington, D.C., USA, pp. 186–203.
Lövei, G., Sunderland, K.D., 1996. Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu Rev Entomol 41, 231–56.
Magrini, M.J., Freitas, A.V.L., Uehara-Prado, M., 2011. The effects of four types of anthropogenic disturbances on composition and abundance of terrestrial isopods (Isopoda: Oniscidea). Zoologia 28, 63–71. https://doi.org/10.1590/S1984-46702011000100010
Mansoor, S., Farooq, I., Kachroo, M.M., Mahmoud, A.E.D., Fawzy, M., Popescu, S.M., Alyemeni, M.N., Sonne, C., Rinklebe, J., Ahmad, P., 2022. Elevation in wildfire frequencies with respect to the climate change. J Environ Manage. https://doi.org/10.1016/j.jenvman.2021.113769
Mlynář, Z., 1977. Revision der Arten und Unterarten der Gattung Molops Bon. (s. str.) (Coleoptera, Carabidae). Folia Entomologica Hungarica 30, 3–150.
Modrić Surina, Ž., 2011. Ecological gradients as determinants of different vegetation types on mires in Croatia (Doctoral Thesis). University of Zagreb, Faculty of Science, Zagreb.
Moretti, M., Obrist, M.K., Moretti, P.D., Duelli, M.K., 2004. Arthropod biodiversity after forest fires: winners and losers in the winter fire regime of the southern Alps. Ecography 27, 173–186.
Naeem, S., Chazdon, R., Duffy, J.E., Prager, C., Worm, B., 2016. Biodiversity and human well-being: An essential link for sustainable development. Proceedings of the Royal Society B: Biological Sciences 283. https://doi.org/10.1098/rspb.2016.2091
Payne, R.J., 2013. Seven reasons why protists make useful bioindicators. Acta Protozool 52, 105–113. https://doi.org/10.4467/16890027AP.13.0011.1108
Perica, D., Orešić, D., 1999. Klimatska obilježja Velebita i njihov utjecaj na oblikovanje reljefa. Senjski zbornik 26, 1–50.
Pešić, K., 2019. Feromonski monotoring smrekovih potkornjaka Ips typographus i Pityogenes chalcographus na području NP “Risnjak” (Professional Thesis). University of Zagreb, Faculty of Forestry, Zagreb, Croatia.
Radchenko, A., Elmes, G.W., 2010. Myrmica ants (Hymenoptera: Formicidae) of the Old World. Natura optima dux Foundation, Warszawa, Poland.
Rainio, J., Niemela, J., 2003. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers Conserv 12, 487–506.
Randić, M., 2018. Osobitosti biljnog svijeta šireg područja Šverde. Subterranea Croatica 16, 31–36.
Regent, 2006. WinSCANOPY for hemispherical image analysis - Manual.
RStudio Team, 2020. RStudio: Integrated Development for R.
Sardet, É., Roesti, C., Braud, Y., 2015. Cahier d’identification des orthoptères de France, Belgique, Luxembourg et Suisse. Biotope Éditions.
Šegota, T., Filipčić, A., 2003. Köppenova podjela klima i hrvatsko nazivlje. Geoadria 8, 17–37. https://doi. org/10.15291/geoadria.93
Mjere prilagodbe klimatskim promjenama za održivo upravljanje prirodnim resursima 100
Seifert, B., 2018. The Ants of Central and Northern Europe. Lutra Verlags und Vertriebsgesellschaft, Boxberg, Baden-Württemberg, Germany.
Sertić, M., 2016. Zaštita šuma od ledoloma na primjeru šumske štete 2014. godine u UŠP Karlovac, UŠP Ogulin, UŠP Delnice i Nacionalnom parku Risnjak (Bachelor Thesis). Karlovac University of Applied Sciences, Karlovac, Croatia.
Šimić Milas, A., Rupasinghe, P., Balenović, I., Grosevski, P., 2015. Assessment of Forest Damage in Croatia using Landsat-8 OLI Images. South-east European forestry 6. https://doi.org/10.15177/seefor.15-14
Šišić, S., Randić, M., Stipanović, M., 2009. Cret / mire Trstenik.
Šoštarić, M., 2016. Prirodno pomlađivanje nakon ledoloma u jelovo-bukovim sastojinama na području Gerova (Master Thesis). University of Zagreb, Faculty of Forestry, Zagreb, Croatia.
ter Braak, C.J.F., Šmilauer, P., 2012. Canoco reference manual and user’s guide: software for ordination. Microcomputer Power, Ithaca, USA.
Topić, J., Stančić, Z., 2006. Extinction of fen and bog plants and their habitats in Croatia. Biodivers Conserv 15, 3371–3381. https://doi.org/10.1007/s10531-005-4874-2
Trautner, J., Geigenmüller, K., 1987. Tiger beetles and ground beetles, illustrated key to the Cicindelidae and Carabidae of Europe. Josef Margraf.
Turin, H., Penev, L., Casale, A., 2003. The Genus Carabus in Europe: A Synthesis, Fauna Euro. ed. Pensoft Publishers & European Invertebrate Survey.
Walther, G.R., 2010. Community and ecosystem responses to recent climate change. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2019–2024. https://doi.org/10.1098/ rstb.2010.0021
Zaninović, K., Gajić-Čapka, M., Tadić, M.P., Vučetić, M., Milković, J., Bajić, A., Cindrić, K., Cvitan, L., Katušin, Z., Kaučić, D., Likso, T., Lončar, E., Lončar, Ž., Mihajlović, D., Pandžić, K., Patarčić, M., Srnec, L., Vučetić, V., 2008. Climate atlas of Croatia: 1961-1990; 1971-2000. Croatian Meteorological and Hydrological Service.
101 BIORAZNOLIKOST
Bilješke
Sveučilište u Zagrebu Fakultet šumarstva i drvne tehnologije
PARTNERI: DIONICI:








Fakultet šumarstva i drvne tehnologije Sveučilišta u Zagrebu
Prirodoslovno-matematički fakultet Sveučilišta u Zagrebu


Agronomski fakultet Sveučilišta u Zagrebu