
History of Pakistan Reinterpreted S M Zafar
Visit to download the full and correct content document: https://textbookfull.com/product/history-of-pakistan-reinterpreted-s-m-zafar/

More products digital (pdf, epub, mobi) instant download maybe you interests ...

Islam in Pakistan A History Muhammad Qasim Zaman
https://textbookfull.com/product/islam-in-pakistan-a-historymuhammad-qasim-zaman/

Making Sense of Pakistan Farzana Shaikh
https://textbookfull.com/product/making-sense-of-pakistanfarzana-shaikh/

Peanut Agriculture and Production Technology Integrated Nutrient Management 1st Edition Zafar Abbas
https://textbookfull.com/product/peanut-agriculture-andproduction-technology-integrated-nutrient-management-1st-editionzafar-abbas/

The Judicialization of Politics in Pakistan A Comparative Study of Judicial Restraint and its Development in India the US and Pakistan 1st Edition Waris Husain
https://textbookfull.com/product/the-judicialization-of-politicsin-pakistan-a-comparative-study-of-judicial-restraint-and-itsdevelopment-in-india-the-us-and-pakistan-1st-edition-waris-
husain/

Pakistan s Inter Services Intelligence Directorate
Covert Action and Internal Operations 1st Edition Owen L. Sirrs
https://textbookfull.com/product/pakistan-s-inter-servicesintelligence-directorate-covert-action-and-internaloperations-1st-edition-owen-l-sirrs/

Courage to Say No A Female Doctor s Battle Against Sexual Exploitation in Pakistan Raana Mahmood
https://textbookfull.com/product/courage-to-say-no-a-femaledoctor-s-battle-against-sexual-exploitation-in-pakistan-raanamahmood/

Pakistan or the Partition of India 2nd Edition B. R. Ambedkar
https://textbookfull.com/product/pakistan-or-the-partition-ofindia-2nd-edition-b-r-ambedkar/

Tomb Robberies at the End of the New Kingdom: The Fayum Case—The Gurob Burnt Groups Reinterpreted Valentina Gasperini
https://textbookfull.com/product/tomb-robberies-at-the-end-ofthe-new-kingdom-the-fayum-case-the-gurob-burnt-groupsreinterpreted-valentina-gasperini/

The Jews: A History John M. Efron
https://textbookfull.com/product/the-jews-a-history-john-m-efron/





























Another random document with no related content on Scribd:
regarded as comprising two distinct types of nervous condition, according as the loop formed by the two visceral nerve-cords is twisted over itself, forming a figure of 8, or continues straight and uncrossed. In the former case, we get the condition known as streptoneurous, in the latter that as euthyneurous. [315] The Euthyneura include the whole of the Opisthobranchiata[316] and Pulmonata, the Streptoneura all the Prosobranchiata.

F�� 101 —Nervous system of the Amphineura: A, Proneomenia; B, Neomenia; C, Chaetoderma; D, Chiton; c, cerebral ganglia; l, l, lateral cords; pc, posterior commissure; s, sublingual commissure or ring, with ganglia; v, v, pedal cords. (Alter Hubrecht.)
The simplest form of nervous system in the euthyneurous Gasteropoda occurs in the Opisthobranchiata. The cerebral, pleural, and pedal ganglia tend to become concentrated in a ring-like form, united by short commissures at the posterior end of the pharynx. The visceral loop is in some cases long, and the two or three visceral ganglia are then situated at its posterior extremity. The nervous system of the Pulmonata is of a similar type, the visceral loop being often much shorter, and tending to draw in towards the central group of ganglia. The tentacular and optic nerves are, as usual, derived from the cerebral ganglion, with which also the octocysts are probably connected by rather long nerves. A pair of buccal ganglia innervate the buccal mass, and are united by commissures with the right and left cerebral ganglia. The osphradial nerve springs from
one of the ganglia on the visceral loop, the osphradium itself being situated (in Limnaea) immediately above the pulmonary orifice and adjacent to the anus (Fig. 102). This massing of the ganglia is still better illustrated by the accompanying figure of Physa (Fig. 103), in which the animal is represented as if transparent, so that the ganglia and nerves are seen through the tissues.

F�� 102 I Nervous system of Limnaea stagnalis L The oesophagus has been cut and pulled forwards through the nerve-collar, so as to expose the lower surface of the buccal mass(dissected by F. B. Stead)
B.M, buccal mass
B.G, buccal, C.G, cerebral, Os.G, osphradial, Pe.G, pedal ganglia
Pl.G, pleural ganglia
Op.N, optic, Os.N, osphradial, Te.N, tentacular nerve
Ot, otocyst; V.L, visceral loop.
R, rectum, dotted in to show its position relative to the osphiadium.
II. Right side of the head of Limnaea stagnalis. The overhanging flap of the
mantle has been cut in the middle line, and the right half twisted back, so as to expose the pulmonary orifice, etc The points A A on the mantle edge were continuous before the mantle was cut; the line BA is part of the free edge of the mantle
An, anus; F, female generative orifice; J, portion of jaw; M, male generative orifice under right tentacle; Os, osphradium; P.O, pulmonary orifice.
Of the streptoneurous Gasteropoda, the nervous system of Fissurella and Haliotis shows distinct points of similarity to that of the Amphineura. The pedal nerves are united by transverse commissures throughout their entire length, while a double commissure unites the cerebral ganglia to the mass from which the pedal nerves proceed. In the great majority of the Streptoneura the ganglia (except the visceral) are more concentrated and the commissures are consequently much shorter. The accompanying figure of Cyclostoma, in which the animal is represented as in that of Physa just described, illustrates this grouping of the ganglia, the twist of the visceral loop, and the position of the visceral ganglia at its posterior end.

F��. 103. Nervous system of Physa acuta Drap., showing the massing of the ganglia at the hinder end of the pharynx: e, e, eyes; m, mouth; m l, m l, mantle lappets; o f, female generative orifice; o m, male generative orifice; os, osphradium (After LacazeDuthiers.)

F��. 104. Example of a streptoneurous Gasteropod (Cyclostoma elegans Drap.): c.g, c g, cerebral ganglia; e, e, eyes; os, osphradium; ot, ot, otocysts; p g, p g, pedal ganglia; pl g, pl g, pleural ganglia; sp g, supraintestinal ganglion; sb g, subintestinal ganglion; t n, tentacle nerve; v g, visceral ganglion (After Lacaze-Duthiers.)
Scaphopoda.—In the Scaphopoda the nervous system resembles that of the Pelecypoda. The cerebral and pleural ganglia lie close together, while the pedal ganglia are placed in the anterior part of the foot, connected with the cerebral ganglia by long commissures; the visceral loop is rather long, and the two visceral ganglia are adjacent to the anus.
Pelecypoda.—The nervous system in the Pelecypoda is the simplest type in which well-marked ganglionic centres occur. The ganglia are few, symmetrically placed, and are usually at a
considerable distance apart. There are, as a rule, three distinct pairs of ganglia, the cerebral (cerebro-pleural), pedal, and visceral. The cerebral are formed by the fusion of the cerebral and pleural ganglia, which, however, in some cases (Protobranchiata) continue distinct. [317] They lie above or on each side of the mouth, united by a commissure of varying length. Another pair of commissures unites them with the pedal ganglia, which are placed at the base of the foot, and are usually very close together, sometimes (as in Anodonta) becoming partially fused. The length of these commissures depends upon the distance between mouth and foot; thus they are very long in Mya and Modiola, and very short in Pecten. In cases where the foot is rudimentary or becomes aborted through disuse (e.g. Ostrea), the pedal ganglia may dwindle or disappear altogether. The visceral ganglia are on the ventral side of the posterior adductor muscle, beneath the rectum, and innervate the branchiae, osphradia, and the whole of the visceral sac. A pair of cerebro-visceral commissures traverses the base of the foot, surrounding it with a comparatively short loop (compare Fig. 106, c.v.c´), while a long commissure, which runs round the entire edge of the mantle, and supplies branching nerves to the mantle border and siphons (Fig. 106, c.v.c), may also connect the visceral and cerebral ganglia.

F�� 105 Nervous system of Pelecypoda: A, Teredo; B, Anodonta; C, Pecten; a, a, cerebral ganglia; b, pedal ganglia; c, visceral ganglia. (After Gegenbaur.)
Cephalopoda.—In the Cephalopoda the concentration of ganglia attains its maximum, and may perhaps be regarded as approaching the point at which a definite brain may be said to exist. Another point of distinction is the formation of special small ganglia upon the nerve-cords in different parts of the body. In the Tetrabranchiata (Nautilus) the cerebral and pedal ganglia form a broad ring which surrounds the oesophagus, the former giving out the optic nerves, with their special optic ganglion, and a pair each of buccal and pharyngeal ganglia, the latter the nerves of the arms and funnel. The visceral loop is still present in the form of a separate band, which innervates the branchiae, osphradia, and viscera generally, forming a special genital ganglion in connexion with the reproductive organs. The principal ganglia of the Dibranchiata are still more concentrated, even the visceral loop being possibly united with the rest in forming an unbroken mass in which scarcely any trace of commissures can be detected. The pedal ganglion becomes separated into two
portions, one of which innervates the arms, the other the funnel. Two peculiar ganglia (the stellate ganglia) supply a number of branching nerves to the mantle.
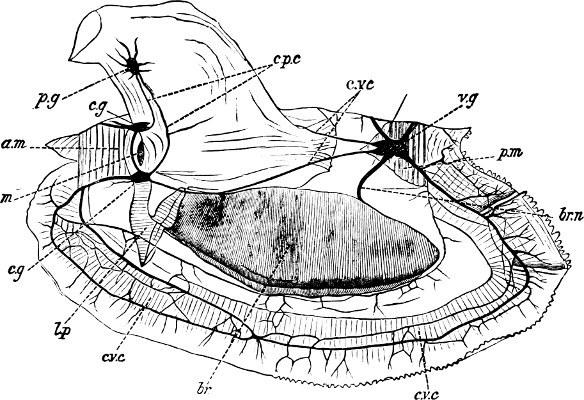
F��. 106. Nervous system of Cardium edule L.: a.m, anterior adductor muscle; br, branchiae; br.n, branchial nerve; c.g, c.g, cerebral ganglia; c.p.c, cerebro-pedal commissure; c.v.c’, cerebro-visceral commissure; c.v.c, cerebro-visceral commissure of mantle; l.p, labial palps: m, mouth; p.g, pedal ganglion; p.m, posterior adductor muscle; v.g, visceral ganglion. (After Drost, × 3.)
E. L. Bouvier, Système nerveux, morphologie générale et classification des Gastéropodes prosobranches: Ann. Sc. Nat. Zool. (7), iii. 1887, pp. 1–510.
J. Brock, Zur Neurologie der Prosobranchier: Zeit wiss Zool xlviii 1889, pp 67–83
O. Bütschli, Bemerkungen über die wahrscheinliche Herleitung der Asymmetrie der Gasteropoda, etc.: Morph. Jahrb. xii. 1886, pp. 202–222
B. Haller, Zur Kenntniss der Muriciden. I. Anatomie des Nervensystems: Denksch. Math. Nat. Kl. Ak. Wien, xlv. 1882, pp. 87–106.
„ Untersuchungen über marine Rhipidoglossen II Textur des„ Centralnervensystems und seiner Hüllen: Morph Jahrb xi 1885, pp 319–436
H. Grenadier, Abhandlungen zur vergleichenden Anatomie des Auges: Abh. Naturf. Gesell. Halle, xvi. 1884, pp. 207–256; xvii. 1886, pp. 1–64.
A. P. Henchman, The Origin and Development of the Central Nervous System in Limax maximus: Bull. Mus. C. Z. Harv. xx. 1890, pp. 169–208.
V. Hensen, Ueber das Auge einiger Cephalophoren: Zeit wiss Zool xv 1865, pp 157–242
C. Hilger, Beiträge zur Kenntniss des Gasteropodenauges: Morph. Jahrb. x. 1885, pp. 352–371.
Lacaze-Duthiers, Otocystes ou Capsules auditives des Mollusques (Gastéropodes): Arch Zool Exp Gén i 1872, pp 97–166
„ „ Du système nerveux des Mollusques gastéropodes pulmonés aquatiques: ibid. pp. 437–500.
P. Pelseneer, Recherches sur le système nerveux des Ptéropodes: Arch. Biol. vii. 1887, pp. 93–130.
„ Sur la valeur morphologique des bras et la composition du système nerveux central des Cephalopodes: Arch Biol viii 1888, pp 723–756
H. Simroth, Ueber die Sinneswerkzeuge unserer einheimische Weichthiere: Zeit. wiss. Zool. xxvi. 1876, pp. 227–348.
J. W. Spengel, Die Geruchsorgane und das Nervensystem der Mollusken: Zeit wiss Zool xxxv 1881, pp 333–383
CHAPTER VIII
THE DIGESTIVE ORGANS, JAW, AND RADULA: EXCRETORY ORGANS
The digestive tract, or, as it is often termed, the alimentary canal or gut, is a very important feature of the Mollusca. It may be regarded as consisting of the following parts: (1) a mouth or oral aperture: (2) a throat or pharynx; (3) an oesophagus, leading into (4) a stomach, (5) an intestine and rectum, ending in (6) an anus.
The primitive positions of mouth and anus were presumably at the anterior and posterior ends of the animal, as in the Amphineura and symmetrical Mollusca generally. But the modifications of original molluscan symmetry, which have already been referred to (p. 154, compare pp. 245, 246), have resulted in the anus becoming, in the great majority of Gasteropoda, twisted forward, and occupying a position on some point in the right side in dextral, and in the left in sinistral species.
The process of digestion, as the food passes from one end of the tract to the other, is performed by the aid of the secretions of various glands, which open into the alimentary canal at different points in its course. The principal of these are the salivary glands, situated on the pharynx and oesophagus, and the liver, biliary or hepatic gland, connecting with the stomach. With these may be considered the anal and ink-glands, which, in certain genera, connect with the terminal portion of the rectum.
1. The mouth is generally, as in the common snail and periwinkle, placed on the lower part of the head, and may be either a mere aperture, circular or semicircular, in the head-mass, or, as is more usual, may be carried on a blunt snout (compare Fig. 6, p. 10, and Fig. 68, p. 159), which is capable of varying degrees of protrusion. From the retractile snout has doubtless been derived the long proboscis which is so prominent a feature of many genera (compare Figs. 1, B, and 99), and in some (e.g. Mitra, Dolium) attains a length exceeding that of the whole body. As a rule, Mollusca provided with a proboscis are carnivorous, while those whose mouth is on the
surface of the head are Vegetable feeders, but this rule is by no means invariable. The mouth is thickened round the aperture into ‘lips,’ which are often extensile, and appear capable of closing upon and grasping the food. In the Pelecypoda the mouth is furnished, on each side, with a pair of special external lobes, the ‘labial palps,’ which appear to be of a highly sensitive nature, and whose object it is to collect, and possibly to taste, the food before it passes into the mouth.
2. The Pharynx, Jaws, and Radula.—Immediately behind the lips the mouth opens into the muscular throat, pharynx, or buccal mass. The pharynx of the Glossophora, i.e. of the Gasteropoda, Scaphopoda, and Cephalopoda, is distinguished from that of the Pelecypoda,[318] by the possession of two very characteristic organs for the rasping or trituration of food before it reaches the oesophagus and stomach. These are (a) the jaw or jaws, and (b) the radula, [319] odontophore, or lingual ribbon. The jaws bite the food, the radula tears it up small before it passes into the stomach to undergo digestion. The jaws are not set with teeth like our own; roughly speaking, the best idea of the relations of the molluscan jaw and radula may be obtained by imagining our own teeth removed from our jaws and set in parallel rows along a greatly prolonged tongue. [320]
In nearly all land Pulmonata the jaw is single, and is placed behind the upper lip. If a common Helix aspersa be observed crawling up the inside of a glass jar, or feeding on some succulent leaf, the position and action of the jaw can be readily discerned. It shows very black when the creature opens its mouth, and under its operation the edge of a lettuce leaf shows a regular series of little curved indentations, in shape not unlike the semicircular bites inflicted by a schoolboy upon his bread and butter. The jaw of Helix (Fig. 107, B) is arched in shape, and is strengthened by a number of projecting vertical ribs. That of Limax (A) is straighter, and is slightly striated, without vertical ribs. In Bulimulus (C) the arch of the jaw is very conspicuous, and the upper edges are always denticulated; in Orthalicus there is a central triangular plate with a number of overlapping plates on either side; in Succinea (E) there is a large
square accessory plate above the jaw proper The form of the jaw is peculiar not only to the genus but to the species as well. Thus the jaw of H. aspersa is specifically distinct from that of H. pomatia, and that of H. nemoralis is distinct from both. Wiegmann has observed[321] that in young Arion, Limax, and Helix, the jaw consists of two pieces, which coalesce by fusion in the adult, thus indicating a stage of development in advance of the double jaw which is found in most of the non-pulmonate Mollusca. In all fresh-water Pulmonata there are two small accessory side plates besides the jaw proper (Fig. 107, F).

F��. 107. Jaws of various Pulmonata: A, Limax (gagates Drap., Lancashire, × 15); B, Helix (acutissima Lam., Jamaica, × 15); C, Bulimulus (depictus Reeve, Venezuela, × 20); D, Achatina (fulica Fér , Mauritius, × 7); E, Succinea (elegans Riss , Aral District, × 30); F, Limnaea (stagnalis L , Cambridge, × 30)
Nearly all the non-carnivorous Prosobranchiata, land, fresh-water, and marine alike, are provided with two large lateral jaws. Many of these are sculptured with the most elaborate patterns, and appear to
be furnished with raised teeth, like a file. In the Nudibranchiata the jaws are of great size and beauty of ornamentation (Fig. 109).

F��. 108. Jaws of A, Triton australis Lam., Sydney; B, Ampullaria fasciata Reeve, Demerara; C, Calliostoma punctulatum Mart., New Zealand; D, Cyclophorus atramentarius Sowb., Sanghir; all × 15.

F�� 109 Jaws of A, Chromodoris gracilis Iher , × 15; B, Scyllaea pelagica L , × 7; C, Pleurobranchus plumula Mont., × 10; D, Pleurobranchaea Meckelii Lam., × 5/2.
The carnivorous genera, whether marine (e.g. Conus, Murex, Buccinum, Nassa) or land (e.g. Testacella, Glandina, Streptaxis, Ennea), are entirely destitute of jaws, the reason probably being that in all these cases the teeth of the radula are sufficiently powerful to do the work of tearing up the food without the aid of a masticatory organ as well. Jaws are also wanting in the Heteropoda, and in many of the Nudibranchiata and Tectibranchiata.
In the Cephalopoda the jaws, or ‘beaks,’ as they are called, are most formidable weapons of attack. In shape they closely resemble the beaks of a parrot, but the hook on the dorsal side of the mouth does not, as in birds, close over the lower hook, but fits under it. Powerful muscles govern these mandibles, which must operate with immense effect upon their prey (Fig. 110).

F��. 110. Jaws of Sepia: A, in situ within the buccal mass, several of the arms having been cut away; B, removed from the mouth and slightly enlarged

F�� 111 Patella vulgata L , showing the normal position of the radula, which is doubled back in a bow; the shell has been removed, and the whole visceral mass is turned forward, exposing the dorsal surface of the muscular foot: gr, longitudinal groove on this surface; i, i, intestine; l, liver; m, m, mantle edge; mu, muscles (cut through) fastening the visceral mass to the upper sides of the foot; ov, ovary; r, radula; u f, upper or dorsal surface of the foot
The Radula.[322]—When the food has passed beyond the operation of the jaw, it comes within the province of the radula, the front part of which perhaps co-operates to a certain extent with the jaw in performing the biting process. The function of the radula as a whole is to tear or scratch, not to bite; the food passes over it and is carded small, the effect being very much the same as if, instead of
dragging a harrow over the surface of a field, we were to turn the harrow points upwards, and then drag the field over the harrow.
The radula itself is a band or ribbon of varying length and breadth, formed of chitin, generally almost transparent, sometimes beautifully coloured, especially at the front end, with red or yellow.[323] It lies enveloped in a kind of membrane, in the floor of the mouth and throat, being quite flat in the forward part, but usually curving up so as to line the sides of the throat farther back, and in some cases eventually forming almost a tube. The upper surface, i.e. the surface over which the food passes, is covered with teeth of the most varied shape, size, number, and disposition, which are almost invariably arranged in symmetrical rows. These teeth are attached to the cartilage on which they work by muscles which serve to erect or depress them; probably also the radula as a whole can be given a forward or backward motion, so as to rasp or card the substances which pass over it.
The teeth on the front part of the radula are often much worn (Fig. 112), and probably fall away by degrees, their place being taken by others successively pushed up from behind. At the extreme hinder end of the radula the teeth are in a nascent condition, and there are often as many as a dozen or more scarcely developed rows. Here, too, lie the cells from which the teeth are originally formed.
The length and breadth of the radula vary greatly in different genera. In Littorina it is very narrow, and several times the length of the whole animal. It is kept coiled away like a watch-spring at the back of the throat, only a small proportion of the whole being in use. I have counted as many as 480 rows in the common Littorina littorea. In Patella it is often longer than the shell itself, and if the radula of a large specimen be freshly extracted and drawn across the hand, the action of the hooks can be plainly felt. In Aerope, the Turbinidae generally, and Haliotis it is very large. In Turritella, Aporrhais, Cylichna, Struthiolaria, and the Cephalopoda it is small in proportion to the size of the animal. In the Pulmonata generally it is very broad, the length not exceeding, as a rule, thrice the breadth; in most other groups the breadth is inconsiderable, as compared to the length.
The Radula is wanting in two families of Prosobranchiata, the Eulimidae and Pyramidellidae, which are consequently grouped together as the section Gymnoglossa. It is probable that in these cases the radula has aborted through disuse, the animals having taken to a food which does not require trituration. Thus several genera contained in both these families are known to live parasitically upon various animals—Holothurians, Echinoderms, etc. —nourishing themselves on the juices of their host. In some cases, the development of a special suctorial proboscis compensates for the loss of radula (see pp. 76–77). In Harpa there is no radula in the adult, though it is present in the young form. No explanation of this fact has yet been given. It is also absent in the Coralliophilidae, a family closely akin to Purpura, but invariably parasitic on corals, and probably nourished by their exudations. There is no radula in Entoconcha, an obscure form parasitic on the blood-vessels of Synapta, or in Neomenia, a genus of very low organisation, or in the Tethyidae, or sea-hares, or in one or two other genera of Nudibranchiata.

F�� 112 Example of a front portion of a radula (Cantharus ringens Reeve, Panama), much worn by use × 70.
The number of teeth in the radula varies greatly. When the teeth are very large, they are usually few in number, when small, they are very numerous. In the carnivorous forms, as a rule, the teeth are comparatively few and powerful, while in the phytophagous genera they are many and small. Large hooked and sickle-shaped teeth, sometimes furnished with barbs like an arrow-head, and poisonglands, are characteristic of genera which feed on flesh; vegetable feeders, on the contrary, have the teeth rounded, and blunter at the apex, or, if long and narrow, so slender as to be of comparatively little effect. Genera which are normally vegetarian, but which will, upon occasion, eat flesh, e.g. Limax and Hyalinia, exhibit a form of teeth intermediate between these two extremes (see Fig. 140, A).
In Chaetoderma there is but one tooth. In Aeolis coronata there are about 17, in A. papillosa and Elysia viridis about 19, in Glaucus atlanticus about 21, in Fiona nobilis about 28. In the common whelk (Buccinum undatum) there are from 220 to 250, in the common periwinkle about 3500. As many as 8343 have been counted in