
10 minute read
Las bases de la sensibilidad al contraste


Apuntes de psicofísica de la visión
Lic. en Opt. Arturo Guilliem Dávila Centro Interdisciplinario de Ciencias de la Salud Unidad Santo Tomás I.P.N.
Advertisement
opti_balbuena@yahoo.com
En el año de 1959 Hubel y Wiesel realizaron experimentos en gatos para detectar la respuesta del córtex a un estímulo determinado colocándoles microelectrodos, estos estímulos eran de tipo puntual y la respuesta que se obtenía era mínima o nula.
forma lineal provocó la respuesta inmediata de una neurona cortical. En la siguiente imagen se observa esta sombra lineal tomada directamente del video Hubel and Wiesel “cat
experiment”.

Pantalla con el estímulo puntual
Dispositivo donde tenían al gato Sombra proyectada por el borde de una diapositiva que

Fue hasta que retiraron una de las diapositivas provocó el disparo de actividad neuronal. Hubel y Wiesel

cuando la sombra que se proyectaba de esta de en su laboratorio de fisiología.
Después de muchos experimentos más, se pudo determinar que la respuesta a un estímulo lineal era óptimo cuando este era vertical y disminuía según se iba alejando de esta hasta ser nulo en posición horizontal y a 45◦.
Abajo: representación de los estudios realizados en gatos. Inferior derecha del trabajo original. reproducción de los resultados


Acercamiento del dispositivo donde eran colocados los gatos para el experimento.

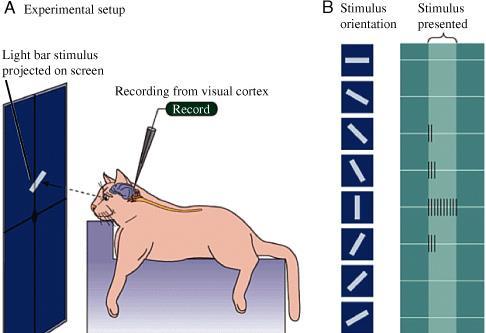
Anotaciones de los resultados obtenidos al aplicar los estímulos en diferentes posiciones. Nótese la respuesta nula el varias de estas. La respuesta de las células corticales simples variaba de acuerdo con su inclinación siendo que a los 20◦ esta disminuía notablemente.
El potencial S: Es una onda eléctrica graduada que se presenta en la retina como respuesta a un estímulo luminoso. El voltaje o amplitud máxima de la respuesta es proporcional a la intensidad del estímulo. Y estos potenciales solo están presentes mientras este aplicándose el estímulo.
Existe dos tipos de potencial S:
Acromático, cuando su potencial
negativo no depende de la longitud de onda del estímulo.
Cromática, esta respuesta es mucho más compleja debido a que puede ser negativo para cierta longitud de onda y positivo para otra. A esta respuesta se le conoce como proceso opuesto y se da cuando las células visuales codifican información de longitudes de onda utilizando dos respuestas antagónicas para dos conjuntos de longitudes de onda.
Actualmente se conocen dos pares de longitudes de onda antagónicas: azul-amarillo y rojo-verde, por ejemplo, en el primer caso existe una onda negativa para el amarillo y una positiva para el azul.
Estudios realizados por Brindley en 1970 con potenciales S sugiere que las respuestas acromáticas provienen de las células horizontales y las cromáticas de las células bipolares.
Los campos receptivos:
Se denomina campo receptivo de una neurona sensitiva a la región del espacio en la cual la presencia de un estímulo altera la respuesta de dicha neurona, esto es, la tasa de impulsos electroquímicos que ésta genera. También se entiende como una función bidimensional que detalla el cambio en la tasa de impulsos que se produce para los diferentes puntos de la región.
La retina es considerada como una especie de mosaico o áreas de proyección para cada nivel del sistema nervioso visual. Cuando se presenta un estímulo la respuesta no es uniforme en estas células,
sino que sucede en zonas específicas solamente.
Así diferentes tipos de células tienen diferentes campos receptivos como las células ganglionares tienen campos receptivos ovales como se observa en la siguiente figura.
En dichos campos la respuesta varía por ejemplo si se aplica un estímulo luminoso a estas células existe respuesta en la parte media (on) y se inhibe la zona periférica (off).
Mientras que las células del cuerpo geniculado lateral (CGL) y las corticales presentan campos receptivos muy diferentes, con zonas off centrales y de formas cuadradas, rectangulares, circulares, muy estrechas, etc.

Campos receptivos en A y B del Cuerpo Geniculado Lateral las restantes son corticales. (Hubel y Wiesel 1962)
Hubel y Wiesel diferenciaron tres tipos de células en el córtex que presentaban campos receptivos retinianos distintos. Estas eran:
Células simples: Presentaban una configuración 'lado a lado' del campo receptivo, de modo que estas células responden de forma óptima a una barra de luz que tenga una orientación determinada. Por tanto, estas células simples vienen a ser una especie de detectores de líneas, de bordes y de barras. Sus campos receptivos son circulares y estrecho en comparación con las células complejas. La respuesta de las células simples aumenta de forma lineal a medida que aumenta la cantidad de luz que llega a una zona excitadora, o disminuye a medida que aumenta la luz que incide en una zona inhibidora. Esta naturaleza lineal, es una de las características más importantes de las células simples. De tal forma estos campos receptivos alargados responden mejor a estímulos lineales o bordes de una determinada orientación y posición en la retina.
Otras tienen campos receptivos con una zona central alargada, excitatoria o inhibitoria, flanqueada a uno y otro lado por zonas antagónicas. Responden preferentemente a líneas claras u oscuras de una determinada orientación y posición en la retina: detectores de líneas o detectores de barras.
Aun no se sabe con certeza como se relacionan con las células ganglionares y las células del cuerpo geniculado lateral.
Células complejas: Estas neuronas producen su respuesta óptima ante barras que se mueven en una dirección determinada (estímulo óptimo). Comparten con las células simples el responder a estímulos lineales de una orientación específica. No responden de forma específica a la posición de una línea dentro del campo perceptivo. Cualquier posición dentro del campo perceptivo es capaz de excitarlas y son muy sensibles al movimiento de las líneas que sirven de estímulo, si una línea se mueve adecuadamente a la largo del campo perceptivo, su respuesta es máxima, y se mantiene desde que el estímulo entra en el campo visual hasta que lo deja.

Existe relación estrecha entre las células simples (arriba) y las células complejas Tienen campos receptivos de un área mayor a las células simples y son más abundantes.
Células hipercomplejas:
estas neuronas responden óptimamente ante ángulos o esquinas que se mueven en una dirección determinada, o ante barras de una longitud específica, que poseen una orientación específica y que se mueven en una dirección específica. Por lo tanto, receptivos más selectivos. tienen campos
Su característica fundamental es una disminución de su respuesta cuando la longitud de la línea que la estimula aumenta por encima de un límite, las células tanto simples como complejas aumentan la respuesta cuando aumenta la longitud de la línea.
En la actualidad se cree que las células hipercomplejas son grupos particulares tantos de células simples como de complejas
presentando una sensibilidad gradual a la presencia de una terminación.
Las vías parvo y magno celular
Los primeros estudios llevados a cabo con primates fueron realizados por Wiesel y Hubel (1966) sobre células del núcleo geniculado lateral, pero fue Gouras (1968,1969) el que estableció la existencia de una división funcional ya desde las células ganglionares de la retina.
La información recibida por los conos pasa a las células ganglionares de las cuales se conocen bien dos tipos de vías paralelas que la conforman. Aunque existe una tercera vía aun no muy bien documentanda, llamada koniocelular.
Estas clases de células dan lugar a dos canales que muestran diferencias anatómicas, fisiológicas y de comportamiento. La corteza visual también se puede subdividir en dos vías, una especializada para el procesamiento del movimiento y el otro para el color.
Cada uno de los caminos visuales conduciría diferente clase de información sobre los objetos, es decir estarían especializados en ciertas facetas de la percepción visual. [Lennie (1980), Livingstone y Hubel (1988), Shapley (1990), Merigan (1991); Kaplan (2004)].
Varias líneas de evidencia indirecta han sugerido una estrecha correspondencia entre las vías subcorticales y corticales, de tal manera que el canal M (magno) proporciona la entrada a la vía de movimiento y el canal P (parvo) impulsa la vía de color.
A través de experimentos con modelos animales, se puede comprobar que los caminos Parvo y Koniocelular (una tercera vía visual aún poco estudiada en la actualidad) conducirían la información necesaria para codificar esencialmente la forma y el color, mientras que el camino Magnocelular conduciría la información necesaria para la localización espacial de los objetos y la medida de la velocidad del movimiento.

Las células ganglionares de conección sináptica parvo celular presentan respuestas selectivas al color, describiendo a continuación tres tipos principales de unidades de respuesta en el campo receptivo:
CELULAS TIPO I: Presentan una disposición estándar del campo receptivo en forma de centro-periferia a diferentes sensibilidades espectrales, ya que reciben aferencias de distintos sistemas de conos. Los componentes “encendido” y “apagado” se encuentran activos a la sumación espacial de colores opuestos. Las
células de tipo I representan aproximadamente el 80% de las unidades parvocelulares.
CELULAS TIPO II: Carece de organización del campo receptivo en centro-periferia, se componen solo de un centro de campo, alimentado por aferencias procedentes de poblaciones de conos antagonistas con sensibilidad espectral diferente. Estas células tienen campos receptivos opuestos cromáticos, pero no espacialmente. Dan una respuesta de encendido a un color y una respuesta de apagado a otro en cualquier parte del campo receptivo.
CELULAS TIPO III: Posee organización del campo en centro-periferia, pero carece de selectividad al color, el centro y la periferia reciben aferencias indiferenciales de todos los tipos de conos.
Mientras que las células ganglionares que reciben aferencias periféricas de identificación magno celular; presentan respuestas receptivas de banda ancha “ciegas al color”.
Estas células poseen campos receptivos en forma de centro-periferia, su fisonomía se asemeja a la capa III del sistema parvo celular.
Otra variedad de células M (tipo IV) típica de las laminas magnocelulares, presentan aferencias de conos de banda ancha en el centro del campo y una periferia tónica supresora alimentada por conos de larga longitud de onda. Estas células responden con un prolongado silencio a las grandes manchas de color rojo.
Vía magnocelular: son células ganglionares grandes, responden con ráfagas breves de disparos y hacen su sinápsis en el Núcleo Geniculado Lateral (NGL) en las capas 1 y 2.

Tabla 1. Clasificación de las células según la vía visual.

Tabla 2 Aporte de diferentes autores características de las células magno y parvo.
Campbell y Robson
En el año de 1966 Campbell y Robson realizaron estudios psicofísicos por primera vez basados en las investigaciones de Hubel y Wiesel con la intensión de poder evaluar subjetivamente la
visión yendo más convencional. allá de la agudeza visual
En 1968 realizaron estudios de umbrales con redes sinusoidales comparándolas con redes cuadradas.
Bibliografía
1. Berry GA. Cited
in Regan D. Low-contrast letter charts and sinewave grating tests in ophthalmological and neurological disorders. Clin Vis Sci 1988; 2: 235-50.
2. Young G. Threshold tests. BrJ Ophthalmol 1918; 2: 384-92.
3. Campbell FW, Green DG. Optical and retinal factors affecting visual resolution.
J Physiol 1965; 181: 576-93. 4. Arden GB, Jacobson JJ. A simple grating test for contrast sensitivity: preliminary results indicate value in screening for glaucoma. Invest Ophthalmol VisSci 1978; 17: 23-32.
5. Banks, Athikinson, “Development of contrast sensitivity over the first 3 month of life in the human infant”, Vision Res., 1997, 17:1037-44
6. Elliott, Whitaker, “Clinical contrast sensitivity chart evaluation”, en: Ophthalmic and Physiological Optics, 1992, 12:275 –280
7. Gwizda, et.al., “Development of Spatial contrast sensitivity from infancy to adulthood: psychophysical data” en: Optometry and Vision Science, 1997, 74:785-789.
8. Lathman, K., “Who uses contrast sensitivity optometric practice?” en: Ophthalmic and Physiological Optics,. 1998, 18:S2-S13.


