Master Thesis ǀ Tesis de Maestría
submitted within the UNIGIS MSc programme presentada para el Programa UNIGIS MSc

at/en
Interfaculty Department of Geoinformatics- Z_GIS Departamento de Geomática – Z_GIS University of Salzburg ǀ Universidad de Salzburg
Evaluando la efectividad de los modelos de distribución de especies desde una perspectiva ecológica en la especie Ischnura capreolus en Colombia
the effectiveness of species distribution models from an ecological perspective in the species Ischnura capreolus in Colombia
by/por
2019C Néstor Fabián Galindo Ruiz
11947763
A thesis submitted in partial fulfilment of the requirements of the degree of Master of Science– MSc
Advisor ǀ Supervisor: Leonardo Zurita Arthos PhD
Bogotá - Colombia, December, 2022
Evaluating
Compromiso de Ciencia
Por medio del presente documento, incluyendo mi firma personal certifico y aseguro que mi tesis es completamente el resultado de mi propio trabajo. He citado todas las fuentes que he usado en mi tesis y en todos los casos he indicado su origen.
Bogotá - Colombia, diciembre de 2022

Agradecimientos
A la naturaleza, por proveer fuentes inagotables de maravilla a los ojos del científico y a mi mentor Emilio Realpe, por motivarme a no perder esa capacidad de asombro.
Si el modelo lo dice, ¿la especie está ahí?
Evaluando la efectividad de los modelos de distribución de especies desde una perspectiva ecológica en la especie Ischnura capreolus en Colombia.
RESUMEN
Los modelos de distribución de especies permiten establecer desde la distribución potencial de una especie hasta establecer rutas de conservación, tanto del hábitat como de la especie. Ha sido empleado como una estrategia para conectar y caracterizar la información de presencia de un grupo biológico con información ambiental, mediante la implementación de herramientas estadísticas. Sin embargo, unos limitantes de este tipo de estudio es que el contexto ecosistémico no es fácil de incorporar y por ende muchas veces no es tenido en cuenta. Esto debido a que la información que se usa para parametrizar el modelo es de tipo continuo, siendo el medio ambiente, difícilmente asociado hacia variables del tipo numérico. La naturaleza de la información biótica, en gran medida la que determina o no la permeabilidad de una especie a un territorio difícilmente se puede asociar a una variable numérica. Por esto, el presente estudio propone una metodología aplicada para evaluar la pertinencia de los modelos de distribución de especies, de tal manera que se permite evaluar y poner en contexto si el área predicha por un modelo presenta la conectividad necesaria para que la especie pueda cruzar, es decir sumando ese faltante de los modelos tradicionales. Para esto, se desarrollaron cinco modelos de predicción de distribución de especies, y para vencer la limitación de estos se evaluó la capacidad que tiene la especie de interés en moverse a través del área predicha. Los modelos se construyeron con la herramienta R-project y el análisis de conectividad ecológica con el programa ArcGIS. En este caso, para la especie I. capreolus, se evidencia una gran aplicabilidad de la metodología, ya que las curvas AUC muestran una gran capacidad predictiva de los cinco modelos analizados Sin embargo, desde una perspectiva de conectividad ecológica se reduce la capacidad predictiva o precisión de estos, esto indica que puede que ambientalmente exista la posibilidad de que habite (nicho potencial), sin embargo, difícilmente las condiciones bióticas (barreras) facilitarían que la especie se desplace hacia toda la zona predicha. Esto indica un potencial uso para determinar la capacidad predictiva de los modelos de distribución de especies, y con esto los análisis de modelamiento de nicho serán más robustos, superando una de las limitaciones que presentan.
Palabras clave:
Modelos de distribución de especies, conectividad ecológica, Ischnura capreolus, Colombia.
ABSTRACT
Species distribution models allow to predict the potential distribution of a species, to build conservation routes, both for the habitat and the species. It has been used as a strategy to connect and characterize the information from the presence data of a taxa linked to environmental information, through the implementation of statistical tools. However, this analysis has some limitations, in this type of studies it is not easy to incorporate categoric data. This is because the information used to parameterize the model has to be continuous, being the environment, hardly associated with variables of the numerical type. Biotic information that determines the permeability of a species through a habitat is hardly associated with a numerical variable. For this reason, the present research proposes an applied methodology to evaluate the relevance of species distribution models, in such a way that it is possible to put in context if the area predicted by a model presents the necessary connectivity so that the species can cross this area, integrating categorical data, solving one of the limitations of species distribution models. For this, five species distribution prediction models were developed, andto overcome their limitation, the ability ofthe species of interest to move through the predicted area was evaluated. The models were built with R-project statistical tool and the ecological connectivity analysis with ArcGIS software In this case, for the species I.capreolus, great applicability of the methodology is evidenced, since the AUC curves show a great predictive capacity of the five models analyzed. However, from an ecological connectivity perspective, the predictive capacity is reduced or precision of these. This indicates that there may be an environmental possibility that it inhabits a potential niche, however, the biotic conditions (barriers) would hardly facilitate the species to move to the entire predicted area. This indicates a potential use to determine the predictive capacity of species distribution models, and with this the niche modeling analyzes will be more robust, overcoming one of the limitations it presents.
Key words:
Species distribution models, ecological connectivity, Ischnuracapreolus, Colombia.
6
RESUMEN 4 ABSTRACT 5 1 INTRODUCCIÓN 14 1.1 Antecedentes------------------------------------------------------------------------------------14 1.2 Objetivos 15 1.2.1 General 15 1.2.2 Específicos 15 1.3 Preguntas de investigación 15 1.4 Hipótesis 15 1.5 Justificación--------------------------------------------------------------------------------------15 1.6 Alcance 16 2 REVISIÓN DE LITERATURA 18 2.1 Grupo de estudio 18 2.2 Modelo de distribución de especie 21 2.3 Convergencia insectos y MDE -------------------------------------------------------------- 25 3 METODOLOGÍA 28 3.1 Zona de estudio 28 3.2 Obtención de las capas ambientales 29 3.2.1 Análisis de correlación 30 3.3 Datos de presencia ----------------------------------------------------------------------------31 3.4 Construcción del modelo de distribución de especies -31
TABLA DE CONTENIDO
7 3.5 Propuesta metodológica 32 3.5.1 Análisis de Resistencia -33 3.5.2 Conectividad entre coberturas 36 3.5.3 Creación de la matriz de resistencia. 37 4 RESULTADOS----------------------------------------------------------------------------------------38 4.1 Registros para la especie Ischnuracapreolus 38 4.2 Distribución potencial de la especie Ischnuracapreolus 39 4.2.1 Resultados BIOCLIM 39 4.2.2 Resultados RPART 42 4.2.3 Resultados GAM 45 4.2.4 Resultados GLM 47 4.2.5 Resultados Maxent 50 4.2.6 Integrado 53 4.3 Análisis de conectividad ecológica 54 4.4 Evaluación de los modelos en contexto ecosistémico---------------------------------56 5 DISCUSIÓN 60 6 CONCLUSIONES 67 7 REFERENCIAS 69
LISTADO DE TABLAS
8
Tabla 1 Variables categóricas involucradas en el análisis ecológico y de resistencia de la especie I.capreolus. 34 Tabla 2 Lineamientos para la recategorización de las variables categóricas en función de la respuesta ecológica de la especie I.capreolus. 35 Tabla 3 Calificación de la resistencia del territorio en función del tipo de cobertura. 35 Tabla 4 Escala de clasificación entre variables 36 Tabla 5 Formato ejemplo para comparar las variables de interés------------------------------ 36 Tabla 6 Interacción entre variables/coberturas, según la preferencia de hábitat. 37 Tabla 7 Resumen resultados de los modelos implementados para la predicción de la distribución de la especie Ischnuracapreolus ------------------------------------------------------54 Tabla 8 Porcentaje de permeabilidad de la especie, respecto al área predicha por cada modelo de distribución presentado. 64
LISTADO DE FIGURAS
9
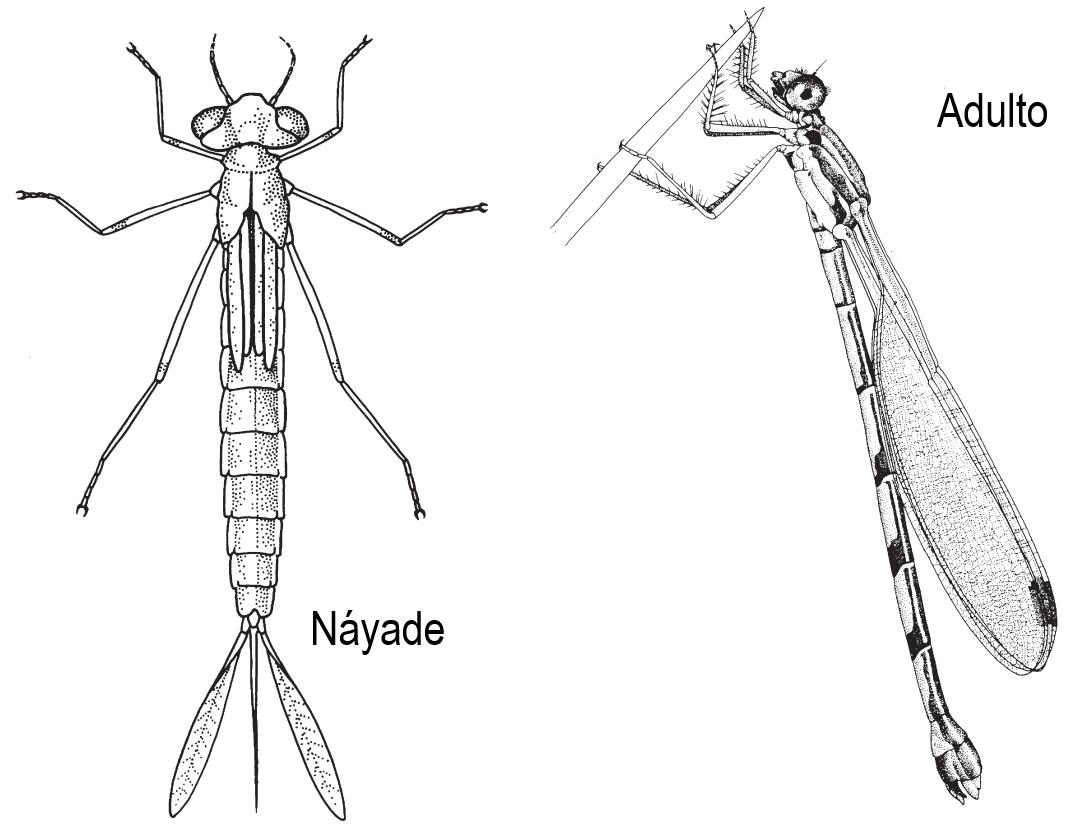
Figura 1 Morfología general del Orden Zygoptera. 12 Figura 2 Morfología general del Orden Anisoptera. 12 Figura 3 Interacción nicho, visto como recursos y distribución geográfica con dos especies diferentes (Fischer y Lindenmayer, 2007). 20 Figura 4 Se muestra en A la estructura en forma de ‘C’ de los paraproctos del macho y en B los tubérculos del pronto de la hembra (modificado de Garrison et al. 2010). 21 Figura 5 Flujo de trabajo de un MDE (J. Brown y Yoder, 2015).-------------------------------- 24 Figura 6 Colombia, delimitada como área de estudio para la presente investigación. 28 Figura 7 Visualización preliminar de las capas ambientales implementadas usadas para la formulación de los modelos de distribución de la especie de interés.-------------------------30 Figura 8 Análisis de correlación entre las capas implementadas para los modelos de distribución de la especie. 31 Figura 9 Esquema de trabajo para la obtención de los modelos de distribución potencial de Ischnuracapreolus. 32 Figura 10 Distribución reportada de la Ischnuracapreolus en Colombia, a partir de información encontrada en el Museo de Historia Natural CJ Marinkelle. 38 Figura 11 Curva ROC para el modelo BIOCLIM.---------------------------------------------------39 Figura 12 Importancia de las variables en la construcción del modelo a partir de BIOCLIM. 40 Figura 13 Caracterización de las condiciones ambientales según el modelo BIOCLIM.--41 Figura 14 Mapa de distribución potencial construido a partir del método BIOCLIM. 42 Figura 15 Curva ROC para el modelo RPART. 42 Figura 16 Importancia de las variables en la construcción del modelo a partir de RPART. 43
Figura 32 Se presentan los análisis de resistencia en función de los resultados de modelo de distribución de especie. A. RPART, B. Maxent, C. Bioclim, D. GLM y E. GAM. Las coloraciones más cálidas indican una mayor resistencia ecológica, es decir presencia de barreras, mientras el verde es menor resistencia, zonas de hábitat permeable para la especie.
Figura 33 Se presentan los análisis de resistencia en función de los resultados de modelo de distribución de especie, como variables categóricas, de permeable o barrera ecológica.
A. RPART, B. Maxent, C. Bioclim, D. GLM y E. GAM. En rojo se presentan las zonas con alta restricción ecológica y en verde las zonas sin restricciones ecológicas
10 Figura 17 Caracterización de las condiciones ambientales según el modelo RPART. 44 Figura 18 Mapa de distribución potencial construido a partir del método RPART. 44 Figura 19 Curva ROC para el modelo GAM. 45 Figura 20 Importancia de las variables en la construcción del modelo a partir de GAM.-46 Figura 21 Caracterización de las condiciones ambientales según el modelo GAM. 46 Figura 22 Mapa de distribución potencial construido a partir del método GAM. 47 Figura 23 Curva ROC para el modelo GLM. 47 Figura 24 Importancia de las variables en la construcción del modelo a partir de GLM. -48 Figura 25 Caracterización de las condiciones ambientales según el modelo GLM. 49 Figura 26 Mapa de distribución potencial construido a partir del método GLM. 50 Figura 27 Curva ROC para el modelo Maxent 50 Figura 28 Importancia de las variables en la construcción del modelo a partir de Maxent. ------------------------------------------------------------------------------------------------------------------51 Figura 29 Caracterización de las condiciones ambientales según el modelo Maxent. 52 Figura 30 Mapa de distribución potencial construido a partir del método Maxent. 52
31 Resultado del análisis de resistencia para todo el territorio colombiano. 55
Figura
59
66
GLOSARIO
AIC: Criterio de Información de Akaike, AIC por sus siglas en inglés Akaike Information Criteria.
GAM: Modelo Aditivo Generalizado, GAM por sus siglas en inglés Generalized Additive Model.
GLM: Modelo Lineal Generalizado, GLM por sus siglas en inglés GeneralizedLinearModel
Especie: Será un taxa que mínimamente sea capaz de reproducirse y producir una progenie fértil, además de contar con características ecológicas y comportamentales particulares y únicas. Sin embargo, esta definición es muy amplia y en la historia de la taxonomía tendrá desviaciones y excepciones particulares a la historia natural de ciertos grupos (por ejemplo, virus, plantas, hongos y bacterias) (Begon et al., 2006).
IDEAM: Siglas del Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia
IGAC: Siglas del Instituto Geográfico Agustín Codazzi de Colombia.
Insecto: De manera genérica, se le define como aquellos organismos que tienen tres tagmas (cabeza, tórax y abdomen), tres pares de patas y dos pares de alas (Brown, 1995)
SDM: Modelo de distribución de especies, en sus siglas en inglés Species Distribution
Model.
Odonata: Grupo de insectos que tiene un estadio netamente acuático y otro terrestre. El adulto macho posee un aparato reproductor secundario en el segundo segmento abdominal. Sus piezas bucales en estadios larvales se caracterizan por la presencia de un labio prominente y proyectable y los adultos poseen mandíbulas altamente desarrolladas (Brown, 1995)
Zygoptera: Este suborden se caracteriza por presentar una torsión torácica que simula un plegamiento de las alas (véase la Figura 1). Adicionalmente, las larvas presentan cuerpos más esveltos y delgados que los Anisoptera, y poseen lamelas caudales para el intercambio gaseoso (Brown, 1995)
11
Anisoptera: Este suborden se caracteriza por no lograr plegar las alas, dejándolas expuestas en todo momento. Por otro lado, presentan cuerpos robustos y grandes en comparación a los Zygoptera. Finalmente, sus larvas no presentan lamelas caudales para el intercambio gaseoso (véase la Figura 2).

Región Geográfica: Hace referencia a una zona espacial, la cual posee particularidades afines, como las condiciones bióticas, abióticas y humanas (pueden ser culturales, espirituales, históricas, etc.) (Olcina Cantos, 1996)
R-Project: Es una herramienta computacional, mediante la cual se pueden realizar análisis estadísticos. Mediante este programa se desarrollaron los modelos de distribución de especie, sin embargo, R no es una herramienta que por defecto tenga múltiples funciones. Al ser un software libre, se puede añadir paquetes o extensiones que le atribuyen análisis
12
Figura 1 Morfología general del Orden Zygoptera.
Figura 2 Morfología general del Orden Anisoptera.
o posibilidades adicionales a la herramienta. En este caso, se descargó la extensión SpeciesDistributionModel – SDM, la cual permite en simultáneo realizar múltiples modelos de distribución de especie. Esto es facilitado porque R, la no tener una interfaz gráfica, es un programa con bajos requerimientos computacionales, haciendo el proceso de modelado más sencillo. Otro paquete usado es ggplot2, el cual es ampliamente usado para la producción de gráficas, el cual es muy versátil y facilita la comunicación científica.
Variable: Variable estadística es un atributo de una muestra o población que puede variar o tomar valores diferentes. Usualmente, es aquellos que se emplea para caracterizar, describir o diferencias en un proyecto de investigación, y puede ser de índole, social, ambiental, entre otros, según el objeto de estudio. Puede agruparse en dos grupos, ser una variable numérica o categórica, las cuales se diferencias a continuación.
• Numérica: Es una variable de tipo cuantitativo, y describe una cantidad, y se diferencia entre discreta, cuando debe tomar valores enteros como número de hijos o número de personas, y continua, cuando puede tomar infinitos valores o intermedios, como la temperatura o la concentración de una sustancia
• Categórica: Esta variable permite la clasificación, la cual puede tomar atributos cualitativos o cuantitativos según el origen de la pregunta de investigación, un ejemplo usado en las métricas poblacionales son los rangos de edad: niño, joven, adulto-joven, adulto, anciano; o de ordenamiento escolar, Primero, Segundo y Tercero.
13
Si el modelo lo dice, ¿la especie está ahí?
Evaluando la efectividad de los modelos de distribución de especies desde una perspectiva ecológica en la especie Ischnura capreolus en Colombia.
1 INTRODUCCIÓN
A continuación, en el presente capítulo se caracteriza la exploración para el establecimiento y consolidación del diseño experimental del proyecto de tesis.
1.1 Antecedentes
Los modelos de distribución de especies han sido una herramienta que, a lo largo del tiempo, han venido siendo muy utilizados, desde aspectos sencillos como caracterizar o entender las condiciones climáticas que facilitan el establecimiento de una especie, hasta entender el área de distribución o riesgo asociado al hábitat de la especie. En ambos sentidos, siendo relevante en aspectos de conservación, o aumento del conocimiento de las especies. Claro está que es una herramienta que ha sido fuente de críticas en la comunidad científica, y todo a partir de la facilidad que hay para su implementación y desarrollo a través de herramientas computacionales. Esa facilidad ha hecho que las publicaciones sean rápidas y a veces descuidadas, lo que hace que la herramienta se asocie con falencias conceptuales y de aplicabilidad en función de la escritura de los investigadores. Sin embargo, esto no es ajeno a otras ramas del conocimiento, donde constantemente las “autoridades” en los temas controversiales deben establecer y homogenizar los conceptos adecuados.
Desafortunadamente, una de las críticas más repetitivas de los modelos de distribución de especie es la falta de conexión que tienen estos estudios con la realidad ecosistémica de una especie. Ya que si bien, ambientalmente, puede habitar un espacio, el modelo no sabe si existe un río que parte en dos el área lo que limita la capacidad de colonización de la especie. Claro está que, por eso, los modelos de distribución de especie evalúan la distribución potencial de la especie, conceptualmente muy diferente al nicho realizado. Sin embargo, el presente estudio busca darle una solución a esta gran problemática y es añadirle un paso a la evaluación de la pertinencia y efectividad de los resultados de los modelos de distribución de especie, en este caso usando como organismo de estudio la especie Ischnuracapreolus, una libélula encontrada en Colombia.
14
1.2 Objetivos
1.2.1 General
Evaluar cinco modelos de distribución de especies de la especie Ischnuracapreolus a partir de curvas de AUC, factores ecológicos y comportamentales, en el territorio colombiano
1.2.2 Específicos
o Describir las condiciones ambientales de la especie Ischnuracapreolus que determina nsu distribución en Colombia
o Predecir la distribución potencial de la especie I. capreolus, para el territorio colombiano.
o Proponer una nueva metodología para evaluar los modelos de distribución de especies incorporando datos ecológicos y comportamentales de la especie objeto de estudio, en Colombia.
1.3 Preguntas de investigación
¿Cuáles son las características ambientales que permiten a la especie I. capreolus potencialmente habitar Colombia?
¿En qué medida lo modelos de distribución de especies permiten aumentar el conocimiento sobre la distribución potencial de la especie I.capreolus en Colombia?
¿Qué diferencias se encuentran frente a la evaluación del modelo a partir de las curvas AUC y la información ecológica y comportamental de la especie I.capreolus?
1.4 Hipótesis
A partir de la información recolectada de la distribución y biología de Ischnuracapreolus en Colombia, es posible poner a prueba los modelos tradicionales de especies a través de la incorporación de un análisis de conectividad ecológica.
1.5 Justificación
El esfuerzo de muestreo para muchas especies de insectos en la mayoría de los casos se ve afectada por factores difíciles de determinar, cómo el clima, el grado de antropización, la época hidrológica, entre otros. Esto hace que encontrar la especie de interés resulte
15
similar a un evento estocástico, que se resume en términos coloquiales a la suerte. Limitando muchas veces la compresión de la ecología y la capacidad de dispersión de muchos grupos, teniendo que asumir ciertos aspectos de su biología.
La modelación de la distribución potencial de las especies es una alternativa matemática, que permite determinar la distribución de una especie según ciertos atributos ecológicos. La ventaja de esto es que se evalúa con un mínimo conocimiento previo, y sin la necesidad de ir a campo. Esto dado que el modelo ayudará a determinar con criterios objetivos los sitios que la especie de interés podría llegar a ocupar. Esto permitiría llegar a entender su distribución y evaluar su fragilidad en un periodo de cambio climático y degradación del ecosistema.
Desafortunadamente, esto ofrece una solución parcial, ya que un ecosistema presenta una inmensa heterogeneidad de microclimas, lo que afecta la distribución y desplazamiento de las especies. Por esto, los modelos de distribución de especie han sido ampliamente criticados, al no tener en cuenta aspectos comportamentales. Lo que ha hecho que caiga en desuso esta herramienta, por eso se plantea proponer una nueva metodología que permita seguir usando estas herramientas para establecer rutas potenciales de conservación o zonas de muestreo para censos poblacionales. Esto a partir de una especie con amplia distribución como Ischnuracapreolus y un país con una inmensa variedad de hábitat como es Colombia.
1.6 Alcance
El presente estudio busca evaluar una nueva metodología para evaluar cinco modelos de distribución potencial de la especie de libélula Ischnuracapreolus en Colombia. Sin embargo, no se pretende evaluar el modelo únicamente desde el punto de vista de efectividad del modelo o probabilidad de presencia; se buscará establecer la correspondencia a partir de un análisis de conectividad y dispersión de la especie. Esto quiere decir que se hará una validación ecológica para determinar si la especie es capaz de llegar a las regiones que el modelo propone.
En las ciencias naturales la premisa de aumentar el conocimiento de los animales sobre la tierra es una motivación constante. Adicionalmente, poner a prueba la efectividad de esta herramienta, en una especie ampliamente conocida, permitirá determinar su efectividad y de esta manera replicarlo en grupos biológicos cuya biología no sea muy clara ni estudiada
16
y así aportar herramientas para describir su distribución y potenciales estrategias de conservación.
El presente trabajo pretende mostrar una metodología complementaria a lo trabajado de manera tradicional. Este se está basando únicamente, en las herramientas estadísticas, las cuales determinan el nivel de similitud de los píxeles y según el modelo incorporar como variable la capacidad de desplazamiento de la especie de interés. Con esto se plantea evaluar el área potencial de distribución y determinar si el espacio modelado, construye un corredor viable para la especie, esto incorporando variables adicionales, como lo son aspectos ecológicos esenciales para el establecimiento de nicho de la especie. Siendo un resultado esperado si el modelo es preciso, pero desde otra perspectiva
17
2 REVISIÓN DE LITERATURA
Se presenta a continuación, los resultados de una revisión bibliográfica construida en función de las preguntas de investigación, anteriormente presentada.
2.1 Grupo de estudio
Los insectos son uno de los taxa que mayor éxito evolutivo han tenido a lo largo de la historia del planeta (Okada et al., 2012). Determinado principalmente por su abundancia en términos de biomasa, y número de especies, además, por la variedad morfológica, ecológica y comportamental, llevándolos a colonizar múltiples hábitats (Bennett, 2010) Adicionalmente, son modelos biológicos1 pertinentes para diversas áreas del conocimiento, dado que están involucrados en los procesos más relevantes sobre la tierra, desde el reciclado de nutrientes, control de grandes vertebrados, vectores, entre otros (Vet y Smid, 2006) Tal ha sido la fama o el repudio de los insectos que muchos avances tecnológicos y militares han girado en torno al problema que atraen los insectos, como las enfermedades que se esparcen, siendo muchas de ellas incurables hasta la fecha (Smith y Kennedy, 2009). Este tradicional miedo ha causado que la fama de los insectos se vea afectada y la cacería hacia ellos sea indiscriminada, por eso pocos estudios o poca publicidad se les hace. Sin embargo, estas características no los ha dejado fuera del declive que está sufriendo el planeta en el actual Antropoceno, alterando las poblaciones de insectos, lo cual afectará servicios ecosistémicos esenciales para el ser humano (entre ellos la polinización y el control biológico) (Keys et al., 2019; Stromberg, s. f.)
Uno de los retos más grandes que se tiene en el estudio de insectos es determinar sus rangos de distribución y nivel de riesgo de conservación (Austin, 2007; Pearson y Dawson, 2003). Debido a la complejidad de los muestreos y un complejo ciclo de vida, en términos de duración y fases que estos tienen, siendo capaces de cambiar totalmente su morfología y medio en el que habitan. Sin embargo, entender y caracterizar este proceso permite entender su rol ecológico y los cambios que un organismo (en general, no sólo los insectos) puede tener a lo largo de su crecimiento y desarrollo, siendo de gran relevancia especialmente en temas de conservación de especies amenazadas, cambio climático, control de enfermedades y patógenos (Ellis y Pfeiffer, 1990; Imai y Yamaguchi, 2012;
1 Modelo biológico: organismos con características genéticas y fisiológicas de los humanos, para así probar de manera previa el potencial médico o de estudio previo a la prueba humana.
18
Paillard, 2016; Walker et al., s. f.) Y es que, si se habla de conservación, los insectos son la base ecosistémica de la mayor parte de los hábitats del mundo, ofrecen la característica de ser consumidores primarios, secundarios, depredadores tope, detritívoros y descomponedores, es decir, en el flujo de energía están involucrados en todo proceso, y por la biomasa que representan (aunque invisible) no puede ser despreciada su importancia (Slobodkin, 1962). Y más ahora que en Colombia, un territorio con gran capacidad de carga para la productividad primaria es víctima de un mal uso del suelo, lo que requiere que para su restauración ecológica mejore la calidad del suelo, y con esto, los insectos, bacterias y hongos, tienen el rol principal (Barrera-Cataño et al., s. f.)
Hablar de insectos es tener que hablar de 32 ordenes diferentes, sin embargo, el presente trabajo, se enfoca en un grupo en especial. Dentro de los insectos, las libélulas, por ejemplo, poseen un ciclo de vida con gran complejidad, ya que a lo largo de su desarrollo deben enfrentarse a dos medios diferentes, acuático y terrestre (Rowe y Ludwig, 1991). Y es debido a su estrecha relación con los diferentes medios lo que ha permitido que sean considerados como bioindicadores, ayudando a determinar el impacto antropogénico empleando su estadio acuático y terrestre, para así establecer zonas potenciales para conservar (Collins y McIntyre, 2015)
El orden Odonata (libélulas y caballitos del diablo) es un clado de insectos alados, caracterizados por no plegar sus alas (Paleoptera) (Borror et al., 1992), su origen se estima hace alrededor de 300 millones de años (MA) durante el periodo Carbonífero (Misof et al., 2014). Este grupo presenta metamorfosis incompleta (es decir, hemimetábola), donde morfológica y estructuralmente la ninfa acuática (náyade) se diferencia del adulto volador. Este orden agrupa libélulas y caballitos del diablo, distribuidos en los subórdenes Anisoptera y Zygoptera respectivamente y un grupo con distribución restringida Anisozygoptera. Los anisópteros se caracterizan por ser robustos y con alas grandes, diferentes entre anteriores y posteriores en tamaño y forma, sus náyades poseen respiración mediante cámara rectal; mientras que los zigópteros son delgados con alas del mismo tamaño, y las náyades poseen lamelas caudales para el intercambio gaseoso (McCafferty, 1983)
Uno de los retos a los que se enfrentan las especies de odonatos, asociados a aguas estancadas es a la pérdida de hábitat. Al estar en zonas estancadas o áreas inundables, para los humanos es un problema (Cardozo et al., 2016). Esto, dado que, en estas zonas, o no es posible cultivar, o el ganado se ahoga en periodos de grandes lluvias, por esto, el territorio se transforma con el objetivo que estas zonas sean drenadas fácilmente y no se
19
acumulen aguas, dejando en riesgo a estas especies. Muchas logran asociarse a estanques artificiales, ya sea decorativos o utilizados como zoo-criaderos de peces y patos. Sin embargo, no hay estudios poblacionales o de censo de este hábitat, ni una evaluación rigurosa del territorio colombiano (Bota-Sierra et al., 2019). Esto no quiere decir que no se estudie la fragmentación en Colombia, sino que esta está enfocada a especies sombrilla o emblemática como las aves o primates, los cuales tienen como hábitat principal los bosques, o coberturas donde los árboles priman (Bennett, 2010; Polhemus, 1993) Esto ha hecho que ecosistemas de aguas estancadas queden rezagados de los estudios, y por ende no se tenga conocimiento de la vulnerabilidad de las especies que allí habitan, siendo un ejemplo la especie I.capreolus.
Uno de los retos más grande que presentan las libélulas de aguas estancadas como se mencionó es la falta de información sobre la pérdida de hábitat, lo que resulta ser un reto, al no ser un foco de estudio. Según la Figura 3, se evidencia como el desequilibrio entre los factores bióticos y abióticos altera la separación de especies, y si permite el sobrelapamiento de nicho, entonces aumentará la competencia interespecífica, y por ende la intraespecífica, llevando a un efecto mariposa (Sánchez-Guillén et al., 2022). Esto significa que los recursos no serán suficientes y el espacio no será suficiente para todas las especies, aumentando la vulnerabilidad y posible extinción (Chichorro et al., 2019; Mace et al., 2008). Esto terminará afectando la capacidad de carga y resiliencia del ecosistema, ya que no tendrá especies que permitan su restauración natural, ante cualquier perturbación, natural o antrópica (Neuhoff y Helm, 2016)
Focalizando el estudio, se describirá la especie objeto de interés. El Género Ischnura, cuya distribución es cosmopolita y cuenta con aproximadamente 70 especies a nivel mundial;
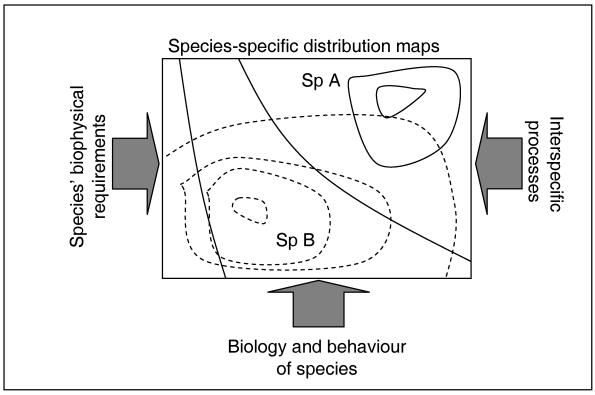
20
Figura 3 Interacción nicho, visto como recursos y distribución geográfica con dos especies diferentes (Fischer y Lindenmayer, 2007).
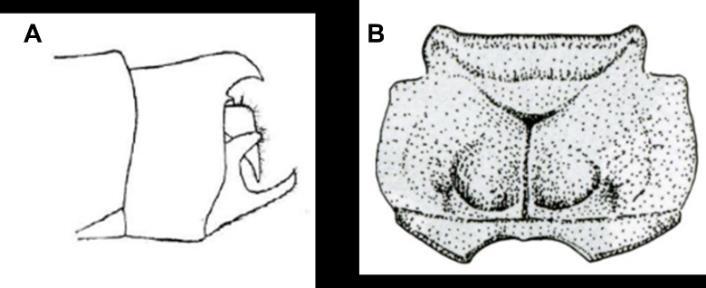
para Colombia, hay 8 especies reportadas en un amplio rango altitudinal (Pérez-Gutiérrez y Palacino-Rodríguez, 2011). El presente trabajo se concentra en la especie Ischnuracapreolus Presenta amplia distribución en Sudamérica (Heckman, 2006; Paulson et al., 2016), sin embargo, poco se sabe sobre su ecología y ciclo de vida. Se puededistinguir esta especie por las dos protuberancias que presenta la hembra en su pronoto, cercos grandesy el ovopositor termina en punta, y el macho posee los paraproctos en forma de “C” (véase la Figura 4). El Género Ischnura se encuentra en aguas estancadas como lagos, estanques y pantanos; siempre asociadas a los pastos, presentes en la región emergente de la zona litoral (Ward, 1992). Los adultos se encuentran asociados a la misma vegetación que el estadio inmaduro, y su tiempo de vida puede variar desde un par de semanas a un par de meses (McCafferty, 1983). En general el adulto no se encuentra muy alejado del agua, por esto se considera que su dispersión no es buena. Por otra parte, las hembras del presentan polimorfismo de color, característica ampliamente estudiada especialmente para el grupo (Cordero-Rivera y Pérez, 1998; Cordero-Rivera y Andrés, 2002; Sánchez-Guillén et al., 2011; Sanmartín-Villar et al., 2016).
Figura 4 Se muestra en A la estructura en forma de ‘C’ de los paraproctos del macho y en B los tubérculos del pronto de la hembra (modificado de Garrison et al. 2010)
2.2 Modelo de distribución de especie
Los modelos de distribución de especies son la representación espacial para las zonas que cumplen ciertos requerimientos de la especie para poder establecerse, basado en información ambiental relevante (Mateo et al., 2011). Esto indica que, a partir de herramientas computacionales, es posible predecir la distribución de una especie, lo que permite caracterizar y entender los hábitats de las especies, y lograr desglosar la heterogeneidad de microclimas que posee un territorio, bioma, o incluso un ecosistema. Antes de empezar a hablar del uso o las características de los modelos de especies, es necesario comprender dos conceptos esenciales en ecología:
21
Nicho Fundamental: Es aquel espacio geográfico que cumple los requerimientos ambientales y bióticos necesarios para el establecimiento de una especie, es decir, es la zona donde potencialmente podría estar la especie de interés (Begon et al., 2006). Este tipo de nicho es el resultadok de un modelo de distribución de especies.
Nicho Realizado: Se le denomina al espacio geográfico en el cual se reporta y observa la especie de interés. Es el insumo para caracterizar los requerimientos de una especie y de esta manera establecer el nicho potencial o fundamental (Begon et al., 2006) Es decir, que es definido por la información de presencia, obtenido en campo o de fuente de datos secundarias.
Esto indica que un territorio debe contar con la caracterización ambiental de este Si bien no hay una regla que indica la información que se debe tener disponible, es importante, que logre dar cuenta de las condiciones ambientales del territorio en función de la necesidad de la especie objeto de estudio (Barve et al., 2011). Claro está, que hay organizaciones que han venido publicando esta información a escala global, pero es importante tener en cuenta qué tipo de información ofrecen y su relevancia en el estudio (Nezer et al., 2017). Para Colombia, por ejemplo, hay una gran cantidad de información disponible, que han venido publicando organizaciones gubernamentales como el Instituto Geográfico Agustín Codazzi – IGAC2 y el Instituto de Hidrología, Meteorología y Estudios Ambientales – IDEAM3 y el Instituto de Investigaciones Biológicas Alexander von Humboldt4, sin embargo, en esta última la información no siempre se encuentra de manera libre para usuarios. Pero a nivel internacional se cuenta con información ambiental global, como WorldClim, que de entrada ofrece cerca de 19 variables ambientales, las cuales deben ser usadas con mucha precaución para modelos de distribución de especies. Esto dado que estas variables construidas por la organización son extrapoladas y en una gran medida son colineales, es decir, que tienen un comportamiento lineal entre las mismas variables, es decir, son redundantes, dicen la misma información del ecosistema, por lo tanto, incorporando variables con este comportamiento, el modelo de vuelve redundante y pierde precisión. Claro está que WorldClim ofrece información de gran utilidad para la investigación, especialmente en la actualidad que surgen más y más estudios que tienen como objetivo estudiar el cambio climático y evaluar el riesgo de los ecosistemas y con esto las especies que lo habitan. Ya que ofrece proyecciones climáticas, de hasta 100 años
2 Página de acceso a la entidad del (IGAC): https://www.igac.gov.co
3 Página de acceso a la entidad (IDEAM): http://www.ideam.gov.co
4 Página de acceso a la entidad (Instituto Humboldt): http://www.humboldt.org.co/es/
22
lo que facilita evaluar aspectos relevantes de los territorios, condiciones y riesgos (Booth, 2018).
Por otro lado, es importante tener claro el objetivo del proyecto de investigación, esto ya que este definirá el área evaluada para la distribución potencial de la especie. Y con esto se puede definir el uso o aplicación de esta metodología. Por ejemplo, en el conocimiento de la distribución potencial de una especie, con el objetivo de conocer su biología, aumentar el conocimiento, entender su riesgo, fortalecer la conectividad o incluso complementar alguna estrategia de conservación. También permite delimitar y establecer límites de ecosistemas frágiles o en riesgo, y así determinar la vulnerabilidad de las especies ante el cambio climático y establecer rutas de manejo o introducción controlada de especies con potencial ecosistémico (Dormann et al., 2012; Gallien, 2012).
Sin embargo, el área de estudio no responde únicamente a la necesidad del investigador o de una población. Esto dado que debe responder de igual manera a la representatividad espacial que se tenga de una especie, es decir, la distribución conocida de esta. No será posible, por ejemplo, estudiar la distribución de una especie en Colombia, cuando se tienen estudios enfocados únicamente en los llanos o en la Amazonía, ya que son territorios con un nivel de especificidad muy alto y no tendría sentido ese esfuerzo computacional. Se podrá aplicar o realizar un modelamiento de nicho a aquellas especies que no cuenten con un sesgo en sus datos, porque obviar esto lleva a que la información del modelo no sea realista (Guisan y Thuiller, 2005; Wiens et al., 2009)
Los estudios que buscan establecer el nicho fundamental de una especie son tan amplios y usados que han hecho que desarrollen diferentes tipos de modelos biológicos. Por esto, es importante mencionar que no es posible responder a la pregunta ¿cuál es el mejor modelo?, la respuesta dependerá de la información existente, los datos, las variables, y posiblemente la respuesta sea vaga, por eso es importante buscar diversas estrategias de evaluación (Brodie et al., 2020).
BIOCLIM: Modelo de similitud climática, este modelo determina si las condiciones climáticas son favorables teniendo en cuenta la distancia que hay entre los datos de presencia.
GAM (Modelo Aditivo Generalizado) y GLM (Modelo Lineal Generalizado): Son modelos lineales no paramétricos, es decir, asumen que las variables tienen una relación lineal entre ellas. Su limitación es que no se ve afectada directamente por la distancia inicial generada por los reportes de la especie de interés.
23
Maxent (Máxima Entropía): Este modelo está basado en máxima entropía, usando la información ambiental, es decir, que les da pesos diferenciales a las variables ambientales para describir la distribución ambiental de la especie de interés.
RPART (Árboles de Decisión): Está basado en eventos logísticos de probabilidad, construyendo árboles de decisión para determinar la distribución potencial de una especie.
Puede que cada uno de estos modelos tenga premisas diferentes y aspectos a favor y en contra. Sin embargo, en los artículos que referencian el uso de modelos de distribución de especies, usualmente no manejan un único modelo, presentan un grupo de modelos, a fin con la información existente y presentan los resultados respetando aquel que mayor precisión llegó a tener con la especie (Elith y Graham, 2009)
Si bien el modelo implementado es un aspecto con una gran relevancia en un estudio de distribución potencial, el esquema de trabajo es el mismo en todos los aspectos (Wunderlich et al., 2022). Siendo de los más relevantes los datos climáticos, por eso se hace énfasis a la calidad y escala obtenida, ya que finalmente este es el insumo para describir las condiciones ambientales aptas para los organismos. Siendo los datos de presencia los que determinen si los resultados son sesgados o no, en función de la representatividad espacial de la información
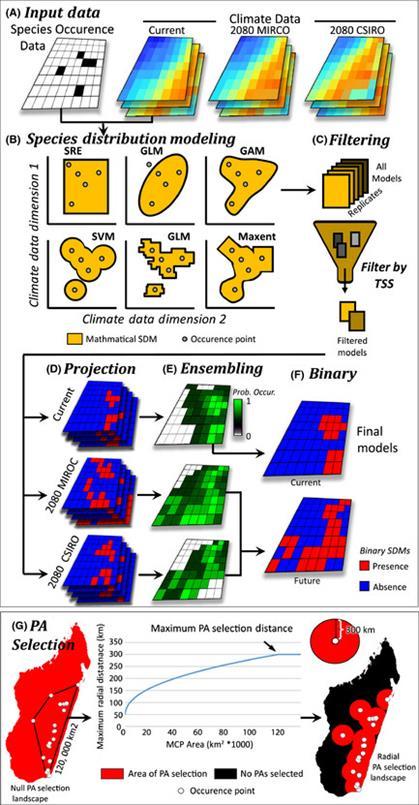
(Araújo y Guisan, 2006). En la Figura 5 se muestra el esquema fundamental de trabajo en modelos de distribución de especies.
Finalmente, a continuación, se presenta el potencial uso y aplicación que tiene los modelos de distribución de especies en insectos.

24
Figura 5 Flujo de trabajo de un MDE1 (J. Brown y Yoder, 2015)
2.3 Convergencia insectos y MDE
Los insectos, día a día han mostrado tener un rol ecológico de gran relevancia, característica que por falta de conocimiento ha hecho que exista un gran desconocimiento asociado, con repercusiones desde mal manejo de los recursos hasta extinciones locales. Sin embargo, pese a que existe una motivación en términos agronómicos (productivos) y naturalistas (aumento del conocimiento) para caracterizar las poblaciones de insectos en campo, es un reto. Siendo un problema encontrar la distribución real de una especie, dado los altos costos de las actividades de campo; se vuelve significativo implementar herramientas computacionales. En este caso específico, a partir de una herramienta de SIG, la cual permite entender y caracterizar el territorio a partir de información cartográfica existente. Facilitada por no requerir un conocimiento amplio o extenso del territorio, pero sí de la especie, adicionalmente contar con información base, o variables ambientales que permitan describir y generar modelos probabilísticos de distribución.
Los Sistemas de Información Geográfica han articulado la estadística con la información cartográfica, de una manera muy armónica, partiendo del hecho que los animales son reflejo de su ambiente. Siendo entonces, la herramienta idónea para entender el ecosistema, la interacción de los taxones con este y lo más importante permitir responder preguntas biológicas a partir del conocimiento espacial o geoestadística. Particularmente, la especie Ischnuracapreolus, este es el primer acercamiento hacia entender su distribución, pese a que no es el único estudio de libélulas como bien los detallan Collins y McIntyre (2015)
Por esto, el entendimiento de la distribución de especies es una herramienta que permite caracterizar y entender el territorio a partir de los taxones vulnerables y/o generalistas, describiendo así los cambios (o posibles cambios) generados por la pérdida de hábitat y el cambio climático. Esto, gracias a que con el conocimiento acerca de la ecología de un taxa se puede modelar las regiones en las que podría encontrarse este grupo, de esta manera aumentar el conocimiento sobre la distribución de las especies y entender los patrones ecológicos de estas, sin tener que dedicar grandes jornadas en campo (Segurado y Araujo 2004)
Si bien, a priori luce como una herramienta muy útil, tiene una limitante, que altera el principio de falsabilidad en cualquier estudio. Su relación sobre la distribución de la especie de interés con las variables ambientales en principio es netamente matemática, es decir, basado en un principio de similitud, sin correlación detallada sobre la capacidad de
25
dispersión del organismo y el hábitat proyectado. Sin embargo, entender los rangos de movimiento de cualquier especie es bastante complejo, especialmente de los insectos, ya que no pueden cargar pesados GPS, como algunos grupos de vertebrados (Granberg et al., 2008). Por esto, el presente estudio busca calibrar el modelo a partir de un análisis de conectividad ecológica, determinando a partir de la biología del grupo y su distribución actual, las potenciales zonas a las que realmente no puede llegar, siendo estas ausencias reales. Aclarando que se mantiene el sesgo que sólo se puede trabajar con la información de distribución disponible y pueda existir errores en la identificación de los individuos o errores en la preparación de la información disponible en las plataformas consultadas (Lobo, 2016; Newbold, 2010)
El estudio de libélulas ha permitido entender la transformación que ha sufrido un ecosistema, sin importar si el medio es terrestre o acuático. Esto, aprovechando que estos insectos presentan una estrecha relación con ambos medios para su desarrollo, siendo los candidatos idóneos para rastrear el grado de antropización en un territorio. Adicionalmente, su heterogénea biología permite cuantificar el grado de afectación dependiendo del ensamblaje de la zona de estudio, dado que ciertos taxa requieren de condiciones específicas en cuanto a la calidad de agua y densidad vegetal (Hardersen, 2000)
Por ejemplo, los modelos de distribución de especies para libélulas han permitido establecer los efectos que tendrá el cambio climático en la diversidad de libélulas del género Ischnura. Estas, al ser un organismo con una importante dependencia a las condiciones climáticas y baja capacidad de dispersión, la pérdida del hábitat ya sea por transformación y degradación o por cambios climáticos, las pone en una situación de vulnerabilidad ante la extinción. Esto, no sólo por el aumento de la competencia interespecífica o intraespecífica por territorio y recursos sino porque aumenta la probabilidad de hibridación en especies del género, esto dado que en el mundo se han reportado múltiples casos de hibridación en este género, incluida Colombia (SánchezGuillén et al., 2016; Galindo-Ruiz et al., 2019). Esto llevaría a una pérdida de la diversidad taxonómica, aunque no se ha evaluado los efectos que tendría sobre la diversidad genética, ya que se ha comprobado que los híbridos en casos específicos no son necesariamente estériles (Sánchez-Guillén et al., 2016)
Adicionalmente, en odonatos, se ha evidenciado la implementación de los modelos de distribución de especies para evaluar nuevos sitios potenciales de distribución. Esto para grupos que se encuentran con pocos registros, y se busca aumentar el conocimiento sobre su distribución (Finch et al., 2006). Sin embargo, estos responden a las necesidades
26
convencionales de los modelos de distribución de especies, es decir son puestos a prueba de la manera tradicional. Suelen usar las curvas AUC y la bondad de ajuste de modelo de paramétricos con el Criterio de Información de Akaike (AIC, por sus siglas en ingles Akaike InformationCriteria).
Por otro lado, el aumento considerable de la fragmentación de hábitat naturales por el emplazamiento de cultivos y ciudades tiene importantes repercusiones en las poblaciones naturales. En insectos, por ejemplo, esta problemática se ve afectada de dos maneras, para insectos especialistas hay una disminución de hábitats por lo tanto un aumento en la competencia, mientras que, en grupos generalistas, hay fragmentación de hábitat, lo que lleva a la disminución de la diversidad genética (Diekötter et al., 2008). Sin embargo, con la implementación de los estudios de conectividad, se ha evidenciado que, dependiendo del tipo de cultivo, la fragmentación se puede vencer. Por ejemplo, si es un monocultivo, aumenta la fragmentación y disminuye la diversidad taxonómica, es decir el número de especies disminuye, contrario en policultivos, donde la diversidad vegetal es mayor, entonces la riqueza de especies aumenta, disminuyendo la fragmentación (Streib et al., 2020)
Algunos conceptos que no están considerados en los modelos de distribución de especie son:
Conectividad Ecológica: Es la capacidad que tiene un organismo para desplazarse de una zona a otra, ese corredor debe favorecer el movimiento, es decir cumplir con algunas de las características ambientales y biológicas de la especies para que se dé (Valencia, 2020)
Ecología Funcional: Rama de la biología que busca entender de manera amplia las características de una especie/ población/ comunidad, para su correcta función ecosistémica. La definición no se limita a las condiciones ambientales, sino a características de interacciones comunitarias y flujo de energía (Calow, 1987)
Estos conceptos permiten y dan luz de una profunda caracterización de la biología de las especies, entendiendo los retos que tienen en el ecosistema. Ya que no se trata únicamente de explorar qué tan similares o diferentes son los territorios asociados en el área de estudio, sino que tan heterogéneo resulta el ecosistema para una especie y por ende qué tan permeables son algunas capas para que esta se pueda desplazar con total facilidad. Tener en cuenta únicamente el clima sería un error, ya que algunas especies no pueden habitar pastos, prefiriendo bosques o viceversa.
27
3 METODOLOGÍA
3.1 Zona de estudio
La zona de estudio para el modelo de distribución de especie será Colombia (véase la Figura 6), esto dado que la información ambiental base para los análisis son creados en este país, de esta manera se asegura la integridad y homogeneidad en la construcción de los modelos y matrices usadas en el estudio.
Colombia por su parte es considerado uno de los países con mayor diversidad de especies, siendo conocido por su diversidad de aves, plantas y anfibios (Bussmann et al., 2018)
Claro está, que la antropización descontrolada ha causado unas altas de extinción local (Solari et al., 2013). Por esto, entender y caracterizar el territorio, no sólo en términos de la diversidad que alberga sino la vulnerabilidad de sus ecosistemas toma relevancia en un contexto de cambio climático (Agudelo, 2016) Adicionalmente, la heterogeneidad del territorio en cuanto a su orogenia hace a Colombia, un país valioso para poner a prueba la metodología presentada en el presente estudio. Esto es el resultado de una gran variedad de formas montañosas y la influencia de los vientos producto de su cercanía al océano pacífico y atlántico.
28
Figura 6 Colombia, delimitada como área de estudio para la presente investigación.
3.2 Obtención de las capas ambientales
El presente trabajo se concentra en determinar la distribución potencial de la especie Ischnuracapreolus, a partir de la unión de herramientas computacionales cuyo enfoque es netamente estadístico y otro biológico. Esto con el objetivo de desarrollar un flujo de trabajo que permita poner un contexto evolutivo y ecosistémico a un modelo de distribución de especies tradicional, e involucrar el concepto matemático con el biológico, de esta manera evaluar los resultados obtenidos de modelos de distribución de especies, con mayor precisión y exactitud. El área de estudio se restringió a Colombia, con el objetivo de ponerlo a prueba en un país con una amplia heterogeneidad de ecosistemas y accidentes geográficos.
Para el análisis se descargaron capas ambientales en formato ráster de la plataforma CliMond (Kriticos et al., 2012), siendo priorizadas aquellas con mayor relevancia ecológica para entender y predecir la distribución potencial de insectos (Collins y McIntyre, 2015)
Las capas contienen información a escala global, por esta razón fueron previamente editadas para restringir el análisis al área de estudio de interés Esta edición se realizó con el software QGIS (QGIS Development Team, 2009).
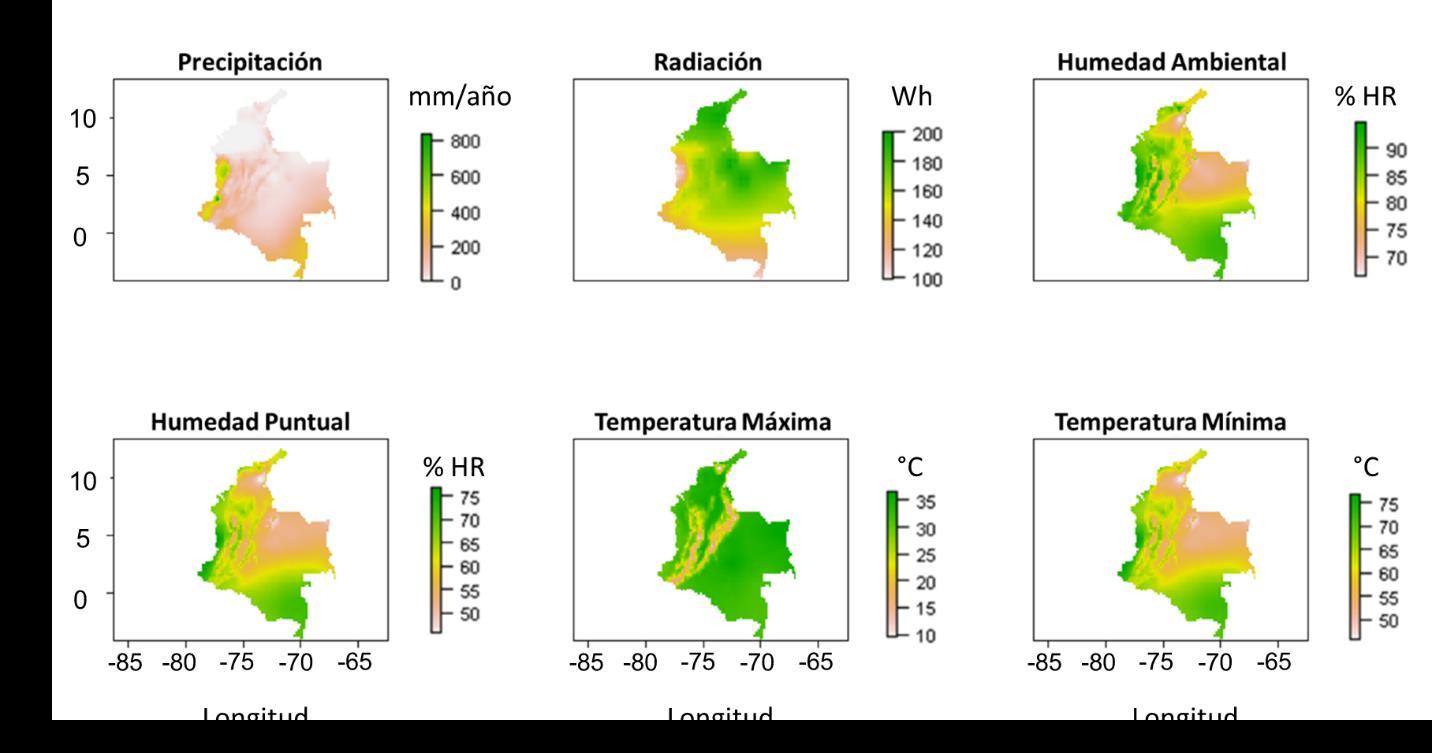
Las capas implementadas con mayor representatividad fueron: precipitación (Prec: mm/año), radiación (Rad: Wh), humedad relativa del aire (Rham: %HR), humedad relativa puntual (RHmp: %HR), temperatura máxima (Tmax: °C) y temperatura mínima (Tmin:°C). Estas capas fueron seleccionadas dado que reflejan las condiciones esenciales de un animal ectotermo; siendo variables que explican de manera adecuada el ambiente que permitiría y daría viabilidad a la colonización o establecimiento del hábitat por parte de Ischnuracapreolus. En la Figura 7, se muestran las capas ajustadas al área de interés Colombia.
29
3.2.1 Análisis de correlación
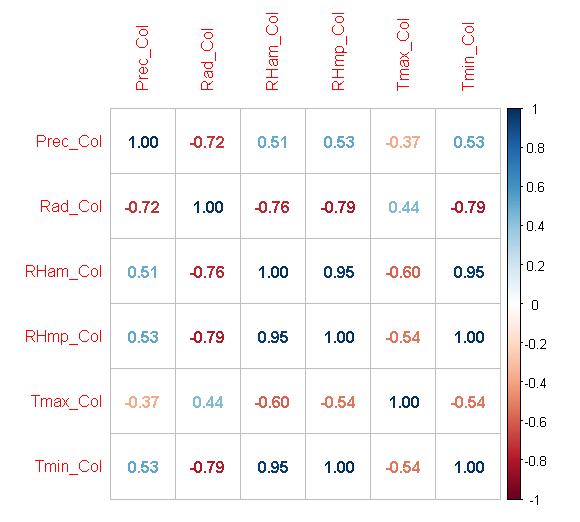
Posteriormente se validó el nivel de correlación que tienen las variables entre sí, con el objetivo de minimizar la redundancia del modelo y así incluir información relevante e informativa. Los resultados se muestran en la Figura 8, se consideró un R2 de ±0.82, como una fuerte correlación y los demás valor como una baja o con poco poder en términos de correlación (Levine y Stephan, 2010). De manera general, se evidencia una baja correlación de las variables indicando una baja redundancia en las variables de interés, excepto para las variables de humedad relativa (ambiental y puntual), temperatura mínima y máxima. Sin embargo, se decide conservar estas variables pese a la correlación, ya que estas describen de manera diferencial las condiciones ambientales, e involucran diferentes perspectivas las condiciones ambientales en términos ecológicos y funcionales de los insectos (Sofaer et al., 2019)
30
Figura 7 Visualización preliminar de las capas ambientales implementadas usadas para la formulación de los modelos de distribución de la especie de interés.
3.3 Datos de presencia
Posteriormente se recopiló la información de presencia de la especie Ischnuracapreolus, con el objetivo de minimizar sesgos y errores en el modelo por errores en la determinación de la especie publicados en plataformas digitales, se usaron datos de presencia de las especies almacenadas en el Museo CJ Marinkelle de la Universidad de los Andes, localizado en la ciudad de Bogotá-Colombia; los cuales fueron previamente validados y confirmados como la especie de interés Posteriormente, se compiló en una base de datos la información de colecta, la cual se validó que las coordenadas correspondieran espacialmente a lo consignado en la etiqueta de la especie.
3.4 Construcción del modelo de distribución de especies
Con esta información recopilada se procedió a desarrollar múltiples modelos de distribución de la especie Ischnuracapreolus a través del paquete Species Distribution Modelling –SDM (Naimi y Araújo, 2020), una extensión del software estadístico R que permite obtener la información de las variables más relevantes en el estudio, caracterizar según el modelo las condiciones idóneas para la distribución de la especie y la efectividad del modelo para predecir la distribución potencial de esta. Finalmente, los gráficos obtenidos fueron
31
Figura 8 Análisis de correlación entre las capas implementadas para los modelos de distribución de la especie.
construidos mediante el paquete SDM y ggplot2 (Wickham, 2016), y con Quantum GIS –QGIS (QGIS Development Team, 2009). El esquema general realizado se presenta en la Figura 9 Esquema de trabajo para la obtención de los modelos de distribución potencial de Ischnuracapreolus.
Intalación de los paquetes en R •install.packages()
Validación de la instalación y llamado del paquete •library()
Importación de ráster y datos de presencia
•raster = raster('path')
•presencia = read.csv(file.choose(), stringsAsFactors=FALS E)
Construcción del modelo de distribución
•sdmData(Especie~., predictors = all_raster, bg = list(n=100))
•Matriz de importancia
3.5 Propuesta metodológica
Generación de resultados
•ROC/AUC
•Raster de dsitribución potencial
El presente estudio busca combinar dos herramientas de trabajo con el objetivo de encontrar modelos de distribución más precisos y por lo tanto más realistas. Como se presentó anteriormente, los modelos de distribución de especies permiten determinar a través de variables continuas la similitud de un territorio, y, por ende, la capacidad de ser colonizada por la especie de interés. Sin embargo, esta información no describe la complejidad de un territorio, y para controlar una de las limitantes, se busca implementar modelos de distribución de especies. Esta herramienta es usada para entender la capacidad que tiene una especie en ambientes fragmentados, ahora será usada para determinar si un área predicha (modelada) es apta para que la especie de interés logre
32
Figura 9 Esquema de trabajo para la obtención de los modelos de distribución potencial de Ischnuracapreolus.
moverse, y llegar hasta donde el modelo de distribución indica. El principio es el mismo, sólo que esta vez el área será un territorio modelado mas no real.
Posteriormente se estableció una nueva ruta metodológica para determinar la efectividad de los modelos ecológicos a partir de información ecológica que no puede ser contemplada en los modelos iniciales al ser variables del tipo categórico y no continuo (numérico) Para esto, fue necesario realizar una búsqueda de información ambiental, complementaria a los factores abióticos, implementados en el modelo anterior para validar que las condiciones ecosistémicas cumplan los requerimientos para el desplazamiento de las especies, en las zonas de distribución potencial y de esta manera establecer si en el polígono resultado la especie tenía facilidad de movilizarse o no. Para esto se hizo una revisión de la información existente en los sistemas de geo-servicios del país, en este caso el IGAC y el IDEAM. Viniendo la totalidad de la información en materia cartográfica de un único punto, facilita la implementación y uso, dado que los métodos de muestreo, tabulación y consolidación de la información cartográfica es constante, facilitando la manipulación de esta. De lo contrario sería necesario una normalización o procesamiento diferencial de los datos, lo que dificultaría la implementación de la información Adicionalmente, al ser una organización gubernamental se asegura que se implementó la misma metodología para la captura y consolidación de los shapefiles. La metodología implementada para hacer este análisis fue la presentada por McRae et al. (2016), quienes presentan la herramienta Circuitscape, la cual puede ser integrada o no con ArcGIS o QGIS, para el presente estudio se vinculó con QGIS.
3.5.1 Análisis de Resistencia
De estas fuentes se usaron las capas ambientales más actualizadas (2019) y aquellas que lograran definir y condicionar los factores ambientales que describen el hábitat de la especie de interés. Es importante tener en cuenta que los odonatos tienen ciclo de vida con fase acuático y terrestre, por esto se deben tener en cuenta ambos tipos de información cartográfica. Para el presente estudio se usó la información hidrológica, para la fase acuática de náyade, esto dado que Ischnuracapreolus tiene una preferencia por los cuerpos de agua lénticos, como charcas temporales o humedales, mientras que cuerpos de agua como ríos o quebradas de nivel 3 o superior funcionan como barrera ecológica para la especie. Por otro lado, la cobertura vegetal del territorio describe de igual manera las preferencias, siendo los adultos voladores. Esto hace que la caracterización sea muy compleja por su doble forma de vida. A continuación, en la Tabla 1 se presentan las
33
variables categóricas descartadas de bases de datos oficiales colombiana. En estas se describen las coberturas y cuerpos de agua asociados al territorio objeto de estudio.
Tabla 1 Variables categóricas involucradas en el análisis ecológico y de resistencia de la especie I. capreolus
Macrovariable Variable
Aguas continentales Ríos (50 m)
Áreas abiertas, sin o con poca vegetación
Áreas agrícolas heterogéneas
Tierras desnudas y degradadas
Mosaico de cultivos y espacios naturales
Mosaico de cultivos, pastos y espacios naturales
Mosaico de pastos con espacios naturales
Mosaico de pastos y cultivos
Áreas con vegetación herbácea y/o arbustiva Vegetación secundaria o en transición
Bosques
Pastos
Zonas industriales o comerciales y redes de comunicación
Zonas urbanizadas
Zonas verdes artificializadas, no agrícolas
Bosque denso
Plantación forestal
Pastos arbolados
Pastos enmalezados
Pastos limpios
Red vial, ferroviaria y terrenos asociados
Zonas industriales o comerciales
Tejido urbano continuo
Tejido urbano discontinuo
Vegetación secundaria o en transición
Zonas verdes urbanas
A partir de esta información se efectúa la clasificación de las coberturas, según la permeabilidad o facilidad del movimiento que puede llegar a tener la especie. Para esta categorización, se aprovecharon las observaciones en campo y la información encontrada en la literatura, aprovechando que I.capreolus es una especie cosmopolita ampliamente estudiada en Sudamérica con reportes que describen la ecología y comportamiento de la especie. Partiendo quetienen preferencia por cuerpos de agua lénticos y zonas inundables, así como ecosistemas artificiales de piscicultura, además de pastos abiertos y arbolados, para el desplazamiento de los adultos (Renner et al., 2015; Sánchez‐Guillén et al., 2020; Vilela et al., 2017) Otras coberturas serán como barrera o de una considerable resistencia al movimiento de la especie, la clasificación se presenta en la Tabla 2.
34
Tabla 2 Lineamientos para la recategorización de las variables categóricas en función de la respuesta ecológica de la especie I. capreolus.
Categoría Calificación Descripción
Resistencia mínima 0 Mejor calidad de hábitat con mayores oportunidades de supervivencia y éxito reproducción.
Resistencia muy baja 1 Hábitats modificados. Conservan altas tasas de supervivencia y éxito reproductivo. Mayores niveles de perturbación antrópica en comparación con categoría 1.
Resistencia baja 15 Hábitats modificados. Conserva moderado éxito reproductivo. Baja supervivencia. Mayores niveles de perturbación antrópica en comparación con categoría 1 y 2.
Resistencia moderada 30 Hábitats modificados. Bajo éxito reproductivo. Baja supervivencia. Mayores niveles de perturbación antrópica en comparación con categoría 1, 2 y 3.
Resistencia alta 60 Sitios de poco frecuente. No hay éxito reproductivo. Mayores niveles de perturbación antrópica en comparación con categoría 4.
Resistencia muy alta 80 Sitios evitados por especie bioindicadora del análisis de conectividad. Mayores niveles de perturbación antrópica en comparación con categoría 5.
Resistencia máxima 100 Sitios no usados por especie bioindicadora del análisis de conectividad dada su máxima resistencia, lo que indica mayores niveles de perturbación antrópica en comparación con todas las demás categorías.
En la Tabla 3 se presentan los resultados de la clasificación, como se puede observar, las mayores calificaciones de ecosistemas terrestres se encuentran en aquellas coberturas que se asocian con grandes densidades de árboles, sin vegetación, ya sea zonas naturales (tierras desnudas) o sitios urbanos. Mientras que, para ecosistemas acuáticos, se encontraron las mayores calificaciones para cuerpos de agua lóticos, la cual agrupa ríos y quebradas. Aquellas coberturas que se caracterizan por poseer una baja densidad arbórea, y estar asociada a cuerpos de agua como charcos o lagunas, presentaron las menores calificaciones, es decir describen la preferencia de hábitat de la especie de interés, como se describió anteriormente.
la resistencia del territorio en función del tipo de cobertura.
35
Macrovariable Variable Calificación Cuerpos de agua Ríos (50 m) 60 Uso agropecuario Mosaico de cultivos y espacios naturales 15 Mosaico de cultivos, pastos y espacios naturales 15 Mosaico de pastos con espacios naturales 15 Mosaico de pastos y cultivos 30 Pastos arbolados 15 Pastos enmalezados 30 Pastos limpios 30 Tierras desnudas y degradadas 60 Uso preservación Bosque denso 100 Plantación forestal 15 Vegetación secundaria o en transición 15 Zonas verdes urbanas 15 Uso urbano Red vial, ferroviaria y terrenos asociados 100 Zonas industriales o comerciales 100 Tejido urbano continuo 80 Tejido urbano discontinuo 80
Tabla 3 Calificación de
Con esta información, se crea la matriz de resistencia, usando la herramienta de álgebra de mapas de QGIS se combinan en un único Ráster, todas las variables presentadas anteriormente. Con esto se logra que aquellas por las que la especie no entra tendrán los valores ponderados más altos, mientras que entre mayor preferencia de hábitat el valor será menor por lo tanto hay menor resistencia al movimiento.
3.5.2 Conectividad entre coberturas
El análisis anterior clasifica de manera individual las coberturas, pero no caracteriza la interacción entre estas, por ejemplo, si un pasto limpio se encuentra colindando con un bosque, dónde se sentirá mejor la especie. Por eso, finalmente, se consolida la matriz de resistencia conforme a una evaluación binaria entre variables, es decir comparando uno a uno, usando el método de ordenamiento, a partir de una estructura jerarquizada, propuesto por Saaty (1987) Según su propuesta, la jerarquía de interacción entre variables no es hecha de manera subjetiva, siempre y cuando exista un conocimiento amplio y suficiente que caracterice el grupo de interés, paso que permite completar el numeral anterior, de resistencia. Mientras que de esta manera se prioriza la interacción entre variables y de esta manera saber cuál de las coberturas de interés resulta ser más relevante para caracterizar la presencia de la especie de interés y en cuál es poco probable encontrarla Como se muestra en la Tabla 4, hay siete posibles valores para adjudicar, los cuales definen la importancia de una variable sobre otra, en este caso sobre una cobertura y otra. Luego, se completa la Tabla 5, de esta manera bajo la diagonal mostrada, se prioriza las variables presentadas en la columna, de tal manera que, si SC2 es de menor habitabilidad para SC1, tendrá un valor del orden de 1/3-9 o menor, es decir que SC1 sería un hábitat preferido sobre SC2.
Tabla 4 Escala de clasificación entre variables
Tabla 5 Formato ejemplo para comparar las variables de interés
Una vez esto fue establecido, se realizó la consolidación de la interacción entre las diferentes agrupaciones de coberturas realizadas, con el fin de optimizar el análisis en

36
Subcriterios SC1 SC2 SC3 SC1 1 5 7 SC2 1/5 1 7 SC3 1/7 1/7 1
función de los resultados de resistencia. Agrupando aquellas coberturas que permiten el establecimiento de los adultos terrestres como zonas habitables, sitios de paso o que permiten la conectividad como bosques y cultivos, y aquellas coberturas que son una barrera total para la especie. Adicionalmente, y dado a su vida acuática larval y para reproducción, se obtuvo tres grupos, el de drenajes dobles (ríos), siendo este una barrera para la especie; sencillos (quebradas), el cual permite en cierta medida el cruce de la especie, aunque no el establecimiento; y finalmente hábitat acuático haciendo completa referencia a ecosistemas lénticos. Con esta información se establece la matriz de importancia y costo en función de las interacciones entre las variables, la cual se presenta en la Tabla 6
Finalmente, con la herramienta Circuitscape se logra consolidar el análisis, combinando la información de presencia de la especie, la matriz de resistencia, que presenta la preferencia de hábitat y la matriz de costo, que relaciona la probabilidad de pasar de un tipo de cobertura a otro. Esto permite entender la conectividad que hay en un área determinada, y con esto se evaluará la pertinencia de cada uno de los modelos de distribución de especies construidos.
37
Subcriterios C1 C2 C3 C4 C5 C6 C7 Bosques C1 1.0 0.7 0.3 0.5 0.3 0.7 0.7 Hábitat C2 0.7 1.0 0.2 0.8 0.3 0.7 1.0 No Hábitat C3 0.3 0.2 1.0 0.4 1.0 0.7 0.8 Cultivos C4 0.5 0.8 0.4 1.0 0.5 0.7 0.9 Drenajes dobles C5 0.3 0.3 1.0 0.5 1.0 0.1 0.6 Drenajes sencillos C6 0.7 0.7 0.7 0.7 0.1 1.0 0.7 Hábitat Acuático C7 0.7 1.0 0.8 0.9 0.6 0.7 1.0
Tabla 6 Interacción entre variables/coberturas, según la preferencia de hábitat.
3.5.3 Creación de la matriz de resistencia.
4 RESULTADOS
A continuación, se describen los resultados de análisis tradicional y con la implementación de la metodología propuesta para poner a prueba los modelos de distribución de especie.
4.1 Registros para la especie Ischnura capreolus
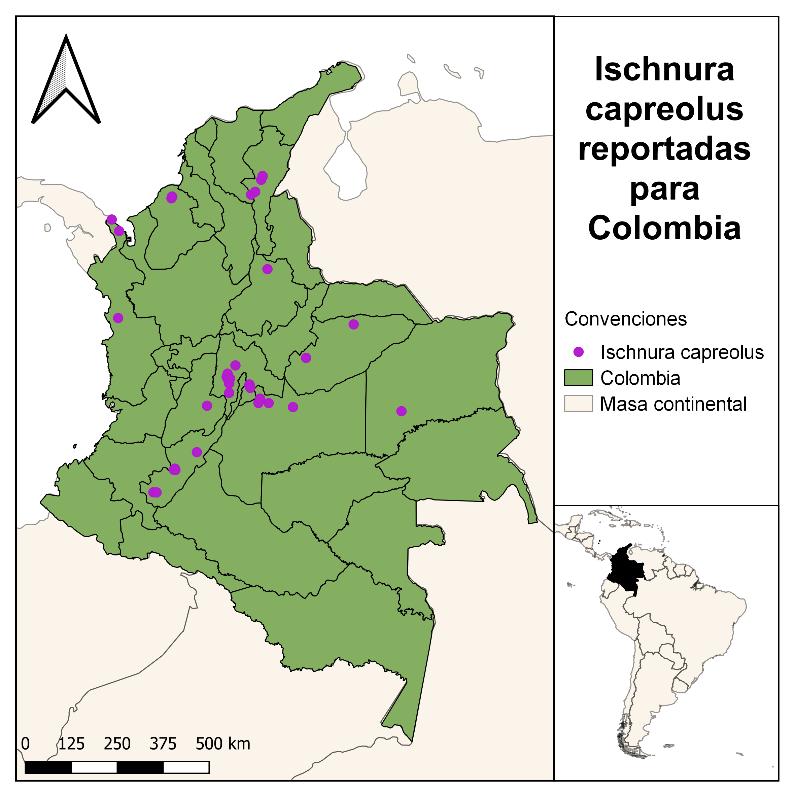
Para la especie Ischnuracapreolus en el Museo de Historia Natural Cornelis Johannes Marinkele de Bogotá-Colombia, se registraron y validaron las determinaciones para 43 registros para Colombia, teniendo una cobertura en diez departamentos, es decir el 31.25% del territorio, con dos registros (~2%) para los departamentos de Santander, Tolima y Vichada; tres registros para Casanare y Chocó (~7%); cuatro registros para Córdoba (~9%); cinco registros para Meta (~12%); seis en el Cesar (~14%); siete en el Huila (~16%) y doce para Cundinamarca (~28%). Los resultados se presentan en la Figura 10, en un mapa generado haciendo uso de la herramienta QGIS.
38
Figura 10 Distribución reportada de la Ischnuracapreolus en Colombia, a partir de información encontrada en el Museo de Historia Natural CJ Marinkelle.
4.2 Distribución potencial de la especie Ischnura capreolus
A continuación, se procederán a presentar los principales resultados estadísticos de los modelos desarrollados para la evaluación biológica de la distribución de la especie Ischnuracapreolus Cada modelo se describirá en primer lugar la efectividad y capacidad predictiva de este, posteriormente, el nivel de importancia de las variables implementadas, luego la respuesta que tuvieron las variables y finalmente, la distribución esperada de la especie de interés.
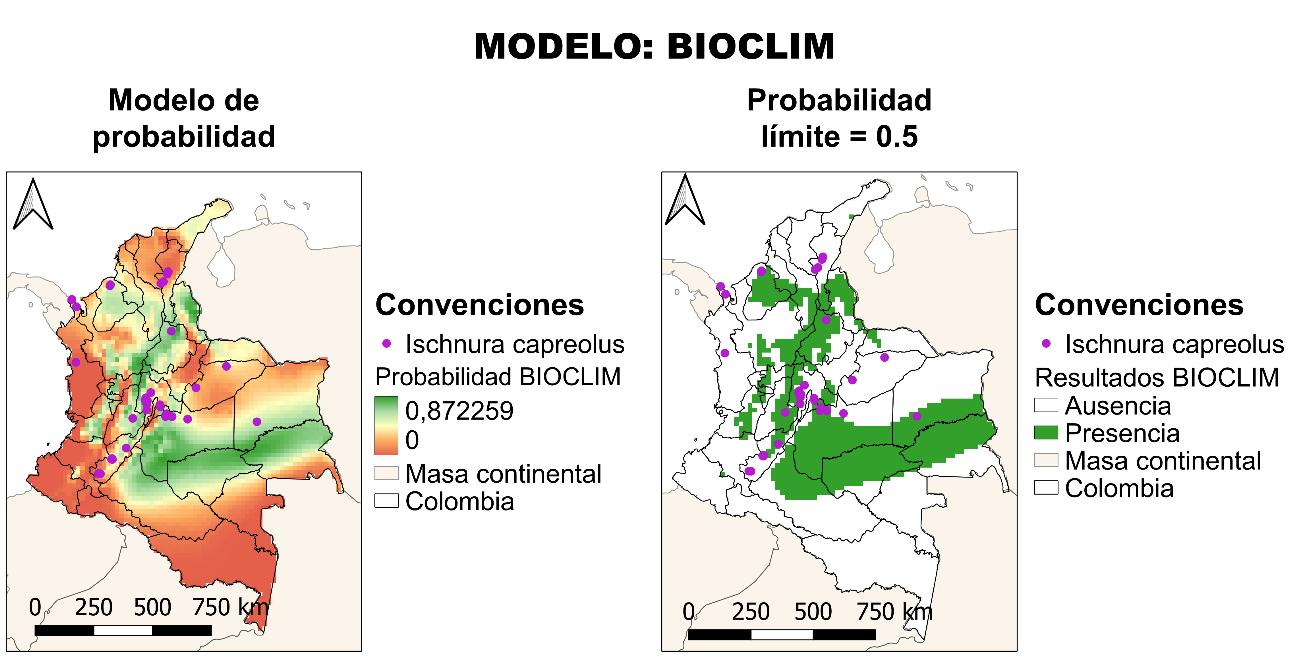
4.2.1 Resultados BIOCLIM
El primero modelo que se describirá es BIOCLIM, el cual no tuvo un resultado positivo, esto medido con la curva AUC-ROC. El resultado fue del 60% (véase la Figura 11), es decir que el modelo tiene una probabilidad de no acertar en la predicción de un 40%, siendo este un valor muy alto, indicando que la distribución real de la especie en términos estadísticos puede ser desacertada.
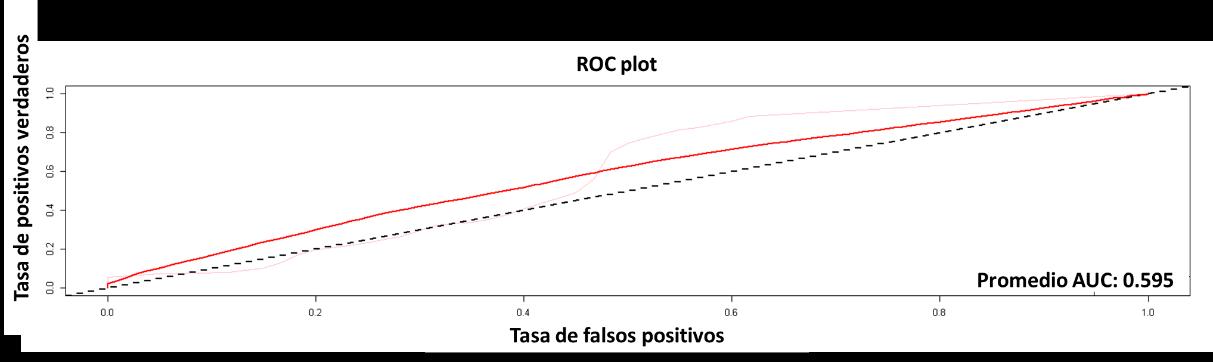
11 Curva ROC para el modelo BIOCLIM
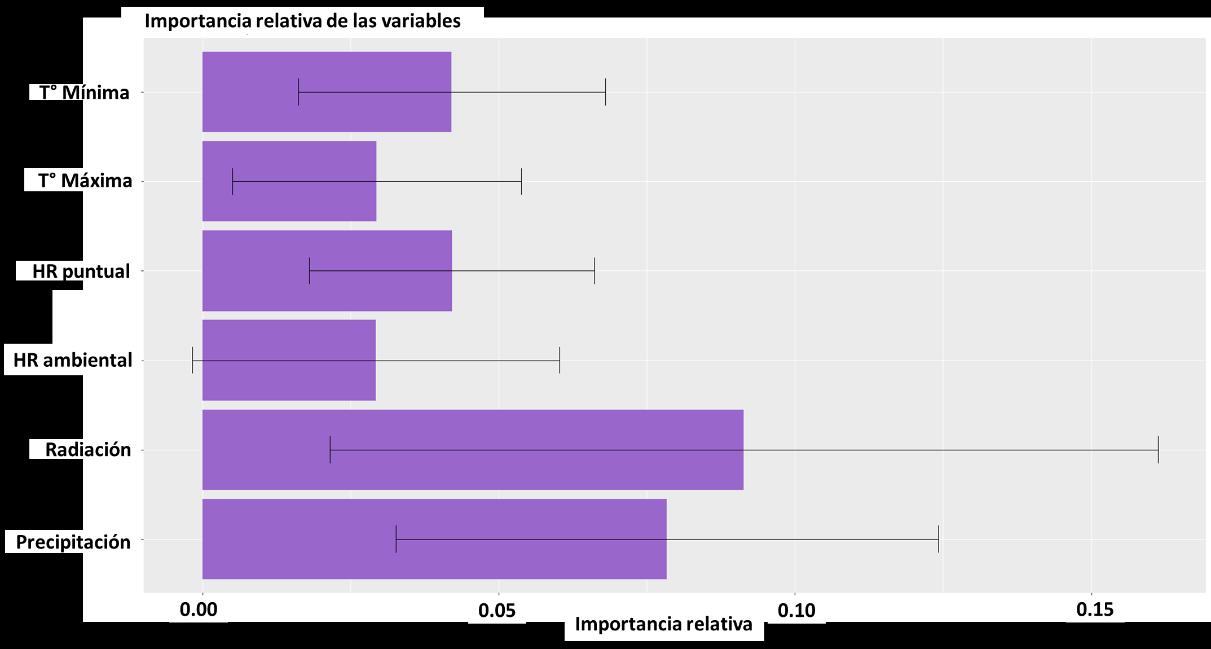
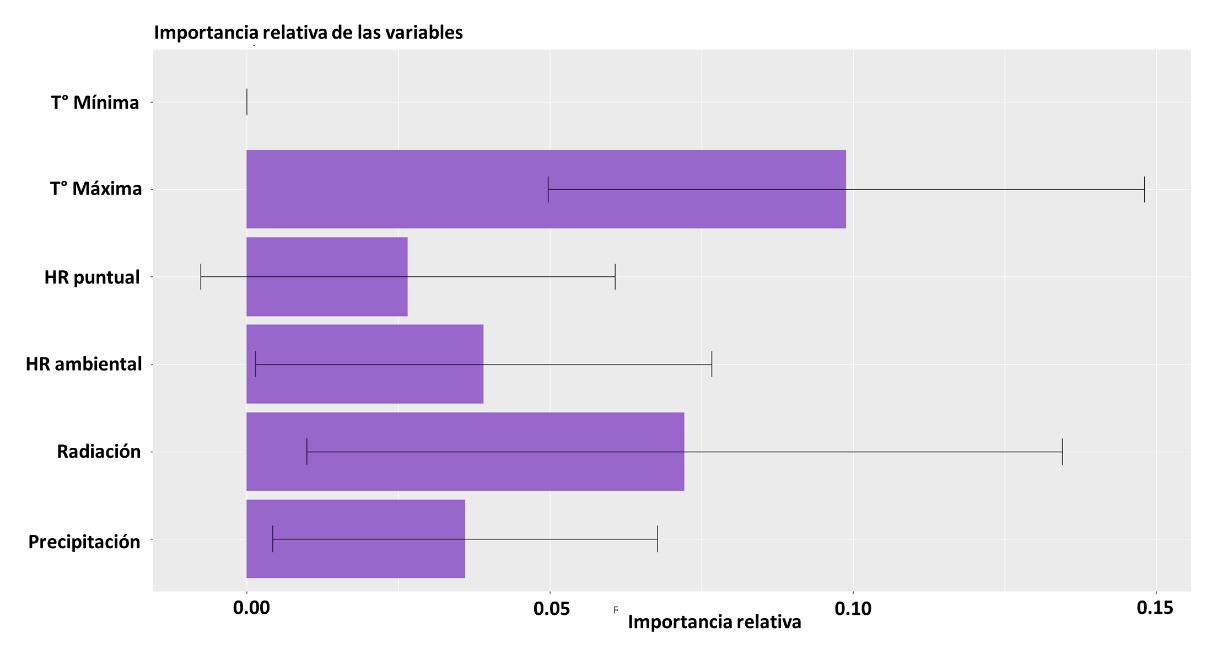
Para la construcción del modelo, las variables implementadas no tuvieron diferencias significativas, como se evidencia en la Figura 12, donde se reportan los intervalos de confianza y peso del efecto de cada una de las capas. Sin embargo, dados los resultados es posible jerarquizar los resultados de la siguiente manera:
1. Radiación
2. Precipitación
3. Temperatura mínima
4. Humedad relativa puntual
5. Temperatura máxima
6. Humedad relativa ambiental
39
Figura
Según los resultados para el modelo BIOCLIM, la precipitación y la radiación son determinantes para predecir la distribución potencial de la especie de interés. Lo cual es válido con la constitución biológica de una libélula, siendo organismos ectotermos
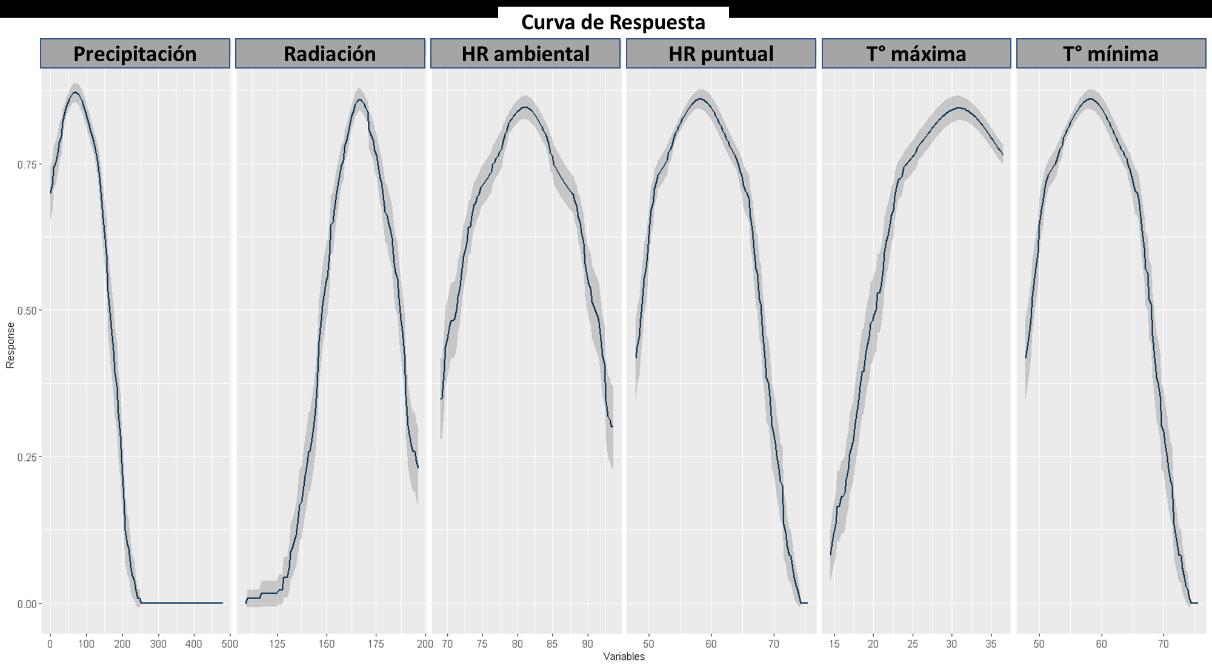
Respecto a la respuesta de cada una de las variables mostrada en la Figura 13, se puede observar los valores más frecuentes que permiten determinar la distribución de la especie. Siendo para la precipitación preferentes los valores bajos, es decir bajas lluvias. Contrario con los valores de radiación, los cuales, si muestran preferencia a valores altos de este, correspondiente con la preferencia de una temperatura máxima elevada. Adicionalmente, la temperatura mínima muestra que hay una mayor presencia en bajas temperaturas, o al menos bajas en referencia a la temperatura máxima, lo cual demuestra una gran resiliencia para resistir y tolerar abruptos cambios de temperatura, sin embargo, esto se asocia con el estadio de resistencia, más conocido como larva o náyade.
40
Figura 12 Importancia de las variables en la construcción del modelo a partir de BIOCLIM.
Finalmente, para el modelo BIOCLIM, en la Figura 14 se muestra la distribución potencial en términos de probabilidad y en términos de presencia ausencia partiendo de un valor límite establecido por el modelo de 0.5. Este resultado, significa en un notable aumento de la distribución potencial de la especie, contrastado con la distribución real, pasando de estar en un 31% del territorio al 53% de este, encontrándose de manera adicional en los departamentos de Antioquia, Arauca, Bolívar, Caldas, Caquetá, Guainía, Guaviare, Norte de Santander, Quindío, Sucre y Valle del Cauca. Sobre las particularidades ecológicas del modelo de predicción, es importante mencionar que la especie se encuentra fuertemente asociada a la cadena montañosa de la cordillera de los Andes, con un potencial movimiento sobre los llanos orientales, en la zona que tiene límite biogeográfico con la Amazonía. Recordando que Ischnuracapreolus es una especie fuertemente ligada con zonas de alta radiación, pero baja precipitación, por lo que los resultados son congruentes.
41
Figura 13 Caracterización de las condiciones ambientales según el modelo BIOCLIM.
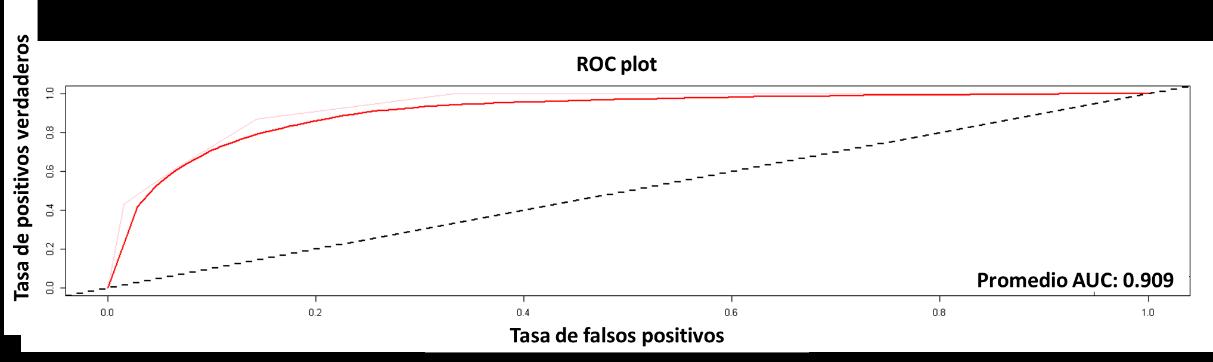
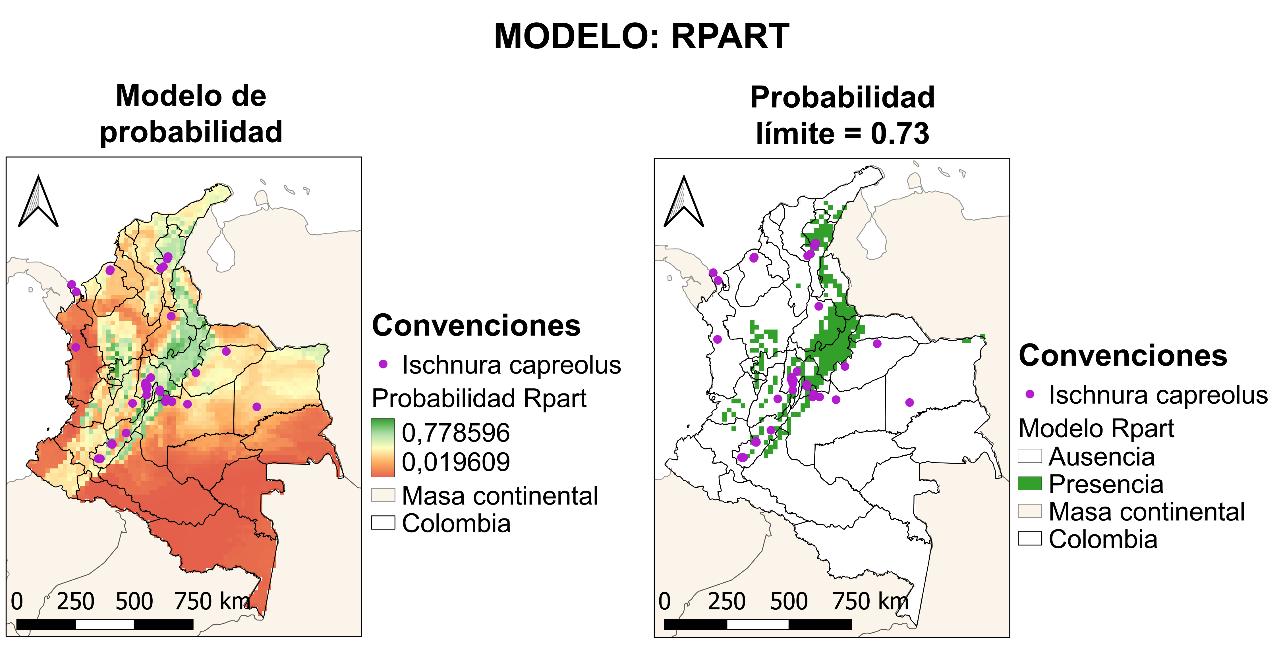
4.2.2 Resultados RPART
Seguido se presentarán los resultados del modeloRPART, cuyo resultado de la curva AUCROC, es del 90% (véase la Figura 15). Este resultado contrario al modelo de BIOCLIM, es muy positivo, quiere decir que existe un alto nivel de confianza y capacidad predictiva por parte de este modelo.
Para la construcción del modelo, en la Figura 16 se muestra la importancia de las variables implementadas. En general no hubo diferencias significativas entre las capas, excepto para la temperatura mínima, la cual, dada su alta correlación con la temperatura máxima, el modelo reduce el efecto de esta variable, lo cual explica el valor negativo en algunas iteraciones del modelo para la variable humedad relativa puntual, la cual se correlaciona con la humedad relativa ambiental. A continuación, se describe la jerarquía de importancia de las variables:
42
Figura 14 Mapa de distribución potencial construido a partir del método BIOCLIM.
Figura 15 Curva ROC para el modelo RPART.
1. Temperatura máxima
2. Radiación
3. Humedad relativa ambiental
4. Precipitación
5. Humedad relativa puntual
6. Temperatura mínima, con valor mínimo
De igual manera, se puede observar que las variables con mayor relevancia son aquellas asociadas a la condición de vida ectotermo, siendo este un factor biológico determinante para establecer y modelar la distribución de una especie.
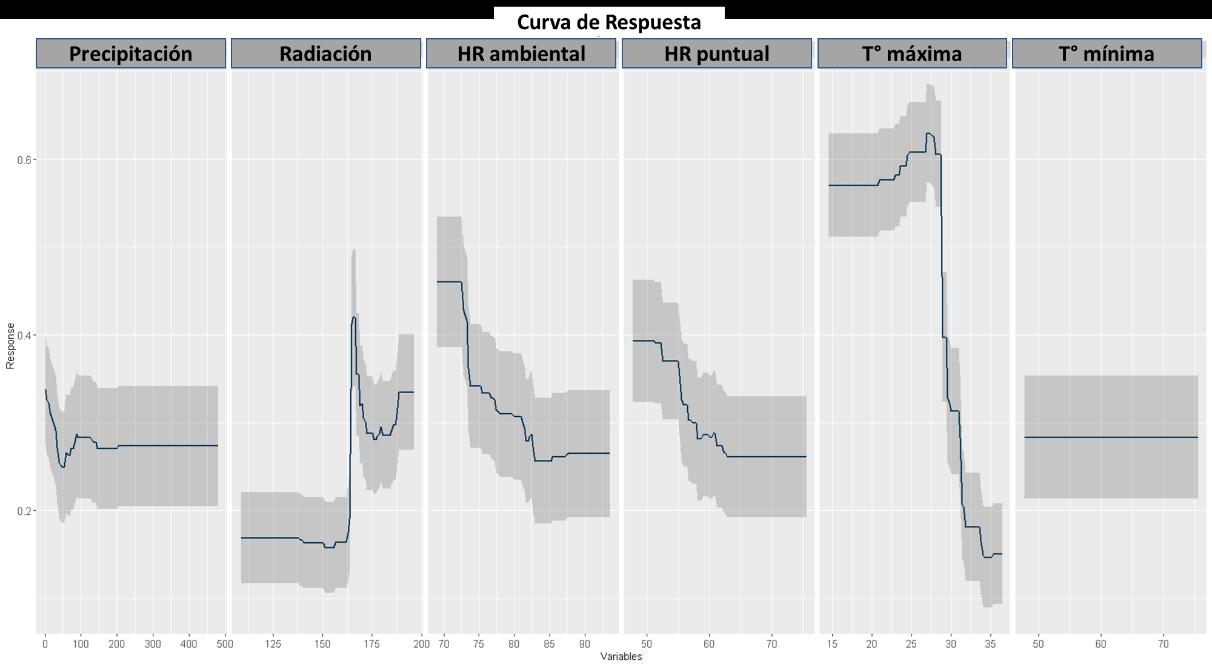
Respecto a la respuesta de las variables analizadas, en la Figura 17 se puede observar que las variables de precipitación y temperatura mínima no tienen una respuesta representativa en algún valor. Esto es contrario en las restantes capas, siendo en la radiación una respuesta para valores altos, mientras que para la humedad relativa tanto ambiental como puntual, se tiene que la especie tiene una mejor respuesta para ecosistemas o nichos secos y para la temperatura máxima se encuentra una alta respuesta a valores por debajo de 27 grados. Algo que se conserva respecto al modelo BIOCLIM es la respuesta a las fluctuantes temperaturas, sin embargo, esta vez la precipitación no tuvo efecto, pero los resultados siguen siendo congruentes en términos biológicos para la especie de interés, pese a las fluctuaciones presentadas.
43
Figura 16 Importancia de las variables en la construcción del modelo a partir de RPART.
Finalmente, para el modelo RPART, en la Figura 18 se presenta la distribución potencial de la especie de igual manera que con el modelo anterior, pero con un valor límite de probabilidad de 0.73. Pese a que en principio se ve una distribución más restringida, se confirma una estrecha relación de la especie con la cadena montañosa de la cordillera de los Andes, ahora reportada en el 56% del territorio colombiano. Se amplía su distribución según el modelo a los siguientes departamentos: Antioquia, Arauca, Bolívar, Boyacá, Caldas, Caquetá, Cauca, La Guajira, Norte de Santander, Quindío, Risaralda y Valle del Cauca.
44
Figura 17 Caracterización de las condiciones ambientales según el modelo RPART.
Figura 18 Mapa de distribución potencial construido a partir del método RPART.
4.2.3 Resultados GAM
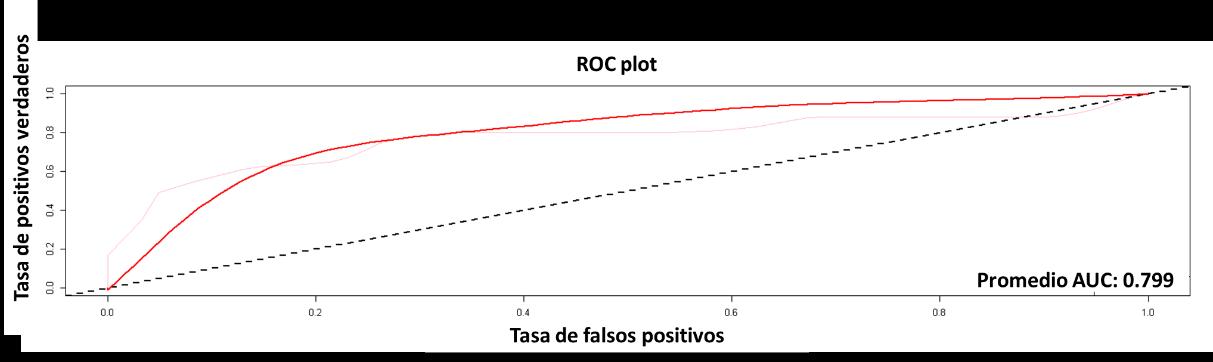
A continuación, se presentan los resultados para el modelo GAM. En la Figura 19, se muestra la curva AUC-ROC, con un valor del 99%, el cual al igual que el modelo RPART, muestra una alta capacidad predictiva por parte del modelo basado en las variables introducidas en este.

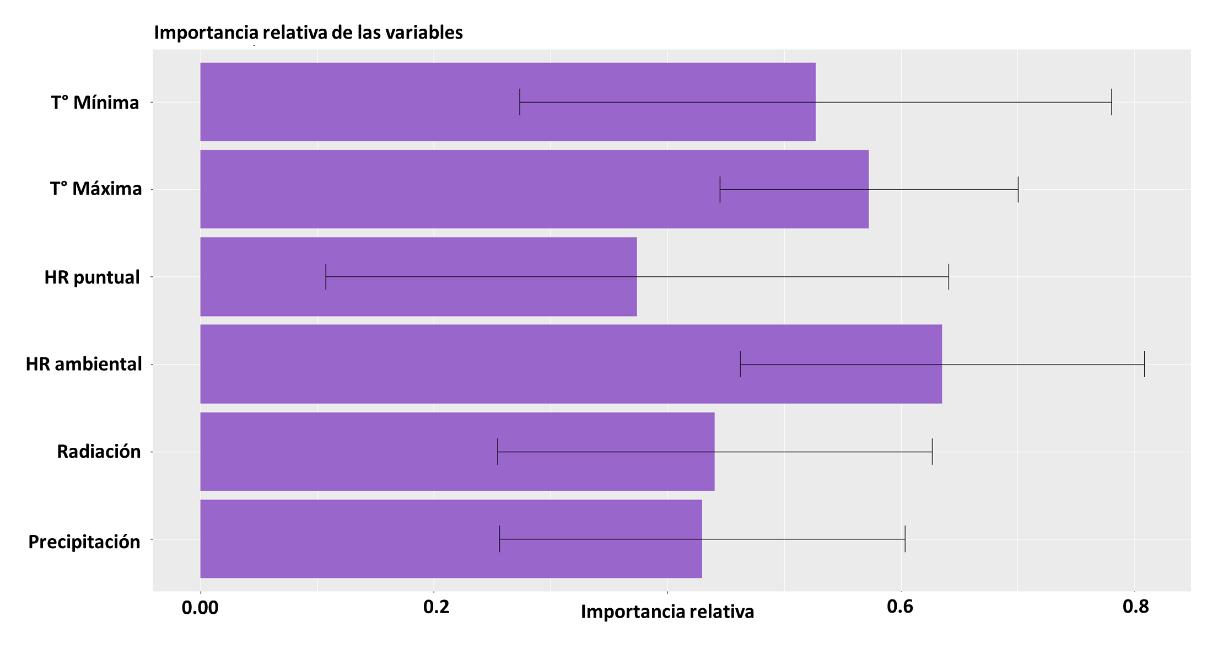
De igual manera, para el modelo GAM se realizó el análisis de importancia para la generación del modelo con las variables implementadas. Para este caso todas tuvieron un peso y a continuación, se presenta la jerarquización de estos:
1. Humedad Relativa Ambiental
2. Temperatura máxima
3. Temperatura mínima
4. Radiación
5. Precipitación
6. Humedad relativa puntual
Los resultados son diferenciales respecto a los otros modelos, sin embargo, son de alguna manera congruentes porque reflejan la estrecha relación que tienen los insectos con la temperatura siendo estos ectotermos. Sin embargo, como se evidencia en la Figura 20, se observa una variable de gran relevancia que en los anteriores modelos no había sido contemplada y es la humedad relativa, esto indica que hay un requerimiento sobre esta variable ambiental condicionando la probabilidad de distribución de la especie.
45
Figura 19 Curva ROC para el modelo GAM.
Las respuestas de las variables según el modelo GAM se presentan en la Figura 21, donde se hace evidente una respuesta diferencial. Por ejemplo, en términos de precipitación, radiación y humedad relativa, la especie tiene una preferencia sobre rangos bajos para estas capas ambientales. Contrario con la humedad relativa puntual, la cual es evidente que prefieren o tienen una mayor abundancia donde esta variable tiene valores superiores a 60% de HR. Un fenómeno constante en los modelos presentados hasta ahora es respecto a la temperatura, el cual a partir del valor de temperatura máxima y mínima se hace evidente la plasticidad de nicho a variaciones considerables de temperatura, mostrando una gran resiliencia cuando varía en el territorio.
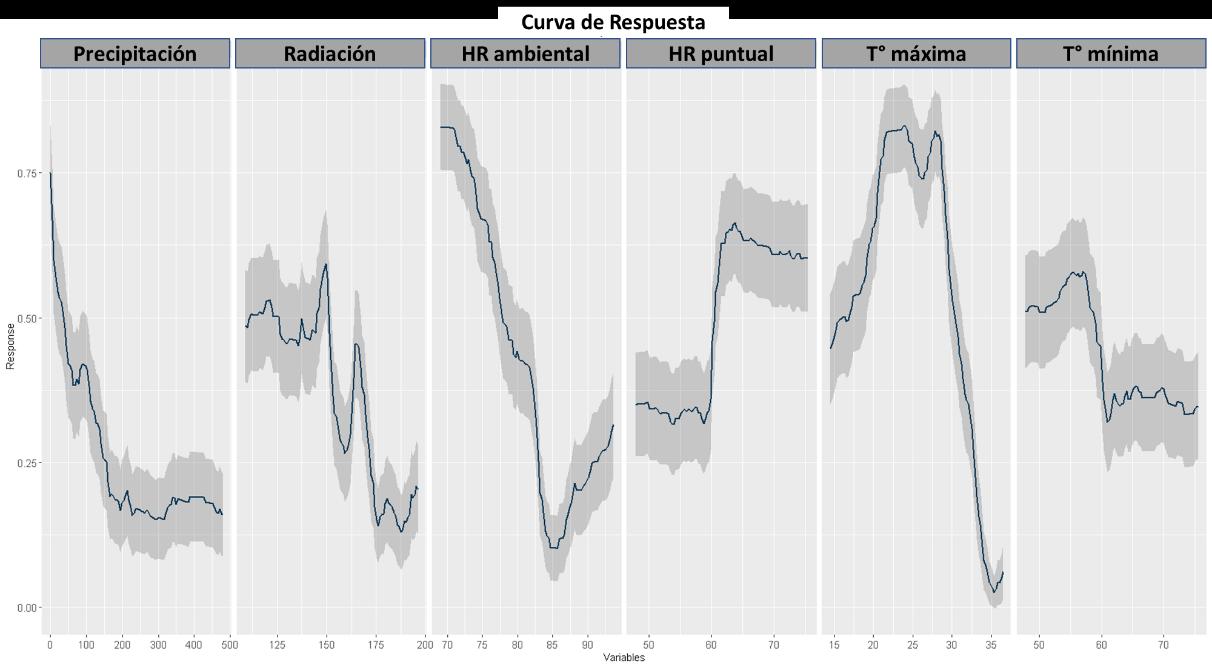
46
Figura 20 Importancia de las variables en la construcción del modelo a partir de GAM.
Figura 21 Caracterización de las condiciones ambientales según el modelo GAM.
Finalmente, para el modelo GAM, en la Figura 22 se muestra el mapa de distribución potencial para la especie con un límite de probabilidad de 0.73 para el modelo. Con este modelo, hay una cobertura de la especie en el territorio equivalente al 66% de los departamentos de este, manteniendo su dependencia a la cadena montañosa de la cordillera de los Andes. Se extiende su distribución potencial a los siguientes departamentos: Antioquia, Arauca, Bolívar, Boyacá, Caldas, Casanare, Cauca, Cesar, Guajira, Meta, Norte de Santander, Quindío, Risaralda, Sucre y Valle del Cauca.
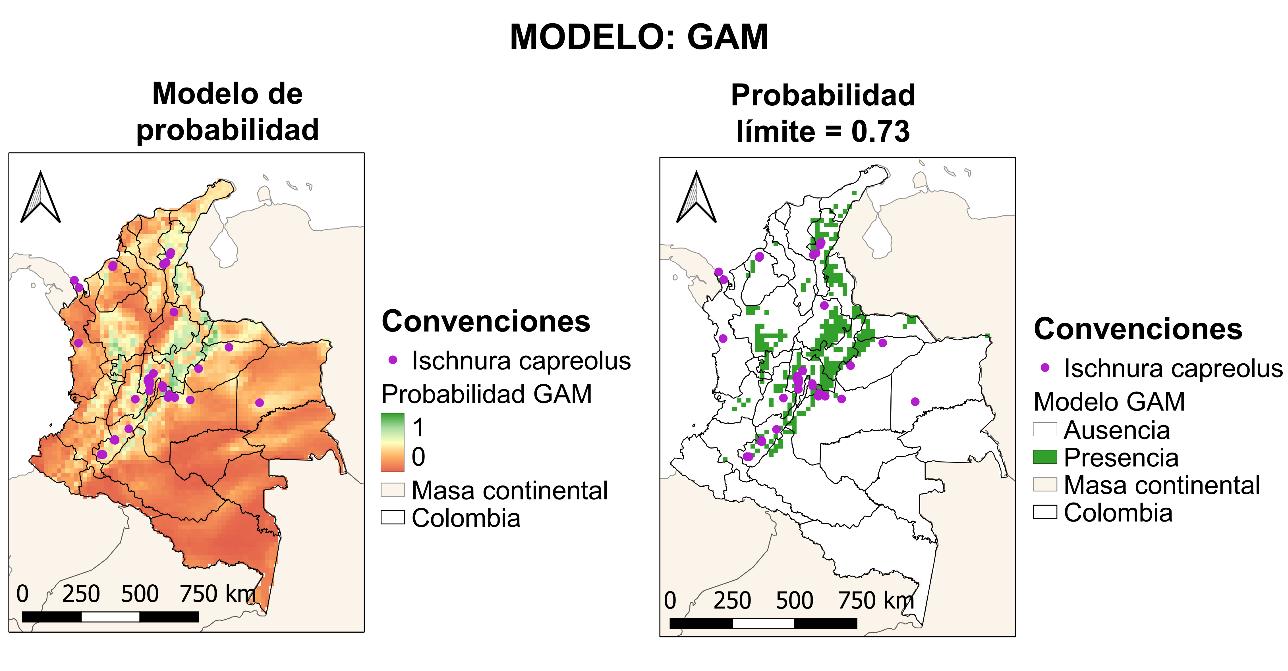
4.2.4 Resultados GLM
El cuarto modelo presentado será GLM, cuyo resultado para la curva AUC-ROC se muestra en la Figura 23. El resultado de este fue del 80%, lo cual indica que el modelo tiene una probabilidad de error del 20% sobre la predicción de la distribución de la especie realizada.
47
Figura 22 Mapa de distribución potencial construido a partir del método GAM.
Figura 23 Curva ROC para el modelo GLM.
Respecto de la importancia de las variables, de acuerdo con los intervalos de confianza presentados en la Figura 24 hay una diferencia significativa entre la humedad relativa ambiental con las demás variables analizadas, de igual manera entre la temperatura máxima y las restantes capas, siendo estas dos las de mayor importancia, sin diferencia significativa entre estas. Adicionalmente, se presenta a continuación, en orden jerárquico la importancia de las variables analizadas:
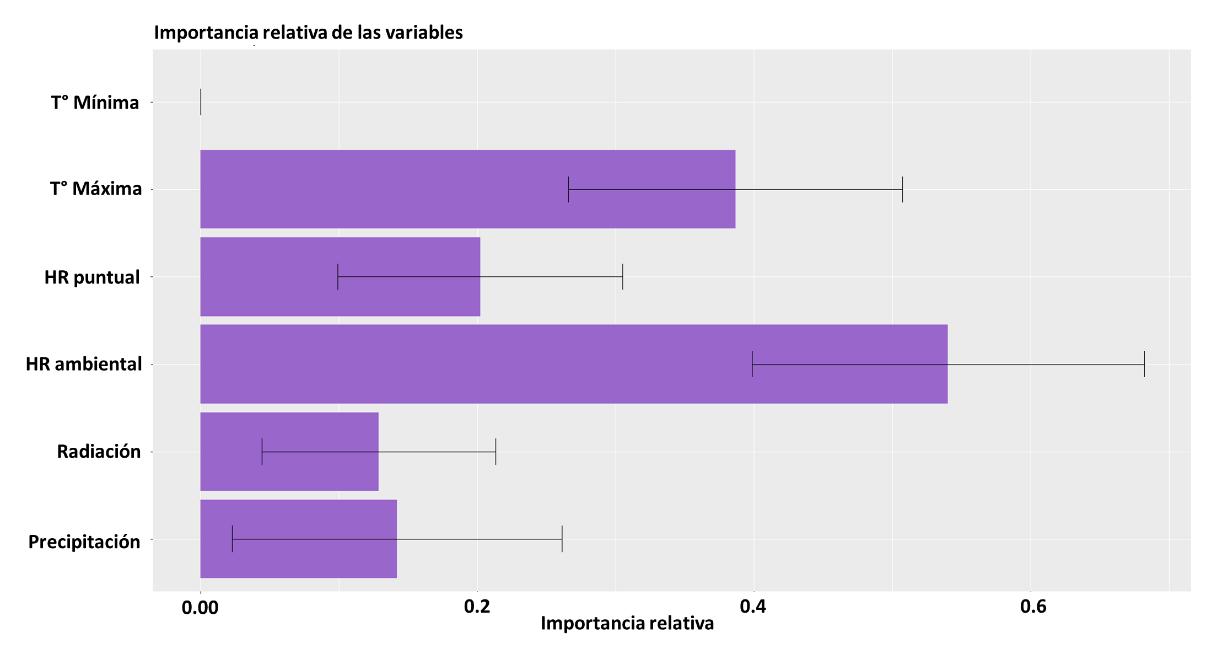
1. Humedad relativa ambiental
2. Temperatura máxima
3. Humedad relativa puntual
4. Precipitación
5. Radiación
6. Temperatura mínima, siendo su relevancia igual a cero
Las variables más importantes en el modelo GLM (véase la Figura 24) son las mismas que en el modelo previamente descrito GAM (véase la Figura 21). Manteniendo la congruencia existente con la biología de los odonatos.
En términos de la respuesta de cada una de las variables (véase la Figura 25) usadas para la construcción del modelo se tiene una baja respuesta de la temperatura mínima, siendo esta una variable que no tuvo ningún peso en la construcción de la predicción de distribución. Contrario a las restantes variables, donde contrario a lo observado en los modelos anteriores la radiación tuvo un factor importante a partir de la presencia de zonas con baja radiación, pero una alta humedad relativa puntual. Particularmente, para este
48
Figura 24 Importancia de las variables en la construcción del modelo a partir de GLM.
modelo no es evidente la mencionada resiliencia de la especie, marcado una afinidad con territorios con temperaturas menores a 25°C. Esto es muy relevante porque habla de la capacidad de dispersión estrictamente del adulto, sin la asunción que donde hay un adulto hay ~100% de encontrar una larva.
Para el modelo GLM presentado en la Figura 26, hay un aumento en la distribución de la especie en un 47%, mostrándose fuertemente restringida en la región montañosa de la cordillera de los Andes, con una pequeña presencia en la región caribe en el norte del país. Adicionalmente, a continuación, se listan los departamentos nuevos según el modelo: Antioquia, Arauca, Atlántico, Boyacá, Caldas, Caquetá, Casanare, Cauca, Cesar, La Guajira, Magdalena, Nariño, Norte de Santander, Risaralda y Valle del Cauca
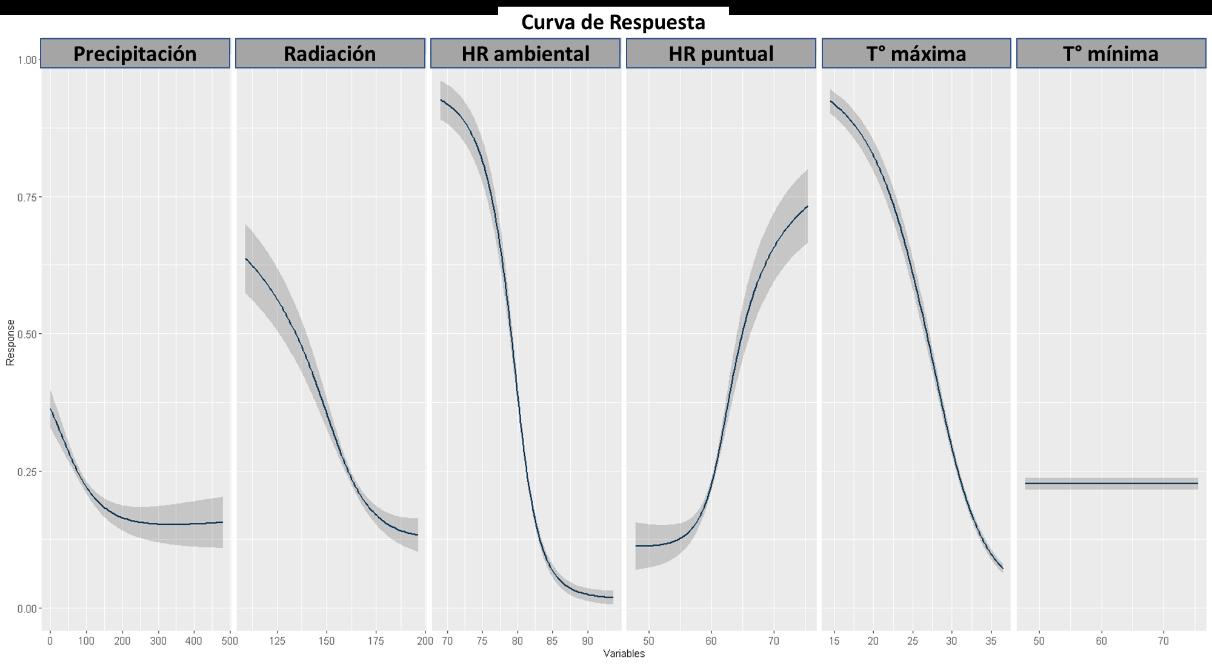
49
Figura 25 Caracterización de las condiciones ambientales según el modelo GLM.
4.2.5 Resultados Maxent
Para finalizar, se presenta el modelo Maxent, cuyos resultados se presentan en la Figura 27 Con un porcentaje del 76%, para la curva AUC-ROC, esto para la capacidad predictiva de la especie de interés.
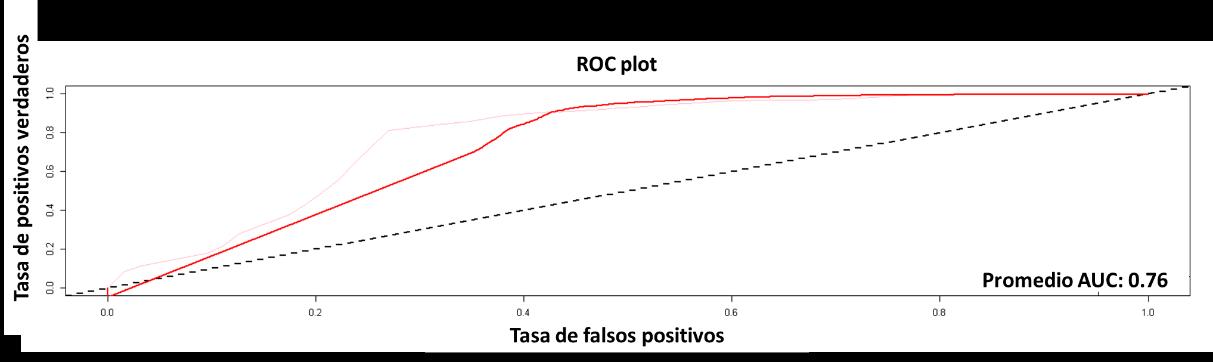
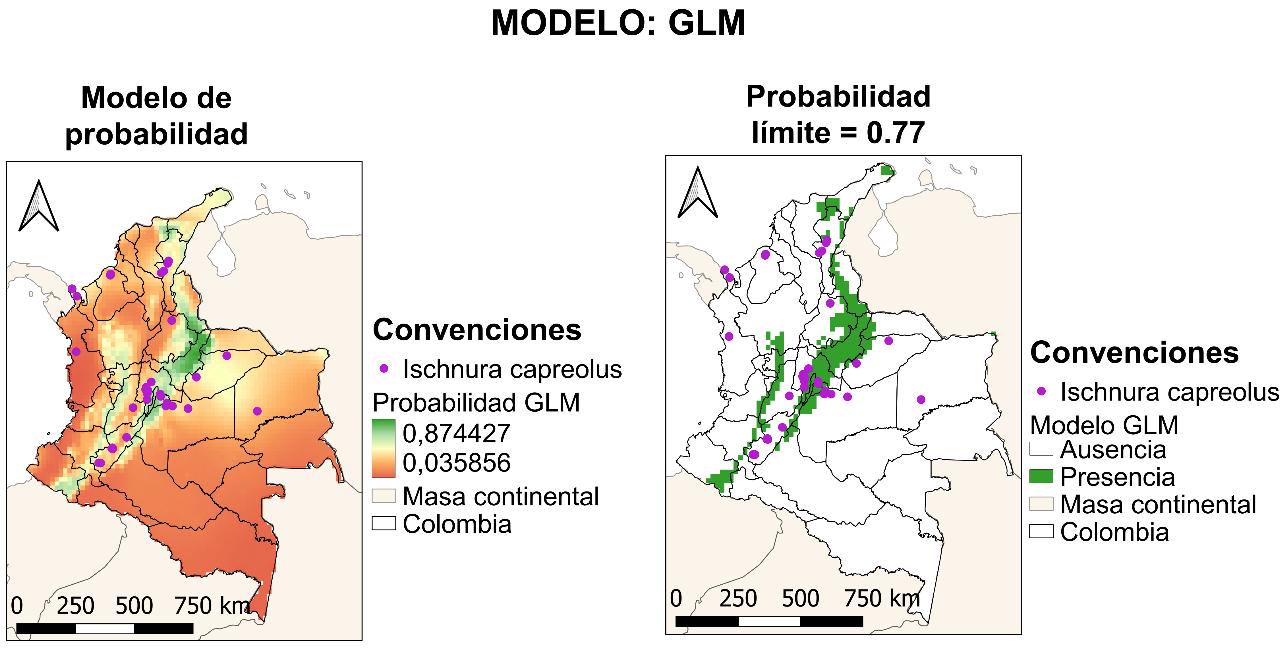
La importancia relativa para el modelo Maxent, se presenta en la Figura 28, donde se evidencia que no hay diferencias significativas entre las variables incorporadas. Sin embargo, hay una jerarquía entre estas la cual se describe a continuación:
1. Temperatura máxima
2. Humedad relativa puntual
3. Temperatura mínima
4. Radiación
50
Figura 26 Mapa de distribución potencial construido a partir del método GLM.
Figura 27 Curva ROC para el modelo Maxent
5. Humedad relativa ambiental
6. Precipitación
La temperatura y humedad relativa han sido constantes en los modelos ejecutados, siendo estos los factores que han estado como las dos variables más importantes. Las posiciones o importancias siguientes muestran una mayor varianza entre los modelos, no siendo consistente entre estos.
Figura 28 Importancia de las variables en la construcción del modelo a partir de Maxent
Sobre la respuesta de las variables presentada en la Figura 29, sigue resaltando la variante respuesta de la temperatura, mostrándose asociada a regiones o territorios que tienen una considerable fluctuación de la temperatura. Adicionalmente, se observa una marcada congruencia entre la respuesta y la importancia, aquellas variables que muestran una respuesta más generalista por parte de la especie serán aquellas variables que no tienen una alta jerarquía dentro del modelo.
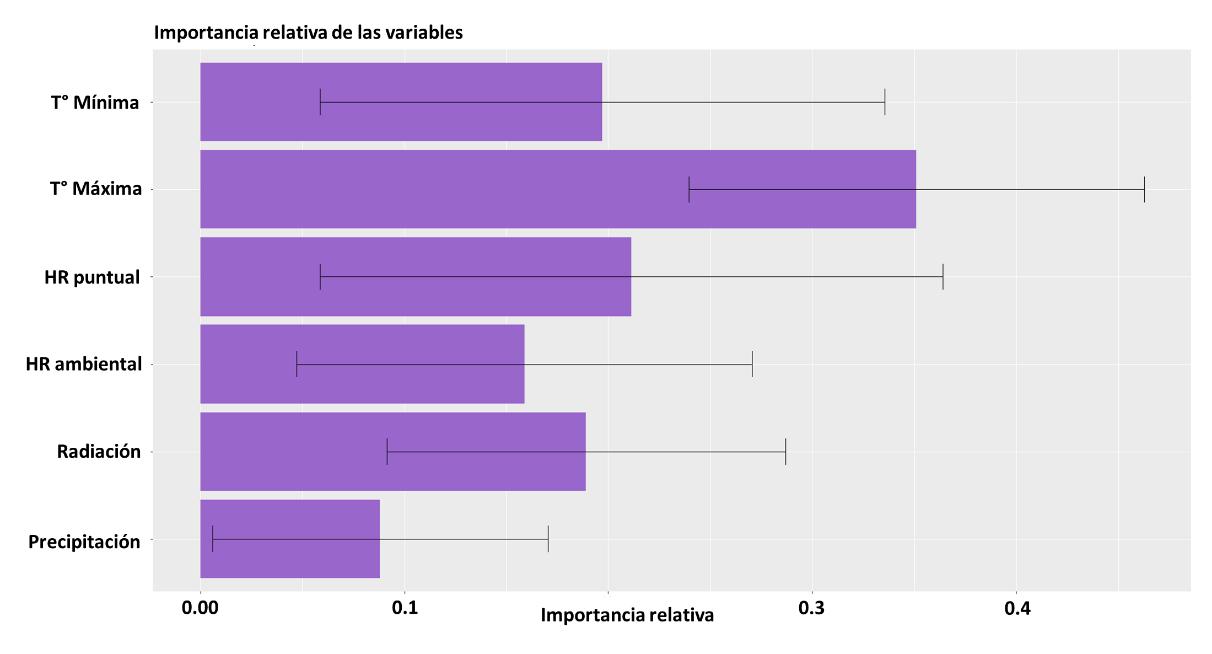
51
Según la predicción realizada por el modelo a partir de Maxent, se puede esperar que haya un aumento del 68% de la especie sobre el territorio colombiano en término de departamentos reportados y colectas realizadas. Se proyecta presencia adicional en los siguientes departamentos: Antioquia, Arauca, Atlántico, Bolívar, Boyacá, Caldas, Caquetá, Casanare, Cauca, Cesar, Chocó, Guainía, Guaviare, La Guajira, Magdalena, Nariño, Norte de Santander, Putumayo, Quindío, Risaralda y Sucre. Según este modelo, se esperaría que la especie no se encuentre restringida a la región montañosa de la cordillera de los Andes, por el contrario, habita un territorio más heterogéneo, como la llanura de la región del Orinoco, la zona Caribe y parte de la región del Pacífico.
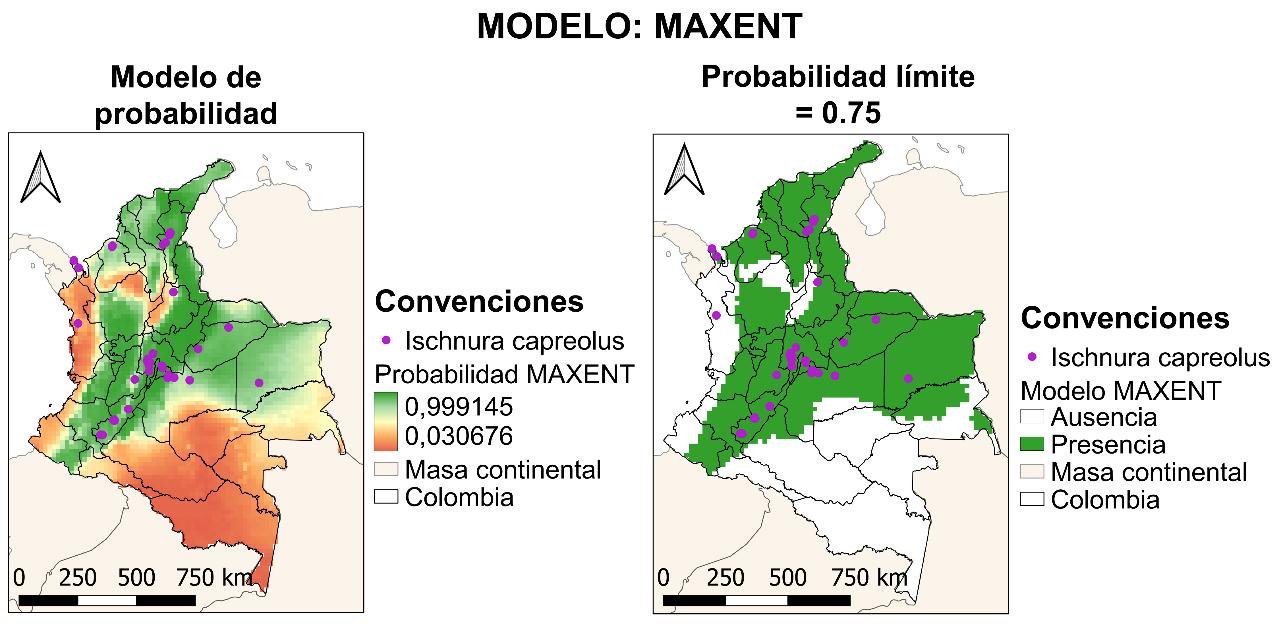
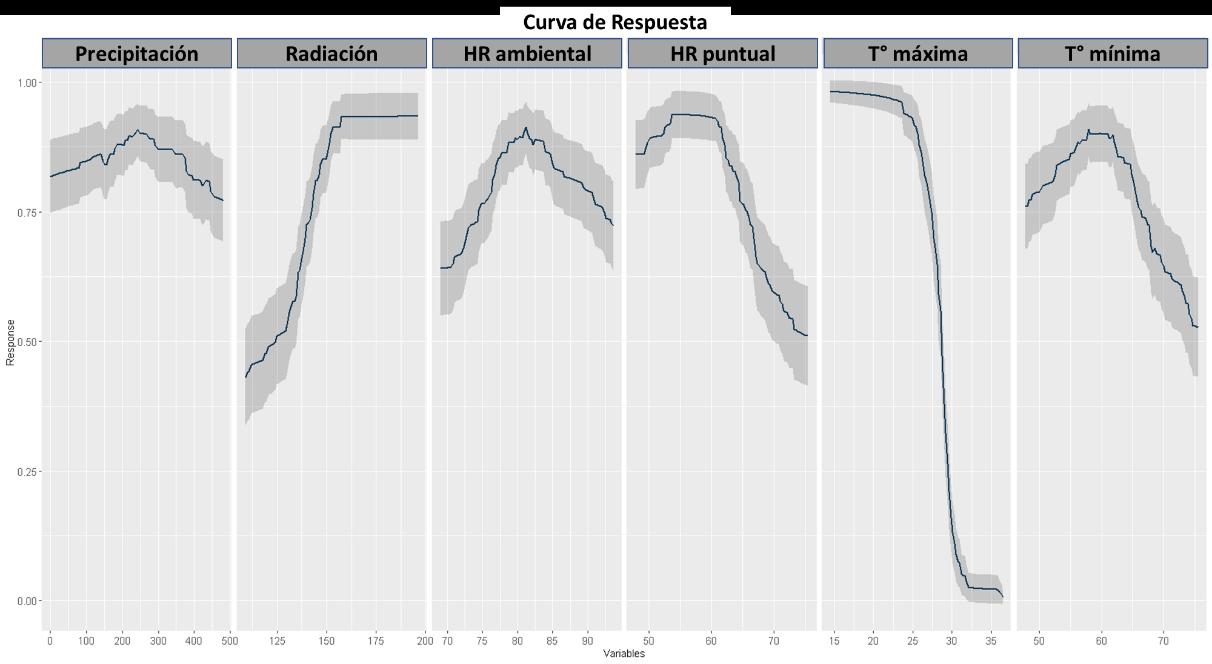
52
Figura 29 Caracterización de las condiciones ambientales según el modelo Maxent
Figura 30 Mapa de distribución potencial construido a partir del método Maxent.
4.2.6 Integrado
A continuación, en la
53
Tabla 7, se muestra en resumen los resultados de los modelos implementados. De estos se rescata que aquel que logró un mejor porcentaje en su capacidad predictiva fue el de GAM, seguido de RPART, ambos con un porcentaje mayor al 90%; contrario los peores modelos fueron el de Maxent y BIOCLIM, con un 76% y 60% respectivamente. Por otro lado, aquellas variables que fueron más constantes en nivel de importancia alta para la constitución de los modelos fue la humedad relativa ambiental y temperatura máxima; mientras que las de menor importancia fueron altamente variantes. Finalmente, de manera general en rangos de distribución, la especie muestra un potencial estadístico de preferencia para la región Andina, Pacífico y Orinoquía; siendo estas regiones altamente heterogéneas, con condicionas ambientales altamente fluctuantes y variables de diversas maneras. Esto recordando las altas lluvias que presenta el pacífico, bajas temperaturas de los Andes, y altas temperaturas de la Orinoquía.
54
Tabla
4.3 Análisis de conectividad ecológica
En la Figura 31, se presenta el mapa de resistencia ecológica para todo Colombia, en el cual se observa que la facilidad de movimiento de la especie no es constante en todo el territorio, sumado a esto, es importante tener en cuenta que la tonalidad cuando va hacia el verde, indica que hay una mayor facilidad de movimiento, y en rojo el desplazamiento es limitado o nulo. Reconociendo que I.capreolus es una especie con una amplia distribución en América del Sur, se evidencia que la cadena montañosa en Colombia es determinante para su desplazamiento y distribución y posiblemente ofrece un punto de conectividad
55
# Modelo Promedio AUC-ROC Variables de mayor importancia Variables de menor importancia Porcentaje de nuevos territorios Regiones geográficas asociadas 1 BIOCLIM 0.595 Radiación Precipitación Humedad relativa ambiental Temperatura máxima 53% Pacífico, Andina, Orinoquía 2 RPART 0.909 Temperatura máxima Radiación Temperatura mínima Humedad relativa puntual 56% Caribe, Pacífico, Andina 3 GAM 0.993 Humedad relativa ambiental Temperatura máxima Humedad relativa puntual Precipitación 66% Caribe, Pacífico, Andina 4 GLM 0.799 Humedad relativa ambiental Temperatura máxima Temperatura mínima Radiación 47% Andina, Caribe 5 Maxent 0.76 Temperatura máxima Humedad relativa puntual Precipitación Humedad relativa ambiental 68% Caribe, Pacífico, Andina, Orinoquía
7 Resumen resultados de los modelos implementados para la predicción de la distribución de la especie Ischnuracapreolus
esencial para el resto del continente. Adicionalmente, se evidencia que es congruente la distribución real con el análisis de resistencia de la especie, ya que esta especie no se muestra en parches aislados, indicando que con la información espacial actualizada a 2017, la especie cuenta con una aceptable conectividad ecológica. Ya que, si bien hay puntos amarillos entre zonas de distribución, este análisis de resistencia más que de la facilidad de movimiento, establece la facilidad de movimiento y el establecimiento de la especie. Por lo tanto, las coloraciones rojas se pueden considerar puntos de aislamiento ecológico total, es decir donde la especie no habita ni puede atravesar, mientras que las zonas amarillas si bien no permiten el establecimiento, tienen cierto gradode permeabilidad al movimiento de la especie.
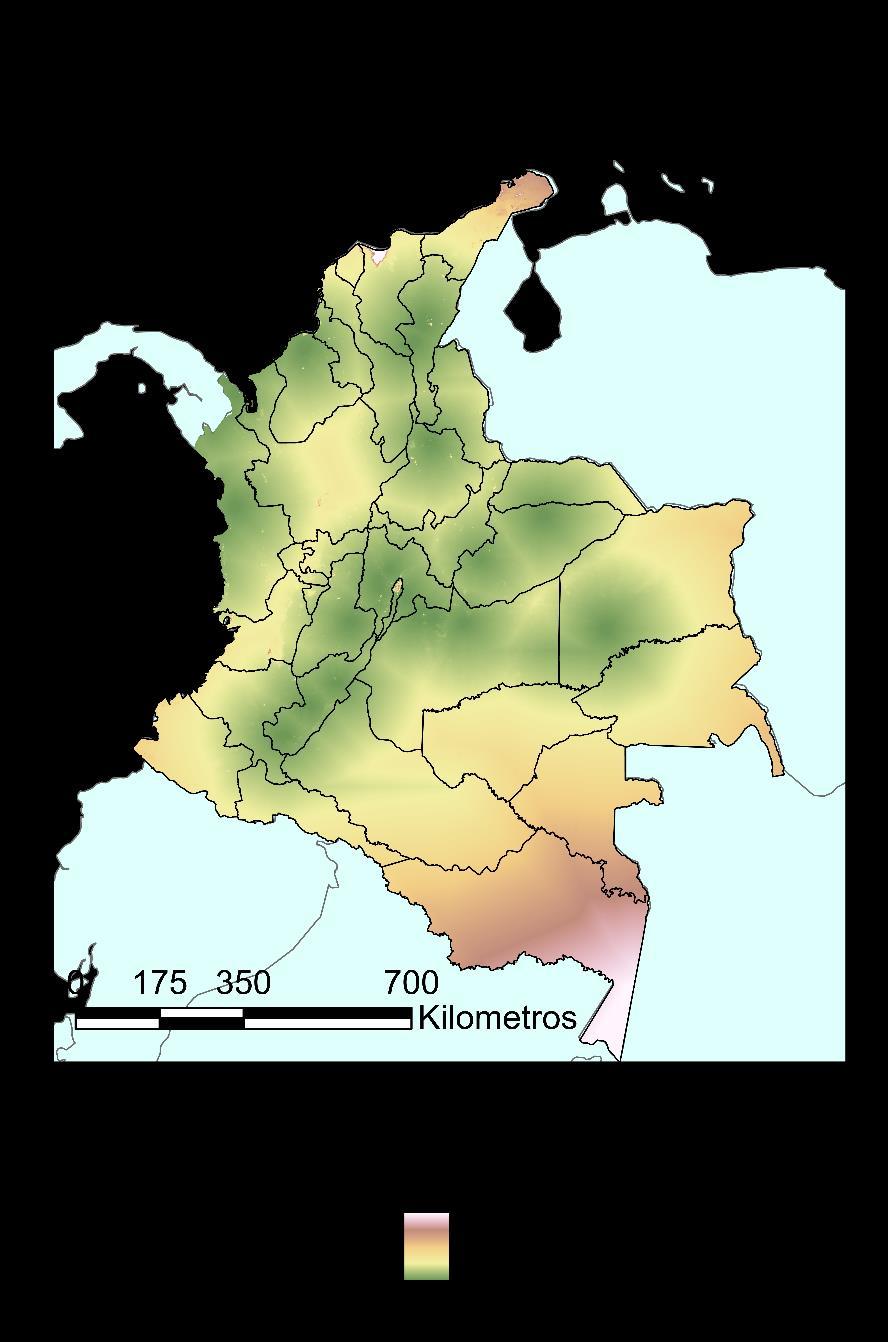
56
Figura 31 Resultado del análisis de resistencia para todo el territorio colombiano.
4.4 Evaluación de los modelos en contexto ecosistémico
El punto anterior presenta de manera general la capacidad de desplazamiento que tiene la especie I.capreolus para la totalidad del territorio colombiano. Sin embargo, este no es el foco principal del trabajo, ya que se busca establecer el potencial de conectividad ecológica que hay en los modelos obtenidos. Para esto, se realizó el álgebra de mapas para cada uno de los resultados de distribución de especies, cuyo resultado se muestra en la Figura 32.
En primer lugar, se puede ver que hay grandes modificaciones entre el análisis de resistencia general para Colombia y los detallados, en función de los modelos de distribución. Esto es un primer indicio que la propuesta metodológica tiene a priori el potencial para la evaluación de los modelos. Si bien, según los modelos el área presentada es la distribución potencial de especies, y de esta manera mediante el análisis de restricción, es posible evaluar la capacidad que tendría la especie si se respetara esta distribución.
Según esto, la efectividad presentada en términos de los resultados estadísticos de los modelos es muy diferente a la encontrada en términos ecológicos. Aquellos modelos que presentan un área más limitada, como GLM, GAM y RPART, tiene una mayor limitación para permitir que la especie se mueva sobre la zona predicha, es decir si bien el modelo tiene unos buenos resultados, cuando se combina con la información ecológica es evidente que las coberturas asociadas al territorio no permiten el desplazamiento, y mucho menos el establecimiento de la especie. Esto indica que el modelo no tiene la capacidad predictiva de la distribución real de la especie, ya que se encuentra asociada con un territorio con baja capacidad de desplazamiento de la especie.
Por otro lado, los modelos de MAXENT y BIOCLIM son los que tienen una extensión más amplia y demuestran una menor restricción general para el desplazamiento de las zonas predichas, indicando, una mayor precisión en la capacidad predictiva del modelo objeto de análisis, ya que estos modelos presentan una mayor congruencia entre la distribución predicha y las características ecológicas. Con esto, se confirma que el área predicha permite que las especies se movilicen según el análisis de conectividad
57
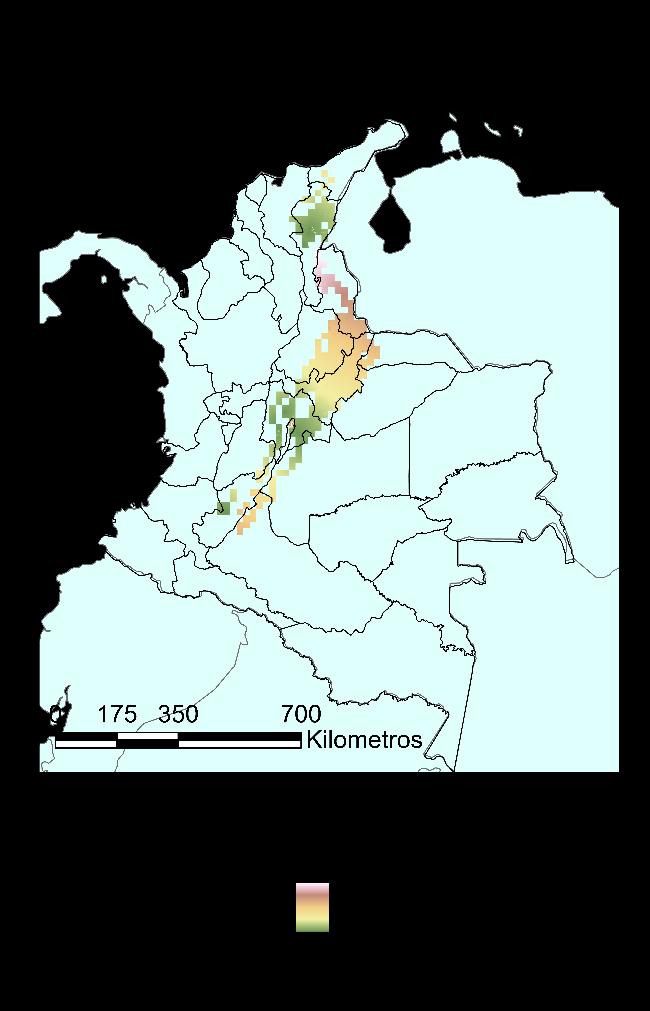
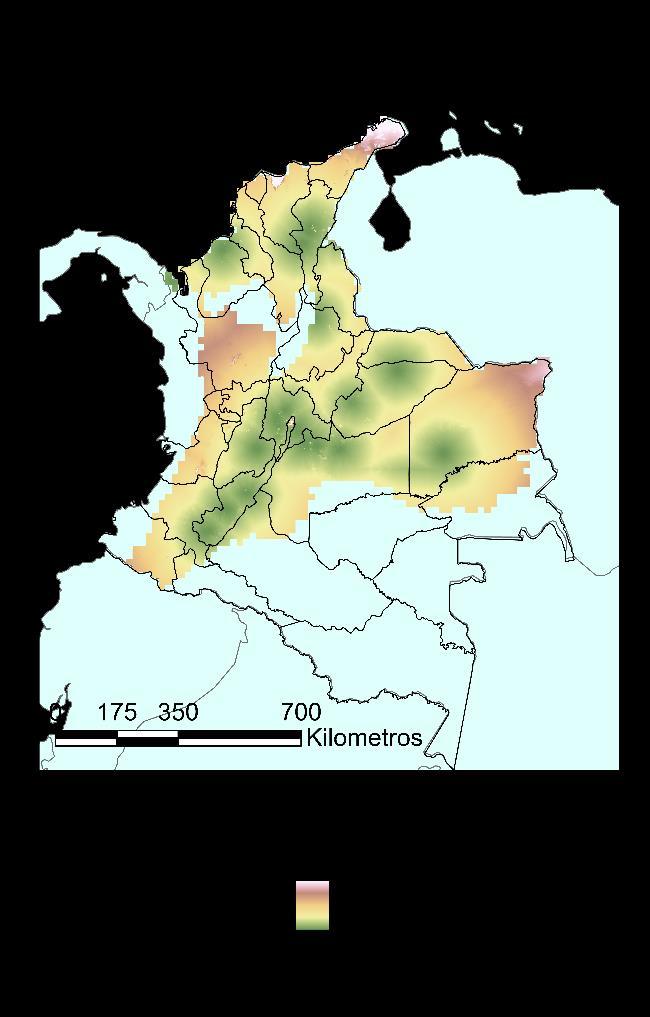
58 A. B.
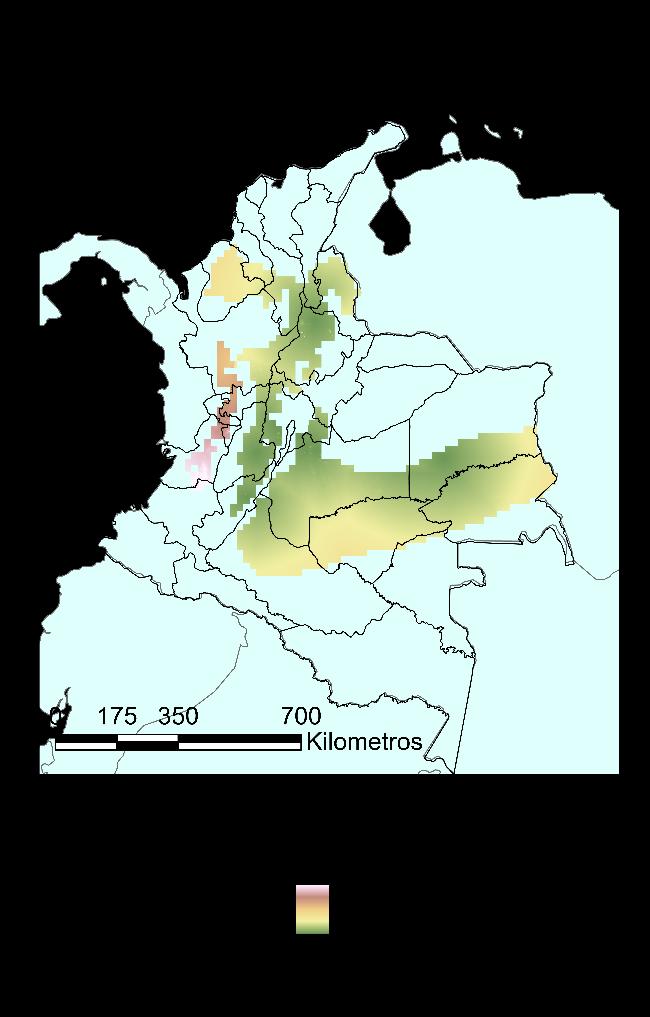
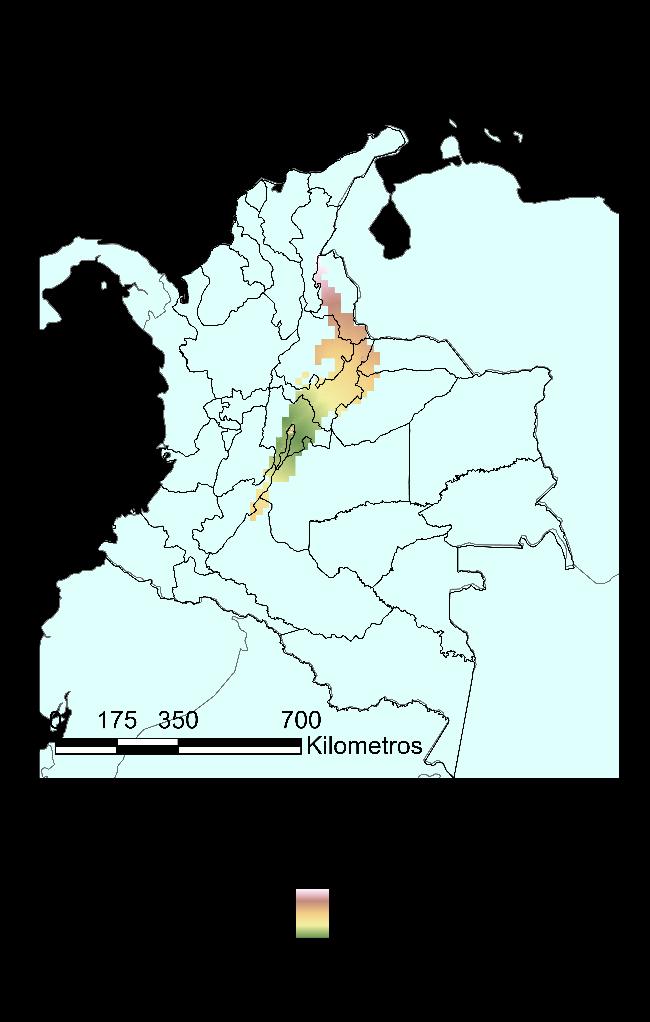
59 C. D.
Figura 32 Se presentan los análisis de resistencia en función de los resultados de modelo de distribución de especie. A. RPART, B. Maxent, C. Bioclim, D. GLM y E. GAM. Las coloraciones más cálidas indican una mayor resistencia ecológica, es decir presencia de barreras, mientras el verde es menor resistencia, zonas de hábitat permeable para la especie.
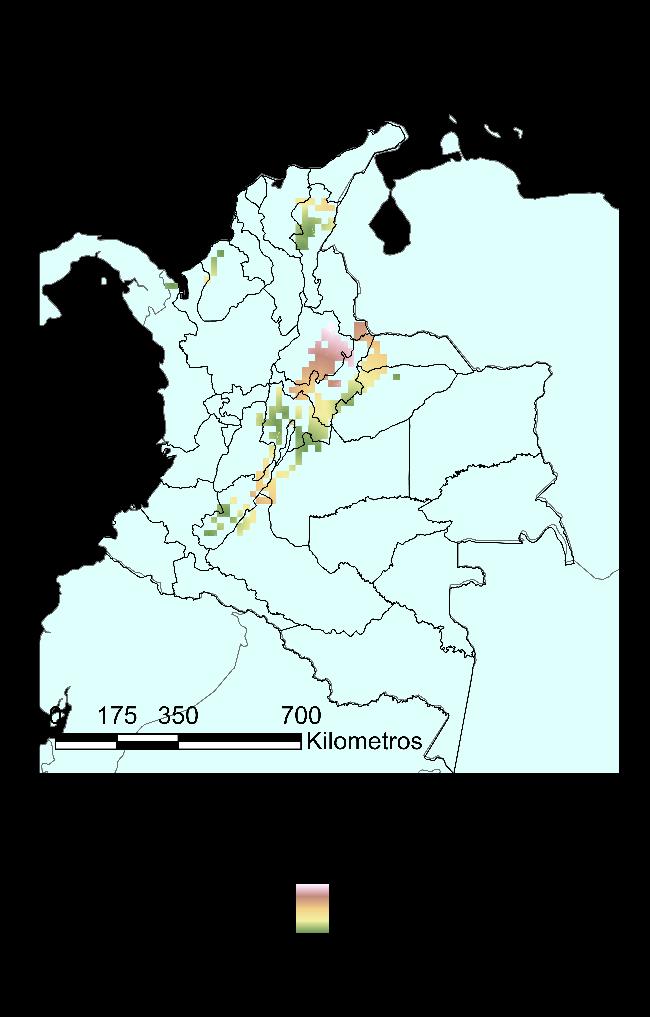
60 E.
5 DISCUSIÓN
El objetivo de esta investigación es proponer una extensión para la evaluación y análisis de los modelos de distribución de especies. Estos permiten de manera rápida y eficiente poner a prueba las condiciones ambientales para determinar la posible distribución de una especie. Y si bien, es posible establecer parámetros de cercanía o continuidad de la distribución, esto no implica que se tenga en cuenta una limitación ecológica, ya que la limitación al movimiento la debe dar la biología de la especie, pero no necesariamente la parametrización del modelo (Lee-Yaw et al., 2022). Con esto, no se busca desestimar o restarles relevancia a los modelos, sino establecer un paso más para su evaluación, es decir, el mecanismo actual (AUC) para evaluarlo en término estadísticos es efectivo, ya que es comparable.
Un modelo de distribución de especies es una herramienta que, si bien luce como una estrategia estadística para entender la ecología de un individuo, por la caracterización de los registros encontrados, el uso de bases de datos debe ser muy cuidadoso. Al comienzo de este estudio, se inició con una consolidación de base de datos secundarias, pero dada la facilidad de acceso al Museo de Historia Natural C.J. Marinkelle de la Universidad de los Andes, se decidió verificar algunos registros que allí reposaban. De la revisión de la colección entomológica, se evidenció que hay un porcentaje considerable de especímenes mal determinados (~35%), por esto al final se logra trabajar con 43 individuos, 19 menos a los encontrados en la base de datos. Esto de manera general para cualquier taxa, debe generar una alta preocupación en tomar la información directamente de fuentes de datos publicadas en línea, dado que estos errores pueden generar sesgos, llevando a conclusiones erróneas sobre la biología e historia natural de una especie. Adicionalmente, representa que se tiene una menor información de las especies, es decir poca representatividad de la especie en el territorio, pese a ser considerada una especie común en el territorio colombiano.
En el caso particular de esta especie para Colombia, se encontró que los errores de clasificación están asociados a la asignación de su especie hermana I. cyane o a un organismo con morfología intermedia, el cual podría llegar a ser un potencial híbrido. Esto indica una inmensa vulnerabilidad en los estudios que implementan reportes en líneas, especialmente cuando hay grupos biológicos con especies crípticas o sin barreras geográficas o reproductivas evolutivamente establecidas, dado que morfológica y filogenéticamente son disruptivas (Newbold, 2010). Para estas especies, es importante no confiar plenamente en las determinaciones o aplicaciones que se implementan de manera
61
comunitaria. Sino tomarse el tiempo de hacer las revisiones en museos, con las limitantes que esto conlleva. Pasar esta consideración por alto podría llevar a sobreestimaciones del rango de distribución potencial de la especie, adicionalmente, se caracterizaría de manera incorrecta las necesidades ambientales de la especie estudiada.
Por otro lado, como parte de la preparación del modelo se suele sugerir en la bibliografía la necesidad de establecer y analizar la colinealidad, para determinar las variables que tienen un alto nivel de correlación, y de esta manera eliminar aquellas variables redundantes, es decir con alto grado de colinealidad. De esta manera, el modelo será más eficiente y con menor redundancia (Feng et al., 2019). Claro está, que esta regla siempre que se tenga criterio ecológico, no es estricto su cumplimiento, por el contrario, es importante incorporarlas. Especialmente, para zonas de estudio como Colombia, el cual tiene una inmensa variedad de microhábitats, además el nivel de escala de las capas implementadas en este estudio ofrece una resolución, que permite tener en cuenta la heterogeneidad característica del territorio colombiano, que, si bien en gran medida respeta factores generalizados como que entre mayor elevación menor temperatura, ciertas zonas tienen un valor intrínseco único que permite que su clima no siga esta constante. Por esta razón, para el presente estudio se respeta la norma de la linealidad, excepto para elevación, temperatura y radiación, que pueden tener variaciones en función de la orogenia del país, pero en mostraron colinealidad entre ellas (Araújo et al., 2019; Sofaer et al., 2019)
Por otro lado, en la evaluación de los modelos se evidencia un área diferencial proyectada para los modelos de distribución. Si bien, son modelos con diferentes fundamentos matemáticos, se usa un dato estadístico comparable, el cual es el valor AUC (Liu et al., 2011), por su traducción, área bajo la curva, donde entre más cercano sea a 1, el modelo presenta una alta precisión y capacidad predictiva. Para los cinco modelos estudiados, el resultado demuestra que son significativamente altos, se evidencia que la proyección de la distribución de especies es precisa en función de los datos de distribución usados para calibrar el modelo. Sin embargo, uno de los riesgos que se tiene en torno a estos resultados es la falta de conocimiento sobre la biología del organismo de estudio. Y, en particular, el conocimiento o antecedentes de la especie permite seleccionar un modelo u otro, más allá de los resultados AUC de cada uno de los modelos Para esto, se debe hacer claridad de la importancia biológica, ya que es común encontrar en la literatura observaciones alrededor de la importancia de un buen número de datos de presencia, o delimitar el área de estudio (Vollering et al., 2019). Sin embargo, es esencial una evaluación en términos ecológicos, por eso, esta investigación busca proponer una metodología que incorpore y establezca la conexión con esta información. Siendo de entrada un factor limitante la falta
62
de conocimiento de una especie. Para el caso de I.capreolus, es una especie que cuenta con información y estudios que permiten entender el comportamiento y requerimientos ecológicos; lo que facilitaría la puesta a prueba de la metodología propuesta para complementar la pertinencia y precisión de los modelos de predicción de especies.
Complementado con el criterio biológico, presentado anteriormente, el mejor modelo predictivo podría ser Bioclim o MAXENT, no sólo porque presenta una mayor área sino porque el área obtenida presenta una mayor continuidad; contrario a los modelos descartados, los cuales son zonas con aislamientos relevantes entre las áreas predichas, causando una gran dificultad o haciendo inviable la conectividad ecológica, lo que de una u otra manera condicionaría la colonización o posibilidad de movilización de una especie sobre un territorio (Booth et al., 2014; Tognelli et al., 2009). Con esta afirmación, entonces es relevante aclarar que el resultado de cualquier modelo de distribución, en realidad establece en función de variables ambientales la distribución potencial, es decir bajo cierto grado de incertidumbre, la especie de interés puede o no habitar la zona, pero este concepto no restringe que la zona modelada o proyectada sea un corredor, es decir, una zona en la que la especie puede cruzar ya sea que habite o no. Por lo tanto, la continuidad indicaría que el área modelada no presenta barreras ecológicas para la especie, en función del área de estudio establecida (Jiménez-Valverde et al., 2008; Miller, 2010). Claro está que la selección del modelo, aunque tenga un principio ecológico, no puede estar fundamentado en un criterio subjetivo del investigador ya que hace que falle el principio de replicabilidad, por esta razón, la metodología propuesta hace a un lado un juicio de valor del investigador y se basa de información verificable y cuantificable (Lieggi et al., 2019).
Retomando la primera pregunta de investigación, la cual busca entender las variables que permiten determinar la distribución de la especie I.capreolus, se evidencia que la totalidad de las variables usadas son relevantes. Dado que, aunque las variables con mayor relevancia no son congruentes entre los diferentes modelos, es un indicio que la información ambiental implementada es relevante para describir y caracterizar las condiciones o lo aspectos relevantes para predecir o estudiar la distribución de la especie Ischnura capreolus Esto dado que para las dos variables con mayor relevancia fueron: radiación y precipitación (BIOCLIM), temperatura y radiación (RPART), humedad y temperatura (GAM), humedad y temperatura (GLM) y temperatura y humedad (MAXENT).
De esto se puede concluir que, pese a que hay una colinealidad aparente, las variables con mayor relevancia no son colineales, es decir, que se corrobora que el criterio ecológico fundamentado en la heterogénea orogenia colombiana para no eliminar variables es válido. Adicionalmente, se tiene como respuesta que las variables con mayor representatividad
63
son aquellas que definen y determinan el ciclo de vida de una especie ectotérmica, con variables como la temperatura y humedad (Koch, 2021; Tognelli et al., 2009). Si la relación lineal hubiera mostrado un inconveniente para el modelo de predicción, variables como la elevación y temperatura se encontrarían juntas o elevación y radiación; siendo esto un argumento esencial donde para Colombia hay una variedad de microhábitats esenciales que no responden de ninguna manera a tendencias generales (Achury et al., 2012), sino que esos aspectos que discrepan la colinealidad son información de la biología de la especie.
Por otro lado, para la segunda pregunta de investigación, sobre el uso de modelos de distribución de especies para aumentar el conocimiento, se evidencia que en los cinco modelos hay un aumento de la distribución conocida de la especie, por lo tanto, estos modelos han permitido conocer más sobre los nichos potenciales en Colombia. Si bien, es un modelo y es esencial validar el modelo con visita en campo, es un punto de partida que optimizaría recursos y tiempo para hacer seguimiento o análisis genéticos según el interés de los investigadores.
Retomando la pregunta de investigación a partir de los resultados obtenidos, es claro que un modelo basado de manera exclusiva en la similitud estadística no es suficiente para determinar la distribución real de una especie. Es importante conectar esta información, con la capacidad o permeabilidad que tiene el área modelada para que la especie se pueda desplazar por el área predicha inicialmente. Por otro lado, con la hipótesis establecida, los cinco modelos evaluados tienen una significativa capacidad predictiva de la especie I.capreolus, sin embargo, no tuvieron la misma capacidad de reconocer la permeabilidad de la especie para el área predicha. En función de ambas metodologías, aquellos modelos que mejor responde a la predicción y evaluación del movimiento de la especie fueron MAXENT y BIOCLIM, esto para la especie I.capreolus. Con esto se establece que la hipótesis no se rechaza, ya que se pudo comprobar que los modelos de distribución de especies no tienen en cuenta las barreras ecológicas en el área modelada, pero con la propuesta metodológica, se puede mitigar esta incertidumbre.
En la Figura 33 se presentan los mapas de resistencia transformados como variables discretas, esto con el objetivo de ver claramente la permeabilidad efectiva del área predicha por los diferentes modelos. Con este complemento visual, se evidencia que, de manera general, las áreas proyectadas por los modelos cumplen con los criterios de permeabilidad, para la especie. Es decir que son zonas de movilidad para la especie en función de criterios ecológicos. Como es usual, se aclara que es imposible predecir si es el nicho o espacio
64
donde la especie pueda establecerse, pero si es una aproximación al nicho fundamental de la especie, es decir donde podría establecerse o dónde podría cruzar.
Uno de los retos para la construcción de estos mapas es el establecimiento del valor límite para categorizar la potencial área habitable en función de la variable ecosistémica, ya que en la literatura no hay información que establezca un sitio potencial. Esto posiblemente se deba a que, en un análisis ecológico, es suficiente e informativo ya que las preguntas de investigación van en torno a la capacidad que tiene la especie de atravesar un área o zona de estudio (Brooks, 2003; Vogt et al., 2009). Sin embargo, para este caso, se estableció un intervalo de confianza del 95%, usado de manera estándar, para así determinar la permeabilidad de la zona. Esto permite soportar que las áreas con mayor permeabilidad son aquellas que son de mayor extensión, ya que presentan el área con mayor continuidad luego del análisis de conectividad (Figura 33-B y Figura 33-C). Adicionalmente, es importante establecer, que el modelo resultado de GLM (Figura 33-D), presenta un problema serio de continuidad, disminuyendo la viabilidad del área realmente que representa el nicho potencial para la especie. Los modelos restantes la continuidad no fue afectada, sin embargo, ya presentaban aislamientos desde la consolidación del modelo, es decir que el mismo modelo establece las limitaciones en la conectividad ecológica.
Como complemente, en la Tabla 8 se presenta el área permeable con que permite la conectividad en función del área total establecida por el modelo. En el cual se evidencia que la mayor parte de la proyección en todos los modelos permite que exista permeabilidad de la especie. Siendo esto un punto importante para establecer que para la especie I.capreolus los modelos de predicción de especie, guardan congruencia con la conectividad ecológica. Con esto, se logra responder la tercera pregunta de investigación, sobre el aporte de los estudios de conectividad ecológica del área predicha, ya que la reducción del área predicha comprueba las limitaciones de los modelos de distribución de especie.
65
Alta restricción ecológica Baja restricción ecológica RPART GLM GAM MAXENT BIOCLIM 90% 97% 100% 88% 96%
Tabla 8 Porcentaje de permeabilidad de la especie, respecto al área predicha por cada modelo de distribución presentado.
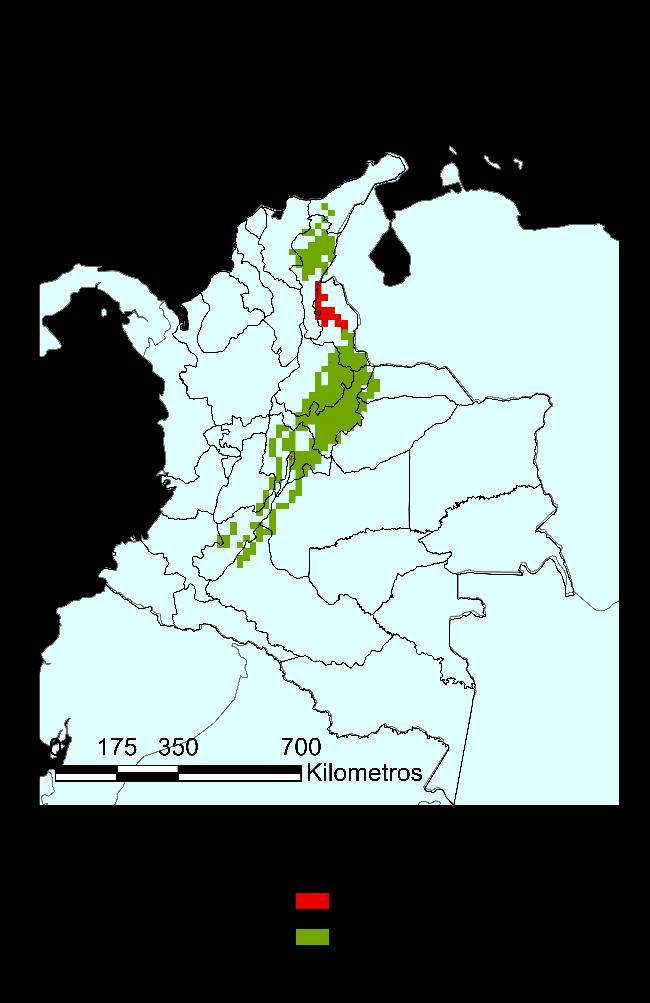
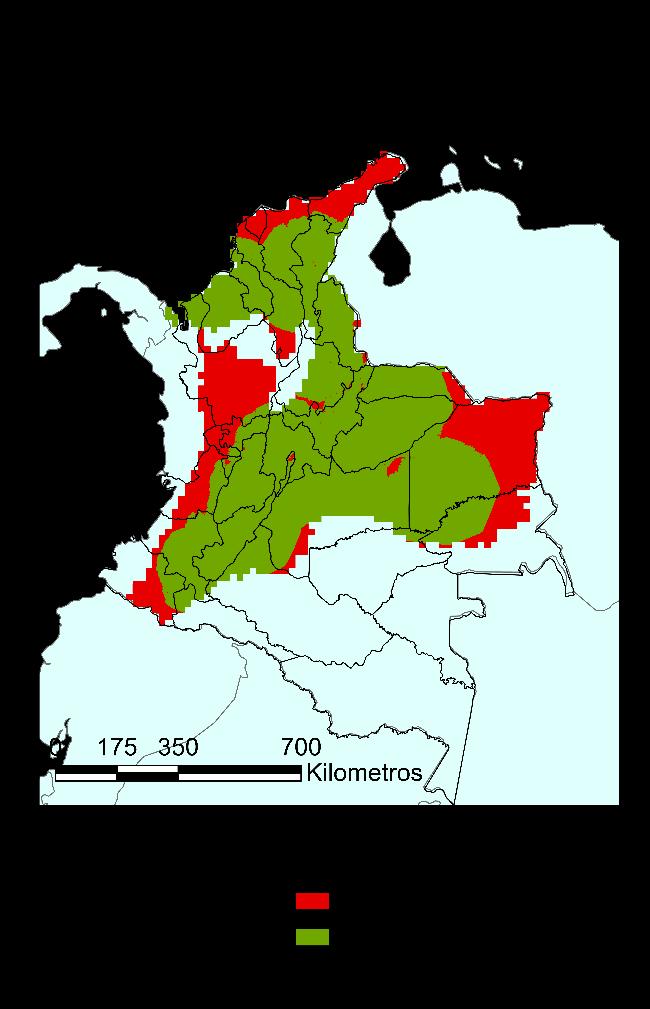
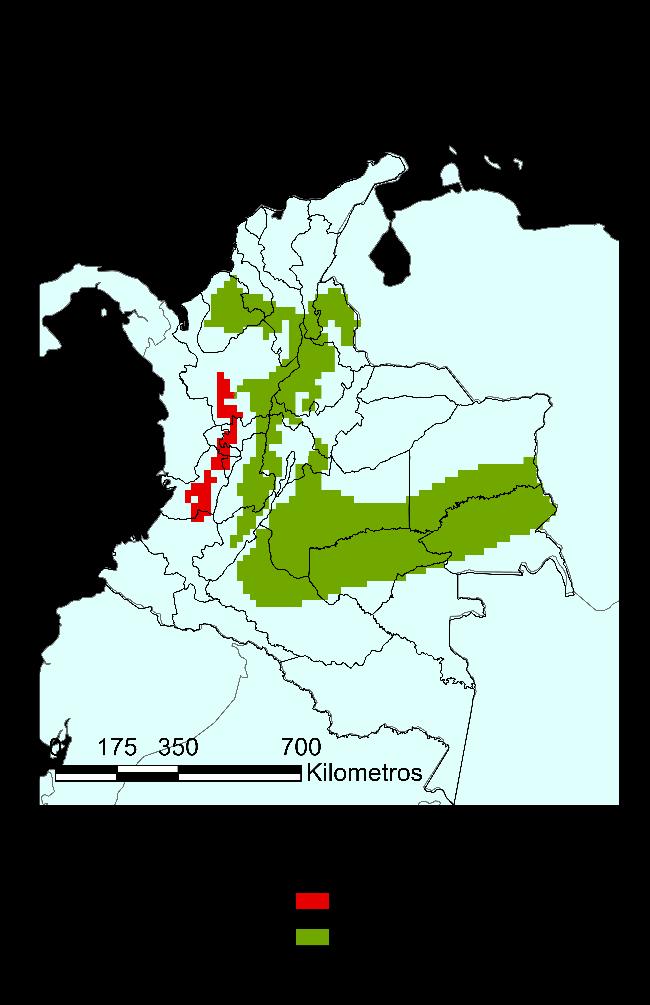
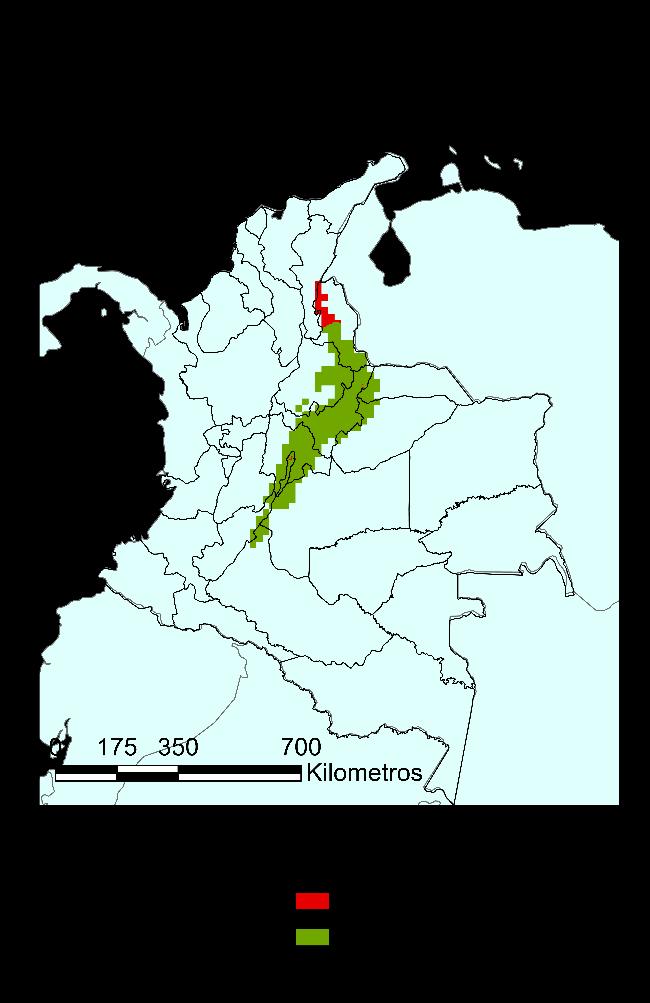
66
A.
B.
C. D.
A. RPART, B. Maxent, C. Bioclim, D. GLM y E. GAM. En rojo se presentan las zonas con alta restricción ecológica y en verde las zonas sin restricciones ecológicas
Finalmente, y conectando el modelo de distribución y el análisis ecológico; la distribución potencial de la especie de interés corresponde estrictamente al área verde presentada en el análisis consolidado presentado en la Figura 33
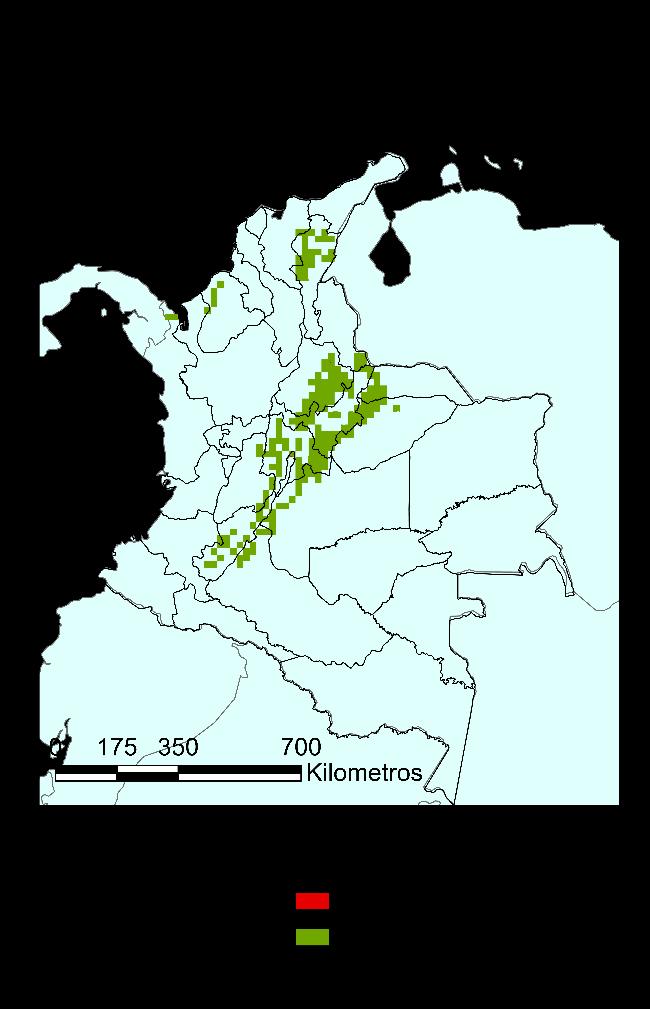
67 E.
Figura 33 Se presentan los análisis de resistencia en función de los resultados de modelo de distribución de especie, como variables categóricas, de permeable o barrera ecológica
6 CONCLUSIONES
Los modelos de distribución de especies implementados de manera tradicional ofrecen información relevante acerca de las características de la especie objeto de interés. Son un mecanismo estadístico falsable, acerca de las características y potenciales limitaciones de la especie. Sin embargo, cuando se tiene una amplia información de la especie, es posible complementar los análisis y de esta manera poner a prueba los modelos de distribución a partir de análisis ecológicos. Claro está, si no hay suficiente información los modelos serán la única manera de predecir el nicho potencial. Para la especie objeto de interés, I.capreolus, al contar con información suficiente y aprovechable, se puso a prueba esta metodología. La cual permitió establecer la importancia de implementar una estrategia complementaria, ya que el área predicha, no necesariamente es un área que la especie pueda cruzar, es decir, hay hábitat que restringe el paso de la especie, por lo que de ninguna manera corresponde el área modelada al nicho fundamental de la especie. Adicionalmente, Colombia cuenta con un gran banco de datos actualizados, que describen las características del territorio, es decir, que la limitación de la aplicación de esta metodología corresponde al conocimiento de la especie de interés y de los estudios previos sobre el área de estudio; es decir, que esta metodología es altamente restrictiva
Por otro lado, el presente documento, logra demostrar que la metodología aquí presentada tiene una importante aplicabilidad en especies con amplia distribución y un conocimiento considerable en su biología, desde el comportamiento de dispersión hasta requerimientos ecológicos, más allá de las variables ambientales usadas como parámetros del modelo de distribución. Siendo esencial el logro de combinar efectivamente variables tanto categóricas como numéricas para así establecer el nicho fundamental de una especie. Ya que como se evidenció, cierta área predicha cumple como barrera para la especie; siendo esto un factor de gran relevancia para territorios con una alta heterogeneidad de hábitat, como Colombia.
Finalmente, se debe probar la herramienta en diferentes grupos biológicos, con especial énfasis en especies que tengan importantes limitaciones de movilidad o de hábitat, esto con el fin de evaluar la metodología en organismos con amplios requerimientos. Por otra parte, es posible que esta metodología no sea aplicable en plantas ni hongos, ya que el análisis puede verse afectado por condiciones in situ muy particulares, lo que sesgaría los resultados y sería un ejercicio sin relevancia. Esto dado que los insectos presentan menos barreras ecológicas y además la especie seleccionada al ser de amplia distribución, puede moverse y establecerse en una gran variedad de hábitats, lo que para un primer ejercicio
68
es conveniente, ya que permitiría evidenciar las limitaciones de los modelos de distribución de especies, es decir que el área proyectada presenta restricciones que por el tipo de parámetros usados son enmascarados.
69
1. Achury, R., Chacón de Ulloa, P., y Arcila, Á. (2012). Effects of the heterogeneity of the landscape and the abundance of Wasmannia auropunctata on ground ant assemblages in a Colombian tropical dry forest. Psyche, 2012
2. Agudelo, M. F. C. (2016). EcohydrologyofparamosinColombia:: vulnerability to climate change and land use. Escuela de Geociencias y Medio Ambiente.
3. Araújo, M.B., Anderson, R. P., Márcia Barbosa, A., Beale, C. M., Dormann, C. F., Early, R., Garcia, R. A., Guisan, A., Maiorano, L., Naimi, B., O’Hara, R. B., Zimmermann, N. E., y Rahbek, C. (2019). Standards for distribution models in biodiversity assessments. ScienceAdvances, 5(1), eaat4858. https://doi.org/10.1126/sciadv.aat4858
4. Araújo, M., y Guisan, A. (2006). Araujo MB, Guisan A. Five (or so) challenges for species distribution modelling. Journal of Biogeography. Journal of Biogeography, 33, 1677-1688. https://doi.org/10.1111/j.1365-2699.2006.01584.x
5. Austin, M. (2007). Species distribution models and ecological theory: A critical assessment and some possible new approaches. EcologicalModelling, 200(1-2), 1-19. https://doi.org/10.1016/j.ecolmodel.2006.07.005
6. Barrera-Cataño, J. I., Garibello, J., Moreno-Cárdenas, C., y Basto, S. (s. f.). Trade offs at applying tree nucleation to restore degraded high andean forests in colombia. RestorationEcology, n/a(n/a), e13753. https://doi.org/10.1111/rec.13753
7. Barve, N., Barve, V., Jiménez-Valverde, A., Lira-Noriega, A., Maher, S. P., Peterson, A. T., y Villalobos, F. (2011). The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecologicalmodelling, 222(11), 1810-1819.
8. Begon, M., Townsend, C. R., y Harper, J. L. (2006). Ecology: From individuals to ecosystems. Blackwell.
9. Bennett, A. (2010). The role of soil community biodiversity in insect biodiversity. Insect Conservation and Diversity, 3(3), 157-171. https://doi.org/10.1111/j.17524598.2010.00086.x
70
7 REFERENCIAS
10. Booth, T. H., Nix, H. A., Busby, J. R., y Hutchinson, M. F. (2014). BIOCLIM: the first species distribution modelling package, its early applications and relevance to most current MAXENT studies. DiversityandDistributions, 20(1), 1-9.
11. Borror, D. J., Triplehorn, C. A., y Johnson, N. F. (1992). Study of insects. Saunders College Publishing Harcourt Brace College Publishers.
12. Bota-Sierra, C., Velasquez-Vélez, M., y Realpe, E. (2019). A new species of Ischnura from the Colombian Central Andes (Odonata: Coenagrionidae). Odonatologica, 48, 115-132.
https://doi.org/10.5281/zenodo.2677697
13. Booth, T. H. (2018). Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral ecology, 43(8), 852-860
14. Brodie, S. J., Thorson, J. T., Carroll, G., Hazen, E. L., Bograd, S., Haltuch, M. A.,y Selden, R. L. (2020). Trade‐offs in covariate selection for species distribution models: a methodological comparison. Ecography, 43(1), 11-24.
15. Brooks, C. P. (2003). A scalar analysis of landscape connectivity. Oikos, 433-439.
16. Brown, V. K. (1995). The insects. An outline of entomology. By P. J. Gullan and P. S. Cranston. (London: Chapman y Hall, 1994). x+491 pp. Soft cover £24.99. ISBN 0-41249360-8. Bulletin of Entomological Research, 85(2), 302-302.
https://doi.org/10.1017/S0007485300034428
17. Bussmann, R. W., Paniagua Zambrana, N. Y., Romero, C., y Hart, R. E. (2018). Astonishing diversity The medicinal plant markets of Bogotá, Colombia. Journal of Ethnobiology andEthnomedicine, 14(1), 43. https://doi.org/10.1186/s13002-018-02418
18. Calow, P. (1987). Towards a Definition of Functional Ecology. FunctionalEcology, 1(1), 57. https://doi.org/10.2307/2389358
19. Cardozo, J. A. C., Guzmán, E. A., Arriero, L. B., Garnica, A. C., Capera, D., Ocampo, C., Ruiz, M. O., Aceldas, S. R., y Sánchez, F. (2016). Composición y riqueza de odonatos en tres humedales artificiales suburbanos en Cajicá, Cundinamarca, Colombia. RevistaBiodiversidadNeotropical, 6(2), 164-170.
71
20. Chichorro, F., Juslén, A., y Cardoso, P. (2019). A review of the relation between species traits and extinction risk. Biological Conservation, 237, 220-229. https://doi.org/10.1016/j.biocon.2019.07.001
21. Collins, S. D., y McIntyre, N. E. (2015). Modeling the distribution of odonates: A review. FreshwaterScience, 34(3), 1144-1158. https://doi.org/10.1086/682688
22. Cordero-Rivera, A., y Andrés, J. A. (2002). Male coercion and convenience polyandry in a Calopterygid damselfly (Odonata). J. Insect Sci, 2, 14.
23. Cordero-Rivera, A. C., y Pérez, F. E. (1998). Mating frequency, population density and female polychromatism in the damselfly Ischnura graellsii: an analysis of four natural populations. Etología, 6, 61-67.
24. Diekötter, T., Billeter, R., y Crist, T. O. (2008). Effects of landscape connectivity on the spatial distribution of insect diversity in agricultural mosaic landscapes. Basic and Applied Ecology, 9(3), 298–307.
25. Dormann, C. F., Schymanski, S. J., Cabral, J., Chuine, I., Graham, C., Hartig, F., Kearney, M., Morin, X., Römermann, C., Schröder, B., y Singer, A. (2012). Correlation and process in species distribution models: Bridging a dichotomy: Bridging the correlation-process dichotomy. Journal of Biogeography, 39(12), 2119-2131.
https://doi.org/10.1111/j.1365-2699.2011.02659.x
26. Elith, J., y Graham, C. H. (2009). Do they? How do they? WHY do they differ? On finding reasons for differing performances of species distribution models. Ecography, 32(1), 6677.
https://doi.org/10.1111/j.1600-0587.2008.05505.x
27. Ellis, D H., y Pfeiffer, T J (1990). Ecology, life cycle, and infectious propagule of Cryptococcus neoformans. The Lancet, 336(8720), 923-925.
https://doi.org/10.1016/0140-6736(90)92283-N
28. Feng, X., Park, D. S., Walker, C., Peterson, A. T., Merow, C., y Papeş, M. (2019). A checklist for maximizing reproducibility of ecological niche models. Nature Ecology y Evolution, 3(10), 1382-1395. https://doi.org/10.1038/s41559-019-0972-5
72
29. Finch, J. M., Samways, M. J., Hill, T. R., Piper, S. E., y Taylor, S. (2006). Application of predictive distribution modelling to invertebrates: Odonata in South Africa. Biodiversity & Conservation, 15(13), 4239-4251.
30. Fischer, J., y Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: A synthesis. Global Ecology and Biogeography, 16(3), 265-280.
https://doi.org/10.1111/j.1466-8238.2007.00287.x
31. Gallien. (2012). Invasive species distribution models – how violating the equilibrium assumption can create new insights Gallien 2012 Global Ecology and Biogeography Wiley Online Library
https://onlinelibrary.wiley.com/doi/full/10.1111/j.1466-8238.2012.00768.x
32. Galindo-Ruiz, N., Velasquez-Velez, M. I., Cano-Cobos, Y., Sanchéz-Guillén, R. A., y Realpe, E. (2019). Description of a putative hybrid between Ischnura cyane and I. capreolus from Colombia (Odonata: Coenagrionidae). Notulae odonatologicae, 9(4), 125-172.
33. Garrison, R. W., Ellenrieder, N. V., y Louton, J. A. (2010). Damselfly genera of the New World. Johns Hopkins University Press.
34. Granberg, M., Lidskog, R., y& Larsson, S. (2008). Dealing with uncertainty: a case study of controlling insect populations in natural ecosystems. Local environment, 13(7), 641652.
35. Guisan, A., y Thuiller, W. (2005). Predicting species distribution: Offering more than simple habitat models. Ecology Letters, 8(9), 993-1009. https://doi.org/10.1111/j.14610248.2005.00792.x
36. Hardersen, S. (2000). The role of behavioural ecology of damselflies in the use of fluctuating asymmetry as a bioindicator of water pollution. Ecological Entomology, 25(1), 45-53.
37. Heckman, C. W. (2006). Encyclopedia of South American aquatic insects: Odonata–Anisoptera: Illustrated keys to known families, genera, and species in South America. Dordrecht: Springer Netherlands.
73
38. Imai, I., y Yamaguchi, M. (2012). Life cycle, physiology, ecology and red tide occurrences of the fish-killing raphidophyte Chattonella. Harmful Algae, 14, 46-70.
https://doi.org/10.1016/j.hal.2011.10.014
39. Jiménez-Valverde, A., Lobo, J. M., y Hortal, J. (2008). Not as good as they seem: The importance of concepts in species distribution modelling. Diversity and Distributions, 14(6), 885-890. https://doi.org/10.1111/j.1472-4642.2008.00496.x
40. Keys, P., Galaz, V., Dyer, M., Matthews, N., Folke, C., Nyström, M., y Cornell, S. (2019). Anthropocene risk Nature Sustainability, 2(8), 667-673.
https://doi.org/10.1038/s41893-019-0327-x
41. Koch, F. H. (2021). Considerations regarding species distribution models for forest insects. AgriculturalandForestEntomology, 23(4), 393-399.
42. Kriticos, D. J., Webber, B. L., Leriche, A., Ota, N., Macadam, I., Bathols, J., y Scott, J. K. (2012). CliMond: Global high resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods in Ecology and Evolution, 3, 53-64.
https://doi.org/10.1111/j.2041-210X.2011.00134.x
43. Lee-Yaw, J. A., L. McCune, J., Pironon, S., y N. Sheth, S. (2022). Species distribution models rarely predict the biology of real populations. Ecography, 2022(6), e05877.
https://doi.org/10.1111/ecog.05877
44. Levine, D., y Stephan, D. (2010). Evenyoucanlearnstatistics:aguideforeveryonewho haseverbeenafraidofstatistics (Second Edition). Pearson Education.
45. Lieggi, C., Kalueff, A. V., Lawrence, C., y Collymore, C. (2019). The influence of behavioral, social, and environmental factors on reproducibility and replicability in aquatic animal models. ILARjournal, 60(2), 270-288.
46. Liu, C., White, M., y Newell, G. (2011). Measuring and comparing the accuracy of species distribution models with presence-absence data. Ecography, 34(2), 232-243.
https://doi.org/10.1111/j.1600-0587.2010.06354.x
47. Lobo, J. M. (2016). The use of occurrence data to predict the effects of climate change on insects. Current Opinion in Insect Science, 17, 62-68.
74
48. Mace, G. M., Collar, N. J., Gaston, K. J., Hilton-Taylor, C., Akçakaya, H. R., LeaderWilliams, N., Milner-Gulland, E. J., y Stuart, S. N. (2008). Quantification of Extinction Risk: IUCN’s System for Classifying Threatened Species. ConservationBiology, 22(6), 1424-1442.
https://doi.org/10.1111/j.1523-1739.2008.01044.x
49. Mateo, R. G., Felicísimo, Á. M., y Muñoz, J. (2011). Modelos de distribución de especies: Una revisión sintética. Revista chilena de historia natural, 84(2), 217-240.
https://doi.org/10.4067/S0716-078X2011000200008
50. McCafferty, W. P. (1983). Aquatic Entomology: The Fishermen’s and Ecologists’ IllustratedGuidetoInsectsandTheirRelatives. Jones y Bartlett Learning.
51. McRae, B. H., Shah, V., y Edelman, A. (2016). Circuitscape: modeling landscape connectivity to promote conservation and human health. The Nature Conservancy, 14.
52. Miller, J. (2010). Species distribution modeling. GeographyCompass, 4(6), 490-509.
53. Misof, B., Liu, S., Meusemann, K., Peters, R. S., Donath, A., Mayer, C., Frandsen, P. B., Ware, J., Flouri, T., Beutel, R. G., Niehuis, O., Petersen, M., Izquierdo-Carrasco, F., Wappler, T., Rust, J., Aberer, A. J., Aspöck, U., Aspöck, H., Bartel, D., … Zhou, X. (2014). Phylogenomics resolves the timing and pattern of insect evolution. Science, 346(6210), 763-767.
https://doi.org/10.1126/science.1257570
54. Naimi, B., y Araújo, M. B. (2020). Species Distribution Modelling SDM (1.0-89) [Rproject].
http://biogeoinformatics.org/
55. Neuhoff, K., y Helm, D. (2016). Review of Natural Capital. Valuing the Planet, HelmDieter. EconomicsofEnergyyEnvironmentalPolicy, 5(1), 158-161.
56. Newbold, T. (2010). Applications and limitations of museum data for conservation and ecology, with particular attention to species distribution models. Progress in Physical Geography: Earth and Environment, 34(1), 3-22.
https://doi.org/10.1177/0309133309355630
57. Nezer, O., Bar-David, S., Gueta, T., y Carmel, Y. (2017). High-resolution speciesdistribution model based on systematic sampling and indirect observations. Biodiversity and Conservation, 26(2), 421-437.
75
58. Okada, Y., Suzaki, Y., Miyatake, T., y Okada, K. (2012). Effect of weapon-supportive traits on fighting success in armed insects. Animal Behaviour, 83, 1001-1006. https://doi.org/10.1016/j.anbehav.2012.01.021
59. Olcina Cantos, J. (1996). La geografía hoy: Reflexiones sobre el pensamiento geográfico, la región y la docencia de la geografía. InvestigacionesGeográficas, 16, 93. https://doi.org/10.14198/INGEO1996.16.06
60. Ortiz-Moreno, M. (2014). ANÁLISE DA INTERAÇÃO ENTRE ORDENAMENTO TERRITORIALEBIODIVERSIDADE:ESTUDODECASOEMVILLAVICENCIO(META, COLÔMBIA) PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E RECURSOSNATURAIS. UNIVERSIDADE FEDERAL DE SAO CARLOS
61. Paillard, C. (2016). An Ecological Approach to Understanding Host-PathogenEnvironment Interactions: The Case of Brown Ring Disease in Clams. En J. L. Romalde (Ed.), Oysters and Clams: Cultivation, Habitat Threats and Ecological Impact Nova Science Publishers.
62. Paulson, D., Schorr, M., y Deliry, C. (2021). World Odonata List. Last revision, 17.
63. Pearson, R. G., y Dawson, T. P. (2003). Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? GlobalEcologyand Biogeography, 12(5), 361-371. https://doi.org/10.1046/j.1466-822X.2003.00042.x
64. Pérez-Gutiérrez, L. A., y Palacino-Rodríguez, F. (2011). Updated checklist of the Odonata known from Colombia. Odonatologica, 40(3), 203.
65. Polhemus, D. A. (1993). Conservation of Aquatic Insects: Worldwide Crisis or Localized Threats? AmericanZoologist, 33(6), 588-598. https://doi.org/10.1093/icb/33.6.588
66. QGIS Development Team. (2009). QGIS Geographic Information System (3.14.16) [Open Source]. Geospatial Foundation. http://qgis.org/
76
67. Renner, S., Sahlén, G., y Périco, E. (2015). Testing Dragonflies as Species Richness Indicators in a Fragmented Subtropical Atlantic Forest Environment. Neotropical Entomology, 45. https://doi.org/10.1007/s13744-015-0355-9
68. Rowe, L., y Ludwig, D. (1991). Size and Timing of Metamorphosis in Complex Life Cycles: Time Constraints and Variation. Ecology, 72, 413-427.
https://doi.org/10.2307/2937184
69. Saaty, R. W. (1987). THE ANALYTIC HIERARCHY PROCESS-WHAT IT IS AND HOW IT IS USED. MathModelling, 9(3-5), 161-176.
70. Sánchez-Guillén, R. A., Arce-Valdés, L. R., Swaegers, J., Chauhan, P., Cávez-Rios, J., Rivas-Torres, A., Wellenreuther, M., y Hansson, B. (2022). Variable genomic patterns of hybridization in an expanding hybrid zone of damselflies [Preprint]. Preprints.
https://doi.org/10.22541/au.164628976.68855627/v1
71. Sánchez‐Guillén, R., Cordero-Rivera, A., Rivas Torres, A., Wellenreuther, M., Bybee, S., Hansson, B., Velasquez-Vélez, M., Realpe, E., Chávez-Ríos, J., Villalobos, F., y Dumont, H. (2020). The evolutionary history of colour polymorphism in Ischnura damselflies (Odonata: Coenagrionidae). Odonatologica, 49, 333-370.
72. Sánchez‐Guillén, R. A., Córdoba‐Aguilar, A., Hansson, B., Ott, J., y Wellenreuther, M. (2016). Evolutionary consequences of climate‐induced range shifts in insects. Biological Reviews, 91(4), 1050-1064.
73. Sánchez-Guillén, R. A., Hansson, B., Wellenreuther, M., Svensson, E. I., y CorderoRivera, A. (2011). The influence of stochastic and selective forces in the population divergence of female colour polymorphism in damselflies of the genus Ischnura. Heredity, 107(6), 513-522.
74. Sánchez-Guillén, R. A., Muñoz, J., Rodríguez-Tapia, G., Feria Arroyo, T. P., y CórdobaAguilar, A. (2013). Climate-Induced Range Shifts and Possible Hybridisation Consequences in Insects. PLoS ONE, 8(11).
75. Sanmartín-Villar, I., Zhang, H. M., y Cordero-Rivera, A. (2016). Colour polymorphism and ontogenetic colour changes in Ischnura rufostigma (Odonata: Coenagrionidae). Odonatologica, 45(1-2), 77-86.
77
76. Segurado, P., y Araujo, M. B. (2004). An evaluation of methods for modelling species distributions. Journal of biogeography, 31(10), 1555-1568.
77. Slobodkin, L. B. (1962). Energy in Animal Ecology. En J. B. Cragg (Ed.), Advances in Ecological Research (Vol. 1, pp. 69-101). Academic Press.
https://doi.org/10.1016/S0065-2504(08)60301-3
78. Smith, E. H., y Kennedy, G. G. (2009). Chapter 119 History of Entomology. En V. H. Resh y R. T. Cardé (Eds.), Encyclopedia of Insects (Second Edition) (pp. 449-458). Academic Press. https://doi.org/10.1016/B978-0-12-374144-8.00128-4
79. Sofaer, H. R., Jarnevich, C. S., Pearse, I. S., Smyth, R. L., Auer, S., Cook, G. L., Edwards, T. C., Guala, G. F., Howard, T. G., Morisette, J. T., y Hamilton, H. (2019). Development and Delivery of Species Distribution Models to Inform Decision-Making. BioScience, 69(7), 544-557.
https://doi.org/10.1093/biosci/biz045
80. Solari, S., Muñoz-Saba, Y., Rodríguez-Mahecha, J. V., Defler, T. R., Ramírez-Chaves, H. E., y Trujillo, F. (2013). Riqueza, endemismo y conservación de los mamíferos de Colombia. Mastozoologíaneotropical, 20(2), 301-365.
81. Stromberg, J. (s. f.). What Is the Anthropocene and Are We in It? 1.INCLUIR DATOS DE PUBLICACIÓN COMPLETOS.
82. Streib, L., Kattwinkel, M., Heer, H., Ruzika, S., y Schäfer, R. B. (2020). How does habitat connectivity influence the colonization success of a hemimetabolous aquatic insect?A modeling approach. Ecological Modelling, 416, 108909.
83. Tognelli, M. F., Roig, S. A., Marvaldi, A., Flores, G. E., y Lobo, J. M. (2009). An evaluation of methods for modelling distribution of Patagonian insects Sociedad de Biología de Chile; Revista Chilena de Historia Natural; 82; 3; 12-2009; 347-360
84. Valencia, M. A. (2020). LA IMPORTANCIA DE LA CONECTIVIDAD ECOLÓGICA ENTRE LOS HUMEDALES DE BOGOTÁ, DISTRITO CAPITAL. Universidad Militar NuevaGranada, 9.
78
85. Vet, L., y Smid, H. (2006). Learning in insects: From behaviour to brain. AnimalBiology, 56(2), 121-124. https://doi.org/10.1163/157075606777304186
86. Vilela, D., Ricioli, L., Del-Claro, K., y Guillermo-Ferreira, R. (2017). Female color polymorphism of Ischnura capreolus Hagen, 1861 (Odonata: Coenagrionidae) with notes on behavior and ontogenetic color changes. InternationalJournalofOdonatology, 20, 191-200. https://doi.org/10.1080/13887890.2017.1373152
87. Vogt, P., Ferrari, J. R., Lookingbill, T. R., Gardner, R. H., Riitters, K. H., y Ostapowicz, K. (2009). Mapping functional connectivity. ecologicalindicators, 9(1), 64-71.
88. Vollering, J., Halvorsen, R., Auestad, I., y Rydgren, K. (2019). Bunching up the background betters bias in species distribution models. Ecography, 42(10), 1717-1727.
89. Walker, K. K., Rudicell, R. S., Li, Y., Hahn, B. H., Wroblewski, E., y Pusey, A. E. (s. f.). Chimpanzees breed with genetically dissimilar mates. Royal Society Open Science, 4(1), 160422. https://doi.org/10.1098/rsos.160422
90. Ward, J. V. 1992. Aquatic Insect Ecology. 1. Biology and Habitat. John Wiley, New York.
91. Wickham, H. (2016). ggplot2:ElegantGraphicsforDataAnalysis [R-project]. SpringerVerlag. https://ggplot2.tidyverse.org/
92. Wiens, J. A., Stralberg, D., Jongsomjit, D., Howell, C. A., y Snyder, M. A. (2009). Niches, models, and climate change: Assessing the assumptions and uncertainties. Proceedings oftheNationalAcademyof Sciences, 106(supplement_2), 19729-19736. https://doi.org/10.1073/pnas.0901639106
93. Wunderlich, R. F., Mukhtar, H., y Lin, Y.-P. (2022). Comprehensively evaluating the performance of species distribution models across clades and resolutions: Choosing the right tool for the job. Landscape Ecology. https://doi.org/10.1007/s10980-02201465-1
79
