Desafíos del camarón ecuatoriano en sus principales mercados
Investigación y Mercado: Sosteniendo el liderazgo del camarón ecuatoriano
Una mirada a la industria del camarón en Guatemala
Las zonas francas: una oportunidad para la exportación del camarón ecuatoriano
Memoria de sostenibilidad: conoce la herramienta que te ayudará a medir tus impactos
Vibriosis en la producción de camarón
Análisis de la disparidad de talla y su relación con el IHHNV en la larvicultura de camarón P. vannamei
Supervivencia de cepas de Vibrio cholerae 01 en camarones sujetos a procesos de congelación y ebullición
¿Cuál es el panorama global de alimentos para el segundo semestre del 2024?
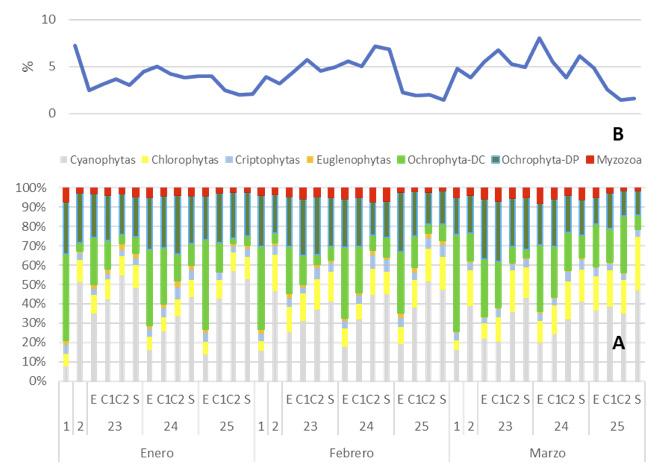
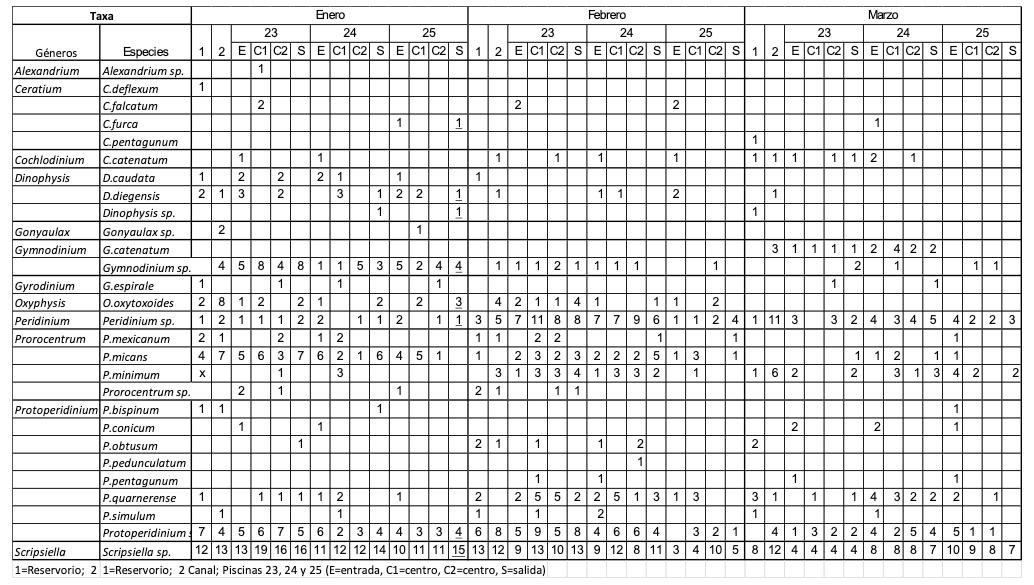
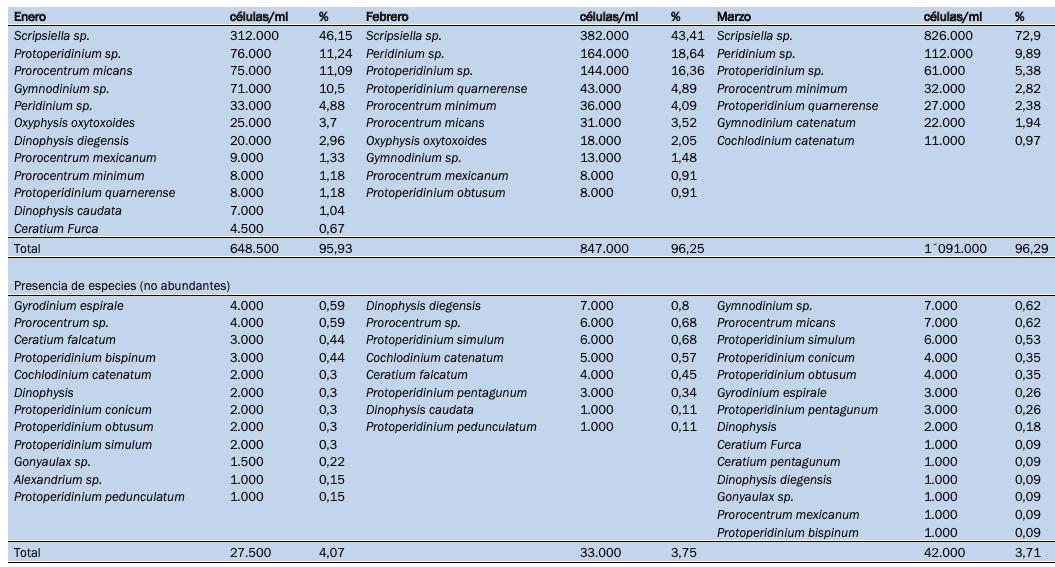
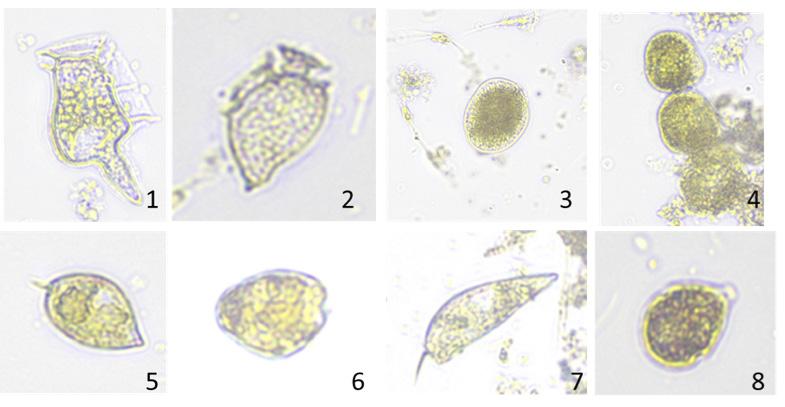
Caracterización de dinoflagelados en un sector de producción de camarón en el golfo de Guayaquil –Ecuador, como un paso inicial a la toma de medidas fitosanitarias
Efecto de la variación temporal de la temperatura sobre la producción del camarón Penaeus vannamei en sistemas de cultivo en piscinas
Bienestar de crustáceos decápodos con especial énfasis en la fisiología del estrés
Niveles de sulfito en langostinos y camarones congelados
Ante el inminente cambio climático: ¿Cuáles son las prácticas a implementar en la acuicultura?
Exportaciones de camarón
Reporte de mercado de China
Reporte de mercado de EE. UU.
Noticias de interés
Noticias empresariales
Presidente Ejecutivo
Ing. José Antonio Camposano
Editora “AquaCultura”
MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita PhD. Leonardo Maridueña Ing. José Antonio Lince Econ. Danny Vélez Ing. Alex de Wind
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Corrección de estilo Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
Gabriela Nivelo gnivelo@cna-ecuador.com
Este mes de junio, la Cámara Nacional de Acuacultura (CNA) ha anunciado logros trascendentales para el sector camaronero, reafirmando su compromiso y determinación.
Primero, hemos asegurado el acceso continuo al crucial mercado de la Unión Europea. La presentación de información técnica a la AESAN de España ha llevado a la actualización de los criterios microbiológicos para determinar la ausencia de cepas patogénicas de Vibrio en el camarón de importación. Este proceso, que tomó cerca de un año, asegura que el camarón ecuatoriano cumple con las más estrictas exigencias de nuestros clientes europeos.
En segundo lugar, tras intensas gestiones técnicas, comerciales y diplomáticas, hemos logrado el levantamiento de las suspensiones a establecimientos ecuatorianos en China. Las exportaciones a nuestro mercado más importante se han restablecido, y estamos trabajando para eliminar por completo las alertas restantes, en colaboración con la Administración General de Aduanas de China y autoridades ecuatorianas.
Sin embargo, estos logros son solo la punta del iceberg. Nuestra labor no se detiene aquí. Continuamos defendiendo al sector ante las infundadas acusaciones de competencia desleal mediante supuestos subsidios y dumping. Desde el
EDITORIAL
José Antonio Camposano Presidente Ejecutivo
La gestión que no se ve, pero que no se detiene
La Cámara Nacional de Acuacultura ha demostrado, a lo largo de más de tres décadas, su incansable compromiso con la resolución de los problemas que afectan a la industria camaronera.
primer día hemos protegido a nuestra industria y a los niveles de competitividad que hemos alcanzado gracias a décadas de esfuerzo privado. ¡Rechazamos categóricamente estas acusaciones y demostraremos nuestra posición ante las autoridades competentes!.
Además, seguimos exigiendo soluciones a la inseguridad que afecta al sector. No dejaremos de demandar resultados concretos por parte del Gobierno, que cobra cuantiosos aportes extraordinarios e impuestos que pagamos puntual y transparentemente. También insistimos en la protección de ecosistemas frágiles, reclamando sanciones ejemplares para los infractores y evitando la impunidad en delitos ambientales. Finalmente, mantenemos una activa participación ante la Asamblea Nacional para asegurar que las leyes reflejen la perspectiva de la producción y exportación de camarón.
La Cámara Nacional de Acuacultura ha demostrado, a lo largo de más de tres décadas, su incansable compromiso con la resolución de los problemas que afectan a la industria camaronera.
En este 2024, seguimos cumpliendo con nuestra misión y la responsabilidad que el gremio camaronero nos ha confiado. ¡Nuestras gestiones, que muchas veces no se ven, no se detienen, pero los resultados están a la vista de todos!•
DIRECTORIO
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Ing. Roberto Aguirre
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Blgo. Carlos Sánchez
Ing. Diego Puente
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Andrés Rivadulla
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León
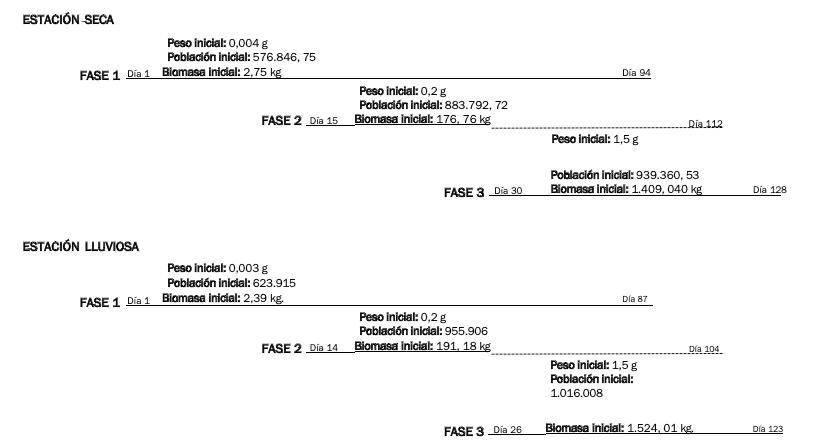
Desafíos del camarón ecuatoriano en sus principales mercados
CHINA
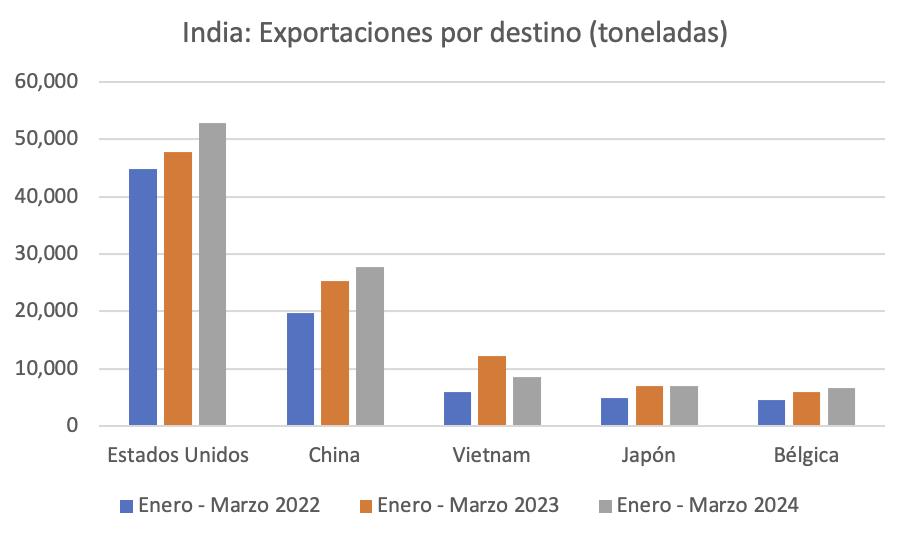
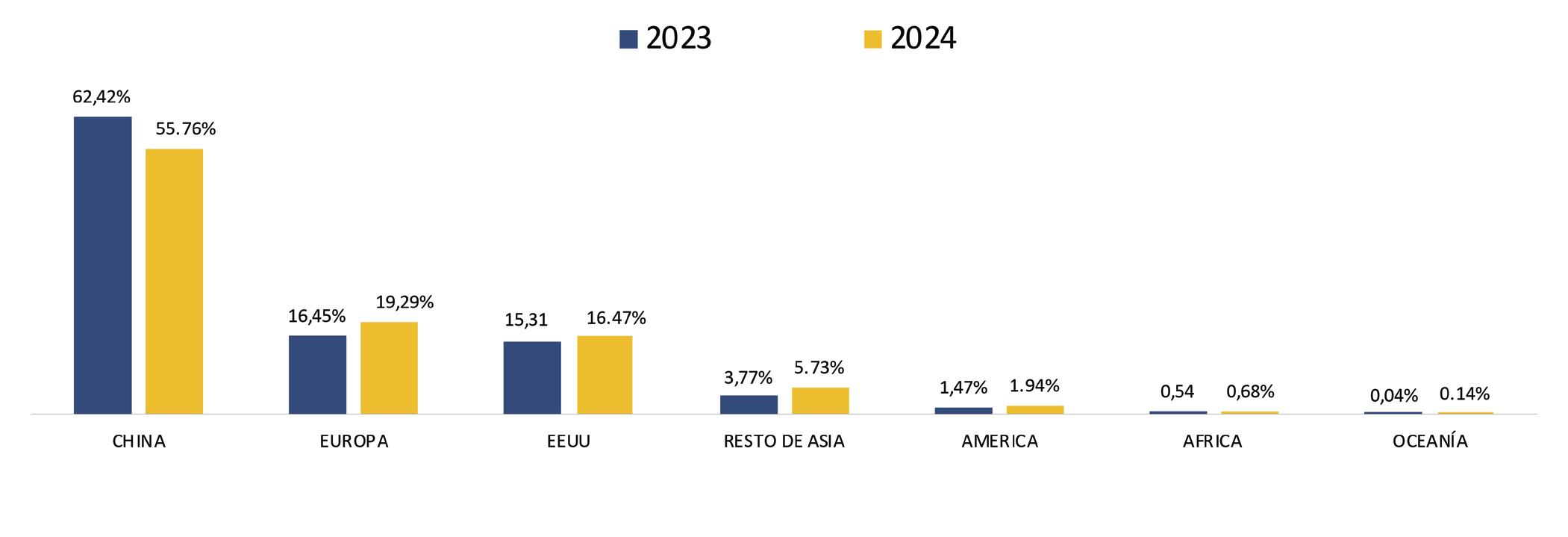
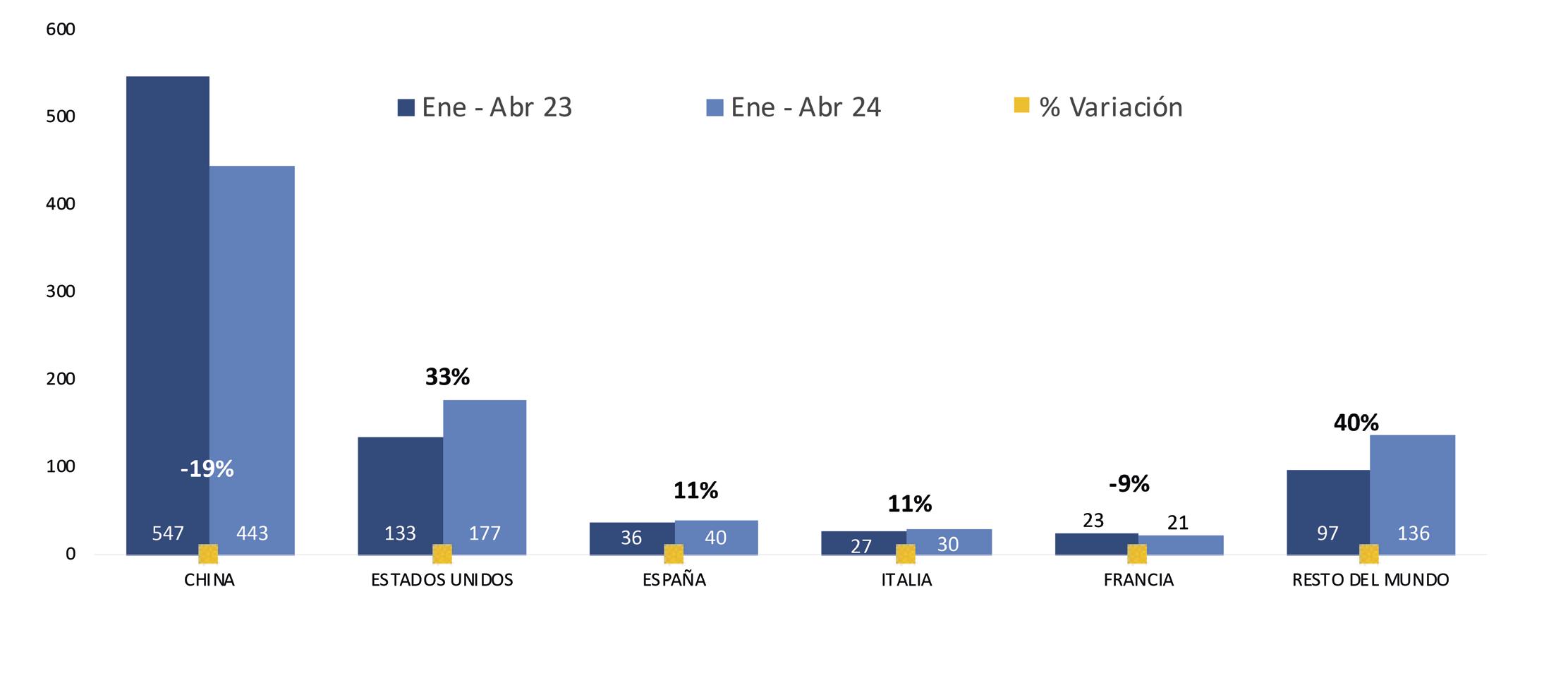
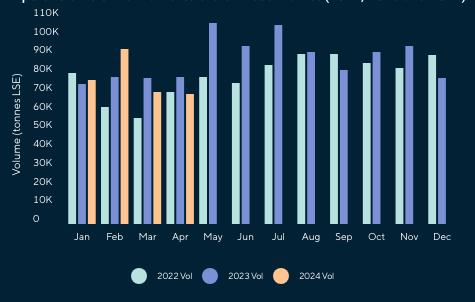
Ecuador es el principal proveedor de camarón al gigante asiático, es decir que 7 de cada 10 libras de camarón que importa China, provienen de nuestro país; sin embargo, el flujo comercial hacia este mercado ha enfrentado graves dificultades en los últimos 5 años.
La más reciente se registró a inicios del 2024, cuando la Aduana de China suspendió temporalmente a nueve establecimientos exportadores de camarón en febrero de este año, por presunta detección de metabisulfito por encima de los límites permitidos. Se trata de un preservante utilizado para evitar coloraciones oscuras en la cabeza o cola del camarón.
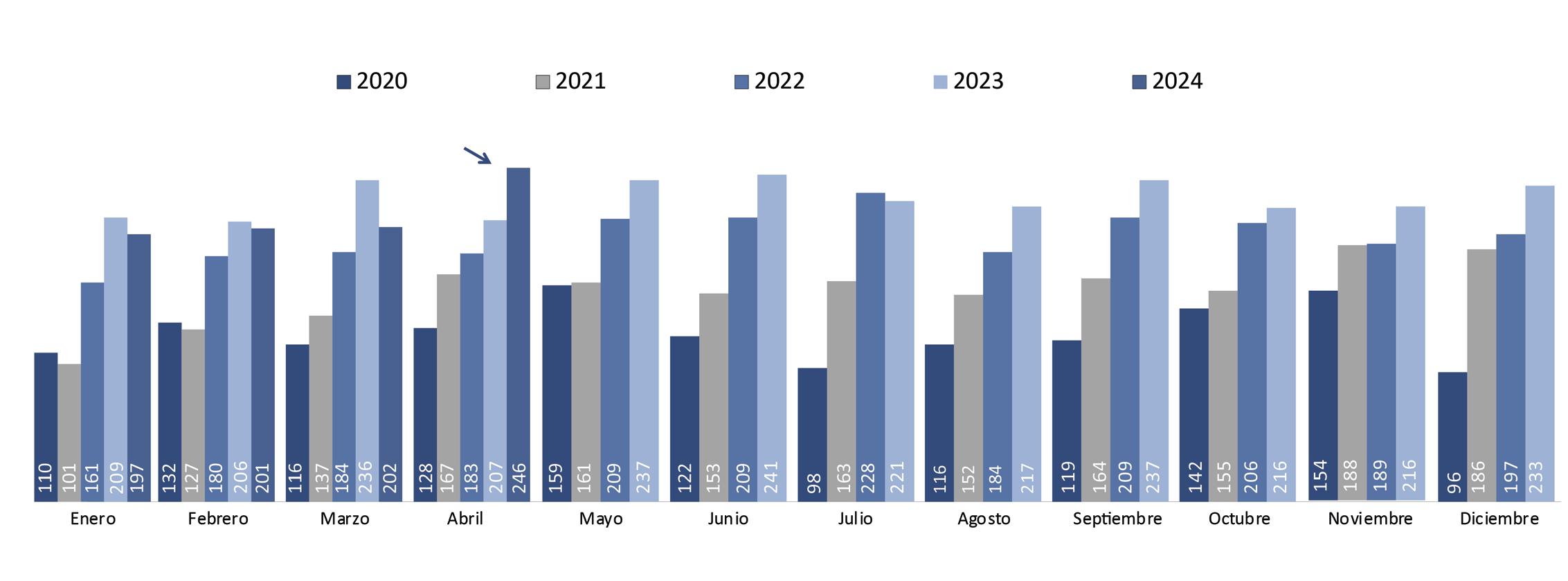
Estas restricciones provocaron que las exportaciones de camarón ecuatoriano a China tuvieran una notable caída del 27% en volumen y del 43% en generación de divisas durante el primer trimestre del 2024, según estadísticas de la Cámara Nacional de Acuacultura.
Para levantar la medida, tanto el sector público como el privado en Ecuador emprendieron todas las acciones necesarias para presentar los argumentos técnicos ante China y elaborar un plan de mejoras.
Como resultado de estos esfuerzos, los establecimientos exportadores ecuatorianos suspendidos pudieron reanudar sus envíos el 5 de junio pasado, presentando un análisis de laboratorio de cada lote, junto con el certificado sanitario correspondiente.
Es importante precisar que China y Ecuador tienen un Tratado de Libre Comercio que entró en vigencia desde el 1 de mayo de 2024, estableciendo un marco legal que busca estabilizar las relaciones comerciales y minimizar los riesgos derivados de cambios abruptos en sus políticas.
En tal sentido, José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, explicó que es prioritario acelerar el diálogo para que fluya mejor el comercio entre ambos países: "Con China hay que profundizar temas de carácter técnico, sanitario y arancelario", dijo en entrevista con RTP Radio Ecuador.
Además, el TLC con este país estipula que los productos estrella como el camarón tengan una desgravación a plazos, que en este caso, es del 0,5% anual. Camposano precisó el arancel que paga actualmente el camarón ecuatoriano y los posibles escenarios: "En 2022, China unilateralmente bajó el arancel para nuestro camarón al 2%, pero con la entrada en vigencia del TLC podría
aumentarlo al 5%, acorde con lo establecido ante la Organización Mundial del Comercio (OMC), pero con la reducción del 0,5% anual que estima el acuerdo, el arancel máximo que podría pagar el camarón ecuatoriano sería de 4,5%. Es decir, el TLC garantiza que en seis años el arancel llegará al 2% y de forma eventual hasta cero, independientemente de las decisiones de China", afirmó Camposano.
Por otra parte, el TLC también busca atraer
Autor: Shirley Suasnavas - Editora general de la Revista Aquacultura - ssuasnavas@cna-ecuador.com
SANCIONES ANTERIORES
Septiembre de 2019: empresas ecuatorianas fueron suspendidas temporalmente, luego de que la Aduana de China detectó la supuesta presencia de los virus de la Mancha Blanca y Cabeza Amarilla. La suspensión de establecimientos exportadores ecuatorianos fue levantada por el gigante asiático el 27 de noviembre de ese año.
Julio de 2020: nuevamente empresas ecuatorianas fueron suspendidas, pero en esta ocasión por detección de trazas de COVID-19 en los contenedores, no en los productos. Un mes después, en agosto de ese año, China empezó a levantar la medida verificando cada caso.
A pesar de los desafíos, existe la capacidad de superar estas situaciones y retomar las exportaciones de camarón ecuatoriano a China para mantener la estabilidad económica del sector y continuar aprovechando las oportunidades que ofrece el gigantesco mercado de ese país, sin perder cuota en su principal destino.
De enero a abril de 2024, Ecuador exportó camarones a China por un valor de $900,824.18, representando el 52% de las exportaciones totales de este crustáceo, según datos de la Cámara Nacional de Acuacultura. Este valor significó $409,660,262.97 menos, en comparación con el mismo período del año pasado.
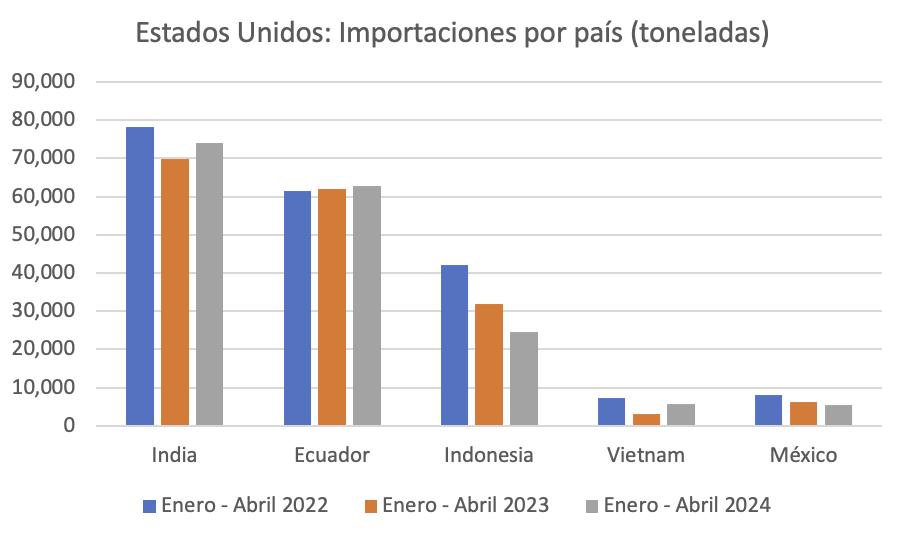
ESTADOS UNIDOS
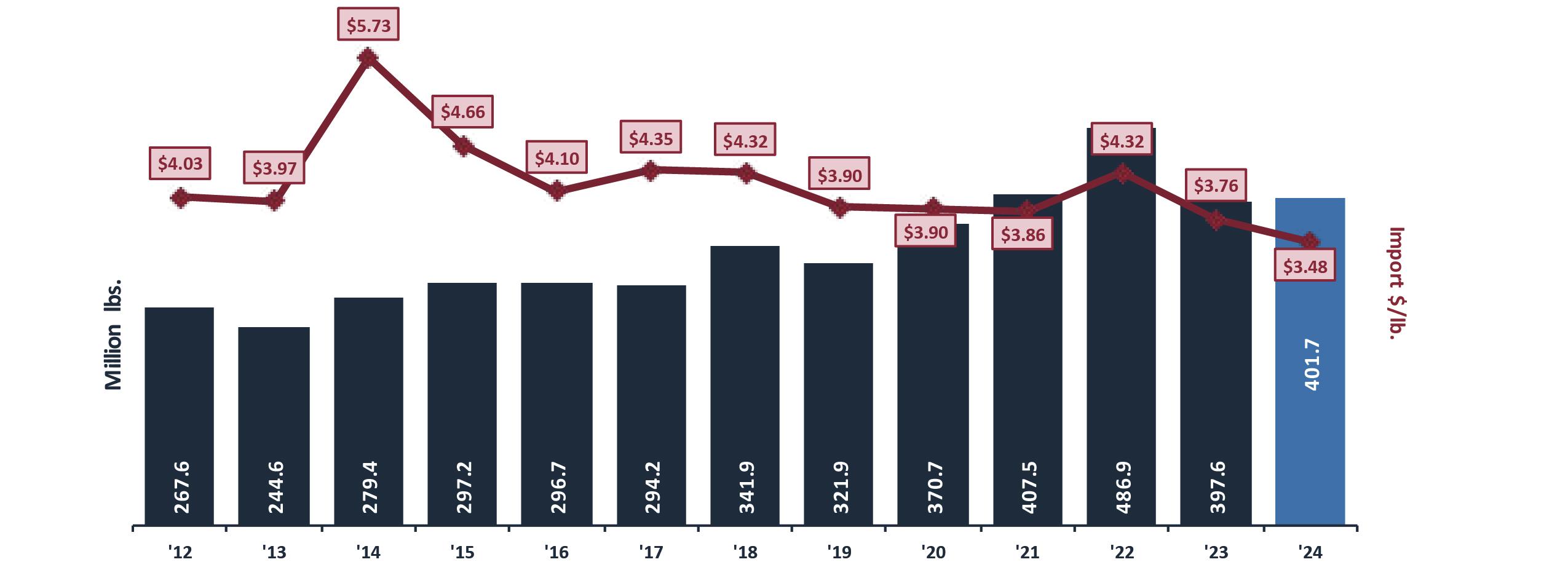
Estados Unidos es el segundo destino de exportación del camarón ecuatoriano. Este año, entre enero y abril de 2024 representó un volumen de 177067,451 libras y generó divisas por $474,573.26.
Mantenerse vigente en ese mercado se atribuye a la reconfiguración de la industria camaronera ecuatoriana, que ha implementado procesos con valor agregado para satisfacer las demandas específicas de este exigente destino.
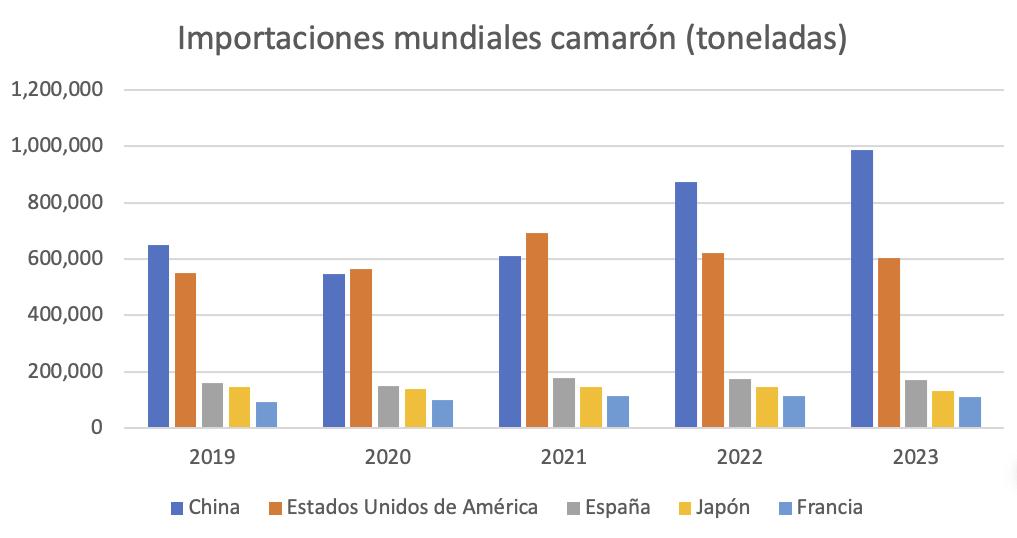
Seis años atrás, Estados Unidos estuvo posicionado como principal destino del camarón ecuatoriano, un mercado históricamente dominado por India e Indonesia. Desde 2017, Ecuador enfocó sus exportaciones hacia Europa y Asia. Como resultado de esta diversificación, Ecuador se convirtió en el segundo proveedor de camarón a Estados Unidos, asegurando una participación del 35% en el mercado. Aunque India sigue liderando con un 42%, Indonesia ha caído al tercer puesto. Actualmente, incluso el principal proveedor de camarón a EE. UU. es un establecimiento ecuatoriano.
Sin embargo, en ese destino el producto enfrenta dos demandas: una por derechos compensatorios y otra por dumping, y en ambos casos ya existen fallos preliminares.
En el marco del proceso de investigación sobre presuntas prácticas comerciales desleales relacionadas con la exportación de camarón ecuatoriano al mercado americano, el Departamento de Comercio de Estados Unidos (DOC) comunicó los resultados de su determinación preliminar por derechos antidumping (AD). El documento especifica porcentajes entre 1.58% y 10.58% para los establecimientos exportadores.
La determinación preliminar de este caso se publicó el 30 de mayo pasado en el Registro Oficial, y con ello entraron en vigencia los requisitos de depósito en efectivo.
Por otra parte, en marzo pasado, el DOC emitió una resolución inicial sobre derechos compensatorios (CVD) que fue impugnada por Ecuador, en una acción público-privada entre el Ministerio de Producción, Comercio Exterior, Inversiones y Pesca y la Cámara Nacional de Acuacultura. Ante el pedido, la autoridad comercial americana corrigió los porcentajes y presentó las nuevas tasas preliminares de depósito que tuvieron una significativa reducción, determinando un arancel preliminar de 2.89% para los establecimientos exportadores de camarón ecuatoriano.
Ante los fallos preliminares de EE. UU., José Antonio Camposano, en una entrevista con el medio televisivo Ecuavisa, señaló que el
COYUNTURA
pago de los aranceles impuestos por Estados Unidos representaría aproximadamente 15 millones de dólares mensuales para el sector exportador camaronero, debido a ambas demandas.
Es importante mencionar que Ecuador ya ha sido demandado en dos ocasiones anteriores por parte del Departamento de Comercio de Estados Unidos, casos que han sido motivados por la Asociación Estadounidense de Procesadores de Camarón, tanto en el año 2003 por antidumping y en el 2013 por medida compensatorias.
“Nuestra postura siempre ha sido la misma: el Ecuador no practica competencia desleal de ningún tipo, y por ello, en ocasiones anteriores, la Corte de Comercio Internacional de los Estados Unidos nos ha dado la razón, en el sentido en que no hemos sido sujetos de arancel en fallos definitivos, pero sí en los fallos preliminares”. Así lo expresó José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, en entrevista con el periodista Gonzalo Ruiz, en Radio Platinum en mayo pasado.
Considerando lo mencionado anteriormente, es fundamental destacar los desafíos que Ecuador ha enfrentado con Estados Unidos.
Actualmente, la CNA continúa articulando
acciones legales para defender los argumentos planteados por Ecuador en ambos casos. La determinación final tanto para el caso AD como para el caso CVD se prevé que sea recibida el martes 15 de octubre de 2024, y el anuncio de las decisiones respectivas se realizará el día siguiente.
SANCIONES ANTERIORES
2007 - EL DEPARTAMENTO DE COMERCIO RECONOCIÓ QUE ECUADOR NO COMETIÓ
DUMPING
Los ministerios de Agricultura y de Relaciones Exteriores informaron la decisión preliminar del Departamento de Comercio de EE.UU. de revocar el arancel de 3,58% impuesto al camarón ecuatoriano que entró a ese mercado por supuestas prácticas de dumping (competencia desleal).
2013 - ECUADOR GANÓ LA DEMANDA ANTE ESTADOS UNIDOS
La apelación a la decisión del Departamento de Comercio de Estados Unidos, de imponer un arancel del 11,68% al camarón ecuatoriano que ingrese a su país, dio resultados favorables ante la Comisión de Comercio Internacional (CCI).
A este panorama se suma la crisis global. Hoy en día, lamentablemente, la demanda no acompaña el importante crecimiento que tiene Ecuador. "Muchas veces se aduce que por su desarrollo, nuestro país es el responsable de la situación de precios", indica
Camposano, "pero si realmente Ecuador no creciera, lo hiciera la India más de lo que crece Indonesia. La industria camaronera se ha vuelto realmente competitiva. ¿Qué hay que hacer? Motivar el consumo".
Por su parte, la Cámara Nacional de Acuacultura continúa defendiendo los intereses del país en estas disputas comerciales, mientras Ecuador busca mantener su competitividad global en un contexto de demanda global fluctuante.
Frente a los desafíos que enfrenta el camarón ecuatoriano en sus principales mercados, como China y Estados Unidos, es fundamental destacar la capacidad de la industria para adaptarse y superar obstáculos.
La reciente suspensión de exportaciones a China y las demandas en EE.UU. han puesto a prueba la resiliencia del sector, pero también han destacado la importancia de la cooperación entre el sector público y privado para enfrentar estos retos. Con la entrada en vigor del Tratado de Libre Comercio entre China y Ecuador, así como la defensa activa en las demandas estadounidenses, la industria camaronera ecuatoriana se encamina hacia la estabilidad y el crecimiento sostenible.
Es crucial mantener el diálogo y la colaboración entre todos los actores involucrados para garantizar un futuro próspero para la acuicultura ecuatoriana•
Investigación y Mercado: Sosteniendo el liderazgo del camarón ecuatoriano
Autor:
Daniel Pesantes
Director Adjunto de la Cámara Nacional de Acuacultura dpesantes@cna-ecuador.com
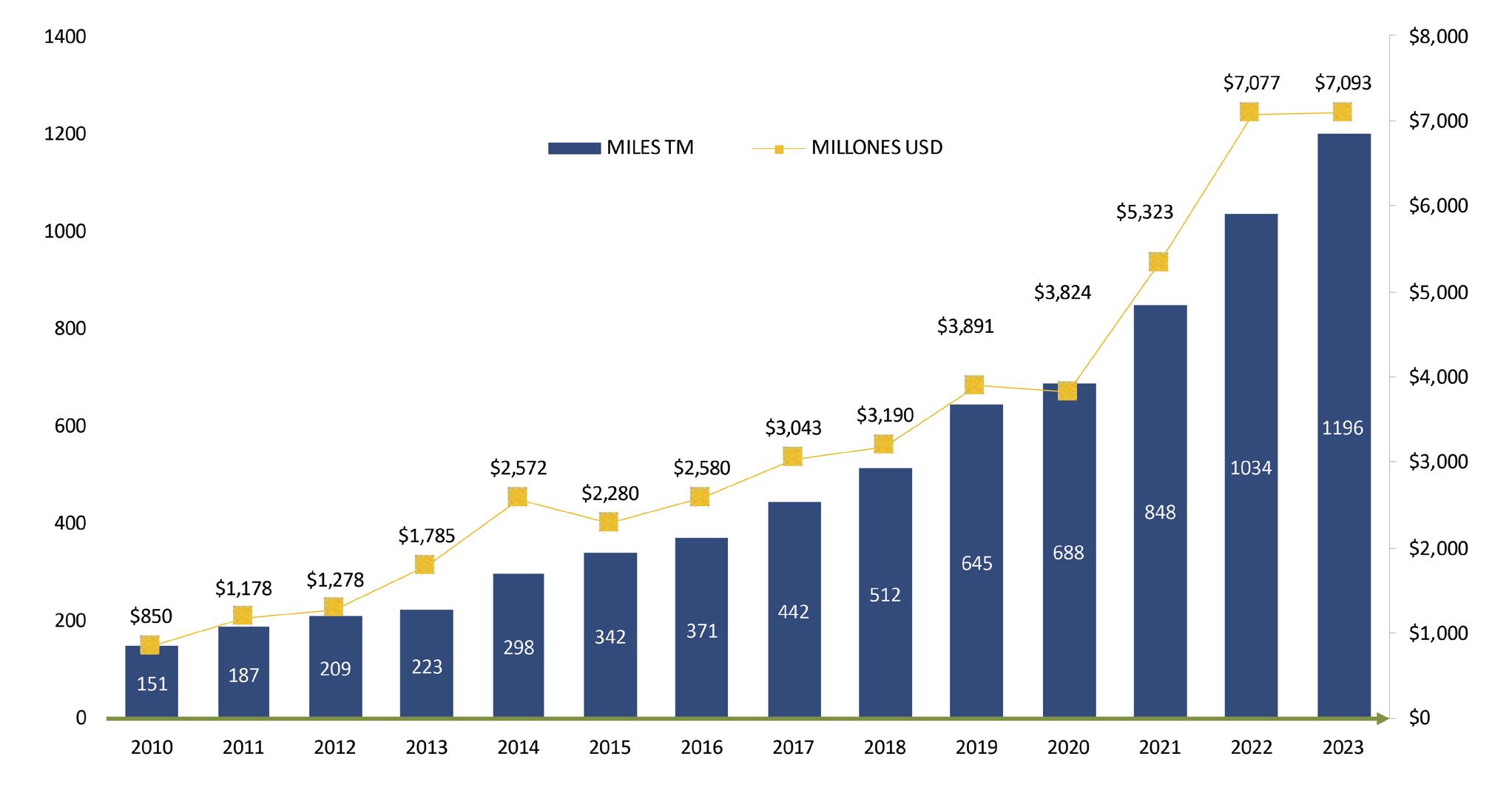
Desde 2021, el camarón ecuatoriano se ha convertido en el principal producto de exportación del país. En 2022, Ecuador se consolidó como el líder mundial en producción y exportación de camarón, alcanzando cifras impresionantes en el año siguiente, con una producción de más de 1.2 millones de toneladas y generación de divisas por encima de los 7 mil millones de dólares.
El éxito del camarón ecuatoriano radica en su capacidad para llegar a más de 60 países alrededor del mundo. Desde los vecinos de la Comunidad Andina, con lazos culturales similares, hasta regiones como el Medio Oriente y Asia, donde las distancias y las diferencias culturales son significativas.
Debido a los altos volúmenes de producción y el bajo consumo local, los mercados internacionales se han convertido en el destino natural para nuestro producto estrella. El camarón ecuatoriano se distingue por su calidad superior, por lo que es reconocido como “el mejor camarón del mundo”. Sin embargo, para acceder a diversos mercados, es crucial cumplir con una amplia gama de requisitos.
Los destinos de exportación varían, y con ellos, los requisitos de acceso. Estas exigencias incluyen barreras arancelarias, normativas sanitarias, habilitación de establecimientos, etiquetado, presentación del producto (entero, pelado, etc.), tendencias de consumo y potencial de importación. Incumplir con alguno de estos requisitos puede resultar en el rechazo del producto, o en el peor de los casos, su destrucción.
Las alertas presentadas a establecimientos procesadores ecuatorianos por parte de la Unión Europea debido a la presencia de Vibrio cholerae, o las medidas impuestas por autoridades chinas ante la presencia de metabisulfito, son ejemplos claros de barreras no arancelarias y su aplicación ante el incumplimiento de la normativa definida en cada uno de los mercados de destino.
La investigación de mercados es vital para identificar y comprender dónde y cómo colocar nuestro camarón. Antes de definir un objetivo, es esencial conocer datos básicos como las cifras de importación de camarón de diferentes países, respondiendo interrogantes como: ¿es el país un productor o depende de las importaciones?, ¿es un productor con capacidad limitada que aún requiere importaciones para satisfacer su demanda interna?. Esta información ayuda a identificar mercados potenciales donde el camarón ecuatoriano puede tener una ventaja competitiva. Otros datos relevantes incluyen el poder adquisitivo, las preferencias de consumo y las barreras arancelarias, lo que permite a los exportadores enfocar sus esfuerzos en los mercados más prometedores.
Conocer las cifras oficiales de importación de un país proporciona una visión general del tamaño del mercado y de los competidores. Analizar a la competencia y sus estrategias ayuda a los exportadores ecuatorianos a diferenciar sus productos, incluyendo aspectos como precios, calidad, presentación, estrategias de marketing y canales de distribución.
También es crucial entender las barreras arancelarias y no
COYUNTURA
arancelarias en el mercado objetivo. Un arancel elevado o competir con mercados que tienen preferencias arancelarias puede influir en la competitividad de nuestro producto. Aspectos técnicos como los requisitos sanitarios (análisis virales, límites de sustancias), inspecciones a establecimientos (fincas o plantas procesadoras), y normas de etiquetado y presentación son fundamentales.
Con el objetivo de proporcionar información y herramientas para la toma de decisiones en la identificación de mercados de exportación y conocer la situación de los diferentes mercados competidores, la Cámara Nacional de Acuacultura (CNA) ha desarrollado nuevos productos y servicios de información para sus usuarios. Estos incluyen boletines de noticias sobre tendencias en mercados productores, exportadores, consumidores e importadores de camarón; reportes mensuales de importaciones y exportaciones de los mercados más importantes, e informes de exportaciones ecuatorianas y fichas de requisitos de acceso a mercados.
La investigación y diversificación de mercados son pilares fundamentales para el éxito y sostenibilidad de las exportaciones de camarón en Ecuador. Estas estrategias no solo permiten identificar y aprovechar oportunidades en el mercado global, sino que también mitigan riesgos y aseguran una posición competitiva a largo plazo. Al continuar invirtiendo en estas áreas, nuestro país puede mantener y fortalecer su liderazgo en la industria camaronera, asegurando el crecimiento económico y el bienestar de sus comunidades.
Para mantener el liderazgo mundial del camarón ecuatoriano, es vital invertir en investigación y diversificación de mercados. Esto implica cumplir con requisitos arancelarios y sanitarios, y analizar tendencias de consumo y poder adquisitivo. La Cámara Nacional de Acuacultura continuará proporcionando herramientas e información a exportadores, como boletines y reportes. También es esencial adaptarse a normativas internacionales para asegurar calidad y seguridad del producto. Estas estrategias permiten aprovechar oportunidades globales y mitigar riesgos, asegurando competitividad y crecimiento económico sostenible del sector camaronero ecuatoriano•
Una mirada a la industria del camarón en Guatemala
Autor: Yahira Piedrahita
Directora Ejecutiva de la Cámara Nacional de Acuacultura
ypiedrahita@cna-ecuador.com
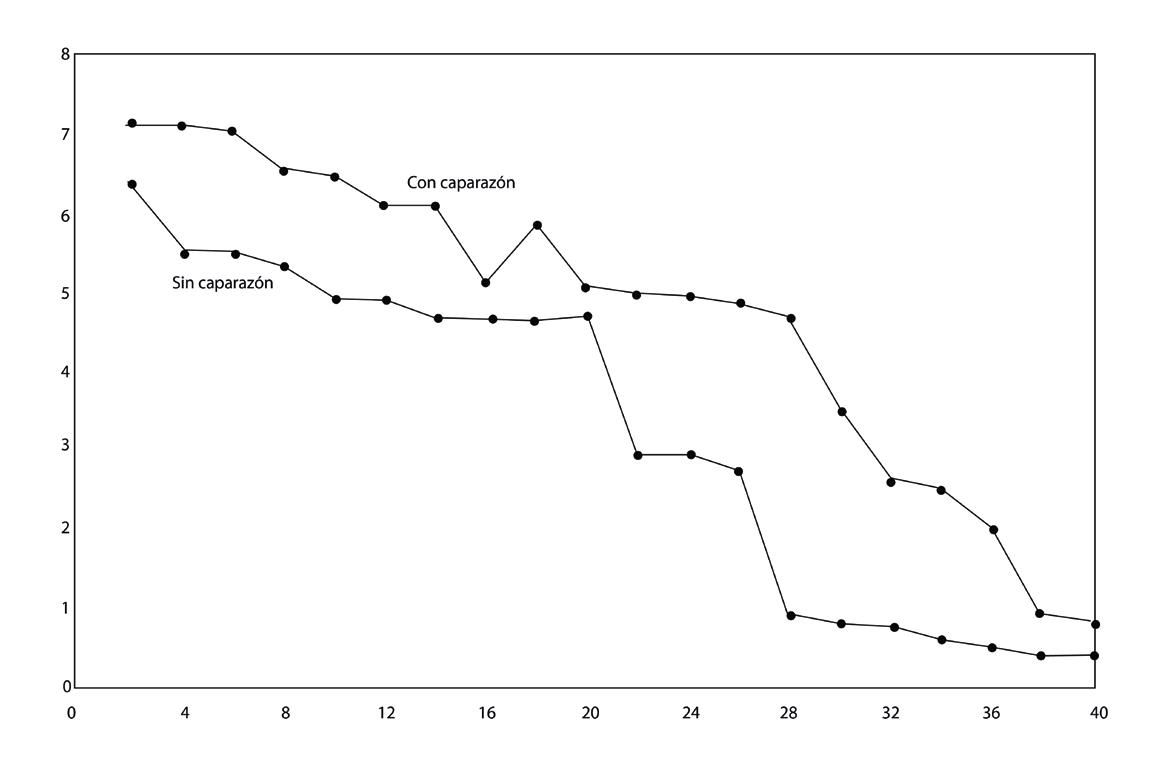
Guatemala inició el cultivo de camarón en 1983, originalmente con un sistema de producción similar al de Ecuador y México. Sin embargo, el acceso limitado a zonas costeras para la construcción de infraestructura camaronera y el alto costo de la tierra (que compite con actividades como turismo, cultivo de caña de azúcar y otros) hicieron que a partir del año 2000 el modelo de producción se tornara más intensivo, con estanques pequeños y densidades de cultivo más altas que los demás países productores de la región. La industria de camarón en Guatemala es descrita por sus actores como altamente innovadora, tecnificada y sostenible, cuyos modelos de manejo están dirigidos a la producción intensiva, alcanzando productividades de talla comercial de hasta 60,000 libras/ha/ciclo de 60 días.
Durante su evolución, la industria guatemalteca se ha visto expuesta a terremotos, erupciones volcánicas, inundaciones y problemas socioeconómicos que han afectado al país. También los patógenos han impactado en diversa medida a los cultivos, y en los últimos años aún se enfrentan a enfermedades ocasionados por el virus de la Mancha Blanca (WSSV), infecciones por Vibrios, microsporidios, entre otros patógenos.
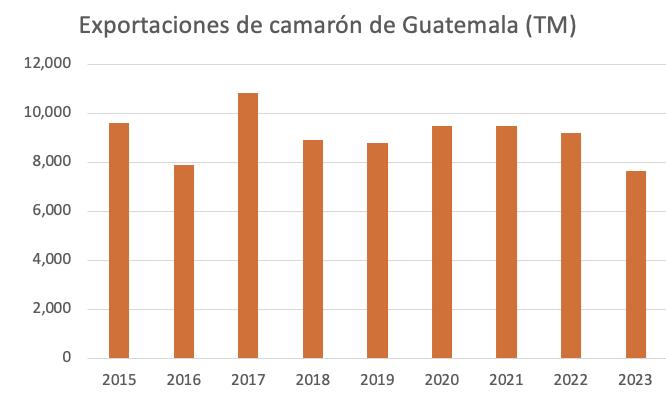
Las exportaciones anuales de Guatemala bordean las 10,000 toneladas y sus principales mercados son Estados Unidos, la Unión Europea y México. La producción de camarón genera alrededor de 15 mil puestos de trabajo directos 1 Sin embargo, en los últimos años las densidades de cultivo se han reducido para poder adaptarse a las circunstancias del mercado, los brotes de enfermedades y los altos costos de producción.
A pesar de ello, los productores se esfuerzan por salir adelante en un mercado de precios bajos y los efectos de la suspensión temporal para el ingreso de su producto a México, derivada de una orden judicial emitida en octubre del 2023, implementada desde enero de 2024 y que finalmente lograron revertir hacia finales de mayo de este año, por lo que al momento, la frontera sur mexicana se ha reabierto al paso del camarón de Guatemala. El gobierno busca abrir el
mercado brasileño a las exportaciones de camarón, por lo que delegados del gigante sudamericano analizarán el sistema de inspección de productos de la pesca y las normativas legales vigentes que regulan al sector2
Durante el Tercer Simposio de Acuicultura realizado el 22 y 23 de mayo por la Asociación Guatemalteca de Exportadores (AGEXPORT), expertos nacionales e internacionales, empresarios y líderes de la industria de la acuicultura se reunieron en Antigua Guatemala para explorar las oportunidades y desafíos actuales que esta industria enfrenta a nivel global.
Para Gabriel Biguria, presidente de AGEXPORT y CEO de Acuamaya, una de las mayores empresas acuícolas del país, la crisis de Guatemala no es muy diferente a la de los demás países productores de camarón.
Para el empresario, en la actualidad es importante entender los límites que la naturaleza impone a los sistemas de cultivo, determinando la capacidad de carga de los estanques para poder optimizar las estrategias productivas mediante el análisis de la información y el manejo de precisión.
Esto se pudo constatar durante la visita a Mayasal, la granja que opera Acuamaya, la misma que cuenta con tres áreas de estanques que producen bajo diferentes densidades y sistemas de alimentación, con una zona de manejo intensivo con estanques pequeños en los que siembran a densidades de 80-120 animales/m2 y en donde la biomasa puede llegar a 2.5 kg/m2 (un equivalente a 25 ton/ha).
“Todos los productores de camarón estamos en la misma tormenta, pero en barcos diferentes, por lo que cada país y empresa debe implementar estrategias acordes a su realidad”
Mencionó Biguria quién indicó además que hay que aprovechar esta crisis como una gran oportunidad para fortalecer a la industria, implementando estrategias de colaboración, resaltando los atributos del camarón como
Figura 1. Exportaciones de camarón de Guatemala (en toneladas) entre 2015 y 2023. Fuente: Trade Data Monitor
una proteína de alta calidad y promoviendo su consumo tanto en el mercado local como internacional.
Reconoce el gran trabajo de Ecuador promoviendo su camarón en los mercados internacionales, ejemplo que debe ser
replicado por los demás productores, y resaltando que “todos debemos convertirnos en embajadores del camarón a nivel mundial”•
de Acuicultura realizado el 22 y 23 de mayo por la Asociación Guatemalteca de Exportadores (AGEXPORT).
Visita a camaronera Mayasal, en el marco del Tercer Simposio
Las zonas francas: una oportunidad para la exportación del camarón ecuatoriano
Autor:
Viviana Vásquez Moreira
Parinpro S.A. – Zona Franca Progreso (Santa Elena – Ecuador)
viviana.vasquez@zedeprogreso.com
COYUNTURA
La aprobación de la Ley Orgánica de Eficiencia Económica y Generación de Empleo por parte de la Asamblea Nacional en diciembre de 2023 ha levantado muchas expectativas en relación con las inversiones que realizan los sectores productivos (o que tienen la intención de realizar). Este interés gira en torno a los esquemas especiales de atracción de inversiones a través de la aplicación de incentivos fiscales, como es el caso del nuevo régimen de zonas francas.
Apreciaciones sobre la estructura especial tributaria han sido vertidas desde antes de la promulgación de la ley en la que está contenida, debido a las experiencias que han tenido otros países de la región latinoamericana. Por ejemplo República Dominicana, con exportaciones que salen de sus zonas francas que sobrepasan el 60% de las exportaciones totales del país; Panamá, país que triplica las exportaciones totales nacionales con aquellas que salen desde sus zonas francas, y Uruguay, cuyo componente de exportaciones desde zonas francas representa más del 30% del total nacional, son ejemplos destacados de cómo la estabilidad de un régimen fiscal contribuye a la sostenibilidad de la inversión, la generación de plazas de trabajo y al crecimiento continuo de los flujos de producción exportable. Son naciones en las que este régimen tiene varias décadas de aplicación y cuyas legislaciones han inspirado a la reciente ley aprobada en el país, adoptando sus conceptos de mejores prácticas internacionales para el perfilamiento de un instrumento legal innovador, versátil y ajustado adecuadamente a la realidad del comercio internacional.
Los incentivos fiscales que trae consigo el nuevo régimen de zonas francas constituyen un atractivo para todos los sectores productivos, tanto exportadores como importadores, y ahora también para los prestadores de diversos servicios, por la adición de este espectro de actividades económicas que se incluyó en la ley para su desarrollo dentro de zonas francas.
En este contexto, se destaca la actividad que de manera expresa detalla el artículo 37 del Código Orgánico de la Producción, en la categoría industrial de bienes. Nos
COYUNTURA
referimos al procesamiento de productos pesqueros o acuícolas, actividad en la que tiene participación estelar nuestro camarón.
Revisemos a continuación, algunos de los aspectos más relevantes.
1. Exenciones e incentivos fiscales. El nuevo régimen de zonas francas ofrece en su paquete de incentivos los siguientes:
a) Exoneración del Impuesto a la Salida de Divisas por todo el tiempo de calificación dentro de la zona franca (Art. 50.3 Código Orgánico de la Producción COPCI).
b) Exoneración del Impuesto a la Renta por los 5 primeros años de la calificación, y una tarifa reducida de este impuesto por el tiempo restante de la misma y sus prórrogas (la calificación dentro de zona franca tiene un plazo de 30 años prorrogables), (Art. 50.3 COPCI).
c) Exoneración de los impuestos al comercio exterior (aranceles, IVA de importación, FODINFA) por todo el tiempo que dure su calificación en la zona franca (Art. 34 COPCI).
d) Devolución de IVA de las compras locales de bienes empleados en el proceso productivo (Art. 50.4 COPCI).
e) Exención del Impuesto a la Renta sobre el pago de dividendos a los accionistas o socios de las empresas que se califican dentro de una zona franca (Art. 50.6 COPCI).
2. nfraestructura e instalaciones. La amplitud del paquete fiscal de incentivos permite que el interesado en instalarse en una zona franca cuente con las condiciones necesarias para establecerse y tenga los elementos de infraestructura apropiados, sea por la construcción a medida de galpones y edificios, o por la importación sin pago de tributos al comercio exterior de toda la maquinaria y equipos requeridos para el desarrollo de su actividad (Art. 50.4 COPCI).
3. Procedimientos aduaneros simplificados. Las zonas francas cuentan con procedimientos aduaneros simplificados, lo que reduce el tiempo y tramitología necesarios para la exportación. Esto constituye un apoyo a los exportadores de camarón a llevar sus productos a los mercados internacionales de manera más rápida y eficiente (Art. 50.18).
4. Acceso a los mercados internacionales. Las zonas francas son naturalmente centros para el comercio internacional, atrayendo compradores e inversores extranjeros. Esto brinda a los exportadores de camarón de nuestro país acceso a nuevos mercados y oportunidades comerciales (Art. 35 COPCI).
5. Promoción de la competitividad. Las zonas francas son espacios que propician el establecimiento de clústeres, en donde se van a agrupar industrias similares y aquellas que pertenecen a su cadena de valor. Así, las zonas francas promueven la competitividad a través de economías de escala, intercambio de conocimientos y colaboración. Todos estos elementos contribuyen al mejoramiento de su eficiencia, calidad y competitividad en el mercado global (Art. 35 COPCI).
Los elementos antedichos proveen un marco con enorme potencial para brindar un escenario positivo para la industria camaronera, por cuanto las ventajas expuestas se traducen en una importante reducción de costos para la cadena de procesamiento del camarón. Desde el transporte, la limpieza, clasificación y procesamiento, mantenimiento en la cadena de frío, empacado, control de calidad, distribución y exportación, son susceptibles de recibir los beneficios de los incentivos fiscales.
Es preciso enfocar el ejercicio de complementación que se concreta entre la aplicación de los incentivos fiscales y ventajas
de las zonas francas con los acuerdos comerciales que ha suscrito nuestro país, entre los que destacamos el convenio con la Unión Europea y el reciente tratado de comercio con China, cuya ejecución arrancó a inicios del mes de mayo.
Los acuerdos comerciales facilitan el acceso a los mercados, al reducir los aranceles y las barreras comerciales entre países. Por tanto, se convierte en un elemento crucial el poder contar con las exenciones fiscales detalladas en párrafos anteriores, y acceder a los mercados internacionales principales de exportación con preferencias o exoneraciones de la carga impositiva de comercio exterior en los mercados de destino. Todo esto generaría un crecimiento exponencial a la competitividad del sector. La combinación citada comporta, sin duda, un atractivo adicional para los inversores nacionales o extranjeros para establecer negocios en las zonas francas.
En consecuencia, los incentivos fiscales que ofrecen las zonas francas complementan los beneficios de los acuerdos comerciales, armando un frente potente para ser aprovechado por el sector camaronero.
En general, las zonas francas desempeñan un papel crucial para brindar un mayor impulso al mercado de exportación de camarón del país. El aprovechamiento estratégico de las ventajas que ofrecen, hará que Ecuador siga construyendo un posicionamiento líder en el mercado mundial del camarón•
Pamela Nath Directora
Una frase común entre los expertos en sostenibilidad es “lo que no se mide, no se puede gestionar”. Solo cuando somos conscientes de un problema, podemos comenzar a abordarlo de manera efectiva. En la actualidad, el mundo enfrenta desafíos críticos en materia de sostenibilidad que requieren una acción inmediata para garantizar el bienestar de las sociedades presentes y futuras. El cambio climático, la pérdida de biodiversidad y el respeto a los derechos humanos son solo algunos de los retos que debemos afrontar.
En este contexto, existe una herramienta muy valiosa que permite a las empresas demostrar su compromiso para contribuir al desarrollo sostenible: la memoria de sostenibilidad. Mediante este instrumento, las organizaciones de todo el mundo, incluídas las de la industria acuícola de camarón, podrán identificar oportunidades de mejora para tener un mejor desempeño en su actividad. Además, a través de este reporte se evidenciarán los esfuerzos y las buenas prácticas que las empresas implementan para tener impactos positivos en su entorno.
Todas las entidades, desde el individuo hasta la corporación, deben ser conscientes de que son parte de una sociedad y que comparten un mismo planeta. Las gestiones que realizan a diario generan impactos positivos y negativos que no se limitan únicamente a las operaciones comerciales, sino que también afectan al medio ambiente y a la sociedad en general. Por lo tanto, las organizaciones no solo deben buscar resultados financieros, sino también sociales y ambientales.
Esta visión consciente implica entender que la sostenibilidad es una oportunidad para innovar, crear valor a largo plazo y contribuir al bienestar global. Pero, sobre todo, implica adoptar la responsabilidad y la transparencia como valores que guían las operaciones empresariales y que permiten brindar información y gestionar los efectos de su actividad en favor de las personas y del mundo.
Memoria de sostenibilidad: conoce la herramienta que te ayudará a medir tus impactos
Autores:
Pamela Nath
Sally Tabares
Sustainable Shrimp Partnership (SSP)
pnath@sustainableshrimp.org
Cada vez más personas consideran el desempeño económico, ambiental y social de las empresas al decidir si invertir en ellas o no. Según la encuesta ‘Global Consumer Insights Pulse Survey’ de PwC del 2023, los consumidores están colocando su dinero donde están sus valores. Cuatro de cada cinco encuestados expresan disposición a pagar más por productos fabricados de manera sostenible, siendo la Generación Z y los millennials los más inclinados a aceptar un precio más alto por la sostenibilidad.
Asimismo, el estudio “Las demandas de sostenibilidad de los consumidores están en aumento”, publicado en Harvard Business Review, señala tres factores clave que están impulsando un cambio significativo en los hábitos de consumo, donde la sostenibilidad se está convirtiendo en un requisito esencial para las compras. Primero, la confianza impulsa el comportamiento del consumidor y, en consecuencia, también los resultados comerciales. Segundo, la sostenibilidad fomenta la confianza, especialmente entre las generaciones más jóvenes. Tercero, estas generaciones son las que pronto tendrán la mayor parte del poder adquisitivo.
Los autores añaden que las empresas que entiendan estas tendencias y desarrollen marcas verdaderamente sostenibles, cumpliendo con sus compromisos hacia las personas y el planeta, tendrán una ventaja sobre aquellas que hacen afirmaciones débiles o no han invertido lo suficiente en sostenibilidad.
¿Qué es una memoria de sostenibilidad? Concepto y beneficios “Normalmente, las organizaciones han estado acostumbradas a llevar a cabo informes de su gestión centrados en brindar resultados a nivel financiero. Los reportes de sostenibilidad, más allá de presentar un informe de lo que la organización hace a diario, permiten comunicar acerca del impacto que una empresa produce sobre el medio ambiente y la sociedad, incluidos los impactos que afectan a los derechos humanos”, explica la experta en Comunicación y Sostenibilidad, Susana Chávez.
La memoria de sostenibilidad, también llamada reporte de sostenibilidad, es un documento que sirve como herramienta para transparentar, comunicar y gestionar el desempeño económico, ambiental y social que tiene una empresa. Al desarrollar este informe, la compañía puede identificar su contribución, positiva o negativa, al desarrollo sostenible.
Chávez, quien es parte de la consultora de responsabilidad social empresarial Con3xión, explica que el reporte de sostenibilidad va dirigido a los grupos de interés de la organización, tales como clientes, proveedores y colaboradores. Les comunica los efectos que la empresa produce sobre ellos y cómo la empresa busca maximizar los impactos positivos y reducir o mitigar los negativos.
Hay varias metodologías de informes que se pueden seguir al elaborar una memoria de sostenibilidad. Aunque en Ecuador estos informes son voluntarios y su contenido depende del nivel de transparencia que una empresa esté dispuesta a adoptar, existen estándares que especifican los contenidos. Entre los principales se encuentran el Global Reporting Initiative (GRI), Sustainability Accounting Standards Board (SASB), Carbon Disclosure Project (CDP), entre otros.
“Cuando elaboras un informe de sostenibilidad basado en un estándar, uno de los beneficios es que te expones a las mejores prácticas. Estos estándares son reconocidos a nivel mundial y se mantienen al tanto de las tendencias y temas importantes a escala global. Esto significa que los indicadores que elijas medirán aspectos relevantes y de actualidad a nivel internacional”, comparte Chávez.
La consultora explica que el proceso de elaborar un informe de sostenibilidad va más allá de recopilar datos y presentarlos en una narrativa. Durante la recolección de información, es probable que se identifiquen deficiencias, ya sea porque los datos no están disponibles, no se ajustan a los estándares, no se desean comunicar, o porque los resultados no son los esperados. El desarrollo del informe permite identificar áreas de mejora, tomar medidas correctivas y establecer metas.
Entre los beneficios para la gestión interna de las organizaciones, Con3xión Consultores destaca que la memoria de sostenibilidad permite determinar riesgos e impactos económicos, sociales y ambientales; sirve como una herramienta de medición, monitoreo y gestión de ASG (Ambiental, Social y de Gobernanza); promueve un proceso de mejora continua, genera sentido de pertenencia y compromiso de los colaboradores; identifica oportunidades para la reducción de costos, y estimula la innovación y la creación de nuevas oportunidades de negocio.
Además, los reportes de sostenibilidad también son útiles en la gestión de las relaciones con los públicos externos de la empresa. Entre los beneficios que resalta la consultora se encuentran: atraer financiamiento y acceso a capital, mejorar la imagen y reputación frente a los grupos de interés, incrementar la lealtad y compromiso de los clientes, reducir el impacto negativo en caso de crisis, responder a las expectativas de comunicación de los stakeholders y crear una ventaja competitiva.
Tendencia de los reportes de sostenibilidad Actualmente, las organizaciones se sienten motivadas a elaborar memorias de sostenibilidad por varios factores. Según Chávez, uno de ellos es la creciente conciencia empresarial sobre la sostenibilidad. El propósito y la visión del negocio impulsan a estas empresas a ser sostenibles y reponsables con sus grupos de interés. Esto conlleva un deseo de transparentar sus acciones con dichos grupos. Además, las instituciones reconocen la importancia de informar a su público sobre sus iniciativas, ya que la manera en que gestionan y comunican estas acciones impacta directamente en su reputación.
Por otro lado, existe una tendencia en crecimiento sobre las regulaciones que obligan a elaborar reportes de sostenibilidad. De acuerdo con el reporte “Beyond Disclosure in ESG and Sustainability Policy” de Carrots & Sticks de 2023, en los últimos 3 años hubo un aumento del 296% en las políticas y regulaciones sobre sostenibilidad corporativa que registran en su base de datos. Además, indica que 113 países, un equivalente al 58% de naciones en el mundo,
ya tienen normativas y regulaciones sobre sostenibilidad corporativa.
Un caso relevante es el de la Unión Europea (UE), donde la ley exige a grandes empresas como a pymes cotizadas que informen sobre lo que consideran los riesgos y oportunidades derivados de cuestiones sociales y ambientales, al igual que sobre el impacto de sus actividades en las personas y el medio ambiente. Algunas empresas no pertenecientes a la UE también deberán informar si generan más de 150 millones de euros en el mercado europeo.
“Los accionistas muestran cada vez más interés no solo en evaluar los resultados financieros, sino también en considerar los riesgos y oportunidades ambientales, sociales y de gobernanza que enfrentan las organizaciones. Por ello, la normativa está comenzando a abordar esta preocupación”, expresa Chávez.
Según la Encuesta de Informes de Sostenibilidad de KPMG de 2022, aunque en ese año la presentación de informes de sostenibilidad se mantuvo principalmente en marcos voluntarios, se espera un cambio en los próximos años. La encuesta indica que “las regulaciones regionales y nacionales sobre la presentación de informes están evolucionando rápidamente, por lo que las empresas deberían comenzar a prepararse para la obligatoriedad de estos informes”
En el 2022, el 96% de las 250 empresas más grandes del mundo informaron sobre su desempeño en sostenibilidad o en temas ambientales, sociales y de gobernanza, donde el 73% de ellas utilizaron los Estándares GRI.
GRI y el estándar sectorial que involucra a la acuicultura
Desde 1997, el Global Reporting Initiative (GRI) ha sido el estándar más ampliamente utilizado en todo el mundo para la elaboración de memorias de sostenibilidad. Actualmente, Latinoamérica es la región de mayor uso de este estándar, según KPMG.
El GRI permite a cualquier organización, grande o pequeña, privada o pública, comprender y presentar información sobre
sus impactos en la economía, el medio ambiente y las personas, incluidos los derechos humanos, así como la forma en la que gestionan estos impactos.
Al emplear estos estándares, las empresas pueden facilitar comparaciones de rendimiento entre compañías distintas y permitir que los inversores y otros interesados tomen decisiones bien informadas. Este enfoque promueve la transparencia, la credibilidad y la comparación en diversas industrias y regiones.
Cada tres años, la Junta de Estándares Globales de Sostenibilidad (GSSB, por sus siglas en inglés) crea un nuevo programa de trabajo para mantener actualizados los estándares GRI. Este plan incluye proyectos para revisar los estándares actuales y también para desarrollar los nuevos. Dentro de este marco está el Programa Sectorial, que se enfoca en desarrollar estándares específicos para 40 sectores diferentes, comenzando por los que tienen el mayor impacto.
Desde el 1 de enero del 2024, entró en efecto el estándar sectorial de Agricultura, Acuicultura y Pesca: el GRI 13.
“Las actividades de este sector tienen impactos ampliamente documentados en las dimensiones económica, medioambiental y
social, incluidos los efectos sobre el cambio climático, la biodiversidad, los derechos humanos, la pobreza y la desigualdad. Estos sectores suministran productos alimentarios y no alimentarios esenciales, cuya demanda se prevé que siga creciendo en el futuro. Por lo anterior, el desarrollo de este estándar es fundamental para avanzar en los esfuerzos del sector en contribuir al desarrollo sostenible”, explica la directora de GRI Latinoamérica, Andrea Pradilla.
Asimismo, Pradilla informa que el GRI 13 ofrece beneficios tanto internos como externos para la industria acuícola. Internamente, permite a las organizaciones rastrear y documentar el origen y destino de sus productos, evitando la entrada de productos de pesca ilegal y no reglamentada. Esto ayuda a combatir la sobrepesca y promover prácticas sostenibles. Además, informar sobre prácticas ambientales ayuda a abordar riesgos como los escapes de explotaciones acuícolas, preservando los ecosistemas marinos.
Externamente, el estándar promueve la divulgación de información sobre prácticas laborales y sociales, como remuneración justa y condiciones de trabajo seguras, fortaleciendo la confianza de los empleados y la sociedad. También impulsa la transparencia en la salud y bienestar animal, lo que mejora la imagen de la industria entre los consumidores preocupados por este factor.
“Además, promueve la conciencia sobre los miembros de la cadena de suministro, sus prácticas y su impacto, contribuyendo así a una mayor seguridad alimentaria y a la disponibilidad de alimentos de alta calidad”, detalla la directora.
El estándar también se centra en proporcionar las “cinco libertades” del bienestar animal: ausencia de hambre y sed; ausencia de incomodidad, ausencia de dolor, lesiones y enfermedad; libertad para expresar comportamientos normales, y ausencia de miedo y angustia. “Este aspecto está estrechamente relacionado con los beneficios anteriores, ya que fomenta una mayor conciencia sobre el trato ético hacia los animales”, añade.
Pradilla sugiere que las empresas de la industria camaronera que estén pensando en elaborar una memoria de sostenibilidad siguiendo el estándar de GRI, empiecen lo antes posible. “No hay que esperar a tener toda la información o los resultados positivos que quisieran evidenciarse, sino atreverse a medir y reportar de manera transparente los impactos positivos y negativos y beneficiarse de ese proceso de transparencia”, indica.
Los desafíos de reportar impactos
1. La cultura de silencio
Durante los años que Con3xión Consultores lleva desarrollando memorias de sostenibilidad bajo el estándar GRI, Chávez ha identificado que entre los principales
desafíos para las empresas se encuentra la cultura de silencio. “Este reto es uno de los más difíciles de trabajar”, reconoce, “porque supone enfrentar el miedo a exponerse. Normalmente, ha existido una tradición de no hablar sobre ciertos asuntos, sean estos impactos que las organizaciones sí están midiendo y gestionando, pero que, generalmente, se trabajan a un alto nivel y que nadie dentro de la organización menciona”
Según Chávez, muchos creen que al comenzar a comunicar sobre sus impactos, sus grupos de interés reaccionarán con preocupación al notar las repercusiones. Sin embargo, aclara que es importante reconocer que todos generan impactos negativos. “Esta realidad es común a todas las organizaciones y sectores, y aunque algunos puedan tener impactos más significativos que otros, es fundamental entender que todos los tenemos”, expresa.
2. El compromiso
El compromiso es otro de los desafíos que menciona la consultora. “Muchas veces, el área encargada de llevar adelante un informe de sostenibilidad es un departamento específico dentro de la empresa. Sin el compromiso de la alta dirección, el proyecto no progresará”, expone Chávez.
En la misma línea, la directora del GRI Latinoamérica comenta: “La capacidad empresarial de medirse y reportar en sostenibilidad debe ser construida, y esto es un proceso en el que es necesario que la alta dirección se involucre y establezca la visión y el seguimiento, y que desde allí se destinen los recursos humanos y financieros para generar dicha capacidad”.
3. Diálogos con los grupos de interés, ¿por qué son importantes?
Al redactar un informe de sostenibilidad basado en el estándar GRI, es esencial mantener conversaciones con los grupos de interés. De acuerdo con Chávez, el reto de algunas empresas es realizar estos diálogos a conciencia y entenderlos como una herramienta valiosa y una oportunidad para el negocio, no solo como un paso a seguir.
Estas conversaciones permiten conocer las preocupaciones, necesidades y expectativas
que tienen los stakeholders respecto a la empresa. Así se puede establecer la materialidad, es decir, los asuntos relevantes tanto para el grupo de interés como para la organización.
Como el reporte de sostenibilidad va dirigido a ellos, se deben comprender las principales preocupaciones de estos grupos y reflejarlas en los informes, junto con las acciones que la empresa está llevando a cabo para abordarlas. La información que se recibe sirve también para elaborar estrategias, programas y planes a futuro.
Ruta para desarrollar una memoria de sostenibilidad
El proceso de elaboración de un informe de sostenibilidad bajo el estándar GRI implica varios pasos esenciales, informa Chávez. En primer lugar, es importante identificar a los stakeholders clave de la organización, aquellos grupos de interés más relevantes. Esto permite entender a quién se dirige el informe y qué información es crucial para ellos.
Posteriormente, se deben llevar a cabo diálogos con estos stakeholders para comprender sus principales preocupaciones y expectativas respecto a la organización. Una vez recopiladas estas perspectivas, es importante realizar un análisis de materialidad para identificar los temas más relevantes y significativos tanto para la organización como para sus grupos de interés. Con esta base, se procede al desarrollo de la memoria
de sostenibilidad, siguiendo la estructura y principios establecidos en el estándar GRI.
Finalmente, la comunicación del informe es fundamental. No basta con enviar el documento; se debe trabajar en un plan de comunicación para asegurar que la información llegue de manera efectiva a todos los grupos de interés pertinentes. Esta etapa es vital para garantizar que el trabajo realizado sea conocido, comprendido y valorado por todos los involucrados.
Casos de empresas ecuatorianas en la elaboración de memorias de sostenibilidad
“Estos reportes han sido la principal herramienta para evolucionar la gestión de sostenibilidad de Vitapro”, expone María Alejandra Rivera, quien desde 2019 es la gerente de Desarrollo Sostenible de esta empresa de alimento balanceado para acuicultura. Gracias a la implementación de la memoria de sostenibilidad en 2017, hoy en día Vitapro cuenta con programas, equipos de trabajo, objetivos cuantitativos y comités dedicados a la sostenibilidad y a su avance.
Lo que comenzó como un proyecto liderado por Rivera y un grupo de consultores externos hace más de siete años, en la actualidad ha logrado involucrar a más de 200 personas de Vitapro en iniciativas de sostenibilidad.
“Cada vez es más necesario poder comunicar de manera clara el progreso, las estrategias, la perspectiva y el entendimiento que las empresas tienen sobre la sostenibilidad. Las memorias de sostenibilidad son importantes porque permiten este ejercicio de transparencia y movilizan a los equipos internos. Son una herramienta de medición por excelencia, en donde se encuentra la mayor parte de la información sobre sostenibilidad de una empresa, lo que facilita su gestión, porque ningún equipo desea encontrarse en la misma situación dos años consecutivos”, añade Rivera.
Por otro lado, Sustainable Shrimp Partnership (SSP) está actualmente desarrollando la primera edición de su memoria de sostenibilidad, que cuenta con la participación de sus trece miembros.
“Al ser una de las plataformas precompetitivas más relevantes a nivel mundial en la industria de mariscos, para nosotros es fundamental elaborar nuestra primera memoria de sostenibilidad. No
solamente para demostrar el cumplimiento de las mejores prácticas en un lenguaje universal, sino también para incentivar a que las empresas en Ecuador, en los diferentes eslabones de la cadena de valor de la industria camaronera, puedan también reportar estos avances y compromiso con la sostenibilidad”, expone la directora de SSP, Pamela Nath.
“Reconocemos que muchas compañías ya están trabajando en numerosas iniciativas de responsabilidad social, pero creemos que
es esencial fomentar el uso de herramientas oficiales y universales para evidenciar este compromiso. Buscamos que estos reportes sean más que un documento bien presentado, sino que sean una herramienta útil para medir la evolución de las empresas en sostenibilidad”, afirma•
Para más información sobre este artículo, contactar a: pnath@sustainableshrimp.org
ARTÍCULOS TÉCNICOS ÍNDICE
Edición 159 - Junio 2024
Vibriosis en la producción de camarón
Análisis de la disparidad de talla y su relación con el IHHNV en la larvicultura de camarón P. vannamei
Supervivencia de cepas de Vibrio cholerae 01 en camarones sujetos a procesos de congelación y ebullición
¿Cuál es el panorama global de alimentos para el segundo semestre del 2024?
Caracterización de dinoflagelados en un sector de producción de camarón en el golfo de Guayaquil – Ecuador, como un paso inicial a la toma de medidas fitosanitarias
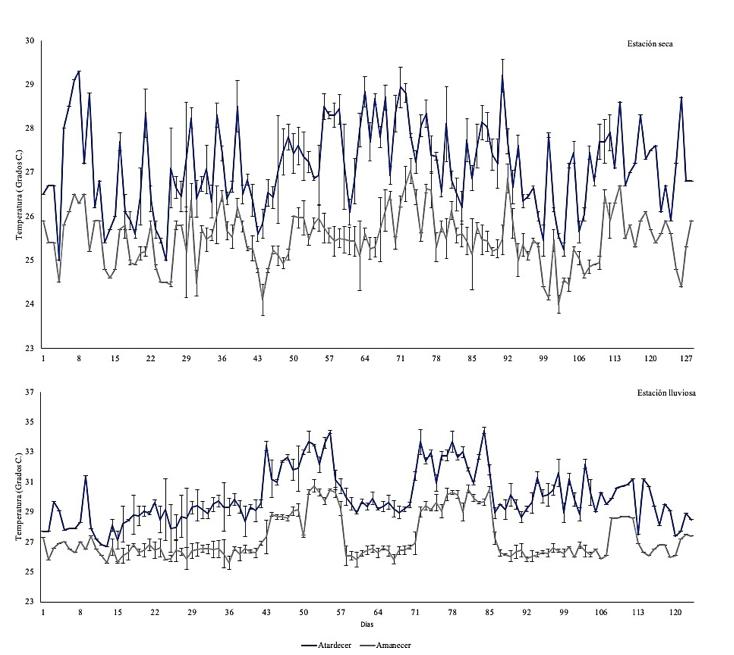
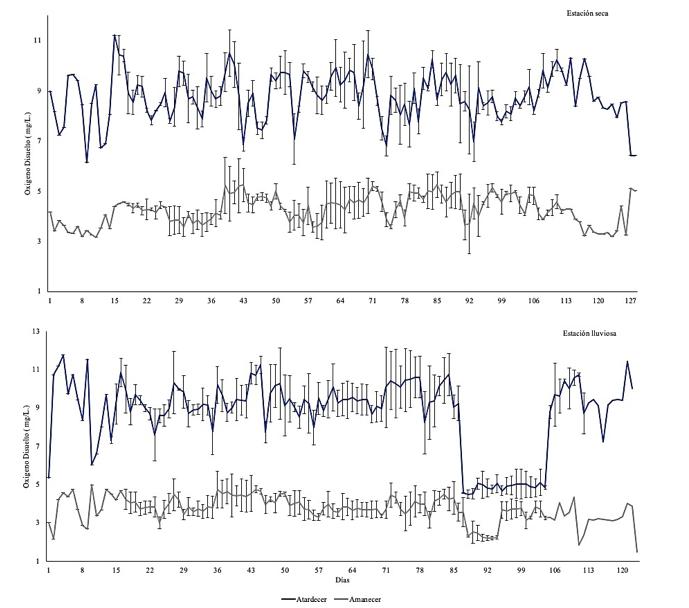
Efecto de la variación temporal de la temperatura sobre la producción del camarón Penaeus vannamei en sistemas de cultivo en piscinas
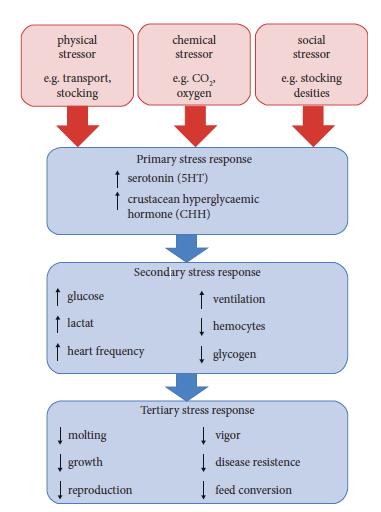
Bienestar de crustáceos decápodos con especial énfasis en la fisiología del estrés
Niveles de sulfito en langostinos y camarones congelados
Ante el inminente cambio climático: ¿Cuáles son las prácticas a implementar en la acuicultura?
Vibriosis en la producción de camarón
Autor:
A. Venkateswara Rao
dravrao@neospark.co.in
La acuicultura es el sector alimentario de más rápido crecimiento a nivel mundial, y se ha consolidado como un recurso de alto valor proteico para satisfacer la demanda de alimentos debido a la sobreexplotación de recursos naturales. Pero en la actualidad, el mayor problema que enfrenta la industria acuícola en todo el mundo son las enfermedades causadas por diversos agentes biológicos y no biológicos.
Entre los grupos de microorganismos que causan graves pérdidas en el cultivo de camarón, los más conocidos son las bacterias, debido a los devastadores efectos económicos que tienen en las granjas afectadas. Se han reportado enfermedades bacterianas, principalmente debidas a Vibrio, en sistemas de producción de peneidos que involucran al menos 14 especies, como: Vibrio harveyi, V. splendidus, V. parahaemolyticus, V. alginolyticus, V. anguillarum, V. vulnificus, V. campbelli, V. fischeri, V. damsella, V. pelagicus, V. orientalis, V. ordalii, V. mediterrani, V. logei, etc.
La vibriosis es una de las afectaciones más preocupantes en la producción de mariscos y peces. Se trata de una enfermedad bacteriana responsable de la mortalidad del camarón que se produce en todo el mundo (Lightner y Lewis, 1975; Adams, 1991; Lightner et al., 1992; Lavilla-Pitogo et al., 1996; Lavilla-Pitogo et al., 1998; Chen et al., 2000), ya que las especies de Vibrio están ampliamente distribuidas en las instalaciones de cultivo del planeta. Las infecciones relacionadas con Vibrio ocurren con frecuencia en los laboratorios, pero las epizootias también se presentan comúnmente en las especies de camarón cultivadas en estanques. La vibriosis es causada por bacterias gramnegativas de la familia Vibrionaceae
Los brotes pueden ocurrir cuando los factores ambientales desencadenan una rápida multiplicación de bacterias que son toleradas en bajos niveles dentro de la hemolinfa del camarón (Sizemore y Davis, 1985), o por penetración bacteriana de las barreras del huésped. El exoesqueleto proporciona una barrera física eficaz para los patógenos que intentan penetrar la superficie externa de los crustáceos, así como el intestino anterior y posterior. Sin
embargo, Vibrio spp. se encuentran entre las bacterias quitinoclásticas asociadas con la enfermedad de la concha (Cook y Lofton, 1973), y pueden entrar a través de heridas en el exoesqueleto o poros (Jiravanichpaisal y Miyazaki, 1994; Alday-Sanz et al., 2002). Las branquias pueden parecer susceptibles a la penetración bacteriana porque están cubiertas por un exoesqueleto delgado (Taylor y Taylor, 1992), pero sus superficies son limpiadas por las “setobranquias” (Bauer, 1998).
El intestino medio, compuesto por la glándula digestiva (DG) y el tronco del intestino medio (MGT, a menudo denominado intestino, véase Lovett y Felder, 1990), no está revestido por un exoesqueleto y, por lo tanto, parece ser un sitio probable para la penetración de patógenos transportados en el agua, alimentos y sedimentos (Ruby et al., 1980; Jayabalan et al., 1982).
Vibrio harveyi, una bacteria luminosa y gramnegativa, es uno de los importantes agentes etiológicos de las mortalidades masivas en los sistemas de cultivo de larvas de Penaeus monodon. Un gran número de criaderos (más de 280) a lo largo de la costa de India, dedicados a la producción de semilla de camarón, a menudo sufren reveses debido a la enfermedad bacteriana luminiscente y sufren enormes pérdidas económicas.
Entre los aislamientos de Vibrio harveyi, algunos son virulentos y otros no, lo que sugiere una gran variación molecular y genética en este grupo de bacterias. El mecanismo patogénico también se ha atribuido recientemente a los bacteriófagos.
La vibriosis se extiende en todo el mundo y en todos los crustáceos marinos, incluyendo los camarones, que son más susceptibles. Las epizootias ocurren en todas las etapas de la vida, pero son más comunes en los laboratorios. Se han reportado epizootias importantes de vibriosis para P. monodon de la región del Indo-Pacífico, P. japonicus de Japón y P. vannamei de Ecuador, Perú, Colombia y América Central (Lightner, 1996). La vibriosis se manifiesta a través de una serie de síntomas, entre los que se incluyen: vibriosis oral y entérica, vibriosis cuticular y de apéndices, vibriosis localizada de
heridas, enfermedad del caparazón, vibriosis sistémica y hepatopancreatitis séptica (Lightner, 1996).
La vibriosis es causada por varias especies de bacterias Vibrio, entre las que se incluyen V. harveyi, V. vulnificus, V. parahaemolyticus, V. alginolyticus y V. penaeicida (Brock y Lightner, 1990; Ishimaru et al., 1995). Se han reportado casos ocasionales de vibriosis causada por V. damsela, V. fluvialis y otras especies indefinidas de Vibrio (Lightner, 1996).
Las especies de Vibrio forman parte de la microflora natural en los camarones silvestres y cultivados (Sinderman, 1990) y se convierten en patógenos oportunistas cuando se suprimen los mecanismos naturales de defensa (Brock y Lightner, 1990). Generalmente se asocian a múltiples agentes etiológicos. Sin embargo, algunas especies de Vibrio, o cepas de ciertas especies, han sido identificadas como patógenos primarios (Owens y Hall-Mendelin, 1989; Owens et al, 1992; Lavilla-Pitogo et al., 1990; de la Peña et al., 1995). Las cepas patógenas de V. harveyi, V. vulnificus y V. parahaemolyticus han causado epidemias masivas en Tailandia (Nash et al., 1992) y Filipinas (Lavilla-Pitogo et al., 1990). V. harveyi luminiscente parece liberar exotoxinas (Liu et al., 1996) y puede causar una mortalidad del 80-100% en los criaderos de P. monodon (Harris, 1995). También se ha reportado de la asociación entre V. anguillarum, V. campbelli, V. nereis, V. cholerae (no 01) y V. splendidus con brotes de enfermedades en camarón (Chen 1992; Lavilla-Pitoga, 1990; Esteve y Quijada, 1993; Sahul-Hameed et al., 1996). Tatsuya Nakayama et al. (2005) investigaron la relación entre la luminiscencia y la toxicidad de Vibrio carchariae en camarón.
Según Jayasree. L. et al. (2006), la aparición de cinco tipos de enfermedades: necrosis de la cola, enfermedad de la cáscara, enfermedad roja, síndrome de la Cáscara Suelta (LSS) y enfermedad del Intestino Blanco (WGD) es causada por Vibrio spp. en Penaeus monodon de estanques de cultivo de la costa de Andhra Pradesh. Entre estas, LSS, WGD y enfermedad roja causaron mortalidades masivas en estanques de cultivo de camarón. Seis especies de Vibrio—V. harveyi, V. parahaemolyticus, V.
alginolyticus, V. anguillarum, V. vulnificus y V. splendidus—están asociadas con camarones enfermos.
La distribución y composición de especies de bacterias luminosas en laboratorios comerciales de camarones peneidos fueron estudiadas por Jawahar Abraham. T y R. Palaniappan (2004). La observación sobre la presencia de V. harveyi (97.30%) y V. orientalis (2.70%) en el contenido intestinal del camarón demostró que la fuente primaria de estas bacterias en un laboratorio de camarón era la materia fecal de los reproductores, posiblemente en el momento del desove.
Signos clínicos
La mortalidad por vibriosis ocurre cuando los camarones se encuentran estresados por factores como: mala calidad del agua, hacinamiento, alta temperatura del agua, bajo DO y bajo recambio de agua (Lewis, 1973; Lightner y Lewis, 1975; Brock y Lightner, 1990). Generalmente, se producen altas tasas de mortalidad en postlarvas y juveniles de camarón.
Las larvas de P. monodon sufrieron mortalidades dentro de las 48 horas posteriores al ensayo de inmersión con cepas de V. harveyi y V. splendidus (Lavilla-Pitogo, et al., 1990). Se han reportado mortalidades que involucran vibriosis en P. monodon de tamaño comercial (Anderson et al., 1988). Los camarones adultos que sufren vibriosis pueden presentar hipoxia, enrojecimiento del cuerpo con branquias de color rojo a marrón, reducir la alimentación y se los puede observar nadando letárgicamente en los bordes y la superficie de los estanques (Anderson et al., 1988; Nash et al., 1992). Vibrio spp. también causa la enfermedad de las patas rojas, caracterizada por la coloración roja de los pleópodos, periópodos y branquias, en camarones desde jóvenes hasta adultos y puede causar una mortalidad de hasta el 95% durante la estación cálida (Chen, 1992). La enfermedad de necrosis de los globos oculares es causada por V. cholerae. Los globos oculares de los camarones infectados se vuelven marrones y se caen, y la mortalidad ocurre en unos pocos días (Chen, 1992).
Seis especies de Vibrio, que incluye a
V. harveyi y V. splendidus, producen luminiscencia, que es fácilmente visible por la noche, en postlarvas infectadas, juveniles y adultos (Ruby et al., 1980; Lightner, et al., 1992). Las postlarvas infectadas también pueden presentar una motilidad reducida, fototaxismo reducido e intestinos vacíos (Chen, 1992).
Patología macroscópica
Los camarones que sufren vibriosis pueden presentar lesiones localizadas en la cutícula típicas de la enfermedad bacteriana del caparazón o “concha”, infecciones localizadas por heridas punzantes, pérdida de extremidades, musculatura turbia, infección localizada del intestino o hepatopáncreas y/o septicemia general (Lightner, 1993). Las lesiones de la enfermedad bacteriana de la concha son marrones o negras y aparecen en la cutícula del cuerpo, los apéndices o las branquias (Sinderman, 1990).
Las postlarvas afectadas pueden presentar hepatopáncreas turbios (Takahashi et al., 1985). Las branquias suelen ser de color marrón (Anderson et al., 1988). La hepatopancreatitis séptica se caracteriza por la atrofia del hepatopáncreas con necrosis multifocal e inflamación hemocítica.
La presencia de altas cantidades de Vibrio parahaemolyticus o V. harveyi indujo el redondeo y desprendimiento de las células epiteliales de la lámina basal del MGT. No se observó desprendimiento de células epiteliales en presencia de bacterias no patógenas (probióticos) (Chen et al., 2000; Gary G. Martin et al., 2004).
Los patógenos como Vibrio spp., que causan el desprendimiento del epitelio en el MGT, pueden producir una alta mortalidad en el camarón al eliminar dos capas que protegen a los camarones de las infecciones: el epitelio y la membrana peritrófica que secreta. Además, la pérdida de epitelio puede afectar la regulación de la absorción de agua e iones en el cuerpo (Mykles 1977, Neufeld y Cameron 1994).
Histopatología
La vibriosis sistémica generalmente produce la formación de nódulos hemocíticos sépticos en el órgano linfoide, el corazón y los tejidos conectivos de las branquias,
el hepatopáncreas, la glándula antenal, el cordón nervioso, el telson y el músculo (Anderson et al., 1988; Mohney et al., 1991; Jiravanichpaisal et al., 1994). Los hepatopancreocitos infectados pueden aparecer poco vacuolados, lo que indica bajas reservas de lípidos y glucógeno (Anderson et al., 1988). La vibriosis en P. monodon está asociada con la formación de “esferoides” en el órgano linfoide (Nash et al., 1992).
Diagnóstico
El diagnóstico de la infección por Vibrio se basa en los signos clínicos y la demostración histológica de la presencia de bacterias Vibrio en forma de bastón en lesiones, nódulos o hemolinfa. Los órganos extirpados y la hemolinfa pueden ponerse en una placa de agar marino general o selectivo para Vibrio (TCBS). Cuando se investigan las postlarvas, se puede triturar el animal entero y luego ponerse en una placa de agar. Se pueden observar colonias luminiscentes después de 12 a 18 horas si se incuban a temperatura ambiente o a una temperatura de 25 a 30 °C.
Los aislamientos de Vibrio pueden identificarse mediante diversos métodos, entre ellos: tinción de Gram, motilidad, prueba de oxidasa, modo de utilización de la glucosa, crecimiento en presencia de NaCl, reducción de nitrato y luminiscencia. Las especies de Vibrio pueden identificarse rápidamente en el campo utilizando el sistema API-20 NFT, que implica el cultivo de colonias de Vibrio en tiras API-NFT y la puntuación de las colonias de acuerdo con las instrucciones del kit (Lightner, 1996) o BIOLOG (un sistema de identificación bacteriana miniaturizado que es una alternativa al sistema API). Se pueden utilizar pruebas de sensibilidad
antimicrobiana para identificar la vibriosis y se pueden implementar utilizando el método del disco Kirby-Bauer (DIFCO, 1986) o el método de concentración inhibitoria mínima (CIM) (Lightner, 1996).
Transmisión
Las especies de Vibrio existen en el agua utilizada en las instalaciones de cultivo de camarón (Lavilla-Pitogo et al., 1990) y en la biofilm, que se forma en diferentes estructuras de contacto con el agua de los laboratorios y las granjas. Las bacterias entran en los camarones a través de heridas o grietas en la cutícula y son ingeridas con los alimentos (Paynter 1989; Lavilla-Pitogo et al., 1990). La fuente principal de V. harveyi en los laboratorios parece ser el contenido del intestino medio de los reproductores hembra, que se desprende durante el desove (Lavilla-Pitogo et al., 1992).
Viabilidad
Se han realizado numerosos estudios sobre el efecto de la congelación en los Vibrio que contaminan los mariscos cosechados. V. vulnificus en la ostra cosechada (Crassostrea virginica) sobrevivió al almacenamiento a -20 °C durante 70 días (Parker et al., 1994). V. parahaemolyticus, aislado de homogeneizados de carne de ostra, se inactivó en 16 días a -15 °C cuando la carga bacteriana era muy alta (10 cfu/gm; Muntada-Garriga et al., 1995). Hay evidencia reciente que sugiere que V. harveyi puede sobrevivir en sedimentos de estanques incluso después de cloración o tratamiento con cal (Karunasagar et al., 1996).
Estado actual de la vibriosis
La vibriosis es un problema común en todo el mundo, particularmente en la India. V. harveyi continúa causando mortalidades crónicas de hasta un 30% entre larvas, postlarvas
Luminescent bacteria (Vibrio harveyi)
y adultos de P. monodon en condiciones estresantes. También está surgiendo una cepa altamente patógena de Vibrio spp. que continúa causando mortalidades entre el camarón cultivado (Le Groumellec et al., 1996). Los problemas causados por la vibriosis secundaria son comunes, pero se consideran menores en comparación con las epidemias virales.
Tratamiento
La vibriosis se controla mediante un manejo riguroso del agua y saneamiento para prevenir la entrada de vibrio en el agua de cultivo (Baticados et al., 1990) y para reducir el estrés en los camarones (Lightner, 1993). La selección del sitio, el diseño y la preparación del estanque también son importantes (Nash et al., 1992). Se recomienda aumentar los recambios diarios de agua y reducir la biomasa del estanque mediante la cosecha parcial para reducir la mortalidad causada por la vibriosis. También se recomienda drenar, secar y administrar cal/dolomita a los estanques después de la cosecha (Anderson et al., 1988).
La vibriosis luminiscente se puede controlar en el criadero, lavando los huevos con yodo
y formaldehído y evitando la contaminación con heces de los reproductores. El V. harveyi en la columna de agua se puede inactivar con dióxido de cloro. Los probióticos se administran directamente en el agua o a través de los alimentos. Los inmunoestimulantes también han tenido éxito en la reducción de la mortalidad de los camarones asociada con la vibriosis (Itami, 1996).
Jiravanichpaisal y Chuaychuwong et al. (1997) reportaron sobre el uso de Lactobacillus spp. como bacteria probiótica en el camarón tigre gigante (P. monodon). Diseñaron un estudio para investigar un tratamiento eficaz de Lactobacillus spp. contra la vibriosis y las enfermedades de la Mancha Blanca en P. monodon. Investigaron el crecimiento de algunas bacterias probióticas y su supervivencia en agua de mar a 20 ppm durante al menos 7 días. Se determinó la actividad inhibidora de dos Lactobacillus spp. contra Vibrio sp., E. coli y Staphylococcus spp.
Nakayama T. et al. (2007) investigaron el efecto de la concentración de cobre en la expresión tanto de la luminiscencia como
de la toxina de V. harveyi. Encontraron que una concentración de cobre de menos de 40 ppm no tenía efecto en el crecimiento del camarón. Mientras que V. harveyi cultivado con una concentración de cobre de 40 ppm mostró una disminución en la luminiscencia. Por lo tanto, la combinación de prebióticos, probióticos, inmunoestimulantes y sustancias no antibióticas tiene una especificidad superior contra la vibriosis y la bacteria luminiscente (LB) junto con buenas prácticas acuícolas (BAP, por sus siglas en inglés), lo que la convierte en una herramienta de gestión eficaz para el control de la toxicidad bacteriana por luminiscencia en la acuicultura•
Para más información sobre este artículo, contactar a: dravrao@neospark.co.in
Análisis de la disparidad de talla y su relación con el IHHNV en la larvicultura de camarón P. vannamei
Autores:
Sebastián Alejandro Ramos Rojas
Leandro Stalin Bajaña Zambrano
Clara Esperanza Lozano Farias
René Orlando Molina Barreto
Mariuxi Jacqueline Mirabá Guerrero
Luis Fernando Aranguren Caro
sebastian.ramos@limbomar.com.ec
El virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa (IHHNV), clasificado como Penstyldensovirus (PstDV1) (Penzes JJ et al., 2020), es una preocupación importante para la industria camaronera debido a las pérdidas económicas significativas que ha ocasionado históricamente (Lightner, 2011). El IHHNV afecta crónicamente a los camarones Penaeus vannamei, causando el síndrome de Deformidad y Enanismo (RDS) (Kalagayan et al., 1991; Primavera & Quinitio, 2000), caracterizado por un retardo en el crecimiento y deformidades cuticulares, lo que genera serias afectaciones relacionadas con el valor comercial al cosechar.
El método de contagio de este virus se da horizontalmente en el agua y verticalmente de generación en generación a través de la transmisión transovárica, es decir, desde la madre al huevo. Este estudio busca determinar si hay correlación entre la presencia de IHHNV y la disparidad de tallas en camarones P. vannamei durante un período de 6 meses en el año 2023.
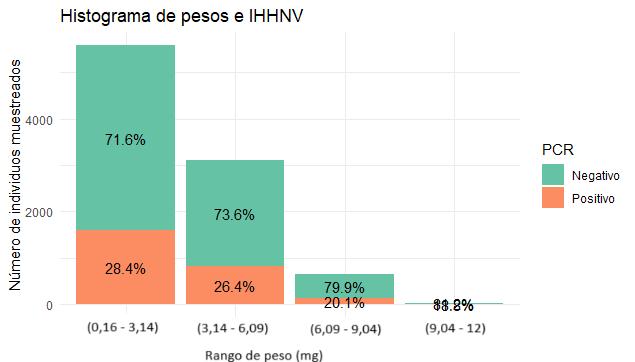
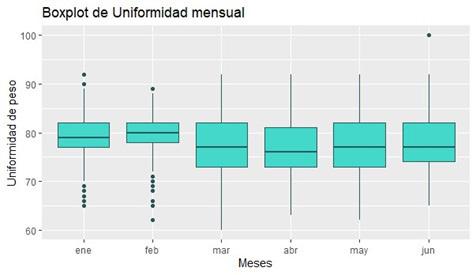
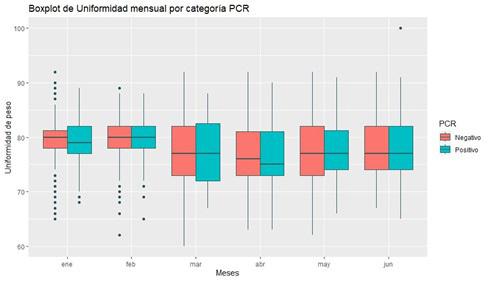
Se determinó la prevalencia de IHHNV mediante PCR en tiempo real (qPCR) en un laboratorio de producción de larvas de camarón durante el 2023. La disparidad de larvas se evaluó desde PL3 a PL14, utilizando el software para estimar peso y uniformidad de las muestras.
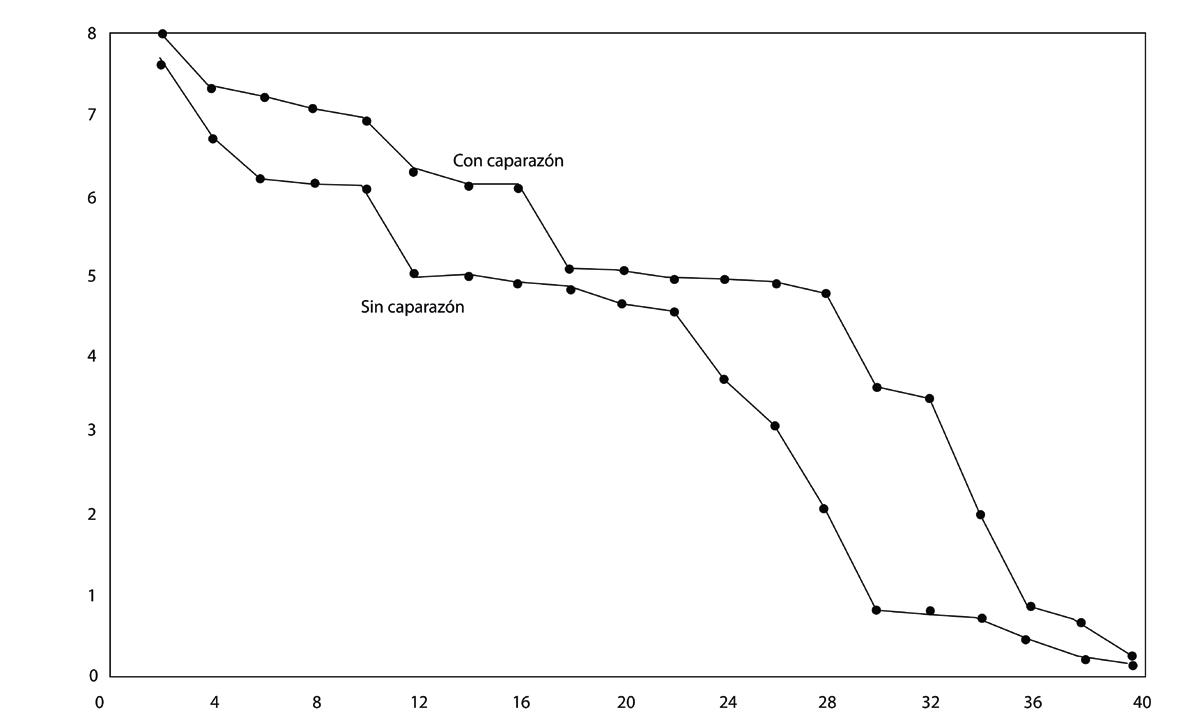
Los datos se analizaron utilizando el software R en RStudio, con el que se observó que los meses con mayor prevalencia fueron enero (28%), mayo (25%), y junio (33%), sin encontrar diferencias significativas en la uniformidad entre muestras positivas y negativas para IHHNV, ni en los rangos de pesos.
A pesar de las variaciones en la prevalencia de la enfermedad en diferentes meses, los resultados no mostraron correlación entre los casos positivos y la disparidad de tamaño. Se realizó un análisis de varianza (ANOVA), que no reveló diferencias significativas en la uniformidad entre muestras positivas y negativas para IHHNV.
Este estudio no encontró relación entre la presencia de IHHNV y la disparidad de tallas en camarones P. vannamei. Estos resultados
contribuyen a la comprensión detallada de la distribución de pesos y la uniformidad a lo largo del tiempo.
El virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa (IHHNV), clasificado como Penstyldensovirus (PstDV1), representa una amenaza significativa para la industria camaronera debido a las pérdidas económicas y productivas que ha ocasionado históricamente. El IHHNV afecta crónicamente a los camarones Penaeus vannamei, causando el síndrome de Deformidad y Enanismo (RDS), caracterizado por un retardo en el crecimiento y deformidades cuticulares que afectan negativamente su valor comercial al momento de la cosecha (Kalagayan et al., 1991; Primavera & Quinitio, 2000).
Este virus pertenece a la familia Parvoviridae y es altamente prevalente en poblaciones de camarones en cultivo. La infección por IHHNV se transmite de dos maneras principales: horizontalmente a través del agua contaminada, donde los camarones entran en contacto con partículas virales liberadas por individuos infectados, y verticalmente de generación en generación mediante la transmisión transovárica, donde las hembras infectadas transmiten el virus a sus descendientes a través de los óvulos (Hasson et al., 1995; Lightner, 2011).
La transmisión vertical del IHHNV es particularmente preocupante debido a su capacidad para persistir en poblaciones de camarones a lo largo de múltiples generaciones, incluso en ausencia de fuentes externas de infección. Esta modalidad de transmisión contribuye significativamente a la propagación y mantenimiento del virus en granjas acuícolas, representando un desafío continuo para la salud y productividad de las poblaciones de camarones cultivados.
Además de la transmisión directa entre individuos infectados, existe una preocupación adicional relacionada con las secuencias no infecciosas de ADN relacionadas con el virus, conocidas como elementos virales endógenos no-retrovirales (EVE). Estas secuencias pueden estar insertadas en el genoma de los camarones, lo que plantea desafíos en el diagnóstico preciso del IHHNV debido a la posibilidad de
falsos positivos (Tang et al., 2003; Tang & Lightner, 2006; Tang et al., 2007).
Los EVE de IHHNV insertados en el genoma de camarones P. monodon han sido ampliamente documentados (Tang et al., 2003; Krabsetve et al., 2004; Tang & Lightner, 2006; Tang et al., 2007). Estas inserciones genómicas son de suma importancia para el diagnóstico, ya que pueden afectar la sensibilidad y especificidad de las técnicas utilizadas para detectar la presencia del virus. La detección precisa del IHHNV es esencial para el comercio internacional de camarón cultivado, dado que este virus está catalogado como un patógeno de reporte obligatorio según la Organización Mundial de Sanidad Animal (OMSA/OIE), y el diagnóstico adecuado es requerido para la exportación e importación de camarones cultivados. Es fundamental garantizar la precisión en la detección de este virus para evitar impactos económicos y sanitarios en la industria camaronera a nivel mundial.
Dado el impacto del IHHNV en la producción de camarones peneidos y la existencia de áreas libres de este virus, es imperativo desarrollar protocolos de PCR específicos para la detección de EVE en P. vannamei Actualmente, los protocolos de PCR más utilizados provienen de estudios en otros hospederos y pueden no ser totalmente sensibles ni específicos para P. vannamei. La OIE enfatiza la importancia de implementar técnicas de diagnóstico avanzadas y sensibles en el sector de cultivo de camarón, lo que incluye la secuenciación del producto de PCR para identificar posibles falsos positivos y mejorar la detección precisa del IHHNV y sus variantes endógenas en camarones cultivados.
El objetivo principal de este estudio fue investigar la posible relación entre la presencia del IHHNV y la disparidad de tallas en larvas de camarón P. vannamei durante un período de seis meses en el año 2023 (enero a junio). La disparidad de tallas, que puede reflejar diferencias en el crecimiento y desarrollo de las larvas, se evaluó meticulosamente desde PL3 a PL14 utilizando el software para estimar peso y uniformidad de las muestras.
La comprensión de cómo la presencia del IHHNV puede influir en la disparidad de tallas
en larvas de camarón es crucial para mejorar las estrategias de manejo de enfermedades en la larvicultura, lo que a su vez podría impactar positivamente en la rentabilidad y sostenibilidad de la industria camaronera. En este contexto, este estudio busca aportar evidencia científica relevante que contribuya a esta comprensión detallada y a la formulación de medidas efectivas para el control y prevención del IHHNV en sistemas de cultivo de camarones.
Materiales y métodos
El muestreo se llevó a cabo en un laboratorio de producción de larvas de camarón. Para obtener los datos de uniformidad de peso para verificar las tallas en la muestra se utilizó una aplicación desde el estadio de PL2 hasta su cosecha (PL8-PL14). Se realizaron mediciones en 700 puntos de muestreo, lo que generó un total de 9300 datos sobre la distribución y uniformidad de peso.
Detección del virus
El método de detección del virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa (IHHNV) se basó en la técnica de PCR en tiempo real (qPCR) utilizando los cebadores IHHNV1608F/IHHNV1608R desarrollados por Tang y Lightner en 2001 y recomendados por la OMSA (OMSA, 2023). En total se analizaron 1133 muestras, las cuales se obtuvieron de los tanques que se encontraban en producción; estas larvas se encontraban en estadio de PL 6 y para realizar la extracción de material genético se realizó un pool de 1 gramo de larva. Se relacionaron los datos de uniformidad de peso obtenidos con la aplicación los resultados de qPCR para determinar la relación entre la presencia de IHHNV y la disparidad de tallas con respecto a la uniformidad de peso en larvas de camarón.
Análisis estadísticos
Para el análisis estadístico, se utilizó el software R en el entorno de desarrollo RStudio (versión 4.3.2, R Core Team, 2023; RStudio Team, Boston, MA, USA). Se realizó una exploración inicial de los datos mediante análisis descriptivos, usando histogramas de pesos divididos en cuatro rangos de frecuencia, y también se realizó un análisis de la uniformidad de peso a través del tiempo usando boxplots. Este enfoque permitió visualizar la distribución de pesos
en relación con la categoría de PCR positivo o negativo en diferentes rangos de pesos y caracterizar la distribución de la uniformidad en los meses estudiados.
Finalmente, se realizó un análisis de varianza (ANOVA) para evaluar las diferencias estadísticas en la uniformidad de peso entre las categorías de PCR. Se llevaron a cabo ANOVAs individuales por mes con énfasis en la variación a lo largo de cada período. Los resultados se visualizaron mediante un gráfico de barras que representaba los valores de p obtenidos en cada prueba de ANOVA, proporcionando información sobre la significancia estadística de las diferencias observadas. Estos análisis permitieron generar una comprensión detalladas de la distribución de uniformidad a través del tiempo bajo la categorización de PCR positivo o negativo para IHHNV.
Resultados
Como análisis descriptivo se realizó un histograma de pesos (Fig. 1), separado por IHHNV positivo y negativo, donde se observa que no existe una tendencia entre animales positivos y un menor peso; de hecho, se ve que la proporción de positivos para IHHNV se mantiene constante entre los diferentes rangos de peso, por lo que no se visualiza una relación entre diferentes pesos con la patología.
Según el análisis mensual de prevalencias (Tabla 1), los meses con mayor porcentaje de IHHNV positivos son junio y enero, mientras que el mes con menor prevalencia es marzo. Al tener los meses con mayor prevalencia en dos distintas épocas del año, esto quiere decir que en este grupo de datos, la patología no está relacionada al efecto de la estacionalidad de invierno o verano.
Se analizó también el efecto mensual del porcentaje de uniformidad de peso en los diferentes meses analizados (Fig. 2), revelando que enero y febrero son los meses con mayor cantidad de datos atípicos para un porcentaje de uniformidad menor, pero con un promedio de porcentaje de uniformidad mayor. A pesar de eso, el mes de febrero no resultó con un porcentaje alto de prevalencia de IHHNV (Tabla 1). La tendencia mensual refleja que todos los meses mantuvieron un
Prevalencias de IHHNV obtenidas en los meses de enero a junio
Mes
promedio de uniformidad cercano al 80% para las muestras de larvas de camarón.
Se volvió a realizar un boxplot, pero esta vez separando las muestras consideradas positivas-negativas para IHHNV (Fig. 3), donde se observa que en los meses con mayor número de datos atípicos (enero y febrero) los que resultaron negativos para la patología son los que tenían mayor variabilidad de datos con respecto a la uniformidad. Mientras que en los demás meses del análisis, los boxplot se mantienen muy parecidos entre positivo y negativo.
Para los análisis estadísticos se realizaron ANOVA entre ambas categorías de IHHNV y otro ANOVA clasificándolos por meses (Fig. 3), donde no se encuentran diferencias significativas entre IHHNV positivo o negativo, con valor p de 0.8292, podemos decir que con respecto a la uniformidad de tallas no hay relación alguna con el virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa. Además, al analizar los ANOVAS entre los
positivos y negativos separados por meses, ningún mes obtuvo un p value menor a 0.05, por lo que no hay diferencias significativas entre positivo o negativo de forma anual ni mensual.
Discusión
El análisis descriptivo de los datos reveló varios hallazgos importantes sobre la relación entre la presencia del virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa (IHHNV) y la disparidad de tallas en larvas de PL2-PL8 de camarón Penaeus vannamei. Observamos que no existe una tendencia clara entre los camarones positivos para IHHNV y un menor peso, como se refleja en el histograma de pesos (Fig. 1). La proporción de camarones positivos se mantuvo constante a través de diferentes rangos de peso, lo que sugiere que la presencia de IHHNV no está relacionada con diferencias en el peso de las larvas.
Al examinar la prevalencia mensual de muestras positivas para IHHNV (Tabla 1), se observó que los meses con mayor prevalencia fueron enero y junio, mientras que marzo
Figura 1. Histograma de rangos de pesos con los porcentajes de negativos y positivos para IHHNV
Tabla 1.
Figura 3. Boxplot con los datos de uniformidad de los meses de enero a junio obtenidas entre muestras positivas y negativas.
mostró la menor incidencia. Esta variabilidad en la prevalencia a lo largo del año sugiere que la patología no está fuertemente influenciada por factores estacionales como el invierno en enero o el verano en junio.
El análisis mensual del porcentaje de uniformidad de peso (Fig. 2) mostró que en enero y febrero hubo una mayor cantidad de datos atípicos con porcentajes de uniformidad más bajos, pero con un promedio de uniformidad generalmente alto. Sin embargo, febrero no presentó una alta incidencia de IHHNV (Tabla 1). Estos resultados indican que la uniformidad de peso no parece ser un predictor confiable de la presencia de IHHNV en las larvas de camarón.
Al analizar los datos separados entre
muestras positivas y negativas para IHHNV (Fig. 3), encontramos que en los meses con más datos atípicos (enero y febrero), las muestras negativas mostraron una mayor variabilidad en la uniformidad de peso en comparación con las muestras positivas. En general, no se observaron diferencias en la uniformidad de peso entre muestras positivas y negativas en otros meses.
Los análisis estadísticos, incluyendo el ANOVA entre muestras positivas y negativas para IHHNV, no revelaron diferencias significativas en la uniformidad de tallas entre estas categorías, con un valor de p de 0.8292. Esto respalda la conclusión de que no hay una relación clara entre la presencia de IHHNV y la uniformidad de tallas en larvas de camarón P. vannamei
Bibliografía
Chayaburakul K, Lightner DV, Sriurairattana S, Nelson KT, Withyachumnarnkul B. Different responses to infectious hypodermal and hematopoietic necrosis virus (IHHNV) in Penaeus monodon and P. vannamei Dis Aquat Organ. 2005 Nov 28;67(3):191-200. doi: 10.3354/dao067191. PMID: 16408834.
Kalagayan, H., Godin, D., Kanna, R., Hagino, G., Sweeney, J., Wyban, J., & Brock, J. (1991). IHHN Virus as an Etiological Factor in Runt-Deformity Syndrome (RDS) of Juvenile Penaeus vannamei Cultured in Hawái. Journal of the World Aquaculture Society, 22(4), 235–243. https://doi.org/10.1111/J.1749-7345.1991. TB00740.X
Lightner, D. V., & Redman, R. M. (1985). A parvolike virus disease of penaeid shrimp. Journal of Invertebrate Pathology, 45(1), 47–53. https://doi. org/10.1016/0022-2011(85)90048-5
Pénzes JJ, Söderlund-Venermo M, Canuti M, EisHübinger AM, Hughes J, Cotmore SF, Harrach B. Reorganizing the family Parvoviridae: a revised taxonomy independent of the canonical approach based on host association. Arch Virol. 2020 Sep;165(9):2133-2146. doi: 10.1007/s00705-02004632-4. PMID: 32533329.
Primavera, J. H., & Quinitio, E. T. (2000). RuntDeformity Syndrome in cultured giant tiger prawn Penaeus monodon. Journal of Crustacean Biology, 20(4), 796–802. https://doi.org/10.1651/02780372(2000)020[0796:rdsicg]2.0.co;2
Tang, Kathy F.J., & Lightner, D. V. (2006). Infectious hypodermal and hematopoietic necrosis virus (IHHNV)related sequences in the genome of the black tiger prawn Penaeus monodon from Africa and Australia. Virus Research, 118(1–2), 185–191. https://doi. org/10.1016/j.virusres.2006.01.003
Tang, Kathy F.J., Navarro, S. A., & Lightner, D. V. (2007). PCR assay for discriminating between infectious hypodermal and hematopoietic necrosis virus (IHHNV) and virus-related sequences in the genome of Penaeus monodon. Diseases of Aquatic Organisms, 74(2), 165–170. https://doi.org/10.3354/dao074165
Tang KF, Poulos BT, Wang J, Redman RM, Shih HH, Lightner DV. 2003. Geographic variations among infectious hypodermal and hematopoietic necrosis virus (IHHNV) isolates and characteristics of their infection. Dis Aquat Organ 53:91–99.
Figura 2. Boxplot con los datos de uniformidad de los meses de enero a junio obtenidas en la aplicación
Además, al analizar los ANOVA separados por meses, no se encontraron diferencias significativas entre muestras positivas y negativas en ningún mes, incluso el mes de enero que es uno de los que tiene mayor prevalencia para IHHNV, es el mes que tienen el p value más bajo y mayor variabilidad en la uniformidad de peso en su rango de negativos para IHHNV, lo que refuerza la falta de asociación entre IHHNV y la uniformidad de tallas a lo largo del año.
En conjunto, nuestros resultados sugieren que la presencia de IHHNV no está relacionada con la disparidad de tallas en larvas de camarón P. vannamei. Es posible que otros factores, no investigados en este estudio, puedan estar contribuyendo a la variabilidad en la uniformidad de peso observada. Futuras investigaciones podrían
explorar estos factores adicionales para obtener una comprensión más completa de la caracterización de uniformidad de tallas en la larvicultura de camarón.
Conclusión
En este estudio, se investigó la relación entre la presencia del virus de la Necrosis Hematopoyética e Hipodérmica Infecciosa (IHHNV) y la disparidad de tallas en larvas de camarón Penaeus vannamei. Nuestros análisis descriptivos y estadísticos no revelaron evidencia de una asociación significativa entre la presencia de IHHNV y la uniformidad de peso en las larvas de camarón, ya que la proporción de camarones positivos para IHHNV se mantuvo constante en diferentes rangos de peso. El análisis de la uniformidad de peso mensual tampoco reveló diferencias entre muestras positivas
y negativas para IHHNV, sugiriendo que la uniformidad no es un buen predictor para la presencia del virus.
Basado en estos hallazgos, se concluye que la presencia de IHHNV no está relacionada con la disparidad de tallas en larvasde camarón P. vannamei en el contexto de nuestro estudio a partir de PL2 hasta su cosecha ~ PL14. Es posible que otros factores, no investigados en este estudio, estén contribuyendo a la variabilidad en la uniformidad de peso observada, pero no podemos decir que el IHHNV sea uno de ellos•
Para más información sobre este artículo, contactar a: sebastian.ramos@limbomar.com.ec
Supervivencia de cepas de Vibrio cholerae 01 en camarones sujetos a procesos de congelación y ebullición
Autores:
Denilde R. Nascumento,1*
Regine H. S. F. Vieira,*
Hauston B. Almeida,2
Thakor R. Patel,3*
Sebatiao T. Iaria4
1Departamento de Tecnología Química, Universidad Federal de Maranhao, Campus do Bacanya s/n 5-Luis, Maranhao, Brasil; 2 Laboratorio de Ingeniería de Ciencias del Mar del Departamento de Pesca, Labomar, Universidad Federal de Ceará, Fortaleza, Brasil;
3Departamento de Biología, Memorial University of Newfoundland, St. John’s, Newfoundland, Canadá AlB 3X9; y 4 Instituto de Ciencias Biomédicas, Universidad de Sao Paulo, Sao Paulo, Brasil.
El cólera es una enfermedad diarreica deshidratante aguda causada por el toxigénico Vibrio cholerae 01. Es muy contagiosa y a lo largo de la historia se han reportado grandes epidemias con una alta mortalidad. En los últimos 30 años, el cólera se ha reportado en más de 100 países de todo el mundo (37), incluyendo a América. El cólera se registró por primera vez en Perú en 1991 (2, 4, 6) y se propagó rápidamente dentro del país y otras naciones sudamericanas, con más de 500,000 casos y 5,600 muertes reportadas (1, 5). Hoy en día, los organismos de la enfermedad se han extendido por todo el mundo y están asociados con el cólera epidémico (3).
Los mariscos fueron un vector de transmisión del V. cholerae 01 que apareció recientemente en América Latina. Estos alimentos corren un alto riesgo de propagar el cólera, porque el pescado y los mariscos recolectados en aguas contaminadas con el patógeno también se infectan. Además, puede producirse una contaminación cruzada durante la cosecha, el procesamiento y la distribución. El V. cholerae también ha sido aislado de ambientes estuarinos y costeros (12, 14, 20, 35). Por tanto, la contaminación fecal en el agua de bahías es un problema de salud pública (22). También existe cierta evidencia de que V. cholerae 01 se encuentra naturalmente en el medio ambiente, independientemente de la contaminación fecal humana (3, 7, 9, 10, 24). Por tanto, el agua ha sido un importante vector de transmisión del cólera en las epidemias latinoamericanas (33, 34).
Este estudio se llevó a cabo para examinar el efecto de la congelación y ebullición de células de V. cholerae 01 inoculadas en muestras de camarón con y sin exoesqueleto.
Materiales y métodos
Bacterias y condiciones de cultivo Los cultivos puros de V. cholerae 01 se obtuvieron de la colección de cultivos del Instituto Oswaldo Cruz (R.J.). Los cultivos se mantuvieron en agar tríptico de soya (TSA) que contenía NaCl al 1.5%. Los medios líquidos y las placas/tubos de cultivo se incubaron a 35 °C durante 24 h. La pureza de las colonias se confirmó primero en medio selectivo, tiosulfato-citrato-sales biliaressacarosa, TCBS (Difco Laboratories, Detroit,
Michigan). Después de una incubación durante la noche a 35 °C, se subcultivaron colonias sacarosa positivas (amarillas) en agar sangre (Oxoid) para determinar su pureza. Se obtuvieron colonias aisladas con morfología uniforme. El subcultivo en placas de agar sangre se realizó transfiriendo una única colonia aislada de las placas TCBS.
Muestras de mariscos
Los camarones (Penaeus schmitti) utilizados en esta investigación fueron capturados en la zona costera del estado de Maranhao (Brasil) durante los meses de julio de 1995 a marzo de 1996. Se recolectaron ocho muestras de camarones, cada una con un peso de 2.5 kg, en cuatro intervalos de tiempo diferentes; es decir, cuatro lotes de 5 kg de camarón por muestreo.
Cuatro muestras estaban con y cuatro sin exoesqueleto; es decir, en cada momento de muestreo 2.5 kg de camarones estaban con y 2.5 kg estaban sin exoesqueleto. Se realizó un análisis microbiológico de estas muestras para determinar la carga microbiana utilizando métodos estándar (9, 17). Cada una de las muestras (25 g) se homogeneizó en 225 ml de agua peptonada alcalina (APW) y se prepararon diluciones en serie en APW. Las muestras diluidas se esparcieron en placas por triplicado de agar TCBS selectivo. Las placas se incubaron durante 18 a 24 h a 35 °C.
Preparación de muestras para tratamientos
Se hirvieron muestras de camarón (ocho), cada una de las cuales pesaba aproximadamente 2.5 kg, durante 15 minutos para eliminar la microflora contaminante. Las muestras que se iban a congelar se trataron de la siguiente manera: se sumergieron cuatro muestras calentadas, cada una de 2.5 kg, en una suspensión de V. cholerae 01 cultivada durante la noche en caldo de soya tripticasa (Difco) que contenía NaCl al 1.5%. La suspensión bacteriana se preparó suspendiendo células recién cultivadas en 4 litros de solución salina fisiológica en un beaker estéril. Después de una inmersión de 5 min, se separaron en 20 porciones de 25 g cada una y se almacenaron a -20 °C. A intervalos de 2 días, se retiraron las muestras, se descongelaron y se contaron las bacterias supervivientes utilizando el método estándar de recuento en placa para vibrios (8). Se
mezclaron las muestras descongeladas (25 g) en 225 ml de APW durante 2 minutos usando un mezclador de acero inoxidable (Waring Blender, Fisher Scientific, Pittsburgh, Pensilvania). Se prepararon diluciones en serie en APW. Luego se esparcieron las diluciones apropiadas en placas de agar TCBS y se incubaron durante 18 a 24 h a 35 °C. Se contaron y registraron las colonias amarillas sacarosas positivas.
Se utilizaron muestras no inoculadas como controles negativos. Cuatro muestras calentadas, cada una de las cuales pesaba 2.5 kg, se separaron en 12 grupos, cada una de los cuales pesaba 25 g, después de la inmersión en una suspensión del organismo de prueba como se describió anteriormente. Cada una de estas muestras divididas se expuso a temperaturas de ebullición en un baño de agua. Después de 1 a 2 minutos de calentarlas, las muestras se enfriaron en hielo (3 a 4 minutos) y se determinó el número de supervivientes utilizando los procedimientos estándar descritos anteriormente. Las muestras no inoculadas formaron los controles negativos.
Análisis estadístico
Los análisis estadísticos se realizaron utilizando el programa estándar de métodos lineales generales (23).
Resultados y discusión
La Tabla 1 muestra la riqueza de la microbiota de camarones con y sin exoesqueleto capturados entre julio de 1995 y marzo de 1996 en aguas costeras alrededor del estado de Maranhao. Los resultados no mostraron diferencias significativas en los recuentos de Vibrio en muestras de camarones con y sin exoesqueleto.
Utilizando pruebas bioquímicas comprobadas fue posible identificar al Vibrio parahaemolyticus y Vibrio alginolyticus entre la microflora de las muestras de camarón. No se hizo ningún intento de llevar a cabo un examen más profundo taxonómico de los aislados.
La aparición simultánea de estas dos especies no es infrecuente (18). Por ejemplo, Molitoris et al. (25) reportaron la distribución de V. alginolyticus y V. parahaemolyticus en muestras de agua de mar a lo largo de
Bibliografía
1. Amako, K., s. Shimodori, S. Miake, and A. Umada 1987. Effect of chitin and its soluble derivatives on survival of Vibrio cholerae 01 at low temperature. Appl. Environ. Microbiol. 53:603-605.
2. Baudry, B., A. Fasano, J. Ketley, and J. B. Kaper. 1992. Cloning of a gene (zot) encoding a new toxin produced by V. cholerae. Infect. Irnrnun. 60:428434.
3. Blake, P. A., D. T. Allegra, J. D. Snyder, T. J. Barett, L. Mcfarland, C. T. Caraway, 1. C. Feeley, J. P. Craig, J. V. Lee, N. D. Puhr, and R. A. Feldman. 1980. Cholera-a possible endemic focus in the United States. N. Engl. J. Med. 302:305-309.
4. Center for Disease Control. 1991. Cholera-Peru 1991. Morbid. Mortal. Weekly Rep. 40: 108-110.
5. Center for Disease Control. 1992. Update--cholera-western hemisphere-1992. Morbid. Mortal. Weekly Rep. 41:667-668.
6. Dalsgaard, A., M. J. Albert, D. N. Taylor, T. Shimada, R. Merzer, O. Serichantalergs, and P. Echeverria. 1995. Characterization of Vibrio cholerae non-01 serogroups obtained from an outbreak of diarrhoea in Lima, Peru. J. Clin. Microbiol. 33:2715-2722.
7. DePaola, A. 1981. Vibrio cholerae in marine foods and environmental waters: literature review. J. Food Sci. 46:66-70.
8. EUist, E. L., C. A. Kayner, and M. L. Tamplin. 1992. V. cholerae, V. parahaemolyticus, V. vulnificus, and other Vibrio species. FDA, p. 111. Bacteriological Analytical Manual, AOAC International.
9. Epstein, P. R. 1992. Cholera and the environment. Lancet 339:1167-1168.
10. Feachem, R. G. 1981. Environmental aspects of cholera epidemiology II. Occurrence and survival of Vibrio cholerae in the environment. Trop. Dis. Bull. 78:865-880.
11. Furniss, A. L., J. V. Lee, and T. J. Donovan. 1978. The vibrios, p.1-18.1n Public Health Laboratory Service, Monograph Series 11, Her Majesty's Stationary Office, London.
12. Garay, E., A. Arnau, and C. Amaro. 1985. Incidence of V. cholerae and related vibrios in a coastal lagoon and seawater influences by lake discharges along an annual cycle. Appl. Environ. Microbiol. 50:426-430.
13. Guthrie, R. K., C. A. Makukutu, and R. W. Gibson. 1985. Recovery of Vibrio cholerae after heating and or cooling. Dairy Food Sanit. 5:427-430.
14. Hackney, C. R., and A. Dicharry. 1988. Seafoodborne bacterial pathogens of marine origin. 42: 104-109.
15. Heinz, J., A. M. Harrison, A. Leiting, and A. Victoria. 1988. Microflora of brown shrimp (Penaeus azeticus) from Georgia waters. Food Microbiol. 5:141-145.
16. Huq, A. E., B. Small, P. A. West, M. I. Huq, R. Rahman, and R. R. Colwell. 1983. Ecological relationships between Vibrio cholerae and planktonic crustacean copepods. Appl. Environ. Microbiol. 45:275-283.
17. International Commission for Microbiological Specifications for Foods. 1978. Microorganisms in foods 1. Their significance and method of enumeration. University of Toronto Press, Toronto, Canada.
18. Joseph, S. w., R. R. Colwell, and J. B. Kaper. 1983. Vibrio parahaemolyticus and related halophilic vibrios. Crit. Rev. Microbiol. 10:77-124.
19. Kaneko, T., and R. R. Colwe11. 1975. Adsorption of Vibrio parahaemolyticus in Chesapeake Bay. Appl. Microbiol. 29:269-274.
20. Kaysner, C. A., C. Abeyta, R. F. Stott, M. H. Krane, and M. M. Weki1l. 1990. Enumeration of Vibrio species, including Vibrio cholerae, from samples of an oyster growing area, Grays Harbor, Washington. J. Food Protect. 53:300-301.
21. Kenyon, J. E., D. R. Piexoto, B. Austin, and D.
la costa de Yakarta, Indonesia. Los altos recuentos bacterianos de vibrios obtenidos en el presente estudio pueden haber incluido otras bacterias que no son Vibrio. Según Furniss et al. (11) el agar TCBS permite el crecimiento de bacterias distintas de las agrupadas dentro de la familia Vibrionaceae En placas TCBS crecen bacterias gramnegativas como Proteus, Pseudomonas y Aeromonas, y algunas cepas grampositivas de Bacillus y Enterococcus (11).
Una de las mayores preocupaciones con respecto a la reciente aparición del cólera en América Latina es la posibilidad de que V. cholerae 01 pueda transmitirse a otros países a través de alimentos en el comercio internacional.
La Tabla 2 muestra cepas de vibrios aislados de muestras de camarón. Se utilizaron pruebas bioquímicas simples (17) para realizar una identificación tentativa de los aislados de Vibrio sin entrar en más detalles. Las pruebas bioquímicas incluyeron pigmentación, arginina hidrolasa, oxidasa, producción de gas a partir de glucosa, producción de acetoína, crecimiento a 30, 35 y 40 °C; utilización de sacarosa y D-gluconato; celobiosa, y-aminobutirato y putrescina.
La falta de V. cholerae en las muestras de camarón puede explicarse por estas pruebas superficiales. El V. cholerae 01 ha sido aislado de ambientes costeros (12, 14, 19, 20, 21, 25), por lo que la microbiota inicial en las muestras de camarón indica el nivel de contaminación en las aguas donde se cosechó el organismo (15).
Las muestras 1, 2 y 4 mostraron la presencia de V. parahaemolyticus y V. alginolyticus, así como otros Vibrio sacarosa positivos que no fueron identificados. Según West (36), el V. alginolyticus es un patógeno débil oportunista implicado en infecciones extraintestinales.
Este organismo se encuentra en grandes cantidades en los mariscos, pero rara vez causa infecciones en los humanos. Se encontró que hasta un 14% de los aislamientos bacterianos de las aguas de la bahía de Sepetiba, Río de Janeiro, (Brasil) eran de V. alginolyticus, que causa infecciones en el camarón cultivado.
Tabla 1. Número de bacterias sacarosa positivas y negativas del camarón, Penaeus schmitti Burkenroad
Conteo bacterianob (log CFU/g)
a. Cada muestra estuvo compuesta por 2.5 kg de camarón.
b. Para el recuento bacteriano se utilizaron placas de tiosulfato-citrato-sal biliar-sacarosa. La incubación fue de 18 a 24 h a 30 °C. La desviación estándar de los valores anteriores se estimó en ser ±0.0-0.34.
Tabla 2. Identificación de cepas de Vibrio aisladas de los camarones con y sin exoesqueleto.
Número de muestra Aislado bacterianoa
V.parahaemolyticus, V. alginolyticus, Vibrio sp
V.parahaemolyticus y V. alginolyticus
V.parahaemolyticus y Vibrio spy (sac +)
V.parahaemolyticus y V. alginolyticus
a Vibrio spp. creciendo en placas de tiosulfato-citrato-sal biliar-sacarosa y encontradas para ser (sue+) no fueron identificadas.
3. Recuperación de Vibrio cholerae 01 de muestras de camarón con exoesqueleto contaminadas artificialmente y almacenadas a -20 °Ca
Almacenamiento (días)
aLas muestras se congelaron en porciones de 25 g y se almacenaron a -20 °C. Los recuentos de V. cholerae se puntuaron en placas de tiosulfato-citrato-sal biliar-sacarosa que contenía NaCl al 1.5%. Las placas se incubaron durante 18-24 h a
Cada valor representa
recuentos. La desviación estándar observada para el log CFU/g fue de ±0.358.
Tabla
Tabla 4. Recuperación de Vibrio cholerae 01 de muestras de camarón con y sin exoesqueleto contaminadas artificialmente y almacenadas a -20 °Ca
Almacenamiento (días)
Número de muestra (log CFU/g)
aLas muestras se congelaron en porciones de 25 g y se almacenaron a -20 °C. Las colonias se puntuaron en placas de tiosulfato-citrato-sal biliar-sacarosa que contenía NaCl al 1.5%. Las placas se incubaron durante 18-24 h a 30 °C. Cada valor representa el promedio de tres recuentos. La desviación estándar observada para el log CFU/g fue de ±0.45.
Las Tablas 3 y 4 muestran los patrones de supervivencia de V. cholerae inoculados en camarones estériles almacenados a -20 °C. La presencia del exoesqueleto protegió al camarón contra las bacterias por un período más largo (36 días); las muestras congeladas sin exoesqueleto tenían células viables que duraban solo 26 días. Las Figuras 1 y 2 ilustran los patrones de supervivencia de V. cholerae 01 en muestras con y sin exoesqueleto (muestras 1 y 4). Reily y Hackney (31) han reportado observaciones similares en muestras de ostras, cangrejos y camarones.
Hubo una disminución gradual en el número de células viables a -20 °C hasta 20 días. Popovic et al. (30) han discutido la supervivencia de V. cholerae en diferentes condiciones, como las temperaturas altas y bajas, el alto contenido orgánico, pH, ausencia y presencia de microflora competidora y una alta humedad. Llegaron a la conclusión de que mueren más organismos entre -2 y -10 °C que a -70 °C. Además, se demostró que el secado y la luz solar son perjudiciales
para el crecimiento de V. cholerae 01. Guthrie et al. (13) también encontraron que la carne de cangrejo contaminada con V. cholerae 01 contenía células viables después de 35 días cuando se almacenó a 5 °C o 20 °C. Las ostras y almejas que han sido esterilizadas o hervidas antes de ser contaminadas artificialmente pueden tener una supervivencia de V. cholerae 01 por más de 3 semanas cuando se enfría y congela (27). En la mayoría de los alimentos, V. cholerae 01 puede sobrevivir de 2 a 4 días y de mejor entre 5-10 °C que a 30-31 °C (28).
Las muestras expuestas a temperaturas de ebullición mostraron una rápida destrucción de las células de V cholerae. En 1 a 2 minutos, los recuentos se redujeron de log 8.87 CFU/g a menos de 1.0 log CFU/g en el caso de muestras con y sin exoesqueleto. Estas observaciones concuerdan con las reportadas por muchos otros (7, 27, 28, 32). Cuando se las expuso a temperaturas de 80 a 100 °C, el V. cholerae murió en unos pocos segundos. A 65 °C muere en 10 min. Guthrie et al. (13) encontraron que una temperatura
C. Gillies. 1984. Seasonal variation in numbers of Vibrio cholerae (non-OJ) isolated from California coastal waters. Appl. Environ. Microbiol. 47:1243-1245.
22. Madden, J. M., B. A. McCardell, and R. R. Read. 1982. Vibrio cholerae in shellfish from U. S. Coastal waters. Food Technol. 36:93-96.
23. Mendel, W. (ed.). 1983. Introduction to probability and statistics, p.17,646. W. S. Publishers, Boston.
24. Miller, C. J., R. G. Feachem, and B. S. Drasar. 1985. Cholera epidemiology in developed and developing countries: new thoughts on transmission, seasonality and control. Lancet 1:261-263.
25. Mo1itoris, E., S. W. Joseph, M. I. Krichersky, W. Sindhuhardja, and R. R. Colwell. 1985. Characterization and distribution of Vibrio alginolyticus and Vibrio parahaemolyticus isolated in Indonesia. Appl. Environ. Microbiol. 50:1388-1394.
26. Nalin, D. K., V. Daya, M. M. Levine, and L. Cisreros. 1979. Adsorption and growth of Vibrio cholerae on chitin. Infect. Irnrnun. 25:768-770.
27. Pan American Health Organization. 1991. Cholera situation in the Americas. Bull. Pan Am. Health Organ. 12: 10-24.
28. Pan American Health Organization. 1991. Risk of cholera transmission by foods. Bull. Pan Am. Health Organ. 25:274-276.
29. Platt, M. K., M. D. Rich, and J. C. McClaughlin. 1995. The role of chitin in the therrnoprotection of Vibrio cholerae. J. Food Protect. 58:513-514.
30. Popovic, T., O. Osvik, P. A. Blake, and K. Wachsmuth. 1993. Cholera in the Americas: foodborne aspects. J. Food Protect. 56:811-821.
31. Reily, L. A., and C. R. Hackney. 1985. Survival of V. cholerae during cold storage in artificially contaminated seafoods. J. Food Sci. 50:838-839.
32. Singleton, F. L., R. Atwell, S. Jangi, and R. R. Colwell. 1982. Effects of temperature and salinity on V. cholerae growth. Appl. Environ. Microbiol. 44:1047-1058.
33. Swerdlow, D. L., E. D. Mintz, M. Rodriguez, E. Tejada, C. Ocampo, L. Epejo, K. D. Greene, W. Saldana, L. Seminario, R. V. Tauxe, J. G. Wells, N. H. Bean, A. A. Ries, M. Pollack, B. Vertiz, and P. A. Blake.
1992. Waterborne transmission of epidemic cholera in Trujillo, Peru: lessons for a continent at risk. Lancet 340:28-32.
34. Tauxe, R. V., and P. A. Blake. 1992. Epidemic cholera in Latin America. J.Am. Med.Assoc. 267:1388-1390.
35. Venkateswaran, K., T. Takai, L. M. Navarro, H. Nakano, H. Hashimoto, and R. J. Sieb1ing. 1989. Eco10y of Vibrio cholerae non-01 and Salmonella sup and role of zooplankton in their seasonal distribution in Fukuyama coastal waters. Jpn. Appl. Environ. Microbil. 55:15911598.
36. West, P. A. 1989. The human pathogenic vibrios. A public update with environmental perspectives. Epidemiol. Infect. 103:1-34.
37. World Health Organization.
de 50 °C era suficiente para eliminar las células viables del arroz contaminado en 20 minutos. Sin embargo, en condiciones similares fueron necesarios 55 °C para eliminar las células viables en peces y 60 °C en las aves de corral. Blake et al. hicieron observaciones contradictorias (3), quienes reportaron sobre la supervivencia de V. cholerae 01 en cangrejos hervidos durante 2 a 8 minutos, pero no de cangrejos hervidos durante 10 a 14 minutos o cocidos al vapor durante 30 minutos.
Debido a que los hallazgos de esta investigación sugieren que grandes áreas de aguas costeras de Brasil albergan Vibrio que incluyen cepas potencialmente patógenas, es importante que el Estado inste al público a hervir el camarón y productos derivados de camarón durante al menos 10 minutos para asegurar la destrucción completa de posibles patógenos.
Estos experimentos han demostrado y confirmado previos hallazgos de que V. cholerae 01 puede persistir durante largos períodos de tiempo durante el almacenamiento en frío. Huq et al. (16) han discutido la importancia de los copépodos y el material quitinoso en la proliferación y supervivencia de Vibrio en ambientes marinos.
La colonización de copépodos y otros organismos quitinosos (zooplancton) puede contribuir a la abundancia de Vibrio en áreas acuáticas (26, 35). La quitinasa producida por especies de Vibrio puede ayudar a la firme absorción de los organismos al material quitinoso (17).
Nuestros hallazgos coinciden con los de Platt et al. (29). Estos autores concluyeron que la quitina tiene un papel en la crioprotección y la resistencia a los ácidos. No tiene ninguna función en la supervivencia bacteriana a las altas temperaturas utilizadas en la preparación de alimentos. Las propiedades crioprotectoras de la quitina han sido descritas por otros (1, 24)•
Días de almacenamiento (-20 °C)
1.
de supervivencia de Vibrio
01 en camarones en condiciones de congelación (-20 °C). Muestra 1. Los valores representan el promedio de tres recuentos con una desviación estándar de ±0.34 para A y ±0,45 para B.
Días de almacenamiento
2. Patrón de supervivencia de Vibrio cholerae 01 en camarones en condiciones de congelación. Muestra 4. Los valores representan el promedio de tres recuentos con una desviación estándar de ±0.34 para A y ±0.45 para B.
Para más información sobre este artículo, contactar a: tpate1@morgan.ucs.mun.ca.
Figura
Patrón
cholerae
Figura
¿Cuál es el panorama
global de alimentos para el segundo semestre del 2024?
Autor: Henry Wong Director de Desarrollo Comercial, Asia Acuicultura, Alltech
hwong@alltech.com
La industria de alimentos ha experimentado una tasa de crecimiento anual promedio del 3% durante la última década, y un crecimiento promedio del 3,7% en los últimos cinco años. Estas cifras subrayan la progresión constante y la resiliencia de la industria en medio de paisajes agrícolas en evolución.
Producción mundial de alimentos, ligeramente baja en 2023
La producción mundial de alimentos alcanzó los 1,29 mil millones de toneladas en 2023, mostrando una disminución marginal de 2,6 millones de toneladas o 0,2% con respecto al año anterior. Esta reducción en la demanda de alimentos se puede atribuir en parte a la adopción de métodos de producción más eficientes en los sistemas de agricultura intensiva. Estos sistemas priorizan tecnologías como la nutrición animal y la gestión de fincas para reducir el consumo de alimentos, mientras se mantiene o se incrementa la producción de proteínas.
Además, la desaceleración en la producción de proteínas animales, impulsada por márgenes de beneficio reducidos en los sectores de alimentos y proteínas animales, también contribuyó a la disminución de la demanda alimenticia. Factores como los cambios en los patrones de consumo debido a la inflación, las tendencias dietéticas, los costos de producción más altos y las tensiones geopolíticas también contribuyeron a las tendencias observadas en la producción de alimentos en 2023.
Dinámicas de la industria de alimento por región
América Latina ha demostrado un crecimiento constante durante la última década, impulsado por exportaciones robustas en la producción de animales monogástricos y la expansión de los mercados de acuicultura y mascotas. En 2023, la producción de alimentos aumentó en casi 2,5 millones de toneladas, liderada principalmente por un aumento del 2,6% en el sector de pollos de engorde y una adición significativa de 1 millón de toneladas.
La región Asia-Pacífico experimentó un crecimiento modesto en la producción de alimentos, con 6,54 millones de toneladas
o 1,40%, impulsado notablemente por el crecimiento constante en los sectores de pollos de engorde, carnes y lácteos.
Europa enfrentó una disminución en la producción de alimentos, especialmente en el sector de la carne de res, debido a las crisis políticas y de mercado en curso, requiriendo soluciones sostenibles de alimentos para abordar preocupaciones regulatorias.
Norteamérica reportó una disminución en el número de molinos de alimentos y un ligero aumento en las plantas de comidas para mascotas, con el sector de la carne de res enfrentando desafíos y una disminución del 1,1% en la producción de alimentos. La sequía persistente y los altos costos de producción plantearon más desafíos, particularmente en Canadá, donde la producción de alimentos para carne de res experimentó una caída significativa del 10,15% en 2023.
Tendencias y desafíos en la producción de alimentos en
Asia-Pacífico
Las tendencias que impactaron la producción de alimentos en Asia-Pacífico en 2023 están estrechamente alineadas con las destacadas en la Perspectiva Global de Proteínas Animales 2024 de Rabobank.
Los factores económicos y los precios de los productos influyeron significativamente en el comportamiento del consumidor, siendo la inflación y las condiciones económicas las principales preocupaciones. Los cambios en las preferencias del consumidor, particularmente entre los millennials que se enfocan en la nutrición, calidad y conveniencia, influyeron en la demanda de proteínas y en las elecciones de ingredientes para alimentos.
Los factores de salud y las tendencias dietéticas, notablemente en Asia-Pacífico, jugaron un papel significativo. Enfermedades como la Fiebre Porcina Africana (ASF) y la Influenza Aviar Altamente Patógena (HPAI) siguieron siendo un desafío en China y el sudeste asiático, afectando significativamente el sector de pollos de engorde. Las tensiones geopolíticas y los desafíos relacionados con el clima también impactaron en los costos de producción y la consistencia de la cadena de suministro, con
Región
Top 10 países productores de alimento accounted 63.1% of the world’s 2023 feed production. (Fuente: Alltech Agri-Food 2024)
eventos climáticos adversos que afectaron la disponibilidad y calidad de los cultivos y forrajes en varias regiones.
Recesión global en alimentos para acuicultura
El sector de la acuicultura global experimentó una disminución notable en la producción de alimentos. Con una producción total que cayó a 52,09 millones de toneladas en 2023, la industria de alimentos para acuicultura enfrentó una disminución de 2,42 millones de toneladas, equivalente a aproximadamente un 4,43%, en comparación con el año anterior. Un factor clave en esta disminución fue la reducción sustancial en la producción de China, como el principal generador de alimentos para acuicultura, que representó más del 40% del mercado mundial de alimentos para acuicultura.
La reducción de 2,398 millones de toneladas en el volumen de alimentos para acuicultura de China en 2023 impactó significativamente el panorama global de alimentos para esta rama. De hecho, la disminución en el volumen de alimentos para acuicultura de China fue influenciada en gran medida por la lenta recuperación económica después de la pandemia de COVID-19.
Con la demanda del mercado lenta para recuperarse debido a las persistentes incertidumbres económicas, los precios de las especies cultivadas se mantuvieron deprimidos. En consecuencia, muchos se vieron obligados a ajustar sus prácticas de cultivo, como reducir la densidad de siembra de peces o suspender temporalmente las actividades de piscicultura.
Panorama de alimentos para acuicultura por región
En 2023, la región de Asia-Pacífico dominó la producción de alimentos para acuicultura con una producción total de 35,7 millones de toneladas, aunque esto marcó una notable disminución respecto a la cifra del año anterior de 38,42 millones de toneladas, representando un crecimiento negativo del 7,08%. Siguiéndola de cerca, América Latina registró una producción de alimentos de 7,17 millones de toneladas, experimentando un ligero aumento del 3,87% en comparación con el 2022.
Europa quedó rezagada con una producción de alimentos de 4,63 millones de toneladas, demostrando una disminución de 0,14 millones de toneladas o un 2,89% menos que el año anterior. Norteamérica mostró
Tabla 1: Producción de alimentos por región 2023 (Fuente: Alltech Agri-Food 2024)
Tabla 2:
Región
Crecimiento (millón Ton)
Crecimiento (%) Rank, 2023 Rank, 2022
2023 total toneladas de alimento (millón Ton)
2022 total toneladas de alimento (millón Ton*)
un crecimiento marginal en la producción de alimentos para acuicultura, con una producción de 1,78 millones de toneladas, reflejando un modesto aumento de 0,03 millones de toneladas o 1,43%. África exhibió una tasa de crecimiento significativa, alcanzando una producción de alimentos de 1,57 millones de toneladas, un aumento de 0,08 millones de toneladas o 5,53% respecto a 2022.
Las regiones de Medio Oriente y Oceanía también experimentaron crecimiento en la producción de alimentos para acuicultura, con Medio Oriente registrando 1,02 millones de toneladas (crecimiento del 5,43%) y Oceanía, 0,22 millones de toneladas (crecimiento del 7,5%). En general, mientras Asia-Pacífico se mantuvo como líder en la producción de alimentos para acuicultura, varias regiones experimentaron fluctuaciones, con América Latina y África mostrando tasas de crecimiento prometedoras, mientras que Europa enfrentó una disminución.
Las dinámicas cambiantes en la producción de alimentos para acuicultura de 2022 a 2023 son intrigantes, con la mayoría de los países asiáticos presenciando una disminución, mientras que las naciones latinoamericanas vieron un aumento. A pesar de las reducciones en los volúmenes de alimentos para acuicultura de aproximadamente el 10% y el 7,8% en China y Vietnam, respectivamente, ambos países retuvieron sus posiciones principales.
•China, a pesar de una desaceleración en la producción de alimentos para acuicultura,
mantuvo su dominio como el mayor productor de alimentos del mundo, contribuyendo con un sustancial 41% al volumen total de alimentos para acuicultura.
•La industria de alimentos para acuicultura de Vietnam enfrentó desafíos debido a los precios más bajos del mercado del camarón, enfermedades y una disminución en los precios de pangasius y especies domésticas.
•Filipinas vio una disminución del 25% atribuida a la reducción de la producción de
leche de pez y un cambio hacia alimentos comerciales de mayor calidad, mejorando la eficiencia de producción.
•Bangladesh presenció un aumento del 16,7% en la producción de alimentos para acuicultura, impulsado por la creciente demanda de pescado doméstico.
Por el contrario, países latinoamericanos como Ecuador y Chile experimentaron crecimiento, con aumentos respectivos del
Tabla 3: Principales regiones productoras de alimentos para acuicultura (Fuente: Alltech Agri-Food 2024)
Tabla 4. Top 10 países productores de alimento acuicola (Fuente: Alltech Agri-Food 2024)
Región
Crecimiento (millón Ton)
Crecimiento (%)
2023 total toneladas de alimento (millón Ton)
2022 total toneladas de alimento (millón Ton*)
Región
Crecimiento (millón Ton)
Crecimiento (%) Rank, 2023 Rank, 2022
2023 total toneladas de alimento (millón Ton)
2022 total toneladas de alimento (millón Ton*)
Producción de alimento extruido
ALIMENTACIÓN
7,38% y 3,26% en 2023. Sus mercados orientados a la exportación se mantuvieron resilientes en medio de tensiones geopolíticas y condiciones climáticas adversas, impulsados por una robusta demanda de productos acuáticos. Ecuador retuvo su posición como el principal productor de camarones, mientras que Chile emergió como el segundo mayor productor de salmón.
Noruega enfrentó desafíos significativos debido a enfermedades en el salmón y la trucha, resultando en tasas de mortalidad aumentadas, índices de conversión alimenticia reducidos y bienestar animal comprometido. Notablemente, tanto Noruega como Ecuador vieron su sector de alimentos para acuicultura convertirse en el mayor sector de alimentos para animales en sus respectivos países, destacando la importancia de la industria.
Perspectivas para el segundo semestre del 2024
En el ámbito de la producción de alimentos para acuicultura, las perspectivas para 2024 presentan un panorama multifacético. Extrayendo ideas de la encuesta cualitativa, la trayectoria de la industria cobra forma, revelando tendencias y tecnologías que marcan su curso. Notablemente, 2023 presenció una disminución en la producción de alimentos para acuicultura, con una caída del 4,4% (2,42 millones de toneladas), marcando una desviación significativa de años anteriores. Sin embargo, en medio de este desafío, hay un rayo de esperanza, con un 30% de los encuestados expresando optimismo por el crecimiento sectorial en el próximo año. La mayoría de los consultados cree que los altos precios de las materias primas siguen dominando la lista de desafíos enfrentados en la agricultura alrededor del mundo, junto con los bajos retornos para los productores y los problemas con enfermedades animales.
Mirando hacia adelante, las proyecciones de la industria para la producción global de pescado y camarón en 2024 ofrecen un panorama medianamente alentador, sugiriendo una posible recuperación en la producción de alimentos para los países que dependen de estos sectores. Sin embargo, las incertidumbres son grandes, derivadas de las fluctuaciones en la demanda global, la volatilidad de los precios y los desafíos persistentes de enfermedades en la industria del camarón. A pesar de estos obstáculos, mantener una perspectiva optimista es crucial, ya que las proteínas acuáticas de la acuicultura siguen siendo un faro de esperanza, ofreciendo un futuro prometedor en comparación con las prácticas tradicionales de pesca salvaje•
Para más información sobre este artículo, contactar a: hwong@alltech.com
Caracterización de dinoflagelados en un sector de producción de camarón en el golfo de Guayaquil – Ecuador, como un paso inicial a la toma de medidas fitosanitarias
Autores:
Gladys Torres, Carla Torres, Manuel Espinoza-Ortega1
Carlos Mora-Pinargote1
Viviam Rugel
César Molina-Poveda*1
Skretting LATAM
1Skretting Aquaculture Innovation
*cesar.molina@skretting.com
La producción de camarón es una de las principales actividades económicas de Ecuador, y su sostenibilidad depende en gran medida de la calidad del ambiente acuático. En este contexto, la presencia de dinoflagelados, que son un grupo diverso de microalgas, ha cobrado especial relevancia debido a su capacidad de proliferar rápidamente y causar efectos negativos en los ecosistemas acuáticos y la salud de los camarones. Su abundancia esporádica se relaciona con las condiciones óptimas para reproducirse en horas, generando cambios del color del agua denominado mareas rojas o floraciones algales nocivas (FANHAB). Además, algunas especies pueden producir toxinas y alterar la calidad del agua, lo que pone en riesgo tanto la producción camaronera como la biodiversidad local. Estos eventos han sido de corta duración, pero en otros casos se prolongaron mientras las condiciones ambientales persistan.
La clara relación entre la calidad del fitoplancton y el desarrollo del camarón (Llario et al., 2019) ha llevado muchas veces al uso excesivo de fertilizantes para fomentar su crecimiento. Sin embargo, esto puede favorecer al desarrollo de especies no deseadas como los dinoflagelados, provocando afloramientos que amenazan la producción del camarón (Alonso-Rodríguez y Páez-Osuna, 2003). Además, algunas especies de estas microalgas producen toxinas que se propagan rápidamente a través de la cadena alimenticia y afectan a los consumidores superiores (Towers, 2014). El camarón P. vannamei aprovecha eficientemente la productividad primaria de los estanques (New, 2009). La presencia de microalgas dañinas en las áreas de acuicultura constituye un riesgo para la producción y la salud humana. Por lo tanto, es esencial controlar las diferentes especies de dinoflagelados especialmente aquellas que puedan causar daños físicos en branquias, disminución de oxígeno, y con ello consecutivamente llevar a mortalidad. En cultivos acuáticos, altas concentraciones de dinoflagelados (>93.600 células/ml) causan estrés, reducen el oxígeno disponible y afectan la supervivencia debido a su toxicidad (Alonso-Rodríguez y Páez-Osuna, 2003; Keawtawee et al., 2012).
Dinoflagelados: características y funciones A fin de entender la rápida reproducción de los dinoflagelados, es necesario conocer su ciclo de vida. Es muy complejo, debido a las condiciones oceanográficas y neríticas locales, y puede variar considerablemente entre especies, pero generalmente incluye fases tanto asexuales como sexuales. Las células hijas mantienen la mitad de la pared celular original, un proceso conocido como desmoquisis, como ocurre en las especies del género Ceratium. En otros casos, las células hijas no comparten la pared parental, un proceso llamado eleuteroquisis. Este tipo de división puede ocurrir dentro de la célula madre (como en Peridinium) o fuera de ella (como en Protoperidinium), donde cada célula hija forma una nueva pared celular. Ecológicamente, los dinoflagelados juegan roles fundamentales en los ecosistemas acuáticos, tanto marinos como de agua dulce. Son importantes productores primarios en las cadenas tróficas, especialmente las especies fotosintéticas que contribuyen significativamente a la biomasa fitoplanctónica.
Además, algunos dinoflagelados pueden formar un quiste temporal o quiste de ecdisis en respuesta a condiciones desfavorables como cambios de temperatura, y estos pasan al sedimento superficial y permanecen latentes por muchos años. Estos quistes también pueden dividirse, y son parte del ciclo asexual en géneros como Pfiesteria, Crypthecodinium y Alexandrium (Garcés et al., 2007).
Los dinoflagelados son un grupo diverso de protistas unicelulares, conocidos por su papel crucial en los ecosistemas acuáticos planctónicos y bentónicos. Se caracterizan por tener dos flagelos que les permiten moverse de manera distintiva en busca de condiciones nutritivas (nutrientes) y ambientales óptimas (luz solar, temperatura). Cuéllar-Martínez et al., (2023) han reportado recientemente la preferencia de los dinoflagelados por temperaturas superficiales del mar más cálidas y condiciones más estables indicadas por bajas velocidades del viento.
Además, algunos dinoflagelados son bioluminiscentes, emitiendo luz cuando se agitan, mientras que otros pueden formar mareas rojas (Valiadi y Iglesias-
Rodriguez, 2013), eventos de proliferación que pueden liberar toxinas dañinas para la vida marina y humana. Su capacidad para realizar fotosíntesis y su participación en las redes tróficas los convierten en elementos esenciales para la dinámica y salud de los ecosistemas acuáticos.
Importancia ecológica y económica
Los dinoflagelados habitan en la columna de agua (planctónicos) pero también habitan en los sedimentos (bentónicos); otros viven adheridos sobre estructuras superficiales como macroalgas o basura plástica, y se denominan dinoflagelados epibentónicos (Mafra et al. 2023). Otras especies viajan en los tanques de agua de lastre de los buques y se los denomina especies invasoras (Anderson et al. 2014).
Ecológicamente, desempeñan un papel crucial en los ecosistemas acuáticos como productores primarios. Algunas especies de dinoflagelados han registrado floraciones en aguas costeras asociadas a la influencia de nutrientes desde origen antropogénico o desde los afloramientos costeros. Sin embargo, los factores que inician estas floraciones aún son en gran medida desconocidos. Estas floraciones pueden ser tóxicas, no tóxicas y nocivas, dependiendo de la especie que las provoque. Por tanto, los dinoflagelados son organismos de gran importancia tanto ecológica como en la bioeconomía azul, afectando a la diversidad marina, a la industria pesquera y acuícola. Su influencia abarca tanto la estabilidad y productividad de los ecosistemas como la salud y seguridad de diversas actividades económicas dependientes del medio acuático.
Los dinoflagelados pueden generar grandes floraciones cuando encuentran nutrientes como nitrógeno y fósforo (Towers, 2014) . Generalmente, estas floraciones pueden ser de corta duración como las mareas rojas con Peridinium balechii que no afectan la supervivencia (Acevedo, 2010). Sin embargo, otras floraciones pueden causar mortalidades y reducir el crecimiento en camarones (Alonso-Rodríguez y Páez-Osuna, 2003; Keawtawee et al., 2012). En Ecuador, las floraciones algales que generan mareas rojas han sido de corta duración pero se han identificado dinoflagelados tóxicos
como Dynophysis caudata y G. catenatum en mareas rojas originadas por Gyrodinium instriatum no tóxico (Reyes, Krauss, Barniol, Espin, & Intriago, 2001). Algunos producen eventos de toxinas paralizantes como en Pfiesteria, Pseudopfiesteria, miembros de la familia Pfiesteriaceae, así como representantes de los géneros Dinophysis, Alexandrium y otros, los cuales pueden causar la muerte de peces (Skarlato et al., 2018).
Bajo este contexto de importancia global con la bioeconomía azul. La FAO y la COI de la UNESCO colaboran para estudiar los efectos ambientales y en la seguridad alimentaria de la proliferación de algas nocivas, creando un comité de expertos para desarrollar programas de vigilancia del fitoplancton, sistemas de alerta temprana en zonas acuícolas y monitoreo del cambio climático.
Este artículo se centra en la caracterización de los dinoflagelados presentes en un sector específico de producción de camarón en el golfo de Guayaquil, con el objetivo de proporcionar una base sólida de la estructura tanto en composición y abundancia de los dinoflagelados como en los factores ambientales del estudio. Se pretende ofrecer una comprensión integral del problema para la implementación necesaria de medidas fitosanitarias y proponer estrategias de manejo que minimicen los riesgos asociados. Este enfoque inicial es crucial para desarrollar políticas y prácticas que aseguren la salud del ecosistema acuático y la viabilidad económica de la industria camaronera en la región.
Materiales y métodos
Área de estudio
Esta investigación se realizó en las cercanías de Posorja, localidad situada en el golfo de Guayaquil, principal estuario del océano Pacífico (Fig. 1), durante enero, febrero y marzo del 2024, meses correspondientes a la época lluviosa y cálida. Las condiciones oceanográficas del Comité ERFEN-Ecuador indicaron que el evento oceanográfico de El Niño se encontró activo en enero y febrero del 2024, así como la Alerta Naranja a nivel nacional (ERFEN, 2024).
La investigación se realizó mediante monitoreo continuo, en donde se analizó la
abundancia de dinoflagelados. Los puntos de muestreo se ubicaron en el reservorio (1), en el canal-estero (2) y en las piscinas 23, 24 y 25 dedicadas a la crianza de camarón en cautiverio; en cada piscina, los puntos estuvieron en la entrada (E), salida (S), a 3 m (C1) y a 300 m (C2), con un total de 498 muestras correspondientes al primer trimestre del 2024. La profundidad del muestreo fue estándar a 50 cm de la superficie, y se lo realizó entre las 10 y 12h00. La turbidez fue medida in situ con un disco Secchi.
La salinidad fue medida con un refractómetro MA887 (Milwaukee Electronics-Hungría) con compensación automática de temperatura, previa calibración a zero con agua desionizada. Las muestras fueron colocadas en frascos de 100 ml y llevadas al laboratorio en hieleras (mantenidas en frío y en oscuridad). A cada muestra se le agregó una gota de solución lugol (diluido), con la finalidad de disminuir la locomoción y observar algunas estructuras de los dinoglagelados. La abundancia se determinó utilizando una cámara de conteo SedgwickRafter en un transecto de la cámara, luego se hizo la revisión de dos transectos más contabilizando especies que no salieron en el transecto inicial, según el método de Boyd y Tucker (1992). La ubicación taxonómica se realizó a nivel de especie y, en algunos casos, a nivel de género, utilizando los trabajos de Pesantes (1983), Jiménez (1983), Balech (1988) y Tomas (1997). Para caracterizar la diversidad diaria de los dinoflagelados se seleccionó el índice de Shannon utilizando el Programa Past, considerando baja diversidad (<2) y alta diversidad (>3).
Resultados y discusión
Abundancia de dinoflagelados y su relación con otros grupos funcionales del fitoplancton
Durante el periodo de esta investigación, los dinoflagelados fueron el grupo funcional de menor abundancia del fitoplancton en todos los sitios de muestreo, mientras que los grupos funcionales más abundantes fueron las Cianophytas y Ochrophytas (Fig. 2A). Por otro lado, las que presentaron mayor diversidad de especies fueron las Ochropytas y los dinoflagelados. La mayor densidad porcentual de abundancia se registró en el reservorio (enero), en C2 y P24-salida
Figura 2. Abundancia (%) de dinoflagelados y su relación con los otros grupos funcionales (A) y la distribución porcentual de los dinoflagelados (B).
(febrero) y en la entrada de la piscina 24 (marzo) observado en la Fig. 2B.
Registro de la estructura funcional de los dinoflagelados
Se registraron 27 especies contenidas en 12 géneros de dinoflagelados durante el periodo de estudio (Tabla 1). En su mayor parte correspondieron a dinoflagelados tecados (Alexandrium, Ceratium, Dinophysis, Gonyaulax, Oxyphysis, Peridinium, Prorocentrum, Protoperidinium y Scripssiella), y escasos dinoflagelados desnudos o atecados (Cochlodinium, Gymnodinium, Gyrodinium).
El registro y número de ocurrencias tanto
en reservorio (1), canal-estero (29) y las piscinas 23, 24 y 25 (Tabla 1), evidenció que Scripssiella sp. fue la más recurrente (418 registros) en todos los puntos de muestreo y durante los 3 meses, seguido de Protoperidinium sp. (164), Peridinium sp. (142), P. micans (90), Gymnodinium sp. (69), P. quarnerence (59), P. minimum (54), O. oxytoxoides (40), D. diegensis (23), G. catenatum (17), P. mexicanum (17), C. catenatum (14); las otras especies registraron un número menor a 10 durante el período de estudio. En la Fig. 3 se agregaron algunas fotos de los dinoflagelados reportados en este estudio.
Figura 1. Ubicación del área de estudio (Sabana Grande, Guayas, Ecuador).
MANEJO
Implicaciones para la producción de camarón y medidas fitosanitarias
Los hallazgos de este estudio evidencian la presencia de dinoflagelados (tecados y desnudos) al interior de piscinas de cultivo de camarón en el golfo de Guayaquil. Algunas de estas especies tienen registros de causar “blooms” (florecimientos) algales o mareas rojas; su mayor ocurrencia ha sido en el golfo de Guayaquil y durante la época lluviosa-cálida (Torres, 2012). La presencia de dinoflagelados y su alta diversidad ha sido registrada en el margen costero (10 millas) frente a las costas de Esmeraldas, Manta, La Libertad y en el golfo de Guayaquil externo (Torres et al., 2023). Sin embargo, las condiciones oceanográficas como El Niño, afloramientos costeros, entrada de nutrientes desde los sistemas fluviales y la influencia del afloramiento costero durante la época lluviosa y cálida, aportan condiciones para el desarrollo de los “blooms” algales en el margen costero, así mismo en la calidad del agua del sector camaronero principalmente en el golfo de Guayaquil.
A pesar de ser un grupo con una abundancia relativamente baja, las consecuencias de factores neurotóxicos producidos por dinoflagelados son particularmente graves. La base de datos de eventos de algas nocivas (HAEDAT, por sus siglas en inglés) disponible en la red (http://haedat.iode.org) registra la ocurrencia y el impacto global de los eventos
de floraciones algales nocivas (HAB). Las ficotoxinas relacionadas con cada síndrome son neurotoxinas termoestables, lo que significa que no se destruyen con la cocción, representando un riesgo significativo para la salud humana (Brown et al., 2020). Aunque no se han registrado casos en la producción de camarón, los mapas globales de intoxicaciones por mariscos se muestran en Manfrin et al., (2012) y referencias sobre síndromes de intoxicación se encuentran en Berdalet et al., (2016).
El seguimiento constante es esencial para la gestión de riesgos. Los afloramientos de algas se detectan principalmente por confirmación visual en la que se observa decoloración y mortalidad, por eso deben aplicarse los programas de seguimiento basados en el estudio del número y la identificación microbiológica de las especies perjudiciales en las muestras de agua. Este monitoreo requiere un alto grado de especialización por parte del personal, y una alta habilidad taxonómica para la identificación (FAO, 2005).
Diversidad de dinoflagelados registrados en este estudio
Los valores de turbidez (Disco Secchi) y salinidad de las piscinas valoradas durante el primer trimestre del 2024 se observan en la tabla 2. En cuanto a turbidez los promedios oscilaron entre 37 cm y 58 cm,
Tabla 1. Géneros y especies de dinoflagelados registrados en el reservorio (1), canal-estero (2), piscinas 23, 24, 25 (entrada, centro 1-2, salida), con su respectivo número de ocurrencia durante el primer trimestre del 2024.
Bibliografía
ERFEN-Ecuador, Instituto Oceanográfico y Antártico de la Armada del Ecuador, BOLETÍN ERFEN Nro. 02-2024. www. inocar.mil.ec
Balech, E., 1988. Los dinoflagelados del Atlántico Sudoccidental. Instituto Español de Oceanografía, 1: 1-300 Jiménez R., 1983. Diatomeas y Silicoflagelados del Fitoplancton del Golfo de Guayaquil. Acta oceanográfica del Pacífico, Ecuador, 2(2): 193-281. Pesantes, F. 1983. Los Dinoflagelados como indicadores de “El Niño” en el mar ecuatoriano. Acta Oceanográfica del Pacífico. Vol. 2(1): 86-117 Tomas C., 1997. Identifying Marine Phytoplankton. Academic Press Int. San Diego, USA.
Acevedo, A. et al. (2010). DINOFLAGELLATES IN SHRIMP CULTURE PONDS UNDER TYPICAL. Alonso-Rodríguez, R., & Páez-Osuna, F. (2003). Nutrients, phytoplankton and harmful algal blooms in shrimp ponds: A review with special reference to the situation in the Gulf of California. Aquaculture 219(1–4), 317–336. https://doi. org/10.1016/S0044-8486(02)00509-4
Barón-Campis, S. A., Hernández-Becerril, D. U., JuárezRuiz, N. O., & Ramírez-Camarena, C. (2005). Marea roja producida por el dinoflagelado Peridinium quinquecorne en Veracruz, México (oct-nov, 2002): morfología del agente causal. Hidrobiológica, 15(1), 73–78. Retrieved from http://148.206.53.230/revistasuam/hidrobiologica/ viewarticle.php?id=90
Berdalet, E., Fleming, L. E., Gowen, R., Davidson, K., Hess, P., Backer, L. C., … Enevoldsen, H. (2016). Marine harmful algal blooms, human health and wellbeing: Challenges and opportunities in the 21st century. Journal of the Marine Biological Association of the United Kingdom, 96(1), 61–91. https://doi.org/10.1017/S0025315415001733
Brown, A. R., Lilley, M., Shutler, J., Lowe, C., Artioli, Y., Torres, R., … Tyler, C. R. (2020). Assessing risks and mitigating impacts of harmful algal blooms on mariculture and marine fisheries. Reviews in Aquaculture, 12(3), 1663–1688. https://doi.org/10.1111/raq.12403
Coello, D. (2010). Marea Roja ocacionada por Scripsiella trochoidea en el estuario del Río Chone. Revistas Ciencias Del Mar y Limnología, 4(2)(2), 93–100.
Cuellar-Martinez, T., Huanca Ochoa, A. del R., Sánchez, S., Aguirre-Velarde, A., Martínez Ocas, E. O., Rodriguez Velasquez, A. M., … Gutiérrez, D. (2023). Abundance and distribution of potentially toxic phytoplankton in aquaculture sites along the Peruvian coast. Journal of Marine Systems, 240(April 2022). https://doi.org/10.1016/j.jmarsys.2023.103865 FAO (Organización de las Naciones Unidas para la Alimentación y la Agricultura). (2005). Estudio FAO: Alimentación Y Nutrición - Biotoxinas marinas. Https:// Openknowledge.Fao.Org/Server/Api/Core/Bitstreams/ E01aa097-46d0-4f32-808c-Bc2854175054/Content, 292. Retrieved from http://www.fao.org/3/a-y5486s.pdf
Fernández, M. L., Reguera, B., Ramilo, I. & Martínez, A. 2001. Toxin content of Dinophysis acuminata, D. acuta and D. caudata from the Galician Rías Bajas. In Hallegraeff, G. M., Blackburn, S. I., Bolch, C. J. & Lewis, R. J. [Eds.] Harmful Algal Blooms. Intergovernmental Oceanographic Commission of UNESCO,Paris, pp. 360–3.
Fernández, M. L., Reguera, B., González-Gil, S. & Míguez, A. 2006. Pectenotoxin-2 in single-cell isolates of Dinophysis caudata and Dinophysis acuta from the Galicia Rías (NW Spain). Revista Toxicon 48:477–90 Garcés, E., Quijano, S., Figuero, R. I., & Camp, J. (2007). Progresos en el conocimiento de los ciclos de vida de dinoflagelados y diatomeas productoras de proliferaciones de algas nocivas. Repositorio Del IEO (Instituto Español de Oceanografía)
Keawtawee, T., Fukami, K., & Songsangjinda, P. (2012). Nutrient , phytoplankton and harmful algal blooms in the shrimp culture ponds in Thailand, 129–136. Llario, F., Rodilla, M., Escrivá, J., Falco, S., & SebastiáFrasquet, M. T. (2019). Phytoplankton evolution during the creation of a biofloc system for shrimp culture. International Journal of Environmental Science and Technology, 16(1), 211–222. https://doi.org/10.1007/s13762-018-1655-5
Manfrin, C., De Moro, G., Torboli, V., Venier, P., Pallavicini, A., & Gerdol, M. (2012). Physiological and molecular responses of bivalves to toxic dinoflagellates. Invertebrate Survival Journal, 9(2), 184–199.
Reguera, B., & Alonso, R. (n.d.). Comisión Oceanográfica Intergubernamental Manuales y Guías.
Reyes, E., Krauss, E., Barniol, R., Espin, M., & Intriago, P. (2001). Gymnodinium catenatum, Microalga Tóxica aislada en la costa ecuatoriana y sus efectos sobre larvas de Litopenaeus vannamei, (October), 2–3. https://doi. org/10.13140/RG.2.1.2592.0405
Skarlato, S. O., Telesh, I. V, Matantseva, O. V, Pozdnyakov, I. A., Berdieva, M. A., Schubert, H., … Pechkovskaya, S. A. (2018). Studies of bloom-forming dinoflagellates Prorocentrum minimum in fluctuating environment: contribution to aquatic ecology, cell biology and invasion theory. Prostistology, 12(3). Towers, L. (2014). Harmful Effects on Toxin Harmful Effects on Toxin Dinoflagellates in Shrimp Culture Ponds. Thefishsite. Com. Retrieved from https://thefishsite.com/articles/ harmful-effects-on-toxin-dinoflagellates-in-shrimp-cultureponds.
Valiadi, M., & Iglesias-Rodriguez, D. (2013). Understanding bioluminescence in dinoflagellates—how far have we come? Microorganisms, 1(1), 3–25. https://doi.org/10.3390/ microorganisms1010003
con una tendencia a valores mayores en el reservorio (58 y 49), mientras que los niveles menores fueron registrados en el canal de distribución (38, 40 Y 37). La turbidez en niveles altos suele afectar negativamente la disponibilidad de luz lo que puede reducir el crecimiento de ciertas microalgas afectando la diversidad (Remy, 2017); sin embargo, si los sólidos suspendidos en el agua están compuestos en su mayoría de nutrientes, pueden liberar a otras especies de fitoplancton de esta limitante y estimular la biomasa, diversidad y productividad algal (Ogilvie y Mitchell, 1998). Nuestros resultados, sin embargo, no mostraron una relación directa entre la diversidad y la turbidez del agua; piscinas con índices de diversidad bajos (0.37) no presentaron necesariamente la mayor turbidez; por lo que es posible suponer que otras variables, como por ejemplo los nitratos y fosfatos, tienen un efecto mayor sobre la diversidad observada.
Respecto a la salinidad, en enero las piscinas presentaron valores de salinidad superiores a los meses de febrero y marzo, con un rango entre ~19 - ~23 ppt, respectivamente. La salinidad en las piscinas camaroneras depende de la entrada de agua y sus cambios se manejan acorde a las condiciones
climáticas (Welch, 1952). Los valores altos de salinidad en enero podrían deberse a un incremento en la evaporación del agua durante meses calurosos. En este estudio, no se observaron diferencias entre los meses de mayor salinidad comparados con los de menor salinidad en términos de diversidad. Por lo que el efecto sobre la diversidad algal en las granjas acuícolas debe estar relacionado a otros factores bióticos o abióticos como el recambio de agua (Tseng et al., 1991) o concentración de nutrientes y sus relaciones (Boyd, 1995; Paerl y Tucker, 1995).
Las especies registradas (Tabla 2a) presentaron un índice con baja diversidad (<2) tanto en piscinas, reservorio y el canal, con un promedio que osciló entre 0.33 a 1.19, con valores mínimos de 0 cuando en algunas muestras solo se encontraba una sola especie (Tabla 3) con un promedio general de 0.82.
Las especies registradas (Tabla 2b) corresponden a dinoflagelados tecados como Scripsiella, Alexandrium, Dinophisis caudata, D. diegensis Gonyaulax sp., Prorrocentrum mexicanun, P. micans, P. mínimum. Otro
Tabla 2b. Abundancia de dinoflagelados durante el primer trimestre del 2024 en Sabana Grande, Guayas, Ecuador.
Tabla 2a. Valores de turbidez (Disco Secchi), salinidad, e índice de Shannon durante el primer trimestre del 2024.
grupo correspondió a dinoflagelados no tecados (desnudos) como Coclodinium catenatum, Gymnodinium, Gymnodinium catenatum, Gyrodinium spirale. Algunas de estas especies tienen referencias de contener toxinas y causar florecimientos algales.
Se evidenció la predominancia de Scripsiella sp. durante los tres meses del período de estudio, tanto en reservorio, piscinas y el canal. En el 2010, Coello reportó la presencia de un evento de marea roja ocasionada por Scripsiella trochoidea en el litoral ecuatoriano, específicamente en la provincia de Esmeraldas, ocasionando la mortalidad de peces como resultado de los descensos de oxígeno.
Respecto al segundo género en abundancia, Peridinium, existe escasa información en aguas de Ecuador. En México, este dinoflagelado ha sido reportado como causante de una marea roja en el año 2002. En octubre de este año, dos muestreos con 12 días de diferencia fueron realizados, identificándose el dinoflagelado tecado Peridinium quinquecorne (Abé, 1927) mediante un microscopio electrónico de barrido. En este evento, la mencionada especie alcanzó una densidad de 2,5 x 10^6 células por litro, mientras que en el segundo muestreo, las concentraciones de P. quinquecorne descendieron a 4.975 células por litro. Otras especies, como Dinophysis caudata y Pseudonitzschia spp.,
se encontraron en menores densidades. Los valores registrados de temperatura, salinidad y oxígeno disuelto del citado estudio fueron 26,3 °C, 34 ups y 6,5 mg/l, respectivamente. Sin embargo, en este evento y al igual que en otras partes del mundo, Peridinium quinquecorne no resultó nociva (Barón-Campis et al., 2005).
Por otro lado respecto a Dinophysis diegensis, reportado por Reguera y Alonso (2007) como sinónimo taxonómico de Dinophysis caudata, ha sido ampliamente encontrado en aguas costeras tropicales y subtropicales. Forma floraciones que pueden decolorar el agua y han sido asociadas con la mortalidad, especialmente en peces. A nivel mundial ha sido reportado estacionalmente en aguas templadas-cálidas junto con Dinophysis acuta, provocando episodios de intoxicación, por altos niveles de pectenotoxinas y trazas de ácido okadaico (Fernandez et al., 2001 y 2006).
Relación entre dinoflagelados y condiciones ambientales
Los dinoflagelados son altamente sensibles a cambios en factores como la temperatura del agua, la salinidad y la disponibilidad de nutrientes. Las corrientes oceánicas, influenciadas por fenómenos climáticos como El Niño y La Niña, pueden alterar significativamente estas condiciones, promoviendo proliferaciones de dinoflagelados que pueden tener efectos nocivos (daños físicos a branquias) y tóxicos
que afectan al rendimiento en operaciones acuícolas por la producción de toxinas.
Conclusiones y recomendaciones
Estos resultados conforman un primer registro de 27 especies de dinoflagelados tecados y no tecados en un sector de producción de camarón situado en el golfo de Guayaquil. Durante este período no se registraron florecimientos algales, a pesar de que algunas de estas especies tienen registros de haberse generado en el área de estudio.
Monitorear y comprender la interacción entre los dinoflagelados, el medio ambiente y la producción es esencial para la gestión eficiente de una unidad de producción de camarón.
Para abordar estos desafíos, importante implementar prácticas de manejo sanitario robustas, incluyendo la bioseguridad, el monitoreo constante de la salud de los camarones, el control de calidad del agua y el uso prudente de aditivos para tratamientos.
Los resultados sugieren la necesidad de ejecutar estrategias de manejo adecuadas para mantener la sostenibilidad de la acuicultura del camarón, una actividad económica crucial para Ecuador.
Mas allá de ello, a nivel de país, son cada vez más necesarios los programas de alerta temprana para algas tóxicas y especies de floraciones algales nocivas. Estos programas estarían basados en una colaboración entre instituciones y organizaciones de gestión marina, involucrando a la industria acuícola y la comunidad interesada en realizar ciencia ambiental. El monitoreo algal y su ambiente a largo plazo, permitirá comprender la interacción de blooms algales generados por los dinoflagelados, factores ambientales "in situ" y eventos oceanográficos costeros en zonas de producción de camarón•
Para más información sobre este artículo, contactar a: cesar.molina@skretting.com
Figura
Efecto de la variación temporal de la temperatura sobre la producción del camarón Penaeus vannamei en sistemas de cultivo en piscinas
Autores:
Ana Noemí Narváez Vega2
Patricio Colón Velásquez López2
Génesis Estefanía Romero Bustamante1
1Universidad Técnica de Machala. Machala, Ecuador
2Universidad Técnica de Machala, Facultad de Ciencias Agropecuarias. Machala, Ecuador
anarvaez08@hotmail.com
Publicación original:
Revista AquaTechnica https://revistas.utm.edu.ec/index.php/ aquatechnica
Según FAO (2022), la producción acuícola mundial en el 2018 alcanzó 114,5 millones de toneladas de peso vivo de organismos acuáticos, con un valor de $263,600 millones, correspondiendo $9,4 millones a la producción de crustáceos. La Cámara Nacional de Acuacultura del Ecuador (2024) reportó que en el período enero-diciembre del 2023 se exportaron un total de 1,21 millones de t, dando un valor total de $6,289 millones, convirtiendo a la industria camaronera en uno de los sectores productivos más importantes del Ecuador.
En Ecuador, cerca del 90% de la producción de Penaeus vannamei se origina en cultivos que se desarrollan bajo el sistema tradicional o abierto, utilizando estanques de tierra que abarcan entre 4 a 10 ha de terreno (Hirono, 2009). Desde el inicio de la actividad camaronera, la estabilidad de la temperatura ambiente ha sido considerada como una ventaja para que el cultivo de camarón se lleve a cabo durante todo el año en Ecuador. Sin embargo, estudios climáticos que investigan el comportamiento del clima en este país proyectan un aumento en los eventos de calentamiento, con un pronunciado incremento de la temperatura en la estación lluviosa (diciembre-abril) y la presencia de extremas bajas temperaturas en estación seca (Montenegro et al., 2022). Este cambio climático plantea un desafío significativo para la industria acuícola, especialmente para el cultivo del camarón (Boyd, 2020).
Para el mundo de la acuicultura, y de manera específica en el cultivo del camarón, el cambio climático es un asunto complejo que requiere adoptar medidas para el manejo del agua; sin embargo, las iniciativas y acciones varían de un productor a otro debido a las diferencias socioeconómicas y criterios entre propietarios de granjas camaroneras (Do y Ho, 2022).
Debido al calentamiento global, los cuerpos de agua de los sistemas de cultivo tienden a incrementar los niveles de temperatura tanto en la estación seca como lluviosa e incidiendo en el nivel del oxígeno disuelto, lo cual afecta a muchas especies acuáticas, incluido el camarón blanco P. vannamei. Para manejar la calidad del agua en el cultivo del camarón en Ecuador, se ha documentado que la
aplicación de un sistema de transferencias es un mecanismo utilizado como medida para sostener la producción durante las dos estaciones climáticas del año en el país (Castillo-Ochoa y Velásquez-López, 2021).
El método de cultivo mediante sucesivas transferencias de una parte de la población de organismos cultivados, es una técnica que permite iniciar estanques con una alta densidad e ir reduciendo las densidades mediante transferencias sucesivas para obtener mejores tallas comerciales del camarón (De Castro Freire et al. 2021; Seabra de Malgahaes, 2004).
Estudios sobre el comportamiento fisiológico del camarón P. vannamei muestran que este organismo es capaz de adaptarse a ciertos intervalos de temperatura por medio de la autorregulación, pero tiene un límite de tolerancia (Wang et al. 2019). Otros estudios recientes desarrollados por Fóes et al., (2021), relacionados a la tolerancia térmica de P. vannamei, reportan que las mejores supervivencias y conversiones alimenticias se obtuvieron a temperaturas de 20 a 26 °C, en comparación con temperaturas de 30 oC. Así mismo, es bien conocido que cambios o variaciones del oxígeno disuelto (OD) en el agua causan alteraciones moleculares y bioquímicas en el organismo. Se ha reportado que las variaciones de temperatura en el agua causan alteraciones a nivel de la expresión génica del camarón P. vannamei (Camacho-Jiménez et al. 2018) y cambios a nivel enzimático que afectan la función branquial (Camacho-Jiménez et al. 2019). Estudios desarrollados por González-Ruiz et al. (2022), determinaron que altas temperaturas provocaron un mayor contenido de glucosa y lactato, ocasionando normoxia e hipoxia, así como cambios en el metabolismo anaeróbico, causando un significativo estrés que puede provocar la mortalidad de los camarones.
A pesar de la extensa información que se ha documentado sobre la capacidad de resistencia de P. vannamei a variaciones de temperatura y OD, la información de lo que ocurre en cuerpos de agua de cultivo es escasa, y más aún en sistemas de cultivo desarrollados en Ecuador, principal país productor de camarón. Frente a la evidente incidencia del cambio climático en este país,
el presente estudio tiene como objetivo analizar las variaciones de temperatura y OD en el agua durante el cultivo de camarón en la estación lluviosa con una salinidad de 18 ppm y en la estación seca con salinidad de 25 ppm, y su posible incidencia en la producción del camarón cultivado en sistemas abiertos en el sur del Ecuador.
Materiales y métodos
Lugar de realización del trabajo El estudio se realizó en una granja camaronera contigua a la ciudad de Machala, provincia de El Oro, Ecuador, durante junio de 2021 a mayo de 2022, realizando un monitoreo de temperatura y OD en el agua y el crecimiento semanal del camarón (Fig. 1).
Todos los estanques analizados siguieron el mismo protocolo de cultivo que generalmente se emplea en la granja camaronera estudiada. Los administradores de la granja facilitaron información sobre la preparación de estanques, adquisición de postlarvas, siembra y alimentación. El estudio se basó en el acompañamiento para el análisis de temperatura y OD en el agua, y el control de crecimiento del camarón.
El protocolo de preparación de estanques antes de la siembra o antes de cada transferencia, consistió en el drenaje y secado del suelo del estanque y aplicación de hidróxido de calcio a razón de 500 kg/ha. En
los charcos de agua remanentes, se adicionó 500 g de barbasco (planta leguminosa que contiene rotenona) disuelto en 20 l de agua con la finalidad de eliminar larvas de peces, moluscos y pequeños crustáceos. En el suelo se aplicó un inhibidor de la cianofitas a una cantidad de 10 kg/ha para enriquecer el fondo del estanque. El tipo de suelo de los estanques es generalmente conocido como suelo limo-arcilloso. No se realizó un análisis de materia orgánica, pero la condición del suelo fue aceptable puesto que permitió caminar sin provocar hundimientos.
Después de la preparación del estanque, se llenó la mesa de la piscina (40% aproximadamente) y se aplicó una mezcla de melaza a razón de 90 kg con 25 kg de Nutrilake y 45 kg de carbonato de calcio para promover la productividad primaria en el agua. Cinco días posteriores a la siembra, el estanque alcanzó el máximo nivel de agua y a partir de este día, se realizaron recambios de agua de alrededor del 15% diario. La salinidad del agua en la estación lluviosa fue de 18 ppm y en la estación seca fue de 25 ppm.
Las postlarvas utilizadas en el cultivo fueron obtenidas de laboratorios localizados en la península de Santa Elena, perteneciente a la provincia homónima de Ecuador, provenientes de reproductores sometidos a maduración sexual controlada.
Figura 1. Ubicación geográfica de la camaronera motivo de estudio en la provincia de El Oro, Ecuador
Sistema de cultivo mediante transferencias
El sistema de cultivo de camarón descrito en este estudio es un sistema adaptado a tres fases sucesivas, cada una con su estanque de cultivo donde se llevan a cabo transferencias de una parte de la población cultivada.
El sistema de cultivo de transferencias consistió en el manejo de una alta población de postlarvas sembradas en un estanque inicial, la cual da inicio a transferencias sucesivas, en las que se realizó un monitoreo en las dos estaciones lluviosa y seca. El estanque inicial (Fase 1) cumplió la de arranque, donde se transfiere una parte de la población a un estanque secundario (Fase 2). En el estanque secundario continúa el período de crecimiento del camarón, pero luego de un tiempo ocurre la transferencia de una parte de la población a un tercer estanque (Fase 3). En el tercer estanque se cumple la fase final del sistema de transferencias. En este sistema, denominado trifásico, en los tres estanques el camarón se desarrolla en crecimiento y engorde hasta la cosecha final; es decir que el monitoreo de parámetros de cultivo fue sostenido en los tres estanques.
La Figura 2 muestra datos de cada estanque. Para la estación seca, las postlarvas sembradas tuvieron un peso de 0,004 g y luego de 15 días de residencia en el primer estanque (Fase 1), cuando los organismos alcanzaron la edad de prejuvenil con un peso alrededor de 0,2 g, fueron transferidos al segundo estanque (Fase 2). Después de aproximadamente 30 días de cultivo en el segundo estanque, una vez que los organismos alcanzan la edad juvenil con un peso de 1,5 g fueron transferidos al tercer estanque (Fase 3).
En la estación lluviosa, el camarón tuvo un tiempo de residencia diferente en cada estanque. En el primero (Fase 1), los organismos permanecieron hasta el día 87; en el segundo (Fase 2) permanecieron hasta el día 104, y en el tercer estanque hasta el día 123 (Fase 3). Los tiempos de transferencia entre los cultivos de las dos estaciones climáticas difieren en alrededor de 1 a 4 días, por lo tanto, el manejo del cultivo fue prácticamente similar en la estación seca y lluviosa.
Tal como se muestra en la Figura 2, en la estación seca los organismos fueron sembrados en la misma edad y peso que la estación lluviosa, pero los días de transferencia variaron ligeramente.
En la estación seca los organismos tuvieron un tiempo de residencia de 94 días en el primer estanque (Fase 1); hasta el día 112 en el segundo estanque (Fase 2); y hasta el día 128 en el tercer estanque (Fase 3).
Alimentación durante el cultivo
Figura 2. Secuencia de los períodos de residencia del camarón en tres estanques de cultivo mediante sistema de transferencia en tres fases. Se muestra el peso inicial promedio de los organismos en la etapa de siembra en el primer estanque, la población inicial, biomasa inicial y días de cultivo en cada estanque o fase de transferencia en la estación lluviosa y estación seca. liberado en cada entrega del equipo estuvo en función de la dosis diaria de alimento. Las dosis fueron proporcionadas según el peso y la población de camarones a la fecha, ajustando la cantidad de alimento cada día según la biomasa estimada y las tablas de alimentación sugeridas por fabricantes de alimento balanceado. Un ajuste a la dosis diaria de alimento fue realizado considerando la observación de los platos testigos, los cuales entregan información de la cantidad de alimento no consumido y lo que permite deducir si se aumenta o se disminuye el alimento balanceado de la dosis diaria. La efectividad de este proceso dependió de los operadores de la granja camaronera.
En el primer estanque de ambas estaciones (seca y lluviosa) en la etapa de postlarvas, se proporcionó alimento balanceado por el método al voleo 35% proteína (0,8 mm). Es decir, este método fue empleado solo durante 15 días de la Fase 1 previo a la trasferencia a la Fase 2. Posteriormente, el alimento balanceado fue distribuido mediante el uso de los alimentadores automáticos tipo “timer”. En la fase juvenil del camarón se proporcionó alimento balanceado 35% de proteína (1,2 mm) y en la fase final de desarrollo del camarón el alimento balanceado 35% de proteína (3,5 mm).
Los alimentadores automáticos fueron programados para distribuir las raciones desde las 10:00 horas hasta las 00:00 horas, distribuyéndose un total de 120 raciones en un intervalo de 7,5 minutos durante el tiempo de 14 horas. El volumen de alimento
Control de población y crecimiento del camarón
El crecimiento del camarón fue determinado mediante muestreos semanales, tomando en cuenta el número de lances aproximados.
Cxl: #Tc #L
Cxl: camarón por lance #Tc: número total de camarones
#L: número de lances (realizados en cada piscina)
Peso (P): PTO (g) #To
PTO: peso total de los organismos (g), #TO: número total de organismos (obtenidos de los lances realizados)
El peso fue determinado utilizando una balanza digital con precisión 0,01 g y aplicando la siguiente fórmula:
Incremento de peso semanal (PS)= ISN − ISA
ISN: incremento de peso de la semana nueva ISA: incremento de la semana anterior
La supervivencia (% Supervivencia) se calculó con los datos de la densidad inicial, biomasa cosechada y el peso final obtenido de los camarones cosechados.
% Supervivencia= Número de camarones cosechados x 100 Número de camarones sembrados
El factor de conversión alimenticia (FCA) expresa la ganancia en peso del organismo que es cultivado con relación al peso del alimento que fue utilizado, para lo cual se basó a partir de esta fórmula:
Los parámetros de oxígeno y temperatura se midieron dos veces al día, al amanecer a las 05:00 horas y al atardecer a las 17:00 horas, con ayuda del instrumento multiparámetro
YSI modelo Pro-20, calibrado a la salinidad del agua. Ambos parámetros fueron medidos sumergiendo el electrodo a 20 cm sobre el fondo del estanque.
Análisis de datos
Como los estanques fueron sembrados de manera secuencial acorde a las fases previamente indicadas en la estación lluviosa y la estación seca, los datos de oxígeno, temperatura, crecimiento, supervivencia y conversión alimenticia fueron organizados en una hoja Excel en la que posteriormente se realizó un análisis de datos para cada estanque o fase de cultivo. El promedio, valor mínimo, valor máximo y el coeficiente de variación fueron determinados de acuerdo con el arreglo de datos de la mañana y de la tarde, a fin de evaluar las variaciones y comparar los resultados entre el amanecer y el atardecer en cada fase o estanque y en
cada estación climática. Adicionalmente, los resultados fueron cotejados con documentación científica que reporta valores permisibles para el camarón P. vannamei, y estableciéndose un análisis comparativo en las variaciones de los parámetros temperatura y oxígeno disuelto, consumo de alimento y producción de camarón entre las dos estaciones climáticas.
Resultados y discusión
Los resultados evidencian variaciones de temperatura durante el cultivo tanto en la estación seca como en la estación lluviosa. Aparentemente, tales variaciones de temperatura en cada estación climática tuvieron un efecto directo sobre la disponibilidad de OD en el agua y consecuentemente sobre la supervivencia de los organismos y la productividad camaronera.
Variación de temperatura del agua
En la Figura 3 se puede observar que en varios días de la estación seca la temperatura del agua estuvo alrededor de 29 °C en las tres fases de cultivo. Así mismo, en la estación seca la temperatura del agua descendió a alrededor de 23,8 °C. En cambio, en la
estación lluviosa, la temperatura del agua alcanzó a 34,6 °C y descendió a 20 °C, es decir, 3 °C de diferencia con relación a la temperatura mínima registrada en estación seca.
En la Tabla 1 se muestran los valores promedio, mínimo, máximo y coeficiente de variación de temperatura en la estación lluviosa y estación seca en las tres fases de cultivo. En general, la temperatura promedio diaria en la estación seca fluctuó alrededor de 26,36 °C ± 0,83. Como era de esperar, los niveles más altos de temperatura se registraron al atardecer y los niveles más bajos fueron registrados al amanecer en ambas estaciones del año.
De los resultados analizados para las dos estaciones climáticas, se puede observar que la diferencia entre los promedios de las temperaturas entre el amanecer y al atardecer en todas las fases de cultivo fue de alrededor de 2 °C durante la estación seca y de alrededor de 3 °C en estación lluviosa. Al comparar las dos estaciones climáticas, se determina que la diferencia de la temperatura mínima y máxima fue de 11°C, aspecto que merece ser tomado en cuanta para el manejo
Figura 3. Variación diaria de la temperatura del agua (mañana y tarde) en las tres fases de cultivo de camarón y durante la estación seca y estación lluviosa. Se muestra el promedio de temperatura diaria y las barras cruzadas muestran la desviación estándar en días cuando las tres fases se juntan.
Tabla 1. Temperatura (°C) durante el cultivo de camarón en la estación seca y lluviosa en un sistema de cultivo abierto
Promedio DE Minimo
Máximo
Coeficiente de
estacional del cultivo. Consecuentemente, el análisis del coeficiente de variación diario de temperatura demuestra que en la estación seca, la temperatura del agua fluctuó alrededor de 2,6% al amanecer y 4,4% al atardecer, mientras que en la estación lluviosa la variación de temperatura al amanecer fue del 7,8% y al atardecer de 6,9%. Evidentemente, la mayor variabilidad de temperatura se registró durante el amanecer en la estación lluviosa.
Se ha reportado ampliamente que la temperatura del agua ejerce efectos sobre la supervivencia, consumo de oxígeno, comportamiento nutricional y metabolismo de los camarones. En el presente estudio los incrementos de temperatura en la estación lluviosa fueron hasta 5 °C por encima del límite permisible de 30 oC para P. vannamei (Ponce-Palafox et al., 1997). Estudios de Wyban et al. (1995) indican que la temperatura óptima de P. vannamei está alrededor de 30 °C. Por otra parte, PiñaValdez et al. (2015) aseguran que el nivel de consumo de oxígeno es normal con menor gasto energético a una temperatura menor a 30 °C. Un estudio que involucró camarones infectados con virus de la Mancha Blanca (WSSV) y sostenidos a 30 °C se reportó mejor supervivencia en aquellos organismos que estuvieron expuestos a temperaturas alrededor de 26 °C (Wigglesworth, 2002).
Para entender la dinámica de la temperatura en el agua de los estanques de cultivo del camarón en Ecuador, datos de la temperatura superficial del agua del Instituto
Oceanográfico de la Armada del Ecuador (INOCAR) demuestran que frente a la costa de la provincia de El Oro, los niveles de temperatura durante la estación seca fluctuaron alrededor de 23,26±0,84 °C y en la estación lluviosa entre 26,22±0,63 °C. La diferencia de temperatura del océano con la del estanque en la estación seca fue de alrededor de 0,7 oC, siendo el promedio de la temperatura del agua superficial del océano inferior a los valores mínimos registrados en los estanques de cultivo de camarón.
Durante la estación lluviosa, la capacidad de absorber calor en los estanques fue más pronunciada, puesto que la temperatura del agua de cultivo de camarón se incrementó en más de 5 °C con relación a la temperatura del agua superficial del océano registrada por el INOCAR en esa misma estación del año. Esto plantea que la eventualidad del aumento de la temperatura del agua en los estanques de cultivo de camarón es un aspecto crítico y requiere control. El aumento de la temperatura del agua en la estación lluviosa en los estanques de cultivo de camarón respecto a la temperatura del océano puede explicarse por el efecto de la radiación y la turbidez generada por las microalgas, con lo cual se produce conservación de calor y “efecto invernadero” en el agua de la piscina, conservando la temperatura y retrasando el enfriamiento de la columna de agua.
La radiación solar tiene enorme incidencia en el calentamiento del estanque. En efecto, la estación seca en el Ecuador está asociada a mayor nubosidad, relativa baja temperatura
y una débil radiación solar, lo cual explica que la escasa radiación recibida en esta temporada reduce la presencia de microalgas y contribuye a un ligero sostenimiento de la temperatura del agua en el estanque. En el presente estudio, la diferencia de temperatura del agua del océano con la del estanque fue de aproximadamente 1 °C. Pero, contrario a este efecto, en la estación lluviosa la contribución del efecto de la radiación solar en el agua del estanque es más severa, puesto que en algunos casos la temperatura excedió los niveles permisibles para los camarones en cultivo. Este aspecto es relevante para futuros estudios respecto a las variaciones de temperatura en el agua en sistemas abiertos, considerando que el 90% de los sistemas de cultivo en Ecuador son llevados bajo este modelo. Lo indicado confirma que el control de los parámetros meteorológicos de temperatura y radiación solar son críticos para la acuacultura costera y el manejo del sistema de cultivo abierto.
En el presente trabajo, durante la estación lluviosa se detectaron mortalidades más altas que en la estación seca. Falta de información no nos permite deducir la razón de las mortalidades, puesto que podrían ocurrir diversos factores. Sin embargo, se ha indicado que la estación lluviosa está asociada a una intensa radiación solar en el Ecuador y se ocasiona un excesivo crecimiento de microalgas y consecuentemente un incremento en la materia orgánica disuelta en el agua (Boyd, 2018). El incremento de la temperatura en el agua desencadena otros efectos en su calidad para el cultivo del
*AM: Amanecer, PM: Atardecer
camarón, con especial referencia al alimento no consumido, el incremento de materia orgánica y aumento del nitrógeno amoniacal (Limsuwan y Ching, 2018). Recientemente, a inicios del 2023, algunas granjas camaroneras del sur de Ecuador reportaron episodios de afloramientos de microalgas toxicas, provocando asfixia y mortalidades de camarón. Algunos empresarios camaroneros también han reportado el incremento de la presencia de protozoarios y afectación de la salud del animal. Un análisis integral de la calidad del agua proveería más datos para entender la ocurrencia del afloramiento de algas toxicas, patologías y mortalidades; sin embargo, de acuerdo con el análisis del presente trabajo, parece ser que en sistemas de cultivo abierto temperaturas extremas que suceden durante la estación lluviosa inciden en el balance metabólico del camarón y en la propagación de microrganismos que podrían estar afectando la salud y supervivencia de las poblaciones en cultivo.
A pesar de la percepción de que las condiciones del clima durante la estación seca, caracterizadas por bajas temperaturas, son poco favorables para el cultivo de camarón, los resultados del presente estudio indican que en esta estación climática las condiciones de temperatura permitieron
mejores supervivencias, lo cual solo podría explicarse por una condición menos incidente en el afloramiento de microorganismos tóxicos y niveles de temperatura más estables en comparación con lo observado en la estación lluviosa.
La estabilidad de la temperatura durante la estación seca permitió un ambiente térmico más aceptable para P. vannamei, resultando en una mejor supervivencia; sin embargo, los resultados indican que la tasa de crecimiento de los organismos fue menor. Implementar mecanismos de control de las variaciones climáticas con especial atención al manejo de la temperatura del agua en estanques de cultivo de camarón sostenidos bajo sistemas abiertos, requeriría adecuar condiciones que permitan mantener el calor en el agua durante la estación seca y mecanismos de refrescamiento durante la estación lluviosa cuando la temperatura exceda los límites permisibles.
Variación de OD en el agua y dinámica en el cultivo
En la Figura 4 y Tabla 2 se muestran los valores de oxígeno, promedio, valor mínimo y máximo y el coeficiente de variación en la estación lluviosa y estación seca, y en las tres fases de cultivo. En concordancia con lo
Figura 4. Fluctuación del OD en el agua en las tres fases de cultivo de camarón en la estación lluviosa y seca. La línea de tendencia muestra el promedio de OD diario y las barras cruzadas muestran la desviación estándar en días en los cuales las tres fases se juntan.
Bibliografía
Avnimelech Y., Ritvo G. (2003). Shrimp and fish pond soils: Processes and management. Aquaculture, 220(1):549-569. http://doi.org/10.1016/S00448486(02)00641-5
Boyd C. E. (2018). Proper management most important aspect of aquaculture pond water quality management. Global Seafood Alliance, 1-6. Boyd C. E. (2020). Efectos del tiempo y el clima en la acuacultura. Responsible Seafood Advocate. https:// www.globalseafood.org/advocate/efectos-del-tiempoy-el-clima-en-la-acuacultura/
Boyd C. E., Tucker C. S. (1998). Pond aquaculture water quality management. Kluwer Academic Publishers. Boyd, C. E., Watten B. (1989). Aeration systems in aquaculture. Review of Aquatic Sciences, 1(3):425-472.
Camacho-Jiménez L., Felix-Portillo M., NuñezHernandez D. M., Yepiz-Plascencia G. (2019). Molecular cloning and modeling of the Tp53-induced glycolysis and apoptotic regulator (TIGAR) from the Pacific white shrimp Litopenaeus vannamei and its expression in response to hypoxia. Fish & Shellfish Immunology, 93:484-491. https://doi.org/10.1016/j. fsi.2019.08.003
Camacho-Jiménez L., Peregrino-Uriarte A. B., Martínez-Quintana J. A., Yepiz-Plascencia G. (2018). The glyceraldehyde-3-phosphate dehydrogenase of the shrimp Litopenaeus vannamei: Molecular cloning, characterization and expression during hypoxia. Marine environmental research, 138:65-75. https:// doi.org/10.1016/j.marenvres.2018.04.003
Cámara Nacional de Acuacultura. (2024). Estadísticas- Reporte de Exportaciones Ecuatorianas Totales. Revista Aquacultura, 157:58. https://www. cna-ecuador.com/revista-acuacultura/ Castillo-Ochoa B. D., Velásquez-Lopez P. C. (2021). Manejo estacional de los sistemas de producción de camarón en el Ecuador. Sociedad & Tecnología, 4(3):447-461. https://doi.org/10.51247/st.v4i3.151
De Castro Freire M. V., Alves Costa M. G., Lopes Ferreira R. (2021). Eficiência dos sistemas de cultivo de camarões marinhos. Pubvet, 1-7. https://doi. org/10.31533/pubvet.v15n2a752.1-7
Do H., Ho T. Q. (2022). Climate change adaptation strategies and shrimp aquaculture: Empirical evidence from the Mekong Delta of Vietnam. Ecological Economics, 196.
FAO. (2022). El estado mundial de la pesca y la acuicultura. Hacia la transformación azul. https:// www.fao.org/3/cc0461es/cc0461es.pdf
Ferreira N. C., Bonetti V., Seiffert W. Q. (2011). Hydrological and Water Quality Indices as management tools in marine shrimp culture. Aquaculture, 318((3-4):425-433. https:// doi.org/10.1016/j.aquaculture.2011.05.045
Fóes G., Wasielesky Junior W., Marchetti I., Rosas V. (2021). Assessing the effect of temperature on FCR in Pacific white shrimp cultured in biofloc systems. Global Seafood Alliance, 1-7.
García A., Brune D. (1991). Transport limitation of oxygen in shrimp culture ponds. Aquacultural Engineering, 10(4):269-279. https://doi.org/10.1016/01448609(91)90016-D
González-Ruiz R., Leyva-Carrillo L., Peregrino-Uriarte A. B., Yepiz P. G. (2022). The combination of hypoxia and high temperature affects heat shock, anaerobic metabolism, and pentose phosphate pathway key components responses in the white shrimp (Litopenaeus vannamei). Cell Stress and Chaperones, 1-17. https:/doi.org/10.1007/s12192-022-01265-1
Tabla 2. Estadística descriptiva de la concentración de OD (mg/L) en el agua durante el cultivo de camarón en estación seca y estación lluviosa en un sistema de cultivo abierto. OD
*AM: amanecer, PM: atardecer
determinado en el registro de temperatura, tanto en la estación seca como en la lluviosa, al amanecer se registraron los niveles más bajos de OD, mientras que al atardecer se obtuvieron los valores máximos de OD. Siguiendo la norma ambiental, al amanecer en algunos días de cultivo se registraron valores críticos de OD en el agua de alrededor de 2 mg/L, mientras que al atardecer los valores estuvieron siempre en intervalos aceptables. Al amanecer en la estación seca, el coeficiente de variación de OD en el agua fue de 16,11% y al atardecer de 12,36%. En la estación lluviosa, el coeficiente de variación de oxígeno al amanecer fue del 18,44% y al atardecer fue de 12,40%. Esto confirma que, tanto en la estación lluviosa como en la seca, las variaciones de OD en el agua son más severas durante la noche evidenciándose el punto crítico en el monitoreo a tempranas horas de la mañana. Así mismo, es relevante resaltar que las variaciones de OD son más críticas en los cultivos de camarón durante la estación lluviosa.
La variabilidad de OD fue similar tanto para la estación seca como para la estación lluviosa, pero las concentraciones máximas y mínimas de OD reportadas al amanecer y al atardecer fueron diferentes en ambas estaciones climáticas. Niveles por debajo de 3 mg/L de OD están asociados a condiciones de estrés y por debajo de 2 mg/L provocan anoxia, lo cual se traduce en un retardo en el crecimiento y mortalidad en el cultivo del camarón (Boyd, 2018; Ferreira et al., 2011). Niveles por encima de 4 mg/l son ideales para la producción en acuicultura (Robertson, 2006). Se ha reportado que el nivel de OD en el agua decrece cuando la temperatura
y la salinidad aumentan (Boyd y Tucker, 1998). De acuerdo con lo registrado en el presente estudio, la condición estuarina del sitio de estudio revela que altas salinidades coincidieron con bajas temperaturas de la estación seca.
El consumo de oxígeno de P. vannamei a una temperatura de 20 °C fue observado por Martínez Palacios et al. (1986), quienes han confirmado que los organismos acuáticos requieren más oxígeno por unidad de peso cuando se exponen a temperaturas extremas, ya sea alrededor de 20°C y superiores a 30°C. Una investigación previa de Villareal et al. (1994) había determinado que la tasa de respiración en postlarvas de P. vannamei se incrementa cuando la temperatura aumenta. Así mismo, Vinatea et al. (2011) estudiaron la demanda de oxígeno en P. vannamei, acorde al peso de los organismos, la densidad poblacional y la salinidad del agua, y concluyeron que este último es un factor crítico en el consumo de oxígeno y que la tasa de respiración se incrementa cuando los organismos aumentan de peso. En este sentido, una combinación de la temperatura en diferentes condiciones de salinidad podría también explicar posibles diferencias en la respuesta de los camarones a las diferentes condiciones de cultivo. Es decir, la temperatura estará asociada a otros factores ambientales que no fueron estudiados en el presente trabajo por falta de información y sugiere mayor análisis en diferentes sectores.
En cultivos tradicionales con densidades de cultivo relativamente no tan altas como las analizadas en el presente trabajo, se ha reportado que los organismos consumen
aproximadamente el 9% del oxígeno disuelto; por tanto, el mayor consumo de oxígeno está en la respiración del agua y los sedimentos, entendiéndose que ese consumo es debido a la actividad metabólica de los microorganismos (Santa y Vinatea, 2007). Los niveles bajos de oxígeno registrados en el presente estudio durante las primeras horas de la mañana son debido a la actividad fitoplanctónica por el consumo del oxígeno durante la noche (Boyd, 1990; García y Brune, 1991).
En el presente estudio se observó que la aireación mecánica se mantuvo solo en los primeros 15 días de cultivo de la primera fase, y posteriormente el manejo del OD en función de los recambios de agua diarios. Los recambios de agua estuvieron alrededor del 15% del volumen total de las piscinas. Aproximadamente en el 80% de granjas camaroneras, el recambio de agua está en función de los niveles de la marea, por lo cual no es un mecanismo seguro para suplir las demandas de oxígeno. Ante estas condiciones, el control del OD al amanecer y al atardecer es posiblemente insuficiente, requiriéndose que este parámetro junto a la temperatura y otras variables críticas como el pH, requieran un control más frecuente.
Se entiende que las condiciones de OD en el agua de un estanque influyen en la transformación de nutrientes a través de la degradación de la materia orgánica, la cual se produce diariamente en el estanque por excreción de los organismos y la acumulación diaria de alimento no consumido. Por esto, la disponibilidad de nutrientes disueltos en el agua en sistemas abiertos de cultivo
Tabla 3. Parámetros de producción en el cultivo del camarón durante las tres fases de cultivo
la estación seca y lluviosa
PARÁMETROS
Población final (N)
Peso final (g)
Tasa de crecimiento (%/ semana)
Biomasa inicial (kg)
Biomasa final (kg)
Alimento consumido (kg)
Conversión alimenticia
Supervivencia (%)
es limitada (Avnimelech y Ritvo, 2003; Velásquez-López et al., 2023). A esto se suma que la descomposición de materia orgánica en un ambiente anaeróbico es más lenta por la ausencia de oxígeno (Sahrawat, 2004). Por tanto, el oxígeno es el principal elemento que limita la producción en acuicultura (Boyd y Watten, 1989; Santa y Vinatea, 2007). En adición, la materia orgánica del fondo y que está en suspensión, sirve como sustrato para bacterias de toda clase, incluso las patógenas. Entre más materia orgánica haya en un estanque de cultivo de camarones, mayor riesgo de bacteriosis habrá, con los subsecuentes brotes de otros microorganismos, enfermedad y eventual mortalidad, bajando el porcentaje de sobrevivencia.
La condición de transferencia de OD en el agua con el uso de aireadores ha sido documentada por Vinatea et al. (2011). En Ecuador, aproximadamente desde el 2010 se implementaron aireadores mecánicos para mejorar las condiciones de oxígeno en los estanques de cultivo de camarón. En el presente estudio, la administración de los aireadores mecánicos fue dedicada exclusivamente a la etapa inicial e intermitentemente en etapas de mayor desarrollo e independientemente de la estación climática. Es decir, no se considera el peso corporal de los organismos durante el ciclo de cultivo, la biomasa y las necesidades de aireación cuando el estanque alcanza la mayor demanda de oxígeno. Lo observado en el presente estudio no puede generalizarse, pero la evidente incidencia de la temperatura demuestra la necesidad de considerar alternativas de manejo de la calidad del agua.
Productividad del camarón
La Tabla 3 muestra el resumen de producción de las tres fases de cultivo en la estación lluviosa y seca. La supervivencia alcanzada en la estación seca fue 1,5 veces superior a la registrada en la estación lluviosa. Así mismo, la biomasa total de camarón producido en las tres fases de cultivo durante la estación seca fue superior a la biomasa reportada para la estación lluviosa.
El peso de los organismos cultivados en la estación lluviosa fue superior, alrededor de 4,8 g al peso registrado en la estación seca. En esta hay temperaturas bajas, pero más estables; el camarón sobrevive más, pero crece menos. Esto puede ser atribuible a la reducción del metabolismo por el agua más fría, así como a la subalimentación al haber mayor biomasa (por mayor supervivencia) y falta en ajustes a la tasa diaria de alimentación. La inanición también tiene un efecto directo en la muda o desarrollo de P. vannamei y en la supervivencia (Ochoa y Velásquez-López, 2023).
Durante la estación seca la conversión alimenticia en las tres fases de cultivo fue de 1,88 y en la estación lluviosa fue de 1,76. Tanto en la estación seca como en la lluviosa, la conversión de alimento más alta fue registrada en el cultivo del tercer estanque o Fase 3, seguida de la conversión obtenida en el segundo estanque o Fase 2 y la menor se registró en el primer estanque o Fase 1. Es importante destacar que la Fase 1 tuvo alimentadores automáticos en las primeras etapas de vida en cautiverio del camarón, aspecto que podría haber incidido en el consumo de alimento. En general,
Hirono Y. (2009). Preliminary Report on Shrimp Culture Activites In Ecuador. Journal of the World Mariculture Society, 14(1-4):451-457. https://doi. org/10.1111/j.1749-7345.1983.tb00097.x
Islam M. A., Akber M. A., Ahmed M., Rahman M. M., Rahman M. R. (2018). Climate change adaptations of shrimp farmers: a case study from southwest coastal Bangladesh. Climate and Development, 11(6):459468. https://doi.org/10.1080/17565529.2018.144 2807
Limsuwan C., Ching C. A. (2018). Temperature affects shrimp survival, feed conversion. Responsible Seafood Advocate. https://www.globalseafood. org/advocate/temperature-affects-shrimp-survivalfeed- conversion/#:~:text=Lab%20trials%20at%20 Kasetsart%20University,white%20shrimp%20 (Litopenaeus% 20vannamei).
Martínez Palacios C. A., Ross L. G. (1986). The effects of temperature, body weight, and hypoxia on the oxygen consumption of the Mexican mojarra, Cichlasoma urophthalmus (Günther). Aquaculture Research, 17(4):243-248. https://doi. org/10.1111/j.1365-2109.1986.tb00110.x Montenegro M., Campozano L., Urdiales-Flores D., Maisincho L., Serrano-Vincenti S., Borbor-Cordova M. J. (2022). Assessment of the Impact of Higher Temperatures Due to Climate Change on the Mortality Risk Indexes in Ecuador Until 2070. Frontiers in Earth Science, 9:1340. https://doi.org/10.3389/ feart.2021.794602
Ochoa-Pereira P. M., Velásquez-López P.C. (2023). Effect of fasting on molting and survival rate in postlarvae of the shrimp Litopenaeus vannamei: Efecto del ayuno sobre la muda y la tasa de supervivencia en postlarvas del camarón. Revista de Biología Marina y Oceanografía, 58(1):10-18. Piña-Valdez P., Arzola G. J., Nieves-Soto M., MedinaJasso M. A. (2015). Efecto combinado de Temperatura y Salinidad en el consumo de oxígeno en postlarvas de Camarón Blanco Litopenaeu vannamei. Boletim do Instituto de Pesca Sao Paulo, 41(1):89-101. Ponce-Palafox J., Martinez-Palacios C. A., Ross L. G. (1997). Los efectos de la salinidad y la temperatura en las tasas de crecimiento y supervivencia de juveniles de camarón blanco, Penaeus vannamei, Boone, 1931. Aquaculture, 157(1-2):107-115. https://doi. org/10.1016/S0044-8486(97)00148-8
Robertson C. (2006). Australian Prawn Farming Manual: Health Management for profit. The State of Queensland, Department of Primary Industries and Fisheries, Brisbane, 159. http://aciar.gov.au/ publication/CoP01
Sahrawat K. L. (2004). Organic matter accumulation in submerged soils. Advances in Agronomy, 81:169-201. https://doi.org/10.1016/S0065-2113(03)81004-0 Santa K., Vinatea L. (2007). Evaluation o respiration rates and mechanical aeration requirements in semiintensive shrimp Litopenaeus vannnamei culture ponds. Aquacultural Egineering, 1:73-80. https://doi. org/10.1016/j.aquaeng.2006.07.002
Seabra de Malgahaes M. E. (2004). Cultivo do camarao marihno Litopenaeus vannamei (Boone, 1931) em sistema multifásico. Masters Thesis. Universidade Federal Rural de Pernambuco, Recife. Velásquez López P., Solórzano J., Ochoa P., Solano G., Quizhpe P., Guillen R. (2023). Caracterización de la calidad del agua durante el cultivo del camarón Litopenaeus vannamei con agua dulce en el Sur del Ecuador. Journal of the Selva Andina Animal Science 10(2):74-87. https://doi.org/10.36610/j. jsaas.2023.100200074.
Villareal H., Hinojosa P., Naranjo J. (1994). Effect of temperature and salinity on the oxygen consumption of laboratory produced Penaeus vannamei postlarvae.
la conversión de alimento resultante de la estación seca fue ligeramente superior a la conversión de alimento obtenida en la estación lluviosa para las tres fases de cultivo.
En concordancia con lo observado en las variaciones de oxígeno y temperatura en el presente estudio, posiblemente las inestables y extremas temperaturas registradas en la estación lluviosa causaron estrés en los organismos sostenidos en las tres fases de cultivo. Esto coincidió con descensos del OD en el agua a niveles por debajo de 2 mg/l en la estación lluviosa, lo cual podría haber afectado el metabolismo energético de los camarones, lo que conlleva a la susceptibilidad de enfermedades y mortalidades que repercuten en la productividad (Xu et al., 2019).
La respuesta fisiológica del camarón se altera considerablemente cuando niveles críticos de temperatura y el OD en el agua de cultivo se combinan. Temperaturas menores a 26 °C dieron como resultado un retardo de un 25% en el crecimiento respecto al crecimiento detectado en estación lluviosa, cuando las temperaturas estuvieron alrededor de 30 °C. Por su parte, temperaturas sobre los 30 °C marcaron una mortalidad alrededor del 30% en la estación lluviosa con relación al número de organismos cosechados en la estación seca.
Respecto al consumo de alimento, Limsuwan y Ching (2018) establecieron que a 33 °C los camarones consumieron mayor cantidad de alimento que a 29 °C y, aunque el crecimiento fue similar en ambas temperaturas, la mortalidad fue más alta a 33 °C. En el estudio de estos investigadores se determinó que el grupo de animales expuestos a 33 °C tuvo la mayor conversión alimenticia, altos niveles de nitrógeno y menor supervivencia. Es decir que se evidencia enlace de la temperatura con otro factor ambiental que también incide en la salud de los organismos. En el presente estudio, se encontraron mortalidades de 46,3±5,9% en la estación lluviosa cuando los organismos estuvieron a temperaturas que fluctuaron entre 24,9 a 34,7 °C. Todas las fases de la estación lluviosa tuvieron la misma respuesta de mortalidad. Al analizar el consumo total de alimento exógeno en las tres fases de la
estación seca, con temperaturas de 23 a 26 °C, se identificó un incremento de 13,334 kg de alimento consumido en comparación con la estación lluviosa. Por tanto, la conversión alimenticia observada en la estación seca tuvo un promedio de 1,88 kg mientras que en la estación lluviosa fue 1,76 kg. Es decir, en la estación seca, con una supervivencia significativamente superior a la estación lluviosa, se consumió 0,12 kg más de alimento por cada kg de camarón producido.
De acuerdo con lo previamente analizado en el presente estudio, la ganancia en peso del camarón no tuvo relación con el consumo de alimento en las tres fases de cultivo. A altas temperaturas, menor consumo de alimento, mayor peso del camarón y menor tiempo de cultivo, determinándose un promedio de peso de 23,87±3,05 g en las tres fases de cultivo y en 91,33±4,19 días de cautiverio. En cambio, en la estación seca con bajas temperaturas, el promedio de peso de las tres fases fue de 19,04±2,42 g y 96,33±1,69 días de cultivo. El mejor rendimiento en peso obtenido en la estación lluviosa podría haber sido por mejor asimilación del alimento exógeno cuando los organismos están a altas temperaturas. Sin embargo en la estación seca, con temperaturas más bajas, el mayor consumo de alimento permitió sostener la población de organismos en los estanques de las tres fases. Se requiere más información de manejo y control del consumo de alimento exógeno junto a parámetros de calidad de agua para deducir con más claridad la influencia en la supervivencia y ganancia en peso del camarón bajo las condiciones de cambios de clima.
Finalmente, de acuerdo con el cálculo de ingreso económico por venta de camarón y considerando el precio del camarón a la fecha de cultivo, en la estación seca se obtuvo un ingreso económico de USD 208,514 en la cosecha de las tres fases de cultivo, mientras que en la estación lluviosa se generó un ingreso económico de USD 183,748, análisis que ratifica que la estación seca fue económicamente más productiva para este caso de estudio.
Algunos productores de camarón no están conscientes del efecto invernadero y las consecuencias en los cambios de temperatura, la disponibilidad del OD en el
agua y la productividad. Consecuentemente, tampoco se presta atención al efecto sobre la calidad del agua, la salud ambiental y bienestar animal. Debido a los cambios del clima y temperaturas extremas, la estación lluviosa o el efecto del cambio climático con altas temperaturas en el Ecuador, se convierte en una condición crítica para el cultivo del camarón, lo que implica que para el manejo de sistemas abiertos es crítico implementar medidas para el manejo de las variaciones de la temperatura del agua, en especial cuando exceden niveles permisibles, así como las consecuencias en otras variables como el OD en el agua y el consumo de alimento de los organismos.
El conocimiento de las proyecciones sobre la variación del clima respecto a los cambios en la temperatura del agua y la afectación a la condición de oxígeno disuelto, podrían ayudar a los acuicultores a tener una mejor planificación y mejores resultados de producción (Islam et al. 2018). Medidas para una acuicultura sostenible deben ser tomadas a nivel de granja, sector o región, puesto que alrededor del 90% del sector camaronero del Ecuador practica el cultivo en sistemas abiertos de cultivo, donde la temperatura del agua depende de las condiciones del clima.
Conclusiones
El presente estudio muestra que, durante el cultivo del camarón P. vannamei en Ecuador, las temperaturas del agua de la estación lluviosa y seca marcan diferencias significativas. Por lo tanto, frente a los eventos de cambio climático, el control de estos parámetros en sistemas de cultivo abierto es crítico para evitar intervalos térmicos fuera de los límites permisibles para la especie. Los resultados del presente estudio se enfocaron a las variaciones de temperatura y OD en el agua de cultivo con salinidades de 18 ppm para la estación lluviosa y 28 ppm para la estación seca.
La temperatura del agua de los estanques de cultivo de camarón fue más baja y estable en la estación seca. En la estación lluviosa, la supervivencia del camarón en cultivo fue menor, compensado por el mejor crecimiento del camarón y menor tiempo de cultivo. Estudios de calidad de agua más integrales que incluyan otras variables tales como pH,
amonio, materia orgánica, productividad primaria y análisis microbiológico, podrían contribuir a un mejor entendimiento de la incidencia de los cambios extremos de temperatura sobre las condiciones de calidad de agua en estanques de cultivo y la respuesta fisiológica del camarón en cautiverio.
Dado que los datos del presente estudio surgen de una granja de cultivo de camarón del Ecuador, los resultados no pueden generalizarse o aplicarse a otras latitudes geográficas o con diferentes modelos de cultivo. Sin embargo, este estudio deja en evidencia que durante los últimos tiempos en el sur del Ecuador, la temperatura en el agua de cultivo sufre variaciones que pueden alcanzar niveles extremos para el camarón,
aspecto crítico para la sostenibilidad del cultivo en sistemas abiertos, así como también para el manejo del alimento exógeno, la supervivencia y la producción de biomasa, problema que los productores de camarón de esta región del país tienen que enfrentar con la implementación de medidas de adaptación al cambio climático acorde a las condiciones de cada granja•
Para más información sobre este artículo, contactar a: anarvaez08@hotmail.com
Comparative Biochemistry and Physiology Part A: Physiology, 108(2-3):331-336. https://doi. org/10.1016/0300-9629(94)90103-1
Vinatea L., Muedas W., Arantes R. (2011). The impact of oxygen consumption by the shrimp Litopenaeus vannamei according to body weight, temperature, salinity, and stocking density on pond aeration: A simulation. Acta Scientiarum Biological Sciences, 33(2):125-132. https://doi. org/10.4025/actascibiolsci.v33i2.7018
Wang Z., Qu Y., Yan M., Li J., Zou J., Fan L. (2019). Physiological responses of Pacific White Shrimp Litopenaeus vanamei to temperature fluctuation in low-salinity water. Frontiers in physiology, 10:450479. https://doi.org/10.3389/fphys.2019.01025
Wigglesworth J. (2002). Temperature effects on shrimp survival to disease. Global Seafood Allience. https:// www.globalseafood.org/advocate/temperatureeffects-on-shrimp-survival-to-disease/ Wyban J., Walsh W. A., Godin D. M. (1995). Temperature effects on growth, feeding rate and feed conversion of the Pacific white shrimp (Penaeus vannamei). Aquaculture, 138(1-4):267-279.
Xu Z., Guan W., Xie D., Lu W., Ren X., Yuan J., Mao L. (2019). Evaluation of immunological response in shrimp Penaeus vannamei submitted to low temperature and air exposure. Dev. Comp. Immunol, 100:103413. https://doi.org/10.1016/j. dci.2019.103413
Bienestar de crustáceos decápodos con especial énfasis en la fisiología del estrés
Autores:
Sven Wuertz,1
David Bierbach,1
Mirko Bögner2
1Departamento de Biología de Peces, Pesca y Acuicultura, Instituto Leibniz de Ecología de Agua Dulce y Pesca Continental, Müggelseedamm 310, Berlín 12587, Alemania
2Instituto Alfred-Wegner, Centro Helmholtz de Investigación Polar y Marina, Am Handelshafen 12, Bremerhaven 27578, Alemania
Hindawi
Aquaculture Research
Volume 2023, Article ID 1307684, 17 pages https://doi.org/10.1155/2023/1307684
sven.wuertz@igb-berlin.de
BIENESTAR ANIMAL
Durante la última década, entre los crustáceos decápodos, el cultivo de camarón duplicó su producción global (2008: 5 millones de t, 2018: 9.4 millones de t) [1, 2]. Entre las especies cultivadas, el camarón blanco del Pacífico Penaeus vannamei es la especie más importante y representa aproximadamente el 53% de la producción total de crustáceos [2]. Junto al salmón, el atún, la merluza y el arenque, el camarón se encuentra entre las especies acuícolas más consumidas en Alemania [3]. Para cubrir esta demanda, se han importado aproximadamente 47 000 de t [4]. La mayoría de estas importaciones se producen en sistemas de piscinas, alcanzando un rendimiento global de 5 millones de t anuales [5]. Por el contrario, existe una pequeña producción emergente basada en un sistema acuícola de recirculación (RAS) de aproximadamente 447 t en Europa [6]. La producción basada en un RAS a menudo se vende mediante marketing directo como producto fresco, publicitando una imagen sostenible. Aunque la producción RAS es tan pequeña, ha sido un núcleo para la integración de dimensiones de bienestar en el cultivo de crustáceos (principalmente en camarón, pero también en langosta o cangrejo de río), pero la evaluación científica sistemática sobre el bienestar animal en camarón es escasa.
De hecho, los principios fundamentales del bienestar animal en criaderos, como el “Concepto de las Cinco Libertades” (Concepto de las Cinco Disposiciones) [7] no han sido evaluados ni adaptados al cultivo de camarón como requisito previo de una evaluación basada en el conocimiento del bienestar animal en el cultivo. A diferencia de otros animales, que incluye a peces, la tecnología de cultivo apenas se ha evaluado en un contexto de criterios de bienestar en procesos de producción rutinarios y, por lo tanto, rara vez puede comunicarse al consumidor. La seguridad de la cadena alimentaria está directamente relacionada con el bienestar de los animales cultivados para la producción de alimentos [8]. Como parte de la Estrategia de Bienestar Animal, la Comisión Europea pidió indicadores medibles de bienestar animal para reforzar la buena cría con base científica [9], pero hasta ahora los invertebrados están excluidos, como se anunció recientemente durante
la revisión de las leyes de bienestar animal de la UE [ 10]. Sin embargo, el parlamento europeo adoptó la resolución “De la Granja a la Mesa”, que pide a la Comisión que apoye y fomente el desarrollo de normas de bienestar más estrictas para los invertebrados marinos como los crustáceos decápodos [11]. En el Reino Unido, los crustáceos decápodos ahora son reconocidos como “sensibles” en el proyecto de ley de sensibilidad y se esperan cambios en la legislación.
Los animales enfrentan peligros nocivos que pueden causar estrés y daño tisular. Para protegerlos de tales efectos adversos, los crustáceos tienen respuestas reflejas nociceptivas. Aun así, existe una diferencia fundamental entre la nocicepción y el dolor: la primera tiene una dimensión fisiológica, mientras que la segunda tiene una dimensión emocional/mental [1215]. Los receptores específicos que son sensibles a las lesiones se denominan nociceptores y se han conservado en todo el reino animal [14, 16]. En principio, son órganos sensoriales que procesan estímulos nocivos, ya sean químicos, mecánicos, térmicos o combinaciones de estos en un reflejo para prevenir lesiones graves [14, 15]. Si la experiencia se procesa en el cerebro y se vincula a una experiencia aversiva, tiene un valor altamente adaptativo que incluye el aprendizaje de evitación para poner fin al estímulo nocivo. Estos intentos de escapar del estímulo van más allá de la respuesta reflexiva vista con la nocicepción y, por tanto, podrían tener más éxito que con la mera respuesta. En consecuencia, esta experiencia se denomina dolor. Debido a su prominencia, es probable que se recuerde, así como también la situación que resultó en la nocicepción. La discusión sobre el dolor y sufrimiento en los crustáceos decápodos es bastante controversial y está lejos de estar resuelta. Algunos autores demostraron que las respuestas de los decápodos no pueden explicarse como reflejos puros, mientras que otros afirman que la literatura no es concluyente y que no existen preocupaciones sobre el bienestar [17, 18].
Por el contrario, el estrés es una respuesta fisiológica de un organismo que ayuda a reconstituir la homeostasis después de la interferencia de un estímulo externo adverso (estresor). Al experimentar un estrés crónico
intenso, la respuesta al estrés puede perder su función adaptativa y volverse disfuncional (angustia). Cuando el estrés es prolongado, puede provocar adaptaciones conductuales a las que a menudo se hace referencia como un estado de mayor alerta. En los vertebrados se ha formulado un concepto de estrés tripartito [19, 20]. Aunque los crustáceos carecen del sistema endocrino de los vertebrados, se puede aplicar un concepto similar a los crustáceos (Figura 1) [21-23, 26]. En una reacción primaria, se liberan factores neuroendocrinos que desencadenan la respuesta secundaria al estrés. En la respuesta secundaria al estrés, se producen cambios metabólicos que cambian la energía para restaurar la homeostasis (alteración de la glucosa, el lactato y el glucógeno). En la respuesta terciaria, después de una exposición prolongada al factor estresante, se observan cambios en el rendimiento del animal, por ejemplo, crecimiento reducido, disminución de la resistencia a enfermedades o cambios de comportamiento [21]. El estrés prolongado con graves efectos en el rendimiento se denomina generalmente “angustia”. En general, se acepta que las condiciones de producción estresantes y prolongadas se traduzcan en una mala salud y, posteriormente, en brotes de enfermedades, que a menudo implican la pérdida total de stocks.
En esta revisión, nos centraremos en los aspectos fisiológicos del bienestar y su seguimiento. El impacto de las enfermedades en el bienestar se ha presentado en detalle recientemente [27]. Revisaremos la literatura existente sobre el dolor y la angustia en crustáceos, particularmente en el camarón, para respaldar un principio de precaución sobre el bienestar animal establecido en peces y el ganado [13]. Además, destacaremos aquellos indicadores de bienestar establecidos que respalden futuros intentos de optimizar las prácticas existentes en el cultivo de crustáceos. Además, identificaremos los riesgos relacionados con el bienestar en la práctica, en particular en laboratorios, en engorde, cosecha, para anestesia, transporte y para el sacrificio compasivo, y señalaremos aquellas cuestiones relevantes para la optimización sostenible de la tecnología, preferiblemente en el camarón blanco del Pacífico.
Concepto de bienestar animal
En el cultivo de animales acuáticos, el número puro de especies cultivadas (>400 especies) supera con creces el de los animales terrestres. La información sobre el bienestar específico de cada especie suele ser limitada. Por ejemplo, aproximadamente el 70% de las especies acuáticas cultivadas no tienen publicaciones sobre su bienestar [28].
Actualmente, el rápido crecimiento de especies como el camarón está incrementando la información disponible sobre bienestar [28]. De hecho, la producción de camarón se está intensificando, mientras que la sensibilidad del camarón todavía no es comúnmente aceptada [29, 30].
Definir bienestar es difícil debido a la complejidad del tema y a la falta de criterios de bienestar. El concepto de las Cinco Libertades se estableció en la década de los 60 en respuesta a un informe gubernamental del Reino Unido sobre la ganadería. Establece que cualquier animal acuático de cultivo debe al menos estar libre de hambre y desnutrición mediante un acceso adecuado a una dieta para mantener plena salud y vigor; libre de molestias y exposición al proporcionarle un ambiente apropiado que incluya refugio y área de descanso; libre de lesiones y enfermedades mediante la prevención o un diagnóstico y terapia rápidas; libre para expresar un comportamiento normal proporcionando suficiente espacio, instalaciones adecuadas y compañía de sus congéneres, y libre de vivir sin miedo, dolor y angustia garantizando condiciones y tratamientos que eviten la angustia y el sufrimiento.
De hecho, no es posible liberarse por completo de estos estados negativos y el objetivo debería centrarse en minimizarlos como parte de una estrategia de buenas prácticas acuícolas. El Sistema Europeo de Evaluación de la Calidad del Bienestar para la cría de animales excluye especialmente la utilización del concepto de las Cinco Libertades y enuncia cuatro “principios de bienestar”, de “buena alimentación”, “buena vivienda”, “buena salud” y “comportamiento apropiado” [31 –33]. De manera similar, Mellor et al. [34-36] defendieron un concepto de cinco disposiciones, que incluían “buena
nutrición”, “buen ambiente”, “buena salud”, “comportamiento apropiado” y “experiencias mentales positivas”, centrando la atención en las medidas prácticas necesarias para lograr resultados de bienestar deseables. Asimismo, el Modelo de Cinco Dominios para el bienestar animal identifica “nutrición”, “medio ambiente”, “salud”, “comportamiento” y “estado mental” como dominios físicos/funcionales que llaman la atención sobre el manejo práctico de los animales [27, 36 –39], minimizando las experiencias negativas y promoviendo logros positivos. Recientemente, se han presentado indicadores no invasivos para el cultivo de camarón, introduciendo un sistema de puntuación integral basado en los dominios nutrición, salud ambiental y comportamiento [29].
En este sistema se puntúan valores de referencia como la temperatura o el pH y los indicadores asignados al dominio respectivo. Broom señaló que no existe un parámetro único para evaluar el bienestar en el cultivo de animales [40]. La necesidad de incorporar un enfoque interdisciplinario basado en parámetros conductuales, fisiológicos y de salud está ampliamente aceptado en la actualidad [41, 42]. Ashley [43] destacó la importancia de los parámetros fisiológicos basados en la sensibilidad y el valor diagnóstico que respalda una evaluación gradual de los aspectos del bienestar.
Nocicepción
La nocicepción es el mecanismo sensorial que permite a los animales detectar y evitar rápidamente estímulos potencialmente dañinos para los tejidos, ya sean térmicos, mecánicos o químicos [14]. Después de la entrada nociceptiva, las señales neuronales desencadenan procesos protectores (nociceptivos) que llegan al sistema nervioso. Posteriormente, se procesan y pueden percibirse como “dolorosos” [16]. La nocicepción es más bien plástica y de ninguna manera vuelve a su función normal después de que se ha producido el daño [44, 45]. En consecuencia, a menudo se observa un aumento de la sensibilidad después de una lesión, lo que se denomina sensibilización. Claramente, los crustáceos decápodos muestran un comportamiento nociceptivo [46-48], pero solo recientemente se han descrito nociceptores primarios
Estresor físico
p.ej. transporte, almacenamiento
Estresor químico
p.ej. CO2, oxígeno
Estresor social
p.ej. densidades de stock
Respuesta primaria al estrés serotonina (5HT)
Hormona hiperglucémica de los crustáceos (CHH)
Respuesta secundaria al estrés
Glucosa
Lactato
Frecuencia cardiaca
Ventilación
Hemocitos
Glucógeno
Respuesta al estrés terciario
Ecdisis
Crecimiento
Reproducción
Vigor
Resistencia a enfermedades
Conversión alimenticia
Figura 1: Descripción general de algunas respuestas fisiológicas en crustáceos decápodos expuestos a factores estresantes físicos, químicos o sociales. Los factores estresantes desencadenan una respuesta neuroendocrina (respuesta primaria) que evoca efectos fisiológicos y conductuales (respuesta secundaria) para restaurar la homeostasis. Si persiste, el factor estresante puede inducir cambios sistémicos que afecten el desempeño del animal [21-25].
en una especie [49]. Cabe señalar que, a diferencia de Drosophila, las langostas o cangrejos de río no responden a sustancias químicas nocigénicas como la capsaicina [49], el isotiocianato [49] o el pH extremo [50].
Hay muchas moléculas y receptores correspondientes vinculados a la nocicepción, como los canales de potencial receptor transitorio (TRP) y los receptores opioides que se han reportado en crustáceos decápodos [24, 51-54]. Los canales TRP controlan la temperatura y la nocicepción mecánica, transduciendo estímulos nocivos en corrientes [54]. Los opioides y los receptores de opioides están intrínsecamente vinculados a la nocicepción y al dolor, modulando el estímulo real a nivel del cerebro. Hay tres receptores opioides acoplados a la proteína G, siendo, μ-, δ- y κ- y los respectivos ligandos encefalina, ß-endorfina y dinorfina [53, 55].
Varios estudios demostraron que la estimulación nociva aguda y crónica inducen la expresión del óxido nítrico sintetasa (NOS), que sintetiza el mensajero neuronal óxido nítrico (NO) en mamíferos e invertebrados [56-58]. En cangrejos de costa, se ha sugerido que el NO no solo participa en el procesamiento sensorial sino también en la modulación del programa motor, como el movimiento de la cola [58]. Por lo tanto, el aumento del NO puede afectar la señalización sensorial, el procesamiento neuronal central y la generación de señales motoras eferentes después de la exposición a un estímulo nocivo, similar a los mecanismos observados en los mamíferos.
Dolor
Aunque la mayoría de los animales tienen respuestas reflejas nociceptivas que los protegen de influencias nocivas, algunos taxones han desarrollado una capacidad de experimentar dolor, presumiblemente para
mejorar una protección a largo plazo basada en el recuerdo de la naturaleza nociva del dolor. Zimmerman afirmó que el dolor puede identificarse como una experiencia sensorial aversiva causada por una lesión que provoca reacciones protectoras motoras y vegetativas, y da como resultado una evitación aprendida en el futuro [59]. Por lo tanto, un animal que sufre dolor debería aprender rápidamente a evitar el estímulo nocivo. Bateson [12] sugirió ocho criterios indicativos con respecto a la sensación de dolor. Los tres primeros definen un conjunto de requisitos previos funcionales: la posesión de nociceptores, una estructura central similar a la corteza cerebral de los humanos, así como moduladores endógenos como los opioides y los receptores de opioides. Además, los analgésicos deberían modificar la respuesta a los estímulos nocivos. Los restantes son puramente conductuales, aseverando que el animal debe evitar los estímulos nocivos experimentados y que esta experiencia debe ser relativamente persistente y que el animal debe asociar los eventos neutrales con los estímulos nocivos. De hecho, si se cumplen muchos de estos criterios, se puede concluir que es probable que la especie respectiva experimente dolor. La evidencia de estudios conductuales sugiere que los crustáceos decápodos tienen la capacidad de “experimentar dolor/sufrir” porque muestran respuestas consistentes con el dolor y tienen una capacidad cognitiva relativamente compleja. Sneddon [14] amplió estos criterios (Tabla 1), pero los criterios han sido cuestionados por “poner el listón demasiado bajo para el dolor”. Hay que tener en cuenta que la introspección es probable y que el dolor animal no es necesariamente el mismo que nuestros propios sentimientos, particularmente en especies filogenéticamente distantes como los crustáceos [60, 61]. De hecho, el dolor describe una experiencia emocional humana y la analogía con cualquier animal no puede probarse en última instancia. Cuanto mayor es la distancia filogenética, es razonable preguntarse hasta qué punto la experiencia de los estímulos nocivos es análoga a la experiencia humana. Pragmáticamente, se podría definir el dolor como una respuesta no reflexiva a un estímulo nocivo, potencialmente dañino para los tejidos, que alteran futuros comportamientos [17]. Esta definición se aleja de la idea de sentimientos
o conciencia, a los cuales es imposible acceder o comprobar.
Los crustáceos heridos muestran actividades como frotarse, cojear o acicalarse que indican conciencia del sitio de la herida y algunos intentos de reducir un daño mayor [15, 24]. De hecho, este comportamiento se ha interpretado como compatible con el dolor [62]. En casos extremos, los apéndices lesionados pueden retirarse (autotomía). Se ha sugerido que la autotomía está mediada por una experiencia similar al dolor [61, 63]. Además, la autotomía aumenta la mortalidad en los crustáceos [64]. Además, la autotomía puede reducir la biomasa recolectada porque los individuos utilizan recursos energéticos para volver a hacer crecer las extremidades a expensas del crecimiento corporal [65]. La langosta de río Procambarus clarkii tiende a preferir ambientes oscuros que claros. Sin embargo, si se exponen por primera vez a una descarga eléctrica que induce respuestas de escape, muestran una evitación mucho mayor de los entornos luminosos y parecen volverse reacios al riesgo, lo que se ha descrito como “mostrar ansiedad”. Esta “ansiedad” va acompañada de niveles más altos de serotonina (5HT) en el cerebro [66, 67]. Al mismo tiempo, la estimulación nociva induce la liberación de la hormona hiperglucémica de los crustáceos (CHH) que es análoga a la hormona del estrés cortisol, en el sentido de que moviliza reservas de energía para restaurar la homeostasis (Figura 1). Además, como consecuencia, el lactato también se elevó en las langostas impactadas, lo que documenta la interrelación entre el dolor y el estrés.
Se ha argumentado que los decápodos no tienen un sistema nervioso complejo de gran tamaño. Aun así, algunos decápodos poseen un cerebro tan grande como el de algunos peces y tienen un alto grado de compartimentación funcional que permite procesos cognitivos sorprendentemente avanzados [68, 69]. Sin embargo, los criterios descritos por Sneddon et al. [15] (Tabla 1), han sido cuestionados [18] y se necesita más investigación. Algunas de las preguntas a abordar están incluso relacionadas con aspectos funcionales como la posesión de un sistema opioide que module la estimulación nociva, una descripción morfológica/funcional de los
nociceptores o la interrelación entre la percepción del dolor y la angustia. Además, se ha planteado el uso de estímulos nocivos verdaderamente relevantes –por ejemplo, en términos ecológicos– [18].
Angustia
Definimos angustia como la exposición prolongada a un factor estresante o a factores estresantes repetidos con un impacto grave en la homeostasis con efectos adversos y duraderos en el desempeño general del animal (respuesta al estrés terciario). De hecho, el sistema de estrés altamente evolucionado de los vertebrados tuvo su origen en el sistema nervioso de los invertebrados [26], que se describirá para el camarón en las siguientes secciones.
Respuesta al estrés primario y secundario en crustáceos. En los invertebrados, numerosos estudios han identificado un sistema neuroendocrino análogo al sistema hipotalámico-hipofisario de los vertebrados, con sustancias muy parecidas a los neuropéptidos y hormonas de los vertebrados, como por ejemplo la oxitocina, la vasopresina, la adrenocorticotropina (ACTH), la hormona estimulante de los melanocitos α (MSH), somatostatina y otros [26, 70–72]. Hace más de 600 millones de años, las células neuroendocrinas especializadas ya estaban presentes en los cnidarios y la señalización neuroendocrina persiste tanto en los linajes de protostomados como en los deuterostomados [72-75]. Sin embargo, los sistemas endocrinos de artrópodos y vertebrados evolucionaron de forma independiente y diferente desde hace más de 540 millones de años [72]. Dado que los artrópodos perdieron la capacidad de sintetizar colesterol [76], los esteroides, presumiblemente, tuvieron oportunidades limitadas de evolucionar dentro de este linaje, ya que el colesterol es el precursor de las hormonas esteroides [72].
Un actor clave del sistema endocrino en decápodos son las hormonas hiperglucémicas de crustáceos (CHH) derivadas del tallo ocular. Esta neurohormona participa principalmente en la homeostasis de la glucosa [77] (Figura 2). Lorenzon et al. reportaron que la CHH está involucrada en una respuesta al estrés después de un estrés ambiental, como temperaturas
elevadas [80], inyección de lipopolisacáridos [81], infección con el virus del Síndrome de la Mancha Blanca [82], transporte [83], cambio de salinidad [80] o una exposición a metales pesados [84]. La CHH se origina principalmente en el pedúnculo ocular, que comúnmente se considera un centro neuroendocrino integrador [78, 85]. Aquí, la CHH es sintetizada por el órgano X del pedúnculo y posteriormente se almacena en la glándula sinusal junto al seno sanguíneo [78]. Se han identificado dos variantes de empalme, una larga (CHH-L) y una corta (CHH), que comparten la misma secuencia para los primeros 40 residuos pero que a partir de entonces difieren considerablemente [78, 86]. La sub o neofuncionalización de estos parálogos aún debe resolverse antes de que la CHH pueda usarse como marcador de estrés.
Durante la ecdisis, la CHH alcanza su punto máximo en la hemolinfa [78, 87]. Aun así, este pico se origina en el intestino, no en el pedúnculo [88]. Tras la secreción de CHH, la movilización de glucógeno en el tejido objetivo aumenta la glucosa y estimula la glucólisis y el ciclo del ácido cítrico (CAC) (Fig. 2). Paralelamente, la lipólisis aumenta y estimula aún más el CAC. Al igual que en los vertebrados, la glucosa y el lactato en la hemolinfa pueden utilizarse como generadores de estrés [89]. De hecho, el lactato se considera el principal producto final en condiciones de estrés [90].
Al mismo tiempo, la CHH restablece las concentraciones de Na+ y K+ en la hemolinfa a los niveles previos al estrés [80]. En el camarón blanco del Pacífico Penaeus vannamei, la CHH aumentó la eliminación de patógenos y la supervivencia de camarones infectados con patógenos [91], elevó el recuento de hemocitos y la actividad fagocítica de los hemocitos [92] y reguló positivamente la expresión de varios genes inmunes, incluyendo los péptidos antimicrobianos y el superóxido dismutasa [92, 93]. Además, la CHH inhibe la síntesis de ecdiesteroides y, por tanto, puede ralentizar el crecimiento [87, 94, 95].
Lorenzon et al. han demostrado que la serotonina (5HT) aumenta los niveles de glucosa, mientras que la dopamina (DA) disminuye los niveles de glucosa en la
Tabla 1: Criterios de dolor según Sneddon et al. [15].
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
Lipólisis
Nocicepción por nociceptores
Procesamiento central de la nocicepción que involucra un órgano similar al cerebro que regula el comportamiento
Procesamiento nociceptivo de moduladores sensibles a endógenos (opioides y receptores opioides)
La nocicepción activa respuestas fisiológicas (cambios en la respiración, la frecuencia cardíaca o los niveles hormonales)
Evidencia de que las respuestas no son solo un reflejo nociceptivo (modulación fisiológica y conductual)
Alteraciones de la conducta que reducen los encuentros con el estímulo (aprendizaje evitativo)
Comportamiento protector, como protegerse las heridas, cojear, frotarse o lamerse
Todo lo anterior reducido por analgesia o anestésicos locales
Autoadministración de analgesia
Hay un costo para acceder a la analgesia
Se observa una atención selectiva después de un estímulo nocivo tiene alta prioridad sobre otros estímulos (por ejemplo, menor rendimiento en tareas de aprendizaje)
Comportamiento alterado después de una estimulación nociva donde se pueden observar cambios en paradigmas de evitación de lugares condicionados y de aprendizaje
Aprendizaje de alivio (“Relief learning”)
Cambio duradero en un conjunto de respuestas, especialmente aquellas relacionadas con evitar la estimulación nociva repetida
Evitación del estímulo nocivo modificado por otros requisitos motivacionales como en las compensaciones
Evidencia de pagar un costo para evitar el estímulo nocivo
Glucógeno
Glucogenólisis
Hepatopáncreas
ADP
Glucosa
órgano X Glándula sinusal
Células neurosecretoras
cHH Transcripción Pedúnculo
Ciclo del ácido cítrico
Acetil-CoA
Piruvato
Glucosa
ADP ATP ATP
Glucólisis
Glucólisis
Músculo
Glucosa
Glucógeno
Glucogenólisis
Figura 2: Papel metabólico de la cHH en el hepatopáncreas y el músculo de los crustáceos. FFA: ácidos grasos libres (modificados de Chen et al. [78] y Li et al. [79]).
Glucosa
mayoría de las especies [85]. Este efecto está mediado por la hipoglucemia [101, 102] de manera específica en cada especie. Después de la ablación del pedúnculo no se observan efectos, lo que confirma firmemente la participación de la CHH del pedúnculo [85]. Los péptidos CHH exhiben una variedad mucho mayor con comúnmente cuatro o más isoformas diferentes [103, 104]. En consecuencia, debido a la subfuncionalización o neofuncionalización, el uso de “bienestar” como indicador necesita una evaluación cuidadosa en las especies respectivas. Los cambios en la 5HT en el cerebro parecen inducir comportamientos similares a la ansiedad en las langostas de río de una manera dependiente de la intensidad [66].
Esto puede sugerir una interrelación entre un supuesto evento estresante y una sensación similar al dolor en este caso. Además, la inyección de 5HT indujo un comportamiento similar a la ansiedad y podría prevenirse mediante la inyección de antagonistas de 5HT [66, 67]. Por lo tanto, al igual que en los mamíferos, los crustáceos parecen exhibir una respuesta de estrés metabólico y conductual [66].
Se sabe que la confrontación con una variedad de sustancias químicas y estímulos ambientales, como destellos de luz o contacto, pueden detener los latidos del corazón y la respiración de los crustáceos durante períodos cortos (bradicardia cardíaca y/o apnea respiratoria), aumentando el lactato como parte de una respuesta antidepredadora normal que podría ocultar al crustáceo de los depredadores que lo atacan y que utilizan campos eléctricos débiles para detectar presas (como los tiburones y rayas) [105, 106].
Respuesta terciaria y angustia
Como se ha señalado, la secreción de CHH puede interferir con la respuesta inmunitaria a gran escala, lo que incluye un recuento reducido de hemocitos totales o un tiempo de coagulación prolongado [107-109]. Además, la CHH alterada puede afectar la muda y, por tanto, inhibir el crecimiento [110]. Los efectos sobre la osmorregulación se han observado en varias especies y pueden aumentar el gasto energético a gran escala [111]. Aun así, rara vez se describen
mecanismos específicos. En este contexto, se ha reportado de una modulación del ARNm de Na+-K+-ATPasa [112]. De forma crónica, el metabolismo energético se ve afectado por este estrés osmorregulador [113]. La baja salinidad y las altas densidades también reducen la capacidad del camarón blanco del Pacífico para hacer frente al estrés agudo [114]. Aunque el camarón blanco tolera una amplia gama de salinidades, la salinidad por debajo del rango óptimo afecta el crecimiento y la supervivencia. Además, la baja salinidad reduce la capacidad del camarón para hacer frente a un factor estresante agudo, por ejemplo, hipoxia o reacción de escape [114].
Altas densidades de población en el cultivo de camarón blanco del Pacífico influyen en la actividad de, por ejemplo, el superóxido dismutasa, la catalasa, glutatión peroxidasa y la proteína de choque térmico para afrontar este estrés crónico [115]. Además, las temperaturas de cultivo subóptimas causan estrés en los camarones: las bajas temperaturas son particularmente problemáticas para el camarón blanco del Pacífico [116] y conducen a un aumento de los niveles de dopamina y CHH en la hemolinfa, mayores recuentos de hemocitos, autofagia y una alteración del metabolismo de las grasas y las proteínas. [117]. En el cultivo de camarón puede ocurrir un suministro de oxígeno subóptimo y la hipoxia recurrente tiene un impacto severo, afectando negativamente la supervivencia, el crecimiento y la susceptibilidad a las enfermedades [118].
Aunque la autotomía a menudo se considera un indicador de bienestar, se ha demostrado que no es más estresante que el manejo normal, exhibiendo niveles similares de lactato y glucosa en el cangrejo comestible Cancer pagurus [119].
El aspecto faltante: medidas de bienestar basadas en expresiones conductuales Como se mencionó anteriormente, la evaluación del estrés, el sufrimiento y el dolor como base para garantizar el bienestar y la salud animal y un cultivo sostenible y apropiado para las especies, se necesita una combinación de parámetros conductuales, morfológicos y fisiológicos. En la práctica, sin embargo, se puede considerar que
los indicadores conductuales de estrés, sufrimiento y dolor promueven la atención sanitaria preventiva y, por tanto, la detección temprana del estrés, las lesiones y las enfermedades, pero aún no se ha logrado su fundamento científico mediante su combinación con indicadores fisiológicos o morfológicos en crustáceos.
Además de la detección del estrés y el sufrimiento, el conocimiento de los patrones de comportamiento expresados por los crustáceos durante todas las etapas de su vida ayuda a promover medidas cruciales en la acuicultura, como la eficiencia alimentaria, la tasa de crecimiento y el éxito reproductivo [120]. Por ejemplo, el conocimiento de los patrones de alimentación de una especie ayuda a perfeccionar las prácticas alimentarias [121]. Verificar el momento del día en el que es más probable que los camarones realicen comportamientos de búsqueda permite suministrar alimento en el momento en que es más probable que los camarones lo coman, reduciendo así el desperdicio de alimento y asegurando una mejor calidad del agua [122]. Para una reproducción exitosa, el conocimiento de los patrones de actividad durante el ciclo de muda ayuda a seleccionar prácticas de manejo apropiadas que reducen el estrés y mejoran la producción de larvas. Por lo tanto, las observaciones de comportamiento son ya una observación de suma importancia para el productor.
Además de los indicadores conductuales de estrés, sufrimiento y dolor o aquellos que mejoran el cultivo, la “libertad de expresar el comportamiento natural” se considera fundamental para el bienestar animal [123]. Estas consideraciones surgen del supuesto de que los animales necesitan expresar su repertorio conductual innato o “natural” para poder vivir en un estado positivo (“ser felices”).
Sin embargo, el comportamiento natural solo puede expresarse en la naturaleza porque los entornos en los que se encuentran los animales en cautiverio se desvían sustancialmente del entorno en el que viven sus ancestros “salvajes” (en cautiverio: falta de depredadores, suficiente comida de alta calidad, espacio restringido, individuos a menudo antinaturales, densidades, etc.).
Así, varios comportamientos naturales o no se expresan (reacciones de evitación de depredadores) o cuando se expresan, se vuelven desadaptativos en cautiverio y luego son incluso perjudiciales para la salud del animal [123]. Por ejemplo, las interacciones agresivas entre congéneres son adaptativas en la naturaleza cuando los individuos defienden ciertos escasos recursos, como parejas para apareamiento o fuentes de alimento [124].
En cautiverio, donde no hay necesidad de defender la comida o las parejas de apareamiento y las limitaciones de espacio impiden escapar, la agresión induce estrés y lesiones y, por tanto, es perjudicial para el bienestar animal. Entonces se requiere que los cuidadores tomen medidas para reducir o suprimir la motivación para expresar estos comportamientos naturales, pero ahora desadaptativos. Por lo tanto, en la producción de organismos (que no son peces), la agresión a menudo se suprime mediante una mayor densidad de cultivo lo que impide que los animales muestren motivación para establecer territorios que quieren defender o proporcionando mejoras estructurales (refugio y espacios para esconderse) que permitan evitar encuentros con sus congéneres.
Para poder identificar estados negativos, promover prácticas culturales beneficiosas y permitir que los animales expresen comportamientos necesarios para su bienestar, defendemos que un primer paso debe ser el establecer qué comportamientos son normales bajo ciertas condiciones ambientales [122]. “Normal” aquí significa que estos comportamientos se expresan de manera predecible en condiciones ambientales similares. En un segundo paso, se deben combinar indicadores fisiológicos y morfológicos para evaluar la importancia de estos comportamientos para el bienestar del animal, por ejemplo, para categorizar ciertos comportamientos como promotores o decrecientes del bienestar animal.
Metodológicamente, los avances en los sistemas de cámaras y el software automatizado de seguimiento de animales nos permiten estudiar los animales de granja con gran detalle [110]. Por ejemplo, las medidas de actividad de movimiento que
informan sobre la fase de muda o la búsqueda de alimento, así como el establecimiento de la prevalencia de encuentros agresivos y las distancias interindividuales, tienen un gran potencial para complementar los indicadores fisiológicos y morfológicos del bienestar animal. En piscicultura, se han establecido con éxito sistemas de monitoreo por video en tiempo real [125-130], pero en su mayoría faltan en el cultivo de camarón [121, 131]. Esto se debe en parte a la alta turbiedad al menos en los sistemas abiertos, mientras que el sistema biofloc depende completamente del monitoreo de los indicadores de bienestar fisiológico [132].
Implicaciones para buenas prácticas acuícolas
El objetivo de las buenas prácticas en la acuicultura es una producción sostenible y segura. Además del uso responsable de los recursos, el bienestar animal es fundamental. Los organismos que se producen en acuicultura pueden experimentar estrés de diversas formas. La comprensión de las interrelaciones es necesaria para reconocer los problemas y, a su vez, compensarlos.
Desde una perspectiva aplicada, el ciclo de producción se divide en períodos, y los períodos respectivos se discutirán con respecto a las principales preocupaciones para el bienestar animal. Se ha afirmado que los estudios sobre el dolor en crustáceos se han basado “en gran medida en unos pocos, dudosos y controvertidos estudios realizados en un pequeño número de animales” [18]. Desgraciadamente, el bienestar de los crustáceos en relación con su relevancia para la acuicultura apenas se ha evaluado sistemáticamente. De hecho, la mayor parte del trabajo se centra en el estrés oxidativo (Tabla 2). Aquí presentamos estudios seleccionados que se han llevado a cabo con respecto a los factores estresantes más comunes. El establecimiento de parámetros confiables para la evaluación del bienestar en la producción puede respaldar el desarrollo futuro de la tecnología existente.
En los crustáceos, se ha llevado a cabo una reducción del estrés para reducir la susceptibilidad a enfermedades [144-146], mejorar el crecimiento y la conversión alimenticia después de una mejor asignación de energía [110, 144, 147, 148], y reducir
el canibalismo y la agresión dentro del sistema de cultivo [149-152]. De hecho, se puede utilizar una evaluación de estrés para optimizar y monitorear integralmente las condiciones de cultivo (parámetros del agua, densidades de población, régimen de alimentación, entre otros) en términos de buenas prácticas acuícolas. Por lo tanto, en el futuro se necesitan evaluaciones en condiciones realistas y a mayor escala. Para hacer cumplir las mejores prácticas acuícolas y de bienestar, Birch et al. [153] recomendaron prohibir la venta viva de decápodos a manipuladores no expertos ni capacitados. Sin duda, poner fin a esta práctica mejoraría el bienestar animal de los decápodos.
Reproductores y reproducción
Los reproductores se mantienen en condiciones óptimas en términos de densidad, parámetros hídricos y alimentación para lograr el mejor rendimiento reproductivo posible. Para una reproducción óptima, la maduración aún se induce mediante la ablación del pedúnculo. Este procedimiento reduce la secreción de la hormona inhibidora de la muda (MIH) del órgano X del pedúnculo y reinicia la maduración de las gónadas.
El uso de pinzas calientes esterilizadas con llama garantiza una ablación y esterilización rápida (<2 s) de la herida [18]. En algunas especies (por ejemplo, Penaeus monodon, Penaeus vannamei), el manejo reproductivo eficiente requiere dicha ablación. Las consecuencias negativas incluyen desequilibrios fisiológicos [154, 155], agotamiento reproductivo [154, 155], trauma físico [156], regulación de genes relacionados con el sistema inmunológico [157, 158], estrés [156, 157] y mortalidad. Desde una perspectiva a largo plazo, la maduración predecible y el desove eficiente se han considerado “objetivos de oro” [159, 160]. Recientemente, se ha logrado un desempeño reproductivo comparable con hembras no sometidas a ablación en Penaeus vannamei, allanando el camino para un mejor manejo reproductivo [160]. De hecho, hay un par de laboratorios de camarón que producen sin realizar una ablación del pedúnculo [27].
Precría
Comúnmente, para aumentar la calidad, el
BIENESTAR ANIMAL
Tabla 2: evaluación de estrés en crustáceos decápodos y sus respectivos parámetros.
Estresor
Exposición al aire
Exposición al aire a mayor T
Marcador SOD
Mortalidad
TAC
Glucosa
Lactato
pH SOD CAT GST
Hemocianina
Glucosa
Lactato
Glucógeno
THC SOD PO
Amoníaco
Nocicepción
pH (ácido, alcalino)
Salinidad
Densidad de población
Temperatura
Turbiedad
Na-K-ATPasa ARNm
THC SOD HSP70 ARNm iNOS
SOD ARNm CAT ARNm
GPx ARNm ROS
THC
Glucosa
HSP70 ARNm
HSP90 ARNm
Na+-K+-ATPasa ARNm
GSH
GPx CAT HSP70
THC PO NOS SOD
Transcriptómica HSPs
THC SOD PO CAT
Hematocrito Glucosa
Lactato, glucosa
Histología de branquias
THC SOD PO
Branquias ensanchadas
Efecto
Aumentó
Aumentó
Aumentó
Ningún cambio
Aumentó
Disminuido
Aumentó
Aumentó
Aumentó
Disminuido
Aumentó
Aumentó
Disminuido
Ningún cambio
Disminuido
Disminuido
Ningún cambio
Disminuido
Aumentó
Aumentó
Aumentó
Aumentó
Aumentó
Aumentó
Ligeramente disminuido
Ligeramente incrementado
Aumentó
Aumentó
Aumentó
Aumentó
Aumentó
Disminuido
Ningún cambio
Aumentó
Primero aumentó y luego disminuyó
Primero aumentó y luego disminuyó
Primero aumentó y luego disminuyó
Diferencialmente regulado
Aumentó
Disminuido
Insignificante
Aumentó
Insignificante
Aumentó
Insignificante
Aumentó
Especies
Camarón blanco del Pacífico Penaeus vannamei
Cangrejo
Liocarcinus depurator
Falsa centolla
Paralomis granulosa
Cangrejo azul asiático
Portunus trituberculatus
Camarón blanco del Pacífico Penaeus vannamei
Camarón de río amazónico
Macrobrachium amazonicum
Jaiba japonesa
Charybdis japónica
Cangrejo costero Hemigrapsus sanguineus
Camarón blanco del Pacífico Penaeus vannamei
Camarón blanco del Pacífico Penaeus vannamei
Cangrejo chino
Eriocheir sinensi
Cangrejo de río del Danubio
Astacus leptodactylus
Camarón blanco del Pacífico Penaeus vannamei
Camarón
Palaemon spp
Cangrejo mediterráneo Carcinus aestuarii
Camarón blanco del Pacífico Penaeus vannamei
Fusión lamelar, hiperplasia, hipertrofia
Aumentó
Aumentó
Aumentó
Aumentó
CAT: catalasa; FCR: índice de conversión de alimenticia; GSH: glutatión; GST: glutatión-S-transferasa; GPx: glutatión peroxidasa; HSP: proteína de choque térmico; NOS: óxido nítrico sintasa; PO: fenoloxidasa; SOD: superóxido dismutasa; TAC: capacidad antioxidante total; THC: recuento total de hemocitos; GT: aumento de peso.
crecimiento y la supervivencia del camarón, se lleva a cabo una fase de precría antes de la fase de engorde. Un método común para evaluar y garantizar la calidad de las postlarvas es realizar “pruebas de estrés” en pequeños lotes de postlarvas [154, 161-163]. De hecho, estas pruebas son bastante letales y el diagnóstico resultado del comportamiento subsecuente de las postlarvas puede ser cuestionado. Las larvas están expuestas a niveles de estrés muy altos (p. ej., salinidad, temperatura, amoníaco y formalina) y se determina el número de supervivientes.
Este procedimiento altera profundamente los mecanismos metabólicos y provoca la muerte de los individuos más débiles. En el futuro, los marcadores moleculares y las pruebas de comportamiento pueden proporcionar herramientas de evaluación más sensibles y significativas. De hecho, el resultado de las pruebas de mortalidad a menudo no está estrechamente correlacionado con el desempeño posterior durante el cultivo, y se requieren ensayos más confiables.
Engorde
Desde el punto de vista del productor, la fase de engorde es sin duda el período más importante en términos de éxito comercial, ya que implica el mayor gasto y determina los ingresos. La interacción entre las condiciones fluctuantes, y en algunos casos subóptimas, y la salud animal desempeña un papel crucial para el éxito económico.
La turbidez del agua en el cultivo intensivo de camarón es un factor estresante importante, particularmente en estanques de tierra con aireadores (Tabla 2). El aumento de partículas de arcilla induce el aumento en los niveles de lactato y glucosa, y derivan en una mayor tasa de mortalidad [89]. Además, la concentración de partículas induce una elevación de los biomarcadores de estrés oxidativo, como la actividad del superóxido dismutasa (SOD), y da lugar a un elevado recuento total de hemocitos (THC).
Realizar la evaluación de estrés basada en marcadores permitió recomendar niveles seguros de turbidez (<30 NTU). En otro estudio, después de la exposición al amoníaco, la SOD disminuyó, mientras que el THC no se vio afectado [136]. Bajo un pH
ácido y alcalino, los marcadores de estrés oxidativo aumentaron sustancialmente, pero volvieron a la normalidad después de un día, lo que sugiere que la restauración adaptativa de la homeostasis se produjo relativamente rápido [139]. En Astacus leptodactylus, el glutatión (GSH) y la SOD, así como la proteína de choque térmico, aumentaron con una mayor densidad de población [140], lo que sugiere una fuerte respuesta al estrés oxidativo ante el hacinamiento.
En juveniles de Penaeus vannamei, se observaron cambios sustanciales en el comportamiento al aumentar la densidad de población [122]. En este caso, se puede utilizar una evaluación basada en etogramas para caracterizar el estrés social y recomendar densidades de población optimizadas [122]. Aunque los camarones se consideran menos caníbales [152], el canibalismo puede exacerbarse cuando enfermedades o condiciones ambientales (por ejemplo, alto nivel de ozono, bajo nivel de oxígeno disuelto) inducen exoesqueletos blandos [149, 164, 165].
Los períodos más prolongados de alimentación subóptimos afectan gravemente al camarón, causando pérdidas económicas potenciales a los acuicultores, lo que debe evitarse [166]. También se ha reportado que la inanición induce inmunidad celular en Penaeus vannamei y afecta la resistencia a las enfermedades. La mayoría de las veces, con excepción del RAS, el crecimiento se produce en sistemas abiertos, por lo que están expuestos a influencias ambientales fluctuantes. Especialmente, los cambios de temperatura y salinidad se identificaron como factores estresantes en estas condiciones. El estrés por baja salinidad influye directamente en el crecimiento y en los genes relacionados con la osmolalidad y el metabolismo [112]. El camarón blanco del Pacífico responde a rápidos cambios de temperatura activando enzimas antioxidantes y biomarcadores relacionados con el estrés [167].
Como conclusión, respecto a los biomarcadores de estrés evaluados se deben analizar cuidadosamente y se necesitan marcadores crónicos específicos de angustia para optimizar la tecnología existente y definir rangos para salvaguardar a los animales. Hasta ahora, se han evaluado
muy pocos parámetros (principalmente biomarcadores de estrés oxidativo) y existe una necesidad urgente de marcadores universales relacionados con el estrés.
Cosecha
La cosecha es uno de los pasos más estresantes durante el ciclo de producción e inevitablemente implica manipulación física. Los animales se juntan y se sacan del agua mediante redes o bombas. Este procedimiento desencadena una conducta de huida y representa estrés para los animales, lo que puede provocar lesiones físicas y reducir la calidad de la carne. En estas circunstancias, la frecuencia cardíaca aumenta sustancialmente [168]. Además, el amoníaco, la glucosa y el lactato aumentan [169, 170]. Al llegar a una fábrica de procesamiento, los crustáceos a menudo (rara vez el camarón) se vuelven a sumergir, lo que permite cierta recuperación [171, 172]. Después de sacarlos nuevamente del agua, los individuos se clasifican inmediatamente y se transfieren a agua helada para aturdirlos y matarlos.
Los crustáceos, como las langostas y los cangrejos, suelen mantenerse vivos durante largos períodos antes de sumergirlos en agua hirviendo. Por lo tanto, es razonable preguntarse si los animales tienen algún grado de conciencia y experimentarán el tratamiento como doloroso.
Anestesia
Los decápodos se pueden insensibilizar con una amplia variedad de anestésicos, que se administran mediante inyección o como baño anestésico con fines experimentales [173, 174]. Dado que los crustáceos procedentes de la acuicultura se producen para el consumo humano, el uso de narcóticos químicos está prohibido por motivos de seguridad humana. Para el aturdimiento de camarones tropicales, actualmente se recomiendan dos métodos estandarizados principales: el enfriamiento y el electroshock [175]. Un método viable para anestesiar langostas (Homarus americanus) es aumentar lentamente la temperatura, ya que la actividad neuronal de los animales se detiene a temperaturas más altas sin provocar anomalías de comportamiento.
El enfriamiento no es efectivo en esta
especie, ya que los animales aún reaccionan a estímulos externos [176]. Por el contrario, el enfriamiento estandarizado se utiliza con éxito en camarones tropicales y generalmente se lleva a cabo sobre hielo. La inmersión en hielo durante >1 min es una forma eficaz de sacrificar la mayoría de las especies de camarón cultivados a temperaturas de agua tropicales. Dado que las bajas temperaturas no activan los nociceptores en algunos crustáceos [49], la suspensión de hielo parece ser un método no químico, muy eficaz y fácil de realizar para el aturdimiento.
Sin embargo, en algunos países, como por ejemplo en Suiza o Italia, este método no está reconocido por la ley. De hecho, se sospecha que el enfriamiento solo puede paralizar a los animales sin anestesiarlos. El “electroaturdimiento” es controversial ya que induce ataques epilépticos en langostas y langostas de río [176]. El electroaturdimiento parece paralizar especies como cangrejos y langostas [176-178], pero la evidencia de su eficacia es escasa [175]. En el caso del camarón, se puede recomendar el aturdimiento eléctrico [178].
Transporte
Varias especies de crustáceos se comercializan como productos vivos (revisión, ver [179]). Se ha reconocido que un refugio adecuado mejora el bienestar de los animales durante el transporte [153]. La variación de temperatura y la exposición al aire son factores estresantes reconocidos para especies transportadas vivas [135]. Además, la manipulación, el aumento de densidades y el deterioro de los parámetros del agua, como la concentración de dióxido de carbono y amonio, son estresantes, especialmente para P. vannamei [110, 180].
Se han establecido varias pautas para el transporte (ver, por ejemplo, [181, 182]), basadas en parte en la evaluación de marcadores relacionados con el estrés. Generalmente, en la mayoría de los países el transporte está permitido en hielo, pero en Suiza debe realizarse en agua de mar.
Aunque algunos crustáceos respiran aire facultativamente, como Carcinus maenas, el transporte sobre hielo es un factor potencialmente estresante que puede alterar el consumo de oxígeno para la mayoría de las
especies, lo que lleva a una acumulación de metabolitos anaeróbicos como el lactato.
La exposición al aire también induce la CHH, lo que desencadena una respuesta de estrés [22, 183]. Para promover el letargo y reducir el estrés, los crustáceos vivos se enfrían antes del transporte.
Durante el enfriamiento, la autotomía ocurre con frecuencia en algunas especies, aunque la interpretación de la autotomía con respecto a su relevancia para su bienestar no está clara. A velocidades de enfriamiento más lentas, la autotomía se reduce [179]. Como recomendación, una temperatura relativamente baja (5-10°C), una atmósfera húmeda y evitar perturbaciones mejoran el bienestar de los animales durante el transporte [179]. En conjunto, el transporte es particularmente desafiante y causa un estrés considerable a los animales transportados. Por lo tanto, los parámetros de cultivo y manipulación durante el transporte deben mantenerse dentro de un rango óptimo para las diferentes especies.
Sacrificio compasivo
Estudios recientes demostraron que algunas respuestas de los decápodos no pueden explicarse como reflejos [17, 61, 184], lo que abre la posibilidad de que puedan experimentar dolor. Recientemente, existe una preocupación sustancial de los consumidores sobre el sacrificio de ciertos crustáceos decápodos; por ejemplo, la langosta y el sacrificio compasivo han sido objeto de estudios recientes [185].
En Suiza y Nueva Zelanda se ha prohibido hervir langosta viva y es necesario aturdirla antes de hervirla, pero en la mayoría de los países los decápodos todavía están excluidos de la legislación sobre bienestar animal.
El método de sacrificio más “humano” o compasivo es probablemente un dispositivo de aturdimiento eléctrico llamado CrustastunTM [175] para animales más grandes como la langosta. Aun así, en animales más pequeños como el camarón, este método no ha sido certificado hasta el momento, lo que requiere más investigaciones [153].
El aturdimiento eléctrico mata al animal en
un segundo y no se ha observado estrés en los animales transferidos al dispositivo (monitoreado como concentración de lactato).
La inmersión en una suspensión de hielo causa igualmente sedación en el camarón y langosta de río en cuestión de minutos, pero los cangrejos permanecen sensibles durante algún tiempo y conservan un circuito neuronal funcional. La división es una técnica de sacrificio compasivo en la langosta, pero requiere experiencia. En este caso, el sistema nervioso se destruye cortando a lo largo de la línea media longitudinal [176].
Conclusiones
Especialmente en el caso de los crustáceos decápodos, el crecimiento de la acuicultura está superando los conocimientos sobre bienestar. Con respecto al debate en curso sobre el bienestar de los decápodos, se requiere una mejor comprensión de los mecanismos que subyacen a la nocicepción, el dolor y la angustia para salvaguardar un buen cultivo de animales de acuerdo con el concepto de las Cinco Libertades.
La evidencia científica sobre la percepción del dolor se basa principalmente en estudios de comportamiento y hasta el momento no se han identificado mediadores relacionados con el dolor, como los receptores opioides o los PRT relacionados con los nociceptores. En gran medida, faltan estudios que midan los parámetros fisiológicos en el seguimiento rutinario de la acuicultura.
Es necesario contar con biomarcadores fisiológicos de fácil acceso y estudios conductuales integrales para monitorear el bienestar en la práctica. Sin duda, se deben utilizar consideraciones de bienestar con base científica para mejorar aún más la tecnología existente, comunicar el bienestar a los clientes y salvaguardar mejor el bienestar en el cultivo de crustáceos•
Para más información sobre este artículo, contactar a: sven.wuertz@igb-berlin.de
Niveles de sulfito en langostinos y camarones congelados
Autor:
Yahira Piedrahita
Directora Ejecutiva de la Cámara Nacional de Acuacultura
ypiedrahita@cna-ecuador.com
El sulfito y sus derivados son comúnmente usados en España como aditivos en una variedad de alimentos, actuando como antioxidantes y conservantes. También se emplean para prevenir la decoloración enzimática y no enzimática. La adición de sulfitos a langostinos y camarones crudos ha sido una práctica global para evitar la melanosis, un defecto conocido como Blackspot. La Tabla 1 presenta un resumen de las concentraciones de sulfitos en camarones y crustáceos frescos, congelados y enlatados presentados en la literatura. Sin embargo, debido a problemas de salud asociados con los sulfitos, se han propuesto alternativas para prevenir la decoloración. Aunque se han sugerido opciones como el 4-hexilresorcinol, los agentes sulfatados siguen siendo los preferidos para prevenir la melanosis en camarones.
Debido a reacciones adversas, especialmente alergias, causadas por los sulfitos en alimentos, se ha regulado su concentración en productos alimenticios. Las personas asmáticas, en particular, corren un mayor riesgo de tener reacciones adversas a los sulfitos, lo que puede desencadenar ataques de asma. La sensibilidad a los metabisulfitos, utilizados como antioxidantes, también puede desencadenar síntomas de asma en algunas personas.
Aunque todavía no se comprende completamente la fisiopatología de las reacciones adversas al sulfito, la literatura ha propuesto tres posibles mecanismos:
1.Estimación directa de los receptores bronquiales aferentes parasimpáticos después de la inhalación de dióxido de azufre. (SO2). 2.Posible mecanismo relacionado con la inmunoglobulina E (Ig E). 3.a deficiencia o disminución de la enzima sulfito-oxidasa, crucial para la conversión de sulfitos a sulfatos.
La presencia de estos aditivos en los alimentos lleva a la destrucción de la tiamina, lo que ha conducido a la prohibición de su uso en carnes y productos cárnicos en los Estados Unidos. Esta cuestión de salud pública ha generado un escrutinio sobre el uso de sulfitos por parte de las autoridades reguladoras. Varios países han establecido "listas positivas" de aditivos que especifican las concentraciones máximas permitidas en los alimentos. En Europa, las concentraciones permitidas de sulfitos en diversos alimentos están reguladas por la Directiva 95/2/CE.
Se ha observado que el uso de sulfitos excede los límites legales, especialmente en langostinos y camarones congelados, donde el contenido de SO2 total supera los 150 mg/kg. Cabe mencionar que los niveles de sulfito residual varían no solo según las condiciones de tratamiento, sino también en función de las características, manipulación y procesamiento de los productos. Por ello, se realizó un estudio para determinar la concentración de sulfitos en las partes comestibles y no comestibles de muestras de langostinos y camarones congelados de España y Venezuela. Además, se calculó la ingesta de sulfitos para un individuo español promedio de 65 kg de peso corporal al consumir diferentes tamaños de porciones de langostinos y camarones congelados (100 g, 150 g y 200 g), y se comparó esta ingesta con la Ingesta Diaria Admisible (ADI, por sus siglas en inglés) establecida por la FAO/OMS en 1990 para este
aditivo (0.7 mg/kg/día = 45.5 mg/día para una persona de 65 kg de peso corporal).
En el estudio se empleó el método MonierWilliams (AOAC, 1984) para analizar 50 muestras de langostinos congelados y 30 muestras de camarones congelados de España y Venezuela. Se llevó a cabo una etapa de destilación para separar el sulfito de la matriz de la muestra en forma de dióxido de azufre. Posteriormente, el dióxido de azufre fue capturado y se hizo reaccionar con peróxido de hidrógeno para convertirlo en ácido sulfúrico, el cual fue cuantificado mediante titulación con hidróxido de sodio. Se tomaron resultados duplicados de las partes comestibles y no comestibles del producto, y se observó consistencia entre estos duplicados en todo momento.
Para verificar la efectividad del método Monier-Williams, se llevó a cabo un estudio de recuperación con dos enfoques:
1. Se utilizó metabisulfito estándar de 50 mg/l en una muestra con una aplicación previa de sulfitos.
2. Se añadió metabisulfito estándar de 50 mg/l a 200-250 g de muestra sin aplicación previa de sulfitos. En la prueba se encontró que el porcentaje de recuperación (96%) era similar al obtenido por Williams et al. (1990) (97%). En la Tabla 2 se proporcionan los valores promedio, la desviación estándar y las recuperaciones de sulfitos agregados a las muestras para mayor referencia.
Las muestras de langostinos y camarones congelados fueron adquiridas a diversos minoristas y proveedores locales de mariscos, asegurando que fueran representativas de los productos vendidos en España. Posteriormente, las muestras fueron transportadas al laboratorio en contenedores refrigerados pequeños con control de temperatura y almacenadas a -30°C hasta su análisis. Al sacarlas del almacenamiento congelado, las porciones se dejaron descongelar a temperatura ambiente. Para los análisis, las muestras se pelaron y se picaron finamente. La parte comestible se obtenía al retirar la cabeza y pelar las colas, mientras que la parte no comestible incluía la cabeza y la cáscara pelada. Cada muestra constaba de 200-250 g de músculo pelado de langostino o camarón, homogeneizado según el método AOAC.
Tabla 1.- Niveles de sulfitos en alimentos presentados en la literatura
Río Utrabo, Peña Rivas, Cuesta Bertomeu, and Gata Díaz (1994)
Food Research Institute (1994)
Lawrence and Chadha (1988)
Lawrence and Chadha (1988)
De Vries, Ge, Ebert, Magnuson, and Ogawa (1986)
De Vries et al. (1986)
Tabla 2.- Valores promedio, desviaciones estándar y recuperaciones
Sulfito añadido
Muestra con aplicación previa de sulfitos
Valor promedio S.D. Recuperación %
0a 20
Muestra sin aplicación previa de sulfitos
a Realizado sobre 10 alícuotas de la misma muestra.
En la sección de Resultados y Discusión, la Tabla 3 muestra los niveles de sulfito encontrados en las partes comestibles y no comestibles de las 50 muestras de langostinos congelados y 30 muestras de camarones congelados analizadas.
Para la porción comestible de los langostinos congelados, las concentraciones de sulfito oscilaron entre 12.8 y 546.0 mg/kg (valor promedio = 115.8 mg/kg; desviación estándar = 155.7 mg/kg). Se observó que quince muestras tenían una concentración de sulfito superior a 300 mg/kg, siendo el máximo encontrado de 546.0 mg/kg. Estos resultados se encuentran ligeramente por debajo de los valores reportados por Armentia et al. (1994) (182–572 mg/kg) y Río Utrabo et al. (1994) (585.79 mg/kg). Se incluye un histograma que muestra la distribución de los diferentes rangos de concentración de sulfito en los langostinos congelados en la Figura 1.
La concentración de sulfito en las partes comestibles de los camarones congelados varió entre 10.7 y 380.7 mg/kg (con un promedio de 105.3 mg/kg y una desviación estándar de 120.6 mg/kg). Se identificaron 10 muestras con concentraciones superiores a 150 mg/kg y dos muestras que excedieron los 300 mg/kg. Estos resultados son más bajos que los informados por Ellin, Steinhart y Cochrane (1994), quienes encontraron valores de 175 mg/kg. El histograma en la Figura 2 ilustra la distribución de los
diferentes rangos de concentración de sulfito en los camarones congelados.
Según la legislación española actual RD 145/1997 (Ministerio de Sanidad y Consumo, 1997), basada en la Directiva 95/2/CE, el límite máximo permitido en las partes comestibles de langostinos y camarones congelados es de 150 mg/kg. Los valores promedio encontrados en las partes comestibles de langostinos y camarones exceden este límite establecido.
Debido a la falta de información concreta sobre la ingesta diaria promedio de langostinos y camarones, los autores realizaron una estimación considerando que el español promedio consume porciones de 100 g, 150 g o 200 g de langostinos y camarones. Según estas estimaciones, la ingesta de sulfitos sería de 11.58, 17.37 y 23.16 mg para langostinos, y de 10.5, 15.8 y 21.06 mg para camarones respectivamente (ver Tabla 4). Aunque no se han tenido en cuenta posibles pérdidas durante la cocción de los productos, las ingestas resultantes de consumir estas porciones (100 g, 150 g y 200 g) están por debajo de la Ingesta Diaria Admisible (ADI) de sulfitos.
Es importante tener en cuenta que en España es común consumir langostinos y camarones en celebraciones como Navidad, considerándose estos alimentos como productos de lujo debido a su costo. En este contexto, si en estas ocasiones la porción
de langostinos consumida supera los 400 g, la ingesta de sulfitos sería de 46.32 mg, excediendo la ADI (45.5 mg/día) para una persona con un peso corporal de 65 kg. Consumir una porción de 400 g de camarón resultaría en una ingesta de sulfitos de 42.12 mg, casi alcanzando la ADI de sulfitos para una persona con un peso corporal de 65 kg. Los niveles de sulfitos en las partes no comestibles de langostinos congelados muestran un rango de valores mínimo y máximo de 80.7 y 8256.3 mg/kg respectivamente, con un valor promedio de 2428.4 mg/kg y una desviación estándar de 2677.2. En comparación con estudios anteriores, el promedio observado en las partes no comestibles de langostinos congelados (2428.4 mg/kg) resultó ser mayor al reportado por Río Utrabo et al (1994) (1681.0 mg/kg) pero se encontró dentro del rango obtenido por Armentia et al. (1994) (971–2399.0 mg/kg).
Por otro lado, la concentración de sulfitos en las partes no comestibles de camarones congelados tuvo un promedio de 1500.3 mg/ kg y una desviación estándar de 1230.7 mg/ kg, con valores mínimo y máximo de 60.2 y 3420.2 mg/kg respectivamente. Dado que es común en España chupar las cabezas de langostinos y camarones (partes no comestibles), es importante tener en cuenta estas concentraciones de sulfitos. Aunque pudiera haber alguna pérdida de sulfitos durante la cocción, en estas ocasiones en particular, se podría sobrepasar la Ingesta Diaria Admisible (ADI) de sulfitos debido a esta práctica.
En base a los hallazgos, se pueden concluir los siguientes puntos:
- Las concentraciones de sulfitos, tanto en las partes comestibles como en las no comestibles, son más altas en langostinos que en camarones.
- La mayor concentración en las partes no comestibles se debe a la adición específica de sulfitos en estas porciones para prevenir el oscurecimiento.
- La variabilidad de resultados sugiere una adición no controlada de sulfitos a estos productos, a veces superando el límite máximo de 150 mg/kg permitido para las partes comestibles.
- Dado que langostinos y camarones se consideran alimentos de lujo consumidos en
Tabla 3.- Niveles de sulfitos en los langostinos y camarones congelados comercializados en España
Langostinos congelados
Porción comestible
Porción no comestible
Camarones congelados
Porción comestible
Porción no comestible
S.D.: desviación estándar; N: número de muestras.
Muestras con una concentración de sulfitos inferior al límite de 150 mg/Kg. Muestras que superan el límite de concentración de sulfito de 150 mg/Kg.
Fig. 2. Rangos de concentración de sulfitos para las muestras totales de camarón congelado.
Muestras con una concentración de sulfitos inferior al límite de 150 mg/Kg. Muestras que superan el límite de concentración de sulfito de 150 mg/Kg.
ocasiones especiales, la cantidad ingerida diariamente suele ser pequeña, y por tanto, la ingesta de sulfitos proviene principalmente de consumir estos mariscos.
- Debido a que los asmáticos pueden experimentar reacciones adversas a los sulfitos, es importante que estén conscientes de que langostinos y camarones son alimentos que a menudo contienen este aditivo•
Este artículo se basa en la publicación Content of sulphite in frozen prawns and shrimps escrito por A. Hardisson, C. Rubio, I. Frías, I. Rodríguez y J.I. Reguera; publicado en Food Control 13 (2002) 275–279 y disponible en https://doi.org/10.1016/ S0956-7135(02)00022-1. El documento ha sido resumido respetando los derechos de autor.
Fig. 1. Rangos de concentración de sulfitos para las muestras totales de langostino congelado.
Ante el inminente cambio
UAutor: Leonardo S. Maridueña
Simone Maridueña
Dirección de Ambiente Cámara Nacional de Acuacultura
lmariduena@cna-ecuador.com
no de los grandes retos que enfrenta el sector camaronero y gran parte de las industrias a nivel mundial, es la rápida adaptación de su sistema para enfrentar el cambio climático. Hoy en día, las condiciones que antes eran cíclicas, repetitivas y bastante predecibles, están siendo muy variables y difíciles de descifrar. Como consecuencia, aumentan las amenazas y el riesgo de pérdidas durante todas las etapas de producción de camarón.
Actualmente, el cambio climático se evidencia cada vez más a través de eventos extremos, que se conocen como condiciones anormales. Durante las últimas décadas, estos eventos se han presentado como: fenómenos meteorológicos extremos, temperaturas por encima de la media establecida, fenómenos de aparición lenta, cambios en cuanto al tiempo y períodos de las estaciones, eventos de lluvia intensa que causan inundaciones, o sequías muy prolongadas.
La predicción en cuanto a intensidad y tiempo de estos eventos anómalos es muy reducida, al generarse de un momento a otro y con una intensidad muy alta sin tener la oportunidad a reaccionar. Por lo general, estos eventos intensos son los más perjudiciales, puesto que ocasionan graves daños a la infraestructura y erosionan la mayoría de los suelos. En cambio, los eventos que se prolongan en el tiempo provocan la reducción o escasez de los servicios ambientales; un ejemplo de esto es la reducción de caudal de agua para la generación de energía eléctrica y las dificultades de riego en momentos de sequía.
Así mismo, las anomalías pronosticadas pueden cambiar su presencia y efectos, como sucedió con El Niño 2023-2024, que como se observó, se dieron todas las condiciones océano-atmosféricas para que se genere un fenómeno de alta intensidad, pero manifestaciones oceanográficas, como la repentina aparición del Índice de Oscilación en el hemisferio norte cerca de las costas colombianas, debilitaron la fortaleza de El Niño, convirtiéndose en una estación invernal casi normal.
Por otro lado, en los últimos diez años se han observado a nivel global eventos como:
calor extremo, frío extremo, huracanes, inundaciones, incendios forestales, sequía, terremotos, tormentas de nieve, tornados, que por su impredecibilidad se los atribuye al cambio climático, teniendo en consideración que los cambios del clima son manifestaciones extraordinarias que generalmente no obedecen o tienen su génesis antrópicas; sin embargo, desde el siglo XIX, las actividades humanas han sido el principal motor del cambio climático, debido principalmente a la quema de combustibles fósiles, como el carbón, el petróleo y el gas, lo que produce emisiones que atrapan el calor. Los cambios bruscos, producto del cambio climático, también tienen un efecto a nivel de fauna y flora. Los cambios de la temperatura en el agua y suelo, o vientos muy secos o con demasiada humedad, hacen que se pierda el equilibrio entre los factores bióticos que interactúan en él, causando estrés a las especies más vulnerables, la presencia de enfermedades no comunes, como fue la gripe aviar, atribuida a especies de aves migratorias, que en la estación invernal 2023 afectó puntualmente la zona de El Morro y Playas y en especial a las fragatas, muriendo miles de ejemplares de esta especie, en la provincia del Guayas en Ecuador.
Riesgos en la acuicultura
En cuanto a la acuicultura, se considera que las amenazas que más riesgo ocasionan son las lluvias intensas, el aumento o decremento de la temperatura del agua, la potencial acidificación del agua de la zona estuarina, proveniente del océano.
Las lluvias intensas pueden romper los muros de las camaroneras, obstruir vías, causar daños graves a la infraestructura, provocar la pérdida de insumos, entre otros. Por otro lado, el aumento de la temperatura del agua produce estrés en el camarón, permite la proliferación de organismos patógenos y algas nocivas, aumenta la salinidad del agua, reduce la disponibilidad de alimento natural, acidifica la calidad del agua, entre otros cambios conducentes al desequilibrio del ecosistema en que se produce la acuicultura. A todo esto, se añade la posibilidad de temblores y terremotos, como el ocurrido el 16 de abril del 2016 que afectó a las provincias de Manabí y Esmeraldas, causando serios daños a la infraestructura camaronera.
Las Naciones Unidas y sus foros han desarrollado procesos que se deben adoptar para prepararnos y reducir el impacto que el cambio climático podría generar:
Fig. 1 Interpretación al proceso de adaptación al cambio climático establecido en el marco de la Convención de las Naciones Unidas para el cambio climático según la “Introducción al manejo del cambio climático”. John C. Shieder y Jean Hetzel (2021)
Por los motivos mencionados, es prudente que todas las actividades productivas apliquen soluciones rápidas y eficaces, que las conviertan en empresas resilientes al cambio climático. De esta forma, se adaptan a situaciones adversas, reducen su estado de vulnerabilidad y obtienen ventaja ante otras empresas.
Los consultores en acuicultura Kyra Hoevenaars y Jonah van Beijnen, con el apoyo de Doris Soto, se unieron en el 2024 para elaborar un artículo sobre la acuicultura y el cambio climático. Basado en trabajos realizados en acuicultura en pequeña escala en el Sudeste Asiático, Medio Oriente y África, el artículo se titula “Seis formas de hacer que sus operaciones acuícolas sean más resilientes al clima”. Este trabajo ofrece consejos prácticos dirigidos a acuicultores ubicados en los trópicos que trabajan a pequeña escala.
De esta forma, brindan las herramientas necesarias para comenzar a ser resistentes al cambio climático y a los fenómenos meteorológicos extremos. A lo largo del artículo, los autores resaltan la importancia de implementar medidas de adaptación
que guarden relación con las amenazas propias del lugar de ubicación. Además, plantean las características de las medidas, las cuales deben ser: eficaces, contar con la contribución o aceptación de la comunidad, priorizadas en cuanto a la urgencia, responder a la viabilidad técnica, ser fáciles de aplicar y factibles con relación a sus costos.
A continuación, se mencionan los titulares de las recomendaciones indicadas en el artículo de Hoevenaars y van Beijnen (2024). Adicionándole el contenido que aplica a la realidad de la situación de la producción acuícola de camarón en el país, tomando en consideración las condiciones en que se desarrolla el cultivo de camarón.
Recomendaciones
Implementar buenas prácticas de gestión El concepto de buenas prácticas de gestión puede sonar bastante amplio, pero estas están dirigidas a la optimización en los procesos productivos para generar una mayor efectividad. Lo que implica el conocimiento a fondo de todas las actividades, factores que pueden poner en riesgo y la implementación de procesos clave para la obtención de los
resultados, tomando en consideración la salud y bienestar de todos los que forman parte de la cadena productiva.
De acuerdo con lo ya mencionado, el cambio climático dificulta el cumplimiento de los objetivos de la actividad camaronera, porque pone en riesgo la salud del camarón. Este genera estrés y por consiguiente el debilitamiento del sistema inmunológico del camarón, haciéndolo susceptible a cualquier patógeno que se encuentre presente en el medio.
Por esto, es necesario desarrollar un plan de bioseguridad, que tome en consideración la realidad del medio geográfico y otras condiciones en que se desenvuelve el cultivo de camarón. Tal como explica Soto, “mejorar las prácticas de gestión es el primer paso hacia la adaptación al clima, especialmente mejorando la bioseguridad, considerando menores densidades de población y garantizando una buena ubicación de las granjas”. (Hoevenaars & Van Beijnen, 2024) El plan de bioseguridad establece una ruta con las actividades que se realizan en la producción camaronera, analiza los posibles patógenos que pueden ingresar en cada etapa y sus formas de propagación, con el fin de determinar los riesgos en cada parte del proceso productivo. De tal forma que se establezcan las medidas necesarias para la minimización y control de los patógenos, el mantenimiento de las condiciones de higiene, la eficiencia alimentaria, la calidad del agua, entre otros factores a evaluarse.
Además, el plan de bioseguridad pretende reducir el riesgo de contaminación o propagación de enfermedad detectada a otras piscinas, otras fincas o al medio ambiente. Por tal motivo, es importante que todos estén al tanto de las medidas establecidas, mediante la socialización del documento con los trabajadores de la camaronera y con la comunidad dentro del área de influencia.
Es importante que estas actividades mantengan indicadores de cumplimiento que puedan ser evaluados con el paso del tiempo. La actualización del documento debe mantenerse de manera anual, especialmente si se han hecho cambios significativos en la producción, con la inclusión de nuevas
medidas y eliminación de actividades que no fueron eficientes para cumplir con los objetivos planteados o que no aplican a la realidad presente.
Realizar un análisis basado en el riesgo En el artículo técnico proponen llevar a efecto un análisis de riesgo de las amenazas que presenta un área específica, de manera que podemos identificar el nivel de riesgos del área con respecto al desarrollo de las actividades productivas para establecer la factibilidad del área para la instalación de la actividad o las medidas estructurales necesarias para contrarrestar los posibles riesgos.
El análisis de riesgo debe tomar en cuenta la exposición del terreno ante una amenaza como cambio climático y fenómenos meteorológicos. Cada amenaza identificada debe ser calificada de acuerdo con el riesgo, la probabilidad de ocurrencia y los impactos que podría causar. En caso de que alguno de los riesgos aún pueda causar impactos, la empresa se encargará de realizar un plan de contingencia que brinde la respuesta adecuada y la forma en que se debe proceder en caso de alguna eventualidad.
Aunque el análisis de riesgos o las medidas adoptadas no impedirán que un evento extraordinario pueda afectar la producción acuícola, sí será útil para gestionar correctamente o contrarrestar los impactos o
problemas que puedan surgir. Además, una empresa que considera estos riesgos brinda mayor confianza a las partes involucradas, ya que asegura que las decisiones y acciones tomadas están siendo evaluadas cuidadosamente.
En nuestro caso, se puede encontrar información de las amenazas climáticas en la página de la Secretaría Nacional de Gestión de Riesgos,en la que proporcionan los mapas de las amenazas (inundación y zonas de derrumbe), reportes del nivel de amenaza por oleaje, niveles de alerta por zona, entre otros documentos útiles para realizar el análisis de riesgo.
Diversificar la producción
La necesidad de adaptarse a una nueva era de irregularidades en que lluvias, sequías y otros fenómenos naturales se presenten, afectarían, según el efecto, a la producción acuícola y a unas especies más que a otras según el evento que se generaría a mediano plazo. La dependencia del monocultivo acuícola produciría que un solo evento puede acabar con la producción, si esta depende del cultivo de una sola especie. Por lo tanto, se hace necesario incursionar en policultivos como alternativa, toda vez que ciertas especies se adaptan más a cambios climáticos que otra, y al experimentar la posibilidad de policultivos con especies comerciales que puedan adaptarse a condiciones adversas deben ser
Fig. 2 Interpretación de riesgos según el clima y los procesos socioeconómicos, según la “Introducción al manejo del cambio climático”. John C. Shieder y Jean Hetzel (2021)
investigadas desde varios puntos, partiendo de la factibilidad económica y biológica.
En cuanto a la diversificación del producto, Hoevenaars y van Beijnen (2024)lo traducen como “una estrategia comúnmente utilizada para distribuir los riesgos contra las pérdidas”. Un ejemplo es que dentro de una finca productora de camarón se produzca uno o dos productos adicionales, con el fin de reducir el riesgo por pérdida.
Siempre y cuando se tome en cuenta lo mencionado por Soto (2024), “la diversificación solo funciona si diferentes especies o productos no están sujetos al mismo peligro”. Esto se realizaría con el fin de que la producción continúe en caso de que el cultivo falle.
La diversificación de la producción puede generar beneficios que contribuyan tanto al crecimiento del acuicultor como a la reducción de los eventos relacionados con el cambio climático. A través de este enfoque, se generará un nuevo ingreso y se reducirán los costos de producción al reutilizar los recursos dentro del mismo sistema.
Policultivo-Acuicultura simbiótica: por ejemplo, el camarón y la tilapia. El camarón se alimenta de los excrementos de la tilapia y la tilapia se alimenta de camarones moribundos o muertos. De tal forma que no solo se disminuyen los costos de producción, sino que funciona como un método eficaz para la eliminación de virus que afecten al camarón.
En este sentido la producción de peces, bivalvos o algas de valor comercial y demanda en los mercados internacionales, contribuyen a la no dependencia de una sola especie, en el caso de verse afectada por algún factor del cambio climático, o epidemia generada.
Todo esto hace necesaria la implementación de Centros de Investigación y Desarrollo, para establecer la factibilidad de policultivos o de cultivos alternativos que permitan la sustentabilidad de la producción acuícola, ante los efectos que generaría el cambio climático.
Implementar y familiarizarse con sistemas de alerta temprana
Los sistemas de alerta temprana contribuyen al trabajo del acuicultor, proporcionando información relacionada con el estado de clima, predicciones y previsiones meteorológicas. De tal forma que estará al tanto de fenómenos significativos, parámetros en cuanto al tiempo, intensidad del evento y el espacio sobre el cual va a actuar.
El acceso a estos sistemas es cada vez más fácil y seguro, ya que, en muchas ocasiones, es un servicio proporcionado por los gobiernos a través de tecnología muy sencilla a la cual se puede acceder desde el teléfono celular. Esto permite la difusión de las alertas y el constante monitoreo por parte del acuicultor para preparar sus piscinas y minimizar pérdidas por cambios en parámetros importantes.
Entre los ejemplos proporcionados por Hoevenaars y van Beijnen (2024), en respuesta a una emergencia citan los siguientes: “la cosecha temprana, el fortalecimiento de la infraestructura acuícola, el aumento de la tasa de intercambio de agua y el suministro de aireación”. A lo que se sumaría la constante capacitación del personal con respecto a la interpretación de mapas y condiciones meteorológicas, y el de preparación y respuesta ante emergencias.
Mejoramiento de las infraestructuras
Las medidas a tomar en cuanto a infraestructura deben ejecutarse con relación con el análisis de riesgo. Generalmente, las amenazas a las que se enfrenta el sector camaronero son las inundaciones, la sequía y el deslizamiento de tierra; por ende, existe el riesgo de fuga de la especie cultivada, cambios en la calidad del agua, la destrucción de las piscinas, el rebose del agua de las piscinas, entre otros. Para el caso de las piscinas para el cultivo de camarón, recomiendan que la infraestructura considere lo siguiente:
•Muros más altos y bien compactados, utilizando proporciones correctas de altura, ancho y ángulo correcto de la pendiente.
•Canales de captación, distribución de agua y descarga de efluentes, lo suficientemente anchos, profundos y en un ángulo correcto de la pendiente, para un desfogue rápido y
evitar el desborde de los canales.
•Implementación de un sistema de rejillas y bolsos para evitar el escape del camarón de las piscinas de cría y cultivo.
Uno de los grandes beneficios que tiene la zona costera del Ecuador es la protección adicional que brinda el bosque de manglar. Este funciona como barrera natural ante el oleaje y las lluvias intensas. Una medida estructural natural sería el reforzar la franja de manglar, mediante la forestación de este. De igual forma, las áreas de campamento, almacenamiento de insumos y desechos peligrosos, así como dormitorios y áreas administrativas, también deben considerar medidas estructurales que permitan el funcionamiento de estas bajo cualquier condición. Esto implica: el techado de todas las instalaciones, la construcción sobre una superficie alta para evitar el ingreso del agua, y un sistema para la recolección y drenaje del agua.
Colaboración
Finalmente, Hoevenaars y van Beijnen (2024), hacen énfasis en la necesidad de la articulación de los diferentes actores para llegar a las recomendaciones propuestas, puesto que el productor acuícola va a necesitar de la colaboración de las comunidades, asociaciones de productores e instituciones públicas para implementar las operaciones descritas. El intercambio de información (especialmente en la prevención de enfermedades, pronósticos del tiempo, mareas) debe ser considerado por las asociaciones de productores y la autoridad competente para que opte por las medidas necesarias. De igual forma, con base en las necesidades de adaptación ante el cambio climático, las asociaciones de productores junto con la autoridad competente y la comunidad estarán a cargo de implementar acciones que sean necesarias para contrarrestar sus efectos.
Conclusión:
El cambio climático es una de las amenazas más significativas que enfrenta nuestro planeta en la actualidad. Reconocer su impacto es crucial para entender la magnitud de los desafíos ambientales, económicos y sociales que trae consigo. Este fenómeno global está provocando un aumento de las
temperaturas, cambios en los patrones de precipitación, el derretimiento de los glaciares y el aumento del nivel del mar, lo que a su vez afecta la biodiversidad, la seguridad alimentaria y el bienestar humano. Adoptar medidas urgentes es indispensable para mitigar estos efectos y adaptarnos a las nuevas condiciones.
La reducción de las emisiones de gases de efecto invernadero, la promoción de energías renovables, la implementación de políticas de sostenibilidad y la conservación de los ecosistemas son acciones esenciales que deben ser llevadas a cabo de manera inmediata y coordinada a nivel global•
Bibliografía
"El planeta nos está enviando varias señales del desequilibrio que está experimentando, y alertando sobre la impredecibilidad de los efectos y consecuencias que generan; por eso, está en nuestras manos contribuir de forma proactiva en mitigar las alteraciones que se gestan. Es imprescindible empezar hoy; probablemente, mañana sea demasiado tarde".
Leonardo Maridueña
Director de Ambiente
Cámara Nacional de Acuacultura
Hoevenaars, K., & Van Beijnen, J. (2024, March 18). Six ways to make your aquaculture operations more climateresilient. The Fish Site. https://thefishsite.com/articles/six-ways-to-make-your-aquaculture-operations-moreclimate-resilient
87 91 94
ESTADÍSTICAS
Edición 159 - Junio 2024
Exportaciones de camarón Reporte de mercado de EE. UU. Reporte de mercado de China
COMERCIO EXTERIOR
CAMARÓN:
EVOLUCIÓN DE EXPORTACIONES 2010 -2023
Fuente: Cámara Nacional de Acuacultura
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: COMPARATIVO MENSUAL
(Millones de Libras) 2020- 2024
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
COMERCIO EXTERIOR
CAMARÓN: PARTICIPACIÓN POR DESTINO (Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: PRINCIPALES PAÍSES DESTINOS DE EXPORTACIÓN (Millones de Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
COMERCIO EXTERIOR
CAMARÓN: EXPORTACIONES POR PARTIDA ARANCELARIA
Enero - Marzo 2024
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO ANUAL DE EXPORTACIÓN (Libras)
El volumen de importación de camarón por parte de China en abril disminuyó con respecto al mes anterior en un 10% a 73,750 toneladas. En comparación con abril del año pasado, esto significa un 14% menos. El volumen de importación hasta la fecha alcanzó las 324,780 toneladas, lo que representa una disminución del 6% en comparación con el mismo período en 2023. El precio de importación continuó aumentando en abril, en otros $0.10 a $5.12/ kg. En comparación con el año anterior, esto fue $ 0.82 menos. Esto ha llevado a que el valor total de las importaciones hasta ahora en 2024 alcance poco más de $1.6 billones, una disminución del 17% en comparación con los primeros cuatro meses del año pasado.
Rendimiento de las especies
Las importaciones de Vannamei no disminuyeron tan significativamente como las importaciones de manera general del camarón, con una disminución del 2% desde marzo, alcanzando 69,210 toneladas LSE. En comparación con el año anterior, esto representó un 12% menos. Esto debido a que para el camarón Vannamei hubo algunos cambios en la participación del volumen de presentación, ya que el camarón HOSO vio su participación caer de 59% el mes anterior a 55% en abril, mientras que el camarón pelado creció de 8% a 12%, siendo el resto de la participación HLSO.
Ecuador siguió siendo el principal proveedor de Vannamei a China en abril, pero el volumen de exportación continuó disminuyendo después de los problemas con los envíos descritos en nuestros reportes previos sobre el mercado chino. El volumen de Vannamei que vino de Ecuador disminuyó un 7% con respecto a marzo y un 22% en comparación con el año anterior, a 47,680 toneladas LSE. Se espera otra disminución en las importaciones de Ecuador en mayo en un 10% aproximadamente, según el Modelo de Mercado Kontali.
Sin embargo, los volúmenes de exportación reportados por la CNA para Ecuador durante abril muestran un aumento significativo en las exportaciones a China, lo que genera una expectativa de repunte de las importaciones durante el mes de junio para China. Durante abril, las importaciones de India también mostraron una disminución del 11% con respecto al mes anterior, hasta las 11,100 toneladas LSE, lo que resalta que no solo dificultades logísticas provocaron la disminución de las importaciones de camarón, sino también la lenta demanda en el mercado. Sin embargo, el volumen fue un aumento significativo en comparación con el año anterior (78%), lo que destaca la capacidad de la India para abastecer al mercado chino cuando surgen problemas de las exportaciones ecuatorianas. Los volúmenes de importación de Vannamei de Vietnam se recuperaron en abril, mostrando un aumento del 84% hasta las 5,050 toneladas LSE, continuando la fuerte tendencia de este año para el camarón vietnamita, ya que fue un aumento del 47% en comparación con el mismo mes del año pasado.
Los volúmenes de importación de Monodon experimentaron un aumento del 18% desde marzo hasta las 1,540 toneladas LSE. Sin embargo, esto fue un 47% menos que en abril de 2023. Los volúmenes de importación de Monodon hasta la fecha han alcanzado las 7,090 toneladas LSE, un 5% menos que en el mismo período del año pasado.
Importaciones de China en 2022, 2023 y 2024 (volúmenes y peso promedio/kg)
Perspectivas
El comienzo del año para las importaciones de Vannamei desde China parecía sólido con un aumento interanual del 3% y el 19% durante enero y febrero, respectivamente, pero hubo una disminución interanual durante marzo y abril. Una de las razones de esta caída fue una perturbación en las importaciones provenientes del principal proveedor de China, Ecuador, lo que resultó en una caída del 14% en el volumen proveniente del país. Parte de esta caída fue absorbida por India y Vietnam, aumentando
las importaciones un 30% y un 68% durante marzo, respectivamente.
Si bien una parte de la disminución en el volumen de importación puede explicarse por esta disminución de Ecuador, también muestra una demanda lenta en el mercado chino. Los precios del camarón en el mercado local también resaltan esta demanda relativamente lenta, ya que están en un nivel muy bajo y no parecen repuntar incluso durante los períodos tradicionalmente de alta demanda, como el Festival Qingming y
el feriado del Primero de Mayo. En China, el camarón de mayor tamaño enfrenta caídas de precios más pronunciadas debido a la baja demanda de los consumidores, lo que lleva a los productores, particularmente en el sur de China, a abandonar sus operaciones. Esto se demuestra por una disminución en la compra de semilla, especialmente en Guangdong y Guangxi, lo que podría conducir a una menor oferta del mercado interno en los próximos meses.
El 1 de mayo entró en vigencia el tratado de libre comercio (TLC) entre Ecuador y China. El TLC elimina los aranceles solo para el camarón sin cabeza congelado, mientras que los aranceles para el camarón con cabeza congelado importado, la mayor parte de las importaciones de Ecuador, se eliminarán para 2033. Los aranceles sobre el camarón ecuatoriano sin cabeza, clasificado como HS 03061730, se reducirán al 0% en virtud del tratado de libre comercio (TLC).
Este cambio permite que parte del camarón ecuatoriano importado compita de manera más efectiva con el camarón producido localmente. La depreciación del yuan (RMB) frente al dólar estadounidense ha encarecido el camarón ecuatoriano para los importadores y consumidores chinos. Como resultado, las importaciones chinas de camarón ecuatoriano durante los primeros cuatro meses de 2024 fueron un 6% más bajas que el año pasado. A pesar de las señales de recuperación económica, se espera que la demanda de camarón por parte de los consumidores chinos en el segundo trimestre sea menor que el año pasado debido al gasto cauteloso•
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
Importaciones de Vannamei al mercado chino (2022, 2023 y 2024)
Importación de camarón de Estados Unidos
Autores: Jim Kenny jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com
Urner Barry
Importaciones de todos los tipos, por tipo
El jueves 23 de mayo de 2024, el Departamento de Comercio de los EE.UU. anunció su determinación preliminar en la investigación de derechos antidumping (AD) de camarón congelado de Ecuador e Indonesia. La determinación se deriva de una petición comercial presentada por la Asociación Estadounidense de Procesadores de Camarón (ASPA) en octubre de 2023.
La ASPA dijo que estimaron márgenes de dumping para Ecuador de hasta 111% y para Indonesia de hasta 37%.
Si analizamos primero a Ecuador, Sociedad Nacional de Galápagos
C.A. / Marina del Rey recibió una tasa de dumping del 10.58%. La tasa de depósito (ajustada por la compensación de subsidios) es del 10.58%. La tasa de dumping para Industrial Pesquera Santa Priscila S.A./Tropical Packing Ecuador Tropack S.A. es del 1.54%, que es mínima ya que es inferior al 2%. Todos los demás tienen una tasa de dumping del 10.58%, mientras que la tasa de depósito es del 10.18%. En cuanto a Indonesia, el exportador PT Bahari Makmur Sejati tiene una tasa de dumping del 0.00%, pero PT First Marine Seafoods/PT Khom Foods se vio afectado por una tasa de dumping del 6.30%. Todos los demás exportadores/productores indonesios tienen una tasa de dumping del 6.30%.
La próxima fecha de interés en la investigación antidumping del camarón congelado de Ecuador e Indonesia es el 4 de octubre de 2024. En ese momento, el Departamento de Comercio publicará su determinación final. Si el Departamento de Comercio toma una decisión afirmativa final, la Comisión de Comercio Internacional de los Estados Unidos (ITC) tomará una decisión final el 18 de noviembre de 2024. Y si tanto el Departamento de Comercio como la ITC toman decisiones afirmativas finales, la Orden de Emisión se emitirá el 25 de noviembre de 2024.
Previo a este anuncio, las importaciones de camarón de los Estados Unidos seguían aumentando año tras año gracias a un aumento significativo de los dos principales socios comerciales. Las importaciones totales de marzo fueron de 139.489 millones de libras, en comparación con los 129.469 millones de libras de 2023. India (+24.6%), Ecuador (+24.6%) y Vietnam (+39.0%) aumentaron. Estos números positivos también hicieron que las cifras del año hasta la fecha fueran al alza para esos tres países. Indonesia (-30.0%) y Tailandia (-25.7%) enviaron menos, completando los cinco primeros puestos. México (-31.8%) y Argentina (-28.8%), aunque enviaron menos, ocuparon un cercano sexto y séptimo lugar.
En términos de forma del producto, fue mixto en las categorías, donde las categorías de mayor cantidad, como camarón sin cabeza y con cáscara, que incluye el de pelado fácil (+12.8%) y pelado (+12.5%), superaron al cocido (-6.7%) y empanizado (-18.7%).
Ciclos de importación mensual por país (todos los tipos)
India: Las importaciones de camarón de marzo de la India (+24.6%) aumentaron fuertemente por segundo mes consecutivo después de las caídas de enero. Las importaciones totalizaron 51.395 millones
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
de libras en comparación con 41.233 millones de libras el año pasado. La cifra del año hasta la fecha es ahora un siete por ciento más alto en el primer trimestre. Hubo más camarón con cáscara (+1.6%), pelado (+30.1%) y cocido (+10.5%).
Ecuador: La brecha con el número uno, India, se redujo considerablemente después de que Ecuador (+24.6%) registrara otra gran ganancia en el mes de marzo. Con 47.959 millones de libras, en comparación con 38.490 millones de libras, esto fue menos de cuatro millones de libras de la India. Al igual que su competidor, las importaciones del primer trimestre son un 9.2 por ciento más altas. Hubo más camarón con cáscara (+37.0%) y pelado (+8.7%), pero menos empanizado (-15.1%).
Indonesia: Con el telón de fondo del AD, Indonesia (-30.0%) siguió rezagada con respecto a las importaciones del año pasado. Las importaciones de marzo cayeron a 20.978 millones de libras. Las pérdidas se debieron principalmente a camarón con cáscara (-31.7%), pelado (-35.5%), cocido (-20.1%) y empanizado (-25.7%).
Vietnam y Tailandia: Vietnam (+39.0%) siguió mostrando un crecimiento interanual. Y se sitúa un 22.8 por ciento por encima del año pasado durante los tres primeros meses. Aunque Tailandia (-25.7%) envió menos, recuperó el quinto lugar en la lista.
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones del camarón con cáscara, que incluye el pelado fácil, aumentaron un 12.8 por ciento en marzo.
Durante el mes, la ganancia fue impulsada principalmente por Ecuador (+37.0%) que envió casi 5 veces más que el número dos, Indonesia (-31.7%). El resto de los socios comerciales fueron poco variados.
Las pérdidas se dieron solo en un tamaño, el de 21-25, también la categoría de mayor volumen. El resto fueron ganancias positivas en marzo.
Los valores de reemplazo del camarón con cáscara (importación $/lb.) continuaron cayendo, calando los mínimos históricos nuevamente. Los precios fueron de $3.33 por libra, $0.03 por libra más bajos mes a mes.
de importaciones
Valor agregado, importación de camarón pelado
Las importaciones de camarón pelado, la categoría más grande por volumen, aumentaron por segundo mes consecutivo. Las importaciones de marzo totalizaron 66.340 millones de libras, o 12.5 por ciento más que el año pasado. El socio comercial más grande por volumen, India (+30.1%), ganó nuevamente, al igual que el número dos, Ecuador (+8.7%). Esto superó las pérdidas de la mayoría de los demás países.
Los valores de reemplazo (importación $/ lb.) para el camarón pelado perdieron los mismos $0.03 por libra al mismo precio de $3.33 por libra que el camarón con cáscara. Esto también se mantuvo en la tendencia de continuar con mínimos cíclicos.
Las importaciones de camarón cocido (de agua tibia) perdieron un 6.7 por ciento en
Fuente: USDOC. Urner Barry
Detalle
de camarón YTD por tipo
Fuente: USDOC. Urner Barry
REPORTE DE MERCADO
marzo, mientras que el camarón empanizado perdió un 18.7 por ciento con respecto al año pasado.
Importaciones de camarón
cocido, empanizado y otros
Los precios de reemplazo del camarón cocido volvieron a bajar, de $4.11 por libra a $4.07. El entorno de demanda en EE.UU. es más débil de lo previsto y la venta al por menor ha sido un desafío y no ha respaldado los precios.
Línea de tiempo del precio del camarón; anuncios minoristas
Comercio minorista: las oportunidades de compra minorista aumentaron por primera vez mes a mes en 2024. En términos relativos y comparativos, el camarón tiene un buen valor. Los minoristas intentaron transmitir ese mensaje a los consumidores, pero pareció no tener demasiado impacto. Hubo un 33.7 % más anuncios que en marzo y un 17% más que el promedio de tres años. El precio promedio del camarón cayó $0.39 por libra a $7.67 por libra.
Suministro de camarón a EE.UU. y situación del Golfo
***La data de desembarques proporcionada por la NMFS ha sido inconsistente y existen lagunas en la data que hace que las comparaciones no sean confiables.
La data de desembarques de febrero de 2024 publicada por el NMFS muestra que se desembarcaron 2.226 millones de libras (total, todas las especies, sin cabeza); por debajo de los 3.191 millones de libras de febrero de 2023.
Durante los primeros dos meses del año, se reportaron desembarques de 5.805 millones de libras, un 20.4 por ciento por debajo de los 7.289 millones de libras de 2023. Los desembarques han estado disminuyendo constantemente año tras año. El mercado sigue siendo muy inestable.
La temporada comercial de camarón del Golfo de México, tanto para las aguas de Texas como para las federales, cerró 30 minutos después del atardecer del miércoles 15 de mayo, hasta una fecha aún por
determinar en julio. La Comisión de Pesca y Vida Silvestre de Luisiana fijó la fecha de apertura de la temporada de camarón costero de primavera; anunciaron que todas las aguas costeras desde la frontera estatal de Mississippi/Luisiana hacia el oeste hasta la frontera estatal de Luisiana/Texas se abrirán a las 6 a.m. del jueves 16 de mayo de 2024.
Exportación del camarón ecuatoriano
El mercado de camarón tigre blanco y negro importado y cultivado en granjas se ha estabilizado. La industria ahora tiene cifras preliminares de CVD y AD, que probablemente afecten el comercio. Además, los costos más altos en toda la cadena de valor y la escasez de contenedores, especialmente en Asia, han alterado la acción de los precios. Pero la demanda sigue siendo más débil de lo esperado y es un factor limitado•
CNA impulsa la agenda diplomática con China
Con el propósito de dar seguimiento a la implementación del acuerdo comercial entre Ecuador y China, que entró en vigencia en mayo de 2024, José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura (CNA) y titular del Directorio de la Corporación de Gremios Exportadores (CORDEX), se reunió con el embajador de la República Popular de China en Ecuador, Chen Guoyou, junto con otros representantes del sector productivo exportador.
Durante la reunión, se abordaron temas importantes para las cadenas de valor como: resolución de restricciones en las exportaciones, reglas de origen, medidas sanitarias y fitosanitarias, cooperación en inversiones, cooperación aduanera, transferencia tecnológica, comercio electrónico, entre otros aspectos.
ASC conoció las observaciones del sector camaronero sobre el nuevo borrador de estándar para fincas
Mediante oficio dirigido a María Filipa Castanheira, gerente de Estándares de Bienestar y Salud Acuícola del Aquaculture Stewardship Council (ASC), y Javier Unibazo, jefe de Estándares de esta institución, la Cámara Nacional de Acuacultura planteó las observaciones del sector camaronero ecuatoriano sobre el borrador del nuevo estándar ASC para la certificación de granjas, que estuvo en consulta pública hasta el 22 de mayo pasado.
El documento expresó preocupación, ya que el borrador para el nuevo estándar supone un cambio significativo que obligaría a las instalaciones certificadas a replantear varios de sus procesos.
Entre los puntos destacados se incluye la falta de reconocimiento de estudios de biodiversidad e impacto social, la falta de claridad en la aplicación del estándar a diversas especies, inconsistencias en principios cruciales como la prohibición de antibióticos, la viabilidad del método de sacrificio por shock eléctrico, y el uso de metabisulfito de sodio. Además, se destaca la necesidad de adaptar los horarios laborales a las realidades de la producción acuícola. Se solicitó una revisión exhaustiva para asegurar la sostenibilidad y el desarrollo de la industria.
Por su parte, el embajador chino se comprometió a actuar como un puente entre ambos países, facilitando la consecución de estos objetivos y promoviendo el comercio bilateral. Esta colaboración promete fortalecer las relaciones comerciales y abrir nuevas oportunidades para los productos ecuatorianos en el gigante asiático.
Estas observaciones fueron recabadas tras una sesión de trabajo con representantes del sector camaronero, y en la que Leonardo Maridueña, director de Ambiente de la CNA, presentó el borrador de la propuesta ASC y se analizaron aspectos relevantes de la nueva versión sugerida.
Comparecencia en Comisión Legislativa sobre proyecto de Ley de Bienestar Animal
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, participó en la sesión No. 176 de la Comisión de la Biodiversidad y Recursos Naturales de la Asamblea Nacional del Ecuador, para dar a conocer a los legisladores las observaciones que tiene el sector camaronero respecto al borrador del Proyecto de Ley para el Bienestar Animal. Subrayó que el sector camaronero ya cumple con las prácticas de protección animal y manejo adecuado de los ecosistemas, cumpliendo con los estándares internacionales exigidos por los mercados. Por otra parte, mencionó que existen autoridades de control que están encargadas de normar y regular al sector a través de parámetros técnicos específicos, y señaló la importancia de no interferir con las facultades de estas autoridades, sino más bien complementarlas.
CNA observó el borrador de Proyecto de Ley de Discapacidades ante Comisión de la Asamblea
Daniel Pesantes, Director Adjunto de la Cámara Nacional de Acuacultura, participó en la mesa territorial de la Comisión Especializada Permanente de Garantías Constitucionales, Derechos Humanos, Derechos Colectivos y la Interculturalidad, realizada en el municipio del cantón Piñas, en El Oro. La reunión se centró en el tratamiento del informe para el segundo debate del Proyecto de Ley Orgánica Reformatoria a la Ley Orgánica de Discapacidades.
El proyecto de ley incluye una propuesta para reformar el Artículo 47 de la ley vigente, aumentando el porcentaje mínimo de contratación de personas con discapacidad del 4% al 8% de la nómina de trabajadores de los empleadores. La Cámara Nacional de Acuacultura ha señalado que no se ha presentado un análisis técnico que justifique este incremento, por lo que se presentó una oposición a este cambio sugerido. Además, se mencionó que las condiciones geográficas en las que se desarrolla la actividad camaronera requieren trabajos al aire libre en áreas remotas y traslados por medios acuáticos que dificultan la integración de personas con discapacidad física.
Además, Camposano sugirió que la ley establezca principios generales en lugar de descripciones técnicas específicas sobre el contenido nutricional del alimento balanceado para camarón, pues se podría dificultar su implementación. Al concluir su intervención, el titular de la CNA felicitó a la comisión y destacó la apertura para recibir aportes desde todos los sectores para la construcción de un proyecto de Ley de Bienestar Animal bien elaborado.
Cámaras de videovigilancia de la Prefectura del Guayas apoyan el control de seguridad del sector camaronero
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, se reunió con la Prefecta del Guayas, Marcela Aguiñaga, para revisar los avances en materia de seguridad gracias a la intervención de la entidad en ejes viales de la provincia. Se intercambió información técnica entre la Dirección de Seguridad de la CNA y la Coordinador de Gestión de Riesgos y Seguridad de la Prefectura.
Durante la mesa de trabajo, también se analizó la contribución del sector privado en términos de información e inteligencia para hacerle frente a la inseguridad que afecta a las actividades productivas, entre ellas la del sector camaronero. Se acordó que se continuará con una estrecha colaboración con la autoridad, a fin de seguir haciendo frente a la delincuencia que opera en zonas por las que se movilizan miembros de nuestro sector.
España actualizó oficialmente sus criterios para la detección de Vibrios en el camarón
Sanidad Exterior de España notificó a Ecuador la actualización del procedimiento para la detección de Vibrio Cholerae, alineado con los criterios del Comité Científico de la Agencia Española de Seguridad Alimentaria y Nutrición (AESAN) y en el que se incorporaron argumentos técnicos presentados por nuestro país meses atrás.
“En 2024, dicho comité, a la luz de la reciente información científica publicada sobre este género, ha emitido un nuevo informe que recoge criterios microbiológicos para V. cholerae, aplicables a langostinos y otros productos de la pesca congelados importados, como medidas adicionales de control en los puestos fronterizos”.
Tras esta disposición, las exportaciones de camarón ecuatoriano a España podrían retornar a la normalidad, siempre que se cumpla con lo establecido en el documento. Es importante recalcar que los controles se mantendrán, y si se encuentran nuevos hallazgos en lotes ecuatorianos, los laboratorios acreditados por la autoridad española serán los encargados de detectar serotipos y el gen que produce la toxina.
“La detección en productos de la pesca de V. cholerae no perteneciente a los serogrupos O:1 y O:139 y que no porten el gen CTX, presenta un bajo nivel de riesgo en la población en general, por lo que no será necesario tomar ninguna medida. Por consiguiente, si se detectan productos de la pesca con la presencia de V. cholerae de los serogrupos O:1 y O:139 o que porten el gen CTX, se considerarán como alimentos no seguros” , y por ende, se destruirá toda la carga correspondiente.
Coordinación para apoyar la desarticulación de bandas delictivas
Con el propósito de combatir la inseguridad en corredores viales por donde se transporta camarón y diversos productos acuícolas, la Dirección de Seguridad de la Cámara Nacional de Acuacultura, junto con equipos de Inteligencia Militar y la Unidad Nacional de Investigación y Apoyo en Seguridad (UNASE) de la Zona 5, intercambiaron información que sirvió para desarticular varias estructuras delictivas en Durán, Km. 26, Yaguachi y Taura en este mes.
En complemento a dichas acciones, mediante oficio de la CNA dirigido a la ministra del Interior, Mónica Palencia Núñez, se señaló que, durante el mes de mayo de 2024, tanto productores de camarón como empresas procesadoras de alimento balanceado han sido objeto de asaltos en la vía Durán - Boliche. Estos incidentes han puesto en riesgo la integridad física tanto de conductores como de guardias de seguridad, generando una creciente preocupación en el sector acuícola.
Insumos fabricados 100% en Ecuador llegan al mercado mexicano
La primera exportación de insumos acuícolas ecuatorianos a México, fabricados con la última tecnología y calidad, se realizó en mayo del 2024 por parte de la empresa Nepropac S.A. Esta expansión empresarial permitirá mejorar la nutrición y salud de los camarones, previniendo enfermedades y promoviendo un crecimiento sostenible, prácticas que mejoran la eficiencia y la productividad.
La empresa ecuatoriana, que cuenta con más de 24 años de experiencia en esta rama, cumple con los protocolos de bioseguridad implementados por las autoridades ecuatorianas, tales como las certificaciones BPM Y HACCP, lo cual es vital para mantener el acceso a mercados internacionales. Además, cuenta con una fábrica de primer nivel con
Nuevo titular de multinacional Danesa
Andrés Rivadulla asumió su nuevo cargo como gerente general de BioMar Ecuador desde el 1 de junio, tras la dimisión de Danny Vélez, quien decidió dar un nuevo paso en su carrera profesional para asumir funciones de asesoría y apoyo al directorio de BioMar Ecuador.
Rivadulla es licenciado en Ciencias Económicas por la Universidad Pompeu Fabra en Barcelona y es magíster en Dirección Financiera en la Universidad Adolfo Ibáñez en Santiago de Chile. Tiene más de 18 años de carrera profesional en áreas de Finanzas Corporativas en España, Chile, Argentina y Ecuador, y es parte del equipo directivo de BioMar Ecuador desde el 2017.
Nuevo Director Comercial y Marketing de empresa de alimentos balanceados
Oscar Segura Modesto, Médico Veterinario Zootecnista por la Universidad del Tolima en Colombia, fue nombrado Director Comercial y Marketing de la empresa Molinos Champion, empresa ecuatoriana de más de 60 años con estándares internacionales de su casa matriz la multinacional Seaboard Corporation de los Estados Unidos.
Segura cuenta con una destacada trayectoria habiendo colaborado con empresas como Avícola Casablanca - Proceal. Posteriormente, desempeñando el cargo de Gerente Regional de Ventas y Operaciones en Contegral - Grupo Bios durante más de 14 años.
capacidad instalada para abastecer el mercado tanto nacional como internacional, cuidando todos los estándares de calidad requeridos por los organismos de control.
Desde 2012, ha estado involucrado en la acuicultura de producción de tilapia en el departamento del Huila, expandiendo su experiencia a los departamentos del Meta y Casanare en Colombia a partir de 2018.