
13 minute read
Résistances de la tomate


Résistances de la tomate aux bioagresseurs Source : INRA France
La première méthode de protection utilisée contre les bioagresseurs a été la résistance génétique. Ainsi, des plantes possédant de meilleures caractéristiques agronomiques et capables de produire en présence de parasites ont été sélectionnées dès le début de l’agriculture. C’est à partir de 1900 que commence à se réaliser une sélection sur des bases scientifiques. Les efforts déployés par les sélectionneurs depuis les années 1950 ont abouti à la création de variétés possédant de plus en plus de gènes de résistance, comme nous le verrons ultérieurement.
Advertisement
Sources de résistance Chez la tomate, la lutte génétique repose sur l’exploitation de résistances généralement monogéniques dominantes provenant d’espèces sauvages affines de l’espèce cultivée. C’est l’une des espèces modèles pour l’utilisation de résistances monogéniques dans les variétés cultivées. Les deux espèces sauvages qui ont jusqu’ici apporté le plus de gènes de résistance dans
les variétés cultivées sont Solanum pimpinellifolium (ex Lycopersicon pimpinellifolium) et S. peruvianum (ex L. peruvianum). En prenant la tomate comme parent femelle, les croisements avec S. pimpinellifolium sont réalisés facilement. En revanche, les croisements avec S. peruvianum sont délicats et nécessitent de recourir à des techniques particulières comme le sauvetage d’embryons in vitro. Il en est de même avec l’espèce S. chilense (ex L. chilense), très voisine de S. peruvianum, et qui est à l’origine de récents travaux sur la résistance à plusieurs Begomovirus. L’hybridation de chacune de ces deux espèces avec la tomate produit des embryons qui avortent
dans les graines bien avant la maturation des fruits. Il faut donc recourir à leur extraction à l’état immature, c’est-à-dire 30 à 34 jours après l’hybridation, sans attendre les 55-60 jours nécessaires à la maturation des fruits de la tomate prise comme femelle. Les embryons sont cultivés in vitro sur milieu nutritif jusqu’à l’obtention de plantules. Une autre technique consiste à polliniser la tomate avec un mélange du pollen de la tomate avec du pollen
de l’espèce sauvage. Cette pollinisation produit de nombreuses graines parmi lesquelles figurent quelques hybrides interspécifiques. Les hybrides F1 obtenus sont presque autostériles. Pour progresser vers le type tomate cultivée, les sélectionneurs doivent réaliser plusieurs générations de recroisements par la tomate. Le premier recroisement nécessite d’utiliser une des techniques employée pour l’obtention des plantes F1. Les recroisements suivants s’effectuent sans difficultés. Une autre espèce sauvage, S. habrochaites (ex L. hirsutum), a apporté des résistances à la tomate cultivée sans grande difficulté d’hybridation. Son rôle est devenu très important comme parent mâle des hybrides F1 utilisés comme porte-greffes, tant de la tomate que de l’aubergine. Ces hybrides sont réalisés en croisant une variété de tomate, porteuse de gènes dominants de résistance à plusieurs maladies notamment d’origine tellurique (« Ve », « I », « I-2 », « Fr », « Tm-2 2 » et « Mi ») avec un écotype de S. habrochaites, apportant de la vigueur et des résistances dominantes, dont une à la maladie des racines liégeuses (corky root) et l’autre à Didymella lycopersici. À ces résistances, il convient d’ajouter la résistance à Alternaria alternata f. sp. lycopersici contrôlée par le gène « Asc » présent dans la quasi-totalité des variétés cultivées, anciennes ou récentes. Des programmes de sélection utilisant d’autres résistances venant des espèces citées arriveront prochainement à leur terme. Des recherches de résistances exploitables sont conduites dans d’autres espèces de Lycopersicon, ainsi que dans des espèces éloignées appartenant au genre Solanum. Quatre espèces appartenant à ce dernier genre (S. lycopersicoides, S. juglandifolium, S. ochranthum et S. sitiens), regroupées dans la série « juglandifolia », présentent de fortes analogies morphologiques et chromosomiques avec des espèces du genre Lycopersicon. Ainsi, des hybrides F1 sont aisément obtenus avec S. lycopersicoides, espèce intéressante pour sa tolérance au froid et ses résistances au Cucumber mosaic virus (CMV), à Clavibacter michiganensis subsp. michiganensis et à Botrytis cinerea. Des problèmes de stérilité et d’incompatibilité rendent toutefois difficiles les processus de rétrocroisement.
Nature, niveau d’efficacité et durabilité des résistances La résistance est une caractéristique héritable qui diminue les effets du parasitisme. Son efficacité dépend de la combinaison de deux facteurs : - le niveau d’expression de la résistance ; - la durabilité, ou stabilité de la résistance dans le temps.

Ces deux facteurs possèdent chacun un déterminisme particulier :
Déterminisme du niveau de la résistance Résistance et sensibilité sont les deux extrêmes d’un ensemble de réactions de la plante hôte. Selon le mécanisme impliqué, le niveau de la résistance apparaîtra plus ou moins élevé, d’où la distinction entre résistance absolue et résistance partielle. La résistance absolue, appelée aussi « verticale », est due à un phénomène d’immunité ou à un mécanisme d’hypersensibilité. Dans le cas d’immunité, la plante est totalement indemne de parasitisme. Cette résistance peut résulter d’une absence de fixation de l’agent pathogène sur l’hôte ou, dans le cas des virus, de l’absence chez l’hôte d’un élément ou d’une fonction essentielle à la réplication virale. La résistance à Mycovellosiella fulva, conférée par le gène « Cf-2 », est un bel exemple d’immunité ; elle est maintenant surmontée par la race 2 présente dans de nombreuses régions. Le terme d’« immunité » est souvent utilisé improprement pour dénommer une résistance qui se manifeste en absence de symptômes visibles, mais qui n’exclut pas la pénétration du bioagresseur. Lorsqu’intervient un mécanisme d’hypersensibilité, le processus d’infection demeure localisé et inactivé par la mort des tissus infectés. À température élevée, le mécanisme est plus lent et des symptômes peuvent apparaître. Deux exemples sont fournis par le gène « Tm-2² » de résistance aux Tomato mosaic virus (ToMV) et au Tobacco mosaic virus (TMV) et par le gène « Mi » conférant la résistance aux Meloidogyne spp. La résistance partielle, appelée aussi « horizontale », est caractérisée par la diminution du nombre de points de fixation des agents pathogènes sur l’hôte, le ralentissement de leur croissance et de leur développement dans les tissus et la diminution du nombre d’unités infectieuses émises. La résistance partielle à Phytophthora infestans, contrôlée par le gène « Ph-2 », illustre cette situation. Il résulte globalement de ces phénomènes une progression plus lente de la maladie sur la plante et de l’épidémie dans la culture. Une résistance partielle soutenue par de bonnes pratiques culturales et une protection phytosanitaire raisonnée peut éviter le développement d’une épidémie. La notion de résistance partielle ne doit pas être confondue avec la tolérance qui est une notion agronomique. Cette dernière caractérise le comportement d’une plante dans laquelle le parasite vit et se reproduit, comme dans une plante sensible présentant des symptômes typiques de maladie, mais dont le rendement n’est pas affecté. Cependant, il est fréquent de parler de « variétés tolérantes » en virologie pour caractériser des plantes qui permettent une multiplication active d’un virus sans extérioriser de symptômes typiques de la maladie, et dont le rendement n’est pas affecté.
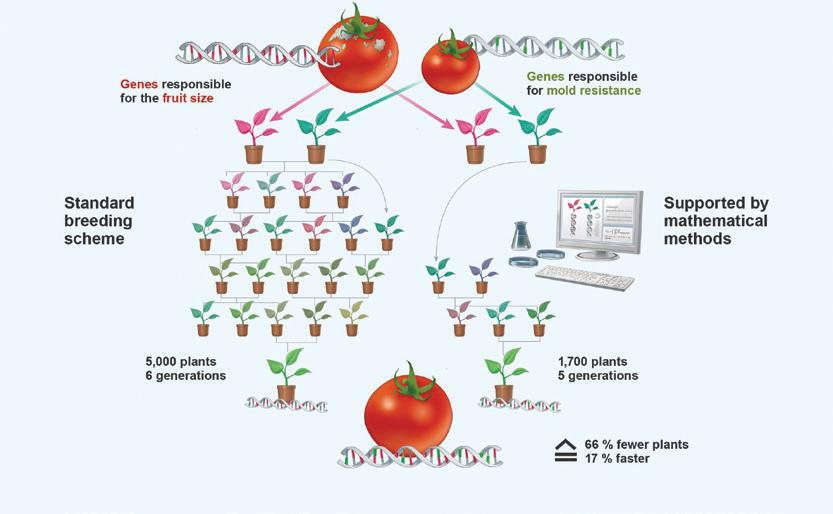
Déterminisme de la stabilité de la résistance dans le temps La stabilité des résistances de haut niveau peut être extrêmement variable selon les gènes les contrôlant. Ce n’est qu’après de nombreuses années d’utilisation que l’on peut évaluer avec fiabilité la durée d’une résistance ou l’importance pratique de l’adaptation d’un agent pathogène à une résistance donnée. La rapidité d’apparition de nouveaux pathotypes est extrêmement grande chez certains agents pathogènes comme Mycovellosiella fulva. À l’opposé, après de nombreuses années d’utilisation dans des conditions environnementales très variées, certaines résistances n’ont jamais été surmontées comme celle aux Stemphylium spp. D’autre part, il existe de nombreux exemples de résistances qui, bien que surmontées, continuent de présenter un intérêt pratique non négligeable dans certains contextes culturaux.
Situation des résistances disponibles chez les variétés cultivées et les porte-greffes Une quinzaine d’agents pathogènes sont maintenant contrôlables par des résistances génétiques chez la tomate. L’efficacité de ces résistances est très variable, que ce soit pour leur niveau d’expression ou leur stabilité dans le temps face à l’évolution de la virulence des agents pathogènes. Les variétés sélectionnées pour leur résistance aux bioagresseurs sont surtout destinées aux cultures abritées, plus fréquemment et sévèrement parasitées. Leur potentiel de production étant très important, le prix élevé des semences résultant de coûteux programmes de sélection est facilement accepté. Certaines résistances mises en évidence depuis longtemps ne sont présentes que dans de rares variétés. Dans le cas de Pyrenochaeta lycopersici, la raison est complexe ; cette résistance est partielle et d’hérédité monogénique récessive, et le géne « pyl » qui la contrôle doit être présent dans les deux parents des hybrides F1. De plus, la pratique du greffage sur porte-greffes multirésistants possédant notamment un haut niveau de résistance à P. lycopersici diminue l’intérêt de la sélection d’hybrides F1 résistants à ce champignon tellurique. La résistance partielle à Phytophthora infestans, difficile à mettre en évidence par des tests précoces de sélection, n’a intéressé que peu de sélectionneurs. De plus, elle concerne une maladie aérienne pour laquelle de nombreux fongicides efficaces sont disponibles sur le marché. Les autres résistances, d’utilisation limitée, intéressent surtout les zones de production particulières (par exemple tropicales humides pour Ralstonia solanacearum) ou font l’objet de programmes relativement récents. La disponibilité de résistances monogéniques dominantes permet de cumuler dans les hybrides F1 un nombre croissant de résistances : les hybrides destinés aux cultures abritées offrent généralement 4 à 5


résistances, certains en cumulent jusqu’à 7. Pour les cultures de plein champ, les variétés fixées possèdent 2 à 4 résistances. De plus en plus d’hybrides F1 de ces différents types de production disposent de ces résistances et contribuent à la protection durable de la tomate. Les porte-greffes disponibles ne sont pas très nom- breux et peuvent être séparés en 2 groupes : - les hybrides F1 dans le type tomate cultivée avec plusieurs résis- tances aux maladies telluriques dont une résistance partielle à Pyre- nochaeta lycopersici ; - les hybrides F1 interspécifiques entre la tomate cultivée et Lycopersicon hirsutum. Ces hybrides ont un système racinaire puissant supportant des températures plus basses que la tomate cultivée. Lycopersicon hirsutum, originaire des hauteurs andines, offre aussi un haut niveau de résistance dominante à Pyrenochaeta lycopersici.

www.agri-mag.com Espoirs de nouvelles résistances à court terme Devant l’efficacité et l’absence de nuisance pour l’environnement de la lutte génétique, d’importants programmes de sélection ont été et sont conduits dans le monde. Ils devraient permettre de contrôler un nombre croissant de bioagresseurs. Les techniques de biologie moléculaire utilisées pour marquer les gènes intéressant les sélectionneurs permettent la sélection de résis- tances difficiles à mettre en évidence par des biotests précoces. Elles facilitent ainsi la sélection de résistances partielles, oligogéniques ou polygéniques et le cumul d’un plus grand nombre de gènes de résis- tance. De sérieux espoirs de résistance sont envisageables pour pratique- ment tous les types de micro-organismes phytopathogènes. Ils concernent pour plusieurs d’entre eux le contrôle de certaines races adaptées aux résistances actuellement disponibles dans des variétés cultivées. À titre d’exemple, les micro-organismes susceptibles d’être contrôlés par des résistances issues des espèces sauvages de Solanum (ex Lycopersicon) sont présentés ci-après. - Un nouveau pathotype de Fusarium oxysporum f. sp. lycopersici qui surmonte les gènes « I » et « I-2 » est maintenant présent dans diffé- rentes régions. Cette nouvelle race, désignée généralement « race 3 », est contrôlée par le gène dominant « I-3 » issu de S. pennellii (ex L. pennellii). Il a été introduit dans des lignées de tomate ; des hybrides F1 commerciaux résistants aux 3 races commencent à apparaître. - De nombreux travaux font état de résistances partielles à Alternaria tomatophila, provenant de diverses espèces sauvages dont S. habro- chaites (ex L. hirsutum) et présentes dans différentes variétés commer- ciales. Cette résistance ne doit cependant pas être confondue avec celle efficace contre A. alternata f. sp. lycopersici qui est contrôlée par le gène dominant « Asc ». Cette résistance, présente chez la quasi-to- talité des variétés cultivées anciennes et modernes, est référencée « résistance à l’Alternaria » dans certains catalogues semenciers. Cette indication prête à confusion : il ne s’agit en aucun cas d’une résistance à l’alternariose du feuillage provoquée par A. tomatophila. - Le gène « Ph-1 » de résistance à Phytophthora infestans a été sur- monté avant même d’avoir été utilisé en sélection, et le gène « Ph-2 », incomplètement dominant et contrôlant une résistance partielle, a connu les mêmes déboires. Les sélectionneurs ont alors recherché une résistance plus efficace. Les espoirs portent maintenant sur le gène « Ph-3 » issu de S. pimpinellifolium (ex L. pimpinellifolium).

- En ce qui concerne Oidium neolycopersici, les sélectionneurs visent à cumuler les gènes de résistance issus d’espèces sauvages afin d’assurer un haut niveau de résistance à différentes races récemment mises en évidence. - Des sources de résistance à Clavibacter michiganensis subsp. michiganensis sont connues depuis longtemps. Il s’agit de résistances partielles, avec des niveaux d’expression très influencés par les conditions de milieu. Issues d’espèces sauvages, elles sont à la base de programmes de sélection conduits dans le monde entier, dont les types variétaux destinés à la transformation industrielle semblent les plus avancés. - Différentes sources de résistance aux Xanthomonas spp. ont été mises en évidence. Les travaux de sélection visent à cumuler les résistances aux différentes races et à en obtenir une s’exprimant tant pour le feuillage que pour les fruits. Une résistance provenant de S. pimpinellifolium est particulièrement travaillée. - En ce qui concerne le Potato virus Y (PVY), les travaux portent essentiellement sur la résistance issue de S. habrochaites contrôlée par deux gènes : « pot1 », récessif, et « Pot-2 », dominant. Cette résistance est efficace tant à l’égard des souches provoquant une mosaïque que des souches nécrotiques. - Un gène dominant, stable à haute température et permettant de contrôler l’Alfalfa mosaic virus (AMV), a été mis en évidence dans une origine de S. habrochaites. Ce gène, dénommé « Am », intéresse les sélectionneurs des pays méditerranéens où l’AMV est parfois grave. - Pour le Tomato yellow leaf curl virus (TYLCV), la sélection tend à augmenter le niveau de résistance des hybrides en cumulant des gènes issus de différentes espèces sauvages. Des programmes de sélection sont également conduits pour contrôler les autres Begomovirus, dont la diversité est importante. - Le gène « Mi », utilisé depuis longtemps dans différents contextes culturaux pour combattre les nématodes à galles appartenant au genre Meloidogyne, est surmonté dans de nombreuses régions. Deux autres gènes issus de S. peruvianum (ex L. peruvianum) ont été introduits dans des lignées de tomate. Le gène « Mi-2 » contrôle les souches surmontant « Mi » mais s’avère, comme ce dernier, inefficace à température élevée. En revanche, « Mi-3 » permet de maîtriser les souches adaptées à « Mi » et se montre stable à haute température. Les sélectionneurs s’intéressent à ce gène, notamment pour les cultures de la tomate en zones chaudes et sols sableux, dans lesquels la température s’élève davantage au niveau des racines. Les programmes de sélection, aidés par le marquage moléculaire, cherchent maintenant à cumuler ces nouvelles résistances avec celles déjà disponibles et ayant fait leurs preuves dans le matériel commercialisé. À plus long terme, des résistances à d’autres bioagresseurs seront introduites dans les programmes de sélection. Des espoirs existent par exemple pour contrôler Verticillium albo-atrum, race surmontant le gène « Ve », Phytophthora nicotianae, Colletotrichum coccodes, Botrytis cinerea, Pseudomonas syringae pv. tomato (race surmontant le gène « Pto ») et le Pepino mosaic virus (PepMV). Des recherches sont également conduites pour contrôler génétiquement différents insectes, mais il est trop tôt pour en pronostiquer les résultats pratiques.









