
14 minute read
Efecto de un limitado suministro de oxígeno disuelto, sobre el uso del alimento y el rendimiento del juvenil Litopenaeusvannamei


Las caídas de oxígeno son evitadas, por los camaroneros, debido a la incidencia directa sobre el consumo del alimento que, en tiempo prolongado, disminuye el crecimiento del camarón e incrementa el factor de conversión alimenticia si no se considera este parámetro al momento de alimentar. De allí, la importancia de investigar acerca del efecto de la composición nutricional como factor influyente en la fisiología del camarón.
El cultivo de camarón es una importante actividad económica a nivel mundial (FAO, 2022), y en el Ecuador proporciona fuentes de ingresos y empleos para miles de personas (CNA, 2021). Los camarones son cultivados en piscinas expuestas, las cuales requieren que las condiciones ambientales sean las adecuadas para lograr tasas óptimas de crecimiento y supervivencia.
El oxígeno disuelto (OD) es un importante parámetro abiótico esencial para muchos procesos fisiológicos en los camarones, que incluyen la respiración, inmunidad, muda y crecimiento (Allan y Maguire, 1991; Ulaje et al., 2020; Wei et al., 2009).
Uno de los principales desafíos que enfrentan los productores de camarón, es la caída de oxígeno durante los ciclos de cultivo, pudiendo alcanzar concentraciones tan bajas como <1.5 mg L-1, considerados críticos (Chen, 1984; Guerrero- Galván et al., 1999). Estas condiciones generan estrés en el organismo, lo cual afecta su rendimiento óptimo e influye en la incidencia y prevalencia de las enfermedades (Alonzo et al., 2017; Muniesa et al., 2017). Por esta razón, el control de los niveles de oxígeno de las piscinas camaroneras es fundamental para mantener la salud y la productividad del cultivo.
El bajo nivel de OD es la principal variable limitante de la calidad del agua en la acuicultura intensiva (Boyd, 1989). En exposición prolongada, a bajas concentraciones de oxígeno, los organismos utilizan sus mecanismos de adaptación para mantener niveles normales de actividad para la búsqueda de alimento y la reproducción. Según Frey (1947), el OD es un factor metabólico regulador en los organismos acuáticos. OD puede limitar la capacidad metabólica y la producción de biomasa. En general, los camarones peneidos son oxireguladores dentro de un limitado intervalo de OD (Villarreal et al., 1994, Villarreal y Ocampo, 1993).
Por otro lado, la producción de OD está determinada por la presencia de microalgas a través de la fotosíntesis, el uso de aireadores y el recambio de agua (Boyd y Tucker, 1998). En promedio, la óptima concentración de OD se encuentra entre 4.4 y 8.6 mg L-1 (Chakravarty et al., 2016; Chen, 1984), donde la solubilidad del oxígeno va a depender también de la salinidad y la temperatura del agua (Boyd y Tucker, 1998).
Las caídas de oxígeno son evitadas por los camaroneros debido a la incidencia directa sobre el consumo del alimento que, en tiempo prolongado, disminuye el crecimiento del camarón e incrementa el factor de conversión alimenticia (FCA) si no se considera este parámetro al momento de alimentar. De allí, la importancia de investigar acerca del efecto de la composi- ción nutricional como factor influyente en la fisiología del camarón.
Descripción de lo realizado
Se colocaron camarones Litopenaeus vannamei, con peso inicial de 5.29 ± 0.02 g, en 24 tanques circulares de 500 L, donde se distribuyeron seis tratamientos por cuadruplicado a una densidad de 18 camarones m-2. Luego de un período de aclimatación a las condiciones del estudio, en los que se incluían niveles óptimos de oxígeno de 6 mg L-1, los camarones fueron alimentados manualmente con las dietas experimentales a saciedad tres veces al día: 09h00, 12h00 y 17h00 durante 8 semanas. Una vez al día, el alimento consumido fue recolectado, secado y posteriormente pesado para calcular el FCA. Los camarones fueron alimentados con una dieta experimental nutricionalmente completa (AC) y otra con menor valor nutricional (AMN), cada una enfrentada a períodos de reducción de oxígeno (<2 mg L-1) durante 4 horas consecutivas una vez a la semana, en diferentes períodos que comprendieron las últimas 2 y 4 semanas de estudio, como se detalla en la Tabla 1.

Al final de la prueba, los camarones fueron contados y pesados después de un día de ayuno, para limpiar el intestino del camarón. Los camarones se capturaron con una red y se recolectaron en baldes (un balde por tanque) que representaban cada réplica de los tratamientos; luego, fueron secados con papel absorbente y pesados de manera individual. A continuación, se evaluaron estadísticamente el peso final, la supervivencia, la biomasa final, la tasa de crecimiento específica (TCE) y FCA, primero mediante un ANOVA de dos vías para encontrar diferencias entre tipos de alimentos, niveles de restricción de oxígeno, además de estimar la existencia de interacciones, y posteriormente con ANOVA simple y prueba post-hoc LSD Fisher para encontrar diferencias significativas (p < 0.05) entre los tratamientos. Todos los análisis se realizaron con Statgraphics® Centurion™ XVII (®Copyright 19822014 Statpoint Technologies, Inc.).
Resultados del estudio
El efecto de las restricciones de oxígeno y el tipo de alimento se presentan en la Tabla 2. Un análisis ANOVA dos vías reveló que existe una diferencia significativa (p < 0.05) en el peso final, TCE dada por el tipo de alimento, las restricciones de oxígeno y las interacciones de estas dos variables.

Con el objetivo de encontrar diferencias entre los tratamientos, se realizó un análisis ANOVA de una vía y, posteriormente, la prueba post-hoc LSD Fisher se llevó a cabo para cada uno de los parámetros zootécnicos que mostraron diferencias estadísticas entre tratamientos.
En la Tabla 3 se observa el efecto que tuvo la reducción de OD en las poblaciones de camarón a lo largo del tiempo. Aquellos camarones alimentados con un misma dieta crecieron de forma similar hasta ser expuestos a caídas de oxígeno durante 2 semanas; mientras que, si se prolongaban a 4 semanas consecutivas, más pronunciada fue la disminución (11%) en rendimiento que se observaba en los camarones alimentados con un alimento completo (AC), comparado con el otro de menor valor nutricional que disminuyó a un 3% con respecto a aquellos camarones que soportaron bajones de oxígenos durante 2 semanas, lo que indicaba que el camarón no lograba sacar el máximo provecho del alimento con mayor contenido nutricional por las continuas caídas de oxígeno. Ocampo et al. (2000) estudiaron los efectos del OD y la temperatura en Farfantepenaeus californiensis, los cuales fueron mantenidos en niveles de oxígeno entre 5.8 a 2.6 mg L-1 y encontraron que el menor crecimiento se obtenía en el nivel más bajo de oxígeno. Estos autores indicaron que los niveles bajos de oxígeno deprimían significativamente el crecimiento. Estos hallazgos concuerdan con los datos presentados en este estudio, el tratamiento de 4 semanas con caídas de OD <2 mg L-1, mostró que los organismos no son capaces de obtener suficiente energía de los alimentos y metabolizar el efecto limitado sobre el nivel crítico de OD.

Los datos de supervivencia no presentan una relación con el tiempo de restricción de oxígeno a los niveles ensayados, tampoco se detectaron tendencias por tipo de alimento durante estas semanas.
Aunque no se detectaron diferencias estadísticas (p > 0.05) en cuanto a eficiencia alimenticia entre tratamientos, una tendencia al aumento del FCA sí fue observada cuando el nivel de OD se restringió en un período de 4 semanas, comparado con la restricción en 2 semanas (Tabla 3). Resultados similares en términos de eficiencia de conversión alimenticia fueron reportados por Wei et al. (2009) en Fenneropenaeus chinensis a niveles de OD de 2 mg L-1. Por otro lado, Allan y Maguire (1991) en Penaeus monodon enfrentado a diferentes niveles de estrés por niveles de OD (0.5 o 1.0 mg O2 L-1) y tiempos de exposición de 4, 8 y 12 h, no observaron diferencias estadísticas en el FCA. Estas diferencias pueden deberse a las distintas especies estudiadas, ya que cada especie de camarón tiene su propia tasa de consumo de oxígeno (Rosas et al., 2001).

Cuando se analizó la biomasa ganada (Figura 1), no se observaron diferencias significativas (p > 0.05) entre tratamientos; sin embargo, cuando el oxígeno fue restringido en 2 semanas, un incremento del 18% fue observado usando alimento AC en comparación al alimento AMN. Una restricción más prolongada a 4 semanas, afectó negativamente la ganancia en biomasa; bajo estas circunstancias, los camarones mantenidos con el alimento completo AC, se vieron afectados al reducir su rendimiento al 8% en comparación con aquellos alimentados con la dieta de menor contenido nutricional. Según Fry (1947), el OD es un factor metabólico regulador en los organismos acuáticos, OD puede limitar la capacidad metabólica y la producción de biomasa.


Se observaron diferencias estadísticas (p > 0.05) cuando se analizaron los datos de TCE correspondientes a las semanas en las que se restringió el oxígeno (Figura 2). La tasa más alta se consiguió con el alimento nutricionalmente completo en el periodo corto de restricción (2 semanas), mientras que, a mayor tiempo de restricción (4 semanas), la tasa de crecimiento con alimento AC dio un menor valor, muy similar a la alcanzada por el alimento AMN.
Conclusiones
En el presente estudio, las restricciones temporales de oxígeno sugieren que existe un 12% de mejor rendimiento del alimento nutricionalmente completo en términos de peso final y TCE comparados con el alimento de menor valor nutricional, hasta cuando hubo restricciones de OD en las últimas 2 semanas. Sin embargo, esta diferencia en peso final cayó al 3% cuando las restricciones temporales de OD fueron durante las últimas 4 semanas del ensayo de alimentación. Esto indicaría que, las exposiciones crónicas a bajas concentraciones no letales de OD por pocas horas afectan el rendimiento de L. vannamei, al destinar más energía para el balance bioenergético y trabajo metabólico que para el crecimiento, como por ejemplo, concentraciones de OD <2 mg L-1 por períodos de 4 horas una vez a la semana, no se traducen necesariamente en mortalidad. No obstante, los efectos se hacen evidentes en los parámetros zootécnicos y, por ende, en la rentabilidad del cultivo, más aún cuando se está suministrando un alimento de mayor valor nutricional.
Guayas
Innovation cesar.molina@skretting.com
Las
La elevada demanda de fuentes proteicas ha llevado a recurrir a fuentes no convencionales. La reutilización de los subproductos bovinos representa una oportunidad para crear diferentes ingredientes destinados a la formulación de dietas para el camarón blanco Litopenaeusvannamei
Por: Redacción de PAM*
Introducción
La elevada demanda de fuentes proteicas ha llevado a utilizar fuentes no convencionales, como la harina de pescado (FM, por sus siglas en inglés). Sin embargo, la harina de subproductos avícolas (PBM) ha ido sustituyendo progresivamente a la FM con excelentes resultados, provocando un aumento de su demanda y, en consecuencia, de su precio. También existen otras fuentes de residuos que podrían aprovecharse, como por ejemplo, la harina de subproductos bovinos (BBM), la cual es una fuente de proteína que contiene alrededor del 50% de proteína bruta y 10% de grasa bruta, lo que permite su uso como alternativa no solo a la FM, sino también a la PBM, en dietas para Litopenaeus vannamei.
La harina de subproductos avícolas es el principal producto de extracción de grasa animal que se vende en el mercado. Sin embargo, su demanda crece en exceso por ser el principal ingrediente proteínico de la mayoría de los alimentos fabricados para la producción animal, en particular, los destinados a las mascotas y la acuicultura. Aunque la industria avícola crece de forma constante, lo hace en una magnitud diferente a la necesaria para cubrir el mercado.
Las harinas de carne y huesos de bovino provienen de los residuos de la producción animal, no aptas para el consumo humano directo, específicamente del ganado vacuno. La BBM se procesa de forma similar a la PBM, y representa más del 10% del total de la materia prima destinada a la producción de carne para consumo humano.
Con base en las oportunidades, que representa la reutilización de los subproductos bovinos, para crear una cartera de diferentes ingredientes para la formulación de alimentos acuícolas, la presente investigación tiene como objetivo evaluar el efecto de la sustitución gradual de PBM por BBM en la supervivencia, el crecimiento y los parámetros bioquímicos de la hemolinfa del L. vannamei
Materiales y método
Camarones y unidades
Experimentales
Un lote de 10,000 camarones blancos, L. vannamei , se transportó a las instalaciones del Instituto de Investigaciones Oceanológicas de la Universidad Autónoma de Baja California (IIO – UABC); los juveniles de L. vannamei fueron proporcionados por el criadero comercial FITMAR en Sinaloa, México, y caracterizados para representar una línea genética mexicana resistente a patógenos locales. A su llegada al IIO, se recibieron en un sistema de recirculación compuesto por dos tanques circulares de

1,000 L, una bomba, un biofiltro y un skimmer de proteínas.

Preparación de dietas y diseño experimental
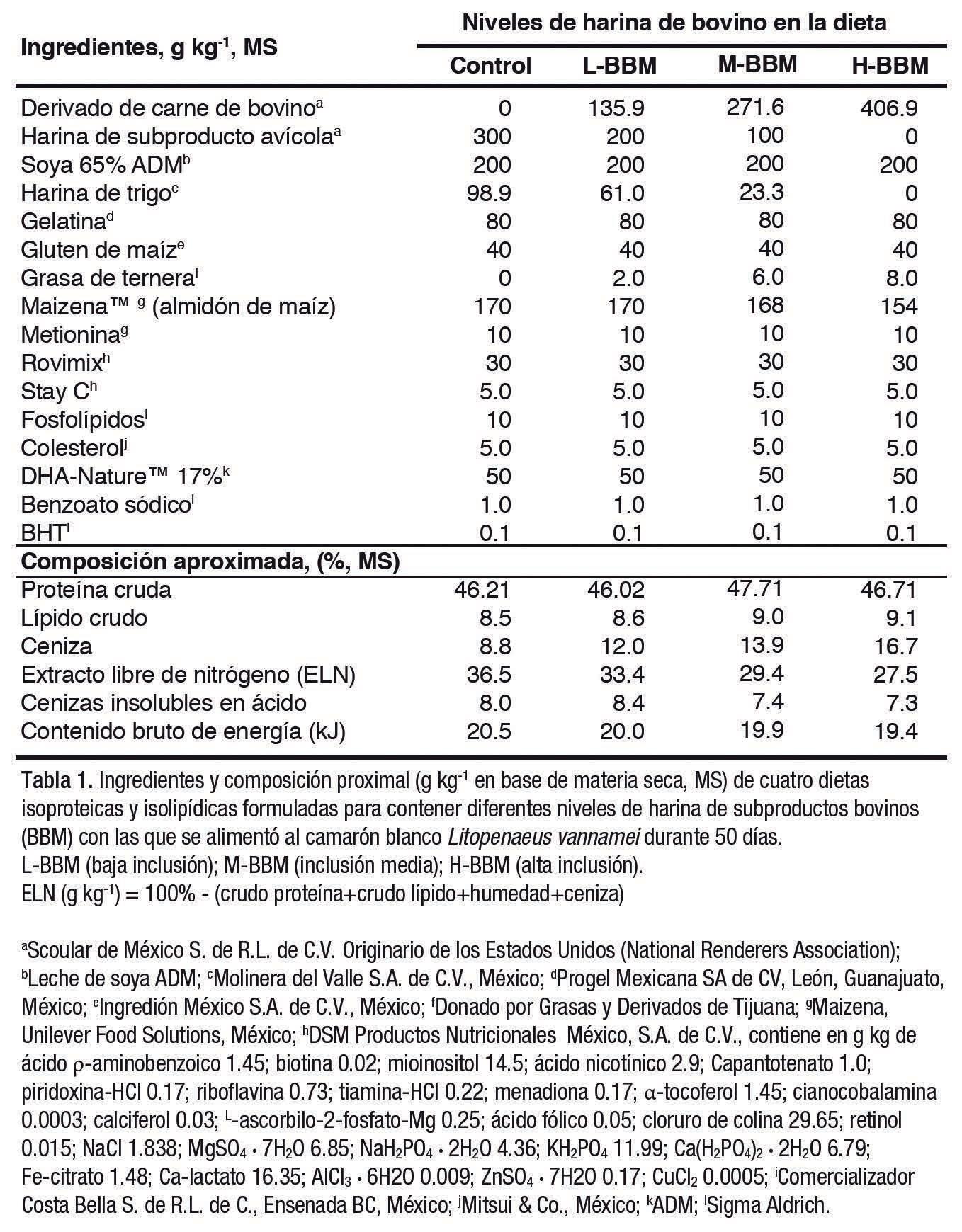
Se formularon cuatro dietas, una de Control [BBM (0%)] y tres con diferentes niveles de inclusión de BBM: L-BBM [baja inclusión (13.6%)], M-BBM [inclusión media (27.2%)] y H-BBM [alta inclusión (40.7%)]. Todas contenían un 46.5% de proteína cruda (PC) y un 8.5% de grasa cruda (GC), empleando BBM en sustitución de PBM en una formulación completamente libre de pescado, usando aceite de microalgas y colesterol para alcanzar los niveles óptimos (Tabla 1). El aceite de microalgas contenía un 17% de ácido docosahexaenoico (DHA, por sus siglas en inglés), y se añadió a partes iguales a cada dieta.
Análisis químico
Se analizó la composición aproximada de todas las dietas experimentales en muestras triplicadas
(Tabla 1). En resumen, el peso seco de las dietas se determinó secando las muestras a 60ºC durante 24 h. Las muestras se incineraron en un horno a 550ºC durante 6 h para determinar las cenizas. A continuación, se analizó la proteína bruta por el método micro-Kjeldahl, y se calculó el contenido de proteína por conversión de nitrógeno, y el contenido en lípidos a través del método Soxhlet, con éter de petróleo como disolvente portador.
La digestibilidad aparente se determinó utilizando la ceniza insoluble en ácido como marcador interno. Se determinó el contenido de cenizas insolubles en ácido de las dietas experimentales y de las heces recogidas durante el bioensayo.
Se analizaron los ácidos grasos de las dietas experimentales y de los tejidos musculares y del hepatopáncreas, según la metodología adaptada de Folch et al. (1957), empleando diclorometano en lugar de cloroformo.
Resultados
La composición aproximada de las dietas experimentales se muestra en la Tabla 1. Los contenidos de proteína bruta y grasa fueron de 46.0-47.7% y 8.5-9.1%, respectivamente. El contenido en cenizas aumentó a medida que se incluía la BBM, desde 8.8 en el grupo Control hasta 16.7 para la H-BBM; mientras que, el contenido de ácidos grasos de las dietas (Tabla 2) mostraron diferencias aparentes en ciertos ácidos grasos, disminuyendo el ácido linoleico a medida que se incorporó la BBM.
En cuanto al rendimiento global, se observaron diferencias significativas en la supervivencia; donde la mayor supervivencia se presentó con la dieta Control (100%), la cual disminuyó uniformemente hasta 88.9 ± 4.6% para el H-BBM (p < 0.05). Se registraron diferencias significativas en todos los parámetros respecto a los tratamientos. Tras siete semanas de procedimiento experimental, el rendimiento global arrojó diferencias. El tratamiento H-BBM presentó la mayor tasa específica de crecimiento (SGR, por sus siglas en inglés) significativo (3.43 ± 0.09%/día) y coeficiente térmico de crecimiento (TGC) (0.44 ± 0.016). Sin embargo, el nivel más alto de BBM resultó en el factor de conversión alimenticia (FCR) más bajo (1.51 ± 0.12) (p < 0.05).

El aumento relativo de peso (%) fue significativamente mayor para el tratamiento H-BBM (457.05 ± 27.0) en comparación con el Control (258.6 ± 6.6). Además de la prueba ANOVA, se realizó una prueba polinómica para observar la correlación entre el rendimiento global y los tratamientos dietéticos, mostrando una baja correlación entre el índice hepatosomático (HSI, por sus siglas en inglés) y los tratamientos dietéticos (R2 = 0.245), una correlación media entre los tratamientos y el peso final (R2 = 0.478), el SGR (R2 = 0.419) y el FCR (R2 = 0.455) (Figura 1).
Discusión
Esta investigación, ha demostrado que la BBM puede sustituir a la PBM en las dietas para camarones, mejorando el rendimiento global. Los resultados obtenidos son significa- tivos, considerando la existencia de otras materias primas que permiten reducir los costos, sin afectar la calidad de la producción de camarón, mediante el uso de BBM en su alimentación. Además, la BBM, al igual que la PBM, proviene de procesos de rendering de la producción de animales terrestres, lo que representa una contribución al objetivo de sostenibilidad de los alimentos acuícolas y una economía circular, reduciendo potencialmente la huella de carbono de los alimentos.

Los resultados sobre los porcentajes de acumulación de ácidos grasos en el hepatopáncreas de los organismos, sugieren que el L. vannamei puede asimilar eficientemente ácidos grasos mono y poliinsaturados en porcentajes superiores respecto al PBM de la dieta Control.
Conclusiones
Se observó que el rendimiento mejoró significativamente, con niveles crecientes de inclusión de BBM. Sin embargo, si la tasa de crecimiento no alcanzó el crecimiento deseable esperado, se debe tener en cuenta que los camarones se cultivaron a 27.8ºC. Además, los camarones del presente estudio provienen de un proyecto de línea genética mexicana, elegida para hacer frente a los patógenos en las costas mexicanas. No obstante, el nivel más alto de inclusión de BBM se comporta mejor que los del grupo Control.
Un alto contenido en cenizas podría afectar negativamente a la estabilidad hídrica de la dieta. Considerando que el grupo de tratamiento H-BBM presentó un mayor índice de conversión alimenticia, no se puede especular sobre la estabilidad de la dieta.
Por lo tanto, se infiere que la respuesta favorable de los alimentos con altos porcentajes de inclusión de BBM en los camarones se debe a la alta calidad de los nutrientes. Araújo et al. (2019) concluyeron que la combinación de grasa de vacuno con aceite de microalgas daba lugar a un mejor rendimiento que el uso de aceite de pescado en las dietas de camarones, y el aceite de microalgas resultaba una combinación más económica.
El camarón no tiene la capacidad de sintetizar ácidos poliinsaturados de cadena larga (LC-PUFA, por sus siglas en inglés). Chen et al. (2014) señalan que se deben incluir ácidos eicosapentaenoico (EPA) y docosahexaenoico (DHA) cuando los camarones se cultivan a salinidades superiores a 20 ppm. Las dietas utilizadas en el presente estudio fueron formuladas con DHA, y se observó que los porcentajes de ácidos grasos en el hepatopáncreas son estadísticamente diferentes (p < 0.05) en ácido araquidónico (ARA), EPA y DHA entre tratamientos. Adicionalmente, la acumulación de LC-PUFA aumentó, correlacionándose de manera directa con la inclusión de DHA, indicando que la acumulación selectiva juega un papel esencial en la acumulación de PUFA.
En cuanto al resto del contenido de ácidos grasos, la reducción significativa de los ácidos grasos n6, en particular del ácido linoleico, refleja su menor contenido en la BBM, 17.1 mg/dL, y todavía no hay ni una sola prueba de que valores de 40.8 mg/ dL en camarones alimentados con H-BBM, como los registrados en este estudio, puedan provocar problemas fisiológicos.
En conclusión, los resultados de esta investigación proporcionan una clara evidencia de que el camarón blanco, L. vannamei , puede ser alimentado con dietas basadas en BBM en sustitución de PBM, sin afectar el rendimiento del crecimiento y la salud, en dietas libres de FM. La inclusión de BBM mejoró los porcentajes de digestibilidad aparente de los organismos, superando los rendimientos productivos de la PBM. Los resultados obtenidos sobre los porcentajes de acumulación de ácidos grasos en el hepatopáncreas de los organismos sugieren que L. vannamei puede asimilar eficientemente ácidos grasos mono y poliinsaturados en porcentajes superiores respecto al PBM de la dieta Control. Sin embargo, la química de la hemolinfa muestra que la dieta H-BBM dio lugar a una mayor acumulación de colesterol, un nutriente esencial del camarón.
Las referencias y fuentes consultadas por el autor en la elaboración de este artículo están disponibles bajo petición previa a nuestra redacción.
Esta es una versión resumida desarrollada por el equipo editorial de Panorama Acuícola Magazine del artículo “EFFECT OF SUBSTITUTION OF POULTRY BY-PRODUCT MEAL BY BOVINE
BY-PRODUCT
MEAL ON OVERALL PERFORMANCE FOR LITOPENAEUS VANNAMEI” escritoporLÓPEZ-ORTIZ,E.-Universidad AutónomadeBajaCalifornia;ARAMBULMUÑOZ,E.-CONACYT;TINAJERO,A.,BASÍLIODEL RIO-ZARAGOZA,O.,GALINDO-VALDEZ,L.yVIANA, M.-InstitutodeInvestigacionesOceanológicas, UniversidadAutónomadeBajaCalifornia. La versión original, incluyendo tablas y figuras, fue publicada en MAYO de 2023 en ANIMAL FEED SCIENCE AND TECHNOLOGY. Se puede acceder a la versión completa a través de https://doi.org/10.1016/j.anifeedsci.2023.11568.









