


Edición 158 - Abril 2024
Sector camaronero ecuatoriano es reconocido por su generación de empleo.
Líder ecuatoriano, destacado entre 30 de los 100 más influyentes de la industria acuícola mundial.
NOAA anuncia finalización del Fenómeno de El Niño (neutral) y alerta sobre la posible presencia de La Niña en la región.
Alimento balanceado: nutriendo la sostenibilidad en la industria camaronera.
Camaroneros de India apuestan por el mercado local para mantener a flote la industria.
Identificación de nuevas cepas de Vibrio campbellii portadoras del plásmido pVA1 aislado de postlarvas de Penaeus vannamei afectadas por brotes de Necrosis Hepatopancreática Aguda (AHPND).
Investigaciones muestran que las pruebas rápidas de campo (POC) solo detectan WSSV cuando hay alto nivel de infección, pero hay una mejor solución para los productores de camarón.
Maximizando la productividad: el papel de los sensores en la acuicultura.
Experiencias y desafíos de la eliminación de la ablación en reproductores de Penaeus vannamei en una maduración de Ecuador.
La acumulación tisular de microplásticos de poliestireno provoca estrés oxidativo, lesión hepatopancreática y alteraciones del metaboloma en Penaeus vannamei.
Aplicación de la secuenciación de nueva generación en el análisis del microbioma asociado a la producción camaronera.

Presidente Ejecutivo Ing. José Antonio Camposano
Editora “AquaCultura” MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial MSc. Yahira Piedrahita PhD. Leonardo Maridueña Ing. José Antonio Lince Econ. Danny Vélez Ing. Alex de Wind
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Reporte de mercado de China.
Reporte de mercado de EE. UU.
Noticias de interés. Exportaciones de camarón.
Corrección de estilo Daniel Ampuero daniel.ampuero@gmail.com
Comercialización Gabriela Nivelo gnivelo@cna-ecuador.com
ÍNDICE
INFORMACIÓN DE COYUNTURA ARTÍCULOS TÉCNICOS ESTADÍSTICAS NOTICIAS 8 12 13 20 15 27 36 40 47 53 66 74 79 82 85
En estas últimas semanas, hemos reflexionado con Marcelo Vélez, Presidente del Directorio de la Cámara Nacional de Acuacultura (CNA), sobre la compleja coyuntura que atraviesan nuestras empresas, así como nuestro sector gremial e institucional en su conjunto.

Por un lado, en un entorno empresarial cada vez más competitivo, es necesario que nuestros colaboradores se mantengan en constante búsqueda de eficiencias, lo que demanda un alto grado de creatividad y adaptabilidad por parte de todos.
Mientras que, a nivel institucional, nos enfrentamos al desafío de discernir entre lo prioritario y lo urgente, una tarea que requiere un criterio cuidadoso para no perder de vista lo que realmente impacta en nuestra industria. El papel del gremialismo es crucial en este escenario, ya que debe abordar las complejidades de los negocios manteniendo un equilibrio entre las diversas perspectivas de las empresas que componen nuestro sector. Esta tarea no es sencilla, pero es fundamental para enfrentar con éxito los desafíos que se nos presentan, especialmente en una industria exigente como la acuacultura.
Si bien es imposible complacer a todos y acoger cada punto de vista, nuestro objetivo es alcanzar metas que reflejen una gestión profesional, técnica y proactiva, generando valor para nuestros asociados.
La CNA se ha destacado por su compromiso en abordar una amplia variedad de temas, con el objetivo de encontrar
EDITORIAL
José Antonio Camposano Presidente Ejecutivo
Reflexiones desde la CNA: Desafíos, nuestro rol y compromisos en tiempos actuales
soluciones a situaciones que demandan una visión unificada, muchas veces complementando la acción gubernamental ante problemas comunes.
Es justamente aquí donde la comunidad acuícola mundial reconoce que la Cámara Nacional de Acuacultura del Ecuador es un ejemplo de cómo el diálogo colaborativo y precompetitivo privado se ha constituido en el mecanismo para que una industria promueva su crecimiento sostenido y sostenible.
A pesar de estas virtudes, reconocemos la necesidad de un proceso continuo de mejora, pues sólo así mantendremos siempre la mirada en el horizonte para anticipar tanto amenazas como oportunidades que puedan afectar a nuestra industria. La retroalimentación de nuestras empresas afiliadas es fundamental, ya que nos brinda la oportunidad de corregir errores y perfeccionar lo que ya está funcionando correctamente. En este sentido, el compromiso del todo el equipo que conforma la CNA radica en su capacidad para cultivar una visión proactiva que permita a los empresarios concentrarse en sus negocios mientras el gremio allana el camino hacia el éxito.
Mientras continuemos cumpliendo con este rol, pueden confiar en que estaremos, como es debido, junto al sector empresarial formal, que no solo genera empleo, sino que también crea más oportunidades para aquellos que buscan en la cría y procesamiento de camarón un medio de vida digno•

PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
DIRECTORIO
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
Ing. Roberto Aguirre
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
Blgo. Carlos Sánchez
Ing. Diego Puente
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Carlos Miranda
Econ. Danny Vélez
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León


SECTOR CAMARONERO ECUATORIANO ES RECONOCIDO POR SU GENERACIÓN DE EMPLEO
El sector camaronero ecuatoriano es considerado un pilar fundamental para la economía del país porque genera más de 290 mil plazas de trabajo directas e indirectas a lo largo de su cadena de valor. A esto se suma que tras la entrada en vigencia de la Ley de Eficiencia Económica y Generación de Empleo promulgada el 20 de diciembre de 2023, empresas del sector camaronero han puesto en marcha proyectos de inclusión laboral para jóvenes.
El 13 de marzo pasado, el presidente de la República, Daniel Noboa Azin, visitó las instalaciones de Industrial Pesquera Santa Priscila (IPSP) en la provincia del Guayas. Esta acción estuvo enmarcada en
el programa gubernamental denominado “Somos El Nuevo Ecuador del Empleo Joven”.
Durante el encuentro, más de mil trabajadores de esa empresa participaron en la actividad que fue transmitida en vivo por las redes sociales de la Presidencia de la República a nivel nacional.
Al inicio del acto, Christina Salem, directora Financiera de IPSP, saludó en nombre de los trabajadores al Primer Mandatario y a la ministra del Trabajo, Ivonne Núñez, y destacó la importancia de su acercamiento para continuar fomentando el empleo digno. Hasta marzo pasado, Santa Priscila contrató a más de 1,100 jóvenes en aplicación a la

"Varios son los sectores que están poniendo de su parte para reactivar la economía, y el sector camaronero es uno de ellos. Agradezco a todas las empresas que con compromiso y valentía apuestan por nuestra gente, invierten aquí. Esto nos permitirá construir un país que nos beneficie a todos”.
Daniel Noboa Presidente de la República
normativa que fue propuesta por el Ejecutivo en diciembre del año pasado.
Por su parte, el presidente Noboa expuso al país los primeros resultados de la iniciativa y resaltó el trabajo conjunto entre el sector público y privado, tomando como ejemplo esta empresa. Anunció que más de 63,890 jóvenes entre 18 y 28 años se han sumado al mercado laboral con un contrato legal hasta marzo pasado, lo que a decir del mandatario demuestra el compromiso del país con su juventud y la reactivación económica, a través de leyes que fomenten la inclusión del trabajo joven.

“Su presencia aquí es un testimonio del compromiso con el desarrollo económico del país y es un estímulo adicional para seguir adelante nuestro proyecto de generar empleo digno y contribuir al progreso del Ecuador”.
Cristina Salem
Directora Financiera de Industria Pesquera Santa Priscila
8
plazas de trabajo genera el sector camaronero +290 mil

Señaló que estas iniciativas mejoran las condiciones laborales y ofrecen oportunidades de vida. Además, subrayó que la ley ha proporcionado empleo a la juventud, reduciendo su vulnerabilidad ante la delincuencia. Sobre este último tema, Noboa hizo énfasis en la importancia de mantener la lucha contra el narcoterrorismo, respaldando las reformas propuestas por el Gobierno para fortalecer la seguridad y la paz del país.
63,890
jóvenes fueron contratados, a partir de la ley
las contrataciones juveniles, sumando un total aproximado de 19,600 empleos creados en promedio. Además se mencionó la disponibilidad de cursos y capacitaciones gratuitas a través del servicio público "Encuentra Empleo", beneficiando a cerca de 5,000 personas en áreas como administración, tecnología y servicios.
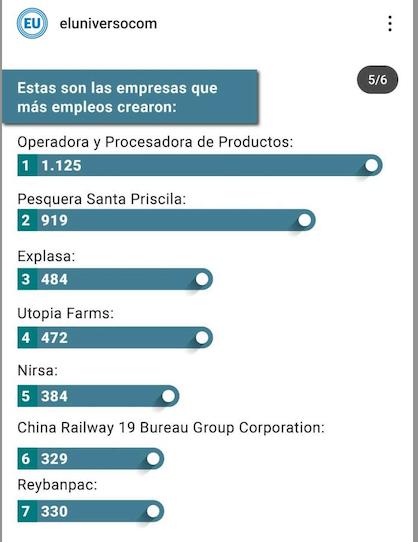
camaronero como principales generadoras de empleo joven en el Ecuador. En su lista figuró: La Operadora y Procesadora de Productos OMARSA, Industrial Pesquera Santa Piscila, Expalsa y NIRSA.
Por su parte, el Ministerio del Trabajo (MDT) destacó que Guayas y Pichincha encabezan
Por otra parte, en una publicación realizada por Diario El Universo en marzo pasado, mencionó a otras empresas del sector

“El presidente dijo: en los jóvenes está la fortaleza, en los jóvenes está la valentía, el presente y el futuro de nuestro país; para ellos debe haber una política social y laboral del Ecuador. Empezó como una idea, se construyó en un sueño y ahora es una realidad”.
Ivonne Núñez Ministra del Trabajo

“Tengo 24 años y actualmente me desempeño en el área de logística. Estoy muy agradecida presidente, especialmente con Ud. por su ley que respalda a los jóvenes, que no tenemos experiencia laboral y a los que no nos tomaban en cuenta, y ahora nos dan la oportunidad para poder desempeñar nuestras capacidades”.
Samantha Navarro Trabajadora de Santa Priscila
9
Publicación de @eluniversocom en Instagram, 15 de marzo de 2024.
COYUNTURA
BENEFICIOS AL GENERAR EMPLEO PARA JÓVENES
La Ley de Eficiencia Económica y Generación de Empleo ofrece diversos incentivos a las empresas para fomentar la contratación de jóvenes y otros grupos específicos:
▪ Aquellas empresas que incrementen netamente el número de empleados jóvenes entre los 18 y 29 años, así como aquellos responsables de pagar pensiones alimenticias, recibirán una deducción adicional del 50% sobre los salarios en los que se hayan realizado contribuciones a la seguridad social.
▪ Si las nuevas contrataciones son jóvenes graduados de universidades públicas o institutos superiores técnicos, tecnológicos, pedagógicos, de artes y conservatorios superiores, o de instituciones educativas públicas, municipales o fiscomisionales, la deducción adicional aumentará al 75%.
▪ Para las empresas que aumenten netamente sus empleados en los sectores de la construcción y la agricultura, la deducción adicional será del 75% sobre los gastos salariales sujetos a aportaciones a la seguridad social.
▪ Además, aquellas compañías que generen un incremento neto en empleos para individuos que hayan cumplido con una pena privativa de libertad superior a un año podrán beneficiarse de una deducción adicional del 75% sobre los gastos salariales sujetos a aportaciones a la seguridad social.
▪ Quienes se acojan al sistema de estabilidad tributaria del régimen general de IR por 5 años, tendrán un incremento del 2% cada año en la tarifa general de la empresa.
CIFRAS
Según el portal web del Instituto Ecuatoriano de Seguridad Social, IESS, en febrero de 2024 había 850,558 afiliados entre personas que tienen hasta 30 años de edad, lo que mostraba una disminución de 13,081 afiliados con relación a la cantidad de afiliados que había a diciembre de 2023.
Según el Instituto Nacional de Estadística y Censos (INEC), la tasa de desempleo en jóvenes de entre 15 y 24 años es de 8,5% a enero de 2024. No solo eso, el empleo adecuado se ubica en apenas 17%, mientras que el informal llega al 65,4%. Es decir, la mayoría de jóvenes labora en empresas sin
Registro Único de Contribuyentes (RUC), por lo que tampoco tienen un contrato laboral ni son afiliados al IESS.
En las personas de entre 25 y 34 años el desempleo está en 5,9%, con lo cual también supera la tasa general de desempleo en el país, de 3,9%. No obstante, en este rango el empleo adecuado llega al 44,7% y el trabajo informal ocupa el 39,3%.
El camino a transitar no es fácil, pero la empresa privada está dispuesta a avanzar en el desarrollo empresarial y la empleabilidad juvenil en Ecuador. Se requiere una formación profesional adaptada a las demandas del mercado laboral y programas que integren aprendizaje y empleo, por eso las empresas desempeñan un papel crucial invirtiendo en este rubro. Para garantizar el bienestar de los jóvenes trabajadores, es necesario mejorar la calidad y las condiciones laborales, fomentar la creación de empleos formales, ofrecer capacitación en habilidades pertinentes y promover el espíritu emprendedor y la innovación•

- ABRIL 2024


Líder ecuatoriano, destacado entre 30 de los 100 más influyentes de la industria acuícola mundial

La revista internacional INTRAFISH, en su especial Seafood Power 100 – 2024, ubicó a José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura (CNA), entre 30 de los 100 ejecutivos más influyentes de la industria acuícola a nivel global. Este significativo logro representa un ascenso notable de 20 puestos con respecto a su posición anterior, colocándolo en una destacada posición en la categoría.
En esta reciente publicación se resalta el “sólido compromiso que tiene su líder con la industria acuícola durante más de una década al frente de la CNA”. La nota menciona además que “Ecuador ha establecido récords en la exportación mundial de camarón, con un aumento significativo del 14,5% en 2023, gracias a sus buenas prácticas, bienestar animal y control de calidad”.
El Seafood Power 100 se enfoca en mostrar el liderazgo y la capacidad de dinamizar los mercados mediante decisiones estratégicas, asumiendo grandes desafíos frente a sus competidores y la ejecución de acciones asertivas que impulsen el avance de toda la industria. Es
por eso que su lista de ejecutivos define el panorama de influencia y dirección en este sector tan dinámico.
Este no es el primer reconocimiento internacional que recibe José Antonio Camposano. En años anteriores también ha sido incluido en el Seafood Power 100 y en el "40 Under 40" de INTRAFISH en 2015. Actualmente, Camposano se mantiene al frente de la Presidencia Ejecutiva de la CNA y se desempeña también como Presidente de la Corporación de Gremios Exportadores del Ecuador (CORDEX), entidad que agrupa a más del 75% de los sectores productivos y de exportación (no petrolero y no minero) del país, reuniendo a más de 1.000 empresas que generan cerca de 900 mil empleos directos. Por otra parte, sigue siendo Presidente del Directorio de la Corporación de Promoción de Exportaciones e Inversiones del Ecuador (CORPEI).
Durante su amplia trayectoria, José Antonio Camposano ha desempeñado roles clave en organizaciones de cooperación para el desarrollo, promoviendo programas de responsabilidad social corporativa y colaboración público-privada, así como promoviendo la participación de Ecuador en ferias comerciales internacionales.
Su contribución como expositor en foros internacionales y como docente en universidades ecuatorianas ha sido notable, compartiendo su experiencia en comercio exterior y marketing internacional. En conjunto, su liderazgo ha sido fundamental para posicionar al país como líder en la producción y exportación de camarón a nivel global, consolidando su influencia en la industria acuícola mundial•
COYUNTURA
12
NOAA anuncia finalización
del Fenómeno de El Niño
(neutral) y alerta sobre la posible presencia de La Niña en la región

La alternancia entre El Niño y La Niña, ha sido una constante comprobada científicamente por más de 70 años, según señala Leonardo Maridueña, Director de Ambiente de la Cámara Nacional de Acuacultura. Este fenómeno debe haber existido durante un período prolongado, pero fue en 1950 cuando la comunidad oceanográfica confirmó su presencia y su alternancia con El Niño. Desde una perspectiva científica, esto tiene sentido, ya que se alinea con la tercera Ley de Newton, que establece que: "toda acción provoca una reacción de igual intensidad pero en dirección opuesta".
El Centro de Predicciones Climáticas de los Centros Nacionales de Predicción Ambiental, en colaboración con la Administración Nacional Oceánica y Atmosférica (NOAA), emitió un comunicado el 11 de abril de 2024 sobre las condiciones oceánicas y atmosféricas levantadas a la fecha y un pronóstico probabilístico de las condiciones climáticas en el océano Pacífico ecuatorial.
Los pronósticos del Instituto Internacional de Investigación (IRI) revelan lo siguiente:
• Existe un 85% de probabilidad de que, para el periodo abril - junio de 2024, exista una transición de fenómeno de El Niño a ENSO neutral, es decir que durante este periodo de transición no prevalecerán características ni del Niño ni de La Niña.
• Por otro lado, existe una probabilidad creciente del 60% de que, para los meses de junio- agosto se desarrolle el fenómeno de La Niña.
Es decir que los efectos de sequía correspondientes a La Niña podrían empezar a notarse a partir del segundo semestre de este año. Recordemos que este es un fenómeno oceánico caracterizado por el
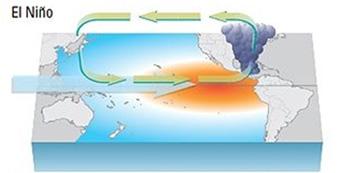
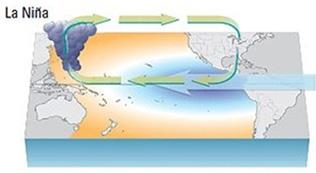
enfriamiento de las aguas superficiales de la parte central y oriental del Pacífico ecuatorial, y por cambios en la circulación atmosférica tropical. Suele ir acompañada de bajas temperaturas y provoca fuertes sequías en las zonas costeras del Pacífico.
Implicaciones para el sector camaronero
Las afectaciones o riesgos que genera este fenómeno dependerán de la severidad y la duración del mismo. Por lo general, las implicaciones que han traído para el sector camaronero han sido:
- El decremento de la temperatura del agua puede afectar directamente a las especies acuáticas, causando estrés térmico y disminución de la calidad del líquido.
- La sequía que produce, especialmente en la región interandina, reduce sustancialmente el aporte de agua de las vertientes hacia la costa, permitiendo mayor intrusión de agua de mar y por consiguiente, el incremento de la salinidad en las áreas estuarinas.
- Aumenta la surgencia de aguas frías y aumenta la producción primaria; esto implica
una mayor presencia de plancton, lo que favorece a la alimentación de las especies acuáticas.
- Estos cambios incrementan la posibilidad de que se desarrollen nuevas enfermedades adaptadas a esas condiciones.
- Las bajas temperaturas reducen el metabolismo y por consiguiente, el crecimiento de los camarones; por lo tanto, tomaría más tiempo y alimento alcanzar las tallas comerciales.
La adaptación y la gestión sostenible de los recursos acuáticos se vuelven fundamentales para mitigar los posibles impactos negativos y aprovechar las oportunidades que estos fenómenos climáticos presentan.
En este sentido, los continuos monitoreos y la colaboración entre instituciones científicas, las cuales deben mantener informados a los organismos gubernamentales y actores del sector, son clave para una respuesta efectiva frente a los cambios en el clima y sus implicaciones en la actividad camaronera y otros sectores relacionados•
- ABRIL 2024 COYUNTURA
13
Esquema océano-atmosférico de El Niño Modoki y La Niña Modoki

MENSAJE
 Pamela Nath Directora
Pamela Nath Directora
En las últimas décadas hemos sido testigos de un cambio climático acelerado, manifestándose a través de fenómenos notables como el incremento de las temperaturas, el deshielo de los glaciares y la presencia de eventos meteorológicos extremos cada vez más frecuentes. Al mismo tiempo, hemos visto avances significativos en la conciencia ambiental, reflejados en un compromiso mundial creciente para reducir las emisiones de gases de efecto invernadero y fomentar prácticas sostenibles en las industrias.
En la acuicultura de camarón, abordamos actualmente el desafío de asegurar que nuestro sector se consolide como una práctica limpia, sostenible y exitosa a nivel global. Por esta razón, valoramos la importancia de identificar, comprender y reflexionar sobre las oportunidades de mejora en el rendimiento ambiental de nuestra industria. Sin embargo, para trabajar en este reto, es fundamental contar con transparencia y colaboración en toda la cadena de suministro.
Solo mediante esfuerzos conjuntos será posible elevar a toda la industria al siguiente nivel en cuanto a prácticas sostenibles. Es por esto que en Sustainable Shrimp Partnership (SSP) exploramos alianzas estratégicas con empresas, ONGs y expertos que compartan nuestra visión y valores. Si bien nuestros miembros asociados no son productores directos de camarón, ellos están comprometidos con impulsar el desarrollo de soluciones, proyectos y prácticas que contribuyan a un futuro más responsable para la acuicultura.
Desde la preservación del medio ambiente hasta la promoción de estándares éticos y responsables, junto con nuestros socios aspiramos a crear un impacto positivo y duradero. Gracias a estas colaboraciones, hoy es posible identificar e impulsar las mejores prácticas para garantizar la sostenibilidad en la acuicultura del camarón y abordar de manera integral soluciones para mitigar el cambio climático.
Alimento balanceado: nutriendo la sostenibilidad en la industria camaronera
Autores:
Pamela Nath
Sally Tabares
Sustainable Shrimp Partnership (SSP)
pnath@sustainableshrimp.org
Hoy en día, al igual que muchos otros sistemas alimentarios, la acuicultura se enfrenta a un doble desafío de sostenibilidad. Por un lado, existe la necesidad de incrementar la producción acuícola para solventar la demanda de alimento de una población en crecimiento. Por otra parte, la industria afronta la tarea de reducir el impacto ambiental de su producción para combatir el cambio climático.
Para el año 2050, se estima que la población mundial alcance los 9700 millones de personas y que la demanda global de proteínas se duplique, según datos de la Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). Además, para 2030 se espera que la acuicultura sea responsable del 59% de los alimentos acuáticos disponibles para consumo humano, según la misma organización.
Sin embargo, el crecimiento del sector acuícola conlleva inevitablemente un aumento en las emisiones de gases de efecto invernadero (GEI), los principales impulsores del cambio climático.
Por este motivo, el Grupo de Trabajo de Acuicultura sobre la Huella Ambiental, liderado por Sustainable Trade Initiative (IDH) y en el que participa Sustainable Shrimp Partnership (SSP), continúa trabajando para comprender, medir y reducir la huella ambiental de la acuicultura en toda la cadena de suministro.
Con el objetivo de evaluar el impacto negativo en la producción acuícola, diferenciando entre aquellos con un mejor desempeño ambiental y los que necesitan mejoras, el grupo de trabajo ha realizado pruebas preliminares para medir la huella de carbono de algunos miembros. En este proceso, se destacó que el alimento balanceado juega un papel fundamental para mejorar el impacto ambiental en el sector.
De acuerdo con el informe “Quantifying greenhouse gas emissions from global aquaculture”, publicado en 2017 por Scientific Reports, la producción de alimento balanceado constituyó el 57% de las emisiones totales de GEI derivadas de la acuicultura. El informe también destaca que dentro de las emisiones asociadas a la acuicultura de moluscos y peces, la del camarón contribuyó con el 21% de las emisiones de GEI pero tan solo el 10% de la producción total.
15 - ABRIL 2024 SOSTENIBILIDAD

Estos resultados subrayan la oportunidad de optimizar el desempeño ambiental de la nutrición en la industria camaronera. Además, destacan el rol de las fábricas de alimento balanceado para contribuir a la disminución de la huella ambiental del camarón cultivado. Por lo tanto, es importante que los profesionales del sector conozcan y fomenten activamente las mejores prácticas entre los productores de pienso, con el fin de abordar eficazmente este desafío.
“La sostenibilidad del alimento balanceado comienza con las materias primas”, informa la gerente global de Sostenibilidad de BioMar, Helen Ann Hamilton. En esta empresa especializada en la producción de alimentos balanceados para acuicultura, el 97% de las emisiones de GEI provienen de la producción, crecimiento y cosecha de estos insumos.
“Para mejorar la sostenibilidad de nuestro alimento balanceado, debemos medir y monitorear estrictamente el rendimiento sostenible de cada ingrediente individual, utilizando herramientas científicas de evaluación ambiental”, sostiene Hamilton, quien es doctora en Ecología Industrial.
Las plantas de alimentos balanceados emplean diversas herramientas científicas para conocer la huella de carbono de sus ingredientes, y una de ellas es el Análisis del Ciclo de Vida (LCA, por sus siglas en inglés). El LCA resulta ser el enfoque más adecuado y completo para identificar los puntos críticos de impacto ambiental a lo largo del ciclo de vida del producto, que podrían mitigarse o compararse con diversas alternativas, de acuerdo con el
informe “Effects of feed formula and farming system on the environmental performance of shrimp production chain from a life cycle perspective”, publicado en el Journal of Industrial Ecology.
En esta misma línea, Hamilton subraya que, desde la perspectiva de las emisiones de gases de efecto invernadero, son principalmente los productos agrícolas los responsables de la huella de carbono. La experta detalla que la mayoría de las recetas de alimento para camarón contienen aproximadamente un 70% de ingredientes vegetales. Además, especifica que las materias primas utilizadas en el alimento balanceado, asociadas con problemas de sostenibilidad, varían según las recetas, la edad del camarón y las prácticas locales de obtención de ingredientes.
Entre los ingredientes que impactan significativamente en la huella de carbono del pienso, Hamilton menciona la soya, el aceite de palma, la harina de pescado, el aceite de pescado y la harina de krill. “Monitoreamos de cerca estas materias primas para asegurarnos de contar con prácticas de obtención adecuadas que garanticen y documenten su adquisición responsable”, explica.
A nivel mundial, durante el año 2022, la totalidad de la soya que utilizaron en BioMar contaba con certificaciones de RTRS, ProTerra, Donau/Europe Soya o SSAP, según señala Hamilton. En relación al aceite de palma, el 100% estaba certificado bajo Roundtable on Sustainable Palm Oil (RSPO) o Green Palm. Además, el 100% de la harina de krill consumida tenía la certificación de
Marine Stewardship Council (MSC); y el 91% de la harina de pescado y el 86% del pescado cumplían con los estándares de Aquaculture Stewardship Council (ASC), MSC, MarinTrust, FIP o equivalentes.
Por otra parte, Hamilton explica que en BioMar utilizan el LCA no solo para calcular las huellas de carbono de las materias primas, sino también otros 17 indicadores, incluido el uso de agua, eutrofización de agua dulce, eutrofización marina y uso del suelo, entre otros. “Con un LCA completo, podemos identificar áreas críticas para mejoras. Por ejemplo, a nivel de procesamiento, el análisis de escenarios nos permite cuantificar los beneficios climáticos de cambiar de una caldera diésel a una eléctrica para el secado”, expresa.
La sostenibilidad del alimento balanceado también depende de conocer los programas y prácticas de producción de la materia prima utilizada, informa la gerente de Calidad de Skretting Latam, Karina Briones. “No es lo mismo hablar con un productor europeo que con uno norteamericano o uno de Sudamérica, porque cada país tiene diferentes prácticas de producción. Para nosotros es importante conocer cómo nuestros proveedores se van alineando a nuestros objetivos de sostenibilidad”, comparte.
En promedio anual, las dietas de Skretting Latam están compuestas en un 65% por ingredientes de origen vegetal, según informa Briones. Como parte de su enfoque en soluciones nutricionales e innovadoras para reducir su huella de carbono, esta empresa incorpora ingredientes novedosos
- FEBRERO 2024 SOSTENIBILIDAD 16

“La sostenibilidad del alimento balanceado comienza con las materias primas”.
Helen Ann Hamilton
Gerente
Global de Sostenibilidad de BioMar
que sustituyen en cierta medida a los ingredientes vegetales convencionales. La gerente de Calidad coincide en que los productos vegetales, especialmente la soya y sus derivados, son ingredientes críticos debido a su alto LCA e impacto en la huella de carbono.
Por esta razón, las certificaciones de sostenibilidad marcan el punto de partida en el proceso de abastecimiento de materias primas. Briones menciona que entre las certificaciones para los ingredientes vegetales se incluyen ProTerra y Round Table on Responsible Soy Association (RTRS), mientras que para los ingredientes de origen marino se encuentran MSC, Marine Trust y Marine Trust Improvement Program.
Adicionalmente, para reducir las emisiones GEI de sus productos, Skretting Latam implementa medidas como la optimización en el uso del agua y el control de consumos; el inicio de un programa de uso de empaques sostenibles y la implementación de un sistema de eficiencia energética en los procesos productivos de las fábricas. “Una de nuestras principales acciones en el 2024 es empezar a migrar del uso de Fuel Oil o búnker ecuatoriano hacia gas natural, lo cual nos permitirá reducir la huella de carbono operacional”, asegura.
Una perspectiva adicional a contemplar en relación con la producción sustentable de alimentos balanceados es la de la gerente de Desarrollo Sostenible y Comunicaciones Externas de Vitapro, María Alejandra Rivera. Ella indica que entre los sistemas que ejecutan para garantizar la sostenibilidad en Nicovita, la marca de Vitapro especializada en crear soluciones nutricionales para camarones y peces, se incluyen estrategias

“Es
importante conocer cómo nuestros proveedores se van alineando a nuestros objetivos de sostenibilidad”.
Karina Briones
Gerente de Calidad de Skretting Latam
tanto a nivel operacional, como de procurar una provisión responsable de materias primas a través de su cadena de valor.
Un ejemplo de ello es el Programa de Abastecimiento Sustentable (PAS), donde Vitapro colabora con sus proveedores para la optimización y certificación de los ingredientes clave. Esto ha traído como resultado el desarrollo de dietas con harinas de pesca entera 100% certificada MarinTrust o FIP.
“Conocer el origen de los ingredientes que utilizamos como materia prima es vital para evaluar y gestionar el impacto ambiental asociado con la producción de piensos. Nos permite trabajar para reducir impactos generados por el transporte, eligiendo fuentes más cercanas si están disponibles, así como optimizando rutas de transporte o promoviendo prácticas agrícolas más sostenibles en nuestros proveedores”, indica.
Asimismo, Rivera menciona que entre los mecanismos que adoptan para impulsar una producción de alimentos balanceados sostenible en el largo plazo se encuentran la incorporación de subproductos en las dietas para reducir la presión sobre recursos escasos; la innovación sostenible para desarrollar productos, prácticas y tecnologías que aporten a disminuir la conversión de alimento (FCA); incrementar la digestibilidad y la eficiencia del alimento; contribuir al cuidado del medio de cultivo, y la adopción y promoción de estándares y certificaciones sostenibles.
En el contexto de la búsqueda continua de soluciones sostenibles en la industria de alimento para acuicultura se destaca Houdek-Prairie Aquatech, una empresa
estadounidense que emplea la fermentación para reciclar subproductos del procesamiento agrícola, como la harina de soya y el grano destilado seco, y así producir un ingrediente para dietas comerciales que es fácilmente digerible y que reduce la dependencia de ingredientes menos funcionales y menos sostenibles, como la harina de pescado.
“Houdek fue fundada con el propósito de lograr la producción sostenible de alimentos. Un diálogo entre innovadores con una visión de conservación y el conocimiento para generar cambios llevó al desarrollo de la tecnología de fermentación, que transforma la harina de soya en una fuente segura y de alta calidad de proteínas para animales terrestres y acuicultura”, expresa la directora de Sostenibilidad en Houdek, Stephanie Armstead.
Este enfoque no solo mejora la sostenibilidad de la cadena de suministro de alimentos para camarón, sino que también contribuye a reducir la presión sobre los recursos marinos y promueve una mayor eficiencia en la producción. Houdek afirma que su ingrediente ME-PRO tiene más del 70% de proteína, reduce los compuestos antinutritivos de las plantas y aumenta la biodisponibilidad de fósforo.
La directora también señala que, dado que la harina de soya es el ingrediente principal, surge la necesidad de abordar las preocupaciones potenciales relacionadas con la deforestación. Es por eso que resalta la importancia de comprender los detalles de origen de los ingredientes para garantizar la trazabilidad y ofrecer un cálculo preciso de la huella de carbono.
“Obtenemos la harina de soya de entidades
17 - ABRIL 2024 SOSTENIBILIDAD

“Conocer
el origen de la materia prima es vital para evaluar y gestionar el impacto ambiental asociado con la producción de piensos”.
María Alejandra Rivera
Gerente de Desarrollo Sostenible
Comunicaciones
Externas de Vitapro
locales, ubicadas a menos de 200 km, que procesan soya cultivada en las Grandes Llanuras. La mayoría de ellas se encuentran dentro de nuestro estado natal de Dakota del Sur, en Estados Unidos. La harina de soya adquirida por Houdek se produce siguiendo el Protocolo de Aseguramiento de Sostenibilidad de la Soya Estadounidense (SSAP)”, comenta.
El Grupo de Trabajo de Acuicultura sobre la Huella Ambiental también ha detectado que ingredientes como la soya pueden contribuir significativamente al impacto ambiental general en la acuicultura. Asimismo, ha identificado que el cambio de uso de suelo, como la tala de bosques o deforestación para convertirlos en tierras agrícolas, puede representar un porcentaje significativo de la huella ambiental de los alimentos.
Sin embargo, no toda la soya es igual en su perfil nutricional e impacto ambiental. Blonk Consultants, una consultora especializada en servicios de investigación en el ámbito ambiental y de sostenibilidad agroalimentaria, realizó un estudio en el que utilizó datos de su base de datos Agri-Footprint™ para evaluar la huella ambiental de la soya de diferentes orígenes. La consultora empleó el LCA, que tiene en cuenta el impacto del cambio en el uso de suelo. Los resultados del análisis determinaron que la soya proveniente de Estados Unidos tiene una huella de carbono más reducida en comparación con la soya de otras procedencias.
El director regional para las Américas del Consejo de Exportación de Soya de Estados Unidos (USSEC), Carlos Salinas, sostiene que varios factores contribuyen a una menor huella de carbono en la producción de soya en el país. Estos incluyen rendimientos

“Houdek fue fundada con el propósito de lograr la producción sostenible de alimentos. Un diálogo entre innovadores con una visión de conservación y el conocimiento para generar cambios llevó al desarrollo de la tecnología de fermentación”.
Stephanie Armstead
Directora de Sostenibilidad en Houdek
más altos, un uso mínimo de fertilizantes, maquinaria eficiente, prácticas agrícolas de precisión y una sólida atención en la conservación del suelo. “Los agricultores de soya estadounidenses están dedicados a soluciones sostenibles, produciendo más mientras usan menos recursos e implementando prácticas agrícolas que reducen la huella de carbono”, comparte.
“Consumidores, empresas y gobiernos de todo el mundo están exigiendo más acción en sostenibilidad, especialmente en la reducción de emisiones de gases de efecto invernadero. Por lo tanto, los productores de alimento deberían considerar la huella de carbono, las credenciales de sostenibilidad y la oferta de soya de varios orígenes que ayuden a avanzar en los Objetivos de Desarrollo Sostenible de la ONU, y con ello ganar con sus clientes y consumidores”, relata Salinas.
En última instancia, la directora de SSP, Pamela Nath, apunta que la elección consciente de productos provenientes de prácticas sostenibles y la presión por parte de los profesionales de la acuicultura son fundamentales para impulsar los cambios necesarios en el desempeño ambiental de los alimentos balanceados en la industria.
“La sostenibilidad en la acuicultura de camarón es un esfuerzo colectivo que abarca toda la cadena de valor. Por este motivo, la colaboración y la transparencia son esenciales para la integración de consideraciones de sostenibilidad en cada etapa, desde la granja hasta el plato”, expone Nath. “En SSP, contamos con miembros asociados que, a pesar de no ser productores directos de camarón, tienen un compromiso con la producción
sostenible que contribuye significativamente a nuestros esfuerzos. Desde proveedores hasta académicos aportan su experiencia y recursos para impulsar la sostenibilidad en la industria”, continúa.
Por este motivo, y para sensibilizar a todos los participantes del sector acerca de los puntos críticos que requieren atención en la producción de alimento balanceado, SSP ha colaborado activamente con sus miembros asociados. A través del conocimiento y la experiencia de estas organizaciones, se busca que los actores de la industria acuícola estén mejor informados sobre las prácticas sostenibles que los fabricantes pueden implementar para minimizar el impacto ambiental de sus productos. Sobre todo, se busca que los consumidores estén concientes de que las plantas de alimento balanceado están preparadas para ofrecer soluciones y adaptar las dietas según las preferencias de los clientes, especialmente de aquellas que priorizan la sostenibilidad.
Para que estas prácticas sostenibles se vuelvan un estándar en la industria, es vital que el mercado respalde y valore las dietas que promueven la sostenibilidad. Esto implica reconocer que, aunque puedan tener un costo inicial mayor debido a sus requisitos específicos, impulsar estas prácticas es una inversión en el futuro del planeta y de la industria, así como en la calidad de nuestros productos•
Para más información sobre este artículo, contactar a: pnath@sustainableshrimp.org
18 - ABRIL 2024 SOSTENIBILIDAD

Camaroneros de India apuestan por el mercado local para mantener a flote la industria
Autor:
Yahira Piedrahita
Directora Ejecutiva de la Cámara Nacional de Acuacultura
ypiedrahita@cna-ecuador.com

Con una producción estimada cercana a las 850,000 TM en 2023, la industria camaronera de India afronta varios desafíos, tanto a nivel de producción (enfermedades, costos elevados) como de mercados (rechazo de producto, bajos precios, demandas legales en su principal mercado); por lo que tanto el gobierno como el sector privado analizan alternativas para afrontar la crisis y mantener la industria a flote. Este artículo recoge parte de la información y datos presentados durante Aqua India 2024, un evento que se desarrolló en Chennai (Tamil Nadu) entre el 15 y 17 de febrero.
India exportó aproximadamente 850,000 TM de camarón en 2023 y contribuye con más del 15% del camarón que se comercializa a nivel mundial. En cuanto a la capacidad instalada, de acuerdo con Madhusudhan Reddy, presidente de la Sociedad de Profesionales en Acuicultura (SAP), en el país existen 5 centros de multiplicación de reproductores (BMC), 600 fincas, 45 fábricas de alimentos balanceados y 667 plantas procesadoras, para soportar cultivos que se desarrollan en cerca de 200,000 hectáreas distribuidas en los estados productores. Los cinco principales estados en 2023 son Andhra Pradesh (78% de la producción total), Bengala Occidental (6.1%), Odisha (6.6%), Gujarat y los estados de Costa Oeste (2.3%) y Tamil Nadu (2.6%).
Se espera que la producción de camarón de cultivo aumente en los próximos años. Sin embargo, existen desafíos como una mayoritaria dependencia de reproductores importados, la escasez de gusanos poliquetos SPF para alimentar a los reproductores, tendencias de población impredecibles y cambios en las preferencias de los camaroneros sobre las líneas de reproductores importados. El cultivo de camarón blanco enfrenta además otras amenazas, incluidas enfermedades como mancha blanca, heces blancas (WFD), microsporidiosis hepatopancreática (causada por Enterocytozoon hepatopenaei, EHP) y mortalidad por goteo (Running Mortality Syndrome, RMS) que pueden ocasionar el fracaso total de un ciclo de cultivo (failure crops). Debido a esto, algunos productores han apostado por volver al cultivo de camarón tigre gigante (Penaeus
- ABRIL 2024 COYUNTURA 20
monodon), por lo que se espera que su producción aumente en los próximos años, con una mayor productividad y rentabilidad debido a factores como mayores tasas de éxito, mayor tamaño de cosecha y mercados potenciales para los camarones de tallas grandes.
La drástica caída de los precios del camarón en mercados internacionales también ha desencadenado una crisis entre los productores de India, con precios locales inestables y fluctuantes. Esta situación ha generado dificultades significativas, ya que no solo ven reducidos sus márgenes de ganancia y limitado el flujo de caja, sino que también se enfrentan a desafíos adicionales, como el incremento del costo de los insumos, las condiciones climáticas impredecibles y la lucha por mantener el liderazgo en su mayor destino, Estados Unidos, donde estiman que la demanda está saturada. Existe la necesidad de explorar nuevos mercados debido a los exigentes requisitos norteamericanos, las cuestiones relacionadas con las demandas de dumping y derechos compensatorios, el costo de los fletes, y la importancia de una percepción mejorada respecto a la calidad de sus productos.
Esta crisis ha puesto de manifiesto la necesidad urgente de buscar salidas sostenibles y estrategias a largo plazo para proteger a la industria camaronera y asegurar su supervivencia. Muy pocos camaroneros están ganando dinero, a pesar de que la producción va en aumento, por lo que, si la situación a nivel internacional no cambia, el 2024 será peor. Varias son las acciones que se proponen para mejorar las perspectivas de la industria; sin embargo, la apuesta más fuerte se dirige a la promoción del camarón para incentivar su consumo, especialmente en el mercado interno. La población de la India es de aproximadamente 1,440 millones de habitantes, la sexta parte de la población mundial, y más del 70% tiene una dieta no vegetariana. Sin embargo, se estima que más del 80% de la población no cubre los requerimientos diarios de proteínas y es preciso mejorar su alimentación. El consumo medio anual de pescado y mariscos per cápita es inferior a 8.0 kg, y el consumo estimado de camarón es de 100,000 toneladas. En comparación,
el ciudadano chino consume cerca de 40.0 kg y el estadounidense 23.0 kg1. Con una población en crecimiento que es cada vez más consciente de la salud y la nutrición, las oportunidades en el mercado interno para incentivar el consumo de pescado y mariscos son promisorias.
Con un incremento en el consumo anual per cápita de camarón de 1.0 kg por parte del 50% de la población india, la demanda anual bordearía las 700,000 TM. Este consumo podría contribuir significativamente a mejorar los precios a nivel internacional. Para lograr este objetivo, los expertos indican que India debe cumplir cuatro condiciones:
Disponibilidad: el camarón debe estar fácilmente disponible para la compra
Accesibilidad: la gente debe poder costearse el camarón
Conciencia: la gente debe estar consciente de que el camarón es una opción alimenticia frente a otras proteínas disponibles
Atracción: a la gente debe gustarle comprar y comer camarón
Para cumplir con lo anterior, se requiere un cambio en las preferencias alimenticias de los consumidores (que actualmente no ven el camarón como una opción), una estrategia para el almacenamiento y distribución del producto en los centros de consumo y campañas masivas de información respecto a los beneficios de consumir camarón.
De acuerdo con lo expuesto en Aqua India 2024, la Revista Aquacultura presenta a continuación la experiencia y desafío de 2
1 https://worldpopulationreview.com/country-rankings/ fish-consumption-by-country
empresas reconocidas en el mercado de India.
Falcon Marine Exports, localizada en Bhubaneswar, Odisha, es una de las principales exportadoras de camarón del país. Priyanka Mohanty, Directora Ejecutiva de la empresa, explicó que han planteado una estrategia de marketing nacional, como una iniciativa social encaminada a la promoción de los beneficios de los productos del mar, volviéndolos disponibles y atractivos para la población local, de modo que pueda incentivar el consumo para sus planes futuros, entre los que se puede mencionar los siguientes:
Diversificación más allá de los productos del mar: La empresa ha ampliado su gama de productos para incluir alimentos congelados como papas fritas y parathas (un tipo de pan indio tradicional). Esta diversificación permite a la empresa atender a una base de clientes más amplia y potencialmente aumentar sus flujos de ingresos.
Promoción de los beneficios de comer productos del mar: La empresa se centra en crear conciencia sobre los beneficios para la salud del consumo de productos del mar. Participan en diversas actividades, como pancartas en las tiendas y campañas en las redes sociales, para educar y atraer clientes.
Disponibilidad y accesibilidad: Garantizan que el pescado y el marisco estén fácilmente disponibles en los pueblos y ciudades costeras. Actualmente tienen presencia en 12 localidades muy cercanas a los mercados diarios locales. Además, ofrece

"Uno de los grandes desafíos para promover el consumo doméstico es cambiar la percepción del consumidor de que el producto fresco es mejor y prefiere comprar de los mercados húmedos y ofrecerles un producto congelado con mejores condiciones sanitarias y nutricionales".
Priyanka Mohanty
Directora Ejecutiva Falcon Marine Exports
- ABRIL 2024 COYUNTURA 21
COYUNTURA
una aplicación en línea y se ha asociado a Zomato y Swiggy (servicios de entrega a domicilio) para que sus productos sean fácilmente accesibles para los clientes.
Ofrecer un valor superior y mantener precios competitivos: El objetivo es brindar a los clientes un valor superior manteniendo sus precios competitivos. Se centran en una exhibición adecuada y un alto atractivo visual en sus puntos de venta, además de mantener la limpieza y ofrecer una mejor experiencia al cliente.
Centrarse en el segmento B2B y conocimiento masivo: Enfocarse en el segmento B2B, particularmente en la industria HORECA (hoteles, restaurantes y catering). También enfatizan la importancia del conocimiento masivo para mejorar el alcance de su marca y aumentar la rentabilidad.
Para Mohanty, el proyecto planteado presenta numerosos desafíos, que se detallan a continuación:
Luchar contra la percepción del consumidor: La acción inicial es cambiar la percepción del consumidor con respecto al precio y el producto. Esto sugiere que los clientes pueden tener nociones preconcebidas sobre el precio y la calidad de los productos ofrecidos por Falcon.
Competir con vendedores ambulantes: Los vendedores ambulantes ofrecen productos similares a precios más bajos; por lo que es preciso encontrar formas de diferenciarse de los vendedores ambulantes y atraer clientes a sus puntos de venta.
Retención de procesadores y cortadores de pescado: Esto implica enfrentar dificultades para encontrar y retener trabajadores calificados que puedan cortar y preparar de manera eficiente los productos.
Tensión financiera: Debido a la situación del mercado de camarón, la empresa enfrenta desafíos para administrar sus finanzas. Se puede lograr rentabilidad con mayores volúmenes, por lo que se espera que la economía mejore junto con los volúmenes de venta.
Por otra parte, Zhingalala es un restaurante especializado en recetas elaboradas con camarón. Está localizado en Surat, en el estado occidental de Gujarat y su propietario es Manoj Sharma, quien lleva más de 30 años en la industria y lidera la producción de camarón en la costa oeste de India. Sharma empezó a percibir la saturación en los mercados internacionales de camarón y decidió apostar por promover el consumo en el mercado doméstico, considerando el enorme potencial de consumo de India, que por su gran población y bajo consumo per cápita ofrece una alternativa para la comercialización del producto.
Zhingalala debe su nombre a dos palabras de la lengua local; zhinga (camarón) y lala (gente, personas), por lo que el nombre nace de una idea de acercar el camarón a la gente. Según Sharma, el camarón es conocido y es consumido por población que vive en las zonas costeras. Sin embargo, 90% de la población que vive en las zonas interiores del país nunca ha visto un camarón, mucho menos lo ha degustado. Junto a Vidya, su esposa, en 2019, empezó a buscar recetas elaboradas con camarón en todas las regiones de India que lo consumen, así como recetas internacionales que pudieran ser aceptadas por los comensales de Surat; y luego de un profundo trabajo probando con más de 300 opciones construyeron un

menú con las mejores recetas elaboradas a base de camarón, con los sabores de diferentes lugares de India y Asia, diseñado científicamente, pero siguiendo las recetas tradicionales, y que ofrece una primera gran experiencia para quienes nunca habían probado camarón.
En principio fue difícil porque, según Sharma, en la región de Gujarat (con uno de los más altos porcentajes de población vegetariana) se consume menos de 100 g de camarón per cápita. Pero la iniciativa ha sido una gran oportunidad para muchos, ya que el éxito de su proyecto significa que los productores de camarón tienen una opción para mantener la industria a flote. Actualmente Zhingalala ya es una marca en el sector de la acuicultura de India y es reconocida por muchos como una gastronomía de primera para promover el consumo de camarón.
Desde el sector productor, la Federación de Productores de Camarón de India ha presentado al gobierno un plan de acción para incentivar el consumo de camarón a nivel local.
La Federación de Productores de Camarón de India (PFFI) está conformada por asociaciones de productores de camarón de 9 estados del país, y su plan contempla tres puntos fundamentales: 1) establecer

- ABRIL 2024
22
Menú a base de camarones
Manoj y Vidya Sharma, propietarios de Zhingalala
un mercado doméstico sustancial para el camarón; 2) establecer redes de distribución fluidas para movilizar grandes volúmenes de camarón; y 3) garantizar una disponibilidad de camarón constante, con excelente calidad y precio, satisfaciendo la demanda del consumidor a lo largo de toda India por camarón de buena calidad. Según Balasubramaniam, Secretario general de PFFI, esto permitiría a los productores estabilizar el precio de venta y asegurar una demanda consistente, a la vez que beneficiaría a los consumidores, incluyendo tanto a los hogares como a la industria hotelera y de consumo de alimentos. El plan pretende atraer traer inversiones por más de 1.000 millones de rupias (USD 12 millones, aproximadamente) en bienes de capital y 10.000 millones de rupias (USD 120 millones, aprox.) en oportunidades de negocio en el sector del camarón.
Para lograrlo, inicialmente la Federación establecerá alianzas con 10 cooperativas de productores en Andhra Pradesh, las cuales permitirán asegurar la disponibilidad constante de camarón para vender un estimado de 100 toneladas diarias. El proyecto contará con centros de procesamiento (en las áreas cercanas a los centros de producción) y centros de distribución satélite (en las áreas cercanas a los centros de consumo) hasta donde el producto congelado llegará en camiones repartidores. De acuerdo con las estimaciones de la PFFI, involucrar directamente a los productores permitirá asegurar el suministro de camarón siempre fresco y sin el uso de aditivos que modifiquen su textura y sabor. Además, se eliminará a los intermediarios que encarecen el producto, manteniendo así el precio más estable. Luego de 5 años esperan expandirse y llegar a vender entre 300 y 500 toneladas diarias, apuntando a establecer el negocio en 100 ciudades a través de 100 cooperativas de productores.
No obstante, lo interesante y ambicioso de este plan, y los recursos que se pondrán desde el sector privado, se requiere un fuerte aporte de la infraestructura y equipamiento, especialmente las facilidades de procesamiento, almacenamiento en frío y camiones que movilizarán la materia prima desde las fincas hasta las plantas y desde estas hacia los centros de distribución; por lo que aún deben lograr el apoyo del gobierno


de India para cristalizar el proyecto y es probable que no empiece a operar durante este año y, hasta entonces, la situación de los productores seguirá siendo complicada.
Lo anteriormente descrito pone en evidencia la difícil situación de los productores de camarón de India, quienes, al igual que en Ecuador, están luchando por sobrevivir en medio de altos costos operativos, problemas de producción e incertidumbre sobre el mercado. Por ello, la opción de promover el consumo interno, en un mercado potencial que comprende la sexta parte de la población mundial, constituye una opción que ya tiene trabajando a varios actores desde diferentes aristas.
Para Ecuador, esta alternativa no tendría el mismo impacto debido al tamaño de la población ecuatoriana, por lo que la mejor opción disponible es incentivar la demanda en los mercados internacionales y seguir explorando nuevos mercados y presentaciones de producto para satisfacer las preferencias de un mayor número consumidores.
"Puede tomar 5 o 10 años lograr que la población india consuma camarón como lo hace con pollo o huevos, pero alguien tenía que empezar a promoverlo y por eso mi decisión de poner un restaurante. Zhingalala es una opción para que la gente tenga una primera experiencia inolvidable comiendo camarón y lo incorpore luego a su alimentación regular".
Manoj Sharma
Propietario de Zhingalala
"India tiene una población inmensa, con ciudades enormes que son centros de consumo. Es un buen momento para promover el consumo de camarón, especialmente de las tallas más pequeñas, preferidas por la población india. Esta iniciativa de venta local permitirá comercializar camarón a un precio justo para el consumidor, a la vez que permitirá ganancias a los productores".
Balasubramaniam
Secretario general de PFFI
Siendo la exportación la opción de Ecuador para poder vender el camarón que se produce en el país, es indispensable que todos los eslabones de la industria estén alineados y trabajando para lograr objetivos comunes, aplicando una estrategia clara que les permita sobrevivir en esta época de desafíos e incertidumbres. Los mecanismos para incentivar el consumo de camarón en los mercados también deben ir de la mano con políticas públicas y normativas claras, así como un firme compromiso de todos los actores con el cumplimiento de los más altos estándares en términos de calidad e inocuidad del producto, responsabilidad social y cuidado ambiental para lograr nuevos espacios de mercado y mantener los que se han logrado hasta el momento•
Vea el resumen de la entrevista a 3 empresarios de India.
Para más información sobre este artículo, contactar a: ypiedrahita@cna-ecuador.com
- ABRIL 2024
23
COYUNTURA


ARTÍCULOS TÉCNICOS
ÍNDICE
Edición 158 - Abril 2024 27 36 40 47 53 66
Identificación de nuevas cepas de Vibrio campbellii portadoras del plásmido pVA1 aislado de postlarvas de Penaeus vannamei afectadas por brotes de Necrosis
Hepatopancreática Aguda (AHPND)
Investigaciones muestran que las pruebas rápidas de campo (POC) solo detectan WSSV cuando hay alto nivel de infección, pero hay una mejor solución para los productores de camarón
Maximizando la productividad: el papel de los sensores en la acuicultura
Experiencias y desafíos de la eliminación de la ablación en reproductores de Penaeus vannamei en una maduración de Ecuador
La acumulación tisular de microplásticos de poliestireno provoca estrés oxidativo, lesión hepatopancreática y alteraciones del metaboloma en Penaeus vannamei
Aplicación de la secuenciación de nueva generación en el análisis del microbioma asociado a la producción camaronera
Identificación de nuevas cepas de Vibrio campbellii portadoras del plásmido pVA1 aislado de postlarvas de Penaeus vannamei afectadas por brotes de Necrosis Hepatopancreática Aguda (AHPND)
Caso: México

LAutores:
Sonia A. Soto-Rodríguez a,*
Bruno Gómez-Gila
Rodolfo Lozano-Olveraa
Karla G. Aguilar-Rendóna
Jean P. González-Gómez b
a CIAD, AC Mazatlán Unidad de Acuicultura y Gestión Ambiental, Av. Sábalo-Cerritos 82112, Mazatlán, México
b CIAD, AC Unidad Culiacán, Carretera a Eldorado Km 5.5, Campo El Diez, 80110 Culiacán, Sinaloa, México
ssoto@ciad.mx
as enfermedades que afectan a las postlarvas o estadios juveniles tempranos del camarón de cultivo son una de las principales limitaciones para la sostenibilidad de la industria camaronera. La Necrosis Hepatopancreática Aguda (AHPND), causada principalmente por Vibrio parahaemolyticus (Vp AHPND), continúa siendo la enfermedad bacteriana de mayor importancia económica que afecta tanto al camarón tigre (Penaeus monodon) como al camarón blanco del Pacífico (Penaeus vannamei) (FAO, 2013). La virulencia de Vp AHPND se debe a plásmidos conjugativos de aproximadamente 70 kpb (pVA1) que expresan la toxina binaria PirAB, que es homóloga a la toxina insecticida Pir (Han et al., 2015a). La toxina PirAB, estable en agua de mar, afecta al hepatopáncreas, órgano objetivo de la enfermedad, que es una glándula digestiva esencial implicada en el metabolismo con múltiples funciones fisiológicas e inmunológicas que se ven afectadas durante la infección (Muthukrishnan et al., 2019).
La AHPND se caracteriza por causar necrosis aguda del hepatopáncreas que conduce a su disfunción (Joshi et al., 2014; Soto-Rodríguez et al., 2015). En camarones juveniles y adultos, los signos clínicos de la enfermedad incluyen letargo, hepatopáncreas pálido, anorexia y tracto digestivo vacío (AguilarRendón et al., 2020). Las observaciones histológicas de camarones afectados por la AHPND incluyen desprendimiento masivo y redondez de las células epiteliales de los túbulos hepatopancreáticos hacia el lumen (lesión patognomónica), reducción de las vacuolas de reserva y, a medida que avanza la enfermedad, se produce una colonización bacteriana secundaria del hepatopáncreas dañado. El diagnóstico confirmatorio de AHPND en camarones también debe incluir, además de la lesión patognomónica observada en la etapa aguda de la enfermedad, la detección molecular de los genes pirA y pirB junto con infecciones experimentales (OIE, 2021). En México, como en otros países, se suele realizar una prueba de laboratorio PCR antes de enviar las postlarvas a una finca para su cultivo. Sin embargo, este procedimiento no puede garantizar que las postlarvas enviadas estén libres de AHPND. Los protocolos de PCR basados en la detección de los genes
- ABRIL 2024 SALUD
pirA y pirB se consideran los más sensibles; sin embargo, se recomienda un paso de enriquecimiento antes de extraer el ADN para muestras ambientales, como sedimentos y biopelículas, donde las bacterias están presentes en pequeñas cantidades (OIE, 2021).
Durante el cultivo de camarón, la AHPND puede alcanzar entre 90 y 100% de mortalidad; sin embargo, los camarones son más susceptibles en sus primeras etapas de vida a la intoxicación por cepas AHPND, con un umbral de densidad infectiva superior a 104 CFU ml—1 (Soto-Rodríguez et al., 2015). Varias especies de Vibrio se han identificado como causantes de AHPND en camarones peneidos, incluido Vibrio harveyi en Tailandia (Kondo et al., 2015), Vibrio owensii en China (Liu et al., 2015) y Vibrio punensis en América del Sur (Restrepo et al., 2018). Las cepas de Vibrio campbellii han sido implicadas con la AHPND en Guanxi, China (cepa 20130629003S01; Dong et al., 2017) y en un país latinoamericano (cepa LA16-V1; Ahn et al., 2017), aisladas en la provincia del Guayas en Ecuador (https://www.ncbi. nlm.nih.gov/bios ample/SAMN06909310/ ). Además, en un “país latinoamericano también se han aislado otras cuatro cepas de V. campbellii que poseen los genes pirAB” (Han et al., 2017), determinado mediante su amplificación por PCR y se demostró que al menos una causa AHPND en camarones. Asimismo, se ha comprobado experimentalmente que el plásmido conjugativo que alberga los toxigenes pirAB puede transferirse de manera horizontal entre la cepa original de V. campbellii y una cepa receptora de V. owensii en condiciones controladas de laboratorio (Dong et al., 2019).
En México, la enfermedad fue detectada en 2013 en camaroneras causando el 70% de pérdidas de producción (Nunan et al., 2014). Actualmente, la AHPND continúa produciendo grandes pérdidas en el cultivo de camarón, pero no se ha publicado ningún informe científico sobre la mortalidad en granjas y laboratorios de este crustáceo. La enfermedad representa una amenaza especial para el cultivo de camarón peneido debido a sus diversos agentes causales, complejidad y patogénesis desconocida, además de la naturaleza generalizada de esta
enfermedad. Recientemente, los laboratorios han sufrido esporádicamente bajas tasas de supervivencia de postlarvas de camarón, lo que ha sido atribuido a la presencia de marea roja en la zona costera; sin embargo, su etiología no ha sido confirmada.
Debido a la información antes mencionada, este trabajo estudió las causas de mortalidad y morbilidad en postlarvas de camarón con signos clínicos de AHPND provenientes de laboratorios comerciales en México, utilizando métodos de secuenciación de genoma completo (WGS).
Materiales y métodos
Condiciones de cultivo
Los días 2, 8 y 30 de junio de 2022, lotes de postlarvas de camarón fueron transportados desde tres laboratorios comerciales locales de camarón (Penaeus vannamei) a las instalaciones del CIAD, AC Unidad Mazatlán, para ser utilizados en experimentos de investigación. En los laboratorios, los camarones se mantuvieron en agua de mar de color marrón debido al uso regular de probióticos durante el cultivo de las larvas. Las postlarvas fueron transportadas en un contenedor plástico redondo de 1000 L lleno de agua de mar (35 g L—1 salinidad, pH 7.8–8.0 y 28–30 °C de temperatura), que también eran similares a las condiciones de transporte utilizadas por los laboratorios.
Luego de su llegada a las instalaciones del CIAD, los camarones fueron inmediatamente trasladados a tanques redondos de fibra de vidrio de 400 L y se mantuvieron acorde a las condiciones de cultivo del laboratorio (agua de mar natural con una salinidad de 35 g L—1, pH de 7.8-8.1, temperatura 30 °C y fotoperiodo de 12 h luz/12 h oscuridad). Se utilizó un sistema de agua de mar de flujo abierto para cada lote y las postlarvas se alimentaron tres veces al día con alimento comercial para postlarvas de camarón. No se emplearon probióticos. Se observaron dos veces al día los signos externos de la enfermedad, como el comportamiento de natación, los cromatóforos, el color del hepatopáncreas, el tracto digestivo vacío y la actividad larvaria. Todos los animales se mantuvieron siguiendo los estándares aceptados de cuidado animal humano. No se utilizaron especies protegidas o en peligro de extinción.
Recolección de muestras y aislamiento de bacterias similares a Vibrio
El estado de salud de las postlarvas de cada lote se comprobó el primer día de aclimatación. Para la detección del WSSV, IHHNV y TSV se siguieron los protocolos de PCR descritos en la OIE (2022). El sistema AP4 en una reacción de PCR anidada (Dangtip et al., 2015) se utilizó como diagnóstico presuntivo de animales clínicamente afectados, según lo recomendado por la OIE (2021). Durante la aclimatación del camarón, ocurrió un brote natural en cada lote entre las 72 a 96 h después de su llegada a las instalaciones, y se observaron mortalidad y signos clínicos característicos de la AHPND (Tran et al., 2013; Soto-Rodríguez et al., 2015). Las mismas pruebas para diagnóstico fueron aplicadas en cada lote. Una vez que se produjo la primera mortalidad, se montaron preparados húmedos a partir del hepatopáncreas de las postlarvas. Además, se pesaron los camarones y se diseccionaron asépticamente muestras de hepatopáncreas y estómago de postlarvas con signos clínicos de AHPND y organismos sin signos. Una parte de cada tejido se fijó en solución de Davidson para análisis histológico; la segunda parte se homogeneizó en 1.0 ml de solución salina estéril (NaCl al 2.5%) y se diluyó 10 veces en solución salina estéril. Se inocularon 100 microlitros de las suspensiones en agar TCBS (Difco™) por triplicado para enumerar las bacterias similares a Vibrio. Todas las placas se incubaron a 30 °C durante 24 h, y se registraron las unidades formadoras de colonias (CFU) por g. Las colonias amarillas y verdes se volvieron a sembrar en TSA (Difco™) + agar NaCl al 2.0 % (TSA+) para obtener cultivos puros. También se inocularon Vibrio aislados en CHROMagar™ y se incubaron a 30 °C durante 48 h. Finalmente, los aislados fueron criopreservados a 80 °C hasta su uso. En el segundo brote se realizó un paso preliminar de enriquecimiento de muestras previo a la extracción de ADN en las postlarvas sin signos clínicos de la enfermedad (infecciones subclínicas) recomendado por la OIE (2021). Diez muestras individuales obtenidas del estómago y hepatopáncreas se homogeneizaron en agua de peptona alcalina + suplemento de NaCl al 2.5% y se incubaron de 8 a 10 h a 30 °C con agitación. Más tarde, las muestras se centrifugaron a 8609 x g durante 10 minutos y el sedimento se analizó para detectar los genes pirAB
- ABRIL 2024 28
SALUD
utilizando el sistema AP4. Para el segundo y tercer brote, se tomaron muestras adicionales para futuros análisis metagenómicos.
Análisis histopatológico
El tejido hepatopancreático de postlarvas con signos clínicos de AHPND y sin signos clínicos se procesó mediante histología de rutina (Bell y Lightner, 1988). Las secciones de tejido incluidas en parafina se cortaron a 5 μm y se tiñeron con hematoxilina-eosinafloxina, según la metodología de Lightner (1996). Los cortes histológicos se observaron al microscopio óptico (Olympus CX31).
Secuenciación del genoma completo
Se obtuvo un total de 29 aislados bacterianos puros. El ADN se extrajo con el kit de extracción Promega Wizard Genomic según las instrucciones del fabricante. El ADN extraído se etiquetó con el kit Illumina Nextera XT, se indexó y se secuenció en la plataforma Illumina Miniseq para obtener una secuenciación a baja profundidad para identificar los aislados.
Las secuencias resultantes se limpiaron con el script propio Nextera Cleaner (https:// github.com/GenomicaMicrob/nextera_ cleaner ) y se ensamblaron con A5-miseq (Coil et al., 2015). La calidad de los genomas ensamblados se evaluó con Checkm2 (Chklovski et al., 2022) y luego se clasificó con GTDB-Tk v2.1.0 (Chaumeil et al., 2022). También se calcularon los valores promedio de identidad de nucleótidos (ANI) con OAT (Lee et al., 2016), comparando las cepas tipo de cada especie. En total, ya están disponibles dos genomas de V. campbellii AHPND+ y en este estudio se incluyen seis nuevos. Comparamos la arquitectura del plásmido pVA1 de estos ocho genomas, incluido un genoma de V. parahaemolyticus (M0904, González-Gómez et al., 2020) como referencia. Se obtuvo un pangenoma de V. campbellii con anvi’o v7.1 (Eren et al., 2015) que, además de los ocho genomas de AHPND+, incluyó la cepa tipo de la especie (ATCC 25920T).
Análisis estadístico
Para la enumeración de bacterias similares a Vibrio y colonias verdes, primero se analizó la data para determinar si estaban distribuidos normalmente utilizando la prueba de normalidad de Shapiro-Wilk, y la
homogeneidad de las varianzas utilizando la prueba de Levene. Si se rechazaba la normalidad, se utilizaban comparaciones post hoc por pares de la prueba de suma de rangos de Mann-Whitney para comparar las medianas entre los grupos. Todos los análisis se realizaron utilizando un grado de significancia de 0.05.
Resultados
En el primer, segundo y tercer brote, el peso de las postlarvas fue de 80 a 190 mg, 180 a 310 mg y 80 a 240 mg, respectivamente. Al inicio de la aclimatación no se observaron signos clínicos de enfermedades y la detección por PCR del WSSV, IHHNV, TSV y AHPND fue negativa en todos los lotes.
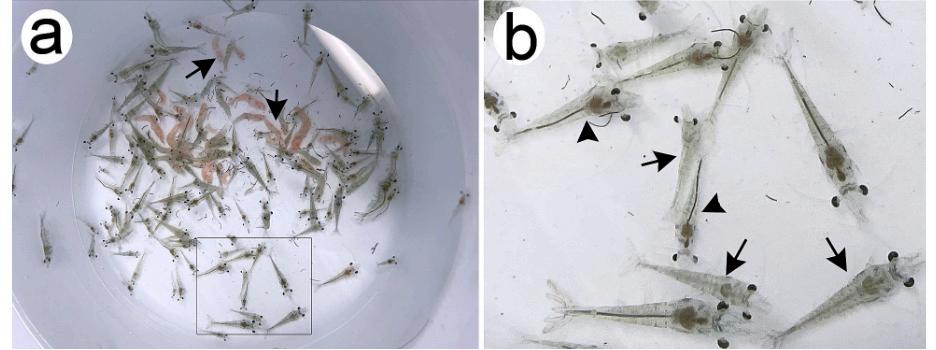
Fig. 1. Fotografías de postlarvas de P. vannamei de un brote natural de AHPND. a. Camarón del primer brote, mostrando camarones muertos (flecha), moribundos y normales. b. En el cuadro de la Fig. 1a, los camarones enfermos con signos clínicos de AHPND (flechas) y los camarones sin signos clínicos exhibieron un tracto digestivo completo (estómago e intestino) y hepatopáncreas normal con la pigmentación típica (punta de flecha).

Fig. 2. Preparaciones húmedas del hepatopáncreas de postlarvas enfermas de P. vannamei del primer y segundo brote con signos clínicos de AHPND (a–c). d. Organismos sin signos clínicos. a. Microfotografía de los túbulos hepatopancreáticos que muestra un desprendimiento masivo y acumulación de células epiteliales en el lumen (flecha). b-c. Falta de vacuolas en las células R y B (flecha), deformidades en los túbulos (punta de flecha) y melanización en los túbulos (asterisco). d. Túbulos con abundantes vacuolas en células epiteliales y morfología normal.
- ABRIL 2024 29 SALUD
Sin embargo, cuando se aplicó el método de enriquecimiento (segundo lote), 7 de cada 10 muestras de postlarvas sin signos clínicos fueron presuntamente positivas para la AHPND.
Durante los primeros días de aclimatación, las postlarvas exhibieron actividad normal en todos los lotes. Sin embargo, después de tres o cuatro días, se produjo una mortalidad que oscilaba entre el 70 y el 80% en todos los tanques (Fig. 1a). Se observaron signos clínicos de AHPND, como tracto digestivo vacío (estómago e intestino), hepatopáncreas pálido y atrofiado, natación errática, anorexia y letargo (Fig. 1b).
Preparado húmedo de hepatopáncreas
El preparado húmedo de postlarvas con signos clínicos de AHPND mostró una menor refringencia a la luz del tejido hepatopancreático que las postlarvas sin signos clínicos (Fig. 2). Los camarones enfermos mostraron un color pálido a blanquecino y pocas vacuolas en las células de los túbulos hepatopancreáticos, un epitelio tubular plano y ausencia de gotitas de lípidos y vacuolas secretoras en las células R y B (respectivamente); además, se observó claramente un desprendimiento masivo de células epiteliales y un túbulo deformado (debido a la acumulación de células necróticas en la luz tubular), lo que sugiere una etapa aguda de AHPND (Fig. 2a). Los camarones con hepatopáncreas blanquecino mostraron la menor refringencia y lesiones necróticas melanizadas en los túbulos (Fig. 2b-c). Los camarones sin signos clínicos de AHPND mostraron más refringencia en el tejido hepatopancreático y abundantes gotitas de lípidos y vacuolas secretoras (Fig. 2d).
Observaciones histopatológicas
Todas las postlarvas con signos clínicos de AHPND de los tres brotes mostraron cambios histopatológicos asociados con la etapa aguda, terminal o de remisión de AHPND (AHPND+) (Fig. 3). Las lesiones del hepatopáncreas en la etapa aguda mostraron desprendimiento masivo de células en la luz del túbulo, infiltración hemocítica leve y falta de vacuolas en las células epiteliales tubulares (Fig. 3a - b). El hepatopáncreas con lesiones en etapa terminal mostró melanización severa y necrosis del tejido
hepatopancreático con abundante masa de bacterias en el tejido necrótico (Fig. 3c). Las postlarvas sin signos clínicos de AHPND no mostraron cambios patológicos (AHPND-) (Fig. 3d).
Enumeración de bacterias similares a Vibrio
Las densidades de Vibrio y colonias verdes en TCBS mostraron resultados diferentes para cada brote de AHPND. Como los datos carecían de normalidad, se utilizó la prueba de suma de rangos de MannWhitney para comparar Log CFU g—1 entre el hepatopáncreas y el estómago, así como el porcentaje de colonias verdes en TCBS de postlarvas AHPND+ y AHPND- para los tres brotes (Tabla 1). El brote 1 mostró diferencias significativas (p = 0.045) entre la densidad de bacterias similares a Vibrio del hepatopáncreas y el estómago de los organismos AHPND+. En los tres brotes, los valores Log CFU g—1 del hepatopáncreas era más alto que el estómago de los camarones AHPND+. Además, el hepatopáncreas de las
postlarvas de AHPND+ tuvo valores más altos que el hepatopáncreas de los organismos AHPND-. El brote 2 mostró diferencias significativas (p = 0.021) entre la densidad de colonias verdes en el hepatopáncreas y el estómago de los organismos AHPND+. Las colonias verdes en el hepatopáncreas de las postlarvas de AHPND+ oscilaron entre el 58 y el 77%, en comparación con las postlarvas de AHPND-, que oscilaron entre el 13 y el 51%.
Identificación de especies de los brotes
Se identificaron un total de 11 especies conocidas (Tabla 2) y dos potenciales nuevas especies de Vibrio también fueron obtenidas; de los 29 genomas, 27 pertenecían a Vibrio y 2 a Photobacterium damselae. Las especies potencialmente nuevas tenían un valor de ANI por debajo del umbral estándar del 95% para delimitar una especie; Vibrio sp. nov. 1 (un genoma) está estrechamente relacionado con Vibrio owensii del clado Harveyi y Vibrio sp. nov. 2 (un genoma) de Vibrio tubiashii, perteneciente al clado Orientalis
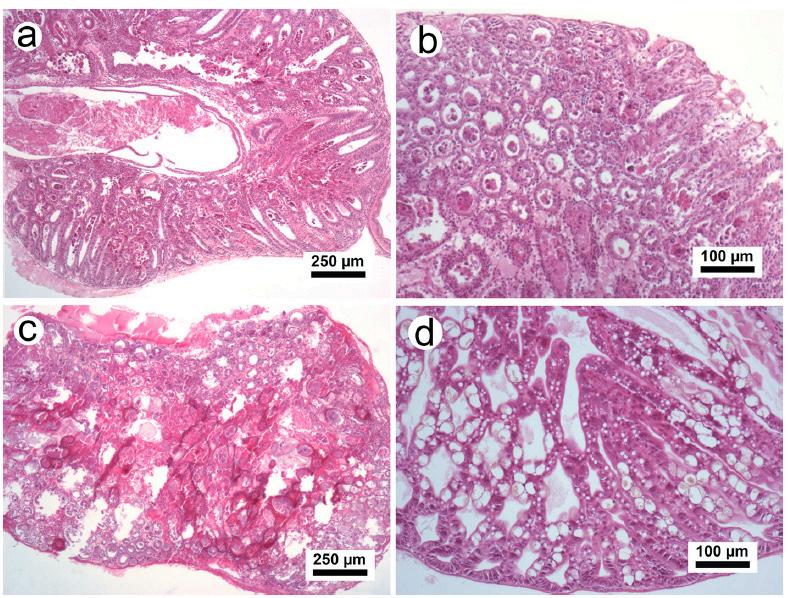
Fig. 3. Microfotografía del hepatopáncreas de postlarvas de P. vannamei procedentes de brotes naturales. a–b. Observación del hepatopáncreas en la etapa aguda de AHPND, desprendimiento masivo de las células epiteliales, baja infiltración hemocítica y acumulación de células en el lumen tubular. c. Hepatopáncreas con necrosis severa, melanización, infiltración hemocítica y proliferación bacteriana asociada a la etapa terminal de AHPND. d. Hepatopáncreas sin cambios patológicos, epitelio tubular normal con abundantes vacuolas en las células R y B. Tinción de hematoxilina-eosinafloxina.
- ABRIL 2024 30
SALUD
Las seis cepas mexicanas de Vibrio campbellii están muy relacionadas (> 0.9996 ANI); aun así, cada una tiene algunos genes, entre 4 y 20, que se encuentran solo en esa cepa (Fig. 4; anvi’o), por lo que no pueden considerarse clones.
Análisis de plásmidos
La arquitectura del plásmido pVA1 de los genomas analizados no difirió en más que unos pocos nucleótidos. El transposón Tn6264 que porta el operón pirAB también es idéntico en todos los genomas, excepto en el de la cepa 20130629003S01 de China, donde se encontró una tercera transposasa de la familia IS5 insertada y alterando la supuesta proteína de la familia de anclaje de la superficie de la pared celular, ubicada en el operón aguas arriba de pirAB (Figs. 5, 6). Esta doble inserción también provocó que parte del primer operón y todo el operón pirAB cambiaran de dirección (Fig. 6), y además el transposón creció de 5537 nucleótidos a 6600. El segundo transposón (Tn3) encontrado en este mismo plásmido está ausente en la cepa china 20130629003S01 (Fig. 5).
Los genes pirAB, responsables de producir la delta-endotoxina que causa la AHPND, solo se detectaron en los seis genomas identificados como Vibrio campbellii, en lo que parece ser un plásmido conjugativo muy similar al descrito previamente para la cepa M0904 de Vibrio parahaemolyticus recopilada en 2013 en una localidad de Sinaloa, México.
Los aislados de V. campbellii fueron las especies más abundantes, seguidos de V. parahaemolyticus con cinco aislamientos (ver Fig. S1 en material complementario). El color de las colonias en CHROMagar™ Vibrio fue diverso; la mayoría de los aislados presentaron color malva (incluido P. damselae), excepto Vibrio fortis y Vibrio rotiferianus, que presentaron un color turquesa; Vibrio sp.nov. 2 era translúcido, Vibrio parahaemolyticus era pálido o azul marino y Vibrio alginolyticus era crema.
Discusión
La aparición de brotes en el cultivo de camarón es compleja; estos están asociados con múltiples factores ambientales, el huésped y la presencia de patógenos. El rápido crecimiento de la producción de camarón
Tabla 1.- Comparación entre medianas de Log CFU g—1 y porcentaje de colonias verdes en TCBS del hepatopáncreas y estómago de postlarvas de Penaeus vannamei para cada brote. Mediana ± error estándar de la mediana, n estuvo entre cinco y diez muestras.
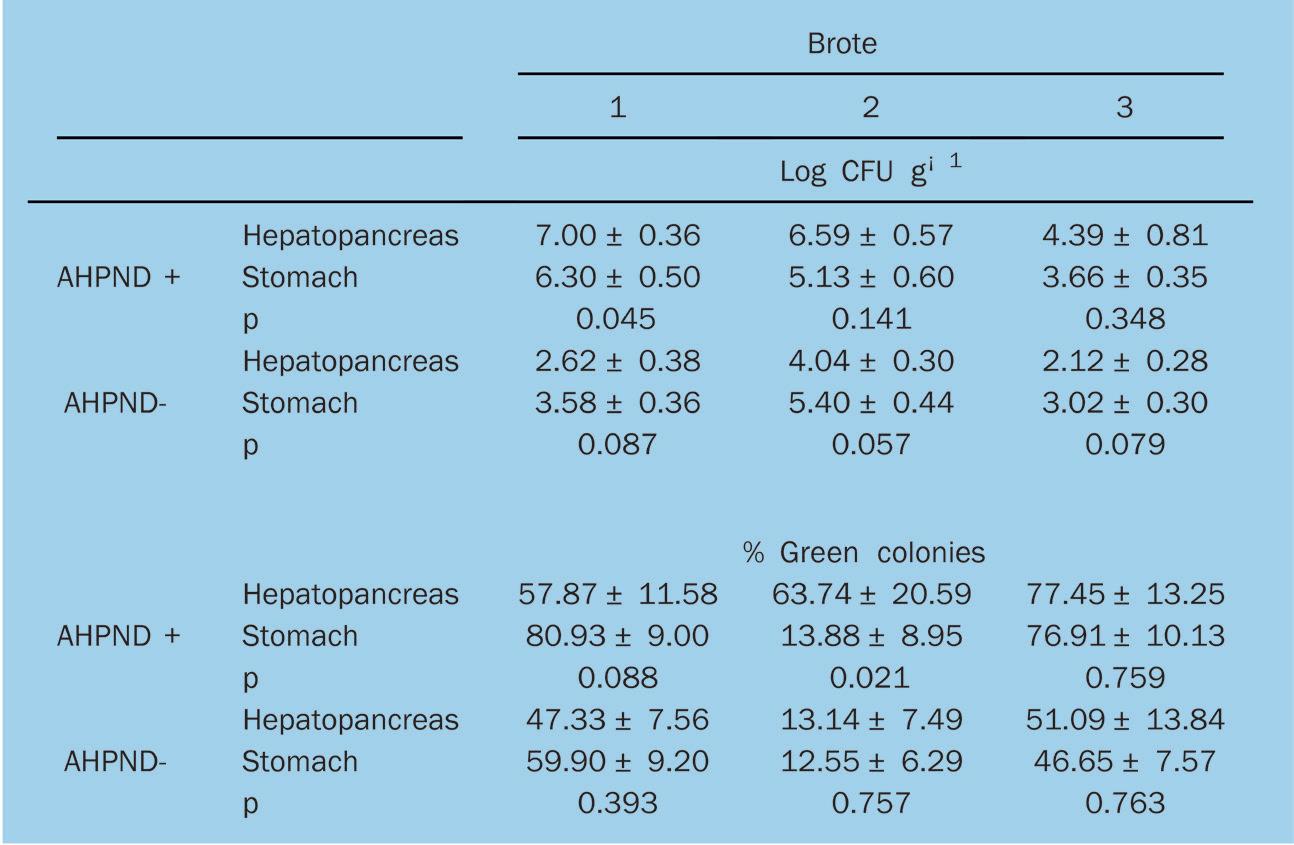
ha promovido el desarrollo de la industria de semillas y la producción anual de larvas de camarón ha superado los 1.5 trillones (Wang et al., 2021). Desgraciadamente, los laboratorios de camarón se encuentran actualmente amenazados por enfermedades frecuentes y graves. El riesgo de desarrollar AHPND aumenta con las condiciones de cultivo de alta temperatura del agua, salinidad y densidad de población de postlarvas (Boonyawiwat et al., 2017). El noroeste de México domina la producción semiintensiva de cultivo de camarón, con la mayoría de los laboratorios comerciales ubicados en la región. Específicamente, el estado de Sinaloa alberga alrededor de 900 camaroneras y 50 laboratorios de precría, todos ubicados estratégicamente dentro de instalaciones de bioseguridad con el objetivo de mejorar la producción. Sin embargo, en 2020 y 2021, el 38% y 31% de las camaroneras, respectivamente, se vieron afectadas por enfermedades infecciosas (https://cesasin.mX/programacrustaceos/ revisado en diciembre de 2022). La AHPND sigue siendo una preocupación importante para el gobierno mexicano y la industria camaronera (SENASICA, 2020). Aunque la mortalidad en los laboratorios de camarón se ha atribuido principalmente a las toxinas de la proliferación de algas en los últimos años, los resultados presentados aquí demuestran que los brotes en postlarvas se asociaron con la presencia de aislados de pirAB+ , incluso en densidades por debajo del umbral
infeccioso (Soto-Rodríguez et al., 2015).
En este estudio, los brotes de AHPND ocurrieron entre 72 y 96 h después de transportar las postlarvas. Los signos clínicos y las lesiones histopatológicas se parecían a los reportados anteriormente para AHPND (Tran et al., 2013; Soto-Rodríguez et al., 2022; Lozano-Olvera y Abad-Rosales, 2023). Los indicadores comunes incluyeron tractos digestivos vacíos, hepatopáncreas pálido a blanquecino y desprendimiento masivo de células epiteliales, característico de la etapa aguda de AHPND. Además, se observaron hepatopáncreas melanizados con necrosis durante la etapa terminal y de remisión. Las cepas de V. campbellii pirAB+ se utilizarán en ensayos con postlarvas y camarón juvenil para registrar el proceso infeccioso de la AHPND en estudios futuros. Aunque la OIE (2021) no considera que la histopatología sea un diagnóstico confirmatorio de AHPND, este análisis muestra evidencia de la enfermedad clínica, incluido el grado y el desarrollo de las lesiones en el tejido objetivo. Por tanto, este método podría utilizarse para la detección y confirmación de infecciones clínicas, basándose en la presencia de lesiones hepatopancreáticas típicas de la fase aguda de la enfermedad, como desprendimiento masivo de las células epiteliales y acumulación de células muertas en el lumen, lo que solo se ha observado en camarones afectados por AHPND (Tran et al., 2013; Joshi et al., 2014; Soto-Rodríguez et
- ABRIL 2024 31 SALUD
al., 2015; Hong et al., 2016; Restrepo et al., 2018; Aguilar- Rendón et al., 2020; Han et al., 2020).
El análisis histopatológico y del preparado húmedo de las postlarvas que mostraban signos de AHPND reveló una progresión variable de la enfermedad dentro de cada brote, lo que sugiere una infección gradual que no afectó a toda la población. Estos eventos pueden haber sido el resultado de la disbiosis del microbioma dentro del ecosistema del tanque, potencialmente causada por la eliminación de los probióticos aplicados en el laboratorio a su llegada; el agua clara podría favorecer a bacterias de rápido crecimiento, incluido Vibrio sp. pirAB+ (De Schryver et al., 2014). En este estudio, los microorganismos benéficos del agua de cultivo se eliminaron con las condiciones de transportación y laboratorio (5–10 μmde filtro agua de mar, 29 ±1°C, aireación y fotoperíodo de 12 h luz/12 h oscuridad). Actualmente en México, una práctica común en los laboratorios de camarón y precría es el uso de sistemas de agua madurada, que significa la aplicación de diversos probióticos comerciales y caseros, así
como “postbióticos”, enzimas, levaduras, melazas y en ocasiones, una combinación con tecnología biofloc. El sistema de cultivo de agua comprende un complejo consorcio de bacterias, levaduras y protozoos que cohabitan con las larvas de camarón. Silva et al. (2012) reportaron que un probiótico comercial no influyó en la supervivencia y el crecimiento de las postlarvas de P. vannamei, aunque hubo una reducción en el recuento de especies similares a Vibrio en el agua y en el camarón. Mientras tanto, Wang et al. (2020) encontraron una alta proporción de Vibrionaceae en zoea1 y postlarva 1 después de cinco probióticos comerciales. Los laboratorios y precría de camarón deben tener cuidado en el uso de productos comerciales durante el cultivo larvario.
En este estudio, los resultados de los vibrios en postlarvas dependieron del laboratorio; sin embargo, se encontró más densidad similar a Vibrio y colonias verdes en el hepatopáncreas de las postlarvas AHPND+ en comparación con los camarones AHPND–, lo que puede indicar que la infección secundaria es más rápida en las postlarvas. Las prácticas actuales en laboratorio,
precría y producción de camarón promueven intercambios genéticos mediante elementos móviles como los plásmidos, lo que ha provocado la aparición de nuevas poblaciones virulentas de Vibrio sp., causando AHPND como se reporta aquí, con consecuencias potencialmente devastadoras para el cultivo de camarón (Fu et al., 2020). Recientemente, Intriago et al. (2023) reportaron especies similares a Vibrio que portaban genes Vp pirAB en postlarvas de P. vannamei de laboratorios de América Latina. Desafortunadamente, estos aislados solo se identificaron bioquímicamente utilizando API20E. El WGS debe usarse como el único método para alcanzar precisión a nivel de especie (Culot et al., 2021), es el método recomendado para una identificación precisa y se utiliza en pocos trabajos para confirmar especies de Vibrio (Gómez-Gil et al., 2014; Gómez-Jiménez et al., 2014; Restrepo et al., 2016; González-Gómez et al., 2020). Este es el primer reporte utilizando WGS de cepas de V. campbellii, portadoras de los genes tóxicos pirA y pirB aislados de postlarvas de camarón en laboratorios comerciales mexicanos; anteriormente, las especies de Vibrio que causaban brotes de AHPND
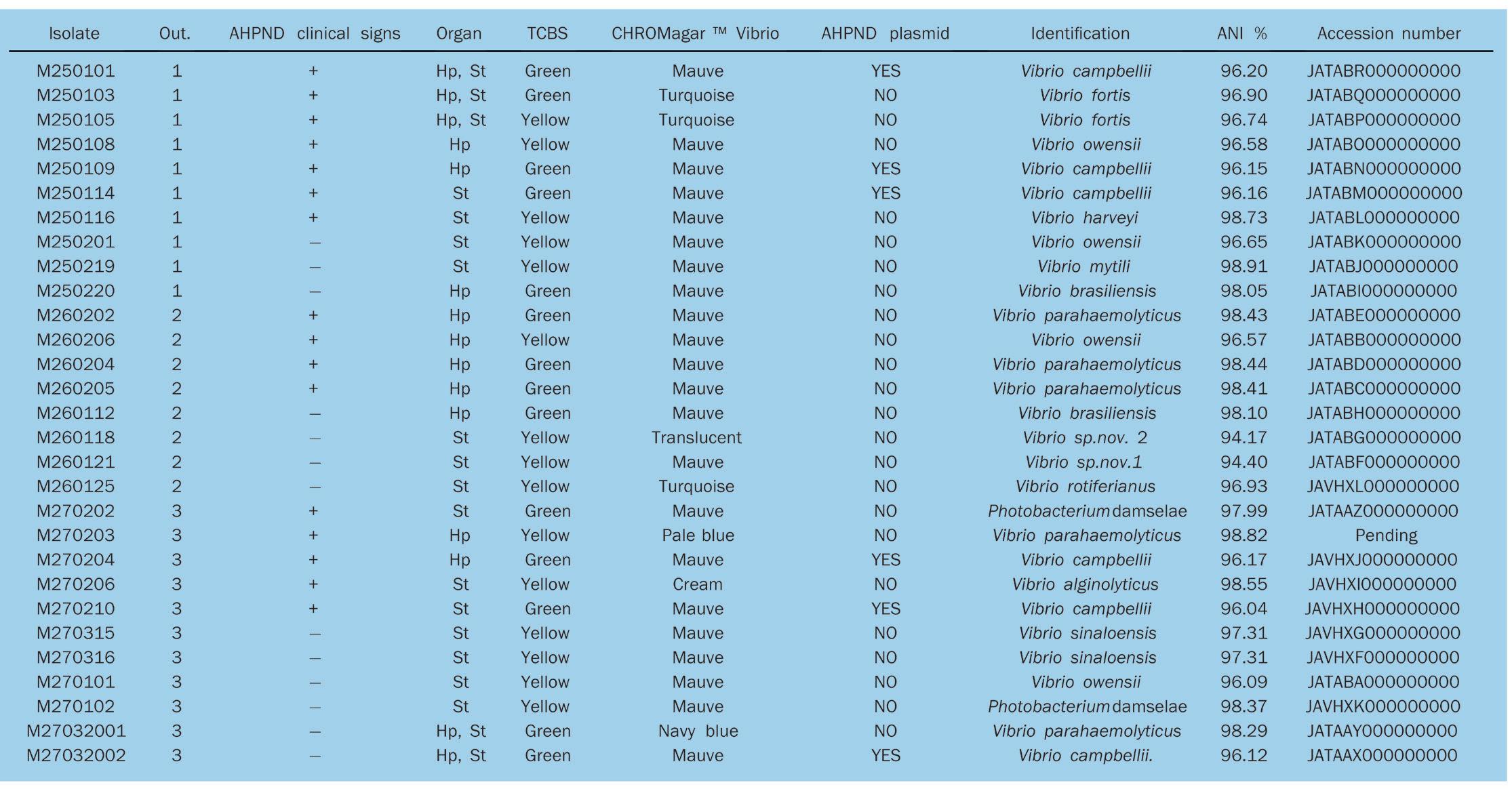
- ABRIL 2024 32 SALUD
Tabla 2.- Secuenciación completo del genoma de los aislamientos de postlarvas de Penaeus vannamei afectadas por AHPND en México.
Out.: brote; Hp: hepatopáncreas; St: estómago; TCBS: agar tiosulfato, citrato, sales biliares y sacarosa; ANI: Identidad media de nucleótidos.
ocurrían solo en camaroneras (Nunan et al., 2014; Soto-Rodríguez et al., 2015; LópezLeón et al., 2016). La identificación confiable de cepas de Vibrio spp. en camaroneras es muy importantes ya que dicha identificación podría usarse para monitorear la propagación y aparición de la AHPND; seguramente seguirán apareciendo nuevas especies de Vibrio pirAB+ en camaroneras de todo el mundo.
Además, se identificaron cuatro aislados de V. owensii y cinco de V. parahaemolyticus sin los genes pirAB como parte de la comunidad bacteriana de postlarvas, aunque esos genes han sido identificados en otras cepas (Liu et al., 2015; Restrepo et al., 2016). Otras especies de Vibrio, comúnmente asociadas con el agua de mar, también fueron identificadas con menor frecuencia, como parte de la microbiota de las postlarvas de los brotes de AHPND. Nuevamente, las colonias similares a Vibrio que causaban AHPND mostraron diversos colores en CHROMagar™ Vibrio, como reportaron Soto-Rodríguez et al. (2019). La mayoría de los aislados eran colonias de color malva a pesar de que crecieron como colonias amarillas en TCBS, como V. owensii, V. harveyi, Vibrio mytili y Vibrio sinaloensis, incluido P. damselae. Para la detección o enumeración de bacterias patógenas potenciales en programas de vigilancia, no recomendamos el uso de criterios de fenotipo de color.
Desde la aparición de AHPND, los estudios epidemiológicos se han centrado principalmente en las cepas de AHPND+ de V. parahaemolyticus, dada su amplia información y la disponibilidad de un número significativo de genomas. Sin embargo, las cepas de V. campbellii han demostrado tasas de mortalidad comparables a las de las cepas de V. parahaemolyticus, lo que justifica una cuidadosa consideración (Dong et al., 2017b). Esta investigación reveló que los seis genomas de V. campbellii recién secuenciados forman un clado distinto de las cepas ecuatorianas y chinas pirAB+ previamente documentadas. Aunque el número de genomas reportados para esta especie ha aumentado, sigue siendo limitado para un análisis evolutivo concluyente. Sin embargo, esta limitación se ve mitigada en cierta medida por el impacto significativo de la transferencia horizontal de plásmidos,
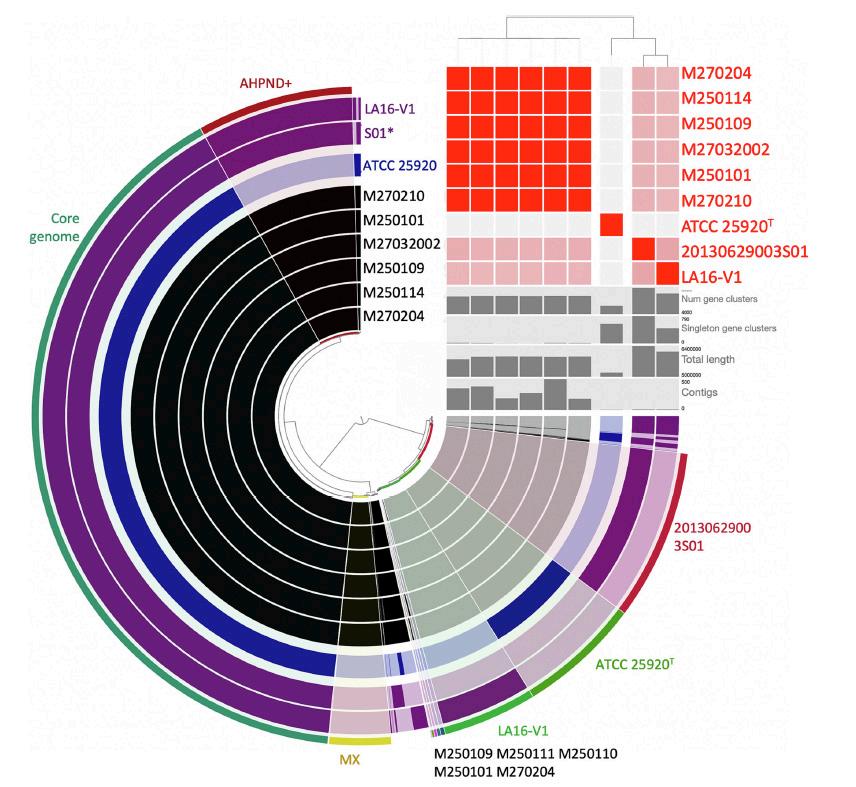
Fig. 4. Representación del pangenoma de las cepas de Vibrio campbellii que causan AHPND. Contenedores (círculo externo): Genoma central = grupos de genes presentes en todos los genomas analizados; AHPND+ = grupos de genes presentes en todas las cepas AHPND+; MX = grupos de genes encontrados solo en las cepas mexicanas; códigos de cepa = grupos de genes que se encuentran solo en esa cepa. Cuadrados rojos = valores ANI (>96%). Genomas ordenados filogenéticamente según los valores ANI (dendrograma). Pangenoma calculado con anvi’o v7.1 utilizando la cepa tipo ATCC 25920T como referencia. (Para la interpretación de las referencias al color en la leyenda de esta figura, se remite al lector a la versión web de este artículo).
ampliamente documentada en la literatura (Dong et al., 2019a; Muthukrishnan et al., 2019; Fu et al., 2020). En consecuencia, los análisis de plásmidos tienen el potencial de brindar información invaluable sobre la dinámica de la enfermedad. Hasta la fecha, se producen brotes de AHPND en todo el mundo y pueden ser causados por múltiples especies de Vibrio. Por lo tanto, se debe enfatizar la importancia de establecer programas de vigilancia sanitaria en los laboratorios de camarón para detectar la presencia del plásmido virulento. Curiosamente, la OIE (2021) no considera al WGS como un diagnóstico confirmatorio de AHPND, aunque este método identifica la presencia de los genes pirAB sin ningún error. Este método debe incluirse como diagnóstico confirmativo de AHPND en camarón.
A lo largo de los años, la arquitectura de los plásmidos pVA1 se ha destacado por su alta conservación. Sin embargo, también han surgido diferencias puntuales, en particular la presencia o ausencia del transposón Tn3, la inclusión de transposasas IS903 en Tn6264 y las deleciones en la región pirAB (Han et al., 2015b; Han et al., 2017; Yan et al., 2019; González-Gómez et al., 2020). El transposón Tn3 se ha identificado como un factor distintivo entre las cepas AHPND+ de V. parahaemolyticus en América y Asia, aunque también se ha reportado su presencia en el plásmido de una cepa vietnamita (Kumar et al., 2018). Esta observación parece extenderse a la cepa LA16-V1 (Ecuador) de V. campbellii que alberga el transposón; mientras que está ausente en 20130629003S1 (China), a pesar de
- ABRIL 2024 33 SALUD
los altos valores de ANI. Este fenómeno se aclara mediante la transferencia horizontal de plásmidos entre especies coexistentes dentro de regiones geográficas (Dong et al., 2019b). En la cepa china de V. campbellii, se observó la inserción de la transposasa IS903, junto con una reorientación del operón pirAB; curiosamente, esta alteración no impidió su capacidad de causar AHPND (Dong et al., 2017a). Se han detectado alteraciones comparables en V. parahaemolyticus, ejemplificadas por la cepa R14 donde la inserción de transposasa ocurre aguas arriba del gen pirA, lo que resulta en la ausencia de signos clínicos de AHPND en P. vannamei (Kanrar y Dhar, 2018). Sorprendentemente, las deleciones en la región pirAB estuvieron ausentes en los plásmidos de V. campbellii, en contraste con las observaciones en los plásmidos de las cepas de V. parahaemolyticus reportadas por múltiples autores (Han et al., 2017; Yan et al., 2019), incluidos casos de resiembras in vitro (Xiao et al., 2017).
Aunque la OIE (2021) recomienda el sistema AP4 como método de PCR para el diagnóstico de la AHPND, en este estudio, las postlarvas al llegar de los laboratorios fueron negativas usando ese método. Sin embargo, el cultivo preliminar en caldo de enriquecimiento permitió la multiplicación de V. campbellii pirAB+, aumentando la sensibilidad de la prueba. Identificamos que sin el cultivo de enriquecimiento existe una alta probabilidad de resultados falsos negativos. Por tanto, este protocolo debe incluirse en la detección de genes pirAB mediante PCR. Se deben revisar los kits comerciales y los diagnósticos oficiales de AHPND para comprobar la confiabilidad de los métodos de PCR.
Conclusiones
Postlarvas de P. vannamei aparentemente sanas de laboratorios de camarón desarrollaron signos clínicos y lesiones histopatológicas características de AHPND después de 72 a 96 h de transporte.
El plásmido tipo pVA1 fue identificado en todos los genomas de las cepas de V. campbellii, lo que confirmó por primera vez la presencia de genes pirAB en postlarvas de laboratorios comerciales mexicanos, lo que representa una gran amenaza para la industria camaronera. La mayoría de los

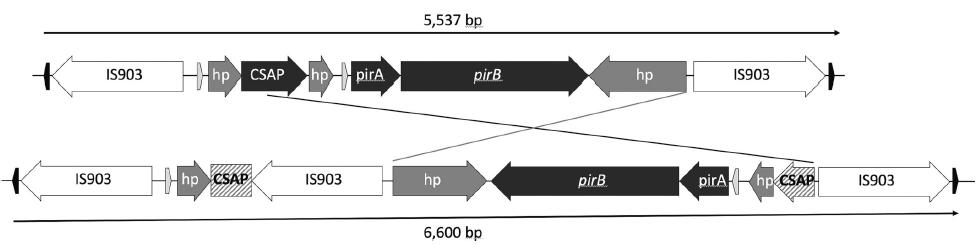
Fig. 6. Arquitectura del transposón Tn6264 del plásmido pVA1 que porta los genes pirAB. Arriba, arquitectura de transposón original presente en la cepa M0904 de V. parahaemolyticus y en todas las cepas de V. campbellii, excepto la cepa 20130629003S01 de Guanxi, China (=CP020078, abajo). Flechas blancas, genes de transposasa (IS903); flechas de color gris oscuro, gen hipotético de proteína (hp); flechas negras, genes con función conocida; pequeñas flechas de color gris claro, promotores potenciales; pequeñas flechas negras, repetir secuencias; cajas texturizadas, gen truncado. CSAP, supuesta proteína de la familia de anclaje de la superficie de la pared celular.
aislamientos de los brotes fueron vibrios (93%), incluidas dos nuevas especies, como parte de la microbiota asociada con las postlarvas.
Los genomas de las cepas de V. campbellii aisladas en este estudio son diferentes de las cepas ecuatorianas y chinas previamente reportadas. Además, estas cepas albergan el transposón Tn3 en el plásmido pVA1 , al igual que la cepa V. campbellii LA16-V1 (Ecuador) y V. parahaemolyticus AHPND+ de América; mientras que está ausente en 20130629003S1 (China). Esto sugiere que los genes pirAB se adquirieron mediante transferencia horizontal de plásmidos entre V. parahaemolyticus y V. campbellii coexistentes dentro de una región geográfica. Finalmente, se debe tener precaución al utilizar métodos de PCR para detectar genes pirAB para el diagnóstico de AHPND en postlarvas de
camarón; recomendamos fuertemente que haya un período de incubación previo del tracto digestivo o de todo el tejido larvario para evitar resultados negativos•
Data complementaria
La data complementaria de este artículo se puede encontrar en línea en https://doi. org/10.1016/j.aquaculture.2023.740221
Para mayor información escriba a: ssoto@ciad.mx
- ABRIL 2024 34
SALUD
Fig. 5. Sintenía del plásmido pVA1 entre los genomas analizados.


Investigaciones muestran
que las pruebas rápidas de campo (POC) solo detectan WSSV cuando hay alto nivel de infección, pero hay una mejor solución para los productores de camarón

Autor:
Dr. Melony Sellars
info@genics.com.au
En 2019, la Organización de Investigación Científica e Industrial del Commonwealth (CSIRO) de Australia llevó a cabo un estudio sobre la eficacia de los kits de pruebas de campo de camarones para detectar el virus del Síndrome de la Mancha Blanca (WSSV) para su posible uso en el manejo de enfermedades. En el momento del análisis, las pruebas de campo se habían convertido en un método común de detección de enfermedades para los productores, una situación que persiste hasta el día de hoy. Pero, con la reciente presentación de los hallazgos de CSIRO en la Conferencia Nacional de Productores de Camarón de Australia, el “Simposio ProAqua 2023”, es hora de una reevaluación urgente de las prácticas de las pruebas de campo a nivel mundial.
El virus de la Mancha Blanca exige las mejores prácticas de controles
El WSSV se mantiene como uno de los patógenos de mayor impacto económico para la industria mundial del camarón. En 2016, graves brotes en Queensland, Australia, pusieron al WSSV en el primer plano de la atención de la industria camaronera nacional. Dado que esta emergencia destruyó las poblaciones y generó pérdidas de millones de dólares, los productores necesitaban urgentemente una manera de monitorear a sus animales para detectar la infección por WSSV en una etapa temprana.
La adopción mundial de kits de pruebas de campo POC, incluso para el WSSV, se había convertido en algo común en la industria del camarón, y se utilizaban para diversos fines, incluida la detección temprana del virus. Con la devastación del 2016, la industria australiana solicitó acceso a dichos kits, que primero requirieron la aprobación del gobierno.
En 2019, a pedido de una industria de camarón en riesgo, CSIRO inició una evaluación de laboratorio de las pruebas de WSSV para determinar la eficacia y la idoneidad de los kits de prueba de campo POC en comparación con las pruebas de PCR de laboratorio.
- ABRIL 2024
CSIRO examina las pruebas rápidas de campo POC para el WSSV
A partir de 2019, CSIRO colaboró con las partes interesadas de la industria del camarón en un estudio integral de los métodos de prueba del WSSV. El informe del estudio denominado Evaluación de pruebas en campo (POC) para el virus del Síndrome de la Mancha Blanca (WSSV) compara las pruebas disponibles comercialmente diseñadas para detectar WSSV en granjas comerciales de camarón con pruebas de PCR WSSV realizadas en laboratorio.
La investigación de CSIRO compara y evalúa cinco pruebas de WSSV, que comprenden tres pruebas rápidas de estilo antígeno y dos pruebas basadas en PCR. Después de pruebas exhaustivas, el estudio muestra claramente que los kits de prueba rápida de antígenos POC producen límites de detección más bajos en comparación con las pruebas de laboratorio de PCR. El grupo de investigación descubrió que las pruebas POC rápidas de estilo antígeno eran una defensa subóptima contra el WSSV porque “las pruebas POC tenían una sensibilidad menor que las pruebas qPCR de WSSV de referencia de laboratorio validadas”.
La falta de sensibilidad de las pruebas rápidas de antígenos POC es tal, que en realidad crea una falsa sensación de seguridad entre los productores. Los hallazgos del estudio de CSIRO son de vital importancia para la industria global porque la expectativa actual entre los productores es que las pruebas de campo POC puedan detectar el WSSV en las primeras etapas de la infección, proporcionando una alerta temprana de la enfermedad. Sin embargo, este estudio identifica problemas vitales con los kits de prueba de campo de antígenos POC, lo que significa que, de hecho, no son una solución para la alerta temprana y la prevención del WSSV.
El peligroso
defecto revelado por el estudio de CSIRO
Se revelaron problemas sustanciales con los kits de prueba de campo POC como medio para prevenir brotes de WSSV. Entre los inconvenientes con los kits POC, CSIRO enfatiza la brecha entre las expectativas de los usuarios y la verdadera sensibilidad de
los kits de prueba en el campo.
La confianza de los productores en los kits de prueba de campo POC está respaldada por la percepción de que pueden detectar el WSSV con la suficiente antelación para evitar pérdidas de población a gran escala. Sin embargo, la investigación de CSIRO revela que los estudios utilizados para proporcionar la capacidad del kit POC se realizaron en condiciones de laboratorio, donde la velocidad de la infección y las vías de transmisión no reflejaban la realidad en las granjas de camarón. Una lectura cuidadosa de las directrices de los fabricantes sobre algunos kits también revela que la validación se produce en plantillas sintéticas, lo que proporciona indicaciones poco realistas para el uso en el mundo real con muestras de camarón.
El equipo de CSIRO escribe en el informe que “en la literatura existente, se informó que todas las pruebas POC podían detectar el WSSV en las primeras etapas de la infección” Sin embargo, continúa: “los experimentos con camarones vivos realizados en este estudio demuestran que la enfermedad de la Mancha Blanca progresa mucho más rápido cuando a los camarones se les inyecta WSSV en comparación con cuando cohabitan con cohortes infectadas con WSSV. Por lo tanto, el período de tiempo previsto desde la exposición al WSSV hasta su detección
mediante pruebas POC en el campo puede estar sobreestimado...”.
Este hallazgo resalta el problema crucial de las pruebas POC como medio de control del WSSV. El virus se desarrolla más rápidamente en camarones que se infectan deliberadamente para la validación de la prueba de campo POC. Sin embargo, cuando se utilizan en entornos comerciales, donde la enfermedad se desarrolla más lentamente en animales individuales, los kits de prueba de campo POC no logran encontrar el virus lo suficientemente temprano como para evitar que la infección se propague a través de las poblaciones en la camaronera. Esta falla fatal en la sensibilidad de los kits POC significa que, si los productores continúan usándolos para la detección del WSSV, no detectarán el virus en sus primeras etapas, lo que provocará brotes continuos de enfermedades.
Protegerse contra el WSSV requiere un cambio estratégico
El virus de la Mancha Blanca es ahora endémico en todo el mundo, por lo que es esencial un seguimiento preventivo con tecnología adecuada. El WSSV puede destruir poblaciones enteras de granjas camaroneras en cuestión de días, lo que sugiere la necesidad de un régimen eficaz de detección temprana.

- ABRIL 2024 37 SALUD
Uno de los hallazgos clave del estudio reciente de CSIRO es que cuando se trata de pruebas confiables para WSSV, los kits POC actuales no pueden igualar la sensibilidad y precisión de las pruebas PCR validadas en laboratorio.
“Para las tres cepas de WSSV evaluadas, las qPCR de referencia del laboratorio fueron 10 veces más sensibles que la prueba rápida de campo POC más sensible”, encontró el estudio de CSIRO. De hecho, las pruebas de PCR son 100 veces más sensibles que uno de los kits de pruebas POC más utilizados en el mercado. CSIRO afirma en sus hallazgos que para “las muestras positivas débiles”, típicas de la infección por WSSV en etapa temprana, la sensibilidad de las pruebas POC es inadecuada, “solo entre el 56% y el 78% de las muestras positivas débiles replicadas dan positivo, en contraste con el 100% de concordancia demostrada por las qPCR de referencia del laboratorio”
Para los productores, la conclusión que se puede extraer de la investigación de CSIRO es enfáticamente clara: para evitar brotes destructivos de Mancha Blanca (WSSV), la industria del camarón necesita urgentemente escoger el uso de pruebas PCR de laboratorio para análisis de rutina en lugar de depender de kits de pruebas rápidas POC. Así mismo, estas pruebas PCR en laboratorio también deben ser validadas adecuadamente por el personal del mismo, ya que el rendimiento de todos los ensayos varía según el equipo, la química, las personas y las condiciones operativas.
Dado que los tiempos de respuesta de las pruebas de laboratorio son tan cortos como
Comparación de la sensibilidad analítica comparativa de las pruebas de punto de atención (POC) para WSSV y qPCRs de referencia de laboratorio para WSSV/China/201
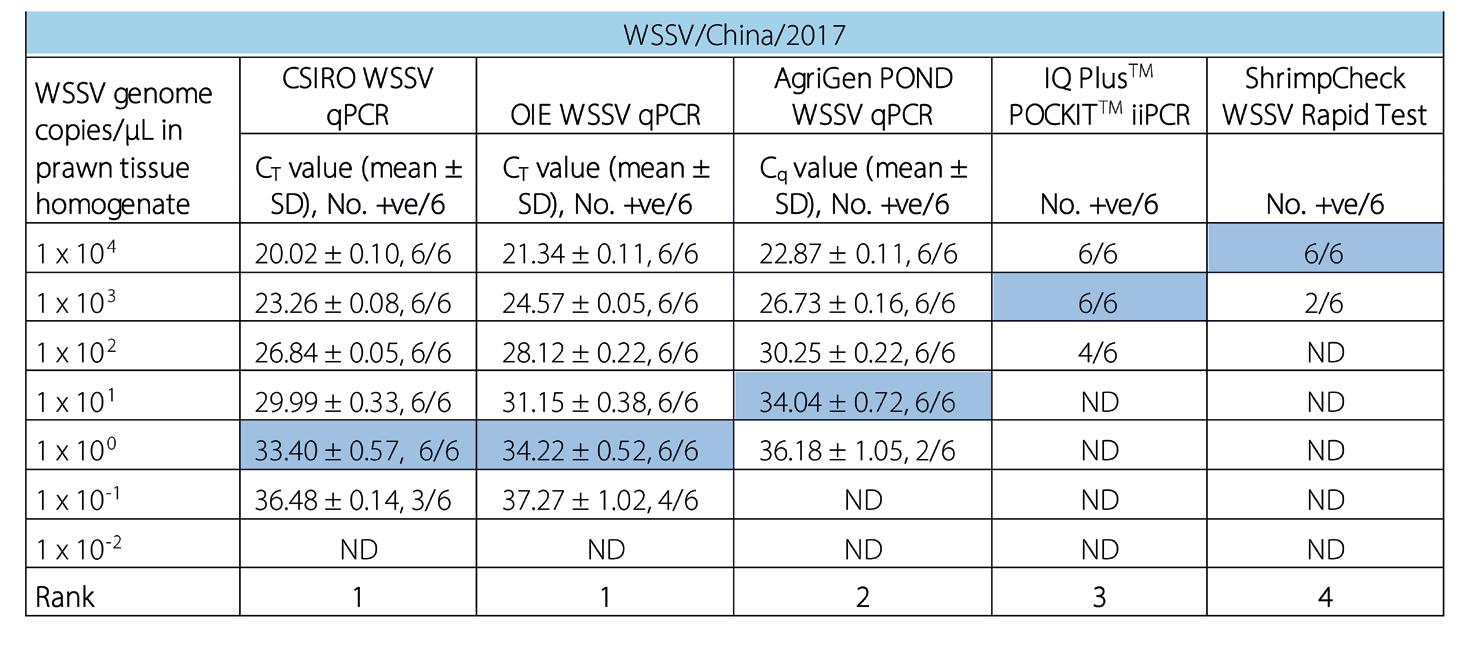
ND: no se detectaron positivos. El resaltado en azul indica el LDM. LDM definido como la dilución final donde todas las 6 repeticiones de la prueba son positivas
24 horas, cualquier preocupación sobre la inmediatez puede dejarse de lado. Realizar una prueba POC brinda la falsa seguridad de obtener resultados rápidos, pero la realidad es que, debido a su falta de sensibilidad, las pruebas POC pasan por alto la enfermedad en sus primeras etapas cuando una prueba de laboratorio sí la detectaría.
Conclusiones para los acuicultores
El informe de CSIRO muestra claramente que las pruebas PCR de laboratorio son más sensibles y confiables que los omnipresentes kits POC. También cabe señalar que las pruebas de laboratorio deben validarse adecuadamente en cualquier instalación determinada para garantizar un rendimiento óptimo y que la validación es un requisito continuo, no un evento aislado.
El aspecto de alerta temprana de las pruebas PCR en laboratorio ofrece una ventaja vital a los productores camaroneros, ya que conocer la infección por WSSV en sus piscinas unos días antes puede evitar pérdidas catastróficas de población.
Cuando los brotes de enfermedades pueden diezmar las granjas camaroneras en cuestión de días y hay millones de dólares en juego, la investigación de CSIRO envía una fuerte señal a la industria camaronera para que deje de depender de los kits POC. Las pruebas PCR en laboratorio, con su sensibilidad y confiabilidad superiores, son la única forma de manejar eficazmente la amenaza de la Mancha Blanca•
Para mayor información escriba a: info@genics.com.au

- ABRIL 2024 38 SALUD

Maximizando la productividad: el papel de los sensores en la acuicultura
Autores:
Manuel Espinoza-Ortega1
Carla Torres
Carlos Mora-Pinargote1
Arturo Moreno
Rafael Samaniego-Camacho
César Molina-Poveda*1.
Skretting LATAM
1Skretting Aquaculture Innovation *cesar.molina@skretting.com

La industria acuícola de camarón se enfrenta constantemente al desafío de mantener un óptimo ambiente de cultivo para sostener el crecimiento y el éxito de la producción. En este contexto, es importante mantener la calidad de agua dentro de ciertos rangos para alcanzar un adecuado crecimiento y salud de los organismos (Rahman et al., 2019). La calidad del agua de las piscinas es monitoreada varias veces al día mediante sensores manuales, análisis de muestras en laboratorio o con sensores montados permanentemente en la piscina (Rahman et al., 2023). Los parámetros de calidad de agua comúnmente medidos son temperatura, oxígeno disuelto (OD), salinidad, pH, turbidez, alcalinidad, amonio total, nitrito y nitrato (Rana et al., 2021). Todos estos parámetros proporcionan información crítica de la condición del ambiente de la piscina camaronera. Por este motivo, la monitorización precisa y exacta de los parámetros es significativa para la detección de desviaciones que puedan comprometer el crecimiento y la salud del camarón, hasta la rentabilidad del cultivo. En los últimos años, el desarrollo tecnológico ha brindado a los acuicultores una variedad de herramientas para realizar este seguimiento, entre las que se destacan el uso de los sensores galvánicos u ópticos conectados a dispositivos remotos que otorgan una vigilancia continua (Abdullah et al., 2021); por ejemplo, de los niveles de OD y la temperatura del agua brindando una visión detallada y precisa del entorno acuático, así permitiendo a los productores tomar decisiones informadas y proactivas en cuanto a recambios de agua o alimentación precisa.
En este artículo, compararemos de manera detallada las características, ventajas y limitaciones de los sensores galvánicos y ópticos en el contexto de su aplicación en la acuicultura. Además, exploraremos cómo los resultados emitidos por los sensores permiten mejoras en la productividad del cultivo de camarón.
El oxígeno disuelto y la temperatura
El OD es uno de los parámetros ambientales más críticos en la acuicultura, porque incide directamente en la producción, ya que dependiendo de su concentración en el agua puede afectar el rendimiento del
- ABRIL 2024
40
camarón, factor de conversión alimenticia (FCA) y la capacidad de carga de la piscina. (Mustafa et al., 2022). En la industria camaronera, a través del uso de sensores, el monitoreo del OD es posible cada 5 - 10 min, ayudando a distribuir y controlar la alimentación en horas o eventos críticos, protegiendo de esta manera el FCA. El papel del oxígeno disuelto en el agua es crucial en el metabolismo de los camarones (Rosas et al., 1998), ya que participa directamente en la producción de energía a través de la respiración, mediante la fosforilación oxidativa. En este proceso, el oxígeno actúa como el último receptor de electrones en la cadena respiratoria, permitiendo a los camarones y otros organismos aeróbicos aprovechar al máximo la energía de las moléculas de carbono metabolizadas por el ciclo de Krebs. La presencia o ausencia de oxígeno como receptor final de electrones marca una gran diferencia en la cantidad de energía disponible para realizar el trabajo. Incluso una ligera disminución en los niveles de oxígeno disuelto, por ejemplo, de 5 a 4 mg/L, puede provocar una reducción de hasta un 25% en la producción de biomasa (Rosas et al., 1998). Esta limitación es significativa, especialmente en ambientes como estanques de cultivo de camarón, lagunas costeras y estuarios tropicales, donde es común encontrar bajos niveles de oxígeno, especialmente después de lluvias intensas de verano que aumentan la carga de materia orgánica en el agua.
Por otro lado, la temperatura es un parámetro influenciado por la estación del año, la latitud, altitud, hora del día, circulación del aire, cobertura de las nubes y el flujo y profundidad del cuerpo de agua (Yamanaka, 2016). Este parámetro ambiental también es clave en el cultivo de camarón, debido a su efecto sobre actividades biológicas como la alimenticia, que puede aumentar o disminuir dependiendo si la temperatura es cálida o fría (Spanopoulos-Hernández et al., 2005). Por esta razón, registrar y supervisar este parámetro es vital para controlar nutrición, crecimiento y conversión alimenticia. La temperatura juega un papel decisivo en el crecimiento de los camarones, ya que por lo general, a temperaturas más altas se observa un mayor crecimiento. La tolerancia a la temperatura, los rangos óptimos y cómo esta influye en el crecimiento dependen
de factores como la especie, la edad, la salinidad y la concentración de OD, entre otros (Bett y Vinatea, 2009; SpanopoulosHernández et al., 2005).
Tipos de sensores: ópticos y galvánicos
Los sensores para medir OD en agua pueden dividirse principalmente en dos categorías: galvánicos y ópticos. Cada tipo tiene sus propias ventajas y desventajas, y la elección entre ellos depende de factores como la aplicación específica, la precisión requerida y el entorno en el que se usarán. Los sensores galvánicos, basados en principios electroquímicos, han sido ampliamente utilizados en aplicaciones acuícolas en la medición de parámetros como la concentración de oxígeno disuelto y la conductividad eléctrica del agua. Por otro lado, los sensores ópticos, que emplean tecnologías como la espectroscopia o la
fluorescencia, han surgido como alternativas prometedoras, ofreciendo ventajas en términos de sensibilidad y capacidad para medir una gama más amplia de parámetros, incluyendo la turbidez y la concentración de nutrientes.
¿Cómo funciona un sensor óptico?
En un sensor óptico, la luz azul excita la película fluorescente (Figura 2) y esta película emite luz roja que es detectada por el fotodetector. Cuanto menor sea el oxígeno, mayor será la intensidad de la luz roja recibida y viceversa.
El fotodetector convierte la luz roja recibida en una señal eléctrica proporcional que se procesa para mostrar el valor del OD. Por otro lado, el capuchón del sensor está cubierto por una capa protectora.
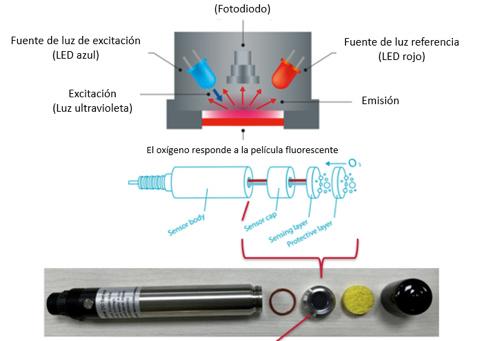
- ABRIL 2024 41 PRODUCCIÓN
Figura 1. Breve ilustración sobre el efecto de las lluvias y el sol sobre la temperatura del agua
Figura 2. Partes y funcionamiento de un sensor óptico (nivel de incertidumbre ±5 %)
¿Cómo funciona un sensor galvánico?
Cuando el sensor galvánico se sumerge en agua, el oxígeno del agua que pasa a través de la membrana permeable al gas se consume en el cátodo. Esto provoca un flujo de electricidad proporcional a la concentración de oxígeno entre el cátodo y el ánodo (Figura 3). Esta señal eléctrica se calibra como nivel de OD. La reacción química provoca la precipitación de un sólido blanco en el electrolito. A partir de cierta cantidad, este sólido blanco interfiere en el funcionamiento de la celda y debe ser sustituido. La membrana permeable al gas también debe reemplazarse con cierta frecuencia para garantizar un funcionamiento correcto.
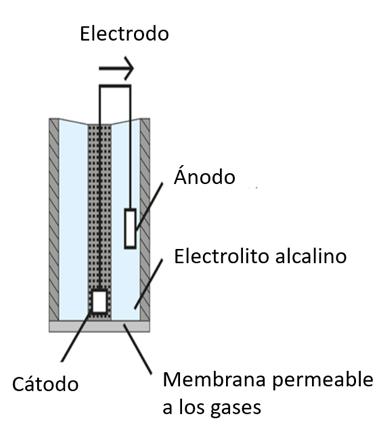
Ventajas y desventajas de los sensores ópticos y galvánicos
Los sensores galvánicos pueden detectar concentraciones de oxígeno muy bajas con alta sensibilidad; en general, el límite de detección suele ser tan bajo como 0,1 mg/L de OD o incluso menor en algunos casos. Estos valores pueden variar dependiendo del diseño o la calidad del sensor (Wei et al., 2019).
En general, los sensores galvánicos requieren calibración y mantenimiento regular debido a la degradación que sufren el electrodo y la membrana con el pasar del tiempo. En cuanto a la velocidad de respuesta pueden ser más lentos en comparación con los sensores ópticos. Pueden ser
susceptibles a interferencias de gases y sustancias químicas presentes en el agua. Por otro lado, los sensores ópticos utilizan la fluorescencia o la absorción óptica para medir la concentración de oxígeno disuelto. Por ejemplo, en un sensor de fluorescencia, un fluoróforo sensible al oxígeno emite luz fluorescente que varía en intensidad en función de la concentración de oxígeno.
Los sensores ópticos tienen tiempos de respuesta más rápidos en comparación con los sensores galvánicos y no requieren partes móviles ni membranas permeables al oxígeno, lo que los hace más duraderos y menos propensos a la degradación; además, por lo general necesitan calibración con menos frecuencia que los sensores galvánicos. Los sensores ópticos tienen una alta precisión en rangos de OD entre 0 y 8 mg/l y un comportamiento más estable si se comparan con los sensores galvánicos, ya que la presencia de sulfuro de hidrógeno y dióxido de carbono no daña este tipo de sensor. Un sensor óptico no necesita un flujo de agua para ser preciso, es fácil de limpiar y mantener, pues no requiere de cambio del electrolito, a la vez que tiene una mayor vida útil. Los sensores ópticos, por tanto, se usan cuando se necesita una respuesta rápida, alta durabilidad y menor mantenimiento.
Tanto los sensores galvánicos como los ópticos son susceptibles a interferencias ambientales, como la turbidez o la presencia de materia orgánica en el agua, por lo que es necesaria la limpieza constante. Por ejemplo, los sensores galvánicos, que se basan en la detección electroquímica, pueden ser sensibles a las variaciones de temperatura, lo que puede afectar su precisión y requerir que el equipo lleve una compensación de temperatura para mediciones más precisas. Por otro lado, los sensores ópticos, que utilizan principios como la extinción de fluorescencia, suelen ser menos sensibles a las variaciones de temperatura y, por lo tanto, pueden proporcionar mediciones más estables en diferentes condiciones de temperatura (Wei et al., 2019). En referencia a los costos, por lo general los primeros son más económicos en comparación con los sensores ópticos.
Variación del nivel de oxígeno en varias locaciones de una piscina
La variación del OD en diferentes zonas de una piscina camaronera puede deberse a varios factores, incluyendo la actividad biológica, la circulación del agua, el recambio y la temperatura.
El recambio es una de las principales variables que impactan en la concentración de oxígeno en una piscina camaronera. El agua que ingresa contiene niveles más altos de OD. El recambio de agua asegura un suministro adecuado de oxígeno para los camarones, lo que es vital para su respiración y metabolismo. En la superficie de la piscina, el oxígeno puede entrar en contacto directo con el aire, lo que facilita su absorción. Aquí, el oxígeno puede estar en niveles más altos debido a la exposición directa al aire y a la agitación causada por la circulación del agua.
Así también, los sistemas de aireación introducen oxígeno en el agua, lo que aumenta los niveles en esas áreas.
En contraposición, en las zonas más profundas de la piscina o en áreas alejadas de la circulación del agua, los niveles de
Figura 4. Variación del nivel de oxígeno en piscinas. Los niveles de oxígeno bajos (rojo) se concentran en las zonas de alimentación solamente las entradas (azul) mantienen un nivel aceptable de oxígeno por sobre 4 ppm.
42 - ABRIL 2024
PRODUCCIÓN
Figura 3. Componentes de un sensor galvánico (nivel de incertidumbre ± 2 – ± 5 %)
oxígeno pueden ser más bajos. La falta de movimiento del agua y la menor exposición al aire pueden reducir la cantidad de OD en el agua. Adicionalmente, en zonas con alta densidad de camarones, como las zonas de alimentación, la actividad biológica puede consumir oxígeno a tasas más altas (Figura 4). Esto puede resultar en niveles más bajos de OD en esas áreas, especialmente si el recambio y la circulación del agua es limitada.
En cuanto a la temperatura, también puede influir en los niveles de OD. El agua más fría puede contener más oxígeno que el agua más cálida. Por lo tanto, las zonas de la piscina con temperaturas más bajas pueden tener niveles de oxígeno ligeramente más altos.
El nivel de oxígeno mínimo para tener una adecuada digestión es 4 mg/l, mientras que el mejor rango de temperatura esta entre 28 y 31 oC; este parámetro tiene un impacto directo en el aprovechamiento del alimento.
El mantenimiento de los sensores: un factor esencial para mediciones precisas
Un monitoreo preciso del oxígeno disuelto y de la temperatura requiere un sensor en óptimas condiciones. La limpieza de un sensor en acuicultura es un proceso importante para garantizar mediciones precisas y confiables.
En una piscina, a medida que los días transcurren sin limpieza del sensor, más allá de 2 o 3 días, las lecturas decrecen si se compara con un sensor externo de referencia y la correlación entre lecturas de los dos sensores desciende drásticamente.
En el ejemplo de la Figura 5, desde el día 8 de abril hasta el 10 de abril, la correlación entre lecturas del sensor de referencia y el sensor óptico va disminuyendo desde un 92% hasta un 54% cuando el sensor permanece sin limpiar. Sin embargo, el día 10 de abril, en cuanto se realizó la limpieza, los parámetros de los sensores volvieron a la normalidad con una correlación de 99%.
Bajo las condiciones de producción de camarón, la frecuencia de limpieza recomendada es dos veces por semana. Sin embargo, esto depende de la turbidez del
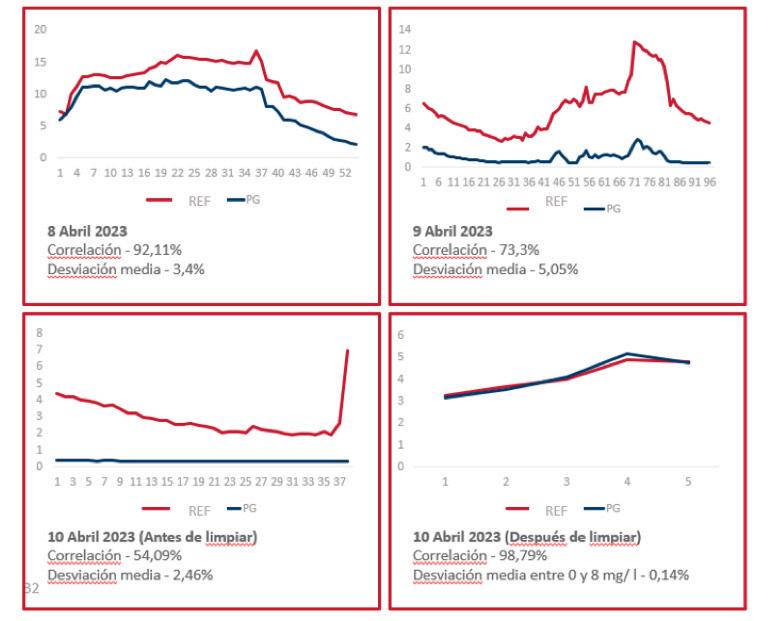
5. Degradación de la señal por falta de limpieza de un sensor óptico (línea azul), frente a un sensor de referencia externo (línea roja) a medida que transcurre el tiempo. REF= sensor de referencia; PG= sensor óptico
estanque y de los aditivos añadidos al agua. Normalmente los sensores se limpian bajo chorro de agua, y si esto no es suficiente, el sensor debe limpiarse suavemente con un paño previamente humedecido en agua, que no desprenda partículas (Figura 6). En el caso de sensores galvánicos, la membrana debe limpiarse muy suavemente, evitando el uso de cualquier material abrasivo. No se deben usar disolventes orgánicos para limpiar ninguna de las superficies del sensor (Figura 7).
En las mismas condiciones ambientales, las lecturas de un sensor óptico no tienen “deriva”, lo que quiere decir que no varían a medida que transcurre el tiempo. Básicamente, una lectura de 6,1 mg/l no se convertirá en 6,2 mg/l o 6,3 mg/l. Mientras que en el sensor galvánico, según la condición del electrolito, estos cambios son bastante frecuentes.
La ubicación de los sensores y su montaje
Un montaje adecuado garantiza que el sensor esté ubicado en una posición óptima para captar las condiciones reales del agua en la piscina. Si el sensor no está correctamente montado, las mediciones pueden ser inexactas, lo que podría conducir a decisiones erróneas en el manejo de la piscina (Figura 8).
Utilidad de los sensores remotos en camaroneras
Control de la alimentación usando sensores. El caso de las altas temperaturas y el FCA.
Para el presente análisis se seleccionaron piscinas con una extensión en un rango amplio de 11 a 34 hectáreas, ubicadas en la provincia del Guayas. Estas piscinas corresponden al período invernal 2023, y fueron alimentadas con una dieta del 35% de proteína, tanto en formato extruido como pelletizado.
Las piscinas fueron seleccionadas con base en los valores históricos de temperatura. La alimentación en la etapa de prueba fue realizada con dispositivos sónicos, los cuales procesan el sonido generado por el camarón cuando se encuentra en actividad alimenticia.
La prueba consistió en identificar los períodos del día en que la temperatura del agua excedió los 32 oC, usando un sensor óptico en tiempo real para de esta manera, realizar pausas en la alimentación durante dicho período. Una vez iniciada la prueba, diariamente se controlaba la alimentación en los rangos de temperatura por sobre los 32 oC. Los resultados mostraron una reducción en el FCA que varió entre 1,41 y 1,57 (Tabla
- ABRIL 2024 43 PRODUCCIÓN
Figura
2) en piscinas que históricamente mantenían un valor de 1,72.
En la Tabla 3.se muestra un extracto de los datos de temperatura registrados diariamente resaltándose en rojo cuando alcanzo o excedió los 32 oC. Las pausas en la alimentación se aplicaban cuando alcanzo y sobrepaso dicha la temperatura; en estos períodos es preferible no dar de comer al animal debido a que, al acelerarse el metabolismo, el tiempo que permanece en el tracto intestinal es tan corto que no permite una adecuada digestión y posterior absorción de los nutrientes liberados del alimento.
La velocidad de las reacciones metabólicas en los organismos vivos está determinada por la temperatura, la cual se manifiesta en procesos como el crecimiento y la muda (Bett y Vinatea, 2009; Spanopoulos-Hernández et al., 2005). Todos los organismos tienen un rango de temperatura en el cual realizan estas funciones de manera óptima, pero fuera de este rango, las funciones se ven afectadas y pueden detenerse o acelerarse en temperaturas extremas.
En este sentido, Limsuwan et al. (2009) reportó los efectos de la temperatura sobre el comportamiento alimentario de juveniles de camarón blanco del Pacífico (Penaeus vannamei) a diferentes temperaturas del agua: 24±1, 26±1, 28±1, 30±1, 32±1 y 34±1°C, en un experimento realizado a 25 ppt de salinidad. Los resultados revelaron que los camarones a 24±1, 26±1 y 28±1°C, comían lentamente en el fondo de los acuarios. Específicamente a 24±1°C, se inició la excreción a los 90-105 minutos, mientras que a 34±1°C este proceso se inició a los 20-35 minutos.
Además, después de alimentar durante 2 h, el alimento no consumido fue aproximadamente del 35 al 70 % por día a 24±1°C, mientras que no hubo sobrante a 30-34 oC luego de alimentar 2 h.
Los tiempos de vaciado del tracto intestinal fueron descendiendo a medida que la temperatura aumentaba, lo que indica un incremento de la actividad metabólica a medida que la temperatura ascendía, desde 4 horas a 24 oC hasta 2,3 horas a 34 oC.
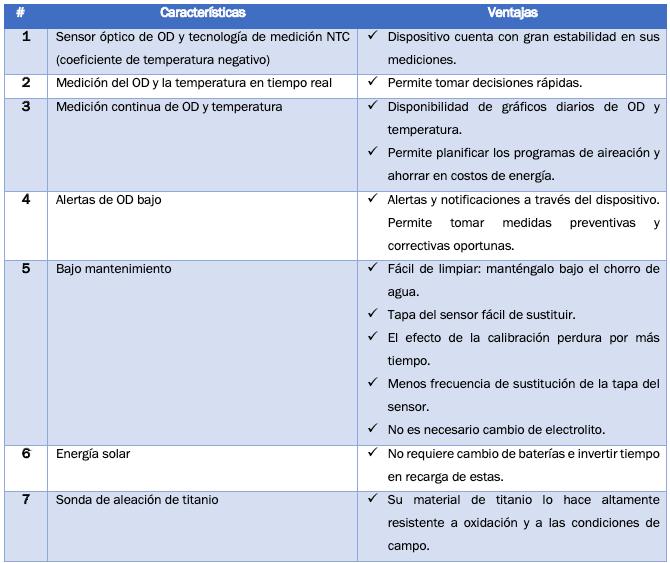


Figura 7. Sensor óptico dañado por una inadecuada limpieza
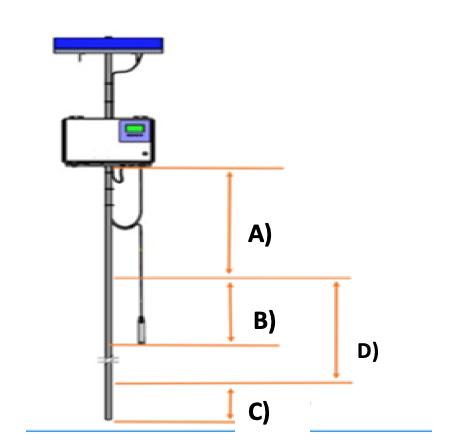
Figura 8. Distancias desde: A: caja del sensor desde el agua: 50 cm. B: profundidad del sensor: a la mitad de la columna de agua. C: profundidad del poste del suelo: 1 metro de profundidad. D: nivel del agua (la profundidad del agua en el estanque)
44 - ABRIL 2024 PRODUCCIÓN
Tabla 1. Características y ventajas de un sensor óptico comercial
Figura 6. Un sensor óptico antes y después de la limpieza
Tabla 2. Piscinas con alimentación automática asistida por sensores, las cuales tuvieron pausas en alimentación alineadas a las lecturas de temperaturas para precautelar el aumento del FCA.
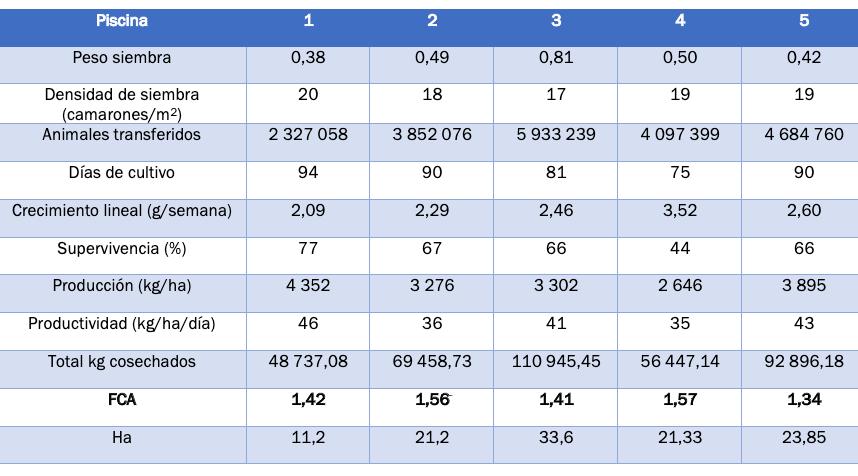
Conclusión
El uso de sensores remotos permite realizar monitoreos 24/7, lo que asegura identificar en tiempo real cómo afecta el nivel de oxígeno disuelto y la temperatura en la demanda de alimento, permitiendo optimizar la dosificación y racionalización del balanceado en la piscina. Un seguimiento constante y preciso de estos parámetros, específicamente de la temperatura, posibilita actuar en los momentos en los que sabemos que el camarón no va a lograr una completa digestión del alimento al mayor ritmo de ingestión y evacuación gástrica al que está sometido, lo cual impacta significativamente en el factor de conversión alimenticia, una variable que incide en el costo de producción del camarón•
Para mayor información escriba a: cesar.molina@skretting.com
Tabla 3. Ejemplo de un monitoreo continuo de temperatura según la hora (los datos en celdas rojas indican un incremento. En horario de 14h00 a 16h00 la alimentación fue pausada.
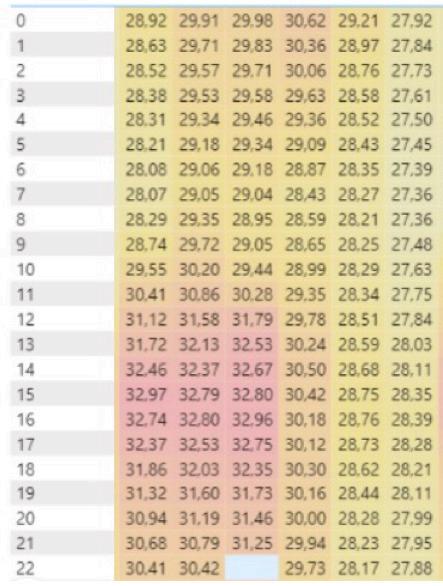
Bibliografía:
Abdullah, M., Idrus, S.M., Yusof, K.M., Azmi, A.I., Ismail, W., Kamaludin, K.H., Ali, N., Rani, A., Yusof, F., 2021. Field Trial and Performance Evaluation of IoT Smart Aquaculture Monitoring System for Brackish Water Shrimp Farm, International Journal of Nanoelectronics and Materials. Bett, C., Vinatea, L., 2009. Combined effect of body weight, temperature and salinity on shrimp Litopenaeus vannamei oxygen consumption rate. Braz J Oceanogr 57, 305–314.
Limsuwan, C., Chuchird, N., Wongmaneeprateep, S., Prasertsri, S., Laisutisan, K., Wiriyapattanasub, P., & Limhang, K. 2009.. Effects of temperature on feeding behavior of Pacific white shrimp (Litopenaeus vannamei). In Proceedings of the 47th Kasetsart University Annual Conference, Kasetsart, 17-20 March, 2009. Subject: Fisheries (pp. 337345). Kasetsart University.
Mustafa, A., Paena, M., Athirah, A., Ratnawati, E., Asaf, R., Suwoyo, H.S., Sahabuddin, S., Hendrajat, E.A., Kamaruddin, K., Septiningsih, E., Sahrijanna, A., Marzuki, I., Nisaa, K., 2022. Temporal and Spatial Analysis of Coastal Water Quality to Support Application of Whiteleg Shrimp Litopenaeus vannamei Intensive Pond Technology. Sustainability 14, 2659. https://doi.org/10.3390/su14052659
Ponce-Palafox, J., Parra-Flores, A., SpanopoulosHernández, M., Martinez- Cardenas, L., 2019. Feeding behavior and ingestion rate of juvenile shrimp of the genus Penaeus (Crustacea: Decapoda). Open Access Journal of Science 3. https://doi.org/10.15406/oajs.2019.03.00140
Rahman, A., Arnold, S., Dabrowski, J.J., 2019. Identification of variables affecting production outcome in prawn ponds: A machine learning approach. Comput Electron Agric 156, 618–626. https://doi.org/10.1016/j.compag.2018.12.015
Rahman, A., Csiro, S.A., Emerenciano, M., Article, R., 2023. Soft sensing of water quality parameters in indoor shrimp farming using machine learning models. https://doi.org/10.21203/ rs.3.rs-3233512/v1
Rana, M., Rahman, A., Dabrowski, J., Arnold, S., McCulloch, J., Pais, B., 2021. Machine learning approach to investigate the influence of water quality on aquatic livestock in freshwater ponds. Biosyst Eng 208, 164–175. https://doi.org/10.1016/j. biosystemseng.2021.05.017
Rosas, C., Martinez, E., Gaxiola, G., Brito, R., DíasIgresia, E., Solo, L.A., 1998. Effect of dissolved oxygen on the energy balance and survival of Penaeus setiferus juveniles. Mar Ecol Prog Ser 174, 67–75.
Spanopoulos-Hernández, M., Martínez-Palacios, C.A., Vanegas-Pérez, R.C., Rosas, C., Ross, L.G., 2005. The combined effects of salinity and temperature on the oxygen consumption of juvenile shrimps Litopenaeus stylirostris (Stimpson, 1874). Aquaculture 244, 341–348. https://doi. org/10.1016/j.aquaculture.2004.11.023
Wei, Y., Jiao, Y., An, D., Li, D., Li, W., Wei, Q., 2019. Review of Dissolved Oxygen Detection Technology: From Laboratory Analysis to Online Intelligent Detection. Sensors 19, 3995. https://doi. org/10.3390/s19183995
Yamanaka, M.D., 2016. Physical climatology of Indonesian maritime continent: An outline to comprehend observational studies. Atmos Res 178–179, 231–259. https://doi.org/10.1016/j. atmosres.2016.03.017
- ABRIL 2024 45 PRODUCCIÓN

Bienestar Animal
Experiencias y desafíos de la eliminación de la ablación en reproductores de Penaeus vannamei en una maduración de Ecuador
Autores:
Jaime García
Juan Sebastian Pérez
Alejandro Pesantes
jgarcia@aquagen.com.ec

La ablación ocular –extirpación quirúrgica o corte del pedúnculo ocular del camarón– ha sido común en los criaderos de todo el mundo durante el último cuarto de siglo. El pedúnculo ocular contiene un complejo de glándulas que influyen en los procesos de muda, maduración sexual y desove. La ablación fue un avance importante en la comercialización del cultivo de camarón en las décadas de 1970 y 1980, ya que las hembras reproductoras con pedúnculos oculares extirpados maduraron más rápido y producían de 10 a 20 veces más huevos, lo que dio a los gerentes de criaderos una producción consistente y confiable (Global Aquaculture Advocate, 2019).
Existen dos técnicas aplicadas comúnmente para la ablación del pedúnculo ocular. La primera es por cauterización, usando una pinza metálica la cual es calentada al rojo vivo y con la cual se le amputa y cauteriza la herida en el ojo. La segunda técnica de ablación consiste en anudar con un hilo el pedúnculo ocular, cortando la circulación en el mismo y provocando su desprendimiento al tercer o cuarto día (Foto 1). Ambas técnicas ocasionan daño físico al animal.
Sin embargo, es posible manejar las hembras sin necesidad de realizar la remoción del tallo o pedúnculo ocular, con la finalidad de que estas logren su ciclo natural de madurez para ser copuladas y de esta manera obtener los nauplios. La importancia de este método radica en que se evita ocasionar daño físico al animal y no se acelera de manera continua su estado de gravidez para la obtención de nauplios.
Existen trabajos previos sobre la no ablación, como la investigación conjunta realizada entre Global Aquaculture Alliance, la Universidad de Stirling, Seajoy Seafood y Lyons Seafood. En esta investigación, ejecutada por Simao Zacarias, un estudiante de doctorado de Stirling, se determinó que las hembras sin ablación producen más huevos, más nauplios y que estos tienen una mayor sobrevivencia en los criaderos, comparados con los que provienen de hembras ablacionadas (Global Seafood Advocate, 2016).
A fin de cumplir con regulaciones de certificación, se inauguró en 2017 una
PRODUCCIÓN
47
sala de maduración orgánica separada completamente de las salas de maduración convencionales. Cabe resaltar que los reproductores con certificación orgánica provienen de camaronera con certificación de producción orgánica, y es requisito que las hembras no sean ablacionadas para la producción. En esta sala se reconfirmó lo que se venía observando, respecto a que las hembras orgánicas no ablacionadas producían más huevos y nauplios que las hembras convencionales ablacionadas.
Las hembras ablacionadas tienen más copulas, pero menor cantidad de huevos y nauplios debido a que se desgastan rápidamente por su acelerada y continua gravidez. En cambio, las hembras no ablacionadas tienen su ciclo de gravidez natural, por lo que producen más huevos y más nauplios, que son más fuertes y por consiguiente, con una mayor tolerancia a las enfermedades y al medio ambiente. En la tabla se aprecia que las hembras sin ablación produjeron un promedio de 208,891 huevos por hembra y 137,571 nauplios por hembra; en comparación con las hembras ablacionadas que tuvieron un promedio de 186,904 huevos por hembra y 120,146 nauplios por hembra (Tabla 1).
El paso de hembras ablacionadas a hembras sin ablación
En el último trimestre del año 2020 la empresa dejó de realizar la ablación a las hembras, por lo que en la actualidad funciona totalmente sin ablación. Antes de tomar esta decisión existía la duda sobre cómo alcanzar la misma cantidad de nauplios para la venta a partir de hembras sin ablación, ya que con estas solo obteníamos 8% de cópulas/día, mientras que con las hembras convencionales ablacionadas obteníamos alrededor del 12%. La relación de hembras - machos para ambos casos era de 1:1. Los cambios que hicimos para obtener la misma cantidad de nauplios fueron aumentar la cantidad de hembras y reducir la cantidad de machos; se incrementó la relación a 1.25 hembras por cada macho (1.25:1).
Por otro lado, se aumentó la cantidad de tanques en producción incorporando los tanques de preproducción que eran usados para colocar las hembras ablacionadas durante un tiempo, antes de entrar a

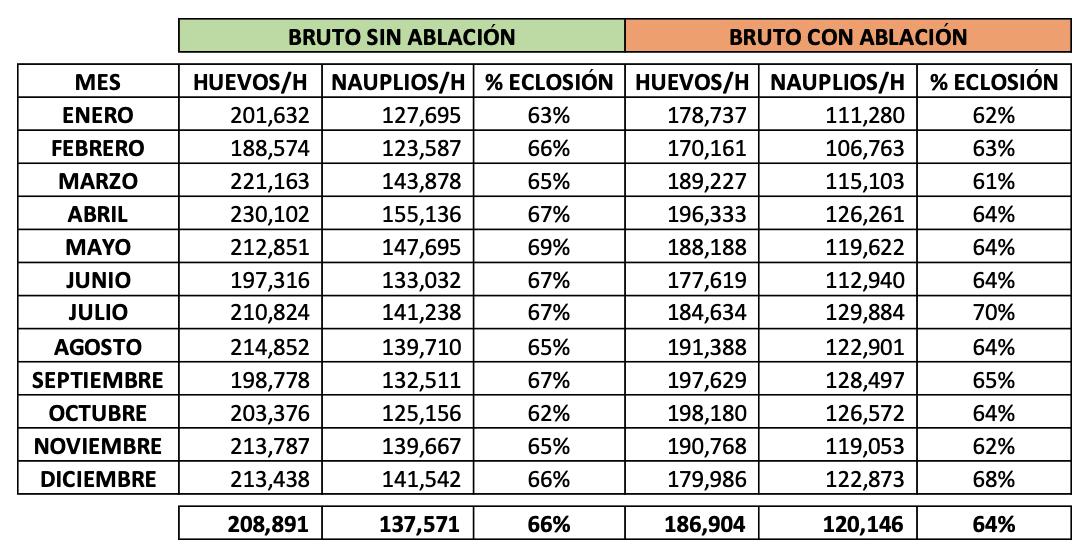
producción, por lo que ya no se necesitarían más para este propósito.
Los resultados obtenidos al dejar de ablacionar fueron los siguientes (Tabla 2):
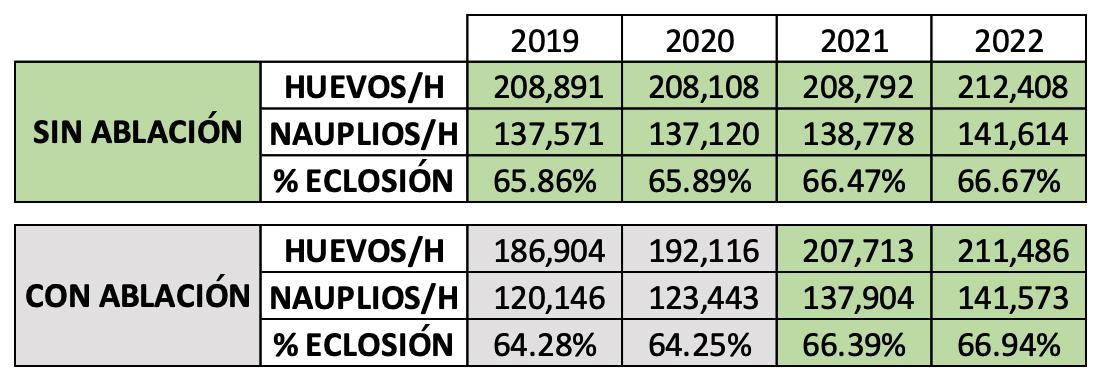
Los datos muestran que para el año 2019, las hembras no ablacionadas tenían mayor cantidad de huevos y nauplios que las hembras ablacionadas. En el año 2020 esta diferencia se acortó, ya que no se realizaron ablaciones de ojo en el último trimestre, y para los años 2021 y 2022 la diferencia prácticamente dejó de existir.
Resultados en larvicultura
En el 2019, el laboratorio sembraba alrededor de 76 millones de nauplios por módulo y el porcentaje de supervivencia promedio era del 68.47% para las larvas provenientes de hembras no ablacionadas y 66.76% para larvas proveniente de hembras con ablación.
48 - ABRIL 2024 PRODUCCIÓN
Foto 1: Técnica de ablación del pedúnculo ocular por cauterización y por amarre de hilo
Tabla 1: Comparativo Huevos y Nauplios por hembra sin y con ablación
Tabla 2: Resumen comparativo Huevos y Nauplios por hembra sin y con ablación
En la siguiente tabla se aprecian los resultados antes mencionados (Tabla 3):
Tabla 3: Resultados en larvicultura años 2019-2022
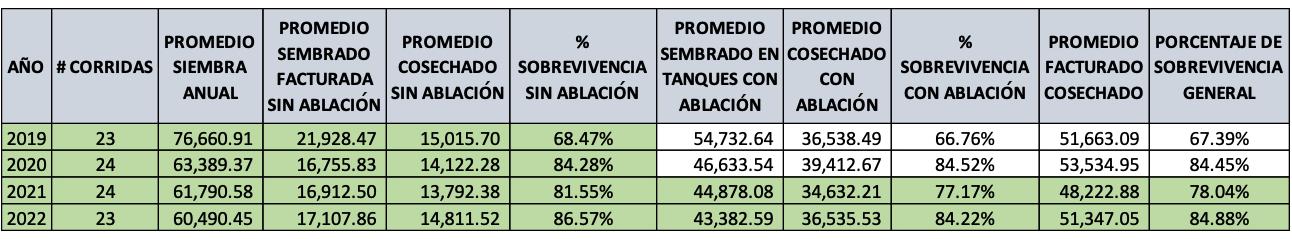
En el 2020, debido a la pandemia del COVID-19, se presentaron problemas de movilización y comercialización por las vías cerradas. En vista de estos inconvenientes, se decidió bajar la densidad de siembra en cada módulo a 60 millones. Los resultados sorprendieron, pues la sobrevivencia subió al 84%, lo que significó que se producía la misma cantidad de larvas que cuando se sembraba mayor cantidad de nauplios. Otra observación fue que mejoró el desarrollo de las larvas y se presentaron menos inconvenientes con enfermedades, ya que la cantidad de nauplios por litro es menor (entre 90 a 110 pl/l), logrando también una mejor calidad de agua.
Resultados en camaronera
Los resultados antes de eliminar totalmente la ablación se pueden observar en el análisis comparativo de piscinas sembradas con larvas provenientes de hembras sin ablación (SA) vs. piscinas sembradas con larvas provenientes de hembras convencionales ablacionadas (Tabla 4).
Resultados muy similares con ligera mejora en el crecimiento se obtuvieron en códigos procedentes de hembras con ablación. Los animales sin ablación tienen una sobrevivencia ligeramente mayor.
La gráfica siguiente es la histórica de producción en lb/ha promedio. A partir del 2020, el 100% de animales sembrados proceden de hembras sin ablación (Gráfico 1).
Bienestar Animal (Buenas Prácticas en Maduración)
-Eliminar la ablación en maduración permite que las hembras tengan su ciclo natural de gravidez y como respuesta produzcan más
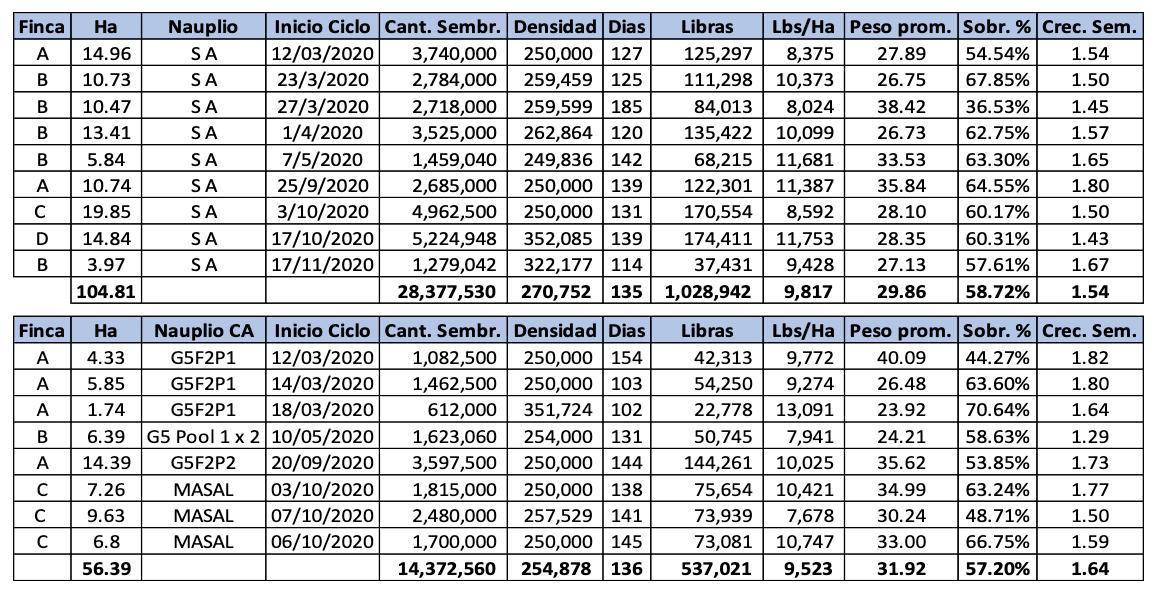
Gráfico 1: Crecimiento de producción en lb/ha con camarones de 28-30 gr. promedio
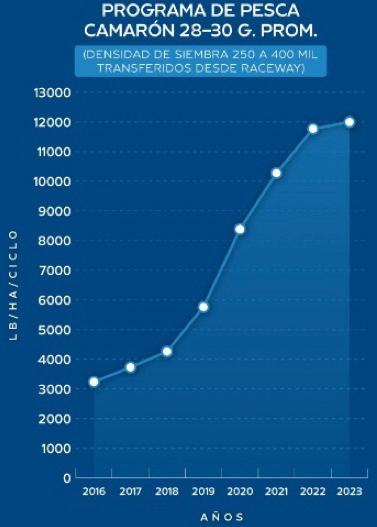
huevos y nauplios más fuertes que aquellos que provienen de hembras ablacionadas. Es una técnica amigable que no infringe ningún daño al animal para su reproducción.
Técnicas de siembra y de transferencia cuidadosas. Para transportar desde los raceways a las hembras que serán sembradas en los tanques de producción, se introduce la hembra en un tubo que tiene una malla en un extremo. El tubo debe estar en posición casi horizontal para que la hembra no choque con la malla y se lastime cuando se introduzca, ya sea que se coloque primero la cabeza o la cola. Esta técnica también se usa para transportar las hembras copuladas a los tanques de desove. Con esto evitamos que las hembras tengan laceraciones en escafoceritos o urópodos (Foto 2). Este tipo de laceraciones son producidas por manipulación inadecuada de los animales e incrementan las mortalidades por infecciones. Los tubos deben manipularse y colocarse en todo momento en un balde con agua de manera horizontal (Foto 3).
Al momento de sembrar los animales en el tanque de producción o en las tinas de desove, los tubos deben ser sacados de manera horizontal y sumergirlos en el agua del tanque de la misma manera; para después, con un movimiento suave
- ABRIL 2024 49 PRODUCCIÓN
Tabla 4: Tabla comparativa de resultados en piscinas transferidos desde raceways
de reversa, hacer que la corriente de agua pase a través de la malla expulsando suavemente al camarón. Este sale nadando tranquilamente sin mostrar ningún tipo de estrés (Foto 4). El porcentaje de dureza de la población (dureza del exoesqueleto) debe ser del 80% para transferir animales a producción. Se transportan 3 reproductores por balde, sumergidos en agua dentro de los tubos.
Mayor columna de agua. Por lo general, en las maduraciones del pasado, los tanques de producción se manejaban con columnas de agua entre 30 - 40 cm. En nuestro caso, la columna de agua en los tanques de producción la hemos subido a 55 o 60 cm. Esto permite que los animales tengan más espacio para el nado y los cortejos. Además, el tener un mayor volumen de agua ayuda a que la temperatura se mantenga estable y que no existan cambios bruscos. El mayor volumen de agua en el hábitat de los animales en producción se interpreta también como una condición de bienestar animal.
Buena calidad de agua. En maduración, el agua utilizada corresponde entre 60 a 65% de agua recirculada y un 35 a 40% de agua nueva. Se emplean filtros ecológicos, los cuales deben ser limpiados con frecuencia, al igual que las líneas de agua y tanques. La buena calidad de agua se refleja en las branquias de los reproductores. Branquias sucias ocasionan mortalidad y disminución de cópulas por estrés.
Beneficios de la no ablación
Con base en la experiencia y la información recopilada en el proceso de eliminar la práctica de ablación del ojo de las hembras, se han identificado los siguientes beneficios:
- Provee 10 a 15% más de huevos y nauplios, y a su vez, nauplios más fuertes.
- Permite ahorrar tiempo, ya que al no realizar ablación no es necesaria la preproducción y los animales pasan directo a producción después de su período de cuarentena.
- Existe menor mortalidad de hembras por no ser ablacionadas.
- Menor necesidad de reproductores ya que las mortalidades son bajas en producción.
- Hay una mayor tendencia en el mercado internacional de consumir animales que provengan de maduraciones que tengan



prácticas amigables con los animales en sus cultivos, por lo que eliminar la ablación se convierte en una ventaja competitiva.
Brindar mejores condiciones en el medio ambiente de producción a los reproductores, teniendo más cuidado en el trato de estos, da como resultado el bienestar de los animales expresado en:
-Menor mortalidad
-Estabilidad de cópulas
-Mayor porcentaje de cópulas
-Mayor cantidad de nauplios
-Mayor producción
-Menor necesidad de reproductores
Cabe mencionar que cuando se eliminó la ablación, la maduración inició con un promedio de 8% de cópulas diarias. En la actualidad, con un mejor manejo se ha llegado al 10% de cópulas diarias, produciendo 10 a 15% más de nauplios en comparación con la época en que se realizaba la ablación.
Resultados en camaroneras
En el siguiente gráfico se puede apreciar el incremento anual en la tasa de crecimiento promedio semanal a partir del 2019, así
50 - ABRIL 2024
PRODUCCIÓN
Foto 4: Siembra de los reproductores en tanques
Foto 2: Laceraciones en escafoceritos y urópodos producidos por choques o roces con la malla del tubo en que se las transportan
Foto 3: Forma incorrecta (izquierda) y correcta (derecha) de introducir las hembras o machos en los tubos
como las libras/ha/día ganadas. En 2019, con crecimiento lineal de 1.39 g/semana se obtenían 44 lb/ha/día. Estos resultados se lograron utilizando raceways en la recepción de las postlarvas.
En los siguientes años, gracias al mejoramiento genético y por mayor presión de selección de crecimiento entre los reproductores, se lograron mejores crecimientos semanales y más lb/ha/día, a pesar de haber incrementado la densidad de siembra. Hasta el 2022 se había obtenido una ganancia anual de año en año de 20 lb/ha/día, con un incremento aproximado del crecimiento lineal semanal de 0.15 g. En el año 2023 se empezaron a utilizar precriaderos como primera etapa en la recepción de las postlarvas en camaronera, obteniendo un incremento notablemente superior a los años anteriores, y pasando de 1.86 a 2.65 g/sem de crecimiento lineal y de 102 a 135 lb/ha/día (Gráfico 2).
Resumiendo, la producción de camarón de las fincas durante el año 2023 ha sido de 11,436 lb/ha. Esto se ha logrado con una densidad de siembra de 234,000 juveniles/ ha, logrando un crecimiento lineal de 2.65 g/sem. El promedio de días de cultivo fue de 85 y la supervivencia promedio 68,87%, obteniendo un peso final promedio de 32.17 g con un factor de conversión alimenticia de 1.56 (Gráfico 3).
La práctica de no ablación, junto con mayores niveles de agua y técnicas de manipulación cuidadosas, dan como resultado mayor bienestar animal.
En respuesta, los animales han incrementado el porcentaje de cópulas, dan mayor cantidad de huevos, mayor cantidad de nauplios, mejores supervivencias y, junto al programa de maduración ininterrumpido durante 15 años, han permitido ofrecer un producto de alta calidad para el Ecuador y el mundo•
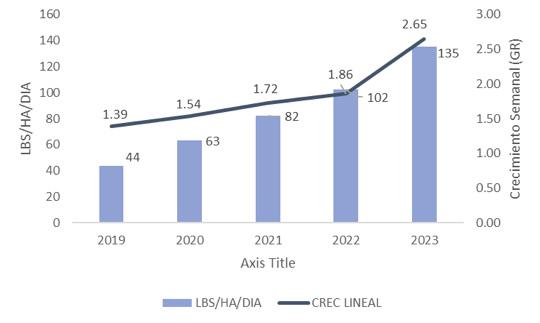
Gráfico 2: Crecimiento lineal semanal e Incremento de ganancia de lb/ha/día
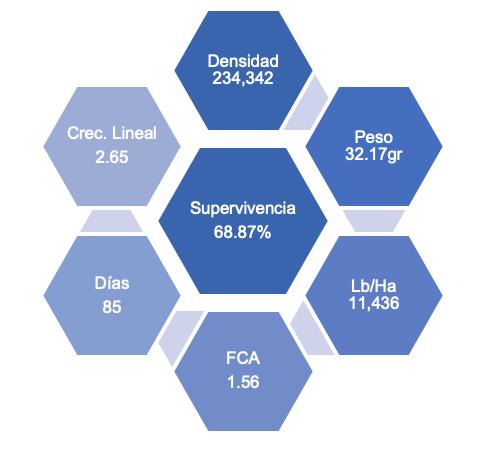
Gráfico 3: Resumen de la producción en camaronera 2023
Referencias
Global Aquaculture Advocate. (2019, Febrero). Investigadores de Stirling identifican alternativas de ablación viables para criaderos de camarón. Retrieved from Global Seafood Alliance: https://www.globalseafood.org/advocate/ investigadores-de-stirling-identifican-alternativas-de-ablacion-viables-para-criaderos-de-camaron/#:~:text=La%20 ablaci%C3%B3n%20ocular%20%E2%80%93%20la%20extirpaci%C3%B3n,muda%2C%20maduraci%C3%B3n%20 sexual%20y%20deso
Para mayor información escriba a: jgarcia@aquagen.com.ec
Global Seafood Advocate. (2016, Mayo). Camarón sin ablación de Seajoy responde a preocupación emergente de bienestar. Retrieved from Global Seafood Alliance: https://www.globalseafood.org/advocate/camaron-sin-ablacionde-seajoy-responde-a-preocupacion-emergente-de-bienestar/
- ABRIL 2024 51 PRODUCCIÓN

La acumulación tisular de microplásticos de poliestireno provoca estrés oxidativo, lesión hepatopancreática y alteraciones del metaboloma en Penaeus vannamei
Autores:
Yingxu Zenga,*
Baichuan Dengb
ZiXin Kanga
Pedro Araujoc
Svein Are Mjøsd
Ruina Liua
Jianhui Lina
Tao Yanga
Yuangao Que
a Laboratorio Clave de Procesos Marino-costeros y Sumidero de Carbono de la provincia de Hainan, Instituto de Innovación de la Bahía de Yazhou, Facultad de Ecología y Medio Ambiente, Universidad del Océano Tropical de Hainan, Sanya 572000, China
b Laboratorio Provincial Clave de Control de Nutrición Animal de Guangdong, Facultad de Ciencias Animales, Universidad Agrícola del Sur de China, Guangzhou 510642, China
c Instituto de Investigación Marina, 5817 Bergen, Noruega
d Departamento de Química, Universidad de Bergen, N-5020 Bergen, Noruega
e Instituto de Ciencia e Ingeniería de Fondos Marinos, Academia China de Ciencias, Sanya 572000, China
https://www.sciencedirect.com/science/article/pii/S0147651323003755?via%3Dihub
zengyx@hntou.edu.cn

En las últimas décadas, se han vertido continuamente grandes cantidades de desechos plásticos al océano debido a la falta de concienciación sobre su disposición final. En 2016, se estimó que la cantidad global de desechos plásticos que ingresaba a los ambientes acuáticos era de 19 a 23 millones de toneladas métricas (tm), con un aumento potencial a 53 millones de tm para 2030 (Borrelle et al., 2020). Se prevé que los microplásticos (MPs) que se producen por la descarga directa de micropartículas de plástico y la fragmentación de desechos plásticos más grandes en el ambiente, representen el 13.5% del presupuesto de plástico marino (Kane et al., 2020). Esto ha llevado a una detección cada vez mayor de altos niveles de MPs en el agua de mar superficial (alrededor de varios miles de partículas m—3), particularmente en zonas costeras, puertos, estuarios, manglares, etc. (Cai et al., 2018; Fu y otros, 2020; Kang et al., 2015; Li et al., 2020a). La amplia presencia de MPs también se confirmó en sedimentos marinos de puertos costeros, lo que equivaldría a concentraciones de MPs en el agua capilar de 2.2 a 5.4 mg L—1 (Claessens et al., 2011). Las enormes cantidades de MPs en el océano han generado preocupación global sobre su potencial impacto sobre la vida y los ecosistemas marinos.
Cada vez hay más pruebas que demuestran la ingesta de MPs por parte de animales acuáticos como peces (Lu et al., 2016), bivalvos (Wang et al., 2021a), crustáceos (Saborowski et al., 2022), zooplancton (Jeong et al., 2016). Los MPs ingeridos podrían acumularse en los tejidos durante un largo período de tiempo debido a su pequeño tamaño y su escasa biodegradabilidad. Las principales vías de ingesta de MPs en organismos acuáticos incluyen la ingesta oral, la respiración y la adherencia a la piel (Kim et al., 2021; Kolandhasamy et al., 2018). La principal vía de absorción de MPs en animales acuáticos se ha identificado como la ingesta gastrointestinal en bivalvos y crustáceos (D’Costa, 2022; ValenciaCastañeda et al., 2022b; Wang et al., 2021a). Los MPs ingeridos pueden acumularse rápidamente en el tracto gastrointestinal, cruzar las barreras biológicas y trasladarse a otros órganos internos a través del sistema circulatorio o linfático (Kim et al., 2021).
53 - ABRIL 2024 AMBIENTE
AMBIENTE
Las branquias son los órganos respiratorios que muchos organismos acuáticos utilizan para oxigenar su sangre extrayendo oxígeno disuelto del agua. Por lo tanto, una gran superficie de branquias proporciona una vía de absorción de MPs transmitidos por el agua a través de la respiración, la actividad de las microvellosidades y la endocitosis (Von Moos et al., 2012). En particular, las variaciones morfológicas y fisiológicas entre las especies acuáticas suelen conducir a discrepancias en las características de biodistribución y bioacumulación de los MPs. Por lo tanto, es fundamental investigar la acumulación característica de MPs en especies acuáticas específicas, identificar órganos con fuerte toxicidad de MPs y evaluar los modos de acción tóxica relevantes.
Estudios han demostrado que la acumulación de MPs en los tejidos podría causar un gran número de efectos adversos sobre los animales acuáticos (D’Costa, 2022; Ko¨gel et al., 2020). Por ejemplo, la acumulación abundante de MPs en el tracto digestivo puede provocar bloqueos físicos y lesiones mecánicas en animales acuáticos (Alimba y Faggio, 2019), lo que provoca además una reducción de la alimentación, un comportamiento deteriorado e inhibición del crecimiento y desarrollo (Wang et al., 2020). La acumulación a largo plazo de MPs en el intestino de los peces puede provocar inflamación intestinal y estrés oxidativo, lo que lleva al desarrollo de trastornos metabólicos intestinales debido a la disbiosis de la microbiota intestinal (Kang et al., 2021; Qiao et al., 2019). Además, la bioacumulación de MPs en el hígado de peces se ha relacionado con la inflamación hepática, el estrés oxidativo elevado (Feng et al., 2021; Lu et al., 2016), así como con la alteración del metabolismo de los glicolípidos hepáticos en varios niveles (Zhao et al., 2020). La acumulación de MPs en los tejidos también afecta significativamente a otros animales acuáticos como los bivalvos y los crustáceos (D’Costa, 2022; Mkuye et al., 2022). Estudios de exposición han observado que la ingesta de MPs puede afectar la funcionalidad del sistema inmunológico de los bivalvos al interferir con los sistemas antioxidantes, reducir la fagocitosis, el recuento de hemocitos y la viabilidad, además de aumentar daños al ADN (Mkuye et al., 2022). Además, los MPs causaron daño histológico
y alteraciones en los biomarcadores enzimáticos del metabolismo energético y de lípidos en órganos importantes, incluyendo las branquias y la glándula digestiva de los bivalvos, así como la alteración de las vías del metabolismo de aminoácidos y los glicerofosfolípidos en la glándula digestiva (Teng et al., 2021).
En cuanto a crustáceos marinos, nuevas investigaciones han comenzado a descubrir efectos potenciales de la acumulación de MPs en la supervivencia, el crecimiento, la alimentación, el comportamiento, la fisiología, el estrés oxidativo, los cambios microbianos, etc. (Bergami et al., 2016; D’Costa, 2022; Wang et al., 2020, 2021b). Por ejemplo, estudios de campo han documentado la ingestión de MPs en camarón Penaeus vannamei y han demostrado la acumulación de MPs en el tracto gastrointestinal y en las branquias, donde los polímeros más abundantes detectados fueron polietileno, poliamida y poliestireno (PS) (ValenciaCastañeda et al., 2022a, 2022b). Varios estudios toxicológicos han reportado que los MPs acumulados en los tejidos pueden provocar un aumento de la mortalidad, una reducción de la tasa de alimentación y una alteración del comportamiento de natación de los crustáceos, según lo revisado por D’Costa (2022). Además, la retención de MPs en los tejidos puede ejercer una influencia sobre el sistema antioxidante, provocando daños oxidativos y lesiones histopatológicas en P. vannamei (Hsieh et al., 2021), así como la activación de la expresión de genes antioxidantes y la regulación de la vía de señalización MAPK en el cangrejo Eriocheir sinensis (Yu et al., 2018). Estudios recientes también encontraron que la exposición a MPs puede inducir una respuesta de defensa inmune y disbiosis de la microflora en P. vannamei (Wang et al., 2021b) y alterar los perfiles proteicos y metabólicos en la hemolinfa de P. vannamei (Duan et al., 2021).
Según algunos estudios sistemáticos centrados en 2020, la producción mundial anual de P. vannamei alcanzó los 5.8 millones de toneladas con un valor estimado de 24.7 mil millones de dólares, lo que representa el 51.7% del total de especies de crustáceos cultivados (FAO, 2022). Los desechos plásticos generados por las actividades
de acuicultura y pesca, junto con otras fuentes antropogénicas terrestres, están ingresando cada vez más a los ecosistemas de producción, lo que representa una gran amenaza para el bienestar y la sostenibilidad de P. vannamei. Por lo tanto, se seleccionó a P. vannamei como organismo de prueba en este estudio. Se seleccionaron microesferas de poliestireno (PS) como MP para los experimentos de exposición, ya que el PS es uno de los plásticos más fabricados y los tipos de polímeros de MPs más predominantes detectados en crustáceos marinos (Alimba y Faggio, 2019; Hara et al., 2020; Valencia-Castañeda et al., 2022b). Este estudio planteó la hipótesis de que P. vannamei puede ingerir y acumular MPs de PS en órganos específicos, lo que produce una variedad de efectos tóxicos a nivel conductual, histológico y bioquímico. En este caso, P. vannamei estuvo expuesto a MPs transmitidos por el agua en concentraciones ambientalmente relevantes. La acumulación y distribución de MPs en los órganos se determinó mediante rastreo de fluorescencia. Se realizó de manera conjunta el monitoreo del comportamiento, ensayo de biomarcadores enzimáticos, análisis histopatológico y perfiles metabolómicos para desentrañar las consecuencias toxicológicas interrelacionadas y los mecanismos relevantes de los MPs en P. vannamei
Experimento
Animales y MPs
Se obtuvieron postlarvas sanas de camarón blanco (P. vannamei, de 1–2 cm de longitud corporal, postlarvas de 25 días (PL25)) de Hainan Zhongzheng Aquatic Products Technology Co., Ltd (Hainan, China) y posteriormente se cultivaron con agua de mar artificial (ASW, composición de ASW se muestra en la Tabla S1) con una salinidad de 20 ± 1‰ mediante el uso de sal marina acuática (Blue Treasure, China) y agua desionizada. Los individuos fueron alimentados diariamente con una dieta comercial formulada para camarón blanco (5% del peso corporal, Guangdong Yuehai Diet Co., Ltd, Guangdong, China) diariamente. Durante el periodo de aclimatación, los organismos fueron mantenidos en acuarios durante tres semanas en condiciones de laboratorio (ciclos luz/oscuridad (12h/12h), temperatura 28 ±1°C; pH 7.6 ±0.1; oxígeno disuelto: 7.4 ±0.3 mgL-1).
54 - ABRIL 2024
Se adquirieron microesferas de poliestireno (PS) con un diámetro uniforme de 2 µm en el Centro de Investigación BaseLine ChromTech (Tianjin, China). Aunque el estudio de campo mostró que las esferas de MPs más abundantes detectadas en camarones tenían entre 10 y 20 µm de diámetro (Curren et al., 2020), muchos MPs en el ambiente se presentan como pequeños MPs secundarios derivados de la degradación de los plásticos, que apenas se detectan en los estudios de campo debido a limitaciones técnicas (Barbosa et al., 2020). Por lo tanto, en este estudio se seleccionó un tamaño de PS pequeño (diámetro de 2 μm) considerando la degradación ambiental de los plásticos, que normalmente exhibe una mayor biodisponibilidad para los organismos que los MPs más grandes, pero sus efectos toxicológicos permanecen en gran medida inexplorados (Dawson et al., 2018; Lee et al., 2013). Las microesferas de PS marcadas con fluorescencia (2 μm de diámetro) se utilizaron para rastrear la bioacumulación y biodistribución de PS en el organismo (longitud de onda de excitación: 488 nm; longitud de onda de emisión: 518 nm). Para la prueba de toxicidad crónica se emplearon microesferas de PS virgen (2 μm de diámetro) sin fluorescencia. La composición polimérica de los MPs se examinó mediante espectroscopía micro-Raman (DXR™, Thermo-Fisher Scientific) (Fig. S1). El tamaño y la morfología de las microesferas de PS se caracterizaron mediante microscopía electrónica de transmisión (Tecnai G2 F20, FEI, Países Bajos) (Fig. S1). La solución stock de MPs de 1.0 mg ml-1 se preparó utilizando ASW. La solución stock preparada se trató en un baño ultrasónico antes de su uso.
Acumulación y distribución de MPs Para evaluar la acumulación, distribución y eliminación de MPs, se cultivaron aleatoriamente juveniles sanos y aclimatados de P. vannamei (4.4 ± 0.4 cm de longitud corporal, 0.49 ± 0.06 g de peso corporal) en 3 tanques de vidrio por triplicado (un control y dos tratamientos). El control estuvo formado por un total de 24 individuos distribuidos en 3 tanques (8 × 3) que contenían 2.5 L de ASW puro cada uno sin MPs y sin ciclo de depuración después de 48 h de exposición. Los dos tratamientos consistieron en 48 individuos distribuidos equitativamente en 6 tanques (8 x 6) que contenían 2.5 L
de ASW cada uno y enriquecidos con MPs fluorescentes a una concentración de 1.0 mg L—1. Esta concentración es la misma que la concentración más alta de MPs (1.0 mg L-1) utilizada en la prueba de toxicidad crónica posterior, que refleja las concentraciones ambientales de MPs en el agua de mar superficial de áreas costeras, puertos, estuarios y manglares (p. ej., 3.7 mg L-1) (Cai et al., 2018; Fu et al., 2020; Kang et al., 2015; Li et al., 2020a) y en agua capilar de sedimentos marinos de puertos costeros (2.2–5.4 mg L—1) (Claessens et al., 2011). Los experimentos para el primer tratamiento (8 x 3) finalizaron después de 48 h de exposición a MPs. Por el contrario, los experimentos para el segundo tratamiento (8 x 3) se sometieron a un ciclo de depuración de 48 h después de la exposición inicial de 48 h a MPs. Durante el experimento, todos los camarones estuvieron en ayunas y la solución en cada tanque se cambió diariamente. Todas las demás condiciones siguieron siendo las mismas que durante el período de aclimatación. Después del experimento, los camarones fueron sacrificados en hielo y se recolectaron diferentes órganos (branquias, intestino, estómago y hepatopáncreas) para análisis posteriores.
Para determinar la ingesta de MPs por parte de P. vannamei y observar la biodistribución de MPs en varios órganos (branquias, intestino, estómago, hepatopáncreas), los órganos recolectados se fijaron en paraformaldehído al 4%, se incluyeron en cera de parafina y se seccionaron en grosor de 4 µm para observación microscópica. Los análisis se realizaron con un microscopio confocal de barrido láser (Nikon, Eclipse Ti, Japón) utilizando una fuente láser de 480 nm. Las imágenes obtenidas fueron procesadas por el software Image J (http:// imagej.net/ versión 1.53k) para calcular el tamaño de partícula.
Para análisis cuantitativo de MPs, los tejidos de camarón se almacenaron a -20°C. Los tejidos de dos individuos se agruparon como una muestra y se realizaron tres réplicas para los análisis. Cada muestra fue pesada y se digirió en 8 mL de solución de KOH al 10% a 60°C durante 48 h. El protocolo de digestión se modificó ligeramente con base en un estudio previo (Dehaut et al., 2016), que ofreció una digestión eficiente de los
tejidos de los mariscos sin una degradación evidente de los polímeros MPs. La solución de digestión resultante se diluyó con agua desionizada hasta 8 mL y sus intensidades fluorescentes se determinaron por triplicado utilizando un espectrofotómetro de fluorescencia (Agilent Cary Eclipse G9800A) con una longitud de onda de excitación de 488 nm. Los contenidos de MPs acumulados en varios tejidos se cuantificaron según la curva estándar construida por una serie de soluciones de MPs fluorescentes (Fig. S2). Los resultados finales se presentaron como concentración de masa μg (partícula)/ mg (peso húmedo) (precisión (CV%), 4.1% –12.4%).
Experimento de toxicidad
Para la prueba de toxicidad, se utilizaron juveniles sanos y aclimatados de P. vannamei (4.4 ± 0.4 cm de longitud corporal, 0.49 ± 0.06 g de peso corporal) que fueron asignados aleatoriamente a un grupo control y tres grupos de exposición a MPs con cinco tanques de vidrio replicados para cada grupo (8 individuos en un tanque). Los camarones se expusieron en tanques de vidrio de 3 L con 2.5 L de solución de prueba durante 8 días. Esta duración de exposición se seleccionó considerando el intervalo de muda de P. vannamei (alrededor de 4 a 6 días en la etapa juvenil), que es un proceso fisiológico crucial en su ciclo de vida, de modo que abarque al menos un ciclo de muda durante el período de exposición (Betancourt-Lozano et al., 2006; Corteel et al., 2012). Los individuos del grupo control fueron expuestos a ASW preparado como se describe anteriormente, mientras que los individuos del grupo expuesto a MPs estuvieron expuestos a MPs PS en ASW en concentraciones de 0.02 mg L—1 (PS-L), 0.2 mg L—1 (PS-M) y 1.0 mg L—1 (PSH), respectivamente. Estas concentraciones de exposición representan niveles de MPs encontrados en aguas marinas superficiales de áreas costeras, puertos, estuarios, manglares, así como el rango más bajo de concentraciones de MPs comúnmente empleadas en experimentos de exposición (Cai et al., 2018; Fu et al., 2020; Lenz et al., 2016; Li et al., 2020a; Phuong et al., 2016). Todas las otras condiciones permanecieron iguales al período de aclimatación. Los individuos fueron alimentados con dietas comerciales formuladas (5% peso corporal) una vez al día como se describió
55 - ABRIL 2024 AMBIENTE
AMBIENTE
anteriormente. Después de 8 días de exposición, los camarones se utilizaron para análisis posteriores que incluyeron pruebas de comportamiento de natación (camarones vivos), análisis histopatológicos (las muestras se fijaron en paraformaldehído al 4%), análisis bioquímicos y metabolómicos (las muestras se congelaron en nitrógeno líquido y transferidos a -80°C), respectivamente.
Prueba de comportamiento de natación Después del período de exposición tóxica, se sacaron camarones al azar de cada grupo para evaluar el comportamiento de natación (se utilizaron 12 individuos para cada grupo, 4 individuos por tanque para cada réplica). Posteriormente se transfirieron a tanques de vidrio de 30 x 18 x 20 cm (L x W x H) que contenían 2 L de solución de ensayo. Para registrar su comportamiento de natación, se fijó una cámara de video (Sony, FDR-AX60, Japón) en el marco de la parte superior del tanque de vidrio para asegurar la captura de todo el fondo rectangular del tanque en la misma posición. La prueba de comportamiento se realizó por triplicado. Para cada prueba, se registró simultáneamente el comportamiento de natación de cuatro camarones de cada grupo durante 4 minutos después de 5 minutos de aclimatación (Wang et al., 2020). Los vídeos grabados fueron procesados por un software de seguimiento automatizado basado en imágenes ToXTrac (Universidad de Umeå, Umeå, Suecia), que puede rastrear múltiples organismos simultáneamente con la preservación de su identidad en el entorno del laboratorio y producir parámetros de comportamiento básicos en un entorno de laboratorio de manera sólida y eficiente en el tiempo (Rodríguez et al., 2018). Se midió la velocidad promedio, la distancia total recorrida y la tasa de exploración de cada individuo en cada tanque y exportado en formato Excel. La velocidad promedio se definió como el promedio de la velocidad instantánea en mm s—1. La distancia total recorrida representó la distancia total nadada en mm. La tasa de exploración se presentó como el porcentaje del número de áreas exploradas dividido por el número de áreas totales. Estos parámetros se visualizaron adicionalmente mediante gráficos de barras y mapas de calor. Los mapas de calor están codificados por colores de acuerdo con la representación normalizada de la frecuencia
de uso en la escala lineal y se muestran superpuestos a la imagen de la arena (es decir, el fondo del tanque rectangular) en escala real (Rodríguez et al., 2018).
Análisis bioquímico
El sistema de defensa antioxidante de P. vannamei fue investigado para medir las respuestas de los biomarcadores relevantes del estrés oxidativo que incluyen a la actividad de superóxido dismutasa (SOD), catalasa (CAT), glutatión peroxidasa (GPX) y malondialdehído (MDA) para la exposición a MPs (n = 3– 6). Además, se midió el indicador de respuesta neurotóxica, es decir, acetilcolinesterasa (AChE). Estos biomarcadores y el contenido de proteína total se analizaron con kits de detección comerciales, de los cuales el kit de ensayo de proteína de ensayo de ácido bicinconínico mejorado (BCA), el kit de ensayo de SOD total con WST-8, el kit de ensayo CAT, el kit de ensayo GPX celular con NADPH y el kit de peroxidación lipídica MDA fueron de Beyotime Biotechnology (Shanghai, China) y el kit de ensayo de AChE fue del Instituto de Bioingeniería Jiancheng (Nanjing, China) (los detalles de estos kits se muestran en el material complementario). Se reunieron tres camarones como una muestra y cada una se homogeneizó en suficiente solución tampón helada mediante un homogeneizador de tejidos (Tissuelyser, Shanghai, China). Los homogeneizados se centrifugaron a 10,000 g durante 15 min a 4°C. Posteriormente se recogieron los sobrenadantes para la determinación de biomarcadores. Las operaciones experimentales se realizaron con base en los protocolos de los fabricantes y las mediciones se realizaron mediante un espectrofotómetro equipado con un lector de microplacas (Thermo Scientific™ Varioskan™ Flash).
Análisis histopatológico
Los hepatopáncreas de P. vannamei de varios grupos (n=3 para cada grupo, 3 cortes de tejido para cada muestra) se diseccionaron y se fijaron inmediatamente en paraformaldehído al 4%. Los tejidos fijados se embebieron en cera de parafina, se seccionaron a 4 µm de espesor y se tiñeron con hematoxilina y eosina (H&E). Los cortes de tejido se observaron bajo microscopio para evaluar el daño del hepatopáncreas mediante análisis histopatológico, que
se evaluaron semicuantitativamente clasificando la gravedad de las lesiones como se describió anteriormente (Lin et al., 2022).
Análisis metabolómico
Se agruparon los hepatopáncreas de tres P. vannamei como una muestra y se realizaron seis réplicas para cada grupo. El protocolo de extracción de metabolitos se adaptó de un estudio anterior (Fu et al., 2018). Brevemente, los tejidos hepatopancreáticos (40 ± 1 mg) se homogeneizaron con 400 μL de agua Milli-Q helada y enriquecida con 800 μL de metanol/acetonitrilo (1:1, v/v), seguido de un tratamiento con ultrasonido para extracción de metabolitos. Los extractos de metabolitos preparados se sometieron al análisis de cromatografía líquida-espectrometría de masas en tándem (LC-MS/MS) utilizando un sistema de cromatografía líquida de rendimiento ultra alto (UPLC) junto con un espectrómetro de masas Orbitrap Q EXactive Focus (Thermo Fisher Scientific, EE.UU.). La separación cromatográfica se realizó en una columna Hypersil Gold C18 utilizando un sistema de fase móvil binario (A: agua, B: acetonitrilo). La data de espectrometría de masas se adquirió mediante ionización por electropulverización en los modos de ionización negativa y positiva. La data sin procesar de LC-MS/ MS fue procesada por Compound Discover 3.1 (Thermo Fisher Scientific, EE. UU.) para la asignación, extracción, alineación, identificación e integración del área de los picos. La preparación de muestras detallada y los parámetros instrumentales se proporcionan en el material complementario.
Análisis estadístico
Los análisis estadísticos univariados se realizaron mediante el software SPSS 21.0. La normalidad de la data y la homogeneidad de las varianzas se evaluaron mediante la prueba de Kolmogorov-Smirnov para una muestra y la prueba de Levene. Posteriormente, se realizó un ANOVA unidireccional seguido de la prueba post hoc de Tukey para evaluar las diferencias estadísticas de varios parámetros entre los distintos tratamientos. Los análisis estadísticos multivariados de la data metabolómica se realizaron mediante SIMCA 14.1.0 (Umetrics, Suecia). La data metabolómica se examinó primero mediante análisis de componentes principales (PCA)
56 - ABRIL 2024
para detectar agrupaciones y valores atípicos. A continuación, se construyó un modelo de análisis discriminante de mínimos cuadrados parciales (PLS-DA) para cuatro grupos para dilucidar las tendencias de las alteraciones metabólicas inducidas por los diferentes tratamientos. El modelo PLS-DA obtenido se validó basándose en las puntuaciones CVANOVA PLS para probar su significancia. Los metabolitos con importancia de variable en la proyección (VIP) > 1 y univariada p < 0.05 se consideraron metabolitos significativamente alterados en los grupos de exposición a MPs en comparación con el grupo control. Se generó un mapa de calor con MetaboAnalyst 5.0 (www.metaboanalyst.ca/ ) para presentar el patrón de cambios metabólicos de metabolitos significativamente diferentes entre los grupos control y tratamiento.
Resultados
Distribución y acumulación de MPs
Se utilizó la técnica del trazador de fluorescencia para rastrear la distribución y acumulación en varios órganos de P. vannamei. Los resultados obtenidos de secciones histológicas de camarones expuestos mediante microscopía confocal de barrido láser mostraron que las partículas de MPs se observaron claramente en branquias, estómago, intestino y hepatopáncreas, y sus características de distribución en varios órganos fueron registrados (Fig. 1, Tabla S2). En las branquias, se dispersaron pequeños MPs con un tamaño promedio de 1.98 ± 0.31 µm en el filamento branquial (Fig. 1A), mientras que varias manchas grandes de MPs fluorescentes (tamaño 14.7–25.5 µm) junto con muchas pequeñas manchas de MPs se visualizaron claramente en el estómago (Fig. 1B). En cuanto al intestino, las manchas de MPs observadas (1.26 ± 0.38 µm de tamaño) eran básicamente más pequeñas que los MPs expuestos (2 µm de tamaño), con la excepción de dos puntos de MPs más grandes (3.5 µm y 2.2 µm de tamaño), que eran ligeramente más grandes que los MPs expuestos (Fig. 1C). En el hepatopáncreas se detectaron muchos MPs diminutos con un tamaño similar al de los MPs expuestos dentro de los túbulos del hepatopáncreas (Fig. 1D); además, se agregaron dos grandes manchas fluorescentes (tamaño 21.0 µm y 19.6 µm) adyacentes a los túbulos del hepatopáncreas.
Los contenidos de MPs en diferentes órganos
se cuantificaron mediante espectrofotómetro de fluorescencia (Fig. 1E). Se puede observar que la acumulación de MPs en el hepatopáncreas fue más abundante que en branquias, estómago e intestino tanto durante el período de exposición como de eliminación (p <0.05) (Figura 1E). El contenido de MPs acumulado en el intestino fue significativamente mayor que en las branquias (p <0.05). Después del período de eliminación, el contenido de MPs en branquias, estómago e intestino disminuyó significativamente (p <0.05), mientras que la acumulación de MPs en el hepatopáncreas no disminuyó significativamente después de la eliminación (Fig. 1E). En particular, la tendencia de distribución de absorción de los MPs en varios órganos permaneció similar antes y después del período de depuración, aunque los MPs ingeridos se depuraron hasta cierto punto.
Supervivencia, crecimiento y comportamiento de natación
Para la prueba de toxicidad, no se encontraron diferencias significativas en la tasa de supervivencia entre varios grupos (Fig. S3). Sin embargo, en comparación con los controles, el crecimiento de P. vannamei se inhibió en un 52.1 % cuando se expuso
Branquia
Estómago
a la concentración más alta de MPs (PSH, 1.0 mg L—1) (Fig. S3). Los efectos de los MPs sobre el comportamiento de natación de P. vannamei se muestran en la Fig. 2. En comparación con los controles, la exposición a los MPs en tres concentraciones disminuyó significativamente la velocidad promedio de natación de P. vannamei en un 43.7 % (PS-L), 57.9 % (PS-M) y 80.2% (PS-H), respectivamente (Fig. 2A).
De manera similar, se detectó un efecto distinto dependiente de la concentración para la distancia total de natación, donde la exposición a los MPs redujo significativamente la distancia total de natación en un 39.6 % (PS-L), un 55.3 % (PSM) y un 79.9 % (PS-H), respectivamente, en relación con el del grupo control (Fig. 2B). En cuanto a la tasa de exploración, solo la exposición a MPs en la concentración más alta disminuyó significativamente la tasa de exploración de los camarones comparable a la del grupo control (Fig. 2C). El rango de nado y la frecuencia de P. vannamei en varios grupos también se monitoreó utilizando el software de seguimiento del comportamiento animal ToXTrac y posteriormente se visualizó mediante mapas de calor (Fig. 2D-G). Los
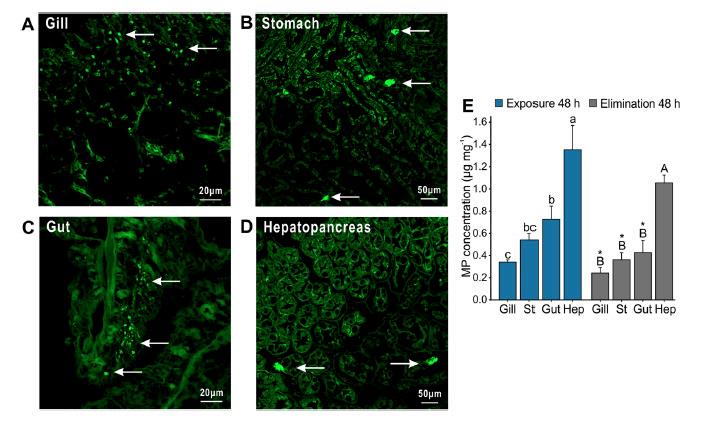
Intestino
Hepatopáncreas
Concentración de MPs (μg mg -1 )
Fig. 1. Distribución y acumulación de MPs fluorescentes en varios órganos de P. vannamei expuestos a 1.0 mg L—1 MP. Se observaron MPs fluorescentes verdes en branquias (A), estómago (B), intestino (C) y hepatopáncreas (D) de P. vannamei mediante microscopía confocal de barrido láser. Las flechas blancas indican manchas típicas de MPs en los órganos. (E) Concentraciones de MPs en varios órganos de P. vannamei después de períodos de exposición y eliminación de 48 h (St: estómago; Hep: hepatopáncreas). Los valores son media ± DE (n = 3). Las diferentes letras representan diferencias significativas entre las concentraciones de MPs en diferentes órganos, y los asteriscos indican diferencias significativas de la concentración de MPs en un órgano específico entre dos puntos temporales (p <0.05, ANOVA seguido de la prueba post-hoc de Tukey).
57 - ABRIL 2024 AMBIENTE
AMBIENTE
camarones en el grupo control generalmente mostraron un rango de nado más amplio y una mayor frecuencia de nado en comparación con los grupos de exposición a MPs. En particular, los camarones del grupo PS-H exploraron el menor espacio entre todos los grupos (Fig. 2G). Además, los camarones en los grupos de exposición a MPs mostraron una tendencia a nadar pegados a la pared.
Respuestas de biomarcadores
Se examinaron los niveles de una serie de biomarcadores en P. vannamei para estudiar el estrés oxidativo, la peroxidación lipídica y la neurotoxicidad inducida por diversas concentraciones de MPs (Fig. 3). En comparación con los controles, la actividad de SOD mostró una tendencia creciente después de la exposición a los MPs, y se elevó significativamente en el grupo PS-H (p <0.05) (Fig. 3A). Por el contrario, la actividad CAT mostró una tendencia a la baja dependiente de la concentración después de la exposición a MPs (p < 0.05) (Fig. 3B). En particular, se observaron fuertes correlaciones positivas entre la actividad CAT y los parámetros de rendimiento en natación, incluyendo la velocidad promedio de natación (r = 0.804, p < 0.01) y la distancia total de natación (r = 0.727, p <0.01) (Tabla S3). La exposición a concentraciones bajas y altas de MPs indujo aumentos significativos en las actividades de GPX en comparación con el grupo control (p <0.05) (Fig. 3C). Además, los niveles de MDA desarrollaron respuestas crecientes dependientes de la concentración después de la exposición a MPs, donde la exposición a la concentración más alta de MPs indujo aumentos significativos en los niveles de MDA en comparación con los controles (p <0.05) (Fig. 3D). Además, el nivel de MDA estuvo altamente correlacionado con la velocidad promedio de natación (r = – 0.748, p <0.01) y la distancia total de natación (r = – 0.762, p <0.01) (Tabla S3). En cuanto a los niveles de AChE, no se observaron diferencias significativas entre los distintos grupos (Fig. 3E).
Observaciones histológicas
El examen microscópico reveló que el hepatopáncreas de P. vannamei de los controles mostró una estructura bien definida de túbulos, lumen y células (Fig. 4A). Después de la exposición a bajas concentraciones de MPs, la estructura
morfológica general del hepatopáncreas era cercana a la del grupo control y el lumen tubular en forma de estrella aún se conservaba en el hepatopáncreas del grupo PS-L; sin embargo, los túbulos mostraron una disposición laxa y algunas de las células epiteliales se rompieron (Fig. 4B). Cuando se expuso a una concentración media de MPs, se observó claramente un lumen distorsionado y deformado, un aumento del tamaño de las células B y una citólisis de las células epiteliales en el hepatopáncreas del grupo PS-M (Fig. 4C). En particular, la exposición a altas concentraciones de MPs causó una acumulación evidente de MPs en las células epiteliales tubulares, lo que eventualmente resultó en cambios histopatológicos severos en el hepatopáncreas del grupo PS-H, incluyendo atrofia tubular, disposición laxa de los túbulos y pérdida de la estructura de lumen en forma de estrella, citólisis de células epiteliales y deterioro de la integridad
de la estructura tubular (Fig. 4D). El grado semicuantitativo de los cambios histológicos en el hepatopáncreas de P. vannamei se resumió en la Tabla 1.
Cambios metabólicos
El enfoque metabolómico LC-MS/ MS caracterizó 119 metabolitos en el hepatopáncreas de P. vannamei, que incluían aminoácidos, ácidos grasos, lípidos, péptidos, acilcarnitinas, ácidos nucleicos y ácidos orgánicos. La inspección exploratoria inicial realizada por PCA presentó una buena descripción general de la estructura de la data, que mostró una agrupación de clases adecuada de cuatro grupos en el gráfico de puntuaciones después de excluir dos valores atípicos (Fig. S4). El modelo PLS-DA se construyó para todos los grupos para dilucidar las alteraciones metabólicas inducidas por las diferentes condiciones de exposición (CVANOVA, p = 0.01). El gráfico de puntuaciones
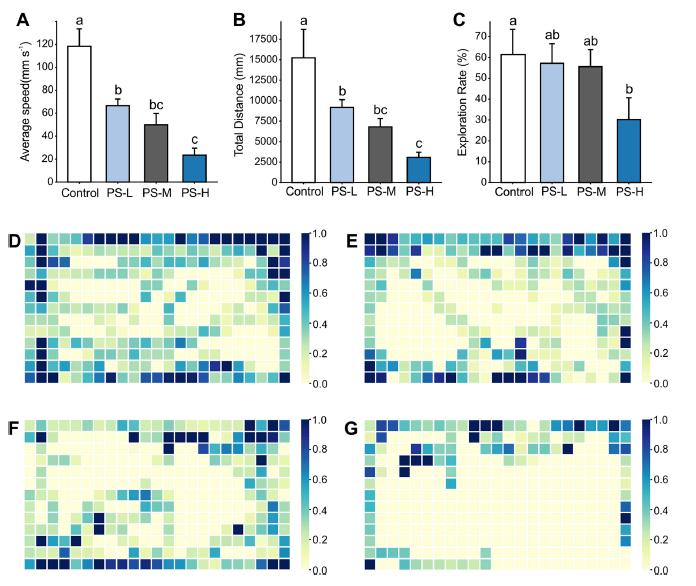
Fig. 2. Comportamiento de natación y rendimiento de P. vannamei después de la exposición a MPs. Velocidad promedio (A), distancia total (B) y tasa de exploración (C) de P. vannamei después de exposición durante 8 días a 0 (control), 0.02 mg L—1 (PS-L), 0.2 mg L—1 (PS -M) y 1.0 mg L—1 (PS-H) MPs. Los valores son media ± DE (n = 3). Las diferentes letras indican diferencias significativas entre las concentraciones de MPs en diferentes órganos (p < 0.05, ANOVA seguido de la prueba post-hoc de Tukey). Los mapas de calor representan el rango de natación y la frecuencia de P. vannamei en los grupos control (D), PS-L (E), PS-M (F) y PS-H (G) medidos durante 4 minutos en el acuario. Se muestran los ejes “x” y el “y” superpuestos a la imagen de la arena (es decir, el fondo rectangular del acuario) en escala real. La barra de intensidad de color indica la frecuencia de nado de P. vannamei
58 - ABRIL 2024
mostró que los perfiles metabólicos entre los grupos control y grupos expuestos a MPs estaban bien separados (Fig. 5A). Esto indicó que la exposición a MPs causó alteraciones metabólicas evidentes en el hepatopáncreas de P. vannamei que variaron con las concentraciones de MPs. El gráfico de cargas reveló los principales metabolitos responsables de la discriminación de los perfiles metabólicos de varios grupos desde los aspectos de las dos cargas (Fig. S5). Los metabolitos diferenciales entre cuatro grupos se identificaron mediante enfoques univariados y multivariados, y sus patrones de alteración dentro de varios grupos se visualizaron mediante mapas de calor (Fig. 5B). Por ejemplo, el gliceraldehído se redujo significativamente en el hepatopáncreas de los grupos expuestos a MPs en comparación con los controles. Los ácidos grasos, incluidos el ácido 3-metiladípico, el ácido azelaico, el ácido subérico, el ácido adípico, el ácido sebácico, el ácido leucínico y el ácido 2-hidroximirístico, se redujeron significativamente debido a la exposición a bajas concentraciones de MPs. Mientras tanto, la exposición a MPs provocó cambios pronunciados en los niveles de varias acilcarnitinas. Además, los niveles de aminoácidos y dipéptidos, como taurina, ácido L-aspártico y N-acetilL-alanina, aumentaron significativamente después de la exposición a MPs en todas las concentraciones. Las principales vías metabólicas perturbadas por la exposición a los MPs se resumen en la Fig. 6. El análisis de las vías metabólicas demostró además que la exposición a los MPs podría influir en las vías del metabolismo de la glucólisis, la lipólisis y los aminoácidos.
Discusión
Distribución y acumulación de MPs en P. vannamei
Para identificar órganos con toxicidad significativa de MPs y explorar los mecanismos subyacentes, es crucial investigar las características de distribución y acumulación de MPs en los organismos. Nuestros resultados demostraron la captación y acumulación de MPs en branquias, estómago, intestino y hepatopáncreas de P. vannamei, pero con diferentes características de distribución de MPs entre varios órganos. En las branquias, el tamaño de los MPs observados es similar al
tamaño de los MPs expuestos (2 µm), lo que sugiere que P. vannamei podría ingerir MPs predominantemente en sus formas prístinas. Esto es consistente con hallazgos previos de acumulación de MPs en las branquias de peces (Lu et al., 2016), bivalvos (Li et al., 2020b; Wang et al., 2021a) y crustáceos (D’Costa, 2022). En el estómago y el hepatopáncreas, el fenómeno más evidente fue el tamaño significativamente mayor de varios puntos de MPs fluorescentes que de los MPs prístinos. Se observaron fenómenos similares en otros estudios que emplearon bivalvos como modelos animales de exposición a MPs (Li et al., 2020b; Wegner et
al., 2012), lo que podría atribuirse a la homoagregación de MPs en organismos y a la hetero-agregación de MPs interactuando con biomacromoléculas en P. vannamei (Alimi et al., 2018; Li et al., 2020b; Saborowski et al., 2022). Además, se produjeron muchos MPs diminutos (< 1 µm) en el sistema digestivo de P. vannamei, lo que podría explicarse por la alteración mecánica de los MPs por los molinos gástricos, así como por la ayuda de las enzimas digestivas (Dawson et al., 2018). Por ejemplo, el krill antártico puede fragmentar la mayoría de los MPs ingeridos (31.5 µm) en trozos más pequeños (< 1 µm) que se dirigen al intestino para su excreción
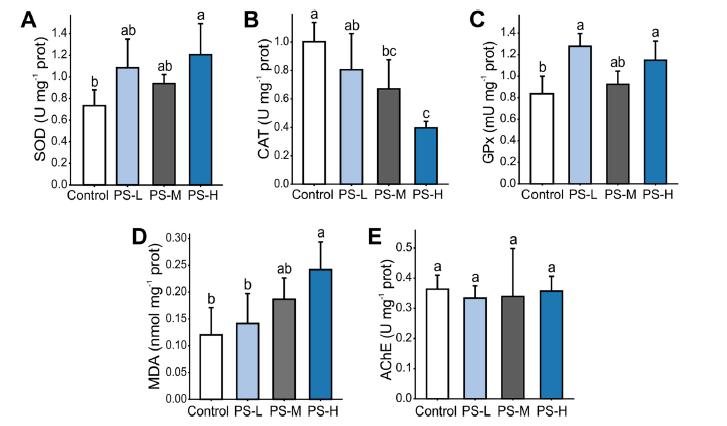
Fig. 3. Efectos de la exposición a MPs sobre biomarcadores de estrés oxidativo y neurotoxicidad en P. vannamei. (A) actividad SOD; (B) actividad CAT; (C) actividad GPX; (D) niveles MDA; (E) Actividad AChE. Los valores son media ± DE (n = 3–6). Las diferentes letras indican diferencias significativas entre tratamientos (p <0.05, ANOVA seguido de la prueba post hoc de Tukey).
Fig. 4. Observaciones histológicas en hepatopáncreas de P. vannamei después de la exposición a MPs. Escala de barra (20 µm). (A) control; (B) PS-L, 0.02 mg L—1; (C) PS-M, 0.2 mg L—1; (D) PS-H, 1.0 mg L—1 . Abreviaturas: B (células B, células secretoras), L (lumen en forma de estrella), ALU (lumen anormal), MP (microplásticos), punta de flecha negra (citólisis).
59 - ABRIL 2024 AMBIENTE
AMBIENTE
(Dawson et al., 2018). Este proceso de degradación podría facilitar la internalización de pequeños MPs mediante endocitosis en células epiteliales y mejorar la capacidad de los MPs pequeños para cruzar barreras biológicas, afectando así la biodisponibilidad y toxicidad de los MPs (Zitouni et al., 2021).
Nuestros resultados cuantitativos revelaron además las características de acumulación de MPs en P. vannamei, es decir, los MPs de 2 µm tendieron a acumularse principalmente en el hepatopáncreas, seguido por el intestino, el estómago y las branquias. El tracto digestivo del camarón consta de esófago, estómago, hepatopáncreas e intestino (Sousa y Petriella, 2006). Por lo tanto, la tendencia de acumulación observada sugiere que el sistema digestivo interno, más que las branquias, es el principal órgano objetivo de la acumulación de MPs, lo que se corresponde bien con estudios de laboratorio sobre la acumulación de MPs en bivalvos y peces mediante cuantificación mediante espectroscopia de fluorescencia (Kolandhasamy et al., 2018; Lu et al., 2016; Wang et al., 2021a). Además, estudios de campo recientes han encontrado que tanto el camarón P. vannamei silvestre como el de cultivo muestran un patrón similar de acumulación de MPs mayor en el tracto digestivo que en otros tejidos (Valencia-Castañeda et al., 2022a, 2022b), donde las concentraciones estimadas de MPs en el hepatopáncreas y las branquias fueron altamente comparables a las de nuestro estudio. Esta tendencia también se ha observado en algunos estudios sobre el proceso de ingestión y translocación de MPs en otras especies de camarón mediante microscopio de fluorescencia. Por ejemplo, la administración de MPs fluorescentes de diferentes tamaños mostró que el camarón del Atlántico (Palaemon varians) podía acumular MPs de 9.9 µm en el estómago y el intestino, mientras que los MPs más pequeños (2.1 y 0.1 µm) podrían viajar a través del filtro pilórico y ser transportados a los tejidos internos, como el hepatopáncreas (Saborowski et al., 2022). Se obtuvieron resultados similares en el presente estudio, donde los MPs pequeños expuestos (2 µm) pueden ingerirse y acumularse fácilmente en el estómago y luego expulsarse a través del intestino después del proceso de eliminación. En el tracto digestivo, los MPs
Tabla 1: Evaluación semicuantitativa de cambios histológicos en el hepatopáncreas de P. vannamei expuesto a 0 (control), 0.02 mg L—1 (PS-L), 0.2 mg L—1 (PS-M) y 1.0 mg L—1 (PS- H) MPs.
Observación histológica
Disposición laxa de los túbulos
Estructura tubular degenerada
Citólisis de células epiteliales hepáticas
Aumento del tamaño de las células B
Pérdida de la estructura del lumen en forma de estrella
Control PS-L PS-M PS-H
Nota: (–) ausente; (+) débil; (++) moderado; (+++) fuerte.
sufren degradación biológica y alteración mecánica, que los fragmentan en MPs más pequeños e incluso nanoplásticos (Dawson et al., 2018). Este proceso facilita la translocación de MPs al hepatopáncreas, lo que lo convierte en el órgano más abundante para la acumulación de MPs. Además, los MPs acumulados tienden a retenerse en el hepatopáncreas incluso después del proceso de eliminación. Dado que el hepatopáncreas funciona como el órgano principal para la resorción de nutrientes y la síntesis de enzimas digestivas, los MPs acumulados en sus túbulos podrían inducir respuestas citotóxicas graves (Saborowski et al., 2022). En consecuencia, nuestra data mostró que la agregación de MPs en el hepatopáncreas condujo a la desintegración de sus túbulos adyacentes, causando daño celular y una mayor necrosis de los túbulos en el hepatopáncreas.
Efectos de los MPs sobre la supervivencia y el crecimiento de P. vannamei
En este estudio, la acumulación de MPs no afectó significativamente la supervivencia de P. vannamei en tres concentraciones (de 0.02 a 1.0 mg L-1) después de 8 días de exposición, de acuerdo con investigaciones previas sobre artemia expuesta a una serie de MPs PS de 0.1 µm (de 0.001 a 10 mg L-1) durante 48 h (Gambardella et al., 2017) y PS carboxilado de 40 nm (PS-COOH) MPs hasta 10 mg L-1 durante 14 d (Bergami et al., 2017). Un estudio previo demostró que la tasa de supervivencia de P. vannamei no se vio afectada por exposición a concentraciones más bajas de MPs de polietileno (0.05 y 0.5 mg L-1) durante 48 h, pero se redujo significativamente cuando se expuso a una alta concentración de MPs de polietileno (5.0 mg L-1) (Wang et al., 2021b).
Estos hallazgos indican que las condiciones
de exposición a los MPs (tiempo y concentración), las propiedades de las partículas (tamaño, tipo y forma del polímero) y las diferencias entre especies son factores importantes que contribuyen a las respuestas tóxicas inconsistentes de los organismos (Kögel et al., 2020).
En particular, la acumulación de MPs causó efectos subletales, como efectos fisiológicos, patológicos y bioquímicos, que pueden afectar la supervivencia de P. vannamei después de un período prolongado. Los resultados mostraron que la inhibición del crecimiento de P. vannamei solo se observó después de una exposición a altas concentraciones de MPs (1.0 mg L-1), lo que puede ocurrir en algunas zonas costeras, puertos y estuarios muy contaminados con concentraciones de MPs en la superficie del agua de mar de hasta 3.7 mg. L-1 (Cai et al., 2018; Fu et al., 2020; Kang et al., 2015). Después de la exposición a los MPs, la abundante acumulación de MPs en el sistema digestivo de P. vannamei puede causar bloqueo del tracto gastrointestinal, lesiones mecánicas y funcionales, reduciendo así la alimentación y la absorción de nutrientes, lo que en última instancia conduce a la inhibición del crecimiento de P. vannamei (Cole et al., 2015; Watts et al., 2015). De manera similar, la ingesta de MPs tanto de PS como de PSCOOH causó inhibición del crecimiento de las larvas de camarón mísido en un modo dependiente de la concentración, lo que podría atribuirse a una ingesta reducida de alimentos y una deficiencia de energía en respuesta a la acumulación de MPs en el estómago (Wang et al., 2020). La exposición a MPs de polietileno degradado también inhibió el crecimiento del camarón blanco, y este impacto se intensificó al aumentar la concentración y la duración (Hariharan et al., 2022).
60 - ABRIL 2024
+
– + + +++ – + ++ +++ + ++ +++ +++ – – ++ +++
++ + +++
Efectos de los MPs sobre el comportamiento de natación de P. vannamei
Se demostró que las respuestas del comportamiento de natación a condiciones de exposición tóxica son un indicador sensible en varios invertebrados, incluyendo a los crustáceos (Gambardella et al., 2017). Nuestros resultados mostraron que 8 días de exposición a MPs en todas las concentraciones ejercen un impacto significativo en todo el rendimiento de natación de P. vannamei (por ejemplo, reducción de la velocidad promedio y la distancia total de natación). Los organismos mostraron cambios de comportamiento anormales caracterizados por forma de nadar pegado a la pared y disminución de la actividad exploratoria, particularmente después de una exposición a altas concentraciones de MPs (1.0 mg L-1), lo que probablemente indica un comportamiento contra un ambiente hostil. Varias razones podrían explicar estos fenómenos mencionados. En primer lugar, la adherencia de los MPs a la superficie de las branquias debido a la exposición transmitida por el agua podría impedir la actividad natatoria de los camarones (Bergami et al., 2016). En segundo lugar, la acumulación de MPs en el sistema digestivo
podría reducir la alimentación y la absorción de nutrientes, provocando una deficiencia de energía y, posteriormente, un debilitamiento de la actividad natatoria de los camarones (Wang et al., 2020). En tercer lugar, la acumulación de MPs en la amplia superficie de las branquias puede afectar la actividad de las microvellosidades, inducir lesiones branquiales y estrés respiratorio, lo que puede reducir aún más el consumo de energía para nadar, con el fin de mantener las funciones fisiológicas (Kim et al., 2021). Del mismo modo, se observaron efectos conductuales negativos después de la exposición a MPs en estudios con varios animales acuáticos. Por ejemplo, 96 h de exposición a MPs PS atenuó las actividades de natación (distancia de movimiento y velocidad máxima) y las actividades de exploración de los camarones mísidos (Neomysis japonica) (Wang et al., 2020). De manera similar, se encontró una disminución de la velocidad y el alcance de la natación en pez roca juvenil (Sebastes schlegelii) expuestos a MPs PS, lo que sugiere que los MPs PS afectaron negativamente el desempeño depredador y la competencia de exploración de los peces (Yin et al., 2018). Además, un estudio sobre la carpa cruciana (Carassius carassius) mostró que
los grupos alimentados con nanopartículas de PS exhibieron un mayor comportamiento de agrupamiento, exploraron menos espacio del acuario y mostraron una disminución pronunciada en la actividad durante la alimentación en comparación con los controles (Mattsson et al., 2015). Los estudios previos respaldan el hallazgo de esta investigación de que la exposición a MPs PS tiene un impacto adverso en el rendimiento y el comportamiento de natación de P. vannamei.
Estrés oxidativo y neurotoxicidad inducida por MPs
La superóxido dismutasa proporciona la primera línea de defensa celular contra el estrés oxidativo al convertir los radicales superóxido en peróxido de hidrógeno (Kim et al., 2021). El peróxido de hidrógeno producido puede ser descompuesto aún más por CAT y GPX en compuestos no tóxicos, que ayudan a eliminar las ROS acumuladas y protegen la célula del daño oxidativo (Prokić et al., 2019). Nuestro estudio reveló que la exposición a MPs causó incrementos significativos en las actividades SOD de P. vannamei, acompañado por el aumento de las actividades GPX (Fig. 3A, C). El aumento de
Fig. 5. Alteraciones metabólicas inducidas por diferentes concentraciones de MPs en hepatopáncreas de P. vannamei. (A) Gráfico de puntuaciones de PLS-DA para los perfiles metabólicos del hepatopáncreas de P. vannamei expuesto a 0 (control), 0.02 mg L—1 (PS-L), 0.2 mg L—1 (PS-M) y 1.0 mg L —1 (PS-H) MPs. (B) Mapa de calor para metabolitos diferenciales en el hepatopáncreas de P. vannamei identificados entre los grupos control y de tratamiento. La barra de color representa el contenido relativo de metabolitos según las puntuaciones z. Los asteriscos indican diferencias significativas en los niveles de metabolitos entre los grupos control y de tratamiento (* p < 0.05 y VIP ≥ 1; ** p < 0.01 y VIP ≥ 1). Abreviaturas: TCDA, ácido tauroquenodesoxicólico.
61 - ABRIL 2024 AMBIENTE
la actividad SOD sugirió la sobreproducción de radicales superóxido inducido por los MPs, que fue compensada por el aumento de las actividades de GPX para neutralizar el impacto de los peróxidos hasta cierto punto (Kim et al., 2021).
Estos fenómenos probablemente indican que los organismos comienzan a desarrollar una estrategia defensiva contra el aumento de ROS al mejorar la capacidad antioxidante (Suman et al., 2021). Por el contrario, otra enzima antioxidante, CAT, mostró una tendencia decreciente dependiente de la concentración en P. vannamei después de la exposición a MPs (Fig. 3B). Esto fue respaldado por la correlación inversa significativa observada entre las actividades de SOD y CAT (Tabla S3). El aumento de las actividades SOD provocó la acumulación de peróxido de hidrógeno, que el CAT no eliminó eficientemente, lo que provocó un desequilibrio del sistema de defensa antioxidante (Kim et al., 2021). La inhibición observada de las actividades CAT podría atribuirse a la acumulación de ROS causada por la acción de los MPs, el consumo de energía para la respuesta al estrés oxidativo o daños en la estructura de la enzima inducida
por ROS (Ghelichpour et al., 2019; Yu et al., 2018). Se encontró una relación inversa similar entre las actividades de CAT y SOD en cangrejos Eriocheir sinensis expuestos a MPs de poliestireno (Yu et al., 2018) y en las larvas de peces Cyprinus carpio var expuestos a MPs de cloruro de polivinilo (Xia et al., 2020). Además, el producto tóxico de la peroxidación lipídica (es decir, MDA) mostró una tendencia creciente dependiente de la concentración en P. vannamei después de la exposición a MPs (Fig. 3D). Por lo tanto, interpretamos que el organismo fue incapaz de manejar el estrés oxidativo causado por la exposición a MPs, y posteriormente se produjo un daño oxidativo celular debido a la peroxidación lipídica (Hariharan et al., 2022; Prokić et al., 2019). La actividad AChE se utiliza inusualmente como biomarcador para evaluar la neurotoxicidad (Suman et al., 2021).
Nuestro estudio no observó la inhibición de las actividades AchE en P. vannamei después de la exposición a MPs, por lo que los cambios de comportamiento observados en los organismos expuestos a MPs podrían estar relacionados con otros factores directos o indirectos responsables de posibles
efectos neurotóxicos, como el porcentaje de cerebro en el cuerpo, el contenido de agua y cambios morfológicos en el cerebro (Mattsson et al., 2015; Yin et al., 2019), así como alteraciones metabólicas (Deng et al., 2017). Además, encontramos que varios biomarcadores, incluidos SOD, CAT y MDA, estaban significativamente relacionados con los parámetros de rendimiento en la natación, lo que indica que la actividad y el comportamiento de la natación se vieron afectados negativamente por el estrés oxidativo celular y la peroxidación lipídica inducidos por MPs.
Alteraciones histológicas en el hepatopáncreas inducidas por MPs Como se manifestó anteriormente, se demostró que el hepatopáncreas de P. vannamei acumula la mayoría de los MPs de todos los órganos estudiados; como resultado, los cambios histopatológicos fueron particularmente evidentes en este órgano, como atrofia tubular, disposición laxa de los túbulos, pérdida de la estructura del lumen en forma de estrella y citólisis de las células epiteliales. El grado de daño patológico se exacerbó al aumentar la concentración de MPs (Tabla 1). Estudios
Fig. 6. Análisis de la ruta de metabolitos diferenciales comunes en el hepatopáncreas de P. vannamei inducido por la exposición a MPs. Los compuestos coloreados en rojo indican los metabolitos significativamente alterados detectados en este estudio. Los procesos resaltados en azul claro representan las principales vías metabólicas vinculadas a estos metabolitos alterados. Abreviaturas: TCA, ácido tricarboxílico; TCDA, ácido tauroquenodesoxicólico.
62 - ABRIL 2024 AMBIENTE
previos también han documentado daños histopatológicos importantes en el hepatopáncreas, el hígado o las glándulas digestivas de varios animales acuáticos inducidos por la exposición a MPs. Por ejemplo, la administración intramuscular de MPs de polietileno provocó deformación y rotura del lumen y pérdida de la membrana basal intercelular en el hepatopáncreas de P. vannamei (Hsieh et al., 2021). Se observó inflamación hepatopancreática en cangrejos de río (Procambarus clarkii) después de la exposición dietética a MPs de polietileno (Zhang et al., 2022). También se revelaron respuestas inflamatorias hepáticas en los hepatocitos del pez cebra (Danio rerio) expuesto a MPs PS (Lu et al., 2016). Además, la exposición a MPs de polietileno y tereftalato de polietileno causó anomalías morfológicas evidentes en las glándulas digestivas de las ostras (Crassostrea gigas), incluida la dispersión del daño citoplasmático y la necrosis (Teng et al., 2021).
Estas anomalías histológicas podrían estar relacionadas con el estrés oxidativo inducido por los MPs, que altera los lípidos, las proteínas y el ADN, promueve la necrosis y la apoptosis en los hepatocitos e intensifica la respuesta inflamatoria (Kang et al., 2021; Sánchez-Valle et otros, 2012). Además, cabe señalar que el hepatopáncreas de los crustáceos desempeña un papel clave en la resorción de nutrientes y la síntesis de enzimas digestivas. Por tanto, las alteraciones histopatológicas graves pueden afectar negativamente la función fisiológica normal y el proceso metabólico del organismo.
Cambios metabólicos en el hepatopáncreas inducidos por MPs
La metabolómica proporciona un medio poderoso para detectar biomarcadores moleculares sensibles de toxicidad, identificar las vías metabólicas alteradas y revelar el mecanismo de toxicidad subyacente (Bhagat et al., 2022). Para revelar aún más el mecanismo de toxicidad de los MPs desde los aspectos metabólicos moleculares, se realizó una metabolómica para caracterizar las respuestas metabólicas en el hepatopáncreas de P. vannamei a la exposición a los MPs.
Los análisis estadísticos sugirieron la regulación negativa del gliceraldehído en el hepatopáncreas es inducida por los MPs. El gliceraldehído se genera mediante la descomposición de carbohidratos, que juega un papel clave en la conexión de varias vías metabólicas, particularmente el metabolismo de los carbohidratos. La regulación negativa del gliceraldehído podría atribuirse al incremento de consumo de carbohidratos de P. vannamei para mejorar las necesidades energéticas contra los MPs en condiciones de exposición, que es consistente con estudios previos que reportan el trastorno del metabolismo de los carbohidratos hepáticos en el pez cebra (Danio rerio) tras la exposición a MPs (Sheng et al., 2021; Zhao et al., 2020).
Esta investigación también reveló que los MPs indujeron cambios en los niveles de metabolitos vinculados al metabolismo de los lípidos, como ácidos grasos y acilcarnitinas en el hepatopáncreas de P. vannamei. Los ácidos grasos libres se producen a partir del proceso de lipólisis, que luego se transportan a las mitocondrias para su β-oxidación mediante acilcarnitinas, liberando así energía para las actividades celulares (McCoin et al., 2015). Las alteraciones de estos

63 - ABRIL 2024
AMBIENTE
metabolitos indican que los MPs pueden alterar el metabolismo de los lípidos en el hepatopáncreas de P. vannamei. De manera similar, se han demostrado cambios de metabolitos relacionados con el metabolismo de los lípidos en camarones (P. vannamei), pez cebra (Danio rerio) y ostras (Crassostrea gigas) expuestos a MPs (Duan et al., 2021; Lu et al., 2016; Teng et al., 2021).
El metabolismo de los lípidos hepáticos es fundamental para la absorción de nutrientes y la producción de energía. Por lo tanto, las anomalías del metabolismo de los lípidos observadas en el estudio actual probablemente indican que la exposición a los MPs provocó un cambio en la estrategia del metabolismo energético de P. vannamei, que ayuda a mejorar la capacidad del organismo para manejar el estrés. Se demostró que, para satisfacer la creciente energía en condiciones estresantes como la exposición a MPs, los organismos podrían adaptarse presentando compensaciones energéticas entre diversas actividades conductuales y fisiológicas (Sussarellu et al., 2016). Este fenómeno también se mostró en medaka marina (Oryzias melastigma) y ostra (Crassostrea gigas) después de la exposición a MPs (Feng et al., 2021; Teng et al., 2021). Además, este estudio observó que los MPs aumentaron significativamente los niveles de varios aminoácidos y dipéptidos en el hepatopáncreas de P. vannamei, lo que indica una alteración en el aminoácido metabolismo.
Este fenómeno también se ha demostrado en el camarón (P. vannamei), pez cebra (Danio rerio) y ostra (Crassostrea gigas) expuestos a MPs (Duan et al., 2021; Qiao et al., 2019; Teng et al., 2021). Los metabolitos de los aminoácidos no solo pueden regular el metabolismo energético, sino que también afectan la función del sistema inmunológico. La taurina, un abundante aminoácido intracelular libre presente en casi todos los tejidos animales, participa activamente en muchas funciones biológicas y fisiológicas (Surai et al., 2021).
Muchos estudios han revelado con frecuencia que la taurina puede actuar como un agente protector contra los impactos nocivos del estrés oxidativo causado por sustancias tóxicas (Naddafi et al., 2022; Surai et al.,
2021). La pronunciada regulación positiva de la taurina hepática en P. vannamei después de la exposición a MPs puede reflejar un intento del organismo de mejorar la capacidad antioxidante de las células para hacer frente a las condiciones de estrés.
El ácido L-aspártico, otro aminoácido importante, funciona como intermediario del ciclo del ácido tricarboxílico (TCA) y del ciclo de la urea y desempeña múltiples funciones en el metabolismo y la función de los leucocitos (Gong et al., 2020). Se ha reportado que el ácido L-aspártico podría aliviar el daño oxidativo y la respuesta inflamatoria en el hepatopáncreas, aumentando así la inmunidad de los peces y mejorando la supervivencia contra la infección bacteriana mediante la fagocitosis inducida por óxido nítrico (Gong et al., 2020; Zhao et al., 2022).
En consecuencia, nuestro estudio encontró que la exposición a MPs causó una mayor producción de ácido L-aspártico en P. vannamei, lo que probablemente implica la promoción de las respuestas inmunes de los organismos contra factores estresantes exógenos. Además, la taurina y el ácido L-aspártico también actúan como neurotransmisores, por lo que la regulación positiva de estos dos metabolitos podría implicar posibles efectos neurotóxicos relacionados con los cambios de comportamiento observados en P. vannamei después de la exposición a MPs (Deng et al., 2017). En resumen, el estudio metabolómico demostró que la exposición a MPs podría alterar las vías de glucólisis, lipólisis y metabolismo de aminoácidos en el hepatopáncreas de P. vannamei
Conclusiones
Caracterizamos los efectos tóxicos de la acumulación de MPs en P. vannamei desde los aspectos de comportamiento, histología tisular, actividad enzimática y metaboloma. La acumulación y distribución de MPs se demostró en múltiples órganos, siendo el hepatopáncreas el que contiene la mayor cantidad de MPs. La acumulación de MPs causó una variedad de efectos subletales en P. vannamei, incluyendo la inhibición del crecimiento, cambios en el comportamiento de natación, estrés oxidativo y daño hepatopancreático. Además, el análisis metabolómico reveló que los MPs interfirieron
con el metabolismo hepatopancreático de P. vannamei incluso a un nivel de exposición muy bajo (0.02 mg L-1), lo que ofreció una nueva perspectiva para delinear los modos de acción tóxicos de los MPs.
Nuestro estudio proporciona data de referencia para futuras evaluaciones de los impactos y efectos nocivos de MPs en crustáceos, lo que tiene implicaciones notables para avanzar en nuestro conocimiento de los mecanismos tóxicos de los MPs en los crustáceos. Los resultados resaltan aún más el riesgo ecológico potencial de la contaminación por MPs para las poblaciones y la función ecológica de los crustáceos marinos.
Importante:
Conforme lo indica el presente artículo, los microplásticos en camarón, se alojan en el tracto digestivo especialmente en el hepatopáncreas, órgano donde se manifiestan gran parte de las enfermedades, por lo que la presencia de micro o nano plásticos que pudieran estar presentes en el hepatopáncreas, tendería a ser confundido con otra patología.
Los micro y nano plásticos se los detecta y evalúa a través de la espectroscopia FTIR, la misma que utiliza energía infrarroja, modulada para analizar una muestra. Este tipo de equipos no se encuentra disponible en el país, España tiene disponible esta tecnología.
Los mecanismos para filtrar el agua para eliminar microplásticos hasta la presente, han sido diseñados para bajos volúmenes de agua:
Filtración mecánica: utiliza medios como arena, cerámica o fibras para atrapar partículas basándose en el tamaño. Filtración por ósmosis inversa: emplea una membrana semipermeable para filtrar una amplia gama de contaminantes, incluidos los micro plásticos, sin embargo, el uso de plantas de osmosis inversa es oneroso y no se podría instalar en una camaronera debido al volumen de agua que maneja. Esta tecnología puede ser mejor aplicada en los laboratorios de producción larvaria•
Para mayor información escriba a: zengyx@hntou.edu.cn
64 - ABRIL 2024

Aplicación de la secuenciación de nueva generación en el análisis del microbioma asociado a la producción camaronera
Autores:
César Bedoya Pilozo1, 2
Joselyn Briceño1
Luis Felipe Urresta1
1Biosequence
2Laboratorio de Biología Molecular, Hospital
Luis Vernaza cesariotito@yahoo.com

Las circunstancias actuales en la producción de camarones han obligado a la optimización de los recursos para mejorar la rentabilidad del negocio. Uno de los aspectos que ofrece un campo amplio de mejoramiento es el manejo de los factores abióticos y bióticos asociados a la acuicultura del camarón. En este ámbito, uno de los factores posiblemente más influyente pero menos estudiado es el microbioma presente tanto en el animal como en el agua y el suelo de las zonas de producción. El microbioma es la suma de todos los organismos unicelulares, de linaje procarionte de manera principal, que interactúa con el camarón y su entorno. Es un factor clave en los procesos de digestión, asimilación de los nutrientes, en la degradación de la materia orgánica tanto en la columna de agua como en el fondo de la piscina, en los procesos de competencia microbiana y al parecer, tendría también un papel en la respuesta inmune del huésped. En recientes años se han publicado varios estudios que han demostrado cómo la composición del microbioma del camarón está constituida por diferentes grupos taxonómicos que varían en proporción a lo largo de las diferentes fases de cultivo. Una de las principales herramientas que ha permitido este aumento en las investigaciones sobre el microbioma en la acuicultura de camarones es el uso de la tecnología de secuenciamiento masivo. La madurez de esta tecnología permite hoy en día, a costos bajos y asequibles, analizar muestras biológicas y ambientales en cuestión de días o semanas para medir la población microbiana presente, posibilitando un seguimiento continuo de sus variaciones y las potenciales consecuencias sobre el animal y el entorno. El propósito de esta revisión es presentar estos avances y el impacto que podrían tener a futuro en la acuicultura.
La producción de camarones desempeña un papel importante en la economía de varios países; así también, en la demanda de productos acuícolas y recursos marinos naturales. Actualmente, la generación de una producción acuícola sostenible se ha convertido en un objetivo prioritario para la industria. Es por esto que la optimización de esta área productiva se basa en la comprensión del sistema de interacción biológica entre los organismos y el medio en
- ABRIL 2024
66
el que desarrollan (Anderson et al., s/f).
El microbioma es una parte esencial de los ecosistemas productivos. Dentro del ambiente acuífero, desempeña una variedad de funciones críticas que impactan en la salud, crecimiento y resistencia a enfermedades de los camarones. Se encarga de estimular el desarrollo gastrointestinal, producir vitaminas, mejorar la absorción de nutrientes, proporcionar actividades enzimáticas adicionales, fortalecer el sistema inmunitario, mantener la tolerancia de las mucosas, proveer de resistencia frente a las enfermedades infecciosas y producir compuestos antiinflamatorios. Además, cada vez se reconoce a las epidemias o a la presencia de microorganismos patógenos como una de las limitaciones más importantes para el cultivo de camarón. (Chaudhari et al., 2023)
A menudo, el microbioma intestinal es considerado como un órgano extra implicado en varios procesos de homeostasis energética, y puede estar influenciado por factores intrínsecos del huésped, como dieta, condiciones y entorno del cultivo, densidad de población, salud del hospedero y la comunidad microbiana intestinal central de la especie, que se traducen en la gestión de diversas actividades fisiológicas de los peces y mariscos. A su vez, los probióticos son microorganismos vivos que desempeñan un papel importante en el equilibrio del microbioma intestinal de los organismos huéspedes. Estos pueden reducir los microbios patógenos, estimular la tasa de crecimiento y mejorar la salud de los animales.
El uso de probióticos ha sido una alternativa sostenible y amigable con el ambiente acuícola, reduciendo la aplicación de antibióticos y aumentando la eficiencia de la producción; aun así, la información de la interacción huésped – microbioma del camarón es escasa. Teniendo en cuenta el importante papel que este juega en toda la vida de los peces y mariscos, es necesario comprender los procesos relacionados entre el huésped y la comunidad microbiana. Esta discusión ha surgido con gran interés en la investigación acuícola, sobre todo para garantizar la salud y el bienestar de los camarones en cultivo y la seguridad alimentaria en su consumo.
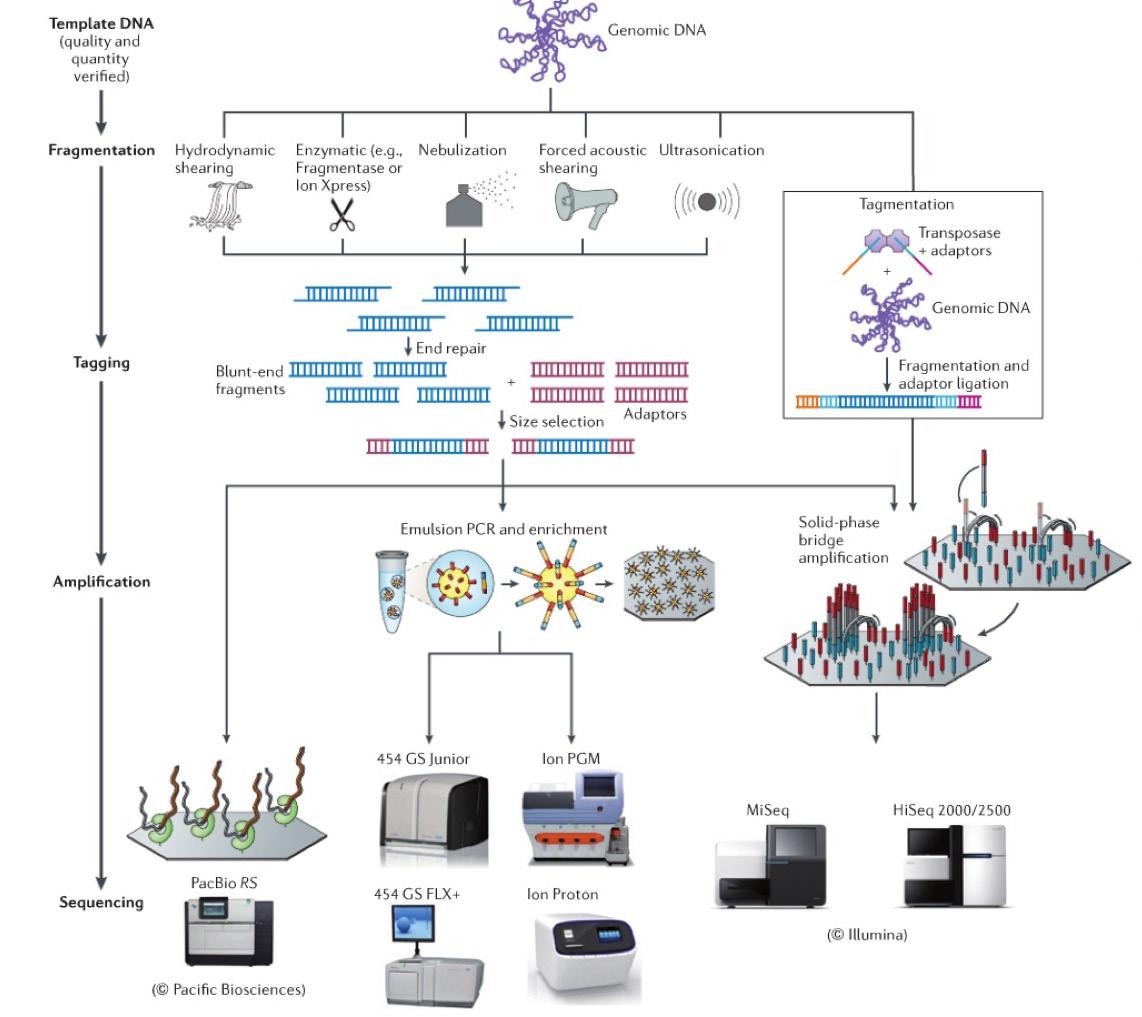
La tecnología de NGS (Next Generation Sequencing, secuenciamiento de nueva generación) se utiliza para investigar la diversidad taxonómica y funcional de los microbiomas en una amplia gama de entornos, incluidos el suelo, el agua, el tracto digestivo de los animales, entre otros. Estas técnicas permiten identificar microorganismos presentes en muestras ambientales o biológicas, caracterizar su abundancia relativa y estudiar sus funciones metabólicas y genéticas. Dentro del NGS encontramos la metagenómica, que es el análisis del material genético extraído de una muestra sin la necesidad de cultivar los microorganismos individualmente. Esto permite analizar la totalidad del material genético presente en la comunidad microbiana, y facilita la exploración de la diversidad genética y funcional de las comunidades microbianas en una escala sin precedentes.
De igual forma, el NGS ayuda a identificar especies microbianas raras o poco conocidas que pueden tener un papel importante en la función y la estabilidad de los ecosistemas. Estas aproximaciones
proporcionan información valiosa sobre la diversidad genética de los microorganismos en su entorno natural.
En la presente revisión, se han realizado esfuerzos para recopilar información sobre diversos aspectos y usos del estudio de microbiomas utilizando la tecnología de nueva generación y la interacción huéspedmicrobioma, en particular sobre el sistema inmunitario y el mantenimiento de la salud, el tratamiento de enfermedades, la absorción de nutrientes, la digestión y absorción, la expresión génica y el metabolismo en peces y mariscos.
Estado actual de la acuicultura de camarones
Penaeus vannamei, también conocido como camarón blanco del Pacífico, camarón patiblanco o langostino blanco, es nativo de la costa del Pacífico, desde el noreste de Perú hasta el norte de México. En Ecuador, desde el 2003 ha tenido un crecimiento constante de exportación, alcanzando los 816,466 t aproximadamente hasta el 2023, llegando a convertirse en una industria de más de 6 billones de dólares. El desarrollo
- ABRIL 2024 67 TECNOLOGÍA
Figura 1: Tecnologías de secuenciamiento masivo
TECNOLOGÍA
de la línea de reproducción tuvo un efecto positivo en la mejora del rendimiento del camarón y el control de enfermedades, pero la tendencia mundial de la acuicultura de una sola especie aumentó el potencial de aparición de enfermedades específicas y la translocación de patógenos peligrosos existentes a nuevas ubicaciones geográficas (Anderson et al., s/f).
Para esto, han existido varios desafíos que el sistema productivo ha tenido que enfrentar, como la demanda mundial en aumento, ya sea por preferencia de productos del mar, crecimiento de población o consumo de proteína, generando una intensificación significativa en la producción a gran escala y la adopción de tecnologías avanzadas.
El rápido crecimiento y expansión de la industria acuícola del camarón estuvo acompañado del brote de varias enfermedades, que causan pérdidas significativas en la producción acuícola y el comercio internacional y, en consecuencia, impiden el desarrollo económico de la zona costera. Así mismo, existen desafíos sanitarios como la aparición de enfermedades virales y bacterianas, así como los problemas ambientales generados por actividades antropogénicas, como la degradación de los ecosistemas costeros, contaminación de aguas, deforestación de manglares y más. La industria se enfrenta al desafío de adoptar prácticas sostenibles para mitigar su impacto ambiental. Aunque es difícil reunir datos económicos precisos
sobre el efecto de las enfermedades en la producción de camarones, se estima que las pérdidas anuales serían de alrededor del 40%. Las consecuencias negativas del brote de la enfermedad del camarón y el comercio internacional irrestricto de camarones han aumentado la conciencia mundial sobre la necesidad de normas de gestión.
El impacto económico de las enfermedades se traduce en pérdidas en la producción, ingresos, escasez de alimento, cierre de negocios, inversión y confianza del consumidor. Son muchos los esfuerzos que se realizan para el control eficaz de la producción camaronera por lo delicado de sus ecosistemas. Estos están sujetos a regulaciones cada vez más estrictas relacionadas con la calidad del agua, el uso de antibióticos, la trazabilidad del producto y el bienestar animal. La comprensión de las tendencias y los desafíos actuales es fundamental para el desarrollo futuro sostenible de la acuicultura de camarones a nivel mundial.
Importancia de los microbiomas en la acuicultura de camarones
Los microbiomas son comunidades microbianas complejas que colonizan el cuerpo y los tejidos de los camarones, como también el entorno acuático en el que habitan. Este juega un papel crucial en la salud y el bienestar de los camarones al contribuir a la digestión de alimentos, la síntesis de vitaminas, la competencia con patógenos y la regulación del sistema inmunológico.
Además, nuestros hallazgos demostraron que los taxones clave suelen desempeñar un papel importante en el mantenimiento de la estabilidad de la comunidad microbiana, lo que implica que también podrían ser cruciales para mejorar la resistencia a la colonización de los camarones (Das et al., 2023).
El microbioma acuático desempeña un rol importante en la calidad del agua al influir en la descomposición de nutrientes, la mineralización de compuestos orgánicos, fijación de nitrógeno, mineralización de fósforo y la remoción de contaminantes. Dentro del ambiente, son indicadores biológicos de la calidad de agua y sostenibilidad de los sistemas acuícolas. Los desequilibrios en el microbioma pueden aumentar la susceptibilidad a enfermedades y afectar negativamente el rendimiento de los camarones (Secuenciación de microbiomas para el manejo adaptativo de la producción en estanques, s/f).
El microbioma de los camarones está compuesto por una variedad de bacterias, virus, hongos y otros microorganismos. Puede variar dependiendo de la especie y ambiente que se analice, aunque se desconocen en gran medida los taxones intestinales específicos que podrían conferir resistencia contra los patógenos en los camarones (Yun et al., 2022).
Las bacterias son los microorganismos dominantes en el microbioma de los camarones, representando una gran
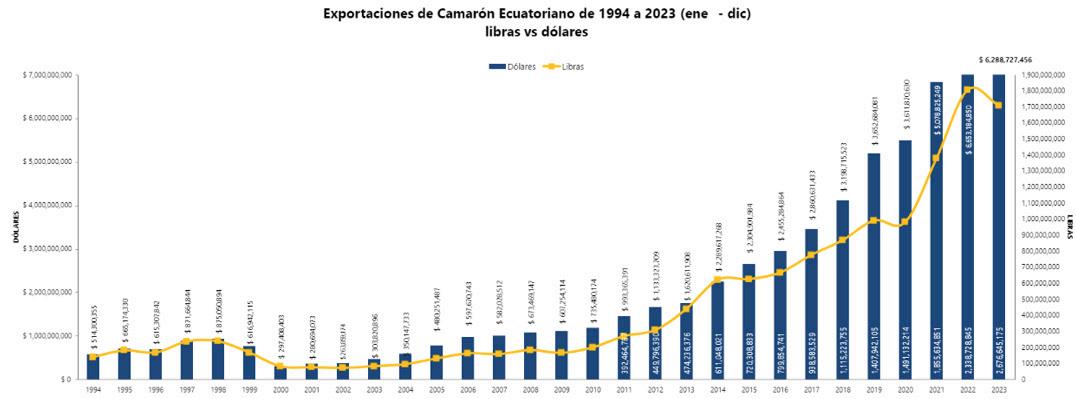
- ABRIL 2024 68
Figura 2: Exportaciones de camarón ecuatoriano en el período 1993 – 2023 (Estadísticas - Cámara Nacional de Acuacultura, s/f)
proporción de la diversidad microbiana total. Su diversidad puede ser extremadamente dinámica con miles de especies diferentes en un solo individuo. Entre estos individuos se encuentran miembros de los filos: Proteobacteria, Firmicutes, Bacteroidetes, Actinobacteria, entre otros (CornejoGranados et al., 2017a).
Los géneros Vibrio, Pseudomonas, Photobacterium y Acinetobacter se encuentran con frecuencia en el hepatopáncreas y el intestino de los camarones, lo que indica su importante papel en estos ambientes. Familias como Enterobacteriaceae, Vibrionaceae, Pseudoalteromonadaceae, Moraxellaceae, Pseudomonadaceae y Rhodobacteraceae se encuentran entre las más abundantes en varias muestras de camarones, lo que sugiere una amplia presencia en sus microbiomas (Z. Huang et al., 2016; Zeng et al., 2017). Además de estos, también prevalecen géneros como Spongiimonas, Candidatus Bacilloplasma, Shewanella, Synechococcus, Aeromonas, Rhodobacter, Propionigenium y Fusibacter, con sus abundancias relativas que varían bajo diferentes condiciones de cultivo de salinidad (Hou et al., 2020).
Algunas bacterias son consideradas beneficiosas para los camarones, ya que pueden estar involucradas en la digestión de alimentos, la síntesis de nutrientes esenciales, la protección contra patógenos y la regulación del sistema inmunológico. Sin embargo, existen otros que pueden resultar altamente patógenos y causar la muerte del individuo.
Las interacciones microbianas dentro de los camarones pueden involucrar competencia por nutrientes, cooperación en la degradación de sustratos, intercambio de metabolitos y comunicación celular. Estas pueden influir en la estabilidad y resistencia del microbioma a cambios ambientales e invasión de patógenos. Estas interacciones son un área de investigación activa y en constante evolución que tiene importantes implicaciones para la salud, el rendimiento y la sostenibilidad de la producción acuícola.
El microbioma desempeña funciones importantes en la digestión, la absorción de nutrientes, la protección contra patógenos
y la regulación del sistema inmunológico de los camarones. Algunas bacterias del microbioma pueden producir enzimas que ayudan en la digestión de compuestos complejos presentes en la dieta de los camarones, como la celulosa y la quitina (Liang et al., 2020).
Un microbioma equilibrado y diverso puede contribuir a la salud y la resistencia a enfermedades de los camarones. Algunas bacterias del microbioma pueden competir con patógenos por espacio y recursos, inhibiendo su crecimiento y reduciendo el riesgo de infecciones. En la actualidad, los probióticos se ven favorecidos por encima de estas sustancias antimicrobianas debido a su capacidad para estimular la resistencia a las enfermedades en las granjas camaroneras mediante el fortalecimiento del sistema inmunitario de forma natural. Se ha demostrado que algunas cepas probióticas, como Bacillus spp, Lactobacillus y Pediococcus pentosaceus mejoran la inmunidad innata de los camarones contra enfermedades como el síndrome de la Mancha Blanca (WSSV), reducen la carga viral, aumentan la digestibilidad y el crecimiento, y apoyan el microbioma intestinal del huésped. En múltiples investigaciones de acuicultura se ha destacado el uso generalizado de probióticos, debido a su capacidad para mejorar la calidad del agua, liberar nutrientes y enzimas digestivas, aumentar las defensas y la inmunidad del huésped y disminuir la incidencia de enfermedades. Exige que la investigación se lleve a cabo a gran escala, así como la creación de instrumentos de seguimiento adecuados y una producción estrictamente regulada (Kesselring et al., 2022; Wang et al., 2020).
Se ha descubierto que los microbios beneficiosos como Lactobacillus pentosus, Pseudomonas putida, Pseudomonas synxantha, Rodopseudomonas palustris y Streptococcus thermophiles aumentan en abundancia con la introducción de agavina, una fibra dietética, en las dietas de los camarones, lo que indica el potencial de las intervenciones dietéticas para modular la microbiota de los camarones (Ochoa-Romo et al., 2022).
Se encontró que los camarones sanos tienen un enriquecimiento de Faecalibacterium prausnitzii y Pantoea agglomerans, lo que
Bibliografía
Alfiansah, Y. R., Peters, S., Harder, J., Hassenrück, C., & Gärdes, A. (2020). Structure and cooccurrence patterns of bacterial communities associated with white faeces disease outbreaks in Pacific white-leg shrimp Penaeus vannamei aquaculture. Scientific Reports, 10(1). https:// doi.org/10.1038/s41598-020-68891-6
Anderson, J., Valderrama, D., & Jory, D. (s/f). GOAL 2019: Revisión de la producción mundial de camarones
Chang, Y.-T., Ko, H.-T., Wu, P.-L., Kumar, R., Wang, H.C., & Lu, H.-P. (2023). Gut microbiota of Pacific white shrimp ( Litopenaeus vannamei ) exhibits distinct responses to pathogenic and non-pathogenic Vibrio parahaemolyticus . Microbiology Spectrum, 11(5). https://doi.org/10.1128/spectrum.01180-23
Chaudhari, H. G., Prajapati, S., Wardah, Z. H., Raol, G., Prajapati, V., Patel, R., Shati, A. A., Alfaifi, M. Y., Elbehairi, S. E. I., & Sayyed, R. Z. (2023). Decoding the microbial universe with metagenomics: a brief insight. Frontiers in Genetics, 14, 1119740. https://doi.org/10.3389/fgene.2023.1119740
Chen, W. Y., Ng, T. H., Wu, J. H., Chen, J. W., & Wang, H. C. (2017). Microbiome Dynamics in a Shrimp Grow-out Pond with Possible Outbreak of Acute Hepatopancreatic Necrosis Disease. Scientific Reports, 7(1). https:// doi.org/10.1038/s41598-017-09923-6
Cicala, F., Lago-Lestón, A., Gomez-Gil, B., GollasGalván, T., Chong-Robles, J., Cortés-Jacinto, E., & Martínez-Porchas, M. (2020). Gut microbiota shifts in the giant tiger shrimp, Penaeus monodon, during the postlarvae, juvenile, and adult stages. Aquaculture International, 28(4), 1421–1433. https://doi.org/10.1007/s10499-020-00532-1
Cornejo-Granados, F., Lopez-Zavala, A. A., GallardoBecerra, L., Mendoza-Vargas, A., Sánchez, F., Vichido, R., Brieba, L. G., Viana, M. T., Sotelo-Mundo, R. R., & Ochoa-Leyva, A. (2017a). Microbiome of Pacific Whiteleg shrimp reveals differential bacterial community composition between Wild, Aquacultured and AHPND/EMS outbreak conditions. Scientific Reports 2017 7:1, 7(1), 1–15. https://doi.org/10.1038/s41598-017-11805-w Cornejo-Granados, F., Lopez-Zavala, A. A., GallardoBecerra, L., Mendoza-Vargas, A., Sánchez, F., Vichido, R., Brieba, L. G., Viana, M. T., SoteloMundo, R. R., & Ochoa-Leyva, A. (2017b). Microbiome of Pacific Whiteleg shrimp reveals differential bacterial community composition between Wild, Aquacultured and AHPND/EMS outbreak conditions. Scientific Reports, 7(1). https://doi.org/10.1038/s41598-017-11805-w Das, S. P., Abidin, Z., Huang, H. T., Lin, Y. R., Huang, C. Y., Wu, Y. S., Hu, Y. F., & Nan, F. H. (2023). Deciphering the influence of dietary synbiotics in white shrimp gut and its effects in regulating immune signaling pathways. Frontiers in Marine Science, 10, 1342708. https://doi. org/10.3389/FMARS.2023.1342708/BIBTEX Ding, Y., Song, X., Cao, X., He, L., Liu, S., & Yu, Z. (2021). Healthier communities of phytoplankton and bacteria achieved via the application of modified clay in shrimp aquaculture ponds. International Journal of Environmental Research and Public Health, 18(21). https://doi.org/10.3390/ijerph182111569 Estadísticas - Cámara Nacional de Acuacultura (s/f). Recuperado el 30 de marzo de 2024, de https://www.cna-ecuador.com/estadisticas/ García-López, R., Cornejo-Granados, F., LopezZavala, A. A., Cota-Huízar, A., Sotelo-Mundo, R. R., Gómez-Gil, B., & Ochoa-Leyva, A. (2021). OTUs
- ABRIL 2024 69 TECNOLOGÍA
TECNOLOGÍA
puede ser indicativo de un estado beneficioso del microbioma (Cornejo-Granados et al., 2017a).
Por el contrario, los camarones enfermos mostraron un enriquecimiento de Aeromonas taiwanensis, Simiduia agarivorans y Photobacterium angustum, lo que sugiere que estas especies podrían estar asociadas con enfermedades (Cornejo-Granados et al., 2017a).
La composición de la comunidad bacteriana en los intestinos de los camarones varía en condiciones de cultivo de baja salinidad (LS) y alta salinidad (HS), con condiciones de HS que favorecen a especies patógenas más oportunistas, como varias especies de Vibrio (Hou et al., 2020).
Las abundancias relativas de ciertos géneros como Vibrio, Propionigenium, Spongiimonas y Synechococcus estaban sobrerrepresentadas en condiciones de HS, mientras que otras como Roseomonas, Rhodobacter, Aeromonas, Pseudomonas, Snowella y Fusibacter eran mayores en condiciones de LS.
Cambios en las condiciones ambientales, como fluctuaciones en la temperatura del agua, salinidad y contaminación, pueden alterar la composición y la función del microbioma de los camarones. Estudios sugieren que un microbioma saludable puede mejorar la capacidad de los camarones para resistir y recuperarse del estrés ambiental (F. Huang et al., 2018).
Avances tecnológicos en el análisis del microbioma
Las técnicas de secuenciación de nueva generación (NGS) han transformado nuestra comprensión de los microbiomas, al proporcionar una visión detallada de la composición, la diversidad y la función de las comunidades microbianas en una amplia variedad de entornos. Es importante conocer que las muestras se analizan de manera individual dentro del estudio general de microbiomas. A partir de esto, es posible realizar estudios comparativos utilizando variables de dieta, entorno, fase de crecimiento, salud, entre otros. Para el proceso de obtención de material genético, se utilizan kits comerciales especializados en obtención de ADN de alta calidad de manera
eficiente, sin contaminantes ni inhibidores que podrían afectar las reacciones para la secuenciación. La calidad de ADN es un factor determinante del éxito de la secuenciación; por lo cual, es importante utilizar kits que garanticen la integridad y pureza del material genético (Robinson & Chin, 2023).
La secuenciación metagenómica de amplicones ha sido ampliamente adoptada en la investigación contemporánea. En esta técnica se emplean regiones específicas del ADN ribosomal (ARNr), como el gen 16S en bacterias y el gen ITS en hongos. Se realiza utilizando la amplificación de las regiones
de interés mediante la reacción en cadena de la polimerasa (PCR). El amplicón V3V4 de la región 16S dirigido al microbioma del camarón ha sido ampliamente utilizado debido a un mayor número de unidades taxonómicas operativas (OTU); esto conlleva a un mayor reconocimiento taxonómico. Estos amplicones generados se someten a secuenciación de nueva generación utilizando a plataforma Illumina MiSeq (Rieder et al., 2023).
Un papel crucial en el análisis biológico es la bioinformática, que permite la interpretación de los datos secuenciados. Esto incluye el filtrado de secuencias de

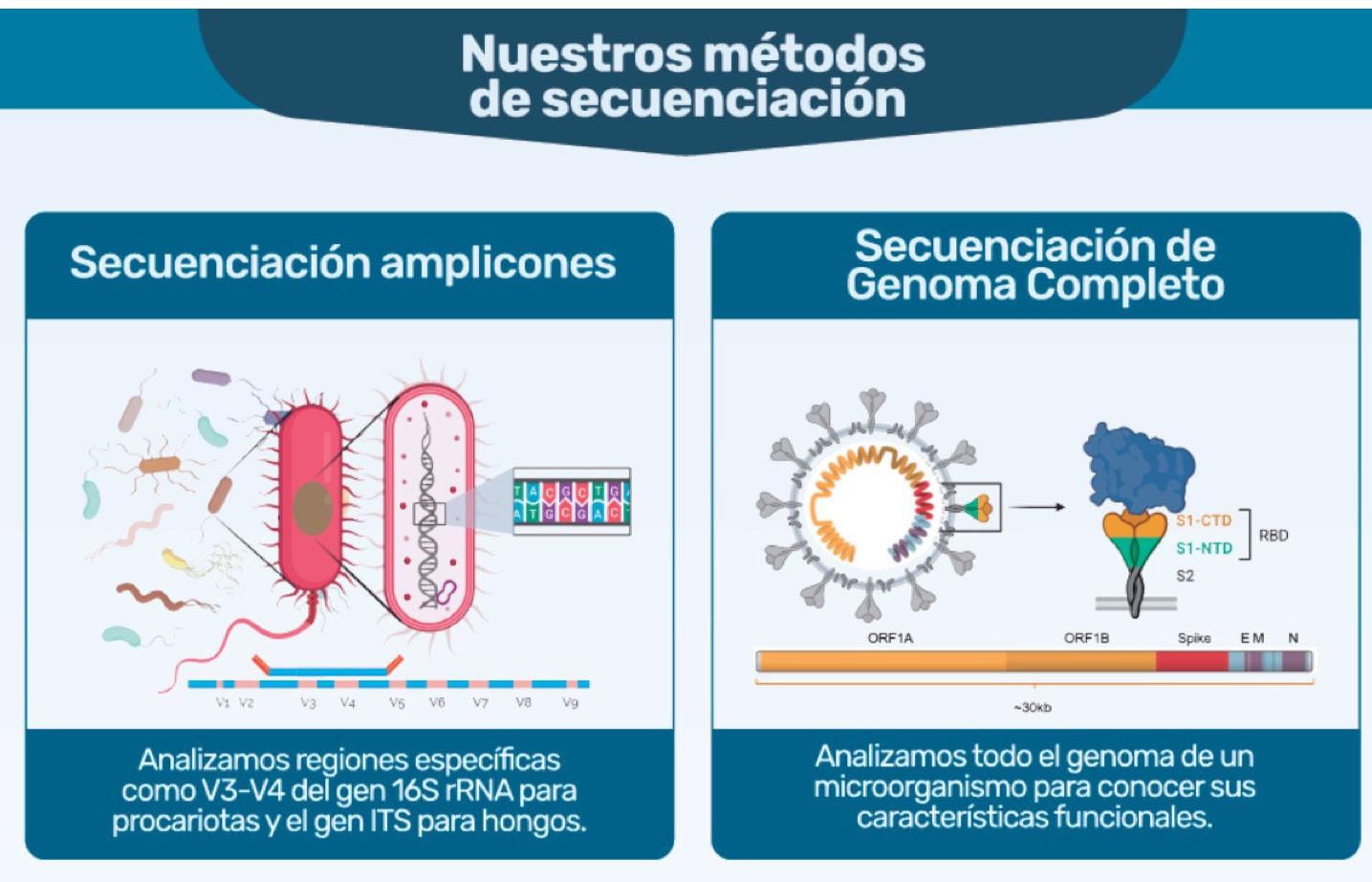
- ABRIL 2024 70
Figura 3: Aplicación de la tecnología NGS en el microbioma del camarón de cultivo
Figura 4: Métodos de secuenciamiento masivo disponible en acuicultura
baja calidad, la agrupación de secuencias en unidades taxonómicas operativas (OTUs) y la asignación taxonómica utilizando bases de datos de referencia como SILVA, UNITE y Greengenes junto con el algoritmo DADA2.
Se necesitan flujos de trabajo bioinformáticos especializados para procesar los datos de secuenciación, que a menudo implican la agrupación de secuencias en unidades taxonómicas operativas (OTU) o variantes de secuencias de amplicones (ASV) (GarcíaLópez et al., 2021), (García-López et al., 2020).
El análisis del microbioma con secuenciación profunda es una herramienta transformadora en la acuicultura de camarones, ya que ofrece una vía para mejorar la salud de los animales, gestionar los sistemas de producción y abordar las preocupaciones ambientales. Al comprender y manipular la microbiota intestinal, los productores pueden mejorar el crecimiento y la inmunidad de los camarones, mientras que un enfoque holístico de la gestión del microbioma puede conducir a prácticas acuícolas más sostenibles y ecoeficientes (García-López et al., 2021).
La integración de herramientas bioinformáticas y análisis estadístico permite la comparación y caracterización de la composición y la estructura del microbioma de los camarones. Mediante estas herramientas es posible la identificación de patrones y asociaciones significativas entre la comunidad microbiana, factores ambientales y el huésped.
Manejo del microbioma
Se debe especificar que las estrategias de manejo nutricional, sanitario y ambiental pueden influir en la composición y la salud del microbioma de los camarones (Zeng et al., 2017).
La selección de dietas balanceadas, el uso de probióticos y prebióticos, así como la mejora de las condiciones de cultivo pueden promover un microbioma saludable y mejorar la producción de camarones. Se ha convertido en una práctica popular el control de la aparición de enfermedades mediante la adición de probióticos en la acuicultura. Sin embargo, las funciones de muchos probióticos comerciales se han visto
limitadas porque generalmente no son las especies dominantes en el sistema de cultivo de camarones y son difíciles de adaptar al entorno acuícola (Chen et al., 2017).
El estudio de microbiomas implica el uso de varios métodos para el análisis de la composición, estructura y comparación de las comunidades microbianas. Estas tienen el objetivo de comprender mejor los diferentes sistemas (Chen et al., 2017).
Cada uno de los sistemas de cultivo tiene su propia composición microbiana influenciada principalmente por factores ambientales y su manejo. Entre estos sistemas se incluyen: estanques, jaulas, sistemas de recirculación y otros. Se han demostrado variaciones en las diferentes etapas del cultivo, siendo afectadas por factores físicos, químicos y biológicos. Por ejemplo, en los sistemas de recirculación pueden presentar microbiomas más estables y homogéneos debido al control riguroso de las condiciones ambientales. En los estanques de cultivo pueden mostrar una mayor variabilidad en la composición microbiana debido a la influencia de factores ambientales fluctuantes y prácticas de manejo menos controladas (Trung Tran et al., 2019).
La comprensión de cómo los factores ambientales y de manejo influyen en la diversidad y estabilidad de los microbiomas de camarones es crucial para optimizar las prácticas de manejo y promover la salud y el rendimiento de los camarones en diferentes sistemas de producción acuícola. Los avances en tecnologías de secuenciación de nueva generación y análisis bioinformáticos han permitido estudios más detallados sobre la dinámica de los microbiomas en diversos ambientes acuáticos, lo que contribuye a un mejor manejo y control de la producción de camarones.
El estudio de microbiomas en camarones tiene diversas aplicaciones que son fundamentales para comprender y mejorar la producción acuícola. Aquí algunas de ellas:
Salud animal
El análisis de microbiomas permite identificar comunidades microbianas asociadas con la salud de los camarones. Detectar desequilibrios en el microbioma externo o intestinal puede ayudar a prevenir
and ASVs Produce Comparable Taxonomic and Diversity from Shrimp Microbiota 16S Profiles Using Tailored Abundance Filters. Genes, 12(4), 564. https://doi.org/10.3390/genes12040564
García-López, R., Cornejo-Granados, F., LopezZavala, A. A., Sánchez-López, F., Cota-Huízar, A., Sotelo-Mundo, R. R., Guerrero, A., MendozaVargas, A., Gómez-Gil, B., & Ochoa-Leyva, A. (2020). Doing more with less: A comparison of 16S hypervariable regions in search of defining the shrimp microbiota. Microorganisms, 8(1). https://doi.org/10.3390/microorganisms8010134
Garibay-Valdez, E., Martínez-Córdova, L. R., Vargas-Albores, F., Gollas-Galván, T., LagoLeston, A., Calderón, K., & Martínez-Porchas, M. (2019). Biofilm consumption shapes the intestinal microbiota of shrimp (Penaeus vannamei). Aquaculture Nutrition, 25(2), 427–435. https://doi.org/10.1111/anu.12868
Guo, H., Huang, L., Hu, S., Chen, C., Huang, X., Liu, W., Wang, S., Zhu, Y., Zhao, Y., & Zhang, D. (2020). Effects of Carbon/Nitrogen Ratio on Growth, Intestinal Microbiota and Metabolome of Shrimp (Litopenaeus vannamei). Frontiers in Microbiology, 11. https://doi.org/10.3389/fmicb.2020.00652
Hembrom, P. S., Barik, S., Deepthi, M., Kannoth, S., & Grace, T. (2024). Influence of gut microbiome on health and development of penaeid shrimps. En Aquatic Sciences (Vol. 86, Número 1). Springer Science and Business Media Deutschland GmbH. https://doi.org/10.1007/s00027-023-01018-x Holt, C. C., Bass, D., Stentiford, G. D., & van der Giezen, M. (2021). Understanding the role of the shrimp gut microbiome in health and disease. Journal of Invertebrate Pathology, 186 https://doi.org/10.1016/j.jip.2020.107387
Hou, D., Zhou, R., Zeng, S., Wei, D., Deng, X., Xing, C., Yu, L., Deng, Z., Wang, H., Weng, S., He, J., & Huang, Z. (2020). Intestine Bacterial Community Composition of Shrimp Varies Under Low- and High-Salinity Culture Conditions. Frontiers in Microbiology, 11, 589164. https:// doi.org/10.3389/FMICB.2020.589164/BIBTEX Huang, F., Pan, L., Song, M., Tian, C., & Gao, S. (2018). Microbiota assemblages of water, sediment, and intestine and their associations with environmental factors and shrimp physiological health. Applied Microbiology and Biotechnology, 102(19), 8585–8598. https://doi.org/10.1007/s00253-018-9229-5 Huang, Z., Li, X., Wang, L., & Shao, Z. (2016). Changes in the intestinal bacterial community during the growth of white shrimp, Litopenaeus vannamei. Aquaculture Research, 47(6), 1737–1746. https://doi.org/10.1111/are.12628 Kesselring, J. J., Kuhn, D. D., Ponder, M. A., Stevens, A. M., & Trushenski, J. (2022). The Influence of Prebiotics, Probiotics, and Exposure to an Opportunistic Pathogen on the Intestinal Microbiome of White Shrimp (Litopenaeus vannamei) Li, E., Xu, C., Wang, X., Wang, S., Zhao, Q., Zhang, M., Qin, J. G., & Chen, L. (2018). Gut Microbiota and its Modulation for Healthy Farming of Pacific White Shrimp Litopenaeus vannamei. En Reviews in Fisheries Science and Aquaculture (Vol. 26, Número 3, pp. 381–399). Taylor and Francis Inc. https:// doi.org/10.1080/23308249.2018.1440530 Liang, F., Li, C., Hou, T., Wen, C., Kong, S., Ma, D., Sun, C., & Li, S. (2020). Effects of Chitosan–gentamicin conjugate supplement on nonspecific immunity, aquaculture water, intestinal histology and microbiota of pacific white shrimp (litopenaeus vannamei) Marine Drugs, 18(8). https://doi.org/10.3390/MD18080419
- ABRIL 2024 71 TECNOLOGÍA
enfermedades y promover prácticas de manejo que favorezcan la salud y el bienestar de los camarones (Alfiansah et al., 2020; Chen et al., 2017; Cornejo-Granados et al., 2017b).
El estudio de microbiomas también facilita la identificación de patógenos y la evaluación de su prevalencia en las poblaciones de camarones. Esto ayuda en el diagnóstico temprano y el control de enfermedades, permitiendo la implementación de medidas preventivas y terapéuticas de manera más eficiente (Chang et al., 2023).
El hepatopáncreas y el intestino medio de los crustáceos son responsables de la digestión, absorción y almacenamiento de los nutrientes necesarios para apoyar el crecimiento de los camarones. En este contexto se destacan la Necrosis Hepatopancreática Aguda (AHPND), considerada como una de las patologías infecciosas bacterianas más virulentas del camarón debido a su rápida aparición y alta tasa de mortalidad, y la Hepatopancreatitis Necrotizante (NHP-B), causada por Vibrio parahaemolyticus y los organismos similares a la rickettsia (RLO), que han generado pérdidas significativas en la producción mundial de camarones (Chen et al., 2017; Cornejo-Granados et al., 2017b).
Selección de probióticos y simbióticos
Mediante el análisis de microbiomas, se pueden identificar microorganismos beneficiosos que promuevan la salud y el crecimiento de los camarones. Estos microorganismos pueden utilizarse como probióticos o simbióticos para mejorar la resistencia a enfermedades y optimizar la digestión y la absorción de nutrientes (Cicala et al., 2020; Garibay-Valdez et al., 2019; Hembrom et al., 2024; Holt et al., 2021; Li et al., 2018; Pinoargote et al., 2018).
Optimización de la alimentación y nutrición
Entender la interacción entre la microbiota intestinal y la dieta de los camarones es crucial para optimizar la formulación de alimentos acuícolas. El análisis de microbiomas puede proporcionar información sobre la capacidad de los camarones para digerir ciertos nutrientes y ayudar a diseñar dietas más eficientes y saludables (Ding et al., 2021; Kesselring et al., 2022; Liang et al., 2020).
Mejora de la calidad del entorno
El microbioma acuático influye en la calidad del agua y en la estabilidad del ecosistema acuático. Comprender la composición y la función de la microbiota acuática puede ayudar a diseñar estrategias para mantener una calidad de agua óptima, reducir la acumulación de contaminantes y prevenir la proliferación de patógenos (Guo et al., 2020).
El estudio de microbiomas puede contribuir a prácticas acuícolas más sostenibles, al ayudar a minimizar el uso de antimicrobianos y promover el uso de enfoques más naturales y respetuosos con el medio ambiente para controlar enfermedades y mantener la salud de los camarones.
Perspectivas futuras y aplicaciones potenciales
Las perspectivas futuras en el estudio de microbiomas de camarones ofrecen un amplio campo de aplicaciones potenciales que pueden revolucionar la acuicultura y la gestión de recursos acuáticos. Basado en el conocimiento de las interacciones microbioma-hospedero, se pueden desarrollar terapias probióticas y simbióticas personalizadas para mejorar la salud y el rendimiento de los camarones, reduciendo así la dependencia de antimicrobianos y químicos.
Estrategias de manipulación del microbioma, como la adición de microorganismos beneficiosos, prebióticos y prebióticos, pueden utilizarse para modular la composición y la función del microbioma intestinal y promover una salud óptima en los camarones.
La identificación de biomarcadores microbianos asociados con la salud, la enfermedad y el rendimiento de los camarones puede facilitar el monitoreo y la evaluación de la salud del hospedero y la eficacia de las intervenciones terapéuticas.
Además, el conocimiento de la microbiota acuática y su papel en la calidad del agua puede aplicarse en programas de biorremediación para mejorar la calidad ambiental de los sistemas acuícolas y restaurar ecosistemas acuáticos degradados•
Para mayor información escriba a: cesariotito@yahoo.com
Ochoa-Romo, J. P., Cornejo-Granados, F., Lopez-Zavala, A. A., Viana, M. T., Sánchez, F., Gallardo-Becerra, L., Luque-Villegas, M., ValdezLópez, Y., Sotelo-Mundo, R. R., Cota-Huízar, A., López-Munguia, A., & Ochoa-Leyva, A. (2022). Agavin induces beneficial microbes in the shrimp microbiota under farming conditions. Scientific Reports 2022 12:1, 12(1), 1–16. https://doi.org/10.1038/s41598-022-10442-2 Pinoargote, G., Flores, G., Cooper, K., & Ravishankar, S. (2018). Effects on survival and bacterial community composition of the aquaculture water and gastrointestinal tract of shrimp (Litopenaeus vannamei) exposed to probiotic treatments after an induced infection of acute hepatopancreatic necrosis disease. Aquaculture Research, 49(10), 3270–3288. https://doi.org/10.1111/are.13791 Rieder, J., Kapopoulou, A., Bank, C., & AdrianKalchhauser, I. (2023). Metagenomics and metabarcoding experimental choices and their impact on microbial community characterization in freshwater recirculating aquaculture systems. Environmental Microbiome, 18(1), 8. https:// doi.org/10.1186/s40793-023-00459-z
Robinson, C., & Chin, Y.-P. (2023). The structure, composition, and predicted microbiome functional genes in Pacific white shrimp (Litopenaeus vannamei) grow-out ponds with different survival rates through high-throughput sequencing. Aquatic Sciences, 85(84), 1–13. https://www.springer.com/journal/27/editors Secuenciación de microbiomas para el manejo adaptativo de la producción en estanques. (s/f). Recuperado el 30 de marzo de 2024, de https://aquatechcenter.com/ news/microbiome-sequencing-in-ponds/ Trung Tran, T., Bott, N. J., Dai Lam, N., Trung Nguyen, N., Hoang Thi Dang, O., Hoang Le, D., Tung Le, L., & Hoang Chu, H. (2019). The Role of Pseudomonas in Heterotrophic Nitrification: A Case Study on Shrimp Ponds (Litopenaeus vannamei) in Soc Trang Province. Microorganisms, 7(6), 155. https://doi.org/10.3390/microorganisms7060155
Wang, R., Guo, Z., Tang, Y., Kuang, J., Duan, Y., Lin, H., Jiang, S., Shu, H., & Huang, J. (2020). Effects on development and microbial community of shrimp Litopenaeus vannamei larvae with probiotics treatment. AMB Express, 10(1). https://doi.org/10.1186/s13568-020-01041-3 Yun, H. S., Kim, D. H., Kim, J. G., Kim, Y. S., & Yoon, H. S. (2022). The microbial communities (bacteria, algae, zooplankton, and fungi) improved biofloc technology including the nitrogen-related material cycle in Litopenaeus vannamei farms. Frontiers in Bioengineering and Biotechnology, 10, 883522.
https://doi.org/10.3389/fbioe.2022.883522
Zeng, S., Huang, Z., Hou, D., Liu, J., Weng, S., & He, J. (2017). Composition, diversity and function of intestinal microbiota in pacific white shrimp (Litopenaeus vannamei) at different culture stages. PeerJ, 2017(11).
https://doi.org/10.7717/peerj.3986
- ABRIL 2024 72 TECNOLOGÍA
ESTADÍSTICAS
Edición 158 - Abril 2024 74 79 82 Exportaciones de camarón Reporte de mercado de EE. UU. Reporte de mercado de China
73
ÍNDICE
COMERCIO EXTERIOR
CAMARÓN:
EVOLUCIÓN DE
EXPORTACIONES
2010 -2023
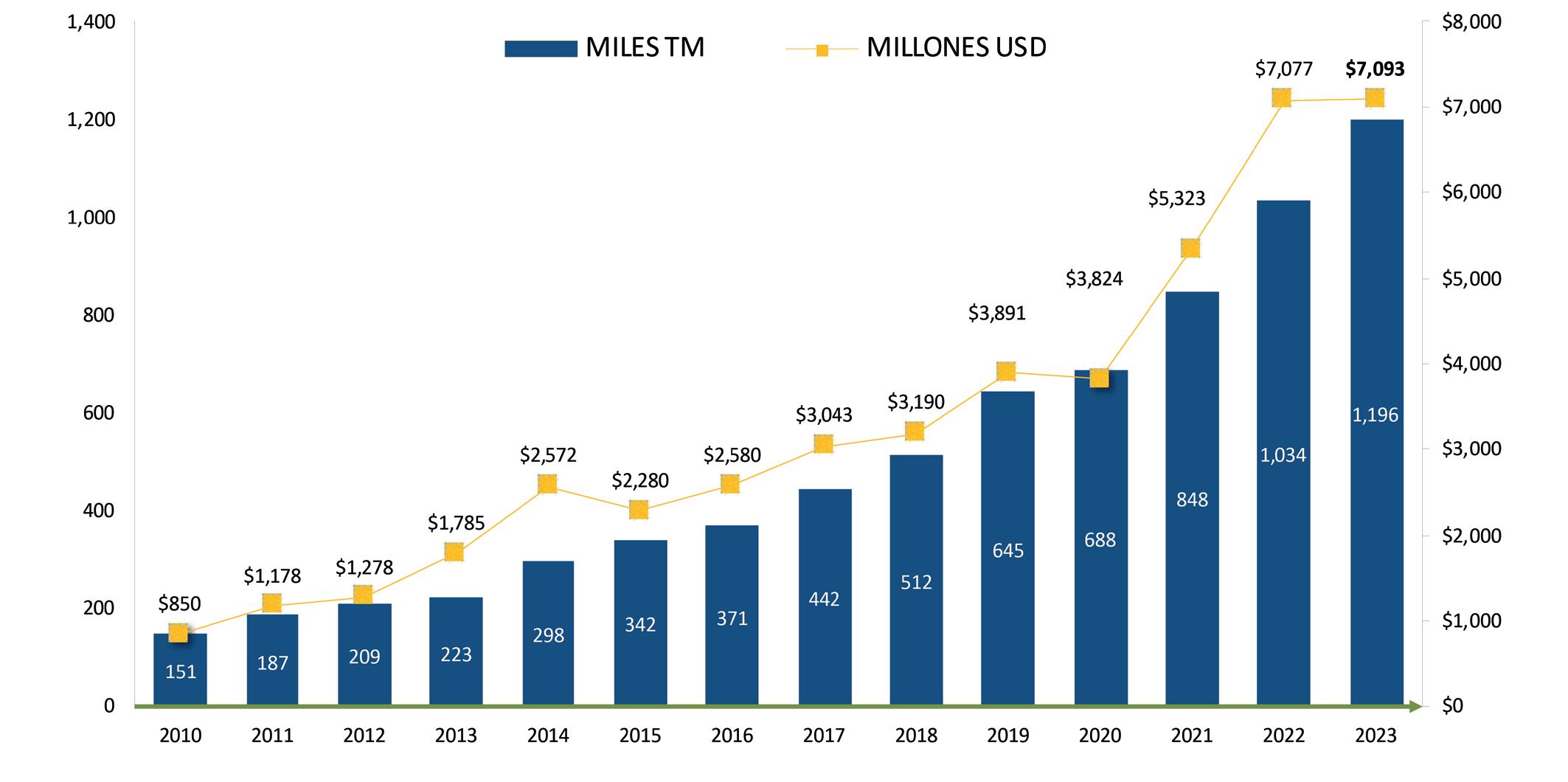
Fuente: Cámara Nacional de Acuacultura
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: COMPARATIVO MENSUAL
(Millones de Libras) 2020- 2024
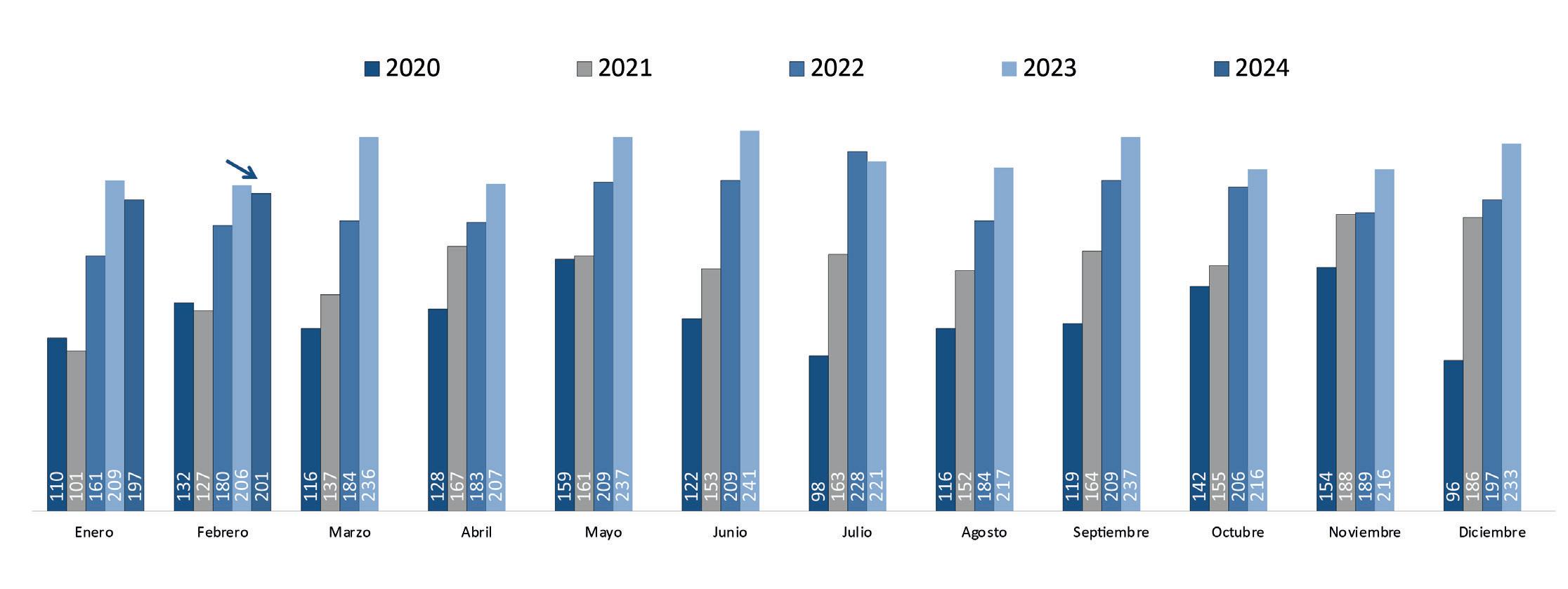
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- ABRIL 2024 74
ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: PARTICIPACIÓN POR DESTINO (Libras)

Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: PRINCIPALES PAÍSES DESTINOS DE EXPORTACIÓN (Millones de Libras)
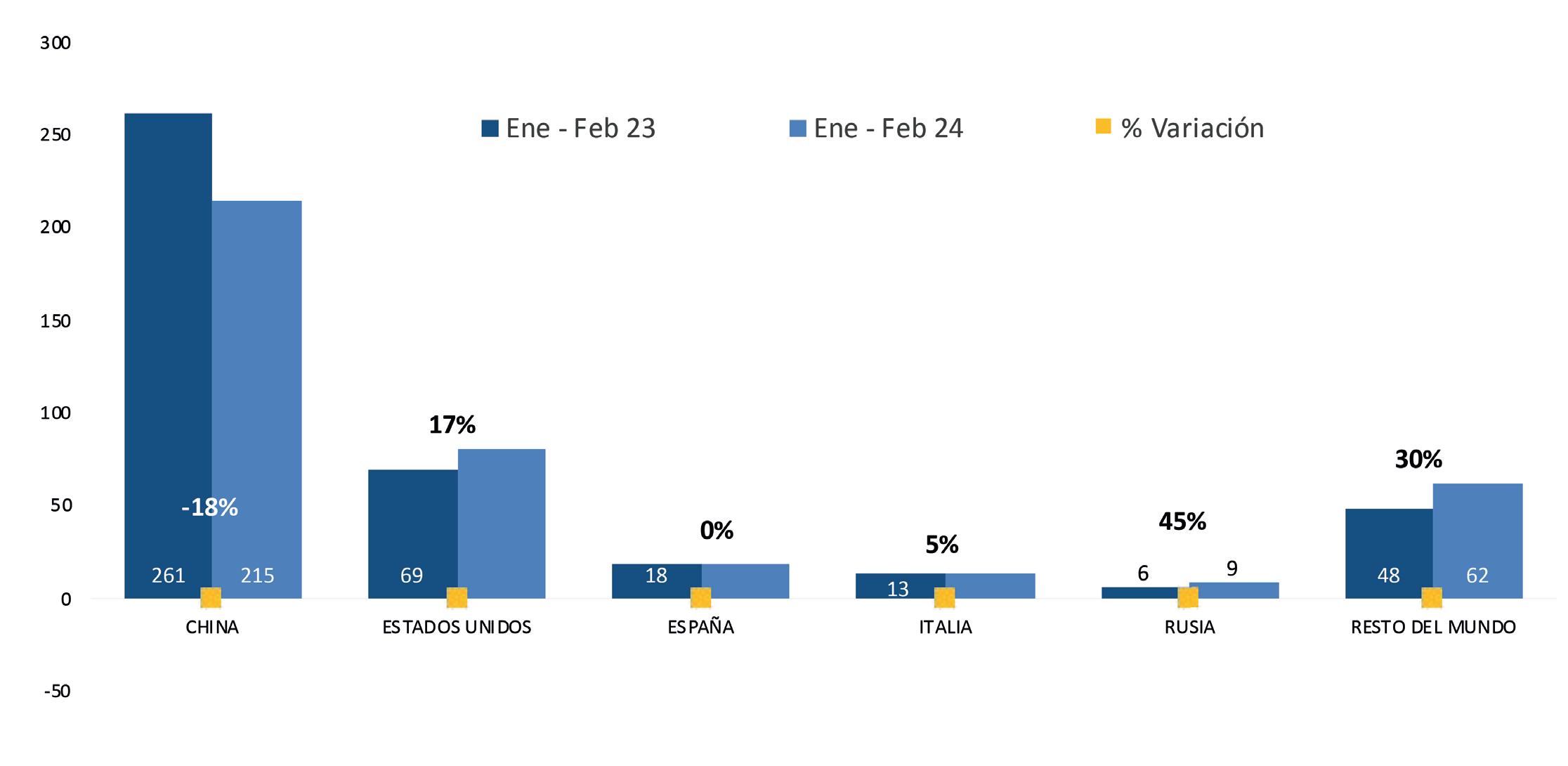
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- ABRIL 2024 75 ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: EXPORTACIONES POR PARTIDA ARANCELARIA
Enero - Febrero 2024
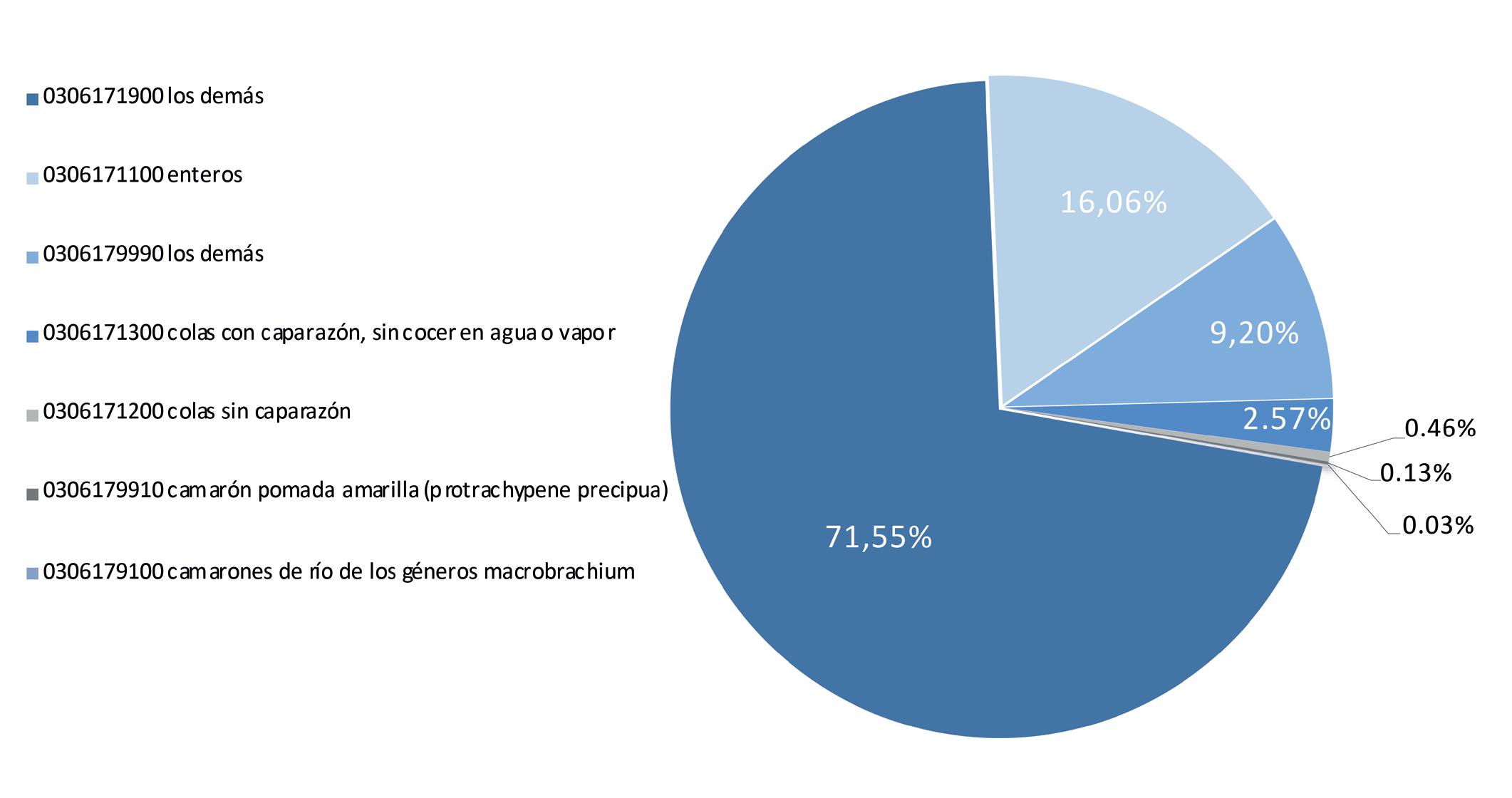
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO MENSUAL DE EXPORTACIÓN (Libras)
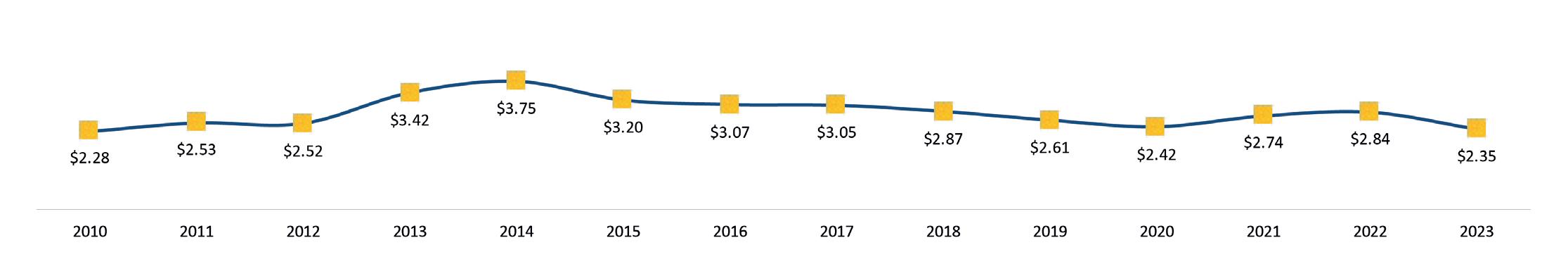
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- ABRIL 2024 76 ESTADÍSTICAS

Importación de camarón de China
 Autor: Sander Visch
Autor: Sander Visch
Kontali Shrimp sander@kontali.no www.shrimp.kontali.no

Después de cierto retraso, la aduana china ha publicado las estadísticas de importación de enero y febrero. Las importaciones durante estos meses aumentaron en comparación con el mismo período del año pasado, mientras que los precios de importación cayeron más durante ambos meses.
El volumen de importación en enero experimentó un aumento impresionante en comparación con diciembre de 2023, alcanzando 104.180 toneladas, un incremento del 31% intermensual y del 56% interanual. Sin embargo, es importante señalar que estas cifras se reportaron junto con la data de febrero, cuando los volúmenes de importación cayeron a 65.240 toneladas, un 37% menos que en enero y un 15% menos que el año pasado. Debido a que estas cifras se reportan simultáneamente, analizaremos ambos meses en conjunto, ya que esperamos que brinden una imagen más realista de las estadísticas comerciales.
La caída está relacionada con el Año Nuevo chino y la festividad asociada. La mayoría de los importadores cierran durante ciertos momentos de este período y, cuando abren, suelen estar más centrados en llevar su camarón al mercado que en adquirir más stock. Debido a que estas cifras se reportan simultáneamente, analizaremos ambos meses juntos, ya que esperamos que brinden una imagen más realista de las estadísticas comerciales.
La combinación de los volúmenes da una cifra hasta la fecha de 169.420 toneladas, un aumento del 18% en comparación con el mismo período en 2023. Si analizamos más de cerca los volúmenes de importación del año pasado, vemos un aumento significativo durante marzo, ya que los efectos finales de los problemas logísticos en el país disminuyeron, lo que significa que el aumento durante los primeros dos meses de este año podría reflejar un aumento de la demanda menos impactante, como se puede esperar si se observan solo las cifras.
El precio de importación durante enero y febrero bajó ambos meses, alcanzando $4.94/kg y $4.92/kg, respectivamente, frente a $5.39/ kg durante el último mes de 2023. El valor total de las importaciones en lo que va de 2024 alcanzó $836 millones, un aumento de 4% en comparación interanual.
La importación de productos de camarón congelados HS 030617 alcanzó 157.340 toneladas hasta la fecha, un 19% más en comparación con el mismo período del año pasado. Los precios de importación cayeron $0.36 en enero y nuevamente $0.08 en febrero, finalizando en $4.68/kg durante ese mes, con un promedio de $4.73/kg durante los dos primeros meses del año.
Ecuador sigue siendo el mayor proveedor de productos de camarón congelados bajo este código HS, con 114.720 toneladas provenientes del país hacia China a principios de año, un aumento del 20% en comparación con el año anterior. Otro aumento significativo se observó en el segundo mayor proveedor del mercado chino, India, donde los volúmenes aumentaron un 25% hasta alcanzar 22.160 toneladas durante enero y febrero. Un notable aumento del 184%
MERCADO
MERCADO 79
Importaciones chinas en 2021, 2022, 2023 y 2024 (volumen y precio medio/kg)
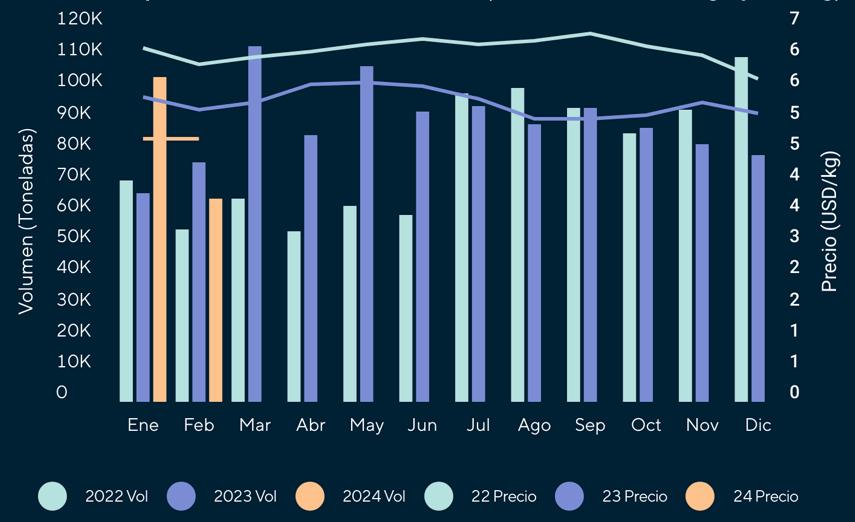
en el volumen suministrado hizo que Arabia Saudita creciera hasta convertirse en el tercer mayor proveedor de productos de camarón congelado con 5.150 toneladas, seguido de Tailandia y Perú. Con volúmenes de 3.890 toneladas y 3.080 toneladas, incrementos del 61% y 64%, respectivamente.
Eventos como el Festival Qingming y el feriado del Primero de Mayo se acercan para el mercado chino, lo que suele impulsar la demanda. Sin embargo, este año ha habido una menor demanda durante las vacaciones de Año Nuevo, pero esto no disminuye el optimismo entre los productores chinos.
Una razón para este optimismo es la suspensión de múltiples envíos de camarón ecuatoriano debido a niveles excesivos de dióxido de azufre. Esto provoca una menor importación en febrero y posibles retrasos en marzo.
Esto podría provocar una escasez de oferta de camarón justo antes del aumento de la demanda en los festivales, lo que elevaría los precios del camarón nacional.
Los comerciantes ecuatorianos prometen controles más estrictos para evitar problemas futuros. Sin embargo, los consumidores chinos
no están muy preocupados, por lo que es posible que la demanda de camarón congelado importado no disminuya significativamente.
En cuanto a la producción, si bien los stocks actuales son bajos, el repentino aumento de precios en finca en el delta del río Perla genera optimismo entre los productores. En la región norte, la disponibilidad de materia prima es aún menor, lo que lleva a un mayor flujo de transporte de larga distancia que viene a recoger camarón a un precio relativamente alto en provincias como Guangdong•
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
- ABRIL 2024 80 MERCADO

Importación de camarón de Estados Unidos


Autores: Jim Kenny jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com
Urner Barry

Importaciones de todos los tipos, por tipo
Con el antecedente de restricción hecho por la Comisión de Comercio Internacional (ITC) de EE.UU. que mantiene derechos antidumping (AD) y compensatorios (CVD) sobre el camarón importado después de haber publicado las primeras importaciones del año, la importación de camarón de enero cayó un 13.5% con respecto al año pasado. Las cifras de enero de 2024 ascendieron a 131.041 millones de libras, frente a los 151.535 millones de libras del año pasado.
Hubo amplias caídas en muchos de los principales socios comerciales. India (-18.1%), Ecuador (-5.7%), Indonesia (-19.6%), Vietnam (-5.7%), Tailandia (-24.4%) y China (-23.8%) quedaron rezagados en enero pasado. México (+7.9%) y Argentina (+6.1%) ganaron.
Una nota importante en el primer informe fue que las ganancias en el camarón mexicano ayudaron a impulsar a este país más allá de Tailandia para ocupar el cuarto lugar con 4.224 millones de libras. Además, la brecha entre India y Ecuador era de apenas 5.307 millones de libras. El año pasado fue de casi 10 millones de libras y promedió una diferencia de 16.569 millones de libras por mes en 2023.
En términos de forma de producto, fue una pérdida generalizada. Hubo menos camarón con cáscara sin cabeza, que incluye el pelado fácil (-6.5%), pelado (-15.5%), cocido (-23.4%) y empanizado (-10.3%).
Ciclos de importación mensual por país (todos
los tipos)
India: Las importaciones de camarón procedente de la India (-18.1%) cayeron bruscamente en enero. El total de 44.213 millones de libras quedaron por debajo de los 54.006 millones de libras del año pasado al comenzar el año. Se produjeron descensos importantes y generalizados en los envíos de camarón con cáscara (-37.3%), pelado (-12.0%) y cocido (-24.5%).
Ecuador: Después de ser el único país entre los cinco primeros con ganancias positivas en 2023, enero comenzó como casi el resto de los socios comerciales de Estados Unidos, con Ecuador (-5.7%) enviando menos camarón. Se enviaron 38.906 millones de libras y, aunque es menos, esto redujo la brecha con la India a poco más de 5 millones de libras. En enero, volvieron a aumentar los productos de camarón con cáscara (+4.7%), pero menos pelado (-22.2%) y empanizado (-15.3%).
Indonesia: En el primer mes del año se enviaron 25.104 millones de libras a Estados Unidos. Esta disminución interanual del 19.6% estuvo en línea con las tendencias que iniciaron en 2024. Hubo grandes caídas de dos dígitos en los productos con cáscara (-17.0%), pelado (-26.8%), cocido (-19.7%) y medianas pérdidas en el empanizado (-6.0%).
Vietnam y Tailandia: Para completar la lista, Vietnam (-5.7%) y Tailandia (-24.4%) exportaron menos. De hecho, la gran pérdida en este último país llevó a Tailandia a caer por debajo de México hasta el sexto lugar al comienzo del año.
- ABRIL 2024
82
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
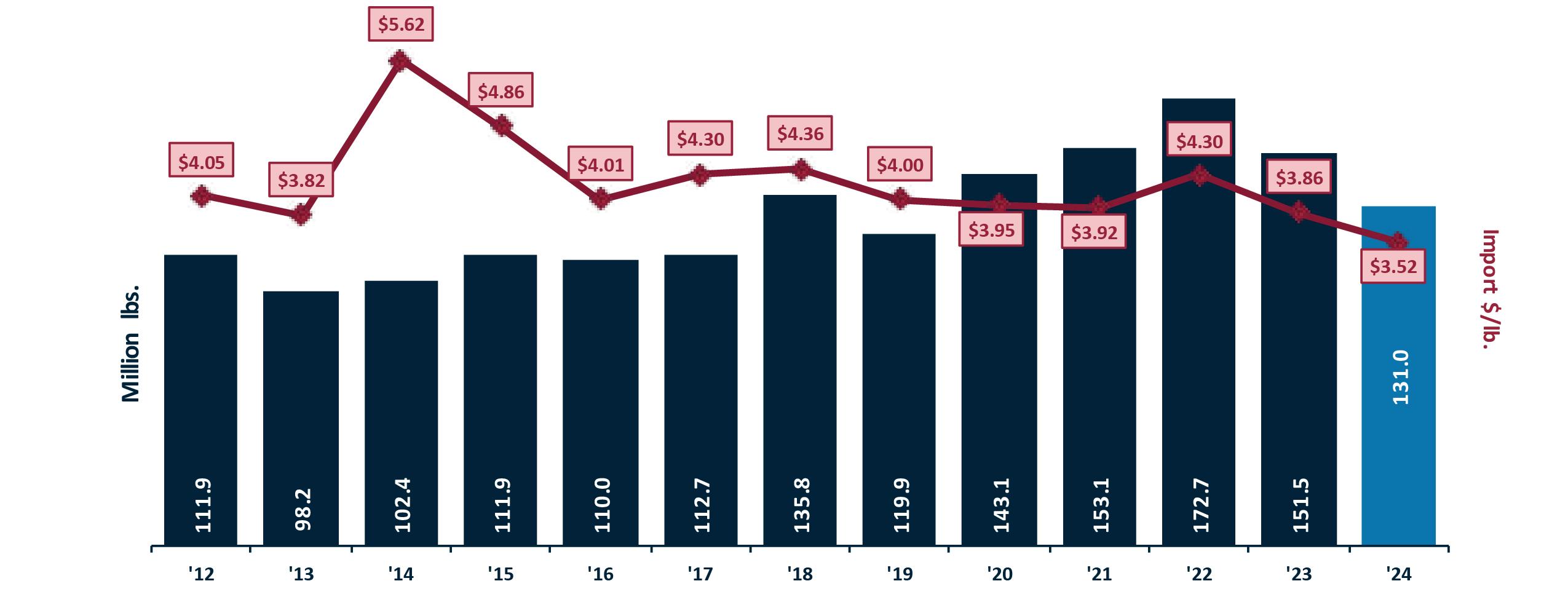
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones de camarón con cáscara, que incluye el pelado fácil, cayeron un 6.5% en enero.
Durante el mes, las mayores pérdidas de Indonesia (-17.0%) e India (-37.3%) compensaron las ligeras ganancias de sus principales socios, Ecuador (+4.7%) y México (+8.4%).
Las ganancias se mantuvieron en el camarón muy grande, U/15 y 41-50. Todos los demás tamaños fueron negativos, algunos de manera espectacular, en una comparación año tras año.
Los valores de reposición del camarón con cáscara (importación dólares/libra) volvieron a caer, pero se mitigaron las pérdidas. Los precios fueron de $3.40 por libra, sólo $0.02 por libra menos, lo que llevó a nuevos mínimos del ciclo.
Valor agregado, importación de camarón pelado
Las importaciones de camarón pelado, la categoría más grande en volumen, cayeron con un enorme 15.5% en enero. Las importaciones del mes totalizaron 57.215 millones de libras. Se observaron grandes pérdidas en los tres principales países: India (-12.0%), Ecuador (-22.2%) e Indonesia (-26.8%). Vietnam (+35.9%) fue el único ganador entre los cinco primeros.
Los valores de reemplazo (importación $/ libra) para el camarón pelado bajaron de $0.08 por libra a $3.38 por libra, el nivel más bajo desde julio de 2010.
Las importaciones de camarón cocido (de agua caliente) cayeron un 23.4% en enero, mientras que el empanizado alcanzó un 10.3% menos que el año pasado.
Importaciones de camarón cocido, empanizado y otros
Los precios de reemplazo del camarón cocido también bajaron ligeramente, a $4.18/ libra desde $4.22/libra. Enero de 2013 fue la última vez que los valores de reposición fueron tan bajos. A pesar de las continuas caídas de las importaciones de producto cocido, la demanda sigue estando en un nivel insuficiente para ofrecer producto.
Línea de tiempo del precio del camarón; anuncios minoristas
Comercio minorista: Las oportunidades de compra minorista cayeron en febrero, una tendencia típica ya que el mes de los productos del mar sigue en marzo y los puntos de venta se preparan para una exposición total. La actividad de funciones continuó situándose por debajo del promedio de tres años. Los precios, sin embargo, subieron $0.62 por libra a $8.01 por libra. La acción de los precios minoristas sigue estando desvinculada de los precios mayoristas.
Suministro de camarón a EE.UU. y situación del Golfo
***La data de desembarques proporcionada por la NMFS ha sido inconsistente y existen lagunas en la data que hace que las comparaciones no sean confiables.
Detalle de importaciones de camarón YTD por tipo
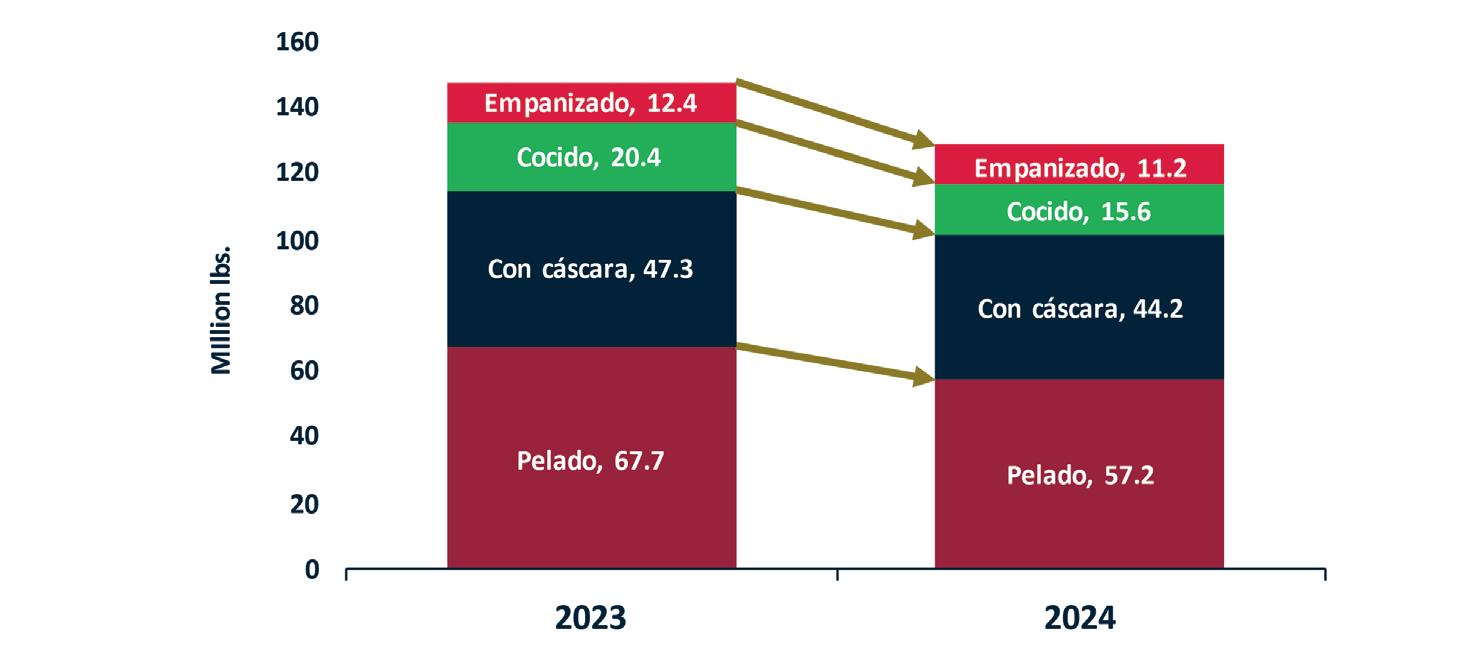
- ABRIL 2024 83
MERCADO
REPORTE DE
Fuente: USDOC. Urner Barry
Fuente: USDOC. Urner Barry
REPORTE DE MERCADO
En noviembre de 2023 la data de desembarque publicada por la NMFS muestra que se desembarcaron 8.863 millones de libras (en total, todas las especies, sin cabeza); poco más del 2% por encima de 2022. La cifra de 84.061 millones de libras todavía se sitúa casi un 19% por debajo del año pasado, en lo que va del año. Se ha observado poca o ninguna acción de precios en esta categoría, pero hemos estado monitoreando la producción y distribución de 7.77 millones de libras de camarones P&D de talla 21-30 a 51-70 que se entregarán del 16 de marzo al 30 de junio de 2024, como parte del Programa de Adquisición de Commodities del USDA y analizar qué impacto puede tener en el mercado.
Exportación del camarón ecuatoriano
El mercado de camarón tigre blanco y negro importado, cultivado en granjas, se encuentra entre estable y firme. Los costos en el extranjero han aumentado junto con el transporte, el almacenamiento y los intereses. Los importadores conservan el producto con confianza y creen que el inventario será cada vez más valioso.
Otro shock para el mercado son las recientes determinaciones positivas preliminares del Departamento de Comercio (DOC) en las investigaciones sobre CVD.
En cuanto a las tasas de subsidio preliminares, el exportador/productor ecuatoriano Industrial Pesquera Santa Priscila ha recibido la mayor tasa de subsidio con un 13.41%. La respuesta inicial de la empresa es que apelarán el fallo. La Sociedad Nacional de Galápagos tiene una tasa de subsidio del 1.69%, mientras que “todos los demás” en Ecuador tienen una tasa de subsidio del 7.55%.En cuanto a la India, a Devi Sea
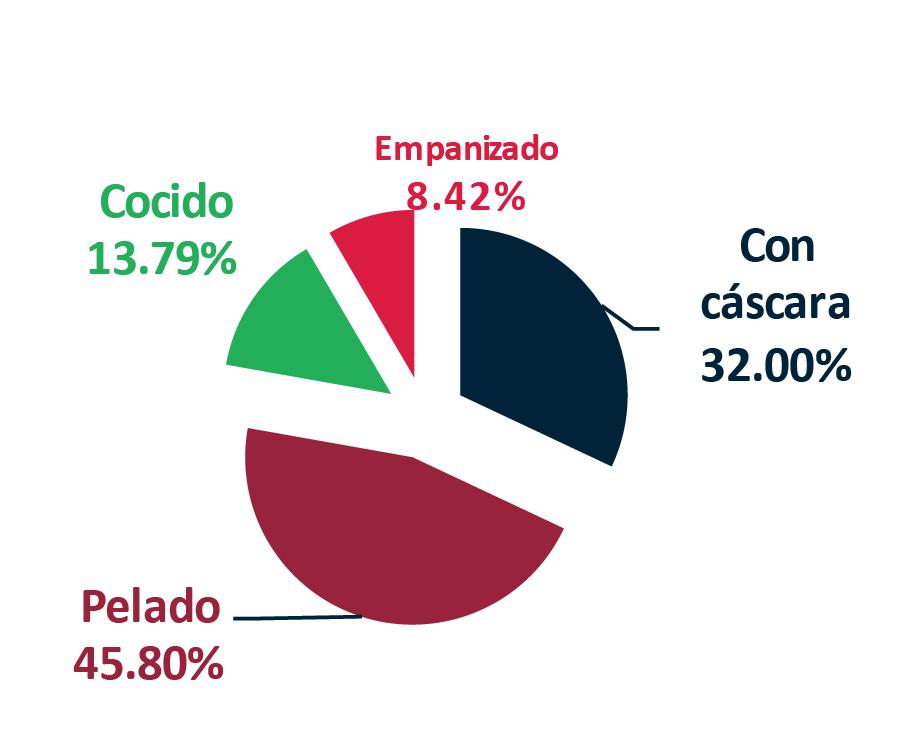
Foods Limited se le ha concedido una tasa de subvención del 4.72%. Se ha determinado que Sandhya Aqua Exports, Neeli Sea Foods Private Limited, Vijay Aqua Processors Private Limited y Neeli Aqua Farms tienen una tasa de subvención del 3.89%. "Todos los demás" en la India tienen una tasa de subsidio del 4.36%.
Fuera de Vietnam, la sociedad anónima Soc Trang Seafood y “todos los demás” tienen una tasa de subvención del 2.84%•
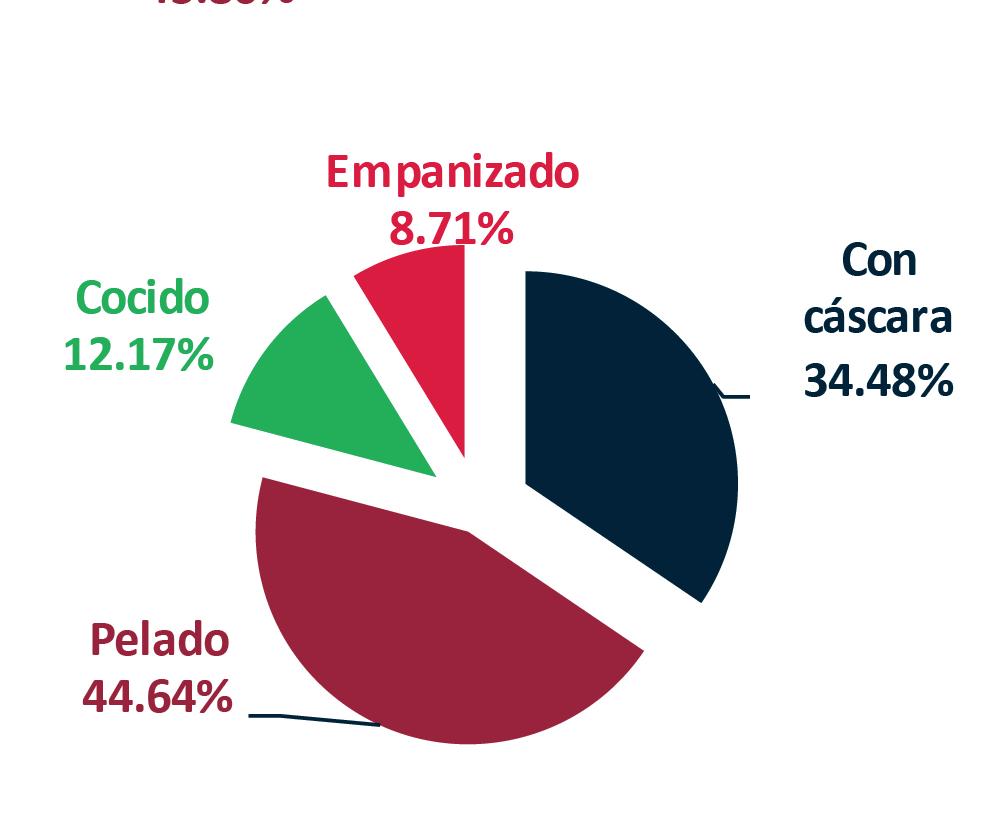

- ABRIL 2024
2023 2024
Nuevas autoridades
La Viceministra de Acuacultura y Pesca, Dra. María Rafaela Hurtado Espinoza, y la Subsecretaria de Calidad e Inocuidad, Mgs. Ana María Maldonado Carrasco, junto con el subsecretario Axel Vedani, visitaron la Cámara Nacional de Acuacultura (CNA) el 19 de marzo pasado.
Las autoridades se reunieron con el Presidente Ejecutivo de CNA, José Antonio Camposano, y ejecutivos de la Cámara para analizar la situación actual del sector y coordinar acciones inmediatas para atender asuntos prioritarios de nuestra actividad.
El propósito de este encuentro fue establecer una plataforma de diálogo permanente para abordar los desafíos críticos que enfrenta la industria acuícola. Entre los temas discutidos se incluyeron aspectos relacionados con las suspensiones a establecimientos ecuatorianos en China, así como la problemática de los controles reforzados en la
Mesa de trabajo por la seguridad con la delegación de la Unión Europea en Ecuador
Con el propósito de analizar la situación actual en materia de seguridad sobre las exportaciones ecuatorianas, Peter Wagner, Director del Servicio de Instrumentos de Política Exterior de la Comisión Europea (Service for Foreign Policy Instruments - FPI), junto con demás funcionarios de la FPI y el Embajador Charles-Michel Geurts, se reunieron con líderes de varios sectores productivos y de exportación, entre ellos la Corporación de Gremios Exportadores del Ecuador, a la que pertenece la Cámara Nacional de Acuacultura.
Este programa es organizado por la Corporación de Promoción de Exportaciones e Inversiones del Ecuador (CORPEI). En su encuentro
Cooperación público –privada para la seguridad
El Director de Seguridad de la Cámara Nacional de Acuacultura, Luis Herrera, participó en una mesa de trabajo liderada por el Director Nacional de la Policía Judicial, Cnel. Gustavo Zárate, y el Jefe de Operaciones de la Zona 8. El objetivo de este encuentro fue actualizar información sobre la problemática de la delincuencia en el país. La iniciativa busca proporcionar a los afiliados y miembros del sector las herramientas necesarias para minimizar riesgos y contrarrestar las acciones delictivas perpetradas por grupos criminales. Lo más destacado de la reunión fue la disposición del Director de la Policía Judicial de instruir a los jefes zonales para fortalecer las investigaciones, localizar y poner a disposición de las autoridades pertinentes a miembros de estructuras criminales identificadas.
Con el levantamiento de información se logró reunir los elementos probatorios necesarios y llevar a cabo las diligencias legales pertinentes, lo que contribuye significativamente a fortalecer la seguridad y proteger los intereses del sector camaronero.

Unión Europea. Adicionalmente, se trató la agenda de habilitación sanitaria de establecimientos exportadores ecuatorianos y el estado del cumplimiento de tiempos de los trámites en la Subsecretaría de Calidad e Inocuidad.

más reciente, la mesa de trabajo contó con la presencia de Dominique Bucas, Jefe de Misión de la Delegación de la Unión Europea, así como consultores de la UE. En el encuentro se revisaron posibles estrategias para mejorar la seguridad de las terminales portuarias con el objetivo de proteger la cadena logística de exportación.
Bioseguridad en establecimientos acuícolas
Ejecutivos de la Cámara Nacional de Acuacultura participaron en la capacitación sobre "Lineamientos para la Implementación de Bioseguridad Aplicables en Establecimientos Acuícolas", organizada por la Subsecretaría de Calidad e Inocuidad (SCI).
En este evento, se logró brindar a los representantes de establecimientos acuícolas información detallada de los protocolos y directrices necesarios para garantizar la bioseguridad en sus operaciones.
La SCI presentó las mejores prácticas y procedimientos que contribuyen a prevenir y controlar enfermedades en la acuicultura, promoviendo así la producción responsable y sustentable en el sector.
- ABRIL 2024 85 NOTICIAS
Vibrio cholerae

La CNA mantuvo reuniones de seguimiento a las acciones desplegadas para dar solución a los controles específicos a tres establecimientos ecuatorianos en la UE. La adopción de un criterio microbiológico diferente y sin sustento técnico por parte de España para la detección de Vibrio cholerae ha generado alertas en tres empresas ecuatorianas, lo que resultó en controles específicos impuestos por la Dirección General de Salud de la UE a estos productos.
La Cámara Nacional de Acuacultura y el Gobierno Nacional están trabajando en conjunto para levantar estas sanciones, formando un grupo de trabajo que incluye a los ministerios de Relaciones Exteriores y Comercio Exterior. Se está buscando una respuesta de las autoridades europeas y se elabora un informe técnico para respaldar la posición ecuatoriana y resolver este problema que afecta al sector.
SSP Y GSSI exploran posibles colaboraciones en Ecuador

Sustainable Shrimp Partnership (SSP) se reunió con Global Sustainable Seafood Initiative (GSSI) durante la Seafood Expo North America en Boston, donde GSSI expresó interés en expandir su alcance en Latinoamérica, considerando a SSP y a Ecuador como aliados estratégicos para implementar proyectos regionales. El propósito principal fue establecer vínculos, presentar a la nueva gerente de Impactos de ODS (Objetivos de Desarrollo Sostenible) de GSSI, Laura del Castillo-Munera, y discutir las posibles colaboraciones entre ambas organizaciones, incluyendo la opción de que SSP se convierta en un socio afiliado de GSSI.
Esta posible asociación beneficiará a los afiliados y miembros del sector camaronero ecuatoriano al ofrecer posibilidades de apoyo para nuevos proyectos de sostenibilidad.
Detección de metabisulfito
La CNA está trabajando en coordinación con las autoridades locales y chinas para levantar las suspensiones impuestas a varios establecimientos ecuatorianos debido al exceso de metabisulfito de sodio en varios lotes de camarón, así como incumplimientos de normas de etiquetado del producto. Se están presentando medidas correctivas mediante los planes de gestión de crisis preparados por las empresas y la SCI, así como otras gestiones al más alto nivel. La Viceministra de Acuacultura, una vez posesionada, ha solicitado una reunión a la Administración General de Aduanas de China (GACC) para revisar el estado del levantamiento de las suspensiones a los establecimientos nacionales. Asimismo, se están realizando gestiones a través de la Embajada de China en Quito y la oficina comercial de ese país para exponer el estado del sector y que puedan transmitir a las autoridades en Beijing sobre los controles en materia de sanidad e inocuidad que nuestro sector aplica al producto que va a ser exportado.
Ecuador
presente
en
la feria de productos del mar más importante de América del Norte

La Cámara Nacional de Acuacultura informó mediante boletín que 18 empresas exportadoras de camarón ecuatoriano estuvieron presentes en la Seafood Expo North America, la feria de productos del mar más destacada de América del Norte, que se llevó a cabo del 10 al 12 de marzo en el Centro de Convenciones y Exhibiciones de Boston, Estados Unidos.
El pabellón ecuatoriano "First Class Shrimp" ocupó 288 m², marcando un hito al ser la extensión más grande en la historia del evento. La participación de estas empresas, entre las que se encontraban Aquagold, Expalsa y Sustainable Shrimp Partnership (SSP), fue coordinada por la CNA, destacando la importancia estratégica de este encuentro global para la promoción del camarón ecuatoriano en el mercado internacional.
La Seafood Expo North America no solo atrajo a más de 20.000 compradores y proveedores de productos del mar de más de 100 países, sino que también ofreció un congreso con más de 30 sesiones educativas presentadas por expertos de la industria pesquera y acuícola.
- ABRIL 2024 86 NOTICIAS


