


The!Cupola!Award!is!presented!to!no!more!than!five!students!whose! research!papers!rank!in!the!top!from!the!papers!submitted.!!Winners! receive!$500!and!their!paper!published!in!this!journal,!CNU’s!web!based! journal!for!undergraduate!research.!!The!top!Cupola!paper!receives!the! Douglas!K.!Gordon!Cupola!Award.
"Research is the essential element of the scholar's craft. It is the means by which scholars pay forward into the fund of human knowledge the debt owed to their predecessors. At its best, research honors the past while enriching the future."
Dr. Richard M. Summerville CNU Provost 1982-1995, 2001-2007Table!of!Contents
Title!and!Author
Integrin!signaling!differentiation!in!Caenorhabditis-elegans.
By Danielle!DaCrema Douglas!K.!Gordon!AwardSwitzerland: The Relationship between Neutrality and European Union Membership
By Kerry CampbellA!Study of!Competition!Between Native!Eastern!Oyster
By Katherine!Frances!HoltIntegrin signaling differentiation in Caenorhabditis elegans
Danielle DaCremaDanielle.dacrema.09@cnu.edu
703-403-1368
Faculty Sponsor: Dr. Christopher Meighan
Department of Molecular Biology and Chemistry
Christopher.meighan@cnu.edu
757-594-7454
Abstract
Integrins are transmembrane receptors containing α- and β- subunits that are responsible for connecting the cell to its environment by directly interacting with the extracellular matrix and connecting to the cytoskeleton through large signaling pathways. Many signaling partners of the intracellular domains of integrins have been identified, but specific pathways are largely unknown. As a complication, different integrins are capable of being expressed in the same tissue, at the same time, and still have different functions. Caenorhabditis elegans, a common model organism for integrin study, has two α-integrins, ina-1 and pat-2, and one β-integrin, pat-
3. RNA interference was used to knock down expression of two potential integrin signaling partners, pat-4 and pat-6, in seven strains of C. elegans. Data were compared to empty vector controls and evaluated for proper distal tip cell migration, egg viability, and body morphology to gain insights into integrin function in three tissue types: distal tip cells, gonad arms, and muscles
It was determined that pat-4 is likely to signal with pat-3 in the distal tip cells, gonad arms, and muscles. Pat-6 was determined to be likely to signal with pat-3 in the distal tip cells, with ina-1 in the gonad arms, and is unlikely to signal with integrins in the muscles. This adds pat-4 and pat-6 to integrin signaling pathways including src-1, pxl-1, nischarin, and talin in these three tissue types.
Introduction
A cell must be able to interact with its environment in order to survive. One of the most understood and studied ways this is done in all animals is through a family of cell-surfaceadhesion receptors known as integrins.1,2,3 Integrins (Figure 1A) are transmembrane receptors with alpha (α-integrin) and beta (β-integrin) subunits that noncovalently associate with each other to form a heterodimer.4 The extracellular “head” domains connect to various components of the extracellular matrix (ECM) while the intracellular “tail” domains connect to the cytoskeleton.3,4,5 Although the integrin heads bind directly to specific components of the ECM, the association between integrin tails and the cytoskeleton occurs through very large signaling pathways (Figure 1B).1,4,6 As of 2007, 159 integrin signaling partners have been identified in humans.1 The control of these pathways is multifaceted, and dependent on ECM components near the cell as well as variable expression of integrins and their signaling partners within the cell 7 Simultaneous connections of the ECM and the cytoskeleton through integrins allow signals to be relayed bidirectionally across the membrane and function to control gene regulation, protein synthesis, cytoskeletal rearrangements, cellular migration, cell trafficking, cell division, differentiation, and cell survival (Figure 1B).1,2,3,8 At the tissue and organismal level these cellular regulations are critical for development, immune response, homeostasis maintenance, and wound repair.2,6 They have also been found to play roles in many diseases, most notably cancer and various autoimmune diseases, and are used by many types of viruses and bacteria as a receptor on the cell surface.2,6 The incredible number of cellular responsibilities and signaling partners of integrins has led them to be considered one of the major signaling hubs of the cell, and their study is of great interest to the scientific community.1,2,3,4,9
Figure 1.4,10 Integrins are heterodimeric transmembrane receptors that are responsible for many cellular activities. (A) Integrin receptors are comprised of noncovalently attached α- and β- subunits.1,4 Each subunit has distinct extracellular, transmembrane, and intracellular domains.4 (B) While attached to the ECM, integrins function through large signaling pathways to control many aspects of cellular life.4 Although many integrin signaling partners have been identified, most of the pathways controlling each activity are presently unknown.8 Integrin study in mammals is complicated by the presence of 24 function integrin heterodimers formed from 18 α-integrins and 8 βintegrins.2,5 To simplify the system, the nematode Caenorhabditis elegans (Figure 2), a common model organism, are often used for integrin studies.3,11,12,13 C. elegans have two functional heterodimers formed from two α-integrins, ina-1 and pat-2, and one β-integrin, pat-3, greatly decreasing the complexity presented by mammalian integrin studies.12 Other advantages of C. elegans include the small (approximately 1 mm in length) and translucent bodies with welldefined organs, rapid and hermaphroditic reproductive cycles, a sequenced genome with 50-80 % homology to humans, and ease of growth and care.11 These characteristics allow for easy study of experimental effects on the nematode body relatively quickly and without worry of


genetic contamination by accidental breeding, while still providing insights for humans and other mammalian models.
Figure 2.14 C. elegans are hermaphrodites with relatively simple anatomy and easily identifiable organs. The head region includes the pharynx, which leads into the intestine. Most of the nematode body is filled by the intestines and the gonad arms fill the remaining half. The whip-like tail is used for locomotion. The vulva lies in approximately the center of the nematode body and is for egg laying. During larval development, the gonad arms grow in opposite directions, change sides near the ends of the nematode body, and grow back towards the vulva. The size bar indicates 0.1 mm.14
Even in the simplified system of C. elegans, integrins have many signaling partners and the understanding of integrins is complicated by the fact that the α-integrins, ina-1 and pat-2, are frequently expressed in the same cells, may have different functions, and can also have different signaling partners in different tissue types.12,13,15 I previously identified three integrin signaling partners and classified them as signaling with ina-1, pat-2, or pat-3 in three different tissue types by knocking down expression of potential signaling partners using RNA interference (RNAi) in various strains, and then analyzing egg viability, body morphology, and distal tip cell (DTC)

migration (Figure 3, data in Table S1 and Figures S1 and S2). This work describes the addition of two other signaling proteins, pat-4 and pat-6, to these pathways.
Figure 3. During C. elegans development, pat-3 signals with talin and src-1 to cause proper DTC migration; in adulthood pat-3 signals with talin to cause proper oocyte formation in the gonad arms; and throughout life pat-3 signals with talin and src-1 while pat-2 signals with pxl-1 to cause proper muscle function. Four targets were analyzed: src-1, pxl-1, nischarin, and talin Although no direct evidence for pat-3 signaling was analyzed, the protein targets were considered likely to work with integrins and found not to signal with α-integrins, they were thus concluded to work with the β-integrin. Nischarin was not found to signal with integrins in these tissues. These conclusions were based on analysis of DTC migration, body morphology, and egg viability (data in Table S1 and Figures S1 and S2) in several strains when RNAi was used to knock down expression of each potential target.

Materials & Methods
In order to study signaling partners of integrins several strains of transgenic nematodes containing chimeric integrins were used. Transgenic nematodes have had extra DNA, in this case a specific integrin and genes for several identifying phenotypes, inserted into their genome.
Chimeric integrins are transgenes with the extracellular domain of one integrin and the intracellular domain of another. By the use of chimeric integrins in dominant-negative integrin strains (strains containing a loss-of-function mutation in one of their integrin subunits), it is possible to ensure expression of only one α-integrin on the inside of the cell, while maintaining normal integrin expression outside of the cell. This is a fairly common and well established method to study transmembrane proteins.2,15,16
Since chimeric integrins are not naturally produced by any organism and transgenic strains have had alterations made to their genome, these studies are done with many controls. For this work, seven different strains were used (Figure 4). Ina-1/pat-2 cyto and pat-2/ina-1 cyto (Figure 4A), are the two strains with a dominant-negative background. Ina-1/pat-2 cyto nematodes express the pat-3 β-integrin and the pat-2 α-integrin normally yet have nonfunctional ina-1. To combat the loss of ina-1, they also have a transgenic chimeric integrin with the extracellular ina-1 domain and the intracellular pat-2 domain, this integrin is called ina-1/pat-2
The pat-2/ina-1 cyto nematodes, on the other hand express functional pat-3 and ina-1, nonfunctional pat-2, and transgenic pat-2/ina-1. The wild type, ina-1 rescue, and pat-2 rescue strains are three control strains effectively express both α-integrins and the β-integrin (Figure 4B). Wild type nematodes have no transgenes and functional copies of all integrins. Ina-1 rescue nematodes have nonfunctional ina-1 but are phenotypically rescued by expression transgenic ina-1. Likewise, pat-2 rescue nematodes have nonfunctional pat-2 but functional transgenic pat-2. Two additional strains, pat-2 WT background and pat-2/ina-1 cyto WT background, contain functional copies of all integrins as well as transgenic integrins, in one case pat-2 and in the other a chimeric integrin (Figure 4C). Main source populations for each strain
were maintained on 6 mm agar plates seeded with the slow growing strain of Escherichia coli, OP50, at room temperature; this is standard for C. elegans maintenance.11
Figure

expression of α-integrins in the seven C. elegans strains studied was varied for control and experimental purposes using transgenic and chimeric integrins. All strains express the βintegrin, pat-3, normally, and every α-integrin must dimerize with pat-3 in order to function, thus pat-3 is not shown in order to simplify the diagrams. (A) Pat-2/ina-1 cyto and ina-1/pat-2 cyto are dominantnegative strains with transgenic chimeric integrins such that extracellular expression is normal and intracellular expression is limited to one type of α-integrin. (B) Wild type, ina-1 rescue, and pat-2 rescue are strains that express both α-integrins. In wild type nematodes this is done naturally while the rescue


strains have one α-integrin that is dominant-negative and the phenotype is rescued by a functional transgenic copy of the gene. (C) Pat-2 WT background and pat-2/ina-1 cyto WT background are strains with wild type backgrounds that also have had transgenes introduced into their genome.
Expression of a potential signaling partner was suppressed using RNAi. This technique takes advantage of a cellular defense against viruses.17 Normal eukaryotic cells, like those found in plants and animals, do not contain double stranded RNA (dsRNA), while certain types of viruses do.17 Without intervention, the virus would highjack the cell, the dsRNA of the virus would be translated by the cell’s own components into viral proteins and dsRNA, which would cause the formation of more virions (virus particles).17 When enough virions have been accumulated, the cell would lyse, releasing the particles to infect neighboring cells.17 As a defense, the cell produces proteins that search for unusual RNA (such as dsRNA) then triggers the degradation of all RNA sequences matching the sequence of any that was found.17 By doing this, the cell prevents the production of viral proteins and replication of the viral genome.17
RNAi is a technique that introduces dsRNA matching the messenger RNA (mRNA) for a specific protein.17 Because of the match to the dsRNA, these mRNAs are degraded and the protein is prevented from being produced, effectively silencing or knocking-down expression of the protein.17 In C. elegans, the introduction of dsRNA is quite simple. Since C. elegans are grown on bacteria, RNAi experiments can be easily performed by prepping nematodes onto agar plates seeded with bacteria that produce dsRNA (Figure 5).18 During digestion, the dsRNA is absorbed and distributed throughout the cells, subsequently triggering the degradation of all matching mRNA.18 For these experiments, expression of the dsRNA in the bacteria is dependent on the presence of isopropyl β-D-1-thigalactopyranoside (IPTG), which is infused into the agar plates used for RNAi. As a control, every RNAi experiment includes an empty vector target. Empty vector bacteria produce dsRNA, but the sequence does not match any protein target.
Effectively, the empty vector works the same way as running a blank through a chemical instrument.
a loss of phenotype associated with that protein.18,19
Analysis was done using two quantitative methods measuring egg viability and DTC migration, as well as a qualitative analysis of body morphology in order to gain insight into three different tissue types: gonads, muscles, and DTCs. Egg viability was measured in percentage of eggs laid that hatched, and provided information on gonad function, primarily oocyte (unfertilized egg) maturation, vulval muscle function, and muscle function in early larval stages.20 Analysis was done through two generations. The parental (P) generation was introduced to the RNAi plates through a procedure known as an egg prep. This is a common, well established method that involves soaking adults in a bleach/NaOH solution to introduce only the eggs of a population onto a new plate.21 Once the P generation had matured (approximately 3 days), individuals were selected based on transgenic phenotypes, were transferred onto a separate RNAi plate, and allowed to lay eggs for 5 hours before being

removed. These eggs hatched to become the first filial (F1) generation. The percentage of the eggs that hatched provided data on egg viability. DTC migration was analyzed in the P generation.
There are several stages of embryonic development, the last, of course, being hatching (Figure 6). Integrins are involved in early embryonic development and cell fusion. In order for the nematode to hatch, it must extend its body to break the protein layer encasing the egg, which requires the use of muscles throughout the body.14,22 In the adult body, the vulval muscles must contract in order for eggs to be laid.23 However, poor vulva muscle function is not the only reason for no eggs to be laid; it is also possible for oocytes to fuse together (Figure 7), which is indicative of poor gonad function. Since integrins are responsible for focal adhesions and connections to the extracellular environment, it follows that, with improper integrin function in muscles, the muscle cells will detach from each other when they try to contract. This results in shredding of the muscle tissue and full-body paralysis. Thus, muscle function was determined by body morphology, primarily the presence or absence of full body paralysis.

Figure 6.14,22 Muscles are required for hatching. (A) The life cycle of a nematode involves four stages of embryonic development after laying, four stages of larval development, and adult life 14 (B) There are four notable stages of embryonic development after being laid: comma (B.i.), 1.5 -fold (not shown), 2-fold


(B.ii.), and 3-fold (B.iii.).14,22 In order to hatch from the 3-fold stage and proceed to the first stage of larval development (B.iv.), the nematode must extend its body, which requires the use of muscles.14,22
causes sterility.
There are two DTCs in a nematode body that migrate throughout larval development.12,24
The gonad arms develop following the pattern of DTC migration, which means that DTC migratory patterns can be analyzed in adult nematodes.24 During normal migration, the DTCs migrate independently of each other in opposite directions starting from the vulva.12 They then turn and migrate back towards the center of the body, forming a notable “U” shaped pattern (Figure 8).12 DTCs express both α-integrins, and migration is dependent on integrin interactions with the ECM, integrin-associated proteins, and the cytoskeleton.12 Deviations are considered to be any migratory patterns not consistent with the U-structure. Normal stopping position is considered to be within 2 egg-lengths of the vulva. Although migratory defects can vary in severity, for the purposes of my studies severity is irrelevant and only the presence or absence of defects was considered.

Figure 8 12 DTC migration pattern and gonad arm development in C. elegans is expected to follow a U-shaped pattern and deviations from that pattern may indicate improper integrin signaling in the DTCs. DTCs are guided through the body by interactions between integrins, integrin associated protein complexes, the ECM, and the cytoskeleton. Dotted white lines highlight the growth of one gonad arm starting from the vulva. Images are taken from previous research. (A,B) Normal DTC and gonad arm development follow U-shaped patterns. The gonad arms grow are led by the DTCs during larval development.12 They are expected to come to a stop in the center of the body, near the vulva, after the Ushaped pattern has developed.12 (C,D) Abnormal growth patterns include premature stopping (not displayed), migration past the vulva (D), sharp turns (C), too many turns (C), and no turns (not displayed).12




By these methods, pat-4 and pat-6 were analyzed for specific α-integrin interactions in the gonads, muscles, and DTCs. Data were collected in terms of percentage of eggs hatched, percentage of DTC migratory defects, and whether or not the adults were paralyzed. These methods were previously validated by RNAi against talin, a known β-integrin scaffold protein (data seen in Table S1 and Figures S1 and S2).25 For each trial, strains were maintained at 23 °C on 6 mm agar plates seeded with bacteria producing dsRNA against the target gene.
Results & Discussion
When RNAi was performed against empty vector controls, the P generation of nematodes were not paralyzed (Table 1, Figure 9), laid viable eggs (Table 2, Figure 9), and had few DTC migration defects (Table 3, Figure 10). However, when RNAi was performed against pat-4, the
P generation of nematodes were paralyzed (Table 1, Figure 9), experienced oocyte fusion causing sterility (Table 2, Figures 9 and 11), and experienced more DTC migration defects than the empty vector controls (Table 3, Figure 10) in all strains. When pat-6 was knocked down by RNAi, the P generation of nematodes were not paralyzed (Table 1, Figure 9) and had more DTC migration defects than the empty vector (Table 3, Figure 10) in all strains. Interestingly, when considering oogenesis and egg viability the ina-1/pat-2 cyto and wild type strains produced normal shaped eggs (Figure 11) that were capable of hatching (Table 2, Figure 9), while the other strains produced eggs that were of abnormal shape (Figure 11) that were not laid (Table 2, Figure 9). The abnormally shaped eggs appeared as if some of the oocytes had fused together and others were completely engulfed by their surroundings.
Table 1. Full body paralysis was observed when pat-4 expression was knocked down by RNAi, but not for pat-6 or empty vector controls.
Table 2. Egg viability comparison of pat-4 and pat-6 RNAi and cumulative empty vector controls within 95 % confidence for the seven strains used.
Figure 9. Pat-4 signals independently of α-integrins for oocyte maturation and muscle function, while pat-6 signals with ina-1 for oocyte maturation. The differences between pat-4 and the empty vector RNAi in all the strains indicates no preference for α-integrin signaling. Pat-6 RNAi show significant differences compared to the empty vector in the first five strains, but the removal of ina-1 from inside the cell rescues the phenotype, indicating a signaling partnership. Error bars represent double the standard error which equates to 95 % confidence.
Table 2. Comparison of gonad arm migration defects for target gene and cumulative empty vector controls within 95 % confidence for the seven strains used.

Figure 10. Pat-4 and pat-6 act independently of α-integrins for DTC migration. The differences between both RNAi targets and the empty vector controls in all the strains indicates no preference for αintegrin signaling. Error bars represent double the standard error which equates to 95 % confidence.

Figure 11. When pat-4 expression is knocked down by RNAi, oocyte fusion is observed; in most strains when pat-6 expression is knocked down by RNAi oocyte maturation is limited, but this is recovered in the ina-1/pat-2 cyto and wild type strains. Empty vector controls were capable of




producing eggs normally. When pat-4 was knocked down, nematodes experienced oocyte fusion. In most strains, when pat-6 was knocked down, nematodes were able to produce eggs, but they were of abnormal shape and were not laid; this phenotypes looked as if some of the oocytes had fused and the others were engulfed completely. In the ina-1/pat-2 cyto and wild type strains when pat-6 was knocked down, eggs were produced and laid normally.
The extreme difference between pat-4 and empty vector RNAi experiments when observing motility, egg formation, and DTC migration indicates that pat-4 signals independently of α-integrins in the muscles, gonad arms, and DTCs. Since pat-4 is thought to signal with integrins and was found to be α-integrin independent in these tissues, it is likely to signal with the β-integrin, pat-3.
The difference between pat-6 and empty vector RNAi experiments when observing DTC migration, indicates signaling independence from α-integrins and implies signaling with pat-3 in the DTCs. There were no observable differences in muscle function between pat-6 RNAi and empty vector controls. This indicates signaling independence of integrins in muscle tissue. In the gonad arm the recovery from the phenotype in the ina-1/pat-2 cyto strain indicates that the loss of ina-1 on the inside of the cell made the nematodes immune to the pat-6 phenotype. This implies a signaling partnership between pat-6 and ina-1 in the gonad arm during oocyte maturation. Interestingly, the recovery of the phenotype was also observed in the wild type strain. The wild type strain was the only strain used that does not have any transgenic markers. The transgenic markers are necessary in the other strains to determine if the transgenic integrin was introduced into the genome of the nematode, but also makes the nematodes less resilient to cellular stress. It is likely that the wild type nematodes were able to signal through other pathways during the stress of the pat-6 loss, thereby effectively recovering from the loss of pat-
6, whereas the cells in the transgenic strains were unable to do this or unable to do it efficiently enough to survive.
Conclusion
The previous signaling pathway (Figure 3) has been modified based on the data presented here (Figure 12). Pat-4 was determined to signal independently of α-integrins in the DTCs, gonad arms, and muscles, and is likely to signal with pat-3 in these tissues. This is because the phenotypes for these tissues were very different in pat-4 knockdowns than in the empty vector controls. Pat-6 was determined to signal independently of α-integrins, and is likely to signal with pat-3, in the DTCs due to the differences from the empty vector controls that were observed when pat-6 is knocked down. Conversely, pat-6 signals with ina-1 in the gonad arm for proper oocyte maturation since the inability to produce and lay eggs properly was recovered in the ina1/pat-2 cyto strain (the strain that does not present cytoplasmic ina-1). Since there were no differences between the empty vector controls and pat-6 RNAi in terms of muscle function, it is likely that pat-6 does not signal with integrins in the muscles.
Figure
and
cause proper DTC migration; in adulthood pat-3 signals with talin and pat-4 while ina-1 signals with pat-6 to cause proper oocyte formation in the gonad arm; and throughout life pat-3 signals with talin, src-1, and pat-4 while pat-2 signals with pxl-1 to cause proper muscle function. The data presented here have added pat-4 and pat-6 to the integrin signaling pathways in these tissues. Pat-4 was determined to signal independently of α-integrins in DTCs, gonad arms, and muscles, and is likely to signal with pat-3 in these tissues. Pat-6 was determined to signal independently of α-integrins in the DTCs, and is likely to signal with pat-3 for proper DTC migration. In the gonad arms pat-6 was determined to signal with ina-1 to cause proper oocyte maturation. Additionally, pat-6 was determined to signal independently of integrins in muscles.

Integrins signal through very large, very robust cytoplasmic signaling pathways. 1,2,3,4,8,9
The proteins identified here as signaling with integrins in these tissues form a very small portion of the larger picture. Much more research is needed to determine which other proteins signal with which integrins, in what tissues, and for which functions. These data also do not attempt to identify the order of the signaling pathways, or even direct interactions between the proteins, and
only serve to identify signaling partners. These interactions could be very far up-stream or down-stream of each other. This experimental design simply cannot provide this information, demonstrating the need for additional designs once each partner has been identified.
Acknowledgements
Many thanks to Dr. C. Meighan for providing materials, supervising the work, and assisting in the interpretation of the data. Many thanks to the Summer 2012 Undergraduate Student Research Award from Christopher Newport University, which made results for talin and nischarin possible.
References
1. Harburger, D. and Calderwood, D. Integrin signaling at a glance. Journal of Cell Science 2009, 122, 159-163.
2 Hynes, R. Integrins bidirectional, allosteric signaling machines. Cell 2002, 110, 673-687.
3. Humphries, J.; Byron, A.; Humphries, M. Integrin ligands at a glance. The Journal of Cell Biology 2006, 119, 3901-3903.
4. Plopper, G. The extracellular matrix and cell adhesion. In Lewin’s Cells, 2nd ed.; Cassimeris, L., Lingappa, V., Plopper, G., Eds.; Jones and Bartlett Publishers: Sudbury, Massachusetts, 2011; pp 821-879.
5. Kim, S.; Turnbull, J.; Guimond, S. Extracellular matrix and cell signaling: the dynamic cooperation of integrin, proteoglycan and growth factor receptor. Journal of Endocrinology 2011, 209, 139-151.
6. Daley, W.; Peters, S.; Larsen, M. Extracellular Matrix dynamics in development and regenerative medicine. The Journal of Cell Science 2008, 121, 255-264.
7. Palecek, S.; Loftus, J.; Ginsberg, M.; Lauffenburger, D.; Horwitz, A. Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature 1997, 385, 537-540.
8. Pines, M.; Fairchild, M.; Tanetzapf, G. Distinct regulatory mechanisms control integrin adhesive processes during tissue morphogenesis. Developmental Dynamics 2010, 240, 36-51.
9. Critchley, D. and Gingras, A. Talin at a glance. The Journal of Cell Science 2008, 121, 1345-1347.
10. Seymour, B. Designed to bind: Penn scientists engineer small molecules to probe proteins deep inside cell membrane. Penn Medicine 2007, http://www.uphs.upenn.edu/news/News_Releases/mar07/transmembrane-proteins.html.
11. Hulme, S.; Whitesides, G. Chemistry and the worm: Caenorhabditis elegans as a platform for integrating chemical and biological research. Angeivandte Chemie International Edition 2011, 50, 4774-4807.
12 Meighan, C.; Schwarzbauer, J. Control of C. elegans hermaphrodite gonad size and shape by vab-3/Pax6-mediated regulation of integrin receptors. Genes & Development 2007, 21, 1615-1620.
13. Ou, G.; Vale, R. Molecular signatures of cell migration in C. elegans Q neuroblasts. The Journal of Cell Biology 2009, 185, 77-85.
14. Herndon, L. (ed.). Introduction to C. elegans anatomy. Wormatlas Caenorhabditis elegans 2009, http://www.wormatlas.org/hermaphrodite/introduction/Introframeset.html
15. Lee, M.; Cram, E.; Shen, B.; Schwarzbauer, J. Roles for βpat-3 integrins in development and function of Caenorhabditis elegans muscles and gonads. Journal of Biological Chemistry 2001, 276, 36404-36410.
16. Liu, S.; Calderwood, D.; Ginsberg, M. Integrin cytoplasmic domain-binding proteins. Journal of Cell Science 2000, 113, 3563-3571.
17. Madigan, M.; Martinko, J.; Stahl, D.; Clark, D. Archaeal and eukaryotic molecular biology. Brock Biology of Microorganisms, 13th Ed.; Benjaminh Cummings: San Francisco, California, 2012; pp 191-208.
18. Timmons, L. and Fire, A. Specific interference by ingested dsRNA. Nature 1998, 395, 854.
19 Ahringer, J. Genome-wide RNAi screening in C. elegans. Ahringer Lab, http://www.gurdon.cam.ac.uk/~ahringerlab/pages/res_rnai.html
20. Schafer, W. Egg-laying. WormBook the Online Review of C. elegans Biology 2005, http://www.wormbook.org/chapters/www_egglaying/egglaying.html.
21. Stiernagle, T. Maintenance of C. elegans. WormBook the Online Review of C. elegans Biology 2005,
http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html
22 Ding, M.; Goncharov, A.; Jin, Y.; Chrisholm, A. C. elegans ankyrin repeat protein VAB-19 is a component of epidermal attachment structures and is essential for epidermal morphogenesis. Development 2003, 130, 5791-5801.
23. Herndon, L. (ed.). Reproductive system egg-laying apparatus. Wormatlas Caenorhabditis elegans 2013,
http://www.wormatlas.org/hermaphrodite/egglaying%20apparatus/Eggframeset.html
24. Herndon, L. (ed.). Reproductive system the germ line. Wormatlas Caenorhabditis elegans
2013, http://www.wormatlas.org/hermaphrodite/germ%20line/Germframeset.html
25. Tanetzafp, G. and Brown, N. An interaction between integrin and the talin FERM domain mediates integrin activation but not linkage to the cytoskeleton. Natural Cell Biology
2006, 8, 601-606.
independently
does not signal with integrins during DTC migration. The differences between the RNAi and empty vector controls for src-1, pxl-1, and talin indicate α-integrin signaling independence, implying pat-3 signaling, in the DTCs.
Note that no data was collected for wild type nematodes exposed to pxl-1 RNAi. Since nematodes exposed to nischarin RNAi have no significant difference from the empty vector controls, it is unlikely to signal with integrins in the DTCs. Error bars represent 95 % confidence.
paralysis.

Figure S2. Talin and src-1 signal independently of α-integrins in the gonad arms and the muscles, nischarin does not signal with integrins in the gonad arms and the muscles, and pxl-1 signals with pat-2 in the gonad arm for proper muscle function and does not signal with integrins in the gonad arm. The differences between the RNAi and empty vector controls for src-1and talin indicate α-integrin signaling independence, implying pat-3 signaling, in the muscles and the gonad arm for oocyte maturation
Since nematodes exposed to nischarin RNAi have no significant difference from the empty vector controls, it is unlikely to signal with integrins in the muscles or in the gonads for oocyte maturation Most of the strains exposed to pxl-1 RNAi are similar to the empty vector controls, but the pat-2 rescue strain has a decreased ability to hatch, but are still capable of producing eggs. This indicates a signaling partnership with pat-2 for proper muscle function, and no integrin signaling dependence for proper oocyte maturation.
Error bars represent 95 % confidence.

Biography
Danielle DaCrema is a May 2013 graduate from Christopher Newport University with a double major in Biology and Chemistry and a minor in Mathematics. She loves learning and is fascinated by the sciences particularly molecular and cellular biology. Her experience working in Dr. Meighan’s lab for the last two years at CNU has driven her to peruse a Ph.D. in the field. She was recently accepted to the Biomedical Sciences Ph.D. Program at the University of Virginia, and, although she will miss CNU, is very excited to begin there over the summer.
Switzerland: The Relationship between Neutrality and European Union Membership
Author: Kerry Campbell
Contact Information: kerry.campbell.10@cnu.edu or (703) 964-7979
Faculty Sponsor: Dr. Tina Kempin-Reuter
Information: Department of Government; tina.reuter@cnu.edu or (757) 594-7821
In this article the author examines the concept of Swiss neutrality and applies international relations theory and public opinion to better understand Switzerland’s relationship with the European Union (EU). The author evaluates three main international relations theories that can be applied, along with an analysis of public opinion, to determine why Switzerland has yet to join the European Union. The article begins by discussing the history of Swiss neutrality before noting the rise of centralized power in Europe and evaluating the compatibility of Switzerland’s neutrality with the framework of the European Union. The author then discusses public opinion within Switzerland regarding EU membership, analyzing public justifications, including neutrality, for remaining non-aligned while focusing on the increasing level of interaction between Switzerland and the European Union. Finally, the article concludes with suggestions for the future of Swiss neutrality and recommendations for a continuation of neutrality policy as an EU member-state.
Introduction
With the rise of international organizations and the success of the European Union (EU), the position of Switzerland in the European Community has gained importance. Located in the middle of Europe and surrounded by member states, Switzerland is the only continental Western European state that has not ascended to the European Union; Norway, however, along with other Baltic States, have also not joined the EU. While Switzerland has yet to join the EU, on many different occasions agreements have been reached between the two entities highlighting the nature of cooperation present between them The lack of membership in the European Union has been attributed to Swiss neutrality. According to Swiss historian and politician Walther Hofer, Swiss neutrality, or non-alignment, can be understood as perpetual and unlimited neutrality.1 Not only is Switzerland neutral in wartime, but they are also neutral concerning their foreign affairs in a time of peace as well. How has Switzerland’s neutrality impacted its relationship with the European community? More specifically, does this neutrality conflict with ascension to the European Union? If not, why has Switzerland been so reluctant to join the European Union as a member state? In addition, what is the future for Swiss neutrality and participation in the European Union?
Research regarding the Swiss refusal to join the European Union can be divided into two theoretical frameworks: realism and liberalism. Additionally, constructivism is used to practically define relations. Related to the constructivist notion, and the true answer as to why Switzerland has yet to join the European Union and has steadfastly maintained its position of neutrality, can be found in the political obstacle known as public opinion. Paul Ramseyer, former French ambassador to Switzerland, highlighted the relationship between neutrality and EU membership when he stated that “neutrality constitutes one of the main stumbling blocks towards
1 Walther Hofer, Neutrality as the Principle of Swiss Foreign Policy (Zurich: Guggenbühl & Huber, 1957), 5.
a possible membership of Switzerland in the European Community.”2 While constructivism seems to be the best theory when applied practically to the situation, as it takes into account public opinion, realism and liberalism have also be used to describe the issue, albeit with less success.
Swiss history, specifically as it relates to neutrality, is very important to understand why Swiss public opinion is irrevocably in favor of neutrality. After presenting a brief synopsis of Switzerland’s history of neutrality, a short analysis will be conducted on the rise of centralized power in Europe. This analysis of both Swiss neutrality and the history of the European Union will be helpful in the discussion of past and current Swiss-EU relations that will follow. There will then be a discussion of realism, liberalism, and constructivism, the international relations theories that are used when trying to understand the complicated relationship between Switzerland and the European Union, before discussing past and current relations between Switzerland and the European Union along with Swiss public opinion regarding neutrality and the possibility of EU membership In order to demonstrate the importance the Swiss people place in neutrality over EU membership a qualitative study conducted by the University of Zurich’s Center for Security Studies will be utilized. It is important to study international relations theory alongside of public opinion polls to more fully understand the complicated relationship between Switzerland and the European Union, and the role neutrality plays in this relationship. While realism and liberalism might be lacking in their understanding of the role neutrality plays in Switzerland, especially in regards to relations with Europe and the EU, the use of constructivism can be justified after looking at opinion polls. In addition, constructivism is best understood with public opinion as it interprets the feelings of the people on neutrality, understanding neutrality to
2 Paul Andre Ramseyer, “Switzerland and Europe – Between the EC, the CSCE and Neutrality” in Neutrals in Europe: Switzerland, eds. Bo Huldt and Atis Lejins (Stockholm: Swedish Institute of International Affairs, 1988), 17.
be one of the reasons why Switzerland has yet to join the European Union. Briefly, a discussion of additional factors, federalism and direct democracy, will follow as these are aspects of Swiss society that work with neutrality to allow the public to prevent Swiss entrance into the EU. Finally, recommendations for the future of Swiss-EU relations, specifically regarding the ability of Switzerland to maintain neutrality while becoming an EU member, will be proposed.
A Brief Synopsis of Swiss Neutrality3
Neutrality has been the central foreign policy principle of Switzerland for centuries While it has been practically observed since 1515, several major events, including the rise of Napoleon during the late 18th Century, caused Switzerland to pursue a more active foreign policy until the Congress of Vienna and subsequent Treaty of Paris from 1814-1815. On November 20th, 1815, the great powers of Europe acknowledged “the neutrality and inviolability of Switzerland and of its independence from all foreign influences as being in the true political interest of the whole of Europe.”4 In such a diverse nation, a policy of neutrality has served to unite the people. Without a foreign policy of neutrality, some believe, there would have been internal struggle throughout the centuries that would have destroyed the Swiss Confederation.5 This would have led to a decisive division of the cantons, and the existence of Switzerland as we know it today is questionable, especially when looking at the aggression which would come to plague Europe in the next century. In 1907, just before the onset of a war that would decimate the territory surrounding Switzerland, Switzerland signed the Hague Conventions on the Rights and Duties of Neutral States. This confirmed Swiss neutrality and allowed Switzerland to remain
3 For a helpful reference point see Stefan Aeschimann, et al. Swiss Neutrality, 4th revised ed. (Bern: Communication DDPS, 2004), 21.
4 Leo Schelbert, Historical Dictionary of Switzerland (Lanham, Maryland: The Scarecrow Press, 2007), 244.
5 Aeschimann et al., Swiss Neutrality, 3
neutral throughout World War I. In the aftermath, however, Switzerland employs a policy known as differential neutrality; under differential neutrality Switzerland reaffirmed its policy of neutrality while joining the League of Nations and accepting the economic sanctions imposed by the League, as differential neutrality involves a country remaining neutral while accepting participation in international organizations. When these economic sanctions were unsuccessful in preventing Italian aggression, Switzerland abandoned differential neutrality for integral neutrality, or a return to traditional neutrality6: while they would remain a member of the League of Nations, Switzerland no longer supported the use of economic sanctions. Integral neutrality is employed throughout World War II. After the collapse of the League of Nations, Switzerland restrains from joining the United Nations, a course of action they would maintain until 2002.
During the Cold War, however, Switzerland maintained active neutrality and participated in many peace-keeping missions, including participation in the North Atlantic Treaty Organization’s (NATO) Partnership for Peace.7 With ascension to the United Nations, Switzerland once again adopted a policy of differential neutrality.
As can be seen, there are many important developments in the history of Swiss neutrality. Most important, however, to the continued cooperation with and possible membership in the European Union, is Switzerland’s membership in the League of Nations. While the League of Nations represented the first attempts at collective security, it also brought to light tensions between neutrality and universal cooperation.8 These tensions can still be seen today in the relationship between the European Union and Switzerland. When Switzerland ascended to the League of Nations, they practiced differential neutrality, a neutrality policy that separated
6 Traditional neutrality and integral neutrality are used interchangeably, and refer to neutrality as un derstood in international law
7 Aeschimann et al., Swiss Neutrality, 4-9.
8 Efraim Karsh, “International Co-Operation and Neutrality,” Journal of Peace Research 25, no. 1 (1988): 58.
economic actions from military neutrality. Furthermore, differential neutrality was a policy that allowed Switzerland to participate in collective security, viewing collective security as different from war.9 Once Switzerland returned to integral neutrality, however, they retained their membership in the League of Nations. After the failure of the League of Nations, Switzerland perceived collective security as a failed means to organize the international system and saw participation in an organization providing for collective security as having too many risks. Furthermore, the failure of the League also affected the Swiss decision to discard differential neutrality.10 With the rise of universal cooperation in Europe, the Swiss looked on with disdain due in part to the failure of the League of Nations. As Switzerland once again practices differential neutrality, there is the opportunity for Swiss ascension to the European Union, conducive to their policy of neutrality.
The Rise of Centralized Power in Europe
After World War II, it became evident that cooperation between the different countries of Europe was necessary. This idea of universal cooperation soon turned into the desire for collective security, or preserving interests and ensuring that another war would not ensue, based first in economics, as seen first with the establishment of the European Coal and Steel Community (ECSC) in 1951 and later with the European Economic Community (EEC) in 1957.11 The building blocks for a European Union were now being set as the ECSC and EEC generated a European common market where goods could be shared more easily, helping to prevent the continent from entering into a depression, as seen after World War I. In 1973 this
10 Ibid , 63.
11 European Union, The History of the European Union
common market expanded further with the ascension of Denmark, Ireland and the United Kingdom as well as the entry into force of the European Free Trade Association (EFTA). The EFTA established trade relationships with non-EEC member states including Austria, Switzerland, Portugal, and Sweden.12 Switzerland was able to benefit from the EFTA while remaining neutral while other countries used it as a path to membership.
After Greece, Spain, and Portugal joined the EEC in the 1980s, it became necessary to combat certain problems that arose with the establishment of a European common market. Relations between member-states had stalled. The Single European Act, which entered into force in 1987, was a major ratification to the Treaty of Rome that had established the EEC. In the Single European Act, countries were propelled closer together as Europe’s Common Market turned into a Single Market. This Single Market is understood as “an area without internal frontiers in which the free movement of goods, persons, services and capital is ensured in accordance with the provisions of this treaty.”13 With the entry into force of the Single European Act, the evolution of universal cooperation from the purely economic realm to becoming a political institution was now possible. From 1992-1993 this became reality with the signature and entry into force of the Maastricht Treaty.
The Maastricht Treaty signaled the official establishment of the European Union by combining the EEC and other policies into one European Community. This community expanded upon the economic union of the past with a monetary union, eventually to be known as the euro, in addition to institutionalizing cooperation foreign policy, justice, and defense.14 While not all countries accepted policies as outlined in the Maastricht Treaty, the United Kingdom and Denmark refused to accept the euro when it was adopted; the Maastricht Treaty demonstrated the
12 Ibid.
13 European Union, “The Single European Act,” Summaries of EU Legislation
14 Ibid.
continued concern for universal cooperation in Europe that would provide all citizens with a collective security that would be beneficial into the future. Switzerland, however, did not view this to be the case. After the entry into force of the Maastricht Treaty, the Swiss still refused to join the European Union, focusing more than ever on their policy of neutrality. An assessment of international relations theory can provide an indication of why Switzerland refused to join the European Union, and how the people are using neutrality as a reason to avoid Europe’s collective security.
International Relations Theory
In international relations, there are two main branches of thought that govern why states undertake certain actions. While realism believes that there is a constant state of anarchy and states do what is in their own interest, liberalism believes that states undertake actions to promote global cooperation, not purely from self-interest. Both of these theories have been applied when discussing Switzerland’s refusal to join the European Union. Constructivism, stemming from liberalism, is often used practically, as constructivists understand the role public opinion plays in Switzerland’s decision. The similarities between the three theories, especially as applied to Swiss neutrality and their impact on membership in the European Union can be understood after each theory is examined separately.
Realism is a dominant perspective in international relations theory and has often been used when describing relations between Switzerland and the European Union. This is problematic, however, when trying to understand the reasons behind the importance of Swiss neutrality. Realists, according to Kate Morris and Timothy White, see neutrality as a rational
calculation, undertaken primarily by small states, to promote national interests, like survival.15
The problem with this theory, however, when applied to neutrality is that neutrality does not often conform to traditional realist principles; especially in the case of Switzerland, Morris and White see neutrality as not about what happens during wartime but as an outline of foreign relations that has shaped Swiss politics for centuries.16 Furthermore, realism is skeptical about the ability of international institutions to impact the relations between states. Therefore, according to Rene Schwok, it would be pointless for a state to join an international organization.17 While the realist perspective has often been used in international relations, and has also been applied to the case of Switzerland, it does not provide the reasoning behind why Switzerland continues refusal of European Membership; Swiss refusal to join the European Union is based mostly on their domestic policy. It does, however, shed light on why the Swiss first decided on a principle of neutrality.
While realists say that anarchy is constantly present, liberalists take a different perspective; liberalism is the opposite of realism. Instead of attributing a nation’s desire for neutrality to external factors, protecting themselves from outside threats, the liberalist approach attributes a nation seeking neutrality based on domestic affairs. Morris and White therefore believe that neutral states would be willing to abandon their neutrality if there is the prospect for economic benefit.18 This economic benefit could therefore be obtained if Switzerland was to join the European Union. However, a flaw in this interpretation of neutrality is that Switzerland, in maintaining neutrality, is trying to avoid interactions between other states. While the EU is
15 Kate Morris and Timothy J. White, “Neutrality and the European Union: The Case of Switzerland,” Journal of Law and Conflict Resolution 3, no. 7 (2011): 2.
16 Ibid.
17 Rene Schwok, Switzerland–European Union: An Impossible Membership (Brussels: P.I.E. Peter Lang, 2009), 101.
18 Morris and White, “Neutrality and the European Union,” 2.
currently a predominantly economic institution, in the future it could turn into a security obligation, something that would conflict with Swiss neutrality. Therefore, liberalism actually provides reasoning for why Switzerland would join the European Union, not why they are using neutrality as the reason they are avoiding membership.
Constructivism is the most helpful of the international relations theories when trying to understand Switzerland’s neutrality policy in relation to the European Union. According to Schwok, “constructivism has rehabilitated concepts such as identity, historic memory, the role of collective ideas, and the importance of ideology.”19 In other words, as indicated by White and Morris, constructivism analyzes neutrality not as a legal notion, but as a political and/or ideological perspective.20 Neutrality is a national symbol to the Swiss and this can be understood under a constructivist lens. Since the social and historical contexts for a nation maintaining neutrality are different for each country, each individual nation has their own way of reconciling neutrality with increased international participation. For Switzerland, this has involved different bilateral agreements with the European Union21 while still not joining the organization. Related to the constructivist approach is the perception that neutrality is a classic political obstacle which is preventing the Swiss from joining the European Union.
With an understanding of these three theoretical approaches, each can be applied directly to the current state of Switzerland’s relationship with the European Union. Realism as conventionally understood does not explain the relationship between Switzerland and the European Union that currently exists. On the other hand, traditional liberalism does not explain Switzerland’s unwillingness to forgo neutrality in favor of the economic benefits granted by full EU membership. Constructivism, by treating neutrality as a political/ideological perspective,
19 Schwok, Switzerland–European Union, 97.
20 Morris and White, “Neutrality and the European Union,” 2.
21 Federal Department of Foreign Affairs, “Switzerland and the European Union,” 2012.
views neutrality in the same manner as the Swiss people. As constructivist theory can vary depending on the social and historical contexts impacting a particular situation, it is the most efficient and effective theory when attempting to understand Switzerland’s affinity for neutrality. For this reason, the remainder of the paper will focus on neutrality as Switzerland’s political stumbling block to EU membership as understood by constructivism.
Neutrality: A Political Stumbling Block
Paul Ramseyer, former ambassador to Switzerland, said it best in that neutrality “constitutes one of the main stumbling blocks towards a possible membership of Switzerland in the European Community.”22 The constructivist approach highlights how neutrality can be understood as stemming from a nation’s history. This is the case in Switzerland. Many scholars go one step further in their interpretation of Swiss neutrality, believing that neutrality is one of a few political obstacles that are preventing Switzerland from joining the European Union. Clive Church describes neutrality as “a basic component of Swiss identity… [That] formed the basis of the strong and popular anti-EU movement.”23 Switzerland is not able to move forward in their relations with the European Union because the people do not want change. Neutrality, as understood by political researchers Laurent Goetschel, Magdalena Bernath, and Daniel Schwarz, is inherently linked with Swiss identity.24 The Swiss have come to expect neutrality, seeing it as the way they are perceived by the international community. Expanding on this notion and the point made by Church, Amy C. Di Stasio characterizes Swiss neutrality as flexible, since it has
22 Ramseyer, “Switzerland and Europe,” 17.
23 Clive H. Church, Introduction to Switzerland and the European Union: A Close, Contradictory, and Misunderstood Relationship (London: Routledge, 2007), 6.
24 Laurent Goetschel, Magdalena Bernath, and Daniel Schwarz, Swiss Foreign Policy: Foundations and Possibilities (London: Routledge, 2005), 16.
adapted based on society’s changing circumstances.25 While neutrality has served its purpose for centuries she suggests that neutrality might have reached its limits. The usefulness of neutrality has rooted itself in the Swiss conscience to such an extent, Di Stasio suggests, that it is difficult for the Swiss to abandon, especially as it has been such a successful policy.26 The Swiss are unable to separate the political concept of neutrality, something which they have known for so long, from the possibilities that being a neutral member of the European Union could provide.
Daniele Ganser and Georg Kreis work to show that it is possible for Switzerland, as a neutral country, to remain neutral as a member-state of the European Union. Historically, the accepted conceptualization of neutrality was influenced by the time period; whether in World War II, the Cold War, or a time of peace, Swiss neutrality has been adapted to fit the situation. This practice, according to Ganser and Kreis, could be employed once more in order to ensure that the Swiss remain neutral as members of the EU. However, as Ganser and Kreis point out, there is a significant attachment to the concept of neutrality currently understood linked to a desire of Swiss citizens to remain a sovereign and independent entity.27 In other words, the Swiss are afraid if they become a member of the EU neutrality will lose its significance.28 Neutrality, even though an element of foreign policy, plays a very important role in domestic politics. In fact, Church presents opposition to neutrality as an important opposition force.29 This political opposition force serves to influence both domestic and foreign relations. This reluctance to
25 Amy C. Di Stasio, “Switzerland: Has Neutrality Reached its Limits?” Perspectives on Business and Economics 18 (2000): 2.
26 Ibid., 6.
27 Daniele Ganser and Georg Kreis, “Swiss Neutrality: Incompatible with EU Membership?” in Switzerland and the European Union: A Close, Contradictory, and Misunderstood Relationship , ed. Clive H. Church (London: Routledge, 2007), 52-73.
28 Ibid , 72.
29 Clive H. Church, “The Contexts of Swiss Opposition to Europe” (Working Paper, Sussex European Institute, 2003), 4.
become a member-state has impacted Swiss relations with Europe 30 However, that relationship is still present as several bilateral agreements between the two entities have been passed
Switzerland and the European Union
While Switzerland has yet to become a member state of the European Union and, as popular opinion (see below) seems to suggest, will not become a member nation of the EU any time in the near future, there is still a strong relationship between the two establishments. Even before the European Union was the EU, Switzerland maintained a strong relationship with the precursor organizations hoping to build cooperation within the region. On January 1st, 1973
Switzerland entered into a free trade agreement with the European community of the period, the European Free Trade Association. This agreement allowed for the abolishment of customs duties on industrial goods and certain agricultural products.31 With the evolution of relations among European nations and the success of the EFTA, the European Free Trade Association soon flourished into the European Economic Area (EEA) The EEA was established in 1992 with the Porto Agreement but did not enter into force until 1994, after the establishment of the European Union. Although Switzerland originally signed the treaty, a national referendum on December 6, 1992 indicated a refusal of the Swiss population to participate as a member of the EEA.32 With the refusal to enter the EEA, Swiss policymakers set to work on negotiations for an agreement between the European community and Switzerland.
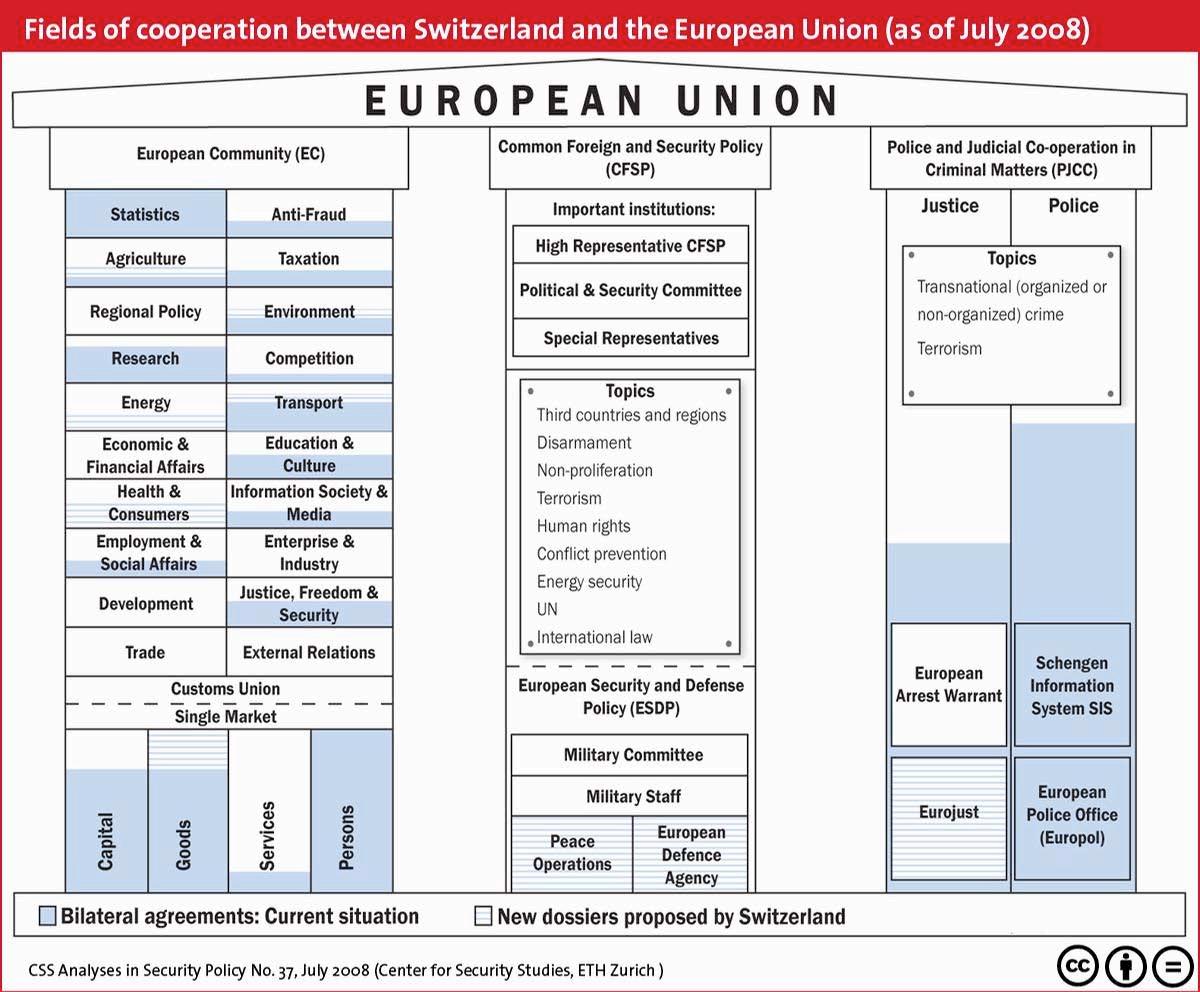
What came to be known as Bilateral Agreements I between Switzerland and the European Union passed referendum and entered into force in 2000. Bilateral Agreements I outlined provisions for a relationship between Switzerland and the EU relating to the following
30 Ibid , 32.
31 Schwok, Switzerland–European Union, 24-25.
32 Ibid , 29.
areas: air transport, public procurement markets, participation in EU research programs, agriculture, elimination of technical barriers to trade, overland transport, and the free establishment of persons.33 Each point, after ratified by the Swiss, soon became a source of open cooperation between Switzerland and the European Union, even as Switzerland continued to refuse ascension based on their principle of neutrality; while the people refused ascension, it is clear they accepted cooperation. In 2004 another series of bilateral agreements, what came to be known as Bilateral Agreements II, was signed by Switzerland and the EU; however, the nine areas of focus presented in the agreement were ratified through referendum at different times. Bilateral Agreements II discuses provisions for cooperation on the following: taxation of savings, the fight against fraud, Schengen/Dublin, processed agricultural products, statistics, pensions, the environment, MEDIA, and education, occupational training, and youth.34 The European Union is clearly working to pursue a bilateral relationship with Switzerland. Schwok has even gone as far as to say that “the alignment of Switzerland’s legislation to that of the EU will continue in the coming years, as there is a mutual interest in concluding agreements.”35
It is clear that Switzerland and the European Union are slowly coming closer together on many important policy issues. All of these issues relate in some way to political, economic, or social concerns held by each party. Therefore, with a return to differential neutrality, these issues in no way compromise Switzerland’s principle of neutrality. A continuation of differential neutrality will allowed for a continued relationship between Switzerland and the EU with possible future membership. However, the public must be willing to ascend. Even with the bilateral agreements that were ratified there is still an innate resistance towards EU membership
33 Ibid , 37-38.
34 Ibid , 53-54.
35 Ibid , 78.
along with a deep skepticism of the euro within Switzerland.36 As was seen in the referendum of the EEA statue, the public are unwilling to enter into an agreement that compromises, or has the possibility to compromise, their neutrality. In the European Report of 2006, the Federal Council decided that instead of treating European membership as a “strategic goal,” it would now be a “longer-term option” for Switzerland. The future of Swiss foreign policy, specifically how neutrality will be treated in relation to increasing alignment with the European Union, is now in the hands of the people
Current Opinions
Just as they were unwilling to accept ascension to the EEA, the Swiss people are also unwilling to enter into the European Union. Even though Switzerland is “a quintessentially European country,”37 there is little hope for ascension to the EU anytime soon. This critical view towards the European Union is in stark contrast to the overwhelmingly positive standpoint held by the people on neutrality.
A long term public opinion poll conducted by the Swiss Military College at the Federal Institute of Technology from 1993 to 2004 on neutrality never dipped below 78% and peaked at almost 90%.38 It is clear that the Swiss people would like to retain their neutrality. Furthermore, the 2012 Sicherheit study conducted by the Center for Security Studies at the Swiss Federal Institute of Technology Zurich indicates that 95% of the Swiss population supports the maintenance of Switzerland’s neutrality policy; this is the highest since the Security study began
in 1989.39 Regarding the Swiss perception of neutrality, the study shows that 83% of the population views neutrality as “inseparable from the Swiss concept of the state.”40 In other words, the Swiss view neutrality as an integral value of their national identity. This confirms the belief, important to the theory of constructivism, that neutrality is more than a legal notion and is actually a part of Swiss ideology.41 This ideology, while instilling in 52% of the population the conviction that armed Swiss neutrality has actually contributed to Europe’s stabilization, has not affected Swiss feelings toward the European Union; actually, only 23% of the population believe that membership in a European defense alliance, the European Union, would provide them with more stability than their current policy of neutrality; this is down 5% from the 2011 figures.42 While the Swiss might have positive feelings towards neutrality, this is not the case when discussing the European Union.
Skeptical attitudes towards the European Union intensified in 201143 and this trend can be seen most clearly in Swiss public opinion regarding EU ascension. A recent survey released in November 2012 by the SonntagsZeitung demonstrates that only 11.5% of the Swiss support membership in the European Union this is the lowest public opinion rating in the history of the EU.44 Furthermore, the Sicherheit study, published earlier in 2012, indicated that only 17% of the population wishes to ascend to the European Union.45 Undoubtedly, as long as a referendum needs to be passed for Switzerland to join the European Union, Switzerland will not become a member with such low public support. On the other hand, though, the Swiss people understand the necessity of maintaining a relationship with the EU. When asked how Switzerland should
39 Tibor Szvircsev, et al., Sicherheit 2012: Aussen-, Sicherheits-, und Verteidigungspolitische Meinungsbidung im Trend (Zurich: Center for Security Studies und Militӓrakademie and der ETH, 2012), 65.
40 Ibid.
41 Morris and White, “Neutrality and the European Union,” 2.
42 Szvircsev et al., Sicherheit 2012, 66.
43 Ibid , 63.
44 “Immer Mehr Schweizer Sprechen Sich Gegen EU-Beitritt Aus,” SonntagsZeitung, November 25, 2012
45 Szvircsev et al., Sicherheit 2012, 64
interact with the European Union, 82% said that they believe “Switzerland should not join the EU but strengthen economical cooperation.”46 Although the Swiss people understand that cooperation with the EU is important, they are not willing to take all the steps necessary to become a full member-state under the EU charter. These feelings on neutrality, in addition to other factors important within Switzerland, magnify the tensions between Switzerland and the European Union.
Additional Factors
As has been demonstrated, neutrality is an important point historically within Switzerland, one which the people do not want to compromise on, and which has impacted the relationship between Switzerland and the European Union. In addition to neutrality, federalism and direct democracy also play a role in the Swiss refusal to accept entrance into the European Union. By providing the people with more of a voice, federalism and direct democracy prevent the government from making a decision that the people will not accept Federalism allows local communities and individual cantons to participate in government while they are able to coexist peacefully, even with the differences they may have.47 However, Swiss federalism has been affected by the rise of centralized power in Europe. The cantons have limited access to government as Swiss dealings are increasingly held at the international level and the people are able to have a say only when items are brought to them by referendum.48 In addition to federalism, therefore, direct democracy is an important way that the Swiss
46 Ibid.
47 Paolo Dardanelli, “Federalism: Institutional Adaptation and Symbolic Constraints” in Switzerland and the European Union: A Close, Contradictory, and Misunderstood Relationship , ed. Clive H. Church (London: Routledge, 2007), 19
48 Ibid , 26-27.
demonstrate their public opinion, especially in regard to the Europeanization process and EU membership.
Direct democracy, especially within Switzerland, provides the people with a voice. Most important is the mandatory referendum, which requires all constitutional amendments to be approved by a majority of the cantons as well as a majority of the people.49 The optional referendum, on the other hand, allows the people or the cantons to request a vote on legislation as long as they earn the required number of signatures within 100 days of the laws’ publication.50
Referendum has become important as the relationship between Europe and Switzerland has gained increasing importance. It has slowed down the Europeanization process in Switzerland while also preventing ascension to the EEA51 and now the European Union.52 Direct democracy allows the people, who have a desire to ensure that the Swiss policy of neutrality remains important, the ability to decide whether or not Switzerland joins the European Union. While the people are unwilling to compromise on neutrality, and public opinion shows that they are unwilling to join the EU, the impact direct democracy has had on EU integration has been mostly indirect.53 The issue of EU ascension has never been directly submitted to Swiss voters; this can be attributed to the lack of support for the EU the government sees in the electorate.54 Instead, the Swiss people have voted on whether or not to open negotiations with the European Union and the government has worked bilaterally with the EU for years without membership.
The Future of Swiss Neutrality and EU Relations
Switzerland can remain neutral and join the European Union, but it is unlikely to occur due in part to the current public opinion on the matter. Even the Swiss government understands that membership in the EU is compatible with neutrality so long as “the EU has no binding mutual military assistance obligation for all members.”55 While this may be the case, the Swiss people stand behind their commitment to neutrality, despite the fact that Swiss ascension to the European Union is included in the official long-term plan of the Swiss government.56 However, few citizens are aware of this.57 Today, neutrality is an integral part of Swiss identity. Divergent from the facts, many Swiss citizens consider neutrality and independence to be “indivisibly intertwined” and equally important pillars within the Swiss Confederation. 58 For Switzerland to become a member of the European Union, they must come to terms with an understanding of neutrality within a world focused on collective security. Unless the EU takes on a military character, similar to that of NATO, Swiss neutrality will remain compatible with membership; this can be seen in the membership of neutral countries such as Austria and Finland, who actively participate in EU activities while remaining exempt from the EU’s defense policy.59 However, even without membership, the Swiss can continue to preserve and expand upon the bilateral relations Switzerland has with the European Union while maintaining a close relationship with EU member states. In fact, these bilateral agreements have “significantly reduced the pressure on Switzerland to join the EU on economic grounds.”60 This is because Switzerland can participate in the European Single Market, selectively, in order to maintain independence and neutrality. Switzerland already works with the European Union in many
55 Aeschimann et al., Swiss Neutrality, 13.
56 Elia Pusterla and Francesca Piccin, “The Loss of Sovereignty Control and the Illusion of Building Walls,” in Journal of Borderland Studies 27, no. 2 (2012): 130.
57 Goetschel, Bernath, and Schwarz, Swiss Foreign Policy, 16.
58 Ibid, 16.
59 Schwok, Switzerland–European Union, 98.
60 Daniel Möckli, “Switzerland and the EU: The Prospects of Bilateralism,” trans. Christopher Findlay, CSS Analyses in Security Policy 3, no. 37 (2008): 2.
sectors, including agriculture, medicine, and labor, through their Mutual Recognition Agreement.61 The problem with this strategy, though, is that Swiss policy becomes more vulnerable both domestically and internationally.62 Several domestic problems arise with the continuity of bilateral associations for a prolonged period of time without membership, especially in regard to direct democracy.63 As each piece of a bilateral agreement is up for referenda, a majority of Swiss citizens must accept the provision. Internationally, bilateral agreements place the future of Swiss-EU relations in the hands of the European Union since bilateral relations are valid as long as the European Union is willing to negotiate. As new, nonWestern European, countries continue to join the EU skepticism grows as they were required to make concessions to the European Community that they see the Swiss as refusing to make.
There is a future for Swiss neutrality within the European Union framework, if the people of Switzerland continue on their policy of differential neutrality. In the near future, a continuance of bilateral relations will allow for further alignment between Swiss and EU policies. This alignment can provide the gateway for Swiss ascension to the European Union. By doing so, Switzerland can maintain their policy of neutrality so long as the European Union remains a political institution for the purpose of achieving collective security within Europe. If the EU takes on a military purpose, then EU policies will no longer be in accordance with Swiss neutrality. As this is not currently the case, it would be prudent for the Swiss people to accept continued relations with Europe, similar to the participation of other neutral nations such as Ireland and Finland, before the European Community is no longer willing to accept Switzerland into their organizations. Pusterla and Piccin observe that “Switzerland is surrounded by the
61 European Union, “Mutual Recognition between the European Union and Switzerland,” Single Market for Goods
62 Möckli, “Switzerland and the EU,” 1
63 Ibid, 3.
Europeanization process and can hardly resist it ”64 However, Möckli notes that the European Community is becoming increasingly tired of Switzerland receiving some of the benefits of Europeanization without contributing to Europe as EU member states must.65 The Swiss government has made it a policy to pursue EU membership in the long-term, ensuring that their neutrality is maintained; only time will tell if the Swiss people are willing to accept this.
Conclusion
In recent years, neutrality has impacted Switzerland’s relationship with the European Union, even though a policy of neutrality is compatible with EU membership. Public opinion on neutrality has been a major political obstacle for European integration. Switzerland has been neutral for centuries, and Swiss neutrality has served as a basis for the evolution of neutrality policy within the international realm. As neutrality has been a part of Swiss identity for so long, it is very difficult for the people to relinquish. This difficulty has led to tensions between Switzerland and other European countries after the rise and expansion of centralized power, culminating in the European Union. Switzerland has yet to ascend to the European Union, citing their policy of neutrality as incompatible with EU policies. However, as has been demonstrated, Swiss neutrality can, in fact, be compatible with current EU policy. After all, Sweden, Finland, Austria, and Ireland are all neutral European countries that have joined the EU.
The history of Swiss neutrality helps to provide an understanding for the importance which the Swiss people place in their neutrality policy; the Swiss see neutrality as an essential aspect of their identity. Along with federalism and direct democracy, neutrality is one of the four core principles of Swiss politics. There are three main international relations theories which have
64 Pusterla and Piccin, “The Loss of Sovereignty Control and the Illusion of Building Walls,” 131
65 Möckli, “Switzerland and the EU,” 2.
been used when analyzing the Swiss desire to maintain a position of neutrality in relation to whether or not Switzerland will become a member state of the European Union Each of these three realism, liberalism, and constructivism help to shed light on why neutrality is important to the Swiss. However, they do not help to explain why the Swiss insist on mainlining their principle of neutrality, and how that neutrality can fit into the potential status of Switzerland as an EU member-state. An aspect of constructivism, treating neutrality as a political obstacle, answers this question. Neutrality has been an aspect of Swiss foreign policy for so long that it has become a part of their national identity. Constructivism provides an explanation for why the Swiss people are unwilling to join the European Union. The Swiss people are reluctant to relinquish their neutrality because they fear that it will relinquish their sovereignty and independence.
For Switzerland to become a member of the European Union in the future, instead of just entering into Bilateral Agreements, the Swiss people must understand the potential for neutrality as a member state of the EU. For this to happen, however, the concept of neutrality might have to be adapted, expanding upon Switzerland’s current differential neutrality As has been demonstrated, while the Swiss people are slowly accepting relations with a collective Europe, they are not yet ready to “abandon” their policy of neutrality, however compatible it may be with the EU, in favor of European Union membership.
References
Aeschimann, Stefan, et al Swiss Neutrality. 4th revised ed. Bern: Communication DDPS, 2004.
Church, Clive H. “The Contexts of Swiss Opposition to Europe.” Working Paper, Sussex European Institute, 2003.
Church, Clive H. Introduction to Switzerland and the European Union: A Close, Contradictory, and Misunderstood Relationship. London: Routledge, 2007.
Dardanelli, Paolo. “Federalism: Institutional Adaptation and Symbolic Constraints.” In Switzerland and the European Union: A Close, Contradictory, and Misunderstood Relationship. Ed. Clive H. Church, 17-36. London: Routledge, 2007.
Di Stasio, Amy C. “Switzerland: Has Neutrality Reached its Limits?” Perspectives on Business and Economics 18 (2000): 1-15.
http://www.lehigh.edu/~incntr/publications/perspectives/v18/distasio.pdf (accessed September 30, 2012).
European Union. The History of the European Union http://europa.eu/about-eu/euhistory/index_en.htm (accessed October 25, 2012).
European Union. “The Single European Act.” Summaries of EU Legislation.
http://europa.eu/legislation_summaries/institutional_affairs/treaties/treaties_singleact_en. htm (accessed October 25, 2012).
European Union. “Mutual Recognition Agreement between the European Union and Switzerland.” Single Market for Goods http://ec.europa.eu/enterprise/policies/singlemarket-goods/international-aspects/mutual-recognitionagreement/switzerland/index_en.htm (accessed May 23, 2013).
Federal Department of Foreign Affairs. “Switzerland and the European Union.” Representations
http://www.eda.admin.ch/eda/en/home/reps/eur/vgbr/ukemlo/ecofin/cheu.html (accessed October 5, 2012).
Ganser, Daniele and Georg Kreis. “Swiss Neutrality: Incompatible with EU Membership?” In Switzerland and the European Union: A Close, Contradictory, and Misunderstood
Relationship. Ed. Clive H. Church, 52-78. London: Routledge, 2007.
Goetschel, Laurent, Magdalena Bernath, and Daniel Schwarz. Swiss Foreign Policy: Foundations and Possibilities. London: Routledge, 2005.
Hofer, Walther. Neutrality as the Principle of Swiss Foreign Policy. Zurich: Guggenbühl & Huber, 1957.
“Immer Mehr Schweizer Sprechen Sich Gegen EU-Beitritt Aus.” SonntagsZeitung, November 25, 2012. http://www.sonntagszeitung.ch/suche/artikel-detailseite-sda/?newsid=236259 (accessed November 25, 2012).
Karsh, Efraim. “International Co-Operation and Neutrality.” Journal of Peace Research 25, no. 3 (1988): 57-67.
Loeffel, Urs. “Swiss Neutrality and Collective Security: The League of Nations and the United Nations.” PhD diss., Naval Postgraduate School, 2010.
Möckli, Daniel “Switzerland and the EU: The Prospects of Bilateralism.” Trans. Christopher Findlay
CSS Analyses in Security Policy 3, no. 37 (2008): 2
Morris, Kate and Timothy J. White. “Neutrality and the European Union: The Case of Switzerland,” Journal of Law and Conflict Resolution 3, no. 7 (2011): 1-8.
http://www.academicjournals.org/jlcr/PDF/pdf%202011/Sep/Morris%20and%20White.p
df (accessed October 1, 2012).
Pusterla, Elia and Francesca Piccin. The Loss of Sovereignty Control and the Illusion of Building Walls.” In Journal of Borderland Studies 27, no. 2 (2012): 121-138.
Ramseyer, Paul Andre. “Switzerland and Europe – Between the EC, CSCE and Neutrality.” In Neutrals in Europe: Switzerland. Eds. Bo Huldt and Atis Lejins, 17-26. Stockholm: Swedish Institute of International Affairs, 1988.
Schelbert, Leo. Historical Dictionary of Switzerland. Lanham, Maryland: The Scarecrow Press, 2007.
Schwok, Rene. Switzerland-European Union: An Impossible Membership Brussels: P.I.E. Peter Lang, 2009.
Szvircsev, Tibor, et al. Sicherheit 2012: Aussen-, Sicherheits-, und Verteidigungspolitische Meinungsbidung im Trend. Zurich: Center for Security Studies und Militӓrakademie and der ETH, 2012.
Biography
Kerry Campbell, of Haymarket, Virginia, is a senior at Christopher Newport University pursuing bachelor’s degrees in Political Science and History in addition to minors in Leadership and Classical Civilizations. Kerry is the President of Alpha Chi (National Honor Society), Model International, the History Club, and Outdoor Adventure and Recreation, in addition to being treasurer of the Women’s Lacrosse Club Team. Furthermore, she is a part of Phi Alpha Theta (History Honor Society), Pi Sigma Alpha (Political Science Honor Society), Omicron Delta Kappa (Leadership Honor Society), and Alpha Mu Gamma (Foreign Language Honor Society). On campus, she has served as the student assistant for the Reiff Center and has participated in several study abroad programs. She would like to thank her parents for supporting her in everything she has undertaken. Additionally, she would like to thank Dr. Tina Kempin-Reuter, her Senior Seminar professor, for serving as a mentor throughout the writing process.
1. Title: A STUDY OF COMPETITION BETWEEN NATIVE EASTERN OYSTER
(CRASSOSTREA VIRGINICA) AND NONNATIVE SUMINOE OYSTER
(CRASSOSTREA ARIAKENSIS) AND IMPLICATIONS FOR THE CHESAPEAKE
BAY
2. Author’s name: Katherine Frances Holt
3. Author’s email address and phone number: holtkf@cox.net; 757 879-8947
4. Faculty sponsor’s name: Dr. Gary Whiting, Professor
5. Faculty sponsor’s department, email address and phone number: Department of Organismal and Environmental Biology; gwhiting@cnu.edu; 757 594-7613
Abstract
For the Chesapeake Bay, the drastic decline in the population of native eastern oyster, Crassostrea virginica, impairs water quality. Introduction of nonnative Suminoe oyster, C. ariakensis, has been proposed to improve water quality and meet consumer demands for oysters. This study explored competition between diploid C. virginica and triploid C. ariakensis when the two species were grown in competitive mixtures in the Piankatank and Rappahannock Rivers. The study also determined water clearance rates and food assimilation efficiencies of the two species. At a stocking density of 1076 juvenile oysters/m2, C. ariakensis grew more robustly than C. virginica and suppressed growth of C. virginica in competitive mixtures. C virginica had little effect on, or enhanced growth of, C. ariakensis. Mean water clearance rates were not significantly different between the two species C. ariakensis showed higher food assimilation efficiencies. Further research is needed to determine threshold stocking density for competitive effects between the two species in rivers of the Chesapeake Bay and to assess likely ecological impacts of competition.
Introduction
Water quality in the Chesapeake Bay is severely impaired (CBF 2000). One reason is the demise of the native oyster, Crassostrea virginica (Hargis 1999). This keystone species ingests polluted water, digests microscopic floating plants, and clarifies the water column through encapsulation of ingested, non-food particles as pseudofeces deposited on the bottom of the
estuary (Shumway 1996) Reasons for the demise include: (1) Over-harvesting (Gross and Smyth 1946; Rothschild et al. 1994), (2) Diseases, such as MSX, caused by Haplosporidium nelsoni and Dermo, caused by Perkinsus marinus (Fisher 1996), (3) Loss of habitat (Hargis and Haven 1988a, b), and (4) Reduction in water quality because of anthropological activity (Lenihan and Peterson 1998). In addition to deteriorated water quality and loss of habitat for other species, the demise of the native eastern oyster has decimated a once lucrative oyster industry, adversely affecting the incomes of watermen, the enterprises of Chesapeake Bay communities, the availability of a desired food for oyster consumers, and the economies of several states (Harwell et al. 2010).
To restore the native oyster, the Virginia Marine Resources Commission (VMRC) built artificial reefs using various substrates, including surplus U.S. Navy ships (Burke 2010).
Beginning in the late 1990s, VMRC sponsored various initiatives to encourage citizens to grow seed oysters for oyster reefs (Brumbaugh et al. 2000) Burke (2010) reported results of an extensive study focused on alternative substrate options for reefs in large-scale restoration initiatives in the Chesapeake Bay Allen (2003) researched improved broodstock and determined efficacy of strains such as CROSBreed and DEBY. Carnegie and Burreson (2009) reported that natural disease resistance may be developing among groups of C. virginica in the Bay. Other researchers reported that C. virginica has improved resistance to MSX and Dermo if grown in protected sanctuaries (Encomio et al. 2005).
Even with promising signs that the demise of C. virginica may have been arrested, some researchers concluded in early 2000 that restorative efforts alone would not meet the environmental goal of increasing the native oyster population in the Chesapeake Bay tenfold by 2010 (Brumbaugh et al. 2000). Given this prediction, attention focused on nonnative oysters,
specifically the Suminoe oyster, Crassostrea ariakensis, as a means of augmenting the native oyster, improving water quality in the Bay, and satisfying market demand for edible oysters.
Calvo et al. (2001) performed an early study to compare survival, growth, and disease resistance of C. virginica and C. ariakensis in three salinity regimes (low, medium, and high) of the Bay. They found that, in all salinities, C. ariakensis had superior survivability and significantly greater growth in medium- and high-salinity estuaries as compared to C. virginica. They also found that C. ariakensis exhibited higher resistance to the diseases MSX and Dermo as compared to C. virginica
Concurrent with the enthusiasm for introduction of C. ariakensis into the Bay, others expressed concern that C. ariakensis would become an invasive species (Harwell et al. 2010).
Acknowledging this concern, the National Academy of Sciences (NAS) assembled a committee of experts in 2002 to review available research on C. ariakensis. After careful consideration of available data, NAS concluded that not enough information existed to support introduction of diploid C. ariakensis into the Bay (NAS 2004).
In 2003, the United States Congress authorized the U.S. Army Corps of Engineers (USACE), Norfolk District, to develop a Programmatic Environment Impact Statement (PEIS).
As lead agency, USACE worked with the Department of Natural Resources (DNR) of the VMRC to develop the PEIS. For the PEIS, the agencies considered eight alternatives involving either C. virginica or C. ariakensis or both in the Bay, but issued a Record of Decision in 2009 that featured only eastern oysters. This decision directed states surrounding the Bay to enhance efforts to restore the eastern oyster, continue restrictions on harvests, and increase cultivation of eastern oysters through state-controlled aquaculture.
Some researchers, believing that actions related to the Record of Decision would not result in required improvement of water quality in the Bay and that the proposal to introduce nonnative oysters into the Bay would re-surface in the future, continued studies related to C. ariakensis (Harwell et al. 2010). Dew et al. (2003) developed a model for assessing the probability of self-sustaining populations resulting from commercial production of triploid Suminoe oysters. Hudson et al. (2005) investigated reversion and gametogensis of C. ariakensis, in addition to growth, mortality and condition index of C. ariakensis and C. virginica grown in eight field sites of the Chesapeake Bay. Ruesink et al. (2005) addressed the ecosystem effects of introduction of nonnative oysters. Paynter et al. (2008) reported a first comparison of triploid C. virginica and triploid C. ariakensis on growth, disease resistance, and mortality in shallow subtidal estuaries of the Chesapeake Bay. Harding (2007) compared growth rates of diploid DEBY eastern oysters, triploid eastern oysters, and triploid Sumino oysters. Harlan (2007) compared the physiology and biochemistry of C. virginica and C. ariakensis under anoxic conditions. Kingsley-Smith et al. (2008) studied the post-settlement growth and survival of C. ariakensis in simulated emersion regimes. Volsad et al. (2008) developed a model to project population growth of C virginica and C. ariakensis in the Bay based on estimation of annual mortality rates.
Research reported in this paper is the first study involving competition between diploid C. virginica and triploid C. ariakensis. In related studies, Kingsley-Smith et al. (2009) explored survival and growth of triploid C. virginica and triploid C ariakensis in bottom environments of four estuaries of the Chesapeake Bay. They found that at low- and medium- salinity subtidal sites, C. ariakensis shell heights were significantly greater in the single-species samples as compared to the mixed-species samples. Harwell et al. (2010) investigated habitat functions of
triploid C. virginica and triploid C. ariakensis grown together as mixed populations in four bottom locations in the Bay. They found that at subtidal sites of higher salinity, the numbers of organisms associated with reefs of triploid C. virginica were significantly greater than the numbers of organisms associated with triploid C. ariakensis reefs and that mixed-oyster reefs were functionally equivalent to monospecific C. ariakensis reefs. A mechanism to explain this result may involve competitive differences between triploid C. ariakensis and triploid C. virginica relative to other organisms of the Bay (Harwell et al 2010).
A literature search revealed no studies comparing water clearance rates and food assimilation efficiencies of the two species in the Chesapeake Bay. However, studies typically show that C. ariakensis grows more robustly than C. virginica in the Bay (e.g., Calvo 2001). An obvious question is: Does C. ariakensis grow faster because it filters more water per unit of time and/or because it has a higher food assimilation efficiency? Whereas no studies previously addressed this question, several studies focused on water clearance rates and metabolic activities of filter feeding species. Newell (1966) covered mechanisms of feeding for bivalves, explaining in-take mechanisms for food and production of pseudofeces. Berg et al. (1986) discussed variations in seston available for feeding of C. virginica. Beals (2004) reported on clearance rates and particle selectivity in the hard clam, Mercenaria mercenaria. Brownlee et al. (2008) determined metabolic responses of C. virginica and C. ariakensis to bloom-forming phytoplankton, including ichthyotoxic Karlodinium veneficum and Prorocentrum minimum.
The purpose of this research was two-fold: (1) Determine competitive effects of growing diploid C. virginica and triploid C. ariakensis together in confined spaces within the Piankatank and Rappahannock Rivers, and (2) Determine the water clearance rates and food assimilation efficiencies of the diploid C. virginica, diploid C. ariakensis, and triploid C. ariakensis.
Methods: Competitive effects
Methods for determining competitive effects were detailed by Holt (2013). Populations of juvenile diploid C. virginica and triploid C. ariakensis, were randomly selected from stocks at Virginia Institute of Marine Science (VIMS). Initial maximum shell heights and wet weights of a sub-sample of 100 oysters of each species, randomly selected from the populations, were determined. Oysters of both species were randomly assigned to single-species samples and competitive mixtures The number of oysters in each single-species sample and competitive mixture was 100, giving a total of 1,000 oysters for each river. The maximum shell heights and wet weights of oysters in the single-species samples and competitive mixtures were measured. The single-species samples and competitive mixtures were constituted as shown in Table 1.
Table 1 Experimental constitution of single-species samples and competitive mixtures
The single-species samples and competitive mixtures were placed in bags constructed of plastic mesh with square openings about 5 millimeters on a side. The height (30.5 cm) and width (30.5 cm) of bags were selected to give a stocking density of 1,076 juvenile oysters/m2, a stocking density designed to produce competition for food in the water column (Rheault and Rice 1996). The bags were secured in wire-mesh trays to prevent loss of oysters Trays were secured on the bottom of each river at 10 meters from the shoreline. Growth of the oysters in

both rivers was assessed at 3, 6, 9, 18, and 25 weeks, starting in September 2000 and ending in March 2001. At each measurement period, water temperatures were determined with an alcohol thermometer suspended near the trays in each river. Salinity was determined using a handheld refractometer filled with river water. Data analysis. Initial shell heights and wet weights of oysters used in the research were assessed using descriptive statistics and Student’s t-test. At each measurement period, maximum shell heights and wet weights of oysters in single-species samples and competitive mixtures were assessed using descriptive statistics and Student’s t-test Growth of oysters in single-species samples and competitive mixtures grown in the two rivers were compared using two-factor Analysis of Variance (ANOVA). Effects related to competition within single-species samples and competitive mixtures in each river were determined using descriptive statistics, single-factor ANOVA, and Student’s t-test. Increases in shell heights and wet weights of oysters in singlespecies samples and competitive mixtures in the two rivers were calculated as percent increases from initial values Methods described by Zar (1999), together with Microsoft Excel Statistical Package, were used for the statistical analyses.
Methods: Determining water clearance rates and food assimilation efficiencies
Methods for determining water clearance rates and food assimilation efficiencies were detailed by Holt (2013). Water clearance rates and food assimilation efficiencies were determined in the laboratory at VIMS using the biodeposition method as recommended by Newell (2001) and described by Iglesias et al (1998). This method uses the fact that oysters ingest inorganic matter with food (organic matter). They eject inorganic matter as pseudofeces, or it goes through their digestive system and comes out unaffected in their feces. The organic matter is digested and used for body functions, or if undigested, comes out in the feces. Feces
and pseudofeces settle to the bottom of the water column, with the net result being a filtration of the water column (Newell et al. 2005). To obtain biodeposits, oysters of the two species were allowed to filter river water with suspended particulate organic matter (POM) and particulate inorganic matter (PIM) for a given period of time. Biodeposits were collected after the feeding period and analyzed for POM and PIM. The POM and PIM found in biodeposits (feces and pseudofeces), when compared to POM and PIM in the river water, allowed calculation of water clearance rates and food assimilation efficiencies. The experimental apparatus is shown in Figure 1.
Figure 1 Experimental apparatus for determining clearance rates and assimilation efficiencies. Water was pumped from the York River (a) to a retention tank (b) which was then passed through a filter (c) into the experimental tank (d). A baffle to stabilize water current was placed in front of the feeding baskets (N = 1 oyster per basket, 30 baskets per trial: 10 diploid C. virginica; 10 diploid C. ariakensis; 10 triploid C. ariakensis; 10 empty baskets for controls). Water was circulated over the feeding baskets and then drained back into the York River (e).

Water clearance rate (CR) was calculated using the formula given for the biodeposition method by Iglesias et al. (1998):
CR = IFR/PIM
where IFR was the rate that inorganic matter moved through the oyster, i.e., the rate of inorganic matter filtration (g/hour), and PIM was the concentration of particulate inorganic matter in the water supply (g/l).
Food assimilation efficiency (AE) was determined, as recommended by Newell (2001), using the ratio method of Conover (1966). By this method, AE is defined as percentage of utilization (U’) and was calculated as
AE = U΄ = !΄ !΄ ! !΄ !΄ ×100 where F’ was the fraction of organic matter in the ingested food and E’ is the same fraction in the feces.
The independent variables were species and ploidy of oysters. The dependent variables were water clearance rate and food assimilation efficiency. Controlled variables were particulate matter in the river water, temperature of the river water, flow rate of the river water over the baskets, salinity of the river water, and light in the feeding trough.
Data analysis. Water clearance rates and food assimilation efficiencies were assessed using descriptive statistics, one-factor ANOVA, and Tukey multiple comparison test for means. Methods described by Zar (1999), together with Microsoft Excel Statistical Package, were used for the statistical analyses.
Results and Discussion for Competitive Effects
Initial mean shell heights and wet weights. The initial mean shell height of C. virginica was 16.5 mm (n = 50, SD = 3.71) and the mean shell height of C. ariakensis was 18.9 mm (n = 50, SD = 4.36). At the beginning of the research, C. ariakensis displayed a significantly greater mean shell height than C. virginica (t = 2.845, p = 0.00646) although the two species were essentially the same age. The initial mean wet weight of C. virginica was 0.61 g, and the
initial mean wet weight of C. ariakensis was 0.60 g. These initial wet weights were not significantly different (t = 0.0712, p = 0.472)
Water temperatures. Differences in water temperature between the two rivers were not significant over the 25 weeks of the research (t = 0.667, p = 0.545). Maximum water temperatures of 12-13 oC occurred at the beginning of the research in September 2000. Minimum temperatures of 0.5-1.6 oC occurred during November and December 2000, corresponding to measurement periods of 9 and 18 weeks (Figure 2)
Salinity. Salinity was significantly different in the two rivers (t = 6.247, p = 0.00154). Salinity ranged from 10.1 to 12.4 parts per thousand (ppt) in the Piankatank River site and from 11.9 to 16.6 ppt in the Rappahannock River (Figure 3). These values classify the Piankatank River site as a low-salinity site and the Rappahannock River site as a medium-salinity site based on information given by Paynter et al. (2008).

Growth of C. virginica and C. ariakensis in single-species samples. For the measurement periods of 3, 6, 9, 18, and 25 weeks, mean maximum shell heights and wet weights for C. virginica and C. ariakensis in single-species samples in the two rivers are given in Figures 4 and 5, respectively.


Measurements were made

Effects of the two independent variables, which were single-species samples (i.e., 100% C. virginica and 100% C. ariakensis) and river, on growth of oysters at 25 weeks were determined using two-factor ANOVA. Comparison of shell heights for oysters in the singlespecies samples of C. virginica and C. ariakensis grown in the two rivers showed a significant difference in shell heights (F = 228.4, p < 0.0005). In the Piankatank River, mean shell height was 2.97 mm for C. virginica and 5.35 mm for C. ariakensis. In the Rappahannock River, mean shell height was 3.89 mm for C. virginica and 5.98 mm for C. ariakensis There was no significant effect on shell heights caused by river (F = 0.481, p = 0.489). In addition, there was no significant interaction between species and rivers on shell heights (F = 2.82, p = 0.0946).
Wet weights at 25 weeks for oysters grown in the two rivers were significantly different for single-species samples (F = 112.7, p < 0.0005) and for river (F = 13.5, p < 0.0005). Mean wet weight of C. virginica was 2.97 g when grown in the Piankatank River and 3.89 g when grown in the Rappahannock River. Mean wet weight of C. ariakensis was 5.35 g when grown in the Piankatank River and 5.98 g when grown in the Rappahannock River. There was no significant interaction between single-species samples and river on wet weights (F = 0.437, p = 0.509).
Effects of competition on growth. For the measurement periods of 3, 6, 9, 18, and 25 weeks, mean shell heights and wet weights of oysters in the single-species samples and competitive mixtures in the two rivers are depicted in Figures 6 through 9.




Effects of the two independent variables, which were single-species samples/competitive mixtures and river, on growth of oysters at 25 weeks were determined using two-factor ANOVA (Holt 2013). Shell heights at 25 weeks for oysters in the single-species samples and competitive mixtures in both rivers showed a significant difference in shell heights based on effect of singlespecies samples/competitive mixtures (F = 228, p < 0.0005). There was no significant effect on shell heights caused by river (F = 0.481, p = 0.489). There was no significant interaction between single-species samples/competitive mixtures and rivers on shell heights (F = 2.82, p = 0.0946). Wet weights at 25 weeks for oysters in the single-species samples and competitive mixtures from both rivers showed a significant difference based on effect of single-species samples/competitive mixtures (F = 112.7, p < 0.0005) and a significant effect based on river (F = 13.5, p = 0.0003). There was no significant interaction between single-species samples/competitive mixtures and river on wet weights (F = 0.437, p = 0.509).
For each measurement period, the effect of competition on growth of oysters in each river was assessed using single-factor ANOVA (Holt 2013). At 25 weeks, C. ariakensis in competitive mixtures with C. virginica had a significant suppressive effect on shell heights of C. virginica (for the Piankatank, F = 13.2, p < 0.0005; for the Rappahannock, F = 12.6, p < 0.0005). In each river, mean shell height of C. virginica grown with C. ariakensis was smaller than mean shell height of C. virginica in the single-species samples, suggesting that significant interspecific competition existed in the competitive mixtures. On the other hand, C. virginica in competitive mixtures with C. ariakensis had no significant effect on shell heights of C. ariakensis (for the Piankatank, F = 0.294, p = 0.830; for the Rappahannock, F = 1.150, p = 0.330).
On wet weight at 25 weeks, C. ariakensis in competitive mixtures with C. virginica had a significant suppressive effect on wet weights of C. virginica (i.e., for the Piankatank, F = 11.2, p < 0.0005; for the Rappahannock, F = 16.4, p < 0.0005). Mean wet weights of C. virginica in competitive mixtures were smaller than the mean wet weight of C. virginica in the single-species samples, again suggesting significant interspecific competition within the competitive mixtures as compared to the single-species samples.
C. virginica in competitive mixtures with C. ariakensis had a significant effect on wet weights of C. ariakensis (i.e., F = 6.486, p = 0.0003 in the Piankatank and F = 3.75, p = 0.0012 in the Rappahannock). With one exception (wet weight of C. ariakensis in the competitive mixture involving 75% C. ariakensis and 25% C. virginica in the Piankatank), all other wet weights of C. ariakensis in competitive mixtures with C. virginica exceeded the wet weights of C. ariakensis in single-species samples. This result suggests that intraspecific competition among C. ariakensis in the competitive mixtures was reduced by the presence of C. virginica
Results of Student’s t-tests, performed at each measurement period to determine if shell heights and wet weights of oysters in competitive mixtures differed from those in single-species samples, were detailed by Holt (2013). In the Piankatank River, the mean shell heights of C. virginica in competitive mixtures with C. ariakensis were suppressed as compared to singlespecies samples of 100% C. virginica (Figure 6). On the other hand, the presence of C. virginica in competitive mixtures with C. ariakensis did not affect the mean shell heights of C. ariakensis as compared with the single-species samples of 100% C. ariakensis. This result, suppression of shell height of C. virginica by C. ariakensis and no effect of C. virginica on shell height of C. ariakensis, was also observed in the Rappahannock River (Figure 7).
In both the Piankatank and Rappahannock Rivers, mean wet weights of C. virginica grown together with C. ariakensis were suppressed as compared to mean wet weights of C. virginica in the single-species samples. Mean wet weights of C. ariakensis grown in the presence of 25% and 50% C. virginica were not affected as compared to mean wet weights of C. ariakensis in the single-species samples. However, in competitive mixtures involving 75% C. virginica with 25% C. ariakensis, the mean wet weights for C. ariakensis were greater than the mean wet weights of C. ariakensis in the single-species samples of 100% C. ariakensis (Figures 8 and 9). This result suggests that intraspecific competition among C. ariakensis was greater in the single-species samples of 100% C. ariakensis than in the competitive mixtures involving 75% C. virginica and 25% C. ariakensis.
Percent increase in growth over 25 weeks. Percent increases between initial and final mean shell heights and wet weights are shown in Figures 10 and 11, respectively. In both rivers,
C. ariakensis in single-species samples grew more robustly on shell heights and wet weights than C. virginica in single-species samples The percent increase in initial-to-final mean shell height of C. ariakensis in the Piankatank was 98.4 % as compared to 72.7 % for C. virginica, and the percent increase in initial-to-final mean wet weight was 791.7 % as compared to 386.9 % for C. virginica. In the Rappahannock River, the percent increase in initial-to-final mean shell height of C. ariakensis in single-species samples was 95.8 % as compared to 80.6 % for
C. virginica, and the percent increase in initial-to-final mean wet weights was 896.7 % compared to 495.1 %. C. virginica in single-species samples grew larger in the Rappahannock River on both shell height and wet weight as compared to the Piankatank River. The opposite was observed for C. ariakensis, which grew larger on both height and wet weight in the Piankatank River.
Figure 10 Percent increase between initial and final mean shell heights of oysters grown in two rivers. N = 50 for single-species samples and all competitive mixtures, except for competitive mixtures containing 25% C. virginica or 25% C. ariakensis. For these mixtures, N for the 25% aliquot was smaller than 50 if oysters died during the research.
Figure 11 Percent increase between initial and final mean wet weights of oysters grown in two rivers. N = 50 for single-species samples and all competitive mixtures, except for competitive mixtures containing 25% C. virginica or 25% C. ariakensis. For these mixtures, N for the 25% aliquot was smaller than 50 if oysters died during the research.


In both rivers, shell heights for C. virginica in competitive mixtures with C. ariakensis were significantly reduced as compared to shell heights of C. virginica in single-species samples. The percentages of C. ariakensis in the competitive mixtures did not make a significant difference in the suppression. In both rivers, shell heights for C. ariakensis in competitive mixtures with C. virginica were not significantly different from shell heights of C. ariakensis in single-species samples These results suggest that interspecific competition existed between C. ariakensis and C. virginica in competitive mixtures with C. ariakensis being the better competitor for food (Figures 10 and 11).
In the Piankatank River, wet weights of C. virginica in competitive mixtures with C. ariakensis were significantly suppressed when compared to wet weights of C. virginica in single-species samples. The percentages of C. ariakensis in the competitive mixtures did not
make a statistically significant difference in the suppression. In the Rappahannock River, the suppression of wet weights of C. virginica by C. ariakensis in competitive mixtures was significant when compared to the wet weights of C. virginica in single-species samples. The percent of C. ariakensis in the competitive mixtures made a significant difference in wet weights of C. virginica. The higher the percent of C. ariakensis, the greater the suppression in wet weights of C. virginica. The percentage of C. virginica in the competitive mixtures made a significant difference in wet weights of C. ariakensis The higher the percent of C. virginica, the greater the wet weights of C. ariakensis as compared to the wet weights of C. ariakensis in single-species samples, suggesting that intraspecific competition between C. ariakensis was reduced by the presence of C. virginica in the competitive mixtures (Figures 10 and 11).
Mortality. Information on mortality of oysters in single-species samples and competitive mixtures in the two rivers was given by Holt (2013). Mortality did not differ significantly for C. virginica and C. ariakensis in the single-species samples and competitive mixtures in either river, based on single-factor ANOVA (for the Piankatank River, F = 0.549, p = 0.487; for the Rappahannock River, F = 3.42, p = 0.114). The highest mortality occurred in the Piankatank River. For C. virginica, 45 oysters died at 18 weeks. For C. ariakensis, 43 oysters died at 9 weeks and 28 died at 18 weeks. These spikes in mortality corresponded closely to minimum water temperatures shown in Figure 2 for both the Piankatank and Rappahannock Rivers. Given the low mortality of C. virginica and C. ariakensis in the Rappahannock River throughout the research, it is likely that, during the periods of 9 and 18 weeks, low-tide freezing affected some bags of oysters in the Piankatank, but not in Rappahannock River.
Effect of C. virginica on growth of C. ariakensis Student’s t-test was used to compare growth of oysters in single-species samples and competitive mixtures at each measurement
period. At week 3, the mean shell height of C. ariakensis grown in competitive mixtures with C. virginica in both rivers was significantly smaller than the mean shell height of C. ariakensis in the single-species samples. The effect was not apparent at weeks 6, 9, 18, and 25. At these periods, the mean shell heights of C. ariakensis in competitive mixtures with C. virginica did not differ significantly from mean shell heights of C. ariakensis in single-species samples. At these measurement periods, C. virginica had essentially no effect on C. ariakensis in the competitive mixtures (Holt 2013)
At week 3, the mean wet weights of C. ariakensis grown in competitive mixtures with C. virginica were significantly smaller than mean wet weights of C. ariakensis in the singlespecies samples. However, by week 25 the mean wet weights of C. ariakensis grown in competitive mixtures with C. virginica were, in general, larger than the mean wet weights of C. ariakensis grown in single-species samples By the end of the research, C. virginica in competitive mixtures with C. ariakensis, except for the competitive mixture of 75%
C. ariakensis/25% C. virginica, enhanced the growth in wet weights of C. ariakensis.
Results and discussion: Water clearance rates and food assimilation efficiencies
Mean water clearance rates were 0.0121 l/h-g for diploid C. virginica, 0.00984 l/h-g for diploid C. ariakensis, and 0.0103 l/h-g for triploid C. ariakensis (Table 2). Results of singlefactor ANOVA show that water clearance rates were not statistically different for the oysters (Table 2). Mean food assimilation efficiencies were 1.572 %/g for diploid C. virginica, 2.075 %/g for diploid C. ariakensis, and 2.197 %/g for triploid C. ariakensis (Table 2). Food assimilation efficiencies were significantly different for the two species (Table 2). As shown by results of Tukey multiple comparison test for means (Table 3), triploid C. ariakensis had a higher
food assimilation efficiency than diploid C. virginica. There was no significant difference in the mean food assimilation efficiencies of diploid C. ariakensis and triploid C. ariakensis (Table 3).
ranked by mean food
The hypothesis that the more robust growth of triploid C. ariakensis as compared to diploid C. virginica, which was observed in the Piankatank and Rappahannock Rivers during the first part of the thesis research and reported by others (e.g., Kingsley-Smith et al. 2008), may be related to higher water clearance rates was not supported. On the other hand, food assimilation efficiencies of the oysters were significantly different (Tables 2 and 3). The hypothesis that the more robust growth of triploid C. ariakensis, which was observed in the Piankatank and Rappahannock Rivers during the first part of the research, may be related to higher food assimilation efficiencies was supported.
Conclusions
In the study of competitive effects, C ariakensis grew more robustly than C. virginica on shell height and wet weight in both rivers. In competitive mixtures, C. ariakensis suppressed growth of C. virginica (interspecific competition). C. virginica initially affected the growth of C. ariakensis (interspecific competition). However, this effect disappeared early in the research. Thereafter, C. virginica did not affect the growth of C. ariakensis, had minimal effect, or enhanced the growth (specifically, growth in wet weight)
Results of this research, which show more robust growth of C. ariakensis as compared to C. virginica in the Piankatank and Rappahannock Rivers, corroborate the findings of KingsleySmith et al. (2009). They also found that C. ariakensis grew significantly larger than C. virginica at four sites of the Chesapeake Bay, which differed in salinity, tidal regime, water depth, predation intensity, and disease pressure. On the other hand, Kingsley-Smith et al. (2009) reported a different result for competition between the two species. In their research, which involved single-species samples of 100% C. virginica and 100% C. ariakensis and one competitive mixture (50% C. virginica/50% C. ariakensis), all at a stocking density of 348 oysters/m2, they found that at low- and medium-salinity subtidal sites, C. ariakensis shell heights were significantly greater in single-species samples as compared with the competitive mixture (50% C. ariakensis/50% C. virginica). These results suggest a possible suppressive effect of C. virginica on growth in shell heights of C. ariakensis.
Harwell et al. (2010), in a study to determine if C. ariakensis would be an ecologically functional equivalent of C. virginica if introduced into the Chesapeake Bay, found that at subtidal sites of higher salinity, the numbers of organisms per unit of oyster biomass associated with C. virginica reefs were significantly greater than the numbers of organisms associated with
C. ariakensis reefs. This result suggests that C. ariakensis may have suppressed the number of organisms growing on reefs with C. ariakensis Harwell et al. (2010) also found that reefs with the competitive mixture of C. virginica and C. ariakensis most often displayed patterns of benthic growth similar to those of monospecific C. ariakensis reefs, suggesting that C. ariakensis may have a competitive effect on benthic populations.
The studies of Kingsley-Smith et al. (2009), which suggested a competitive effect of C. virginica on C. ariakensis, and Harwell et al. (2010), which suggested a possible competitive effect of C. ariakensis on other organisms, both featured similar stocking densities for the competitive mixtures of C. virginica and C. ariakensis. The stocking density used by KingsleySmith et al. (2009) was 348 juvenile oysters/m2. The stocking density used by Harwell et al. (2010) was 400 juvenile oysters/m2. The stocking density used in this research was much higher at 1,076 juvenile oysters/m2.
The definite competitive effects on growth of oysters observed in this research raise questions about whether a threshold stocking density exists for competitive mixtures of
C. virginica and C. ariakensis. A wide range of densities have been reported for C. virginica growing in the wild in the Chesapeake Bay. Schulte et al. (2009) reported densities ranging from an average of 2 oysters/m2 for pre-restoration bottom sites, an average of 250 oysters/m2 on lowrelief reefs, and an average of 1,027 oysters/m2 on high-relief reefs. Burke (2010) reported mean oyster densities of 978 oysters/m2 and peak densities of 2,000 oysters/m2 growing on riprap made of granite and concrete along shorelines of the Lynnhaven and Rappahannock Rivers. Thus, the stocking density of 1,076 juvenile oysters/m2 used in the research is not unrealistic for an in situ setting, raising again the need to determine if there is a threshold stocking density at which competitive effects between C. virginica and C. ariakensis appear. These competitive effects
may manifest, according to Bergon et al. (1996), as growth effects, which were demonstrated in the current research; survivability effects, which were not demonstrated in this research; and effects on fecundity, which were not investigated in this research.
Implications for the Chesapeake Bay
This research is the first study of competitive effects between diploid C. virginica and triploid C. ariakensis and the first study to compare water clearance rates and food assimilation efficiencies of diploid C. virginica and triploid C. ariakensis As such, the research has important implications for improving water quality of the Chesapeake Bay, preserving the native oyster as a species, and reviving a languishing oyster industry.
The research has both positive and negative implications. C. ariakensis grew more robustly than C. virginica in both the Piankatank and Rappahannock Rivers and showed equivalent survivability with C. virginica when low-tide freezing was not an issue. This outcome supports early conclusions that C. ariakensis, grown in controlled aquaculture or released into the Bay, may help to improve water quality, revive a dying fishery, restore cultural traditions for towns dependent on oyster commerce, and satisfy customer demand for oysters as food. On the other hand, this research showed a definite competitive effect of C. ariakensis on C. virginica at the stocking density (i.e., 1076 juvenile oysters/m2) used for the competitive mixtures. Additional research is needed to determine if there is a threshold stocking density at which competitive effects of C. ariakensis on C. virginica are first observed. If such a threshold stocking density is affirmed, additional modeling is needed to determine the probability of such a stocking density occurring naturally within the Chesapeake Bay and to assess probable ecological outcomes.
Acknowledgements
The author gratefully acknowledges the encouragement and support of Dr. Gary Whiting, Dr. Russell Burke, and Dr. Michael Meyer of Christopher Newport University; Dr. Standish K. Allen, Jr., Ms. Amiee Howe, and Ms. Elizabeth Walker of the Virginia Institute of Marine Science; Dr. RIE Newell of the University of Maryland; Dr. James Wesson of the Virginia Marine Resources Commission; and Mr. Jeff Kellum of the Virginia Fisheries Council.
REFERENCES
Allen SK Jr. Current and future roles of genetics in oyster restoration or recovery. Paper presented at: Workshop on oyster research and restoration in U. S. coastal waters: Strategies for the future; 2003 Sep 8-9; Annapolis, MD.
Beals CD. 2004. Clearance rates and particle selectivity in the hard clam, Mercenaria mercenaria, from warm water habitats [thesis]. [Gainsville (FL)]: University of Florida. 56 pp.
Berg JA, Newell RIE. 1986. Temporal and spacial variations in the composition of seston available to suspension feeder Crassostrea virginica. Estuarine, Coastal and Shelf Science. 23: 375-386.
Brownlee EF, Sellner SG, Sellner KG, Nonogaki H, Adolf JE, Bachvaroff TR, Place AR. 2008 May. Responses of Crassostrea virginica (Gmelin) and C. ariakensis (Fujita) to bloom-forming phytoplankton including ichthyotoxic Karlodinium veneficum (Ballantine). Journal of Shellfish Research 27(3): 581-591.
Brumbaugh RD, Sorabella LA, Garcia CO, Goldsborough WJ, Wesson JA. 2000. Building a case for community-based oyster restoration: An example from Hampton Roads, Virginia, U.S.A. Journal of Shellfish Research. 10(1): 467-472.
Burke RP. 2010. Alternative substrates as a native oyster (Crassostrea virginica) reef restoration strategy in Chesapeake Bay [dissertation]. [Gloucester Point (VA)]: The College of William and Mary. 286 pp.
Calvo GW, Luckenbach MW, Allen SK Jr, Burreson EM. 2001. A comparative field study of Crassostrea ariakensis (Fujita 1913) and Crassostrea virginica (Gmelin 1791) in relation to salinity in Virginia. Journal of Shellfish Research. 20(1): 224-229.
Chesapeake Bay Foundation [CBF]. 2010. The state of the Bay: Chesapeake Bay Foundation Save the Bay Publication, 4 pp.
Conover RJ. 1966. Assimilation of organic matter by zooplankton. Limnology and Oceanography. 11: 338-354.
Dew JR, Berkson J, Hallerman EM, Allen SK Jr. 2003. A model for assessing the likelihood of self-sustaining populations resulting from commercial production of triploid Suminoe oysters (Crassostrea ariakensis) in Chesapeake Bay Fishery Bulletin. 101(4): 758-769.
Encomio VG, Stickler SM, Allen SK Jr, Chu F-L. 2005. Performance of “natural Dermoresistant” oyster stocks: Survival, disease, growth, condition and energy reserves. Journal of Shellfish Research. 24: 143.
Fisher WS. 1996. Preface. Journal of Shellfish Research. 15: 1
Gross F, Smyth RC. 1946. The decline of oyster populations. Nature. 157: 540-542.
Harding JM. 2007. Comparison of growth rates between diploid DEBY eastern oysters (Crassostrea virginica, Gmelin 1791), triploid eastern oysters, and triploid Suminoe oysters (C. ariakensis, Fugita 1913). Journal of Shellfish Research. 26(4): 961-972.
Hargis WJ, Haven DS. 1988. Rehabilitation of the troubled oyster industry of the lower Chesapeake Bay. Journal of Shellfish Research. 7(2): 271-279.
Hargis WJ, Haven DS. 1988. The imperiled oyster industry of Virginia. A critical analysis with recommendations for restoration. Special Report No. 290 in applied marine science and ocean engineering. Gloucester Point (VA): Virginia Institute of Marine Science. 130 pp.
Hargis WJ. The evolution of the Chesapeake oyster reef system during the Holocene epoch. In: Luckenbach MW, Mann R, Wesson JA, editors. Habitat restoration: a synopsis and synthesis of approaches. Gloucester Point (VA): Virginia Institute of Marine Science Press; 1999. p. 5-23.
Harlan NP. 2007. A comparison of the physiology and biochemistry of the eastern oyster, Crassostrea virginica, and the Asian oyster, Crassostrea ariakensis [thesis]. [College Park (MD]: University of Maryland. 62 pp.
Harwell HD, Kingsley-Smith PR, Kellogg ML, Allen SM, Allen SK Jr, Meritt DW, Paynter KT Jr, Luckenbach MW. 2010. A comparison of Crassostrea virginica and C. ariakensis in Chesapeake Bay: Does oyster species affect habitat function? Journal of Shellfish Research. 29(2): 253-269.
Holt KF. 2013. A study of competition between native eastern oyster (Crassostrea virginica) and nonnative Suminoe oyster (Crassostrea ariakensis) and implications for the Chesapeake Bay [thesis]. [Newport News, (VA)]: Christopher Newport University. 134 pp.
Hudson KL, Kozlowski A, Erskine AJ, Allen SK Jr. 2005. Comparative field trials of triploid Crassostrea ariakensis with C. virginica at eight field sites in the Chesapeake Bay: growth, mortality, condition, reversion and gametogenesis. Journal of Shellfish Research. 24: 658-659.
Igelesias JIP, Urrutia MB, Navarro E, Ibarrola I. 1998. Measuring feeding and absorption in suspension-feeding bivalves: An appraisal of the biodeposition method. Journal of Experimental Marine Biology and Ecology 219: 71-86.
Kingsley-Smith PR, Harwell HD, Kellogg ML, Allen SM, Allen SK Jr, Merritt DW, Paynter KT Jr, Luckenbach MW. 2008. Survival and growth of triploid Crassostrea virginica (Gmelin, 1791) and C. ariakensis (Fujita, 1913) in bottom environments of Chesapeake Bay: Implications for an introduction. Journal of Shellfish Research. 28: 169-184.
Kingsley-Smith PR, Luckenbach MW. 2008. Post-settlement survival and growth of the Suminoe oyster, Crassostrea ariakensis, exposed to simulated emersion regimes. Journal of Shellfish Research. 27(3): 609-618.
Lenihan HS, Peterson CH. 1998. How habitat degradation through fishery disturbances enhances impacts of hypoxia on oyster reefs. Ecology applications. 8: 128-140.
National Academy of Sciences. [NAS] 2004. Nonnative oysters in Chesapeake Bay. National Academies Press. [Internet]. Available from: www.nap.edu
Newell RIE, Langdon CJ. Mechanisms and physiology of larval and adult feeding. In: Kennedy VS, Newell RIE, editors. The eastern oyster Crassostrea virginica. College Park (MD): Maryland Sea Grant College; 1966. p. 185-230.
Newell RIE. 2001 30 Dec. Consultation meeting between Newell RIE and Holt KF at Horn Point Laboratory, University of Maryland Center for Environmental Science, Cambridge, MD (unpublished notes).
Newell RIE, Fisher TR, Holyoke RR, Cornwell JC. Influence of eastern oysters on nitrogen and phosphorus regeneration in Chesapeake Bay, USA. In: Dame R, Olenis S, editors. The comparative roles of suspension feeder in ecosystems. Springer (Netherlands): NATO Science Series: IV – Earth and Environmental Sciences. 47. 2005. p. 3-120.
Paynter KT, Goodwin JD, Chen ME, Ward NJ, Sherman MW, Meritt DW, Allen SK Jr. 2008. Crassostrea ariakensis in Chesapeake Bay: Growth, disease and mortality in shallow water environments. Journal of Shellfish Research. 27: 509-515.
Rheault RB, Rice MA. 1996. Food limited growth and condition index in the eastern oyster, Crassostrea virginica (Gmelin 1791), and the bay scallop, Argopecten irradians (Lamarch 1819). Journal of Shellfish Research. 15(2): 271-283.
Rothschild BJ, Ault JS, Goulletquer P, Heral M. 1994. Decline of the Chesapeake Bay oyster population: A century of habitat destruction and overfishing. Marine Ecology Program Series. 111: 29-39.
Ruesink SL, Lenihan HS, Trimble AC, Heiman KW, Micheli F, Byers JE, Kay MC. 2005. Introduction of nonnative oysters: Ecosystem effects and restoration implications. Annual Review of Ecology Evolution Systems. 36: 643-689.
Schulte DM, Burke RP, Lipcius RN. 2009. Unprecedented restoration of a native oyster metapopulation. Science. 325: 1124-1128
Shumway SE. 1996. Natural environmental factors. In: Kennedy VS, Newell RIE, editors. The eastern oyster, Crassostrea virginica. Annappolis (MD): Maryland Sea Grant. 467-513.
United States Army Corps of Engineers [USACE]. 2009. Final Programmatic Environmental Impact Statement for oyster restoration in Chesapeake Bay including the use of a native and/or nonnative oyster [Internet]. Available from: www.nao.usace.army.mil/OysterEIS/
United States Army Corps of Engineers [USACE]. 2009. Record of Decision for Final Programmatic Environmental Impact Statement for oyster restoration in Chesapeake Bay including the use of a native and/or nonnative oyster [Internet]. Available from: http://www.nao.usace.army.mil/Portals/31/docs/civilworks/oysters/oysterdecision.pdf
Volstad JH, Dew J, Tarnowski M. 2008. Estimates of annual mortality rates for eastern oysters (Crassostrea virginica) based on box counts and application of those rates to project population growth of C. virginica and C. ariakensis. Journal of Shellfish Research. 27(3): 525-533.
Zar JH. 1999. Biostatistical analysis. Upper Saddle River (NJ): Prentice-Hall.
Zhou M, Allen SK Jr. 2003. A review of published work on Crassostrea ariakensis. Journal of Shellfish Research. 22: 1-20.
Biography for Katherine Frances Holt
Katherine Frances Holt is a 2002 honor graduate of Bruton High School in Williamsburg, Virginia. She is a 2007 graduate of James Madison University with a Bachelor of Science Degree in Biology with a minor in Art. She received her Master of Science Degree in Environmental Science from Christopher Newport University in May 2013. She worked for the Virginia Institute of Marine Science (VIMS) in the Aquaculture, Genetic, and Breeding Technology Center as a flow cytometry technician and hatchery assistant. While at VIMS she performed laboratory tests to access ploidy status of native and non-native oysters via the flow cytometry; performed tasks necessary to sustain controlled growth environments for oyster spat (e.g., cleaning tanks, changing water, monitoring temperature and nutrients); assisted Ph.D. and master’s degree candidates in their research; collected and maintained on- and off-site research projects; assisted in promoting and executing Virginia /National Environmental Oyster initiatives; attended and presented individual research poster presentation at C. ariakensis Symposium held at the College of William and Mary; worked with members of the Virginia Marine Resources Commission; worked with Virginia Shellfish Council and members of the oyster industry on conservation and restoration initiatives; and worked with the Chesapeake Bay Foundation on promotion of oyster restoration/conservation initiatives. Katherine currently works for Technology Commercialization Center as an operations manager for projects related to technology transfers to industry.