
Ths

ÔN TẬP NÂNG CAO
CÁC CHỦ ĐỀ SINH HỌC 12
MÔN SINH HỌC (TÀI LIỆU LƯU HÀNH NỘI BỘ)
MỤC LỤC STT NỘI DUNG TRANG
Chuyên đề: Di truyền học
2 Chương 1. Cơ sở vật chất của hiện tượng di truyền và biến dị
3 Chương 2. Tính quy luật của hiện tượng di truyền
4 Chương 3. Di truyền học quần thể
5 Chương 4. Ứng dụng di truyền học
6 Chương 5. Di truyền học người Chuyên đề: Tiến hoá
7 Chương 1. Bằng chứng tiến hóa
8 Chương 2. Nguyên nhân và cơ chế tiến hóa
9 Chương 3. Sự phát sinh và phát triển của sự sống trên trái đất Chuyên đề: Sinh thái học
10 Chương 1. Cơ thể và môi trường
11 Chương 2. Quần thể sinh vật
12 Chương 3. Quần xã sinh vật
13 Chương 4. Hệ sinh thái, sinh quyển và sinh thái học với quản lí tài nguyên thiên nhiên
CHUYÊN ĐỀ DI TRUYỀN HỌC
CHUƠNG 1. CƠ SỞ VẬT CHẤT CỦA HIỆN TƯỢNG DI TRUYỀ
A - CƠ SỞ VẬT CHẤT CỦA HIỆN TƯỢNG DI TRUYỀ
PHÂN TỬ
I. CẤU TRÚC CỦA AXIT NUCLEIC
1. Cấu trúc ADN
a) Cấu tao hóa học của ADN
- ADN luôn tồn tại trong nhân tế bào và có mặt ở cả ti thể, lạp thể. ADN chứa các
nguyên tố hóa học chủ yếu C, H, O, N và P.
- ADN là đại phân tử, có khối lượng phân tử lớn, chiều dài có thể đạt tới hàng trăm micromet khối lượng phân tử có từ 4 đến 8 triệu, một số có thể đạt tới 16 triệu đvC.
- ADN cấu tạo theo nguyên tắc đa phân, mỗi nucleotit có ba thành phần, trong đó thành phần cơ bản là bazơnitric. Có 4 loại nuleotit mang tên gọi của các bazơnitric, trong đó A và G có kích thước lớn, T và X có kích thước bé.
- Trên mạch đơn của phân tử ADN các đơn phân liên kết với nhau bằng liên kết hoá trị là liên kết được hình thành giữa đường C5H10O4 của nucleotit này với phân tử H3PO4 của nucleotit kế tiếp. Liên kết hoá trị là liên kết rất bền đảm bảo cho thông tin di truyền trên mỗi mạch đơn ổn định kể cả khi ADN tái bản và phiên mã.
- Từ 4 loại nucleotit có thể tạo nên tính đa dạng và đặc thù của ADN ở các loài sinh vật bởi số lượng, thành phần, trình tự phân bố của nucleotit.
b) Cấu trúc không gian của ADN (Mô hình Oatxơn và Crick)
+ ADN là một chuỗi xoắn kép gồm 2 mạch đơn (mạch polinucleotit) quấn song song
quanh một trục tưởng tượng trong không gian theo chiều từ trái sang phải (xoắn phải) như một thang dây xoắn: tay thang là phân tử đường (C5H10O4) và axit photphoric sắp xếp xen kẽ nhau, mỗi bậc thang là một cặp bazơnitric đứng đối diện và liên kết với nhau bằng liên kết hiđro theo nguyên tắc bổ sung (NTBS). Đó là nguyên tắc A của mạch đơn này có kích thước lớn bổ sung với T của mạch đơn kia có kích thước bé và nối với nhau bằng 2 liên kết hiđro. G của mạch đơn này có kích thước lớn bổ sung với X của mạch đơn kia có kích thước bé và nối với nhau bằng 3 liên kết hiđro và ngược lại.
+ Trong phân tử ADN, do các cặp nucleotit liên kết với nhau theo NTBS đã đảm bảo cho chiều rộng của chuỗi xoắn kép bằng 20 Ǻ, khoảng cách giữa các bậc thang trên các
chuỗi xoắn bằng 3,4 Ǻ, phân tử ADN xoắn theo chu kì xoắn, mỗi chu kì xoắn có 10 cặp nucleotit, có chiều cao 34 Ǻ.
- ADN của một số virut chỉ gồm một mạch polinucleotit. ADN của vi khuẩn và
ADN của lạp thể, ti thể lại có dạng vòng khép kín.
c) Tính đặc trưng của phân tử ADN
+ ADN đặc trưng bởi số lượng, thành phần trình tự phân bố các nucleotit, vì vậy từ 4 loại nucleotit đã tạo nên nhiều loại phân tử ADN đặc trưng cho mỗi loài.
+ ADN đặc trưng bởi tỉ lệ A+T G+X
+ ADN đặc trưng bởi số lượng, thành phần trình tự phân bố các gen trong từng nhóm gen liên kết.
2. Cấu trúc ARN
- ARN là một đa phân tử được cấu tạo từ nhiều đơn phân.
- Có 4 loại ribonucleotit tạo nên các phân tử ARN: Ađenin, Uraxin, Xitozin, Guanin, mỗi đơn phân gồm 3 thành phần: bazơnitric, đường ribozơ (C5H10O5) và H3PO4.
- Trên phân tử ARN các ribonucleotit liên kết với nhau bằng liên kết hoá trị giữa đường C5H10O5 của ribonucleotit này với phân tử H3PO4 của ribonucleotit kế tiếp.
- Có 3 loại ARN: rARN chiếm 70-80%, tARN chiếm 10-20%, mARN chiếm 5-10%.
- Mỗi phân tử mARN có khoảng 600 đến 1500 đơn phân, tARN gồm 80 đến 100 đơn phân, trong tARN ngoài 4 loại ribonucleotit kể trên còn có 1 số biến dạng của các bazơnitric (trên tARN có những đoạn xoắn giống cấu trúc ADN, tại đó các ribonucleotit liên kết với nhau theo NTBS (A-U, G-X). Có những đoạn không liên kết được với nhau theo NTBS vì chứa những biến dạng của các bazơnitric, những đoạn này tạo thành những thuỳ tròn. Nhờ cách cấu tạo như vậy nên mỗi tARN có hai bộ phận quan trọng: bộ ba đối mã và đoạn mang axit amin có tận cùng là adenin.
- Phân tử rARN có dạng mạch đơn, hoặc quấn lại tương tự tARN trong đó có tới 70% số ribonucleotit liên kết với nhau theo nguyên tắc bổ sung. Trong tế bào nhân sơ có 3 loại rARN (23S, 5S và 16S); ở sinh vật nhân thật có tới 6 loại rARN (28S, 23S, 18S, 16S, 5,8S, 5S) với số ribonucleoti từt 120 đến 5000/1 phân tử
- Ngoài ba loại ARN tồn tại trong các loài sinh vật mà vật chất di truyền là ADN thì ở những loài virut vật chất di truyền là ARN thì ARN của chúng có dạng mạch đơn, một vài loại có ARN 2 mạch.
II. CẤU TRÚC PROTEIN
1. Cấu trúc hoá học
- Là hợp chất hữu cơ gồm 4 nguyên tố cơ bản C, H. O, N thường có thêm S và đôi lúc có P.
- Thuộc loại đại phân tử, phân tử lớn nhất dài 0,1 micromet, phân tử lượng có thể đạt tới 1,5 triệu đvC.
- Thuộc loại đa phân tử, đơn phân là các axit amin.
- Có 20 loại axit amin tạo nên các protein, mỗi axit amin có 3 thành phần: gốc cacbon (R), nhóm amin (-NH2), nhóm cacboxyl (-COOH), chúng khác nhau bởi gốc R. Mỗi axit amin có kích thước trung bình 3Ǻ
- Trên phân tử protein, các axit amin liên kết với nhau bằng liên kết peptit đó là liên kết giữa nhóm amin của axit amin này với nhóm cacboxyl của axit amin bên cạnh cùng nhau mất đi một phân tử nước. Nhiều liên kết peptit tạo thành một chuỗi polipeptit. Mỗi phân tử protein có thể gồm một hay một số chuỗi polipeptit cùng loại hay khác loại.
- Từ 20 loại axit amin đã tạo nên khoảng 1014 – 1015 loại protein đặc trưng cho mỗi loài. Các phân tử protein phân biệt với nhau bởi số lượng thành phần, trình tự phân bố các axit amin.
2. Cấu trúc không gian
Có 4 bậc cấu trúc không gian
- Cấu trúc bậc I: do các axit amin liên kết với nhau bằng liên kết peptit, đứng ở đầu mạch polipeptit là nhóm amin, cuối mạch là nhóm cacboxyl.
- Cấu trức bậc II: có dạng xoắn trái, kiểu xoắn anpha, chiều cao một vòng xoắn 5,4 A0 , với 3,7 axit amin/1 vòng xoắn còn ở chuỗi bêta mỗi vòng xoắn lại có 5,1 axit amin. Có những protein không có cấu trúc xoắn hoặc chỉ cuộn xoắn ở một phần của polipeptit.
- Cấu trúc bậc III: là hình dạng của phân tử protein trong không gian ba chiều, do xoắn cấp II cuộn theo kiểu đặc trưng cho mỗi loại protein, tạo thành những khối hình cầu.
- Cấu trúc bậc IV: là những protein gồm 2 hoặc nhiều chuỗi polipeptit kết hợp với nhau. Ví dụ, phân tử hemoglobin gồm 2 chuỗi anpha và 2 chuỗi bêta, mỗi chuỗi chứa một nhân hem với một nguyên tử Fe.
3. Tính đặc trưng và tính nhiều dạng của protein
- Protein đặc trưng bởi số lượng thành phần, trình tự phân bố các axit amin trong chuỗi polipeptit. Vì vậy, từ 20 loại axit amin đã tạo nên 1014 – 1015 loại protein rất đặc trưng và
đa dạng cho mỗi loài sinh vật.
- Protein đặc trưng bởi số lượng thành phần trình tự phân bố các chuỗi polipeptit trong mỗi phân tử protein.
- Protein đặc trưng bởi các kiểu cấu trúc không gian của các loại protein để thực hiện các chức năng sinh học.
III. CƠ CHẾ TỔNG HỢP ADN, ARN
1. Cơ chế tổng hợp ADN
VÀ PROTEIN
- Dưới tác động cửa enzim ADN – polimeraza, các liên kết hiđro trên phân tử ADN bị cắt, 2 mạch đơn của ADN tách nhau ra, trên mỗi mạch đơn các nucleotit lần lượt liên kết với các nucleotit tự do của môi trường theo NTBS. Kết quả từ một phân tử ADN mẹ hình thành 2 phân tử ADN con, trong mỗi ADN con có một mạch là nguyên liệu cũ, một mạch là nguyên liệu mới được xây dựng nên, theo nguyên tắc bán bảo toàn.
- Sự tổng hợp ADN là cơ sở hình thành NST, đảm bảo cho quá trình phân bào nguyên phân, giảm phân, thụ tinh xảy ra bình thường, thông tin di truyền của loài được ổn định. Ở cấp độ tế bào và cấp độ phân tử qua các thế hệ. Nhờ đó con sinh ra giống với bố mẹ, ông bà tổ tiên.
2. Cơ chế tổng hợp mARN
- Dưới tác dụng của enzim ARN – polimeraza. Các liên kết hiđro trên một đoạn phân tử ADN ứng với một hay một số gen lần lượt bị cắt đứt, quá trình lắp ráp các ribonucleotit tự do của một trường nội bào với các nucleotit trên mạch mã gốc của gen (mạch 3’ – 5’) theo
NTBS A-U, G-X xảy ra. Kết quả tạo ra các mARN có chiều 5’-3’. Sau đó 2 mạch gen lại liên kết với nhau theo NTBS. Sự tổng hợp tARN và rARN chũng theo cơ chế trên. - Ở sinh vật trước nhân sự phiên mã cùng một lúc nhiều phân tử mARN, các mARN được sử dụng này trở thành bản phiên mã chính thức. Còn ở sinh vật nhân chuẩn sự phiên mã từng mARN riêng biệt, các mARN này sau đó phải được chế biến lại bằng cách loại bỏ
các đoạn vô nghĩa, giữ lại các đoạn có nghĩa tạo ra mARN trưởng thành.
- Sự tổng hợp ARN đảm bảo cho quá trình dịch mã chính xác ở tế bào chất để tạo nên các protein cần thiết cho tế bào.
3. Cơ chế tổng hợp protein
Gồm 2 giai đoạn:
Giai đoạn 1: Tổng hợp ARN để chuyển thông tin di truyền từ gen sang sản phẩm protein
Giai đoạn 2: Tổng hợp protein ở tế bào chất gồm 4 bước cơ bản.
+ Bước 1: Hoạt hoá các axit amin: Các axit amin được hoạt hoá bằng nguồn năng lượng ATP rồi mỗi axit amin được gắn vào một tARN để đi vào riboxom thành dòng liên tục.
+ Bước 2: Mở đầu chuỗi polipeptit: Có sự thanh gia của riboxom, bộ ba mở đầu AUG, tARN mang axit amin mở đầu tiến vào riboxom đối mã của nó khớp với mã mở đầu trêm mARN theo NTBS.
+ Bước 3: Kéo dài chuỗi polipeptit: tARN vận chuyển axit amin thứ nhất tiến vào riboxom đối mã của nó khớp với mã thứ nhất trến mARN theo NTBS, một liên kết peptit được hình thành giữa axit amin mở đầu với axit amin thứ nhất. Riboxom chuyển dịch sang bộ ba thứ 2 đẩy tARN axit amin mở đầu ra ngoài. Lập tức tARN axit amin thứ 2 tiến vào riboxom đối mã của nó lắp ráp với mã bộ ba trên mARN theo NTBS. Cứ tiến hành theo phương thức đó cho đến tận bộ ba tiếp giáp với bộ ba kết thúc chuỗi polipeptit lúc này có cấu trúc
+ Bước 4: Kết thúc chuỗi polipeptit: Riboxom chuyển dịch sang bộ ba kết thúc lúc này ngừng quá trình dịch mã 2 tiểu phần của riboxom tách nhau ra tARN axit amin cuối cùng được tách khỏi chuỗi polipeptit. Một enzim khác loại bỏ axit amin mở đầu giải phóng chuỗi polipeptit.
Trên mỗi mARN cùng lúc có thể có nhiều riboxom trượt qua với khoảng cách là 51Ǻ → 102 Ǻ, nghĩa là trên mỗi mARN có thể tổng hợp nhiều protein cùng loại.
Sự tổng hợp protein góp phần thực hiện chức năng biểu hiện tính trạng, cung cấp nguyên liệu cấu tạo nên các bào quan và đảm nhận nhiều chức năng khác nhau.
IV. CHỨC NĂNG CỦA ADN, ARN VÀ PROTEIN
1. Chức năng của ADN
+ Chứa thông tin di truyền, thông tin đặc trưng cho mỗi loại bởi trình tự phân bố các nucleotit trên phân tử ADN
+ Có khả năng nhân đôi chính xác để truyền thông tin di truyền qua các thể hệ.
+ Chứa các gen khác nhau, giữ chức năng khác nhau.
+ Có khả năng đột biến tạo nên thông tin di truyền mới.
2. Chức năng của các loại ARN được tổng hợp từ ADN
- Chức năng của mARN: bản phiên thông tin di truyền từ gen cấu trúc, trực tiếp tham gia tổng hợp protein dụa trên cấu trúc và trình tự các bộ ba trên mARN.
- Chức năng của tARN: vận chuyển lắp ráp chính xác các axit amin vào chuối polipeptit dựa trên nguyên tắc đối mã di truyền giữa bộ ba đối mã trên tARN với bộ ba mã phiên trên mARN.
- Chức năng của rARN: liên kết với các phân tử protein tạo trên các riboxom tiếp xúc với mARN và chuyển dịch từng bước trên mARN, mỗi bước là một bộ ba nhờ đó mà lắp ráp chính xác các axit amin vào chuỗi polipeptit theo đúng thông tin di truyền được quy định từ gen cấu trúc.
3. Chức năng của protein
- Là thành phần cấu tạo chủ yếu chất nguyên sinh hợp phần quan trọng xây dựng nên các bào quan, màng sinh chất…
- Tạo nên các enzim xúc tác các phản ứng sinh hoá.
- Tạo nên các hoocmon có chức năng điều hoà quá trình trao đổi chất trong tế bào, cơ thể
- Hình thành các kháng thể, có chức năng bảo vvệ cơ thể chống lại các vi khuẩn gây bệnh.
- Tham gia vào chức năng vận động của tế bào và cơ thể.
- Phân giải protein tạo năng lượng cung cấp cho các hoạt động sống của tế bào và cơ thể.
-Tóm lại protein đảm nhận nhiều chức năng liên quan đến toàn bộ hoạt động sống của tế bào, quy định tính trạng của cơ thể sống.
V. SỰ ĐIỀU HOÀ HOẠT ĐỘNG CỦA GEN
Cơ chế điều hoà sinh tổng hợp protein của gen rất phức tạp, có sự khác biệt rõ rệt giữa sinh vật trước nhân và sinh vật nhân chuẩn. Sau đây là cơ chế điều hoà ở sinh vật trước nhân:
- Trong tế bào có rât nhiều gen cấu trúc, không phải các gen đó đều phiên mã, tổng hợp protein đồng thời. Sự điều hoà hoạt động của gen được thực hiện qua cơ chế điều hoà. Vào năm 1961, F.Jacop và J.Mono đã phát hiện sự điều hoà hoạt động của gen ở E.coli
- Một mô hình điều hoà bao gồm các hệ thống gen sau:
+ Một gen điều hoà (R), gen này làm khuôn sản xuất một loại protein ức chế có tác dụng điều chỉnh hoạt động của một nhóm gen cấu trúc qua tương tác với gen chỉ huy.
aaMD – aa1 – aa2 … aan vẫn còn gắn với tARN axit amin thứ n. - 8
+ Một gen chỉ huy (O) nằm kề trước nhóm gen cấu trúc, là vị trí tương tác với chất ức chế.
+ Một gen khởi động (P) nằm trước gen chỉ huy và có thể trùm lên một phần hoặc toàn bộ gen này, đó là vị trí tương tác của ARN – polimeraza để khởi đầu phiên mã.
+ Một nhóm gen cấu trúc liên quan với nhau về chức năng, nằm kề nhau cùng phiên mã
tạo ra một sợi mARN chung đối với sinh vật trước nhân, còn sinh vật nhân chuẩn phiên mã
chỉ tạo ra 1 mARN riêng biệt.
Một operon chỉ gồm có gen chỉ huy và các gen cấu trúc do nó kiểm soát.
- Cơ chế điều hoà diễn ra như sau:
Gen điều hoà chỉ huy tổng hợp một loại protein ức chế, protein này gắn vào gen chỉ huy (O) làm ngăn cản hoạt động của enzim phiên mã. Vì vậy ức chế hoạt động tổng hợp ARN của các gen cấu trúc. Khi trong môi trường nội bào có chất cảm ứng, chất này kết hợp với protein ức chế làm vô hiệu hoá chất ức chế, không gắn vào gen chỉ huy. Kết quả là gen chỉ huy làm cho nhóm gen cấu trúc chuyển từ trạng thái ức chế sang trạng thái hoạt động. Quá trình phiên mã lại xảy ra.
Cơ chế điều hoà ở sinh vật nhân chuẩn rất phức tạp đến nay còn nhiều vấn đề chưa rõ.
VI. MÃ DI TRUYỀN, ĐẶC ĐIỂM CỦA MÃ DI TRUYỀN
1. Khái niệm mã bộ ba
Cứ 3 nucleotit cùng loại hay khác loại đứng kế tiếp nhau trên phân tử ADN hoặc trên mARN mã hoá cho 1 axit amin hoặc làm nhiệm vụ kết thức chuỗi polipeptit gọi là mã bộ ba.
2. Mã di truyền là mã bộ ba
- Nếu mỗi nucleotit mã hoá 1 axit amin thì 4 loại nucleotit chỉ mã hoá được 4 loại axit amin.
- Nếu cứ 2 nucleotit cùng loại hay khác loại mã hoá cho 1 axit amin thì chỉ tạo được 42 = 16 mã bộ ba không đủ để mã hoá cho 20 loại axit amin.
- Nếu theo nguyên tắc mã bộ ba sẽ tạo được 43 = 64 mã bộ ba đủ để mã hoá cho 20 loại axit amin.
- Nếu theo nguyên tắc mã bộ bốn sẽ tạo được 44 = 256 bộ mã hoá lại quá thừa. Vậy về mặt suy luận lí thuyết mã bộ ba là mã phù hợp.
Trong nghiên cứu, khi thêm bớt 1, 2, 3 nucleotit trong gen, người ta nhận thấy mã bộ ba là mã phù hợp và đã xác định được có 64 bộ ba được sử dụng để mã hoá axit amin. Trong đó có Metionin ứng với mã mở đầu TAX đó là tín hiệu bắt đầu sự tổng hợp chuối polipeptit. Ba bộ ba còn lại ATT, ATX, AXT là mã kết thúc.
Hai mươi loại axit amin được mã hoá bởi 61 bộ ba. Như vậy mỗi axit amin được mã hoá bởi 1 số bộ ba. Ví dụ, lizin ứng với 2 bộ ba AAA, AAG, một số axit amin được mã hoá bởi nhiều bộ ba như alanin ứng với 4 bộ ba, lơxin ứng với 6 bộ ba.
3. Những đặc điểm cơ bản của mã di truyền
- Mã di truyền được đọc theo một chiều 5’ – 3’ trên phân tử mARN.
- Mã di truyền được đọc liên tục theo từng cụm 3 nucleotit, các bộ ba không đọc gối lên nhau.
- Mã di truyền là đặc hiệu không một bộ ba nào mã hoá đồng thời 2 hoặc một số axit amin khác nhau.
- Mã di truyền có tính thoái hoá có nghĩa là mỗi axit amin được mã hoá bởi một số bộ ba khác loại trừ metionin, Triptophan chỉ được mã hoá bởi một bộ ba). Nhờ đó mà gen đảm bảo được thông tin di truyền và xác nhận trong bộ ba 2 nucleotit đầu là quan trọng còn nucleotit thứ 3 có thể linh hoạt . Sự linh hoạt này có thể không gây hậu quả gì. Nhưng cũng có thể gây nên sự lắp ráp nhầm các axit amin trong chuỗi polipeptit.
- Mã di truyền có tính phổ biến: tất cả các loài sinh vật đều được mã hoá theo một nguyên tắc chung (các từ mã giống nhau), Điều này phản ảnh nguồn gốc chung của các loài.
- Mã di truyền có mã mở đầu, có mã kéo dài chuỗi polipeptit và mã kết thúc
VII. ĐỘT BIẾN GEN
1. Khái niệm Đột biến gen là những biến đổi trong cấu trúc phân tử của gen liên quan tới một hay một số cặp nucleotit xảy ra tại một điểm nào đó của phân tử ADN biểu hiện ở các dạng: mất, thêm, thay thế 1 cặp nucleotit.
2. Nguyên nhân và cơ chế a) Nguyên nhân
- Đột biến gen phát sinh do tác nhân gây đột biến lí hoá trong ngoại cảnh hoặc rối loạn trong các quá trình sinh lí, hoá sinh của tế bào gây nên những sai sót trong quá trình tự sao của ADN hoặc trực tiếp biến đổi cấu trúc của nó.
- Đột biến gen phụ thuộc vào loại tác nhân, liều lượng, cường độ của tác nhân, đặc điểm cấu trúc của gen.
b) Cơ chế
Sự biến đổi của một nucleotit nào đó thoạt đầu xảy ra trên một mạch của ADN dưới dạng tiền đột biến. Lúc này enzim sửa chữa có thể sửa sai làm cho tiền đột biến trở lại dạng ban đầu. Nếu sai sót không được sửa chữa thì qua lần tự sao tiếp theo nucleotit lắp sai sẽ liên kết với nucleotit bổ sung với nó làm phát sinh đột biến gen.
3.Sự biểu hiện của đột biến gen
- Đột biến gen khi đã phát sinh sẽ được tái bản cùng với sự tái bản của phân tử ADN mang đột biến. - Nếu đột biến phát sinh trong giảm phân sẽ tạo đột biến giao tử qua thụ tinh đi vào hợp tử Đột biến trội sẽ biểu hiện ngay ở kiểu hình của cơ thể mang đột biến. Đột biến lặn sẽ đi vào hợp tử ở dạng dị hợp qua giao phối lan truyền dần trong quần thể, trải qua nhiều thế hệ được nhân lên ngày một nhiều, tới một thời điểm nào đó các đột biến lặn trong các giao tử gặp gỡ nhau trong giao phối, hình thành tổ hợp đồng tử lặn, lúc này kiểu hình đột biến lặn mới xuất hiện.
- Khi đột biến xảy ra trong nguyên phân, chúng sẽ phát sinh ở một tế bào sinh dưỡng rồi được nhân lên trong một mô. Nếu là đột biến trội sẽ biểu hiện ở một phần của cơ thể, tạo nên thể khảm.
- Đột biến soma có thể di truyền bằng sinh sản sinh dưỡng nhưng không thể di truyền qua sinh sản hữu tính.
- Đột biến cấu trúc của gen đòi hỏi một số điều kiện mới biểu hiện trên kiểu hình của cơ thể. Vì vậy cần phải phân biệt đột biến là những biến đổi trong vật chất di truyền với thể đột biến là những cá thể mang đột biến đã biểu hiện ở kiểu hình
4. Hậu quả của đột biến gen
- Sự biến đổi trong dãy nucleotit của gen dẫn đến biến đổi trong dãy ribonucleotit của mARN làm biến đổi dãy axit amin của protein tương ứng. Cuối cùng biểu hiện thành một biến đổi đột ngột, gián đoạn về một hoặc một số tính trạng nào đó trên một hoặc một số ít cá thể trong quần thể - Đột biến gen gây rối loạn trong quá trình sinh tổng hợp protein, đặc biệt là đột biến ở các gen quy định cấu trúc của các enzim nên đa số đột biến thường có hại cho cơ thể, cũng có nhứng đột biến gen trung tính, một số đột biến lại có lợi.
B – CƠ SỞ VẬT CHẤT CỦA HIỆN TƯỢNG DI TRUYỀN VÀ BIẾN DỊỞ CẤP ĐỘ TẾ BÀO
I. CẤU TRÚC VÀ TÍNH ĐẶC TRƯNG CỦA NST
1. Khái niệm NST
NST là thể vật chất di truyền tồn tại trong nhân tế bào bị bắt màu bằng thuốc nhuộm kiềm tính, có số lượng, hình dạng, kích thước, cấu trúc đặc trưng: NST có khả năng tự nhân đôi, phân li, tổ hợp ổn định qua các thế hệ.
2.Cấu trúc của NST
- Ở virut, thể ăn khuẩn, NST chỉ là một phân tử ADN trần. Ở sinh vật có nhân, NST cấu
trúc phức tạp.
- Sau khi nhân đôi mỗi NST có 2 cromatit, mỗi cromatit có 1 sợi phân tử ADN mà có một nửa nguyên liệu cũ và một nửa nguyên liệu mới được lấy từ môi trường tế bào. Các cromatit này đóng xoắn cực đại ở kì giữa nên chúng có hình dạng và kích thước đặc trưng. Mỗi NST có 2 cromatit đính nhau ở tâm động tại eo thứ nhất. Một số NST còn có eo thứ 2 là nơi tổng hợp rARN. Các rARN tích tụ lại tạo nên nhân con. Lúc bước vào phân bào, NST ngừng hoạt động, nhân con lại tái hiện.
- NST có nhiều hình dạng khác nhau: hình hạt, hình que, hình chữ V, hình móc. Ở một số loài sinh vật trong vòng đời có trải qua giai đoạn ấu trùng có xuất hiện các NST với kích thước lớn hàng nghìn lần gọi là NST khổng lồ (như ở ấu trùng ruồi giấm và các loài thuộc bộ 2 cánh). Điển hình là NST có hình chữ V với 2 cánh kích thước bằng nhau hoặc khác nhau. Chiều dài của NST từ 0,2 đến 50 μm, chiều ngang 0,2 đến 2 μm.
- NST được cấu tạo bởi ADN và protein. Phân tử ADN quấn quanh khối cầu protein tạo nên nucleoxom. Mỗi nucleoxom gồm 8 phân tử protein histon tạo nên khối hình cầu dẹt phía ngoài được bao bọc bởi 1 3 4 vòn xoắn ADN khoảng 146 cặp nucleotit. Các nucleoxom nối với nhau bằng các đoạn ADN và một protein histon H1. Mỗi đoạn có khoảng 15 – 100 cặp nucleotit. Tổ hợp ADN với histon trong chuỗi nucleoxom tạo thành sợi cơ bản có chiều ngang 100 Ǻ, sợi cơ bản cuộn xoắn thứ cấp tạo nên sợi nhiễm sắc có chiều ngang 250 –
300 Ǻ. Sợi nhiễm sắc tiếp tục đóng xoắn tạo nên ống rỗng với bề ngang 2000 Ǻ, cuối cùng hình thành cromatit có đường kính tới 6000Ǻ.
- Do có cấu trúc xoắn cuộn như vậy nên chiều dài của NST đã được rút ngắn 15000 –
20000 lần so với chiều dài phân tử ADN, NST dài nhất của người chứa phân tử ADN dài 82 mm, sau khi xoắn cực đại ở kì giữa chỉ dài 10 μm. Sự thu gọn cấu trúc không gian như thế thuận lợi cho sự phân li, tổ hợp các NST trong chu kì phân bào.
3. Tính đặc trưng của NST
Mỗi loài sinh vật đều có bộ NST đặc trưng:
- Đặc trưng về số lượng, hình dạng, kích thước và cáu trúc. Ở những loài giao phối, tế bào sinh dưỡng mang bộ NST lưỡng bội (2n), NST tồn tại thành cặp tương đồng, trong đó một NST có nguồn gốc từ bố, một NST có nguồn gốc từ mẹ. Tế bào giao tử chứa bộ NST đơn bội.
Ví dụ: Ở người 2n = 46, n = 23; Ở ngô 2n = 20, n = 10; Ở lúa 2n = 24, n = 12;
Ở đậu Hà Lan 2n = 14, n = 7…
- Đặc trưng bởi số lượng, thành phần, trình tự phân bố các gen trên mỗi NST.
- Đặc trưng bởi các tập tính hoạt động của NST tái sinh, phân li, tổ hợp, trao đổi đoạn, đột biến về số lượng, cấu trúc NST.
II. CƠ CHẾ HÌNH THÀNH CÁC LOẠI BỘ NST TỪ TẾ BÀO 2n
1. Cơ chế hình thành bộ NST n
- Một nhóm tế bào sinh dưỡng ở các cơ thể trưởng thành được tách ra làm nhiệm vụ sinh sản, gọi là tế bào sinh dục sơ khai. Các tế bào này lần lượt trải qua 3 giai đoạn:
+ Giai đoạn sinh sản: nguyên phân liên tiếp nhiều đợt tạo ra các tế bào sinh dục con.
+ Giai đoạn sinh trưởng: các tế bào tiếp nhận nguyên liệu từ môi trường ngoài để tạo nên các tế bào có kích thước lớn (kể cả nhân và tế bào chất).
+ Giai đoạn chín: các tế bào sinh tinh trùng, sinh trứng bước vào giảm phân gồm 2 lần phân bào liên tiếp để tạo ra các giao tử đơn bội.
+ Giai đoạn sau chín: ở thực vật khi kết thúc giảm phân mỗi tế bào đơn bội hình thành từ tế bào sinh dục đực tiếp tục nguyên phân 2 đợt tạo ra 3 tế bào đơn bội hình thành hạt phấn chín. Mỗi tế bào đơn bội ở mỗi tế bào sinh dục cái lại nguyên phân 3 đợt tạo ra 8 tế bào đơn bội hình thành noãn.
- Giảm phân I:
+ Ở kì trung gian ADN nhân đôi, mỗi cặp NST tương đồng nhân đôi thành cặp NST tương đồng kép.
+ Ở kì trước I: NST tiếp tục xoắn lại, kì này tại một số cặp NST tương đồng có xảy ra trao đổi đoạn giưuã 2 cromatit khác nguồn gốc. Cuối kì trước I, màng nhân biến mất, thoi tơ vô sắc bắt đầu hình thành.
+ Ở kì giữa I: thoi tơ vô sắc hình thành xong. Các NST tương đồng kép tập trung thành cặp trên mặt phẳng xích đạo và nối với thoi tơ vô sắc tại tâm động theo nhiều kiểu sắp xếp.
+ Ở kì sau I: mỗi NST ở dạng kép trong cặp tương đồng kép phân li về 2 cực tế bào, hình thành các tế bào có bộ NST đơn ở trạng thái kép.
+ Ở kì cuối I: tạo 2 tế bào con chứa bộ NST đơn ở trạng thái kép, khác nhau về nguồn gốc, chất lượng NST.
- Giảm phân II: ở lần này, kì trung gian trải qua rất ngắn ở kì giữa II, các NST đơn ở trạng thái kép trong mỗi tế bào tập trung trên mặt phẳng xích đạo nối với thoi tơ vô sắc. Kì sau II, mỗi cromatit trong mỗi NST đơn ở trạng thái kép phân li về 2 cực. Kì cuối II tạo ra các tế bào đơn bội. Từ một tế bào sinh tinh trùng tạo ra 4 tinh trùng, từ 1 tế bào sinh trứng tạo ra 1 trứng và 3 thể định hướng.
2. Cơ chế hình thành bộ NST 2n
- Qua nguyên phân:
+ Ở kì trung gian: mỗi NST đơn tháo xoắn cực đại ở dạng sợi mảnh, ADN nhân đôi để tạo ra các NST kép.
+ Kì trước: NST xoắn lại, cuối kì trước màng nhân mất, thoi vô sắc bắt đầu hình thành.
+ Kì giữa: thoi vô sắc hình thành xong, NST kép tập trung trên mặt phẳng xích đạo nối với dây tơ vô sắc tại tâm động.
+ Kì sau: mỗi cromatit trong từng NST kép tách nhau qua tâm động phân chia về 2 cực tế bào.
+ Kì cuối: các NST đơn giãn xoắn cực đại, màng nhân hình thành, mỗi tế bào chứa bộ NST lưỡng bội (2n)
- Qua giảm phân không bình thường:
Các tế bào sinh tinh trùng hoặc sinh trứng nếu bị tác động của các nhân tố phóng xạ, hoá học… làm cắt đứt thoi tơ vô sắc hoặc ức chế hình thình thoi tơ vô sắc trên toàn bộ bộ NST sẽ tạo nên các giao tử lưỡng bội.
- Qua cơ chế thụ tinh:
Sự kết hợp giữa tinh rùng đơn bội và trứng đơn bội qua thụ tinh sẽ tạo nên hợp tử lưỡng bội (2n).
3. Cơ chế hình thành bộ NST 3n, 4n
- Tế bào 2n giảm phân do rối loạn phân bào (thoi tơ vô sắc bị cắt hoặc được hình thành)
xảy ra trên tất cả các cặp NST sẽ tạo nên giao tử 2n. Giao tử này kết hợp với giao tử bình
thường n sẽ tạo nên hợp tử 3n.
- Các giao tử không bình thường 2n kết hợp với nhau sẽ tạo nên hợp tử 4n.
- Ngoài ra dạng 3n còn được hình thành trong cơ chế thụ tinh kép (ở thực vật) do nhân
thứu cấp 2n kết hợp với một tinh tử n trong hạt phấn chín tạo nên nội nhũ 3n.
- Dạng tế bào 4n, còn được hình thành do nguyên phân rối loạn xảy ra trên tất cả các cặp NST sau khi nhân đôi.
III. Ý NGHĨA SINH HỌC VÀ MỐI LIÊN QUAN GIỮA NGUYÊN PHÂN, GIẢM PHÂN, THỤ TINH.
1. Ý nghĩa sinh học của nguyên phân, giảm phân và thụ tinh
a) ý nghĩa của nguyên phân và giảm phân
- Nguyên phân: ổn định bộ NST qua các thế hệ tế bào của cùng một cơ thể, tăng nhanh
sinh khối tế bào đảm bảo phân hoá mô, cơ quan tạo ra cơ thể
- Giảm phân: đảm bảo sự kết tục vật chất di truyền ổn định tương đối qua các thế hệ
b) Ý nghĩa của thụ tinh
Phục hồi lại bộ NST lưỡng bội do sự kết hợp giữa giao tử đực (n) với giao tử cái (n).
Mặt khác trong thụ tinh do sự phối hợp ngẫu nhiên của các loại giao tử khác giới tính mà
cũng tạo nên nhiều kiểu hợp tử khác nhau về nguồn gốc và chất lượng bộ NST làm tăng tần số các loại biến dị tổ hợp.
2. Mối liên quan giữa nguyên phân, giảm phân và thụ tinh trong quá trình truyền đạt thông tin di truyền
- Nhờ nguyên phân mà các thế hệ tế bào khác nhau vẫn chứa đựng các thông tin di truyền giống nhau, đặc trưng cho loài.
- Nhờ giảm phân mà tạo nên các giao tử đơn bội để khi thụ tinh sẽ khôi phục lại trạng thái lưỡng bội.
- Nhờ thụ tinh đã kết hợp bộ NST đơn bội trong tinh trùng với bộ NST đơn bội trong trứng để hình thành bộ NST 2n, đảm bảo việc truyền thông tin di truyền từ bố mẹ cho con cái ổn định tương đối.
- Nhờ sự kết hợp 3 quá trình trên mà tạo điều kiện cho các đột biến có thể lan rộng chậm chạp trong loài để có dịp biểu hiện thành kiểu hình đột biến.
IV. ĐỘT BIẾN CẤU TRÚC NST
1. Khái niệm Đột biến cấu trúc NST là những biến đổi đột ngột trong cấu trúc của NST do tác nhân gây đột biến làm thay đổi cấu trúc NST tạo ra những tính trạng mới.
2. Nguyên nhân
Do tác nhân gây đột biến lí hoá trong môi trường hoặc những biến đổi sinh lí nội bào làm phá vỡ cấu trúc NST ảnh hưởng tới quá trình tái bản, tiếp hợp, trao đổi chéo của NST.
3. Cơ chế và hậu quả
Đột biến cấu trúc NST gồm các dạng : mất đoạn, lặp đoạn, đảo đoạn, chuyển đoạn.
- Mất đoạn: Một đoạn NST bị đứt ra làm giảm số lượng gen trên NST. Đoạn bị mất có thể ở phía ngoài hoặc phía trong của cánh. Đột mất đoạn thường giảm sức sống hoặc gây chết. Ví dụ, mất đoạn cặp 21 ở người gây ung thư máu.
- Lặp đoạn: Một đoạn NST nào đó được lặp lại một lần hay nhiều lần làm tăng số lượng gen cùng loại. Đột biến lặp đoạn có thể do đoạn NST bị đứt được nối xen vào NST tương đồng hoặc do NSt tiếp hợp không bình thường, do trao đổi chéo không đều giữa các
cromatit. Đột biến lặp đoạn có thể làm tăng cường hay giảm sút sức biểu hiện tính trạng. Ví dụ, lặp đoạn 16A ở ruồi giấm làm cho mắt hình cầu thành mắt dẹt, càng lặp nhiều đoạn mắt càng dẹt. - Đảo đoạn: Một đoạn NST bị đứt rồi quay ngược lại 180° và gắn vào chỗ bị đứt làm thay đổi trật tự phân bố gen trên NST. Đoạn bị đảo ngược có thể mang tâm động hoặc không. Đột biến đảo đoạn NST ít ảnh hưởng tới sức sống của cơ thể vì vật chất di truyền không bị mất đi. Sự đảo đoạn NST tạo nên sự đa dạng giữa các nòi trong phạm vi một loài.
- Chuyển đoạn: Một đoạn NST này bị dứt ra và gắn vào một NST khác hoặc cả 2 NST khác cặp cùng đứt một đoạn nào đó rồi lại trao đổi đoạn bị đứt với nhau, các đoạn trao đổi có thể tương đồng hoặc không tương đồng. Như vậy có thể thấy có 2 kiểu chuyển đoạn là chuyển đoạn tương hỗ và chuyển đoạn không tương hỗ. sự chuyển đoạn làm phân bố lại các gen trong phạm vi một cặp NST hay giữa các NST khác nhau tạo ra nhóm gen liên kết mới. Chuyển đoạn lớn thường gây chết hoặc làm mất khả năng sinh sản. Người ta gặp sự chuyển đoạn nhỏ ở đầu lúa, chuối, đậu trong thiên nhiên. Trong thực nghiệm người ta đã chuyển gen cố định nitơ của vi khuẩn vào hệ gen hướng hương tạo ra giống hướng hương có nitơ cao trong dầu.
V. ĐỘT BIẾN SỐ LƯỢNG NST
1. Khái niệm Đột biến số lượng NST là hiện tượng bộ NST của loài tăng lên một số nguyên lần bộ đơn bội (tạo thể đa bội) hoặc tăng lên hay giảm đi một hay một số cặp NST sẽ tạo nên thể dị bội.
2. Thể dị bội Thể dị bội gồm có: thể ba nhiễm, thể đa nhiễm, thể một nhiễm, thể khuyết nhiễm. Các đột biến dị bội đa phần gây nên hậu quả có hại ở động vật. Ví dụ, ở người có 3 NST 21, xuất hiện hội chứng Đao, tuổi sinh đẻ người mẹ càng cao tỉ lệ mắc hội chứng Đao càng nhiều Ngoài ra, còn gặp hội chứng XXX, XO, XXY, OY đều gây nên hậu quả có hại.
3. Thể đa bội
Có 2 dạng đa bội : đa bội chẵn và đa bội lẻ
- Đa bội chẵn được hình thành bằng cơ chế nguyên phân rối loạn trên toàn bộ bộ NST 2n sẽ tạo nên dạng 4n, hoặc do kết hợp giữa 2 loại giao tử lưỡng bội không bình thường với nhau.
- Đa bội lẻ được hình thành là do sự kết hợp giữa giao tử 2n không bình thường với giao tử n hình thành thể đa bội lẻ 3n.
- Cơ thể đa bội có hàm lượng ADN tăng gấp bội dẫn tới trao đổi chất tăng cường, cơ thể đa bội tế bào kích thước lớn, cơ quan sinh dưỡng, sinh sản to, chống chịu tốt với điều kiện bất lợi của môi trường.
- Cơ thể đa bội lẻ không có khả năng sinh sản hữu tính vì quá trình giảm phân bị trở ngại. Muốn duy trì phải nhân bằng con đường sinh sản sinh dưỡng.
- Thể đa bội khá phổ biến ở thực vật, ở động vật giao phối thường rất ít gặp.
HỆ THỐNG CÂU HỎI ÔN TẬP
Câu 1. Tại sao nói ADN là cơ sở vật chất và cơ chế di truyền ở cấp độ phân tử? Tính đặc trung và đa dạng của ADN được thể hiện ở chỗ nào?
Câu 2: Sự giống nhau và khác nhau cơ bản về cấu trúc giữa ADN và mARN ở những điểm nào?
Câu 3: ADN và protein giống và khác nhau về cấu trúc ở những điểm nào? Những chức năng cơ bản của prôtêin ?
- 14 -
Câu 4: Điểm giống nhau va khác nhau giữa cơ chế tổng hợp ADN ở sinh vật nhân chuẩn với E.coli. Ý nghĩa của cơ chế tổng hợp ADN
Câu 5: Mối quan hệ giữa ADN và prôtêin trong cấu trúc và cơ chế di truyền.Tính đặc trưng cuẩ prôtêin do yếu tố nào quy định?
Câu 6: Tại sao nói đột biến gen thường có hại, ít có lợi, tần số thấp, nhưng lại là nguồn nguyên liệu chủ yếu của chon lọc tự nhiên?.
Câu 7: Điểm giống và khác nhau cơ bản giữa nguyên phân và giảm phân. Ý nghĩa của chúng trong di truyền và tiến hoá..
Câu 8: Trình bày cơ chế hình thành các dạng tế bào n, 2n, 3n, 4n từ dạng tế bào 2n. Ý nghĩa của việc hình thành các loại tế bào nói trên
Câu 9: Các cơ chế sinh học xảy ra như thế nào đối với 1 cặp NST tương đồng ở cấp độ tế bào?.
Câu10. Cấu trúc và chức năng của NST thường và NST giới tính giống nhau và khác nhau ở điểm nào?
Câu11. Trình bày các loại biến dị làm thay đổi về số lượng, cấu trúc NST?
Câu12. Điểm giống và khác nhau cơ bản giữa biến dị tổ hợp và biến dị đột biến. Vai trò của các loại biến dị đó trong tiến hoá và chọn giống.
CÁC BÀI TẬP RÈN KỸ NĂNG
Bài 1. Một gen có chiều dài 5100 Ăngstron. Hiệu số phần trăm giữa adenin với một loại nucleotit khác bằng 10% số nucleotit của gen. Trên phân tử mARN được tổng hợp từ gen đó có 10% uraxin. một mạch đơn của gen có 16% xitozin, số timin bằng 150 nucleotit.
a) Tỉ lệ phần trăm và số lượng từng loại nucleotit của gen.
b) Tỉ lệ phần trăm và số lượng từng loại ribonucleotit của một phân tử mARN bằng bao nhiêu?
c) Nếu gen đó sao mã 6 lần và trên mỗi phân tử mARN có 10 riboxom trượt qua không lặp lại thì số lượng axit amin mà môi trường nội bào cung cấp cho quá trình tổng hợp protein là bao nhiêu?
d) Nếu thời gian giải mã một axit amin la 0,1 giây, thời gian tiếp xúc của một phân tử mARN với các riboxom là 58,1 giây, khoảng cách giữa các riboxom kế tiếp khoảng bao nhiêu Ăngstron?
Bài 2: Một gen dài 5100 Ắngtron. Khi gen tự sao liên tiếp hai đợt, môi trường nội bào đã cung cấp 2700 ađênin. Phân tử mARN được tổng hợp từ gen đó có 600 adenin và 240 guanin. vận tốc giải mã là 10 axit amin/ giây. Tính từ lúc ribôxôm thứ nhất trượt qua phân tử mARN cho đến khi hết phân tử mARN đó là 55,6 giây.
a) Xác định số lượng từng loại nuclêôtit ởtrong toàn bộ các gen được hình thành sau hai đợt tự sao liên tiếp
b) Xác định số lượng từng loại nuclêôtit ở mỗi mạch đơn của gen
c) Tính khoảng cách theo ăngtron giữa ribôxôm thứ nhất với ribôxôm cuối cùng khi chúng đang tham gia giải mã trên một phân tử mARN
Bài 3: mạch thứ nhất của gen có tỉ lệ các nuclêôtit là 10% adenine, 20% timin và 25% xitozin. Phân tử mARN được sao từ gen đó có 20% uraxin.
a) Tính tỉ lệ từng loại nuclêôtit của gen và từng loại ribônuclêôtit của phân tử mARN
b) Nếu gen đó dài 0,306 micromet thì nó chứa boa nhiêu liên kết hidro?
c) Một phân tử mARN sinh ra từ gen có chiều dài nói trên và có một số riboxom cùng hoạt động trong quá trình giải mã, thời gian riboxom thứ nhất trượt qua hết phân tử là 30
giây và thời gian tính từ lúc bắt đầu có sự giải mã đến khi riboxom cuối cùng trượt qua hết phân tử mARN đó là 35,4 giây. Hỏi có bao nhiêu riboxom tham gia vào quá trình giải mã?
Biết rằng các riboxom cách đều nhau một khoảng bằng 61,2Ǻ
Bài 4: Mạch thứ nhất của gen có 240 timin, hiệu số giữa guanin với adenine bằng 10% số nucleotit của mạch. Ở mạch thứ hai, hiệu số giữa adenin và xitozin bằng 20% số nucleotit của mạch. Khi gen đó tổng hợp phân tủe mARN thì mội trường nội bào đã cung cấp 360 uraxin.
a) Tỉ lệ phần trăm và số lượng từng loại nucleotit của gen và của từng mạch là bao nhiêu?
b) Hãy tính chiều dài của phân tử mARN, tỉ lệ phần trăm và số lượng mỗi loại ribonucleotit của nó.
c) Trên mỗi phân tử mARN có 8 riboxom cùng giải mã, tính từ lúc riboxom bắt đâu ftrượt trên phân tử mARN thì thời gian để riboxom thứ nhất trượt qua hết sphân tử là 20 giây, còn riboxom cuói cùng thì phải cần đén 26,3 giây mới hoàn tất việc giải mã. khoảng cách đều giữa các riboxom là bao nhiêu Ắngtron? Biết rang các riboxom trượt với vận tốc bằng nhau
Bài 5: Trên một phân tử mARN có một số riboxom trượt qua với khoảng cách đều bằng nhau. Riboxom thứ nhất trượt qua hết phân tử mARN đó hết 50 giây. Tính từ lúc riboxom thứ nhất trượt qua và tiếp xúc với phân tử mARN đó thì riboxom cuối cùng ppjải mất 57,2 giây mới hoàn thành việc đi qua phân tử mARN . Biết rằng phân tử prôtêin thứ hai được tổng hợp chậm hơn phân tử prôtêin thứ nhất 0,9 giây.
Gen điều khiển việc tổng hợp các phân tử prôtêin nói trên có mạch 1 chứa 10% adeini và 20% guanine, mạch 2 chứa 15% adenine. Quá trình sao mã của gen đã đòi hỏi mội trường nội bào cung cấp 150 uraxin và 155 adenin để góp phần tổng hợp một phân tử mARN
a) Tính chiều dài của gen
b) Tính số lượng từng loại ribônuclêôtit của một phân tử mARN
c) Số riboxom đã tham gia vào qua trình giải mã trên một phân tử mARN đó là bao nhiêu? Biết rằng mỗi riboxom chỉ trượt qua một lần.
d) Khoảng cách giữa 2 riboxom kế tiếp và khoảng cách giữa riboxom thứ nhất với riboxom cuối cùng tính theo ăngtron là bao nhiêu?
e) Toàn bộ quá trình giải mã nới trên đã cần bao nhiêu axit amin của mội trường nội bào và trong tất cả các prôtêin hoàn chỉnh chứa bao nhiêu axit amin?
Bài 6: Ở một loài một tế bào sinh dục 2n thực hiện sự nguyên phân liên tiếp một số lần, đòi hỏi môi trường nội bào cung cấp nguyên kiệu để hình thành nên 4826 nhiễm sắc thể đơn mới. các tế bào con sinh ra từ lần nguyên phân cuối cùng đều giảm nhiễm bình thường cho 256 tinh trùng chưa nhiễm sắc thể giới tính Y
a) Bộ nhiễm sắc thể lưỡng bội của loài ? số lần nguyên phân liên tiếp của tế bào sinh dục 2n đầu tiên ? để tạo ra các tế bào con 2n đã có bao nhiêu thoi dây tơ vô sắc được hình thành trong các lần nguyên phân ấy ?
b) Nếu có 3 tinh trùng được thụ tinh với 3 trứng khác nhau tạo ra các hợp tử thì có bao nhiêu cromait trong các tế bào sinh dục cái sinh ra các trứng đó, vào lúc mà các tế bào bắt đầu thực hiện sự phân bào giảm nhiễm? Các hợp tử vừa được hình thành có bao nhiêu NST đơn?
c) Giả thiết rằng sự giảm phân xảy ra trong các tế bào sinh dục của cá thể cái, mỗi cặp tương đồng đều gồm hai nhiễm sắc thê cấu trúc khác nhau, mỗi cặp nhiễm sắc thể tương đồng đều gồm hai nhiễm sắc thể cấu trúc khác nhau, sự trao đổi đoạn chỉ xảy ra một cặp
nhiễm sắc thể thường,sự
đột biến dị bội chỉ xảy ra ở cặp nhiễm sắc thể giới tính. Khả năng
cá thể cái có thể cho bao nhiêu loại trứng?
Bài 7: giả thiết trong các cặp nhiễm sắc thể tương đông của một bộ nhiễm ssắc thể lưỡng
bội đều chứa cá cặp gen dị hợp tử và hiện tượng trao đổi đoạn tại một điểm chỉ xảy ra với một cặp nhiễm sắc thể tương đồng. Cho biết không có hiện tượng đột biến và số loại giao tử
đực sinh ra từ các điều kiên trên là 32.
Giả thiết trung bình mỗi kì trong phân bào nguyên phân hết 5 phút, hai lần phân bào (kì trung gian) hết 10 phút, quá trình nguyên phân diễn ra liên tục, các tế bào con sinh ra đều tiếp tục nguyên phân.
a) Bộ nhiễm sắc thể lưỡng bội nói trên ở trạng thái chưa nhân đôi là bao nhiêu ?
b) Để hợp tử thực hiện được quả trình nguyên phân thì môi trường nội bào đã cung cấp nguyên kiệu tương đương với bao nhiêu NST đơn vào các thời điểm:
- Kết thúc 20 phút
- Kết thúc 32 phút
- Kết thúc 100 phút
Biết rằng khi hợp tử bước vào kì trước được tính là thời gian bắt đầu
Bài 8: Ở gà khi quan sát một tế bào sinh dục đực đang ở kì giữa của nguyên phân, người ta đếm được 78 nhiễm sắc thể kép
a) Tế bào đó nguyên phân 5 đợt liên tiếp đã đòi hỏi mội trường cung cấp nguyên liệu để tạo ra bao nhiêu nhiễm sắc thể đơn mới ?
b) Loại tế bào này giảm phân bình thường, khả năng nhiều nhất có thể cho bao nhiêu loại tinh trùng trong trường hợp không có hiện tượng trao đổi đoạn giữa các nhiễm sắc thể kép trong cặp tương đồng? Điều kiện để cho số loại tinh trùng nhiều nhất là gì?
c) Giả thiết rằng có 1000 tế bào sinh tinh trùng giảm phân bình thường, hiệu suất thụ tinh của tinh trùng là 1/1000, còn của trứng là 20%, mỗi tinh trùng chỉ thụ tinh với một trứng. Xác định số tế bào sinh trứng.
d) Các hợp tử được tạo thành đã nguyên phân liên tiếp nhiều đợt với số lần bằng nhau, môi truờng nội bào đã cung cấp nguyên kiệu để tạo ra 2184 nhiễm sắc thể đơn mới. Xác
định số tế bào con sinh ra và số đợt nguyên phân của mỗi hợp tử.
Bài 9: Một tế bào sinh duc đực 2n và một tế bào sinh dục cái 2n đều nguyên phân một số đợt bằng nhau (các tế bào con sinh ra đều tiếp tục nguyên phân). Giả thiết rằng các tế bào con sinh ra từ đợt nguyên phân cuối cùng đều giảm nhiễm cho tổng số 80 giao tử binh thường. Cho biết số lượng nhiễm sắc thể đơn trong các giao tử đực nhiều hơn số lượng nhiễm sắc thể đơn trong các giao tử cái là 192.
1. Loài đó tên là gì?
2. Mô tả hình dạng và số lượng bộ nhiễm sắc thể lưỡng bộ trong loài đó Bài 10: Một gà mái đẻ đươc một số trứng, nhưng khi ấp chỉ có 12 trứng nở thành gà con. Các hợp tử nở thành gà con có 936 nhiễm sắc thể đơn ở trạng thái chưa tự nhân đôi. Số trứng còn lại không nở thành gà con. Số tinh trùng được sinh ra phục vụ cho gà giao phối có 624000 nhiễm sắc thể đơn. Giả thiết số tinh trùng được trực tiếp thụ tinh với các trứng nói trên chiếm tỉ lệ 1/1000 so với tổng số tinh trùng được hình thành. Mỗi tinh trùng thụ tinh với 1trứng
a) Số trứng được thụ tinh
b) Trứng gà không nở thành gà con có bộ nhiễm sắc thể như thế nào?
c) Số gà trống và gà mái trong đàn gà con nói trên có tuân theo tỉ lệ 1: 1 không?
GỢI Ý ĐÁP ÁN CHO CÁC CÂU HỎI VÀ BÀI TẬP TỪNG PHẦN
A- PHẦN : CÂU HỎI
Câu1. Tại sao nói ADN là cơ sở vật chất và cơ chế di truyền ở cấp độ phân tử ? Tính đặc trung và đa dạng của ADN được thể hiện ở chỗ nào?
Trả lời 1. ADN là cơ sở vật chất vầ cơ chế di truyền ở cấp độ phân tử :
- ADN là thành phần chính của nhiễm sắc thể (NST), mà nhiễm sắc htể là cơ sở vật chất của tính di truyền ở cấp độ tế bào, vì vậy ADN là cấp độ di truyền ở cấp độ phân tử.
- ADN chứa thông tin di truyền đặc trưng cho mỗi loài ở số lượng, thành phần và trật tự sắp xếp các nuclêôtit.
- ADN có khả năng tự nhân đôi, đảm bảo cho NST hình thành quá trình nguyên phân, giảm phân diễn ra bình thường, thong tin di truyền của loài được ổn định ở cấp độ tế bào và cấp độ phân tử.
- ADN chứa các gen, mỗi gen thực hiên một chức năng di truyền khác nhau thông qua cơ chế dịch mã và phiên mã.
- ADN có khả năng đột biến về cấu trúc: mất, thêm, thay thế, đảo vị trí các nuclêôtit tạo nên các alen mới.
- Nhiều bằng chứng đã chứng minh vai trò mang thông tin di truyền của axit nuclêôtit:
+ Khả năng hấp thụ tia ngoại tử cực đại ở bước sóng 260 nm.
+ Thí nghiệm biến nạp của F.Griffith (1928), của O.T.Avery, C.M.Macleod…(1994) và Fraenket-Conrat, Singer (1957) đã chứng minh được axit nuclêôtit là vật chất mang thông tin di truyền.
2. Tính đặc trưng và tính đa dạng của ADN
- Đặc trưng bởi số lượng, thành phần và trình tự sắp xếp các nuclêôtit. Vì vậy từ 4 loại nuclêôtit tạo nên tính đặc trưng và tính đa dạng của ADN.
- Đặc trưng bởi tỉ lệ A+T/G+X cho mỗi loài.
Đặc trưng bởi số lượng, thành phần và trình tự phân bố các gen trên phân tử ADN. Câu 2: Sự giống nhau và khác nhau cơ bản về cấu trúc giữa ADN và mARN ?
Trả lời
1. Điểm giống nhau về cấu trúc ADN và mARN
- Đều cấu tạo theo nguyên tắc đa phân.
- Mỗi đơn phân đều gồm 3 thành phần cơ bản trong đó thành phần quan trọng nhất là bazơnitric.
- Trên mạch đơn ADN và trên phân tử mARN các đơn phân được liên kết với nhau bởi liên kết hoá trị bền vững.
- Đều có cấu tạo xoắn.
- Đặc trưng bởi số lượng, thành phẩn và trật tự phân bố các đơn phân.
2. Điểm khác nhau giữa cấu trúc ADN và mARN
ADN mARN
- 18 -
- Đại phân tử có kích thước và khối lượng
rất lớn
- Có cấu trúc mạch kép
- Xây dựng từ 4 loại nuclêôtit
- Có bazơnitric Timin
- Trong mỗi nuclêôtit có đường đêôxiribôza
(C5H10O4)
- Liên kết hoá trị trên mạch đơn của ADN là liên kết giữa đường C5H10O4 của nuclêôtit với phân tử H3PO4 của nuclêôtit bên cạnh. Nhiều liên kết hoá trị tạo nên chuỗi pôlinuclêôtit.
- Đa phân tử có khối lượng và kích thước rất
bé.
- Có cấu trúc mạch đơn
- Xây dựng từ 4 loại ribônuclêôtit
- Có bazơnitric U là dẫn xuất của T
- Trong mỗi ribônuclêôtit có đường ribôza
(C5 H10 O5)
- Liên kết hoá trị trên mạch mARN là liên
kết được hình thành giữa đường C5H10 O5
của ribônuclêôtit này với phân tử H3 PO4
của ribônuclêôtit bên cạnh. nhiều liên kết hoá trị tạo nên chuỗi pôliribônuclêôtit.
bằng 3 liên kết hiđrô, và ngược lại tạo nên
cấu trúc ADN chiều rộng khoảng 20Ǻ, khoảng cách mỗi bậc thang bằng 3,4Ǻ. Mỗi chu kì xoắn gồm 10 cặp nuclêôtit có chiều cao 34Ǻ (xoắn phải)
Câu 3: ADN và protein giống và khác nhau về cấu trúc ở những điểm nào? Những chức năng cơ bản của prôtêin ?
Trả lời
1. Điểm giống nhau về cấu trúc giữa ADN và prôtêin
• Đều được cấu tạo theo nguyên tắc đa phân
• Đều được cấu tạo từ các nguyên tử, phân tử C, H, O, N.
• Các đơn phân được liên kết với nhau bằng các liên kết hoá học.
• Đều có cáu trúc xoắn.
• Đều được đăc trưng bởi số lượng, thành phần và trình tự phân bố các đơn phân.
• Đều là 2 thành phần cơ bản tạo nên cấu trúc nhiễm sắc thể.
2. Điểm khác nhau giữa cấu trúc của ADN và prôtêin ADN Prôtêin
- ADN có cấu trúc xoắn kép, gồm 2 mạch đơn.
- ADN là đại phân tử chiều dài tới hàng trăm micrômet, khối lượng phân tử từ 4 triệu đến 8 triệu, thậm chí có thể tới 16 triệu đ.vC
- ADN được cấu tạo từ 4 loại nuclêôtit, thành phần cơ bản của mỗi nuclêôtit kà bazơnitric.
- Liên kết trên mỗi mạch đơn ADN là liên kết phôtphođieste (giữa đường C5 H10 O4 của nuclêôtit này với phân tử H3 PO4 của nuclêôtit bên cạnh). nhiều liên kết photphođieste tạô thành mạch polinuclêôtit.
- Trên mạch kép phân tử ADN các cặp nuclêôtit giữa 2 mạch đơn liên kết với nhau theo nguyên tắc bổ sung (NTBS) A liên kết với T bằng 2 liên kết hiđrô, G liên két với X
- Prôtêin có cấu tạo xoắn, mức độ xoắn tuỳ thuộc vào các bậc cấu trúc
- Prôtêin cũng là đại phân tử có kích thước bé hơn ADN, phân tử prôtêin lớn nhất cũng chỉ tới 0,1 micrômét, khối lượng phân tử
1,5 triệu đvC
- Prôtêin được cấu tạo từ 20 loại axitamin, thành phần cơ bản của mỗi axitamin là gốc cacbon (R)
- Trong phân tử prôtêin các axitamin liên kết với nhau bằng liên kết peptit (giữa nhóm amin của axit amin này với nhóm cácbôxin của axit amin bên cạnh cùng nhau giải phóng 1 phân tử nước). Nhiều kiên kết peptit tạo thành chuỗi pôlipeptit. Mỗi phân tử prôtêin có thể gồm 1 hoặc 1 số chiuỗi pôlipeptit.
- Mỗi phân tử ADN gồm nhiều gen - Cấu trúc hoá học của phân tử ADN quy
định cấu trúc hoá học của các prôtêin tương ừng.
kết với nhau bằng liên kết pepit; bậc 2 xoắn theo hình lò xo (xoắn anơpha) ( xoắn trái), chiều cao mỗi vòng xoắn 5,4Ǻ, với 3,7aa/vòng. Trong chuỗi xoắn bêta mỗi vòng xoắn có 5,1aa; cấu trúc bậc 3 là hình dạng phan tử prôtêin trong không gian 3 chiều tạo thành những khối hình cầu’ cấu trúc bậc 4 là những prôtêin gồm 2 hay nhiều pôlipeptit liên kết với nhau. Ví dụ phân tử hêmôglôbin có 2 chuỗi anpha và 2 chuỗi bêta, mỗi chuỗi chứa một nhân kèm với một nguyên tử Fe
- Mỗi phân tử prôtêin gồm nhiều chuỗi pôlipeptit
- cấu trúc hoá học của prôtêin phụ thuộc vào cấu trúc hoá học của các gen trên phân tử ADN
3. Những chức năng cơ bản của prôtêin
- Kiến tạo nên các bào quan, các tổ chức cơ quan của cơ thể, quy định các đặc điểm hình thái, giải phẫu của cơ thẻ.
- Tạo nên các enzim, mà bản chất là prôtêin xúc tác các phản ừng sinh hoá tổng hợp vật chất di truyền, các bản phiên mã, tham gia tổng hợp prôtêin và các hợp chất hữu cơ quan trọng khác. hiện nay đã biết khoảng 3500 loại enzim.
- Tạo nên các kháng thể có chức năng bảo vệ cơ thể chống lại các vi khuẩn gây bệnh.
- Các hoocmôn phần lớn là prôtêin có chức năng điều hoà các quá trình trong tế bào và cơ thể
- Nhiều loại prôtêin tham gia vào chức năng vận đông tế bào cơ thể
- Khi thiếu hụt gluxit, lipit, prôtêin tự phân huỷ giải phóng năng lượng dưới dạng ATP cung cấp cho quá trình hoạt hoá các nguyên liệu.
Prôtêin đảm nhận nhiều chức năng liên quan đến toàn bộ hoạt động sống của tế bào, quy định các tính trạng, tính chất của cơ thể sống.
Câu 4: Điểm giống nhau và khác nhau giữa cơ chế tổng hợp ADN ở sinh vật nhân chuẩn với E.coli. Ý nghĩa của cơ chế tổng hợp ADN
Trả lời
1. điểm giống nhau giữa cơ chế tổng hợp ADN của sinh vật nhân chuẩn và E,coli :
- Đều dựa trên khuôn mẫu của ADN mẹ
- Cần nguyên kiệu là ribônuclêôtit, các nuclêôtit
- Cần có ezim xúc tác để mở xoắn, tách 2 mạch đơn lắp ráp các nuclêôtit
- Cần tổng hợp đoạn mồi để tạo ra nhóm 3’OH
- Có một mạch tổng hợp gián đoạn (mỗi đoạn là một đoan okazaki)
- Đều dựa vào nguyên tắc bổ sung khi lắp ráp các nuclêôtit trên khuôn mẫu của từng mạch đơn ADN mẹ
- Kết quả đều tạo ra những ADN con giống ADN mẹ theo nguyên tắc bán bảo toàn.
2. Điểm khác nhau giữa tổng hợp ADN ở sinh vật nhân chuẩn với E.coli
- Trên phân tử prôtêin tạô nên 4 bậc cấu trúc không gian: bậc 1 các axit amin kiên -
Tổng hợp ADN ở E.coli Tổng hợp ADN ở sinh vật nhân chuẩn
- Toàn bộ ADN chỉ có một đơn vị tái bản
- sự tổng hợp xảy ra trên hai phiễu tái bản
- ADN- pôlimêraza gồm 3 loại enzim có chức năng khâc nhau đều tham gia tổng hợp ADN, trong đó ADN- pôlimêraza III đóng vai trò chủ yếu.
- có nhiều đơn vị tái bản
- Sự tổng hợp xảy ra trên nhiều đơn vị tái bản, những đơn vị tái bản nào có nhiều G==X được tổng hợp trước, nhiều AT
được tổng hợp sau.
- ADN- pôlimêraza có 5 loại enzim: ADNpôlimêraza γ tham gia tái bản ADN của ti thể. Enzim phôi ס, enzim ε là 2 enzim chính tham gia tổng hợp ADN. Pôli β và kiểu đơn vị bé ס có hoạt tính đọc sửa
- Về cấu trúc hoá học: do gen quy định.
- về cấu tạo không gian: do chức năng sinh học của các laọi prôtêin trong tế bào quy định.
Câu 6: Tại sao nói đột biến gen thường có hại, ít có lơi, tần số thấp, nhưng lại là nguồn nguyên liệu chủ yếu của chon lọc tự nhiên?
Trả lời
1. Khái niệm đột biến gen Đột biến gen là những biến đổi đột ngột trong cấu trúc phân tử của gen liên quan tới một hoặc một vài cặp nuclêôtit xảy ra tại một điểm nào đó của phân tử ADN. Dẫn tới làm xuất hiện một vài đặc điểm mới trên cơ thể sinh vật.
2. Nói đột biến gen thường có hại, ít có lợi, tần số thấp, nhưng lại là nguồn nguyên liệu chủ yếu của chon lọc tự nhiên vì:
3. Ý nghĩa của cơ chê tổng hợp Nhờ khả năng tổng hợp, ADN đã truyền đạt được thông tin di truyền của nó qua các thế hệ tế bào và qua các thế hệ khác nhau của loài.
Câu 5: Mối quan hệ giữa ADN và prôtêin trong cấu trúc và cơ chế di truyền.Tính đặc trưng cuẩ prôtêin do yếu tố nào quy định?
Trả lời
1. Mối quan hệ giữa ADN và prôtêin trong cấu trúc di truyền
- Prôtêin và ADN là hai thành phần cơ bản cấu trúc nên nhiễm sắc thể, trong nhiễm sắc thể
ADN và prôtêin có tỉ lệ tương đương. Prôtêin liên kết với các vòng xoắn của ADN giữ cho
cấu trúc ADN ổn định, thông tin di truyền trên ADN được điều hoà.
- Prôtêin của ADN tổ hợp với nhau tạo nên chất nhiễm sắc hình thành nhiễm sắc thể. Phân tử ADN quấn quanh một khối hinh cầu dẹt (gồm 8 phan tử prôtêin híton) tạo nên nuclêôxom. Các nuclêôxôm nối với nhau bằng các đoạn ADN dài 15-100 cặp nuclêôtit và một phân tử prôtêin histon. tổ hợp ADN với histon trong chuỗi nucleôxom tạo thành các sợi cơ bản.
- Ở tế bào có nhân ribôxôm gồm có một hạt lớn và một hạt bé. Hạt lớn gồm 45 phân tử prôtêin và 3 phân tử rARN. Lúc tổng hơp prôtêin hai hạt này liên kết với nhau, tiếp xúc với mARN và chuyển dịch từng bước trên mARN để thực hiên quá trình dịch mã.
- Cấu trúc hoá học ADN quy định cấu trúc hoá học của prôtêin (trình tự phân bố các nuclêôtit tren ADN quy định trình tự các axit amin trên phân tử prôtêin)
2. Mối quan hệ giữa ADN và prôtêin trong các cơ chế di truyền
- ADN là khuôn mẫu tổng hợp mARN từ đó quy định cấu trúc prôtêin
- ADN chứa nhiều gen cấu trúc, mỗi gen cấu trúc mang thông tin về một loại prôtêin.
- Prôtêin ức chế được tổng hợp từ khuôn mẫu của gen điều hoà gắn vào gen vận hành cản trở hoạt động của enzim phiên mã, do vậy gen cấu trúc được duy trì ở trạng thái không hoạt động.
- Prôtêin tham gia toạ nên các enzim tham gia vào tổng hợp ADN, ARN, prôtêin
- Prôtêin còn tham gia tạo nên các yếu tố mở đầu, kéo dài, kết thúc quá trình sinh tổng hợp prôtêin tù bản phiên mã mARN.
- Prôtêin tạo nên thoi tơ vô sắc, các dây tơ nối với các nhiếm sắc thể ở tâm động, đảm bảo cho sưn phân li nhanh và chính xác ổn đinh vật chất di truyền ở cấp độ tế bào.
3. Tính đăc trung của prôtêin do các yếu tố sau quy định:
- Phần lớn đột biến gen trong tự nhiên là có hại cho cơ thể sinh vật vì chúng phá vỡ mối quan hệ hài hoà trong kiểu gen, trong nội bộ cơ thể, giữa cơ thể với mội trường đã được hình thành qua chọn lọc tự nhiên. Trong môi trường quen thuộc, đa số thể đột biến thường tỏ ra có sức sống kém hoặc kém thích nghi so với dạng gốc. Đặt vào điều kiện mới, nó có thể tỏ ra thích nghi hơn, có sức sống cao hơn. Thí dụ trong mội trưòng không có DDT thì dạng ruồi mang đột biến kháng DDT sinh trưởng chậm hơn dạng ruồi bình thường, nhưng khi phun thuốc DDT thì lại có lợi cho ruồi.
- Tuy đột biến thường có hại nhưng phần lớn alen đột biến là alen lặn, xuất hiện ở một giao tử nào đó, alen lặn sẽ đi vào hợp tử và tồn tại bên cạnh alen trội tương ứng ở thể dị hợp, do đó không biểu hiên thành kiểu hình. Trải qua nhiều thế hệ giao phối các alen lặn được nhân lên, có điều kkiện gặp gỡ nhau trong giao phối tạo nên thể đồng hợp và biểu hiên thành kiểu hình. Giá trị thích ứng của một đột biến có thể thay đổi tuỳ tổ hợp gen. một đột biến nằm trong tổ hợp này có hại nhưng tồn tại trong tổ hợp gen khác lại trở nên có lợi.
Vì vậy có thể nói tính lợi, hại của đột biến gen chỉ là tương đốí.
- Nhờ quá trình giao phối các đột biến được phát tán trong quần thể tạo ra vô số biến dị tổ hợp. Có thể nói đột biến là nguồn nguyên kiệu sơ cấp còn biến dị tổ hợp là nguồn nguyên liệu thứ cấp cho chọn lọc tự nhiên do đó đều tạo nên vốn gen của quần thể. Sự tiến hoá không chỉ sử dụng các đột biến mới xuất hiện mà còn huy động kho dự trữ các gen đột biến đã phát sinh từ lâu nhưng tiêm ẩn trong trạng thái dị hợp.
Câu 7: Điểm giống và khác nhau cơ bản giữa nguyên phân và giảm phân. Ý nghĩa của chúng trong di truyền và tiến hoá.
Trả lời
1. Điểm giống nhau giữa nguyên phân và giảm phân
- Có sự nhân đôi của nhiễm sắc thể mà thức chất là sự nhân đôi của ADN ở kì trung gian.
- Trải qua các kì phân bào tương tự nhau
- đều có sự biến đổi hình thái nhiễm sắc thể theo chu kì đóng và tháo xoắn đảm bảo cho nhiễm sắc thể nhân đôi và thu gon cấu trúc để tập trung trên mặt phẳng xích đạo ở kì giữa.
- Ở lần phân bào II của giảm phân giống phân bào nguyên phân.
- Đều là cơ chế sinh học đảm bảo ổn định vật chất di truyền qua các thế hệ
2. Điểm khác nhau giữa nguyên phân và giảm phân
Nguyên phân Giảm phân
- Xảy ra một lần phân bào gồm 5 kì
- Mỗi nhiễm sắc thể tương đồng được nhân
- Xảy ra 2 lần phân bào lilên tiếp. lần phân bào I là phân bào giảm phân, lần phân bào II là phân bàonguyên phân
- Mỗi nhiễm sắc thể tương đồng được nhân
- 22 -
đôi thành 2 nhiễm sắc thể kép, mỗi nhiễm sắc thể kép gồm 2 crômatit
- Ở kì trước không xảy ra trao đổi chéo giữa 2 crômatit cùng nguồn gốc
- Tại kì giữa các nhiễm sắc thể tập trung thành từng nhiêm sắc thể kép
- Ở kì sau của nguyên phân có sự phân li các crômatit trong từng nhiễm sắc thể kép về 2 cực của tế bào
- Kết quả mỗi lần phân bào tạo ra 2 tế bào con có bộ nhiễm sắc thể lưỡng bội ổn định
- Xảy ra trong tế bào sinh dưỡng và mô têd bào sinh duc sơ khai
đôi thành một cặp nhiễm sắc thể tương đồng
kép gồm 4 crômatit tạo thành một thể thống
nhất.
- Ở kì trước I một số cặp nhiễm sắc thể có
xảy ra hiện tượng tiếp hợp và xảy ra trao đổi
đoạn giữa 2 crômatit khác nguồn gốc, tạo nhóm gen liên kết mới.
- tại kì giữa I các nhiễm sắc thể tập trung
thành từng nhiễm sắc thể tương đồng kép.
- Ở kì sau I của giảm phân có sự phân li các nhiếm sắc thể đơn ở trạng thái kép trong
từng cặp nhiễm sắc thể tương đồng kép để
tạo ra các tế bào con có bộ nhiễm sắc thể
đơn ở trạng thái kép khác nhau về nguồn
gốc nhiễm sắc thể
- Kết quả qua 2 lần phân bào tạo ra các tế bào giao tử có bộ nhiễm sắc thể giảm đi một
nửa khác biệt nhau về nguồn gốc và chất
lượng nhiễm sắc thể
- Xảy ra ở tế bào sinh dục sau khi các tế bào
đó kết thúc giai đoạn sinh trưởng
2. Ý nghĩa của nguyên phân và giảm phân
- Ý nghĩa của nguyên phân
+ Ổn định bộ nhiễm sắc thể của loài qua các thế hệ tế bào của cùng một cơ thể
+ Tăng nhanh sinh khối tế bào, đảm bảo sự phân hoá mô, cơ quan tạo lập nên một cơ thể hoàn chỉnh
+ Tạo điều kiện cho các đột biến tế bào sinh dưỡn có thể nhân lên qua các thế hệ tế bào, tạo nên thể khảm.
- Ý nghĩa của giảm phân
+ Giảm bộ nhiễm sắc thể trong giao tử, nhờ vậy kkhi thụ tinh khôi phục được trạng thái
lưỡng bội của loài trong quá trình tạo giao tử + Trong giảm phân có xảy ra hiện tượng phân li độc lập, tổ hợp tự do của các nhiễm sắc thể, sự trao đổi đoạn tại kì trước I của giảm phân đã tạo nên nhiều loại giao tử khác nhau về nguồn gốc và chất lượng.. Đây là cơ sở tạo nên các biến dị tổ hợp cung cấp nguyên liệu cho quá trình chọn lọc tự nhiên, tạo nên tính đa dạng của sinh giới.
+ Nhờ giảm phân các đột biến được nhân lên dần trong quần thể, trong loài biểu hiên thành kiểu hình đột biến.
Câu 8: Trình bày cơ chế hình thành các dạng tế bào n, 2n,3n,4n từ dạng tế bào 2n. Ý nghĩa
của việc hình thành các loại tế bào nói trên
Trả lời
1. Cơ chế hình thành các dạng tế bào
a. Cơ chế hình thành tế bào n:
- Ở các cơ thể trưởng thành có một nhóm tế bào sinh dưỡng được tách ra làm nhiệm vụ sinh sản, gọi là tế bào sinh dục sơ khai. Các tế bào lần lượt trải qua 3 giai đoạn:
+ Giai đoạn sinh sản: nguyên phân liên tiếp nhiều đợt tạo ra các tế bào sinh dục con
+ Giai đoạn sinh trưởng: các tế bào tiếp nhân nguyên liệu mội trường ngoài tạo nên các tế bào con có kích thước lớn (kể cẩ nhân và tế bào chất)
+ Giai đoạn chín: các tế bào sinh tinh trùng, sinh trứng bước vào giảm phân gồm 2 lần phân bào liên tiếp: lần 1: giảm phân, lần 2: nguyên phân để tạo ra các giao tử đơn bội
- Giảm phân I:
+ Ở kì trung gian ADN nhân đôi, mỗi cặp nhiễm sắc thể tương đồng nhân đôi thành cặp tương đồng kép.
+ Ở kì trước I: nhiễm sắc thể tiếp tục xoắn lại, kì này tại một số cặp nhiễm sắc thể có xảy ra trao đổi đoạn giữa 2 crômatit khác nguồn gổc trong cặp tương đồng. Cuồi kì trước I, màng nhân mất, bắt đầu hình thành dây tơ vô sắc.
+ Ở kì giữa I: thoi tơ vô sắc hình thành xong. Các nhiễm tương đồng kép tập trung thành cặp trên mặt phẳng xích đạo nối với dây tơ vô sắc tại tâm động
+ Ở kì cuối I: tạo 2 tế bào con chứa bộ nhiễm sắc thể ở trang thái kép, khác nhau về nguồn gốc, chất lượng nhiễm sắc thể
- Giảm phân II: ở lần phân bào này giống phân bào nguyên phan, kì trung gian trải qua rất ngắn ở kì giữa II, các nhiễm sắc thể đơn ở trạng thái kép trong mỗi tế bào tập trung trên mặt phẳng xích đạo nối với dây tơ vô sắc. Ở kì sau II, mỗi crômatit trong mỗi nhiễm sắc thể đơn ở trngj thái lép phân li về hai cực. Kì cuối II tạo ra các tế bào đơn bội. Từ một tế bào sinh tinh trùng tạo ra 4 tinh trùng, từ 1 tế bào sinh trứng tạo ra 1 trứng và 3 thể định hướng.
b. Cơ chế hình thành tế bào 2n:
- Qua nguyên phân
+ Ở kì trung gian mỗi nhiễm sắc thể đơn tháo xoắn cực đại ở dạng sợi mảnh, ADN nhân đôi để tạo ra các nhiễm sắc thể kép + Kì trước: nhiễm sắc thể tiếp tục xoắn lại, cuối kì trước màng nhân mất, thoi vô sắc bắt đầu hình thành.
+ Kì giữa: thoi vô sắc hình thành xong, nhiễm sắc thể kép tập trung trên mặt phẳng xích đạo nối với dây tơ vô sắc tại tâm động
+ Kì sau: mỗi crômatit trong từng nhiễm sắc thể kép tập trung trên mặt phẳng xích đạo nối với dây tơ vô sắc tại tâm động
+ Kì sau: mỗi crômatit trong từng nhiễm sắc thể kép tách nhau qua tâm động phân chia về 2 cực tế bào
+ Kì cuối: các nhiễm sắc thể đơn giãn xoắn cực đại, màng nhân hình thành, mỗi tế bào chứa bộ nhiễm sắc thể lưỡng bội (2n) Nhân đôi phân chia
2n ►4n ────────►2n
- Qua giảm phân không bình thường
Các tế bào sinh tinh trùng hoặc sinh trứng nếu bi tác động của các tác nhân phóng xạ, hoá học ... làm cắt đứt dây tơ vô sắc trên toàn bộ bộ nhiễm sắc thể sẽ tạo nên các giao tử lưỡng bội.
- Qua cơ chế thụ tinh:
Sự kết hợp giữa tinh trùng đơn bội và trưng đơn bội qua thụ tinh sẽ tạo nên hợp tử lưỡng bội (2n)
c. Cơ chế hình thành dạng tế bào 3n, 4n:
- Tế bào 2n giảm phân không bình thường xảy ra trên tất cả các cặp nhiễm sắc thể sẽ tạo nên giao tử 2n. Giao tử này kết hợp với giao tử binh thường n sẽ tạo nên hợp tử 3n.
- 24 -
- Giao tử không bình thường 2n kết hợp với nhau sẽ tạo nên hợp tử 4n
- Ngoài ra dạng 3n còn được hình thành trong cơ chế thụ tinh kép ở thực vật do nhân
thứ cấp 2n kết hợp với một tinh tử n tạo nên nội nhũ 3n.
- dạng tế bào 4n còn đuợc hình thành do nguyên phân rối loạn xảy ra trên tất cả các cặp
nhiễm sắc thể sau khi nhân đôi.
2. Ý nghĩa của việc hình thành các dạng tế bào n, 2n ,3n, 4n
a. Ý nghĩa việc hình thành dạng tế bào n
b. Ý nghĩa việc hình thành dạng tế bào 2n, 3n, 4n:
- Kế tục vật chất di truyền ổn định trong sinh sản sinh dưỡng và sinh sản hữu tính
- Nếu tế bào 2n được hình thành do giảm phân rối loạn là cơ sơ tạo nên các dạng 3n, 4n qua thụ tinh hình thành những đặc trưng mới về các tính trạng hình thái, sinh lí, hoá sinh cho cơ thể.
- Ở thực vật dạng 3n còn được hình thành do nhân thứ cấp kết hợp với 1 tinh tử tạo nên nội nhũ 3n
- Việc tạo nên các tế bào có bộ nhiễm sắc thể không bình thường về số lượng, đây cũng là một hướng tiến hoá cơ bản ở thực vật trong tự nhiên.
Câu 9: Các cơ chế sinh học xảy ra như thế nào đối với 1 cặp NST tương đồng ở cấp độ TB?
Trả lời
Có nhiều cơ chế sinh học xảy ra đối với 1 cặp NST tương đồng. Mỗi cơ chế đều có ý nghĩa khác nhau góp phần ổn định bộ NST của loài.
1. Cơ chế nhân đôi của nhiễm sắc thể:
Thực chất mỗi NST đơn trong cặp NST tương đồng là sự nhân đôi ADN trên NST vào kì trung gian. Nhờ đó mỗi NST đơn tạo ra 1 NST kép.
2. Cơ chế trao đổi đoạn: Ở kì trước I có xảy ra hiện tượng tiếp hợp và trao đổi chéo giữa 2 crômatit khác nguồn gốc trong cặp NST tuơng đồng. Sự trao đổi chéo góp phần tạo ra nhiều kiểu giao tử, tạo sự đa dạng, phong phú của loài.
3. Cơ chế phân li:
- Trong nguyên phân, các NST trong cặp NST tương đồng đã phân li đồng đều về mỗi cực của TB để góp phần tạo bộ NST 2n trong các TB con.
- Trong giảm phân I bình thường, mỗi NST trong cặp NST tương đồng đã phân li về 2 cực của TB tạo ra bộ NST đơn bội ở thể kép. Ở mỗi TB con có 1 kiểu sắp xếp NST khác nhau trên mặt phẳng xích đạo ở kì giữa I. Tại kì sau II ở lần phân bào II, mỗi crômatit trong từng NST kép tách nhau qua tâm động, kết quả mỗi giao tử chỉ chưa 1 NST đơn trong cặp tương đồng.
4. Cơ chế tổ hợp tự do của NST: Tại kì giữa I do mỗi TB có 1 kiểu sắp xếp nên khi phân li sẽ tạo ra các loại giao tử khác nhau.
- Nhờ thụ tinh phối hợp ngãu nhiên các giao tử đực với giao tử cái đã tạo lại cặp NST tương đồng, ổn định ở thế hệ sau.
5. Cơ chế đột biến dị bội thể: Do nguyên nhân phóng xạ, hoá chất, cơ học, sức li tâm, sốc nhiệt hoặc quá trình trao đổi chất nội bào bị rối loạn. Các tác nhân làm đứt day tơ vô sắc, hoặc ức chế hình thành day tơ vô sắc xảy ra trong nguyên phân sẽ tạo nên TB chứa cả 2 NST, TB không chứa NST của cặp tương đồng. Nếu xảy ra trong giảm phân sẽ tạo nên giao tử di bội n-1, n+1. Các giao tử
này kết hợp với giao tử bình thường tạo hợp tử chứa 3 NST trong cặp tương đồng đó. Thí dụ người mắc bệnh Đao có 3 NST thứ 21. Các đột biến dị bội thường gây hậu quả có hại.
6. Cơ chế đột biến cấu trúc NST: Do nguyên nhân bên trong TB như rối loạn trao đổi chất nội bào, biến đổi sinh lí, hoá sinh hay do tác nhân phóng xạ, hoá học tác động vào NST lúc chúng đang nhân đôi sẽ tạo nên các đột biến mất đoạn, lặp đoạn, chuyển đoạn, xảy ra trong phạm vi 1 cặp NST. Các đột biến cấu trúc NST thường gây ra hậu quả có hại. Đột biến mất đoan thường làm giảm sức sống hoặc gây chết. Ở người, mất đoạn ở NST 21 gây ung thư máu. Đột biến lặp đoạn thường gây hậu quả khác nhau, hoặc tăng cường hoặc giảm bớt mức biểu hiện của tính trạng. Ở ruồi giấm lặp đoạn 2 lần ở NST X làm cho mắt lồi thành mắt dẹt. Ở đại mạch ,lặp đoạn làm tăng hoạt tính enzim amilaza rất có ý nghĩa trong công nghiệp SX bia. Đảo đoạn thường ít ảnh hưởng tới sức sống của cơ thể vì vật chất di truyền không bị mất mát. Chuyển đoạn có sự phân bố lại các gen giữa các NST khác nhau, 1 số gen trong nhóm liên kết này chuyển sang nhóm liên kết khác. Sự chuyển đoạn thường gây chết hoặc mất khả năng sinh sản. Tóm lại đột biến cấu trúc NST thể sẽ làm rối loạn sự liên kết các cặp NST tương đồng trong giảm phân, làm thay đổi tổ hợp gen trong giao tử dẫn tới biến đổi kiểu gen và kiểu hình.
Câu 10: Cấu trúc và chức năng của NST thường và NST giới tính giống nhau và khác nhau ở điểm nào?
Trả lời 1. Điểm giống nhau của NST thường và NST giới tính về cấu trúc
- Trong TB sinh dưỡng đều tồn tại thành cặp, mỗi cặp NST gồm 2 NST đơn thuộc 2 nguồn gốc. Trong TB giao tử tồn tại từng chiếc.
- Các thành phần cơ bản cấu trúc nên NST thường và NST giới tính đều là ADN và prôtein (chủ yếu là histon)
- Đơn vị cơ bản tạo nên NST đều là nuclêôxôm, mỗi nuclêôxôm gồm 8 phân tử prôtein histon phía ngoài bọc bởi 4 3 1 vòng xoắn gồm khoảng 146 cặp nuclêôtit. Các nuclêôxôm nối với nhau bằng các đoạn nối ADN (có kích thước 15-100 cặp nuclêôtit và 1 phân tử H1 (Histôn H1) tạo nên sợi cơ bản có chiều rộng 100 0 A . Sợi cơ bản tiếp tục xoắn thành sợi nhiễm sắc. Sợi nhiễm sắc đóng xoắn tiếp thành crômatit.
- Có kích thước, hình dạng đặc trưng cho mỗi loài.
- Đều chứa các nhóm gen liên kết hoàn toàn hay liên kết không hoàn toàn.
- Có khả năng đột biến làm thay đổi số lượng và cấu trúc NST
2. Điểm khác nhau cơ bản
- NST thường gồm nhiều cặp NST, luôn tồn tại thành từng cặp NST tương đồng. Còn NST giới tính chỉ tồn tại có 1 cặp khi thì đồng dạng ở giới đực, khi lại đồng dạng ở giới cái tuỳ thuộc vào loài.
-Gen trên NST thường phân bố thành từng cặp gen tương đồng. Gen trên NST giới tính XY phân bố thành nhiều vùng. Ví dụ ở người có 3 vùng gen:
+ Vùng gen có cả trên X và Y
+ Vùng gen chỉ có trên X
+ Vùng gen chỉ có trên Y
- Đại bộ phận các gen phân bố trên NST thường chỉ có 1 số gen được phân bố trên NST giới tính.
3. Điểm giống nhau về chức năng giữa NST thường với NST giới tính.
- 26 -
- Đều góp phần quy định tính đặc trưng bộ NST loài.
- Đều có khả năng nhân đôi, phân li, tổ hợp tự do, trao đổi đoạn trong nguyên phân, giảm phân và thụ tinh đảm bảo di truyền ổn định tương đối qua các thế hệ.
- Các gen trên NST có khả năng điều hoà, tổng hợp ARN, chỉ huy tổng hợp prôtein, để hình thành tính trạng đặc trưng cho loài.
- Các đột biến số lượng, cấu trúc NST đều có thể hình thành các tính trạng không bình thường, ảnh hưởng tới sự tồn tại và phát triển của cơ thể
4. Điểm khác nhau về chức năng:
- NST thường mang gen xác định việc hình thành các tính trạng thường, NST giới tính quy dịnh hình thành tính trạng đực, cái ở sinh vật, các tính trạng sinh dục phụ và gen xác định hình thành 1 số tính trạng liên quan đến giới tính, tuân theo quy luật di truyền chéo (gen nằm trên X), tuân theo quy luật di truyền thẳng (gen nằm trên Y).
- Các đột biến gen lặn xuất hiên trên NST thường biểu hiện tính trạng chậm hơn. Còn gen đột biến lặn trên NST X, Y có thể biểu hiện kiểu hình ngay trong đời cá thể và di truyền cho thế hệ sau.
Câu 11. Trình bày các loại biến dị làm thay đổi về số lượng, cấu trúc NST?
Trả lời
1. Đột biến số lượng NST:
- Sự thay đổi số lượng NST ở 1 hay 1 số cặp NST tạo nên thể dị bội, ở toàn bộ cặp NST tạo nên thể đa bội.
- Có 2 nguyên nhân gây ra đột biến số lượng NST đó là tác nhân lí hoá trong ngoại cảnh hoặc MT nội bào lam cản trở sự phân li của các cặp NST ở kì sau quá trình phân bào.
- Có 2 loại đột biến số lượng NST:
+ Thể dị bội (lệch bội): TB sinh dưỡng đáng nhẽ chúa 2 NST ở mỗi cặp tương đồng thì lại có thể chứa 3 NST ( thể 3 nhiễm), nhiều NST (thể đa nhiễm), 1 NST (thể 1 nhiễm) hoặc thiếu hẳn 1 NST (thể khuyết nhiễm). Các đột biến này thường gây ra những hậu quả có hại, ví dụ 3 NST 21 ở người gây bệnh Đao. Dạng XO biểu hiện hội chứng tơcnơ, XXY hội chứng claiphentơ.
+ Thể đa bội, người ta chia thành 2 loại: đa bội chẵn (4n, 6n, ..) và đa bội lẻ (3n, 5n, ….).
- Các thể đa bội, dị bội là nguồn nguyên liệu cho chọn lọc tự nhiên có ý nghĩa trong chọn giống cây trồng.
2. Biến dị làm thay đổi cấu trúc NST:
- Trao đổi đoạn xảy ra ở kì trước I giảm phân giữa 2 cromatit khác nguồn gốc trong cặp tương đồng, chúng đứt ra các đoạn tương ứng hoặc không tương ứng và chuyển đổi cho nhau. Trao đổi đoạn NST dẫn tới sắp xếp lại gen trong phạm vi 1 cặp NST tạo ra nhóm gen liên kết mới, làm thay đổi cấu trúc NST.
- Đột biến cấu trúc NST bao gồm mất đoạn, đảo đoạn, lặp đoạn NST, chuyển đoạn NST.
Các đột biến này làm thay đổi cấu trúc NST ở các mức độ khác nhau.
Câu 12 Điểm giống và khác nhau cơ bản giữa biến dị tổ hợp và biến dị đột biến. Vai trò
của các loạ biến dị đó trong tiến hoá và chọn giống.
Trả lời
1. Điểm giống
- Cả 2 đều là biến dị di truyền, qua giao tử vào hợp tử để di truyền qua các thế hệ
- Cả 2 đều thuộc biến dị vô hướng có thế có lợi, có hại hay trung tính.
Có thể xuất hiện những biến dị mới chưa có ở bố mẹ hoặc tổ tiên.
2. Điểm khác nhau:
- Về nguyên nhân: Biến dị tổ hợp xuất hiện nhờ quá trình giao phối, biến dị
t biến xuất hiên do tác động của MT trong và ngoài cơ thể.
- Về cơ chế: biến dị tổ hợp phát sinh do cơ chế phân li độc lập, tổ hợp tự do, hoán vị gen trong quá trình tạo giao tử và sự kết hợp ngẫu nhiên giữa giao tử đực và cái trong quá trình thụ tinh. Còn biến dị đột biến phát sinh do rối loạn quá trình phân bào hoặc do rối loạn quá trình tái sinh NST, đã làm thay đổi số lượng, cấu trúc vật chất di truyền.
- Về tính chất biểu hiện:
+ Biến dị đột biến biểu hiện 1 cách đột ngột, ngẫu nhiên, cá biệt, không định hướng. Phần lớn đột biến là có hại, tần số đột biến có lợi là rất thấp. Còn biến dị tổ hợp dựa trên cơ sở tổ hợp lại các gen vốn có ở bố mẹ và tổ tiên, vì thế có thể làm xuất hiện các tính trạng đã có hoặc chưa có ở thế hệ trước. Sự phân li độc lập, sự hoán vị gen, tương tác qua lại giữa các gen đã sắp xếp lại tính trạng vốn có hoặc xuất hiên tổ hợp kiểu hình mới ở thế hệ sau.
+ Biến dị tổ hợp có thể dự đoán được quy mô xuất hiện, tần số xuất hiện nếu biết trước được kiểu di truyền của bố mẹ. Còn biến dị đột biến không thể chủ động xác định trước khả năng xuất hiện ở đời con với loại đột biến và tần số đột biến là bao nhiêu.
+ Biến dị tổ hợp thường có giá trị không đặc sắc như đột biến, chúng thường xuất hiện ở mức độ nhỏ, đa dạng tạo ra nguồn biến dị thương xuyên, vô tận cho chọn lọc tự nhiên.
3. Vai trò
- Biến dị tổ hợp là nguồn ngưyên liệu biến dị di truyền thứ cấp cung cấp cho tiến hoá. Nhờ các biến dị này mà trải qua lịch sử dài từ 1 vài loài ban đầu có thể tạo ra nhiều loài mới. Trong chon giống dựa trên cơ chế xuất hiện biến dị tổ hớp đề xuất các phương pháp lai giống nhằm nhanh chóng tạo giống có giá trị
- Biến dị đột biến là nguồn nguyên liệu biến dị di truyền sơ cấp cung cấp cho tiến hoá. Đặc biệt đột biến gen là nguồn nguyên liệu cơ bản. Trong chọn giống dựa trên cơ chế xuất hiện, giá trị của các loại đột biến, người ta xây dựng các phương pháp gây đột biến nhằm nhanh chóng tạo ra các đột biến có giá trị, góp phần tạo ra giống cây trồng và sinh vật có năng suất, phẩm chất cao, thích nghi tốt.
B -PHẦN : BÀI TẬP
Bài 1. Một gen có chiều dài 5100 Ăngstron. Hiệu số phần trăm giữa adenin với một loại nucleotit khác bằng 10% số nucleotit của gen. Trên phân tử mARN được tổng hợp từ gen đó có 10% uraxin. một mạch đơn của gen có 16% xitozin, số timin bằng 150 nucleotit.
e) Tỉ lệ phần trăm và số lượng từng loại nucleotit của gen.
f) Tỉ lệ phần trăm và số lượng từng loại ribonucleotit của một phân tử mARN bằng bao nhiêu?
g) Nếu gen đó sao mã 6 lần và trên mỗi phân tử mARN có 10 riboxom trượt qua không lặp lại thì số lượng axit amin mà môi trường nội bào cung cấp cho quá trình tổng hợp protein là bao nhiêu?
h) Nếu thời gian giải mã một axit amin la 0,1 giây, thời gian tiếp xúc của một phân tử mARN với các riboxom là 58,1 giây, khoảng cách giữa các riboxom kế tiếp khoảng bao nhiêu Ăngstron?
GIẢI
a) Tỉ lệ % và số lượng từng loại nucleotit
- Số nucleotit của gen l 5100
- Đều là những biến dị có liên quan đến biến đổi vật chất di truyền. - 28
N= 2 . —— = 2 . —— = 3000Nu
3,4Ǻ 3,4
- Tỉ lệ % từng loại nuclêôtit của gen:
N
A + G = — = 50% (1)
2
A − G = 10% (2)
=> A = T = 30% ; G = X = 20%
- Số lượng từng loại nuclêôtit của gen
A = T = 3000 . 30% = 900Nu
G = X = 3000 . 20% = 600Nu
b) Tỉ lệ % và số lượng từng loại ribônuclêôtit 3000
- Số uraxin của mARN : rU = —— . 10% = 150Nu
2
- Số nuclêôtit từng loại ở mỗi mạch:
T1 = A2 = 150 ; A1 = T2 = A − A2 = 900 − 150 = 750Nu
300
X1 = G2 = —— . 16% = 240Nu
2
G1 = X2 = G − G2 = 600 − 240 = 360Nu
- U của mARN được tổng hợp từ A của mạch gốc
rU = A gốc => rU = A2 = 150. Vậy mạch 2 là mạch gốc
- Tỉ lệ % và số lượng từng loại ribônuclêôtit
750
rA = T2 = 750 => —— . 100% = 50%; rU = 150 −>10%
1500
360
rX = G2 = X1 = 240 −> 16%; rG = X2 = 360 −> . 100% = 24% 1500
c) Số axit amin tự do
Số phân tử prôtêin : kn = 6.10 = 60
- Số axit amin tự do cần dùng: rN 1500
∑aatd = Số P ( — − 1) = 60 ( —— − 1) = 29940 3 3
d) Khoảng cách giữa các ribôxôm l
- Vận tốc trượt của ribôxôm : v = — .3.3,4 = 102 Ǻ/ s
0,1
- Thời gian kể từ RB1 tiếp xúc cho đến RB cuối cùng hết tiếp xúc mARN : l ∆l
T = t + t’ = — + ( n- 1) — v v 5100 ∆l
=> Phương trình —— + (10-1) —— = 58,1
102 102
=> ∆l = 91,8Ǻ Bài 2: Một gen dài 5100 Ắngtron. Khi gen tự sao liên tiếp hai đợt, môi trường nội bào đã cung cấp 2700 ađênin. Phân tử mARN được tổng hợp từ gen đó có 600 adenin và 240 guanin. vận tốc giải mã là 10 axit amin/ giây. Tính từ lúc ribôxôm thứ nhất trượt qua phân tử mARN cho đến khi hết phân tử mARN đó là 55,6 giây.
d) Xác định số lượng từng loại nuclêôtit ởtrong toàn bộ các gen được hình thành sau hai đợt tự sao liên tiếp
e) Xác định số lượng từng loại nuclêôtit ở mỗi mạch đơn của gen
f) Tính khoảng cách theo Ắngtron giữa ribôxôm thứ nhất với ribôxôm cuối cùng khi chúng dang tham gia giải mã trên một phân tử mARN
GIẢ
I
a) Số lượng từng loại nuclêôtit trong tất cả các gen con: L 5100
- Số nuclêôtit của gen N = 2. —— = 2. —— = 3000Nu
3,4Ǻ 3,4
- Số nuclêôtit tưng loại của mỗi gen: 2700
∑Atd = A (22 – 1) = 2700 => A = T = ——— = 900Nu
22 – 1 N 3000
G = X = — - A = —— - 900 = 600 Nu 2 2
- Số nuclêôtit từng poại trong toàn bộ gen con:
∑A = ∑T = 900 . 22 = 3600Nu
∑G = ∑X = 600 . 22 = 2400Nu
b) số lượng từng loại nuclêôtit ở mỗi mạch gen:
Quy ước mạch gốc là mạch thứ nhất
T1 = A2 = rA = 660; A1 = T2 = A – A2 = 900 – 600 = 240Nu
X1 = G2 = rG = 240; G1 = X2 = G – G2 = 600 – 240 = 360Nu
c) khoảng cách giữa ribôxôm thứ nhất với ribôxoom cuối cùng
- Vận tốc trượt của RB: v = 10 . 3 . 3,4 = 102Ǻ/s
- Thời gian kể từ RB1 tiếp xúc khi nó trượt qua mARN l 5100
t = — = —— = 50 giây v 102
- Thời gian kể từ RB1 trượt qua mARN đến khi RB cuối cùng trượt hết mARN = khoảng cách về thời gian giữa RB1 với RB cuối cùng
t’ = T – t = 56,5 – 50 = 5,6 giây
- Khoảng cách giữa RB1 với RB cuối cùng
102 . 5,6 = 571,2Ǻ
Bài 3: mạch thứ nhất của gen có tỉ lệ các nuclêôtit là 10% adenine, 20% timin và 25% xitozin. Phân tử mARN được sao từ gen đó có 20% urãin.
d) tính tỉ lệ từng loại nuclêôtit của gen và từng loại ribônuclêôtit của phân tử mARN
e) nếu gen đó dài 0,306 micromet thì nó chứa boa nhiêu liên kết hidro?
- 30 -
f) một phân tử mARN sinh ra từ gen có chiều dài nói trên và có một số riboxom cùng hoạt động trong quá trình giải mã, thời gian riboxom thứ nhất trượt qua hết phân tử là 30 giây và thời gian tính từ lúc bắt đầu có sự giải mã đến khi riboxom cuối cùng trượt qua hết phân tử mARN đó là 35,4 giây. Hỏi có bao nhiêu riboxom tham gia vào quá trình giải mã?
Biết rang các riboxom cách đều nhau một khoảng bằng 61,2Ǻ
GIẢI
a) tỉ lệ từng loại nuclêôtit và tưng loại ribônuclêôtit
- Tỉ lệ từng loại nuclêôtit cau rmỗi mạch gen
A1 = T2 = 10% ; T1 = A2 = 20%
X1 = G2 = 25% ; G1 = X2 = 100% - ( 10% + 20% + 25%) = 45%
- Tỉ lệ từng loại ribônuclêôtit của cả gen
%A1 + %A2 10% + 20%
A = T = —————— = ——————— = 15%
2 2
%G1 + %G2 45% + 25%
G = X = —————— = ——————— = 35%
2 2
- U của mARN được sao từ A gốc của gen
rU = A gốc => rU = A2 = 20%
Vậy mạch 2 là mcạh gốc và tỉ lệ 20% tưng loại ribônuclêôtit ccủa mARN :
rA = T2 = 10%; rU = A2 = 20%
rG = X2 = 45%; rX = G2 = 25%
b) số liên kết hidro của gen L 3060
- Số nuclêôtit của gen: N = 2 . —— = 2 . —— = 1800Nu 3,4Ǻ 3,4
- Số nuclêôtit tưng loại của gen:
A = T = 1800 . 15% = 270; G = X = 1800 . 35% = 630Nu
- Số liên kết hidro của gen:
H = 2A + 3G = 2 . 270 + 3 . 630 = 2430liên kết
c) Số riboxom l 3060
- Vận tốc trượt của RB: v = — = —— = 102Ǻ/s t 30
- Thời gian tính từ lúc RB1 trượt qua mARN đến khi RB cuối cùng trượt qua mARN = khoảng cách về thời gian giữa RB1 với RB cuối cùng
t’= T – t = 35,4 – 30 = 5,4 giây
- Số riboxom tham gia:(n )
∆l 61,2
(n – 1 ) — = t’ => (n – 1 ) —— = 5,4 => n = 10 v 102
Bài 4: Mạch thứ nhất của gen có 240 timin, hiệu số giữa guanin với adenine bằng 10% số nucleotit của mạch. Ở mạch thứ hai, hiệu số giữa adenin và xitozin bằng 20% số nucleotit của mạch. Khi gen đó tổng hợp phân tủe mARN thì mội trường nội bào đã cung cấp 360 uraxin.
d) Tỉ lệ phần trăm và số lượng từng loại nucleotit của gen và của tưng mạch là bao nhiêu?
e) Hãy tính chiều dài của phân tử mARN, tỉ lệ phần trăm và số lương mỗi loại ribonucleotit của nó.
f) Trên mỗi phân tử mARN có 8 riboxom cùng giagỉ mã, tính từ lúc riboxom bắt đâu ftrượt trên phân tử mARN thì thời gian để riboxom thứ nhất trượt qua hết sphân tử là 20 giây, còn riboxom cuói cùng thì phải cần đén 26,3 giây mới hoàn tất việc giải mã. khoảng cách đều giữa các riboxom là bao nhiêu Ắngtron? Biết rang các riboxom trượt với vận tốc bằng nhau
GIẢI
a) tỉ lệ và số lượng từng loại nuclêôtit của gen và của từng mạch đơn
- tỉ lệ % từng loại nuclêôtit mỗi mạch
G1 = A1 = 10% => X2 – T2 = 10% => T2 = X2 – 10% (1)
A2 – X2 = 10% => A2 = 10% + X2 (2)
X2 – G2 = 20% => G2 = X2 – 20% (3)
từ (1) (2) (3) => (X2 – 10%) + ( 10% + X2) + (X2 – 20%) + X2 = 100%
X2 = 30%
Suy ra G1 = X2 = 30%; X1 = G2 = 10%
A1 = T2 = 20%; T1 = A2 = 40%
- Số lượng tưng loại nuclêôtit mỗi mạch
240
T1 = A2 = 240; A1 = T2 = —— . 20% = 120Nu 40% 240 240
G1 = X2 = —— . 30% = 180; G2 = X1 = —— . 10% = 60Nu 40% 40%
- Tỉ lệ và số lượng từng loại nuclêôtit tương ứng của cả gen:
%A1 + %A2 20% + 40%
A = T = —————— = —————— = 30% 2 2
A =T = A1 + A2 = 120 + 240 = 360
%G1 + %G2 30% + 10%
G = X = —————— = —————— = 20% 2 2
G =X = G1 + G2 = 180 + 60 = 240
b) Chiều dài, tỉ lệ % và số lượng từng loại ribônuclêôtit của mARN
- Chiều dài của mARN N
l = L = — . 3,4Ǻ = (210 + 240 + 180 + 60) . 3,4 = 2040 Ǻ 2
- k số phân tử mARN (nguyên, dương)
- U của mARN tổng hợp từ A gốc của gen => ∑rU = k . A gốc 360
- Nếu mạch 2 là mạch gốc : k = —— = 1,5 ( loại) 260
- 32 -
Vậy mạch 1 là mạch gốc với số lần sao mã k = —— = 3 120
- Tỉ lệ % và số lượng ribônuclêôtit từng loại của mARN
240 120
rA = T1 = 240 => —— = 40%; rU = A1 = 120 => —— = 20%
600 600 60 180
rG = X1 = 60% => —— = 10%; rG = G1 = 180 => —— = 30%
600 600
b) Khoảng cách giữa các riboxom l 2040
- Vận tốc trượt của RB: v = — = —— = 102Ǻ/ s t 20
- Gọi ∆l : khoảng cách giữa các RB
Thời gian lúc RB1 bắt đầu trượt cho đến khi RB cuối cùng trượt hết phân tử mARN
∆l ∆l
T = t + t’ = t + (n – 1 ) — => 20 + (8 – 1 ) —— = 26,3
∆l = 91,8Ǻ
v 102
Bài 5: Trên một phân tử mARN có một số riboxom trượt qua với khoảng cách đều bằng nhau. Riboxom thứ nhất trượt qua hết phân tử mARN đó hết 50 giây. Tính từ lúc riboxom thứ nhất trượt qua và tiếp xúc với phân tử mARN đó thì riboxom cuối cùng ppjải mất 57,2 giây mới hoàn thành việc đi qua phân tử mARN . Biết rang phân tử prôtêin thứ hai được tổng hợp chậm hơn phân tử prôtêin thứ nhất 0,9 giây.
Gen điều khiển việc tổng hợp các phân tử prôtêin nói trên có mạch 1 chứa 10% adeini và 20% guanine, mạch 2 chứa 15% adenine. Quá trình sao mã của gen đã đòi hỏi mội trường nội bào cung cấp 150 uraxin và 155 adenin để góp phần tổng hợp một phân tử
mARN
f) tính chiều dài của gen
g) Tính số lượng từng loại ribônuclêôtit của một phân tử mARN
h) Số riboxom đã tham gia vào qua trình giải mã trên một phân tử mARN đó là bao nhiêu? Biết rằng mỗi riboxom chỉ trượt qua một lần
i) khoảng cách giữa 2 riboxom kế tiếp và khoảng cách giữa riboxom thứ nhất với riboxom cuối cùng tính theo Ắngtron là bao nhiêu?
j) Toàn bộ quá trình giải mã nới trên đã cần bao nhiêu axit amin của mội trường nội bào và trong tất cả các prôtêin hoàn chỉnh chứa bao nhiêu axit amin?
GIẢI
a) Chiều dài của gen
- Số nuclêôtit loại A,T của gen
A = T = rA + rU = 225 + 150 = 375
- Tỉ lệ % tưng loại nuclêôtit của gen
%A1 + %A2 10% + 15%
A = T = ————— = ————— = 12,5%; G = X = 37,5%
375. 100
- Số nuclêôtit của gen: N = ———— = 3000 12,5
b) Số kượng từng loại ribônuclêôtit
- U của mARN được tổng hợp từ A gốc 3000
rU = A gốc => rU = A1 = —— . 10% = 150 2
Vậy mạch một là mạch gốc
- Tỉ kệ % từng loại nuclêôtit ở mạch gốc
A1 = 10%; T1 = A2 = 15%; G1 = 30%
X1 = 2 . %G - %G1 = 2 . 37,5% - 30% = 45%
Số ribônuclêôtit từng loại của mARN : rA = 225rU = 150 3000 3000
rG = X1 = —— . 45% = 675; rX = G1 = —— . 30% = 450 2 2
c) Số riboxom: gọi n : số riboxom
Thời gian kể từ RB1 tiếp xúc mARN đến khi RB cuối cùng trượt qua hết mARN :
T = t + t’ = t + (n – 1 ) ∆t => 50 + (n – 1 )0,9 = 57,2 => n = 9
d) Khoảng cách giữa 2RB, giữa RB1 với RB cuói cùng l 5100
- Vận tốc trượt của RB: v = — = —— = 102Ǻ/ s t 50
- Khaỏng cách giữa 2RB: ∆l = ∆t . v = 0,9 . 102 = 91,8Ǻ
- Khoảng cách giữa RB1 với RB cuối cùng: (n – 1 ) ∆l = (9 – 1) 91,8 = 734,4Ǻ
e) Số axit amin
- Tổng số axit amin tự do cần dùng:
rN 1500
∑ aatd = Số P (— - 1) = 9 . (—— - 1 ) = 4491
3 3
- Tổng số axit amin của các phân tử prôtêin hoàn chỉnh:
rN 1500
∑ aap = Số P (— - 2 ) = 9. ( —— - 2) = 4482
3 3
Bài 6: Ở một loài một tế bào sinh dục 2n thực hiện sự nguyên phân liên tiếp một số lần, đòi hỏi môi trường nội bào cung cấp nguyên kiệu để hình thành nên 4826 nhiễm sắc thể đơn mới. các tế bào con sính ra từ lần nguyên phân cuối cùng đều giảm nhiễm bình thường cho 256 tinh trùng chưa nhiễm sắc thể giới tính Y
d) Bộ nhiễm sắc thể lưỡng bội của loài? số lần nguyên phân liên tiếp của tế bào sinh dục 2n đầu tiên? để tạo ra các tế bào con 2n đã có bao nhiêu thoi dây tơ vô sắc được hình thành trong các lần nguyên phân ấy?
e) Nếu có 3 tinh trùng được thụ tinh với 3 trứng khác nhau tạo ra câc hợp tử thì có bao nhiêu cromait trong các tế bào sinh dục cái sinh ra các trứng đó, vào lúc mà các tế bào bắt đàu thực hiện sự phân bào giảm nhiễm? Các hợp tử vừa được hình thành có bao nhiêu NST đơn?
đồng đều gồm hai nhiễm sắc thê cấu trúc khác nhau, mỗi cặp nhiễm sắc thể tương
f) Giả thiết rằng sự giảm phân xảy ra trong các tế bào sinh dục của cá thể cái, mỗi cặp tương
đồng
đều gồm hai nhiễm sắc thể cấu trúc khác nhau, sự trao đổi đoạn chỉ xảy ra một cặp nhiễm sắc thể thường,sự đột biến dị bội chỉ xảy ra ở cặp nhiễm sắc thể giới tính. Khả năng
cá thể cái có thể cho bao nhiêu loại trứng?
GIẢI
a) Bộ nhiễm sắc thể lưỡng bội - số lần nguyên phân - số thoi vô sắc
- Số lượng tinh trùng X = số lượng tinh trùng Y —> Tổng số tinh trùng hình thành:
256 + 256 = 512
Số tế bào con sinh ra sau các lần nguyên phân : 512 : 4 = 128 = 27
số l ần nguyên phân = 7 4826
- Bộ nhiễm sắc thể 2n = ——— = 38 27 – 1
- Mỗi tế bào sau khi nguyên phân thì hình thành 1 thoi vô sắc + Lần nguyên phân thứ 1: 1 tế bào nguyên phân —> 1 thoi vô sắc
+ Lần nguyên phân thứ 2: 2 tế bào nguyên phân —> 2 thoi vô sắc
+ Lần nguyên phân thứ 3: 4 tế bào nguyên phân —> 4 thoi vô sắc
Vậy tổng số thoi vô sắc là tổng một dãy cấp số nhân với sô shạng đầu a1 = 1 thoi vô sắc
ở tế bào ban đầu
Số hạng của dãy n = số lần nguyên phân x
Công bội q = 2 qx – 1
Sn = a1. ——— = 1(2x – 1 ) = 27 – 1 = 127 q – 1
b) Số cromatit – số NST đơn
- Số tế bào sinh duc cái sinh ra trứng: 3
- Bắt đầu giảm phân, mỗi nhiễm sắc thể đơn của từng tế bào sinh dục cái đầu đã tự
nhân đôi thành 1 NST kép gồm 2 cromatit —> Số cromait của tế bào sinh dục cái lúc
bấy giờ: 2 .83 . 3 = 228
- Số hợp tử tạo thành: 3
- Số nhiễm sắc thể đơn: 38 . 3 =114
c) Số loại trứng
- Cá thể đực cho 2 lạo tinh trùng X và Y => Cá thể đực mang cặp NST giới tính XY.
Vậy cá thể cái mang cặp NST giới tính XX.
- 2n = 38 => 19 cặp gồm 18 cặp NST thường và 1 cặp NST giới tính
+ 18 cặp NST thưpờng trong đó có 1 cặp TĐĐ => 218+1
+ Cặp NST giới tính XX có đột biến dị bội => 2 loại giao tử bất thương (XX và O)
Vậy số loại trưng có thể có:
218+1 . 2 = 220 = 1048576
Bài 7: giả thiết trong các cặp nhiễm sắc thể tương đông của một bộ nhiễm sắc thể lưỡng
bội đều chứa cá cặp gen dị hợp tử và hiện tượng trao đỏi đoạn tại một điểm chỉ xảy ra với một cặp nhiễm sắc thể tương đồng. Cho biết không có hiện tượng đột biến và số loại giao tử đực sinh ra từ các điều kiên trên là 32.
Giả thiết trung bình mỗi kì trong phân bào nguyên phân hết 5 phút, hia lần phân bào (kì trung gian) hết 10 phút, quá trình nguyên phân diễn ra liên tục, các tế bào con sinh ra đều tiếp tục nguyên phân.
c) Bộ nhiễm sắc thể lưỡng bội nói trên ở trạng thái chưa nhân đôi là bao nhiêu?
d) Để hợp tử thực hiện được quả trình nguyên phân thì môi trường nội bào đã cung cấp nguyên kiệu tương đương với bao nhiêu NST đơn vào các thời điểm:
- Kết thúc 20 phút
- Kết thúc 32 phút
- Kết thúc 100 phút
Biết rằng khi hợp tử bước vào kì trước được tính là thời gian bắt đầu
GIẢI
a) Bộ NST lưỡng bội
n = số cặp NST trong bộ NST lưỡng bội
m = số cặp NST có trao đổi đoạn tại một điểm
- Số loại giao tử 2n+mARN
2n+1 = 32 = 25 => n = 4; 2n = 8
b) NST được cung cấp
- thời gian của một chu kì nguyên phân : 4 .5 +10 = 30 phút
- Số NST tương đương với nguyên liệu được cung cấp:
+ Ở thời điẻm 20 phút
Chưa qua 1 chu kì nguyên phân nhưng NST đều đã tự nhân đôi 1 lần: 8(21 – 1) = 8
+ Ở thời điểm 32 phút
Qua 1 chu kì nguyên phân và đang ở kì trước của chu kì thứ 2: NST đã tự nhân đôi 2 lần: 8(22 – 1) = 24
+ Ở thời điểm 100 phút
Qua 3 chu kì nguyên phân và đang ở kì giữa của chu kì thứ 4: NST đã tự nhân đôi 4 lần: 8 (24 – 1) = 120
Bài 8: Ở gà khi quan sát một tế bào sinh dục đực dang ở kì giữa của nguyên phan, người ta đếm được 78 nhiễm sắc thể kép e) Tế bào đó nguyên phân 5 đợt liên tiếp đã đòi hỏi mội trường cung cấp nguyên liệu đẻ tạo ra bao nhiêu nhiễm sắc thể đơn mới ?
f) Loại tế bào này giảm phân binhd thường, khả năng nhiều nhất có thể cho bao nhiêu loại tinh trùng trong trường hợp không có hiện tượng trao đỏi đoạn giữa các nhiêm sắc thể kép trong cặp tương đồng? Điều kiện để cho số loại tinh trùng nhiều nhất là gì?
g) Giả thiết rằng có 1000 tế bào sinh tinh trùng giảm phân bình thường, hiệu 1
suất thụ tinh của tinh trùng là ——, còn của trứng là 20%, mỗi tinh trùng chỉ 1000
thụ tinh với một trứng. Xác định số tế bào sinh trứng.
h) Các hợp tử được tạo thành đã nguyên phân liên tiếp nhiều đợt với số lần bằng nhau, môi truờng nội bào đã cung cấp nguyên kiệu để tạo ra 2184 nhiễm sắc thể đơn mới. Xác định số tế bào con sinh ra và số đợt nguyên phan của mỗi hợp tử.
GIẢI
a) Số NST cung cấp:
- Ở kì giữa của nguyên phan, mỗi NST trong bộ NST 2n của tế bào đều tự nhân đôi thành một NST kép. Đếm được lúc đó có 78 bộ NST kép thì => 2n = 78
- Số NST tương đương với nguyên liệu được cung cấp:
2n (2x – 1 ) = 78(25 – 1 ) = 2418
b) Số loại tinh trùng
- Số loại tinh trùng trong điều kiện không có TĐĐ 2n = 239
- Cho số loại tinh trùng nhiều nhất trong trường hợp không có trao đổi đoạn nói trên, khi mỗi cặp NST phải cho 2 loại tinh trùng => 39 cặp NST cho 239 loại tinh trùng
Muốn vậy 2 NST thuộc cùng một cặp phải có cấu trúc khác nhau
c) Số tế bào sinh trứng
- Số tinh trùng hình thành: 4 . 1000 = 4000
- Số tế bào trứng thụ tinh = số tinh trùng thụ tinh: 4000.1/4000 = 4
- Số tế bào sinh trứng = số trứng hình thành: 4.100/20 = 20
d) Số tế bào con - số đợt nguyên phân
- Số hợp tử tạo thành = số trứng thụ tinh = số tinh trùng thụ tinh = 4
G
ọi x là số đợt nguyên phân của mỗi hợp tử
số nhiễm sắc thể tương đương mội trường nội bào cung cấp
4 .2n(2x – 1) = 2148 => phương trình 4.78(2x – 1 ) = 2148 ; giải ra ta được x = 3
- Số tế bào con sinh ra: 4.23 = 32
Bài 9: Một tế bào sinh duc đực 2n và một tế bào sinh duc cái 2n đều nguyên phân một số
đợt bằng nhau (các tế bào con sinh ra đều tiếp tục nguyên phân). Giả thiết rằng các tế bào con sinh ra từ đợt nguyên phân cuói cùng đều giảm nhiễm cho tổng số 80 giao tử binh thường. Cho biết số lượng nhiễm sắc thê đơn trong các giao tử đực nhiều hơn số lượng nhiễm sắc thể đơn trong các giao tử cái là 192.
3. Loài đó tên là gì?
4. Mô tả hình dạng và số lượng bộ nhiêm sắc thể lưỡng bộ trong loài đó
GIẢI
1. Tên loài
Gọi 2n: Bộ NST lưỡng bội của loài; x: số lần nguyên phân
Số giao tử đực: 4.2x; số giao tử cái: 2x
- Tổng số giao tử: 4.2x + 2x = 80 (1)
- Số NST trong các giao tử đực nhiều hơn sô NST trong các giao tử cái:
n . 4 . 2x – n . 2x = 192 (2) Giải ra ta được x = 4 => 2n = 8 ; đây là bộ NST của ruồi giấm
c) Mô tả bộ NST
Khi NST xoắn tối đa:
- 3 cặp NST thường: 2 cặp hình chữ V, 1 cặp hình hạt
- 1 cặp NST giới tính: 2 chiếc hình que XX ở con cái, chiếc hình que X, chếc hình móc Y ở con đực
Bài 10: Một gà mái đẻ được một số trứng, nhưng khi ấp chỉ có 12 trứng nở thành gà con. Các hợp tử nở thành gà con có 936 nhiễm sắc thể đơn ở trạng thái chưa tự nhân đôi.
Số trứng còn lại không nở thành gà con.
số tinh trùng được sinh ra phuc vụ cho gà giao phối có 624000 nhiễm sắc thể đơn. Giả thiết số tinh trùng được trực tiếp thụ tinh với các trứng nói tren chiếm tỉ lệ 1/1000 so với tổng số tinh trùng được hình thành. Mỗi tinh trùng thụ tinh với 1trứng
a) Số trứng được thụ tinh
b) Trúng gà không nở thành gà con có bộ nhiễm sắc thể như thế nào?
c) Số gà trống và gà mái trong đàn gà con nói trên có tuân theo tỉ lệ 1: 1 không?
GIẢI
a) Số trứng được thụ tinh
- Bộ NST lưỡng bội của gà = số NST trong mỗi hợp tử : 2n = 936/12 = 78
- Số tinh trùng được hình thành 62400 : (78 : 2) = 16000
- Số trứng thụ tinh = số tinh trùng thụ tinh 16000. 1/1000 = 16
b) Bộ NST của trứng không nở
Ở gà trứng thụ tinh hoặc không đều được đẻ ra
- Trứng không được thụ tinh thì luôn luôn không nở => bộ NST n = 39
- Trứng được thụ tinh nhưng không gặp điều kiện thuận lợi của mọi trường ấp thì không nở => bộ NST 2n = 78
c) Tỉ lệ trống mái
- Đàn gà con có thể gồm: 6 gà trống và 6 gà mái tuân theo tỉ lệ 1:1
- Đàn gà con có thể có số gà trống không bằng số gà mái không tuân theo tỉ lệ 1:1. Bởi vì tỉ lệ này chỉ nghiệm đúng trên số lượng lớn cá thể
Một số nội dung nâng cao
Nội dung1. SO SÁNH CẤU TRÚC, ĐẶC ĐIỂM DI TRUYỀN CỦA SVNS VÀ SVNT
CẤU TRÚC CHUNG CỦA GEN CẤU TRÚC
Vùng điều hoà Vùng mã hoá (Bộ mở đầu…....mã hóa…….kết thúc)
* Lưu ý: Bộ ba mở đầu, bộ ba kết thúc nằm trong vùng mã hóa
1. Cấu trúc gen
- Gen không phân mảnh
- Gen đa cistron
2. Nhân đôi AND - Ở sinh vật nhân sơ chỉ có 1 điểm khởi đầu sao chép (Ori C), - Hệ enzim đơn giản hơn: (1). Protein B: Nhận biết điểm khởi đầu tái bản

(2). Gyraza (còn được gọi là topoisomeraza II): Làm duỗi thẳng phân tử DNA.
(3). Hêlicaza (helicase): dãn xoắn và tách hai mạch đơn do cắt các liên kết hydro.
(4). Prôtêin SSB: giúp hai mạch đơn không bị dính
(5). RNA polymeraza (Primaza): Tổng hợp đoạn mồi.
(6). DNA polymeraza III: lắp ráp nu, kéo dài mạch đơn mới.
(7). DNA polymeraza I: cắt RNA mồi, tổng hợp mạch polinucleotide mới.





(9). DNA polymeraza II: sửa sai sau khi nối các đoạn okazaki.
(10). Ligaza: nối các đoạn okazaki.
(11). Primaza (RNA polymeraza): Tổng hợp đoạn mồi.

(12). Ngoài ra còn có Telomeraza: hạn chế sự cố đầu mút. Chỉ có trong tinh hoàn và buồng trứng, ở tất cả các tế bào sinh dưỡng enzim này không hoạt động
- Nhìn chung, tốc độ nhân đôi ở sinh vật nhân sơ nhanh hơn ở sinh vật nhân thực
Vùng kết thúc
- Gen không phân mảnh
- Gen đơn cistron
- Đoạn Okazaki dài 1000- 2000 nucleoti - Đoạn Okazaki kích thước lớn hơn
3. Dịch mã
- Do hệ gen lớn, nên có rất nhiều điểm khởi đầu tái bản.
- Hệ enzim tham gia phức tạp hơn so với nhân sơ. Hệ enzim ADN pol có nhiều loại alpha, beta, gama... và cơ chế hoạt động phức tạp hơn.
- Phiên mã đến đâu riboxôm tiến đến và quá trình dịch mã diễn ra tới đó
-
- Gen thuôc dạng đa Cistron nên một Riboxom trược qua có thể tạo ra nhiều loại Protein khác nhau
- Trước khi dịch mã Axitamin mở đầu là Metionin được kết hợp với axit foocmic để tạo thành fmet
- Sau khi phiên mã mARN mới tạo ra phải trải qua giai đoạn sửa đổi: Cắt bỏ các đoạn Intron nối các đoạn Exon để tạo ra mảN trưởng thành. Các mARN trưởng thành phải di chuyển ra TBC lúc đó Riboxom mới đến tiếp xúc để thực hiện dịch mã
- Gen thuộc dạng đơn Cistron nên 1 Riboxom trượt qua 1 phân tử mARN chỉ tạo ra một phân tử Protein
- Trước khi dịch mã Axitamin mở đầu là Metionin không kết hợp với axit foocmic
- 40 -
- Cơ chế điều hòa phức tạp, diễn ra qua nhiều giai đoạn khác nhau + Điều hòa nhân đôi AND + Điều hòa phiên mã và
phiên mã
+ Điều hòa vận chuyển: Có cho
mARN ra khỏi nhân hay không
+ Điều hòa dịch mã và sau dịch mã
- Chứa và truyền đạt thông tin di truyền: Đòi hỏi trước tiên đối với vật chất DT là có khả năng chứa thông tin DT. ADN có chiều ngang thì giới hạn nhưng chiều dài thì không hạn chế. Trình tự sắp xếp các Nu theo chiều dài có thể phản ánh những thông tin nhất định. ADN có 4 loại Nu nhưng số trình tự khác nhau là con số khổng lồ. Khả năng chứa thông tin đó làm cho phân tử ADN là phân tử dài nhất trong tự nhiên. Ngoài ra nó còn được mở rộng do các thay đổi cấu trúc, do gãy nối lại và lắp ghép giữa các đoạn ADN. Thông tin chứa trên ADN được sử dụng và hiện thực hóa nhờ các chất trung gian ARN, rồi đến tổng hợp các protein là những công cụ phân tử thực hiện chức năng của TB.
- Tự sao chép chính xác:
Mô hình Watson - Crick cũng thỏa mãn ở mức lí tưởng yêu cầu thứ 2 của vật chất di truyền. Chuỗi xoắn kép gồm 2 sợi bổ sung cho nhau theo nguyên tắc A-T và G-X , làm cho phân tử như có một bản âm và một bản dương. Mỗi bản có thể làm khuôn tạo ra bản kia để từ một phân tử ban đầu có 2 phân tử giống hệt nó.
- Có khả năng biến dị di truyền:
Trên phân tử ADN có thể xảy ra nhiều biến đổi. Các biến đổi có thể được di truyền. VD: cặp
Nội dung2: Trình bày những những tiêu chuẩn của vật chất di truyền?
Vật chất DT trong TB và cơ thể phải có đầy đủ các tiêu chuẩn cơ bản sau đây:
1. Mang TTDT, đặc trưng của từng loài:
Tính chất này bao gồm:
*Tính chất của gen cấu trúc:
VCDT phải có khả năng, bằng cách này hay cách khác, đặc trưng cho cấu trúc của các phân tử khác trong TB.
VD: Nếu thừa nhận các gen nào đó qui định cấu trúc protein, thì VCDT phải có khả năng mã hoá một cách chính xác một khối lượng lớn các đoạn khác nhau mang axit amin trong các chuổi polypeptit.
*Tính chất của gen điều hoà:
Gen điều hoà chứa thông tin đáp ứng yêu cầu điều hoà hoạt động trong cơ thể. Một số vùng của ADN trên Operon, có gen điều hoà và gen khởi động, hoạt động như "khoá hãm" để ngắt hoặc mở hoạt động của gen.
Các đặc điểm riêng của từng loài, như phát triển phôi, phân hoá cơ quan, có sai khác giữa loài này với loài khác, đều được chương trình hoá về mặt DT.
VCDT phải có khả năng, bằng nhiều cách, điều hoà hoạt động của gen.
2. Tính tự sao, có khả năng tái bản một cách chính xác:
Đây là khả năng hình thành các bản sao, truyền lại cho TB con trong quá trình phân bào, với tất cả TTDT của loài chứa trong đó.
Qua quá trình nguyên phân, mỗi TB con được hình thành, nhận một bản sao toàn bộ TTDT.
Qua giảm phân VCDT giảm đi một nửa ở mỗi giao tử.
3. TTDT chứa trong VCDT phải được sử dụng để tạo ra những phân tử cần thiết cho TB.
4. VCDT phải có khả năng biến đổi: Cơ chế tự sao của VCDT có thể chính xác đến mức các sai sót hiếm xảy ra, nhưng không phải là tuyệt đối chính xác để không xảy ra đột biến.
Chính ĐB của VCDT là một nguồn chủ yếu của biến dị, rất cần cho tiến hoá của các loài.
Nội dung3: ADN có những đặc điểm gì để thoả mãn là vật mang TTDT?
*ADN thỏa mãn các yêu cầu đối với vật chất di truyền:
A-T trên ADN được thay bằng cặp G-X thì sự thay thế được truyền cho các phân tử con.
- Có tiềm năng tự sửa sai:
Các nhà di truyền học còn phát hiện thêm một tính chất nữa của ADN là tiềm năng tự sửa sai. Do cấu trúc mạch kép nên sai hỏng ở một mạch có thể bị cắt bỏ và dựa vào mạch nguyên vẹn để làm khuôn tổng hợp lại cho đúng.
- Có khả năng tạo ra một số sản phẩm phục vụ cho hoạt động sống của TB
Nội dung 4: Những bằng chứng để chứng minh ADN là vật mang TTDT:
* Các bằng chứng để chứng minh ADN là chất di truyền: a. Các chứng minh gián tiếp: ADN có trong tất cả TB sinh vật, chỉ giới hạn trong nhân và là thành phần chủ yếu của NST. Một cấu trúc TB mang nhiều gen xếp theo đường thẳng. Tất cả các TB sinh dưỡng của bất kì một loại sinh vật nào đều chứa một lượng ADN rất ổn định không phụ thuộc vào sự phân hóa chức năng hay trạng thái trao đổi chất. Ngược lại ARN biến đổi tùy thuộc vào trạng thái sinh lí TB.
Số lượng ADN tăng theo số bội thể của TB ở TB đơn bội ADN số lượng ADN là 1 thì TB sinh dưỡng lưỡng bội số lượng ADN tăng gấp đôi.
Tia tử ngoại có hiệu quả gây đột biến cao nhất ở bước sóng 260nm; đây chính là bước sóng mà ADN hấp thụ tia tử ngoại nhiều nhất. b. Hiện tượng biến nạp: Truyền thông tin di truyền nhờ ADN.
Hiện tượng biến nạp được Gripffith phát hiện ở Diplococcus Pneumoniae vào năm 1928. Vi khuẩn này có 2 dạng khác nhau:

- Dạng S, gây bệnh có vỏ TB bằng polisaccharid cản trở bạch cầu phá vỡ TB. Dạng này có khuẩn lạc láng mọc trên môi trường aga.
- Dạng R, không gây bệnh, không có vỏ bao, tạo khuẩn lạc nhăn.
Thí nghiệm biến nạp ở chuột.
+ Tiêm vi khuẩn S sống gây bệnh cho chuột → chuột chết.
Tiêm vi khuẩn R sống không gây bệnh → chuột sống.
Tiêm vi khuẩn S bị đun chết cho chuột → chuột sống.
Hỗn hợp vi khuẩn S bị đun chết trộn với vi khuẩn R sống đem tiêm cho chuột → chuột chết.
Trong xác chuột có vi khuẩn S và R.
Hiện tượng trên cho thấy vi khuẩn S không thể tự sống lại được sau khi bị đun chết, nhưng các TB chết này đã truyền tính gây bệnh cho TB R. Hiện tượng này gọi là biến nạp.
Năm 1944, T.Avery, Mc Leod và Mc Carty đã tiến hành thí nghiệm xác định rõ tác nhân gây biến nạp. Nếu sử lý TB S bằng proteaza hoặc ARNaza hoạt tính biến nạp vẫn còn. Chứng tỏ protein và ARN không phải là tác nhân gây biến nạp. Nhưng nếu TB S chết bị sử lí bằng
ADNaza thì hoạt tính biến nạp không còn nữa. Chứng tỏ ADN là tác nhânbiến nạp. Kết quả thí nghiệm có thể tóm tắt như sau:
ADN của TB S + các TB R sống → chuột → chuột chết (có cả S +R).
CHƯƠNG 2. TÍNH QUI LUẬT CỦA HIỆN TƯỢNG DI TRUYỀN
A. CƠ SỞ LÝ THUYẾT
I. một số khái niệm và thuật ngữ
1. Phương pháp phân tích giống lai:
- Lai các cặp bố mẹ thuần chủng khác nhau về 1 hoặc vài cặp tính trạng, theo dõi riêng con cháu của từng cặp bố mẹ
- Dùng toán thống kê và lý thuyết xác suất để phân tích QLDT các tính trạng của bố mẹ cho các thế hệ sau.
2.Tính trạng là những đặc điểm cụ thể về hình thái, cấu tạo sinh lý riêng cho 1 cơ thể nào đó, giúp ta dễ dàng phân biệt nó với những cơ thể khác. Vd:đậu thân cao, hạt vàng, ...
a. Tính trạng tương ứng là những biểu hiện khác nhau của cùng 1 loại tính trạng.VD:hạt vàng, hạt xanh,hạt đen, hạt trắng.
b. Tính trạng tương phản là 2 tính trạng tương ứng có biểu hiện trái ngược nhau.VD: hạt màu đen và hạt màu trắng.
3. Cặp gen tương ứng:là cặp gen nằm ở vị trí tương ứng trên cặpNST tương đồng và quy định cặp tính trạng tương ứng .VD: cặp gen tương ứng A,a nằm ở vị trí đối diện trên cặp NST số 5 ở cây cà chua quy định dạng quả không múi hoặc có múi.
4. Alen là những trạng thái khác nhau của cùng 1 gen. Alen này khác alen kia ở 1 hoặc một số cặp nu nào đó, sản phẩm của quá trình đột biến gen.
5. Kiểu gen là tổ hợp toàn bộ các gen trong tế bào cơ thể. Thông thường khi nói tới KG của 1 cơ thể người ta chỉ xét 1 vài xặp gen nào đó liên quan với các tính trạng đang được quan tâm .VD:AABB, AaBB..
6. Kiểu hình là toàn bộ các tính trạng của cơ thể. Thưc tế khi nói tới kiểu hình của 1 cơ thể, người ta chỉ xét 1 vài tính trạng mà người ta quan tâm .VD: Đậu thân cao, hoa trắng, hạt vàng .
7. Thể đồng hợp: Thể đồng hợp về một gen nào đó là trường hợp 2 alen trong cặp tương ứng giống nhau.VD:aa, BB,...
8. Thể dị hợp: Thể dị hợp về 1 gen nào đó là trường hợp 2 alen trong cặp tương ứng là khác nhau.VD:Aa, Bb,...
9. Giống thuần chủng là giống có đặc tính di truyền đồng nhất và ổn định, các thế hệ con cháu không phân ly, có KH giống bố mẹ. Trên thực tế, nói tới giống thuần chủng là nói tới sự thuần chủng về 1 hoặc vài tính trạng nào đó đang được nghiên cứu, cặp gen quy định tính trạng này là đồng hợp.
10. Một số ký hiệu thường dùng:
P: cặp bố mẹ xuất phát, x : Ký hiệu cho phép lai.
II. Quy luật phân ly
1. Phương pháp nghiên cứu khoa học của MenĐen
1.1. Đối tượng nghiên cứu : đậu Hà Lan
Vậy hiện tượng biến nạp là một minh chứng sinh hóa xác nhận rằng ADN mang tín hiệu di truyền. -
G: giao tử
F: thế hệ con.
1.2. Tạo các dòng thuần : Bằng phương pháp tự thụ phấn bắt buộc qua nhiều thế hệ
1.3. Theo dõi sự di truyền của từng tính trạng
1.4. Sử dụng toán thống kê và lý thuyết xác suất
1.5. Tuân thủ một quy trình nghiên cứu khoa học độc đáo
- Quan sát thu thập số liệu: Lai các dòng thuần chủng khác nhau về kiểu hình tương phản rồi theo dõi tỉ lệ phân li kiểu hình ở các đời con
- Đưa ra giả thuyết khoa học giải thích những gì quan sát được:
c
- Làm thí nghiệm kiểm tra tính đúng đắn của giả thuyết mà mình đưa ra: Làm thí nghiệm lai phân tích để kiểm tra tính đúng đắn của giả thuyết
2. Nội dung.
2.1.Thí nghiệm:SGK
2.2. Giải thích: Mỗi tính trạng ở cơ thể do 1 cặp nhân tố DT quy định mà sau này gọi là gen. Sự phân ly và tổ hợp của các nhân tố DTđã chi phối sự DT và biểu hiện của các cặp tính trạng tương phản qua các thế hệ
2.3. Quy luật phân ly: Mỗi tính trạng được quy định bởi 1 cặp alen. Do sự phân ly đồng đều
ủa cặp alen trong giảm phân nên mỗi giao tử chỉ chứa 1 alen của cặp.
3.Cơ sở tế bào học Trong tế bào sinh dưỡng, NST tồn tại thành từng cặp, do đó gen cũng tồn tại thành từng cặp alen tương ứng trên cặp NST tương đồng.
Khi giảm phân mỗi NST trong cặp phân ly về mỗi giao tử. Vì vậy mỗi giao tử chỉ mang
A hoặc a. Sự tổ hợp của cặp NST tương đồng trong thụ tinh hình thành F1 có KG Aa.
F1 giảm phân hình thành nên 2 loại giao tử A và ađược tạo thành với xác suất ngang nhau. Sự thụ tinh của 2 loại giao tử đực và cái tạo F2 có tỷ lệ KG:1Aa:2Aa: 1aa. Vì A trội át hoàn toàn alen lặn a nên thể đồng hợp trội và thể dị hợp có KH như nhau. Do đó F2 có tỷ lệ
3 đỏ: 1 trắng, tính trạng lặn được biểu hiện ở thể đồng hợp lặn.
4. Cơ sở phân tử của tính trội - lặn
4.1. Cơ sở phân tử của tính trạng lặn Một alen đột biến nào đó được coi là lặn khi cá thể dị hợp tử về alen đột biến có kiểu hình giống như kiểu hình của cá thể có kiểu gen đồng hợp trội. Nói cách khác, trường hợp
cơ thể chỉ cần 1 alen cũng tạo ra đủ sản phẩm để duy trì 1 chức năng sinh học nhất định thì gen đó khi bị đột biến thường tạo ra các alen lặn. Xét trường hợp sản phẩm của gen là 1 E xúc tác cho 1 phản ứng nhất định trong tế bào, khi có 2 alen bình thường thì lượng E được tạo ra trong tế bào sẽ cao gấp đôi so với trường hợp ở cơ thể có kiểu gen dị hợp. Tuy nhiên trong nhiều trường hợp, E không hoạt động với tốc độ cực đại khi nồng độ cơ chất trong tế bào ở mức độ bình thường. Cụ thể là khi cơ thể có kiểu gen đồng hợp trội thì lượng E tạo ra nhiều nên cơ chất sẽ chuyển ngay thành sản phẩm và các E chỉ cần hoạt động ở tốc độ bình thường, còn ở thể dị hợp thì lượng E chỉ bằng 1 nửa so với cơ thể đồng hợp nên cơ chất của E được tích tụ lại trong tế bào ở mức cao hơn bình thường, do đó làm cho E hoạt động ở tốc độ tối đa. Như vậy, dù chỉ có 1 lượng E bằng 1 nửa so với bình thường nhưng do hoạt động
ở tốc độ tối đa nên lượng sản phẩm tạo ra vẫn duy trì ở mức bình thường như ở các cơ thể có
kiểu gen đồng hợp trội. Sơ đồ như sau:
Cơ thể có kiểu gen AA: S E P Kiểu hình bình thường
Cơ thể có kiểu gen Aa: 2S 0,5E P Kiểu hình bình thường
Có thể nói phần lớn các bệnh di truyền do gen lặn gây nên ở người đều thuộc loại thiếu hụt E như: Bệnh tay - sach, bênh phenilketo niệu...
4.2. Cơ sở phân tử của tính trội
Ví dụ: 1 alen bình thường tạo ra sản phẩm là chất kích thích phân chia tế bào vốn chỉ tồn tại trong 1 thời gian ngắn nay bị đột biến tạo ra sản phẩm có thời gian tồn tại lâu hơn nhiều so với sản phẩm của alen bình thường. Do thời gian tồn tại lâu nên chúng kích thích tế bào đích phân chia nhiều hơn mức bình thường dẫn đến hình thành tế bào khối u. Trường hợp này alen đột biến được xem là alen trội.
b) Sự thiếu hụt sản phẩm gen ở các cơ thể dị hợp tử (thiếu hụt đơn bội).
Một loại bệnh di truyền trội phát sinh do các enzim bị đột biến làm cho hoạt tính của nó chỉ còn bằng một nửa hoạt tính của enzim bình thường nên không đủ để tạo ra một kiểu hình bình thường. Hiện tượng này gọi là sự thiếu hụt đơn bội.
- Nhiều con đường chuyển hóa thường có bước hạn chế tốc độ, tại đó enzim xúc tác cho phản ứng hoạt động ở mức cực đại hoặc gần cực đại đối với những cơ thể có hai alen bình thường. Nếu alen đột biến không tạo ra sản phẩm (một enzim nào đó), thì gen còn lại không thể tạo ra đủ enzim để chuyển hóa cơ chất ngay cả khi nồng độ cơ chất đã tăng cao.
VD: Bệnh chuyển hóa porphyrin cấp tính từng cơn ở người
c) Alen đột biến làm tăng hoạt tính enzim
Khi hoạt tính của một enzim tăng lên sẽ làm cho nồng độ cơ chất của enzim đó giảm quá mức. Nếu cơ chất của enzim đó được sử dụng cho nhiều phản ứng khác nhau thì việc giảm nồng độ của cơ chất này có thể gây trở ngại cho chức năng bình thường vì cơ thể sẽ thiếu một số sản phẩm khác và cho vậy alen đột biến sẽ là trội.
Một số đột biến làm tăng hoạt tính của enzim và kéo theo làm tăng cả sản phẩm của phản ứng. Nếu việc tăng nồng độ của sản phẩm lại làm rối loạn cơ chế ức chế ngược thì đột biến này cũng được xem như đột biến trội vì sản phẩm của đột biến hoạt động như một chất cơ hại cho cơ thể
VD: Enzim PRPP điều khiển tốc độ tổng hợp purin được qui định bởi một gen nằm trên NST X. Khi một người phụ nữ bị hội chứng 3X, có hoạt tính của enzim PRPP cao rõ rệt, làm cho lượng purin tăng lên nhiều hơn lượng cần thiết sử dụng cho quá trình đồng hóa. Khi đó, lượng purin dư thừa đã nhanh chóng bị khử thành axit uric. Axit uric chỉ bị hòa tan một phần, một phần bị kết tủa tạo nên những tinh thể gây sưng viêm, gây đau chủ yếu ở khớp xương và các mao mạch ngoại vi đối với những người bị bệnh gút.
d) Các đột biến biểu hiện nhầm
Đây là loại đột biến làm cho một gen nào đó được biểu hiện nhầm vị trí hoặc nhầm thời điểm mà đáng ra nó không được biểu hiện. Kiểu đột biến này ảnh hưởng lên các yếu tố điều khiển quá trình phiên mã của gen nhiều hơn là tác động lên cấu trúc của một chuỗi polipeptit. Kết quả hoặc là tạo ra sự biểu hiện sai lệch về vị trí trong cơ thể (sản phẩm của gen được tổng hợp trong những tế bào không cần sản phẩm của gen đó) hoặc là sai lệch về thời điểm biểu hiện kiểu hình (sản phẩm của gen biểu hiện đúng vị trí nhưng không đúng thời điểm). Loại đột biến này là những đột biến trội.
VD: ở ruồi giấm Drosophila, đột biến ở gen bithorax làm cho con ruồi xuất hiện bốn cánh thay vì hai cánh. Hay đột biến ở gen antennapedia làm xuất hiện chân trên đầu nơi là đáng ra sẽ phát triển ăngten.
Bài tập vận dụng: Bài 4 trang 45(SGK NC)
a) F1 đồng tính, F2 có tỷ lệ 3:1 => lông xám là tính trạng trội, lông trắng là tính trạng lặn.
Quy ước: Xám : A; trắng : a.
a) Các alen đột biến tạo ra sản phẩm có hại Nếu sản phẩm của alen đột biến trực tiếp hoặc gián tiếp gây hại làm cho kiểu hình của alen trội được biểu hiện ngay cả khi cơ thể chỉ có 1 alen đột biến thì alen đó được xem là trội -
Ta có SĐL:
P t/c: Xám x Trắng
AA aa
F1: Aa (xám) x Aa ( xám)
G: 1A : 1a 1A : 1a
F2:
- Tỷ lệ kiểu gen 1AA : 2Aa : 1 aa
- Tỷ lệ kiểu hình: 3 xám : 1 trắng.
b) F1: Xám x Trắng
Aa aa (Học sinh tự viết)
Bài5 trang 45 (SGK NC)
T
ừ cặp 4 và 5 => lông đen là trội , lông trắng là lặn.
Quy ước: A - đen; a - trắng.
Ta có con 1, con 3, con 6 cùng có KG là aa. Các con (2) và (5) có KG Aa.
C. Quy luật phân ly độc lập
I. Nội dung:
1. Thí nghiệm :SGK
2. Nhận xét: Xét riêng từng tính trạng ở F2 :
Vàng / xanh = 3: 1
Trơn / nhăn = 3: 1
vàng , trơn là những tính trạng trội
xanh , nhăn là những tính trạng lặn Kết quả phân tích cho thấy xác suất xuất hiện mỗi KH ở F2 bằng tích xác suất của các tính trạng hợp thành nó. Từ đó Men Đen thấy các cặp tính trạng di truyền độc lập với nhau
3.Giải thích: Mỗi cặp tính trạng do 1 cặp nhân tố DT (gen) quy định. Các cặp nhân tố
này đã PLĐLvà tổ hợp tự do trong quá trình phát sinh giao tử và thụ tinh đã chi phối sự
DT và biểu hiện của các tính trạng .
4.Nội dung:SGK
II. Cơ sở tế bào học: Mỗi cặp alen quy định 1 cặp tính trạng nằm trên 1 cặp NST tương đồng. Trong quá trình phát sinh giao tử F1 có sự PLĐL của các cặp NST tương đồng dẫn tới sự PLĐL của các gen tương ứng tạo nên các loại giao tử khác nhau với xác suất ngang nhau. Các loại giao tử này kết hợp ngẫu nhiên với xác suất ngang nhau trong thụ tinh tạo nên F2.
SĐL: SGK
III. Công thức tổng quát: SGK
IV. Ý nghĩa của các quy luật Men Đen:
Quy luật phân ly của Men Đen có ứng dụng thực tế là nếu biết được các gen quy định
các tính trạng nào đó phân ly độc lập thì có thể dự doán được kết quả phân ly kiểu hình ở đời sau.
Quy luật phân ly độc lập cho thấy quá trình sinh sản hữu tính sẽ tạo ra một số lượng lớn biến dị tổ hợp, tạo ra nhiều tổ hợp gen khác nhau
V. Sử dụng phép thử X2 trong đánh giá tỉ lệ phân li kiểu hình
- Để xác định số lượng cá thể của các loại kiểu hình ở một phép lai có đúng với tỉ lệ phân li Menđen như dự đoán hay không thì chúng ta phải sử dụng phép thử X2.
- Giả sử trong phép lai giữa đậu Hà lan hoa tím, hạt tròn với đậu hoa trắng, hạt nhăn được F1 có tỉ lệ phân li kiểu hình như sau: 140 cây hoa tím, hạt tròn
135 cây hoa trắng, hạt nhăn
110 cây hoa tím, hạt nhăn
115 cây hoa trắng, hạt tròn
Tỉ lệ phân li kiểu hình trên có đúng là 1 : 1: 1:1 hay không ?
- Đầu tiên ta xây dựng giả thuyết Ho, cho rằng tỉ lệ phân li kiểu hình trong phép lai là 1:1:1:1 và sự sai khác mà ta thu được trong phép lai là hoàn toàn do các yếu tố ngẫu nhiên.
Sau đó ta tính giá trị X2 theo công thức: ( O E )2
Trong đó : O là số liệu quan sát (tỉ lệ phân li kiểu hình của phép lai) E là số liệu lí thuyết (tỉ lệ phân li kiểu hình theo lí thuyết)
- Ta có thể lập bảng tính X2 cho trường hợp trên như sau:
Tỉ lệ kiểu hình O E (O – E)2 (O – E)2 E
Tím, tròn 140 125 225 1,8
Trắng, nhăn 135 125 100 0,8
Tím, nhăn 110 125 225 1,8
Trắng, tròn 115 125 100 0,8
Tổng 500 500 X2 = 5,2
- Để biết được sự sai khác giữa số liệu lí thuyết và thực nghiệm có hoàn toàn do các lí do ngẫu nhiên hay không ta cần tra bảng phân bố của các giá trị X2 ở các mức xác suất khác nhau tương ứng với số bậc tự do
- Từ kết quả thu được X 2 = 5, 2 đối chiếu trên bảng phân bố giá trị X 2
- P chỉ mức xác suất, người ta thường dùng là 0,05 còn n là số bậc tự do (số loại kiểu hình) trừ 1. (4 loại kiểu hình trừ 1 = 3)
- Giá trị trong bảng là 7, 815. Ta thấy giá trị X 2= 5, 2 nhỏ hơn thì ta chấp nhận giả thuyết trên nghĩa là tỷ lệ 140:135:110:115 tương ứng với tỷ lệ 1:1:1:1
- Còn nếu giá trì X 2 lớn hơn thì kết quả thực nghiệm không đáng tin cậy. Sự sai khác giữa thực nghiệm và lý thuyết không phải là do yếu tố ngẫu nhiên mà có thể do 1 nguyên nhân nào đó.
Bài tập vận dụng:
Bài tập 1: Trong 1 phép lai người ta thu được tỷ lệ 165 quả tròn: 28 quả dài. Đây có phải là tỷ lệ 3:1 hay không ?
- Số lượng nghiên cứu là 165 + 28 = 193. Nếu là tỷ lệ 3: 1 thì số lượng chiếm tỷ lệ 3 theo lý thuyết là [193: (3+1)] X 3 = 145 và số lượng chiếm tỷ lệ 1 là 193 – 145 = 48.
- Lập bảng tính X 2
Tỷ lệ kiểu hình O
( O – E )
Quả dài 28 48 400 8, 33
193 193 X 2 = 11,09
- Đối chiếu với bảng phân bố giá trị X 2 là 3, 481. Như vậy kết quả X 2 tính được (X 2 = 11, 09) lớn hơn ( 3,481)→ Đây không phải là tỷ lệ 3 : 1.
Bài tập 2: Trong 1 phép lai giữa các cây đậu Hà lan người ta thu được 315 hạt vàng, trơn:
108 hạt vàng, nhăn: 101 hạt xanh, trơn: 32 hạt xanh, nhăn. Đây có phải là tỷ lệ 9:3:3:1
không ? (Mức độ tin cậy của tỷ lệ trên)
( Bài này học sinh tự làm)
Bài 3 SGK trang 49
a) F2 có đen : trắng = 3: 1
ngắn: dài = 3: 1
tính trạng đen, ngắn là những tính trạng trội trắng , dài là những tính trạng lặn
Quy ước: Đen: A Ngắn: B Trắng: a Dài: b
Ta thấy tỷ lệ phân ly KH ở F2 là 9:3:3:1=(3:1)(3:1)=> các tính trạng trên DT theo
quy luật PLĐL
Ta có SĐL: học sinh tự viết
b) tương tự ý (a), học sinh xét tỷ lệ phân ly từng tính trạng ở F3=> KG của từng phép
lai rồi viết SĐL
D. Sự tác động của nhiều gen và tính đa hiệu của gen
I. Tác động của nhiều gen lên một tính trạng
1. Tác động bổ sung giữa các gen không alen
a) Ví dụ: SGK
b) Giải thích: F2 có 16 tổ hợp = 4 x 4 => F1 dị hợp về 2 cặp gen (AaBb)
Ta có F1: AaBb x AaBb
GF1: 1AB: 1Ab : 1aB:1ab 1AB: 1Ab : 1aB:1ab
KG: F2: 9 A-B- : 3A-bb : 3aaB- : 1aabb
KH: 9 đỏ : 7 trắng .
Kiểu tương tác: Màu hoa do 2 cặp gen không alen quy định, trong đó sự có mặt của 2 loại gen trội A và B => đỏ thẫm, sự có mặt của 1 trong 2 loại gen trội A hoặc B => trắng, không có gen trội nào => trắng .
Ngoài ra tương tác bổ sung này còn có các tỉ lệ khác như:9:3:3:1, 9:6:1
Kiểu bổ sung này có thể giải thích qua sơ đồ dưới đây: Gen A Gen B
Enzim A Enzim B
Chất A (trắng) Chất A (trắng) Sản phẩm P (đỏ)
Cây dị hợp Aa chỉ cần một alen A đã tổng hợp đủ một lượng enzim cần thiết để chuyển chất A thành chất B. Chỉ cần một alen B cũng đủ tạo ra lượng enzim cần thiết chuyển chất B thành sản phẩm B (màu đỏ). Cây có kiểu gen aaBB không sản xuất được enzim chuyển hóa chất A thành B nên dù có tạo ra được enzim b cũng không có cơ chất (chất B) để chuyển
thành sản phẩm P, nên hoa của chúng có màu trắng. Tương tự cây có kiểu gen Aabb chỉ dừng lại ở việc tổng hợp chất B màu trắng tích lũy lại trong tế bào nên hoa có màu trắng và cây aabb cho hoa màu trắng vì không thể tạo ra được chất P.
T
ỉ lệ 9:7 trên đây có thể giải thích cách khác là do 2 gen lặn có tác dụng át chế. Sự hỗ trợ hoặc làm gián đoạn chuỗi phản ứng cho hiệu quả tương tác gen.Ví dụ các gen góp thêm cho đủ phản ứng thì có tác động bổ trợ, làm gián đoạn thì gây át chế
2. Tác động cộng gộp
a) Ví dụ: SGK
b) Giải thích tương tự như trên
Kiểu tương tác: Màu đỏ đậm nhạt khác nhau tùy thuộc vào số lượng gen trội, số lượng gen trội trong KG càng nhiều thì màu đỏ càng đậm, mỗi gen đóng góp 1 phần như nhau vào sự biểu hiện của tính trạng
3. Tác động át chế (1gen này kìm hãm hoạt động của 1 gen khác không cùng locut)
a. Ví dụ: Ptc: Ngựa xám x Ngựa hung
F1: Xám
F2: 12 xám : 3 đen : 1 hung
b. Giải thích: tương tự như trên
Kiểu tương tác: B át C: đen b không át c: hung
GV có thể giải thích kiểu átchế do gen trội C. Ngoài ra còn có kiểu át chế theo tỷ lệ
13:3; 9:3:4
II. Một gen chi phối nhiều tính trạng
1. Ví dụ: Khi lai đậu Men Đen thấy: giống đậu hoa tím thì hạt có màu nâu, nách lá có chấm đen, thứ hoa trắng có hạt màu nhạt, nách lá không có chấm.
Khi nghiên cứu ruồi giấm Mooc gan thấy gen quy định cánh cụt đồng thời quy định 1 số tính trạng khác như đốt thân ngắn, lông cứng, tuổi thọ ngắn
2. Ý nghĩa: Gen đa hiệu là cơ sở để giải thích hiện tượng biến dị tương quan. Khi 1 gen đa hiệu bị đột biến thì nó sẽ đồng thời kéo theo sự biến dị ở 1 số tính trạng mà có chi phối.
Bài tập vận dụng: Bài 4 trang 53 SGK
F2 có tỷ lệ: 9 dẹt:6 tròn: 1 dài→F2 có 16 tổ hợp=4x4→F1dị hợp về 2 cặp gen→ hình dạng quả bí bị chi phối bởi sự tương tác của 2 gen không alen , trong đó:
Kiểu gen có mặt 2 loại gen trội A và B cho quả dẹt, có mặt 1 trong 2 loại gen trội A hoặc B cho quả tròn, có mặt toàn gen lặn cho quả dài.
Bài 5 trang 53 SGK Ở gà: C: lông màu I: át chế màu
c:lông trắng i : không át màu
Ptc: Lông màu x Lông trắng
Ccii ccI I
F1: CcIi ( lông trắng ) X CcIi ( lông trắng )
F2: 9( C-I-), 3( C-ii), 3( ccI-), 1ccii.
KH: 13 trắng : 3 màu
Câu hỏi: làm thế nào để phân biệt được 1 số tính trạng do 1 gen chi phối và số tính trạng di truyền liên kết ( gây đột biến gen, nếu có hiện tượng các tính trạng đó đều bị biến đổi thì các tính trạng đó do cùng 1 gen chi phối)
E. Di truyền liên kết
I. Di truyền liên kết hoàn toàn:
1. TN của Moocgan :SGK
2. Giải thích: Kết quả phép lai cho thấy:
+ Ruồi cái chỉ cho 1 loại giao tử, 2 loại KH thu được chứng tỏ ruồi đực F1 dị hợp về
2 cặp gen nhưng chỉ cho 2 loại giao tử + Tính trạng mình xám luôn đi cùng cánh dài, thân đen luôn đi kèm cánh ngắn→ có sự di truyền liên kết giữa 2 tính trạng màu sắc thân và hình dạng cánh
3. Cơ sở tế bào học của DTLK:
Các gen trên cùng 1 NST sẽ phân ly và tổ hợp cùng nhau trong quá trình giảm phân và thụ tinh đưa đến sự di truyền đồng thời của nhóm tính trạng do chúng quy định.
Quy ước: B: xám V: cánh dài
b: đen v: cụt
Ptc: Xám dài x Đen cụt
GV hướng dẫn HS tự viết
II. Di truyền liên kết không hoàn toàn
1. TN: SGK
2. Giải thích:
Ruồi đực đen ngắn chỉ cho 1 loại giao tử nên sự phân ly ở Fb chứng tỏ ruồi cái F1
cho 4 loại giao tử nhưng khong phải là 1:1:1:1 mà là 0,415: 0,415: 0,085: 0,085
Như vậy trong quá trình phát sinh giao tử cái đã xảy ra đổi chỗ( HV) giữa V và v
dẫn đến sự xuất hiện thêm 2 loại giao tử Bv và bV , do đó có sự tổ hợp lại các tính trạng của bố và mẹ là thân đen, cánh dài và thân xám , cánh cụt (BDTH)
3. Cơ sở tế bào học của HVG:
+ Trong kỳ đầu của GP1 có sự TĐC ở từng đoạn tương ứng giữa 2 trong 4 croma tit
của cặp NST kép tương đồng . Sự TĐC trên đã tạo ra các loại giao tử mang gen hoán vị
có tỷ lệ bằng nhau (Bv = bV) và các loại giao tử có gen liên kết bằng nhau (BV = bv)
+ Tỷ lệ các loại giao tử phụ thuộc vào tần số hoán vị gen. TSHVGđược tính bằng tổng tỷ lệ các loại giao tử mang gen hoán vị + Tần số hoán vị gen thể hiện khoảng cách tương đối giữa các gen trên cùng NST.
Khoảng cách càng lớn thì TSHVG càng cao. TSHVG không vượt quá 50% .
III. Lập bản đồ gen (Bản đồ di truyền)
1. Khái niêm: -Lập bản đồ gen là xác định trình tự và khoảng cách của các gen nhất định trên từng NST. Có 2 loại bản đồ gen đó là bản đồ di truyền và bản đồ tế bào(bản đồ vật lí)
-Bản đồ di truyền là sơ đồ về trình tự sắp xếp và vị trí tương đối của các gen được xây dựng dựa trên tần số hoán vị gen
- Bản đồ tế bào là bản đồ về trình tự và khoảng cách vật lí giữa các gen trên NST. Khoảng cách giữa các gen trong bản đồ di truyền được đo bằng tần số hoán vị gen
2. Cách lập BDDT
- Khi lập BĐDT, cần phải xác định số nhóm gen liên kết cùng với việc xác định trình tự và khoảng cách phân bố của các gen trong nhóm liên kết trên NST.
- Dựa vào việc xác định TSHVG, người ta xác lập trình tự và khoảng cách phân bố của các gen trên NST.
- Các nhóm gen liên kết được đánh số theo thứ tự của NST trong bộ đơn bội của loài . Các gen trên NST được ký hiệu bằng các chữ cái của tên các tính trạng bằng tiếng Anh
- Đơn vị khoảng cách trên bản đồ là cM ứng với TSHVG 1% . Vị trí tương đối của các gen trên NST thường được tính từ đầu mút của NST.
- Để xác định trình tự các gen trên NST người ta thường sử dụng phép lai phân tích giữa các cá thể dị hợp tử về 3 cặp gen với các cá thể đồng hợp tử lặn về cả 3 cặp gen. Sau đó tiến hành phân tích tần số hoán vị gen giữa 2 gen một
3. Ý nghĩa của bản đồ di truyền: Nếu ta biết được tần số hoán vị gen giữa 2 gen nào đó ta có thể tiên đoán được tần số các tổ hợp gen mới trong các phép lai. Điều này là cực kì quan trọng trong công tác chọn giống cũng như trong nghiên cứu khoa học
IV.Ý nghĩa:
1. DTLK hoàn toàn:
-Hạn chế xuất hiện BDTH, đảm bảo sự DT bền vững của từng nhóm tính trạng.
- Trong chọn giống người ta có thể chọn được những tính trạng tốt luôn đi cùng nhau
2. DTLK không hoàn toàn:
- Làm tăng số BDTH, nhờ đó các gen quý trên các NST tương đồng có dịp tổ hợp với nhau làm thành nhóm gen liên kết mới
- Có ý nghĩa thong chọn giống và tiến hóa
- Thông qua việc xác định TSHVG người ta lập bản đồ di truyền
Bài tập vận dụng: Bài 4 trang 58
a) F2 có tỷ lệ trơn: nhăn = 3:1, Có tua cuốn : không tua cuốn = 3:1→ trơn, có tua cuốn là những tính trạng trội và nhăn, không tua cuốn là những tính trạng lặn.
Quy ước: A: trơn B: có tua cuốn
a: nhăn b: không tua cuốn
Ta thấy tỷ lệ KH F2 khác tích xác suất của các tính trạng hợp thành nó → các tính trạng trên di truyền theo quy luật LKG.
HS tự viết SĐL
b) Để F1 có tỷ lệ trơn : nhăn = 1:1→ P: Aa x aa
có tua cuốn :không tua cuốn = 1:1→P: Bbxbb
F1 có 4 loại KH → mỗi bên P cho 2 loại giao tử
→ P: Trơn, không tua cuốn x nhăn, có tua cuốn
Ab//ab aB//ab
HS tự viết : Bài 5 trang 58
a) F1 đồng tính, F2 có tỷ lệ xám : đen = 3:1, dài : cụt = 3:1 → xám, dài là những tính trạng trội, đen , cụt là những tính trạng lặn.
Quy ước: B:xám V:dài
B: đen v: cụt
Ta thấy tỷ lệ KH F2 khác tích xác suất của các tính trạng hợp thành nó→ các tính trạng trên di truyền theo quy luật HVG.
- 62 -
Ta có : 0,205bv//bv = 0,41bv x 0,5bv → TSHVG = 0,18
HS tự viết SĐL
b) Có 2 trường hợp KG con cái xám,dài là dị hợp đều và dị hợp chéo: HS tự viết
G. Di truyền liên kết với giới tính
I. NST giới tính:
1. Khái niệm: NST giới tính là NST chứa các gen quy định tính đực cái, ngoài ra còn có các gen quy định các tính trạng thường
2. Đặc điểm:
- Cặp NST giới tính có thể tương đồng hoặc không tương đồng tùy giới tính và tùy nhóm loài
- Trong cặp XY có đoạn tương đồng và có đoạn không tương đồng
3. Các dạng NST:
- Dạng XX/XY:
+ Cái XX, đực XY : Người, động vật có vú, ruồi giấm, cây gai,cây chua me.
+ Cái XY, đực XX: Chim, bướm,ếch nhái, bò sát, dâu tây .
- Dạng XX/XO:
+ Cái XX, đực XO: Châu chấu, bọ xít.
+ Cái XO, đực XX: bọ nhậy.
II. Gen trên NST giới tính X:
1. Thí nghiệm: SGK
2. Giải thích:
Kết quả cho thấy mắt đỏ là tính trạng trội, mắt trắng là tính trạng lặn(Quy ước: W: mắt đỏ, w:mắt trắng). Theo Moocgan các gen này nằm trên X. Màu mắt được di truyền chéo (phép lai nghịch).Tỷ lệ KH phân bố không đồng đều ở 2 giới( phép lai thuận) và đồng đều ở 2 giới ( phép lai nghịch)
3. Cơ sở tế bào học:
+ Do sự phân ly của các cặp NST giới tính trong giảm phân và sự tổ hợp của chúng qua thụ tinh dẫn đến sự phân ly và tổ hợp của cặp gen quy định màu mắt
+ NST Y không mang gen quy định màu mắt nên ruồi đực chỉ cần NST X mang gen lặn
là biểu hiện mắt trắng.Vì vậy ruồi cái mắt trắng thường hiếm
+SĐL:Lai thuận : Ptc: ♀ mắt đỏ x ♂ mắt trắng
X WXW XwY
F1 : XWXw, XWY( Mắt đỏ)
F1: XWXw x XWY
F2: X WXW , XWXw, XWY , XwY
KH: 3 mắt đỏ: 1 mắt trắng(toàn là ruồi ♂)
Lai nghịch : Ptc: ♂ mắt đỏ x ♀ mắt trắng
XWY XwXw
F1: XWXw , XwY(1 ♀ mắt đỏ,1♂ mắt trắng)
F1: XWXw x XwY
F2: XWXw, XWY, XwXw, XwY
KH: 1♀ mắt đỏ, 1 ♂ mắt đỏ, 1♀ mắt trắng, 1 ♂ mắt trắng
- Kết quả của phép lai thuận nghịch la khác nhau
- Tỉ lệ phân li kiểu hình ở đời F2 có sự khác biệt ở 2 giới
- Có sự di truyền chéo: trong phép lai nghịch, mẹ “ truyền’’kiểu hình mắt trắng cho con trai , bố“truyền’’kiểu hình mắt đỏ cho “con gái’’
III. Gen trên Y
1. VD: Ở người : Gen a quy định tật dính ngón 2,3 nằm trên NST Y, không có alen tương ứng trên X.
P: Mẹ bình thường x Bố dính ngón
XX XYa
F1: XX, XYa
KH: con gái bình thường, con trai dính ngón 2,3
2. Đặc điểm di truyền: Gen trên Y không có alen tương ứng trên X truyền trực tiếp cho giới có cặp XY nên tính trạng do gen trên Y được truyền cho 100% số cá thể có cặp XY (di truyền thẳng)
IV. Ý nghĩa của di truyền liên kết với giới tính
Dựa vào những tính trạng liên kết với giới tính để sớm phân biệt đực cái, điều chỉnh tỷ lệ đực, cái theo mục tiêu sản xuất
VD: ở gà người ta sử dụng gen trội trên NST X xác định lông vằn để phân biệt trống mái từ khi mới nở. Gà trống con mang 2 gen trội trên cặp XX có khoang vằn ở đầu rõ hơn gà mái con chỉ có 1 gen trội trên cặp XY
Bài tập vận dụng: Bài 4 trang 63
Ta nhận thấy tính trạng lặn thường được biểu hiện ở giới có cặp XY nên tính trạng mù màu do gen lặn nằm trên NST quy định .
Con trai mù màu(3) có kiểu gen XmY nhận Xm từ mẹ nên mẹ(1) bình thường có KG là XMXm , bố (2) bình thường có KG là XMY.
Con gái(4) bình thường lấy chồng (5) bị mù màu sinh được con gái (7) mù màu. Con gái (7) mù màu có KG là XmXm nhận Xm từ bố và Xm từ mẹ nên người (4) bình thường có KG là XMXm, người (5)có KG là XmY , người con gái (6) có KG là XMXm. Vậy KG của 7 người trong gia đình đó là:
Người 3,5,có KG là XmY, người 2 có KG là XMY, Người 1,4,6, có KG là XMXm, người 7 có KG là XmXm
Bài 5 trang 63
a) Ở phép lai trên có hiện tượng di truyền chéo nên tính trạng dạng lông do gen trên NST X quy định , tính trạng lông không vằn được biểu hiện ở giới có cặp XY nên tính trạng lông không vằn là do gen lặn quy định, tính trạng lông vằn là do gen trội quy định.
Quy ước: A: lông vằn
A: lông không vằn.
Ta có SĐL:
P: ♂ lông không vằn x ♀lông vằn
XaXa XAY
F1 : XAXa , XaY
KH: 1 ♂ lông vằn, 1♀lông không vằn
b) F1: ♂ lông vằn x ♀lông không vằn
XAXa XaY
F2: XAXa , XaXa , XAY, XaY
KH: 1 ♂ lông vằn, 1 ♂ lông không vằn, 1♀lông vằn, 1♀ lông không vằn
H. Di truyền ngoài NST
I. Di truyền theo dòng mẹ:
1.VD: Lai thuận: P: ♀ xanh lục X ♂ Lục nhạt => F1: 100% xanh lục
Lai nghịch P: ♀ Lục nhạt X ♂ xanh lục => F1: 100% Lục nhạt
2. Nhận xét: Hai hợp tử do lai thuận và lai nghịch tạo thành đều giống nhau về nhân nhưng
khác nhau về tế bào chất. Trong tế bào con lai mang chủ yếu tế bào chất của mẹ, do đó tế bào chất đã có vai trò đối với sự hình thành tính trạng của mẹ ở cơ thể lai
3. Giải thích: Trong thí nghiệm trên , sự di truyền tính trạng xanh lục liên quan với tế bào chất ở tế bào trứng của cây mẹ xanh lục( lai thuận), còn sự di truyền tính trạng lục nhạt chịu ảnh hưởng của tế bào chất ở tế bào trứng của cây mẹ xanh lục nhạt( lai nghịch). Vì vậy hiện tượng di truyền này là di truyền tế bào chất( hay di truyền ngoài nhân hoặc ngoài NST). Do con lai mang tính trạng của mẹ nên di truyền tế bào chất được xem là di truyền theo dòng mẹ.Nhưng không phải mọi hiện tượng di truyền theo dòng mẹ đều là di truyền tế bào chất
II. Sự di truyền của các gen trong ti thể và lục lạp
1. Đặc điểm của gen tế bào chất(gen ngoài nhân hay gen ngoài NST)
-Bản chất của gen này cũng là ADN.
-Lượng ADN trong ti thể và lục lạp ít hơn nhiều so với ADN trong nhân
- Đều chứa ADN xoắn kép trần mạch vòng tương tự ADN vi khuẩn
- Cũng có khả năng đột biến
2. Sự di truyền ti thể ( mtADN: gen ti thể)
- Mã hóa nhiều thành phần của ti thể: hai loại rARN, tất cả tARN trong ti thể và nhiều loại protein có trong thành phần của màng bên trong ti thể .
- Mã hóa cho 1 số protein tham gia chuỗi truyền electron
3. Sự di truyền lục lạp (cpADN: gen lục lạp)
- Chứa gen mã hóa rARN và tARN lục lạp
- Mã hóa cho 1 số protein của riboxom, của màng lục lạp cần thiết cho việc vận chuyển e trong quá trình quang hợp
III. Đặc điểm di truyền ngoài NST
- Kết quả lai thuận nghịch khác nhau, trong đó con lai thường mang tính trạng của mẹ nghĩa là di truyền theo dòng mẹ
- Trong di truyền tế bào chất, vai trò chủ yếu thuộc về tế bào chất của giao tử cái được tạo ra từ mẹ
- Các tính trạng di truyền không tuân theo các quy luật di truyền NST vì tế bào chất không được phân phối đều cho các tế bào con như đối với NST
- Tính trạng do gen trong tế bào chất quy định vẫn sẽ tồn tại khi thay thế nhân tế bào bằng
1 nhân có cấu trúc di truyền khác
IV. Kết luận:
Trong tế bào có 2 hệ thống di truyền : di truyền qua nhân và di truyền qua tế bào chất ( di truyền ngoài NST), trong đó nhân có vai trò chính, tế bào chất cũng có vai trò nhất định
Bài tập vận dụng:
Bài
4 trang 68
Có thể giải thích hiện tượng lá đốm các màu ở 1 số loài thực vật như sau: gen ngoài NST cũng có đột biến. Chẳng hạn ADN của lục lạp có đột biến làm mất khả năng tổng hợp chất diệp lục, làm xuất hiện các lạp thể màu trắng.Lạp thể trắng lại sinh ra lạp thể trắng.Do vậy trong cùng 1 tế bào lá có thể có cả 2 loại lạp thể màu lục và màu trắng. Sự phân phối ngẫu nhiên và không đều của 2 loại lạp thể này qua các lần nguyên phân sinh ra hiện tượng lá có các đốm trắng, có khi cả mảng lớn tế bào lá không có diệp lục. Việc nghiên cứu di truyền tế bào chất có giá trị thực tiễn là: Có thể giải thích lá đốm nhiều màu ở 1 số loài cây cảnh. Hiện tượng bất thụ đực được sử dụng trong chọn giống cây trồng để tạo hạt lai mà khỏi tốn công hủy bỏ phấn hoa cây mẹ. Các dòng bất thụ đực sẽ nhận phấn hoa từ cây bình thường khác
Bài 5 trang 68:
a) Khi cho cây xanh lục F1 ( theo ví dụ ở mục I) giao phấn với nhau thì F2 cho 100% xanh lục vì F2 chứa tế bào chất của cây xanh lục
b) Khi cho cây lục nhạt F1 ( theo ví dụ ở mục I) giao phấn với nhau thì F2 cho 100% lục nhạt vì F2 chứa tế bào chất của cây lục nhạt
K. Ảnh hưởng của môi trường đến sự biểu hiện của gen
I. Mối quan hệ giữa kiểu gen, môi trường và kiểu hình
1. Ví dụ: Cây hoa anh thảo có giống hoa đỏ với kiểu gen AA và giống hoa trắng với kiểu gen aa. Khi đem cây thuộc giống hoa đỏ thuần chủng (AA) trồng ở 35oc thì ra hoa trắng . Thế hệ sau của cây hoa trắng này trồng ở 20oc lại cho hoa đỏ. Trong khi giông hoa trắng trồng ở 20oc hay 35oc đều chỉ ra hoa màu trắng
2. Nhận xét: - Giống hoa đỏ thuần chủng cho ra hoa màu đỏ hay trắng phụ thuộc vào nhiệt độ môi trường, còn giống hoa trắng chỉ cho ra màu trắng, không phụ thuộc vào nhiệt độ môi trường
3.Kết luận:
-Bố mẹ không truyền đạt cho con những tính trạng đã hình thành sẵn mà truyền đạt một kiểu gen. Kiểu gen quy định khả năng phản ứng của cơ thể trước môi trường. Kiểu hình là kết quả sự tương tác giữa kiểu gen và môi trường.
- Trong quá trình biểu hiện kiểu hình, kiểu gen còn chịu tác động khác nhau của môi trường trong và môi trường ngoài cơ thể
+ Tác động của môi trường trong được thể hiện ở mối quan hệ giữa các gen với nhau, giữa gen nhân và gen tế bào chất, hoặc giới tính
+ Các yếu tố môi trường ngoài đó là:Ánh sáng, nhiệt độ, PH trong đất, chế độ dinh dưỡng
- Tác động của môi trường còn tùy thuộc từng loại tính trạng. Loại tính trạng chất lượng phụ thuộc chủ yếu vào kiểu gen, ít chịu ảnh hưởng của môi trường. Các tính trạng số lượng thường là những tính trạng đa gen, chịu ảnh hưởng nhiều của môi trường II. Thường biến 1.Ví dụ: Một số loài thú ở xứ lạnh về mùa đông co bộ lông dài màu trắng lẫn với tuyết, mùa hè lông thưa hơn và chuyển sang màu vàng hoặc xám Sự thay đổi bộ lông của các loài thú này đảm bảo cho sự thích nghi theo mùa
2. Khái niệm: là những biến đổi của kiểu hình của cùng một kiểu gen, phát sinh trong đời cá thể dưới ảnh hưởng của môi trường, không do sự biến đổi trong kiểu gen
3. Đặc điểm :
- Là loại biến dị đồng loạt theo cùng 1 hướng xác định đối với 1 nhóm cá thể có cùng kiểu gen và sống trong điều kiện giống nhau.
- Các biến đổi này tương ứng với điều kiện môi trường.
- Không do những biến đổi trong kiểu gen nên không di truyền.
4. Ý nghĩa: Nhờ có những thường biến mà cơ thể phản ứng linh hoạt về kiểu hình, đảm bảo
sự thích nghi trước những thay đổi nhất thời hoặc theo chu kỳ của môi trường
III. Mức phản ứng
1. Ví dụ: Kiểu gen 1+ môi trường 1 Kiểu hình 1
Kiểu gen 2+ môi trường 2 Kiểu hình 2
Kiểu gen 3+ môi trường 3 Kiểu hình 3
Kiểu gen n+ môi trường n Kiểu hình n
2. Khái niệm: Tập hợp các kiểu hình1,2,3...n nói trên của một kiểu gen tương ứng với các
môi trường khác nhau gọi là mức phản ứng
3. Đặc điểm:
- Mức phản ứng được di truyền
- Trong 1 kiểu gen mỗi gen có mức phản ứng riêng:
+ Tính trạng chất lượng có mức phản ứng hẹp. Ví dụ tính trạng tỉ lệ bơ trong sữa bò
+ Tính trạng số lượng có mức phản ứng rộng. Ví dụ tính trạng sản lượng sữa bò
- Mức phản ứng về mỗi tính trạng thay đổi tùy kiểu gen của từng cá thể
Ví dụ : Trong điều kiện thích hợp, giống lúa DR2cho năng suất tối đa 9,5 tấn/ha, trong khi
đó giống tám thơm đột biến chỉ cho 5,5 tấn/ha
Bài tập vận dụng:
Khi giao phấn 2 dòng cùng loài thân đen và thân xám với nhau thu được F1. Cho F1 tiếp
tục giao phối với nhau được F2 có tỉ lệ:
- Ở giới đực: 3 đen: 1 xám
- Ở giới cái: 3 xám: 1 đen.
Giải thích kết quả phép lai và viết sơ đồ lai từ P đến F2.Biết AA:đen, aa: xám
Giải: Từ tỉ lệ kiểu hình ở F2 cho thấy màu sắc thân bị ảnh hưởng bởi giới tính, kiểu gen
ở thể dị hợp biểu hiện thân đen ở giới đực và biểu hiện thân xám ở giới cái.
Sơ đồ lai:
Ptc: Thân đen x Thân xám
AA aa
F1: Aa
KH: ♂ đen, ♀ xám
F2:1AA:2Aa:1aa
KH: Ở giới đực: 3 đen: 1 xám
Ở giới cái : 3 xám: 1 đen
Trong chương trình sinh học phổ thông, di truyền học là một phần rất quan trọng chiếm 50% thời lượng của chương trình sinh học lớp 12, không những vậy nó còn chiếm một cơ số điểm không nhỏ trong các cuộc thi như: thi Đại học, thi học sinh giỏi Tỉnh và thi học sinh giỏi Quốc gia...
Trong phần Di truyền học của chương trình sinh học lớp 12 thì phần "Các quy luật di truyền" là phần không thể thiếu trong các kỳ thi cấp quốc gia. Đã có nhiều chuyên đề khai thác về phần cơ sở lý thuyết hay các phương pháp giải bài tập của các quy luật di truyền. Tuy nhiên, qua giảng dạy học sinh chuyên Sinh, bồi dưỡng học sinh giỏi... Tôi thấy học sinh còn lúng túng khi giải bài tập di truyền liên kết với giới tính, đặc biệt là các dạng bài tập có sự kết hợp của các quy luật di truyền khác như phân ly độc lập, tương tác gen, hoán vị gen... với di truyền liên kết với giới tính. Ở đây không phải là các em chưa nắm chắc cơ sở lý thuyết hay phương pháp giải mà là do các em chưa có tài liệu riêng có tính hệ thống về các dạng bài tập trên. Vì vậy rất khó khăn cho học sinh rèn luyện để giải các dạng bài tập quy luật di truyền này một cách thành thạo. Mặt khác qua tổng hợp các đề thi học sinh giỏi quốc gia từ năm gần đây, phần bài tập quy luật di truyền đa số có bài tập về sự di truyền liên kết với giới tính, gen trên NST giới tính X, ...
Từ những cơ sở trên, tôi đã sưu tầm một số bài tập chọn lọc có sự kết hợp giữa các quy luật di truyền khác với di truyền liên kết với giới tính nhằm mục tiêu vô cùng thiết thực là giúp cho học sinh thi chuyên Sinh học tập và nghiên cứu về phần bài tập dạng này tốt hơn và đồng thời cũng giúp chính bản thân mình giảng dạy tốt hơn.
I. Phân loại, phương pháp giải chung
1. Phân loại.
- Dựa trên một loạt các tiêu chí sau: Số tính trạng trong phép lai, số gen chi phối một tính trạng, sự phân bố của gen trên NST... có thể phân loại các dạng bài tập di truyền liên kết
với giới tính như sau:
+ 1 gen quy định 1 tính trạng
+ 2 hoặc nhiều gen quy định 1 tính trạng
- Lai 2 tính trạng trở lên có tính trạng liên kết giới tính trong đó có:
+ Sự di truyền của 2 tính trạng tuân theo quy luật phân ly độc lập
+ Sự di truyền của 2 tính trạng tuân theo quy luật liên kết gen (hoàn toàn hoặc không hoàn toàn)
2. Phương pháp giải chung
2.1. Để giải được các dạng bài tập trên yêu cầu học sinh phải nắm được các kiến thức cơ bản và nâng cao sau đây:
- Cách giải các dạng bài tập riêng của từng quy luật di truyền như phân ly độc lập, tương tác gen, hoán vị gen...
- Ngoài ra đây là dạng bài tập kết hợp với di truyền liên kết giới tính nên cần phải nắm vững đặc điểm của riêng dạng bài tập này, cụ thể như
- Cơ sở xác định giới tính khác nhau ở một số loài và tỉ lệ giới tính trong quần thể sinh vật xấp xỉ 1 đực : 1 cái.
- Những đặc điểm của sự di truyền liên kết với giới tính khi gen nằm trên X không có đoạn tương đồng trên Y hoặc ngược lại. 2.2. Phương pháp giải chung
- Quan sát sự khác biệt về kiểu hình giữa giới đực và giới cái hay tính trạng biểu hiện không đồng đều ở 2 giới, tính trạng lặn chủ yếu ở giới dị giao tử XY, XO. Nếu có biểu hiện cùng giới thì cách đời và thường là do gen lặn quy định.
- Mẹ dị hợp tử sẽ sinh ra các con đực có tỷ lệ phân ly về kiểu hình là 1:1.
- Bố truyền nhiễm sắc thể X cho con gái và nhiễm sắc thể Y cho con trai (Quy luật di truyền chéo và di truyền thẳng).
- Một gen trên nhiễm sắc thể thường và một gen liên kết với nhiễm sắc thể X sẽ cho tỷ lệ phân ly 3:3:1:1 ở đời con.
- Gen gây chết liên kết với giới tính sẽ làm giảm số con đực sinh ra.
II. Bài tập rèn kỹ năng.
1. Bài tập cơ bản. Bài 1:
Ở Drosophila, một ruồi ♀ lông ngắn được lai với ruồi ♂ lông dài. Ở đời con có 42 ruồi ♀ lông dài, 40 ruồi ♂ lông ngắn và 43 ruồi ♂ lông dài.
a) Hỏi kiểu di truyền của tính trạng lông ngắn?
b) Hỏi tỷ lệ phân ly kiểu hình ở đời con nếu bạn lai hai ruồi lông dài
Gợi ý giải
a) Lông ngắn là tính trạng liên kết với giới tính nhưng là một gen gây chết. Chúng ta thấy có sự khác biệt cả về kiểu hình cả về số lượng giữa ruồi ♂ và ruồi ♀ ở đời con; điều đó
cho thấy gen liên kết với giới tính là gen gây chết bán hợp tử (không có ruồi ♂ lông ngắn).
Vì giới cái có hai kiểu hình cho nên lông ngắn phải là tính trạng trội và phép lai sẽ là:
XSXs x XsY
(lông ngắn) (lông dài) ↓
XSXs XsXs XSY XsY
(lông ngắn) (lông dài) (chết) (lông dài)
b) Ở đời con tất cả đều có lông dài và phân đều ở cả hai giới. Để có ruồi ♀ lông dài, ruồi mẹ phải đồng hợp tử và phép lai sẽ là:
XsXs x XsY (lông dài) (lông dài)
Bài 2:
Một mèo ♀ lông khoang đen vàng được lai với một mèo ♂ lông vàng. Ở đời con nhận
được:
♀ : 3 vàng, 3 khoang đen vàng
♂ : 3 đen, 3 vàng
Hãy giải thích những kết quả này.
Gợi ý giải
Mèo ♀ dị hợp tử về gen quy định màu lông liên kết với giới tính. Chúng ta thấy có sự khác nhau về kiểu hình ở con ♂ và con ♀ chứng tỏ tính trạng liên kết với giới tính. Mèo ♀ phải dị hợp tử và phải có nhiễm sắc thể X bất hoạt.
Quy ước Xa = màu đen và Xb - màu vàng. Phép lai sẽ là: XaXb x XbY
XbXb XaXb XaY XbY (vàng) (đen và vàng) (đen) (vàng)
Bài 3: Phép lai giữa một chim hoàng yến ♂ màu vàng với một chim ♀ màu xanh sinh ra tất cả chim ♂ có màu xanh và tất cả chim ♀ có màu vàng. Hãy giải thích các kết quả này.
Gợi ý giải
Màu sắc lông là tính trạng liên kết với giới tính và giới ♂ là giới đồng giao tử. Chúng ta thấy có sự khác biệt về kiểu hình giữa giới ♂ và giới ♀ cho thấy có sự liên kết với giới tính.
Vì tất cả các cá thể của mỗi giới giống nhau về kiểu hình nên bố mẹ không thể là dị hợp tử. Ta lập phép lai theo cách thông thường (A: xanh; a: vàng):
XAXA x XaY (xanh) ↓ (vàng)
XAXa , XAY (tất cả xanh)
Trong trường hợp này thì cả chim trống và chim mái đều có màu xanh, vì chim ♀ con là XAXa và chim ♂ con là XAY. Kết quả này không phù hợp với kết quả thực tiễn. Do vậy có thể có sai lầm khi chúng ta đã cho rằng giới ♀ là giới đồng giao tử. Vì giới ♂ là giới đồng giao tử nên phép lai bây giờ sẽ là:
ZAW x ZaZa (♀ xanh) (♂ vàng) ↓ ZaW ZAZa (♀ vàng) (♂ xanh)
Bài 4:
Lai gà trống mào to, lông vằn thuần chủng với gà mái lông không vằn, mào nhỏ thuần chủng, được gà F1 có lông vằn, mào to.
a) Cho gà mái F1 lai với gà trống lông không vằn, mào nhỏ, được F2 phân ly như sau: 1 gà trống mào to, lông vằn: 1 gà trống mào nhỏ, lông vằn: 1 gà mái mào to, lông không vằn: 1 gà mái mào nhỏ, lông không vằn. Biết rằng mỗi gen quy định một tính trạng. Hãy biện luận và lập sơ đồ lai giải thích cho phép lai trên.
b) Phải lai gà trống F1 với gà mái có kiểu gen và kiểu hình như thế nào để ngay thế hệ sau có tỷ lệ phân ly kiểu hình theo giới tính 1:1:1:1:1:1:1:1.
c) Muốn tạo ra nhiều biến dị nhất, phải chọn cặp lai có kiểu gen và kiểu hình như thế nào?
Gợi ý giải
a) Kích thước mào do gen trên NST thường quy định; dạng lông liên kết giới tính. A: mào to, a: mào nhỏ; B: lông vằn, b: lông không vằn. Sơ đồ lai:
P: Trống AAXBXB x Mái aaXbY => Fl: AaXBXb, AaXBY. Mái F1 lai với trống mào nhỏ, lông không vằn: AaXBY x aaXbXb
b) Tỷ lệ 1:1:1:1:1:1:1:1 = (1:1:1:1)(1:1) cho thấy tính trạng liên kết giới tính phân ly 1 : 1 : 1 : 1, còn tính trạng do gen trên NST thường quy định phân ly 1 : 1 => P: AaXBXb x aaXbY
- 70 -
c) Để tạo ra nhiều biến dị nhất, bố mẹ phải sinh ra nhiều loại giao tử nhất. Vậy P phải có
kiểu gen: AaXBXb x AaXbY.
Bài 5:
Bệnh sắc tố từng phần trên da người là một hiện tượng hiếm có, trong đó melanine
không được chuyển hoá bởi tế bào sắc tố, gây ra những dòng tế bào sắc tố dạng xoáy trên da. Một người phụ nữ bị bệnh lấy một người đàn ông bình thường. Cô ta có 3 đứa con gái bình thường, 2 đứa bị bệnh và 2 con trai bình thường. Ngoài ra, cô ta có 3 lần sảy thai mà thai đều là nam giới bị dị tật. Hãy giải thích những kết quả trên.
Gợi ý giải
Tính trạng đó là tính trạng trội liên kết nhiễm sắc thể X và gây chết ở nam giới. Chúng ta thấy sự khác nhau về kiểu hình ở đời con gợi ý đến sự liên kết giới tính. Chúng ta không thấy những người con trai bị bệnh nhưng lại có nhũng người con trai có kiểu hình bình thường; điều đó chứng tỏ những người con trai đó đã nhận được gen lặn. Vậy người mẹ phải có kiểu gen dị hợp tử về tính trạng này. Gọi XA: gen gây bệnh và Xa: gen quy định kiểu hình bình thường. Phép lai khi đó sẽ là: XAXa x XaY ↓
XAXa XaXa XaY XAY (bị bệnh) (bình thường) (bình thường) (chết)
40 ruồi cái thân xám, cánh dài, mắt đỏ : 20 ruồi đực thân xám, cánh dài, mất đỏ
20 ruồi đực thân xám, cánh dài, mắt trắng : 40 ruồi cái thân đen, cánh cụt, mắt đỏ
20 ruồi đực thân đen, cánh cụt, mắt đỏ : 20 ruồi đực thân đen, cánh cụt, mắt trắng
10 ruồi cái thân xám, cánh cụt, mắt đỏ : 5 ruồi đực thân xám, cánh cụt, mắt đỏ
5 ruồi đực thân xám, cánh cụt, mắt trắng : 10 ruồi cái thân đen, cánh dài, mắt đỏ
5 ruồi đực thân đen, cánh dài, mắt đỏ : 5 ruồi đực thân đen, cánh dài, mắt trắng
Biện luận xác định quy luật di truyền của các tính trạng trên, kiểu gen của cá thể đực chưa biết và lập sơ đồ lai.
Gợi ý giải Mỗi tính trạng do 1 gen quy định. Gen quy định màu mắt liên kết X. Gen quy định màu thân và hình dạng cánh liên kết không hoàn toàn trên nhiễm sắc thể thường, tần số trao đổi chéo = 20%. Phân tích từng tính trạng cho thấy ruồi đực chưa biết kiểu gen có kiểu gen: bv/bvXWY.
Phép lai là: BV/ bv XWXw x bv/bv XWY
2. Bài tập nâng cao.
2.1. Hoán vị gen - Di truyền liên kết với giới tính
2.1.1. Kiến thức cơ bản.
* Tóm tắt cách giải chung về bài tập hoán vị gen: Có nhiều đấu hiệu cho thấy các tính trạng nghiên cứu được xác định bởi các gen liên kết với nhau như:
- Tỷ lệ phân ly ở đời lai khác với tỷ lệ mong đợi đối với hai bên phân ly độc lập cho thấy các gen di truyền liên kết với nhau.
- Các tính trạng được xác định bởi các gen liên kết luôn được di truyền cùng nhau.
- Liên kết gen hoàn toàn làm giảm số kiểu gen và kiểu hình ở đời con lai. Ngược lại, trao đổi chéo giữa các gen làm táng số kiểu gen và kiểu hình ở thế hệ sau.
- Tỷ lệ của các loại giao tử mang gen liên kết luôn bằng nhau, tỷ lệ của các giao tử mang gen trao đổi chéo cũng bằng nhau và nhỏ hơn tỷ lệ của các giao tử mang gen liên kết.
- Trong một phép lai phân tích, việc có hai lớp kiểu hình có tần số lớn bằng nhau và hai lớp kiểu hình có tần số nhỏ bằng nhau cho biết trong đó có gen liên kết không hoàn toàn.
* Với các gen liên kết không hoàn toàn nằm trên nhiễm sắc thể giới tính X mà không có các gen tương ứng trên Y, tần số hoán vị gen hoặc tỷ lệ kiểu gen, kiểu hình được xác định như trong trường hợp có trao đổi chéo một bên
2.1.2. Bài tập.
Bài 1:
Lai ruồi giấm cái thuần chủng thân xám, cánh dài, mắt đỏ với ruồi đực thân đen, cánh cụt, mắt trắng, được F1 đồng loạt thân xám, cánh dài, mắt đỏ. Cho ruồi cái F1 lai với ruồi đực khác chưa biết kiểu gen, được thế hệ lai gồm:
Bài 2: Lai ruồi giấm cái cánh bình thường, mắt trắng với ruồi giấm đực cánh xẻ, mắt đỏ, người ta thu được toàn bộ ruồi cái F1 có cánh dài bình thường, mắt đỏ và ruồi đực có cánh bình thường, mắt trắng. Lai phân tích ruồi cái F1, được đời con gồm bốn nhóm kiểu hình, trong đó ruồi cánh bình thường, mắt trắng và cánh xẻ, mắt đỏ chiếm 80% còn ruồi cánh bình thường, mắt đỏ và cánh xẻ, mắt trắng chiếm 20%. Biết rằng mỗi gen quy định một tính trạng và hai gen quy định hai tính trạng trên nằm trong cùng một nhóm liên kết và tính trạng mắt đỏ trội so với mắt trắng. Hãy biện luận và lập sơ đồ lai giải thích cho kết quả thu được ở phép lai trên.
Gợi ý giải Cánh bình thường trội (A: cánh bình thường, a: cánh xẻ). F1 cho thấy tính trạng màu mắt liên kết X. Hai gen nằm trong cùng nhóm liên kết => hai gen cùng liên kết X. Phép lai là:
Tần số trao đổi chéo giữa hai gen là 20%.
Bài 3: Ở gà gen S quy định tính trạng lông mọc sớm trội hoàn toàn so với gen s quy định tính trạng lông mọc muộn. Gen B quy định tính trạng lông đốm trội hoàn toàn so với gen b quy định tính trạng lông đen. Các gen s và b liên kết với giới tính, có tần số hoán vị gen ở gà trống là 30%. Đưa lai gà mái đen lông mọc sớm với gà trống thuần chủng về 2 tính trạng lông đốm, mọc muộn được F1 cho F1 giao phối với nhau được F2
a) Viết sơ đồ lai của P và F1 trong trường hợp cấu trúc NST không thay đổi trong giảm phân.
b) Tỉ lệ phân li kiểu hình ở F2 trong trường hợp cấu trúc NST thay đổi trong giảm phân? Gợi ý giải a) Nếu cấu trúc NST không đổi trong giảm phản nghĩa gì không có trao đổi chéo và đột biến cấu trúc NST. Theo giả thiết có sơ đồ lai : ( ♂ , ♀ )
b) Cấu trúc NST thay đổi trong giảm phân nghĩa là có trao đổi chéo. Ở gà trao đổi chéo chỉ xảy ra ở gà trống. Ta có sơ đồ lai :
a) Theo giả thiết ta có sơ đồ lai sau:
Bài 4:
Ở ruổi giấm gen A quy định cánh thường, gen a quy định cánh xẻ, gen B quy định mắt đỏ, gen b quy định mắt trắng. Các gen này nằm trên NST giới tính X.
a) Lai ruồi cái dị hợp đều về 2 gen với ruồi đực có kiểu hình cánh xẻ, mắt trắng. Nêu phương pháp xác định tần số hoán vị gen.
b) Lai ruồi cái dị hợp về 2 gen trên với ruồi đực có kiểu hình cánh bình thường, mắt đỏ Trình bày phương pháp xác định tần số hoán vị gen? So với trường hợp trên phương pháp này khác ở điểm nào? Tại sao có sự khác nhau đó?
Gợi ý giải
b) Kết quả lai giữa ruồi cái F1 với ruồi đực có kiểu gen
Ở đây ruồi cái F1 đều có kiểu hình giống nhau, nên việc tính tần số hoán vị gen phải dựa vào số cá thể có kiểu hình khác bố mẹ của các cá thể đực F1
Khác với phương pháp xác định tần số hoán vị gen ở sơ đồ lai thứ nhất là ở sơ đồ lai thứ hai, việc xác định tần số hoán vị gen chỉ dựa vào cá thể đực F1 có sự khác nhau đó, vì ở sơ đồ lai thứ nhất ruồi đực và ruồi cái F1 có kiểu hình khác P. Sơ đồ lai thứ 2 chỉ có ruồi đực F1 mới có kiểu hình khác P.
2.2. Tương tác gen - Di truyền liên kết với giới tính
2.2.1. Kiến thức cơ bản.
Các gen có thể tương tác với nhau để quy định một tính trạng. Phổ biến là hai gen không alen (và thường nằm trên các nhiễm sắc thể tương đồng khác nhau) tương tác với nhau. Vì vậy, tỷ lệ phân ly ở F2 thường là tỷ lệ biến đổi của phép lai hai tính (9:3:3:1) của Mendel. Ví dụ tỷ lệ 9:6:l. Có các kiểu tương tác chủ yếu sau:
- Tương tác bổ trợ: Hai gen trội cùng có mặt trong một kiểu gen tương tác với nhau làm xuất hiện tính trạng mới, khác bố mẹ. Ngoài cơ chế tương tác, các gen còn có thể có các chức năng riêng. Vì vậy, kiểu tương tác bổ trợ có thể cho các tỷ lệ phân ly 9:3:3:1, 9:6:1 hoặc 9:7.
- Tương tác át chế. Kiểu tương tác trong đó một gen ức chế sự biểu hiện của gen kia. Gen ức chế được gọi là gen át, còn gen bị ức chế dược gọi là gen khuất. Tuỳ thuộc vào gen át là gen trội hay gen lặn mà F2 có thể có các tỷ lệ phân ly 13:3, 12:3:1 hoặc 9:3:4.
- Tương tác cộng gộp: Kiểu tương tác trong đó mỗi alen trội (hoặc lặn) của mỗi gen đóng góp một phần vào sự hình thành tính trạng. Kiểu tương tác này đặc trưng cho các tính trạng số lượng. Với hai gen tương tác cộng gộp, F2 sẽ có tỷ lệ phân ly kiểu hình là 1:4:6:4:1.
Nếu kiểu hình không phụ thuộc vào số lượng alen trội trong kiểu gen, ta có tỷ lệ phân ly
15:1 ở F2. Tuy nhiên cũng có những trường hợp hai gen tương tác nhưng lại cùng nằm trên một nhiễm sắc thể. Khi đó, ngoài quy luật tương tác, các gen còn chịu sự chi phối của quy luật liên kết và hoán vị gen.
2.2.2. Bài tập. Bài 1: Có những con chuột rất mẫn cảm với ánh sáng mặt trời. Dưới tác động của ánh sáng mặt trời, chúng có thể bị đột biến dẫn đến ung thư da. Người ta chọn lọc được hai dòng chuột thuần chủng, một dòng mẫn cảm với ánh sáng mặt trời và đuôi dài, dòng kia mẫn cảm với ánh sáng và đuôi ngắn. Khi lai chuột cái mẫn cảm với ánh sáng, đuôi ngắn với chuột đực mẫn cảm với ánh sáng đuôi dài, người ta thu được các chuột F1 đuôi ngắn và không mẫn cảm với ánh sáng. Lai F1 với nhau, được F2 phân ly như sau: Chuột cái Chuột đực
ẫn cảm, đuôi ngắn 42
Hãy xác định quy luật di truyền của hai tính trạng trên và lập sơ đồ lai.
Gợi ý giải Tính mẫn cảm ánh sáng do tương tác bổ trợ hai gen trội cho tỷ lệ 9:7; độ dài đuôi liên kết giới tính. Nếu cho hai gen A và B tương tác quy định tính mẫn cảm ánh sáng, D quy định đuôi ngắn thì ta có sơ đồ lai: AAbbXDXD x aaBBXdY => F1: AaBbXDXd và
AaBbXDY.
Bài 2:
Một ruồi đực mắt trắng được lai với ruồi cái mắt nâu. Tất cả ruồi F1 có mắt đỏ kiểu dại. Cho F1 nội phối. Kết quả thu được:
Ruồi cái Ruồi đực
Mắt đỏ: 450 Mắt đỏ: 230
Mắt nâu: 145 Mắt trắng: 305 Mắt nâu: 68
Hãy giải thích các kết quả này
Gợi ý giải Có hai gen, một gen lặn trên nhiễm sắc thể thường quy định mắt màu nâu và một gen lặn liên kết với giới tính quy định mắt màu trắng. Bất cứ ruồi đồng hợp tử/bán hợp tử về gen quy định màu trắng nào cũng sẽ cho mắt màu trắng, dù có mặt các gen khác. F1 biểu hiện kiểu dại chứng tỏ có hai gen và F2 có sự khác nhau về tỷ lệ phân ly ở giới ♂ và giới ♀ chứng tỏ rằng ít nhất có một gen liên kết với giới tính. Một nửa số con ♂ ở F2 có mắt màu trắng, đây là tỷ lệ phân ly của một gen lặn liên kết với giới tính. Chúng ta nhận được tỷ lệ phân ly 3 đỏ: 1 nâu, là tỷ lệ phân ly của một gen trên nhiễm sắc thể thường. Tỷ lệ phân ly ở ruồi ♀ F2 là 3 đỏ: 1 nâu, cho thấy tất cả ruồi ♀ có ít nhất một nhiễm sắc thể X bình thường (X+)
Quy ước: X+-A- : đỏ; X+-aa : nâu; Xw : trắng. Phép lai sẽ là:
X+X+aa x X+YAA
↓
X+XwAa X+YAa
(Tất cả đỏ tự phối)
3 X+-A- : đỏ 3 X+YA- : đỏ
1 X+-aa : nâu
1 X+Yaa : nâu
3 XwYA- : trắng
1 XwYaa : trắng
Chúng ta nhận được một tỷ lệ phân ly biến đổi của tỷ lệ 3:3:1:1 trong số ruồi ♂ ở F2 cho thấy có một gen trên nhiễm sắc thể thường và một gen liên kết với giới tính. Số ruồi ♂ nhận được gần với tỷ lệ 4:3:1.
Bài 3:
Cho hai nòi chim thuần chủng lai với nhau được F1 đều lông vàng, dài. Cho con cái F1 lai phân tích thu được tỉ lệ : 1 con cái lông vàng, dài : 1 con cái lông xanh, dài : 2 con đực lông xanh, ngắn. Cho con đực F1 lai phân tích thu được tỉ lệ : 9 con lông xanh, ngắn : 6 con lông xanh, dài : 4 con lông vàng, dài : 1 con lông vàng, ngắn.
a. Nêu các quy luật di truyền tham gia để tạo nên các kết quả nói trên.
b. Xác định kiểu gen và kiểu hình của P và viết sơ đồ lai từng trường hợp từ P đến Fa.
Biết rằng kích thước lông do 1 gen quy định.
Gợi ý giải
a. Các quy luật : Tính trội, tương tác gen không alen, di truyền giới tính, di truyền liên kết với giới tính, liên kết gen và hoán vị gen.
b.
P : Lông vàng, dài x Lông xanh, ngắn AAXBDXBD aaXbdY ↓
P : Lông xanh, dài Lông xanh, ngắn
Bài 4:
aaXBDXBD AAXbdY
Cho nòi lông đen thuần chủng giao phối với nòi lông trắng được F1 có 50% con lông xám và 50% con lông đen. Cho con lông xám (F1) giao phối với con lông trắng (P) được tỉ lệ : 3 con lông xám : 4 con lông trắng : 1 con lông đen. Trong đó lông đen toàn là đực.
a. Biện luận và viết sơ đồ lai cho kết quả nói trên.
b. Cho con mắt đen (F1) giao phối với con lông trắng (P) thì kết quả phép lai sẽ thế nào ?
Gợi ý giải
a. P : AAXbXb x aaXBY
b. 4 con lông trắng : 2 con lông xám : 2 con lông đen. Bài 4:
Cho cá thể mắt đỏ thuần chủng lai với cá thể mắt trắng được F1 đều mắt đỏ. Cho con cái F1 lai phân tích được ta có tỉ lệ 3 mắt trắng : 1 mắt đỏ, trong đó mắt đỏ đều là con đực.
a. Biện luận và viết sơ đồ lai từ P đến Fa
b. Khi cho các con F1 tiếp tục giao phối với nhau thì kết quả ở F2 như thế nào?
Gợi ý giải
a. P : AAXBXB x aaXbY
b. F2 : 9 mắt đỏ : 7 mắt trắng
Bài 5:
Khi lai con cái (XX) mắt đỏ, tròn, cánh dài thuần chủng với con đực (XY) mắt trắng, dẹt, cánh cụt được F1 gồm các con cái đều mắt đỏ, tròn, cánh xẻ và các con đực đều mắt đỏ, tròn, cánh dài. Cho con cái F1 giao phối với con đực ở P thì được
- Ở giới cái có : 48 con mắt đỏ tròn, cánh xẻ ; 51 con mắt nâu, tròn cánh cụt, 52 con mắt nâu, dẹt cánh xẻ, 49 con mắt trắng, dẹt, cánh cụt.
- Ở giới đực có : 49 con mắt đỏ, tròn cánh dài : 48 con mắt nâu, tròn, cánh cụt ; 51 con mắt nâu, dẹt, cánh dài ; 52 con mắt trắng, dẹt, cánh cụt.
a. Từ kết quả phép lai trên hãy cho biết quy luật tác động của gen và quy luật vận động của NST như thế nào đối với sự hình thành và tỉ lệ phân li của kiểu hình ?
b. Viết sơ đồ lai từ P đến Fb.
Biết rằng hình dạng mắt và cánh đều tuân theo quy luật 1 gen chi phối 1 tính.
Gợi ý giải
a.
- Quy luật tác động của các gen alen : át hoàn toàn và không hoàn toàn.
- Quy luật tác động của các gen không alen theo kiểu bổ trợ
- Quy luật phân li độc lập của các cặp NST đã chi phối từ tỉ lệ phân li kiểu hình cùng với quy luật tác động của gen: b.
P : Con mắt đỏ, tròn, cánh dài x Con mắt trắng, dẹt, cánh cụt
AD/ADXBEXBE Ad/adXbeY
CHƯƠNG 3. DI TRUYỀN HỌC QUẦN THẺ
A. ĐẠI CƯƠNG VỀ QUẦN THỂ
I. ĐỊNH NGHĨA
II. PHÂN LOẠI QUẦN THỂ
III. QUẦN THỂ LÀ ĐƠN VỊ TIẾN HÓA CƠ SỞ.
B. CẤU TRÚC DI TRUYỀN CỦA QUẦN THỂ
I. SỰ CÂN BẰNG CỦA QUẦN THỂ VỚI NHỮNG GEN NẰM TRÊN NHIỄM SẮC THỂ
GIỚI TÍNH
1. Cơ sở lí luận: Xét một gen có 2 alen là A và a nằm trên nhiễm sắc thể giới tính X (Y không mang gen tương ứng), con đực là XY, con cái là XX thì trong quần thể sẽ hình thành 5 kiểu gen là:XAXA, XAXa, XaXa, XAY, XaY.
Gọi p là tần số alen A, q là tần số alen a.
NST X phân bố không đồng đều: 2/3 ở cơ thể ♀, 1/3 ở cơ thể ♂.Cho nên, các alen tương ứng trong quần thể cũng phân bố không đồng đều ở cơ thể đực và cái.
+ Cơ thể ♀: XAXA, XAXa, XaXa với tần số alen tương ứng là p2, 2pq, q2
+ Cơ thể ♂: XAY, XaY có tần số tương ứng là p,q.
+ Tần số alen A ở cá thể ♀: p2 + pq; Tần số alen a ở cá thể ♀: pq + q2
+ Tần số alen A ở cá thể ♂: p; Tần số alen a ở cá thể E: q
Tần số chung của alen trong quần thể ở cả giới cái và đực là:
pA = 3 1 pA♂ + 3 2 pA♀ = (p♂ + 2p♀)/3 => qa = 1 - pA
+ Nếu giá trị pA♂ = pA♀ => thì quần thể đạt trạng thái cân bằng hoặc cân bằng sau một thế hệ ngẫu phối.
+ Nếu pA♂╪ pA♀ => thì quần thể sẽ không đạt trạng thái cân bằng ngay ở thế hệ thứ nhất, thứ hai mà phải qua nhiều thế hệ ngẫu phối mới đạt trạng thái cân bằng.
Lưu ý: Giá trị chung pA trên NST giới tính không thay đổi nếu thỏa mãn các điều kiện nghiệm đúng của định luật Hardy – Weinberg. Tuy nhiên, tần số alen ở mỗi giới bị dao động qua các thể hệ và sự giao động này diễn ra theo quy luật: p ♂,q ♂ (con) = p,q (mẹ).
p ♀,q ♀ (con) = 2 1 (p♂ + p♀), 2 1 (q♂ + q♀),
2. Các dạng bài tập
2.1. Bài tập về sự cân bằng di truyền của quần thể với những gen nằm trên NST giới tính trong trường hợp tần số alen trội và lặn giống nhau ở hai giới
Ở phần này có thể có những dạng bài tập
- Xác định tần số alen, tần số phân bố các kiểu gen trong quần thể
- Xác định trạng thái cân bằng di truyền của quần thể.
- Xác định số lượng gen lặn trong quần thể.
- Xác định tỉ lệ kiểu hình, số lượng cá thể đực, cái trong quần thể…
BÀI TẬP CÓ LỜI GIẢI
Ở một quần thể côn trùng ngẫu phối, giới đực có 10% con mắt trắng, ở giới cái có 1% con mắt trắng, còn lại là những con mắt đỏ.
Hãy xác định tần số tương đối của các alen và tần số phân bố của các kiểu gen trong quần thể. Biết giới đực là XY.
Giải:
Theo bài ra trong quần thể côn trùng kiểu hình mắt trắng biểu hiện nhiều ở con đực (XY) → chứng tỏ sự di truyền màu mắt liên kết với giới tính và gen quy định tính trạng mắt là gen lặn.
Quy ước: Gen A quy định mắt đỏ
Gen a quy định mắt trắng
- Trong quần thể có 10% con đực mắt trắng có kiểu gen XaY; 1% con cái mắt trắng có kiểu gen XaXa. Ta có 10%XaY = 0,1Xa x Y (1)
1%XaXa = 0,1Xa x 0,1Xa (2)
- Từ (1) và (2) suy ra: Tần số alen a ở giới đực và giới cái đều là 0,1,
Tần số alen A là: 1 – 0,1 = 0,9.
Cấu trúc di truyền của quần thể côn trùng trên là:
0,9XA 0,1Xa
0,9XA 0,81XAXA 0,09XAXa
0.1Xa 0,09XAXa 0,01XaXa
Y 0,9XAY 0,1XaY
+Tỉ lệ kiểu gen ở giới đực là: 0,9XAY : 0,1XaY
+ Tỉ lệ kiểu gen ở giới cái: 0,81XAXA : 0,18XAXa : 0,01XaXa
+ Tỉ lệ kiểu gen chung ở cả hai giới : 0,45XAY + 0,05XaY + 0,405XAXA + 0,09XAXa + 0,05XaXa = 1.
BÀI TẬP TỰ LUYỆN
Bệnh mù màu ( mù màu đỏ và mù màu lục) do gen lặn trên nhiễm sắc thể X quy định.
Cho biết trong một quần thể người tần số nam bị bệnh mù màu là 0,08. Hãy tính tần số nữ bị mù màu và tần số nữ bình thường nhưng không mang alen gây bệnh.
(Đề thi chọn HSG quốc gia năm 2003)
2.1. Bài tập về sự cân bằng di truyền của quần thể với những gen nằm trên NST giới tính trong trường hợp tần số alen trội và lặn khác nhau ở hai giới
BÀI TẬP CÓ LỜI GIẢI
Một quần thể có cấu trúc di truyền: pAE= 0,8; qaE= 0,2. pA♀= 0,4; qa♀= 0,6
Xác định cấu trúc di truyền của quần thể ở thế hệ thứ nhất.
Sau bao nhiêu thế hệ thì quần thể đạt trạng thái cân bằng di truyền.
Giải:
Tần số chung của các alen trong quần thể là: qa= 3 2 .0,6 + 3 1 .0,2 = 0,467 => pA = 0,533
pAE= 0,8 qaE= 0,2
pA♀= 0,4 0,32AA 0,08Aa
qa♀= 0,6 0,48Aa 0,12aa
Tần số kiểu gen ở giới đực (con) bằng tần số alen ở alen ở giới cái (mẹ):
pAE = pA♀ = 0,4; qaE = qa♀ = 0,6.
Tần số kiểu gen ở giới cái: 0,32 AA + 0,56 Aa + 0,12 aa = 1
+ Tần số alen a ở thế hệ con của giới cái là: q' a♀= 0,12 + 0,56/2 = 0,4.
+ Tần số alen A ở thế hệ con của giới cái là: p A♀= 0,6.
Như vậy, qa♀ ở thế hệ bố mẹ = 0,6 => q a♀ ( con) = 0,4 = 2 1 (0,2 + 0,6).
qaE ở thế hệ bố mẹ = 0,2 => q aE ( con) = 0,6 .
qa chung = 0,467 =>q a chung = 3 2 .0,4 + 3 1 .0,6 = 0,467
Kết luận:
+ Tần số alen chung không thay đổi. Vì vậy qa = q a
+ q E = q♀ ; q ♀ ( con) = 2 1 ( qE + q♀ ).
+ q E > 0,467 > q ♀ , ngược lại q♀ >0,467 > qE. Ở những thế hệ tiếp theo tần số alen ở
hai giới đều bị dao động.
+ Hiệu giá trị q E với q ♀ ở thệ hệ con là 0,2 = 2 1 so với qE với q♀ ở thế hệ bố mẹ.
+ Như vậy, sau mỗi thế hệ thì sự chênh lệch tần số giữa hai giới giảm 2 1 và tiến tới
0 – khi đó quần thể đạt trạng thái cân bằng.
BÀI TẬP TỰ LUYỆN
Một quần thể có cấu trúc di truyền: pAE= 0,7; qaE= 0,3. pA♀= 0,5; qa♀= 0,5
Xác định cấu trúc di truyền của quần thể ở thế hệ thứ nhất.
Sau bao nhiêu thế hệ thì quần thể đạt trạng thái cân bằng di truyền.
II.THIẾT LẬP TRẠNG THÁI CÂN BẰNG CHO HAI HAY NHIỀU LOCUT GEN Xét 2 locut den dị hợp tử Aa và Bb, thì trong quần thể sẽ có 9 kiểu gen (gen cặp gen nằm trên hai cặp nhiễm sắc thể tuơng đồng khác nhau là :AABB, AABb, AaBB, Aabb, aaBB, aaBb, Aabb, aabb.
Nếu gọi p, q, r và s là tần số của các alen A, a ,B, b thì tần số các kiểu gen ở trạng thái cân bằng sẽ là kết quả triển khai của đa thức: (p + q)2 (r + s)2 = (pr + ps + pr + qs)2
Ta có: (p2AA + 2pqAa + q2aa) (r2BB + 2rsBb + s2bb) = p2 r2 AABB + 2 p2rsAABb + p2s2AAbb + 2pqr2AaBB + 4pqrsAaBb + 2s2pqAabb + q2r2aaBB + 2q2rsaaBb + q2s2aabb = 1
- pr, ps, qr, qs là tần số tương ứng của các giao tử AB, Ab, aB, ab. Khi tần số các giao tử này đạt trạng thái cân bằng thì tần số các kiểu gen cũng ở trạng thái đó.
Nếu quần thể khởi đầu là các cá thể dị hợp tử AaBb với tần số các alen như nhau (nghĩa là p = q = r = s = 0,5) thì bốn kiểu giao tử (AB,Ab,aB,ab) được sinh ra với tần số cân bằng
(AB = Ab = aB = ab = 0,25) và chỉ sau 1 thế hệ ngẫu phối là quần thể đạt trạng thái cân bằng di truyền.
Xét 1 ví dụ khác: Nếu quần thể khởi đầu có các kiểu gen AABB và aabb thì chỉ có 2 loại giao tử được sinh ra (AB và ab), cho nên trạng thái cân bằng di truyền cho mọi kiểu gen không thể đạt ngay ở thế hệ sau và thiếu hầu hết các kiểu gen (như AAbb, aaBB…).
Như vậy, nếu quần thể khởi đầu có tần số các alen không bằng nhau thì cần nhiều thế hệ mới thiết lập được tần số cân bằng cho các giao tử và trạng thái cân bằng di truyền cho quần thể. Trong trường hợp này có hai câu hỏi đặt ra là: Tần số nào là tần số cân bằng cho các giao tử? Tần số này đạt được sau bao nhiêu thế hệ ngẫu phối?
Các loại giao tử có thể được chia thành hai nhóm: giao tử “đồng trạng thái” ( như AB và ab), giao tử “đối trạng thái”(như Ab và aB). Vì tần số gen ở các giao tử “đồng trạng thái” bằng tần số gen ở các giao tử “đối trạng thái” khi chúng đạt trạng thái cân bằng, vậy ta có: AB x ab = Ab x aB.
Ví dụ, nếu tần số của mỗi alen A và B là 0,6; a và b là 0,4 thì tần số giao tử ở trạng thái cân bằng là: 0,36 x 0,16 = 0,24 x 0,24 hay 0,0576 = 0,0576
Nếu có sự sai khác giữa các giao tử “đồng trạng thái” và “đối trạng thái” thì ngay ở quần thể đầu sự khác biệt đó chính là sự khác biệt về tần số giao tử phải loại bỏ để đạt trạng thái cân bằng. Nếu ta kí hiệu độ khác biệt đó là K và là số dương, nghĩa là (Ab)(aB) –(AB)(ab) = K, thì để cân bằng, mỗi loại giao tử trong nhóm “đồng trạng thái” phải được thêm vào một lượng giao tử bằng K. Nếu K âm thì ngược lại.
Như vậy, nếu K = 0 thì tần số giao tử đạt trạng thái cân bằng.
BÀI TẬP CÓ LỜI GIẢI
Bài 1: Một quần thể có cấu trúc di truyền là 30%AABB : 30%AAbb : 30%aaBB :
10%aabb. Xác định tần số giao tử cân bằng? Giải:
Quần thể khởi đầu Kiểu giao tử Tần số khởi đầu Tần số cân bằng
AB
ab
Trong trường hợp này K= (Ab) x ( aB) – (AB) x (ab) = 0,06 và trạng thái cân bằng được thiết lập như sau:
Thế hệ Lượng giao tử thêm vào(AB và ab) hoặc bớt đi (Ab và aB) AB Ab aB ab
Nhận xét:
+ Sau mỗi thế hệ, mức độ khác biệt về tỉ lệ giao tử so với tỉ lệ cuối cùng khi đạt trạng
thái cân bằng giảm đi một nửa.
+ Nếu gặp bài tập tương tự, ta chỉ việc xác định tỉ lệ các loại giao tử khởi đầu, xác định
K → giao tử ở trạng thái cân bằng.
Bài 2: Ở một loài thực vật, alen A quy định hạt vàng, alen a quy định hạt xanh, alen B quy định hạt trơn, alen b quy định hạt nhăn.
T
ỉ lệ giao tử: AB = Ab = 0,3
aB = ab = 0,2
Cho biết các gen nằm trên các nhiễm sắc thể khác nhau. Hãy xác định tần số các alen và tỉ lệ hạt vàng nhăn và xanh trơn.
Giải:
-Tần số các alen là:
+Tần số alen A: pA = 0,3 + 0,3 = 0,6; Tần số alen a : pa = 0,2 + 0,2 = 0,4
+Tần số alen B: rB = 0,3 + 0,2 = 0,5; Tần số alen b : sb = 0,2 + 0,3 = 0,5
-Tỉ lệ các loại kiểu hình:
+ Kiểu hình vàng, nhăn có kiểu gen là : A-bb
Ta có: (0,3Ab + 0,2ab) ( 0,3Ab +0,2ab)
→ 0,09AAbb + 0,06Aabb + 0,06Aabb = 0,21
+Kiểu hình xanh, trơn có kiểu gen là aaB-
Ta có: (0,2aB + 0,2ab) (0,2aB + 0,2ab) → 0,04aaBB + 0,04aaBb + 0,04aaBb = 0,12
Vậy, tỉ lệ hạt vàng, nhăn là 0,21 và xanh trơn là 0,12.
BÀI TẬP TỰ LUYỆN
Bài 1: Một quần thể ngẫu phối có tần số alen là A= 0,6; a = 0,4 ; B = 0,5 ; b=0,5 Xác định tần số các loại giao tử AB, Ab, aB, ab và cấu trúc di truyền của quần thể ở thế hệ thứ nhất sau ngẫu phối. Cấu trúc di truyền của quần thể ở thế hệ thứ nhất có đạt trạng thái cân bằng không? Biết các alen nằm trên các nhiễm sắc thể khác nhau.
Bài 2: Một quần thể cấu trúc di truyền là: 0,4AABB : 0,6AaBb. Hãy tìm số tần số cân bằng của giao tử AB, Ab, aB, ab.
III.CÁC NHÂN TỐ LÀM THAY ĐỔI TẦN SỐ ALEN CỦA QUẦN THỂ:
A. Quá trình đột biến
1. Cơ sở lí luận: Đột biến làm cho mỗi gen phát sinh ra nhiều alen (A độtbiến A1, A2, A3 ... An) và đây chính là nguồn nguyên liệu sơ cấp cho quá trình tiến hoá.
Giả sự 1 locut có hai alen A và a. Trên thực tế có thể xảy ra các trường hợp sau:
Gen A đột biến thành gen a (đột biến thuận) với tần số u. A u a. Chẳng hạn, ở thế hệ xuất phát tần số tương đối của alen A là po. Sang thế hệ thứ hai có u alen A bị biến đổi thành a do đột biến. Tần số alen A ở thế hệ này là: p1 = po – upo = po(1-u)
Sang thế hệ thứ hai lại có u của số alen A còn lại tiệp tục đột biến thành a. Tần số alen A ơ thế hệ thứ hai là: P2 = p1 – up1 = p1(1-u) = po(1-u)2
Vậy sau n thế hệ tần số tương đối của alen A là: pn = po(1-u)n
Từ đó ta thấy rằng: Tần số đột biến u càng lớn thì tần số tương đối của alen A càng giảm nhanh.
Như vậy, quá trình đột biến đã xảy ra một áp lực biến đổi cấu trúc di truyền của quần thể. Áp lực của quá trình đột biến biểu hiện ở tốc độ biến đổi tần số tương đối của các alen bị đột biến.
Alen a cũng có thể đột biến thành A (đột biến nghịch) với tần số v.
a v A
+ Nếu u = v thì tần số tương đối của các alen vẫn được giữ nguyên không đổi.
+ Nếu v = 0 và u > 0 → chỉ xảy ra đột biến thuận.
+ Nếuu≠ v; u> 0, v > 0 → nghĩalà xảy ra cả đột biến thuận và đột biến nghịch.
Sau một thế hệ, tần số tương đối của alen A sẽ là:
p1 = po – upo + vqo
Kí hiệu sự biến đổi tần số alen A là ∆p
Khi đó ∆p = p1 – po = (po – upo + vqo) – po = vqo - upo
Tần số tương đối p của alen A và q của alen a sẽ đạt thế cân bằng khi số lượng đột
biến A→ a và a → A bù trừ cho nhau, nghĩa là ∆p = 0 khi vq = up. Mà q = 1- p.
→ up = v(1 – p) ↔ up + vp = v ↔ v u u q v u v p + = → + =
2. Các dạng bài tập
- Biết tỉ lệ kiểu hình → xác định tần số alen, tần số phân bố kiểu gen và trạng thái cân bằng của quần thể sau khi xảy ra đột biến.
- Dạng 2: Biết số lượng alen và số lượng các alen đột biến → xác định tần số đột biến gen thuận và nghịch.
- Dạng 3: Biết tần số đột biến thuận và nghịch, tổng số cá thể → Xác định số lượng đột biến.
BÀI TẬP CÓ LỜI GIẢI
Bài 1: Một quần thể động vật 5.104 con. Tính trạng sừng dài do gen A quy định, sừng ngắn do gen a quy định. Trong quần thể trên có số gen A đột biến thành a và ngược lại, với số lượng bù trừ nhau. Tìm số đột biến đó. Biết A đột biến thành a với tần số v, với u = 3v =
3.10-3
Giải:
Gọi : p là tần số của alen A và q là tần số của alen a
-Tổng số alen trong quần thể: 5.104 x 2 = 105 (alen)
-Tần số alen trội, lặn khi có cân bằng mới được thiết lập:
+Tần số alen a : qa = 3 3 uv uvvu = ++ = 0,75
+Tần số alen A : pA = 1- 0,75 = 0,25
-Số lượng mỗi alen trong quần thể:
+Số lượng alen A là: 0,25 . 105 = 2,5.104
+Số lượng alen a là: 0,75 . 105 = 7,5.104
-Số lượng đột biến thuận bằng đột biến nghịch và bằng.
3.10-3 x 2,5.104 = 75 (alen) hoặc 10-3 x 7,5.104 = 74 (alen)
BÀI TẬP TỰ LUYỆN
Bài 1:Quần thể ban đầu có 1000000 alen A và a. Tốc độ đột biến của alen A là 3.10-5, còn của alen a là 10-5. Khi cân bằng thì quần thể có số lượng của từng alen là bao nhiêu?
Cho biết không tính áp lực của các nhân tố khác làm biến đổi cấu trúc di truyền của quần thể?
Trong một quần thể gồm 2.105 alen. Tần số alen a bằng 25%. Khi quần thể có 7 alen A bị đột biến thành a và 11 alen a đột biến thành A thì tần suất đột biến trong mỗi trường hợp bằng bao nhiêu?
Bài 2: Trong một quần thể có 106 cá thể. Tần số alen a = 15 %. Trong quần thể có 5 alen A bị đột biến thành a và 7 alen a đột biến thành A thì tần số đột biến trong mỗi trường hợp bao nhiêu. Giả thiết quần thể ban đầu cân bằng
B.Quá trình chọn lọc tự nhiên (CLTN):
1. Cơ sở lí luận: a. Giá trị thích nghi và hệ số chọn lọc Mặt chủ yếu của chọn lọc tự nhiên là sự phân hoá khả năng sinh sản tức là khả năng truyền gen cho thế hệ sau. Khả năng này được đánh giá bằng hiệu suất sinh sản, ước lượng bằng con số trung bình của một cá thể trong một thế hệ.
So sánh hiệu suất sinh sản dẫn tới khái niệm giá trị chọn lọc hay giá trị thích nghi (giá trị chọn lọc hay giá trị thích ứng), kí hiệu là w), phản ánh mức độ sống sót và truyền lại cho thế hệ sau của một kiểu gen (hoặc của một alen).
Ví dụ: kiểu hình dại trội (AA và Aa để lại cho đời sau 100 con cháu mà kiểu hình đột biến lặn (aa) chỉ để lại được 99 con cháu, thì ta nói giá trị thích nghi của alen A là 100% (wA = 1) và giá trị thích nghi của các alen a là 99% (wa = 0,99).
Sự chênh lệch giá trị chọn lọc của 2 alen (trội và lặn) dẫn tới khái niệm hệ số chọn lọc (Salective coeffcient), thường kí hiệu là S.
Hệ số chọn lọc phản ánh sự chênh lệch giá trị thích nghi của 2 alen, phản ánh mức độ ưu thế của các alen với nhau trong quá trình chọn lọc
Như vậy trong ví dụ trên thì thì S = wA – wa = 1 – 0,99 = 0,01
+ Nếu wA = wa → S = 0, nghĩa là giá trị thích nghi của alen A và a là bằng nhau và tần số tương đối của alen A và a trong quần thể sẽ không đổi.
+ Nếu wA = 1, wa = 0 → S=1, nghĩa là các cơ thể có kiểu gen aa bị đào thải hoàn toàn vì đột biến a gây chết hoặc bất dục (không sinh sản được).
Như vậy, giá trị của S càng lớn thì tần số tương đối của các alen biến đổi càng nhanh hay nói cách khác, giá trị của hệ số chọn lọc (S) phản ánh áp lực của chọn lọc tự nhiên. b. Chọn lọc alen chống lại giao tử hay thể đơn bội.
- Giả sử trong 1 quần thể chỉ có 2 loại giao tử là A và giao tử mang alen a.
- Nếu CLTN chống lại giao tử mang mang alen a với hệ số chọn lọc S => Giá trị thích nghi Wa = 1 - S.
+ Tần số alen A trước chọ lọc: p
+ Tổng tần số các giao tử trước chọn lọc: p + S
+ Tổng tần số các giao tử sau chọn lọc: p + q(1 - S) = p + (1 - p)(1 - S) = p + 1 - S - p + Sp = 1 - S(1 - p) = 1 - Sq.
+ Tần số alen sau chọn lọc = Tần số alen trước chọn lọc/ Tổng tần số alen sau chọn lọc.
- 84 -
ị thíchnghi 1 1 1-S
- Đóng góp vào vốn genchungtạorathế hệ sau: p2 2pq q2(1-S) =p2+2pq+q2(1-S) =1-Sq2
0 0 0 0 0 0 0 0 0 0 0 0 0 3 0 0 0 0 0 0 0 0 0 0 1 1 2 0 0 1 1 1 . . . ( . ( 1 ; 3 1 ; 2 1 1 2 1 1 1 1 1 1 ; 1 q q q q q q n q q q q n q q q n q q q n nq q q q q q q q q q q q q q q q q q q q q q n n n n n n n n = = ⇔ = ⇔ = + ⇔ = + + = + = + = + + + = + + + = + = + =
e. Sự cân bằng giữa đột biến và chọn lọc:
* Trường hợp 1: Alen đột biến trội tăng lên với tần số u và chịu tác động của áp lực chọn lọc S.
Thế cân bằng các alen trong quần thể đạt được khi số lượng alen đột biến xuất hiện bằng số alen A bị đào thải đi, hoặc tần số các alen đột biến A xuất hiện phải bằng tần số alen A bị đào thải đi, tức là:
u = p.S → p = u S . Nếu S = 1 → p = u nghĩa là A gây chết. Lúc này tần số kiểu hình xuất hiện ra cũng biểu thị đột biến.
-
2 Sq S) -(1 q 1 - Tần số alen A sau chọn lọc:
p
Sq p
Sq - 1 Sq - 1 ) ( Sq - 1 p p q p p pq = + = + - Tốc độ biến đổi tần số alen A:
1
Sq Sq Sq Spq Spq p p p p p p p = + =
2 2 2 2 2
2 2
2
- Tốc độ biến đổi tần số alen a sau chọn lọc:
* Trường hợp 2: Các alen đột biến lặn tăng. Nếu các alen lặn không ảnh hưởng đến kiểu hình dị hợp một cách rõ rệt, thì chúng được tích luỹ trong quần thể cho đến lúc có thể
biểu hiện ra thể đồng hợp.
Thế cân bằng đạt được khi tần số alen xuất hiện do đột biến bằng tần số alen bị đào thải đi mà cá thể bị đào thải có kiểu gen aa chiếm tỉ lệ là q2 → tần số alen a bị đào thải là: q2 . S
Vậy quần thể cân bằng khi: u = q2 . S → q2 = uu q SS →=
2. Các dạng bài tập
BÀI TẬP CÓ LỜI GIẢI
Bài 1: Một quần thể ở trạng thái cân bằng về 1 gen có 2 alen A, a. Trong đó tần số p = 0,4. Nếu quá trình chọn lọc đào thải những cơ thể có kiểu gen aa xảy ra với áp lực S = 0,02. Hãy xác định cẩu trúc di truyền của quần thể sau khi xảy ra chọn lọc.
Giải:
2 2
(Giá trị âm vì chọn lọc chống lại alen a)
2
3 2 1 Sq ) Sq q Sq Sq q Sq q q q q = + = = ∆
d. Số thế hệ cần thiết để thay đổi tần số gen a từ q ở thế hệ khởi đầu thành qn :
Trường hợp S = 1
q q q q Sq q q + = = = 1 q ) Sq ) 2 2 1
- Quần thể cân bằng di truyền, nên ta có: pA + qa = 1 → qa = 1 – 0,4 = 0,6
- Cấu trúc di truyền của quần thể cân bằng là:
(0,4)2AA + 2(0,4 x 0,6)Aa + (0,6)2aa = 1 → 0,16AA : 0,48Aa : 0,36aa
-Sau khi chọn lọc thì tỉ lệ kiểu gen aa còn lại là: 0,36 (1 – S) = 0,36(1 – 0,02) = 0,3528.
Mặt khác, tổng tỉ lệ các kiểu gen sau chọn lọc là: 0,16 + 0,48 + 0,36(1 – S) = 0,9928
- Vậy cấu trúc di truyền của quần thể khi xảy ra chọn lọc là: 0,16 0,9928 AA : 0,483Aa : 0,3528 0,9928 aa ↔ 0,161AA : 0,483Aa : 0,356aa
- Các thế hệ kế tiếp 0,1,2,...,n. - 86
Câu 2: Trên một quần đảo biệt lập có 5800 người sống, trong dó có 2800 nam giới. trong số này có 196 nam bị mù màu xanh đỏ. Kiểu mù màu này là do 1 gen lặn r nằm trên NST X. kiểu mù màu này không ảnh hưởng tới sự thích nghi của cá thể. Khả năng có ít nhất 1 phụ nữ của hòn đảo này bị mù màu xanh đỏ là bao nhiêu?
Giải
Gọi p là tần số alen A (p +q = 1; p, q > 0); q là tần số alen a.
Cấu trúc di truyền ở nam: pXAY + qXaY = 1
Theo bài: qXaY = 07,0 2800 196 = => p = 1 – 0,07 = 0,93.
Cấu trúc di truyền ở nữ: p2XAXA + 2pqXAXa + q2XaXa = 1
0,8649.XAXA + 0,1302.XAXa + 0,0049XaXa = 1
Tần số cá thể nữ bình thường là: 0,8649 + 0,1302 = 0,9951
=> Tần số để 3000 cá thể nữ đều bình thường là: 0,99513000
=>Tần số để có ít nhất 1 phụ nữ bị bệnh mù màu là: 1 - 0,99513000 .
BÀI TẬP TỰ LUYỆN
Bài 1: Giả sử một quần thể động vật ngẫu phối có tỉ lệ các kiểu gen:
- Ở giới cái: 0,36 AA : 0,48 Aa : 0,16 aa
- Ở giới đực: 0,64 AA : 0,32 Aa : 0,04 aa
a) Xác định cấu trúc di truyền của quần thể ở trạng thái cân bằng.
b) Sau khi quần thể đạt trạng thái cân bằng di truyền, do điều kiện sống thay đổi, những cá thể có kiểu gen aa trở nên không có khả năng sinh sản. Hãy xác định tần số các alen của quần thể sau 5 thế hệ ngẫu phối. (Đề thi học sinh giỏi Quốc gia năm 2010)
Bài 2: Trong 1 quần thể thực vật lưỡng bội sống 1 năm ở trên đảo, tần số alen năm 1999 là
p(A) = 0,90 và q(a) = 0,10. Giả sử rằng quần thể đó có 50 cây vào thời điểm năm 2000. vậy khả năng alen a bị mất đi (nghĩa là p(A) = 1) do ngẫu phối giữa năm 1999 và 2000 là bao nhiêu?
IV. NGẪU PHỐI KHÔNG HOÀN TOÀN.
1. Cơ sở lí luận: Ngẫu phối không hoàn toàn là quần thể vừa ngẫu phối vừa nội phối. Nội phối làm tăng tỷ lệ đồng hợp tử bằng với mức giảm tỷ lệ dị hợp tử. Nội phối có thể làm thay đổi tần số kiểu gen, nhưng không làm thay đổi tần số alen. Tần số các thể đồng hợp tử cao hơn lý thuyết là kết quả của nội phối.
Nếu trong một quần thể có f cá thể nội phối thì tần số các kiểu gen bằng
(p2 + fpq)AA + (2pq – 2fpq)Aa + (q2 + fpq)aa
Hệ số nội phối được tính bằng:
1- [(tần số dị hợp tử quan sát được)/(tần số dị hợp tử theo lý thuyết)]
Hay bằng (tần số dị hợp tử theo lý thuyết – tần số dị hợp tử quan sát được)/tần số dị hợp tử theo lý thuyết.
2. Các dạng bài tập
BÀI TẬP TỰ LUYỆN
Bài 1: Trong một quần thể yến mạch hoang dại, tần số đồng hợp tử trội, dị hợp tử và đồng hợp tử lặn tương ứng là: 0,67; 0,06 và 0,27. Hãy tính hệ số nội phối trong quần thể. Giải
Tần số các alen: p = 0,67 + (1/2)(0,6) = 0,7; q = 1 – 0,7 = 0,3
Tần số dị hợp tử theo lý thuyết: 2pq = 2(0,3)(0,7) = 0,42
Bài 2: Một quần thể có tần số alen A là 0,6. Giả sử ban đầu quần thể đang đạt trạng thái cân bằng di truyền. Sau một số thế hệ giao phối thấy tần số kiểu gen aa là 0,301696. Biết trong quần thể đã xảy ra nội phối với hệ số là 0,2. Tính số thế hệ giao phối? Giải
Tần số alen a là 0,4. Do quần thể đạt trạng thái cân bằng nên cấu trúc của quần thể là: 0,301696AA+ 0,48Aa + 0,16aa = 1. Sau một số thế hệ giao phối, tần số aa là: 0,301696 =>
Tần số kiểu gen aa tăng là:
0,301696 - 0,16 = 0,141696
=> Tần số Aa đã giảm là: 0,141696 x 2 = 0,283392. Tần số Aa sau n thế hệ giao phối là: 2pq(1 - f)n = 0,48(1 - f)n = 0,48.0,8n
Tần số Aa giảm là: 0,48 – 0,48.0,8n = 0,283392
n = 4. Vậy hệ số giao phối là 4.
BÀI TẬP TỰ LUYỆN
Một quần thể ngẫu phối có tần số các alen như sau: p(A) = 0,7; q(a) = 0,3.Giả sử quần thể ban đầu đang đạt trạng thái cân bằng di truyền. Sau 3 thế hệ giao phối cấu trúc di truyền của quần thể như sau: 0,65464 AA + 0,09072 Aa + 0,25464 aa = 1. Biết rằng đã xảy ra hiện tượng nội phối. Tính hệ số nội phối?
Hệ số nội phối = 1 – (0,06/0,42) = 0,86 - 88 -
CHƯƠNG 4. DI TRUYỀN HỌC ỨNG DỤNG
I- MỘT SỐ VẤN ĐỀ VỀ CHỌN GIỐNG
- Giống là một tập hợp cá thể sinh vật do con người chọn tạo ra, có phản ứng như nhau trước cùng 1 điều kiện ngoại cảnh, có những đặc điểm di truyền đặc trưng, chất lượng tốt, năng suất cao và ổn định; thích hợp với điều kiện khí hậu, đất đai, kĩ thuật sản xuất nhất định.
- Nhiệm vụ của ngành chọn giống là cải tiến các giống hiện có, tạo giống mới có năng suất cao phẩm chất tốt nhằm đáp ứng yêu cầu ngày càng tăng của sản xuất và đời sống.
- Quy trình tạo giống mới bao gồm các bước:
+ Tạo nguồn nguyên liệu là biến dị di truyền (biến dị tổ hợp, đột biến và ADN tái tổ hợp).
+ Đánh giá kiểu hình để chọn ra kiểu gen mong muốn.
+ Tạo và duy trì dòng thuần có tổ hợp gen mong muốn.
+ Đưa giống tốt ra sản xuất đại trà.
II- NGUỒN NGUYÊN LIỆU CỦA CHỌN GIỐNG
1. Nguồn gen tự nhiên và nhân tạo
1.1. Nguồn gen tự nhiên
Đặc điểm của nguồn gen này là có sẵn trong tựnhiên. Các giống địa phương có tổ hợp
nhiều gen thích nghi với điều kiện môi trường nơi chúng sống. Trên thế giới có nhiều trung tâm phát sinh giống cây trồng. Ví dụ: Trung tâm phát sinh giống ngô và khoai tây hoang dại
ở Mehicô và Bắc Mỹ
1.2. Nguồn gen nhân tạo Đặc điểm của nguồn gen này là do con ngưới chủ động tẩo, mang tính toàn cầu. Viện nghiên cứu lúa quốc tế (IRRI) ở Philipin hằng năm thu nhận được hơn 60 000 tổ hợp mới, nơi cung cấp nhiều giống lúa năng xuất cao cho các nước sản xuất nông nghiệp.
III. CHỌN GIỐNG VẬT NUÔI, CÂY TRỒNG DỰA TRÊN NGUỒN BIẾN DỊ DI
TRUYỀN
1. Tạo giống thuần dựa trên nguồn biến dị tổ hợp
1.1. Cơ sở: Các gen nằm trên các NST khác nhau sẽ phân li độc lập với nhau, nên các tổ hợp gen mới luôn được hình thành trong sinh sản hữu tính.
1.2. Các khâu:
+ Tạo ra các dòng thuần chủng khác nhau.
+ Lai giống để tạo các biến dị tổ hợp.
+ Chọn lọc ra những tổ hợp gen mong muốn.
+ Tự thụ phấn hoặc giao phối cận huyết để tạo ra các giống thuần thuần.
1.3. Ví dụ: Các giống lúa lùn năng suất cao được tạo ra bằng cách lai các giống địa phương khác nhau.
Ví dụ: Giống lúa Peta (Indoanexia) x Giống lúa Dee-geo woo- gen (Đài Loan)
Takudan x Giống lúa IR 8 x IR – 12 – 178
IR 22 CICA4
2. Tạo giống lai có ưu thế lai cao
2.1. Khái niệm ưu thế lai
Hiện tượng con lai có năng suất, sức chống chịu, khả năng sinh trưởng và phát triển cao vượt trội so với các dạng bố mẹ được gọi là ưu thế lai.
ƯTL biểu hiện rõ nhất ở lai khác dòng và thể hiện cao nhất ở F1
Ví dụ: Lai giữa lợn Ỉ và lợn Đại Bạch cho thế hệ F1: 10 tháng tuổi nặng 100 kg, tỷ lệ nạc trên 40%.
2.2. Cơ sở di truyền của hiện tượng ưu thế lai
Giả thuyết siêu trội cho rằng ở trạng thái dị hợp tử về nhiều cặp gen khác nhau, con lai có kiểu hình vượt trội về nhiều mặt so với bố mẹ thuần chủng do có sự tác động giữa hai alen khác nhau về chức phận trong cùng một locut tạo thành hiệu quả bổ trợ, mở rộng phạm vi biểu hiện kiểu hình. (AaBbCc có kiểu hình vượt trội so với AABBCC, aabbcc, AAbbCC, AABBcc)
Ví dụ: Ở thuốc lá, aa: quy định khả năng chịu lạnh 10 oC
AA: quy định khả năng chịu nóng 35 oC.
Aa: quy định khả năng chịu nhiệt độ từ 10 oC → 35 oC. - Ưu thế lai biểu hiện rõ nhất trong lai khác dòng vì: trong mỗi dòng thuần các gen đều ở trạng thái đồng hợp tử, nên ở F1 đại bộ phận các gen đều ở trạng thái dị hợp, khi đó các gen trội (phần lớn quy định các tính trạng tốt) được biểu hiện, vì vậy F1 có ưu thế lai cao, có độ đồng đều cao về phẩm chất và năng suất.
- Ưu thế lai thể hiện rõ nhất ở F1 và giảm dần qua các thế hệ vì tỉ lệ kiểu gen dị hợp trong quần thể cao nhất ở F1, các thế hệ sau tỉ lệ kiểu gen dị hợp giảm dần nên ưu thế lai cũng giảm dần.
2.3. Phương pháp tạo ưu thế lai
a. Lai khác dòng đơn: Tự thụ phấn liên tục qua 5 – 7 thế hệ để tạo ra các dòng thuần. Lai 2 dòng thuần khác nhau sẽ được dạng ưu thế lai khác dòng A × B → C
b. Lai khác dòng kép Để tạo ra giống lai mới có đặc tính tốt của nhiều dòng thường dùng ghép lai khác dòng kép gồm nhiều dạng khởi đầu tham gia :
ABC CGH DEG ×→ ×→ ×→
c. Lai thuận và lai nghịch
Ưu thế lai phụ thuộc vào cả đặc tính của tế bào chất. Vì vậy, phép lai thuận và lai nghịch cho hiệu quả ưu thế lai không giống nhau → lai thuận và lai nghịch để xác định xem hướng lai nào tạo ra cá thể lai có giá trị nhất.
2.4. Biện pháp duy trì và củng cố ưu thế lai
+ Đối với cây trồng có thể sử dụng sinh sản sinh dưỡng thay thế cho sinh sản hữu tính. + Ở vật nuôi, ưu thế lai được duy trì, củng cố bằng lai luân phiên, con lai tạo ra trong mỗi thế hệ được lần lượt cho lai trở lại với dạng bố, mẹ ban đầu.
3. Tạo giống bằng phương pháp gây đột biến nhân tạo
3.1. Quy trình: gồm 3 bước
+ Xử lí mẫu vật bằng tác nhân đột biến
+ Chọn lọc các cá thể đột biến có kiểu hình mong muốn
+ Tạo dòng thuần chủng
- Lưu ý: phương pháp này đặc biệt có hiệu quả đối với vi sinh vật
3 2. Gây đột biến bằng các tác nhân vật lý
Các tia phóng xạ Tia tử ngoại Sốc nhiệt Loại tác nhân Tia X, tia γ, tia β, chùm nơtron
Là loại bức xạ có bước sóng ngắn từ 1000- 4000A0 Nhiệt độ tăng hoặc giảm đột ngột
Cơ chế
Nguyên tắc sử dụng
Kích thích và ion hóa các
nguyên tử khi chúng đi qua các tổ chức, tế bào sống → thay đổi cấu trúc
của phân tử ADN→ gây
ra đột biến gen, đột biến NST.
Chiếu xạ với cường độ,
liều lượng phù hợp để xử
lý lên hạt khô; hạt nảy
mầm; đỉnh sinh trưởng
của thân, cành hoặc hạt phấn; bầu nhụy.
Kích thích (không gây ion
hoá) phân tử ADN làm thay
đổi cấu trúc của phân tử ADN → gây ra đột biến gen, đột biến NST.
Đặc biệt tia có bước sóng 2570Ao (tia ADN hấp thu)
Không có khả năng xuyên
sâu nên chỉ dùng để xử lí vi sinh vật, bào tử và hạt phấn
3.3. Gây đột biến bằng tác nhân hóa học:
Chất hóa học gây đột biến gen
Loại tác nhân 5BU, EMS, MMS, NMU, nhiều loại thuốc diệt cỏ, thuốc trừ sâu, thuốc nhuộm...
Một số hoá chất khi thấm vào tế bào có khả năng thay thế, làm mất đi hoặc thêm 1 nuclêôtit vào
Làm cơ chế nội cân bằng của cơ thể để tự bảo vệ không khởi động kịp, gây chấn thương bộ máy di truyền.
- Xử lí các tác nhân lí hoá thu được các giống lúa, đậu tương...có nhiều đặc tính quý
- Sử dụng cônxisin tạo được cây dâu tằm tứ bội
- Táo gia lộc xử lí NMU → táo má hồng cho năng suất cao...
* Đối với vật nuôi: Phương pháp gây đột biến chỉ được sử dụng hạn chế ở một số nhóm động vật thấp, khó áp dụng cho các nhóm động vật bậc cao do chúng phản ứng rất nhạy và
dễ bị chết khi xử lí bằng các tác nhân lí hóa.
4. Tạo giống bằng công nghệ tế bào
4.1. Công nghệ tế bào thực vật: có bốn kĩ thuật nuôi cấy tế bào thực vật
Vấn đề phân biệt Nuôi cấy hạt phấn
Nguồn nguyên liệu
Nuôi cấy tế bào TV in -vitro tạo mô sẹo (Nuôi cấy mô)
Chọn dòng tế bào xôma có biến dị
Lai tế bào sinh dưỡng (Dung hợp tế bào trần)
Hạt phấn (n) Tế bào (2n) Tế bào (2n) 2 dòng tế bào có bộ NST 2n của hai loài khác nhau.
Cơ chế
ADN → gây đột biến gen. Mỗi chất chỉ làm mất
hoặc thay thế 1 loại nuclêôtit nhất định.
(Ví dụ: 5BU: thay A - T = G-X;
EMS : cặp G = X bị thay thành T =A hoặc X- G)
Chất hóa học gây đột biến đa bội thể
Cônxixin...
Làm rối loạn cơ chế hình thành
thoi vô sắc, dẫn đến NST đã
nhân đôi nhưng không phân li → bộ NST tăng gấp 2→ tạo tế bào đa bội.
Cách tiến hành
Nuôi hạt phấn trên môi trường nhân tạo, chọn lọc các dòng tế bào đơn bội có biểu hiện tính trạng mong muốn khác nhau. Lưỡng bội hoá các dòng đơn bội tạo các cây lưỡng bội.
Nuôi cấy tế bào hoặc mô trên môi trường nhân tạo, tạo mô sẹo rồi bổ sung hoocmôn kích thích sinh trưởng cho phát triển thành cây trưởng thành.
Nuôi cấy tế bào (2n) trên môi trường nhân tạo, chọn lọc các dòng tế bào có đột biến gen và biến dị số lượng NST khác nhau.
Tạo tế bào trần, cho dung hợp hai khối nhân và tế bào chất thành một, nuôi trong môi trường nhân tạo rồi cho phát triển thành cây lai.
Nguyên
tắc sử dụng
- Đối với thực vật: Ngâm hạt khô hay hạt nảy mầm trong dung dịch hoá chất có
nồng độ thích hợp hoặc quấn bông có tẩm dung dịch hoá chất vào đỉnh sinh trưởng thân hay chồi. Có thể dùng hoá chất ở trạng thái hơi.
- Đối với vật nuôi: Cho hoá chất tác động lên tinh hoàn hoặc buồng trứng.
3.4. Một số thành tựu tạo giống ở Việt Nam
* Trong chọn giống vi sinh vật: Phương pháp gây đột biến và chọn lọc đóng vai trò chủ yếu.
Xử lí các tác nhân lí, hoá đã thu được nhiều chủng vi sinh vật có các đặc tính quý.
Ví dụ: Xử lí bào tử của nấm Penicillium bằng tia phóng xạ rồi chọn lọc, người ta đã tạo được chủng Penicillium có hoạt tính pênixilin tăng gấp 200 lần dạng ban đầu. Trên nấm men, vi khuẩn, người ta đã chọn tạo được các thể đột biến sinh trưởng mạnh để sản xuất sinh khối→ chọn được những chủng vi sinh vật không gây bệnh, đóng vai trò một kháng nguyên, gây miễn dịch ổn định cho kí chủ chống loài vi sinh vật đó, trên nguyên tắc này đã tạo được những vacxin phòng bệnh cho người và gia súc.
* Trong chọn giống cây trồng
Ưu thế
Cây lưỡng bội tạo ra có kiểu gen đồng hợp tử về tất cả các gen.
Nhân nhanh các giống cây quý hiếm từ một cây có kiểu gen quý, tạo nên một quần thể cây trồng đồng nhất về kiểu gen.
4.2. Công nghệ tế bào động vật a. Sản xuất vacxin tổng hợp bằng công nghệ tế bào:
Tạo các giống cây mới có kiểu gen khác nhau từ một giống ban đầu
Tạo ra ra cơ thể lai mang bộ NST của 2 loài khác xa nhau mà bằng phương pháp lai hữu tính không thể thực hiện được.
- Nguyên tắc nuôi cấy tế bào tạo kháng thể cho sản xuất vacxin:
+ Sử dụng loại tế bào ung thư có dòng tế bào phân chia liên tục.
+ Cho lai tế bào ung thư với tế bào động vật có vú có chức năng sản sinh kháng thể.
+ Nuôi cấy tế bào lai để chúng sinh sản liên tục, lâu dài, tạo ra khối lượng lớn kháng thể
- Ưu điểm: Tạo kháng thể có độ tinh khiết tuyệt đối b. Sản xuất vật nuôi bằng công nghệ tế bào:
- 92 -
Áp dụng công nghệ tế bào trong sản xuất vật nuôi chủ yếu là hình thức cấy truyền hợp tử và nhân bản vô tính.
* Cấy truyền hợp tử: còn gọi là công nghệ tăng sinh sản ở động vật. Sau khi phôi đượclấy ra từ động vật và trước khi khi cấy phôi vào động vật cần trải qua các bước sau:
- Bằng kĩ thuật chia tách phôi động vật thành hai hay nhiều phôi rồi cấy các phôi này vào
tử cung của các con vật khác nhau, có thể tạo ra được nhiều con vật có kiểu gen giống nhau.
- Phối hợp hai hay nhiều phôi thành một thể khảm.
- Làm biến đổi các thành phần trong tế bào của phôi khi mới phát triển theo hướng có lợi cho con người.
* Nhân bản vô tính: Điển hình cho kĩ thuật này là nhân bản thành công con cừu Đôli (Dolly).
- Các bước tiến hành:
+ Tách tế bào tuyến vú của cừu cho nhân, nuôi trong phòng thí nghiệm
+ Tách tế bào trứng của cừu khác, loại bỏ nhân của tế bào này
+ Chuyển nhân của tế bào tuyến vú vào tế bào trứng đã bỏ nhân
+ Nuôi cấy trên môi trường nhân tạo để trứng phát triển thành phôi
+ Chuyển phôi vào tử cung của một cừu mẹ để nó mang thai
Cừu con sinh ra có kiểu hình giống hệt kiểu hình của cừu cho nhân tế bào.
- Ý nghĩa:
+ Nhân nhanh giống vật nuôi quý hiếm hoặc tăng năng suất trong chăn nuôi.
+ Tạo ra các động vật mang gen người nhằm cung cấp cơ quan nội tạng để thay thế, ghép
nội quan cho người bệnh.
5. Tạo giống bằng công nghệ gen
5.1. Khái niệm công nghệ gen
Công nghệ gen hay kĩ thuật di truyền bao gồm các kĩ thuật thao tác trên vật liệu di truyền để điều chỉnh, sửa chữa, tạo ra gen mới, từ đó tạo ra cơ thể mới với những đặc điểm mới.
Hiện nay công nghệ gen được thực hiện phổ biến là tạo ra các phân tử ADN tái tổ hợp để
chuyển gen.
5.3. Các khâu cơ bản trong kĩ thuật chuyển gen a. Tạo ADN tái tổ hợp
- Tách chiết thể truyền và gen cần chuyển ra khỏi tế bào (phân lập gen).
- Xử lí bằng một loại enzim giới hạn để tạo ra cùng 1 loại đầu dính.
- Dùng enzim nối để gắn chúng tạo ADN tái tổ hợp b. Đưa ADN tái tổ hợp vào tế bào nhận
- Dùng muối CaCl2 hoặc xung điện cao áp làm giãn màng sinh chất của tế bào để ADN tái tổ
hợp dễ dàng đi qua
c. Phân lập dòng tế bào chứa ADN tái tổ hợp
- Chọn thể truyền có các dấu chuẩn dễ nhận biết hoặc dùng gen “đánh dấu” hay gen “thông báo”.
- Bằng các kỹ thuật nhất định nhận biết được các tế bào có ADN tái tổ hợp dựa vào sản phẩm đánh dấu
5.2. Các công cụ và kĩ thuật của công nghệ gen
5.2.1.
VECTƠ TÁCH DÒNG (THỂ TRUYỀN): là phương tiện chuyển gen
Thường là các phân tử ADN nhỏ, cho phép gắn các gen (ADN) ngoại lai, có khả năng
tự sao chép, tồn tại độc lập trong tế bào chủ và đặc biệt phải mang tín hiệu nhận biết trong tế bào đã mang vectơ tái tổ hợp.
Các loại vectơ tách dòng (thể truyền) thường dùng là:
- Plasmit: có nguồn gốc vi khuẩn
- Phage: có nguồn gốc virut
- Cosmit: Thể truyền lai, có cả thuộc tính của plasmit và phage.
- Ti-plasmit: dùng để nhân dòng và chuyển gen ở thực vật.
- Các NST nhân tạo...
Các thể truyền khác nhau về một số thuộc tính phân tử, loại tế bào chủ và kích thước tối đa các đoạn ADN mà chúng có thể mang.
a. Plasmit: Plasmit là những cấu trúc nằm trong tế bào chất của vi khuẩn. Tùy loài vi khuẩn, mỗi tế bào chứa vài đến vài chục plasmit. Plasmit chứa ADN dạng vòng, có khả năng nhân đôi độc lập với hệ gen của tế bào và trong một tế bào, mỗi loại plasmit thường có nhiều bản sao.
- Đặc điểm của các thể truyền có nguồn gốc từ E.coli plasmit:
+ Có một trình tự khởi đầu tái bản ADN (ori). Trình tự này là thiết yếu để plasmit có thể tái bản trong tế bào E.coli
+ Có một hoặc một số vị trí giới hạn đặc thù. Đây là điểm để cài các đoạn ADN (hoặc gen) cần chuyển vào thể truyền.
+ Có một gen chỉ thị chọn lọc. Gen này biểu hiện như một gen trội, nhờ vậy nó giúp phân biệt được dễ dàng tế bào E.coli mang ADN tái tổ hợp.
- Ưu điểm:
+ Cấu trúc tương đối đơn giản, kích thước nhỏ.
+ Dễ tinh sạch và phân tích sản phẩm ADN tái tổ hợp
+ Có thể nhân lên một số lượng lớn trong tế bào chủ với tốc độ nhanh, do vậy hiệu suất nhân dòng cao.
- Nhược điểm:
+ Hiệu suất biến nạp ADN tái tổ hợp ở nhiều loại tế bào chủ (không phải là vi khuẩn) thấp.
+ Không mang được các đoạn ADN lớn (>10kb)
b. Phage: Phần lớn các thể truyền phage có nguồn gốc từ ADN hệ gen của phage λ, phage M13.
- Ưu điểm:
+ Hiệu quả biến nạp ADN tái tổ hợp vào tế bào chủ thường cao hơn thể truyền plasmit (do phage khả năng tự lây nhiễm, đóng gói phân tử ADN tái tổ hợp và giải phóng khỏi tế bào chủ).
+ Có thể chuyển gen hiệu quả vào cả tế bào vi khuẩn và một số tế bào sinh vật nhân thực.
+ Có thể mang được các đoạn ADN có kích thước lớn hơn thể truyền plasmit (tới ∼ 30kb)
+ Rất bền khi được bảo quản ở nhiệt độ 4oC. Các dòng virút mang ADN tái tổ hợp có thể bảo quản một thời gian dài (nhiều năm) khi chưa có nhu cầu sử dụng.
- Nhược điểm:
+ Kích thước lớn (so với thể truyền plasmit) làn việc phân tích trình tự phức tạp hơn.
- 94 -
+ Số bản sao của phage trong mỗi tế bào chủ thường thấp hơn so với thể truyền plasmit.
c. Vectơ nhân dòng Ti-plasmit: là một plasmit kích thước lớn ( 200kb) tìm thấy ở vi khuẩn Agrobacterium tumefaciens gây nốt sần ở thực vật.
Ti-plasmit là vectơ chuyển gen vào tế bào thực vật d. Các vectơ NST nhân tạo
- NST nhân tạo nấm men (YAC): Là thể truyền có khả năng tự tái bản trong tế bào nấm men.
Cấu trúc gồm:
+ Đầu mút NST (TEL) có ở 2 đầu của mỗi vectơ. Cấu trúc đầu mút này là cần thiết để NST
có thể bền vững và ổn định trong tế bào nấm men.
+ Tâm động của nấm men (CEN). Trình tự này là thiết yếu để NST nhân tạo có thể phân li chính xác về các tế bào con khi tế bào nấm men phân chia.
+ Mỗi vai của NST nhân tạo có một gen chỉ thị để phát hiện được tế bào nấm men mang YAC.
+ Một trình tự khởi đầu tái bản. Trình tự này giúp vectơ có thể tái bản trong tế bào nấm men.
+ Một vị trí đa nhân dòng mang các trình tự nhận biết duy nhất của các enzim giới hạn được dùng làm điểm gắn ADN cần nhân dòng.
- NST nhân tạo vi khuẩn (BAC): được dùng để nhân dòng các đoạn ADN kích thước lớn đến
300kb trong tế bào E.coli. BAC chứa:
+ Một trình tự khởi đầu sao chép của một plasmit có trong tự nhiên
+ Một vị trí đa nhân dòng
+ Một gen chỉ thị chọn lọc
+ Ngoài ra còn có thêm một số trình tự chức năng khác .
e. Cosmit:
- Là 1 vectơ nhân tạo gồm 1 phần có cấu trúc plasmit với hai đầu cos của phage λ giúp vectơ
có thể tự đóng gói thành phân tử ADN vòng sau khi được biến nạp vào tế bào chủ.
- Vectơ này có thể mang các đoạn ADN ngoại lai có kích thước lớn khoảng 40 - 50kb.
- Vectơ này vừa được thừa hưởng khả năng tự tái bản của plasmit vừa có khả năng tự đóng gói của phage.
f. Vectơ con thoi: là nhóm vectơ vừa có khả năng tái bản trong tế bào vi khuẩn vừa có khả năng biến nạp và biểu hiện chức năng trong các tế bào động vật có vú và các tế bào nhân thực khác. Nói cách khác chúng có thể dùng để biến nạp vào hai hay nhiều loại tế bào chủ khác nhau.
5.2.2. ENZIM CẮT, ENZIM NỐI ENZIM CẮTGIỚI HẠN (restrictaza): là các enzim có khả năng nhận biết một đoạn trình tự trên phân tử ADN và cắt ADN ở những điểm đặc hiệu.
Enzim cắt giới hạn chia làm 2 nhóm chính tùy thuộc vào kiểu cắt ADN của chúng:
- Enzim cắt giới hạn đầu so le: là enzim có khả năng nhận biệt đoạn trình tự nhưng lại cắt ở các vị trí lệch nhau giữa 2 mạch đơn theo kiểu cắt chữ Z. Khi sử dụng enzim cắt giới hạn đầu so le cắt 2 nguồn ADN tạo ra các đầu dính, từ đó có thể nối các đoạn ADN bị cắt lại với nhau.
- Enzim cắt giới hạn đầu bằng: là enzim có khả năng nhận biết đoạn trình tự và cắt ngay tại vị trí giới hạn đó để tạo đầu bằng nhau.
- Cần phải sử dụng enzim nối ligaza hoặc các đoạn nối chuyên dụng cho mỗi loại enzim.
ENZIM NỐI (ligaza): xúc tác phản ứng nối tạo liên kết photphodieste giữa hai nuclêôtit liên tiếp. 5.2.3. TẾ BÀO CHỦ: Mục đích là sử dụng bộ máy di truyền của tế bào chủ để sao chép ADN tái tổ hợp thành một lượng lớn bản sao Người ta thường dùng 2 loại tế bào chủ chính:
- Vi khuẩn E.coli: dễ nuôi cấy, dễ thao tác, ít tốn kém lại sinh sản nhanh tạo dòng ADN tái tổ hợp nhanh.
- Tế bào động vật, thực vật nuôi cấy hoặc tế bào nấm men. Loại tế bào chủ này thường dùng vào các mục đích cụ thể 5.2.4. PHƯƠNG PHÁP CHUYỂN GEN
a. Chuyển gen trực tiếp: là phương pháp sử dụng các kĩ thuật như kĩ thuật siêu âm, kĩ thuật xung điện, vi tiêm, bắn gen... để nạp gen là vào tế bào chủ.
- Kĩ thuật siêu âm: là kĩ thuật dùng máy siêu âm để đưa ADN ngoại lai xâm nhập vào bộ gen tế bào trần của vật chủ
- Kĩ thuật xung điện: là kĩ thuật sử dụng dòng điện cao áp khoảng 500V/cm với thời gian 45%o giây tạo các lỗ thủng trên tế bào trần làm cho gen lạ bên ngoài dễ xâm nhập vào bộ gen của tế bào chủ
- Kĩ thuật vi tiêm: là kĩ thuật sử dụng một lượng nhỏ ADN (các gen) tiêm vào tế bào chủ hoặc tiêm vào tế bào trứng đã thụ tinh ở giai đoạn phôi có 4-8 tế bào.
- Kĩ thuật bắn gen: là kĩ thuật sử dụng sử dụng thiết bị bắn vi đạn mang gen cần chuyển (súng bắn gen) vào bộ gen của tế bào chủ. Vi đạn là các hạt vonfram hoặc vàng trộn với gen cần chuyển và phụ gia làm gen chuyển bao quanh vi đạn. Vi đạn được gắn vào đầu viên đạn lớn hơn, sau đó được nạp vào súng bắn gen. Súng bắn gen có lưới thép mịn ở đầu nòng cho phép khi bắn thì viên đạn lớn được giữ lại, còn vi đạn được bắn vào tế bào với gia tốc lớn...
Gen cần chuyển được bắn vào có thể tái tổ hợp với bộ gen của tế bào chủ, tạo nên tế bào mang gen được chuyển, từ đó tạo ra cơ thể chuyển gen.
b. Chuyển gen gián tiếp: là kĩ thuật chuyển gen (tải nạp) nhờ các nhân tố trung gian như vi khuẩn Agrobacterium hoặc virút và phage
- Chuyển gen nhờ vi khuẩn Agrobacterium: Là phương pháp sử dụng vi khuẩn gây các khối u ở thực vật. Vi khuẩn Agrobacterium chứa một plasmit lớn tạo nên khối u ở thực vật gọi là Ti-plasmit. Cây bị nhiễm Agrobacterium qua vết xước thì vi khuẩn truyền một đoạn ADN của Ti-plasmit có mang các gen sản sinh auxin, opin... cho cây, làm cây hình thành khối u. Ti-plasmit được biến đổi bằng cách loại bỏ gen gây khối u mà lại được gắn gen mới để chuyển gen vào cây trồng.
- Chuyển gen nhờ virút và phage: là phương pháp sử dụng các virut (SV 40, BPV...) hoặc các phage (phage λ, phage M13) để chuyển gen vào vi khuẩn hoặc tế bào thực vật.
V. Ứng dụng công nghệ gen trong tạo giống biến đổi gen
- Thành tựu nổi bật nhất trong ứng dụng kĩ thuật di truyền là khả năng cho tái tổ hợp thông tin di truyền giữa các loài đứng xa nhau trong bậc thang phân loại mà lai hữu tính không thể thực hiện được
- Công nghệ gen đã tạo ra các sinh vật biến đổi gen
1.Khái niệm sinh vật biến đổi gen
- 96 -
- Khái niệm: là sinh vật mà hệ gen của nó làm biến đổi phù hợp với lợi ích của con người.
- Cách làm biến đổi hệ gen của sinh vật:
+ Đưa thêm một gen lạ vào hệ gen của sinh vật.
+ Loại bỏ hoặc làm bất hoạt một gen nào đó trong hệ gen.
Ví dụ: cà chua biến đổi gen - có gen làm chín quả bị bất hoạt, vì thế quả cà chua có thể vận chuyển đi xa hoặc bảo quản lâu, không bị thối.
2. Một số thành tựu tạo giống biến đổi gen:
- Tạo các chủng vi sinh vật biến đổi gen: nhằm phục vụ các mục đích khác nhau của con người.
Công nghệ gen đã tạo các chủng vi sinh vật mới có khả năng sản suất nhiều loại sản phẩm sinh học có giá trị như axít amin, prôtêin, vitamin, enzim, hoócmôn, kháng sinh...trên quy mô công nghiệp với số lượng lớn và giá thành rẻ. ...; Công nghệ gen còn tạo các chủng vi sinh vật làm sạch môi trường như phân hủy rác thải, dầu loang...
Ví dụ:Tạo chủng vi khuẩn E.Coli sản suất hoócmôn insulin để chữa tiểu đường ở người.
Tạo chủng vi khuẩn E.Coli sản suất hooc môn somatostatin.
Chuyển gen tổng hợp kháng sinh của xạ khuẩn (sinh sản chậm) vào các chủng vi khuẩn (sinh sản nhanh) nhằm mục đích hạ giá thành thuốc kháng sinh.
Tạo chủng vi khuẩn chuyển gen có khả năng phân hủy nhanh rác thải
- Tạo động vật chuyển gen: Công nghệ gen đã tạo ra những động vật mới có năng suất và chất lượng sản phẩm cao hơn, đặc biệt tạo ra động vật chuyển gen có thể sản xuất thuốc chữa bệnh cho con người.
Ví dụ: Tạo giống cừu sản xuất prôtêin của người, tạo giống bò chuyển gen mà sữa có thể sản xuất prôtêin C chữa bệnh máu vón cục gây tắc mạch máu ở người.
- Tạo giống cây trồng biến đổi gen: bằng kĩ thuật gen người ta đã đưa nhiều gen quy
định các đặc tính quý như năng suất và hàm lượng dinh dưỡng cao, kháng sâu bệnh, kháng thuốc diệt cỏ dại và chịu được các điều kiện bất lợi, tăng thời hạn bảo quản, khó bị dập nát khi vận chuyển... vào cây trồng.
Ví dụ: Chuyển gen trừ sâu vào cây bông tạo giống cây bông kháng sâu hại.
Tạo giống lúa “gạo vàng„ có khả năng tổng hợp β– carôten (tiền chất tạo vitamin A) trong hạt.
B. CÂU HỎI VÀ BÀI TẬ
P
I. Câu hỏi và bài tập tự trả lời Câu 1. Vai trò của thể dị hợp tử trong tiến hóa và chọn giống ? Có thể dùng con lai F1 làm giống được không? Tại sao?
Câu 2. Ưu thế lai là gì? Cho ví dụ? Giải thích cơ sở di truyền của ưu thế lai? Phương pháp tạo ưu thế lai? Vì sao biểu hiện rõ nhất trong lai khác dòng? Vì sao ưu thế lai giảm dần qua các thế hệ? Nêu các biện pháp duy trì và củng cố ưu thế lai?
Câu 3. Phân biệt các phương pháp chọn giống thực vật bằng kĩ thuật nuôi cấy tế bào.
Câu 4. Trình bày quy trình tạo giống bằng phương pháp lai tế bào xôma
Câu 5. So sánh phương pháp cấy truyền phôi và nhân bản vô tính bằng kĩ thuật chuyển nhân
ở động vật.
Câu 6. Người ta có thể tái tổ hợp thông tin di truyền giữa các loài đứng xa nhau trong bậc thang tiến hoá mà lai hữu tính không thể thực hiện được bằng những cách nào?
Câu 7. Vectơ tái tổ hợp là gì? Người ta thường sử dụng những loại vectơ nào?
Câu 8. Trình bày cách sàng lọc và theo dõi sự hoạt động của gen được chuyển vào tế bào chủ?
Câu 9. Sinh vật chuyển gen là gì? Lợi ích của sinh vận chuyển gen? Cho ví dụ
Câu 10. Hoàn thành các bảng so sánh sau:
Bảng 1: Nguồn vật liệu và các phương pháp chọn tạo giống
Đối tượng Nguồn vật liệu Phương pháp
Vi sinh vật
Thực vật
Động vật
Bảng 2: Điểm khác nhau giữa tạo giống thuần dựa trên nguồn BDTH và tạo giống có ưu thế lai cao
Điểm phân biệt Tạo giống thuần dựa trên nguồn BDTH Tạo giống có ưu thế lai cao
Cách tiến hành
Cơ sở di truyền học
Ưu điểm
Nhược điểm
Thành tựu
Bảng 3: Điểm khác biệt nhau giữa chọn giống bằng phương pháp lai hữu tính và phương pháp gây đột biến
Chọn giống bằng phương pháp lai hữu tính
Chọn giống bằng phương pháp gây đột biến
Đối tượng PP tiến hành
Lịch sử
Cơ chế
Hiệu quả Đặc điểm
II. Câu hỏi và bài tập có hướng dẫn Câu 1. Nguồn biến dị di truyền của quần thể cây trồng được tạo ra bằng những cách nào?
Gợi ý trả lời: Nguồn biến dị di truyền của quần thể cây trồng được tạo ra bằng những cách tạo biến dị tổ hợp (các phương pháp lai); gây đột biến nhân tạo bằng các tác nhân vật lý và các tác nhân hoá học; tạo ADN tái tổ hợp.
Câu 2:
- 98 -
a. Có thể tạo ra dòng thuần chủng bằng những cách nào? Tại sao việc duy trì dòng thuần thường rất khó khăn?
b. Vì sao việc chọn lọc trong dòng thuần không mang lại hiệu quả?
Gợi ý trả lời:
Có thể tạo ra dòng thuần bằng những cách sau:
- Cho giao phối gần hoặc tự thụ phấn bắt buộc qua nhiều thế hệ.
- Bằng kĩ thuật nuôi cấy mô tế bào: Từ tế bào hạt phấn (n), người ta lưỡng bội hóa tạo ra tế bào (2n) và cho tái sinh cây.
Việc duy trì dòng thuần thường rất khó khăn vì các dòng thuần thường có sức sống kém do nhiều gen lặn có hại đã được đưa vào thể đồng hợp và rất khó ngăn ngừa sự giao phấn.
b. Việc chọn lọc trong dòng thuần thường không mang lại hiệu qủa vì các gen quan tâm đều ở trạng thái đồng hợp. Sự sai khác về kiểu hình lúc đó chỉ là thường biến.
Câu 3: Vì sao tự thụ phấn bắt buộc và giao phối cận huyết qua nhiều thế hệ sẽ dẫn tới thoái hóa giống? Kiểu gen như thế nào thì tự thụ phấn sẽ không gây thoái hóa? Trong chọn giống, người ta dùng phương pháp tự thụ phấn bắt buộc và giao phối cận huyết vào mục đích gì?
Gợi ý trả lời:
-Tự thụ phấn bắt buộc và giao phối cận huyết qua nhiều thế hệ sẽ dẫn tới thoái hóa giống: con cháu có sức sống kém dần biểu hiện ở sinh trưởng, phát triển chậm, chống chịu với môi trường kém, bộc lộ các tính trạng xấu, năng suất, phẩm chất giảm; ở động vật thường xuất hiện quái thai, dị hình, giảm tuổi thọ - Nguyên nhân: do tỷ lệ kiểu gen dị hợp tử trong quần thể giảm, tỉ lệ đồng hợp tử tăng dần, trong đó các gen lặn có hại được biểu hiện.
- Tự thụ phấn không dẫn đến thoái hoá khi: dòng tự thụ có nhiều cặp gen đồng hợp trội có lợi hoặc mang những đột biến lặn có lợi. VD: Trong tự nhiên có nhiều loài tự thụ phấn như đậu, lạc, lúa mì, lúa mạch... không những không tuyệt chủng mà vẫn phát triển.
- Mục đích:
+ Củng cố các tính trạng mong muốn do các gen xác định chúng ở thể đồng hợp.
+ Kiểm tra, đánh giá kiểu gen của từng dòng nhằm phát hiện, loại bỏ gen xấu, xác định
dòng ưu việt nhất làm cơ sở khoa học cho tạo giống tốt thuần chủng.
+ Tạo dòng thuần để chuẩn bị lai khác dòng tạo ưu thế lai.
Câu 4: Từ sự hiểu biết về các pha của kì trung gian hãy đề xuất thời điểm dùng tác nhân gây
ĐB gen, ĐB NST?
Gợi ý trả lời:
- Các pha của kì trung gian:
- Thời điểm xử lý đột biến:
+ Tác động vào pha S dễ gây đột biến gen (giải thích đúng)
+ Tác động vào pha G2 dễ gây đột biến số lượng NST (giải thích đúng)
Câu 5: Trình bày các bước chính sử dụng kĩ thuật cấy gen vào E.coli để sản xuất vacxin tái tổ hợp phòng chống bệnh lở mồm, long móng ở động vật móng guốc. Biết hệ gen của loại virut này có bản chất ARN và vacxin phòng bệnh là prôtêin kháng nguyên (VP1) do chính hệ gen của virut mã hóa.
Gợi ý trả lời:
- Tách ARN của virut mang gen kháng nguyên VP1
- Phiên mã ngược tạo cADN-VP1.
- Tách plasmit từ E.coli.
- Dùng enzim giới hạn cắt plasmit và VP1.
- Nối pasmit của E.coli với đoạn cADN-VP1 tạo ra plasmit tái tổ hợp.
- Biến nạp plasmit tái tổ hợp vào E.coli.
- Nuôi E.coli có plasmit tái tổ hợp để vi khuẩn sản xuất vacxin.
Câu 6: Trong kỹ thuật di truyền, việc lựa chọn vectơ plasmit cần quan tâm đến những đặc điểm nào?
Gợi ý trả lời:
- Plazmit có kích thước ngắn.
- Có gen chuẩn (gen đánh dấu).
- Có điểm cắt của enzym giới hạn.
- Có thể nhân lên nhiều bản sao trong tế bào nhận.
Câu 7: Plasmid là gì? Để có thể dùng làm thể truyền (vector) cần phải biến đổi plasmid như thế nào?
Gợi ý trả lời:
- Plasmid là những phân tử ADN, vòng, sợi kép, tự tái bản, được duy trì trong vi khuẩn như các thực thể độc lập ngoài nhiễm sắc thể
- Một số plasmid mang thông tin về việc di chuyển chính nó từ tế bào này sang tế bào khác (F plasmid), một số khác mã hóa khả năng kháng lại kháng sinh (R plasmid), một số khác mang các gen đặc biệt để sử dụng các chất chuyển hóa bất thường (plasmid phân huỷ).
- Để được dùng làm vector plasmid cần phải có:
+ Vùng nhân dòng đa vị chứa các điểm cắt cho các endonucleaza giới hạn, dùng để chèn các ADN nhân dòng.
+ Plasmid chứa gen để chọn (như gen kháng ampicillin,... )
+ Điểm khởi động sao chép hoạt động trong E. coli.
Câu 8: Trong kĩ thuật cấy gen, hãy cho biết:
- Thể truyền là gì? Vì sao thể thực khuẩn được xem là một trong các loại thể truyền lý tưởng?
- Thế nào là ADN tái tổ hợp? Nêu tóm tắt các bước tạo ADN tái tổ hợp.
Gợi ý trả lời:
a. Trong kỹ thuật cấy gen...
- Thể truyền là một phân tử ADN nhỏ có khả năng tự nhân đôi một cách độc lập với hệ gen của tế bào. Thể truyền có thể là plasmit hoặc virut
- Thể thực khuẩn được xem là loại thể truyền lý tưởng vì nó thoả mãn mọi tiêu chuẩn của thể truyền và có khả năng biến nạp vào tế bào nhận
- ADN tái tổ hợp là một phân tử ADN nhỏ được lắp rap từ các đoạn ADN lấy từ các tế bào khác nhau (thể truyền và gen cần chuyển)
- Các bước tạo ADN tái tổ hợp: Tách chiết và tinh sạch ADN các nguồn khác nhau; Cắt và nối...
Câu 9: a) Trong kĩ thuật di truyền, người ta cần phải tách được dòng tế bào mang ADN tái tổ hợp ra khỏi các loại tế bào khác. Hãy mô tả qui trình chọn lọc dòng tế bào mang ADN tái tổ hợp.
- 100 -
b) Vectơ biểu hiện dùng trong công nghệ sinh học là loại vectơ có thể giúp tạo ra nhiều sản phẩm của gen là protêin. Để đáp ứng điều này vectơ biểu hiện cần có đặc điểm gì?
Gợi ý trả lời:
a) Để tách được dòng tế bào có chứa ADN tái tổ hợp ra khỏi các loại tế bào khác người ta thường phải dùng plasmit có chứa các gen đánh dấu như các gen kháng kháng sinh. Một plasmit được dùng làm thể truyền cần phải chứa 2 gen kháng lại hai chất kháng sinh khác nhau còn tế bào nhận thì không chứa gen kháng kháng sinh. Tại một trong hai gen kháng chất kháng sinh phải chứa trình tự nhận biết và cắt của enzym cắt giới hạn. Như vậy khi dùng enzim cắt giới hạn cắt plazmit để gắn gen tạo ADN tái tổ hợp thì gen kháng kháng sinh đó sẽ bị hỏng và ADN tái tổ hợp chỉ có thể kháng lại một loại kháng sinh mà thôi.
Như vậy nếu xử lí dòng tế bào bằng loại kháng sinh sau thì có thể tách được các tế bào có
ADN tái tổ hợp b) - Vectơ biểu hiện cần có một promotơ khoẻ, tức là có ái lực cao với ARN polymeraza.
Nhờ vậy gen được phiên mã nhiều cho ra nhiều sản phẩm (protein).
- Vectơ biểu hiện là loại có khả năng tạo ra nhiều bản sao trong tế bào (véctơ đa phiên bản).
Câu 10: Trước kia người ta hay chuyển gen của người vào tế bào vi khuẩn để sản sinh ra
những protein nhất định của người với số lượng lớn. Tuy nhiên, các nhà sinh học phân tử hiện nay lại ưa dùng tế bào nấm men làm tế bào để chuyển gen của người vào hơn là dùng tế bào vi khuẩn. Giải thích tại sao?
Gợi ý trả lời: Vì tế bào nấm men là tế bào nhân chuẩn nên có enzym để loại bỏ intron khỏi ARN trong quá trình tinh chế để tạo mARN, còn tế bào nhân sơ như vi khuẩn do chúng không có gen phân mảnh nên không có enzim cắt intron
Câu 11: Trong công nghệ sinh học, người ta đã tạo được các nhiễm sắc thể nhân tạo. Theo em, cần lắp ráp các trình tự nucleotit nào để tạo nên một nhiễm sắc thể nhân tạo dạng thẳng, sao cho nó có thể hoạt động như nhiễm sắc thể bình thường trong tế bào nhân thực?
Gợi ý trả lời:
- Phải có ít nhất một trình tự khởi đầu sao chép (xuất phát tái bản) – trình tự giúp enzim nhận biết và khởi đầu quá trình tự nhân đôi ADN.
- Có trình tự nucleotit làm nhiệm vụ của tâm động (liên kết với thoi vô sắc trong quá trình phân bào).
- Có trình tự đầu mút ở 2 đầu nhiễm sắc thể để duy trì sự ổn định của nhiễm sắc thể nhân tạo, để các nhiễm sắc thể không dính vào nhau.
Câu 12: Để tổng hợp một loại prôtêin đơn giản của người nhờ vi khuẩn qua sử dụng kỹ thuật ADN tái tổ hợp, người ta có hai cách:
1) Cách thứ nhất: Tách gen mã hóa prôtêin trực tiếp từ hệ gen trong nhân tế bào, rồi cài đoạn gen đó vào plasmit của vi khuẩn nhờ enzim ligaza.
2) Cách thứ hai: Tách mARN trưởng thành của gen mã hóa prôtêin đó, sau đó dùng enzim phiên mã ngược tổng hợp lại gen (cADN), rồi cài đoạn cADN này vào plasmit nhờ enzim ligaza. Trong thực tế, người ta thường chọn cách nào? Tại sao?
Gợi ý trả lời:
Trong thực tế, người ta chọn cách thứ hai. Bởi vì:
- ADN (gen) tách trực tiếp từ hệ gen người thường mang intron, còn cADN (được tổng hợp
từ mARN trong tế bào chất) không mang intron.
- Các tế bào vi khuẩn không có khả năng cắt bỏ các intron của các gen eucaryote, nên đoạn ADN cài tách trực tiếp từ nhân không tạo ra được prôtêin bình thường.
- Đoạn ADN phiên mã ngược (cADN) chính là bản sao tương ứng của mARN dùng để dịch mã prôtêin, có kích thước ngắn hơn nên dễ tách dòng và biểu hiện gen trong điều kiện in-vitro.
Câu 13: Trong công nghệ gen, người ta có thể sản xuất được các prôtêin đơn giản của động vật có vú nhờ vi khuẩn, chẳng hạn như E. coli. Trên cơ sở các đặc điểm khác nhau về cấu trúc gen ở sinh vật nhân sơ và nhân thực, hãy nêu những cải biến cần được thực hiện ở gen được cấy, để tế bào vi khuẩn có thể sản xuất được prôtêin của động vật có vú.
Gợi ý trả lời: + Cấu trúc gen của sinh vật nhân thực khác của sinh vật nhân sơ ở chỗ:
1) có chứa các intron.
2) trình tự ADN khởi đầu phiên mã.
3) trình tự kết thúc phiên mã.
4) trình tự tín hiệu khởi đầu dịch mã.
+ Vì vậy, để tế bào vi khuẩn có thể sản xuất được protein của động vật có vú, gen động vật có vú trước khi được cấy vào E. coli thường
1) được dùng ở dạng cADN (không chứa intron)
2) cải tiến phần trình tự khởi đầu phiên mã
3) cải tiến phần trình tự kết thúc phiên mã
4) cải tiến phần trình tự khởi đầu dịch mã.
- 102 -
CHƯƠNG 5. DI TRUYỀN HỌC NGƯỜI
A. CÁC PHƯƠNG PHÁP NGHIÊN CỨU DI TRUYỀN HỌC NGƯỜI
I. NHỮNG KHÓ KHĂN TRONG NGHIÊN CỨU DI TRUYỀN Ở NGƯỜI
Nghiên cứu DT học người có những khó khăn do:
- Người sinh sản chậm, số con ít.
- Không thể áp dụng phương pháp phân tích giống lai, phương pháp gây đột biến.
- NST người có số lượng khá nhiều, kích thước nhỏ, ít sai khác về hình dạng, kích thước.
II. PHƯƠNG PHÁP PHÂN TÍCH PHẢ HỆ
1. Phả hệ là gì?
Phả hệ là các thế hệ nối tiếp nhau trong cùng 1 dòng họ
2. Mục đích của nghiên cứu phả hệ
Phương pháp phân tích phả hệ có vai trò quan trọng, dùng để theo dõi sự DT của tính trạng nhất định trên những người thuộc cùng dòng họ, qua nhiều thế hệ, xác định tính trạng là trội hay lặn, do 1 hay nhiều gen chi phối, có liên kết giới tính hay không,...
3. Phương pháp phân tích một phả hệ
Theo dõi sự di truyền một tính trạng nào đó trên những người thuộc cùng 1 dòng họ qua nhiều thế hệ; từ đó rút ra quy luật di truyền của tính trạng đó.
4. Các kí hiệu thường dùng trong nghiên cứu phả hệ
III. PHƯƠNG PHÁP LẬP BẢN ĐỒ GEN CỦA NGƯỜI
1. Phương pháp dùng đoạn khuyết Phương pháp này dựa trên nguyên tắc là sự có mặt của bất kì đoạn nào trên NST đều phải có liên quan với sự biểu hiện của một số gen trên đoạn ấy.
2. Phương pháp lai phân tử axit nuclêic

Từ tế bào người được nuôi cấy, người ta làm tiêu bản NST kì giữa.
Dùng nhiệt làm biến tính phân tử ADN trong NST.
Dùng 1 gen được tách dòng đánh dấu phóng xạ và làm biến tính ủ với tiêu bản NST trên. Các sợi đơn đánh dấu của gen tách dòng sẽ bắt cặp với gen trên NST có trình tự bazơ phù hợp. Dùng phóng xạ tự chụp người ta xác định được vị trí của gen trên NST.
3. Phương pháp lai tế bào xoma
Qua thực nghiệm lai tế bào xôma, người ta phát hiện qua 1 số lần phân bào của tế bào lai thì tế bào lai mất đi 1 cách rất nhanh chóng NST của 1 trong 2 loài.
Nguyên tắc lập bản đồ gen qua lai tế bào xôma là sự có mặt sản phẩm của 1 gen nào đấy, có tương quan với sự có mặt của NST trong tế bào.
4. Phương pháp phát hiện các dòng cần tìm ở người
Phương pháp thông dụng hiện nay là sử dụng mẫu dò oligonucleotide, nhằm xác định vài trình tự a.a ngắn (15 - 20 a.a) rồi đánh dấu phóng xạ, cho lai với thư viện gen. Dòng tái tổ hợp sẽ được phát hiện bằng phóng xạ tự ghi.
IV. NGHIÊN CỨU NST VÀ KIỂU NHÂN - KĨ THUẬT HIỆN BĂNG
V. PHƯƠNG PHÁP NGHIÊN CỨU QUẦN THỂ
VI. PHƯƠNG PHÁP NGHIÊN CỨU TRẺ SINH ĐÔI
B. DI TRUYỀN Y HỌC
I. BỆNH DI TRUYỀN PHÂN TỬ
1.Rối loạn bẩm sinh về trao đổi chất
a. Alcaptonuria
b. Phenylketonuria (PKU)
c. Bệnh Tay - Sachs
2. Bệnh di truyền về hemoglobin
Gồm các bệnh về hemoglobin dị dạng, điển hình là bệnh hồng cầu lưỡi liềm; đây là dạng Hbs , khác với HbA bình thường ở chỗ a.a glutamic ở vị trí số 7 bị thay bằng valin làm hemôglobin của nó bị khử oxi trở thành không hoà tan, hình thành những bó sợi hình ống đặc quánh, gây biến dạng hồng cầu, làm hồng cầu dễ vỡ, khó lưu thông,...
Một dạng khác là HbC, do 1 biến đổi phân tử axit glutamic của HbA được thay bằng lizin .
Bệnh Thalasemia do gen trội trên NST thường , không liên quan đến giới tính.
II.BỆNH DI TRUYỀN NST
1. Bệnh do thể ba nhiễm
a. Hội chứng Down Ở bệnh nhân Down, NST 21 không phải ở dạng cặp như người bình thường mà thuộc thể ba nhiễm. Biểu hiện bệnh lý là ngu đần bẩm sinh, trí lực giảm, không có khả năng sinh dục, vóc dáng bé, lùn, cổ rụt, má phệ,...95% trường hợp Down đều có nguyên nhân là sự không phân li của cặp NST 21 ở bố hoặc mẹ, kết quả con sinh ra thừa 1 NST ở cặp 21. Trong 1 số trường hợp, hội chứng Down còn là kết quả của chuyển đoạn trong NST của bố hoặc mẹ, con sinh ra có 3 NST 21, hoặc tối thiểu là 1 phần lớn của NST này như NST 15. Bố hoặc mẹ bệnh nhân bề ngoài thì bình thường nhưng chỉ có 45 NST riêng biệt, còn 1 NST còn lại là do các phần của NST 15 và 21 được nối với nhau. NST này không thể cặp đôi bình thường trong giảm phân.
b. Thể ba nhiễm ở cặp 13, cặp 18
Thể ba nhiễm ở cặp 13 thường gây chết sơ sinh phần lớn ở tuổi 3 tháng sau sinh. Triệu chứng bệnh là não teo đi, mất trí, điếc và nhiều dị hình khác bên ngoài. Thể ba nhiễm ở cặp 18 biểu hiện mất trí và nhiều dị tật bẩm sinh, phần lớn chết sớm.
2. Bệnh nhân NST giới tính
a. Hội chứng Turner
Bệnh nhân có 44 NST thường và chỉ 1 NST X. thường chết ở giai đoạn thai, bệnh nhân trưởng
thành thường có các dị hình: không có buồng trứng, thiếu các tính trạng thứ cấp, mất trí,...
Cơ chế hình thành: Do 1 tế bào trứng hoặc 1 tinh trùng không có NST giới tính, hoặc do mất đi 1 NST giới tính trong nguyên phân ở những lần phân cắt đầu tiên sau khi hình thành 1 hợp tử XX hoặc XY. Trường hợp thứ 2 trên thường là thuộc dạng thể khảm, có ít dị hình hơn dạng 1 . Ở dạng thể khảm, sự tạo tính cái bình thường hơn, kinh nguyệt ít rối loạn hơn và có thể có con.
Trường hợp khác lại do bệnh nhân có 1X bình thường và 1 đoạn của X thứ hai. Cả 2 cánh của NST X thứ 2 đã được chứng minh là rất cần để có sự phân hoá buồng trứng 1 cách bình thường. Bệnh nhân chỉ có cánh dài của X thứ 2 vóc dáng lùn, có các triệu chứng xôma của Turner, trong khi đó bệnh nhân chỉ có cánh ngắn của X thứ 2 lại có vóc dáng bình thường và không có nhiều các biểu hiện dị hình. Từ các sự kiện trên cho thấy Turner là do gen trên cánh ngắn của NST X quy định. b. Hội chứng Klinefelter
Bệnh nhân là nam nhưng không bình thường về tuyến sinh dục, có 1 số nét giống nữ, không có con, trí nhớ kém, thân hình cao không cân đối, chân quá dài.
Cấu trúc XXY có thể do thụ thai từ 1 tế bào trứng đặc biệt XX với tinh trùng Y hoặc từ trứng X với tinh trùng XY.
Người ta đã phát hiện nhiều dạng thừa nhiều NST giới tính hơn XXYY, XXXY, XXXYY,...
III. DI TRUY
ỀN HỌC UNG THƯ
1. Lý thuyết virut di truyền của ung thư
Virut gây ung thư khi xâm nhập vào tế bào không làm tan tế bào mà chia làm biến đổi nhiều tính chất trong tế bào, làm tế bào có khả năng phân chia không giới hạn, không chịu sự kiểm soát của cơ thể, biến đổi số lượng NST không theo quy luật , thành u ác tính.
Cơ chế sao chép ngược trong lý thuyết virut di truyền của ung thư :
Các virut chứa ARN ung thư mang mã của 1 enzim sao chép ngược, enzim này sử dụng
ARN sợi đơn của virut làm khuôn mẫu, tổng hợp nên 1 sợi ADN bổ sung, xâm nhập vào
ADN của tế bào vật chủ tạo ra 1 tiền virut.
2. Đột biến gen, đột biến NST được xem là những nguyên nhân gây ung thư Nhiều số liệu cho thấy khối u thường phát triển từ 1 tế bào bị đột biến nhiều lần làm cho tế bào không còn khả năng đáp ứng lại cơ chế điều khiển phân bào của cơ thể dẫn đến phân chia liên tục.
3. Gen ung thư
- Các gen quy định các yếu tố sinh trưởng, bình thường hoạt động chịu sự điều khiển của cơ thể để chỉ tạo ra 1 lượng sản phẩm vừa đủ đáp ứng lại nhu cầu phân chia tế bào bình thường. Khi bị đột biến, gen trở nên hoạt động mạnh hơn và tạo ra quá nhiều sản phẩm làm tăng tốc độ phân bào dẫn đến khố u tăng sinh quá mức mà cơ thể không kiểm soát nổi. ĐBG
này là ĐB trội. Những gen ung thư loại này thường không được di truyền vì chúng xuất hiện ở tế bào sinh dưỡng.
- Trong tế bào của cơ thể người bình thường còn có các gen ức chế khối u làm cho các khối u không thể hình thành được. Tuy nhiên nếu bị đột biến làm cho các gen này mất khả năng kiểm soát khối u thì các tế bào ung thư xuất hiện tạo nên các khối u. Loại này là đột biến lặn.
IV. DI TRUYỀN HỌC BỆNH AIDS
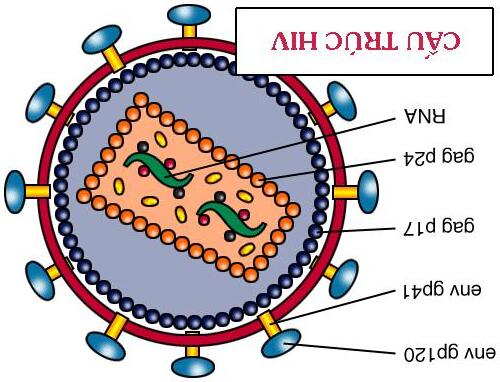
1. Cấu trúc HIV - Virut gây bệnh AIDS
Bao ngoài là 1 lớp màng lipit kép, dưới màng có gai là các protein trọng lượng phân tử 120KD kí hiệu là gp120 hay còn gọi là protein màng ngoài. Xuyên qua các lớp lipit kép là glicoprotein gp41 (Protêin xuyên màng), gp120 và gp41 gắn với nhau thành các phân tử gp160. Vỏ protein có dạng hình cầu gồm các phân tử protein 17KD .
Lõi có dạng hình trụ được bao bọc bằng 1 lớp protein p24. Trong lõi có 2 sợi ARN đơn, có enzim phiên mã ngược và 1 số phân tử protein phân tử lượng nhỏ. Trong mỗi sợi ARN của virut này có 3 gen cấu trúc là gen GAG - gen mã hoá cho các protein trong cuat virut; gen pol - mã hoá cho các enzim phiên mã ngược và gen env - mã hoá cho protein bao ngoài của virut.
2. Sự nhân lên của virut HIV trong tế bào chủ
Khi virut HIV xâm nhập vào tế bào vật chủ, nó gắn lên bề mặt của tế bào tiếp nhận hay điểm thụ thể. Phần gp20 bao ngoài virut sẽ gắn vào thụ thể trên bề mặt tế bào. Nhiều trường hợp thụ thể tế bào là CD4 có ở limpho bào T hoặc có ở 1 số tế bào khác nhau: đại thực bào, bạch cầu đơn nhân hay tế bào limpho B.
Sau đó virut gắn vào màng tế bào và hoà tan màng. Ở đây phân tử gp41 của receptor của virut cắm sâu vào màng tế bào làm cho màng của virut hoà vào màng rồi "bơm" gen của virut vào trong tế bào.
Tiếp theo là quá trình phiên mã. Nhờ enzim phiên mã ngược transcriptase, từ sợi ARN của virut tạo thành sợi ADN bổ sung (cADN).
Sự gắn kết gen: Từ cADN này làm khuôn tổng hợp thành 2 sợi ADN thẳng sau đó chuyển thành ADN xoắn vòng chui qua màng nhân gắn vào bộ gen của tế bào vật chủ nhờ
- 106 -
enzim gắn của virut integrase. Giai đoạn này tiềm tàng không biểu hiện triệu chứng nhưng chính nó đã làm thay đổi gen của tế bào vật chủ
Nhờ enzim ARN polimease tổng hợp ARN của virut từ khuôn ADN. Protein của virut cũng được tổng hợp nhờ riboxom của tế bào chủ, chúng lắp ráp tạo thành các thành phần có thể ra lưới nội chất hay tiến tới màng tế bào tạo thành các virion nằm trên màng hoặc giải phóng ra ngoài.
3. Phòng và điều trị HIV/AIDS
Hiện nay chưa có thuốc đặc trị đối với bệnh AIDS. Phòng bệnh là giải pháp tốt nhất.
Nhiều công trình nghiên cứu đã tổng hợp thành công 1 số loại protein giúp hạn chế sự xâm nhập, nhân lên của virut HIV.
Trong quá trình thử nghiệm, các prôtein tổng hợp này đã cản trở gp41 (một loại protein chủ chốt trên bề mặt của HIV), làm cho nó không thể tương tác được với các prôtein trên màng tế bào chủ (mà nó định xâm nhập), do đó HIV không thể “bám” và “chui” vào bên trong tế bào được.
V.DI TRUYỀN HỌC CHỈ SỐ THÔNG MINH IQ (INTELLIGENCE QUOTIENT)
1. Công thức IQ
Chỉ số IQ được xác định qua công thức: AM
IQ = — × 100 AR
AM :Tuổi khôn
AR : Tuổi thực
Tuổi khôn được xác định qua các test đối chiếu với bản định chuẩn để cho điểm.
2. Các phương pháp nghiên cứu IQ
a. Phương pháp nghiên cứu trẻ đồng sinh (twin studies)
Đây là phương pháp thích hợp để phân tích các hiện tượng di truyền và yếu tố môi trường trong sự hình thành tính trạng.
Một tính trạng có thể được biểu hiện ở các trẻ đồng sinh , đấy là trạng thái có sự tương hợp ; hoặc chỉ biểu hiện ở 1 trong các trẻ đồng sinh , đấy là trạng thái không tương hợp.
Có nhiều công thức khác nhau để tính tỉ lệ tham gia của yếu tố di truyền trong sự hình
thành tính trạng. Sau đây là công thức đơn giản nhất do Holzinger đề xuất:
% tương hợp cùng trứng - % tương hợp khác trứng
H% = 100 - % tương hợp khác trứng
Nếu H = 1 , tính trạng do di truyền quyết định hoàn toàn.
Nếu H = 0 , tính trạng hoàn toàn do môi trường.
Các nghiên cứu điều tra thực nghiệm đã nêu lên được số liệu tổng hợp về sự di truyền của trí thông minh qua các môi tương hợp của chỉ số IQ trong các trường hợp đồng sinh cùng trứng và khác trứng, hoặc khác môi trường sống.
Mối quan hệ Hệ số tương hợp của IQ
Đồng sinh cùng trứng
Sống chung
Sống riêng 0,87
Đồng sinh khác trứng 0,45
Anh chị em ruột Sống chung
Sống riêng 0,53 0,44
Các trẻ em không có quan hệ huyết thống sống chung 0,27
Số liệu trên cho ta khẳng định cơ sở di truyền của trí thông minh, đồng thời cũng cho thấy môi trường có vai trò không kém quan trọng.
b. Phương pháp nghiên cứu gia đình
Đây là thực tế làm cơ sở để đi sâu làm rõ 2 mặt trong sự hình thành và phát triển tài năng.
- Mặt di truyền, cơ sở gen của sự chi phối các quy luật di truyền từ tổ tiên, quy luật quay về giá trị trung bình, biến dị đột biến mới và biến dị tổ hợp.
- Mặt môi trường, bao gồm các lĩnh vực vi mô và vĩ mô, từ giai đoạn bào thai (tâm lý, tình cảm của người mẹ mang thai, dinh dưỡng, sức khoẻ người mẹ,...) đến tuổi sơ sinh, nhà trẻ mẫu giáo cho đến tuổi học đường, tuổi trưởng thành,...
Có thể thấy, thông minh, tài năng phải có nền sinh học của nó; nhưng cũng phải cần đi kèm cơ hội phát triển, ra hoa, kết quả
c. Nghiên cứu mối tương quan giữa các chỉ số huyết học với IQ
3. Cơ sở di truyền học trí thông minh
a. Quy luật di truyền nhiều nhân tố Trí thông minh là tính trạng không phải chỉ do 1 gen quy định tại riêng lẻ 1 locut, mà được chi phối bởi nhiều gen tại nhiều locut khác nhau. Các alen tương ứng của từng gen trong các locut có sự tác động lẫn nhau và tương tác với môi trường.
b. IQ và bộ não * Những vùng não tương ứng với IQ Nhiều nguồn thông tin khác nhau cùng đồng ý ở một điểm là não trước đóng một vai trò quyết định trong việc hình thành những "dòng suy nghĩ".
* Cấu trúc của bộ não và IQ
Nghiên cứu cho thấy có một sự liên quan nào đó giữa IQ và cấu trúc của vỏ não – những người có trí thông minh cao thường có vỏ não mỏng khi nhỏ và dày dần thêm khi lớn lên.
4. Quy luật quay lại giá trị trung bình
Sự di truyền của chỉ số IQ cho thấy khả năng IQ của một đứa trẻ bị ảnh hưởng từ IQ của cha mẹ. Có thể nói, một cặp bố mẹ có chỉ số thông minh cao có nhiều khả năng có những đứa con kém thông minh hơn họ.Trong khi đó, những cặp bố mẹ có chỉ số IQ có chỉ số thông minh thấp lại có xu hướng có những đứa con thông minh hơn. Hiệu ứng trên có thể được biểu diễn bởi công thức:
Trong đó:
• : chỉ số IQ dự đoán của đứa bé
• : chỉ số IQ trung bình của xã hội
• h2: hệ số di truyền của chỉ số IQ

• m và b: chỉ số thông minh của mẹ và bố đứa bé.
Vì vậy, nếu hệ số di truyền là 50%, một cặp có IQ trung bình là 120 và sống trong xã hội có IQ trung bình là 100 thì con của họ có nhiều khả năng có IQ là 110.
Đây là phương pháp thích hợp để phân tích các hiện tượng di truyền và yếu tố môi trường trong sự hình thành tính trạng.
Người ta nêu lên 2 giả thuyết về mặt di truyền:
- Giả thuyết về sự tổ hợp ngẫu nhiên các gen.
Người ta cho rằng thông minh là do nhiều gen. Có gen IQ bình thường, có gen IQ đặc biệt làm trí tuệ tăng cao. Sự tổ hợp của gen IQ bình thường dẫn đến IQ 100, nhưng nếu có 1
số gen đặc biệt, IQ được đẩy lên. VD bố có toàn gen IQ bình thường, IQ = 100, mẹ có thêm
5 gen IQ đặc biệt, IQ được đẩy lên 140, con nhận 1/2 số gen IQ từ bố và 1/2 số gen IQ từ

mẹ. Xác suất mẹ truyền lại cho con là:
1 gen IQ đặc biệt = 1/2
2 gen IQ đặc biệt = 1/2 × 1/2 = 1/4
3 gen IQ đặc biệt = 1/2 × 1/2 × 1/2 = 1/8
4 gen IQ đặc biệt = 1/16
5 gen IQ đặc biệt = 1/32
Người ta cũng giả thiết trong gia đình có gen IQ đặc biệt ở dạng lặn. Sự tổ hợp trong 1 trường hợp nào đó tạo ra thể đồng hợp về gen IQ đặc biệt.
- Giả thuyết về đột biến gen và các tổ hợp gen đột biến.
Các quy luật và giả thiết nêu trên giải thích cho các hiện tượng xảy ra trong thực tế. Có
gia đình bố hoặc mẹ thông minh nhưng con lại bình thường; hay ngược lại, bố mẹ bình thường nhưng con lại rất thông minh xuất sắc.
5. Sự phân bố IQ trong quần thể
a. Mô hình phân bố IQ
Các công trình nghiên cứu, điều tra đã cho thấy 95% dân số chung có chỉ số IQ giữa 70 - 130; khoảng 2,3 - 2,5% có IQ dưới 70 và 2,3 - 2,5% có IQ trên 130. Chỉ 1 số rất ít có IQ dưới 60 hoặc trên 150.
Như vậy, IQ trung bình trong quần thể người là 100, độ lệch chuẩn là 15IQ.
Sau đây là phân loại IQ theo Binet: IQ
≥ 140
120 - 140
90 - 110
80 - 90
70 - 80
50 - 70
25 - 50
Thiên tài
Rất thông minh
Trung bình
Hơi kém
Kém
Dốt
Đần
0 - 25 Ngu
Biểu đồ phân bố IQ trong quần thể:
Biểu hiện
b.Phân bố IQ trong thực tế
Trong thực tế sự phân bố IQ phần nào không đối xứng hoàn toàn như trong mô hình. Sự phân bố của 100 - 15 cao hơn so với 100 + 15, đường biểu diễn phía trên 100 dốc hơn phía dưới 100.
Trong các nguyên nhân thì có thể thấy hiện tượng sai khác về IQ giới tính. Chỉ số IQ của phụ nữ có phần thấp hơn nam giới nhưng chỉ số IQ ở nam giới thường dao động nhiều hơn, số nam IQ trên 130 và dưới 70 cũng nhiều hơn nữ
6. Môi trường với trí thông minh
Chính các gen cung cấp nền tảng cho trí thông minh, quy định tiềm năng IQ, nhưng môi trường lại quyết định phương hướng, khả năng biểu hiện và phát triển của tiềm năng ấy.
7. Điều kiện cực thuận cho biểu hiện, phát triển trí thông minh
Ngay sau khi mới sinh ra, não người đã ổn định về số lượng tế bào thần kinh, nhưng hệ mạng thần kinh còn rất đơn giản. Hệ mạng thần kinh này ở 15 tháng tuổi đã phức tạp hơn lên và lúc 2 tuổi hệ mạng này đã vô cùng phức tạp. Từ đó cho thấy vai trò quyết định nhất đối với sự phát triển trí tuệ trẻ thuộc giai đoạn nói trên, giai đoạn mà hệ mạng thần kinh não bộ phát triển cực nhanh.
Các thực nghiệm khách quan khoa học đã cho thấy tầm quan trọng và có tính quyết định để tạo môi trường cực thuận cho tiềm năng thông minh biểu hiện và phát triển, đó là giai đoạn trẻ từ sơ sinh đến 3 tuổi về cả 2 mặt:
- Nuôi dưỡng trẻ : chế độ dinh dưỡng đầy đủ, cân đối.
- Quan hệ tình cảm gia đình cần được quan tâm thích đáng và khoa học.
8. Các khuyết tật về trí thông minh
a. Các khuyết tật trí tuệ do rối loạn trao đổi chất.
VD: Galactosemia, Phenylketonuria, Bệnh về chuyển hoá đường.
b. Tâm thần phân liệt (Schizophrenia)
Bệnh này do 1 alen đột biến trội dạng đơn gen.
Về bản chất sinh hoá của bệnh này, người ta cho có thể do máu của người bị tâm thần phân liệt có hoạt tính monoamineoxydase thấp hơn người bình thường.
VI. DI TRUYỀN HỌC NẾP VÂN TAY
1. Cơ sở phôi sinh học nếp vân tay
Sự phân hoá nếp vân da xảy ra sớm, từ trong quá trình phát triển thai. Các dạng vân da được xác định về mặt di truyền và có chịu ảnh hưởng bởi các nhân tố mạnh của môi trường.