259 minute read
Valentina Fernandez '24


Understanding e Connection Between Diabetes and Kidney Disease: Are SGLT-2 Inhibitors the “Magic Bullet”?
BY VALENTINA FERNANDEZ '24
Advertisement
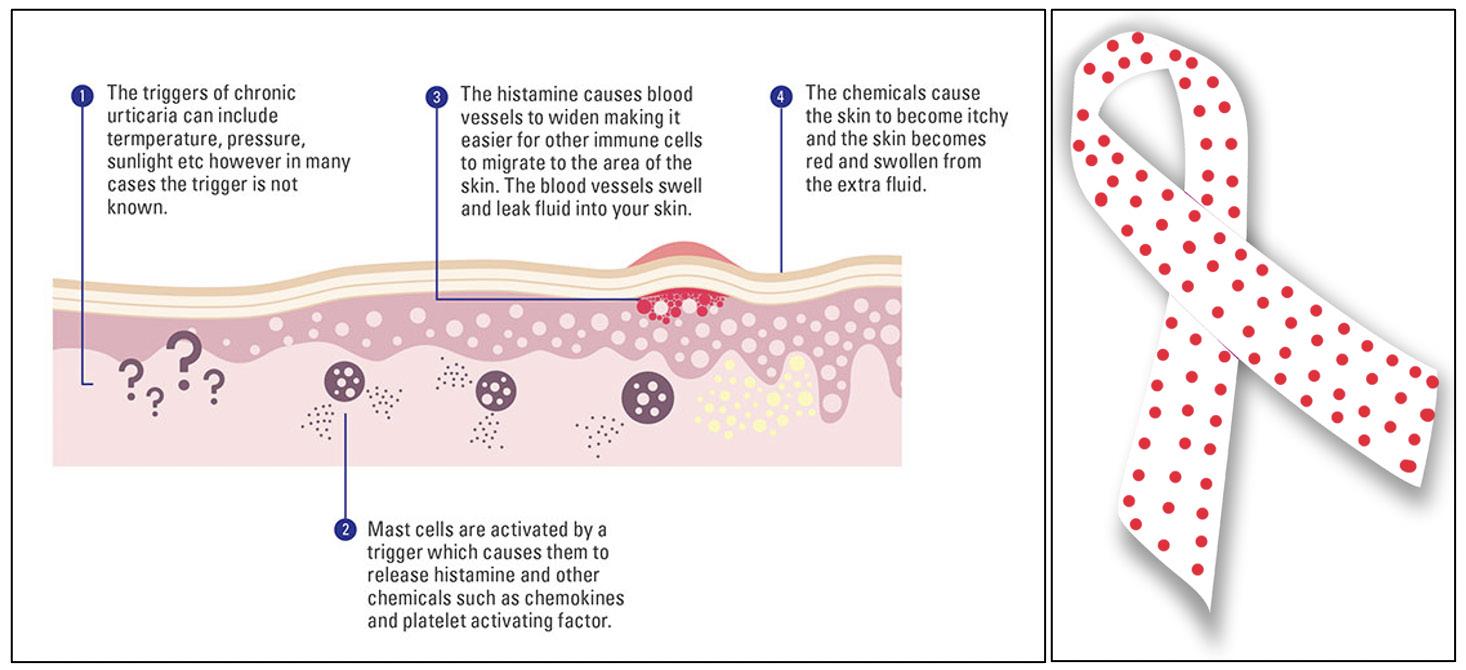
Cover Image: 1916 Schematic of a longitudinal section of a kidney. One of the hallmark symptoms in diabetes mellitus is polyuria, or excessive urination, which results from abnormally high levels of sugar in the blood, causing the kidneys to retain more water. Image Source: Wikimedia Commons Introduction
Diabetes has been called the epidemic of the century, while kidney disease has been called the under-recognized public health crisis (Kharroubi & Dariwsh, 2015; NVS 2021 report of 2018 data). Kidney disease causes more death than breast cancer or prostate cancer, and diabetes a ects over 460 million people worldwide. (Kharroubi & Dariwsh, 2015; NVS 2021 report of 2018 data). But, seldom is the connection between these two considered in-depth. Diabetes is the leading cause of kidney disease, accounting for nearly half of all causes of kidney failure resulting in a kidney transplant (Tuttle et al., 2021). Just in the United States, 34.2 million adults (~10.5% of the population) are thought to have diabetes, with 9095% of cases being type 2 diabetes. It is estimated that the economic burden of diabetes costs the United States about $327 billion USD per year, with reduced productivity accounting for $90 billion of the total, and the rest due to direct medical costs (American Diabetes Association, 2018). Diabetic Kidney Disease (DKD), which is chronic kidney disease for people with diabetes, is estimated to occur in ~30% of people with type 1 diabetes and in ~40% of people with type 2 diabetes (Tuttle et al., 2021). Even though the interconnectedness of diabetes and kidney disease has long been known, the emergence of a new class of drugs called SGLT-2 inhibitors has reignited interest in addressing DKD. SGLT-2s were originally developed for people with type 2 diabetes to control their glycemia levels. However, they were found to have both positive cardiovascular and renal e ects. e development of these drugs provides a promising future for people living with cardiometabolic and renal diseases, such as diabetes, hypertension, and chronic kidney disease.
is review article will focus on the connection between diabetes and kidney disease. is paper will begin by providing background on the molecular mechanisms at play in diabetes mellitus and nephropathy (kidney disease). Next, the paper will turn its attention to the postulated reasons why diabetes causes kidney failure. e paper will then transition to review the current interventions in place to mitigate the burden of these disease, with a speci c focus on the emergence of SGLT-2 inhibitors. To conclude, this paper will explore the limits of SGLT2 therapy right now, and what we can expect from this class of drugs looking forward.
Molecular Mechanisms of Diabetes: e Di erence Between Type 1 and Type 2 Diabetes
Diabetes Mellitus is de ned as a group of metabolic diseases characterized by defective insulin secretion, insulin action, or both, which results in chronic hyperglycemia, or an abnormally high blood sugar level (Kharroubi & Darwish, 2015). ere are various forms of diabetes mellitus, including gestational diabetes and rare-diabetes disorders, such as maturity-onset diabetes of the young (MODY) and latent autoimmune diabetes in adults (LADA). Classifying diabetes remains a controversial issue, with various scientists, physicians, and regulatory bodies disagreeing on the distinct categories for grouping diabetes. For the purposes of this paper, I will follow the American Diabetes Association’s traditional classi cation, which lists four forms of diabetes: gestational diabetes mellitus (GDM), other types (including, but not limited to, LADA and MODY), type 1 and type 2 diabetes (Kharroubi & Darwish, 2015).
Type 1 Diabetes, or insulin-dependent diabetes, is an autoimmune disorder in which the body attacks its own pancreatic beta cells, which are responsible for endogenous insulin production (Kaufman, 2006). Type 1 diabetes constitutes about 5-10% of all diabetes cases and 80-90% of all cases in children (Maahs et al., 2010). Due to its historical prevalence in children, teens, and young adults, type 1 diabetes has been previously called juvenile diabetes, even though diagnosis can occur at any age (CDC, 2021). Although a genetic basis predisposes certain populations to type 1 diabetes, the speci c causes and mechanisms of its inheritance patterns remain unknown. Several risk factors have been identi ed; for example, having certain variants of the HLA-DQA1, HLA-DQB1, and HLA-DRB1 genes may increase the likelihood of developing type 1 diabetes (Type 1 Diabetes, n.d.). ese genes belong to a family of genes known as the human leukocyte antigen (HLA) complex, which play a role in the immune system’s ability to distinguish self from non-self (Overview of the Immune System - Immune Disorders, n.d.). HLA proteins and their combination of variations (haplotypes) can increase the risk of acquiring an autoimmune response, resulting in malfunction of insulin-producing beta cells and leading to the type 1 diabetes phenotype (Rosen & Ingel nger, 2019). Speci cally, the HLA DR3-DQ2 haplotype has been linked to almost half of all patients diagnosed with type 1 diabetes. As such, therapies focused on targeting this haplotype have emerged as candidates for a type 1 diabetes cure; to give just one example, Swedish biopharmaceutical company Diamyd Medical is undergoing a phase 3 clinical trial to develop a vaccine against type 1 diabetes (Hannelius, 2021). To date, there is no cure for type 1 diabetes, and all people with this disease must administer insulin every day, multiple times a day, to maintain their blood glucose levels, as was demonstrated by the Diabetes Control and Complications Trial (DCCT) (Nathan et al., 2005; Wilson, 2011).
Type 2 diabetes is the most common form of diabetes and it is characterized by insulin resistance; pancreatic beta cells can produce insulin, but the body is unresponsive to it. In the United States alone, more than 34 million people (about 1 in 10 Americans) have type 2 diabetes, and the amount of people with type 2 diabetes is expected to continue growing (CDC, 2019). Not all patients with type 2 diabetes will become insulin dependent, and most are treated with a combination of therapies to promote weight loss and normoglycemia, which aims to bring blood sugar levels down to a normal range (these speci c therapies will be discussed in detail below). Although type 2 diabetes is largely due to environmental and lifestyle factors, it also has a signi cant genetic basis. Genome wide association studies (GWAS) have been pivotal in identifying about 70 loci associated with type 2 diabetes in various populations (Kharroubi & Darwish, 2015). Several loci positioned in and around the CDKAL1, CDKN2A/B, HHEX/ IDE and SLC30A8 genes are responsible for increasing type 2 diabetes risk (Zeggini et al., 2007). HHEX/IDE speci cally is related to the insulin-degrading enzyme, while CDKAL1 and CDKN2A/B regulate cell expression, and therefore are linked to beta cell dysfunction (Zeggini et al., 2007). On the other hand, SLC30A8 encodes for a protein involved in the intracellular accumulation of zinc, speci cally in the pancreas, meaning that it colocalizes with insulin in various insulin secretory pathways. e speci c mechanisms involved in these pathways remain elusive, and further analysis is necessary for full comprehension of the signaling present. Recently, a variant of the HNF1A gene was shown to increase the risk of developing type 2 diabetes among the Latino population, suggesting it may serve as a screening tool in the future (SIGMA Type 2 Diabetes Consortium et al., 2014). All in all, there are a variety of genes at play for increasing type 2 diabetes risk, some speci c to certain populations or subgroups (Kharroubi & Darwish, 2005). Since most (but not all) patients with type 2 diabetes have excess weight or obesity, the available therapies target weight loss, blood sugar management, and promote healthy eating
Table 1: e blood glucose levels that correspond to each condition, according to guidelines from the American Diabetes Association’s 2021 Standards of Care.
Data Source: ADA
Table Source: Created by author.
Screening for diabetes is crucial for early identi cation of the disease. And, given the increasing incidence and prevalence of diabetes, screening is now more important than ever (Lynam et al., 2019). For type 1 diabetes, one screening method involves testing for the presence of a few islet autoantibodies that have been identi ed as risk factors for the disease (American Diabetes Association, 2021). Islet cells are found in clusters throughout the pancreas; alpha cells (which produce glucagon, the hormone that raises blood sugar) and beta cells (which produce insulin) are two subtypes of islet cells (American Diabetes Association, 2021). Currently, the American Diabetes Association’s 2021 Standards of Care does not recommend this type of clinical testing for lowrisk populations, citing an insu cient amount of evidence con rming its clinical signi cance and verity (American Diabetes Association, 2021). However, they do recognize the validity of measuring the islet antibodies in individuals at risk for type 1 diabetes (e.g., relatives of those with type 1 or individuals from the general population with type 1 diabetes-associated genetic factors), pointing to a few European and Americans studies that reported a 70% likelihood of developing type 1 diabetes a er testing positive for two or more autoantibodies (Ziegler et al., 2013). For type 2 diabetes, screening measures involve informal assessment of risk factors, such as obesity or hypertension, and is recommended for all individuals beginning at 45 years of age (American Diabetes Association, 2021). Since all types of diabetes are ultimately diagnosed based on glycemic levels, there are three main exams that con rm the prognosis: (i) a fasting plasma glucose test (FPG), (ii) an oral glucose tolerance test (OGTT), and (iii) an A1C glycosylated hemoglobin test (Mayo Clinic, 2021). An FPG test measures blood sugar a er an overnight fast, and results are usually corroborated through an OGTT or an A1c test (American Diabetes Association, 2021). An OGGT, also known as the “glucose challenge,” gives the test-taker a glucose infusion (typically 75g of glucose in solution) and then tracks the test-taker’s glucose metabolism for two hours to achieve a nal blood glucose reading (American Diabetes Association, 2021). An A1c test, on the other hand, does not measure glucose levels directly and instead relies on hemoglobin. Hemoglobin is the protein in red blood cells that carries oxygen throughout the body. Glucose glycates (sticks) to hemoglobin, and so by measuring the amount of glycosylated hemoglobin in the blood, one can measure blood sugar over an extended period. An A1c is given as a percentage of the glucosebound hemoglobin molecules in the body, and it is directly proportional to average glucose levels over a 3-month period, making it a signi cant marker for diabetes diagnosis (Sun et al., 2014). In fact, A1c is such a potent indicator that most clinical trials testing therapies for diabetes will use changes in A1c levels as a marker for the e cacy of the drug. For all exams, blood glucose levels below 100mg/dL (5.6 mmol/L) are normal; those ranging from 100-125 mg/dL (5.6-6.9 mmol/L) indicate prediabetes, and any above 126 mg/dL (7 mmol/L) on two separate tests con rm the presence of diabetes (American Diabetes Association, 2021).
Defects in Insulin: e Culprit for all Types of Diabetes
All forms of diabetes are characterized by de ciencies relating to the insulin hormone. Insulin is an endocrine peptide hormone that binds to its receptors in the plasma membrane and triggers a signaling pathway to move sugar from the blood to the inside of cells, where it can be used for metabolic duties, such as glucose regulation or suppression of triglyceride production, among many others (Petersen & Shulman, 2018). In other words, higher circulating insulin levels are followed by a decrease in blood glucose, meaning that insulin is necessary to achieve and maintain normoglycemia for all populations. Insulin or insulin-like peptides (ILPs) have been identi ed in all animals, even invertebrates, indicating their evolutionary signi cance in harnessing the energy from glucose for use by the cell (Petersen & Shulman, 2018).
Insulin exerts its e ects by binding to the insulin receptor (INSR) on the plasma membrane of its target cells, triggering several signaling cascade


pathways reliant on phosphorylation (Haeusler et al., 2018). e INSR belongs to the receptor tyrosine kinase (RTK) superfamily, which are characterized by their phosphorylation activity (De Meyts, 2000). e INSR is a heterotetrametric (a dimer of heterodimers) RTK, composed of two extracellular α subunits (which bind insulin) and two membrane-spanning β subunits, each which contain a tyrosine kinase domain (Hubbard, 2013).
e tyrosine kinase domain of the INSR is inactive in the absence of a ligand, but once insulin binds to two distinct sites on each α subunit of its receptor, this RTK autophosphorylates to become an activated dimer (De Meyts, 2000).
Unlike most RTKs, the ISNR does not bind signaling proteins directly upon activation but instead binds docking proteins called IRS (Insulin Receptor Substrate) 1-6, which then recruit various other proteins to trigger intracellular signaling cascades (De Meyts, 2000). Phosphorylation of the receptor causes a conformational change that triggers a signal transduction cascade inside the target cell, beginning with the recruitment of insulin receptor substrates (IRS) (De Meyts, 2000). e IRS proteins were identi ed and sequenced in the 1980s, following the development of molecular cloning technologies (Guo, 2014). One of the main pathways involving the IRS proteins is the P13K/AKT pathway, which mediates the metabolic e ects of insulin/ When defective, it is linked to the diabetes phenotype (De Meyts, 2000; Guo, 2014). Activation of this pathway is triggered by the binding of p85 or p55 (two of the regulatory subunits of PK13) to the IRS-1 and IRS-2 proteins (which are bound to the activated ISNR dimer) (De Meyts, 2000). Following this initial binding, PK13 is activated through phosphorylation, allowing it to ultimately generate PIP3, a secondary messenger which then activates PDK1 and PDK 2 (3-phosphoinositidedependent-protein kinase), two kinases that mediate the e ect of insulin on metabolism (Guo, 2014). PDK1 then activates AKT, which will have four critical substrates that serve as downstream targets: (i) mTOR (important for protein synthesis), (ii) GSK 3 (glycogen synthase kinase 3, important for the regulation fo glycogen synthesis), (iii) FoxO transcription factors (important for the regulation of gluconeogenic and adipogenic genes), and (iv) AS160 (involved in glucose transport) (De Meyts, 2000). Glucose transport from the bloodstream into its target cell (whether it be a muscle, skeletal, or fat cell) is precisely what people with diabetes have di culty with. erefore, de ciencies in the part of the pathway involving AS160 and its downstream targets results in postprandial glycemic excursions and dysregulation of glycemia. In normal (non-diabetes) circumstances, AKT will activate AS160. Activation of AS160 is required for GLUT4 translocation, where GLUT4 vesicles localize glucose channels to the plasma membrane, enabling the cell to remove glucose from the bloodstream (Guo, 2014). In its basal state, the GAP activity of AS160 maintains its target Rab in an inactive, GDP-bound form, which retains GLUT4 in intracellular compartments ( ong et al., 2007). A er insulin stimulation, AKT phosphorylates AS160, deactivating its GAP activity, which then allows the target Rab to shi to its active GTP-bound form, thereby relieving an inhibitory e ect on GLUT4 tra c and thus
Image 1: Schematic representation of the insulin receptor depicting the extracellular and intracellular subunits of the molecule
Image Source: Hubbard, 2013
Image 2: Schematic depicting the process of blood ltration and urine formation via the kidney’s nephrons. Speci cally, the kidneys rely on ion concentration gradients to properly lter the blood’s waste, mostly nitrogenous compounds. Image Source: Wikimedia Commons allowing the GLUT4 vesicles to translocate to the plasma membrane ( ong et al., 2007).
e Kidneys
e kidneys, along with the ureters, urinary bladder, and urethra, are part of the body’s excretory system and are responsible for disposing metabolic wastes and regulating the osmotic balance of blood. Each kidney, about 10 cm in length, has an outer renal cortex and an inner renal medulla, which contain tightly packed excretory tubules and their associated blood vessels. As blood enters the kidneys, the excretory tubules form and process the ltrate, composed of salts, sugars, amino acids, and nitrogenous waste. e kidney’s functional units, the nephrons, weave back and forth across the renal cortex and medulla and play a pivotal role in producing urine hyperosmotic to body uids. e nephrons are composed of a single long tubule, a ball of capillaries (referred to as the glomerulus), a cup-shaped swelling in the tubule surrounding the glomerulus (called Bowman’s capsule), and the macula densa, which are a set of salt sensors that generate paracrine chemical signals, such as changes to renal blood ow, glomerular ltration, and renin release to control kidney function (Urry et al., 2017). e kidney has roughly 1 million nephrons, and each is supplied with blood by an a erent arteriole. As blood passes down the nephron, a series of absorption and reabsorption events lter the blood and expel its waste through urine. One of the earliest reported studies linking diabetes with kidney disease was in 1981, when RA DeFronzo and colleagues observed insulin resistance in patients with uremia, a condition indicative of defective ltering of urine. ey noted that the basal insulin concentration of the uremic subjects (125.01 ±1 pmol/L) was higher than insulin concentration in the control subjects, (97.23 ±1pmol/L; p<0.01). Additionally, they reported that the average rate of glucose utilization from minute 20 to 120 of the study period in the 17 uremic subjects (3.71+0.20 mg/kg-min) was 50% lower than in the 36 controls (7.38+0.26 mg/kg min, P < 0.001). is study concluded that while insulin secretion was not impaired in most patients with renal failure, patients exhibited hyperglycemia due to decreased tissue sensitivity to insulin, thus con rming the link between kidney disease and insulin resistance (DeFronzo et al., 1981). What remained elusive a er this study, though, was how exactly hyperglycemia damaged the kidney’s function.
e processes of absorption (from bloodstream to the nephron’s tubule) and reabsorption (back to bloodstream) in the kidneys are determined by ion concentrations; for people with diabetes, the excessive glucose ions in the blood (hyperglycemia) o set these balances and damage the blood vessels’ linings, thus preventing the kidney’s nephrons from properly ltering waste. e tubular hypothesis of nephron ltration and diabetic kidney disease postulates that elevated glucose levels in the glomerular ltrate drive an

increased reabsorption of glucose and sodium by the sodium-glucose cotransporters, SGLT2 and SGLT1, in the proximal tubule, thus resulting in elevated blood glucose levels (Vallon & omson, 2020). SGLT2 and SGLT1 proteins are expressed in the apical membrane of the early and late proximal tubule, respectively. As symporters, these membrane channels contribute to the renal absorption and reabsorption of glucose (Pittampalli et al., 2018). ey mediate active transport of glucose against its concentration gradient by means of cotransport with sodium (sodium’s concentration gradient is created by the Na-K-ATPase pump, which forces intracellular Na to exit the cell across its basolateral membrane) (Vallon & omson, 2020). Speci cally, SGLT2 uses one sodium ion to transport one glucose molecule and is responsible for 90% of the glucose reuptake in the rst segment of the proximal tubule, while SGLT1 uses two sodium ions to transport one glucose molecule and is responsible for the remaining 10% (Pittampalli et al., 2018).
Because people with diabetes have elevated glucose levels in their blood, they experience what is called a “hyper-reabsorption” of glucose and sodium in the proximal tubule (Vallon & omson, 2020). e increase in absorption of glucose and sodium enhances the glomerular ltration rate (GFR), since the glomerulus and its protein channels (SGLTs) are overworking to move the higher-than-normal levels of glucose into the bloodstream; this results in a state of “hyper ltration” for the kidney. e increase in tubular glucose reabsorption capacity helps to conserve the ltration function of the kidney, but it also maladaptive because it sustains hyperglycemia (Hotstetter, 1992). Eventually, the kidneys respond to the chronic increase in tubular reabsorption by overexpressing SGLT2s, an adaption which further promotes chronic hyperglycemia. In mouse models of T1DM and T2DM, SGLT2 expression has been found to increase by 40-80% during the early stages of hyperglycemia (Kajiwara & Sawa, 2021; Vallon et al., 2013, 2014). ese ndings supported the tubular hypothesis of nephron ltration and diabetic kidney disease, suggesting that diabetes increases renal SGLT2 expression and glucose reabsorption (from nephron to bloodstream).
Over time, the kidney’s hyper ltration behavior becomes dangerous; even though overexpression of SGLT2 initially helped the kidney to adapt to increased sugar levels in the blood, it inadvertently promoted hyperglycemia in doing so. In short, a seemingly bene cial adaption becomes maladaptive. Chronic hyperglycemia promoted by overexpression of SGLT2 has a variety of adverse e ects for the body. For the kidney speci cally, studies have shown that the prevalence of hyper ltration is proportional not only to blood glucose levels, but also to blood pressure, suggesting that hyper ltration (resulting in hyperglycemia) may result in or exacerbate hypertension. Over time, hypertension constricts the a erent arterioles in the kidneys, decreasing ltration by the glomerulus; what begins as hyper ltration eventually results in hypo ltration, characterized by a decreased eGFR indicative of a failing kidney (Palatini, 2012).
e adverse e ects associated with kidney failure, which are exacerbated by diabetes, have made diabetic kidney disease a priority to address. Currently, the US Food and Drug Administration (FDA) has approved four SGLT-2 inhibitors for the treatment of hyperglycemia in type 2 diabetes: canagli ozin, dapagli ozin, empagli ozin, and ertugli ozin (Pittampalli et al., 2018; Tuttle et al., 2021). ese agents are all part of the same class (Sodium Glucose Transporter 2 Inhibitors), so their molecular structure and mechanisms of action are very similar (Hsia et al., 2017). eir main di erences lie in which biopharmaceutical company produce the drug, which results in slight di erences on how sensitive or selective the agents are (Hsia et al., 2017). In addition, the drugs di er in in the doses that are available in the market, and slightly in their administration patterns. For example, canagli ozin is only available in 100mg or 300mg doses, while dapagli ozin is a much smaller dose, only available as 5mg or 10 mg (Hsia et al., 2017).
SGLT-2 inhibitors have demonstrated tremendous success in their ability to combat hyperglycemia, these class of drugs have been found to confer cardiorenal protection as well, a er a variety of cardiovascular disease (CVD) outcome trials (CVOT) were carried out in people with type 2 diabetes and established ASCVD (Merck Sharp & Dohme Corp., 2021; Neal et al., 2017; Wiviott et al., 2019; Zinman et al., 2015). For instance, in the Johnson&Johnson sponsored CANVA trial, canagli ozin demonstrated an 18% reduction in urinary albumin-to-creatinine ratio (ACR) and a 27% risk reduction for progression of albuminuria (95% CI), two important markers of kidney disease (Neal et al., 2017). Previously, canagli ozin had demonstrated its safety and e cacy in controlling hyperglycemia for people with type 2 diabetes; speci cally, the 300mg dose of canagli ozin conferred a 1.06% reduction in
Table 2: e risk-bene t pro le of the SGLT-2 inhibitor class, based on results from the DAPA-CKD, CREDENCE, and EMPA-REG trials testing dapagli ozin, canagli ozin, and empagli ozin, respectively. Data Source: Pittampalli et al., 2018
Table Source: Created by author.
A1c at 26 weeks, compared to placebo, and these reductions were sustained at week 52. In a metaanalysis and systematic review done by Zelniker et al (2019), all the results from the CVOTs until 2018 were combined, and the cardiorenal bene ts of SGLT2 for people with diabetes were once again validated. Speci cally, this meta-analysis pooled data from 34, 322 patients and reported that SGLT2 inhibitors reduced major adverse cardiovascular events (MACE) by 11% and reduced the risk of progression of renal disease by 45% (p<0.0001) (Zelniker et al., 2019). Notably, the extent to which SGLT2s conferred renal protection varied with baseline renal functions, with lesser reductions in progression of renal disease in patients that had more severe kidney disease at baseline. is suggested that the drugs are e cacious, early intervention with SGLT2s will maximize their ability to confer protective bene ts.
e future for SGLT2 inhibitors will likely depend on the risk-bene t ratio of the drug class. In other words, for SGLT2 inhibitors to solidify their dominance of the market as the most e cient therapy for type 2 diabetes and kidney disease, the bene ts they confer to the patient will have to strongly outweigh any risks associated with their use. Fortunately, because sodium-glucose co-transporters 1 and 2 function independent of insulin action, they have demonstrated little to no adverse events like hypoglycemia or weight gain. For example, in the DAPA-CKD trial testing dapagli ozin in patients with chronic kidney disease, the incidence of adverse events were similar in both the experimental and control group (dapagli ozin vs placebo) and low overall (Heerspink et al., 2020). In fact, the trial was stopped early due to “overwhelming e cacy” and topline results granted the drug Fast Track designation by the FDA in August 2019 (FDA Grants Fast Track Designation for Farxiga in Chronic Kidney Disease, 2019). Likewise, in the CREDENCE trial, canagli ozin demonstrated similar rates of adverse events compared to the placebo group, with a rate of 12.3 versus 11.2 per 1,000 patient-years for risk of lower-limb amputation (95% CI, 079 to 1.56). Despite these initial positive results, all SGLT-2 inhibitors carry a red ag for people with diabetes: a risk of diabetic ketoacidosis (DKA) (Zelniker et al., 2019). DKA is a serious complication of diabetes caused by an excessive production of blood acids (ketones) due to a lack of insulin circulating in the body. Although the rates of DKA in these trials have been overall very low, they are still present, and certainly a cause for concern. e risk to bene t ratio of SGLT-2 inhibitors are summarized in Table 2.
e era of SGLT-2 inhibitors looks promising; a er all, these classes of drugs are rather novel in this space, and in just a decade or so have managed to establish themselves as rst-line therapies for hyperglycemia, and now CVD and renal disease. As we’ve learned, maintaining glucose homeostasis is vital to preserving a reliable and consistent source of glucose to all the body’s organs. Diabetes is ubiquitous, not only in the world as a global health problem, but also in the body— its comorbidities travel in conjunction with the disease. SGLT-2 inhibitors provide an avenue for better care, and along with lifestyle changes and other self-care measures (i.e. physical activity and nutritional care) will continue to facilitate normoglycemia and minimize adverse cardiorenal outcomes. Whether SGLT-2 inhibitors are the “magic bullet” for diabetes, CVD, and renal disease or not remains unknown, but hopefully over time, we will have an accumulation of more real-world evidence validating its e cacy, and allowing for wider adoption of these agents.
References
American Diabetes Association. (2018). Economic Costs of Diabetes in the U.S. in 2017. Diabetes Care, 41(5), 917–928. https://doi. org/10.2337/dci18-0007
Association, A. D. (2021). 2. Classi cation and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2021. Diabetes Care, 44(Supplement

CDC. (2019, May 30). Type 2 Diabetes. Centers for Disease Control and Prevention. https://www. cdc.gov/diabetes/basics/type2.html
FDA grants Fast Track designation for Farxiga in chronic kidney disease. (2019, August 27). https:// www.astrazeneca.com/media-centre/pressreleases/2019/fda-grants-fast-track-designationfor-farxiga-in-chronic-kidney-disease-27082019. html
Guo, S. (2014). Insulin signaling, resistance, and metabolic syndrome: Insights from mouse models into disease mechanisms. Journal of Endocrinology, 220(2), T1–T23. https://doi. org/10.1530/JOE-13-0327
Haeusler, R. A., McGraw, T. E., & Accili, D. (2018). Biochemical and cellular properties of insulin receptor signalling. Nature Reviews Molecular Cell Biology, 19(1), 31–44. https://doi. org/10.1038/nrm.2017.89
Hannelius, U. (2021, September 27). Diamyd Medical receives rst regulatory approval to start the Phase III trial DIAGNODE-3 with the diabetes vaccine Diamyd®. https://www.diamyd. com/docs/pressClips.aspx?ClipID=4070572
Heerspink, H. J. L., Stefánsson, B. V., CorreaRotter, R., Chertow, G. M., Greene, T., Hou, F.-F., Mann, J. F. E., McMurray, J. J. V., Lindberg, M., Rossing, P., Sjöström, C. D., Toto, R. D., Langkilde, A.-M., & Wheeler, D. C. (2020). Dapagli ozin in Patients with Chronic Kidney Disease. New England Journal of Medicine, 383(15), 1436–1446. https://doi.org/10.1056/NEJMoa2024816
Hsia, D. S., Grove, O., & Cefalu, W. T. (2017). An Update on SGLT2 Inhibitors for the Treatment of Diabetes Mellitus. Current Opinion in Endocrinology, Diabetes, and Obesity, 24(1), 73–79. https://doi.org/10.1097/ MED.0000000000000311
Hubbard, S. R. (2013). e Insulin Receptor: Both a Prototypical and Atypical Receptor Tyrosine Kinase. Cold Spring Harbor Perspectives in Biology, 5(3), a008946–a008946. https://doi. org/10.1101/cshperspect.a008946
Kajiwara, K., & Sawa, Y. (2021). Overexpression of SGLT2 in the kidney of a P. gingivalis LPSinduced diabetic nephropathy mouse model. BMC Nephrology, 22(1), 287. https://doi. org/10.1186/s12882-021-02506-8 Kaufman, F. R. (2006). Diabesity: A doctor and her patients on the front lines of the obesitydiabetes epidemic. Bantam Books.
Kharroubi, A. T., & Darwish, H. M. (2015). Diabetes mellitus: e epidemic of the century. World Journal of Diabetes, 6(6), 850–867. https:// doi.org/10.4239/wjd.v6.i6.850
Lynam, A., McDonald, T., Hill, A., Dennis, J., Oram, R., Pearson, E., Weedon, M., Hattersley, A., Owen, K., Shields, B., & Jones, A. (2019). Development and validation of multivariable clinical diagnostic models to identify type 1 diabetes requiring rapid insulin therapy in adults aged 18–50 years. BMJ Open, 9(9), e031586. https://doi.org/10.1136/bmjopen-2019-031586
Maahs, D. M., West, N. A., Lawrence, J. M., & Mayer-Davis, E. J. (2010). Epidemiology of type 1 diabetes. Endocrinology and Metabolism Clinics of North America, 39(3), 481–497. https://doi. org/10.1016/j.ecl.2010.05.011
Mayo Clinic. (2021, January 20). Type 1 diabetes—Diagnosis and treatment—Mayo Clinic. https://www.mayoclinic.org/diseasesconditions/type-1-diabetes/diagnosis-treatment/ drc-20353017
Merck Sharp & Dohme Corp. (2021). Randomized, Double-Blind, Placebo-Controlled, ParallelGroup Study to Assess Cardiovascular Outcomes Following Treatment With Ertugli ozin (MK8835/PF-04971729) in Subjects With Type 2 Diabetes Mellitus and Established Vascular Disease, e VERTIS CV Study (Clinical Trial Registration study/NCT01986881). clinicaltrials. gov. https://clinicaltrials.gov/ct2/show/study/ NCT01986881
Neal, B., Perkovic, V., Maha ey, K. W., de Zeeuw, D., Fulcher, G., Erondu, N., Shaw, W., Law, G., Desai, M., & Matthews, D. R. (2017). Canagli ozin and Cardiovascular and Renal Events in Type 2 Diabetes. New England Journal of Medicine, 377(7), 644–657. https://doi. org/10.1056/NEJMoa1611925
Overview of the Immune System—Immune Disorders. (n.d.). Merck Manuals Consumer Version. Retrieved October 30, 2021, from https://www.merckmanuals.com/home/ immune-disorders/biology-of-the-immunesystem/overview-of-the-immune-system
Pittampalli, S., Upadyayula, S., Mekala, H. M., &
Lippmann, S. (2018). Risks vs Bene ts for SGLT2 Inhibitor Medications. Federal Practitioner, 35(7), 45–48.
Rosen, C. J., & Ingel nger, J. R. (2019). Traveling down the Long Road to Type 1 Diabetes Mellitus Prevention. New England Journal of Medicine, 381(7), 666–667. https://doi.org/10.1056/ NEJMe1907458
SIGMA Type 2 Diabetes Consortium, Estrada, K., Aukrust, I., Bjørkhaug, L., Burtt, N. P., Mercader, J. M., García-Ortiz, H., HuertaChagoya, A., Moreno-Macías, H., Walford, G., Flannick, J., Williams, A. L., Gómez-Vázquez, M. J., Fernandez-Lopez, J. C., Martínez-Hernández, A., Jiménez-Morales, S., Centeno-Cruz, F., Mendoza-Caamal, E., Revilla-Monsalve, C., … MacArthur, D. G. (2014). Association of a lowfrequency variant in HNF1A with type 2 diabetes in a Latino population. JAMA, 311(22), 2305–2314. https://doi.org/10.1001/jama.2014.6511
Sun, X., Du, T., Huo, R., & Xu, L. (2014). Hemoglobin A1c as a marker for identifying diabetes and cardiovascular risk factors: e China Health and Nutrition Survey 2009. Acta Diabetologica, 51(3), 353–360. https://doi. org/10.1007/s00592-013-0515-5
ong, F. S. L., Bilan, P. J., & Klip, A. (2007). e Rab GTPase-Activating Protein AS160 Integrates Akt, Protein Kinase C, and AMP-Activated Protein Kinase Signals Regulating GLUT4 Tra c. Diabetes, 56(2), 414–423. https://doi. org/10.2337/db06-0900
Tuttle, K. R., Brosius, F. C., Cavender, M. A., Fioretto, P., Fowler, K. J., Heerspink, H. J. L., Manley, T., McGuire, D. K., Molitch, M. E., Mottl, A. K., Perreault, L., Rosas, S. E., Rossing, P., Sola, L., Vallon, V., Wanner, C., & Perkovic, V. (2021). SGLT2 Inhibition for CKD and Cardiovascular Disease in Type 2 Diabetes: Report of a Scienti c Workshop Sponsored by the National Kidney Foundation. American Journal of Kidney Diseases, 77(1), 94–109. https://doi. org/10.1053/j.ajkd.2020.08.003
Type 1 diabetes: MedlinePlus Genetics. (n.d.). Retrieved October 30, 2021, from https:// medlineplus.gov/genetics/condition/type-1diabetes/
Vallon, V., Gerasimova, M., Rose, M. A., Masuda, T., Satriano, J., Mayoux, E., Koepsell, H., omson, S. C., & Rieg, T. (2014). SGLT2 inhibitor empagli ozin reduces renal growth and albuminuria in proportion to hyperglycemia and prevents glomerular hyper ltration in diabetic Akita mice. American Journal of Physiology. Renal Physiology, 306(2), F194-204. https://doi. org/10.1152/ajprenal.00520.2013
Vallon, V., Rose, M., Gerasimova, M., Satriano, J., Platt, K. A., Koepsell, H., Cunard, R., Sharma, K., omson, S. C., & Rieg, T. (2013). Knockout of Na-glucose transporter SGLT2 attenuates hyperglycemia and glomerular hyper ltration but not kidney growth or injury in diabetes mellitus. American Journal of Physiology. Renal Physiology, 304(2), F156-167. https://doi. org/10.1152/ajprenal.00409.2012
Vargatu, I. (2016). WILLIAMS TEXTBOOK OF ENDOCRINOLOGY. Acta Endocrinologica (Bucharest), 12(1), 113. https://doi.org/10.4183/ aeb.2016.113
Wiviott, S. D., Raz, I., Bonaca, M. P., Mosenzon, O., Kato, E. T., Cahn, A., Silverman, M. G., Zelniker, T. A., Kuder, J. F., Murphy, S. A., Bhatt, D. L., Leiter, L. A., McGuire, D. K., Wilding, J. P. H., Ru , C. T., Gause-Nilsson, I. A. M., Fredriksson, M., Johansson, P. A., Langkilde, A.M., & Sabatine, M. S. (2019). Dapagli ozin and Cardiovascular Outcomes in Type 2 Diabetes. New England Journal of Medicine, 380(4), 347–357. https://doi.org/10.1056/NEJMoa1812389
Zeggini, E., Weedon, M. N., Lindgren, C. M., Frayling, T. M., Elliott, K. S., Lango, H., Timpson, N. J., Perry, J. R. B., Rayner, N. W., Freathy, R. M., Barrett, J. C., Shields, B., Morris, A. P., Ellard, S., Groves, C. J., Harries, L. W., Marchini, J. L., Owen, K. R., Knight, B., … Hattersley, A. T. (2007). Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science (New York, N.Y.), 316(5829), 1336–1341. https://doi.org/10.1126/science.1142364
Zelniker, T. A., Wiviott, S. D., Raz, I., Im, K., Goodrich, E. L., Bonaca, M. P., Mosenzon, O., Kato, E. T., Cahn, A., Furtado, R. H. M., Bhatt, D. L., Leiter, L. A., McGuire, D. K., Wilding, J. P. H., & Sabatine, M. S. (2019). SGLT2 inhibitors for primary and secondary prevention of cardiovascular and renal outcomes in type 2 diabetes: A systematic review and meta-analysis of cardiovascular outcome trials. e Lancet, 393(10166), 31–39. https://doi.org/10.1016/ S0140-6736(18)32590-X
Ziegler, A. G., Rewers, M., Simell, O., Simell, T., Lempainen, J., Steck, A., Winkler, C., Ilonen, J., Veijola, R., Knip, M., Bonifacio, E., & Eisenbarth,
G. S. (2013). Seroconversion to multiple islet autoantibodies and risk of progression to diabetes in children. JAMA, 309(23), 2473–2479. https:// doi.org/10.1001/jama.2013.6285
Zinman, B., Wanner, C., Lachin, J., & Fitchett, D. (2015). Empagli ozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes | NEJM. https://www.nejm.org/doi/full/10.1056/ NEJMoa1504720

Climate Change and its Implications for Human Health
STAFF WRITERS: LAUREN FERRIDGE '23, KEVIN STAUNTON '24, VAISHNAVI KATRAGADDA '24, ROHAN MENEZES '23, TANYAWAN WONGSRI '25, SABRINA BARTON '24, SOYEON (SOPHIE) CHO '24, EMILY BAROSIN '25
TEAM LEADS: ANAHITA KODALI '23, DINA RABADI '22
Cover Image: Global warming and climate change will signi cantly change the Earth in coming years; these changes may have signi cant implications for human health in a variety of ways. Image Source: Wikimedia Commons Introduction
Climate change, the gradual shi in earth’s climate due to global warming, is an undeniable reality. ere has long been widespread consensus in the global scienti c community that the primary cause of this warming is anthropogenic, driven by the accumulation of heat-trapping greenhouse gases in the atmosphere due to fossil fuel emissions. A recent survey of peer-reviewed climate science literature published from 2012 to 2020 determined the consensus in the scienti c community that climate change is driven by anthropogenic factors is now over 99% (Lynas et al., 2021).
Greenhouse gases, or gases in the atmosphere that trap heat, have long been emitted by natural sources like volcanoes and forest res, but the rate of these natural emissions is at a similar level to earth’s natural capacity to absorb them through greenhouse gas sinks such as the ocean and terrestrial ecosystems (Yue and Gao, 2018). Anthropogenic emissions have exceeded this capacity almost twice over, overriding the ability of earth’s natural systems to regulate global temperatures (Yue and Gao, 2018).
As previously mentioned, these emissions are primarily caused by the consumption of fossil fuels such as coal, oil and natural gas, which emit greenhouse gases when burnt – in particular carbon dioxide, followed by methane (Perera, 2017). ese fossil fuels are human society’s primary source of electricity and heat generation, and are the primary means of powering industrial activities and buildings, as well as for transportation vehicles like planes and cars (Lamb et al., 2021). According to Lamb et al. (2021), these activities collectively accounted for 86% of all anthropogenic greenhouse gas emissions between 1990 and 2018. e remaining 14% came from the agriculture, forestry and land use sector, primarily in the form of methane as a byproduct of livestock digestive processes, as well as nitrous oxide from fertilizer application and carbon dioxide from clearing of carbonsequestering natural vegetation (Lamb et al., 2021; Lynch, 2019).
disproportionate. e countries with the least emissions, largely poorer, developing countries with lower economic outputs, are the most vulnerable to climate change (Althor et al., 2016; Di enbaugh and Burke, 2019; Ahmed et al., 2009). is vulnerability may be due to the fact these countries are largely in warmer parts of the world, and therefore increased warming may cause them to su er decreased economic productivity. Furthermore, these countries lack the resources to adapt to climate change (Di enbaugh and Burke, 2019). ese impacts are now widening historic international inequalities, as many richer, developed countries have less of an incentive to reduce emissions and restrict their economic growth than their poorer, developing counterparts (Althor et al., 2016; Di enbaugh and Burke, 2019). is begs the question of what is at the heart of these striking di erences in health inequity and climate change response.
e upstream root causes of climate change and health inequity usually stem from socioeconomic inequities. Sites of institutional power such as governments, corporations, and schools can directly impact the way social structures are created; in America, the predominant social structures are related to socioeconomic status and divide the nation into lower, middle, and upper classes. As will be discussed throughout this article, it is clear that climate change disproportionately a ects those in lower economic classes. O en, institutional decisions are in uenced by notions of race, class, and sex, which contribute to the disproportionate impacts of climate change not just on lower economic classes but on people of colour and women and children (Rudolph & Gould, 2015).
ere are myriad concrete examples that show that the causes and drivers of climate change and health inequity are the same - these span energy infrastructure, transportation, housing, food, agriculture, and land use. For example, studies have shown that environmental health hazards are not distributed evenly across the American landscape. In California, the pollution burden (which can be worsened by climate change) of pesticides and toxic chemical contaminants was found to be unequally distributed throughout the state, with just 10% of the state carrying a majority of the burden. e zip codes that encapsulated these areas were found to contain higher proportions of people of colour; Hispanics, African Americans, Native Americans, and Asian/Paci c Islanders all were more likely to live in a highly polluted zip code than their nonHispanic white counterparts. As such, the burden of pollution is signi cantly higher for people of colour (Cushing et al., 2015). Other issues include redlining in urban environments, lack of public transportation from rural to urban environments, and food deserts, preventing access to clean food and water for residents (Rudolph & Gould, 2015). Poorer living conditions due to increased exposure to air pollution, increased violence, or a lack of greenery can lead to disability, death, or chronic illnesses (Rudolph & Gould, 2015). Typically, those that experience health inequities are those that are discriminated against in any way, such as racial minorities, gender minorities, or even those that live in poverty. Unfortunately, these are also the very people who do not have the political, legal, or economic power to ght against the larger institutions who play a role in controlling climate change policy as well as in decreasing health inequities ( e Lancet, 2018). us, this lack of power directly leads to greater climate change e ects as well as greater adverse health e ects faced by minorities.
Inequities in Health:
Disproportionate Impacts of Climate Change on Racial/Ethnic Groups
Many studies have continuously shown that racial and ethnic minorities will experience the worst e ects of climate change. A recent report conducted by the Environmental Protection Agency (EPA) has shown how an individual’s racial and ethnic identity directly relates to heat associated morbidity and mortality in the United States. As compared to their white counterparts, racial minorities are 35% more likely to lose working hours to heat waves, leading to the inability to a ord healthcare (EPA, 2021). ere is a strong association of heatassociated morbidity and mortality with racial and ethnic minorities, with Black and Hispanic individuals experiencing these adverse health e ects at much greater rates than their White counterparts (Gronlund, 2014). In many cases, these disproportionate e ects are due to an increase in heat vulnerability among minorities, due to factors such as housing and redlining, neighborhood crime, heat perception, or cultural or linguistic isolation. ese vulnerability factors then lead to poorer physical health and are founded in systemic discrimination, as factors like lower incomes result in minorities having to nd housing in areas with less vegetation, a greater presence of heat absorbing surfaces, or less air conditioning (Gronlund, 2014).
Hispanic and Latino individuals are at the highest risk of climate change related health-impacts, with a 43% higher chance of these individuals living in areas most a ected by climate change. Black, African American, and Paci c Islander individuals are 10% more likely to live in areas of increasing temperatures (EPA, 2021). As the threat of global warming heightens, heat waves will increase in intensity, duration, and frequency, and the impact of the weather on minorities will also become more severe. Among Hispanic individuals, studies in New York City and Phoenix have shown that in predominantly Hispanic neighborhoods, there is an increased vulnerability and increase in adverse health e ects due to heat because of linguistic isolation that may make it di cult for some to understand heat warnings or health-education messages (Gronlund, 2014). While there is the possibility for individuals in these ethnic or minority neighborhoods to travel to cooler places, there is a reluctance to do so due to cultural isolation, a lack of familiarity with activities in cooler places, and concerns with immigration or possible deportation. As ethnic and racial minorities are forced into these neighborhoods that face greater impacts of climate change related crises, health inequities faced by racial groups will continue to be perpetuated.
A fascinating case study of disproportionate climate e ects can be seen in analyzing the impact of Hurricane Harvey in Houston during 2017. A er concerns of how Hurricane Katrina disproportionately impacted Black residents of New Orleans, researchers began to study how Harvey may have had a similar impact. Using FEMA’s Inundation Footprint and other data collected on ooding from the hurricane, it was found that even a er controlling for explanatory variables, racial and ethnic minorities experienced signi cantly more ooding from the hurricane (Chakraborty et al., 2019). In areas where the population of Black residents was one standard deviation higher than the mean, there was a 4.5% increase in the mean proportion of the area that would be ooded. Increasing the number of Hispanic residents by 1 standard deviation similarly led to an increase in land ooded, by 2.6%. is suggests that ooding is localized to areas with higher proportions of minorities. Similar to the instance of increased adverse e ects due to heat, increased exposure to issues like ooding led to greater health e ects, as ooding results in immediate threats such as hypothermia, long-term physical illnesses due to poverty such as malnutrition, or mental health illnesses due to stress (Du et al., 2010). Flooding is expected to increase as climate change occurs in areas such as Houston, where storms that bring more than 20 inches of rain are already six times more likely to occur in 2019 than they were in 2000. Heavy storms are expected to increase twenty-fold by 2081 at the current rate (Chakraborty et al., 2019). is issue and others will then lead to an even greater increase in health disparities among racial groups, as they will disproportionately bear the burden of health consequences from climate change in addition to systemic inequities that already exist. Disproportionate Impacts of Climate Change on Lower Socioeconomic Classes
e various consequences of climate change o en result in crises that greatly diminish the health of lower socioeconomic classes, building upon the existing health inequity in those communities (Friel et al., 2011). People in lower socioeconomic classes generally live in warmer urban areas, which become more susceptible to extreme temperatures due to the absence of natural greenery and shade, the use of heatabsorbing building materials, and the lack of air conditioning in many of the buildings. ese environmental conditions can add to the typically hot and humid local climates in these cities, raising maximum temperatures during the day by 1 to 3°C. When this increase in temperature is added to the 2°C temperature increase expected by 2050 1.8 to 4.0°C warming by 2100 due to climate change, these urban areas will become dangerous to live in. Climate change-related heat waves around the world increase morbidity and mortality due to heat-related diseases, and humid conditions o en leave people exposed to communicable disease. ese worsening climate conditions can easily become hazards for those working outside or in con ned spaces in manufacturing and construction (Friel et al., 2011).
Rising temperatures, along with insu cient sanitation, water treatment, and drainage, also increase infectivity of diseases. Due to their living conditions, many low-income areas of the world are already extremely susceptible to malaria, dengue, diarrhea and other diseases. Flooding and other extreme weather events, which are projected to become more frequent and severe as climate change worsens, strain already poor infrastructure and thus increase the risk of contracting these diseases. More than 25% of Latin America and the Caribbean and around 60% of slums in Bangladesh lack the drainage needed to combat ooding, and safe and secure water and sanitation is absent from

around 50% of urban Asian and Africa. is lack of infrastructure increases the risk for disease in areas already very prone to disease (Friel et al., 2011).
Extreme weather events and the poor infrastructure have even more direct health consequences than increasing disease due to the extreme danger of these events. As climate change worsens, the severity of these weather events, poorer neighborhoods of the world not only have inadequate infrastructure to protect them, but also lack the funding to support evacuation e orts or rebuilding of said infrastructure. e storms and oods that have ravaged Manila as recently as 2013 exhibit the direct e ect of natural disasters on infrastructure and the health of people in those areas. Hurricane Katrina demonstrated how even living in a richer nation - like the US - does not protect those in lower economic classes from the devastating e ects of climate change. Additionally, the impending sea-level rise can raise the likelihood of these events for 13% of the urban population of the world who live in the low elevation coastal zone (Friel et al., 2011).
Climate change is particularly devastating for the 100 million people, or 2% of the world’s population, who are homeless. e lack of resources and the associated stigma limiting support makes individuals experiencing homelessness unable to protect themselves from any sort of disaster. Consistently, individuals experiencing homelessness are victims of relocation and displacement in response to disasters, which can cause all sorts of health consequences, both physical and mental. Additionally, as housing is destroyed or damaged due to natural disasters, secure housing for homeless individuals becomes even more scarce. For example, even two years a er Hurricane Katrina, over 30,000 people were still living in government-funded emergency housing. (Gibson, 2019). With limited amounts of organizations working to support these people, their health needs are not addressed or can become more severe because disaster e orts cannot reach them (Gibson, 2019). Clearly, displacement of people due to climate change disasters will result in the growth of homelessness and its subsequent health inequities.
Disproportionate Impacts of Climate Change on Women/Children
While climate change is a phenomenon that impacts everyone, women and children are disproportionately a ected compared to men. A 2014 review spanning 141 countries found that more women were killed than men by the e ects of natural disasters (WHO, 2014). Results become more disproportionate in countries where women have fewer rights than men and less disproportionate in countries where men and women were of comparable status. Hence, societal roles and responsibilities clearly place women at a health disadvantage.
Women are traditionally seen as providers and caretakers of a household. As providers, especially in rural areas, they are tasked with securing food, water, and fuel for their families (UN Women Watch, 2009). ese expectations can cause perilous consequences, especially in the event of extreme weather and natural disasters. In Bangladesh, changes in the hydrologic cycle and groundwater recharge due to climate change result in women experiencing greater exposure to waterlogged areas. is leads to the women being heavily exposed to unhygienic water and su ering from increasing gynecological issues (Neelormi, 2009). Additionally, as caretakers, women are responsible for tending to other members of the family and could, as a result, be forbidden from leaving the house unattended.
Image 1: is image shows the percent of the population below the poverty line in the US based on 2015-2019 data. e disparities in the distribution of impoverished people throughout the US is clear - certain states carry a higher burden of poverty than others.
Image Source: Wikimedia Commons
When cyclones struck Bangladesh in 1991, many women died waiting for their family members to return home before departing together to a secure location, despite many cyclone warnings (Aguilar, 2004). is resulted in a death rate that was 71 per 1000 women, but only 15 per 1000 men. ese disproportionate e ects are also seen in developed countries, such as with Hurricane Katrina in Louisiana. e population that was most severely a ected by ooding as a result of the hurricane were women (Butterbaugh, 2005). In the chaos created by power outages and displacement, multiple women claimed that they had been sexually assaulted while sheltering at the Superdome; their claims have yet to be taken seriously by authorities. Hurricane Katrina was also particularly devastating as single-mother households made up 56 percent of families in New Orleans. It is only logical that any harm endured by mothers would go on to a ect their children. It is suggested by multiple studies that children who experience hurricanes are at higher risk for developing depression, anxiety, or posttraumatic stress disorder (Goenjian et al., 2001; Kessler et al., 2006).
Looking at the more general e ects of climate change on children’s health, we can also deduce that the poor health of mothers will lead to the low health of infants. In fact, children are predicted to be the population most severely disadvantaged by climate change, as they are expected to bear 88 percent of the burden of disease (Zhang et al., 2007; Patz et al., 2007). According to a comprehensive review by the Harvard School of Public Health, global warming allows insects to migrate to new places, taking insect-borne diseases such as malaria, zika, dengue with them (Harvard T.H. Chan School of Public Health, 2021). Heavier rainfall that comes with climate change can increase oods, leading to diarrheal diseases, which are particularly harmful in young infants. Air pollution, which currently accounts for 20 percent of global infant deaths, will increase as a result of the root causes of climate change (State of Global Air, 2020). e current e ects of climate change on children’s physical and mental health are already devastating; if no action is taken to combat this issue, the scale of destruction caused by climate change will only increase.
Disproportionate Impacts of Climate Change based on Geographical Location
e consequences of climate change have impacted disproportionately impacted populations in di erent geographical locations. For example, rural and urban populations have been impacted by climate change in di erent ways.
Climate change has increased the severity and frequency of droughts around the world. In particular, according to the 1997 study by the World Resources Institute, 29% of the world population live in dry land, or arid or semi-arid zones which are more vulnerable to droughts and extreme temperatures. Additionally, the geographical distribution of dry land disproportionately impacts di erent continents more than others, as 45% of Africa’s population, 43% of Asia’s population, and 44% of developing regions’ population live in dry land, compared to 17% of the population in the Americas and the Caribbean (World Resources Institute, 1997). ese patterns support the same data that rural areas are more vulnerable to droughts: 43% of rural populations are exposed to droughts, compared to 32% of urban populations. Given that disadvantaged groups like cattle farmers, ethnic minorities and populations under the poverty line are more likely to live in rural areas than urban areas, rural populations are disproportionately impacted by the increases in droughts due to climate change (Islam and Winkel, 2017). Furthermore, rural households with lower income are o en limited to keeping livestock as their primary asset, which are vulnerable to droughts due to their continued dependence on water and food (Nkedianye et al., 2011). Wealthier households can own di erent types of assets that are less a ected by droughts or extreme climate hazards, adding to the unequal impact of climate change on the rural poor.
e consequences of extreme temperatures have also disproportionately impacted the vulnerable groups in urban areas. Disadvantaged groups like ethnic minorities, slum dwellers, and other groups with lower socioeconomic status will be more likely to live in areas with poor ventilation and poor heat management infrastructure, meaning they would be more a ected in their workplaces and homes by the extreme heat (Kovats and Akhtar, 2008). Furthermore, in these urban areas, maximum temperatures are 1 to 3°C higher than cities with more parks, because they have higher population density and fewer trees to absorb the carbon dioxide produced by vehicles and facilities (Ferguson et al., 2008).
Extreme weather events also cause unequal consequences for the at-risk populations. To focus on speci c case studies, disadvantaged groups like slum dwellers live in lower elevation
Image 2: is graph shows various populations in the US that lack access to food and transport to grocery stores or other sources of food as of 2010.
Image Source: Wikimedia Commons
areas in Dhaka, Bangladesh, which put them at greater risk in oods (Braun and Abheure, 2011). In Latin American and Caribbean countries, the dwellings of disadvantaged groups are in hilly slopes that are much more vulnerable to mudslides, which are increasing in frequency with climate change, and more than 25% of them also lack appropriate drainage systems for oods (Painter, 2007; Bartlett et al., 2009). Lower income households in urban areas are o en limited to keeping assets in the form of housing, which are at risk of damage by oods (Moser, 2007; Islam and Winkel, 2017).
Furthermore, these consequences of climate change impact urban food security because extreme temperatures and extreme weather events like storms or oods can damage crops, livestock, or infrastructure for the food industry, increasing food prices. Higher food prices disproportionately a ect low-income urban populations, since they have limited access to non-market food sources and limited budgets for their food (Cohen and Garrett, 2010). ese groups within a city will tend to rely on inexpensive food that t their budgets, which would cause malnutrition due to the food’s poor nutritional value (Brinkman et al., 2010).
Vulnerable groups like poorer urban and rural populations are not only a ected by the consequences of climate change but also have limited options of migrating to safer regions. Case studies of Manila’s oods and storms and the 2005 New Orleans oods have demonstrated that poorer urban households cannot easily migrate to higher elevation regions or cities with safer buildings and infrastructure that prevent damage from extreme weather events (Costello et al., 2009). ese cases demonstrate that vulnerable populations in rural and urban areas are disproportionately impacted by climate change in various aspects.

Climate Change and Health:
Extreme Weather Events
Climate change brings on a lot of occurrences that can a ect the likelihood of extreme weather events. Based on the measurable human impact on the environment, especially the increase of greenhouse gasses, it has become likely that there will be a warming of temperatures across the board and an rise in sea level, with a slightly lower likelihood of an increase in the intensity of extreme precipitation (Sauerborn & Ebi, 2012). ese changes are likely to cause increases in occurrences or intensity of cyclones, droughts, oods, wild res and other events. Extreme weather events are natural occurrences that are generally unexpected or unpredictable, can cause a lot of destruction, and are known to become more common with the onset of climate change. Due to their unpredictability, they can greatly damage populations, communities, and infrastructure. ese extreme natural disaster events can cause death and severe injuries, but they can also increase the risk of disease, communicable and noncommunicable, food
Image 3: Hurricanes are one type of extreme weather event that may become more common due to climate change. Image Source: Wikimedia Commons scarcity, shelter loss and forced migration (Sauerborn & Ebi, 2012). Additionally, the potential disruption of pregnancy and delivery, education, clean water, pregnancy and delivery, sanitation and health infrastructure can have serious adverse health e ects on all those a ected by the event (Sauerborn & Ebi, 2012).
In addition to the immediate injuries people accumulate during natural disaster events, there are many health consequences that can arise for individuals undergoing stress associated with such a traumatic event, especially coronary ailments like potential heart attacks. e stress from extreme weather events can also a ect the ability to ght o disease or infection and can even are up allergies and asthma. Psychologically, stress and trauma can also have extreme mental health consequences that may not appear for weeks or months, including PTSD, anxiety, depression and others like them. All of these happening simultaneously can clog up medical centers in the areas around extreme weather events, which can prevent many, especially those who have preexisting conditions like diabetes, from receiving adequate care (Hidalgo & Baez, 2019).
e stress of traumatic events can majorly psychologically a ect the well-being of victims. To this end, researchers conducted a study focused on mental health following extreme weather events in Australia, an area which is prone to wild res, earthquakes, landslides, oods, cyclones and more (Morrissey & Reser, 2007). ey found that individuals with anxiety or previous traumatic experiences can be more seriously a ected a er a new traumatic occurrence, which aligns with the consensus that what makes traumatic events more impactful is vulnerability, in this case, psychological vulnerability. Just as more stable infrastructure can help minimize the destruction, increasing government funding for adequate mental health care and widening associated programs can help prevent worse mental health issues (Morrissey & Reser, 2007).
e damage that comes from extreme weather events is all related to the quality of the response, but with climate change making these events worse and worse, the response shouldn’t be limited to that particular event. By combining disaster risk reduction with climate change adaptation, organizations (like NGOs) looking to provide support can help minimize the health damage among the population a ected and limit the vulnerability across the board. Limiting vulnerability could be accomplished through building and rebuilding infrastructure as well through changing policy, strengthening and streamlining the healthcare system, and teaching emergency preparedness and response and locating other areas of risk. Since a disaster completely disrupts daily life, a proper and e ective health response should be holistic and far-reaching (Banwell et al., 2018).
Rising Sea Levels
Another major consequence of climate change is posed by rising sea levels, primarily caused by the melting of large terrestrial ice sheets and the thermal expansion of the ocean as it absorbs atmospheric heat (Mimura, 2013). Levitus et al. (2012) found that sea levels have risen by an average 0.54mm per year from 1955 to 2010. is rate of sea level rise has been increasing over time (Mimura, 2013). While this may not seem signi cant, if serious mitigation measures are not taken, sea levels are predicted to rise 0.5

to 2m above sea level by 2100 (Nicholls et al., 2011). is could cause the forced displacement of as many as 187 million people, equivalent to 1 out of every 40 people worldwide (Nicholls et al., 2011). ere is also a more immediate threat posed by rising sea levels; ood risk. Sea level rises of as little as 1-10cm can exponentially increase rates of extreme ooding (Taherkhani et al., 2020). In the US for example, it is estimated that rates of extreme ooding are expected to double every ve years due to climate changeinduced sea level rise (Taherkhani et al., 2020). Using this estimation, within 50 years, risks of extreme ooding will increase by a factor of over a thousand (Taherkhani et al., 2020).
Sea level rise and the accompanying increased frequency of extreme ooding has major consequences for global health. Such e ects include the direct threat higher sea levels pose to poor and vulnerable communities worldwide, but particularly in developing countries in the global South which lack the resources or infrastructure to adequately adapt (Byravan, 2010). Such communities are also less likely to have disaster insurance or other means of recovering from losses due to sea-level rise and extreme weather events, rendering them even more vulnerable to displacement (Public Health institute/Center for Climate Change and Health, 2016). However, there are other signi cant impacts to consider. ese include the increase of saltwater contamination of freshwater sources, the consumption of which may lead to hypertension and other diseases like diarrhea (Wong et al., 2014). is is particularly a problem for groundwater pools which island communities rely on, since they lack easy access to alternate sources of freshwater (Mimura, 2013).
Sea level rise and saltwater intrusion also indirectly a ect communities’ health by degrading populations of mangroves in tropical and subtropical areas, primarily in Asia, where they are most likely to grow (Mimura, 2013; Barbier et al., 2011). Many mangrove species depend on aerial roots which extend above the water’s surface for much of their respiration, which higher sea levels prevent (Mimura, 2013). Among other ecosystem sources, Mangroves mitigate the e ects of storms and other extreme weather events on coastal communities by acting as a bu er, so their loss renders such communities more vulnerable to extreme weather events (Barbier et al., 2011). e same e ects have been observed for wetland ecosystems (which cannot survive in highly saline environments) with similar consequences (Barbier et al., 2011; Ward Additional e ects of sea level rise on global health include soil inundation and salinization, which may turn agricultural land permanently infertile (Gomall, 2010). Temporary storm surges, whose level is increased by sea level rise, may also temporarily degrade or inundate agricultural land (Gomall, 2010). Both of these e ects impact food security for communities living along the coast or river deltaic regions, which are particularly vulnerable to ooding (Gomall, 2010). As such, countries such as India, Pakistan, Myanman and Bangladesh are particularly vulnerable, as these countries have increasingly relied upon such ood-prone areas in an e ort to feed their growing populations (Gomall et al., 2010; Webster, 2008). us, sea level rise is detrimental to food security, thereby impacting global health as well.
Ice Melts
Ice melt as both a general scienti c concept and as a known consequence of climate change has rapidly entered into the public consciousness over the course of the past few decades, largely as a result of public awareness campaigns centered on melting ice caps and rising sea levels. Ice melt can be de ned generally as the ice mass lost from either the surface or base of an ice shelf or sheet, which almost always enters the surrounding ocean as meltwater (Mackie et. al, 2020). Both melting ice caps and rising sea levels contribute to ice melt. Ice melt is neither a negative or positive phenomenon – it has always occurred, as it serves to regulate the size of ice sheets and plays an important role in the circulation of various Arctic water currents. Ice melt has raised concerns in recent history because of the rate at which ice melt is occurring.
Increasing levels of ice melt and Arctic warming have particularly devastating consequences for the residents of these areas; many Arctic residents rely on hunting, shing, and gathering for food and consistent climate conditions for safe food storage. As the climate warms, sh and meat may not be able to dry, and food stored below ground may go bad as permafrost melts. Additionally, outbreaks of food-related bacterial diseases and illnesses will likely increase as temperatures warm, as bacteria like Clostridium botulinum (responsible for botulism) and Vibrio parahemolyticus (responsible for gastrointestinal illness) are able to germinate at warm temperatures. Warmer temperatures may also change migratory patterns of animals and allow
animals to carry disease for longer, both of which ultimately a ect the sources of food available to Arctic residents (Parkinson & Evengård, 2009). As it is predominantly Indigenous peoples that reside in the Arctic, Indigenous peoples - already an underserved population by healthcare systems - will disproportionately take on the brunt of the impact of ice melt and Arctic warming (Cochran & Geller, 2002).
Beyond impacting food sources, ice melt may result in the release of microorganisms, many of which can be pathogenic in humans. Permafrost and glaciers, both of which are usually frozen constantly, both contain a myriad of microorganisms, which are typically dormant; however, as global warming continues to increase levels of ice melt, these microorganisms are released into the natural ecosystems that surround typically frozen structures. e massive release of microorganisms into the environment has already contributed to increases in disease - for example, the 2016 anthrax outbreaks in Siberia were associated with permafrost melt (Yarzábal et al., 2021; Stella et al., 2020). As more disease-causing bacteria, viruses, and other microbes are released, scientists hypothesize that new epidemics may appear in the coming years.
Ice melts, in addition to directly contributing to human disease, can also have downstream e ects that can further impact health. For scientists to develop a complete and nuanced understanding of ice melt and its implications in the broader scheme of climate change, it has been crucial to create advanced methods of modeling ice melt and its e ects. e algorithms used to model ice melt have rapidly advanced within the last few years. Ice melt models are o en used in conjunction with either ocean forcing models (models that show how the ocean responds to various real world forces) or earth system models (models that show how carbon moves through the Earth’s atmosphere) to better acknowledge the far-reaching e ects of ice melt (Goldberg et al., 2018). Current models, while accurate, o en fail to account for more nuanced factors that can alter ice melt: many models oversimplify ocean forcings, while others are too low-resolution to draw accurate conclusions concerning ice melt and potential positive feedback loops along glacial coasts. (Moorman et. al, 2020). Traditional models that incorporated the e ects of melt ponds (pools of open water that form on the surface of Arctic ice during the summers) utilized satellite imagery yet failed to take into account color saturation. is resulted in the variability of di erent ice ponds’ e ects on albedo and solar radiation not being acknowledged (Mingfeng et al., 2020). Mingfeng et al. were able to develop a method that takes color saturation into account when looking at melt pond data, underscoring the need and urgency of obtaining data that is reliable and accurate.
Despite their aws, ice melt models have yielded valuable insights into just how ice melt interacts with other aspects of the climate system, particularly with ocean circulation. Increasing ice melt causes a considerable in ux of freshwater into the ocean, so it has been important to investigate how coastal freshening (the addition of freshwater to the coasts along glaciers) is a ecting water circulation patterns. It has been found that Antarctic Bottom Water (AABW), a dense water mass, is negatively impacted by coastal freshening; this is due to the fact that the formation of the source from which it originates, Dense Shelf Water (DSW) is sensitive to freshwater forcings (Moorman et. al, 2020). Models have also revealed that ice shelves are most prone to melt near grounding lines, the delineation between where they are attached to bedrock and where they become free-standing ice sheets. (Goldberg et al., 2018). It is thought that Circumpolar Deep Water (CDW) breaching ice shelves at over ow sites could be the driving force of basal glacial melt beneath ice shelves, particularly near already weakened grounding lines (Moorman et al., 2020). Studies completed in the past with less accurate ice melt models came to the conclusion that coastal freshening would invariably lead to warming trends in coastal waters – this has recently been disproven with the use of a more accurate model. e updated model showed that coastal freshening can lead to both warming and cooling trends, and that coastal meltwater can either accelerate or inhibit ice shelf melt. (Moorman et al., 2020). Another study, which experimented with modeling by combining an ESM (earth system model) with a dynamic ice sheet model, set out to nd out more about the climate response to increasing melt from the Antarctic shelf. ey found that freshwater entering as meltwater tends to form a buoyant layer on top of the saline water below it, increasing the heat content of the midlayer ocean water and preventing ventilation. is in turn increases strati cation, which, as previously mentioned, can reduce AABW formation (Mackie et al., 2020). In sum, increasing ice melt disrupts the established patterns of oceanic circulation. ese changes, in turn, can result in chemical and bacterial contamination of waters that are used for shing or other sources of food production, potentially contaminating many sources of food

for marine animals and humans. Changes in oceanic circulation also result in changes in heat distribution throughout the ocean, which can cause harmful algal blooms which can release extremely potent natural toxins into the water; these can be ingested by humans or by animals that humans go on to consume (Fleming et al., 2006).
Overall, it is important to our growing understanding of ice melt and its implications on the climate to continue to develop more accurate and nuanced models of ice melt, which ideally will be used in conjunction with models of other parts of the climate system. Ice melts lead to rising sea levels, extreme weather events, and disproportionate impact on marginalized populations.
Food and Waterborne Disease
Food- and water-borne diseases are signi cantly in uenced by climate change. e WHO estimates an “additional 48,000 deaths in children under 15 years old mostly due to diarrheal diseases by 2030 and 33,000 deaths by 2050” (Cissé, 2019). On a broader scale, the impact of diarrheal related diseases, the leading cause of food and water-borne disease burden in human health, is projected to have the greatest burden of mortality impacts with respect to climate change in sub-Saharan Africa by 2030 but leading burden is projected to shi to Southeast Asia by 2050 (Cissé, 2019). Before fully understanding the impact of climate change on food and water borne disease, it is helpful to rst know the scope and severity of current food and water borne disease burden.
Unsafe water used for the cleaning and processing of food is a key risk factor contributing to foodborne diseases. While children under the age of 5 represent 9% of the global population, 43% of food borne disease burden is within this group (Kirk et al., 2015). Salmonella, norovirus, cholera and typhoid are the leading food-borne disease threats; however it is important to recognize that di erent diseases present more of a threat in di erent areas. For example, foodborne cholera presents the biggest burden in Africa and Asia whereas brucellosis and M. bovis have the highest burden in the Middle East and some African regions (Kirk et al., 2015). On a global scale, food borne disease burden is high generating over 600 million illnesses and 420,000 deaths worldwide (Cissé, 2019). Norovirus is the leading cause of food-born illnesses causing 125 million cases and of all food-borne diseases, diarrheal and invasive infections due to nontyphoidal Salmonella causes the highest burden
Image 4: Ice melts can be seen in many bodies of water, including Lake Baikal, located in Russia. Image Source: Wikimedia Commons
a
leading to 4.07 million disability-adjusted life years (DALYs). In the African region, there were 91 million food-borne disease cases and 137,000 deaths per year. Furthermore, diarrheal diseases contributed to 70% of those deaths. (Cissé, 2019). As climate change is leading to increased facilitation of contamination and transmission of food-borne viruses and pathogens, mortality and case incidence rates are expected to increase. To further understand the impact of climate change on food-borne diseases, it is also necessary to understand the climate change relation to waterborne diseases as water plays a major role in both food and waterborne diseases and the separation of food and water exposure is di cult.
Human exposure to water-born infections occurs by contact with or ingestion of contaminated recreation and/or drinking water. Drinking water containing infectious pathogens is the main driver of the burden of water-borne diseases. e most burdensome water-borne diseases are diarrheal diseases, cholera, shigella, and typhoid. Just like food-borne diseases, low-and middle-income countries (LMICs) have the highest waterborne disease burden which is estimated at 842,000 deaths a year including 361,000 in children under the age of 5 (Cissé, 2019). Lack of basic hygiene and sanitation and failing health and water delivery infrastructure are the leading challenges in the ght against water borne diseases. Climate change is projected to exacerbate risk of diarrheal diseases and other water-borne diseases in LMICs as repercussions of climate change intensify.
Climate change has a wide variety of e ects including “rising temperature, soil degradation, loss of productivity or agricultural land, deserti cation, loss of biodiversity, degradation of ecosystems, reduced fresh-water resources, acidi cation of oceans, and the disruption and depletion of stratospheric ozone” (Rossati, 2016). All of these consequences impact human health, increasing frequency of distribution, timing, and intensity of infectious disease and noncommunicable diseases, malnutrition in famine, and increased mortality from complications with heat waves (Rossati, 2016). All environmental e ects disproportionately impact children. Diseases with the greatest environmental contribution in children under the age of 5 include lower respiratory infections (32%), diarrheal diseases (22%), neonatal conditions (15%) and parasitic and vector-borne diseases (12%) (Cissé, 2019). Quality and quantity of water is important to gauging the burden of infectious diseases in LMICs as its e ects go beyond the food chain. Water-related infectious diseases are already a major cause of mortality and morbidity; these diseases are exacerbated by climate change, posing new challenges to the public and global health sector for food and water borne diseases. Trends in Africa and Asia reveal that extreme climate change induced events such as oods will increase risk of infectious diseases spreading through water systems, and, conversely, improvements related to drinking water, sanitation and hygiene are e ective methods to signi cantly reduce intestinal parasitic infections in school-aged children (Cissé, 2019).
Food and water borne diseases are inextricably connected because of the in uence of contaminated and unclean water on food supplies. In low- and middle-income countries (LMICs) the increased frequency of oods because of increasing global temperatures exacerbates challenges with water pollution, which subsequently escalates risk for foodand water- borne diseases, disproportionately impacting people in low socioeconomic communities (Cissé, 2019). ere are direct and indirect ways in which climate change a ects food- and water-borne diseases. Direct impacts refer to extreme climate or environmental events such as ooding or sea-level rise which leads to increased water contamination due to presence of fecal-oral pathogen presence in the environment. Indirect impacts are mostly climatic factors such as temperature and humidity that in uence processes of pathogen replication and survival and rising frequency and relevance of related con icts such as agriculture, water resource management, and population displacements (Walker, 2018). Water- and food- borne diseases are linked to the ingestion of pathogens via contaminated water or food. e diseases are further connected as contaminated water can contaminate food and increase risk of transmission.
Vector Borne Disease
A vector borne disease is a “disease that results from an infection transmitted to humans and other animals by blood-feeding arthropods, such as mosquitoes, ticks, and eas” (VectorBorne Diseases, n.d.). Speci cally, a vector is “an organism that transmits an infectious pathogen from an infected human or animal host to an uninfected human” (Rocklöv & Dubrow, 2020). Global disease burden of vector-borne diseases is led by malaria, dengue, schistosomiasis, leishmaniasis, Chagas disease and African trypanosomiasis which infect more than one billion people and kills over a million people every year (Campbell-Lendrum et al., 2015).
In tandem with a global temperature increase from 1.5 °C to 2 °C, risk and incidence of malaria and dengue fever, two of the most pertinent vector-borne diseases are also projected to increase and shi geographical regions of interest. (Cissé, 2019). Additionally, if global temperature increases by 2 to 3°C as expected, the population at risk for malaria would increase by as much as 3 to 5% (Rossati, 2016). A key factor in understanding and responding to vector-borne diseases is the modes and intricacies of transmission. ese are the multifactorial pathways through which climate change and climate variability a ect human health including social, environmental, ecological, and economic factors, as each of them impact survival and growth of human and pathogen populations (Cissé, 2019).
Vector borne diseases have one of the highest disease burdens globally but disproportionately impact the global south. is is because “vectorborne diseases have wider socioeconomic impacts, increasing health inequities, and acting as a brake on socioeconomic development” (Campbell-Lendrum et al., 2015). ese burdens and inequities are exacerbated by climate change as the mortality rate from vector borne diseases is almost 300 times greater in developing countries than it is in developed countries (Campbell-Lendrum et al., 2015). e burden is exponentially larger due to increased frequency of vector-borne diseases in tropical climates that is common to many developing countries and because of concurrent low levels of socioeconomic development and health service coverage in these areas. On an individual level, the burdens are greater in impoverished populations because individuals are subject to poorer environmental and social conditions such as poor quality housing and increased proximity to vector breeding sites; and lack of access to preventative health interventions and disease treatments (Campbell-Lendrum et al., 2015).
Climate change presents a threat to global health and an already severe vector-borne disease global burden. is risk for vector-borne disease is increasing, and “put simply, vectors, which are ectotherms (that is, cold-blooded animals), do better in a warmer world” (Rocklöv & Dubrow, 2020). A warmer climate is more favorable to the survival and completion of vector life cycles and, in the case of mosquitos, is even capable of speeding it up. is has enabled extension of areas of disease distribution in direct correlation with increasing temperatures (Rossati, 2016). Extension of vector regions and subsequently infections can be observed in changing tick behaviors. Tick-borne diseases have increased over past years because rising temperatures in cold regions have “accelerated the cycle of development, the production of eggs, and the density and distribution of the tick population” (Rossati, 2016). Due to climate change, ticks have been found in more regions than ever before and are also found at higher altitudes, presenting new and increased climate change burden.
Precipitation is another important environmental in uence on vector transmission and breeding. It mostly a ects vectors that have aquatic developmental stages such as mosquitos. Humidity, which is related to precipitation, creates a better environment for diseases transmitted by vectors without aquatic developmental stages such as ticks or sand ies (Campbell-Lendrum et al., 2015).
As many impacts of climate change, including increased precipitation, humidity, ooding, and increased areas with warmer climates are becoming more frequent, so are the frequency of vector-diseases transmissions, infections, and numbers and expansion of burdens. is is in part because of the socioeconomic factor related to disease burden which cannot be excluded when considering remediation methods or global health strategies. As threat of increased infections in more places rise an epidemiological approach to vector-borne diseases must include attention to ecology and behavior of the host, the ecology and behavior of the carrier, and the level of immunity of the population” (Rossati, 2016) and historical and social relationship of impacted communities with such diseases.
Occupational Health and Productivity Risks
Climate change poses a risk to the occupational health and safety of workers. As a result of rising temperatures, many workers working outdoors or in hot indoor conditions will be at an increased risk of su ering from heat-related disorders. Due to these heat-related ailments, workers are more prone to lapses in judgment and reduced alertness, increasing their chances of su ering a workplace injury (EPA, 2016; Tawatsupa, 2016). Other e ects of climate change on the health of outdoor workers could include increased poor air quality and diseases transmitted by ticks and mosquitoes (EPA, 2016). Depending on the type of job they hold, workers might also be more frequently exposed to occupational hazards. For example, as climate change causes warmer and drier conditions in forests, the intensity "Climate change poses a risk to the occupational health and safety of workers."
Image 5: Mosquitos are a common vector for malaria across the world.
Image Source: Wikimedia Commons and frequency of which wild res happen will increase. Studies have correlated climate change to increasing instances of occupational health and safety hazards - such as burns and heat exhaustion - of re ghters (Adetona et al., 2016; Britton et al., 2013).
Looking beyond health e ects, climate change will also negatively a ect our economy as it decreases productivity and the supply of workers. Revisiting rising temperatures, this can cause workers to feel more fatigued and a ects the supply of resources, such as crops, that could be cultivated. Additionally, if more workers su er from workplace injuries caused by climate change, this will decrease the number of workers and a lost work capacity (Ebi et al., 2017). In a more indirect manner, rising temperatures and heavier rainfalls will also lead to increased intergroup con ict in the workplace (National Bureau of Economic Research, 2015). is could also a ect productivity as workers will become preoccupied with resolving con ict rather than focusing on their work.
While discussing occupational health and productivity risks, we must also consider that workers are already a vulnerable population. Due to factors such as socioeconomic status, race, and immigration status, many who have healthendangering jobs do so because of necessity (Levi & Patz, 2015). rough evaluating the e ects of climate change on this area, we can see how it a ects those who are least fortunate.
Conclusion
Climate change has major implications for human health and will result in the exacerbation of health inequities across the globe. Events that will impact health include rising sea levels, ice melts, extreme weather events, airway diseases and increased presence of allergens due to pollution, food and water borne diseases, vector borne diseases, and occupational risks. ese incidents will disproportionately a ect vulnerable populations such as those who are part of racial or ethnic minorities, those in lower socioeconomic classes, and women and children. Addressing climate change with a sense of urgency is key in adequately slowing down progression of such events to minimize drastic health repercussions, particularly in vulnerable populations.
References
Ahmed, S. A., Di enbaugh, N. S., & Hertel, T. W. (2009). Climate volatility deepens poverty vulnerability in developing countries. Environmental Research Letters, 4(3), 034004. https://doi.org/10.1088/1748-9326/4/3/034004
Althor, G., Watson, J. E. M., & Fuller, R. A. (2016). Global mismatch between greenhouse gas emissions and the burden of climate change. Scienti c Reports, 6(1), 20281. https://doi. org/10.1038/srep20281
Ambrey, C., Byrne, J., Matthews, T., Davison, A., Portanger, C., & Lo, A. (2017). Cultivating climate justice: Green infrastructure and

suburban disadvantage in Australia. Applied Geography, 89, 52–60. https://doi.org/10.1016/j. apgeog.2017.10.002
Banwell, N., Rutherford, S., Mackey, B., & Chu, C. (2018). Towards Improved Linkage of Disaster Risk Reduction and Climate Change Adaptation in Health: A Review. International Journal of Environmental Research and Public Health, 15(4), 793. https://doi.org/10.3390/ ijerph15040793
Barbier, E. B., Hacker, S. D., Kennedy, C., Koch, E. W., Stier, A. C., & Silliman, B. R. (2011). e value of estuarine and coastal ecosystem services. Ecological Monographs, 81(2), 169–193. https:// doi.org/10.1890/10-1510.1
Bartlett, S., Dodman, D., Hardoy, J., Satterthwaite, D., & Tacoli, C. (2009). Social Aspects of Climate Change in Urban Areas in Low- and Middle Income Nations.
Braun, B., & Aßheuer, T. (2011). Floods in megacity environments: Vulnerability and coping strategies of slum dwellers in Dhaka/Bangladesh. Natural Hazards, 58(2), 771–787. https://doi. org/10.1007/s11069-011-9752-5
Brinkman, H.-J., de Pee, S., Sanogo, I., Subran, L., & Bloem, M. W. (2010). High Food Prices and the Global Financial Crisis Have Reduced Access to Nutritious Food and Worsened Nutritional Status and Health. e Journal of Nutrition, 140(1), 153S-161S. https://doi.org/10.3945/ jn.109.110767
Byravan, S., & Rajan, S. C. (2010). e Ethical Implications of Sea-Level Rise Due to Climate Change. Ethics & International A airs, 24(3), 239–260. https://doi.org/10.1111/j.17477093.2010.00266.x
Campbell-Lendrum, D., & Corvalán, C. (2007). Climate Change and Developing-Country Cities: Implications For Environmental Health and Equity. Journal of Urban Health, 84(S1), 109–117. https://doi.org/10.1007/s11524-007-9170-x
Campbell-Lendrum, D., Manga, L., Bagayoko, M., & Sommerfeld, J. (2015). Climate change and vector-borne diseases: What are the implications for public health research and policy? Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1665), 20130552. https://doi.org/10.1098/rstb.2013.0552
Chakraborty, J., Collins, T. W., & Grineski, S. E. (2019). Exploring the Environmental Justice Implications of Hurricane Harvey Flooding in Greater Houston, Texas. American Journal of Public Health, 109(2), 244–250. https://doi. org/10.2105/AJPH.2018.304846
Cissé, G. (2019). Food-borne and water-borne diseases under climate change in low- and middle-income countries: Further e orts needed for reducing environmental health exposure risks. Acta Tropica, 194, 181–188. https://doi. org/10.1016/j.actatropica.2019.03.012
Climate Change and Social Vulnerability in the United States: A Focus on Six Impacts. (n.d.). 101. Cochran, P. L., & Geller, A. L. (2002). e Melting Ice Cellar. American Journal of Public Health, 92(9), 1404–1409.
Cohen, M. J., & Garrett, J. L. (2010). e food price crisis and urban food (in)security. Environment and Urbanization, 22(2), 467–482. https://doi. org/10.1177/0956247810380375
Costello, A., Abbas, M., Allen, A., Ball, S., Bell, S., Bellamy, R., Friel, S., Groce, N., Johnson, A., Kett, M., Lee, M., Levy, C., Maslin, M., McCoy, D., McGuire, B., Montgomery, H., Napier, D., Pagel, C., Patel, J., … Patterson, C. (2009). Managing the health e ects of climate change. e Lancet, 373(9676), 1693–1733. https://doi.org/10.1016/ S0140-6736(09)60935-1
Cushing, L., Faust, J., August, L. M., Cendak, R., Wieland, W., & Alexee , G. (2015). Racial/ Ethnic Disparities in Cumulative Environmental Health Impacts in California: Evidence From a Statewide Environmental Justice Screening Tool (CalEnviroScreen 1.1). American Journal of Public Health, 105(11), 2341–2348. https://doi. org/10.2105/AJPH.2015.302643
Di enbaugh, N. S., & Burke, M. (2019). Global warming has increased global economic inequality. Proceedings of the National Academy of Sciences of the United States of America, 116(20), 9808–9813. https://doi.org/10.1073/ pnas.1816020116
Dokken, D. (n.d.). 5—Coastal Systems and LowLying Areas. 49.
Du, W., FitzGerald, G. J., Clark, M., & Hou, X.Y. (2010). Health Impacts of Floods. Prehospital and Disaster Medicine, 25(3), 265–272. https:// doi.org/10.1017/S1049023X00008141
Ferguson, B., Fisher, K., & Golden, J. (2012).
Reducing Urban Heat Islands: Compendium of Strategies—Cool Pavements. U.S. Environmental Protection Agency. https://www.epa.gov/sites/ default/files/2017-05/documents/reducing_ urban_heat_islands_ch_5.pdf
Ferring, D., & Hausermann, H. (n.d.). Full article: e Political Ecology of Landscape Change, Malaria, and Cumulative Vulnerability in Central Ghana’s Gold Mining Country. Retrieved October 14, 2021, from https://www.tandfonline. com/doi/full/10.1080/24694452.2018.1535885
Fleming, L. E., Broad, K., Clement, A., Dewailly, E., Elmir, S., Knap, A., Pomponi, S. A., Smith, S., Gabriele, H. S., & Walsh, P. (2006). Oceans and human health: Emerging public health risks n the marine environment. Marine Pollution Bulletin, 53(10–12), 545–560. https://doi.org/10.1016/j. marpolbul.2006.08.012
Friel, S., Hancock, T., Kjellstrom, T., McGranahan, G., Monge, P., & Roy, J. (2011). Urban Health Inequities and the Added Pressure of Climate Change: An Action-Oriented Research Agenda. Journal of Urban Health, 88(5), 886–895. https:// doi.org/10.1007/s11524-011-9607-0
Gibson, A. (2019). Climate Change for Individuals Experiencing Homelessness: Recommendations for Improving Policy, Research, and Services. Environmental Justice, 12(4), 159–163. https:// doi.org/10.1089/env.2018.0032
Goldberg, D. N., Gourmelen, N., Kimura, S., Millan, R., & Snow, K. (2019). How Accurately Should We Model Ice Shelf Melt Rates? Geophysical Research Letters, 46(1), 189–199. https://doi.org/10.1029/2018GL080383
Gornall, J., Betts, R., Burke, E., Clark, R., Camp, J., Willett, K., & Wiltshire, A. (2010). Implications of climate change for agricultural productivity in the early twenty- rst century. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1554), 2973–2989. https://doi. org/10.1098/rstb.2010.0158
Gronlund, C. J. (2014). Racial and Socioeconomic Disparities in Heat-Related Health E ects and eir Mechanisms: A Review. Current Epidemiology Reports, 1(3), 165–173. https:// doi.org/10.1007/s40471-014-0014-4
Hidalgo, J., & Baez, A. A. (2019). Natural Disasters. Critical Care Clinics, 35(4), 591–607. https://doi.org/10.1016/j.ccc.2019.05.001 Kirk, M. D., Pires, S. M., Black, R. E., Caipo, M., Crump, J. A., Devleesschauwer, B., Döpfer, D., Fazil, A., Fischer-Walker, C. L., Hald, T., Hall, A. J., Keddy, K. H., Lake, R. J., Lanata, C. F., Torgerson, P. R., Havelaar, A. H., & Angulo, F. J. (2015). World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Medicine, 12(12), e1001921. https://doi.org/10.1371/ journal.pmed.1001921
Kovats, S., & Akhtar, R. (2008). Climate, climate change and human health in Asian cities. Environment and Urbanization, 20(1), 165–175. https://doi.org/10.1177/0956247808089154
Lamb, W. F., Wiedmann, T., Pongratz, J., Andrew, R., Crippa, M., Olivier, J. G. J., Wiedenhofer, D., Mattioli, G., Khourdajie, A. A., House, J., Pachauri, S., Figueroa, M., Saheb, Y., Slade, R., Hubacek, K., Sun, L., Ribeiro, S. K., Khennas, S., Can, S. de la R. du, … Minx, J. (2021). A review of trends and drivers of greenhouse gas emissions by sector from 1990 to 2018. Environmental Research Letters, 16(7), 073005. https://doi. org/10.1088/1748-9326/abee4e
Levitus, S., Antonov, J. I., Boyer, T. P., Baranova, O. K., Garcia, H. E., Locarnini, R. A., Mishonov, A. V., Reagan, J. R., Seidov, D., Yarosh, E. S., & Zweng, M. M. (2012). World ocean heat content and thermosteric sea level change (0–2000 m), 1955–2010. Geophysical Research Letters, 39(10). https://doi.org/10.1029/2012GL051106
Lynas, M., Houlton, B. Z., & Perry, S. (2021). Greater than 99% consensus on human caused climate change in the peer-reviewed scienti c literature. Environmental Research Letters, 16(11), 114005. https://doi.org/10.1088/17489326/ac2966
Lynch, J. (2019). Availability of disaggregated greenhouse gas emissions from beef cattle production: A systematic review. Environmental Impact Assessment Review, 76, 69–78. https:// doi.org/10.1016/j.eiar.2019.02.003
Mackie, S., Smith, I. J., Ridley, J. K., Stevens, D. P., & Langhorne, P. J. (2020). Climate Response to Increasing Antarctic Iceberg and Ice Shelf Melt. Journal of Climate, 33(20), 8917–8938. https:// doi.org/10.1175/JCLI-D-19-0881.1
MIMURA, N. (2013). Sea-level rise caused by climate change and its implications for society. Proceedings of the Japan Academy. Series B,
Physical and Biological Sciences, 89(7), 281–301. https://doi.org/10.2183/pjab.89.281
Moorman, R., Morrison, A. K., & Hogg, A. M. (2020). ermal Responses to Antarctic Ice Shelf Melt in an Eddy-Rich Global Ocean–Sea Ice Model. Journal of Climate, 33(15), 6599–6620. https://doi.org/10.1175/JCLI-D-19-0846.1
Morrissey, S. A., & Reser, J. P. (2007). Natural disasters, climate change and mental health considerations for rural Australia. Australian Journal of Rural Health, 15(2), 120–125. https:// doi.org/10.1111/j.1440-1584.2007.00865.x
Moser, C. O. N. (Ed.). (2007). Reducing global poverty: e case for asset accumulation. Brookings Institution Press.
Nicholls, R. J., Marinova, N., Lowe, J. A., Brown, S., Vellinga, P., de Gusmão, D., Hinkel, J., & Tol, R. S. J. (2011). Sea-level rise and its possible impacts given a ‘beyond 4°C world’ in the twenty- rst century. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 369(1934), 161–181. https://doi.org/10.1098/rsta.2010.0291
Nkedianye, D., de Leeuw, J., Ogutu, J. O., Said, M. Y., Saidimu, T. L., Kifugo, S. C., Kaelo, D. S., & Reid, R. S. (2011). Mobility and livestock mortality in communally used pastoral areas: e impact of the 2005-2006 drought on livestock mortality in Maasailand. Pastoralism: Research, Policy and Practice, 1(1), 17. https://doi.org/10.1186/20417136-1-17
Parkinson, A. J., & Evengård, B. (2009). Climate change, its impact on human health in the Arctic and the public health response to threats of emerging infectious diseases. Global Health Action, 2(1), 2075. https://doi.org/10.3402/gha. v2i0.2075
Perera, F. (2018). Pollution from Fossil-Fuel Combustion is the Leading Environmental reat to Global Pediatric Health and Equity: Solutions Exist. International Journal of Environmental Research and Public Health, 15(1), 16. https:// doi.org/10.3390/ijerph15010016
Rignot, E., Xu, Y., Menemenlis, D., Mouginot, J., Scheuchl, B., Li, X., Morlighem, M., Seroussi, H., van den Broeke, M., Fenty, I., Cai, C., An, L., & de Fleurian, B. (2016). Modeling of oceaninduced ice melt rates of ve west Greenland glaciers over the past two decades. Geophysical Research Letters, 43(12), 6374–6382. https://doi. Rocklöv, J., & Dubrow, R. (2020). Climate change: An enduring challenge for vector-borne disease prevention and control. Nature Immunology, 21(5), 479–483. https://doi.org/10.1038/s41590020-0648-y
Rossati, A. (2016). Global Warming and Its Health Impact. e International Journal of Occupational and Environmental Medicine, 8(1), 7–20. https://doi.org/10.15171/ijoem.2017.963
Rudolph, L., & Gould, S. (2015). Climate Change and Health Inequities: A Framework for Action. Annals of Global Health, 81(3), 432. https://doi. org/10.1016/j.aogh.2015.06.003
S. Nazrul Islam, & Winkel, J. (2017). Climate Change and Social Inequality. United Nations Department of Economic & Social A airs. https:// www.un.org/development/desa/publications/ working-paper/wp152
Sauerborn, R., & Ebi, K. (2012). Climate change and natural disasters – integrating science and practice to protect health. Global Health Action, 5(1), 19295. https://doi.org/10.3402/gha. v5i0.19295
SeaLevelRise.pdf. (n.d.). Retrieved November 29, 2021, from https://climatehealthconnect.org/wpcontent/uploads/2016/09/SeaLevelRise.pdf
Stella, E., Mari, L., Gabrieli, J., Barbante, C., & Bertuzzo, E. (2020). Permafrost dynamics and the risk of anthrax transmission: A modelling study. Scienti c Reports, 10(1), 16460. https:// doi.org/10.1038/s41598-020-72440-6
Taherkhani, M., Vitousek, S., Barnard, P. L., Frazer, N., Anderson, T. R., & Fletcher, C. H. (2020). Sea-level rise exponentially increases coastal ood frequency. Scienti c Reports, 10(1), 6466. https://doi.org/10.1038/s41598-02062188-4
e Lancet Planetary Health. (2018). Environmental racism: Time to tackle social injustice. e Lancet Planetary Health, 2(11), e462. https://doi.org/10.1016/S25425196(18)30219-5
United Nations Development Programme. (2007). Deglaciation in the Andean Region— Fighting climate change: Human solidarity in a divided world. In United Nations Development Programme, Human Development Report
2007/2008 (pp. 1–18). Palgrave Macmillan UK. https://doi.org/10.1057/9780230598508_1
Vector-borne diseases. (n.d.). Retrieved October 17, 2021, from https://www.who.int/news-room/ fact-sheets/detail/vector-borne-diseases
Walker, J. (2018a). e in uence of climate change on waterborne disease and Legionella: A review. Perspectives in Public Health, 138(5), 282–286. https://doi.org/10.1177/1757913918791198
Walker, J. (2018b). e in uence of climate change on waterborne disease and Legionella: A review. Perspectives in Public Health, 138(5), 282–286. https://doi.org/10.1177/1757913918791198
Wang, M., Su, J., Landy, J., Leppäranta, M., & Guan, L. (2020). A New Algorithm for Sea Ice Melt Pond Fraction Estimation From High-Resolution Optical Satellite Imagery. Journal of Geophysical Research: Oceans, 125(10), e2019JC015716. https://doi.org/10.1029/2019JC015716
Ward, R. D., Friess, D. A., Day, R. H., & MacKenzie, R. A. (2016). Impacts of climate change on mangrove ecosystems: A region by region overview. Ecosystem Health and Sustainability, 2(4), e01211. https://doi.org/10.1002/ehs2.1211
Webster, P. J. (2008). Myanmar’s deadly da odil. Nature Geoscience, 1(8), 488–490. https://doi. org/10.1038/ngeo257
World Resources Institute. (1997). Aridity zones and dryland populations: An assessment of population levels in the world’s drylands. United Nations Development Programme. https:// digitallibrary.un.org/record/432312?ln=en
Yarzábal, L. A., Salazar, L. M. B., & Batista-García, R. A. (2021). Climate change, melting cryosphere and frozen pathogens: Should we worry…? Environmental Sustainability, 4(3), 489–501. https://doi.org/10.1007/s42398-021-00184-8
Yue, X.-L., & Gao, Q.-X. (2018). Contributions of natural systems and human activity to greenhouse gas emissions. Advances in Climate Change Research, 9(4), 243–252. https://doi. org/10.1016/j.accre.2018.12.003

History and Current State of Virology
STAFF WRITERS: LORD CHARITE IGIRIMBABAZI '24, CAMERON SABET '24, VAISHNAVI KATRAGADDA '24, MATTHEW LUTCHKO '23, DECLAN O’SCANNLAIN '24, JUSTIN CHONG '24, CAMILLA LEE '22, CAROLINE CONWAY '24, SOPHIE (SOYEON) CHO '24, FRANK CARR '22, JULIAN FRANCO JR. '24, DANIELA ARMELLA '24, MIRANDA YU '24, BROOKLYN SCHROEDER '22, ABIGAIL FISCHER '23, VALENTINA FERNANDEZ '24, CALLIE MOODY '24, ANYOKO SEWAVI '23, LAUREN FERRIDGE '23, VAANI GUPTA '24
TEAM LEADS: NISHI JAIN '21, CAROLINA GUERRERO '23
Cover Image: COVID-19 is potentially the most signi cant viral disease of the 21st century. Image Source: Pixabay Introduction
Virus Overview As the world becomes increasingly connected, the threat of infectious diseases, especially those caused by viruses, becomes more prevalent. Namely, the COVID-19 pandemic has shed light on the importance of studying both how viruses work to cause disease and how researchers can devise therapeutic solutions. However, viruses are not only agents of disease – they can also be used as therapeutics themselves as will be explored later in this paper. As a whole, devoting research toward viral mechanisms of action and understanding past pandemics is critical to future prevention.
Viruses are microscopic parasites that cannot survive nor reproduce without a host cell. ey are quite a bit smaller than bacteria, with the measles virus being around ⅛ the size of Escherichia coli (E. coli) bacteria. In one striking study at Davidson College, Dr. David R. Wessner (2010) found that the polio virus is around 10,000 times smaller than a single grain of salt (~30 nm across), demonstrating how incredibly small viruses can be.
Experts debate whether viruses are alive. On one hand, viruses possess nucleic acids like deoxyribonucleic acids (double-stranded) or ribonucleic acids (single-stranded), just like living cells. Both deoxyribonucleic acid (DNA) and ribonucleic acid (RNA) are found in living organisms, like humans. On the other hand, they cannot read this information on their own and require a host organism to replicate. Since viruses require a host, this parasitic biochemical machine might appear to be just that—a machine. Additionally, viral genomes are incredibly small. ey only encode enough amino acids to produce enzymes required for entry and replication within host cells and a capsid, which is the outer protein

shell protecting the virus (Villarreal, 2008). By comparison, the human genome is comprised of about 3.2 billion nucleotides and contains all of the information that the body needs to sustain itself (Brown, 2002).
Mechanisms of Entry and Replication
Viruses may enter the body via numerous pathways, including respiratory passages or open wounds. Some viruses even stay dormant in an insect’s saliva and infect immediately a er the insect bites an animal. en, like puzzle pieces, viruses bind to the host cell’s surface receptors (proteins unique to a certain cell). A er this, there are a few di erent ways the virus can enter; this entry is o en determined by if a virus has an envelope or not; viral envelopes consist of a phospholipid bilayer and membrane proteins that are derived from the host cell, and serve to promote fusion with the host cell For instance, HIV already has an envelope, so it can just fuse with the cell membrane and gain entry. e in uenza virus also has an envelope, and the cell engulfs it. However, when a given virus is nonenveloped, like the polio virus, that virus can cut through the membrane by creating a porous channel since the membrane is le exposed. en, the viruses can spew out their genetic materials and disrupt cellular processes to produce viral proteins. is o en tampers with the host cell’s ability to produce its own proteins or RNA, o en leading to host cell death. Meanwhile, the virus makes cellular conditions favorable for further infection and reproduction. For example, it has been estimated that a single sneeze from someone infected with the novel coronavirus contains ~20,000 droplets of viral particles. In many cases, merely breathing in such particles is enough to prompt further infection, demonstrating the power of virus’ abilities to not only acclimate to di erent hosts, but also to capitalize upon certain behaviors to continue reproducing (Villarreal, 2008).
Mechanisms Treatment and Prevention of Viral Infections
Existing Antiviral Drugs and How ey Work
Some viral infections can be dangerous, so devising treatments is of great signi cance to the medical research community. Beyond simply treating symptoms of infections, there are two broad classes of drugs (di ering in whether they are immunologically derived or chemically synthesized) used to combat viruses. Firstly, there are treatments involving the infusion of monoclonal antibodies (i.e., many antibodies with the same speci city and structure). ese o en bind to free virions, or virus particles that have yet to infect a host cell, preventing them from entering cells as well as targeting them for destruction by one’s immune system. Before the development of these treatments, the use of animal plasma/antibodies was similar, but the foreign nature of these antibodies meant an inevitable immune response against the treatment. So, the quest to develop “humanized” antibodies that would not elicit an immune response began (Riechmann et al., 1988). e rst development of such antibodies was performed
Figure 1: e structure of a virion. Image Source: Wikimedia Commons
in 1973 by creating hybrid cells of cancerous antibody-producing cells from mice and human immune cells (Schwaber & Cohen, 1973); these cells are biologically immortal, and once they develop their antibody speci city can produce seemingly endless amounts of the proteins. Since this breakthrough, new treatments have applied this technology to cancers, autoimmune disorders, and viral infections.
One of the most recent and well-documented uses of this kind of therapy is REGN-COV2 (also called “Regeneron,” a er the company that developed it), which consists of a solution of two di erent antibodies, each having speci city for a di erent part of the SARS-CoV-2 Spike Protein (Weinreich et al., 2020). Other monoclonal antibodies used to treat viral infections include Palivizumab and, more recently, Suptavumab ( e Impact-RSV Study group, 1998; Simões et al., 2020); both are used to treat and prevent infections caused by Respiratory Syncytial Virus (RSV) by binding to its F-Protein, a viral membrane protein used for fusion with target host cells. Bavituximab targets the membrane phospholipid Phosphatidylserine that is present on the external face of human cell membranes only if the cell is cancerous and/or it is infected by Hepatitis C. While this antibody does not directly bind to free Hepatitis C virions, it causes the phagocytosis and immunological destruction of viral-infected cells (Ahn & Flamm, 2011).
Alternatively, antiviral medicines are o en taken as pills or oral solutions and are generally small molecules developed in a lab setting. ese medicines have diverse mechanisms, depending on both the drug and its target. Like the monoclonal antibodies discussed above, many antiviral medicines are speci c to a single virus or a family of viruses. ere are a few drugs, however, that are capable of targeting multiple types of viruses by targeting general structures–those that can are called broad-spectrum antivirals (Vardanyan & Hruby, 2016).
An example of antiviral drugs is the in uenza neuraminidase inhibitors, such as Tami u, which prevent the action of the neuraminidase enzyme, preventing the release of progeny virions. Another class of drugs used to combat the in uenza virus are the adamantanes, which inhibit an ion channel in the In uenza A viral membrane that is necessary for the release of the virus’s genetic payload a er it has entered a cell (Ison, 2011). An example of a broad-spectrum antiviral is Remdesivir. Initially developed to treat Hepatitis C infections, it has proven to be an e ective treatment of many viruses including the Ebola Virus, coronaviruses (including SARSCoV-2), paramyxoviruses (such as the Measles and Mumps viruses), and RSV. Notably, it ended up not being e ective in treating Hepatitis C. It acts by inhibiting the viral RNA polymerase, which is a general structure among many viruses, preventing them from reproducing their genomes (Aleem & Kothadia, 2021).
ese examples showcase the overall trend in antiviral medicine: therapies are designed to target a speci c part of a virus (and sometimes that part is shared by many viruses, resulting in broad-spectrum drugs). us, as further research is done on viruses and new viruses are discovered, new therapies, be they antibodies or small molecules, can be developed, and their usefulness against viruses beyond their initial targets can be determined.
Existing Vaccines and How they Work
Unlike antiviral therapies that treat viral infections a er they occur, vaccines work by preventing them from taking place. Vaccines can prevent a range of the most damaging viral infections, including smallpox, in uenza, measles, Hepatitis B, and HPV (Graham, 2013). e rst vaccine developed was Edward Jenner’s 1796 smallpox vaccine. His vaccine and later ones have played a crucial role in containing and preventing outbreaks. For instance, before the measles vaccine was licensed in 1963, almost all children were infected by the age of 15; measles infections have dropped over 98% since then (Ravanfar et al., 2009).
ese vaccines all share the ability to induce a natural immune response against the viral pathogen, which then grants immunity to the vaccinated individual (Ellebedy & Ahmed, 2016). e earliest antiviral vaccines were derived from live animals or eggs, while recent ones have been created through cell culturing and other advanced molecular biology techniques (Graham, 2013). A er identifying the genetic sequence of a virus and the structure of its surface proteins, researchers develop a vaccine that mimics it.
Historically, vaccines have come in ve main types. Live viral vaccines contain an attenuated (weakened) version of a given virus, while inactivated whole viral vaccines are treated with heat or UV light to damage the virus. Subunit vaccines (containing the surface glycoproteins, which are proteins at the surface of viruses that help viruses enter cells), recombinant

viral proteins (manufactured to contain main surface glycoproteins), and virus-like particles (assembled from viral structural proteins) all contain just parts of the virus. Each method has its advantages and disadvantages. For example, inactivated whole viral forms are one of the most e ective methods but involve safety concerns if not completely inactivated, (Ellebedy & Ahmed, 2016).
ough vaccines are one of the most e cient ways to prevent viral infections, there remain viruses for which no vaccines exist. In 2019, COVID-19 was among them. e process of developing and implementing a vaccine typically takes place over many years, involving vaccine design, testing, and manufacturing on a large scale (Zhou et al., 2020). is lengthy process o en makes it di cult for vaccines to be created in time to stop an epidemic. In the case of COVID-19, especially because of the devastating e ects of the pandemic, scientists were successful in speeding up the steps to generate vaccines quickly (Zhou et al., 2020). Because of the signi cant bene ts conferred by antiviral vaccines, researchers continue to work on new vaccines while improving existing ones.
Viruses, Human Behavior, and the Environment
How do diverse zoonotic viruses develop and how do they jump to humans?
Zoonotic viruses are viruses that can jump from animals to humans via a number of methods, including direct and indirect contact (i.e. bodily uids and contact with an infected animal’s habitat), vectors (i.e. ticks), food, and water (Zoonoses, 2020).
Many recent pandemics have originated in animals, and zoonotic diseases have a high chance of causing the next pandemics due to the ability of zoonoses, or animal-borne diseases, to cause interspecies transmission (Zoonoses, 2020). Of the total emerging infectious diseases (EID), 60.3% have been caused by zoonoses (Zoonoses, 2020). e origins of EIDs are correlated with socioeconomic, environmental, and ecological factors that predict where they are likely to originate. is explains why in regions where disease reporting e orts and resources are low, a greater risk of zoonotic and vector-borne viruses exists (Zoonoses, 2020). From an analysis of viral discovery data, Carroll et. al. (2018) estimate that about 1.67 million undiscovered viruses with zoonotic origins are present in bird and mammal hosts. Additionally, Carroll et al. (2018) expect that 631,000 - 827,000 of these not-yet-discovered viruses have the potential for zoonotic transmission based on analysis of viral-host relationships, the history of zoonotic viruses, and patterns of viral emergence.
e COVID-19 pandemic has stressed the urgency of better understanding zoonotic spillover, the process by which viruses jump from animals to humans. ere are many obstacles and factors relating to ecology, virology, evolution, and human immunity that prevent zoonotic
Figure 2: A digital representation of a generic in uenza virus. Image Source: Flickr
Figure 3: Wuhan, China, is the starting point of the COVID-19 pandemic. Image Source: Wikimedia Commons

spillover, making the mechanism an enigma to scientists. For instance, the disease must unlock receptors on the cell surfaces of the target hosts and must learn to replicate itself without signaling the host’s immune system (Singer et al., 2021). Viruses that manage to make the jump are rare already, and when they are successful, most spillover events do not trigger large-scale outbreaks (Singer et al., 2021).
To make the jump, the virus must somehow be equipped to infect the new host before even coming into contact with said host. Coronavirus research indicates that the current host pressures the virus into mutations that will allow it to infect other hosts that it has not had contact with yet (Singer et al., 2021). In regard to the recent SARS-CoV-1 and SARS-CoV-2 outbreaks, spike proteins unlock ACE2 receptors (which are expressed in various human cells) of new host cells, explaining how bat coronaviruses can infect human cells (Petrosillo et al., 2020).
Plowright et al. (2017) propose a conceptual and quantitative framework in which data has been integrated to address gaps in research on barriers and determinants of zoonotic spillover. is framework can be grouped into three functional phases that describe the major routes of transmission. In phase 1, pathogen pressure (the amount of pathogen available to the human host at any point in time and space) depends on interactions among reservoir host distribution, pathogen prevalence, and release from the reservoir host. ese factors are followed by pathogen survival and the development and dissemination of the pathogen a er leaving the reservoir host. e second phase is determined by how human and vector behavior a ect pathogen exposure. In the third phase, the probability and severity of infection are determined by the genetic, physiological, and immunological components of the human host as well as the dose and route of exposure of the pathogen (Plowright et al., 2017).
Epidemics and Pandemics
Overview of Epidemics and Pandemics and How ey Start
Before COVID-19, a public understanding of pandemics and epidemics existed but was simply too general and incomprehensive. Most knew them to be the spread of an atrocious, natural phenomenon that could wipe out entire populations due to a lack of immunity. In uenza, cholera, malaria, and the u, for example, struck us as examples of epidemics and pandemics that took place decades ago but would never strike us again. But today, as we nd ourselves amidst one of the worst pandemics that mankind has ever seen, we have been forced to understand how epidemics begin and work, and consequently, how they can escalate into a pandemic.
Epidemics occur at a local level; they are the widespread infection of a disease within a
particular community at a particular time and continue to spread as the disease travels into larger geographical scope (Miller, 2014). For example, think of Wuhan, China back in December 2019. SARS-CoV-2, the classi ed virus name for COVID-19, spread in the city of Wuhan and its communities at a rapid rate, culminating in the infection of thousands of people within a matter of weeks. An epidemic consists of a local outbreak, does not have a predictable occurrence rate, and spreads to larger geographical areas (Miller, 2014).
Pandemics are a consequence of epidemics; they are the global spread of a disease that begins locally and, in turn, a ect a very large percentage of the global population (What is a Pandemic?, 2021). Taking COVID-19 as a prime example, SARS-CoV-2 began locally in Wuhan, China and began to spread across Asia and Europe as a result of travel and mobilization, eventually reaching North and South America. e spread of an epidemic depends on 1) how easily the disease is transmitted from one individual to another and 2) the movement of those who carry it (What is a Pandemic?, 2021). For the COVID-19 infection, traveling via airplane made the transmission of the virus exponentially faster; within a matter of hours, the virus had reached another continent and infected hundreds of people along the way. Further characterization of pandemics includes 1) prone age group target of the disease, 2) self-limited illness and recovery of infected individuals, 3) fatality and death count, and 4) seasonal in uenza. e 1918 Spanish In uenza and the bubonic plague of the 14th century serve as examples of world-wide pandemics in human history (What is a Pandemic?, 2021).
COVID-19/SARS/MERS
Coronaviruses (CoV) have been around for the past few decades, o en causing respiratory tract infections, with the viruses rst being discovered in the 1960s. Especially in the last few years, these viruses have exponentially grown in threat and prevalence, with a Severe Acute Respiratory System (SARS-CoV) virus epidemic in 20022003 and the Middle East Respiratory Syndrome (MERS-CoV) epidemic in 2012. In 2019, a novel coronavirus emerged in Wuhan, China, initially referred to as SARS-CoV-2 due to its similarity to SARS. e clinical presentation of this virus is known as the COVID-19 disease. ese RNA viruses have some of the largest genomes and are o en passed to humans through animal intermediates. Once in humans, the disease spreads rapidly through contact and through airborne droplets from sneezing, coughing, or breathing. CoV belong to the order Nidovirales and under the subfamily Coronavirinae, with a few major characteristics: large genomes, high replication rates, unique enzymatic activity, and extensive ribosomal frameshi ing (a process used by viruses to have many proteins encoded by one piece of mRNA) due to a variety of nonstructural genes that are encoded in the RNA (Umakanthan et al., 2019).
CoV are typically enveloped and contain positive single-stranded RNAs of about 8.4-12 kDa in size. e 5’ end of the genome contains the majority of information necessary for viral replication while the 3’ end contains ve important structural proteins: spike protein, membrane protein, nucleocapsid protein, envelope protein, and haemagglutinin-esterase protein. Each of these proteins plays an integral role in the CoV’s life. e spike protein is necessary in attaching and fusing the envelopes to the host cell, the membrane protein de nes the envelope shape, the nucleocapsid contains RNA complexes important for RNA transcription and assembly, the envelope protein constructs the envelop, and the haemagglutinin-esterase is important for receptor binding on host cells (Umakanthan et al., 2019). e spike glycoproteins present on CoV are especially essential in promoting entry of the virus into host target cells (Samudrala et al., 2020).
While SARS, MERS, and SARS-CoV-2 all fall under the category of CoV, each virus presents with slightly di erent characteristics. SARS was rst recognized in Guangdong, China, eventually leading to the virus spreading across 30 countries and a ecting 79,000 people total with a 9.5% fatality. e virus was traced back to Himalayan palm civets from a livestock market, con rming that CoV viruses are indeed zoonotic in origin. MERS presents with pneumonia-like features as well as renal failure. While this epidemic was much smaller, with a total of 91 infected patients, the disease was much more fatal at 34% fatality. is virus was identi ed in bats and Arabian dromedary camels, as well as in goats, sheep, and cows, which acted as intermediate host disease reservoirs before passing the virus on to humans (Umakanthan et al., 2019).
SARS-CoV-2 presents with many similar clinical features but a much smaller fatality rate of 2.3%, as well as a less severe clinical presentation. MERS was much more fatal, with more patients developing acute respiratory distress syndrome (ARDS), potentially because MERS binds to a "While SARS, MERS, and SARS-CoV-2 all fall under the category of CoV, each virus presents with slightly di erent characteristics."
Figure 4: A transmission electron image of the novel SARS-CoV-2 virus.
Image Source: Wikimedia Commons

di erent receptor than SARS and SARS-CoV-2. While MERS binds to dipeptidyl peptidase 4 (DPP4) receptors, SARS and the novel CoV bind to angiotensin converting enzyme (ACE) receptors. Other clinical features are very similar, such as low platelet count and decreased albumin levels. However, the disease reproductive number (which estimates the number of new cases that can be directly related to one original case), was estimated to be 2.0-2.5 for SARS-CoV2????, much higher than MERS (<1) and slightly higher than SARS (1.7-1.9). SARS-CoV-2 and SARSCoV are more closely related than SARS-CoV-2 and MERS-CoV. While the route of transmission has been assumed to be through airborne droplets or contact, it’s also possible that there is a gastrointestinal route of transmission, based on the hypothesis that other CoVs like MERS could have spread through drinking the milk of infected camels (Petrosillo et al., 2020).
Studies speci c to COVID-19 in patients have found that the median age of infected patients is around 56 years, with males more a ected than females due to a higher concentration of angiotensin enzyme 2 (Umakanthan et al., 2019). In many cases, the disease correlates highly with previous factors of susceptibility, such as smoking, hypertension, diabetes, or other similar health conditions. Symptoms o en include milder conditions with nonspeci c symptoms, such as a fever, cough, myalgia, throat ache, or nausea. However, the more dangerous pneumonia classi cation of the virus presents with severe disease, with dyspnoea and dangerously low blood-oxygen saturation level (Umakanthan et al., 2019).
Despite the similarities to SARS-CoV and MERS-CoV, there are several di erences with the novel SARS-CoV-2 (Umakanthan et al., 2019). Preliminary studies of SARS-CoV-2 suggest that signi cant mutations present on the virus membrane proteins and receptor binding sites lead to its extreme transmission and pathogenicity. e receptor-binding domain (RBD) contains many of these mutations. Based on genomics understanding of SARS-CoV, a series of six amino acids on the spike protein have been identi ed as crucial for binding to the ACE2 receptor. However, SARS-CoV-2 presents with ve unique amino acids in those places, causing an abnormally high a nity for the ACE2 receptors. While the pathogenesis of SARS-CoV-2 is not entirely understood yet, the many similarities between the virus and SARSCoV as well as MERS-CoV provide a basis for a likely hypothesis. As ACE2 receptors are typically present in the lungs (speci cally in type-2 pneumocytes), the virus binds to many of these cells, causing the subsequent downregulation of ACE2 receptors. at downregulation leads to an increase in angiotensin-2 through ACE1 (an enzyme). is could then potentially lead to pulmonary vascular permeability and lung injury. As the body attempts to combat this through an immune response, in ammatory cytokines and chemokines are released, which o en lead to more damage (Samudrala et al., 2020).

While the zoonotic origin of SARS-CoV-2 is expected to be through bats, past experience with CoV has shown that the disease o en jumps to an animal intermediate before moving to humans. Based on genomic and evolutionary evidence of SARS-CoV-2-like CoV in pangolins (a type of scaly mammal), the disease is hypothesized to have transmitted from bats to pangolins and then to humans. e Pangolin-CoV virus found is 91.02% identical to SARS-CoV-2 and 90.55% identical to Bat-CoV (RaTG14), suggesting that this is a possibility. e spike protein encoded by the Pangolin-CoV is very closely related to SARSCoV-2 as well. Additionally, 5 key amino acids necessary for the novel virus binding to human ACE2 receptors is consistent with the PangolinCoV whereas the Bat-CoV only shares four of those mutations (Zhang et al., 2020).
Prior to the development of the vaccine, no appropriate medicine or treatment had been FDA-approved. While the race for a vaccine continued in 2020, the e cacy of ve di erent FDA-approved drugs were studied. Cytotoxicity and drug e ect was studied in vitro, with some treatments eventually progressing to phase III clinical trials. Remdesivir is one such treatment that was found to signi cantly reduce the mortality rate of COVID-19 upon 14 days of treatment and showed improvement in 64% of cases. Other treatments, such as chloroquine, showed promise but could lead to toxicity with overdose, posing complications. Protease inhibitors were studied as well, but unsuccessfully, as their e cacy was inconclusive (Samudrala et al., 2020). However, late in 2020, the rst vaccines were announced and approved by the FDA for emergency use. e e cacy of mRNA vaccines has been an increasingly valuable area of research in the past few decades, especially in combating cancer. However, the emphasis on vaccine development was mostly on DNAbased approaches up until the late-2000s due to the many di culties in working with RNA (Pardi et al., 2020). Designed to be a temporary molecule, RNA is very unstable; furthermore, it can elicit unnecessarily excessive immune responses, making it di cult to deliver safely in patients. However, recent advancements in optimizing RNA stability and in uencing the rate of translation as well as the half-life of the transcript by modifying the 5’ and 3’ untranslated regions improve the safety and stability of RNA. Additionally, large-scale RNA puri cation methods have been developed, allowing vaccines to be cheaper and more resource-e ective. E cient delivery molecules have also been recently designed, using substances that protect the mRNA as well as the patient. Polymers, peptides, and lipid nanoparticles are being studied further as potential delivery agents for more e ective mRNA vaccines. Since many of these technologies are relatively novel, as is the use of mRNA vaccines, much more research needs to be done (Pardi et al., 2020).
1918 Flu Pandemic
e 1918 Spanish u epidemic is one of the most
Figure 5: A model representation of the chemical structure of Remdesivir.
Image Source: Wikimedia Commons
infamous pandemics in history. It is estimated to have killed 50-100 million people in total, including approximately 675,000 individuals in the United States. In the absence of the technology necessary to identify viruses, some scienti c communities thought the bacterium Haemophilus in uenzae was responsible for the disease referred to as the “Spanish u.” However, the true culprit was a virus belonging to the in uenza A (H1N1) subtype (Wever & van Bergen, 2014).
ough society has now identi ed the virus responsible, the origin of the 1918 in uenza virus remains something of a mystery; there is no consensus on where the virus rst appeared in a human population. However, the sequencing of the virus’s eight-segment genome has provided more insight into its development. Some scientists initially assumed that the 1918 in uenza virus resulted from gene reassortment between human and animal viruses, but more recent evidence casts doubt on this hypothesis. Because all eight of the sequenced gene segments are avian-like and waterfowl enteric tracts are known reservoirs of in uenza A viruses, an updated hypothesis posits that the 1918 in uenza virus resulted from the adaptation of an existing avian virus to a human host (Morens et al., 2007).
It is impossible to fully discuss the 1918 in uenza pandemic without acknowledging the context of the First World War. In Europe and in American training camps, the 1918 pandemic killed an estimated 45,000 American soldiers in total. According to some sources, the Spanish u led to more American burials in France than the war itself. e disease infected over one million men in the United States Army, 700,000 men in the German Army, and 313,000 men in the British Expeditionary Forces. e rst substantial outbreak of the 1918 in uenza pandemic has been traced to a Kansas military camp called Camp Funston (Wever & van Bergen, 2014). e First World War likely contributed to the spread and severity of the Spanish u, as overcrowding has been associated with a ten times higher risk of the u and an increased severity of the disease. Overcrowding was such a concern in the U.S. Army that in January 1918 (before the pandemic), the Army Surgeon General William Gorgas testi ed before the Senate that U.S. troops required more oor space. e overcrowding and movement of soldiers during the First World War likely contributed to the 1918 pandemic (Aligne, 2016).
Overall, the 1918 pandemic had three distinct waves and stood out from other in uenza outbreaks in its mortality patterns. While in uenza strains tend to be most dangerous for the very young and very old individuals within a population, the 1918 u hit those of middle age (20-50 years) hard as well, resulting in an atypical w-shaped mortality curve. e pandemic’s mortality peaked in October 1918 in the U.S. and Italy, and most victims died of pneumonia or other respiratory system complications (Gavrilova & Gavrilov, 2020). ough relegated to the pages of history textbooks today, the infamous Spanish u lives on, as ve of the genes in the common (H3N2) in uenza virus originated from the 1918 pandemic (Belshe, 2009).
Ebola
Ebola virus disease (EVD) is a severe, o en lethal, infection caused by a zoonotic virus which is a member of the loviruses and causes an acute hemorrhagic fever (Jacob et al., 2020). EVD epidemics typically start by a single case of probable zoonotic transmission (wildlife to human) followed by human-to-human transmission (Groseth et al., 2007). e rst recorded Ebola human outbreak took place in 1976 in Sudan, when an individual came into contact with the blood of a guinea pig infected by the Ebola virus (EBOV) (Emond RT et al., 1977). e virus simultaneously spread to Zaire, which is now called the Democratic Republic of Congo. During the outbreaks, 284 cases and 318 cases were con rmed in Sudan and Zaire respectively. e EBOV Sudan strain (SEBOV) had a Case Fatality Rate (CFR) of 53% and the Zaire strain (ZEBOV) had a high CFR of 89% (Groseth et al., 2007). Subsequent outbreaks of EBOV showed the successful replication of the virus by revealing the presence of other EBOV strains. e Reston Ebola virus was genetically discovered in 1989 in the United States of America from Macaques imported from the Philippines; however, it was proven non-infectious to humans (Miranda et al., 1999). In 1994, scientists discovered the Ivory Coast Ebola virus (ICEBOV) and another strain was found in Bundibugyo, Uganda (BEBOV) in 2007 and has a CFR of 26% (Muyembe-Tamfum et al., 2012). From all the EBOV species, ZEBOV was the most common and highly pathogenic with 30706 con rmed cases as of 2021 with an average lethality rate between 25% and 90% (Ebola Virus Disease, 2021). All known sources of EBOV human infection have involved contact with dead or butchered wildlife such as apes and chimpanzees, or the exploration of natural sites that house bats (Groseth et al., 2007; Emond RT et al., 1977).
Ebola virus disease, like other lovirus infections, is contracted from direct contact with bodily uids of infected humans or animals (dead or alive) and makes its way into the body through the skin and mucous membranes (Beeching et al., 2014). It takes about 2-21 days with an average of 7 days before the onset of rst symptoms a er infection. Early stages of the disease are non-speci c. First symptoms may include acute fever, headache, fatigue, vomiting, and muscle pain (Beeching et al., 2014). e late stages of the disease o en include multi-organ dysfunction, which is the major cause of death of patients infected by EBOV. e virus has a CFR ranging between 30% and 90% of all con rmed cases (Ebola Virus Disease, 2021).
Given the high mortality rate of patients with EVD, the World Health Organization declared the Ebola epidemics as Public Health emergencies to prevent the virus from disseminating across the globe and to help scientists develop an understanding of the mechanisms of the Ebola virus (which could be used as a potential biological weapon). Researchers have attempted to identify the reservoir species or natural hosts of EBOV—i.e., animals that can carry the virus without exhibiting clinical symptoms. Di erent studies suggest that bats are putative reservoir hosts of EBOV, because the virus was successfully isolated from multiple bats species with no clinical signs of the illness (Pourrut et al., 2005). Despite this major discovery, researchers have not yet con rmed or determined the natural reservoir host of the virus; further research needs to be conducted to con rm the theory. Studies to determine the virus reservoirs are important to understand how the virus emerged and to develop risk-reduction measures that will prevent future EBOV outbreaks.
Another scienti c quest was to uncover the viral genetic footprints that might help explain the virulence of the ZEBOV. e severity of the virus is largely understood through its remarkable ability to interfere with the immune response of the host (Wang et al., 2019). It inhibits the expression of genes involved in the innate immune response against viral infections, such as IRF3 (Interferon Regulatory Factor 3) gene—which is an important transcriptional factor for the induction of early antiviral immunity (Wawina-Bokalanga et al., 2019). EBOV is a negative-strand (antisense strand) RNA virus which encodes for eight viral proteins, including four structural proteins: VP35 (viral protein 35), VP40, VP24, NP (nucleoprotein) (Wawina-Bokalanga et al., 2019). ese four viral proteins are produced by the virus as it hijacks the cellular machinery of the host cell, and each protein plays an important role in the pathogenic mechanisms of the virus. VP35 is crucial for the EBOV interferon antagonism (signaling protein produced and released by the host cell in response to viral infections) (Hartman et al., 2008). VP35 inhibits the activation of IRF3 gene by impairing or blocking IRF3 phosphorylation—an important biochemical process that regulates protein function and signal transmitting throughout the cell (Hartman et al., 2008). Without IRF 3 and its dependent genes to initiate innate immunity against the virus, the Ebola virus replicates drastically (Hartman et al., 2008). is uncontrollable viral proliferation prompts an overload of virions harbored by the host that causes multiorgan failure or dysfunction and cellular death or apoptosis, which lead to clinical complications or death in EVD patients. Scientists have found that the carboxyl-terminus or carboxyl-group of VP35 protein is responsible for its immunosuppressive capacity, as this region of the protein can physically inhibit IRF3 activation (Hartman et al., 2008). erefore, environmentally or human induced mutations at a speci c position on the IRF3 inhibitory domain could decrease considerably the ability of VP35 to act as an interferon antagonist or to silence the IRF3 gene (Hartman et al., 2008).
Scientists have also been looking for environmental and biological patterns that could serve as the basis of the resurgence of the Ebola virus. Researchers have demonstrated that environmental changes or seasonal patterns contribute to the preservation of EBOV in nature (Groseth et al., 2007). One of the scenarios that explains the resurgence of EBOV is that the virus is asymptotically harbored by reservoir species and arises seasonally in nature depending on tting environmental conditions. Using geographical modeling and bioinformatics, Allison Groseth et al. (2007) found that ZEBOV, ICEBOV, and SEBOV occupy di erent geographical areas. ZEBOV and ICEBOV outbreaks happened in dry seasons whereas SEBOV outbreaks occurred during the seasonal periods of wetness. is observation coincides with the data from ZEBOV outbreaks in 1996 and 1997 in Gabon whereby a high number of dead great apes was recorded as a result of ZEBOV infection among the population between November and February—a period that marks the dry season in Gabon (Groseth et al., 2007). Another scenario that might explain the recurrence of EBOV in human populations could be the persistence of the virus in some bodily uids even a er complete clinical recovery and "Given the high mortality rate of patients with EVD, the World Health Organization declared the Ebola epidemics as Public Health emergencies to prevent the virus from disseminating across the globe and to help scientists develop an understanding of the mechanisms of the Ebola virus."
Figure 6: e Ebola virus. Image Source: Flickr

clearance in the blood (Dukobu et al., 2018). However, there is not yet evidence that could establish risk of transmission in convalescence.
Clinical advancements of EBOV
Many treatments and vaccines against Ebola virus are in development to reduce the severity of the disease and prevent future outbreaks. Certainly, the most remarkable advancement of EBOV research in 2019 is the development of a vaccine against the ZEBOV species—Recombinant Vesicular Stomatitis virus-Zaire Ebola Virus (rVSV-ZEBOV) (Ehrhardt et al., 2019; Metzger et al., 2018). However, the duration of protective e cacy of the vaccine is highly disputed.
Polio
Poliomyelitis, commonly referred to as the Polio virus, or for short, Polio, is historically known for its long-lasting presence and e ect on populations worldwide. While there is literature and artistic evidence of Polio’s presence as early as 1403BC, the rst clinical description of polio did not occur until the late 1700’s (Mehndiratta et al., 2014). It was in the 1900s that Polio became prevalent in the United States and was acknowledged for the rst time as an existing epidemic in the year 1916 by U.S. public health authorities (Mehndiratta et al., 2014). Due to the extremely early presence of Polio in human history, there is still no distinct origin to pinpoint how its transmission transpired (Lacey, 1949). However, what di erentiates the historical presence of Polio is the recurrence of an epidemic each summer (Mehndiratta et al., 2014).
e near eradication of Polio was due to the rapid study and vaccine development performed by both Jonas Stalk, who produced the rst Polio vaccination in 1955, and Albert Sabin, who developed another form of vaccination shortly a er in 1963 (De Jesus, 2007). e Salk method, recognized as the Inactivated Polio Vaccine (IPV), was given by injection and worked to stimulate serum IgM, IgG, IgA, but not secretory IgA: in such case, immunity had been induced by antibody transduction into the oropharynx (Howard, 2005). Sabin’s method, known as the Oral Polio Vaccine (OPV), replaced Stalk’s vaccination in 1963. is had been deemed advantageous over the IPV due to its cheaper cost, easier administration, and capability of causing an active infection of the oropharynx in addition to the intestinal endothelium, inducing a greater immune response (Howard, 2005).
In the United States, there are fewer than 10 Polio cases that occur annually, all of which are a result of back mutations (Melnick, 1996). However, despite successes in eradication in parts of the world, Polio is still endemic in six countries - Nigeria, India, Pakistan, Niger, Afghanistan, and Egypt (Howard, 2005). is presence is partially attributed to the di culty of providing a heat-stable oral vaccine to ensure su cient seroconversion in these tropical locations (Howard, 2005). Smallpox
Smallpox is a highly contagious disease which is caused by the variola virus. e variola virus comes in two forms: the more common, lifethreatening variola major with a mortality rate of 30% and the milder version- variola minor or alastrim (Geddes, 2006). Its origins are obscured due to its prevalence throughout early world history, but the World Health Organization (WHO) reported ndings of smallpox-related skin rashes on Egyptian mummies around 3000 years ago, suggesting that ancient Egypt could have been the earliest instance of this disease. is report attempted to nd the “original home” of
smallpox, but in practicality, smallpox could have developed anywhere with an irrigated agricultural society and larger populations (Geddes, 2006). By the end of the 1500s, smallpox’s notoriety rose as the virus was a signi cant cause of death in Europe, southwestern Asia, India, and China, and it went global during the age of exploration and colonization, reaching the Caribbean and Americas.
It has been speculated that the variola major and minor viruses evolved from a version of the virus that a ected animals. Variola viruses are a part of the poxviridae group of viruses that includes the vaccinia virus and animal poxviruses such as monkeypox (Geddes, 2006). Phylogeny evidence further solidi es this hypothesis: variola viruses were found to be descendants of an ancestral African rodent-borne variola-like virus as the variola virus and this rodent-a ecting virus are in a monophyletic group with one another. e time frame for this convergence is estimated to be between 16,000 and 68,000 years ago (Li et al., 2007).
In early 1000 A.D China, individuals were putting smallpox pus or scabs by nasal canals or through cuts, so that the body would be exposed to a weakened form of smallpox. is process, called variolation, was the earliest treatment to smallpox and foreshadowed what would be the rst invented vaccine in history. Edward Jenner, an English physician, noticed milkmaids who caught cowpox were resistant to smallpox. Edward removed material from a cowpox lesion on the hand of a milkmaid and inoculated it into a young boy. A er a few days of mild symptoms, the boy became immune to smallpox (Parrino & Graham, 2006). Over time and the eventual replacement of the cowpox with the vaccinia vaccine, this method by Jenner was widely acceptable.
is vaccination, although e ective, required

Figure 7: e Poliovirus. Image Source: Wikimedia Commons
Figure 8: An artistic rendering of a nanoparticle. Image Source: Flickr

booster shots, as its immunity wore o a er 3 years. A er some breakouts throughout the globe, smallpox cases dwindled down, and eventually its existence ceased a er its properformed worldwide and the last cases of major and minor variola was in Bangladesh in 1975 and Somalia in 1977 respectively (Parrino & Graham, 2006). Today, there are few people who are vaccinated for smallpox, but there are two known countries (USA and Russia) who o cially have stocks of the virus in laboratories. ey are used for study if a similar virus ever breaks out, but many have questioned why it still has not been destroyed.
Viruses as erapeutics
Viruses as Delivery Mechanisms
Gene therapy is the introduction of genetic material into a human’s cells to replace a malfunctioning gene or make a protein that can compensate for the e ects of a certain disease or condition (Sung & Kim, 2019). One of the mechanisms by which this genetic material is inserted into cells is viruses. Researchers around the globe have honed and re ned the use of viral vectors, such that current treatment mitigates the systematic in ammation and organ failure that marked earlier attempts (Lundstrom, 2018). e bene t of this mechanism is that it provides continued, long-term expression of the corrected gene at physiologically e ective levels. e most commonly used viruses for viral delivery are adenoviruses (AAV), which provide advantages such as infection of a vast range of host cells, including dividing and non-dividing cells, and their maintenance as an episome, meaning that the inserted genetic material behaves as an extrachromosomal element in the targeted cell’s nucleus, reducing the risk of mutagenesis (McCa rey et al., 2008). Adenoviruses can either be replication-de cient (RD) or replicationcompetent (RC). RD adenovirus vectors have certain genes deleted to prevent replication of the virus, which would exponentially increase the lethal immunogenic response of the host, and to prevent transduced cells from undergoing apoptosis. RD adenoviruses are especially useful for gene therapy to promote continued expression of the foreign transgene. RC adenoviruses, on the other hand, replicate more e ciently since they have the necessary genes coding for replicative proteins and are important factors for lysing and destroying cancer cells.
Retroviruses, unlike adenoviruses, are considered the optimal standard for long-term gene therapy as they can carry up to eight kilobases of foreign inserts and can replicate their single-stranded RNA into double-stranded DNA, which is then permanently inserted into the human genome (Anson, 2004). One of the major downsides of retroviruses is their inability to infect nondividing cells, but one class of retroviruses called lentiviruses circumvents this problem and has
provided a quantum leap in mediating high levels of gene transfer and cell transduction. Bacteriophages, viruses that infect bacteria, are promising viral classes that provide the advantage of easy expression of foreign molecules on the outer surface of the phage, allowing for high levels of targeting (Hosseinidoust, 2017). Downsides of using bacteriophages include undesirable immune responses and lower gene delivery e ciency.
Viruses are an excellent vehicle for targeted gene therapy, immunotherapy, cancer therapy, and even treatment for infectious diseases. eir innovative transformation into therapeutic drugs has revolutionized the elds of cell biology, genetics, and clinical treatment (Hosseinidoust, 2017).
Oncolytic Virus erapy
Oncolytic viruses infect and kill cancer cells either naturally or due to modi cation in a lab (Using Oncolytic Viruses to Treat Cancer - National Cancer Institute, 2018). Patients who developed a naturally-acquired viral infection as early as the late 1800s were observed to have tumor regression either during or a er the infection (Using Oncolytic Viruses to Treat Cancer - National Cancer Institute, 2018). is led to multiple clinical trials between 1950 and 1980 to test the e cacy of naturally occurring viruses in treating cancer, but these e orts were largely unsuccessful (Fukuhara et al., 2016). e challenge currently facing researchers is nding a way to edit virus genomes such that the virus reproduces in cancer cells without also infecting healthy cells. Viruses can also be modi ed to trigger a systemic tumorspeci c immune response (Fukuhara et al., 2016).
Only one oncolytic virus therapy, talimogene laherparepvec (T-VEC), is currently approved by the FDA for clinical use (Using Oncolytic Viruses to Treat Cancer - National Cancer Institute, 2018). T-VEC treats metastatic melanoma by both selectively targeting cancerous cells and promoting a regional and systemic immune response to suppress tumors (Andtbacka et al., 2015). T-VEC was developed from a modi ed herpes simplex virus (HSV) type 1 (Andtbacka et al., 2015). Two genes were deleted: the herpes virus neurovirulence factor gene, ICP34.5, and the ICP47 gene. Additionally, the gene encoding for human granulocyte macrophage colonystimulating factor (GM-CSF, a cytokine that helps produce white blood cells) was inserted into the viral genome to help produce a tumorspeci c immune response (Andtbacka et al., 2015). Currently, T-VEC is given via injection to patients with advanced melanoma—either Stage IIIB, IIC, or IV—that cannot be surgically removed (Andtbacka et al., 2015).
Viruses and Nanoparticles
E ective drug delivery and targeting are essential for e ective medical treatment. However, because drugs are poorly soluble in water, most of them are lost in the body during administration and only a small portion gets localized to the targeted site (van Kan-Davelaar et al., 2014). Nanomedicine has made tremendous leaps in compensating for these de ciencies, as the use of semisynthetic carriers like quantum dots, liposomes, and vesicles has increased the capacity for cellular uptake, intracellular accumulation, and physiological retention. Viruses have also emerged as an optimal nanoparticle biodelivery system, primarily because of their high biocompatibility and biodegradability.
As nanocarriers, viruses can target proteins, polymers, and enzymes to speci c sites and cells in the body. A viral nanocarrier is made by removing the viral genome and reconstructing the outer capsid viral proteins into a shell that can disassemble and release the product in response to pH, chemical stimuli, and temperature (van Kan-Davelaar et al., 2014). erefore, viral nanoparticles allow site-speci c targeting of desired products in a context-dependent regulation.
Viral nanoparticles can also be used for the treatment of metastatic cancer. In comparison to traditional treatments like radiation and chemotherapy, which can be highly lethal and cytotoxic, viral nanotechnology can mediate e cient levels of molecular tra cking of proteins, antibodies, uorescent dyes, and drugs to tumor cells (Grasso & Santi, 2010). When these viral nanoparticles are labeled with a uorescent tag, some classes of these viruses like cowpea mosaic virus (CPMV) can interact with the intermediate lament protein vimentin, which is overexpressed in cancer cells (Steinmetz et al., 2011). rough these particles, we can better detect the localization of tumors and metastatic cancer cells in vitro and in vivo. In addition to imaging, viruses can be used to deliver anticancer agents to only cancer cells instead of normally a ecting healthy cells. eir highly symmetrical structure allows researchers to conjugate and present multiple targeting molecules on its surface to mediate highly speci c cell targeting with high payload capacities (Grasso & Santi, "Patients who developed a naturallyacquired viral infection as early as the late 1800s were observed to have tumor regression either during or a er the infection."
e most e ective types of viruses for nanoparticle targeting are plant-based viruses and bacteriophages. Mammalian viruses are not optimal vehicles because they proliferate in humans, which can trigger downstream negative e ects (Steinmetz et al., 2011). While viral nanoparticles o er exciting potential for drug delivery and disease treatment, many of these technologies are still in their nascent stages and few are in preclinical trials. Yet, the groundbreaking work and bleeding-edge research with viral nanoparticles thus far has paved a new avenue of research that can transform the eld of pharmacokinetics and drug delivery.
Viruses for Imaging
e application of recent advancements in viral nanotechnology and functional viruses as therapeutics relies on sophisticated imaging techniques. Imaging viruses has a wide range of uses. It enables scientists to utilize viruses as calibration tools, using their regular identical structures as control specimens for testing parameters (Goldsmith & Miller, 2009). Primarily, though, imaging is used to study virus structure and function. is includes studying the assembly and infection processes. Virus imaging has also recently been used to study viruses as functional nanoparticles in medicine and nanotechnology. e functional applications also range from materials science to biophysics and electrochemistry (Gulati et al., 2019).
One type of microscopy used to study viruses is Atomic Force Microscopy, which has resolution on the nanometer scale and can observe samples in both liquid and air mediums (Goldsmith & Miller, 2009). is form of microscopy works by shining a laser on a cantilever (a beam that is xed at one end) with a tip. e tip is pressed into the sample and the structure of the sample bends the cantilever. e bends are measured and recorded by the laser focused on the cantilever. One mode, called contact mode, ends in the destruction of the sample. Another mode, tapping mode, is gentler and results in less specimen deformation.
Another type of microscopy frequently used to study viruses is electron microscopy. is type of microscopy works by using electrons to form an image of the specimen instead of using light. One type of electron microscopy is called negative staining. is method produces an image with a dark background and white specimen (Goldsmith & Miller, 2009). ese images come from supernatant, so the specimens are in uid. e specimen is stabilized using a support lm to hold the particles. en, a thin carbon coat is evaporated over the lm so the specimen does not melt from the electron beam. is technique is quick and can take as few as een minutes (Goldsmith & Miller, 2009).
in sectioning is another technique used with electron microscopy. is type is used for thin sheets of cells or tissues. Like in the previously discussed types of microscopy, these samples are xed, so living tissues and cells cannot be viewed. ough it is a powerful way to visualize tissue samples, the main limitation for this type of microscopy is that the sample could miss the portion that contains the virus, (Gulati et al., 2019).
Another type of electron microscopy that allows scientists to understand viruses, speci cally virus localization, is immunogold labeling. is technique uses antibodies that bind to viruses and secondary gold labeled antibodies that bind to the primary antibodies (Gulati et al., 2019). e areas that contain the virus show up easily through electron microscopy because the electron-dense gold appears dark against the white portions of the cell. is is useful for qualitative observations about localization of virus, virus parts, or viruslike particles (Gulati et al., 2019).
Lastly, Cryo electron microscopy is another useful tool for visualizing viruses. e method works by rapidly freezing samples with liquid nitrogen, viewing them with a special electron microscope equipped with a cryo stage (a specimen stage that cools the sample down using either liquid nitrogen or liquid helium), and reconstructing samples. Many di erent angles are viewed and the sample is reconstructed in 3D via computer (Gulati et al., 2019). is powerful tool allows for exact three-dimensional models of samples to be made.
Antibiotic Resistance/Viruses
Antibiotic resistance refers to infectious microorganisms evolutionarily gaining the ability to circumvent the consequences of antibiotics, the medicines commonly used to treat their e ects by killing them. e use of viruses to combat such microbes is an area of great interest, especially as antibiotic resistance increases at an alarming rate and some infections (e.g. pneumonia and tuberculosis, among others) o en become very di cult to treat (Zaman et al., 2017). For context, current threats include multidrug-resistant

(MDR) bacteria—which are resistant to a few of the most powerful antibiotics and kill 25,000 patients in European hospitals annually—and extensively drug-resistant (XDR) bacteria, which are resistant to several of the most e ective drugs and result in mortality for more than half of those infected (Moghadam et al., 2020).
e mechanism predominating the use of viruses against antibiotic microbes is called phage therapy. Bacteriophages are viruses able to infect and kill bacteria, notably without negative implications for the human or animal host (Principi et al., 2019). Bacteriophage therapy includes infection of speci c bacterial hosts by phages, which essentially function to hijack the cellular machinery and induce lysis (cell rupturing) of the host bacteria (Nikolich & Filippov, 2020). Although phage biologists have recognized that phage life cycles exist on a spectrum and are composed of many classi cations, the two conventionally known categories are lysogenic phages and lytic phages. Lysogenic phages integrate their genetic material into the bacterial host chromosomes as prophages, thus replicating with each cell division. Some environmental stimuli can trigger the induction of a transition to the lytic cycle and subsequent release of phage o spring from outside of the chromosome.
However, phage therapy relies on lytic phages; these inject their genetic material into bacterial cells and take control of the replication machinery. is way, the phages can replicate—viruses are unable to do so without a host—and produce increasing quantities of progeny. Eventually, a critical mass of progeny is reached, and certain lytic proteins are activated, which hydrolyze the cell wall, resulting in rupture of the bacterial cell. Also, when this cell ruptures, existing phage progeny are free to infect other bacterial cells and repeat the cycle. is way, the bacteria are ultimately cleared from the system (Lin et al., 2017).
Among the bene ts of bacteriophage therapy is the fact that phages can be engineered. For instance, the OMKO1 phage infects only bacterial cells presenting a certain cell surface protein that is important to the bacteria’s system for evading antibiotics. us, pressure will result in bacterial cells that are phage-resistant by nature of mutating the protein important for antibiotic resistance (hence, their resistant ability will be eliminated). Interestingly, and somewhat ironically, phage therapy can be more e cacious in preventing antibiotic resistance when delivered in combination with antibiotics. Since phages and antibiotics present di erent selective pressures, evolutionary tradeo yields minimal amounts of resistance mechanisms (Torres-Barcelo, 2018). ere are several other advantages to phage therapy, including the fact that it operates via mechanisms di erent from antibiotics and thus antibiotic-resistant bacteria do not present an already-phage-resistant threat (Loc-Carrillo & Abedon, 2011).
Unfortunately, there are also downfalls to consider in the use of phage therapy against antibiotic-resistant microbes. Notably, in order for a phage to be e ective and safe it must have a range of characteristics (i.e. be constitutively lytic, survive in the host and reach its target, have the ability to clear its target, and not induce, a harmful response in the host organism) which may be di cult to achieve in combination. Other problems include the phages’ narrow host range, which limits them from eliminating all targets, as well as the possibility that they may be more prone to negative consequences like those of other pharmaceuticals than scientists currently think (Loc-Carrillo & Abedon, 2011).
Applications in the Gut Microbiome
Our gastrointestinal tract is inhabited by a plethora of di erent viruses. To put things into perspective, the gut microbiome has DNA and RNA viruses that collectively outnumber bacterial cells by as many as 10:1 (Mukhopadhya et al.,
Figure 9: An electron microscope. Image Source: Wikimedia Commons
Figure 10: An artistic representation of the gut microbiome.
Image Source: Flickr 2019). Each gram of human gut content is said to contain at least 108- 109 virus-like particles, termed VLPs, with the majority belonging to the family Podoviridae (Mukhopadhya et al., 2019). e human gut microbiome is thus a very complex ecosystem, with organisms ranging from bacteria to yeast, fungi, and even viruses inhabiting it. anks to recent technological advances such as high-throughput and nextgeneration sequencing, entire viral genomes have been sequenced and analyses of microbial communities (metagenomics) completed, collectively revealing new insights into the role of human gut virome composition and how it functions, as well as its potential clinical applications as a therapeutic method. e viruses that inhabit the gut microbiome have been separated into ve virotypes: eukaryotic viruses, plant-derived viruses, giant viruses (larger than 300kb), prophages, and small viruses (smaller than 145 kb) (Scarpellini et al., 2015).
In contrast to the bacteria gut microbiome, the gut virome is more stable and doesn’t uctuate as much in response to environmental factors (Mukhopadhya et al., 2019). e gut microbiome can be in uenced by a variety of factors, including diet, smoking, and antibiotics whereas the gut virome is not. While much remains unknown, there is hope that the gut virome may be an undiscovered entity related to in ammation processes. e gut virome, being very underresearched, holds signi cant promise in future therapeutic applications (Mukhopadhya et al., 2019). Viruses and Machine Learning
e diverse applications of viruses in various elds have prompted researchers to experiment with novel methods of data processing: machine learning and arti cial intelligence. Machine learning is a type of arti cial intelligence that trains a “machine,” or a set of algorithms, to learn from existing data sets and nd patterns (Dhall et al., 2019). Due to the large scale of data sets about viral genomes, machine learning mechanisms such as support vector machines (SVM), deep neural networks, and random forests have been applied into the eld of virology.
A major application of machine learning to virus research is the recovery of viral genomes from existing metagenomic data sets, which contain both host and viral genetic sequences. For example, researchers used a machine learning method to recover genomes of the Inoviridae virus families, which infect bacteria like Vibrio cholerae and intensify diseases like cholera (Roux et al., 2019). ey trained the algorithm using sequences from known Inoviridae and those from other bacteria or viruses to identify Inoviridae sequences, which later recovered more than 10,000 Inoviridae genomes from metagenomic data sets (Roux et al., 2019).
Another example is MARVEL, a random forest machine learning method that makes prediction using many individual decision trees. Amgarten et al. (2018) trained this system using three factors: density of genes compared to the length of the genome, strand shi s between neighboring

genes, and fraction of signi cant hits from the Prokaryotic Virus Orthologous Groups (pVOGs) database. A erwards, MARVEL identi ed 58 new viral genomes from metagenomic datasets of compost samples, only one of which had bacterial marker genes, demonstrating high accuracy. Virsorter2 is another machine learning system that uses automatic classi ers to identify viral genomes from metagenomic datasets (Guo et al., 2021).
Machine learning can extend from identifying genomes to associating particular viral sequences with speci c diseases. Unlike MARVEL, which uses automatic machine learning classi ers like density of genes, VirFinder (Ren et al., 2017) identi es k-mers, combinations of DNA letters (“DNA words”) with a certain length of k, within sequences, rather than speci c genes. Ren et al. (2017) tested VirFinder with human gut metagenomic data, identifying the viral sequences for people with and without liver cirrhosis. e study identi ed types of viruses more prevalent in healthy or diseased people (Ren et al., 2017), supporting the previously suggested correlation between changes in the human gut microbiome and liver cirrhosis (Qin et al., 2014).
Combinations of machine learning have been implemented for viruses as therapeutics. Multiple machine learning methods, including the random forest classi er, were used to predict human adaptation of swine/avian in uenza A viruses (IAVs) from large data sets (Li et al., 2019). Additionally, machine learning systems like neural networks or SVMs have predicted adeno-associated virus (AAV) capsids, or viral protein shells, that would form viral structures and possibly become AAV vectors for gene therapy vectors (Marques et al., 2020). In these ways, the role of machine learning in virology continues to evolve and grow, and it is likely to contribute to the expansion of virology in many di erent settings.
Conclusions
e COVID-19 pandemic has brought the eld of virology to the public eye. However, viruses and viral diseases have made signi cant impacts across human history, from the Black Plague of the 1300s to the Spanish Flu of the 1900s. As we look back at the history of virology, we can gain better understanding of the advances in treating viruses that have been made in recent years. Hopefully, with more study, we can develop more treatments against viral infections and begin to utilize viral vectors ourselves to tackle more References
Ahn, J., & Flamm, S. L. (2011). Hepatitis C therapy: Other players in the game. Clinics in Liver Disease, 15(3), 641–656. https://doi. org/10.1016/j.cld.2011.05.008
Aleem, A., & Kothadia, J. (2021). Remdesivir. StatPearls. https://www.statpearls.com/ ArticleLibrary/viewarticle/122861
Aligne, C. A. (2016). Overcrowding and Mortality During the In uenza Pandemic of 1918. American Journal of Public Health, 106(4), 642–644. https://doi.org/10.2105/AJPH.2015.303018
Amgarten, D., Braga, L. P. P., da Silva, A. M., & Setubal, J. C. (2018). MARVEL, a Tool for Prediction of Bacteriophage Sequences in Metagenomic Bins. Frontiers in Genetics, 9, 304. https://doi.org/10.3389/fgene.2018.00304
Anson, D. S. (2004). e use of retroviral vectors for gene therapy-what are the risks? A review of retroviral pathogenesis and its relevance to retroviral vector-mediated gene delivery. Genetic Vaccines and erapy, 2(1), 9. https://doi. org/10.1186/1479-0556-2-9
Andtbacka, R. H. I., Kaufman, H. L., Collichio, F., Amatruda, T., Senzer, N., Chesney, J., Delman, K. A., Spitler, L. E., Puzanov, I., Agarwala, S. S., Milhem, M., Cranmer, L., Curti, B., Lewis, K., Ross, M., Guthrie, T., Linette, G. P., Daniels, G. A., Harrington, K., … Co n, R. S. (2015). Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. Journal of Clinical Oncology: O cial Journal of the American Society of Clinical Oncology, 33(25), 2780–2788. https://doi. org/10.1200/JCO.2014.58.3377
Beeching, N. J., Fenech, M., & Houlihan, C. F. (2014). Ebola virus disease. BMJ (Clinical Research Ed.), 349, g7348. https://doi. org/10.1136/bmj.g7348
Belshe, R. B. (2009). Implications of the Emergence of a Novel H1 In uenza Virus | NEJM. Retrieved September 8, 2021, from https://www. nejm.org/doi/full/10.1056/NEJMe0903995
Brown, T. A. (2002). e Human Genome. In Genomes. 2nd edition. Wiley-Liss. https://www. ncbi.nlm.nih.gov/books/NBK21134/
Carroll, D., Watson, B., Togami, E., Daszak, P., Mazet, J. A., Chrisman, C. J., Rubin, E. M., Wolfe, N., Morel, C. M., Gao, G. F., Burci, G. L., Fukuda, K., Auewarakul, P., & Tomori, O. (2018). Building a global atlas of zoonotic viruses. Bulletin of the World Health Organization, 96(4), 292–294. https://doi.org/10.2471/BLT.17.205005
De Jesus, N. H. (2007). Epidemics to eradication: e modern history of poliomyelitis. Virology Journal, 4(1), 70. https://doi.org/10.1186/1743422X-4-70
Dhall, A., Dai, D., & Van Gool, L. (2019). Realtime 3D Tra c Cone Detection for Autonomous Driving. 494–501. https://doi.org/10.1109/ IVS.2019.8814089
Dokubo, E. K., Wendland, A., Mate, S. E., Ladner, J. T., Hamblion, E. L., Ra ery, P., Blackley, D. J., Laney, A. S., Mahmoud, N., Wayne-Davies, G., Hensley, L., Stavale, E., Fakoli, L., Gregory, C., Chen, T.-H., Koryon, A., Allen, D. R., Mann, J., Hickey, A., … Fallah, M. P. (2018). Persistence of Ebola virus a er the end of widespread transmission in Liberia: An outbreak report. e Lancet Infectious Diseases, 18(9), 1015–1024. https://doi.org/10.1016/S1473-3099(18)30417-1 Ebola Virus Disease—WHO. (2021). Retrieved September 8, 2021, from https://www.who.int/ westernpaci c/health-topics/ebola
Ehrhardt, S. A., Zehner, M., Krähling, V., CohenDvashi, H., Kreer, C., Elad, N., Gruell, H., Ercanoglu, M. S., Schommers, P., Gieselmann, L., Eggeling, R., Dahlke, C., Wolf, T., Pfeifer, N., Addo, M. M., Diskin, R., Becker, S., & Klein, F. (2019). Polyclonal and convergent antibody response to Ebola virus vaccine rVSV-ZEBOV. Nature Medicine, 25(10), 1589–1600. https://doi. org/10.1038/s41591-019-0602-4
Ellebedy, A. H., & Ahmed, R. (2016). Chapter 15 - Antiviral Vaccines: Challenges and Advances. In B. R. Bloom & P.-H. Lambert (Eds.), e Vaccine Book (Second Edition) (pp. 283–310). Academic Press. https://doi.org/10.1016/B978-012-802174-3.00015-1
Emond, R. T., Evans, B., Bowen, E. T., & Lloyd, G. (1977). A case of Ebola virus infection. British Medical Journal, 2(6086), 541–544.
Fukuhara, H., Ino, Y., & Todo, T. (2016). Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Science, 107(10), 1373–1379. https://doi.org/10.1111/cas.13027 Gavrilov, L. A., & Gavrilova, N. S. (2020). What Can We Learn about Aging and COVID-19 by Studying Mortality? Biochemistry. Biokhimiia, 85(12), 1499–1504. https://doi.org/10.1134/ S0006297920120032
Geddes, A. M. (2006). e history of smallpox. Clinics in Dermatology, 24(3), 152–157. https:// doi.org/10.1016/j.clindermatol.2005.11.009
Goldsmith, C. S., & Miller, S. E. (2009). Modern Uses of Electron Microscopy for Detection of Viruses. Clinical Microbiology Reviews, 22(4), 552–563. https://doi.org/10.1128/CMR.00027-09 Graham, B. S. (2013). Advances in antiviral vaccine development. Immunological Reviews, 255(1), 230–242. https://doi.org/10.1111/ imr.12098
Grasso, S., & Santi, L. (2010). Viral nanoparticles as macromolecular devices for new therapeutic and pharmaceutical approaches. International Journal of Physiology, Pathophysiology and Pharmacology, 2(2), 161–178.
Groseth, A., Feldmann, H., & Strong, J. E. (2007). e ecology of Ebola virus. Trends in Microbiology, 15(9), 408–416. https://doi. org/10.1016/j.tim.2007.08.001 Gulati, N. M., Torian, U., Gallagher, J. R., & Harris, A. K. (2019). Immunoelectron Microscopy of Viral Antigens. Current Protocols in Microbiology, 53(1), e86. https://doi. org/10.1002/cpmc.86
Guo, J., Bolduc, B., Zayed, A. A., Varsani, A., Dominguez-Huerta, G., Delmont, T. O., Pratama, A. A., Gazitúa, M. C., Vik, D., Sullivan, M. B., & Roux, S. (2021). VirSorter2: A multi-classi er, expert-guided approach to detect diverse DNA and RNA viruses. Microbiome, 9(1), 37. https:// doi.org/10.1186/s40168-020-00990-y
Hartman, A. L., Bird, B. H., Towner, J. S., Antoniadou, Z.-A., Zaki, S. R., & Nichol, S. T. (2008). Inhibition of IRF-3 Activation by VP35 Is Critical for the High Level of Virulence of Ebola Virus. Journal of Virology, 82(6), 2699–2704. https://doi.org/10.1128/JVI.02344-07
Hosseinidoust, Z. (2017). Phage-Mediated Gene erapy. Current Gene erapy, 17(2), 120–126. https://doi.org/10.2174/15665232176661705101 51940
Howard, R. S. (2005). Poliomyelitis and the postpolio syndrome. BMJ : British Medical Journal, 330(7503), 1314–1318.
Ison, M. G. (2011). Antivirals and resistance: In uenza virus. Current Opinion in Virology, 1(6), 563–573. https://doi.org/10.1016/j. coviro.2011.09.002
Jacob, S. T., Crozier, I., Fischer, W. A., Hewlett, A., Kra , C. S., Vega, M.-A. de L., Soka, M. J., Wahl, V., Gri ths, A., Bollinger, L., & Kuhn, J. H. (2020). Ebola virus disease. Nature Reviews Disease Primers, 6(1), 1–31. https://doi.org/10.1038/ s41572-020-0147-3
Lacey, B. W. (1949). THE NATURAL HISTORY OF POLIOMYELITIS. e Lancet, 253(6560), 849–859. https://doi.org/10.1016/S01406736(49)92350-8
Li, Y., Carroll, D. S., Gardner, S. N., Walsh, M. C., Vitalis, E. A., & Damon, I. K. (2007). On the origin of smallpox: Correlating variola phylogenics with historical smallpox records. Proceedings of the National Academy of Sciences, 104(40), 15787–15792. https://doi.org/10.1073/pnas.0609268104
Li, J., Zhang, S., Li, B., Hu, Y., Kang, X.-P., Wu, X.Y., Huang, M.-T., Li, Y.-C., Zhao, Z.-P., Qin, C.-F., & Jiang, T. (2019). Machine Learning Methods for Predicting Human-Adaptive In uenza A Viruses Based on Viral Nucleotide Compositions. Molecular Biology and Evolution, 37(4), 1224–1236. https://doi.org/10.1093/molbev/msz276
Lin, D. M., Koskella, B., & Lin, H. C. (2017). Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World Journal of Gastrointestinal Pharmacology and erapeutics, 8(3), 162–173. https://doi.org/10.4292/wjgpt. v8.i3.162
Loc-Carrillo, C., & Abedon, S. T. (2011). Pros and cons of phage therapy. Bacteriophage, 1(2), 111–114. https://doi.org/10.4161/bact.1.2.14590 Lundstrom, K. (2018). Viral Vectors in Gene erapy. Diseases (Basel, Switzerland), 6(2), E42. https://doi.org/10.3390/diseases6020042
Marques, J., Dillingham, M., Beckett, P., Cherradi, Y., Paun, A., Boumlic, A., & Carter, P. (2020). Optimizing Viral Vector Manufacturing for Gene erapy. PharmTech, 2020 eBook(2), 24–29.
McCa rey, A. P., Fawcett, P., Nakai, H., McCa rey, R. L., Ehrhardt, A., Pham, T.-T. T., Pandey, K., Xu, H., Feuss, S., Storm, T. A., & Kay, M. A. (2008). e host response to adenovirus, helper-dependent adenovirus, and adeno-associated virus in mouse liver. Molecular erapy: e Journal of the American Society of Gene erapy, 16(5), 931–941. https://doi.org/10.1038/mt.2008.37
Mehndiratta, M. M., Mehndiratta, P., & Pande, R. (2014). Poliomyelitis: Historical Facts, Epidemiology, and Current Challenges in Eradication. e Neurohospitalist, 4(4), 223–229. https://doi.org/10.1177/1941874414533352
Melnick, J. L. (1996). Current status of poliovirus infections. Clinical Microbiology Reviews, 9(3), 293–300. https://doi.org/10.1128/CMR.9.3.293
Metzger, W. G., & Vivas-Martínez, S. (2018). Questionable e cacy of the rVSV-ZEBOV Ebola vaccine. e Lancet, 391(10125), 1021. https:// doi.org/10.1016/S0140-6736(18)30560-9
Miller, J. C. (2014). Epidemics on Networks with Large Initial Conditions or Changing Structure. PLOS ONE, 9(7), e101421. https://doi. org/10.1371/journal.pone.0101421
Miranda, M. E., Ksiazek, T. G., Retuya, T. J., Khan, A. S., Sanchez, A., Fulhorst, C. F., Rollin, P. E., Calaor, A. B., Manalo, D. L., Roces, M. C., Dayrit, M. M., & Peters, C. J. (1999). Epidemiology of Ebola (Subtype Reston) Virus in the Philippines, 1996. e Journal of Infectious Diseases, 179(Supplement_1), S115–S119. https://doi. org/10.1086/514314
Moghadam, M., Amirmozafari, N., Shariati, A., Hallajzadeh, M., Mirkalantari, S., Khoshbayan, A., & Masjedian Jazi, F. (2020). How Phages Overcome the Challenges of Drug Resistant Bacteria in Clinical Infections. Infection and Drug Resistance, 13, 45–61. https://doi. org/10.2147/IDR.S234353
Morens, D. M., Folkers, G. K., & Fauci, A. S. (2009). What Is a Pandemic? e Journal of Infectious Diseases, 200(7), 1018–1021. https:// doi.org/10.1086/644537
Mukhopadhya, I., Segal, J. P., Carding, S. R., Hart, A. L., & Hold, G. L. (2019). e gut virome: e ‘missing link’ between gut bacteria and host immunity? https://journals.sagepub.com/ doi/10.1177/1756284819836620
Muyembe-Tamfum, J. J., Mulangu, S., Masumu, J., Kayembe, J. M., Kemp, A., & Paweska, J. T. (2012). Ebola virus outbreaks in Africa: Past and present. Onderstepoort Journal of Veterinary Research, 79(2), 06–13.
Nikolich, M. P., & Filippov, A. A. (2020).
Bacteriophage erapy: Developments and Directions. Antibiotics (Basel, Switzerland), 9(3), E135. https://doi.org/10.3390/antibiotics9030135 Pardi, N., Hogan, M. J., & Weissman, D. (2020). Recent advances in mRNA vaccine technology. Current Opinion in Immunology, 65, 14–20. https://doi.org/10.1016/j.coi.2020.01.008
Parrino, J., & Graham, B. S. (2006). Smallpox vaccines: Past, present, and future. e Journal of Allergy and Clinical Immunology, 118(6), 1320–1326. https://doi.org/10.1016/j.jaci.2006.09.037
Petrosillo, N., Viceconte, G., Ergonul, O., Ippolito, G., & Petersen, E. (2020). COVID-19, SARS and MERS: Are they closely related? Clinical Microbiology and Infection: e O cial Publication of the European Society of Clinical Microbiology and Infectious Diseases, 26(6), 729–734. https://doi.org/10.1016/j.cmi.2020.03.026
Plowright, R. K., Parrish, C. R., McCallum, H., Hudson, P. J., Ko, A. I., Graham, A. L., & LloydSmith, J. O. (2017). Pathways to zoonotic spillover. Nature Reviews Microbiology, 15(8), 502–510. https://doi.org/10.1038/nrmicro.2017.45
Pourrut, X., Kumulungui, B., Wittmann, T., Moussavou, G., Délicat, A., Yaba, P., Nkoghe, D., Gonzalez, J.-P., & Leroy, E. M. (2005). e natural history of Ebola virus in Africa. Microbes and Infection, 7(7), 1005–1014. https://doi. org/10.1016/j.micinf.2005.04.006
Principi, N., Silvestri, E., & Esposito, S. (2019). Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections. Frontiers in Pharmacology, 10, 513. https://doi.org/10.3389/ fphar.2019.00513
Qin, N., Yang, F., Li, A., Pri i, E., Chen, Y., Shao, L., Guo, J., Le Chatelier, E., Yao, J., Wu, L., Zhou, J., Ni, S., Liu, L., Pons, N., Batto, J. M., Kennedy, S. P., Leonard, P., Yuan, C., Ding, W., … Li, L. (2014). Alterations of the human gut microbiome in liver cirrhosis. Nature, 513(7516), 59–64. https://doi. org/10.1038/nature13568
Ravanfar, P., Satyaprakash, A., Creed, R., & Mendoza, N. (2009). Existing antiviral vaccines. Dermatologic erapy, 22(2), 110–128. https:// doi.org/10.1111/j.1529-8019.2009.01224.x
Ren, J., Ahlgren, N. A., Lu, Y. Y., Fuhrman, J. A., & Sun, F. (2017). VirFinder: A novel k-mer based tool for identifying viral sequences from assembled metagenomic data. Microbiome, 5(1), 69. https://doi.org/10.1186/s40168-017-0283-5 Riechmann, L., Clark, M., Waldmann, H., & Winter, G. (1988). Reshaping human antibodies for therapy. Nature, 332(6162), 323–327. https:// doi.org/10.1038/332323a0
Roux, S., Adriaenssens, E. M., Dutilh, B. E., Koonin, E. V., Kropinski, A. M., Krupovic, M., Kuhn, J. H., Lavigne, R., Brister, J. R., Varsani, A., Amid, C., Aziz, R. K., Bordenstein, S. R., Bork, P., Breitbart, M., Cochrane, G. R., Daly, R. A., Desnues, C., Duhaime, M. B., … EloeFadrosh, E. A. (2019). Minimum Information about an Uncultivated Virus Genome (MIUViG). Nature Biotechnology, 37(1), 29–37. https://doi. org/10.1038/nbt.4306
Samudrala, P. K., Kumar, P., Choudhary, K., akur, N., Wadekar, G. S., Dayaramani, R., Agrawal, M., & Alexander, A. (2020). Virology, pathogenesis, diagnosis and in-line treatment of COVID-19. European Journal of Pharmacology, 883, 173375. https://doi.org/10.1016/j. ejphar.2020.173375
Scarpellini, E., Ianiro, G., Attili, F., Bassanelli, C., Santis, A. D., & Gasbarrini, A. (2015). e human gut microbiota and virome: Potential therapeutic implications. Digestive and Liver Disease, 47(12), 1007–1012. https://doi.org/10.1016/j. dld.2015.07.008
Schwaber, J., & Cohen, E. P. (1973). Human × Mouse Somatic Cell Hybrid Clone secreting Immunoglobulins of both Parental Types. Nature, 244(5416), 444–447. https://doi. org/10.1038/244444a0
Simões, E. A. F., Forleo-Neto, E., Geba, G. P., Kamal, M., Yang, F., Cicirello, H., Houghton, M. R., Rideman, R., Zhao, Q., Benvin, S. L., Hawes, A., Fuller, E. D., Wloga, E., Pizarro, J. M. N., Munoz, F. M., Rush, S. A., McLellan, J. S., Lipsich, L., Stahl, N., … Sivapalasingam, S. (2020). Suptavumab for the Prevention of Medically Attended Respiratory Syncytial Virus Infection in Preterm Infants. Clinical Infectious Diseases, ciaa951. https://doi.org/10.1093/cid/ciaa951
Singer, B. J., ompson, R. N., & Bonsall, M. B. (2021). e e ect of the de nition of ‘pandemic’ on quantitative assessments of infectious disease outbreak risk. Scienti c Reports, 11(1), 2547. https://doi.org/10.1038/s41598-021-81814-3
Sung, Y., & Kim, S. (2019). Recent advances in the development of gene delivery systems. Biomaterials Research, 23(1), 8. https://doi.
e IMpact-RSV Study Group. (1998). Palivizumab, a Humanized Respiratory Syncytial Virus Monoclonal Antibody, Reduces Hospitalization From Respiratory Syncytial Virus Infection in High-risk Infants. Pediatrics, 102(3), 531–537. https://doi.org/10.1542/peds.102.3.531
Torres-Barceló, C. (2018). e disparate e ects of bacteriophages on antibiotic-resistant bacteria. Emerging Microbes & Infections, 7(1), 168. https://doi.org/10.1038/s41426-018-0169-z Umakanthan, S., Sahu, P., Ranade, A. V., Bukelo, M. M., Rao, J. S., Abrahao-Machado, L. F., Dahal, S., Kumar, H., & Kv, D. (2020). Origin, transmission, diagnosis and management of coronavirus disease 2019 (COVID-19). Postgraduate Medical Journal, 96(1142), 753–758. https://doi. org/10.1136/postgradmedj-2020-138234
Using Oncolytic Viruses to Treat Cancer— National Cancer Institute. (2018, February 9). [CgvBlogPost]. https://www.cancer.gov/newsevents/cancer-currents-blog/2018/oncolyticviruses-to-treat-cancer
van Kan-Davelaar, H. E., van Hest, J. C. M., Cornelissen, J. J. L. M., & Koay, M. S. T. (2014). Using viruses as nanomedicines. British Journal of Pharmacology, 171(17), 4001–4009. https:// doi.org/10.1111/bph.12662
Vardanyan, R., & Hruby, V. (2016). Chapter 34— Antiviral Drugs. In R. Vardanyan & V. Hruby (Eds.), Synthesis of Best-Seller Drugs (pp. 687–736). Academic Press. https://doi.org/10.1016/ B978-0-12-411492-0.00034-1
Villarreal, L. P. (2008). Are Viruses Alive? Scienti c American. https://www.scienti camerican.com/ article/are-viruses-alive-2004/
Wang, W., Wu, C., Amarasinghe, G. K., & Leung, D. W. (2019). Ebola Virus Replication Stands Out. Trends in Microbiology, 27(7), 565–566. https://doi.org/10.1016/j.tim.2019.05.004
Weinreich, D. M., Sivapalasingam, S., Norton, T., Ali, S., Gao, H., Bhore, R., Musser, B. J., Soo, Y., Rofail, D., Im, J., Perry, C., Pan, C., Hosain, R., Mahmood, A., Davis, J. D., Turner, K. C., Hooper, A. T., Hamilton, J. D., Baum, A., … Yancopoulos, G. D. (2020). REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19. New England Journal of Medicine. https://doi. org/10.1056/NEJMoa2035002 Wessner, D. R. (2010). Mimivirus | Learn Science at Scitable. Discovery of the Giant Mimivirus. https://www.nature.com/scitable/topicpage/ discovery-of-the-giant-mimivirus-14402410/
Wever, P. C., & van Bergen, L. (2014). Death from 1918 pandemic in uenza during the First World War: A perspective from personal and anecdotal evidence. In uenza and Other Respiratory Viruses, 8(5), 538–546. https://doi.org/10.1111/ irv.12267
What is a Pandemic? (2021). http://www.who. int/csr/disease/swineflu/frequently_asked_ questions/pandemic/en/
Zaman, S. B., Hussain, M. A., Nye, R., Mehta, V., Mamun, K. T., & Hossain, N. (2017). A Review on Antibiotic Resistance: Alarm Bells are Ringing. Cureus, 9(6), e1403. https://doi.org/10.7759/ cureus.1403 Zhang, T., Wu, Q., & Zhang, Z. (2020). Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Current Biology, 30(7), 1346-1351.e2. https://doi.org/10.1016/j. cub.2020.03.022
Zhou, X., Jiang, X., Qu, M., Aninwene, G. E., Jucaud, V., Moon, J. J., Gu, Z., Sun, W., & Khademhosseini, A. (2020). Engineering Antiviral Vaccines. ACS Nano, 14(10), 12370–12389. https://doi.org/10.1021/acsnano.0c06109 Zoonoses. (2020, July 29). https://www.who.int/ news-room/fact-sheets/detail/zoonoses

HIV/AIDS and eir Treatments
STAFF WRITERS: CAROLINE CONWAY '24, SREEKAR KASTURI '24, FRANK CARR '22, LAUREN FERRIDGE '23, SOYEON (SOPHIE) CHO '24, ZOE CHAFOULEAS '24, JUSTIN CHONG '24, SARAH LAMSON '24 , JOHN ZAVRAS '24
TEAM LEADS: ANAHITA KODALI '23, MATTHEW LUTCHKO '23
Cover Image: Scanning electron image of HIV. Image Source: Wikimedia Commons Introduction to HIV/AIDS
ough the most commonly discussed viral disease in the current day is COVID-19, there have been many other viral diseases that have forever changed the elds of virology, immunology, and clinical medicine. Perhaps the most well-known of these is the HIV/AIDS epidemic.
HIV, or human immunode ciency virus, originated from chimps in Central Africa. It is likely that the chimpanzee version of the virus – simian immunode ciency virus (SIV) – was passed to humans who hunted chimpanzees and were exposed to contaminated chimpanzee blood. is viral crossover event led to the introduction of HIV to the human world and one of the deadliest pandemics in history (About HIV/AIDS CDC, 2021). Since emerging on a large scale in 1981, HIV has proven to be a deadly virus that has ba ed scientists and required concerted e orts from doctors, epidemiologists, and law-making o cials to understand and control. HIV attacks the human immune system and leads to AIDS (acquired immunode ciency syndrome) if le untreated. If HIV progresses to AIDS, there is no cure. Patients are more prone to multiple infections as a result of a severely weakened immune system.
Since HIV’s emergence in the early 1980’s, over 60 million people have been infected and 25 million people have died because of it or related illnesses. HIV disproportionally impacts developing countries and minority communities because they lack educational resources, screening tools, and access to the necessary medical treatment. HIV was ‘discovered’ when gay men started becoming unusually susceptible to illnesses their immune systems would usually be able to ght o . A icted persons also noticed dark purple lesions on their arms and face, which signaled the rare and aggressive Kaposi’s sarcoma. ese symptoms became the trademark for the “gay plague” in HIV hotspots such as New York and San Francisco. An increase in the number of HIV positive individuals marked an increase in social stigma towards gay communities and ampli ed social paranoia with respect to this new unknown virus. Likewise, doctors did not know how to treat this new mystery disease and all they could do at that time was treat the various “opportunistic infections” (Greene, 2007). It took at least a year for the medical eld to learn more about the virus and its transmission. Epidemiological evidence concluded that HIV and AIDS was transmitted
through contaminated bodily uids and blood. is meant it was a sexually transmitted disease where people sharing drug needles were also susceptible to elevated risks of infection. In addition to these two modes of transmission, pre- and post-natal routes of exposure were found to be a possibility, meaning an infected mother could transmit it to her infant during pregnancy or through breastfeeding. Infection rates rose through the 80s, 90s, and 2000s: from 683,000 people living with HIV at the end of the 80s to an estimated 1,172,700 people in 2010. (About HIV/ AIDS CDC, 2021). Case counts peaked in 1992 when AIDS became the leading cause of death for men aged 25-44 and no signi cant decrease in cases was observed until 1997. is success in preventing an increase in cases was accredited to education e orts on modes of transmission and increased screening e orts in all communities.
HIV is a retrovirus characterized by a chronic course of disease, a long latency period, and persistent viral replication. It seeks out and destroys cells that orchestrate the immune response, speci cally CD4 T lymphocytes (Greene, 2007). ere are two types of HIV: HIV-1 and HIV-2. HIV-1 is spread throughout the world and HIV-2 is contained to Western and Central Africa. Although largely similar and sharing the three basic structural genes, HIV-1 and HIV-2 di er in their internal organization. Furthermore, HIV-1 infection course takes longer to progress, while HIV-2 appears less virulent and is more commonly associated with nervous system diseases in addition to AIDS (Emanuele, 2010). Both types of HIV have an extremely high mutation rate, which makes them even more virulent. is is because of a high error rate in its reverse transcriptase and recombination events, which results in a high rate of mutation, thereby making it possible for patients to be infected with several di erent forms of HIV at the same time (Greene, 2007).
HIV infection is separated into 3 stages. e rst stage, Acute HIV Infection, is the stage where patients have high amounts of HIV in their blood and are very contagious. ey may experience u-like symptoms or no symptoms at all. Next, chronic HIV infection or asymptomatic HIV infection of clinical latency is when HIV is still active in the patient but reproduces at lower levels. It is possible to transmit HIV at this stage, but it is a crucial stage for medication to prevent progression to stage three. Without medication, one could be in this stage for up to a decade. e transition from stage 2 to 3 occurs when HIV in the blood increases and CD4 cell count falls below 200cell/mm. At this point Stage 3 – Acquired Immunode ciency Syndrome (AIDS) – is experienced. is is a critical stage in which the immune system is at its most damaged and there are an increased number of opportunistic infections. e development of AIDS implies both high viral load and high virulence. Without treatment at this stage, patients typically survive for only about 3 years (About HIV/AIDS CDC, 2021). e emergence of HIV has de ned an era of epidemiological trial and error. Although HIV is still a threat and a cure is yet to be discovered, increased education and screening, which are already being implemented, is the rst step to defeating this virus.
Disparities in HIV/AIDS
Racial Disparities
ere are many disparities that exist in the distribution of HIV/AIDS across the US. For one, there are clear racial divides in the prevalence of HIV/AIDS. For example, Black Americans bare the largest burden of HIV infection in the US. ough they account for only 12% of the total American population, they account for 46% of the HIV-infected people in the country. 1,715 Black people per 100,000 individuals have HIV, a gure almost 8 times higher than that of White people. In particular, Black women are disproportionately a ected by HIV infection; the HIV diagnosis rate for Black women is over 14 times higher than the rate of diagnosis for white women (Moore, 2011). e Latinx community is also disproportionately a ected by HIV/AIDS, as the AIDS case rate across the Latinx community is 3 times higher than that of White Americans (Gonzalez et al., 2009). For Indigenous populations, a host of HIV-related behaviors – including high levels of domestic violence, discrimination from other racial/ethnic groups, and high levels of intravenous substance abuse – coupled with mistrust of health services have poor healthcare and outcomes for Indigenous peoples with HIV (Negin et al., 2015). Finally, though Asian Americans are o en considered to be “model” minority groups that typically do not engage in risky sexual or drug related behaviors, they are the only ethnic group in the US that has had a continued increase in HIV infection rates across the 2010 – 2020 decade (Kim & Aronowitz, 2019).
Socioeconomic Disparities
HIV is clearly connected to both social and economic disparities. For urban communities, there is a clear inverse relationship between "Since HIV’s emergence in the early 1980’s, over 60 million people have been infected and 25 million people have died because of it or related illnesses."
socioeconomic status and rate of HIV infection – in other words, the lower socioeconomic status a certain urban community has, the higher the rate of HIV infection is (“Economically Disadvantaged,” 2019). ere are many reasons for this – a lack of socioeconomic resources, including education and nancial security, is linked to HIV-risk behaviors, like unsafe sex or intravenous drug use (“HIV/AIDS and Socioeconomic Status,” n.d.). Critically, though the overall rate of mortality due to HIV/AIDS has dropped in recent years, the decline in mortality for lower socioeconomic groups has been signi cantly slower than the decline in mortality for more privileged groups (Singh et al., 2013). ere are many potential reasons for this – one is that though poorer individuals have been shown to start HIV/AIDS treatment earlier than richer individuals, they have less compliance with treatment regimens. Strict compliance to the recommended drug treatments is essential to the treatments’ success (Tran et al., 2016).
Disparities in the LGBT Community
Historically, HIV/AIDS have both disproportionately a ected the LGBT community. In fact, the rst American o cially diagnosed with AIDS in the US was Ken Horne, a gay sex worker (Ayala & Spieldenner, 2021). Even though there has been signi cant progress made in treating and preventing the spread of HIV/AIDS since 1980 when Horne was rst diagnosed with AIDS, the progress has been uneven, particularly for queer men. As of 2019, gay and bisexual men accounted for about 55% of individuals with HIV/AIDS in the US, though they only account for about 2% of the entire US population (“HIV in the United States and Dependent Areas,” 2021). In part, these disparities are due to healthcare policies that overlook the LGBT community. Studies have shown that the topic of safe sex practices is uncomfortable for healthcare workers to discuss with their patients – one study found that only 68% of medical residents felt comfortable discussing inclusive sexual history, and only 26% of this 68% felt comfortable discussing the topic with LGBT patients (Frasca et al., 2019). Reasons behind these issues include the fact that policies may overlook the existence of LGBT people when developing questionnaires or screenings and that members of the community may be frightened to share their sexual orientation in fear of discrimination (Wheeler, et al., 2011). Social and structural conditions in the US perpetuate the epidemic, too. LGBT individuals are more likely to drink and use illicit substances, are more likely to delay receiving healthcare, are more likely to report poor quality care by healthcare providers, and are less likely to have adequate healthcare than their heterosexual cisgender peers (“LGBTQIA+ Youth and Mental Health,” 2021); each of these factors can increase the risk of contracting HIV.
Disparities Associated with Substance Abuse
Substance abuse disorders, such as addictions to alcohol, crack cocaine, and heroin, are associated with sexually transmitted diseases like HIV because they can also be transmitted through contaminated syringes or needles (“Substance Use,” 2021), such as those used to inject drugs. In 2016, nearly 20 percent of HIV diagnoses among men and 21 percent of diagnoses among women were caused by drug usage (“Drug Use and Viral Infections,” 2020). For drugs that are not injected, the lack of judgement caused by drug use may lead people to partake in risky sexual behavior, making the transmission of HIV more likely (“Substance Use,” 2021). Not only can drugs contribute to the spread of HIV, but they can also worsen the symptoms and progression of the virus. For example, use of drugs like cocaine increases the permeability of the blood brain barrier to viruses, making it easier for HIV to enter the brain, which causes increased nerve cell injury and a ects thinking, learning, and memory (Norman et al., 2009; “Drug Use and Viral Infections,” 2020).
Speci c populations are a ected more severely by substance abuse and HIV. For example, the incarcerated population is disproportionately a ected. e criminal justice system’s population of HIV-infected individuals is 2 to 5 times larger than the outside community. In fact, it is estimated that 1 in 7 of HIV positive people in the US are in the prison system. Additionally, almost half of federal and state prisoners are reported to meet the criteria for drug abuse or dependence, yet few of these prisoners are screened for HIV or receive treatments for substance abuse. Ethnic minorities also appear to be more a ected by both substance abuse and HIV infections. For example, in 2009, the rate of infection within the American Hispanic was three times that of the white communities (“Who Is at Risk,” 2012).
Virology Overview
HIV is a lentivirus within the larger family of retroviruses, which are known to hold their single stranded RNA (ssRNA) genome with a viral envelope and a capsid (Chinen & Shearer, 2002). Two major types of HIV have been discovered: HIV-1 and HIV-2. e three broad
strains of the HIV-1 virus are the following: M (Main)—globally prevalent, O (Outlier)—mostly in Africa, and N (non-M, non-O)—only found in the west-central African country of Cameroon (Simon et al., 1998).
Like other retroviruses, HIV synthesizes DNA copies of its RNA genome and inserts its viral DNA into the host cell’s DNA, a er which the host cell produces viral RNA that form retroviral virions (Nisol & Saib, 2004). e HIV RNA genome encodes and produces 15 proteins— which are divided into regulatory, accessory, and structural proteins—that contribute to the replication process. e regulatory proteins include reverse transcriptase, integrase, and protease, which contribute, respectively, to the reverse transcription of viral RNA into DNA, integration of viral DNA into the host cell’s genome, and cleavage of precursor proteins like gp160 (Gelderblom et al., 1989). e accessory proteins increase the infectivity of the released virions and help with transport at di erent stages (Emerman & Malim, 1998).
e structural proteins are produced by the env, gag, and pol genes and are common to both HIV1 and HIV-2. e env gene produces two envelope glycoproteins called gp120 and gp41 that result from the cleavage of a larger precursor protein gp160 (Fanales-Belasio et al., 2010; Chinen & Shearer, 2002). gp120 is attached to gp41, which is embedded in the lipidic membrane of the viral envelope. Envelope glycoproteins, especially gp 120, contain more variable regions than other parts of an HIV virion. e variable regions allow HIV to bypass the immune system (Wyatt et al., 1998).
Beneath the viral envelope are core proteins produced by the gag gene. One core protein is p17, which comprises the matrix that surrounds core parts like the capsid and the RNA genome. Core proteins called p24 and p6 form the capsid that encloses the RNA. e nucleocapsid protein p7 is attached to the ssRNA itself. e pol gene forms replication enzymes that are used to produce copies of the HIV virions (Fanales-Belasio et al., 2010; Chinen & Shearer, 2002).
An HIV virion enters a human cell as the gp120 and gp41 glycoproteins bind to the CD4 receptors on the human cell membrane. rough one of the variable regions on gp120, called V3, the membrane undergoes conformational change upon binding with CD4 receptors (Kwong et al., 1998), allowing either of the chemokine coreceptors called CCR5 and CXCR4 to bind and A er an HIV virion binds to the host cell, it enters the human cytoplasm and its capsid unpacks the viral RNA and proteins. Here, the viral reverse transcriptase transcribes single stranded RNA into double stranded DNA (dsDNA) through reverse transcription. In this process, cellular lysine tRNA is used as a primer for the viral RNA (Cen et al., 2001), contributing to the errors made by the viral reverse transcriptase, one nucleotide per 1500 to 4000 bases. Due to these errors, many mutants arise from replication that cause drug resistance problems (Tantillo et al., 1994). e viral dsDNA is transported from the cytoplasm to the nucleus, possibly by HIV’s Vpr (Zhang et al., 2001) and Vif proteins (Miller & Sarver, 1997). In the nucleus, the viral integrase integrates dsDNA into a random location on the human genome, a er which the host is likely infected for the rest of their life.
Following integration, the integrated viral DNA is transcribed by human RNA polymerase II to produce three regulatory proteins: Tat, Rev, and Nef. In particular, Tat facilitates faster transcription through transactivation, and Rev regulates the export of di erent lengths of RNA transcripts, with partial RNA transcripts for structural proteins and full-length transcripts for the viral RNA genome of future virions (Parada & Roeder, 1996; Emerman & Malim, 1998). Nef is able to sequester a variety of cell surface proteins by engaging with host tra cking proteins; this disrupts host immune systems and promotes the replication cycle of the virus (Bu alo et al., 2019). As di erent RNA transcripts are transcribed, viral proteases cleave precursor proteins, such as gag-pol, which produce reverse transcriptase, integrase, and protease (Weller & Williams, 2001). Cellular proteases also cleave precursor proteins like gp160, which produce gp120 and gp41, the two envelope glycoproteins, along with other regulatory proteins like Vpr, Vpu, and Vif (Chinen & Shearer, 2002). More research is needed on the late stage of HIV replication, and the assembly of these proteins, which seem to involve cellular components and require energy (Tritel & Resh, 2001). A viral envelope originating from the host cell’s membrane forms around these proteins, which are released into newly generated HIV virions that undergo maturation.
Immunology Overview
e human immune system is responsible for defending the body against outside invaders and harmful molecules and thus serves to combat
Image 1: Image of the HIV capsid. Image Source: Wikimedia Commons pathogens and toxins. It consists of two di erent branches: the innate immune system and the adaptive immune system.
e innate immune system consists of cells and responses that are quick and non-speci c. First, there are the barriers that prevent the entry of pathogens into the body. ese barriers include the skin, the digestive system, and the mucosal barriers which guard openings into the body such as the mouth, nostrils, and the open portions of the genitalia. rough physical and chemical means (such as the enzymes secreted by mucosa and skin, and the gastric acid in the digestive system), many pathogens are prevented from ever posing a threat to the body (Abbas et al., 2020; Agerberth & Guðmundsson, 2006; Boyton & Openshaw, 2002; Smith, 2003). If a threat such as a bacteria or virus is able to slip past these barriers, they then face the cells of the innate immune system. ese include the important phagocytic cells (macrophages, dendritic cells, and neutrophils) which recognize, engulf, and digest many types of pathogens. Once broken into little pieces called antigens, these destroyed pathogens are presented on extracellular proteins called MHC molecules to the T cells of the adaptive immune system located throughout the blood and lymphatic systems. Other innate immune cells include the mast cells, basophils, and eosinophils that are responsible for ghting o larger pathogens, especially parasitic worms. In modern times, they o en are the cause of allergies, which are simply cases of the immune system responding to a non-dangerous pathogen as if it were dangerous. Pathogens are recognized by these cells using sets of proteins on their external and internal membranes called Pattern Recognition Receptors (PRRs). Di erent PRRs are speci c for di erent kinds of pathogen markers, such as double-stranded RNA present in viruses and the agella of bacteria (Abbas et al., 2020; Agerberth & Guðmundsson, 2006).
e adaptive immune system consists of cells that are highly speci c. Each line of cells is speci c for only a single antigen and takes time to fully launch a response. It consists of two di erent classes of cells: B and T cells, named based on their site of nal maturation; B cells emerge complete from the bone marrow while T cells emerge complete from the thymus (Abbas et al., 2020; Janeway, Shlomchik, et al., 2001; Janeway, Travers, et al., 2001).
T cells bind to antigens that are presented to them on the MHC molecules of various cells using their T cell Receptors (TCRs). T cells are further divided into classes based on the di erent cell surface proteins they display. CD8+ T cells are also known as Cytotoxic T cells (CTLs) and are responsible for detecting and destroying cells displaying antigens on MHC Class I molecules. ese o en include cells that are cancerous, infected with a virus, or infested with some sort of intracellular bacteria. CD4+ T cells are also known as Helper T cells ( s) and have a variety of functions, including recognizing


antigens presented on MHC Class II molecules. 1 cells are responsible for activating and supporting CTLs, while 2 cells are responsible for activating and supporting B cells. ere is one other lineage of CD4+ T cells that are not s. ese also display CD25 and are known as Regulatory T cells (Tregs). Tregs are involved in preventing the actions of adaptive immune cells with antigen speci city for self-antigens (and thus preventing autoimmune disorders). ey also ramp down the immune response a er the antigen has been cleared. Because of these various supportive and regulatory functions, CD4+ T cells are some of the most integral cells in the body’s arsenal (Abbas et al., 2020; Janeway, Shlomchik, et al., 2001).
B cells recognize their speci c antigens via binding to them with their B cell Receptors (BCRs). ese antigens can either be 1) presented to them by 2s or cells of the innate immune system or 2) free oating antigens in the blood or lymph systems. is recognition results in the activation of the B cell and the initiation of rapid division, producing many progeny B cells. During replication, these progenies undergo a process known as somatic hypermutation by which they produce slight variations in their BCRs until they produce one with incredibly high speci city. en, the daughter cells develop into plasma cells and memory B cells. Plasma cells secrete free oating versions of their BCRs into the blood and lymph known as antibodies. ese antibodies can be of di erent “classes” based on the type of plasma cell they are, with each class having a slightly di erent function. Antibodies bind to antigens present on infected cells or on the pathogens themselves. Once bound, the antibodies may signal the antigens for phagocytosis, group the antigens together and precipitate them out of the solution, prevent them from undergoing their mechanism of action, or conduct various other functions that result in the destruction of the antigen (Abbas et al., 2020; Janeway, Travers, et al., 2001).
Upon activation, both B and T cells also produce progeny that are known as “memory cells.” ese cells reside within the circulatory systems for weeks to even years, waiting for the antigen they possess a speci city for to reappear in the body. When that happens, the memory cell binds to the antigen and activates to produce more cells with its speci city, causing a more rapid version of the adaptive response generated during the rst activation of the memory cell’s ancestor (Abbas et al., 2020; Janeway, Travers, et al., 2001).
e Biology of HIV/AIDS
Structural Biology of HIV
HIV is composed of a lipid bilayer membrane, two strands of RNA, and diverse types of viral proteins that allow the virus to invade host cells and grow (RCSB, 2000). e structural biology of any virus is crucial to study as information on its structure and function allows targeted drug development to slow or eliminate the virus in infected patients. Additionally, understanding the way HIV operates may provide headway in the development of a vaccine (Engelman & Cherepanov, 2012). Within the viral proteins of HIV, there are subcategories of structural proteins, viral enzymes, and accessory proteins (RCSB, 2000). Each group has a speci c function that could be targeted in the future of HIV treatment.
e structural proteins of HIV include the surface protein, transmembrane protein, matrix protein, capsid protein, and nucleocapsid protein. e surface protein and transmembrane protein are found on the outer layer of the membrane and are coated in carbohydrates to allow them to invade un-noticed, posing as non-threatening
Image Source: Wikimedia Commons
antibodies (Engelman & Cherepanov, 2012). e proteins have jagged structures to penetrate the surface of potential host cells (RCSB, 2000). e matrix protein is located in the inner surface of the lipid bilayer membrane and assists new viruses in budding from the surface of infected host cells (RCSB, 2000). e capsid protein forms a cone shape around the viral RNA and transports the genetic material during infection. Finally, the nucleocapsid protein forms a complex with the RNA to stabilize and protect it during transport (RCSB, 2000).
In addition to the proteins mentioned, HIV has three viral enzymes that are essential for its spread and maturation. First, reverse transcriptase builds DNA using the viral RNA genome (Engelman & Cherepanov, 2012). is enzyme is vital in replicating genetic material to create new viruses. Additionally, the enzyme integrase combines the genetic material of HIV with that of its host cell, which allows HIV to remain dormant in cells for long periods of time. Lastly, protease cleaves large proteins into smaller, functional pieces so that HIV may mature (RCSB, 2000). ere have been drugs developed to block the function of each of these enzymes in an attempt to slow and eliminate HIV (Engelman & Cherepanov, 2012).
In addition to structural proteins and viral enzymes, HIV has accessory proteins that allow the virus to thrive and take part in infection progression. e negative regulatory factor protein attaches to proteins in the infected cell to stop the production of proteins important to the host cell’s defense (RCSB, 2000). is is a crucial factor in how an HIV infection progresses to AIDS (acquired immune de ciency syndrome). To assist the spread of HIV, the viral protein “u” weakens the interaction between envelope proteins and their cell receptors to allow viruses to escape in the cell budding process. Additionally, the viral infectivity factor protein attaches to the host cell’s defense proteins, causing the cell to destroy its own defense proteins in the process of destroying the virus. Lastly, there are accessory proteins that protect HIV’s genetic material. e trans-activator of transcription accelerates viral protein production while the regulator of the virion binds to viral RNA to regulate the slicing and transport of genetic material (RCSB, 2000).
Clearly, each component of HIV is crucial in its ability to spread quickly, maintain infection progression, or even hide dormant in the body for long periods of time (Engelman & Cherepanov, 2012). e future of HIV medicine lies in understanding its structural biology. Acute HIV infection (AHI), also known as primary HIV infection, is when an individual rst becomes infected with HIV. A er the virus enters the human body, there is a period of viral replication that causes the level of HIV in the blood to rise, which greatly increases the risk of future HIV transmission. HIV has two main assets. e rst is mutability of the virus. Reverse transcriptase, the enzyme through which HIV replicates itself, sometimes makes mistakes during transcription. Numerous mistakes accumulate and allow the virus to rapidly mutate over time in response to immunological responses. Studies have shown that when an immune response is targeted toward a particular HIV amino acid sequence, the virus has the ability to change the targeted sequence and become invisible to immune cells. Secondly, HIV can cause the death of CD4+ (helper) T cells, resulting in progressive dysregulation of the immune system. HIV attacks CD4+ cells and infects them, which triggers an immune system response that causes a rise in the number of killer T cells. ese killer T cells kill the infected CD4+ T cells. Consequently, the HIV viral load begins to decline and the number of CD4 cells begins to recover. However, the virus is not completely eliminated from the body and still remains (Cohen et al., 2013).
e detection of AHI is very important for HIV prevention and treatment implementation. However, clinical diagnosis of AHI is extremely di cult because the symptoms that occur during the transition from seronegative to seropositive are o en not recognized as an indicator of AHI (Hoenigl et al., 2016). Previous screening programs rely on point-of-care (POC) HIV antibody testing. POC HIV antibody testing is a technology that allows patients to get tested for HIV and know their status in one visit in under an hour. ough they give rapid results, POC HIV antibody tests o en do not indicate AHI. e Centers for Disease Control and Prevention began addressing this problem by including fourth generation HIV-1 p24 antigen-based immunoassays in laboratory diagnosis of HIV (Hoenigl et al., 2016). Of the current diagnostic tests for AHI, HIV RNA viral load testing seems to be the most useful diagnostic test. is is because HIV antibody testing results are generally negative or indeterminate during AHI (Chu & Selwyn, 2016).
Many people with acute HIV infection commonly experience a fever, swollen lymph nodes, and joint and muscle aches. ese symptoms can
begin just a few days a er an individual has been exposed to HIV and can usually last anywhere from 2 weeks to several months (Gay et al., 2011). However, the signs and symptoms of the infection can feel like any other common virus infection, leaving individuals unable to realize their illness is actually an acute HIV infection.
Chronic HIV Infection
Chronic HIV infection involves a gradual deterioration of the humoral immune response. More speci cally, HIV attacks and depletes antigen-speci c CD4 T cells, which are necessary for lasting B cell memory, which results in B cell dysfunction (Lindqvist et al., 2012). e exact molecular mechanisms leading to this dysfunction remain largely unknown. Lindqvist et al. (2012) proposed that B cell di erentiation is disrupted by HIV-driven increases in populations of TFH (CD4 T follicular helper) cells, which interact with antigen-speci c B cells to instigate the processes of antibody a nity maturation and B cell memory development. NakayamaHosoya et al. (2015), on the other hand, found that broad B cell dysfunction during chronic HIV infections could be due to DNA methylation at the interleukin 2 (IL-2) locus in CD4 T cells. is theory is compatible with that of van Grevenynghe et al. (2011), who found that memory B cells survived at lower rates in cases of disrupted IL-2 signaling. ey implicated IL-2 signaling disruption because IL-2 typically phosphorylates Foxo3a, a transcription factor associated with proapoptotic genes. is phosphorylation results in the degradation of Foxo3a in the cytoplasm, preventing the transcription of the proapoptotic genes. In cases of methylation at the IL-2 locus as Nakayama-Hosoya et al. (2015) described, IL-2 is ultimately unable to phosphorylate Foxo3a, allowing the proapoptotic genes to undergo transcription. is, in turn, reduces the survival of memory B cells during chronic HIV infection by increasing rates of apoptosis (van Grevenynghe et al., 2011). Another molecular factor to consider in chronic HIV infection is the SLAMF7 immune cell receptor, which is generally upregulated in cases of chronic immune activation. Unsurprisingly, SLAMF7 appears to play a role in the immune activity of HIV-infected individuals as well as elevated levels of SLAMF7+ peripheral blood mononuclear cells (specialized immune cells) (O’Connell et al., 2019). Chronic immune activation itself may in part drive mortality in patients living with chronic HIV and could be one reason modern HIV treatments like antiretroviral therapy, while able to signi cantly extend the lifespan of HIV+ patients, have failed to completely extend patient life expectancy to the average for non-infected individuals (Rajasuriar et al., 2013).
Aside from the pathology and symptoms of HIV itself, patients living with chronic HIV face additional challenges. For example, individuals with chronic HIV are especially susceptible to chronic obstructive pulmonary disease (COPD), due in part to severe depletion of lung mucosal CD4 T cells (Popescu et al., 2014). Chronic pain is also incredibly prevalent among people living with HIV, with studies estimating that up to 85% of HIV-infected patients experience chronic pain. While this pain can result from in ammationcaused tissue damage or infection, almost half of reported chronic pain is neuropathic, meaning it results from nervous system dysfunction. is dysfunction might be caused by any number of factors, including HIV viral infection, secondary pathogens, and medications. Generally, disadvantaged groups such as women, people of low socioeconomic status, and drug users do not receive adequate treatment for chronic pain (Bruce et al., 2017). ese treatment disparities are especially concerning since individuals with both chronic HIV and chronic pain tend to report more severe depressive symptoms than individuals coping with either condition alone, possibly due to the internalization of stigmas associated with chronic pain and HIV (Goodin et al., 2018).
Various factors in uence what a patient with chronic HIV considers most crucial for their well-being. In one study, homeless substance users emphasized the need for a 24-hour hotline to provide motivation and support in the face of extreme isolation and the fear of dying. For homeless ex-o enders, housing concerns took precedence over the health concerns associated with chronic HIV itself. In the same study, women stressed the necessity of women-only support groups in order to avoid unwelcome advances from men. Many interviewees referenced feeling shame due to the stigma surrounding HIV and shared that they had faced rejection from family, friends, and spiritual communities (Sankar & Luborsky, 2003). Overall, disparities between demographic groups can a ect how equipped patients are to cope with chronic HIV. In addition to being a matter of physical health, living with HIV can profoundly in uence mental, social, and spiritual wellness.
Comorbidities of HIV/AIDS
e comorbidities of HIV can be considered
as diseases outside the scope of AIDS and its associated illnesses. e average number of HIVassociated comorbidities among HIV patients is 1.1 (Lorenc et al., 2016), and the most common ones include cardiovascular disease, respiratory diseases, and hepatic diseases. Moreover, psychiatric disorders are also extremely prevalent among HIV positive individuals. However, studies show that mortality among HIV infected individuals is primarily due to liver disease, including hepatitis B and C and antiretroviral toxicity, vascular disease, lung disease, cancer, and violence. Even though these comorbidities can occur by chance, most are o en due to the infection itself and its risk factors. As HIV severity increases, comorbidity increases as well.
Overlapping risk factors can also lead to potential coinfection, causing comorbidity. Because of the e ects the HIV infection have on the immune system, older HIV positive individuals have a higher disease burden than those who do not have the infection. is is simply because of the natural decline the immune system experiences as one ages. Negative lifestyles, such as alcoholism, can also a ect increase the risk for overlapping diseases. Some associated disorders are o en associated with ethnicity, gender, and socioeconomic status. Recent literature suggests that patients living with HIV should be assessed independently for common medical conditions. e presence of comorbidities necessitates looking into the associated diseases and the signi cance of HIV care, so that healthcare services can respond adequately to patients who have both HIV and a comorbidity. In the event that patients express symptoms of various associated diseases related to HIV, it may be possible to diagnose the virus early, which can lead to antiretroviral therapy.
Treatment of HIV/AIDS
Current treatments for HIV/AIDS
ough not perfectly e ective, thanks to antiretroviral therapy, HIV-positive patients can lead relatively ordinary lives with life expectancies near (though still below) average. Antiretroviral therapy can help patients continue working; highly active antiretroviral therapy increases the probability of remaining employed from 58% to 94% (Goldman & Bao, 2004). However, HIV treatment should not be confused with HIV elimination. HIV can remain dormant in cells, forming viral reservoirs that can reactivate should daily treatment cease at any point. Due to this fact, antiretroviral therapy is only considered a treatment, not a cure. Despite this shortcoming, antiretroviral therapy is highly e ective and can reduce the amount of HIV RNA in the blood plasma to an undetectable level, which, when maintained for at least six months, practically eliminates the risk of sexually transmitting HIV (National Institute of Allergy and Infectious Diseases [NIAID], 2018). For most individuals, this undetectable level of HIV RNA can be reached within six months of starting antiretroviral therapy. “Blips” in which HIV RNA levels temporarily rise above the undetectable threshold are common even with daily antiretroviral therapy, and the cause of these blips remains unknown. An HIV-negative individual can take antiretroviral medication as a preventative measure; this is known as preexposure prophylaxis (NIAID, 2020).
As for the details of antiretroviral therapy itself, there are ve approved medication classes. e rst class consists of HIV entry inhibitors, which prevent HIV from entering cells by impeding conformational changes in receptors, acting as antagonists by binding to the HIV receptors, or inhibiting fusion between HIV and host membranes. e second and third classes of antiretroviral medication are nucleoside and non-nucleoside reverse transcriptase inhibitors, which both prevent the enzyme reverse transcriptase from successfully completing DNA polymerization, thus stopping the reverse transcription process that would ordinarily convert the HIV single-stranded RNA into double-stranded DNA. e fourth class contains integrase strand transfer inhibitors, which prevent the HIV enzyme integrase from joining the HIV DNA resulting from reverse transcription with the host DNA. e nal class is HIV protease inhibitors. ese medications limit HIV infection by inhibiting the HIV protease enzyme, which is essential to HIV replication (Spach, 2021).
Demographic group disparities are re ected in the distribution of antiretroviral therapy. For instance, women tend to receive later diagnoses despite having more clinical symptoms than men with comparable levels of HIV RNA in their plasma. Generally, HIV+ women access antiretroviral therapy later and die sooner than male HIV patients. Furthermore, HIV+ African Americans are less likely to access antiretroviral therapy than white patients (Sankar & Luborsky, 2003).
Because chronic HIV is o en accompanied by conditions of chronic pain, treating HIV can also involve pain management strategies. Depending

on the patient and their pain severity, chronic pain might be treated using cognitive behavioral therapy, yoga, physical or occupational therapy, hypnosis, acupuncture, medical cannabis, or medications (Bruce et al., 2017). However, HIV treatment can also extend beyond physical needs. For example, antiretroviral therapy in Mombasa, Kenya and in other areas of sub-Saharan Africa emphasizes self-care in addition to the use of medications, particularly highlighting the importance of managing stress (de Klerk & Moyer, 2017). Treatment for HIV/AIDS must incorporate options to address related mental health conditions such as depression in order to truly promote overall patient wellness.
Long-Acting Drugs
As of now, almost all antiretroviral treatments for the HIV virus involve taking medication at least once a day. is is not only exhausting for people living with HIV, but it can also be di cult to maintain. People living with the virus must remain extremely disciplined. Studies have shown that up to 55% of people would prefer not to take medication every day if they had the option to. It also found that 58% of people view taking daily medications as a constant reminder of the virus and many also revealed anxiety and nervousness about the fact that taking the medication could reveal to others that they are infected with HIV (“HIV treatment: Are long-acting therapies the future?”, 2021). us, it is clear that it is both dangerous and inconvenient to require people living with HIV/AIDS to take medication every day for the rest of their lives. Although it has taken scientists long enough to reach this stage, there is hope for a better future without daily medication. Pharmaceutical companies have shi ed their attention to creating long-acting therapies that would eliminate the need for daily medications.
Long-acting antiretroviral (LA ART) therapies are injections that scientists hope can be given to HIV/AIDS patients every 1-2 months. e hope is that these therapies will reduce daily pill burdens for patients, which could increase compliance with treatment regimens. Recently, the FDA approved the rst LA ART: cabotegravir and rilpivirine combination therapy. is treatment consists of 6 intramuscular injections per year ( oueille et al., 2021). Cabotegravir is an HIV integrase inhibitor; this class of drugs work to block the activity of HIV integrase, which e ectively stops HIV from multiplying in the bloodstream. Rilpivirine is a non-nucleoside reverse transcriptase inhibitor (NNRTI); NNRTIs bind to the HIV-1 reverse transcriptase and block its activity, which prevents HIV from replicating. Together, the drugs work to decrease the amount of HIV in a patient’s blood (Cabotegravir and Rilpivirine Injections, n.d.). Other companies are looking into developing more LA ARTs in the future.
Broadly Neutralizing Antibodies
Broadly neutralizing HIV-1 antibodies (bNAbs) are a nascent approach to treating individuals infected with HIV, as conventional antiretroviral therapy only slows down replication of the virus in the body and is not curative (Liu et al., 2020). bNAbs have shown potential in treating and even eliminating HIV infection, with the additional advantages of higher safety and activating the host immune response.
bNAbs function by binding to speci c sequences,
Image Source: Wikimedia Commons
called epitopes, of viral surface envelope proteins that the HIV virus uses to bind to host cell receptors to gain entry into host cells (Kumar et al., 2018). By binding to these proteins that facilitate cell entry, bNAbs can stop infection at the very beginning, inoculating healthy patients from potential HIV infection and stopping viral spread in infected patients. bNAbs are advantageous because unlike traditional neutralizing antibodies, these broadly neutralizing antibodies bind to conserved epitopes among a diverse collection of genotypically di erent HIV viruses, ensuring that the mutagenic nature of the virus do not a ect the long-term e cacy of the treatment (Rusert et al., 2016).
While generally, bNAbs are ine ective in curing existing HIV infection, recent studies have demonstrated that bNAbs can destroy latent reservoirs of HIV by recruiting e ector cells. For example, next-generation bNAbs like VRC01 and PGT121 can recruit immune cells to block HIV replication in infected cells in the viral reservoir (Halper-Stromberg & Nussenzweig, 2016). Others like 3BNC117 can kill latently infected cells because they bind to epitopes of viral envelope proteins expressed on the surface of the cell membrane of infected host cells, triggering the recruitment of natural killer cells. By neutralizing these viral reservoirs, bNAb treatment in combination with other chemodrugs can signi cantly reduce viral load and the frequency and probability of viral rebound (Chun et al., 2014).
While cell entry is an important facet to address, HIV infection is much more robust and signi cant in cell-to-cell transmission. is perhaps explains why some bNAbs tested in vitro neutralized the majority of free viruses, but in vivo, these antibodies only had a small or moderate e ect in suppressing viremia (Malbec et al., 2013). bNAbs also hold much potential to be modi ed and improved in terms of e cacy, longevity, and safety. For example, modifying the Fc receptor – a receptor involved in antigen recognition – of bNAbs has been shown to prolong the duration of the immunoglobulin serum in humans. Bispeci c and trispeci c bNAbs have recently been at the forefront of research for their ability to signi cantly arrest viral transfer and infection because they can bind to both the HIV envelope protein and surface molecules of highly permissive and vulnerable cellular targets of HIV (Monte ori, 2016).
Overall, these broadly neutralizing antibodies are paving the way for a new avenue of cutting-edge research as more potent and broad monoclonal antibodies are being harvested from infected individuals. ese antibodies have the potential to both halt HIV spread in the body and destroy active reservoirs of HIV, serving as an expansive new therapeutic eld of virological and epidemiological research.
Conclusion
HIV/AIDS has been a devastating American epidemic for the past several decades. With better understanding of its structural biology and impacts on the immune system, in combination with advances made in modern day medicine, new treatments may prove to be e ective in treating the disease. Despite the progress made, there are still limitations in all of the treatments available for the disease today that prevent patients with HIV/AIDS from living their lives completely normally. Hopefully, with more work done in the future, a true cure to HIV/AIDS will be found.
References
10 ings to Know About HIV Suppression | NIH: National Institute of Allergy and Infectious Diseases. (n.d.). Retrieved July 17, 2021, from https://www.niaid.nih.gov/ diseases-conditions/10-things-know-about-hivsuppression
A Mixed Methods Evaluation of an Inclusive Sexual History Taking and HIV Prevention Curriculum for Trainees | SpringerLink. (n.d.). Retrieved November 14, 2021, from https:// link.springer.com/article/10.1007/s11606-01904958-z
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020a). Antigen Capture and Presentation to Lymphocytes. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 51–73). Elsevier.
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020b). E ector Mechanisms of Humoral Immunity. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 158–177). Elsevier.
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020c). E ector Mechanisms of T-Cell Mediated Immunity. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 119–137). Elsevier.
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020d). Humoral Immune Responses. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 137–158). Elsevier.
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020e). Innate Immunity. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 23–51). Elsevier.
Abbas, A. K., Lichtman, A. H., Pillai, S., Baker, D. L., & Baker, A. (2020f). T Cell-Mediated Immunity. In Basic immunology: Functions and disorders of the immune system (6th edition, pp. 96–119). Elsevier.
About HIV/AIDS | HIV Basics | HIV/AIDS | CDC. (2021, June 1). https://www.cdc.gov/hiv/ basics/whatishiv.html
Abuse, N. I. on D. (2020, July 30). Drug Use and Viral Infections (HIV, Hepatitis) DrugFacts. National Institute on Drug Abuse. https://www. drugabuse.gov/publications/drugfacts/drug-useviral-infections-hiv-hepatitis
Abuse, N. I. on D. (--). Who Is at Risk for HIV Infection and Which Populations Are Most A ected? National Institute on Drug Abuse. https://www.drugabuse.gov/publications/ research-reports/hivaids/who-risk-hivinfection-which-populations-are-most-a ected
Agerberth, B., & Guðmundsson, G. H. (2006). Host Antimicrobial Defence Peptides in Human Disease. In W. M. Shafer (Ed.), Antimicrobial Peptides and Human Disease (pp. 67–90). Springer. https://doi.org/10.1007/3-540-299165_3
Ayala, G., & Spieldenner, A. (2021). HIV Is a Story First Written on the Bodies of Gay and Bisexual Men. American Journal of Public Health, 111(7), 1240–1242. https://doi.org/10.2105/ AJPH.2021.306348
Boyton, R. J., & Openshaw, P. J. (2002). Pulmonary defences to acute respiratory infection. British Medical Bulletin, 61(1), 1–12. https://doi. org/10.1093/bmb/61.1.1
Broadly neutralizing antibodies that inhibit HIV-1 cell to cell transmission | Journal of Experimental Medicine | Rockefeller University Press. (n.d.). Retrieved July 12, 2021, from https://rupress. org/jem/article/210/13/2813/41501/BroadlyBruce, R. D., Merlin, J., Lum, P. J., Ahmed, E., Alexander, C., Corbett, A. H., Foley, K., Leonard, K., Treisman, G. J., & Selwyn, P. (2017). 2017 HIVMA of IDSA Clinical Practice Guideline for the Management of Chronic Pain in Patients Living With HIV. Clinical Infectious Diseases, 65(10), e1–e37. https://doi.org/10.1093/cid/ cix636
Bu alo, C. Z., Iwamoto, Y., Hurley, J. H., & Ren, X. (n.d.). How HIV Nef Proteins Hijack Membrane Tra c To Promote Infection. Journal of Virology, 93(24), e01322-19. https://doi.org/10.1128/ JVI.01322-19
Bu alo, C. Z., Iwamoto, Y., Hurley, J. H., & Ren, X. (2019). How HIV Nef Proteins Hijack Membrane Tra c To Promote Infection. Journal of Virology, 93(24), e01322-19. https://doi.org/10.1128/ JVI.01322-19
Cabotegravir and Rilpivirine Injections: MedlinePlus Drug Information. (n.d.). Retrieved November 14, 2021, from https://medlineplus. gov/druginfo/meds/a621009.html
Cen, S., Khorchid, A., Javanbakht, H., Gabor, J., Stello, T., Shiba, K., Musier-Forsyth, K., & Kleiman, L. (2001). Incorporation of lysyltRNA synthetase into human immunode ciency virus type 1. Journal of Virology, 75(11), 5043–5048. https://doi.org/10.1128/JVI.75.11.50435048.2001
Chinen, J., & Shearer, W. T. (2002). Molecular virology and immunology of HIV infection. Journal of Allergy and Clinical Immunology, 110(2), 189–198. https://doi.org/10.1067/ mai.2002.126226
Chu, C., & Selwyn, P. A. (2010). Diagnosis and initial management of acute HIV infection. American Family Physician, 81(10), 1239–1244.
Chun, T.-W., Murray, D., Justement, J. S., Blazkova, J., Hallahan, C. W., Fankuchen, O., Gittens, K., Benko, E., Kovacs, C., Moir, S., & Fauci, A. S. (2014). Broadly neutralizing antibodies suppress HIV in the persistent viral reservoir. 111(36), 13151–13156.
Cohen, M. S., Shaw, G. M., McMichael, A. J., & Haynes, B. F. (2011). Acute HIV-1 Infection. e New England Journal of Medicine, 364(20), 1943–1954. https://doi.org/10.1056/NEJMra1011874
Core Concepts—Antiretroviral Medications and Initial erapy—Antiretroviral erapy— National HIV Curriculum. (n.d.). Retrieved July 17, 2021, from https://www.hiv.uw.edu/go/ antiretroviral-therapy/general-information/coreconcept/all
Economically Disadvantaged | HIV by Group | HIV/AIDS | CDC. (2019, December 11). https:// www.cdc.gov/hiv/group/poverty.html
Emanuele. (2010). HIV virology and pathogenic mechanisms of infection: A brief overview. Ann Ist Super Santia, 46(1), 5–14. https://doi. org/10.4415/ANN_10_01_02
Emerman, M., & Malim, M. H. (1998). HIV1 Regulatory/Accessory Genes: Keys to Unraveling Viral and Host Cell Biology. Science, 280(5371), 1880–1884. https://doi.org/10.1126/ science.280.5371.1880
Engelman, A., & Cherepanov, P. (2012). e structural biology of HIV-1: Mechanistic and therapeutic insights. Nature Reviews. Microbiology, 10(4), 279–290. https://doi. org/10.1038/nrmicro2747
Fanales-Belasio, E., Raimondo, M., Suligoi, B., & Buttò, S. (2010). HIV virology and pathogenetic mechanisms of infection: A brief overview. Annali Dell’Istituto Superiore Di Sanita, 46(1), 5–14. https://doi.org/10.4415/ANN_10_01_02
Frasca, K., Castillo-Mancilla, J., McNulty, M. C., Connors, S., Sweitzer, E., Zimmer, S., & Madinger, N. (2019). A Mixed Methods Evaluation of an Inclusive Sexual History Taking and HIV Prevention Curriculum for Trainees. Journal of General Internal Medicine, 34(7), 1279–1288. https://doi.org/10.1007/s11606-019-04958-z
Gelderblom, H. R., zel, M., & Pauli, G. (1989). Morphogenesis and morphology of HIV structure-function relations. Archives of Virology, 106(1–2), 1–13. https://doi. org/10.1007/BF01311033
Goldman, D. P., & Bao, Y. (2004). E ective HIV Treatment and the Employment of HIV+ Adults. Health Services Research, 39(6p1), 1691–1712. https://doi.org/10.1111/j.14756773.2004.00313.x
Gonzalez, J. S., Hendriksen, E. S., Collins, E. M., Durán, R. E., & Safren, S. A. (2009). Latinos and HIV/AIDS: Examining Factors Related to Disparity and Identifying Opportunities for Psychosocial Intervention Research. AIDS and Behavior, 13(3), 582–602. https://doi. org/10.1007/s10461-008-9402-4
Goodin, B. R., Owens, M. A., White, D. M., Strath, L. J., Gonzalez, C., Rainey, R. L., Okunbor, J. I., Heath, S. L., Turan, J. M., & Merlin, J. S. (2018). Intersectional health-related stigma in persons living with HIV and chronic pain: Implications for depressive symptoms. AIDS Care, 30(sup2), 66–73. https://doi.org/10.1080/09540121.2018.1 468012
Greene, W. C. (2007). A history of AIDS: Looking back to see ahead. European Journal of Immunology, 37(S1), S94–S102. https://doi. org/10.1002/eji.200737441
HIV and Substance Use | HIV Transmission | HIV Basics | HIV/AIDS | CDC. (2021, April 21). https://www.cdc.gov/hiv/basics/hivtransmission/substance-use.html
HIV in the United States and Dependent Areas | Statistics Overview | Statistics Center | HIV/ AIDS | CDC. (2021, August 9). https://www.cdc. gov/hiv/statistics/overview/ataglance.html
HIV treatment: Are long-acting therapies the future? (n.d.). Retrieved December 6, 2021, from https://www.pharmaceutical-technology.com/ features/hiv-treatment-long-acting-therapiesfuture/
HIV Treatment, the Viral Reservoir, and HIV DNA | NIH: National Institute of Allergy and Infectious Diseases. (n.d.). Retrieved July 17, 2021, from https://www.niaid.nih.gov/diseasesconditions/hiv-treatment-viral-reservoir-hivdna
HIV/AIDS and Socioeconomic Status. (n.d.). Https://Www.Apa.Org. Retrieved September 1, 2021, from https://www.apa.org/pi/ses/resources/ publications/hiv-aids
Hoenigl, M., Green, N., Camacho, M., Gianella, S., Mehta, S. R., Smith, D. M., & Little, S. J. (2016). Signs or Symptoms of Acute HIV Infection in a Cohort Undergoing Community-Based Screening. Emerging Infectious Diseases, 22(3), 532–534. https://doi.org/10.3201/eid2203.151607
Human Immunode ciency Virus (HIV). (2016). Transfusion Medicine and Hemotherapy, 43(3), 203–222. https://doi.org/10.1159/000445852
Janeway, C. A. Jr., Shlomchik, M. J., Walport, M.,
& Travers, P. (2001). T Cell Mediated Immunity. In C. Janeway (Ed.), Immunobiology: e immune system in health and disease (5. ed, pp. 295–341). Garland Publ. [u.a.].
Janeway, C. A. Jr., Travers, P., Walport, M., & Shlomchik, M. J. (2001a). Adaptive Immunity to Infection. In C. Janeway (Ed.), Immunobiology: e immune system in health and disease (5. ed, pp. 381–425). Garland Publ. [u.a.].
Janeway, C. A. Jr., Travers, P., Walport, M., & Shlomchik, M. J. (2001b). Innate Immunity. In C. Janeway (Ed.), Immunobiology (5th ed., pp. 35–93). Garland Science.
Janeway, C. A. Jr., Travers, P., Walport, M., & Shlomchik, M. J. (2001c). e Humoral Immune Response. In C. Janeway (Ed.), Immunobiology: e immune system in health and disease (5. ed, pp. 341–381). Garland Publ. [u.a.].
JCI - Towards HIV-1 remission: Potential roles for broadly neutralizing antibodies. (n.d.). Retrieved July 12, 2021, from https://www.jci.org/articles/ view/80561
Kim, B., & Aronowitz, T. (2019). Invisible Minority: HIV Prevention Health Policy for the Asian American Population. Policy, Politics, & Nursing Practice, 20(1), 41–49. https://doi. org/10.1177/1527154419828843
Klerk, J. de, & Moyer, E. (2017). “A Body Like a Baby”: Social Self-Care among Older People with Chronic HIV in Mombasa. Medical Anthropology, 36(4), 305–318. https://doi.org/10 .1080/01459740.2016.1235573
Kumar, R., Qureshi, H., Deshpande, S., & Bhattacharya, J. (2018). Broadly neutralizing antibodies in HIV-1 treatment and prevention. erapeutic Advances in Vaccines and Immunotherapy, 6(4), 61–68. https://doi. org/10.1177/2515135518800689
Kwong, P. D., Wyatt, R., Robinson, J., Sweet, R. W., Sodroski, J., & Hendrickson, W. A. (1998). Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature, 393(6686), 648–659. https://doi.org/10.1038/31405
Lindqvist, M., van Lunzen, J., Soghoian, D. Z., Kuhl, B. D., Ranasinghe, S., Kranias, G., Flanders, M. D., Cutler, S., Yudanin, N., Muller, M. I., Davis, I., Farber, D., Hartjen, P., Haag, F., Alter, G., zur Wiesch, J. S., & Streeck, H. (2012). Expansion of HIV-speci c T follicular helper cells in chronic HIV infection. Journal of Clinical Investigation, 122(9), 3271–3280.
Liu, Y., Cao, W., Sun, M., & Li, T. (2020). Broadly neutralizing antibodies for HIV-1: E cacies, challenges and opportunities. Emerging Microbes & Infections, 9(1), 194–206. https://doi.org/10.10 80/22221751.2020.1713707
Lorenc, A., Ananthavarathan, P., Lorigan, J., Banarsee, R., Jowata, M., & Brook, G. (2014). e prevalence of comorbidities among people living with HIV in Brent: A diverse London Borough. London Journal of Primary Care, 6(4), 84–90. https://doi.org/10.1080/17571472.2014.1149342 2
Miller, R. H., & Sarver, N. (1997). HIV accessory proteins as therapeutic targets. Nature Medicine, 3(4), 389–394. https://doi.org/10.1038/nm0497389
Monte ori, D. C. (2016). Bispeci c Antibodies Against HIV. Cell, 165(7), 1563–1564. https:// doi.org/10.1016/j.cell.2016.06.004
Moore, R. D. (2011). Epidemiology of HIV Infection in the United States: Implications for Linkage to Care. Clinical Infectious Diseases: An O cial Publication of the Infectious Diseases Society of America, 52(Suppl 2), S208–S213. https://doi.org/10.1093/cid/ciq044
Nakayama-Hosoya, K., Ishida, T., Youngblood, B., Nakamura, H., Hosoya, N., Koga, M., Koibuchi, T., Iwamoto, A., & Kawana-Tachikawa, A. (2015). Epigenetic Repression of Interleukin 2 Expression in Senescent CD4+ T Cells During Chronic HIV Type 1 Infection. e Journal of Infectious Diseases, 211(1), 28–39. https://doi. org/10.1093/infdis/jiu376
Negin, J., Aspin, C., Gadsden, T., & Reading, C. (2015). HIV Among Indigenous peoples: A Review of the Literature on HIV-Related Behaviour Since the Beginning of the Epidemic. AIDS and Behavior, 19(9), 1720–1734. https:// doi.org/10.1007/s10461-015-1023-0
Nisole, S., & Saïb, A. (2004). Early steps of retrovirus replicative cycle. Retrovirology, 1, 9. https://doi.org/10.1186/1742-4690-1-9
Norman, L. R., Basso, M., Kumar, A., & Malow, R. (2009). Neuropsychological consequences of HIV and substance abuse: A literature review and implications for treatment and future research.
Current Drug Abuse Reviews, 2(2), 143–156. https://doi.org/10.2174/1874473710902020143
O’Connell, P., Pepelyayeva, Y., Blake, M. K., Hyslop, S., Crawford, R. B., Rizzo, M. D., Pereira-Hicks, C., Godbehere, S., Dale, L., Gulick, P., Kaminski, N. E., Amal tano, A., & Aldhamen, Y. A. (2019). SLAMF7 Is a Critical Negative Regulator of IFN-α–Mediated CXCL10 Production in Chronic HIV Infection. e Journal of Immunology, 202(1), 228–238. https:// doi.org/10.4049/jimmunol.1800847
Parada, C. A., & Roeder, R. G. (1996). Enhanced processivity of RNA polymerase II triggered by Tat-induced phosphorylation of its carboxyterminal domain. Nature, 384(6607), 375–378. https://doi.org/10.1038/384375a0
Popescu, I., Drummond, M. B., Gama, L., Coon, T., Merlo, C. A., Wise, R. A., Clements, J. E., Kirk, G. D., & McDyer, J. F. (2014). Activation-induced Cell Death Drives Profound Lung CD4^sup +^ T-Cell Depletion in HIV-associated Chronic Obstructive Pulmonary Disease. American Journal of Respiratory and Critical Care Medicine, 190(7), 744–755.
Rajasuriar, R., Khoury, G., Kamarulzaman, A., French, M. A., Cameron, P. U., & Lewin, S. R. (2013). Persistent immune activation in chronic HIV infection: Do any interventions work? AIDS, 27(8), 1199–1208. https://doi.org/10.1097/ QAD.0b013e32835ecb8b
Rusert, P., Kouyos, R. D., Kadelka, C., Ebner, H., Schanz, M., Huber, M., Braun, D. L., Hozé, N., Scherrer, A., Magnus, C., Weber, J., Uhr, T., Cippa, V., orball, C. W., Kuster, H., Cavassini, M., Bernasconi, E., Ho mann, M., Calmy, A., … Trkola, A. (2016). Determinants of HIV1 broadly neutralizing antibody induction. Nature Medicine, 22(11), 1260–1267. https://doi. org/10.1038/nm.4187
Sankar, A., & Luborsky, M. (2003). Developing a community-based de nition of needs for persons living with chronic HIV. Human Organization, 62(2), 153–165. http://dx.doi.org.dartmouth.idm. oclc.org/10.17730/humo.62.2.695j11t5pmpmljr2
Simon, F., Mauclère, P., Roques, P., LoussertAjaka, I., Müller-Trutwin, M. C., Saragosti, S., Georges-Courbot, M. C., Barré-Sinoussi, F., & Brun-Vézinet, F. (1998). Identi cation of a new human immunode ciency virus type 1 distinct from group M and group O. Nature Medicine, 4(9), 1032–1037. https://doi.org/10.1038/2017 Singh, G. K., Azuine, R. E., & Siahpush, M. (2013). Widening Socioeconomic, Racial, and Geographic Disparities in HIV/AIDS Mortality in the United States, 1987–2011. Advances in Preventive Medicine, 2013, e657961. https://doi. org/10.1155/2013/657961
Smith, J. L. (2003). e Role of Gastric Acid in Preventing Foodborne Disease and How Bacteria Overcome Acid Conditions†. Journal of Food Protection, 66(7), 1292–1303. https://doi. org/10.4315/0362-028X-66.7.1292
Tantillo, C., Ding, J., Jacobo-Molina, A., Nanni, R. G., Boyer, P. L., Hughes, S. H., Pauwels, R., Andries, K., Janssen, P. A. J., & Arnold, E. (1994). Locations of Anti-AIDS Drug Binding Sites and Resistance Mutations in the ree-dimensional Structure of HIV-1 Reverse Transcriptase. Journal of Molecular Biology, 243(3), 369–387. https://doi.org/10.1006/jmbi.1994.1665
e Structural Biology of HIV. (n.d.). Retrieved November 14, 2021, from https://cdn.rcsb.org/ pdb101/learn/resources/structural-biology-ofhiv/index.html
e structural biology of HIV-1: Mechanistic and therapeutic insights | Nature Reviews Microbiology. (n.d.). Retrieved November 14, 2021, from https://www.nature.com/articles/ nrmicro2747
oueille, P., Choong, E., Cavassini, M., Buclin, T., & Decosterd, L. A. (2021). Long-acting antiretrovirals: A new era for the management and prevention of HIV infection. Journal of Antimicrobial Chemotherapy, dkab324. https:// doi.org/10.1093/jac/dkab324
Tran, B. X., Hwang, J., Nguyen, L. H., Nguyen, A. T., Latkin, N. R. K., Tran, N. K., uc, V. T. M., Nguyen, H. L. T., Phan, H. T. T., Le, H. T., Tran, T. D., & Latkin, C. A. (2016). Impact of Socioeconomic Inequality on Access, Adherence, and Outcomes of Antiretroviral Treatment Services for People Living with HIV/AIDS in Vietnam. PLOS ONE, 11(12), e0168687. https:// doi.org/10.1371/journal.pone.0168687
Tritel, M., & Resh, M. D. (2001). e late stage of human immunode ciency virus type 1 assembly is an energy-dependent process. Journal of Virology, 75(12), 5473–5481. https://doi. org/10.1128/JVI.75.12.5473-5481.2001
van Grevenynghe, J., Cubas, R. A., Noto, A.,
DaFonseca, S., He, Z., Peretz, Y., Filali-Mouhim, A., Dupuy, F. P., Procopio, F. A., Chomont, N., Balderas, R. S., Said, E. A., Boulassel, M.R., Tremblay, C. L., Routy, J.-P., Sékaly, R.-P., & Haddad, E. K. (2011). Loss of memory B cells during chronic HIV infection is driven by Foxo3a- and TRAIL-mediated apoptosis. Journal of Clinical Investigation, 121(10), 3877–3888.
Weller, I. V., & Williams, I. G. (2001). ABC of AIDS. Antiretroviral drugs. BMJ (Clinical Research Ed.), 322(7299), 1410–1412. https://doi. org/10.1136/bmj.322.7299.1410
Wheeler, D. P., & Dodd, S.-J. (2011). LGBTQ Capacity Building in Health Care Systems: A Social Work Imperative. Health & Social Work, 36(4), 307–309.
Wilson, C., & Cariola, L. A. (2020). LGBTQI+ Youth and Mental Health: A Systematic Review of Qualitative Research. Adolescent Research Review, 5(2), 187–211. https://doi.org/10.1007/ s40894-019-00118-w
Wyatt, R., Kwong, P. D., Desjardins, E., Sweet, R. W., Robinson, J., Hendrickson, W. A., & Sodroski, J. G. (1998). e antigenic structure of the HIV gp120 envelope glycoprotein. Nature, 393(6686), 705–711. https://doi.org/10.1038/31514
Zhang, S., Feng, Y., Narayan, O., & Zhao, L. J. (2001). Cytoplasmic retention of HIV1 regulatory protein Vpr by protein-protein interaction with a novel human cytoplasmic protein VprBP. Gene, 263(1–2), 131–140. https:// doi.org/10.1016/s0378-1119(00)00583-7

e Psychedelic Renaissance
STAFF WRITERS: CAROLINE CONWAY ‘24, BENJAMIN BARRIS ‘25, ZACHARY OJAKLI ‘25, ANDREA CAVANAGH ‘24, EVAN BLOCH ‘24, ETHAN LITMANS ‘24, SHAWN YOON ‘25, NATHAN THOMPSON ‘25, ELAINE PU ‘25, DAVID VARGAS ‘23, BROOKLYN SCHROEDER ‘22
TEAM LEADS: DEV KAPADIA '23, DANIEL CHO '22
Cover Image: An image of psilocybin, also known as magic mushrooms; these are known to have hallucinogenic e ects on those who ingest the mushroom, which has been observed to have clinical bene ts.
Image Source: Wikimedia Commons Introduction
e usage of psychedelics has garnered wider acceptance in the eyes of the public recently, especially during the a ermath of the COVID-19 pandemic. With the increase in mental-health diagnoses because of quarantine and increasing positive support behind psychedelics, clinicians have looked to psychedelics as an e ective means of treating patients (Nochaiwong et al., 2021). Physicians have also recently found promising e ects of psilocybin (psychedelic mushrooms that have become the face of psychedelic culture) and other psychedelic drugs as a means of treating diabetes and heart disease.
Psychedelics are a class of drug that have psychoactive e ects on users, including changes in perception, mood, and cognitive processes. Some psychedelics can be found in nature, like psilocybin or ayahuasca, while others are made synthetically in labs, like lysergic acid diethylamide (LSD). In the US, the height of psychedelics came in the 1960s during the counterculture movement, though this period was also the reason of why many of these drugs were pushed to the top of regulators’ hit lists at the onset of the war on drugs. And for many nonWestern cultures, psychedelic plants have been used as sacramental tools for thousands of years, shaping the course of many established religions. However, these drugs have demonstrated clinical bene ts and a much-improved safety pro le relative to other illicit drugs. Some psychedelics are now only considered Schedule III drugs, meaning moderate to low physical dependence or psychological dependence. As such, biotech companies have started to experiment not only with psychedelics themselves but also many of the pathways that the drugs target in the body, blossoming what is becoming a Psychedelic Renaissance (Cohen, 2021). For example, for mental health treatment, recent studies from Alan Davis et al. of John Hopkins University found that psilocybin-assisted therapy was up to two times more e ective than psychotherapy, and four times more e ective than traditional prescribed antidepressants. Out of a randomized clinical trial of 24 participants, over 71% of their participants showed improvement with clinically signi cant decreases in mental health symptoms as evaluated by blinded clinician rater-assessed depression severity and self-reported secondary outcomes. (Davis et al., 2020).

further than just mental health. To test the e ects of psychedelics on disease reduction, Otto Simonsson and his team at the University of Oxford studied the e ects of psychedelics on cardiometabolic health, analyzing data from 375,000 participants in the National Survey on Drug Use and Health. ey found that users of psychedelics showed a 2.2% reduced chance of heart disease and a 3.75% reduced chance of diabetes, indicating that psychedelics could potentially be used as an e ective means of treatment for patients su ering from other cardiometabolic diseases as well (Simonsson, 2021).
ere are several legal obstacles that lie in the way of allowing psychedelics to become a mainstream medicinal practice. Despite the FDA listing psilocybin as a “breakthrough therapy” in 2019, as of 2021, the U.S. Congress has not yet made funds available for research dedicated to psychedelic usage. Currently, only Oregon is pursuing the administration of psychedelic services with licensing by the Oregon Health Authority through licensing by the Oregon Psilocybin Advisory Board (Marks & Cohen, 2021).
With this in mind, we aim to give an analysis on the history, science, cultural context, and ethics surrounding psychedelics.
History of Psychedelics
Today, many people might recoil at the idea of using psychedelics medically. In the words of Sessa (2007), psychedelics have been “demonised in the West” since the 1960s. However, the use of psychedelics is nothing new. Psychedelics have an especially well-documented history in religious contexts. For instance, early Sanskrit texts reference an Indian fermented psychoactive juice called “soma,” and records indicate that Athenian philosopher Plato consumed hallucinogenic fungus for the Eleusinian ceremonies, secret religious rites tied to the cult of Demeter and Persephone, in the 5th century BCE (Sessa, 2007).
Psychedelics were thrust into the Western medicinal spotlight with the initial synthesis of lysergic acid diethylamide (LSD) in 1943. Psychologists and psychiatrists began to investigate LSD’s potential therapeutic e ects in the treatment of mood disorders and alcoholism. Due to this surge in psychedelic-driven research throughout the 1950s, estimates suggest that tens of thousands of people were treated with psychedelics over a 15-year-period (Gardner et al., 2019). is was evidenced in the Canadian province of Saskatchewan, where LSD came to be considered the default treatment for alcoholism thanks to the research of Humphry Osmond and Abram Ho er during the 1950s, among others. In attempting a biochemical treatment for alcoholism, Osmond hoped to combat some of its associated stigmas by providing evidence that it was a disease rather than a matter of individual character. Interestingly, Osmond and Ho er were more interested in the subjective experience induced by LSD than in the chemical e ects of the drug itself. Speci cally, they were fascinated by how similar reports of LSD experiences sounded to descriptions of delirium tremens: alcoholic experiences of “hitting bottom” that were fatal 10% of the time but could o en otherwise lead to a turning point in an individual’s struggle with alcoholism. Osmond and Ho er hypothesized that in mimicking the experience of delirium tremens with LSD, they might be able to help patients reach turning points in their disease without risking their lives. Overall, they found that one-time treatment with LSD aided in recovery from alcoholism roughly 50% of the time (Dyck, 2006). More recent metaanalyses reviewing studies done in the 1950s and 1960s on LSD as an alcoholism treatment support the claim that LSD can have a positive impact on patients, and meta-analyses covering research from 1949 to 1973 on psychedelic treatments of mood disorders have found that of 423 individuals across 19 studies, 79% of patients showed clinical improvement. However, many such psychedelic studies were broadly criticized for failing to adhere to conventional empirical standards, like the inclusion of control conditions (Carhart-Harris & Goodwin, 2017).
It is worth noting that many researchers investigated the e ects of psychedelic substances
Image 1: Molecular structure of lysergic acid diethylamide (LSD), the drug that thrust psychedelics into the spotlight of public opinion. is drug was one of the most frequently used recreational drugs during the 1960s counterculture movement by the youth of the time. Image Source: Wikimedia Commons
not only to test treatments for various clinical conditions but also to study the general limits of the human mind, since drugs like LSD were thought to remove mental barriers against common mental predispositions enabling people to make new thoughts (Elcock, 2021). Perhaps this curiosity was partially responsible for the surge of recreational LSD use in the 1960s. Regardless of the origins of recreational use, it had dire consequences for the position of psychedelic treatments in the medical community. By 1965, psychedelics were prohibited within the United States, and the 1970 Controlled Substances Act placed both LSD and psilocybin in the most restrictive category of drugs. Similar changes followed abroad, and psychedelic-driven studies ceased (Gardner et al., 2019).
However, the psychedelic story in medicine is far from over. e 1990s and 2000s witnessed something of a renaissance in psychedelic research on healthy individuals and those with mood disorders or addictions. With shi s in modern attitudes about other controlled substances—like cannabis—and about the United States’ war on drugs, medicinal psychedelics are receiving renewed attention (Gardner et al., 2019). Sessa (2007) notes that this is especially true considering the development of the psychedelic substance MDMA (3,4-Methylenedioxymethamphetamine). While the legal status of psychedelics remains a barrier to widespread psychedelic-based treatment, the tide seems to be turning in favor of psychedelicdriven medical research.
Cultural Context of Psychedelics
Before psychedelics were adopted into modern Western societies, indigenous cultures used psychedelics for both spiritual and health reasons as the plants from which certain psychedelics are derived were thought to possess sacred healing properties. Many of these cultures continue to use psychedelics today. As aforementioned, South American Indigenous societies use Ayahuasca for various spiritual ceremonies and rituals. And, Mexico has bolstered its psychedelic tourism sector as more tourists arrive to Mexico to experience novel encounters of consuming psychedelic plants and substances. Aside from Mexican and South American Indigenous societies, other civilizations worldwide have utilized psychedelics for religious purposes. In ancient Indian religion, psychedelics played a crucial role as individuals consumed soma, a psychedelic plant, as a ritualistic practice (Williams et al., 2020). In addition, historians believed that various religious gures found in the Old Testament of the Bible used psychedelics as a means of divine communication. African cultures have also been heavily in uenced by psychedelic use, as in Western African and Ethiopian societies, plants like Çaate (Katha edulis) were a central xture in healing practices and were believed to ward o evil. Ubulawu, a healing foam created from grinding several plant roots together, speci cally has widespread usage in southern Africa for traditional medicine, ancestral communication, and shamanic practices (Williams et al., 2020).
In the United States, the use of psychedelics was woven into a more politically motivated narrative in the late 60s and early 70s. Nixon’s presidential term saw the criminalization of Black individuals through the War on Drugs, though the racial group used drugs at a lower rate than other racial groups. Groups that opposed the ongoing Vietnam War, like Hippies, were told their opinions were in uenced using psychedelics like LSD. Psychedelics also had a profound in uence on the arts, with music, literature, and visual art featuring new genres and styles inspired by certain drugs. Since the 1960s, the stigma around psychedelics has signi cantly decreased, paving the way for its emerging usage in medicine (Williams, 2018).
During the 1950’s and 1960’s, psychedelics were popular and widely accepted for their promising e ects. More than 40,000 were administered LSD before it became illegal due to the misuse of psychedelics by high-pro le researchers, lack of knowledge, and a rise of conservatism. However, there has been a resurgence of interest in the eld of psychedelics for legally sanctioned research in the twenty- rst century. ere are several reasons this has happened. Over the past couple decades, there has been little improvement in the research of improving depression and anxiety, and psychedelics prove a potential route to explore (dos Santos et al., 2021). Widespread media coverage on this topic has been generally positive, which has gained political and public acceptance in the topic. is has been coupled with the pro ts and recent large-scale acceptance of cannabis legalization (Petranker et al., 2020). However, it is important to note that any emergence of clinically adverse e ects of psychedelics could quickly turn mainstream media against them again.
ere are various other cultural factors to consider. Apart from the social stigma of psychedelics, another large obstacle is its legal status. As of now, compounds like Psilocybin,
DMT, LSD, and MDMA are classi ed as Schedule I in United Nations Convention on Psychotropic Substances. is decision was originally made in 1971, where they described these psychedelics as not having any therapeutic bene ts and a high abuse/dependency rate (dos Santos et al., 2021).
One breakthrough of psychedelics is that they do not have a high abuse potential. erefore, only small doses of psychedelics are used in current studies as participants are already unlikely to become addicted. In fact, one study involving ritual ayahuasca users reported signi cant reductions in previous drug use rather than any social or psychological measures associated with drug dependence (dos Santos et al., 2021).
While psychedelic drugs can generally be used in trials, they are di cult to conduct due to a lack of funding. Since psychedelics are given in small doses, many pharmaceutical companies are wary of investing and supporting these studies. However, this perception has changed in recent years, and many pharmaceutical companies have emerged to develop psychedelics as prescription medicines (dos Santos et al., 2021). For example, the breakthrough therapy of psilocybin used in major depression has been granted by the US FDA and funded by Usona Institute and Compass Pathways/Atai (dos Santos et al., 2021).
One common misconception is that psychedelics would become universally accessible to anyone needing them, giving the patient full range on when to use them. However, a discussion in the medical community is that doctors should have the capacity to prescribe psychedelics to their patients without having to fail all previous treatments. More research is needed to examine puri ed and synthetic compounds to accurately establish dosing (Allen, 2021).
e recent surge of acceptance in this topic has increased support for legally sanctioned research. e topic of psychedelics within medicine will likely become more widely accepted with positive results for treating resistant depression, anxiety, and substance use disorders. is is a delicate topic though, as most psychedelics (i.e., ecstasy and psilocybin) are considered schedule I, meaning that they have a high potential for abuse and a lack of accepted safety for use under medical supervision, and have developed a negative connotation due to their misuse recreationally.
Science of Psychedelics
Psilocybin Psilocybin, more commonly known as “shrooms,” is one of the more well-known psychedelic drugs, especially considering its current relevance in the medical eld and its potential as a therapeutic agent. Mushrooms containing psilocybin fall under a group of psychoactive fungi (Dasgupta & Wahed, 2014). Chemically, psilocybin is a tryptamine alkaloid that possesses an extra phosphoryloxy group located at the fourth carbon (ChEBI, 2013). e chemical composition of psilocybin is signi cant in its functionality as a psychedelic. Once psilocybin is ingested, it is converted to psilocin through the dephosphorylation of the phosphoryloxy substituent. Physiologically, psilocybin is rapidly dephosphorylated in the body to psilocin, which serves as an agonist for several serotonin receptors.
Psilocybin alters the functioning of the human brain by activating serotonin 5-HT2A receptors. is then triggers increased striatal dopamine concentrations which could explain the correlated increase in euphoria and depersonalization (Vollenweider & Kometer, 2010). e heightened attention toward psilocybin can be attributed to its potential as a therapeutic agent for conditions such as anxiety and depression. rough activation of the serotonin 5-HT2A receptors, there is subsequent modulation of multiple brain regions, including the amygdala, the prefrontal, and limbic regions (Lowe et al., 2021).

MDMA
MDMA is a psychoactive drug whose properties do not fall neatly into a prede ned category, having mixed e ects of both stimulants and hallucinogens. MDMA’s formal name is 3,4-Methylenedioxymethamphetamine, but its common street names are ecstasy and molly. Ecstasy is o en taken for the sudden and intense
Image 2: Molecular structure of psilocybin, a psychoactive fungus and one of the more well-known psychedelic drugs. is drug has been implicated in potential therapeutic e ects by the medical community. Image Source: Wikimedia Commons
high that it yields: a perfect “euphoria.” But many criticisms concern the recreational use of ecstasy, such as its serotonergic neurotoxicity, a phenomenon that leads to a reduction in markers for serotonin across higher brain regions. It has been shown to damage the prefrontal cortex and hippocampus, impairing higher-level functioning, explicit memory, and implicit memory (Blagrove et al., 2011). When animals go through MDMA withdrawal, there exists lower serotonin transporter (SERT) binding, meaning that serotonin activity decreases in the hippocampus (Parrott, 2013a). In humans, MDMA also leads to a cascade of physiological and psychological problems, including tremors, abnormal hormone activity, pain perception, and uctuating mood (Parrott, 2013a). In addition, prenatal exposure to MDMA causes developmental problems which may lead to early death of the fetus.
However, MDMA also has a history of medical uses, such as its rst use in psychotherapy in the early 1900s. In addition, it has been investigated as a potential aid in cancer therapy, as long-term exposure to MDMA causes apoptosis in cultured human cells (Parrott, 2013b). Speci cally, it causes lymphoma cells in vitro to lyse, but only with a concentration so large that it induces even more harmful side e ects; thus, further research is needed to separate its potential uses in cancer treatment with its negative e ects (Wasik et al., 2012).
DMT
DMT (N, N-dimethyltryptamine) is a hallucinogen that is naturally found in plants and animals. One of the most distinctive characteristics of DMT is that its e ects can be felt within minutes and can dissipate in less than an hour (Strassman & Qualls, 1994). Biologically, DMT is formed from tryptophan, an amino acid found in food. First, the enzyme AADC catalyzes the decarboxylation of tryptophan; the INMT-facilitated methylation of the resulting product produces NMT and then DMT (Barker, 2018). DMT can also be synthesized chemically (Cameron & Olson, 2018).
A small molecule, DMT can a ect the brain directly by crossing the blood-brain barrier that protects the brain from pathogens and toxic chemicals in the blood (Cameron & Olson, 2018; Pajouhesh & Lenz, 2005). However, the e ects of this psychedelic, when administered into the veins or muscles, are diminished because monoamine oxidase A (MAO-A) readily metabolizes DMT into indoleacetic acid, dropping it to undetectable concentrations within one hour of administration (Barker, 2018). erefore, DMT is o en administered with monoamine oxidase inhibitor or in a way that can avoid metabolism and allow for the penetration of the blood-brain barrier (Riba et al., 2015). Once in the brain, DMT acts as an agonist to 5-HT2A receptors that play a role in causing hallucinogenic e ects (Barker, 2018). 5-HT2A receptors are the main excitatory receptor subtype for serotonin. However, they may also have an inhibitory e ect on certain areas of the visual and orbitofrontal cortexes. DMT also binds to Sigma-1 receptors, which is speculated to increase production of antistress and antioxidant proteins by aiding in protein folding and defending against structural hypoxia (a state in which oxygen is not available at the tissue level to maintain homeostasis) or oxidative stress (a state caused by an imbalance between production and accumulation of oxygen reactive species) that can damage protein structure (Carbonaro & Gatch, 2016; Szabo et al., 2016). In addition, animal studies suggest that DMT may protect astrocytes, which are glial cells that hold neurons together and facilitate nutrient exchange in the brain (Szabó et al., 2021).
DMT can give rise to a range of sensations in humans. ese include out of body experiences, perception of visual or auditory changes in the environment, thoughts about death and a erlife, and communication with otherworldly beings. ese symptoms may be signs that DMT induces near-death experiences associated with increased value of self and others and reduced anxiety about death (Groth-Marnat & Summers, 1998; Timmermann et al., 2018). DMT may also have adverse e ects, including nausea, increased heart rate, and increased blood pressure (Cameron & Olson, 2018; Strassman & Qualls, 1994). It is di cult to isolate the e ects of DMT because recreational DMT users o en use other substances such as narcotics, depressants, and alcohol (Cakic et al., 2010; Cameron & Olson, 2018). However, studies on religious users of this psychedelic through the consumption of ayahuasca show that controlled use of DMT is non-addictive, relatively safe, and bene cial to mental and physical health by improving mood and cognition (Cameron & Olson, 2018).
Due to its positive e ects on mental wellbeing, DMT is widely researched for its potential as therapeutics in depression, though such studies o en focus on DMT analogs or ayahuasca (Barker, 2018). For example, clinical testing of psilocybin, a compound with similar structure
to DMT, showed potential for the treatment of depression, anxiety, and addiction (Cameron & Olson, 2018). In addition, compared to current antidepressants, which have slower onset and are o en less e ective, a study showed that even one dose of ayahuasca produced long term improvements in depressive symptoms (Osório et al., 2015). Despite these promising results, clinical testing of DMT alone was not approved until 2020. Current clinical trials are focusing on DMT as intervention for depression and major depressive disorder by changing neuroplasticity (D'Souza and Flynn, 2021).
Researchers envision the applications of DMT to extend beyond treatment for mood disorders. As a Sigma-1 agonist, DMT may have the potential to treat neurodegenerative disorders such as Alzheimer’s disease by reducing neuroin ammation and suppressing ER stressrelated apoptosis (Ruscher & Wieloch, 2015; Szabó et al., 2021). e e ectiveness of triptan drugs that have a similar structure to that of DMT in treating migraines also presents the possibility of using nonpsychedelic modi cations of DMT for therapeutic purposes (Cameron & Olson, 2018).
Ayahuasca
Ayahuasca is a psychoactive substance commonly used by Indigenous populations throughout Amazonian regions. Although Brazil signed the Convention on Psychotropic Substances of 1971 in Vienna, Austria, which restricts the usage of DMT and puts the substance under federal control, the Brazilian government has sanctioned the use of ayahuasca for religious use. Its application in shamanic ceremonies and rituals is integral to the experience of the ceremonies themselves; considered a sacrament, ayahuasca is widely used as both a mestizo folk curative and spiritual tool. Ayahuasca is commonly ingested as a tea by boiling the leaves of the Psychotria viridis shrub along with the stalks of the Banisteriopsis caapi vine in water (Labate & Goldstein, 2009).
Ayahuasca is extremely pharmacologically complex compared to most psychedelics. Not only does it contain the psychoactive compound DMT, but it also contains other chemicals known as monoamine oxidase inhibitors, or MAOIs, which block enzymes in the human body commonly responsible for breaking down DMT. In other words, β-carbolines in MAOIs interrupt the process of deamination of DMT by inhibiting gut monoamine oxidase enzymes, thus making the substance orally active and allowing it to A er oral administration, hallucinogenic e ects rst occur 30 to 60 minutes a er intake. ese e ects peak around 60 to 120 minutes a er ingestion, and resolve a er around 250 minutes (Aronson, 2014). While there are occasionally dysphoric reactions, such as nausea, anxiety, disorientation, diarrhea, and vomiting, this is largely uncommon, especially since the substance is administered by practiced shamans. e e ects of ayahuasca have been described as a transcendental circle, in which there is a cycle of experiences consistent across all instances of use/ Subjects rst feel extremely vulnerable with feelings of confusion, paranoia, and possibly fear. While this can be overwhelming, this agitating and fear-inducing state is interrupted by a shi of mood and feeling akin to that of a spiritual experience, as participants feel as though they have connected with the universe and a higher power (Kjellgren et al., 2009). roughout the entire experience, time feels altered, although subjects are aware of their surroundings and can speak.
As aforementioned, DMT was classi ed as a Schedule I substance in the United States during the Convention on Psychotropic Substances of 1971. is classi cation is largely un tting due to its contemporary medicinal use and low potential for abuse. For instance, even though ayahuasca has intense e ects on thought processes, emotion, and perception, the drug does not cause addiction or dependence in a similar manner to commonly abused drugs. As a result, it has been described as having properties and uses of a “nondrug” in a court setting. For example, when the DEA seized ayahuasca from the União Do Vegetal (UDV), a church from Brazil with about 130 members in the United States, the UDV subsequently petitioned the federal government for religious use of the substance through citation of the Religious Freedom Restoration Act of 1993. When the Supreme Court asked for compelling interest that demonstrated the dangers and negative e ects of ayahuasca that would thus justify the obstruction of free exercise of religion, the government was unable to produce su cient evidence. As a result, the Court upheld the sacramental use of ayahuasca under the First Amendment’s free exercise of religion clause (Rainey, 2009).
In South America, ayahuasca has been used in rituals and has served as a form of traditional medicine and psychiatry. It has been tested clinically as a form of therapy due to its strong serotonergic e ects (Frecska et al., 2016). Other "Due to its positive e ects on mental wellbeing, DMT is widely researched for its potential as therapeutics in depression..."
Image 3:Ayahuasca, one of the oldest used psychedelics that is part of the culture of many indigenous populations scattered throughout South America, is commonly ingested through tea made from the boiling of the leaves.
Image Source: Wikimedia Commons progressive e orts in the use of ayahuasca include its application to the treatment of addictions, in which some researchers suggest breaking down taboos against the drug and mimicking the practices of Indigenous peoples (Mabit, 2007). Ayahuasca has strong biological e ects on the user, but with the right supervision and state of mind, a substance addict may see meaningful progress in becoming less dependent on other addictive drugs due to its ability to calm the nervous system through reducing excitotoxicity, in ammation, and oxidative stress, factors that are correlated with neurodegeneration (Dos Santos & Hallak, 2017). Ayahuasca also has been shown to have unique neurological bene ts, including the generation of neurons by reducing oxidative stress and in ammation (Dos Santos & Hallak, 2017). Further, some people struggling with depression who were resistant to customary forms of treatment made remarkable strides in their recovery a er taking just a single dose of ayahuasca (Soler et al., 2016). It has produced similar, incredible results in the recovery of people struggling with anxiety, mood disorders, and PTSD (Dos Santos et al., 2018; Inserra, 2018).
Ayahuasca’s fascinating applications seem boundless. One study found that sustained use of the drug improved mindfulness substantially: a weekly dose for four weeks proved equally as e ective as an eight-week mindfulness-based stress reduction course (Soler et al., 2018). Other research on ayahuasca’s e ects on personality has discovered its advantageous e ects on the user’s mental health, con dence, and optimism (Bouso, 2012). Finally, a remarkable study displayed the strong bene ts ayahuasca can have on self-identity. In interviews with many gay and lesbian people who had been socialized in their communities to view their sexual orientation as unacceptable, researchers found that ayahuasca had incredible, positive e ects on their perceptions of themselves as well as their a rmations of their sexuality (Cavnar, 2014). While these ndings appear to portray the drug in a strictly pragmatic, advantageous manner, it is important to note that methodological bias may contribute to scienti c studies reporting the bene ts of ayahuasca far more o en than its adverse e ects.
Ketamine
Ketamine is a chemically synthesized anesthetic drug with mild hallucinogenic e ects when taken in sub-sedative doses. e drug was originally discovered in the 1960’s and FDA-approved for medical use in 1970 (Pribish et al., 2020). e synthesis process begins with a cyclohexanone molecule treated with a 2-chlorophenyl magnesium bromide reagent and high temperatures in a hydrocarbon solvent resulting in an oxidation reaction that produces ketamine (Pribish et al., 2020). Ketamine functions as a non-competitive antagonist for the NMDAglutamate receptor in the brain, which plays a key role in central sensitization and the transmission of pain signals. By holding the NMDA calcium channel open, glutamate is continually absorbed by postsynaptic neurons, producing the drug’s hallucinogenic e ects (Pribish et al., 2020). Firms like Algernon Pharmaceuticals are investigating drugs targeting the receptor in disease treatment, suggesting that ketamine might have clinical bene ts that are currently being underutilized due to its Schedule III drug status in the United States that gives it a negative light in the public (Pribish et al., 2020).
Despite Ketamine being a Schedule III drug, recent research has discovered that it has numerous medical bene ts. For the past 70 years, ketamine


has been used primarily as an anesthetic drug. Particularly, it can induce dissociation, analgesia (the inability to feel pain), sedation, catalepsy (a trance-like state in which consciousness and feeling are lost), and bronchodilation (the dilation of airways). More recently, though, it has been found that ketamine can be used to treat depression, acute and chronic pain, seizures, headaches, and substance disorders. Furthermore, ketamine has been shown to act as a neuroprotector, which can have a wide variety of positive e ects. Namely, neuroprotectors helps promote the preservation of neuronal structures and/or function that protect against neuronal injury.
Numerous studies have found that ketamine is a promising alternative to traditional antidepressants due to its ability to rapidly onset, be e ective, and yield lasting impacts. A single dose of ketamine has been shown to have an antidepressant e ect, which begins as early as 2 hours a er administration, peaks at 24 hours, and lasts for up to 7–14 days (Shiroma et al., 2020). is has been found to be especially useful for speci c groups, such as soldiers who are especially subject to increased mental and physical trauma and feelings of hopelessness. ese conditions signi cantly decreased a er a single dose. is is most likely due to its ability to enhance the brain’s neuroplasticity. Scientists have also found positive correlations with ketamine’s treatment of other psychological disorders, including anxiety and obsessive-compulsive disorder (Kryst et al., 2020).
Ketamine has also been shown to have similar analgesic e ects to opiates but without the serious risk of addiction that plagues traditional opioids that has led to the opioid problem in the United States (Abdollahpour et al., 2020). Ketamine also has well-documented anti-in ammatory properties by interacting with in ammatory cells recruitment and cytokines production, further contributing to its e cacy. is opens a signi cant opportunity to pivot much of the opiate use towards drugs like ketamine that are less likely to be abused (Abdollahpour et al., 2020).
However, treatment applications are not the only area where ketamine has been observed to be useful. Research has shown that the psychedelic has had utility in the surgical setting; unlike most anesthetics typically used in surgery, ketamine does not shut down breathing re exes, allowing patients to maintain normal patterns. is lets patients anesthetized with ketamine to receive operations without the need for intubation which o en causes throat problems and pain post-op (Sassano-Higgins et al., 2016). However, ketamine does not produce as uniform of an anesthetic response in everyone as compared with other more common anesthetics. is means patients anesthetized with ketamine may stay under for longer or shorter than expected – especially in larger doses – making it di cult to anesthetize with ketamine for long intense operations (Sassano-Higgins et al., 2016).
While ketamine has been shown to have a variety of bene ts, its drawbacks must be considered. Some of the side e ects include cognitive impairments, abdominal pain, liver injury, and dose-dependent urogenital pathology. A er repeated doses, another risk includes its neurotoxicity and long-term episodic and semantic memory impairment. More immediate risks include tachyarrhythmias, which is a rapid abnormal heart rate, hallucinations, and ashbacks. Finally, ketamine has been shown to lead to apoptotic cell death in neurons in the cerebral cortex and hippocampal region, causing long-term de cits in cognitive processing. And so, future research aims to catalogue more accurately both the bene ts and the drawbacks of the drug to nd areas where it can be e cacious and safe; soon, the drug may see its delisting from the schedule III drug list and more widespread acceptance by the medical community (SassanoHiggins et al., 2016).
Ethics of Psychedelics
rough the years, society has had an everevolving view about controlled substances. As the conversation shi ed to allow for greater use of substances, such as a ground-based movement for the legalization of marijuana, psychedelics became the next topic of debate. e premise of this debate was centered around
Image 4: Ketamine, as opposed to psychedelics like ayahuasca and psilocybin, is not a hallucinogenic plant, but a synthesized chemical that has hallucinogenic e ects; nevertheless, despite its synthetic origin, it still has important clinical bene ts, though this same origin has led many to treat the drug like other synthetic street drugs, causing the schedule III drug distinction of ketamine. Image Source: Wikimedia Commons
healthcare applications. Some recent examples of legislators that have taken the step to legalize psychedelics stem from the west coast of the United States. e rst state to take this jump was Oregon, when on November 4th, 2020, through Measure 109, the medical supervised use of psilocybin was approved. is measure was supported by a clear majority of citizens (nearly 60%) and began what has been termed a psychedelic renaissance (Romero, 2021). An extremely recent example of the shi ing views on these chemicals is California’s advancement towards decriminalization of psychedelics for those 21 and older. As a forerunner in marijuana legalization, California may be attempting to take the next steps in discussing drug criminality. While the United States is beginning to discuss the merits of decriminalization, many other countries lack laws that prohibit such activities. Brazil, for example, has no laws or restrictions on psilocybin, and this substance does not have the same taboo that it does in the States. Jamaica, Netherlands, and Portugal are just a few of the many countries in the world that do not have strict laws against the use of psychedelics, providing a variety of templates and examples for how we can create our own laws in the future (Feuer, 2020).
Conclusion
Admittedly, psychedelics are a complicated topic both socially and medically. However, there is a rich history surrounding their advent and usage in many cultures throughout the world. While the status of psychedelics for medicinal/therapeutic uses is hazy given its classi cation as schedule I substances under the Controlled Substances Act, there are clear mental and physiological bene ts that have been documented in studies that use psychedelics as a treatment option. With the current paradigm shi occurring with drugs such as marijuana and e orts from citizens and lawmakers to reschedule such drugs, there is a clear direction that is being paved for a similar rescheduling for psychedelics. While still highly stigmatized, it will be interesting to see the research that comes out regarding psychedelics and to see its status in society in the coming decades.
References
Abdollahpour, A., Sa arieh, E., & Zoroufchi, B. H. (2020). A review on the recent application of ketamine in management of anesthesia, pain, and health care. Journal of Family Medicine and Primary Care, 9(3), 1317–1324. https://doi. Allen, T. (2021, October 8). Psychedelic Science Holds Promise for Mainstream Medicine. University of Nevada, Las Vegas. http://www. unlv.edu/news/release/psychedelic-scienceholds-promise-mainstream-medicine
Aronson, J. K. (2014). 76—Plant Poisons and Traditional Medicines. In J. Farrar, P. J. Hotez, T. Junghanss, G. Kang, D. Lalloo, & N. J. White (Eds.), Manson’s Tropical Infectious Diseases (Twenty-third Edition) (pp. 1128-1150.e6). W.B. Saunders. https://doi.org/10.1016/B978-0-70205101-2.00077-7
Barbosa, P. C. R., Mizumoto, S., Bogenschutz, M. P., & Strassman, R. J. (2012). Health status of ayahuasca users. Drug Testing and Analysis, 4(7–8), 601–609. https://doi.org/10.1002/dta.1383
Barker, S. A. (2018). N, N-Dimethyltryptamine (DMT), an Endogenous Hallucinogen: Past, Present, and Future Research to Determine Its Role and Function. Frontiers in Neuroscience, 12, 536. https://doi.org/10.3389/fnins.2018.00536
Blagrove, M., Seddon, J., George, S., Parrott, A. C., Stickgold, R., Walker, M. P., Jones, K. A., & Morgan, M. J. (2011). Procedural and declarative memory task performance, and the memory consolidation function of sleep, in recent and abstinent ecstasy/ MDMA users. Journal of Psychopharmacology (Oxford, England), 25(4), 465–477. https://doi. org/10.1177/0269881110372545
Bouso, J. C., González, D., Fondevila, S., Cutchet, M., Fernández, X., Barbosa, P. C. R., Alcázar-Córcoles, M. Á., Araújo, W. S., Barbanoj, M. J., Fábregas, J. M., & Riba, J. (2012). Personality, Psychopathology, Life Attitudes and Neuropsychological Performance among Ritual Users of Ayahuasca: A Longitudinal Study. PLOS ONE, 7(8), e42421. https://doi.org/10.1371/ journal.pone.0042421
Cakic, V., Potkonyak, J., & Marshall, A. (2010). Dimethyltryptamine (DMT): Subjective e ects and patterns of use among Australian recreational users. Drug and Alcohol Dependence, 111(1), 30–37. https://doi.org/10.1016/j. drugalcdep.2010.03.015
Cameron, L. P., & Olson, D. E. (2018). Dark Classics in Chemical Neuroscience: N,NDimethyltryptamine (DMT). ACS Chemical Neuroscience, 9(10), 2344–2357. https://doi. org/10.1021/acschemneuro.8b00101
Carbonaro, T. M., & Gatch, M. B. (2016). Neuropharmacology of N,N-dimethyltryptamine. Brain Research Bulletin, 126, 74–88. https://doi. org/10.1016/j.brainresbull.2016.04.016
Carhart-Harris, R. L., & Goodwin, G. M. (2017). e erapeutic Potential of Psychedelic Drugs: Past, Present, and Future. Neuropsychopharmacology, 42(11), 2105–2113. https://doi.org/10.1038/npp.2017.84
Cavnar, C. (2014). e e ects of ayahuasca ritual participation on gay and lesbian identity. Journal of Psychoactive Drugs, 46(3), 252–260. https:// doi.org/10.1080/02791072.2014.920117
Cohen, J. (2021). Psychedelic Drugs Are Moving From e Fringes Of Medicine To e Mainstream. Forbes. https://www.forbes.com/ sites/joshuacohen/2021/07/05/psychedelicdrugs-are-moving-from-the-fringes-ofmedicine-to-the-mainstream/
Dasgupta, A., & Wahed, A. (2014). Chapter 17 - Challenges in Drugs of Abuse Testing: Magic Mushrooms, Peyote Cactus, and Designer Drugs. In Clinical Chemistry, Immunology and Laboratory Quality Control (pp. 307–316). Elsevier. https://doi.org/10.1016/B978-0-12407821-5.00017-6
Davis, A. K., Barrett, F. S., May, D. G., Cosimano, M. P., Sepeda, N. D., Johnson, M. W., Finan, P. H., & Gri ths, R. R. (2021). E ects of PsilocybinAssisted erapy on Major Depressive Disorder: A Randomized Clinical Trial. JAMA Psychiatry, 78(5), 481–489. https://doi.org/10.1001/ jamapsychiatry.2020.3285
dos Santos, R. G., Bouso, J. C., Rocha, J. M., Rossi, G. N., & Hallak, J. E. (2021). e Use of Classic Hallucinogens/Psychedelics in a erapeutic Context: Healthcare Policy Opportunities and Challenges. Risk Management and Healthcare Policy, 14, 901–910. https://doi.org/10.2147/ RMHP.S300656
Dos Santos, R. G., & Hallak, J. E. C. (2017). E ects of the Natural β-Carboline Alkaloid Harmine, a Main Constituent of Ayahuasca, in Memory and in the Hippocampus: A Systematic Literature Review of Preclinical Studies. Journal of Psychoactive Drugs, 49(1), 1–10. https://doi.or g/10.1080/02791072.2016.1260189
Dos Santos, R. G., Osório, F. L., Crippa, J. A. S., Riba, J., Zuardi, A. W., & Hallak, J. E. C. (2016). Antidepressive, anxiolytic, and antiaddictive e ects of ayahuasca, psilocybin and lysergic acid diethylamide (LSD): A systematic review of clinical trials published in the last 25 years. erapeutic Advances in Psychopharmacology, 6(3), 193–213. https://doi. org/10.1177/2045125316638008
D’Souza, D. C., & Flynn, L. T. (2021). Fixed Order, Open-Label, Dose-Escalation Study of DMT in Humans—Tabular View—ClinicalTrials. gov. https://clinicaltrials.gov/ct2/show/record/ NCT04711915
Dyck, E. (2006). ‘Hitting Highs at Rock Bottom’: LSD Treatment for Alcoholism, 1950–1970. Social History of Medicine, 19(2), 313–329. https://doi.org/10.1093/shm/hkl039
Feuer, W. (2020, November 4). Oregon becomes rst state to legalize magic mushrooms as more states ease drug laws in “psychedelic renaissance.” CNBC. https://www.cnbc.com/2020/11/04/ oregon-becomes-first-state-to-legalize-magicmushrooms-as-more-states-ease-drug-laws.html
Frecska, E., Bokor, P., & Winkelman, M. (2016). e erapeutic Potentials of Ayahuasca: Possible E ects against Various Diseases of Civilization. Frontiers in Pharmacology, 7, 35. https://doi. org/10.3389/fphar.2016.00035
Gardner, J., Carter, A., O’Brien, K., & Seear, K. (2019). Psychedelic-assisted therapies: e past, and the need to move forward responsibly. International Journal of Drug Policy, 70, 94–98. https://doi.org/10.1016/j.drugpo.2019.05.019
Groth-Marnat, G., & Summers, R. (1998). Altered Beliefs, Attitudes, and Behaviors Following Near-Death Experiences. Journal of Humanistic Psychology, 38(3), 110–125. https:// doi.org/10.1177/00221678980383005
Inserra, A. (2018). Hypothesis: e Psychedelic Ayahuasca Heals Traumatic Memories via a Sigma 1 Receptor-Mediated EpigeneticMnemonic Process. Frontiers in Pharmacology, 9, 330. https://doi.org/10.3389/fphar.2018.00330
Kjellgren, A., Eriksson, A., & Norlander, T. (2009). Experiences of encounters with ayahuasca—"the vine of the soul". Journal of Psychoactive Drugs, 41(4), 309–315. https://doi.org/10.1080/0279107 2.2009.10399767
Kryst, J., Kawalec, P., Mitoraj, A. M., Pilc, A., Lasoń, W., & Brzostek, T. (2020). E cacy of
single and repeated administration of ketamine in unipolar and bipolar depression: A meta-analysis of randomized clinical trials. Pharmacological Reports, 72(3), 543–562. https://doi.org/10.1007/ s43440-020-00097-z
Labate, B. C., & Goldstein, I. (2009). Ayahuasca–From Dangerous Drug to National Heritage: An Interview with Antonio A. Arantes. International Journal of Transpersonal Studies, 28(1), 53–64. https://doi.org/10.24972/ijts.2009.28.1.53
Leonard, J. (2020, January 31). Ayahuasca: What it is, e ects, and usage. https://www. medicalnewstoday.com/articles/ayahuasca
Lowe, H., Toyang, N., Steele, B., Valentine, H., Grant, J., Ali, A., Ngwa, W., & Gordon, L. (2021). e erapeutic Potential of Psilocybin. Molecules, 26(10), 2948. https://doi.org/10.3390/ molecules26102948
Mabit, J. (2007). Published in the book “Psychedelic Medicine (Vol. 2): New Evidence for Hallucinogic Substances as.
Malcom, K. (2019). ‘Mystical’ Psychedelic Compound Found in Normal Brains. University of Michigan. https://labblog.uofmhealth.org/labreport/mystical-psychedelic-compound-foundnormal-brains
Marks, M., & Cohen, I. G. (2021). Psychedelic therapy: A roadmap for wider acceptance and utilization. Nature Medicine, 27(10), 1669–1671. https://doi.org/10.1038/s41591-021-01530-3
Nochaiwong, S., Ruengorn, C., avorn, K. et al. (2021) Global prevalence of mental health issues among the general population during the coronavirus disease-2019 pandemic: a systematic review and meta-analysis. Sci Rep 11, 10173. https://doi. org/10.1038/s41598-021-89700-8
Osório, F. de L., Sanches, R. F., Macedo, L. R., dos Santos, R. G., Maia-de-Oliveira, J. P., WichertAna, L., de Araujo, D. B., Riba, J., Crippa, J. A., & Hallak, J. E. (2015). Antidepressant e ects of a single dose of ayahuasca in patients with recurrent depression: A preliminary report. Brazilian Journal of Psychiatry, 37, 13–20. https:// doi.org/10.1590/1516-4446-2014-1496
Pajouhesh, H., & Lenz, G. R. (2005). Medicinal chemical properties of successful central nervous system drugs. NeuroRX, 2(4), 541–553. https:// doi.org/10.1602/neurorx.2.4.541 Parrott, A. C. (2013a). Human psychobiology of MDMA or “Ecstasy”: An overview of 25years of empirical research. Human Psychopharmacology, 28(4), 289–307. https://doi.org/10.1002/hup.2318
Parrott, A. C. (2013b). MDMA, serotonergic neurotoxicity, and the diverse functional de cits of recreational ‘Ecstasy’ users. Neuroscience & Biobehavioral Reviews, 37(8), 1466–1484. https://doi.org/10.1016/j.neubiorev.2013.04.016
Passie, T., Seifert, J., Schneider, U., & Emrich, H. M. (2002). e pharmacology of psilocybin. Addiction Biology, 7(4), 357–364. https://doi. org/10.1080/1355621021000005937
Petranker, R., Anderson, T., & Farb, N. (2020). Psychedelic Research and the Need for Transparency: Polishing Alice’s Looking Glass. Frontiers in Psychology, 11, 1681. https://doi. org/10.3389/fpsyg.2020.01681
Pribish, A., Wood, N., & Kalava, A. (2020). A Review of Nonanesthetic Uses of Ketamine. Anesthesiology Research and Practice, 2020, 5798285. https://doi.org/10.1155/2020/5798285
Rainey, J. G. (2009). Gonzales v. O Centro Espirita Bene cente Uniao Do Vegetal. https://www.mtsu. edu/ rst-amendment/article/745/gonzales-v-ocentro-espirita-bene cente-uniao-do-vegetal
Riba, J., McIlhenny, E. H., Bouso, J. C., & Barker, S. A. (2015). Metabolism and urinary disposition of N,N-dimethyltryptamine a er oral and smoked administration: A comparative study. Drug Testing and Analysis, 7(5), 401–406. https://doi. org/10.1002/dta.1685
Romero, D. (2021, September 18). California moves closer to decriminalizing psychedelic drugs. NBC News. https://www.nbcnews. com/news/us-news/california-moves-closerdecriminalizing-psychedelic-drugs-n1279509
Ruscher, K., & Wieloch, T. (2015). e involvement of the sigma-1 receptor in neurodegeneration and neurorestoration. Journal of Pharmacological Sciences, 127(1), 30–35. https://doi.org/10.1016/j. jphs.2014.11.011
Sassano-Higgins, S., Baron, D., Juarez, G., Esmaili, N., & Gold, M. (2016). A Review of Ketamine Abuse and Diversion. Depression and Anxiety, 33(8), 718–727. https://doi.org/10.1002/ da.22536
Shiroma, P. R., uras, P., Wels, J., Albott, C. S., Erbes, C., Tye, S., & Lim, K. O. (2020). A randomized, double-blind, active placebocontrolled study of e cacy, safety, and durability of repeated vs single subanesthetic ketamine for treatment-resistant depression. Translational Psychiatry, 10(1), 1–9. https://doi.org/10.1038/ s41398-020-00897-0
Simonsson, O., Osika, W., Carhart-Harris, R., & Hendricks, P. S. (2021). Associations between lifetime classic psychedelic use and cardiometabolic diseases. Scienti c Reports, 11(1), 14427. https://doi.org/10.1038/s41598021-93787-4
Soler, J., Elices, M., Dominguez-Clavé, E., Pascual, J. C., Feilding, A., Navarro-Gil, M., García-Campayo, J., & Riba, J. (2018). Four Weekly Ayahuasca Sessions Lead to Increases in “Acceptance” Capacities: A Comparison Study With a Standard 8-Week Mindfulness Training Program. Frontiers in Pharmacology, 9, 224. https://doi.org/10.3389/fphar.2018.00224
Soler, J., Elices, M., Franquesa, A., Barker, S., Friedlander, P., Feilding, A., Pascual, J. C., & Riba, J. (2016). Exploring the therapeutic potential of Ayahuasca: Acute intake increases mindfulnessrelated capacities. Psychopharmacology, 233(5), 823–829. https://doi.org/10.1007/s00213-0154162-0
Strassman, R. J., & Qualls, C. R. (1994). DoseResponse Study of N,N-Dimethyltryptamine in Humans: I. Neuroendocrine, Autonomic, and Cardiovascular E ects. Archives of General Psychiatry, 51(2), 85–97. https://doi.org/10.1001/ archpsyc.1994.03950020009001
Szabo, A., Kovacs, A., Riba, J., Djurovic, S., Rajnavolgyi, E., & Frecska, E. (2016). e Endogenous Hallucinogen and Trace Amine N,N-Dimethyltryptamine (DMT) Displays Potent Protective E ects against Hypoxia via Sigma-1 Receptor Activation in Human Primary iPSC-Derived Cortical Neurons and MicrogliaLike Immune Cells. Frontiers in Neuroscience, 10, 423. https://doi.org/10.3389/fnins.2016.00423
Szabó, Í., Varga, V. É., Dvorácskó, S., Farkas, A. E., Körmöczi, T., Berkecz, R., Kecskés, S., Menyhárt, Á., Frank, R., Hantosi, D., Cozzi, N. V., Frecska, E., Tömböly, C., Krizbai, I. A., Bari, F., & Farkas, E. (2021). N,N-Dimethyltryptamine attenuates spreading depolarization and restrains neurodegeneration by sigma-1 receptor activation in the ischemic rat brain. Neuropharmacology, 192, 108612. https://doi. org/10.1016/j.neuropharm.2021.108612
Timmermann, C., Roseman, L., Williams, L., Erritzoe, D., Martial, C., Cassol, H., Laureys, S., Nutt, D., & Carhart-Harris, R. (2018). DMT Models the Near-Death Experience. Frontiers in Psychology, 9, 1424. https://doi.org/10.3389/ fpsyg.2018.01424
Vollenweider, F. X., & Kometer, M. (2010). e neurobiology of psychedelic drugs: Implications for the treatment of mood disorders. Nature Reviews Neuroscience, 11(9), 642–651. https:// doi.org/10.1038/nrn2884
Wasik, A. M., Gandy, M. N., McIldowie, M., Holder, M. J., Chamba, A., Challa, A., Lewis, K. D., Young, S. P., Scheel-Toellner, D., Dyer, M. J., Barnes, N. M., Piggott, M. J., & Gordon, J. (2012). Enhancing the anti-lymphoma potential of 3,4-methylenedioxymethamphetamine ('ecstasy’) through iterative chemical redesign: Mechanisms and pathways to cell death. Investigational New Drugs, 30(4), 1471–1483. https://doi.org/10.1007/s10637-011-9730-5
Williams, H. (2018, October 17). How LSD in uenced Western culture. https://www. bbc.com/culture/article/20181016-how-lsdin uenced-western-culture
Williams, M. T., Reed, S., & George, J. (2020). Culture and psychedelic psychotherapy: Ethnic and racial themes from three Black women therapists. Journal of Psychedelic Studies, 4(3), 125–138. https://doi. org/10.1556/2054.2020.00137

Vegetarianism Debate
STAFF WRITERS: OWEN SEINER ‘24, LILY DING ‘24, CARSON PECK ‘22, JULIETTE COURTINE ‘24, ETHAN WEBER ‘24, CALLIE MOODY ‘24
TEAM LEAD: KRISTAL WONG '22
Cover Image: Amongst all the diet trends and fads, one of the most prominent includes the vegetarian diet, in which individuals abstain from consumption of meat. is article will explore possible bene ts and consequences of following such a diet. Image Source: Wikimedia Commons Introduction
Omnivore. Carnivore. Pescatarian. Vegetarian. Modern diet culture is surrounded by various dietary practices that can embrace or restrict the variety of food available. Vegetarianism is described as a plant-based dietary practice that abstains from meat and meat by-products. Individuals decide to follow vegetarian lifestyles for a multitude of reasons, including health bene ts, environmental concerns, moral biases, and religion.
In the US, about 4-5% of individuals follow a vegetarian diet; this gure includes vegans, a subset of vegetarians who don’t eat any animal byproducts (e.g. eggs and cheese) (Hrynowski, 2021; Staher, 2020). Interestingly, a 2016 survey found that 37% of the US population sometimes or always orders vegetarian options when eating out (Hrynowski, 2021). In comparison, a 2014 study found that 31% of the global population follows a vegetarian diet, with the largest percentage of the world’s vegetarians from India (Figus, 2014). What does this say about the US? One thing this statistic signi es is that Americans eat less fruits and vegetables. According to the CDC, only 1 in 10 Americans get enough fruits and vegetables (CDC, 2021).
is article will explore the origins of vegetarianism and analyze its current trends, follow the climate/environmental impacts of vegetarian diets, and expose health impacts of pursuing vegetarian lifestyles.
Background
Anthropologic origins of carnivorous consumption begin as early as 2 million years ago. Early hominids practiced scavenging ( nding an already killed carcass) and hunting (killing the prey) as t for their ecological environment, but evolutionary advantage caused meat consumption to gain popularity. Carnivorous consumption of meat and bone marrow proved to be a bene cially e cient source of carloric resources and nutrients. ese alternative means of consumption allowed hominins to increase body size without sacri cing mobility or dexterity (Pobliner, 2013).
e origin of vegetarianism traces back to as early as 500 BC (Spencer, 1993). e beginning of the movement is evident from the days of Buddha in ancient India. e vegetarian philosophy was
not originally created for the same reasons that inspire contemporary vegetarian ideals, such as personal health concerns or the environmental impact of meat consumption, but rather a spiritual need coupled with unique societal foundations. In ancient India, the Hindu belief in reincarnation supported the emergence of the vegetarian philosophy. e Hindus believed that the pain and terror of the animal that was killed lived on in one’s reincarnated soul. erefore, their belief that consciousness lives on a er death discouraged members of the Hindu religion from meat diets (Rosen, 2006). It was not enforced in their scriptures, yet the language was unambiguous in stating that true peace could not be obtained if one killed animals and consumed meat.
In addition to spiritual reasons for being vegetarian, India’s relative wealth played a unique role in supporting widespread vegetarianism. e concept of a “taboo,” especially one concerning food, could only be present in a civilization where food was not scarce; basic needs must have been met for the consideration of limiting one’s food source to be entertained. Ancient India relied on cows for resources more valuable than meat, which made them too valuable to eat and enabled a taboo against killing cows. e milk and urine of cows were important for dairy and cleanser, and cow dung in agriculture allowed for vegetation growth even with little rainfall (Rosen, 2006). ese societal circumstances and beliefs allowed for a philosophy similar to modern vegetarianism to take root. e term “vegetarianism” has only been in use since the 1840s, but the moral creed to not kill animals for food was formed in this time period (Spencer, 1993).
Much has changed since then, perhaps most notably the traction and credibility of the vegetarian movement itself. Research on vegetarianism didn’t begin until the 1800s, and by this point strict spiritual or religious reasoning for the diet had grown uncommon. Morality continued to be an important motivator, but because of the more abstract nature of this incentive, as well as a decrease in the acceptance of the authority of religion, vegetarianism was a fringe movement. However, the rising in uence of science throughout the late 1800s into the 20th century created a new wave of vegetarians with a desire for improved bodily health (Whorton, 1994). Research on nutrition also began to appear at this time, which supported the diet with empirical evidence and allowed it to be recognized as a healthy alternative. Beginning in the early 21st century, the vegetarian diet became a larger minority in western countries (reaching up to 8% in Canada and 3% in the United States), and therefore, a need for the formalization of the study of vegetarianism was recognized (Ruby, 2012).
Historically, the vegetarian diet has been more popular in less developed countries where reductions in meat consumption help alleviate poverty or are more tting for those in poverty. More a uent countries, such as the US and Europe, incorporate more meat into their diets. And although greater consumption of meat and animal products is correlated with greater wealth, it is important to note that these carnivorous dietary practices aren’t necessarily followed by greater health outcomes (Godfray et al., 2010). In fact, the more developed countries have higher rates of coronary heart disease, diabetes, and atherosclerosis.
In addition, the lifestyles of vegetarians have been a source of prejudice and stereotyping. e infamous term “vegaphobia” is o en associated with those discriminating against individuals who adopt plant-based diets. While typically not thought of as a legitimate social issue, social stigma against vegetarians and vegans exists and are o en perpetuated by the media. In a 2011 study at the London School of Economics, researchers analyzed mentions of veganism in UK newspapers throughout 2007 and found 397 mentions of veganism or vegetarianism with 74.3% being negative, 20.2% neutral—mostly in the case of food and travel reviews—and only 5.5% positive (Cole & Morgan, 2011). Of the negative mentions of veganism, 29.2% ridiculed veganism, 28.8% portrayed it as an overly ascetic lifestyle, 27.8% described it as overly di cult or as a fad, and 13.5% represented vegans as overly sensitive or hostile (Cole & Morgan, 2011). Additionally, complex social issues are intertwined with vegetarianism, with another study conducted in the UK nding that those who follow plant-based diets were consistently perceived as less masculine ( omas, 2016).
In further touching upon the social issues related to the adoption of vegetarian diets, the topic of accessibility must be addressed, especially given that vegetarian diets are o en perceived as being more expensive than standard diets. In a study performed in the United States, women were sorted into two categories: plant-based (n=1109) or control (n=1145). Women in the plant-based group only spent an additional $1.22 per week compared to their counterparts who received no special dietary instructions (Hyder et al., 2009). While statistically signi cant, an additional $1.22 spent on groceries per week likely is not a huge " e origin of vegetarianism traces back to as early as 500 BC."
Image 1: Cultivated livestock such as the cattle pictured above are the largest producers of greenhouse gas (GHG) emissions amongst the global agricultural sector. Image Source: Wikimedia Commons Climate/Environmental E ects
Can vegetarian diets mitigate the ills of climate change? e answer is complex: one of the main causes of global climate change over the last 150 years, and especially over the last six decades, can be traced to agricultural activity, such as deforestation and crop irrigation (Wuebbles et al., 2017; Ruddiman, 2005). Evidence suggests that deforestation and irrigation even dating back to the origins of agriculture 11,000 years ago have in uenced warming patterns (Ruddiman, 2005). Agriculture not only changes land use and cover, but also changes the rate by which greenhouse gases are being released into the atmosphere (Smith et al., 2014). ough its emissions have decreased 24% since 2006, the agriculture, forestry, and other land use sector is still responsible for around 25% of anthropogenic greenhouse gas emissions, thus signi cantly contributing to global warming (Smith et al., 2014). Animal agriculture contributes signi cantly to climate change due to high demands for both land and food.
In fact, the livestock sector is one of the greatest contributors to the global anthropogenic greenhouse gas emissions, composing almost 80% of the agricultural sector’s emissions (McMichael et al., 2007). Animal husbandry, the management and production of domestic (farm) animals, has been an integral part of human history since long before the rise of the modern agricultural system; yet, increases in global household income, along with unprecedented population growth, has created a commensurate increase in the demanded quantity of meat (Rojas-Downing et al., 2017). Farmers now carry a larger herd size, which requires larger amounts of feedstock, grazing pastures, and water. While these e ects increase climate impacts, farmers o en adopt large herds as a bu er against the consequences of climate change, creating a cycle in which the impacts of climate change necessitate a greater herd size, which further engenders more climatic change (Næss and Bårdsen, 2013). Such practices continue to negatively impact the climate, and farmers respond by intensifying their already damaging practices. It is estimated that, collectively, the livestock supply chains emitted around 8.1 gigatons of carbon dioxide equivalent in 2012 (FAO, 2021).
e two most signi cant types of greenhouse gases that are emitted from animal agriculture are methane and nitrous oxide. Mainly produced by enteric fermentation and manure storage, methane has 28 times greater e ect on global warming than carbon dioxide per unit. Nitrous oxide, which can also arise from manure storage and fertilizer use, has a global warming potential that is 265 times higher than carbon dioxide (Grossi et al., 2019). And although some may say that using manure as fertilizer o sets the GHG emissions footprint, the o set is not substantial enough (Godfray et al., 2010). Among all domesticated animals, cattle were the greatest contributor to total emissions with approximately 5.0 gigatonnes of carbon dioxide equivalent, which accounts for 62% of the animal husbandry sector’s total emissions (FAO, 2021). Pigs and poultry, the other most popular domesticated

types of agricultural animals, emit around 7-11% of the sector’s total emissions (FAO, 2021)
Another way that the livestock sector contributes to greenhouse gas emissions is through feed production and water demands. Feed production includes the greenhouse gas emissions, not only from feed processing and transport, but also land use changes, manufacturing, use of fertilizers and pesticides, and manure excreted and applied to elds. Feed production and processing contribute to around 45% of the sector’s greenhouse gas emissions, which is greater than the number of emissions from enteric fermentation and over four times that of manure storage (Grossi et al., 2019).
Beyond basic livestock contributions to greenhouse gas emissions, the animal feed industry contributes greatly to climate change through its mass agricultural production of o en non-organic crops. Organic agriculture, far more commonly used when growing produce intended for humans than in livestock, is far more energy e cient and conservative than non-organic agriculture. Organic agriculture uses no arti cial pesticides or fertilizers, two of the biggest agricultural contributors to climate change. For instance, the N2O released by mineral fertilizers alone accounts for almost 20% of global agricultural greenhouse gas emissions, and N2O emissions overall account for 38% of all agricultural greenhouse gas emissions. e necessary reduction of mineral fertilizer, however, requires other practices to optimize nutrients in the soil, without arti cially adding them using fertilizers. One of the best manners to do this is to diversify crops and employ crop rotation techniques. is is feasible when producing food for human consumption, because there are a variety of common crops produced for that purpose. However, there are very few crops produced as animal feed, and thus crop rotation and diversi cation become far more di cult (Scialabba et al., 2010). us, production of organic crops, intended for direct human consumption, is far more e cient and ecofriendlier than mass-producing crops for animal consumption.
Animal husbandry’s contribution to greenhouse gas emissions is further exacerbated when forests are converted into pasture or cropland. Deforestation is one of the main contributors towards greenhouse gas emissions in the agricultural sector. O entimes, the slash-andburn clearing technique whereby vegetation is cut (slashed) and burned down to create agricultural lands for usages such as plantations and pastures (Tinker et al., 1996). ese changes in land-use and cover not only degrade the land, but also have important impacts on climate change. Deforestation causes large releases of carbon dioxide from the soil and vegetation, which are carbon sinks that store carbon dioxide (Ekblad and Bastviken, 2019). rough natural growth processes, plants take up carbon dioxide from the atmosphere and nitrogen from the soil; however, the decomposition of dead plant biomass that o en occurs as a result of deforestation releases signi cant amounts of greenhouse gases (Smith et al., 2014). In addition, overgrazing is an important environmental issue tied to land use from omnivorous diets. Overgrazing results in decreased biodiversity and vegetation cover, leading to soil erosion, which prevents water from permeating the soil (Kosmas et al., 2015). Without moist soil, the land undergoes deserti cation and loses the ability to become agriculturally productive.
Water consumption is another issue. roughout the world, around one third of all water goes directly towards the production of animal products, mostly meat and dairy. While ovo-lacto vegetarians continue to consume dairy products, in choosing to avoid meat products, vegetarians conserve large quantities of water. Generally, animal products require more water to produce the same caloric content compared to crop-based foods that vegetarians consume more frequently. As a result of the excess water required to produce food for livestock, meat makes up 37% of the food-related water footprint of the average American. If all meat were to be replaced with crop food, the average water footprint would decrease by 30% (Meknonnen andHoekstra, 2012).
Biological E ects
Are vegetarian diets biologically nutritious? Are they sustainable? When considering the viability of vegetarian or vegan diets, it is important to consider not only the common dietary de ciencies associated with these diets but also the typical nutritional value of plants as well. For digestion, plants are essential for ber. Di erences between the digestion of plants and animal products are mainly due to the di ering chemical compositions of these two food categories. Most notably, the presence of ber in plant material is of critical importance for a number of gastrointestinal (GI) conditions. According to the British Journal of Nutrition, dietary ber a any indigestible carbohydrate-based plant
Image 2: Legumes, a healthy plant-based food can be consumed in place of meat protein sources. Image Source: Wikimedia Commons material and is known to in uence nutrient bioaccessibility, microbial fermentation, GI hormone signaling, metabolizable energy, and postprandial metabolism (Grundy et al., 2016). Because ber is indigestible by endogenous enzymes, humans rely on the diverse microbiota of the GI tract to ferment ber into short-chain fatty acids such as acetate, propionate, and butyrate (Tomova et al., 2019). Fiber is also known to accelerate the speed of digestion, allowing for signi cantly reduced digestion times for plant material as compared to meats. Conversely, animal protein relies extensively on a cohort of digestive enzymes in order to be broken down into amino acids. is process is slowed signi cantly by the presence of high fat content within the meat. A slower metabolism may be unfavorable because the slower expenditure of calories and energy converts excess calories into fat in the body (Harvard Health Publishing, 2021).
Generally, plant-based diets tend to be much higher in vitamin C, magnesium, folate, manganese, thiamin, potassium, and vitamin E (van Vliet et al., 2020). A 2003 study of the diets of over 65,000 UK citizens revealed signi cantly higher intakes of these vitamins among vegans and vegetarians as compared to meat-eaters (Davey et al., 2003). Each of these compounds are vital for human health and can be readily obtained by an exclusively plant-based diet.
However, some vital nutrients are either much more readily or exclusively obtained from animal sources, including vitamins A (retinol), B12 (adenosyl- and hydroxocobalamin), D (cholecalciferol), K2 (menaquinone-4), minerals such as iron and zinc, and long-chain polyunsaturated fatty acids. Precursors to these nutrients may be sourced from plants, but many people lack the metabolic e ciency to achieve su cient rates of enzymatic conversion of these precursors to satisfy their own nutritional demands (van Vliet et al., 2020). As such, dietary supplements are o en advised as a complement to plant-based diets.
A common source of controversy with respect to the nutritional discrepancy between plant-based and omnivorous diets is the amount and quality of protein that each diet o ers. According to the National Academy of Medicine, the current recommended dietary allowance (RDA) protein for an adult is 0.8 g/kg (Institute of Medicine, 2005). Animal proteins are most commonly recommended to meet this daily allowance because of their high quality and modest caloric load (van Vliet et al., 2020). Using only plants to meet this daily allowance poses two problems for vegetarians and vegans: rstly, that the amount of easily digestible protein in plants is signi cantly lower than that in animal sources, and secondly, that the quality of raw plant protein is o en lower due to reduced amounts of the essential amino acids such as methionine and lysine (Sarwar et al., 2012; van Vliet et al., 2015). Despite the complications associated with obtaining a su ciently large and diverse daily intake of protein from plants alone, modern plant-based meat alternatives enable vegans and vegetarians to adhere to their diets without risking any nutritional de ciencies. Such alternatives include but are not limited to tofu, beans, nuts, seeds, and quinoa.
Health E ects
Switching to a vegetarian diet is commonly cited as a way to improve one’s general health. Some of the strongest evidence in support of this idea relates to the reduction of diseases like heart disease, diabetes, and cancer. Multiple studies have shown that vegetarians are generally less likely to su er from these ailments than meateaters. For example, one long-term study on heart disease followed 76,000 men and women


in the UK, USA, and Germany for about 10 years. Adjusting for factors like age, sex, and smoking, researchers found that people who did not consume meat or sh were 24% less likely to die from ischemic heart disease than those who did (Key et al., 1998). e gap in heartdisease death rate between vegetarians and nonvegetarians was higher the younger the subjects were. Furthermore, Qian et al.’s study of over 300,000 people found links between plant-based diets and lower risk of type 2 diabetes (2019). ese researchers determined that people who ate a mostly vegetarian diet were 23% less likely to develop this disease (Qian et al., 2019). Finally, Lanou et al. suggested that vegetarian diets are modestly preventative of cancer (2011). Although the results regarding vegetarianism and speci c cancers were less clear, the report stated that most large observational studies showed vegetarianism correlates to a 10-12% reduction in overall cancer risk (Lanou et al., 2011).
Scienti c research also lends support to the idea that vegetarianism can lead to weight loss. In 2014, researchers analyzed a collection of clinical trials on this subject. ey found that those on vegetarian diets lost an average of 4.4 pounds more than those on diets with meat, controlling for obvious factors like exercise and overall calorie intake (Huang et al., 2014). Other research has shown similar implications. For example, dieters who went vegetarian were showed to able to lose twice as much weight as dieters who continued to eat meat (Kahleova et al., 2017). e vegetarians also improved their metabolism more by reducing subfascial and intramuscular fat more e ectively. Tonstad et al. found that people who don’t eat meat tend to have lower BMIs in general (regardless of whether they are trying to lose weight) (2009).
Conclusion
Evidently, there are many reasons for and against following vegetarian diets. In making decisions on whether to follow such a diet, it is important to consult the science and personal values. is article has brie y outlined arguments pertaining to the environment, biology, and health, but is not an exhaustive list. Other possible areas to consider are the issue of water usage/droughts that come with livestock cultivation and emergence of zoonotic diseases. It is also important to consider the barriers to following vegetarian diets as fruit and vegetable lled diets have been seen to correlate with lack of accessibility and lower incomes (CDC, 2021).
References
Ahammad, H., Clark, H., Dong, H., & Tubiello, F. (2014). Climate Change 2014 Mitigation of Climate Change: Working Group III Contribution to the Fi h Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. https://doi. org/10.1017/CBO9781107415416
CDC. (2021, February 16). Only 1 in 10 Adults Get Enough Fruits or Vegetables. Centers for Disease Control and Prevention. https:// www.cdc.gov/nccdphp/dnpao/images/news/
Image 3: Human-animal contact, which increase with livestock cultivation, increases the likelihood of the emergence of zoonotic diseases such as the SARS-CoV-2 virus.
Image Source: Wikimedia Commons
Davey, G. K., Spencer, E. A., Appleby, P. N., Allen, N. E., Knox, K. H., & Key, T. J. (2003). EPIC–Oxford:lifestyle characteristics and nutrient intakes in a cohort of 33 883 meat-eaters and 31 546 non meat-eaters in the UK. Public Health Nutrition, 6(3), 259–268. https://doi.org/10.1079/ PHN2002430
Diamond, J. (n.d.). e Worst Mistake in the History of the Human Race. Discover Magazine. Retrieved April 11, 2021, from https://www. discovermagazine.com/planet-earth/the-worstmistake-in-the-history-of-the-human-race
Ekblad, A., & Bastviken, D. (2019). Deforestation releases old carbon. Nature Geoscience, 12(7), 499–500. https://doi.org/10.1038/s41561-0190394-7
Figus, C. (n.d.). 375 million vegetarians worldwide. All the reasons for a green lifestyle. EXPONet. Retrieved September 13, 2021, from http://www.expo2015.org/magazine/en/ lifestyle/375-million-vegetarians-worldwide. html
Food and Agriculture of the United Nations (FAO). (2021). GLEAM 2.0—Assessment of greenhouse gas emissions and mitigation potential. Fao.Org. https://www.fao.org/gleam/ results/en/
Gardos, G., & Cole, J. O. (1976). Maintenance antipsychotic therapy: Is the cure worse than the disease? e American Journal of Psychiatry, 133(1), 32–36. https://doi.org/10.1176/ ajp.133.1.32
Godfray, H. C. J., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., Pretty, J., Robinson, S., omas, S. M., & Toulmin, C. (2010). Food Security: e Challenge of Feeding 9 Billion People. Science, 327(5967), 812–818. https://doi.org/10.1126/science.1185383
Grossi, G., Goglio, P., Vitali, A., & Williams, A. G. (2019). Livestock and climate change: Impact of livestock on climate and mitigation strategies. Animal Frontiers, 9(1), 69–76. https://doi. org/10.1093/af/vfy034
Grundy, M. M.-L., Edwards, C. H., Mackie, A. R., Gidley, M. J., Butterworth, P. J., & Ellis, P. R. (2016). Re-evaluation of the mechanisms of dietary bre and implications for macronutrient bioaccessibility, digestion and postprandial metabolism. British Journal of Nutrition, 116(5), 816–833. https://doi.org/10.1017/ S0007114516002610
Grunert, K. G. (2006). Future trends and consumer lifestyles with regard to meat consumption. Meat Science, 74(1), 149–160. https://doi.org/10.1016/j.meatsci.2006.04.016
Harvard Health. (2018, April 1). e truth about metabolism. Harvard Health. https://www. health.harvard.edu/staying-healthy/the-truthabout-metabolism
Harvard Health. (2021, October 6). Does Metabolism Matter in Weight Loss? Harvard Health. https://www.health.harvard.edu/dietand-weight-loss/does-metabolism-matter-inweight-loss
Hrynowski, G. (2019, September 27). What Percentage of Americans Are Vegetarian? Gallup. Com. https://news.gallup.com/poll/267074/ percentage-americans-vegetarian.aspx
Huang, R.-Y., Huang, C.-C., Hu, F. B., & Chavarro, J. E. (2016). Vegetarian Diets and Weight Reduction: A Meta-Analysis of Randomized Controlled Trials. Journal of General Internal Medicine, 31(1), 109–116. https://doi. org/10.1007/s11606-015-3390-7
Kahleova, H., Klementova, M., Herynek, V., Skoch, A., Herynek, S., Hill, M., Mari, A., & Pelikanova, T. (2017). e E ect of a Vegetarian vs Conventional Hypocaloric Diabetic Diet on igh Adipose Tissue Distribution in Subjects with Type 2 Diabetes: A Randomized Study. Journal of the American College of Nutrition, 36(5), 364–369. https://doi.org/10.1080/0731572 4.2017.1302367
Key, T. J., Fraser, G. E., orogood, M., Appleby, P. N., Beral, V., Reeves, G., Burr, M. L., ChangClaude, J., Frentzel-Beyme, R., Kuzma, J. W., Mann, J., & McPherson, K. (1998). Mortality in vegetarians and non-vegetarians: A collaborative analysis of 8300 deaths among 76,000 men and women in ve prospective studies. Public Health Nutrition, 1(1), 33–41. https://doi.org/10.1079/ PHN19980006
Lanou, A. J., & Svenson, B. (2010). Reduced cancer risk in vegetarians: An analysis of recent reports. Cancer Management and Research, 1. https://doi.org/10.2147/CMAR.S6910
McMichael, A. J., Powles, J. W., Butler, C. D., &
Uauy, R. (2007). Food, livestock production, energy, climate change, and health. e Lancet, 370(9594), 1253–1263. https://doi.org/10.1016/ S0140-6736(07)61256-2
Næss, M. W., & Bårdsen, B.-J. (2013). Why Herd Size Matters – Mitigating the E ects of Livestock Crashes. PLoS ONE, 8(8), e70161. https://doi. org/10.1371/journal.pone.0070161
Panel on Macronutrients, Panel on the De nition of Dietary Fiber, Subcommittee on Upper Reference Levels of Nutrients, Subcommittee on Interpretation and Uses of Dietary Reference Intakes, Standing Committee on the Scienti c Evaluation of Dietary Reference Intakes, Food and Nutrition Board, & Institute of Medicine. (2005). Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids (p. 10490). National Academies Press. https://doi.org/10.17226/10490
Pobiner, B. (2013). Evidence for Meat-Eating by Early Humans | Learn Science at Scitable. Nature Education Knowledge, 4(6). https://www.nature. com/scitable/knowledge/library/evidence-formeat-eating-by-early-humans-103874273/
Qian, F., Liu, G., Hu, F. B., Bhupathiraju, S. N., & Sun, Q. (2019). Association Between Plant-Based Dietary Patterns and Risk of Type 2 Diabetes: A Systematic Review and Meta-analysis. JAMA Internal Medicine, 179(10), 1335. https://doi. org/10.1001/jamainternmed.2019.2195
Quinton, A. (2019, June 27). Cows and climate change. UC Davis. https://www.ucdavis.edu/ food/news/making-cattle-more-sustainable
Rojas-Downing, M. M., Nejadhashemi, A. P., Harrigan, T., & Woznicki, S. A. (2017). Climate change and livestock: Impacts, adaptation, and mitigation. Climate Risk Management, 16, 145–163. https://doi.org/10.1016/j.crm.2017.02.001
Rosen, S. (2006). Essential Hinduism. Greenwood Publishing Group.
Ruby, M. B. (2012). Vegetarianism. A blossoming eld of study. Appetite, 58(1), 141–150. https:// doi.org/10.1016/j.appet.2011.09.019
Ruddiman, W. F. (2005). How Did Humans First Alter Global Climate? Scienti c American, 292(3), 46–53. https://doi.org/10.1038/ scienti camerican0305-46
Sarwar Gilani, G., Wu Xiao, C., & Cockell, K. A. (2012). Impact of Antinutritional Factors in Food Proteins on the Digestibility of Protein and the Bioavailability of Amino Acids and on Protein Quality. British Journal of Nutrition, 108(S2), S315–S332. https://doi.org/10.1017/ S0007114512002371
Schahczenski, & Hill, H. (2009). Agriculture, Climate Change and Carbon Sequestration (No. 012309; pp. 1–16). National Center for Appropriate Technology (NCAT). https://www. canr.msu.edu/foodsystems/uploads/files/agclimate-change.pdf
Scialabba, N. E.-H., & Müller-Lindenlauf, M. (2010). Organic agriculture and climate change. Renewable Agriculture and Food Systems, 25(2), 158–169. https://doi.org/10.1017/ S1742170510000116
Spencer, C. (1996). e Heretic’s Feast: A History of Vegetarianism. UPNE.
Stahler, C. (n.d.). How Many Adults in the U.S. are Vegetarian and Vegan | e Vegetarian Resource Group (VRG). Retrieved September 13, 2021, from https://www.vrg.org/nutshell/ Polls/2019_adults_veg.htm
Tinker, P. B., Ingram, J. S. I., & Struwe, S. (1996). E ects of slash-and-burn agriculture and deforestation on climate change. Agriculture, Ecosystems & Environment, 58(1), 13–22. https://doi.org/10.1016/0167-8809(95)00651-6
Tomova, A., Bukovsky, I., Rembert, E., Yonas, W., Alwarith, J., Barnard, N. D., & Kahleova, H. (2019). e E ects of Vegetarian and Vegan Diets on Gut Microbiota. Frontiers in Nutrition, 6, 47. https://doi.org/10.3389/fnut.2019.00047
Tonstad, S., Butler, T., Yan, R., & Fraser, G. E. (2009). Type of Vegetarian Diet, Body Weight, and Prevalence of Type 2 Diabetes. Diabetes Care, 32(5), 791–796. https://doi.org/10.2337/ dc08-1886
van Vliet, S., Burd, N. A., & van Loon, L. J. (2015). e Skeletal Muscle Anabolic Response to Plant- versus Animal-Based Protein Consumption. e Journal of Nutrition, 145(9), 1981–1991. https:// doi.org/10.3945/jn.114.204305
van Vliet, S., Kronberg, S. L., & Provenza, F. D. (2020). Plant-Based Meats, Human Health, and Climate Change. Frontiers in Sustainable Food Systems, 4, 128. https://doi.org/10.3389/ fsufs.2020.00128
Whorton, J. C. (1994). Historical development of vegetarianism. e American Journal of Clinical Nutrition, 59(5), 1103S-1109S. https://doi. org/10.1093/ajcn/59.5.1103S
Wuebbles, D. J., Easterling, D. R., Hayhoe, K., Knutson, T., Kopp, R. E., Kossin, J. P., Kunkel, K. E., LeGrande, A. N., Mears, C., Sweet, W. V., Taylor, P. C., Vose, R. S., Wehner, M. F., Wuebbles, D. J., Fahey, D. W., Hibbard, K. A., Dokken, D. J., Stewart, B. C., & Maycock, T. K. (2017). Ch. 1: Our Globally Changing Climate. Climate Science Special Report: Fourth National Climate Assessment, Volume I. U.S. Global Change Research Program. https://doi.org/10.7930/ J08S4N35
DARTMOUTH UNDERGRADUATE JOURNAL OF SCIENCE Hinman Box 6225 Dartmouth College Hanover, NH 03755 USA
http://dujs.dartmouth.edu
dujs@dartmouth.edu