


En el horizonte de la producción animal en Latinoamérica se presenta una oportunidad invaluable para generar productos de origen animal de alta calidad que respondan a las necesidades de nuestro tiempo: naturales, con bajos niveles de emisiones de gases de efecto invernadero y profundamente respetuosos con el entorno.
En un contexto donde la agenda ambiental cobra protagonismo global, nuestras tierras ricas y diversas pueden ser la cuna de un modelo productivo que enfatice la sostenibilidad.
En lugar de ser vista únicamente como productora de commodities, Latinoamérica puede convertirse en un referente de producción diferenciada por su calidad y su compromiso con el medio ambiente.
Los productos de origen animal del futuro serán valorados por estas cualidades, nuestros países pueden aprovechar su biodiversidad y cultura agrícola para generar productos que reflejen estos principios y contribuyan al desarrollo sostenible.
Para avanzar este camino, es imperativo reimaginar el papel de nuestra región como productora y defensora de un futuro donde el bienestar social y ambiental estén entrelazados. Así, podremos alimentar a nuestras comunidades e inspirar a un mundo ávido de cambios significativos hacia un desarrollo más justo y sostenible.
En este contexto, el conocimiento se convierte en la herramienta más poderosa.
Los nutricionistas basamos nuestro trabajo en el entendimiento de la biología animal y la comprensión de los sistemas de producción. Tenemos la responsabilidad de ofrecer respuestas basadas en criterios científicos para aumentar la producción, reduciendo ineficiencias, mitigando los impactos negativos sobre el ambiente y la sostenibilidad de los sistemas. Nuestro foco son los componentes principales de la dieta: forrajes, granos, concentrados y subproductos.
Asimismo, resulta fundamental prestar atención a ciertos compuestos que, aunque se utilicen en pequeñas cantidades, pueden influir significativamente en los sistemas productivos.
En este grupo de compuestos, incluimos a los fitogénicos: mezclas y extractos derivados de plantas. En esta edición abordaremos aquellos que cuentan con evidencia científica de su efecto en la producción, la salud y el bienestar animal. Es fundamental contar con ellos como grandes aliados en nuestra actividad.
Proporcionaremos a nuestros lectores elementos basados en las investigaciones más recientes, señalando también lo que aún se desconoce. Nuestra intención, como siempre, es enriquecer la comprensión del tema con criterios fundamentados. Ustedes, los lectores, utilizarán la información de la mejor manera posible.
¡Buena lectura!
Dra. Cecilia Cajarville. Directora técnica sección rumiantes nutriNews Latam
EDITOR
GRUPO DE COMUNICACIÓN AGRINEWS S.L.
PUBLICIDAD
Simone Dias +55 11 985852436 nutribr@grupoagrinews.com
Luis Carrasco +34 605 09 05 13 lc@agrinews.es
Victoria Domingues +55 41 992091549 nutrisocial@grupoagrinews.com
DIRECCIÓN TÉCNICA
Bernardo Fabricio Iglesias
Cecilia Cajarville
Juan Gabriel Espino
COORDINACIÓN TÉCNICA
Facundo Apecetche
REDACCIÓN
Amanda Abalo
Daniela Morales
Facundo Apecetche
Osmayra Cabrera
COLABORADORES
Carlos De Blas (UPM)
Gonzalo Glez. Mateos (UPM)
Xavier Mora (Consultor)
Alba Cerisuelo (CITA-IVIA)
Carlos Fernández (UPV)
Luis Miguel Gómez Osorio (CES)
Edgar Oviedo-Rondón (USSEC)
ADMINISTRACIÓN
Merce Soler
Tel: +34 677518854 admin@agrinews.es
www.nutrinews.com
Precio de suscripción anual: 90 USD
ISSN (Revista impresa) 2696-810X
ISSN (Revista digital) 2696-8118
GRATUITA PARA FABRICANTES DE ALIMENTO BALANCEADO, EMPRESAS DE PREMIXES Y NUTRICIONISTAS
Depósito Legal Nutrinews B-17990-2015
Ficha de materia prima: guisantes y subproductos derivados

Alba Cerisuelo
Investigadora Alimentación Animal en el IVIA
Elija correctamente su fuente de potasio
Jorge Castro
Servicio Técnico para Latinoamérica, Arm & Hammer, Animal Nutrition
Tablas nutriNews Latam Actualizaciones 2024
FITOGÉNICOS
Tablas nutriNews Latam Actualizaciones 2024 ADITIVOS ENZIMAS
BRONCHOVEST®: la solución natural para soportar los desafíos respiratorios y térmicos en la producción animal
Equipo técnico Biochem
Efectos del estrés crónico y la inflamación intestinal en la salud y el rendimiento de las aves de corral: Revisión
Tellez-Isaias G.
Department of Poultry Science, University of Arkansas
La suplementación con CELMANAX en dietas de reproductoras y pollos de engorde redujo la prevalencia de Salmonella
Equipo técnico Arm & Hammer
Por qué son importantes las micotoxinas en la producción de pollos de engorde 52
Lorran Baeumle Gabardo1 , Elle Chadwick 2, Shelby
Ramirez 3
1Global Product Manager, dsm-firmenich Nutrición y Salud Animal,
2Global Poultry Marketing Specialist, dsm-firmenich Nutrición y Salud Animal, 3Global Poultry Technical Manager, dsm-firmenich Nutrición y Salud Animal

Los secretos detrás de una fitasa
Juan Gabriel Espino
Nutricionista especialista en monogástricos

Salud hepática y metabolismo: ¿cómo optimizarlos para mejorar el rendimiento de los lechones en la fase de cría?
Eduardo Raele de Oliveira, MV, Msc, PhD; Ana Paula Pinoti Pavaneli, MV, Msc, PhD
Asesores técnicos de porcino - Nuproxa Switzerland Ltd

El uso de grasas oxidadas en la alimentación porcina: riesgos y consideraciones (Parte 1/3)
Gerardo Ordaz Ochoa, María
Alejandra Pérez Alvarado, Luis Humberto López
Hernández
Centro Nacional de Investigación Disciplinaria en Fisiología y Mejoramiento Animal, INIFAP
Taninos en la nutrición de rumiantes: de obstáculo a oportunidad

Cecilia Cajarville, José Luis Repetto
Departamento Producción Animal y Salud de Sistemas Productivos - IPAV, Facultad de Veterinaria, UdelaR, San José, Uruguay.
ANAVRIN®: Una solución innovadora para la optimización de la producción ganadera
Departamento científico VetosEurope
El Grupo de Comunicación Agrinews quisiera resaltar y distinguir el notable empeño y apreciable aporte y colaboración de los autores de los artículos. El esfuerzo compartido hace posible que podamos ofrecer a nuestros lectores un contenido técnico de calidad. Reiteramos por tanto nuestro más sincero agradecimiento.

Alba Cerisuelo, Investigadora Alimentación Animal en el IVIA
Los guisantes son plantas leguminosas de la especie Pisum sativum, de cultivo anual, que pueden alcanzar hasta 2 m de altura.
Esta especie está integrada por 2 subespecies: Pisum sativum hortense y Pisum sativum arvense. Las flores de la primera son blancas y las de la segunda son de diferentes colores.
En general, las semillas de las variedades de la subespecie Pisum sativum hortense son las que se utilizan en piensos para animales, mientras que las variedades de Pisum sativum arvense se utilizan en forma de forraje (guisante forrajero).


Dentro de la subespecie Pisum sativum hortense, la mayoría de las variedades son de primavera, aunque también existen variedades de invierno.
Las semillas de estas variedades pueden ser de color blanco, azul-verde o verde y de aspecto liso o rugoso.
En las variedades de Pisum sativum arvensese, las semillas son de color oscuro y la planta se consume como hierba verde, hierba seca o ensilado.
La calidad nutricional de los guisantes depende de la variedad. Los guisantes son, en general, ingredientes ricos en almidón y proteína de elevada calidad.
Además, por sus características agronómicas (plantas fijadoras de nitrógeno con bajas necesidades de fertilización, resistentes a la sequía, locales…) se consideran una alternativa muy interesante tanto a nivel económico como medioambiental a las fuentes de proteína habituales como la harina de soja.
Entre las diferentes variedades, se considera que el grano de guisante de primavera es el ingrediente de mayor calidad para la alimentación animal.
Además de las semillas (guisantes) y las plantas (forrajes), existen una gran variedad de subproductos derivados del procesado de guisantes para consumo humano o animal, con diferentes características nutricionales.
Estos subproductos son hoy en día minoritarios en comparación con las semillas enteras y los forrajes, aunque se espera que con el aumento del uso de proteína de origen vegetal para alimentación humana este tipo de subproductos estén cada vez más disponibles para alimentación animal.
En la Tabla 1 se muestran las materias primas clasificadas como guisantes o derivados que se recogen en el Catálogo de materias primas (Reglamento UE 68/2013). Estas materias primas se clasifican dentro del grupo 3 referente a “Semillas de leguminosas y sus productos derivados”.
Número Denominación

Descripción
3.11.1 Guisantes Semillas de Pisum ssp.
3.11.2 Salvado de guisantes
3.11.3 Copos de guisante
3.11.4 Harina de guisantes
3.11.5. Cáscaras de guisantes
Producto de la elaboración de harina de guisantes que está constituido principalmente por hollejos desprendidos durante la deshollejadura y limpieza de los guisantes.
Producto obtenido por cocción al vapor o mediante micronización por infrarrojos y aplastamiento de semillas de guisante descascarilladas.
Declaraciones obligatorias
Fibra bruta
Almidón
Producto obtenido durante la molturación de guisantes. Proteína bruta
Producto de la fabricación de harina de guisantes a partir de guisantes que está constituido principalmente por hollejos desprendidos al pelar y limpiar los guisantes y, en menor medida, de endospermo.
3.11.6. Guisantes descascarados Semillas de guisante descascaradas.
3.11.7 Harinillas de guisantes
3.11.8. Residuos del cribado de guisantes
3.11.9. Proteína de guisantes
3.11.10 Pulpa de guisantes [fibra interna del guisante]
3.11.11 Solubles de guisantes
3.11.12 Fibra de guisantes
3.11.13 Pasta de guisante
Producto de la fabricación de harina de guisantes que está constituido principalmente por partículas del cotiledón y, en menor medida, por pieles.
Producto obtenido a partir del cribado mecánico y constituido por fracciones de granos de guisante separados antes de un nuevo tratamiento.
Producto obtenido del agua extraída de los guisantes al producir el almidón, o después de la molturación y fraccionamiento por aire, que pueden ser parcialmente hidrolizados.
Producto obtenido de la extracción por vía húmeda de almidón y proteína a partir de guisantes que está constituido principalmente por fibra interna y almidón.
Producto obtenido de la extracción por vía húmeda de almidón y proteína a partir de guisantes que está constituido principalmente por proteínas solubles y oligosacáridos.
Producto obtenido por extracción tras la molturación y el tamizado del guisante descascarado.
Producto obtenido de la extracción por vía húmeda de almidón y proteína a partir de guisantes que está constituido principalmente por proteínas solubles, fibras internas, almidón y oligosacáridos. Además, puede contener hasta un 1 % de ácidos orgánicos.
Fibra bruta
Proteína bruta, fibra bruta
Proteína bruta, fibra bruta
Fibra bruta
Proteína bruta
Humedad, cuando sea < 70 % o > 85 %, almidón, fibra bruta, ceniza insoluble en HCl, cuando sea > 3,5 % sobre materia seca.
Humedad, cuando sea < 60 % o > 85 %, azúcares totales, expresados en sacarosa, proteína bruta
Fibra bruta
Contenido de humedad, cuando sea < 50 % o > 85 %, proteína bruta, fibra bruta, almidón
Tabla 1. Clasificación de guisantes y sus productos derivados según el Catálogo de Materias Primas (Reglamento UE 68/2013).
Debido a su heterogeneidad, los nutrientes que es necesario declarar de cada uno de ellos son diferentes, aunque, en general, la proteína y la fibra son los más destacados.

El proceso de obtención de los guisantes para consumo animal es sencillo. En el caso de los guisantes secos, la cosecha comienza cuando la planta y la vaina están bien secas, lo que, dependiendo de la zona, ocurre entre el mes de mayo, hasta mediados de agosto.
En este caso, se arranca la mata entera y luego se trilla para separar el grano (MAPA, 2018). En las variedades forrajeras, la parte aérea de estas plantas, una vez cosechadas, puede utilizarse en verde (frescas o ensiladas) o en seco.
Para el resto de los subproductos derivados del guisante el proceso de obtención dependerá del procesado industrial al que se sometan las semillas.
COMPOSICIÓNQUÍMICAY
En la Tabla 2 se muestra la composición (en materia seca) de las semillas crudas o procesadas por calor y de los concentrados o aislados de proteína de guisante disponibles para alimentación animal según diferentes fuentes (FEDNA, CVB e INRAE).
En general, los granos de guisante se consideran una buena fuente de almidón y proteína. El porcentaje de almidón se encuentra alrededor del 50% y el de proteína alrededor del 23-24% en materia seca.
En cuanto a los aminoácidos totales que contiene, su perfil es interesante para animales monogástricos ya que contiene un elevado porcentaje de lisina, mayor que el que aportan otras fuentes de proteína con mayor contenido proteico como son la harina de girasol o DDGS de cereales.
Sin embargo, al igual que otras leguminosas, su contenido en aminoácidos azufrados es bajo.
Su contenido en grasa es generalmente bajo, siendo esta principalmente insaturada y el porcentaje de fibra (FND y FAD) es similar al de la harina de soja y notablemente menor al que presentan otras fuentes de proteína como la harina de girasol, colza y DDGS de cereales.

Además, la fibra que contiene es muy poco lignificada. Esta característica junto a la elevada digestibilidad del almidón (90-97%) y de la proteína (>77% en rumiantes y >80% en porcino y aves) hace que su contenido energético sea elevado, similar al de la harina de soja y los cereales.

En el rumen, la velocidad de degradación del almidón es lenta pero la solubilidad de la proteína es elevada, lo que puede ser un factor limitante para rumiantes.
La composición de las semillas tratadas por calor (o extrusionadas) es similar a las crudas, aunque su aporte energético se espera algo más elevado.
A diferencia de otras leguminosas, los guisantes no suelen presentar niveles elevados de factores antinutricionales.
Sin embargo, su contenido en inhibidores de tripsina (TIA) puede ser problemático sobre todo en animales jóvenes (porcino y aves), ya que puede reducir la digestibilidad de su proteína (Stein et al., 2004).
La aplicación de tratamientos por calor (peletizado o extrusión) parece ser efectiva para reducir el contenido de TIA y aumentar la digestibilidad de los nutrientes en porcino (Hugman et al., 2021).
Su contenido en minerales es bajo en comparación con otras fuentes de proteína.
Además de las semillas, en la Tabla 2 se muestra la composición de los concentrados o aislados de proteína de guisante, que, aunque debido a su coste y baja disponibilidad no se usan habitualmente en piensos sí que pueden ser interesantes en ciertas fases del crecimiento como por ejemplo en lechones.
Estos ingredientes presentan una elevada cantidad de proteína (>50%) y aminoácidos y son altamente digestibles, aportando una elevada cantidad de energía en todas las especies.

Una elevada cantidad de estudios corroboran que los guisantes son una buena fuente de proteína y energía en vacas lecheras, terneros y pequeños rumiantes, y que presentan una elevada palatabilidad en estas especies (Anderson et al., 2007; Vander Pol et al., 2009, Greenwell et al., 2018; Lobón et al., 2020).


Ingredientes

Esenciales
No esenciales
Ingredientes

Coeficiente de digestibilidad de la proteína rumiantes
Coeficiente de digestibilidad de la proteína porcino crecimiento
Coeficiente de digestibilidad de la proteína porcino adulto
Coeficiente de digestibilidad de la proteína ponedoras
Coeficiente de digestibilidad de la proteína adultos
1 http://www.fundacionfedna.org/ingrediente; valores expresados en materia seca
2 https://www.cvbdiervoeding.nl/pagina/10021/home.aspx.
CVB Feed Table 2023. Chemical composition and nutritional values of feedstuffs; valores expresados en materia seca
3 https://feedtables.com/content/table-dry-matter; valores expresados en materia seca
Tabla 2. Composición química (en materia seca) de los guisantes y concentrados de proteína.
Estos estudios sugieren niveles de inclusión óptimos de entre 15-25% en vacas lecheras, 20-40% en terneros o hasta 30% en corderos.
Sin embargo, debido a la elevada solubilidad de la proteína de estos ingredientes se recomienda su uso junto a otras fuentes de proteína menos degradables para aumentar la calidad de la proteína de la ración, sobre todo en las fases productivas más exigentes como puede ser el inicio de la lactación. Otra vía para reducir la solubilidad de la proteína es aplicar tratamientos por calor o molienda grosera.
En monogástricos como el porcino, Stein et al. (2004) demostró que era posible incluir niveles superiores al 30% de guisantes en dietas para porcino de cebo y hasta el 18% en lechones sin consecuencias negativas sobre el crecimiento (Stein et al., 2004). Este menor nivel de inclusión en lechones se debe a la presencia de TIA.
En aves, aunque tradicionalmente la utilización de esta materia prima ha sido menor, estudios recientes indican que es posible incorporar niveles de hasta un 30% de guisantes de las variedades de flores blancas sin efectos negativos sobre el crecimiento y el rendimiento en matadero de pollos broiler (Bellof and Freitag, 2021; Kirn et al., 2024).
Al formular piensos con guisante para estas especies, es importante tener en cuenta el bajo contenido en aminoácidos azufrados de éste y sus derivados, para poder complementar los piensos adecuadamente.
A nivel tecnológico es relevante mencionar que, como otros granos de leguminosas, los guisantes son de fácil manejo y conservación en fábrica, y su inclusión en el pienso favorece el proceso de granulación.
Por otro lado, su composición química es relativamente poco variable y relativamente fácil de controlar ya que depende, en gran medida, de la variedad utilizada.
En conclusión, los guisantes son ingredientes locales que pueden incorporarse a niveles moderados-altos en dietas tanto de rumiantes como de monogástricos como el porcino y las aves.
Por su composición, en los piensos son capaces de sustituir tanto fuentes proteicas (soja) como energéticas (cereales), y con ello reducir la dependencia de materias primas no locales y el impacto ambiental (huella hídrica entre otros) y económico de los piensos.
Sin embargo, y sobre todo cuando se incorporan a niveles elevados, es importante tener en cuenta algunos limitantes como la solubilidad de la proteína en rumiantes, los TIA en fases tempranas y el bajo contenido en aminoácidos azufrados en aves.
Referencias disponibles en la versión web del artículo en nutrinews.com
Guisantes y subproductos derivados (field peas and by-products)
DESCÁRGALO EN PDF

Jorge Castro
Servicio Técnico para Latinoamérica, Arm & Hammer Animal Nutrition

El potasio es un nutriente esencial que requieren los animales para mantener la productividad y mitigar el estrés calórico. También es un nutriente móvil, lo que significa que no deja reservas en el cuerpo y debe reponerse en la ración diaria.
Sin esa reposición, los animales pueden presentar deficiencia y experimentar una reducción del crecimiento, debilidad o rigidez muscular, menor ingesta de alimento, trastornos nerviosos, palpitaciones cardíacas y otras manifestaciones.
La adición de fuentes de potasio a la ración requiere un manejo cuidadoso. El potasio es parte de un equilibrio electrolítico crítico, que debe mantenerse cuando se agrega potasio suplementario a la dieta. Ciertas fuentes de potasio también pueden provocar el calentamiento de la ración y la pérdida de nutrientes, por lo que es importante minimizar el riesgo de calentamiento tanto como sea posible.
Encontrar el equilibrio adecuado depende de tu fuente de potasio.


Existen dos fórmulas comunes para el equilibrio de electrolitos en la nutrición animal, que nos ayudan a comprender el importante papel del potasio en las raciones.
La fórmula del equilibrio electrolítico dietético (dEB) es:
(Sodio + Potasio) – Cloro
La fórmula de diferencia de cationes y aniones en la dieta (DCAD) es:
(Sodio + Potasio) – (Cloro + Azufre)
La relación entre estos elementos está estrechamente vinculada, lo que significa que no se puede analizar ningún elemento individualmente, ya que se necesita un equilibrio entre todos para mantener y optimizar la salud animal.
El cuerpo del animal tiene un sistema de amortiguación para mantener un pH fisiológico casi normal y evitar la mayoría de los desequilibrios. Sin embargo, muchos factores pueden afectar los niveles de elementos en un momento dado.
Por ejemplo, las necesidades de mantenimiento diarias, la producción y los factores ambientales como el estrés por calor drenan el potasio, por lo que a menudo se requiere potasio dietético adicional para mantener los niveles.
Ambas fórmulas se manipulan comúnmente para maximizar la salud y la productividad de los animales. Los niveles dietéticos de cationes (sodio y potasio), que son iones con carga positiva, y aniones (cloro y azufre), que son iones con carga negativa, se pueden ajustar para crear un equilibrio que sea positivo o negativo.
Por ejemplo, en el ganado lechero, las dietas previas al parto están diseñadas para DCAD negativo para prevenir la hipocalcemia, mientras que las dietas de lactancia están diseñadas para DCAD positivo para apoyar la producción de leche.





Las dietas a base de trigo, maíz y soya suelen considerarse “suficientes” en cuanto a aporte de potasio. Sin embargo, los animales suelen necesitar un dEB/DCAD más alto que la que se proporciona en la ración sin suplementos de potasio.
Para aprovechar los beneficios de producción de un dEB/DCAD más alto, se necesitan suplementos de sodio y potasio.

El carbonato de potasio, que se encuentra en Arm & Hammer DCAD Plus™, es la mejor opción para agregar potasio a la ración.
Muchos nutricionistas ganaderos y productores avícolas utilizan cloruro de potasio; sin embargo, este último tiene un claro inconveniente en lo que se refiere al equilibrio electrolítico. La carga iónica positiva del potasio y la carga iónica negativa del cloruro se anulan entre sí en el equilibrio electrolítico. En otras palabras, el dEB o la DCAD no pueden volverse más positivas para satisfacer las necesidades de producción.
El carbonato de potasio funciona de manera diferente porque solo aporta potasio. Se puede utilizar para crear un dEB/DCAD más positivo, lo que ayuda a los animales a mantener la producción.
Las fuentes de potasio suelen generar calor cuando se exponen a la humedad. El calor generado termina calentando toda la ración, lo que puede provocar que el alimento se “cocine” y pierda nutrientes esenciales. Las proteínas son particularmente susceptibles.
Los silos o tolvas de pienso pueden incluso generar vapor durante la cocción. Se debe extender una pila de pienso humeante en el suelo para permitir que el pienso se enfríe. Otro riesgo es el fuego.
Los animales que comen alimentos calientes también sufren los efectos. Reducir el calentamiento de las raciones es una estrategia necesaria para reducir el estrés térmico.
Elegir la fuente de potasio adecuada puede ayudar a minimizar el riesgo de calentamiento de la ración, junto con la añadidura de potasio a la ración cerca del final de la mezcla de alimentos.
¿QUÉ FUENTES DE POTASIO MINIMIZAN EL CALENTAMIENTO DE LA RACIÓN?
Hay tres fuentes principales de potasio en la dieta:
Bicarbonato de potasio
Carbonato de potasio anhidro
Carbonato de potasio sesqui-hidratado (DCAD Plus™)
El bicarbonato de potasio no calienta la ración, pero la forma de bicarbonato solo contiene la mitad de potasio en comparación con su contraparte de carbonato. Como resultado, el bicarbonato de potasio cuesta aproximadamente el doble, lo que hace que su uso sea poco práctico.
El carbonato de potasio anhidro genera mucho calor cuando se expone a la humedad, lo que hace que el calentamiento de las raciones sea una gran preocupación.
DCAD Plus™ utiliza carbonato de potasio sesqui-hidratado. Esta forma de potasio no genera calor porque ha sido parcialmente hidratada, lo que elimina la posibilidad de que se caliente en una ración húmeda.
Muchas especies pueden beneficiarse del carbonato de potasio en la dieta. La mayoría de las investigaciones se han realizado en vacas lecheras en período de lactancia y fuera del período de lactancia, y los nutricionistas se sienten cómodos con el manejo de los niveles de DCAD en las raciones para vacas lecheras.

Cuando los niveles de cloruro o azufre son altos, el equilibrio electrolítico general es especialmente importante, y se necesitan cationes (sodio y potasio) para compensar los aniones con carga negativa.
Si bien su adopción es menos generalizada, las operaciones avícolas pueden aplicar los mismos principios al manejo de la dieta de los pollos de engorde y ponedoras con resultados similares en términos de producción y mitigación del estrés térmico.
Los niveles bajos de potasio son peligrosos para las aves y pueden causar presión arterial alta, enfermedades cardíacas y muerte súbita, especialmente en dietas con alto contenido de sal. Aumentar el potasio y reducir la sal puede ayudar a reducir la presión arterial y el riesgo de accidente cerebrovascular.
Los niveles bajos de potasio también pueden reducir el calcio necesario para tener huesos y cáscaras fuertes.
Además, el desequilibrio electrolítico puede causar problemas metabólicos como discondroplasia tibial y alcalosis respiratoria en ponedoras. Un exceso de sodio en la dieta en relación con el potasio, junto con altos niveles de cloruro, puede causar discondroplasia tibial.

Las aves también son susceptibles a la pérdida de potasio durante épocas de estrés por calor, y el bicarbonato de sodio por sí solo puede no mitigar este estrés.
El carbonato de potasio se puede agregar a la dieta durante todo el año y ajustar cuando sea necesario para equilibrar los electrolitos y optimizar la producción.
¿Dónde puedo obtener más información?
Visita el portal de Arm & Hammer en: ahfoodchain.com para saber más acerca del carbonato de potasio y DCAD Plus™, y para ubicar a tu representante local.
Elija correctamente su fuente de potasio DESCÁRGALO EN PDF
Equipo técnico Biochem

Biochem tiene en su portafolio el producto BronchoVest®, un aditivo compuesto por los aceites esenciales de eucalipto, menta, y por cristales de mentol. BronchoVest® ha sido cuidadosamente diseñado para apoyar las funciones respiratorias de los animales.

Puede ser administrado por spray o agua de bebida, con rápida absorción vía mucosas. El producto es ideal para aplicaciones puntuales, en momentos de desafío para los animales.
Los componentes activos de BronchoVest® actuan de forma preventiva, apoyan al sistema respiratorio de los animales y contribuyen a reducir los efectos negativos de los desafíos respiratorios y del estrés por calor.
Por tanto, queda clara la acción sinérgica de los ingredientes de BronchoVest®, que poseen características que se complementan para apoyar las funciones respiratorias y el bienestar animal.

El aceite de eucalipto, con alta concentración del principio activo 1,8-cineol, ayuda a mejorar las funciones pulmonares: restaura la actividad natural del epitelio respiratorio y disminuye la acumulación de moco en los bronquiolos.
El aceite de menta regula la producción excesiva de moco, aumentando el ujo de aire hacia los pulmones.
Los cristales de mentol tienen efectos en receptores sensoriales especí cos, trayendo una sensación refrescante, especialmente importantes en situaciones de estrés por calor.
Los primeros estudios con BronchoVest® fueron realizados en aves. Estos demostraron entre los principales beneficios, una reducción de los efectos negativos de los desafios respiratorios, estandarización de la titulación de anticuerpos cuando suministrado 1 día después de vacunas respiratorias, reducción de las reacciones post-vacunales, reducción de pérdidas en el frigorífico y mejora del rendimiento zootécnico de pollos de corte, reproductoras pesadas y postura comercial.


Recientemente, en Brasil, se realizaron estudios con cerdos en crianza, crecimiento y terminación, donde hubo reducción de la frecuencia respiratoria posparto de las gestantes, reducción de la necesidad de medicación de los lechones en crianza, reducción de los efectos negativos respiratorios frente a desafíos para animales en crianza y engorde, reducción de la mortalidad de los lechones y mejor rendimiento zootécnico para las categorías evaluadas.
De forma similar, para los terneros, el uso de BronchoVest® llevó a un mejor estado general de salud y reducción de la necesidad de medicación.
Como conclusión general, BronchoVest®, aplicado tanto por agua de bebida o spray, tiene eficacia probada, pudiendo llevar a la mejora del bienestar de los animales y de esta forma favorecer la expresión del potencial genético para el rendimiento.
BRONCHOVEST®: la solución natural para soportar los desafíos respiratorios y térmicos en la producción animal DESCARGAR PDF















PAÍSES EN LATINOAMÉRICA DONDE EL PRODUCTO ESTÁ DISPONIBLE
ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS)
COMPONENTES ACTIVOS FITOGÉNICOS
ACCIÓN
PRODUCTO
Aves de corral, incorporar al alimento de forma continua o en aquellos periodos de mayor riesgo o desafío.
Aves de corral, incorporación continua al alimento.
Complejo de ácidos orgánicos
Scutellaria baïcalensis patentada y curcuma longa
Feedstim Poultry ayuda a las aves a enfrentarse a múltiples retos en la cría y, por lo tanto, garantiza un rendimiento y una salud óptimos bajo diferentes parámetros ambientales, condiciones sanitarias o estrés térmico.
Feedstim® Poultry
Productos de transformación de plantas
ProActiv’® Poultry está formulado a partir de un complejo de fitosoluciones activas cuidadosamente seleccionadas con un efecto sinérgico probado sobre la integridad y el equilibrio del tubo digestivo de las aves.
ProActiv'® Poultry
Monogástricos, incorporación continua al alimento o en cura.
Combinación de plantas estandardizada Alcachofa, Boldo, Silbato de Leche y Romero L-Carnitina
Brio Liver C es una fito-solución compleja dedicada a los animales monogástricos para apoyar el rendimiento cuando el hígado está sobre exigido.Se compone de L carnitina, implicada en el metabolismo de las grasas en el hígado. Además, una mezcla de materias primas vegetales para apoyar y mejorar el consumo.
BrioLiver
Argentina, Bolivia, Brazil, Chile, Colombia, Costa Rica, Ecuador, El Salvador, Guatemala, Mexico, Panama, Paraguay,Peru, Rep Dominicana, Uruguay, Venezuela.
Cerdos, incorporar en el alimento de manera continua durante los períodos de constante o crónico estrés por calor.
Sustancias tampón
Capsaicina, antioxidantes
Thermo® Control actua sobre las consecuencias del estrés calórico. Fortaleciendo los sistemas naturales involucrados en las pérdidas de calor, compensa el bicarbonato y estimula la ingestión.
Thermo®Control

Rumiantes
Rumiantes
Scutellaria baïcalensis, anti-oxydants
Feedstim(r) Dairy está especialmente formulado con una mezcla de vitaminas y productos de origen vegetal, ricos en polifenoles, incluida la scutellaria baïcalensis. Este último ha sido patentado por el grupo CCPA para mejorar la lactancia en animales productores. La Scutellaria contribuye a la defensa celular y a la lucha contra contra el estrés oxidativo y la inflamación. Feedstim® Dairy actúa en todo el cuerpo, especialmente en la ubre.
Feedstim® Dairy
tampón
Sustancias
Capsaicina, antioxidantes
Para ayudar a los rumiantes durante la temporada de calor, el Grupo CCPA ha desarrollado Thermo® Plus. Thermo® Plus actúa sobre las consecuencias del estrés térmico. Fortalece los sistemas naturales involucrados en la evacuación del calor, compensa el electrolito y estimula la ingestión. De este modo, Thermo® Plus garantiza el confort y el bienestar de los animales y, por lo tanto, optimiza la producción de leche y el rendimiento del crecimiento
Thermo®Plus
ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES ACTIVOS FITOGÉNICOS
Combinación de extractos de plantas estandarizado en Flavonoides.
Combinación de extractos de plantas estandarizado en Flavonoides + terpenos.
Combinación de extractos de plantas estandarizados en Flavonoides + ácidos cafeoilquínicos.
Combinación de extractos de plantas estandarizados en Flavonoides + ácidos cafeoilquínicos + Betaina.
Combinación de extractos de plantas estandarizados en Flavonoides + terpenos + ácidos cafeoilquínicos
Combinación de extractos de plantas estandarizados en Flavonoides + ácidos cafeoilquínicos
Todas las especies animales (monogástricos, rumiantes, acuicultura y PETFOOD). Dosificación recomendada: consultar con Departamento Tecnico de ENA.
Combinación de extractos de plantas estandarizados en flavonoides, terpenos y betaina
Combinación de extractos de plantas estandarizados y Ácidos orgánicos.
Combinación de extractos de plantas estandarizada, Ácidos orgánicos, monogliceridos y galactomananos
Combinación de extractos de plantas estandarizada en terpenos, sulfuros y compuestos fenolicos.
Combinación de extractos de plantas estandarizados en aldehidos, flavonoides y compuestos fenolicos.
Combinación de extractos de plantas estandarizados en alcaloides, saponinas, lactona y compuestos fenolicos.
Combinación de extractos de plantas estandarizados en terpenos, alcaloides y flavonoides.
Combinación de extractos de plantas estandarizados e glicocomponentes y saponinas.
ACCIÓN
PRODUCTO
ENACOX2® Reduce la respuesta inflamatoria y su coste metabólico . Sin efectos secundarios, lo que permite suministrarlo por tiempos prolongados.
ENARELAX® Reduce el estrés, inhibiendo la transmisión nerviosa de las señales de ansiedad o excitación. Efectivo contra caudofagia, picaje o montas.
ENADETOX® Hepatomodulador. Incrementa la producción y liberación de bilis. Aumenta la absorción de ácidos grasos, vitaminas liposolubles y calcio.
ENABOOST® Incrementa productividad y prolificidad mediante una mejora en la digestibilidad de los nutrientes y su eficacia antioxidante.
ENALACT® Aumenta la producción láctea y su % de grasa. Mejora peso al destete, y previene mamitis.
ENATERM® Optimiza el Índice de Conversión y la ganancia de peso durante la etapa de cebo. Mejora redimiendo y calidad de la canal.
ENASTART® Para un buen arranque, aumenta la capacidad del animal de adaptarse a los cambios, mejorando la producción. (ej: destete, cambios en el pienso, agrupamientos).
ENASSENCE® Actúa contra bacterias patógenas y coccidios, sin perjudicar a la flora intestinal endógena.
ENAGUT® Estrategia multidiana para la mejora de la salud intestinal mediante un efecto bacteriostático, prebiotico e inmunomodulador.
ENACOLIT® Diseñado para la prevención y tratamiento de diarreas en avicultura y porcino.
ENAPREVEN® Estabiliza la microbiota intestinal, evitando así la proliferación del Clostridium, causante de la enteritis necrótica en las aves.
ENAPROOT® Actividad antiprotozoaria que evita la proliferación de coccidios.
ENARESP® Eficaz frente a los transtornos respiratorios mediante su efecto bacteriostático, inmunomodulador y expectorante.
ENAPURE® Mejora las condiciones ambientales mediante una reducción y control del amoniaco producido, además de otros gases y malos olores.
Empresa del grupo

ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES ACTIVOS FITOGÉNICOS
ACCIÓN
PRODUCTO
Porcino: En lactación, diluir
800 g en 20 de agua y administrar 0,5 de esta solución por camada, del 2º al 8º día de vida y los dos días previos al destete En destete, junto con el pienso, añadir 1,5 de esta solución durante 2 días.
Todas las especies (monogástricos, rumiantes y acuicultura): Dosis entre 75-500 g/T dependiendo de la especie y fase productiva del animal. Consultar con el dep. técnico
Todas las especies (monogástricos, rumiantes y acuicultura): Consultar con el dep. técnico
Todas las especies (monogástricos, rumiantes y acuicultura): Consultar con el dep. técnico
Porcino: Como preventivo: 1kg/T de pienso. Para control: hasta 3 kg/T de pienso
Compuestos bioactivos de algas
Combinación específica de aceites esenciales microencapsulados con tecnología propia de Igusol
Mezcla de extractos de plantas entre ellos alcachofa.
Extractos de plantas
Mezcla de extractos de plantas
Actúa sobre la pared del intestino aumentando la capacidad de absorción de los nutrientes. Estimula el consumo voluntario, fortalece el aparato digestivo y aporta vigor e inmunoestimulación
Combinación de aceites esenciales a una alta concentración, con propiedades antibacterianas, antiinflamatorias e inmunoestimulantes, que actúan modulando la microbiota intestinal y previniendo trastornos digestivos.
Activador de la función hepática que fomenta una actividad antioxidante, antiinflamatoria y protectora de los hepatocitos, con funciones coleréticas y colagogas
Fuente nutricional de energía que maximiza el aprovechamiento energético de la ración en las fases de máxima exigencia productiva
Pienso complementario natural para el control y prevención de la necrosis auricular en cerdos
GreenTONIC
Emerald
Igusafe
Solergy
NecroSTOP
PAÍSES EN LATINOAMÉRICA DONDE EL PRODUCTO ESTÁ DISPONIBLE
ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS)
COMPONENTES ACTIVOS FITOGÉNICOS
Monogástricos. Uso en mezcladora de 1 a 3 kg/t de pienso.
Pocino. Uso continuado a 1kg/t de pienso. Uso en eventos a 2kg/t de pienso.
Porcino. Uso en mezcladora a 1kg/t de pienso de cerdas en periparto y lactantes. Uso a 1kg/t en Lechones hasta 70 días de vida. Utilizar en el caso de riesgo de Estreptococias.
Porcino. Uso en mezcladora a 2kg/t de pienso. Utilizar en caso de riesgo durante los trastornos digestivos y en su convalecencia.
Porcino y cunicultura: De 0,25 a 0,5 cc/L en agua de bebida. Aves: De 0,15 a 0,3 cc/L en agua de bebida. Rumiantes y pequeños rumiantes: De 0,25 a 0,5 cc/L en agua de bebida.
Porcino: De 250 a 700ml/Mt de agua de bebida
Consultar al departamento técnico-comercial de Liptosa.
Aves: De 150 a 300ml/Mt de agua de bebida Hasta 7 días consecutivos
Monogastricos: De 0,25 a 1L/Mt de agua
Rumiantes: De 0,25 a 0,5L/Mt de agua de bebida
Porcino y cunicultura: De 0,5 a 1 cc/L en agua de bebida durante 7-10 días.
Aves: De 0,5 a 0,7 cc/L en agua de bebida durante 7-10 días. Rumiantes y pequeños rumiantes: de 1 a 1,5cc/L en lactorremplazante reconstituido y de 0,3 a 0,6 cc/L en agua de bebida durante 10-15 días en cebo.
ACCIÓN
Aditivo equilibrador metabólico que contribuye a un rendimiento óptimo en producción animal. Aceites esenciales protegidos por componentes técnicos. Ácidos orgánicos
Ingredientes vegetales activos, seleccionados con precisión, que ofrecen un efecto sinérgico comprobado para apoyar la integridad del sistema digestivo en cerdos. Aceites esenciales y botánicos protegidos por componentes técnicos. Minerales quelados
PRODUCTO
HYGEN PRO
HYGEN PRO LAW
Monoglicéridos y ácidos grasos
Aceites esenciales y botánicos protegidos por componentes técnicos.
Formulado a partir de un complejo de fitosoluciones activas cuidadosamente seleccionadas con un efecto sinérgico probado, que mejora los rendimientos productivos a través de la modulación de microbioma respiratorio en porcino.
HYGEN PRO STREPT
Aceites esenciales y botánicos protegidos por componentes técnicos. Minerales quelados
Formulado a partir de un complejo de fitosoluciones activas cuidadosamente seleccionadas con un efecto sinérgico probado, que mejora los rendimientos productivos a través de la modulación de microbioma digestivo en porcino.
HYGEN PRO COMPLET
Monoglicéridos, ácidos grasos, vitaminas, provitaminas y fuente de salicilatos
Aceites esenciales y botánicos protegidos por el sistema LSP (Liptosa Smart Protection)
Ayuda a mitigar los efectos del estrés térmico, reforzando los sistemas fisiológicos que facilitan la termoregulación.
HYGEN PRO RESPIRFRESH
Monoglicéridos y ácidos grasos
Apoya el equilibrio de los procesos entéricos y favorece un buen desempeño zootécnico. "Aceites esenciales y botánicos protegidos por componentes técnicos”
LIPTOCOL
Botánicos Ácidos grasos, productos de levadura
Enfoque completo para ayudar a preservar el óptimo funcionamiento de los órganos clave y las funciones metabólicas frente a los efectos de micotoxinas y desbalances nutricionales.
LIPTOSAFE L
Monoglicéridos y ácidos grasos.
Apoyo en la superación de diversos desafíos (ambientales y sanitarios) durante la etapa de cría, asegurando un rendimiento y condiciones óptimas. Aceites esenciales y extractos de plantas protegidos por el sistema LSP (Liptosa Smart Protection).
COCCILIP
Monogástricos
0.51L / 1000L de agua de bebida
Monoglicéridos, ácidos grasos de cadena corta y media, propilenglicol, aromatizantes y conservantes.
"Mezcla sinérgica de ácidos grasos y botánicos diseñada para favorecer el desarrollo de la microbiota saprófita ejerciendo así mismo un control efectivo de bacterias Gram +, a lo largo de todo el tracto intestinal. En agua potable:Para la recepción, post-vacunación y el estrés del transporte.Para los problemas con calidad de agua y otros.Para los cambios de pienso.Es compatible con el uso de antibióticos.
LIPTODAL G+
*Cierta información asociada a los productos, su composición y alegaciones puede ser diferente según la región geográfica y no ser aplicable en todos los países. Liptosa se reserva el derecho de adaptarse a los requisitos y legislación de cada caso.
Así mismo, la marca y nombre del producto puede diferir de un país a otro. Contacte con nuestro departamento técnico-comercial para obtener información adicional.
ESPECIES DE DESTINO Y DOSIFICACIÓN PAÍSES EN LATINOAMÉRICA DONDE EL PRODUCTO ESTÁ DISPONIBLE
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS)
COMPONENTES ACTIVOS FITOGÉNICOS
Colombia, República Dominicana, Mexico
Cerdos y aves de corral: 100200 g por tonelada de alimento completo
Diversos
Pollitos: 100 ml Miarom Intest L por 1.000 de agua de bebida Aves de corral: 100-250 ml Miarom Intest L por 1.000 de agua potable
Lechones destetados: 50100 ml Miarom Intest L por 1.000 l de agua de bebida
Engorde: 50100 ml Miarom Intest L por 1.000 l de agua de bebida
Bolivia, República Dominicana
Cerdos: 200500 g por tonelada de pienso completo Aves de corral: 150350 g por tonelada de pienso completo
Boliva, Chile, Colombia, Costa Rica, República Dominicana, Equador, El Salvador, Guatemala, Honduras, Mexico, Nicaragua, Perú
Pollitos: 100 ml Miarom Classic L cada 1.000 l de agua de bebida Aves de corral: 100-250 ml Miarom Classic L cada 1.000 de agua de bebida
Lechones: 150 ml Miarom Classic L cada 1.000 l de agua de bebida Engorde/ cerdas: 200 ml Miarom Classic L cada 1.000 l de agua de bebida
Chile, Colombia, República Dominicana, Ecuador, El Salvador, Guatemala, Honduras, México, Perú
Lechones: Administración oral 2 ml (1 dosis)
ACCIÓN
Aceites esenciales recubiertos y fitógenos molidos
Alimento complementario, que es una combinación especial de aceites esenciales de alcaravea, anís estrellado, manzanilla, jengibre, canela y orégano. Aumenta la ganancia diaria de peso y mejora la conversión alimenticia, al influir positivamente en la salud intestinal.
Miarom Intest C
Mezcla de aceites esenciales
Miarom Intest L Miarom Intest L es un líquido complementario que proporciona una combinación específica de compuestos fitogénicos variados, entre los que están los componentes activos de alcaravea, anís estrellado, camomila, jengibre, canela y orégano. El sinergismo perfecto de hierbas beneficiosas garantiza la salud intestinal, tanto en la prevención como en caso de problemas digestivos. En la granja, la administración es fácil y flexible gracias a su aplicación en el agua de bebida.
Mezcla de aceites esenciales
Alimento complementario que es una combinación sinérgica de aceites esenciales de tomillo, anís, menta y eucalipto, entre otros. Favorece la salud respiratoria y, además, mejora la calidad del aire en las instalaciones.
Classic P
Miarom

Mezcla de aceites esenciales
Miarom Classic L es un complemento alimenticio líquido que contiene una combinación especial de varios aceites esenciales. Sus beneficios duraderos provienen de una combinación sinérgica y estandarizada de aceites esenciales de tomillo, anís, menta y eucalipto, entre otros. Favorece la salud respiratoria y alivia el estrés térmico. En la granja, la administración es fácil y flexible gracias a su aplicación en el agua de bebida.
Classic L
Miarom
Electrolitos
Taninos del extracto de castana y aceites esenciales
Alimento complementario dietético líquido para estabilizar el equilibrio hídrico y electrolítico de los lechones lactantes durante los episodios de diarrea. En caso de tratamiento y convalecencía de díarreas inespecífícas.
MiaFirstAid L
PAÍSES EN LATINOAMÉRICA DONDE EL PRODUCTO ESTÁ DISPONIBLE
ESPECIES DE DESTINO Y DOSIFICACIÓN
ACTIVOS FITOGÉNICOS
COMPONENTES
Aves: 200-500 g por tonelada de alimento
Brazil, Perú, Chile, Ecuador, Guatemala, México, Honduras, Argentina, El Salvador, Costa Rica
Porcinos: 500 g1 kg por tonelada de alimento
Rumiantes: 50-75 g por animal por día o según recomendación del nutricionista
Aves: 300-500 g por tonelada de alimento
Porcinos: 400-600 g por tonelada de alimento
Brasil, Perú, Chile, Ecuador, Guatemala, México, Argentina
Rumiantes: 750 g1 kg por tonelada de alimento. 1 kg por tonelada para animales de alta producción láctea o según recomendación del nutricionista
Aves, para prevención: 1 kg por tonelada de alimento o según indicación del nutricionista
Aves, como soporte: 2-3 kg por tonelada de alimento
Porcinos, para prevención: 1-2 kg por tonelada de alimento Porcinos, como soporte: 2-3 kg por tonelada de alimento La tasa de mezcla puede ser modificada por un nutricionista Brasil, Perú, Chile, Ecuador, Guatemala, México
Aves: 300-500 g por tonelada de alimento
Porcinos: 400-600 g por tonelada de alimento
Perú
ACCIÓN
Acacia nilotica Curcuma longa Soya Lecithin
Alternativa más ecológica a la colina sintética
Emblica officinalis Ocimum sanctum Withania somnifera
Adaptógeno y antiestresante
Acacia arabica
Andrographis paniculata Holarrhena antidysenterica Punica granatum
Terminalia bellerica
Rumiantes: 750 g1 kg por tonelada de alimento. 1 kg por tonelada para animales de alta producción láctea o según recomendación del nutricionista
Andrographis paniculata Boerhavia diffusa Solanum nigrum Phyllanthus amamrus
Agente aglutinante para heces sueltas
Optimizador del rendimiento hepático
PRODUCTO
Kolin Plus
Phytocee
Stodi
Zigbir
DONDE EL PRODUCTO ESTÁ DISPONIBLE
PAÍSES EN LATINOAMÉRICA
ESPECIES DE DESTINO Y DOSIFICACIÓN
ACTIVOS FITOGÉNICOS
COMPONENTES
ACCIÓN
Brasil, Chile, Colombia, Ecuador, Mexico, Perú.
Timol y Carvacrol, Mono, Diglicéridos, Triglicéridos de ácidos grasos, ácido silícico precipitado y seco. Como componente de una premezcla o la ración completa. Tasa recomendada de 15 a 60 g/t Bovinos, cerdos, aves
Aditivo zootécnico, mezcla de aceites esenciales que contiene timol y carvacrol
150
PRODUCTO
ENHANCE®
Todas las especies: 100-350g/tn alimento balanceado. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada. Argentina, Bolivia, Chile, Costa Rica, Guatemala, Honduras, Panama, Ecuador, México, Peru, Colombia, Republica Dominicana, Paraguay
Fosfatidicolina, rutina, terpenoides
Aditivo modulador metabólico natural, que mejora el desempeño y la calidad de canal a través de la activación de los receptores PPARα. Participación en procesos metabólicos a nivel del uso de la energía de la dieta. Su contenido en fosfatidilcolina tambien contribuye a sustituir el cloruro de colina de la dieta.
BioCholine®
Todas las especies: reemplazo del 50% de metionina sintética. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada. Argentina, Costa Rica, Ecuador, México, Peru
Todas las especies: 500g/tn alimento balanceado. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada. Argentina, Brasil, Bolivia, Chile, Costa Rica, Guatemala, Honduras, El Salvador, Panama, Ecuador, México, Peru, Colombia, Republica Dominicana, Paraguay
Todas las especies: 250g/tn alimento balanceado. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada.
Aves, cerdos, conejos y rumiantes. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada. Argentina, Brasil, Bolivia, Chile, Costa Rica, Ecuador, México, Colombia, Paraguay
Chile, Argentina, El Salvador, Ecuador, Peru
Todas las especies: 100-350g/tn alimento balanceado. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada.
Todas las especies: 100-200g/tn alimento balanceado. Consulte con su técnico de Nuproxa para establecer una dosificación personalizada. Argentina, Brasil, Bolivia, Chile, Costa Rica, Honduras, El Salvador, Panama, Ecuador, México, Colombia, Paraguay
Chile, Argentina, Bolivia, Costa Rica, Guatemala, Honduras, El Salvador, Panama, Ecuador, Peru, Colombia, Republica Dominicana, Paraguay
Consulte con su técnico de Nuproxa para establecer una dosificación personalizada.
Organosulfurados
Mejora la eficiencia nutricional a través de la optimizacion del metabolismo de la metionina. Sustituye parcialmente a la Metionina sintética reduciendo el coste de formulación.
OptiMethione™
Taninos, saponinas, polifenoles, terpenoides
Mejora del rendimiento productivo de los animales a través de la modulación de la microbiota intestinal, reducción de la inflamación crónica y del estrés oxidativo. Además, contribuye en gran medida al control de protozoos intestinales.
PeptaSan™
Terpenoides y flavonoides
Aditivo natural que mejora los rendimientos productivos de los animales por su efecto protector de la función hepática gracias a su actividad antioxidante, detoxificante y estimulante de la secrección biliar.
LivoLiv 250™
Polifenoles
Antioxidante metabólico natural para la proteccion del estrés oxidativo de los animales contribuyendo para la mejora de parámetros productivos, reproductivos y de calidad de carne. Sustituto parcial de la Vitamina E sintética reduciendo el coste de formulación.
NuxaFen™
Tocoferoles
Producto natural que contiene diferentes isómeros de la Vitamina E altamente biodisponibles, con rápida absorción y lenta excreción.
Herbal-E™
Polifenoles, ácido gálico y sus derivados
Poderoso antioxidante, utilizado para frenar el estrés oxidativo y el estrés por calor. Es estable en medio acuoso, a temperatura ambiente y a temperaturas de granulación o extrusión.
C-Power™
Único aditivo 100% natural fuente del metabolitito activo de la Vitamina D 1,25 (OH)2D3. 1,25(OH)2D3
Panbonis®
PAÍSES EN LATINOAMÉRICA DONDE EL PRODUCTO ESTÁ DISPONIBLE
ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS)
COMPONENTES ACTIVOS FITOGÉNICOS
ACCIÓN
Avicultura y porcino: 20 ‐50 g / t de pienso
Vacuno de leche y vacuno de engorde: 250 –300 mg / animal día Ovinos y caprinos: 40 –50 mg / animal día
Avicultura: 100200 g / t de pienso
Porcino: 150 –200 g / t de pienso
Corderos: 250 –500 g / t de pienso
Vacuno de engorde: 8g/animal/día
Avicultura: 150 –250 ml / 1000 L de agua
Conejos: 150 –250 ml / 1000 L de agua
Porcino: 250 –350 ml / 1000 L de agua
Rumiantes: 150 –250 ml / 1000 L de agua
Terneros lactantes: 1 –2 ml / animal / día (añadir a la leche en una sola toma)
Todas las especies: 0,25-1kg/t de pienso (Naturcox Powder)
Chile, Argentina, Ecuador, Colombia, Panama, Mexico
Todas las especies. 250ml/1000L de agua de bebida ó 350 -800g/t de pienso (Licorol Powder)
PRODUCTO
Oleorresina de capsicum
Aumenta el rendimiento productivo, estimula la digestión y reduce el impacto del estrés térmico
PHYTMAX CAPSICUM ENCAPS
Oleorresina de capsicum, cinamaldehído,cúrcuma y extracto de pimienta negra
Bioprotector natural con alta capacidad antioxidante e inmunomoduladora
Oleoressina de capsicum, cinamaldehído y eugenol
Aumenta el rendimiento productivo, estimula el sistema digestivo y el apetito, modifica el patrón de fermentación ruminal y mejora el sistema inmunitario previniendo el riesgo de sufrir acidosis y problemas de laminitis.
Compuestos organosulfurados derivados de Allium sativum , timol, carvacrol.
Modula la microbiota intestinal, genera un ambiente hostil para el desarrollo de bacterias y protozoos, mejora la respuesta inmunitaria y tiene propiedades antiinflamatorias
Purifica las vías respiratorias, promueve la activación ciliar, diluye la concentración de moco y mejora el bienestar animal en épocas de estrés. Mentol y Eucalyptus globulus
Porcino: 1 –2 Kg /t de pienso
Monoglicéridos de ácidos láurico, cáprico y caprílico
Taninos purificados de castaño, carvacrol y extracto de Glycyrrhiza glabra
Controla las diarreas de origen bacteriano
Todas las especies: 0,5-1L/1000L de agua
Avicultura: 0,25 –1,5 Kg / t de pienso
Porcino: 1 –3 Kg /t de pienso
Rumiantes: 1 –3 Kg /t de pienso
Monoglicéridos de ácido butírico, ácido caprílico, ácido cáprico y ácido láurico (y manano-oligosacáridos en Phytobutyrin plus Powder)
Incrementa el rendimiento productivo a través de la mejora de la integridad intestinal y la modulación del sistema inmunitario. Controla el desarrollo de bacterias Gram –negativas” Carvacrol y cinamaldehído
Porcino: 0,8 –1,5 L/ 1000L de agua de bebida.
Avicultura: 0,8-1,5 L/1000L de agua de bebida
Conejos: 0,8-1,5L/1000L de agua de bebida
Ternero de engorde: 5-10ml/animal/día
Vacuno de engorde: 35ml/animal/día
Vacuno de leche: 40-50ml/animal/día
Todas las especies: 1-2kg/t de pienso
Aceite esencial de Gaultheria procumbens y extracto de Withania somnifera
Modula el proceso inflamatorio y minimiza los efectos negativos del estrés reduciendo la agresividad
Aceite esencial de Gaultheria procumbens y extractos de Filipendula ulmaria y Ribes nigrum
Modula el proceso inflamatorio a través de la gestión de la situación de malestar relacionada con la fiebre/dolor/estrés con el fin de recuperar los niveles productivos normales
PHYTMAX COX
PHYTMAX BEEF CATTLE
COXSAN
LICOROL
GASTROHERB PLUS
PHYTOBUTYRIN PLUS
THERMUM
PIRESTOP
PAÍSES EN LATINOAMÉRICA
DONDE EL PRODUCTO ESTÁ DISPONIBLE
ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS)
COMPONENTES ACTIVOS FITOGÉNICOS
ACCIÓN
Vacunos lecheros: 2-4 g/animal/día
Ovinos: 0,3 g/animal/día Uruguay, Brasil y Colombia Argentina y México bajo solicitud.
Vacunos de carne: 1-3 g/animal/día
Porcino, Aves, Ganado lechero, Ganado Vacuno, Rumiantes. Consulte la etiqueta para conocer los niveles de dosificación recomendados.
México, El Salvador, Guatemala, Honduras, Nicaragua, Costa Rica, Panamá, Colombia, Ecuador, Perú, Bolivia, Brasil, Uruguay, Paraguay, Chile, Argentina, República Dominicana, Puerto Rico.
Porcino. Mezclar a razón de 2 lbs. por tonelada con otros ingredientes para hacer un alimento completo.
Ganado vacuno, Rumiantes. Suministrar a todas las clases de rumiantes a razón de 1/2 gramo por 100 libras de peso corporal.
Capsicum encapsulado, Actifibe® Prebiótico
Mezcla de aceites esenciales: clavos (Syzygium aromaticum) coriandro (Coriandrum sativum) , geranio (Pelargonium cucullatum) taninos de castaño (castanea sativa) , bioflavonoides de la oliva (Olea europaea)
Reduce las emisiones de metano en rumiantes, modulador de la microbiota ruminal, promotor de crecimiento en rumiantes, antiinflamatorio y antioxidante, control de la degradación nitrogenada, acción anti estrés
El aditivo alimentario natural más confiable para ayudar a los animales a responder a eventos de estrés; contiene aceites esenciales
El aditivo natural para el agua más confiable para ayudar a los animales a responder a eventos de estrés; contiene aceites esenciales
Aditivo alimentario natural de uso diario, que contiene aceites esenciales, que ayuda a enfrentar una amplia gama de problemas de salud
Aditivo natural de uso diario para el agua, que contiene aceites esenciales, que ayuda a enfrentar una amplia gama de problemas de salud
Suplemento alimentario natural que ayuda a respaldar su programa de bioseguridad alimentaria
Ganado vacuno, Rumiantes. Administrar por vía oral 2 cápsulas por cabeza y día durante las épocas de desafío. Repetir a las 24 horas si es necesario.
Electrolitos, Actifibe® Prebiótico
Ganado vacuno, Rumiantes. Administrar por vía oral 2 cápsulas por cabeza y día durante las épocas de desafío. Repetir si es necesario.
Mezcla de electrolitos, complejo vitamínico y mineral, prebiótico Actifibe®
Microbial Catalyst®, prebiótico Actifibe®
Tecnología
Ganado vacuno, Rumiantes. Alimentar a razón de 20 gramos por 100 libras de peso corporal. PRODUCTO
ANAVRIN®
Regano® Dry
Regano® Liquido
ProsperEO™ Dry
ProsperEO™ Liquido
Dual Defender™
Aceites esenciales
Microfused®: Mezcla específica de fitonutrientes
Aditivo alimentario de uso diario que ayuda al ganado en épocas de desafíos ambientales
Comfort™
Cápsulas naturales que favorecen la salud digestiva en épocas de malestar gastrointestinal
Strong™ For Calf Stress
Fight
Cápsulas naturales que ayudan a que el ganado vuelva a alimentarse
Fight Strong™ For Cow Stress
Aditivo natural para piensos granulados que hace que el ganado coma y ayuda a reforzar la inmunidad
Fight Strong™ Pellet
PAÍSES EN LATINOAMÉRICA
DONDE EL PRODUCTO ESTÁ DISPONIBLE
COMPONENTES SECUNDARIOS (NO FITOGÉNICOS) ESPECIES DE DESTINO Y DOSIFICACIÓN
COMPONENTES ACTIVOS FITOGÉNICOS
ACCIÓN
Ganado vacuno, Rumiantes. Alimente 1 gramo por 100 libras de peso corporal continuamente. Alimente 2 gramos por 100 lbs. de peso corporal durante épocas de estrés.
Ganado vacuno, Rumiantes. Administrar por vía oral 10 ml (1 bomba = 5 ml) dentro de las 12 horas del nacimiento. Repetir durante 1-2 días si el ternero no tiene energía.
Ganado vacuno, Ganado lechero, Rumiantes. Utilizar 2 cápsulas intrauterinas 12-24 horas después del parto. Puede repetirse 2 días adicionales si es necesario.
México, El Salvador, Guatemala, Honduras, Nicaragua, Costa Rica, Panamá, Colombia, Ecuador, Perú, Bolivia, Brasil, Uruguay, Paraguay, Chile, Argentina, República Dominicana, Puerto Rico.
Ganado vacuno, Ganado lechero, Rumiantes. Consulte las instrucciones en la etiqueta.
Ganado vacuno, Ganado lechero, Rumiantes. Alimente 1 onza por cabeza por día.
Ganado vacuno, Ganado lechero, Rumiantes. Administrar 1 lb. por vaca a través de agua, drench o aplicación de alimento inmediatamente después del parto.
Acuícola. Consulte las instrucciones en la etiqueta.
Porcino. Administrar por vía oral 2 ml (1bomba) por lechón dentro de las primeras 24 horas después del nacimiento. Repetir a las 24 horas según sea necesario. También puede utilizarse para cerdos rezagados. Administrar por vía oral 2 ml (1 bomba) por cerdo.
PRODUCTO
Aditivo alimentario natural para ganado para ayudar a mantener la salud y la vitalidad Tecnología Microbial Catalyst® -
Stand Strong™ For Ruminants
Microfused®: Mezcla específica de fitonutrientes Fuente de energía compleja, prebiótico Actifibe
Suplemento oral natural que apoya el vigor y el apetito de los terneros
Start Strong™ For Calves
Yucca Schidigera, Actifibe® Prebiótico
Actifibe® Prebiótico
Aceites esenciales
Fight Strong™ For Uterine Balance Cápsulas naturales que apoyan los desafíos uterinos después del parto
Un aditivo lácteo de avanzada que promueve la inmunidad, estimula el apetito y facilita la transición a una dieta de granos
Stay Strong™ For Dairy Calves
Catalyst® IntegraFlo™
Microbial
Tecnología
Aditivo natural para alimentación de vacas lecheras que ayuda a mantener la salud y la vitalidad con IntegraFlo™
Stand Strong™ For Dairy Cows
3 Calcuim
Blend, Vitaminas A, B, C, D, E, K y Niacina, Actifibe® Prebiótico
Paquete para rendimiento natural para vacas inmediatamente después del parto
Start Strong™ For Fresh Cows
Aceites esenciales
Un aditivo alimentario natural de uso diario y muy sabroso que estimula el apetito, promueve la salud intestinal y fortalece la inmunidad Yucca Schidigera, Actifibe® Prebiótico
Bioactivos del calostro, prebiótico Actifibe®
Vitaminas D y E, mezcla de electrolitos Porcino. Véanse las instrucciones en la etiqueta.
Microfused®: Mezcla específica de fitonutrientes
One Current™ Liquid
Suplemento oral natural para que los lechones se levanten y coman rápidamente
First Pulse® D
Actifibe®
Mega Pro™ Un suplemento oral que contiene anticuerpos de huevo que ayuda a reducir los desafíos gastrointestinales en los cerdos antes o después del destete Tecnología prebiótica

¿Quién dijo que la nutrición animal era aburrida?




PAÍSES EN DISTRIBUCIÓN
RECOMENDACIONES DE USO
DOSIS ESPECIES DE DESTINO
50g/T
Incorporación en mezclas de alimentos
Monogástricos y acuaculturas
25g/500 FTUs
COMPOSICIÓN
PRODUCTO ACTIVIDAD ENZIMÁTICA
Endo-1,3(4)-β-glucanase Arabinofuranosidasa a Complejo multi-enzimático concentrado compuesto por xilanasa, β-glucanasa, celulasa, arabinofuranosidasa termoestable.
Endo-1,4-xilanasa
Rovabio Advance
Rovabio PhyPlus
20.000 6-fitasa a Fitasa
100g/T
Enzima para uso en alimentos de Aves y Cerdos. -
Aves y Cerdos
Aves: Como componente de la ración completa en un nivel de inclusión de 0.250.5 kg/t de alimento
Cerdos: Como componente de la ración completa en un nivel de inclusión de 0.5 kg/t de alimento
Recomendado en sistemas de producción intensivos. La incorporación de RUMINOZYME al alimento mejora la performance productiva de rumiantes lecheros y de engorde, incrementando la producción de leche y la ganancia de peso respectivamente.
Rumiantes
Endo-1,3(4)-β-glucanase 6-fitasa Arabinofuranosidasa a Complejo multi-enzimático compuesto por xilanasa, β-glucanasa, celulasa, arabinofuranosidasa y 1.000FTU de fitasa
Extracto seco de los solubles de la fermentación de Bacillus licheniformis PWD-1 mezclado con carbonato de calcio y saborizante natural.
Endo-1,4-xilanasa
Rovabio Advance Phy
Proteasa mín 600,000 U/g
CIBENZA® DP100
Producto seco de fermentación de Aspergillus oryzae , 5 g Marlo de maíz molido c.s.p, 100g Xilanasas (500 UI/G) Aspergillus oryzae 2x10⁴ UFC/g 1 Kg por Tn de alimento
Producto enzimático con actividad xilanasa, efectiva para la degradación de material fibroso vegetal en alimentos para bovinos lecheros y de engorde. Mejora la digestibilidad de la materia seca y de la fibra detergente neutra.
Rumino-Zyme
Tellez-Isaias G 1*, Eisenreich W 2*, Petrone-Garcia VM 3 , Hernandez-Velasco X 4, Castellanos-Huerta I 1,5, Tellez Jr. G 6 , Latorre JD 1, Bottje WG 1, Señas-Cuesta R 1, Coles ME 1, Hargis BM 1, El-Ashram S 7, Graham BD 1, Shehata AA 8, Iglesias BF 9
1. Department of Poultry Science, University of Arkansas Agricultural Experiment Station, Fayetteville, AR, USA.
2. Structural Biochemistry of Membranes, Bavarian NMR Center, Technical University of Munich (TUM), Garching, Alemania.
3. Facultad de Estudios Superiores Cuautitlan, Universidad Nacional Autónoma de México (UNAM), Cuautitlan Izcalli, Estado de México, México.
4. Departamento de Medicina y Zootecnia de Aves, Facultad de Medicina Veterinaria y Zootecnia (FMVZ), UNAM, Cd. De México, México.
5. Facultad de Ciencias Naturales, Licenciatura en Medicina Veterinaria y Zootecnia, Universidad Autónoma de Querétaro, Querétaro, México.
6. Department of Developmental Biology, Roslin Institute, University of Edinburgh, UK.
7. College of Life Science and Engineering, Foshan University, Foshan, Guangdong, China.
8. Prophy-Institute for Applied Prophylaxis, Bönen, Alemania.
9. Sección Avicultura, EEA Pergamino, INTA, Argentina.
Los pollos de engorde modernos son, sin duda, la manifestación más visible de los avances genéticos que, acompañados de la alimentación, las medidas sanitarias y las prácticas de manejo intensivo, han dado lugar al éxito de esta producción. Sin embargo, para alcanzar estos objetivos, es necesario mantener el tracto gastrointestinal (TGI) en buen estado.
El TGI es el órgano responsable de la digestión y absorción de nutrientes. El costo de la coccidiosis subclínica o la enteritis necrótica en los pollos de engorde es, probablemente, mayor que el de las infecciones agudas clínicamente identificadas en aves de corral, dado que la conversión alimenticia representa más del 70% del costo total de producción (Aggrey et al., 2010).
A medida que se acorta el período de crecimiento de los pollos de engorde y mejora la eficiencia alimenticia, la necesidad de programas mejorados de salud y nutrición se vuelve más evidente.
Debido a que los cambios durante el desarrollo intestinal son pequeños, a menudo se pasan por alto, por lo que, la salud intestinal impacta en la salud general y la productividad.
La sustitución de antibióticos en los sistemas de producción por productos alternativos, mejores prácticas de manejo, estricta bioseguridad, ingredientes de calidad, ausencia de enfermedades (Mycoplasma/Salmonella) y programas efectivos de inmunización son estrategias eficaces para alcanzar los objetivos de salud y productividad.
En este trabajo, nos centramos en revisar las repercusiones significativas del estrés crónico y la inflamación intestinal en la salud y el rendimiento de las aves comerciales.
Además de digerir alimentos y absorber agua y nutrientes, el tracto intestinal contiene una comunidad microbiana diversa y compleja (Celluzzi & Masotti, 2016), así como un sistema nervioso entérico (SNE) que es considerado el “segundo cerebro” del organismo (Schneider et al., 2019).
Esta población microbiana puede igualar en número a las células somáticas encontradas en este tejido (Wallis et al., 2004; Zhu et al., 2010; Li et al., 2014; Sender et al., 2016)
Aparte de la complejidad en la estructura y las relaciones microbianas, en los pollos, el tejido linfoide asociado al intestino (GALT, por sus siglas en inglés) contiene la mayor concentración de células inmunitarias en el organismo, lo que demuestra su gran importancia (Peralta et al., 2017; Casteleyn et al., 2010).
Asimismo, el aparato digestivo tiene órganos linfoides primarios como la bolsa de Fabricio, donde se originan y multiplican los linfocitos B; este componente del GALT, en las especies aviares, es esencial para proteger al tracto digestivo (Bar-Shira et al., 2003).
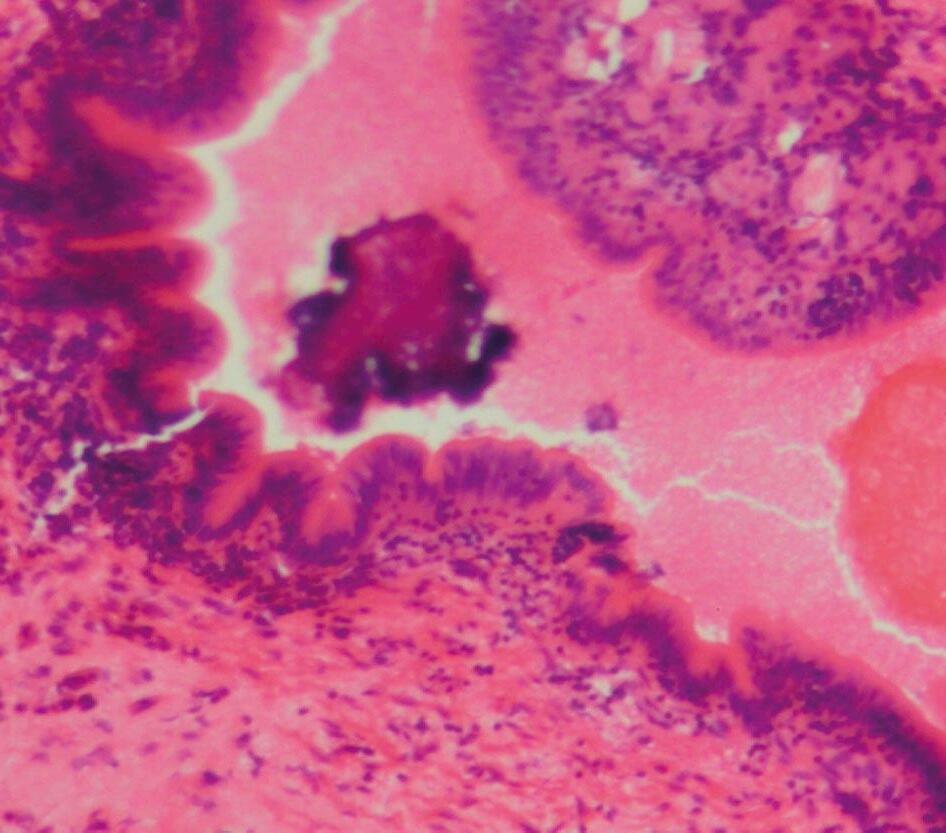
El microbioma intestinal puede influir en la biología, nutrición, inmunidad y sistema neuroendocrino del huésped (Dimitrov, 2011).
La función del TGI parece estar mediada por ácidos grasos de cadena corta (AGCC) generados por la fermentación bacteriana (Wu et al., 2017), la comunicación entre la microbiota y las neuronas (Megur et al., 2020), el sistema endocrino (Fukui et al., 2018), el sistema inmunológico (Maslowski & Mackay, 2011) y la modulación de la barrera epitelial intestinal (Sharma et al., 2010). El SNE y la red endocrina intestinal controlan la motilidad del TGI y su disrupción genera trastornos funcionales en el mismo (Fukui et al., 2018).
La serotonina tiene múltiples funciones biológicas relacionadas con la psicología y las actividades fisiológicas en procariotas y eucariotas, como la regurgitación, la vasodilatación, la motilidad del TGI, la cicatrización de heridas, la reproducción y la vasoconstricción, entre otras (Negri et al., 2021; Berger et al., 2009).
A su vez, la microbiota intestinal controla parcialmente la secreción de serotonina, dopamina, oxitocina y endorfinas producidas por las células enteroendocrinas (CEE) que se encuentran en el epitelio del TGI (Forsythe et al., 2010; Liang et al., 2014; Mayer et al., 2014).
La compleja red neuroendocrina que conecta el cerebro, el SNE, la microbiota intestinal y el GALT (eje microbiota-cerebro-intestino) tiene un impacto sustancial en la frágil barrera epitelial del intestino (Neuman et al., 2015; Megur et al., 2020), que también mantiene la tolerancia y la respuesta inmune contra antígenos extraños y el equilibrio de la microbiota (Maslowski & Mackay, 2011).
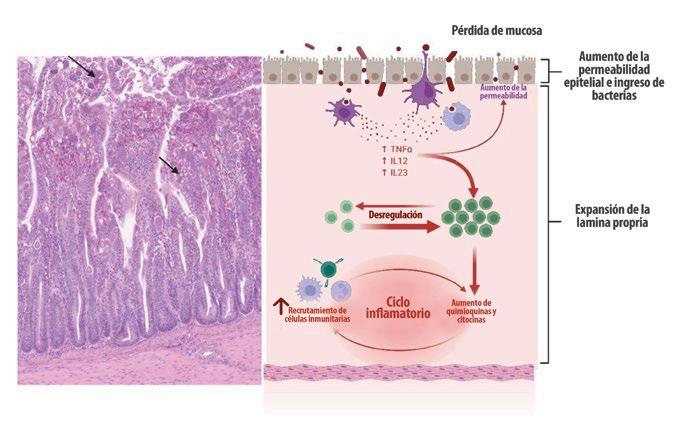
Por lo tanto, el estrés prolongado y la inflamación impactan negativamente en el eje microbiota-cerebro-intestino, causando disbiosis y alterando las proteínas de unión estrecha con la translocación sistémica de bacterias y otros antígenos (Figura 1).
Durante el estrés crónico y, como resultado, la inflamación intestinal crónica, la energía para crecimiento y reproducción se desvía para sostener la respuesta inflamatoria, y las aves no son una excepción a esta regla.
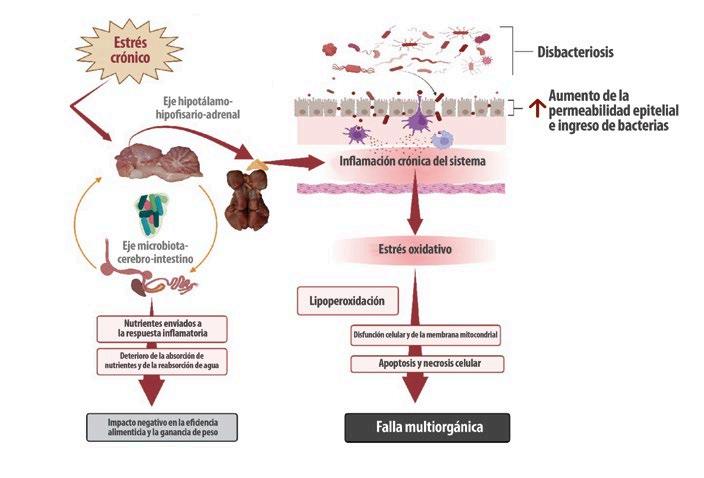
Figura 1. Impacto del estrés crónico: Patogénesis y consecuencias. El estrés crónico (independientemente de su origen) tiene un impacto directo en el eje hipotálamo-hipofisario-adrenal (eje HPA), el eje microbiota-cerebro-intestino (eje MCI) y los sistemas endocrino e inmunológico. Las inflamaciones intestinales y sistémicas crónicas se originan a partir de alteraciones en el delicado equilibrio y entorno de la microbiota intestinal (disbacteriosis) y alteraciones en las proteínas de unión estrecha entre los enterocitos, que causan permeabilidad intestinal. El estrés oxidativo prolongado inducido por el proceso inflamatorio causa peroxidación de fosfolípidos en las membranas celulares y mitocondriales, lo que conduce a apoptosis, necrosis celular y falla multiorgánica (creada con BioRender.com).
Por otra parte, las infecciones bacterianas del TGI, debido a su presencia y mecanismo de colonización, son inductoras de procesos inflamatorios agudos o incluso crónicos (Yamamoto et al., 2013).
En modelos murinos, el crecimiento de Salmonella es ayudado, irónicamente, por respuestas inflamatorias agudas a bacterias patógenas en el intestino, ya que se produce una mayor migración de neutrófilos y producción de especies reactivas de oxígeno (ROS, por sus siglas en inglés) y especies reactivas de nitrógeno (RNS, por sus siglas en inglés) como resultado de la infección por Salmonella, lo que interrumpe el equilibrio de la microbiota (Winter et al., 2010a).
Un aumento del oxígeno molecular en el lumen intestinal deprime importantes anaerobios comensales como Bacteroidetes y Clostridiales, que son bacterias esenciales productoras de ácido butírico (Rigottier-Gois, 2013). La oxidación de tiosulfato a tetrationato es también un subproducto de las ROS (Winter et al., 2010b).
En modelos murinos, se ha demostrado que Salmonella utiliza tetrationato para fortalecer su desarrollo (Winter et al., 2010b); de ahí, el uso de caldo de tetrationato como medio enriquecido para el cultivo de Salmonella en laboratorios de diagnóstico.
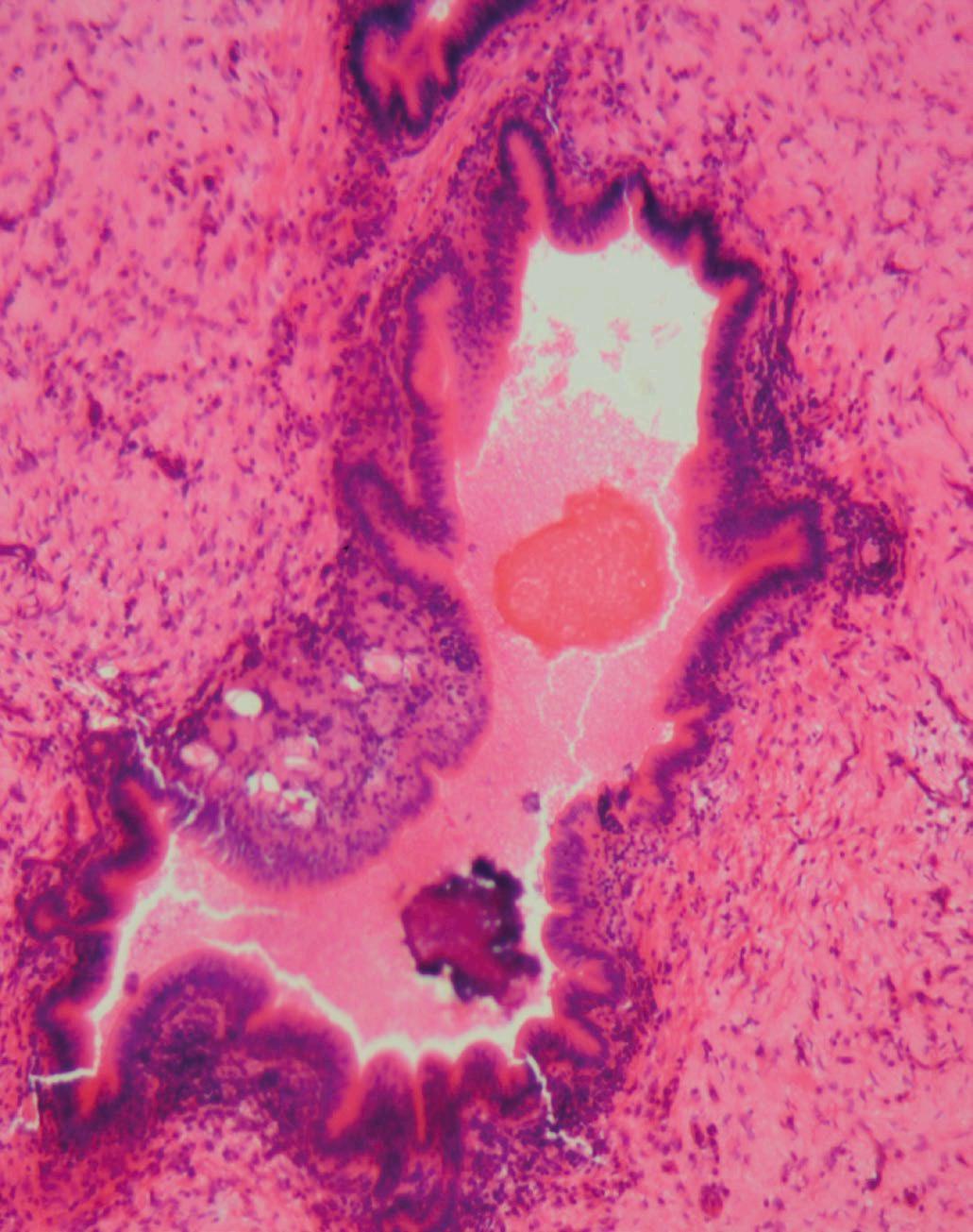
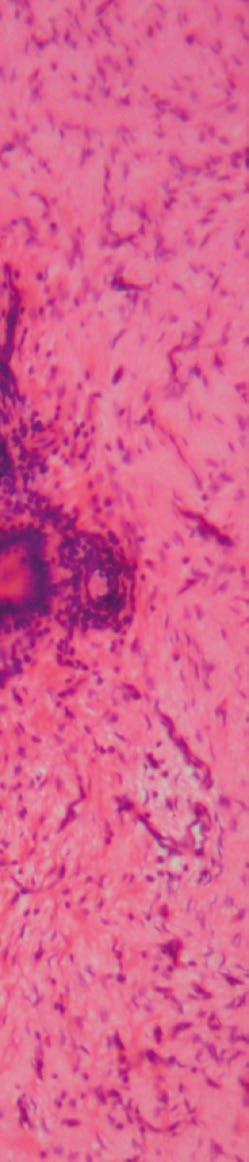
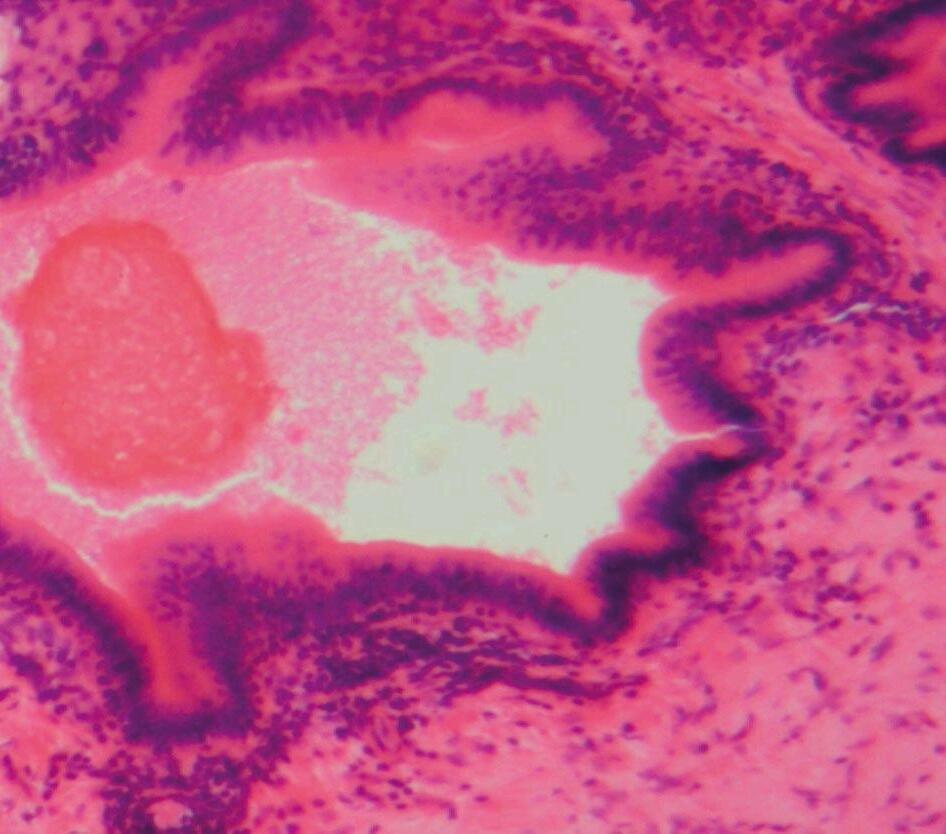
Generalmente, los agentes infecciosos (bacterianos, protozoarios, virales, helmintos) estimulan respuestas proinflamatorias del huésped. Por ejemplo, en aves domésticas, la coccidiosis puede causar necrosis e inflamación en el intestino, resultando en fiebre, depresión, reducción del rendimiento y muerte, dependiendo de la especie de Eimeria y la dosis infectante (Rose et al., 1975) (Figura 2).
Figura 2. Falla de la barrera intestinal. Los agentes infecciosos (bacterianos, protozoarios, virales, helmintos) en aves de corral estimulan las respuestas proinflamatorias del huésped. Falla de la barrera intestinal causada por Eimeria tenella. Mucosa y submucosa de los ciegos con infiltración de células inflamatorias, ulceración y necrosis. Las flechas muestran la presencia del parásito. Tinción con hematoxilina y eosina (creado con BioRender.com).
En el TGI, la inflamación crónica afecta la integridad de la barrera intestinal al alterar las proteínas de las uniones estrechas, lo que lleva a un aumento en la permeabilidad intestinal (“intestino permeable” o “leaky gut”) (Fasano, 2020), causando translocación bacteriana e inflamación sistémica (Ilan, 2012).
La respuesta inflamatoria puede ayudar al animal a recuperar la homeostasis tisular si se controla adecuadamente. Sin embargo, los tejidos y órganos se dañan si el estrés y la inflamación persisten (Dal Pont et al., 2020, 2021).
Los investigadores pueden usar modelos de inflamación entérica en un entorno de laboratorio para examinar alternativas a los antibióticos promotores de crecimiento (APC) y suplementos dietéticos para aves, por lo que se han desarrollado varios modelos de inflamación intestinal, incluidos dietas altas en NSP (Tellez et al., 2014, 2015; Dal Pont et al., 2021), dexametasona (Vicuña et al., 2015), sulfato de dextrano sódico (Menconi et al., 2015; Zou et al., 2018; Gilani et al., 2017a), restricción/ayuno alimentario (Kuttappan et al., 2015) y estrés térmico (Ruff et al., 2020; Rostagno, 2020).
La integridad intestinal depende de la función de la barrera. El estrés oxidativo, los factores antinutricionales de la soya, las fuentes de proteína o energía indigestibles, el estrés térmico y la histomoniasis son ejemplos de estresores que pueden resultar en la ruptura de la barrera intestinal (Williams, 2005; Sharma et al., 2007; Pastorelli et al., 2013; Tellez et al., 2014; Biswal et al., 2022).
Sin embargo, como resultado de la eliminación de los APC, han evolucionado nuevas enfermedades multifactoriales que causan enteritis y trastornos intestinales desconocidos en pollos de engorde, lo que afecta negativamente la salud y el rendimiento de las aves (Dahiya et al., 2006; Gholamiandehkordi et al., 2007; Yegani & Korver, 2008).
La disbiosis, definida como una microbiota anormal a nivel cuali y cuantitativo, desencadena una cascada de eventos en el TGI, incluyendo una disminución en la digestibilidad de nutrientes, inflamación e intestino permeable (Teirlynck et al., 2011; Weiss y Hennet, 2017).
A su vez, una mala salud intestinal se ha relacionado con condronecrosis bacteriana, osteomielitis y cojeras en pollos de engorde (Wideman & Prisby, 2012; Wideman et al., 2015). El correcto funcionamiento de la barrera intestinal es fundamental para la salud general y la homeostasis, y actúa como la primera línea de defensa contra los antígenos ambientales (Jeon et al., 2013).
digestibilidad


La primera capa de la barrera intestinal es la capa de moco extrínseca. Esta capa consta de dos partes: una capa externa de bacterias, que se adhiere ligeramente al epitelio, y una capa interna compuesta por IgA secretora y mucina, que se adhiere a la segunda capa de la barrera intestinal, las células epiteliales intestinales (IECs, por sus siglas en inglés).
Estas células epiteliales forman una única capa que separa el lumen intestinal de la lámina propia subyacente (Sakamoto et al., 2000; Kim & Ho, 2010). Estas células epiteliales deben tener la capacidad de recuperarse rápidamente en caso de daño tisular (Audy et al., 2012).
Los enterocitos en el epitelio apical son responsables de la absorción de nutrientes y las uniones estrechas cierran el espacio paracelular entre las células epiteliales adyacentes en la superficie apical, regulando la permeabilidad de la barrera intestinal al prevenir el ingreso de bacterias y antígenos de manera paracelular a través del epitelio (Ulluwishewa et al., 2011).
A pesar de su origen no hematopoyético, las IECs son un componente crucial de la inmunidad innata en el GALT, realizando una amplia gama de funciones inmunológicas. De hecho, las IECs son capaces de reconocer patógenos mediante la expresión de receptores inmunitarios innatos, liberando moléculas antimicrobianas y secretando una variedad de hormonas, neurotransmisores, enzimas, así como citoquinas y quimiocinas que median la interacción entre las respuestas inmunitarias innatas y adaptativas (Alverdy et al., 2005; Edelblum y Turner, 2009).
Por lo tanto, cualquier daño a las IECs, ya sea directo o indirecto, puede resultar en un colapso de la barrera intestinal y una posterior alteración de la homeostasis inmune de la mucosa, lo que resulta en una inflamación intestinal crónica y sistémica no controlada (Schulzke et al., 2009; Ilan, 2012).
Numerosos investigadores han revelado las vías detrás de la disrupción de las redes proteicas que conectan las células epiteliales mediante mediadores inflamatorios (hormonas, ROS, enzimas y diversas citoquinas proinflamatorias) inducidos por infecciones, elementos del alimento o cualquier otro estrés (Steed et al., 2010; Hu et al., 2013).
Otros factores también pueden contribuir a la ruptura de la barrera intestinal: la alimentación con grasa oxidada/no preservada en pollos y cerdos ha incrementado las tasas de recambio del epitelio intestinal y apoptosis en las puntas de las vellosidades (Dibner et al., 1996).
Los metales, al ser pro-oxidantes, pueden producir un estrés oxidativo y ruptura de la barrera (Teirlynck et al., 2009). Simultáneamente, el zinc regula varias proteínas esenciales para la formación de uniones estrechas entre las células del epitelio intestinal, y se ha encontrado que la deficiencia de zinc afecta la función de la barrera (Debon & Tester, 2001; Ulluwishewa et al., 2011).
El daño e inflamación de bajo grado en el tracto intestinal pueden causar una baja eficiencia alimenticia, lo cual es extremadamente costoso para la industria avícola (Porter, 1998). Factores endógenos y exógenos influyen en el equilibrio homeostático del TGI del huésped (Figura 3).
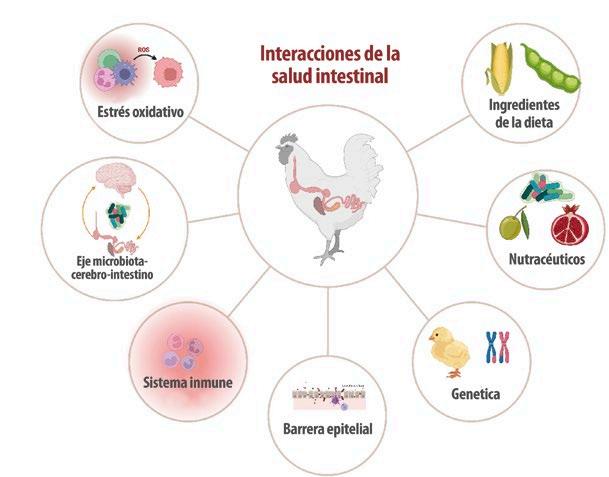
Estos factores representan diferentes tipos de estrés capaces de inducir inflamación y alterar la microflora, causando disbiosis (Iebba et al., 2016). Además, el estrés crónico y la inflamación causan malabsorción, al afectar gravemente la adquisición de nutrientes y la reabsorción de agua (Mishra & Jha, 2019).
Determinar el mejor microbioma para los pollos es un área de investigación continua para científicos avícolas, nutricionistas y gerentes de producción. Un microbioma saludable y diverso puede mejorar la salud general, el crecimiento y la productividad de los pollos.

Figura 3. Interacciones entre el huésped y factores intrínsecos o extrínsecos que influyen en la salud intestinal.
La homeostasis intestinal se define como la ausencia de inflamación y la ausencia de secreciones excesivas (agua o moco) dentro del tracto intestinal (Gabriel et al., 2006). La inflamación fisiológica está en curso durante la homeostasis intestinal como una respuesta controlada para mantener la tolerancia a los antígenos dietéticos y la microbiota comensal y para prevenir daños al epitelio (Kogut et al., 2018).
Sin embargo, cuando esta se induce, una cascada proinflamatoria promueve un flujo de células inmunitarias y proteínas plasmáticas al área afectada, lo que puede causar secreción excesiva de fluidos, especialmente durante la inflamación crónica (Noguchi et al., 1998).
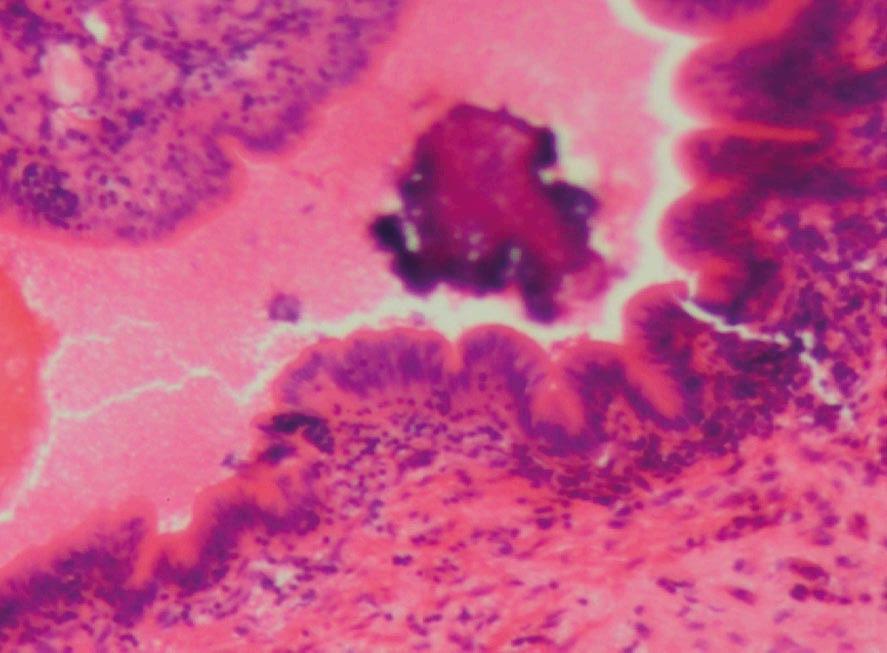
La respuesta inflamatoria es un proceso energéticamente costoso, dado que, los nutrientes se desvían hacia la respuesta inflamatoria del huésped, en lugar de destinarse al crecimiento o la acumulación de proteínas (Korver, 2012).
Se ha encontrado que la inflamación, la fiebre y el metabolismo alterado de los órganos resultan en catabolismo muscular y anorexia durante las respuestas de fase aguda en pollos desafiados con lipopolisacáridos (LPS), reduciendo la ganancia de peso en un 22%, pero solo el 59% de esta reducción se debió a una disminución en la ingesta calórica durante el desafío con LPS.
Por lo tanto, mecanismos adicionales, como la respuesta inmunológica, fueron responsables del 41% de la depresión en la ganancia de peso (Jiang et al., 2010).
Por otro lado, factores que ralentizan el tiempo de tránsito intestinal e incrementan la viscosidad de la digesta, como las dietas altas en polisacáridos no amiláceos (NSP, por sus siglas en inglés) sin la adición de enzimas exógenas, están asociados con disbiosis e inflamación intestinal (Tellez et al., 2014).
La presencia de alimento no digerido en las heces indica un mal funcionamiento del tracto digestivo (Yegani y Korver, 2008).
La inflamación entérica en aves está a menudo relacionada con el paso de alimento no digerido y el aumento de la permeabilidad intestinal. El estrés por calor, los enteropatógenos o las alteraciones en la nutrición pueden causar inflamación intestinal crónica. La inflamación entérica, la muerte de células epiteliales y el intestino permeable afectan la eficiencia alimenticia, la humedad de la cama y el consumo de agua.
La capacidad de inducir inflamación intestinal in vivo y las herramientas (biomarcadores) para evaluar tales cambios fisiológicos en aves son de suma importancia para considerar los nutracéuticos como aditivos alimentarios con capacidades prometedoras antioxidantes, antiinflamatorias y moduladoras del sistema inmunológico para ayudar a eliminar las moléculas redox.





Evaluación
Permeabilidad intestinal sérica
Permeabilidad intestinal
Actividad antioxidante intestinal sérica
Inflamación sérica y tisular
Inflamación intestinal celular e inmunológica (ARNm)
Biomarcadores séricos de enterocitos
Biomarcadores séricos inmunes
Marcadores inflamatorios y redox circulantes
Método indirecto para evaluar ácidos grasos de cadena corta
Inflamación intestinal sérica y fecal
Isotiocianato de fluoresceína-dextrano (FITC-d)
Azúcares manitol y ramnosa
Baxter et al. (2017); Gilani et al. (2017a); Vuong et al. (2021)
Gilani et al. (2017b) d(-)-Lactato
Fibronectina, fosfatasa alcalina intestinal y lipocalina-2
Ovotransferrina fecal
Translocación bacteriana hepática
Griess, superóxido dismutasa, sustancias reactivas al ácido tiobarbitúrico, capacidad antioxidante total
Isoprostano 8-iso-PGF2α
Prostaglandina F2α
Endotoxina y glicoproteína α1-ácida
Interleucina (IL)-1β, factor de necrosis tumoral-α e IL-10
IL-8, IL-1β, factor de crecimiento transformante (TGF)-β4 y proteína de unión a ácidos grasos
Quinasa regulada por señales extracelulares
Citrulina
IFN-γ
IgA secretora
IL-3, IL-6, IL-4, IL-18 y factor de necrosis tumoral-α, quimiocinas (CCL-20) y receptores de la familia NOD con dominio de pirina que contienen tres inflamasomas
Péptido YY
Calprotectina
Tabla 1. Biomarcadores para evaluar integridad intestinal en pollos
Zou et al. (2018)
Barekatain et al. (2020)
Goossens et al. (2018)
Latorre et al. (2014, 2015)
Baxter et al. (2019)
Petrone-Garcia et al. (2021)
Petrone-Garcia et al. (2021)
Chen et al. (2015)
Zou et al. (2018)
Chen et al. (2015)
Baxter et al. (2019)
Baxter et al. (2019)
Mullenix et al. (2021)
Tellez et al. (2020)
Dal Pont et al. (2021)
Numerosos enfoques, en rápida expansión, están siendo utilizados para crear biomarcadores. En la mayoría de los casos, se deben usar varios biomarcadores para evaluar la funcionalidad del TGI debido a la complejidad de las conexiones entre los componentes fundamentales del rendimiento gastrointestinal.

Dado que el origen de la inflamación intestinal y el estrés oxidativo es multifactorial, es importante mencionar que su prevención debe incluir diversos enfoques. El uso de probióticos, prebióticos y sustancias fitogénicas es una de las principales estrategias para mejorar la utilización de nutrientes (Shehata et al., 2022).
Esta estrategia se centra en reducir la colonización de patógenos asociados con la inducción de procesos inflamatorios en el TGI. La compleja interacción de estos microorganismos en relación con la microbiota intestinal juega un papel decisivo en su uso y el efecto correcto en la modulación de la microbiota natural del huésped.
Junto con el uso de componentes dietéticos como aceites esenciales (Zou et al., 2016) y minerales traza (Chen et al., 2022), resultan ser estrategias posiblemente exitosas para reducir estos efectos negativos de la inflamación crónica y el estrés oxidativo en el TGI.
El estudio y la comprensión de la compleja relación entre la microbiota, la dieta, el ambiente, los factores genéticos, así como los componentes de la dieta de las aves de producción, hacen de este enfoque multidisciplinario una de las ramas de la nutrición con mayor impacto en el futuro ante los desafíos de la producción de alimentos a nivel mundial.
Referencias disponibles en la versión web del artículo en nutrinews.com
Efectos del estrés crónico y la inflamación intestinal en la salud y el rendimiento de las aves de corral: Revisión DESCÁRGALO EN PDF

INTRODUCCIÓN
RESUMEN DEL ESTUDIO
Equipo técnico Arm & Hammer
CELMANAX™ es un suplemento alimenticio multicomponente totalmente natural que contiene carbohidratos funcionales refinados™ (RFC™) que tienen el estado de Generalmente reconocido como seguro (GRAS) como ingrediente alimenticio.
Se realizaron dos estudios independientes1,2 para evaluar el efecto de la suplementación con CELMANAX en las dietas de reproductoras de pollos y pollos de engorde sobre la prevalencia de Salmonella.
ESTUDIO 11
Se criaron un total de 1.040 reproductoras de pollos de engorde de un día de edad en 16 corrales (60 a 65 hembras por corral y 8 a 18 machos por corral; 8 corrales por tratamiento)
Las reproductoras de pollos de engorde fueron alimentadas con 0 o 50 g/TM de CELMANAX SCP
Se recolectaron y eclosionaron huevos de gallinas reproductoras de 51 semanas de edad, y los pollos de engorde machos de la progenie fueron alimentados con dietas suplementadas con 0 o 50 g/TM de CELMANAX SCP.
Todos los pollos de engorde (n=192) fueron examinados para detectar Salmonella spp. cecal a los 34 días de edad.
Se recolectaron ciegos de gallinas reproductoras (n = 28 – 30) de cada grupo de tratamiento a las 23 y 64 semanas de edad y se analizaron para determinar la prevalencia de Salmonella.
Se midió el rendimiento de las reproductoras.
Los pollitos de engorde Ross de un día de edad clasificados por sexo se asignaron a 24 corrales replicados con 12 pollos de engorde cada uno por dieta por sexo y se alimentaron con 0 o 50 g/TM de CELMANAX SCP.
Los pollos de engorde fueron alimentados con una dieta estándar de inicio, crecimiento y finalización.
Se analizaron los corrales de pollos de engorde para detectar la prevalencia de Salmonella en la cama. También se probó la prevalencia de Salmonella en ciegos de pollos de engorde.
Se midió el rendimiento de los pollos de engorde.
1 Walker et al. Poult Sci 2017;96(8):2684-2690.
2 Walker et al. Poultry Science 2018;97:1412–1419.
En el estudio 1, las gallinas reproductoras de pollos de engorde de control tuvieron una prevalencia de Salmonella en los ciegos de 71,43% y 40,00%, mientras que las gallinas alimentadas con CELMANAX tuvieron una prevalencia cero de Salmonella (P<0,01) (Figura 1)
Reproductoras a las 23 semanas
Reproductoras a las 64 semanas P<0,01
Figura 1. Efectos del tratamiento sobre la prevalencia porcentual de Salmonella en ciegos de gallinas reproductoras en el Estudio 1
En el estudio 1, la progenie de pollos de engorde de gallinas alimentadas con la dieta de control y que recibieron dietas de control para pollos de engorde tuvo una prevalencia de Salmonella en los ciegos del 12,5%, mientras que los pollos de engorde de gallinas alimentadas con CELMANAX y que recibieron 0 o 50 g/TM de CELMANAX en las dietas para pollos de engorde tuvieron una prevalencia cero de Salmonella en el muestreo de 34 días (Figura 2)
Reproductoras alimentadas con control
Reproductoras alimentadas con CELMANAX
Dieta control de pollos de engorde
Dieta CELMANAX 50 g/MT en pollos de engorde
Figura 2. Efectos del tratamiento de reproductores y pollos de engorde en el % de prevalencia de Salmonella en ciegos de pollos de engorde en el Estudio 1.
En el estudio 2, se aisló Salmonella spp. de la cama de 7 de 48 (14,58 %) corrales de pollos de engorde alimentados con control, pero ninguno (0 %) se aisló de los corrales alimentados con CELMANAX (P≤0,05) (Figura3)
CONCLUSIÓN
FIGURA 3: Efectos del tratamiento de pollos de engorde sobre el porcentaje de incidencia de Salmonella en la cama en el Estudio 2
El muestreo cecal de pollos de engorde para Salmonella a los 44 y 55 días de edad confirmó una prevalencia del 45,83% y 29,17% respectivamente en los pollos de engorde alimentados con control, pero ninguna en los corrales alimentados con CELMANAX (Figura 4).
Las hembras de engorde alimentadas con CELMANAX tuvieron una tendencia a mejorar el peso corporal y la relación de conversión alimenticia (datos no presentados).
FIGURA 4: Efectos del tratamiento sobre la prevalencia porcentual de Salmonella en ciegos de pollos de engorde en el Estudio 2
La suplementación con CELMANAX en las dietas de reproductoras y pollos de engorde redujo significativamente la prevalencia de Salmonella en la cama y los ciegos, lo que respalda su beneficio en una estrategia multifactorial de mitigación de Salmonella en la producción avícola. DESCÁRGALO

La suplementación con CELMANAX en dietas de reproductoras y pollos de engorde redujo la prevalencia de Salmonella

Lorran Baeumle Gabardo - Global Product Manager, dsm-firmenich Nutrición y Salud Animal
Elle Chadwick - Global Poultry Marketing Specialist, dsm-firmenich Nutrición y Salud Animal
Shelby Ramirez - Global Poultry Technical Manager, dsm-firmenich Nutrición y Salud Animal

Los efectos negativos de las micotoxinas en las aves pueden ser de gran impacto, ya que disminuyen la integridad gastrointestinal, la inmunidad y el rendimiento de los pollos de engorde y provocan pérdidas económicas.
Dadas los elevados niveles de micotoxinas en los alimentos para aves, se necesita una estrategia de gestión del riesgo de micotoxinas para reducir los retos en la producción avícola y garantizar la rentabilidad.


Los productores avícolas se preguntan con frecuencia: ¿Cuál es el impacto real de las micotoxinas en la productividad de los pollos de engorde? Lo que parece una pregunta fácil es, en el caso de las micotoxinas, no tan fácil de responder.

Tres consideraciones clave para evaluar el impacto de las micotoxinas en una granja avícola:
Incluir las micotoxinas como posibles factores predisponentes a problemas de salud
Medir los niveles de contaminación en los ingredientes y en los alimentos terminados
La exposición a varias micotoxinas al mismo tiempo puede tener efectos más graves en la salud y el rendimiento de las aves. Por lo tanto, es de suma importancia medir un conjunto de micotoxinas para disponer de una evaluación de riesgos más sólida (Figura 2).
Evaluar el efecto de la contaminación con bajos niveles de micotoxinas sobre el rendimiento y la rentabilidad
Las micotoxinas se encuentran con frecuencia en los alimentos para aves. Según la Encuesta mundial sobre micotoxinas de dsm-firmenich (dsmfirmenich World Mycotoxin Survey), más de 33 000 análisis de alimentos para aves dieron positivo en micotoxinas en los últimos diez años (2013 - 2023).
Un análisis más detallado de estos datos muestra que el 83% de estas muestras están contaminadas con más de una micotoxina (Figura 1) y se encontraron hasta 50 micotoxinas en una sola muestra.
Figura 2. Presencia de micotoxinas en 33.000 alimentos para aves, mostrando el % de muestras positivas para cada micotoxina (Fuente: dsm-firmenich World Mycotoxin Survey)
Analizar el alimento y/o sus ingredientes ayuda a evaluar el riesgo para la salud del ave. La presencia frecuente de micotoxinas afecta a la salud intestinal y al sistema inmunitario de los animales, reduciendo así el rendimiento de los lotes, lo que puede traducirse en una pérdida de beneficios para los productores avícolas.
La falta de signos clínicos visibles se debe principalmente al corto ciclo de vida de los pollos de engorde. Esto significa que el desafío puede estar presente, pero no tener tiempo de manifestarse exteriormente. Los efectos combinados de más de una micotoxina dificultan el diagnóstico.
Sin embargo, los principales efectos de las micotoxinas en la salud de las aves están claramente demostrados: destrucción de las vellosidades intestinales, lo que perjudica la absorción de nutrientes y la digestión, al tiempo que aumenta la permeabilidad intestinal, y modulación de la respuesta inmunitaria local y de la microbiota, lo que aumenta las posibilidades de fallo vacunal y disbiosis.
Disminución de la absorción de nutrientes: Las micotoxinas, especialmente el deoxinivalenol (DON) y las fumonisinas (FUM), afectan a varios aspectos de la integridad intestinal. Un metaanálisis (Grenier y Applegate, 2013) mostró una clara influencia de DON y FUM (figura 3).
Las micotoxinas reducen la funcionalidad de las uniones estrechas entre las células intestinales, abriendo la oportunidad para que los patógenos y los factores antinutricionales pasen de la barrera intestinal al torrente sanguíneo. Esto desafía al sistema inmunitario y al hígado, desviando nutrientes y energía del crecimiento.
El hígado se ve directamente afectado por la mayoría de las micotoxinas. En los pollos de engorde, se ha observado una degeneración grasa visible y un aumento de aproximadamente el 15% del peso del hígado.
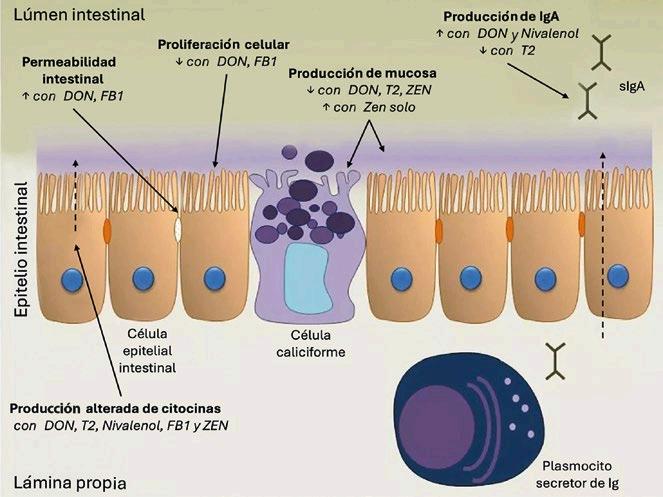
Figura 3. Efecto de DON, NIV, FB1, toxina T-2 y ZEN en el epitelio intestinal. Alteran los diferentes mecanismos de defensa intestinal, incluyendo la integridad epitelial, la proliferación celular, la mucosa, las inmunoglobulinas (Ig) y la producción de citoquinas. (Fuente: Antonissen et al., 2014)

Esto refleja un mayor costo de nutrientes y aminoácidos, especialmente de metionina, que es el primer aminoácido limitante para los pollos de engorde.
Una estrategia que desactive directamente las micotoxinas es una gran oportunidad para prevenir las pérdidas nutricionales sin tener que gestionar cambios nutricionales en la dieta.
Fallas vacunales: La modulación de la respuesta inmunitaria es uno de los principales modos de acción de las micotoxinas, que puede influir en la respuesta de las aves a las enfermedades y en las prácticas de vacunación.
Las micotoxinas actúan como un factor predisponente que reduce la inmunidad frente a enfermedades virales en pollos de engorde (Kamalavenkatesh et al., 2005; Hanif y Muhammad, 2015 y Yunus et al., 2012).
La ocratoxina, DON, T2 y el ácido ciclopiazónico redujeron significativamente el título de anticuerpos para el virus de la enfermedad de Newcastle (NDV), virus de la bronquitis infecciosa (IBV), virus de la bursitis infecciosa (IBDV), y el síndrome de hidropericardio (HPS). Aumenta la susceptibilidad de las aves a enfermedades infecciosas que podrían evitarse en condiciones normales.
Predisposición a la coccidiosis y la enteritis necrótica (EN): Antonissen et al. (2014) evidenciaron que dosis bajas a moderadas de diferentes micotoxinas de Fusarium (DON, NIV, FB1, T-2, ZEN) predisponen a las aves a una respuesta más severa frente a un desafío de coccidiosis y EN, como se muestra con la reducción de la respuesta inmune y la eficacia del tratamiento anticoccidial.
Además, la contaminación por FB1 y DON dio lugar a un desafío de EN más grave en dos estudios distintos (Antonissen et al., 2012 y Antonissen et al., 2015). La coccidiosis es una de las enfermedades más costosas en la producción avícola. Mediante el control y la reducción de las micotoxinas, las estrategias de gestión de la coccidiosis pueden ser más eficaces.

Contaminación alimentaria: También se informó de una mayor susceptibilidad a Salmonella typhimurium y Campylobacter jejuni en presencia de tricotecenos (DON y T2), lo que sugiere que estos compuestos pueden modular el metabolismo bacteriano.
Como se ha descrito anteriormente, las micotoxinas provocan un “intestino permeable”. Una vez interrumpidas las uniones entre las células intestinales, se produce un intercambio de moléculas entre el intestino y el torrente sanguíneo.
Esta situación puede influir en la contaminación de la canal durante el procesamiento:
El exceso de proteínas en el lumen intestinal favorece una mayor contaminación de bacterias patógenas como Salmonella sp y Campylobacter jejuni;
La mayor cantidad de agua en el lumen intestinal conduce a excrementos más fluidos.
Al mitigar el riesgo del efecto de las micotoxinas, se reduce al mínimo el efecto del “intestino permeable”, lo que favorece aún más a la obtención de productos avícolas seguros.
El rendimiento es uno de los factores más importantes de la rentabilidad de la producción avícola.
Una recopilación de datos de ensayos científicos ha demostrado que la presencia de micotoxinas en las dietas de las aves redujo significativamente (P<0,05) la ingesta de alimentos en un 12% y la ganancia de peso corporal (PC) en un 14%, lo que resulta en una deficiente conversión alimenticia (CA) del 7% en comparación con los grupos no contaminados (Andretta et al, 2011).
Además, los resultados de un proyecto de cooperación con la Universidad de Gante (Antonissen et al., 2018) muestran un efecto negativo sinérgico frente a una dieta desafiada por disbiosis y la presencia de DON (5 mg/kg) y FB1 (20 mg/kg) sobre el rendimiento de los pollos de engorde. Estos datos demuestran que la reducción del rendimiento se mantiene durante la fase final de la producción (Figura 4).
Peso corporal
Consumo de alimentos

+ DON
+ FBs
+ DON
Figura 4. Parámetros de rendimiento en el día 39 de pollos de engorde alimentados con una dieta de control negativo, un control con disbiosis, una dieta con disbiosis contaminada con DON o una dieta con disbiosis contaminada con FB1. Las barras representan las medias de las 7 réplicas (corrales) por tratamiento ± DE. Dentro del mismo período, las barras con letras diferentes (a-b) difieren significativamente (P ≤ 0,05). (Fuente: Antonissen et al., 2018)
En una evaluación a largo plazo (18 ensayos sucesivos) en una granja comercial, se demostró que la contaminación natural con niveles inferiores a las recomendaciones de la UE para micotoxinas tiene un impacto significativo de 2,5 puntos en la CA (Figura 5), lo que confirma los efectos sinérgicos de las micotoxinas (fumonisinas, zearalenona, DON y DAS) y la disminución del rendimiento de los pollos de engorde en condiciones comerciales (Kolawole et al., 2020).
Las micotoxinas tienen efectos directos e indirectos sobre la coccidiosis, la seguridad alimentaria y el rendimiento de los pollos de engorde, que influyen directamente en la rentabilidad del productor.
Concentración de micotoxina (µg/kg)
Conversión alimentaria (CA) Diferencia de 2,5 puntos en CA
Micotoxina
Figura 5. Diferencia entre la CA (línea negra) en contaminaciones altas y bajas de micotoxinas (líneas de color) en pollos de engorde (Adaptado de Kolawole et al., 2020).
Debido a la frecuente presencia de micotoxinas en los alimentos para aves, se necesita una estrategia de gestión del riesgo de micotoxinas para proteger a los pollos de engorde en todas las fases y garantizar la rentabilidad.
Por qué son importantes las micotoxinas en la producción de pollos de engorde
DESCARGAR EL PDF dsm-firmenich ofrece la solución más completa del mercado: Nuestro estudio, que es el más antiguo y el conjunto de datos más completo sobre la presencia de micotoxinas para saber qué micotoxinas pueden estar presentes en sus alimentos, nuestros servicios analíticos para apoyar la detección de micotoxinas en sus ingredientes, y nuestro portafolio de aditivos nutricionales Mycofix®, que representa la solución más avanzada para proteger la salud de las aves desactivando las micotoxinas que contaminan los alimentos.




La liberación de fósforo a partir del fitato se produce por escisión secuencial de los grupos fosfatos del anillo de inositol. Se creía que, independientemente de las propiedades de las fitasas, la velocidad de desfosforilación del fitato está limitada por la primera escisión de cualquier grupo fosfato.



La posición del primer grupo fosfato escindido depende de la especificidad de la fitasa. La inhibición del inicio de la desfosforilación no está asociada al mecanismo de acción de la enzima y puede deberse más bien a una actividad insuficiente de la fitasa o a una baja disponibilidad del sustrato.
Los animales no rumiantes no digieren eficientemente al fitato contenido en los piensos debido a la escasa presencia de enzimas endógenas en el tracto gastrointestinal (TGI).

El análisis de las transformaciones en la cadena de reacción inositol hexafosfato (IP6) inositol (I) muestra que la desfosforilación de IP6 en su conjunto limita la eliminación del grupo fosfato de I(1,2,5,6)P4 (tercera reacción desde el inicio de la hidrólisis de los enlaces fosfato el ácido fítico).
La menor disponibilidad de nutrientes en presencia de fitato no se debe a la acción de los fitasa, sino que está causada por los aniones del fósforo fítico (IP6-3), que se unen a iones metálicos cargados positivamente, aminoácidos y proteínas.
El fitato es una forma de reserva de fósforo en las semillas de las plantas. Como resultado de la activación de la fitasa durante la germinación de las semillas, el fitato libera fosfato inorgánico (Pi) para su uso por las plantas en crecimiento.
El término fitato incluye muchas sustancias definidas por la presencia en su composición del anión del ácido fítico (AF).
Las enzimas que escinden el ácido fítico se denominan mioinositol hexakisfosfato 3- y 6-fosfohidrolasas.
El pH del medio es un factor determinante que afecta a la solubilidad del producto de reacción. Así, un aumento del pH del medio hasta 4-5 disminuye la solubilidad de las sales resultantes, a excepción del fitato de magnesio que es soluble a pH 7,5.

En soluciones con pH < 1,1, el ácido fítico tiene una carga neutra y permanece débilmente activo. Un aumento del pH del medio a 2 conduce a la ionización de tres grupos fosfatos; un aumento adicional del pH resulta en la pérdida de los protones restantes.

El anión de fósforo fítico es capaz de retener su carga negativa en el tracto gastrointestinal y reaccionar con cationes.
En el estómago, la lisina, la histidina y la arginina, así como las proteínas y los polipéptidos que contienen estos aminoácidos, pueden formar complejos binarios proteína-IP6 que inhiben la digestión de las proteínas.
La unión de ácido fítico a los aminoácidos cargados positivamente limita su absorción.
Las más comunes y mejor estudiadas son las fitasas ácidas de histidina, que difieren en el átomo de carbono del anillo de mio inositol del que se elimina el primer grupo fosfato.
La subsiguiente eliminación de los grupos fosfato da lugar a la liberación de fósforo disponible para plantas y animales y a la formación de ésteres de inositol en menor número de fosfato solubles.
La solubilidad de los isómeros depende de la proporción de cationes metálicos y de la presencia de aminoácidos libres con propiedades quelantes. Se cree que las tres primeras etapas de la desfosforilación del ácido fítico (IP6 IP4) juegan un papel importante en la creación de condiciones para la hidrólisis completa de IP.
La carga negativa de la molécula de ácido fítico y su capacidad para unirse a otros nutrientes disminuye con la reducción del número de grupos fosfatos.
Las fosfatasas alcalinas y las fitasas alcalinas también participan en la escisión de isómeros.
Recientemente, se ha establecido que cuando se incluye fitasa ácida de histidina en la dieta (500 unidades/kg-1000 unidades/ kg), esta aumenta en el yeyuno la actividad de la fitasa alcalina endógena, una enzima que rompe el fitato de metales divalentes difíciles de disolver.

La acción de cada tipo de fitasa y su actividad máxima dependen del pH del medio.

Sobre esta base, las fitasas se clasifican en ácidas o alcalinas. Las fosfatasas ácidas muestran una amplia especificidad de sustrato hacia los fitatos libres de metales, mientras que las fosfatasas alcalinas son específicas hacia el ácido fítico unido a metales.

Figura 1. En esta figura se observa el tracto gastrointestinal de un cerdo, los valores de acidez en el estómago y el duodeno, y el tiempo de retención del bolo alimenticio en el estómago. En la porción inferior se observa la solubilidad del fitato (in vitro) a diferentes valores de pH.

La solubilidad del fitato es de 97,9% cuando está sumergido en una suspensión de 3,5 de acidez. Esta acidez se alcanza cuando el fitato está en la porción estomacal del tracto gastrointestinal.
Las fitasas ácidas son sumamente efectivas a nivel estomacal, por lo tanto debemos promover el tiempo de retención adecuado del bolo alimenticio. Si la acidez aumenta podemos observar, la precipitación del fitato (suspensión de color blanquecina).
En esta prueba podemos observar, que el estado insoluble del fitato comienza a aparecer a pH 4. En este estado protonado, el fitato comienza a ligar minerales (Zn, Cu ,Co Mn), volviéndose un complejo fitato mineral insoluble para la absorción.
Las proteínas también se pueden ver afectadas con aquel fitato que no fue solubilizado correctamente, provocando un sustrato fitato-mineral-proteína insoluble para la absorción del cerdo.
El aumento del contenido de fitasa en el pienso a 1500 FTU/ kg garantizó la máxima escisión de IP6, mientras que el aumento posterior del contenido de enzima a 3000 FTU/kg no tuvo ningún efecto sobre la hidrólisis.
Los estudios in vitro realizados con el sustrato libre (fitato de sodio), cuya disponibilidad no estaba limitada por la estructura celular de los componentes del alimento, confirmaron que la desfosforilación del IP4 es la etapa limitante en la cadena de desfosforilación secuencial del IP6 original.
Se supone que la acumulación de I(1,2,5,6)P4 indica que su conversión es una reacción de “cuello de botella” en la cadena de reacciones secuenciales de desfosforilación IP6→IP1.
La disminución de la concentración de I(1,2,3,4,6)P5, que retiene el grupo fosfato en el sexto átomo de carbono en presencia de 6-fitasa, indica que múltiples factores pueden activar las fosfatasas alcalina y/o inhibir la 6-fitasa.
Los estudios sobre el contenido de grupos de metabolitos de IP6 en las heces de cerdos confirmaron la suposición de que el catabolismo de IP6, que se produce como una cadena de reacciones sucesivas de desfosforilación, se ralentiza a nivel de I(1,2,4,6)P4.

Basándonos en diferentes investigaciones, podemos concluir que, en los experimentos in vitro donde la disponibilidad del sustrato para la enzima no era restringida, el IP6 sufrió una hidrólisis completa y su conversión a I(1,2,4,5,6)P5 (es decir, la escisión del primer grupo fosfato) no fue la etapa limitante en la cadena de reacciones de desfosforilación secuencial IP6 IP2. En cambio, el proceso se inhibió en la etapa de desfosforilación de I(1,2,5,6)P4.
Las paredes celulares de las células de los cereales están formadas por celulosa y hemicelulosas, principalmente arabinoxilanos, con pequeñas cantidades de β-glucanos y mananos.

Los monogástricos carecen de enzimas capaces de hidrolizar estos polisacáridos de la pared celular, por lo que una parte del contenido intracelular de las células de los cereales permanece físicamente inaccesible a las fitasas.

La adición de carbohidrasas al pienso facilita la destrucción de la pared celular y la formación de canales a través de los cuales el agua y las enzimas pueden entrar en las células, lo que garantiza la digestión de proteínas, almidón y fitato.
Cuando se añadió celulasa al pienso, la disponibilidad de fósforo y proteína aumentó un 30,4 y un 21,8%, respectivamente.
Debido a la actividad de las proteasas exógenas, la liberación de fósforo aumentó un 15% y la de proteína un 14,5%.
El tamaño de las moléculas de fitasa es suficiente como para que puedan penetrar en las paredes vegetales, a través de los poros naturales, y catalizar la desfosforilación del fitato en el interior de las células.
Las fitasas bacterianas suelen tener moléculas más pequeñas que las enzimas fúngicas. Esto sugiere que la mayor eficacia de las fitasas bacterianas puede deberse al tamaño de sus moléculas, que facilita su acceso a los sustratos.
Un factor importante en la actividad de las fitasas es la disolución del fitato, que les permite diseminarse más allá de las células vegetales y quedar disponibles para su acción.
En el caso de los cerdos, el alimento permanece más tiempo en el ambiente ácido del estómago que en las aves de corral, de modo que la cantidad de fitato soluble debería ser mayor. Esto explica que los cerdos utilicen mejor el fósforo que las aves de corral.
La escisión incompleta del fitato de los alimentos naturales in vivo puede explicarse por la disponibilidad limitada de las enzimas para el sustrato, siendo la etapa limitante la deforilación de IP4.



La misma regularidad se observó en otras condiciones experimentales en las que se utilizaron fitasas de Escherichia coli, Aspergillus niger y Buttiauxella sp.
Numerosos artículos de investigación han demostrado que las fitasas presentan un efecto extra-fosfórico, aumentando la digestibilidad de las proteínas y otros nutrientes.
Estas conclusiones se basaban en la interpretación errónea de los resultados de estudios sobre la utilización de nutrientes, en los que se identificaba el proceso fisiológico de la digestión con los procesos de absorción de aminoácidos y metabolización del nitrógeno.
El efecto extra-fosfórico de las fitasas se entiende como un aumento de la disponibilidad de nutrientes distintos del fósforo durante la digestión, es decir, como una manifestación del efecto no asociada a la función bioquímica de las fitasas.
Las fitasas no pueden afectar directamente a la digestibilidad de las proteínas: aumentan la disponibilidad de los aminoácidos ya digeridos al impedir su unión a los fosfatos, como resultado de la reducción de la concentración de fosfatos debido a su escisión.
Por lo tanto, el concepto de efecto extra-fosfórico de las fitasas no es del todo exacto, ya que la fitasa solo tiene un efecto.
El fitato tiene un fuerte impacto en la nutrición de un lechón, provocando una pérdida importante sobre la ganancia de peso si este no es inactivado por fitasas eficientemente.
El lechón presenta una capacidad reducida para producir acidez a nivel estomacal, puesto que la dieta de este animal está basada en carbohidratos (lactosa). Por lo tanto puede haber mucho sustrato insoluble en el tracto gastrointestinal. Otras consecuencias, aunque asociadas a la adición de fitasas exógenas al pienso, son consecuencia de su efecto sobre el fitato. Son indirectos y surgen como resultado de la disminución de la concentración de aniones reactivos IP6-4.

Una estrategia para ayudar al lechón a solubilizar eficientemente el fitato, es con la aplicación de súper dosis.
La aplicación de súper dosis de fitasa, asegura una adecuada solubilización de este sustrato, evitando así efectos adversos en el crecimiento de un lechón.

La digestión de los fitatos depende de dos factores principales: la presencia de cantidades suficientes de fitasa y la disponibilidad de fitato para la acción enzimática.
1
El primer problema se puede resolver añadiendo cantidades suficientes de fitasas exógenas en proporciones adecuadas al pienso.
2
El segundo problema se supera aumentando la digestibilidad del pienso en su conjunto.
3
En casos especiales, pueden añadirse carbohidrasas al pienso, ya que estas enzimas alteran las membranas celulares y facilitan el acceso de las hidrolasas a los nutrientes del citoplasma
La escisión del primer grupo fosfato por parte de 3 o 6 fitasas da lugar a la formación de IP, que incluyen I(1,2,4,5,6)P5 e I(1,2,3,4,5)P5. En comparación con el IP6, estos metabolitos se unen a los nutrientes de manera menos eficiente, pero el I(1,2,4,5,6)P5 forma complejos más fuertes con las proteínas que el I(1,2,3,4,5)P5.
Además, el I(1,2,4,5,6)P5 es un precursor del I(1,2,5,6)P4, que inhibe las etapas posteriores de la escisión del IP. Esto corrobora la preferencia por las 6-fitasas. El efecto extra-fosfórico de las fitasas, es decir, el efecto que va más allá de la acción fosfohidrolasa, se manifiesta como un aumento de la disponibilidad de microelementos, macroelementos y aminoácidos.
Las fitasas no tienen actividad proteolítica ni amilolítica, por lo que la suposición común en la literatura popular de que las fitasas aumentan la digestibilidad de las proteínas y el almidón es incorrecta.

Los secretos detrás de una fitasa DESCÁRGALO EN PDF
Gerardo
Ordaz Ochoa, María Alejandra Pérez Alvarado, Luis Humberto López Hernández
Centro Nacional de Investigación Disciplinaria en Fisiología y Mejoramiento Animal, INIFAP, México
Las grasas y los aceites son componentes esenciales en la dieta de los cerdos, aportan energía, ácidos grasos esenciales, contribuyen a características sensoriales del alimento e inciden en la calidad de la carne (Chen et al., 2018).
Sin embargo, cuando se exponen al oxígeno, la luz y el calor, las grasas pueden sufrir oxidación, lo que provoca cambios en su composición química, sabor y valor nutricional (Nourooz-Zadeh, 1999). Lo cual repercute de manera directa sobre la composición de la dieta, salud de los animales, rendimiento productivo y calidad del producto.

No obstante, al ser la alimentación de los cerdos un aspecto crítico de la producción porcina desde las perspectivas de producción y economía. En los últimos años, ha habido un interés creciente en la utilización de grasas oxidadas como ingredientes en dietas para cerdos (Chen et al., 2018).
La oxidación de las grasas se produce mediante una compleja serie de reacciones químicas que implican la reacción de ácidos grasos insaturados con moléculas de oxígeno.
El proceso se acelera por factores como el calor, la luz y la presencia de catalizadores como metales y enzimas.
El oxígeno reacciona con los dobles enlaces de los ácidos grasos insaturados, lo que lleva a la formación de hidroperóxidos lipídicos, que se descomponen aún más en aldehídos, cetonas y otros compuestos volátiles (Frankel, 1980).


La oxidación de las grasas da como resultado la formación de diversos compuestos químicos:
Hidroperóxidos
Aldehídos
Cetonas
Radicales libres
Estos compuestos pueden alterar el sabor, el aroma y las propiedades nutricionales de las grasas, provocando sabores desagradables, rancidez y una vida útil reducida.
Además, la oxidación puede disminuir la calidad nutricional de las grasas al reducir los niveles de ácidos grasos esenciales y vitaminas liposolubles (Nourooz-Zadeh, 1999).

De acuerdo con lo reportado sobre el uso de grasas oxidadas en la alimentación porcina existen controversias sobre su aplicación. En la literatura se pueden encontrar resultados tanto que promueven como que limitan su utilización:

Entre los beneficios que se han reportado sobre el uso de grasas oxidadas en la alimentación del cerdo se encuentran:
Las grasas oxidadas pueden servir como fuente de energía alternativa en las dietas para cerdos.
A pesar de la formación de subproductos oxidativos, estas grasas retienen una parte importante de su valor calórico, proporcionando a los cerdos energía para el crecimiento y el mantenimiento (Gourle et al., 2020).
La utilización de grasas oxidadas puede ser rentable para los productores de cerdos, especialmente cuando las fuentes de grasa fresca son costosas o su oferta es limitada.
No obstante, al incorporar grasas oxidadas en las dietas de los cerdos se podría estar comprometiendo la calidad nutricional del alimento de acuerdo con las modificaciones ya mencionadas que sufren las grasas al oxidarse.

Las grasas oxidadas, que pueden no ser aptas para el consumo humano debido a sus malos sabores o su rancidez, pueden reutilizarse como ingredientes para la alimentación de los cerdos. Esto reduce el desperdicio en la industria alimentaria y contribuye a la utilización sostenible de los recursos (Gourle et al., 2020).
Los riesgos reportados sobre el uso de grasas oxidadas en la alimentación porcina recaen en:
Las grasas oxidadas pueden contener niveles reducidos de vitaminas liposolubles y antioxidantes debido al proceso oxidativo.
Los cerdos alimentados con dietas ricas en grasas oxidadas pueden correr riesgo de sufrir deficiencias de nutrientes, en particular de vitamina E, que desempeña un papel crucial en la defensa antioxidante y la función inmune (Nguyen et al., 2019).

Palatabilidad y digestibilidad
Las grasas oxidadas pueden impartir sabores y olores desagradables al alimento, afectando potencialmente la palatabilidad y el consumo de alimento en los cerdos.
La presencia de subproductos oxidativos puede dificultar la digestión y absorción de grasas, lo que lleva a una reducción de la utilización de nutrientes y del rendimiento del crecimiento (Chen et al., 2018).
El consumo de grasas oxidadas se ha asociado con efectos adversos para la salud en los animales, incluido el estrés oxidativo, la inflamación y la función inmune comprometida.
Los cerdos alimentados con dietas que contienen altos niveles de grasas oxidadas pueden ser más susceptibles al daño oxidativo y a los trastornos metabólicos (Jha & Berrocoso, 2016).
Las grasas oxidadas exhiben características distintas que resultan de la oxidación de ácidos grasos insaturados.
Los cambios químicos durante la oxidación conducen a la formación de compuestos volátiles que afectan el sabor, el aroma y las propiedades nutricionales de las grasas.
Comprender las características de las grasas oxidadas es esencial para garantizar la calidad y seguridad de los alimentos y para desarrollar estrategias para minimizar los problemas relacionados con la oxidación.
En lo que concierne en la alimentación porcina, el uso de grasas oxidadas en la industria porcina presenta tanto oportunidades como desafíos para los productores de cerdos:
Las grasas oxidadas ofrecen una fuente de energía rentable y contribuyen a la reducción de desechos.
El uso de grasas oxidadas en la alimentación porcina: beneficios, riesgos y consideraciones. Parte I.
DESCÁRGALO EN PDF
La inclusión de grasas oxidadas en la alimentación del cerdo requiere cuidadosas consideraciones debido a sus efectos sobre la calidad nutricional, la palatabilidad, la digestibilidad y los problemas de salud.
Se deben equilibrar los beneficios y riesgos asociados con el uso de grasas oxidadas e implementar medidas de control de calidad adecuadas.
Cabe destacar que independientemente de los beneficios y repercusiones que tiene el uso de grasas oxidadas en la industria porcina se debe establecer límites a la inclusión de grasas oxidadas en los alimentos para cerdos para garantizar la seguridad y calidad del producto.
Los productores de cerdos deben cumplir con estas regulaciones y monitorear la calidad de la grasa para mitigar los posibles riesgos para la salud de los cerdos.

Eduardo Raele de Oliveira, MV, Msc, PhD
Asesor técnico porcino - Nuproxa Switzerland Ltd
Ana Paula Pinoti Pavaneli, MV, Msc, PhD
Asesora técnica porcino - Nuproxa Switzerland Ltd
La salud del hígado es crucial para la eficiencia productiva de los animales, mejorando la digestión, absorción y utilización de los nutrientes de la dieta.
El hígado desempeña funciones vitales en el organismo, como la desintoxicación, el almacenamiento de energía y nutrientes esenciales, la regulación del sistema endocrino y el metabolismo de proteínas, carbohidratos y lípidos (Liu et al., 2017).
En los sistemas de producción intensiva, en los que se busca el máximo rendimiento productivo, la salud del hígado debe cuidarse y mantenerse para no comprometer la salud metabólica del animal.



En el caso de los animales jóvenes, además de una mayor tasa metabólica debido a su rápido crecimiento y desarrollo, factores relacionados con la producción, como el uso de antimicrobianos, la presencia de micotoxinas en las dietas y la mayor generación de radicales libres debido a agentes estresantes, pueden comprometer la salud y el perfecto funcionamiento del hígado (Cui et al., 2016; Hasuda et al., 2023), además de requerir una mayor tasa metabólica para ser eliminado del organismo.
Por otro lado, la disponibilidad y utilización de nutrientes para el crecimiento y desarrollo de los animales puede verse perjudicada en tales circunstancias.
Buscando ofrecer tecnologías naturales y sostenibles para la producción animal, Nuproxa trae a su portafolio el producto LivoLiv®, un modulador hepático para optimizar el metabolismo y el rendimiento productivo de los animales, mejorando la digestión, absorción y utilización de nutrientes.
Con la presencia de Quercetina y Andrographolide en su composición, dos componentes conocidos por sus propiedades hepatoprotectoras en la medicina tradicional, LivoLiv® se destaca por la acción sinérgica de estos ingredientes.
El producto actúa reduciendo el daño hepático, optimizando la capacidad de desintoxicación del hígado, acelerando sus procesos regenerativos y estimulando la producción de enzimas hepáticas del animal, que a una edad temprana está relacionada con su rápido crecimiento y desarrollo.
Para validar la acción de LivoLiv como acelerador del metabolismo, se decidió evaluar su uso en la fase final de transición de los lechones, a partir de los 35 días de edad, cuando el animal inicia su demanda de crecimiento proteico tras el periodo de destete.
Para la evaluación se seleccionaron 288 lechones (genética DanBred, macho Duroc × hembra DB90).
El estudio se realizó en condiciones experimentales en la granja Folhados (Patrocínio, Minas Gerais, Brasil), una granja comercial de ciclo cerrado y referencia de productividad en el país.
El experimento siguió un diseño de bloques aleatorizados, según el sexo y el peso medio de entrada a los 35 días de edad, en un esquema factorial 2x2.
Se estudiaron dos factores y sus interacciones:
Factor Dieta, dieta de máximo rendimiento utilizada convencionalmente en la granja (Rostagno et al., 2017), o dieta de máximo rendimiento enriquecida con + 10% de aminoácidos y energía;
Factor LivoLiv®, uso o no de LivoLiv® en las dietas (250 g/tonelada).



Los lechones se alojaron en 24 corrales con 12 animales cada uno, con un total de 12 unidades experimentales para cada factor, y 6 unidades experimentales para cada interacción en la evaluación del rendimiento.
Los animales fueron alojados a los 35 días de edad y evaluados zootécnicamente durante las fases de alimentación (35 y 50 días; 50 y 70 días) hasta su salida de la transición (70 días de edad). Para los análisis complementarios, se seleccionaron 20 animales para la evaluación de la producción de enzimas hepáticas y bilirrubina a los 35, 50 y 70 días.

35 días
50 días
70 días
Como resultado, los animales del grupo LivoLiv® presentaron una respuesta de ganancia de peso (GP) superior a la del grupo control a los 50 días y durante todo el estudio, independientemente del uso de una dieta más enriquecida en aminoácidos y energía.
El peso final de los animales a los 50 y 70 días también fue superior para el uso de LivoLiv®, con 435 y 628 g de peso vivo, respectivamente, en comparación con el grupo control, independientemente del factor de la dieta. No se observaron diferencias estadísticas para la interacción de los factores en ninguna de las características zootécnicas evaluadas (Tabla 1)
Tabla 1. Rendimiento en transición según los factores estudiados
IC: Índice de conversión; CMD: consumo medio diario; D: factor de dieta; D x L: interacción de factores; EEM: error estándar de la media; GMD: ganancia media diaria ; L: factor LivoLiv; MD: dieta de máximo rendimiento; MD + 10%: dieta de máximo rendimiento enriquecida con + 10% de aminoácidos y energía.

En las evaluaciones del suero, se observó que los animales suplementados con LivoLiv® desde los 35 hasta los 70 días de edad mostraron una mayor producción de la enzima gamma glutamil transferasa (GGT) a los 50 días en comparación con el grupo no suplementado (Gráfico 1).
Este mismo efecto se observó cuando se ofreció una dieta de máximo rendimiento enriquecida con un 10% de aminoácidos y energía, pero sólo a los 70 días (Gráfico 2). En ambos casos, los valores observados estaban dentro de lo que se considera normal para la especie porcina (González & Silva, 2022).
El uso de la dieta enriquecida también resultó en un aumento de los niveles de bilirrubina total (BT) y bilirrubina directa (BD), también a los 70 días (Gráfico 3) –valores que también están dentro del rango normal para la especie porcina (González & Silva, 2022).
Dado el importante papel que desempeña la bilirrubina en los procesos de digestión y absorción de nutrientes, este resultado muestra que su producción se ve estimulada como consecuencia de la utilización de una dieta más rica en aceite y aminoácidos, lo que genera una mayor demanda sobre el sistema digestivo.

Gráfico 1. Enzima gamma glutamil transferasa (GGT) a los 50 días de edad en lechones suplementados o no con LivoLiv®.
Se evaluaron muestras de 10 animales por grupo experimental.
Gráfico 2. Enzima gamma glutamil transferasa (GGT) a los 70 días de edad en lechones alimentados con diferentes dietas.
Se evaluaron muestras de 10 animales por grupo experimental. MD, dieta de máximo rendimiento; MD + 10%, dieta de máximo rendimiento enriquecida con + 10% de aminoácidos y energía.
Gráfico 3. Bilirrubina total y bilirrubina directa a los 70 días de edad en lechones alimentados con diferentes dietas.
Se evaluaron muestras de 10 animales por grupo experimental. MD, dieta de máximo rendimiento; MD + 10%, dieta de máximo rendimiento enriquecida con + 10% de aminoácidos y energía.

Las principales conclusiones de este estudio son que la suplementación de las dietas de transición con LivoLiv® son capaces de estimular anticipadamente la producción de la enzima hepática GGT, que en animales jóvenes y plenamente desarrollados está relacionada con el metabolismo de los aminoácidos y la energía para un crecimiento y rendimiento máximos.
En la práctica, el uso de LivoLiv® en las condiciones experimentales aquí presentadas se traduce en un mayor peso final de los lechones a los 50 y 70 días de edad, con una ganancia de peso diaria máxima durante los primeros días de utilización del suplemento poliherbal.
LivoLiv® es una solución natural eficaz y comprobada para modular el hígado, actuar como estimulante y acelerador metabólico y mejorar el rendimiento en la fase de transición. Sin duda, un producto más de Nuproxa comprometido con una producción animal más eficiente, sostenible y rentable en toda América Latina.
Salud hepática y metabolismo: ¿cómo optimizarlos para mejorar el rendimiento de los lechones en la fase de cría? DESCARGAR PDF





Cecilia Cajarville y José Luis Repetto Departamento Producción Animal y Salud de Sistemas Productivos – IPAV, Facultad de Veterinaria, UdelaR, San José, Uruguay. (Dirección actual: Departament de Ciència Animal i dels Aliments, Facultat de Veterinària, UAB, Bellaterra, Barcelona)


Frecuentemente se habla de los efectos adversos de los taninos, y existen razones para ello. Estos compuestos tienen la capacidad de unirse a las proteínas, precipitándolas y reduciendo su digestibilidad o incluso volviéndolas indigestibles.



En ocasiones también interfieren en la digestión de carbohidratos valiosos como el almidón, al envolver sus corpúsculos, lo que ya hemos discutido en anteriores artículos cuando describíamos las características del grano de sorgo.
Esta es la principal razón por la que los alimentos ricos en taninos no son bien aprovechados por los animales en general, y los rumiantes en particular.
Sin embargo, los taninos también presentan una faceta positiva que puede ofrecer numerosos beneficios si se emplean adecuadamente.
Los taninos son compuestos naturales presentes en numerosas plantas, especialmente en la corteza, hojas y frutos. Estos compuestos son una forma de polifenoles, caracterizados por tener múltiples grupos hidroxilo (-OH) en su estructura.
Existen dos categorías principales de taninos:
Los taninos correctos, en las dosis apropiadas, pueden mejorar la utilización de las proteínas en el rumen y contribuir a la reducción de las emisiones de gases de efecto invernadero (GEIs). ¿Cómo? En las siguientes páginas, explicaremos el mecanismo de acción de los taninos y proporcionaremos consejos prácticos para su mejor utilización.

Derivados del ácido gálico, un compuesto fenólico que se presenta como un sólido cristalino blanco o amarillento. Estos taninos se descomponen en el organismo bajo ciertas condiciones, como el medio ácido del abomaso. Además, poseen propiedades antioxidantes. Se encuentran en diversas plantas, siendo el té, la castaña de indias y algunas frutas sus fuentes más comunes (ver figura 1)
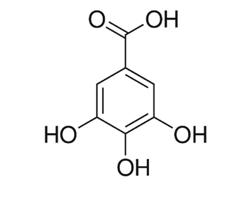
Figura 1. Estructura del ácido gálico, molécula básica en la constitución de los taninos hidrolizables.

También conocidos como proantocianidinas, son oligómeros de flavonoides que no se descomponen fácilmente en el organismo. Estos taninos se caracterizan por su sabor astringente, que provoca una sensación seca y rugosa en la boca al consumir alimentos o bebidas que los contienen, como el vino, el té y ciertas frutas (ver figura 2)
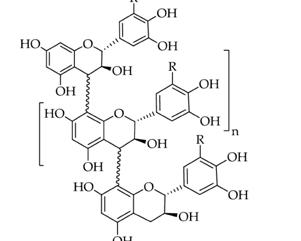
Figura 1. Ejemplo de estructura de taninos condensados.
1. Ejemplos de plantas productoras de taninos (resumido de Besharati et al., 2022)
En las plantas, los taninos desempeñan funciones de defensa contra herbívoros, hongos, bacterias y virus. En situaciones de estrés, incrementan la producción de taninos, lo que les sirve como mecanismo de supervivencia (Iqbal y Poór, 2024).
Además, se les atribuyen propiedades beneficiosas para la salud de los animales, tales como efectos antioxidantes y antiinflamatorios. Sin embargo, en exceso, pueden interferir en la absorción de algunos nutrientes.
La inclusión de taninos en la dieta de los rumiantes, en concentraciones adecuadas, puede ofrecer importantes beneficios para su salud y rendimiento. En la tabla 1 ejemplificamos algunos de los taninos que han sido empleados o tienen potencial para ser empleado en alimentación animal.
Castaña Castanea sativa Tronco, hojas condensados
Quebracho colorado Schinopsis forentzil troncos condensados
Acacia negra Acacia mearnsii cortezas condensados
Roble europeo Quercus robur corteza condensados / hidrolizables
Lotus (bird’s foot) Lotus corniculatus hojas, flores condensados
Vid común Vitis vinifera frutos condensados
Alcachofa Cynara cardunculus hojas condensados
Mirto Myrtus communis hojas condensados / hidrolizables
Nogal común Juglans regiabarks hojas, flores hidrolizables
Café Coffea arabica pulpa condensados
Té Camelia sinensis hojas condensados / hidrolizables

1


2

Figura 3. Árboles y plantas que dan origen a taninos de uso en alimentación animal. 1: Quebracho colorado; 2: Lotus corniculatus; 3: Castaño; 4: Acacia Mearnsii
Estos compuestos polifenólicos tienen la capacidad de unirse a las proteínas, lo que ralentiza su degradación en el rumen y permite que una mayor cantidad de proteínas llegue intacta al intestino delgado. Esto incrementa la disponibilidad de aminoácidos esenciales, mejorando la utilización del nitrógeno. Esta capacidad puede ser interesante especialmente en dietas con exceso de proteína degradable, como las que se basan en pasturas de alta calidad (Orlandi et al., 2015).
3

4
Un estudio de Min et al. (2003) analizó el impacto de los taninos concentrados en los forrajes destinados a rumiantes. El impacto positivo de los taninos en la digestión proteica se ha demostrado con concentraciones bajas (20-45 g de taninos condensados/kg de materia seca).
A estas concentraciones, los taninos presentes en ciertas plantas forrajeras, como Lotus corniculatus, han mostrado aumentar la producción de leche, favorecer el crecimiento de la lana, mejorar la tasa de ovulación y el porcentaje de partos, además de reducir el riesgo de timpanismo y la carga de parásitos internos (Frutos et al., 2004).

Sin embargo, cuando la concentración de taninos condensados supera los 55 g/kg de materia seca, la ingesta voluntaria y la digestibilidad de los nutrientes pueden verse comprometidas, lo que provoca un efecto adverso en la producción. La disminución en la ingesta puede atribuirse a una menor palatabilidad del alimento ocasionada por el sabor astringente de los taninos.
Dado que no todos los rumiantes son sensibles a la astringencia, se han desarrollado también otras teorías que relacionan las reducciones del consumo con factores más vinculados a posibles efectos adversos de un exceso de taninos sobre la digestión.
En ese sentido, es posible que alteraciones en la microbiota ruminal deriven en enlentecimiento de la fermentación con sensaciones de saciedad, o que los animales desarrollen aversiones y rechazo hacia los taninos (Frutos et al, 2004), provocadas por malestares digestivos sufridos en experiencias anteriores al consumir alimentos altos en taninos (Salem et al., 2009).
Continuando con los efectos positivos, además de sus efectos sobre la degradación proteica, los taninos poseen propiedades antiinflamatorias, ya que pueden inhibir la activación de células inmunitarias y reducir la producción de citoquinas y enzimas proinflamatorias, contribuyendo así al control de la inflamación y a la promoción de la salud general del animal.

Son potentes antioxidantes, neutralizando los radicales libres y previniendo la peroxidación de lípidos (Fraga del Corral et al., 2021). En dosis bajas, los taninos tienen la capacidad de modular la microbiota ruminal, como veremos más adelante. Adicionalmente, se ha demostrado que los taninos tienen efectos antiparasitarios, siendo eficaces contra parásitos internos comunes en ovinos, como Haemonchus contortus, y en bovinos, como Ostertagia ostertagi y Cooperia oncophora (Hoste et al., 2006; Naumann et al., 2017). Esto resalta aún más su utilidad en el manejo sanitario de los rumiantes.
Finalmente, algunos estudios sugieren que los taninos podrían extender la vida útil de los productos cárnicos de animales suplementados al mejorar la estabilidad oxidativa y reducir la decoloración de la carne (Luciano et al., 2009; Jenko et al., 2018).



La inclusión de taninos en las dietas de rumiantes ofrece un potencial significativo para promover la sostenibilidad en la producción animal al reducir las emisiones de gases de efecto invernadero.
Este enfoque ha suscitado un creciente interés en la investigación científica en los últimos años, a medida que se buscan soluciones efectivas en base a productos naturales o determinados alimentos, para minimizar el impacto ambiental de la ganadería.
Min y Solaiman (2018) reportaron reducciones lineales en la producción de metano (CH₄) en cabras y ovejas a medida que aumentó la concentración de taninos en sus dietas. Este hallazgo fue respaldado por varias revisiones de literatura (CardozoGutiérrez et al., 2021) y metaanálisis recientes (Scholz et al., 2023).
En ellas se comunican que numerosos resultados in vitro e in vivo respaldan el concepto de que el incremento del nivel de taninos condensados condujo a una disminución de las concentraciones de CH₄ ruminal y a un aumento en las proporciones molares de propionato, butirato y ácidos grasos volátiles totales.
El mecanismo a través del cual los taninos reducen la emisión de metano estaría relacionado con la modulación de la microbiota ruminal, ya que tienen un efecto inhibitorio sobre ciertos microorganismos ruminales metanógenos, protozoarios y otros productores de hidrógeno (Patra y Saxena, 2010). Sin embargo, la efectividad de los taninos puede variar según su tipo y concentración.
Fonseca et al. (2023) observaron que, aunque algunos estudios plantean que los taninos condensados son más efectivos para reducir la producción de metano. Los taninos hidrolizables tienden a ser más fácilmente degradados en el rumen, lo que puede llevar a efectos menos predecibles en la producción de metano.


Dada la propiedad de los taninos para reducir degradación proteica, el consumo de alimentos ricos en taninos (Eugène et al., 2021) o la adición de taninos a las dietas (Pozo et al., 2022; Batelli et al., 2024) produce un cambio en la excreción de nitrógeno. Hay más proteína de la dieta que pasa al intestino delgado y se absorbe menos NH3 a través de la pared ruminal.
Lo anterior se traduce en que una mayor proporción del nitrógeno no aprovechado se elimina a través de las heces en lugar de eliminarse por la orina. Dado que el nitrógeno fecal es más estable, se reduce la liberación de amoníaco (NH₃) y óxido nitroso (N₂O) en la atmósfera. Por ello, en general se considera que una mayor eliminación de nitrógeno a través de las heces es favorable para el medio ambiente.
Para lograr efectos positivos sobre las emisiones de gases de efecto invernadero sin reducir la producción, la dosis de tanino administrada es fundamental. Una dosis baja puede no generar efectos significativos, mientras que una dosis demasiado alta puede tener efectos anti nutricionales como comentamos anteriormente, disminuyendo la ingesta de alimento y la digestibilidad.
Por ejemplo, un estudio realizado por Aguerre et al. (2016) con vacas lecheras evaluó el efecto de adicionar dosis progresivas de una mezcla de extracto de taninos de quebracho y castaño. Los resultados fueron por demás prometedores.
Pequeñas cantidades (0,45%) mejoraron la eficiencia alimentaria, reduciendo el amoníaco ruminal, el nitrógeno urinario y el metano, además de observarse un

Sin embargo, dosis mayores resultaron en una disminución del consumo y de la producción de proteína láctea, lo que resalta la necesidad de un balance adecuado en el uso de taninos en las dietas de rumiantes para maximizar sus beneficios ambientales y nutricionales.
Queda mucha investigación por delante, pero estos resultados sugieren que los taninos condensados pueden ser una alternativa natural eficiente para mitigar el impacto ambiental de los sistemas de producción de rumiantes.

Los taninos pueden ser beneficiosos para la salud de los rumiantes, pero su inclusión en la dieta debe manejarse con cuidado para evitar efectos negativos. Como se mencionó, además de interferir con la digestión y absorción de nutrientes valiosos como proteínas y carbohidratos, también pueden perjudicar el aprovechamiento de otros nutrientes.
Los taninos condensados pueden unirse a minerales como aluminio, calcio, cobre, hierro y zinc, lo que puede reducir su biodisponibilidad (Naumann et al., 2017).
El consumo excesivo puede causar irritación gastrointestinal, manifestándose como diarrea, cólicos y sequedad de boca. En casos severos, se ha asociado la ingestión excesiva de taninos con toxicidad hepática y renal (Mbatha et al., 2002).
La toxicidad de los taninos varía según el tipo, siendo los taninos hidrolizables generalmente más tóxicos que los condensados (Naumann et al., 2017). La sensibilidad a los taninos también varía entre especies; algunos rumiantes pueden
Los taninos son compuestos que, cuando se encuentran en cantidades adecuadas, pueden ofrecer una serie de beneficios para los rumiantes, reduciendo a la vez las emisiones GEIs que estos generan. Mejoran el estatus sanitario e inmunológico del animal, controlan los parásitos, modulan la microbiota e incrementan la utilización proteica, aumentando con todo ello la producción e incluso los productos animales.
Si a ello le sumamos la característica de ser productos directamente extraídos de la naturaleza pensamos que los taninos podrán jugar un papel cada vez mayor en lograr sistemas de alimentación más eficientes y sostenibles. La investigación sobre las fuentes de taninos y sus dosis es necesaria para poder optimizar su uso y transformar estos compuestos en aliados para la nutrición de rumiantes.


Taninos en la nutrición de rumiantes: de obstáculo a oportunidad DESCÁRGALO EN PDF
Departamento científico VetosEurope

La creciente demanda mundial de productos lácteos y cárnicos exige una intensificación de la producción ganadera. Sin embargo, esta intensificación debe ser sostenible, minimizando el impacto ambiental y asegurando la salud animal.
ANAVRIN®, desarrollado por VetosEurope, se presenta como una solución innovadora que aborda ambos desafíos, mejorando la eficiencia productiva y reduciendo la huella de carbono de las explotaciones ganaderas.
Este aditivo es una mezcla sinérgica de aceites esenciales, taninos y bioflavonoides de origen natural, cuidadosamente seleccionados para optimizar la función ruminal.


VetosEurope, fundada hace 11 años, combina la experiencia ganadera de la familia
Ghizzoni con el conocimiento científico en complementos alimenticios naturales para rumiantes.
Tras utilizar internamente sus productos en más de 1.000.000 de animales durante 10 años, y respaldados por más de 20 pruebas científicas, decidieron comercializar ANAVRIN®, un producto diseñado para mejorar el rendimiento productivo de los rumiantes y reducir simultáneamente sus emisiones de metano y CO2, basado en una filosofía de sostenibilidad mediante el uso de ingredientes naturales.
ANAVRIN® actúa sobre la fermentación ruminal a través de un mecanismo multifacético. Los aceites esenciales, provenientes de Syzygium aromaticum (clavo), Coriandrum sativum (cilantro) y Pelargonium cucullatum (geranio), modulan la actividad de la microbiota ruminal, favoreciendo el crecimiento de bacterias benéficas y disminuyendo la producción de metano (CH4), un potente gas de efecto invernadero.
El eugenol (4-alil-2-metoxifenol), presente en el clavo, exhibe propiedades antimicrobianas de amplio espectro, mientras que los aceites de cilantro y geranio contribuyen a una mejor digestibilidad.
Los taninos de Castanea sativa (castaño), por su parte, impactan positivamente en el metabolismo proteico, reduciendo las pérdidas de nitrógeno y ofreciendo un efecto antiinflamatorio que favorece la salud intestinal.
Finalmente, los bioflavonoides de Olea europaea (aceituna) aportan beneficios antioxidantes y antimicrobianos, contribuyendo al bienestar general del animal.
Impactos positivos en la producción y en la reducción de emisiones de metano.
Empleo exclusivo de materias primas y componentes naturales e idénticos a los naturales.
Uso de las más sofisticadas tecnologías productivas e industriales certificadas Made in Suiza.
Capacidad de control de toda la cadena productiva, desde el productor hasta la venta al consumidor final.
Presencia en el territorio a nivel global a través de una red de venta internacional.
Perfecto conocimiento del mercado bovino a nivel mundial.
Una de las fortalezas de ANAVRIN® es que sustenta su desarrollo y su posicionamiento sobre rigurosos trabajos científicos realizados en universidades y centros de investigación.
Numerosas investigaciones científicas realizadas en prestigiosas instituciones internacionales, han demostrado la eficacia de ANAVRIN®. De hecho, en los últimos años se han desarrollado estudios tanto in vitro como in vivo, y en diferentes especies animales (vacas lecheras, novillos de engorde, ovejas) en la Universita degli studi di Milano y Universita degli studi di Sassari (Italia), Universidad de la República (Uruguay), Wageningen University-Research (Países Bajos), Universidade Federal de Santa María (Brasil) y Animal New Tech (Israel).
Complementan estos resultados, pruebas de campo realizadas en alrededor de 40 establecimientos de

Imagen 1. Empleo de SF6 para medición de metano en vacas lecheras (IPAV-FVet, UdelaR, Uruguay)
ANAVRIN® reduce significativamente las emisiones de metano, sin afectar negativamente el consumo de alimento ni la eficiencia alimenticia.
Este efecto se ha observado en vacas lecheras (Parand, 2024; Sgoifo Rossi et al., 2022), novillos de engorde (Montini et al., 2024, Figura 1) y ovejas gestantes (Atzori et al., 2022).
Estados Unidos


Figura 1. Reducción porcentual del metano emitido en términos de emisión total diaria (g/día), con relación a la materia seca (MS) ingerida diariamente, y en relación al peso del animal (Montini et al., 2024).
ANAVRIN® incrementa la productividad ganadera, mejorando el consumo de alimento, la conversión alimenticia, y la producción de carne y leche. Estudios en terneros Holstein Friesian (Grossi et al., 2024) y toros Charolaise (Sgoifo Rossi et al., 2023) mostraron mayor ganancia de peso, peso final y consumo de alimento, así como mejor eficiencia alimenticia.
En novillos de carne, ANAVRIN® mostró un rendimiento similar a la monensina (Ciancio et al., 2022), un reconocido promotor del crecimiento, siendo ANAVRIN® un aditivo natural, que no contiene antibióticos.
En vacas lecheras mientras tanto, en un experimento realizado con 140 vacas Holstein Friesian se observaron mejoras en la ingesta, en la conversión alimenticia y en la producción de leche (Sgoifo Rossi et al., 2022), atribuidas a una mayor digestibilidad.
Imagen3. Empleo de cámaras respiratorias para medición de las emisiones de metano en Wageningen, Países Bajos.
Imagen 2. Empleo de la técnica de SF6 para medición de metano en condiciones de campo (IPAV-FVet, UdelaR, Uruguay)
ANAVRIN®, gracias a componentes vegetales con probada acción sobre la inmunidad, el estrés oxidativo y la salud en general, presenta beneficios en la salud animal. Ello se constató en estudios específicos que han demostrado una disminución en la incidencia de enfermedades respiratorias y una mejora del estado inmunitario en ganado de carne (Sgoifo Rossi et al., 2023).
Además, en novillos de engorde, ANAVRIN® condujo a un pH ruminal más alto, reduciendo el riesgo de acidosis (Ciancio et al., 2022).

La sólida evidencia científica que respalda la eficacia de ANAVRIN® ha resultado en la obtención de importantes certificaciones internacionales, reconociendo su contribución a la sostenibilidad ambiental.
Reducción de Emisiones de Metano:
El 11 de julio de 2022, Carbon Trust Assurance Limited certificó a ANAVRIN® como un agente eficaz en la reducción de emisiones de metano en rumiantes, validando rigurosos estudios de laboratorio y campo.
Generación de Créditos de Carbono:
ANAVRIN® abre nuevas oportunidades en el mercado de carbono. La capacidad comprobada de ANAVRIN® para reducir emisiones de metano permite la generación de créditos de carbono, financiando proyectos de mitigación climática.
En Uruguay, en colaboración con productores ganaderos y bajo la supervisión de Verra, se ha iniciado este proceso, generando un valor comercial adicional para la producción ganadera sostenible.
ANAVRIN® se presenta como una herramienta valiosa para la ganadería moderna, que busca eficiencia y sostenibilidad. Su acción multifacética, basada en componentes naturales, permite optimizar el proceso digestivo ruminal, mejorando la eficiencia productiva y reduciendo las emisiones de metano.
Los resultados de numerosos estudios científicos avalan su eficacia y prometen un futuro más sostenible para la industria ganadera.
Referencias disponibles en la versión web del artículo en nutrinews.com

ANAVRIN®: Una solución innovadora para la optimización de la producción ganadera DESCÁRGALO EN PDF



