AQUAEXPO El Oro 2024 marcó el camino hacia la acuacultura del futuro
La sostenibilidad empieza en el primer eslabón de la cadena
El secreto del camarón ecuatoriano: innovación + compromiso
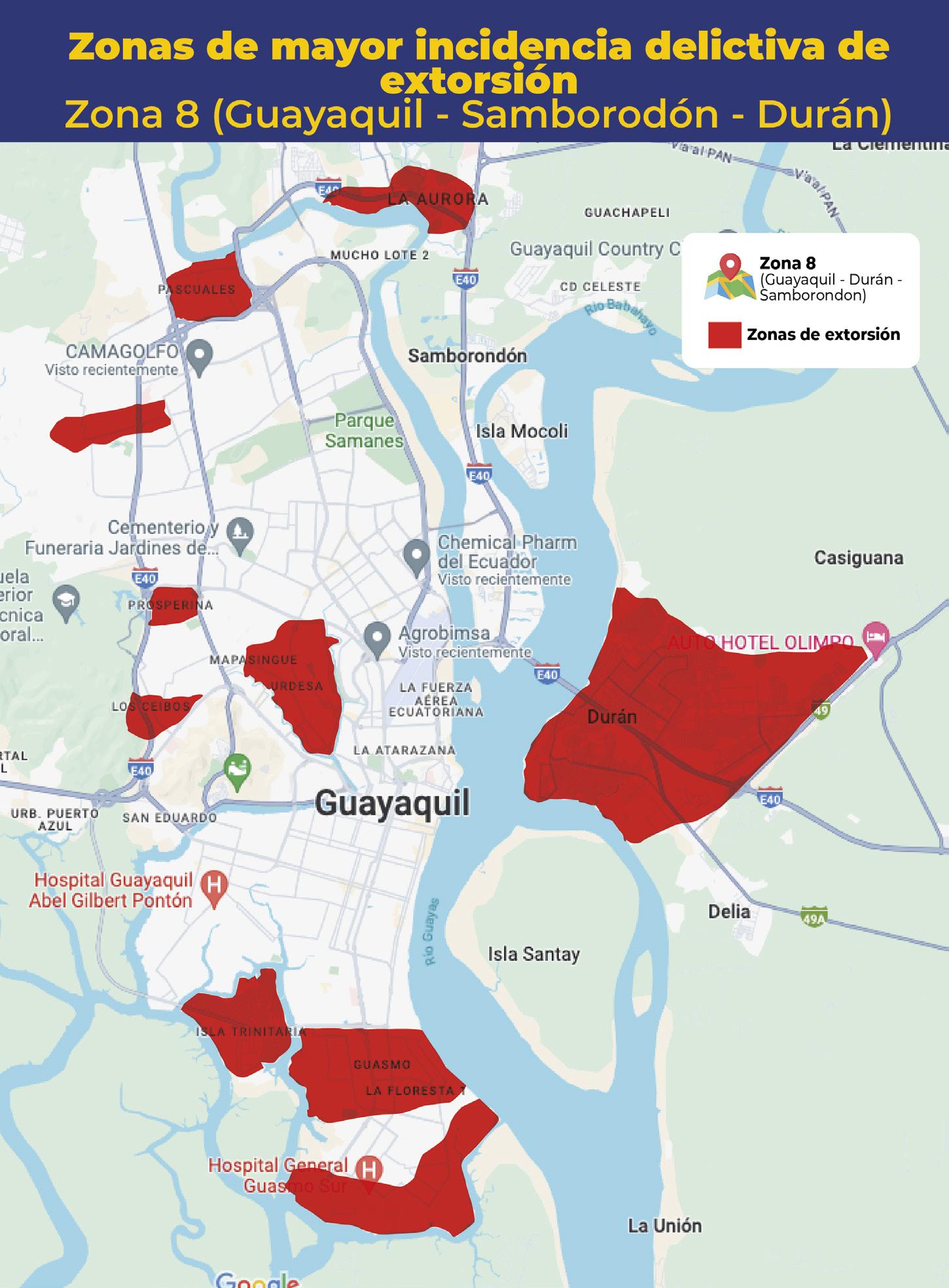
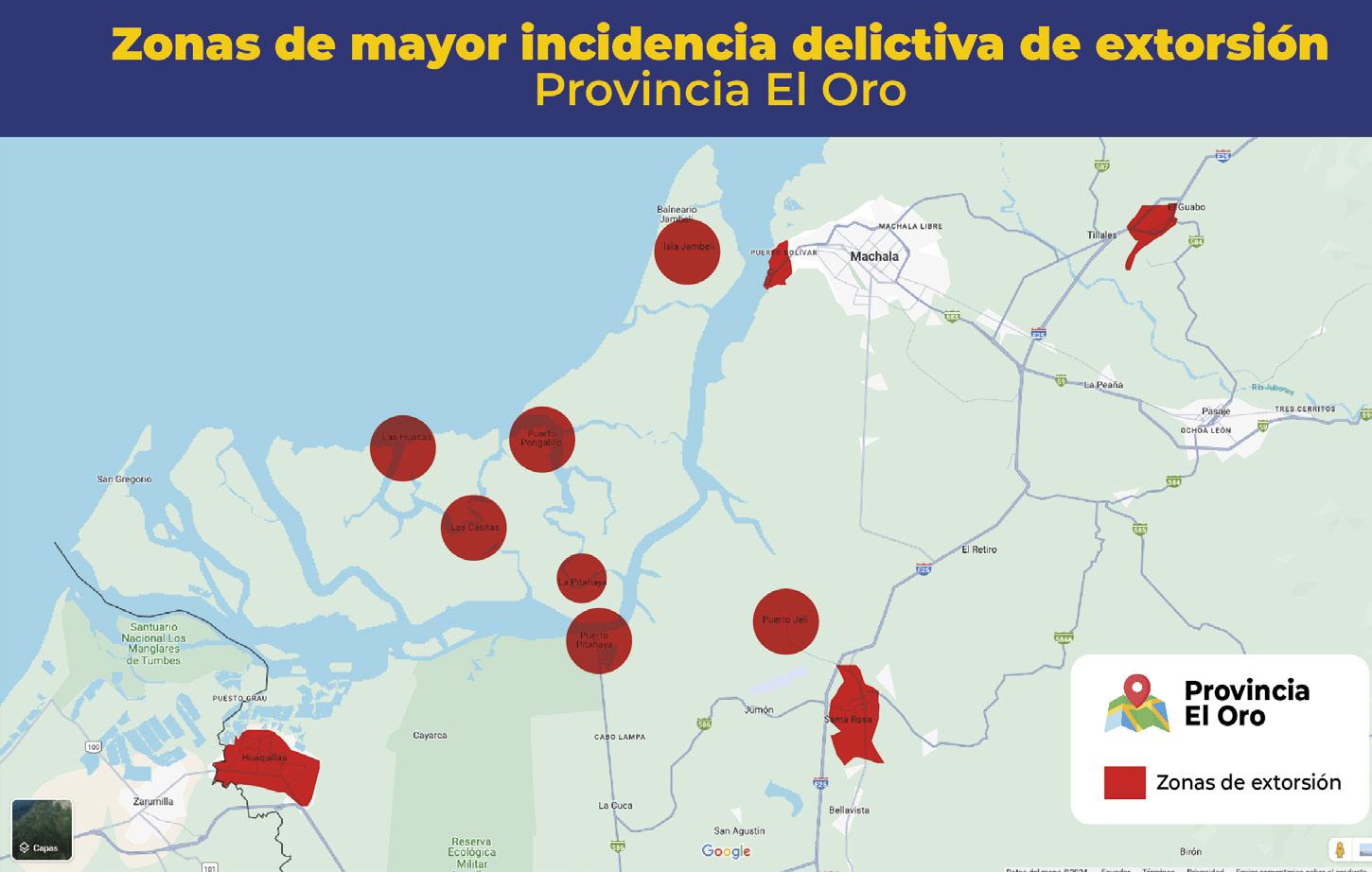
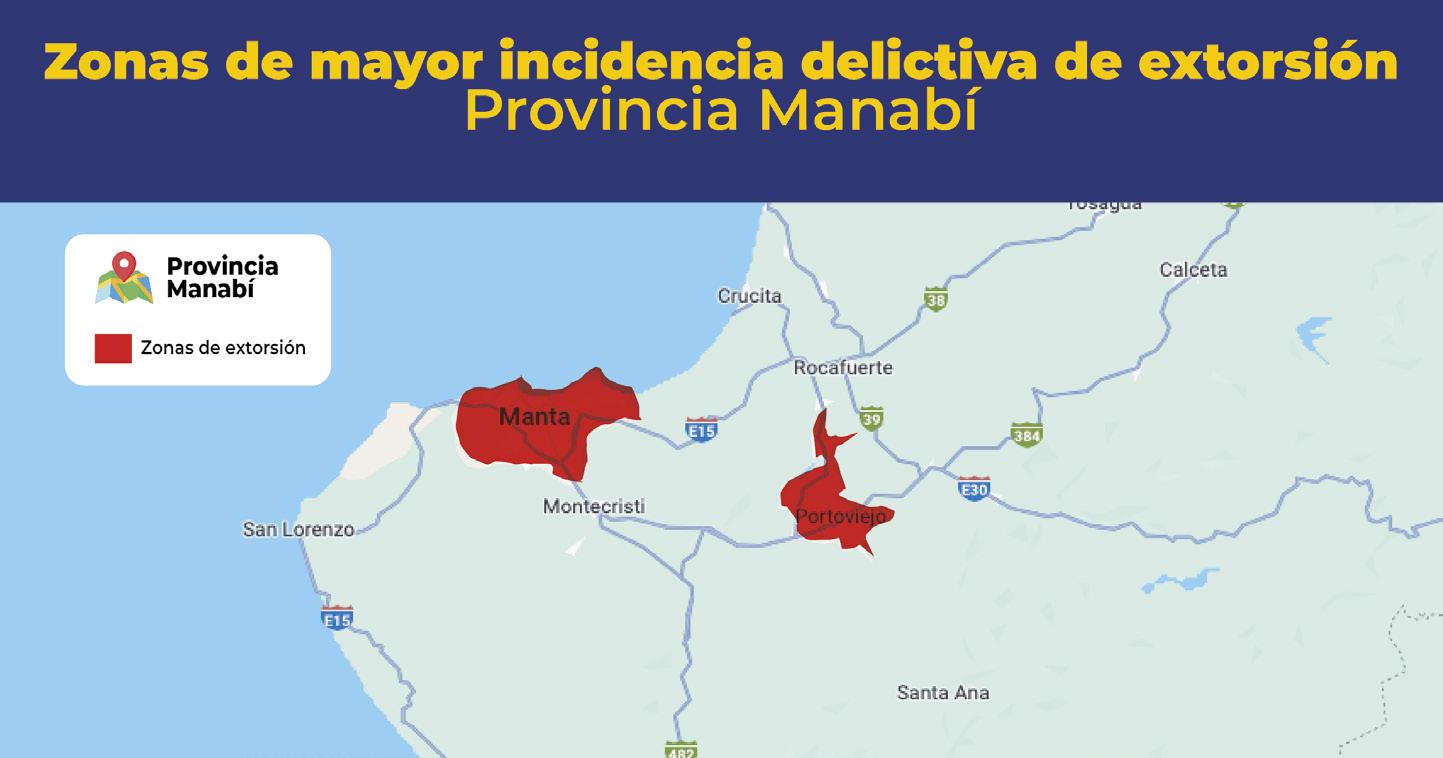
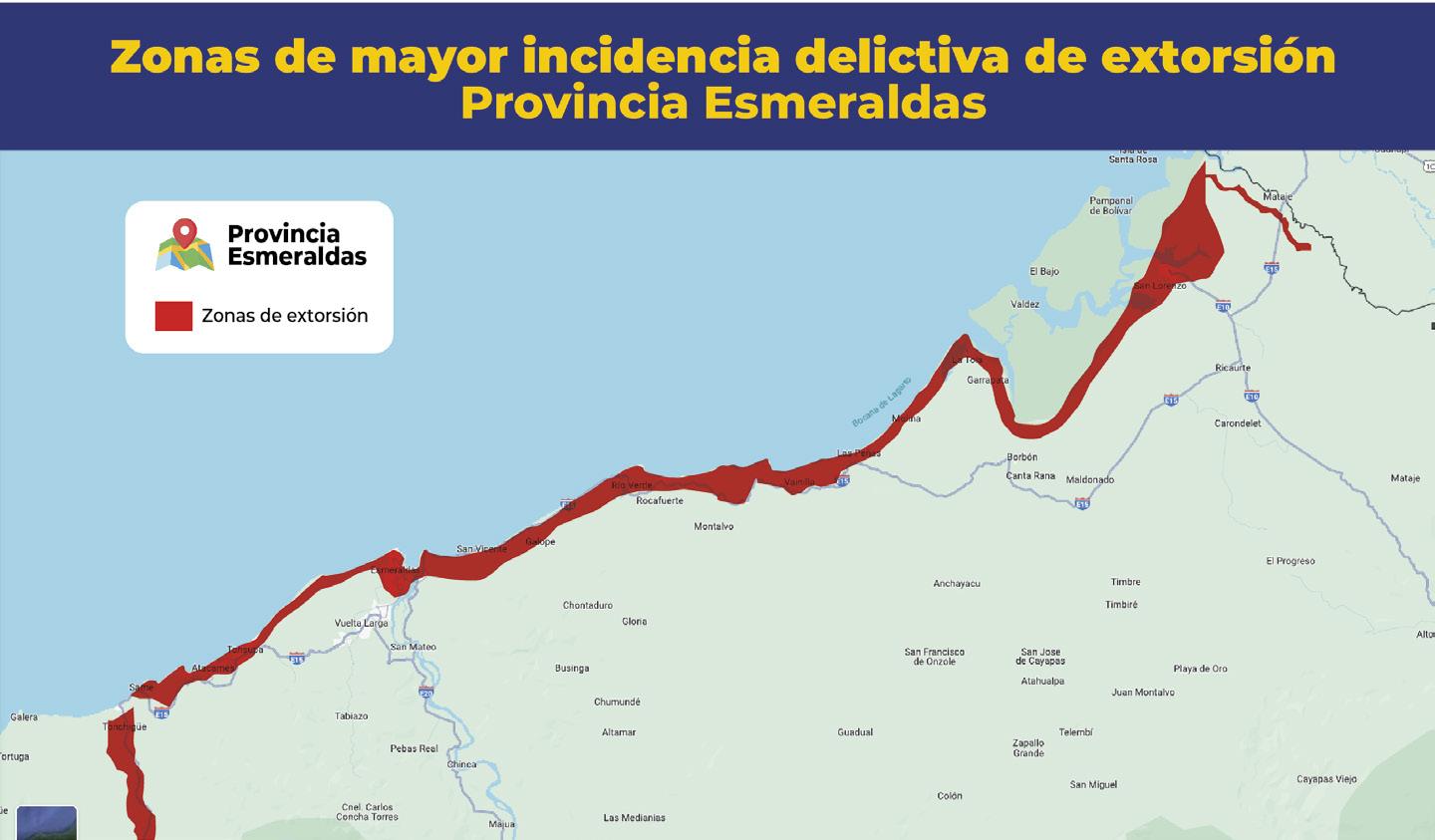
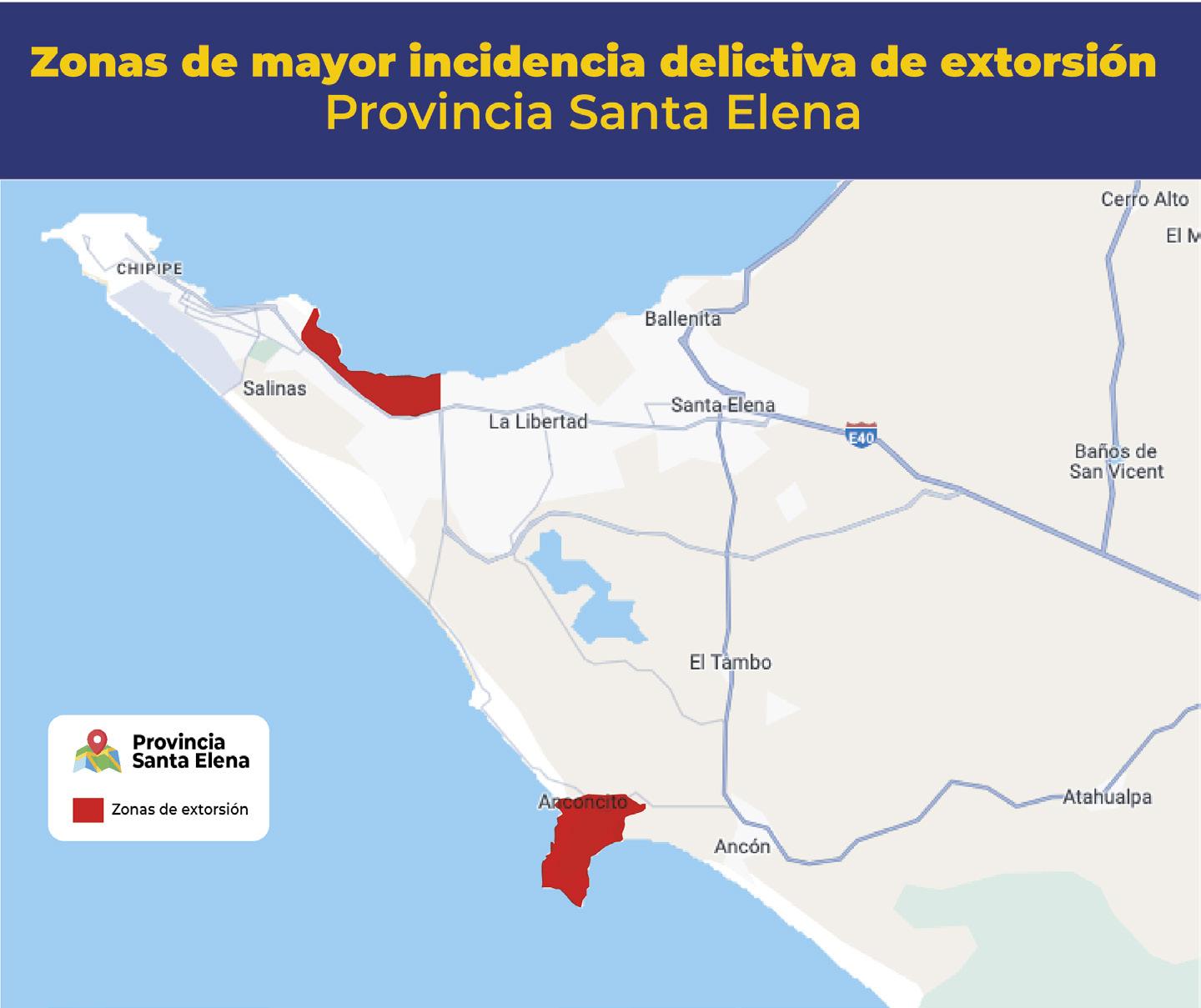
Al menos 20 alertas diarias por extorsión reporta el sector acuícola y agropecuario en zona costera ecuatoriana
Síndrome de las Heces Blancas, un trastorno multifactorial del camarón cultivado: una breve revisión
La diversificación ecológica revela rutas de aparición de patógenos en poblaciones endémicas de Vibrio vulnificus
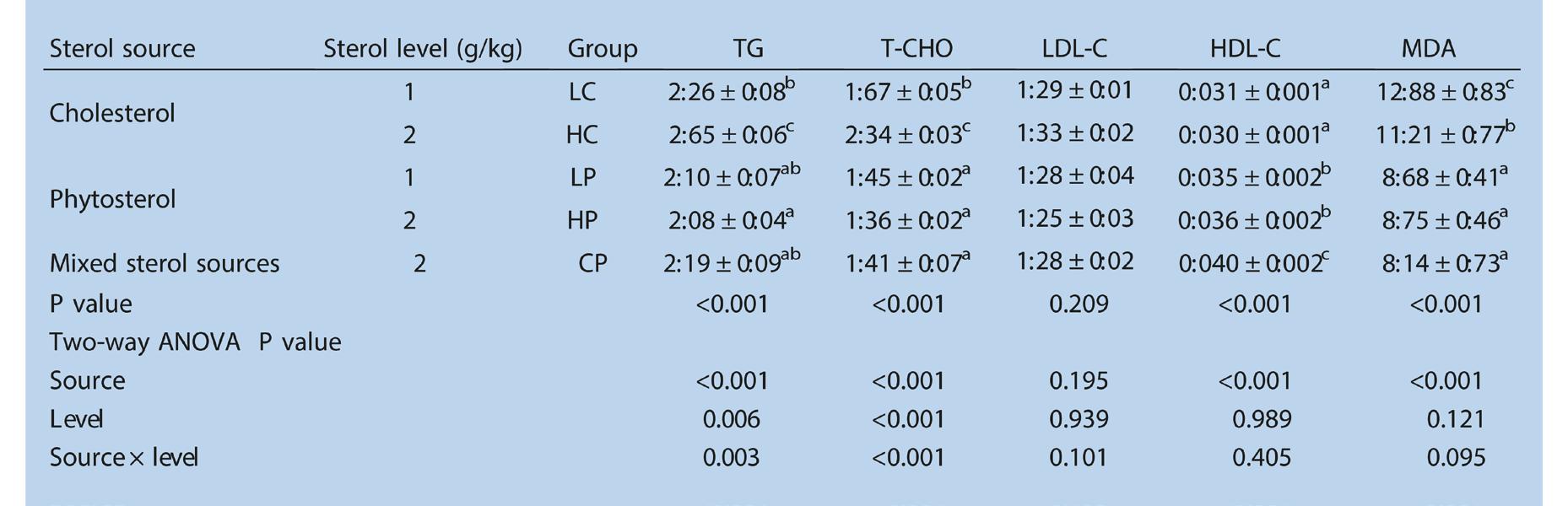
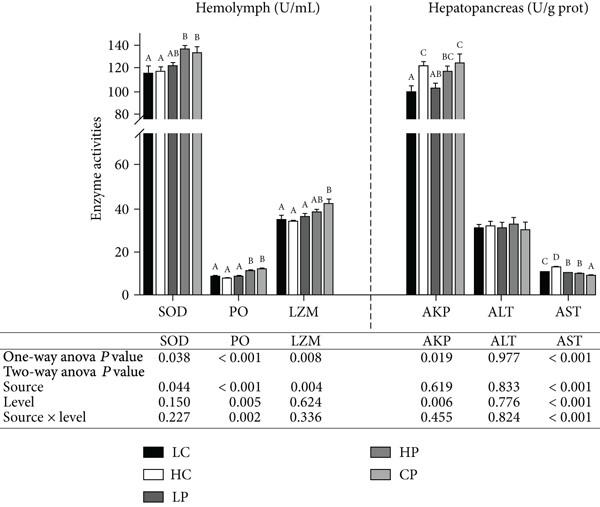
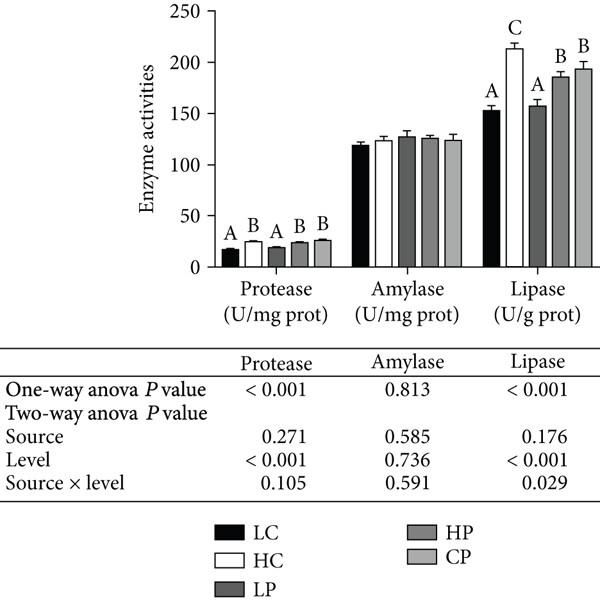
Evaluación de fitoesteroles como alternativa al colesterol en dietas prácticas sobre el crecimiento y la inmunidad no específica de Penaeus vannamei
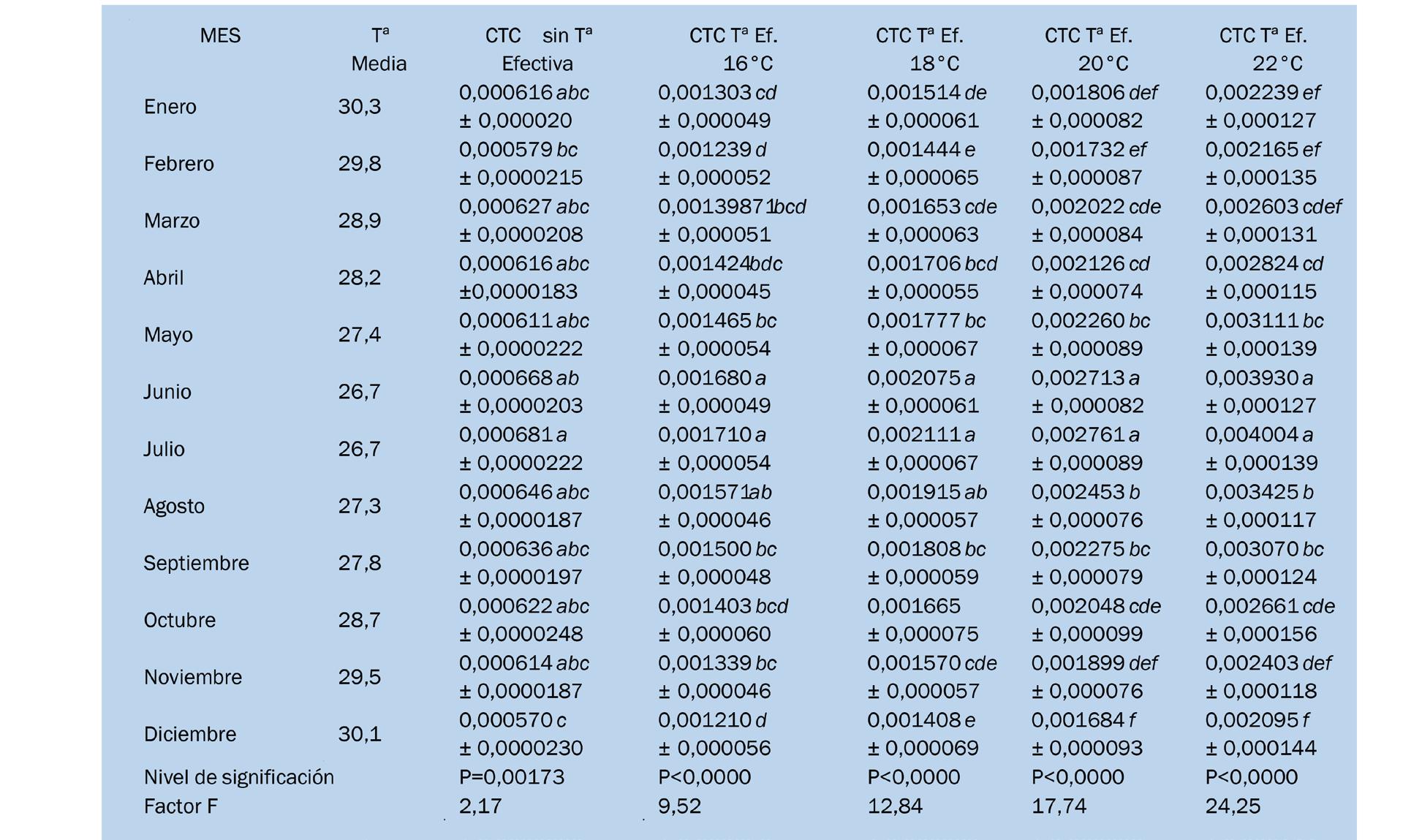
Comparación del coeficiente térmico de crecimiento en el camarón Penaeus vannamei en Ecuador con diferentes sistemas de alimentación
Estandarización de concentraciones de solución de metabisulfito de sodio y tiempo de inmersión para el camarón de cultivo Litopenaeus vannamei
Presidente Ejecutivo
Ing. José Antonio Camposano
Editora “AquaCultura”
MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita PhD. Leonardo Maridueña Ing. José Antonio Lince Ing. Alex de Wind
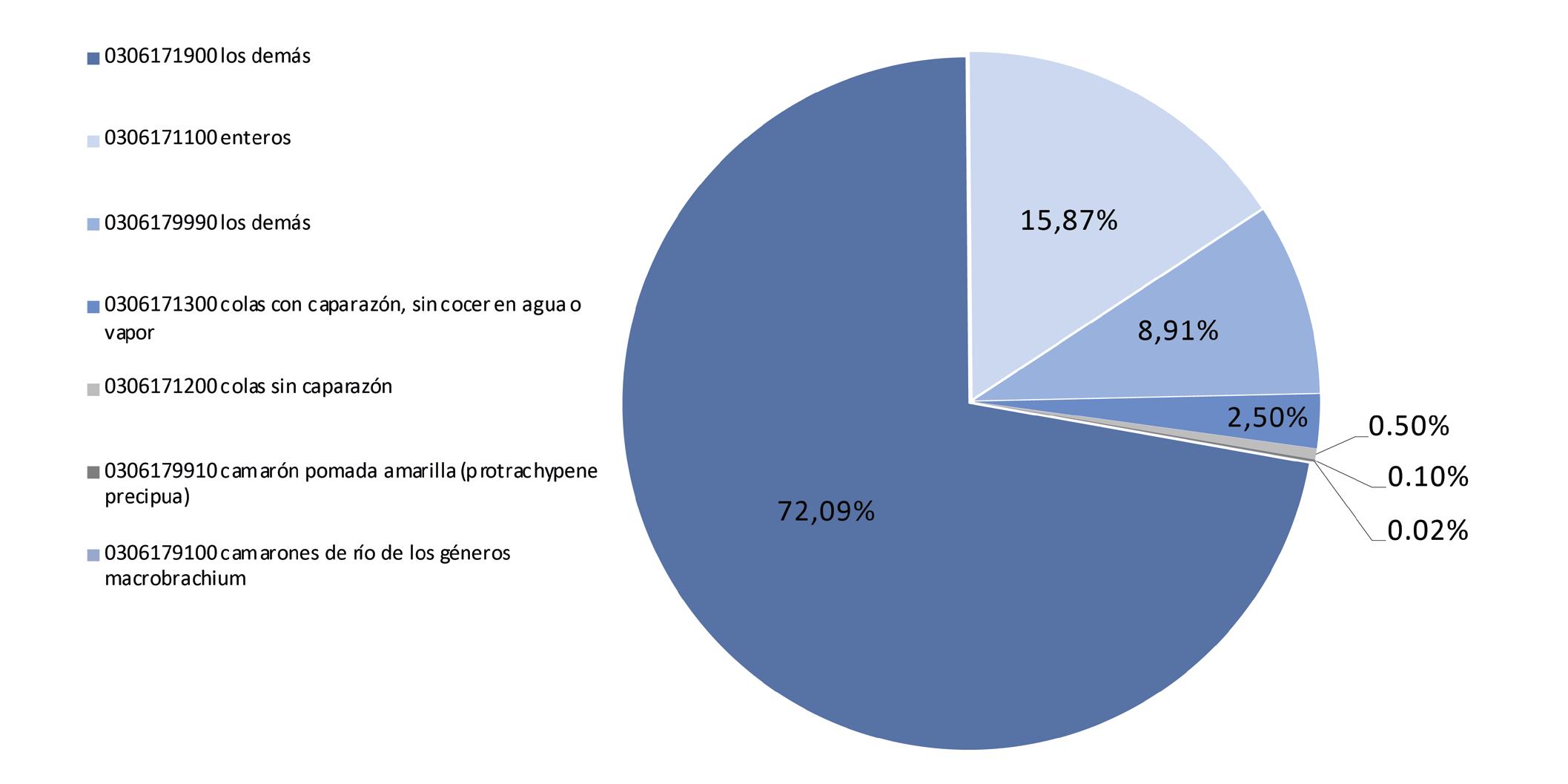
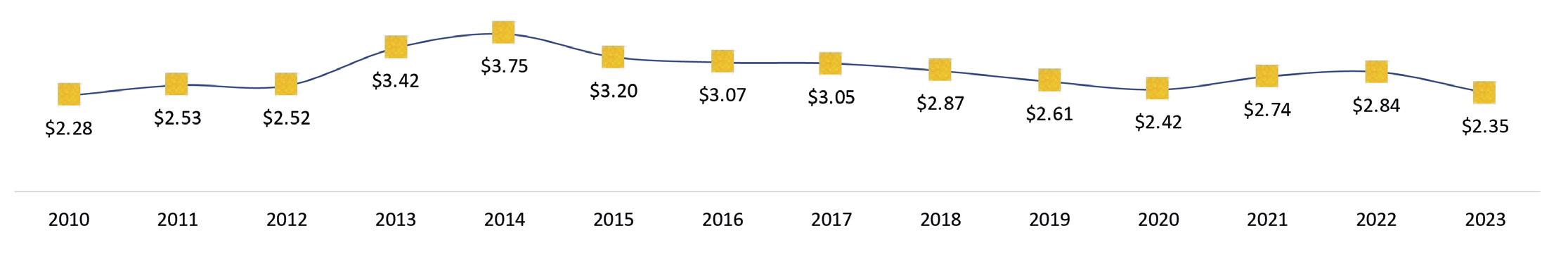
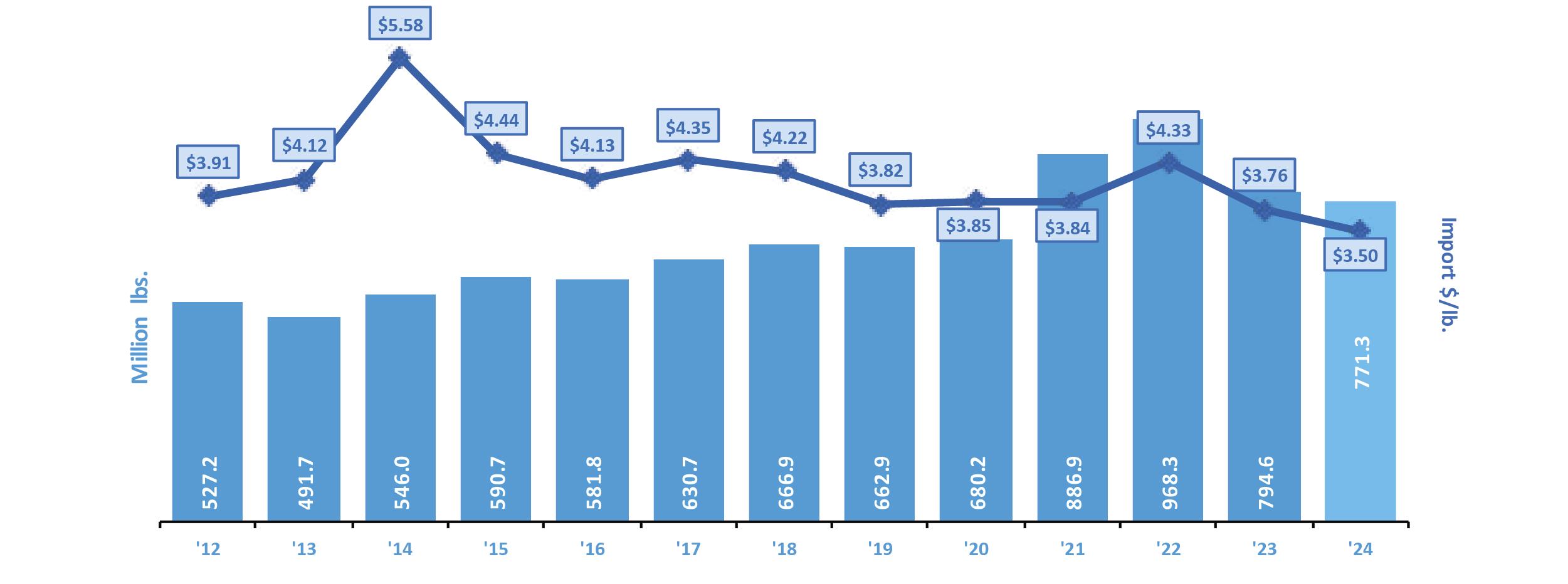
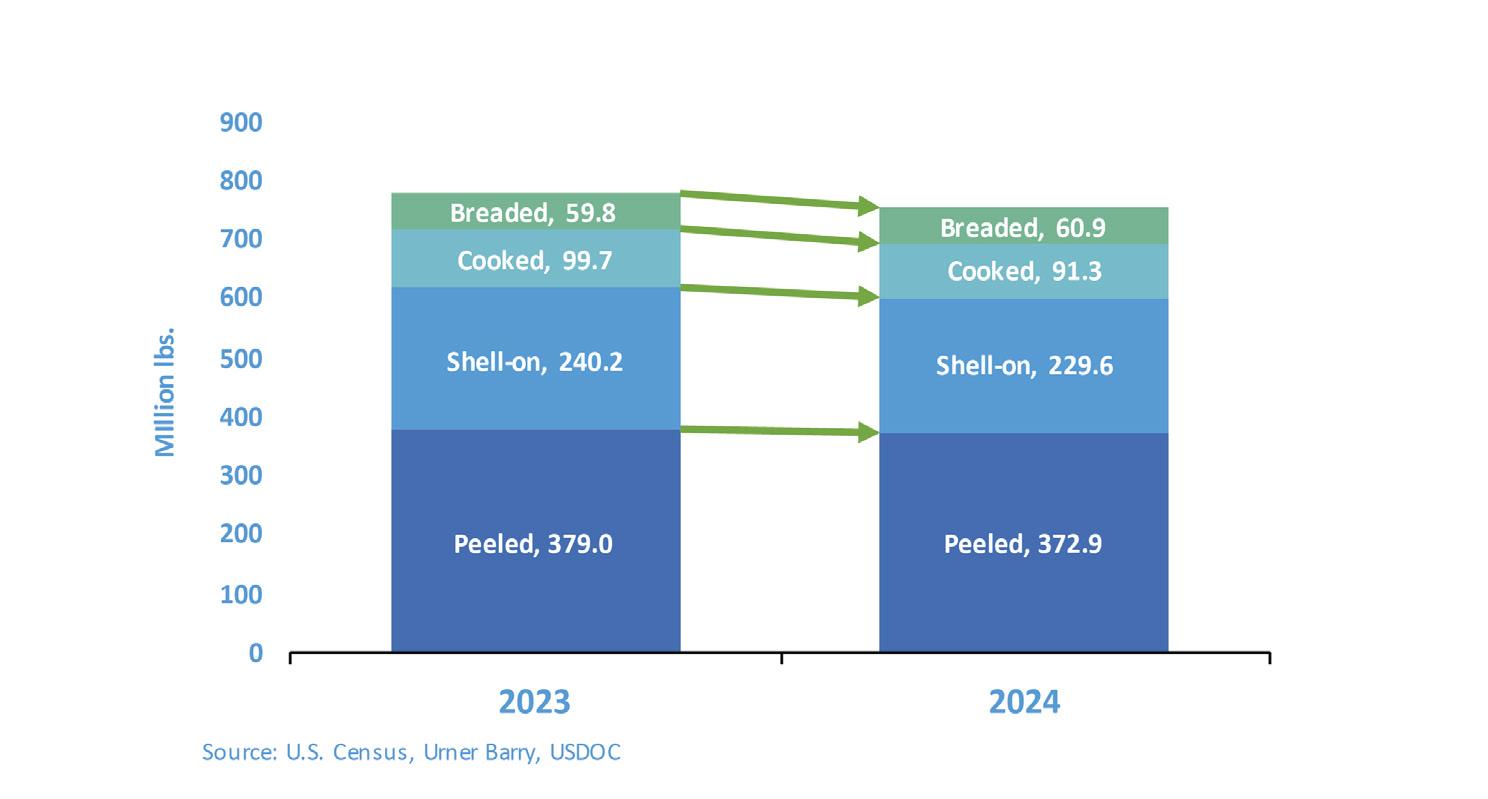
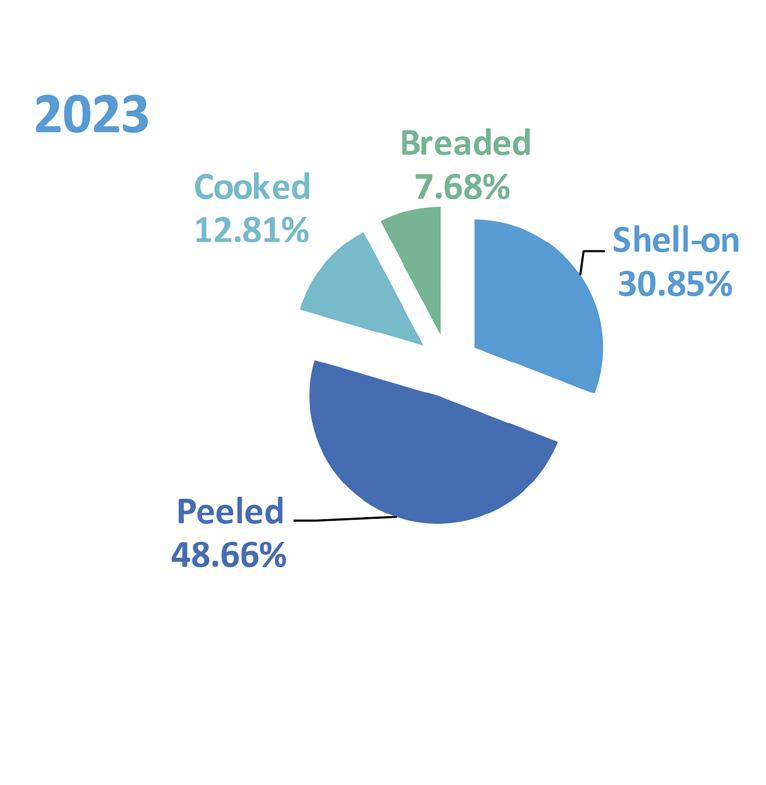
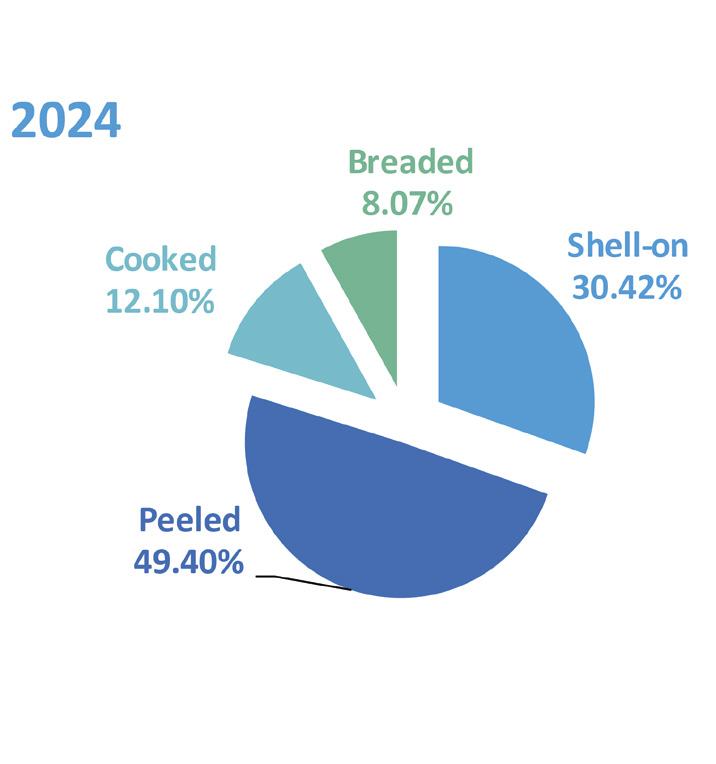
Exportaciones de camarón
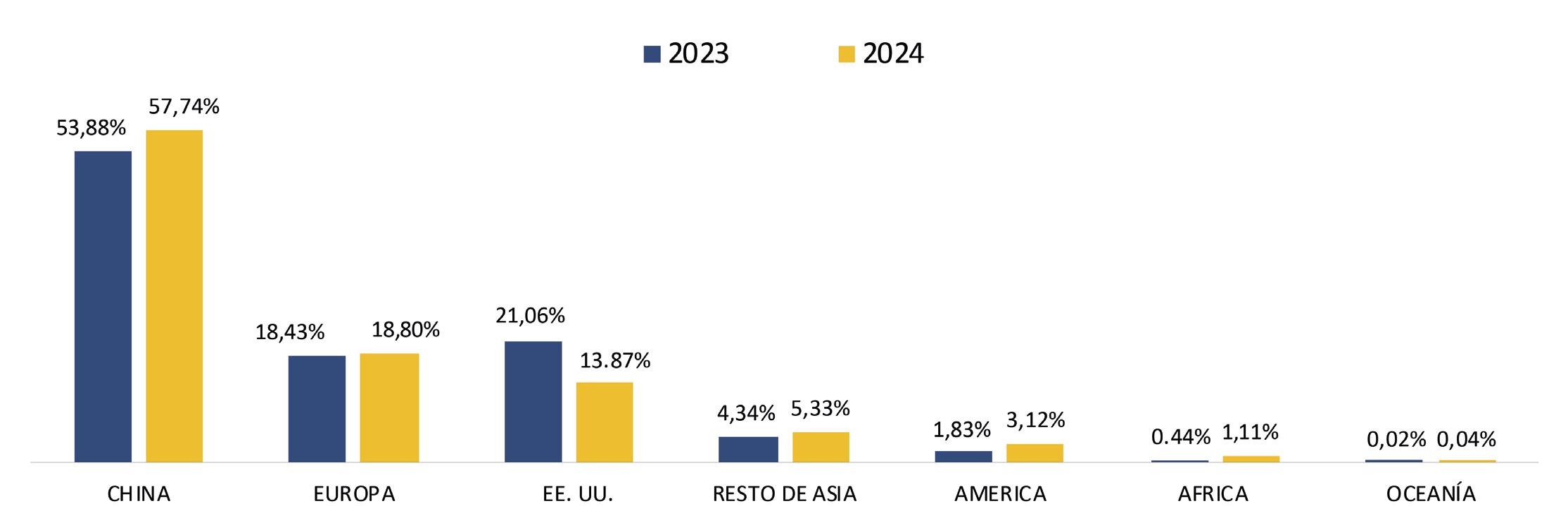
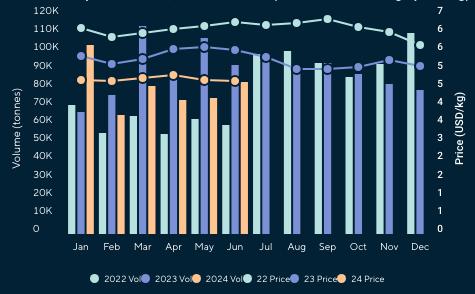
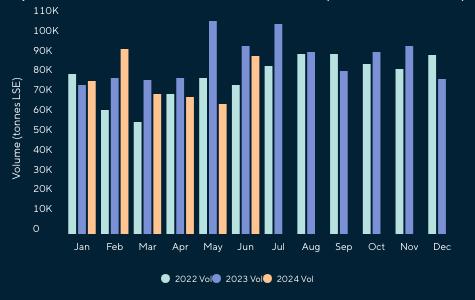
Reporte de mercado de China
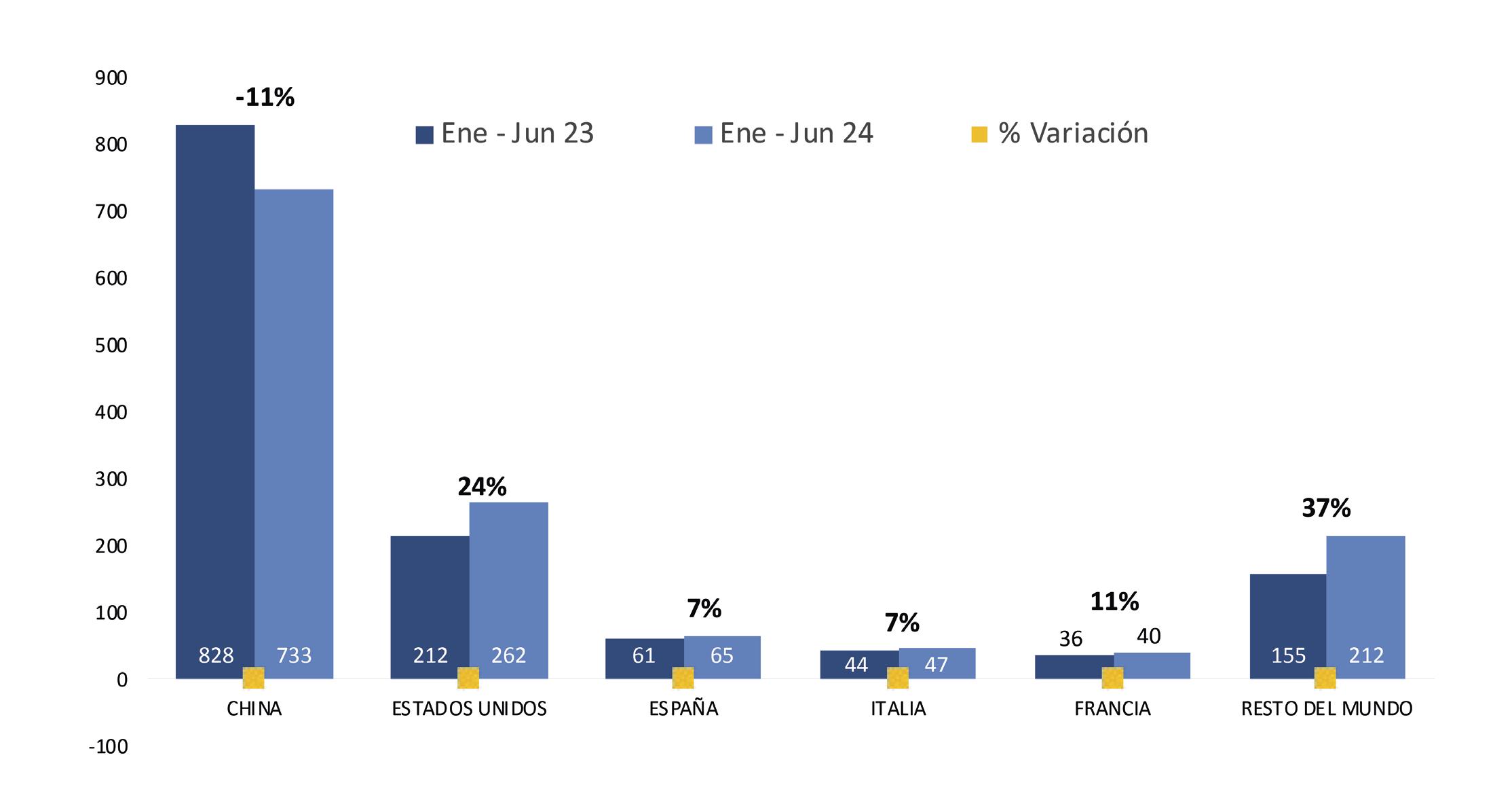
Reporte de mercado de EE. UU.
Noticias de interés
Noticias empresariales
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Corrección de estilo Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
Gabriela Nivelo gnivelo@cna-ecuador.com
El bienestar animal ha sido históricamente un pilar en la cría del camarón, no solo por razones éticas, sino también por su impacto directo en la productividad. Un camarón sano y libre de estrés es la base de un cultivo eficiente, con menores tasas de mortalidad y una mejor calidad de producto final. Este enfoque ha sido parte de nuestros protocolos de manejo, reconociendo que la salud del camarón es clave para alcanzar un rendimiento óptimo de nuestros cultivos.
Sin embargo, la noción de bienestar animal ha evolucionado, ampliándose para incluir aspectos como la reducción del estrés durante el manejo, el transporte y hasta el sacrificio. Estos elementos, que anteriormente se consideraban menos relevantes, ahora están ganando terreno en las discusiones internacionales sobre estándares de producción.
Los mercados internacionales, especialmente Europa, han sido fundamentales para impulsar la adopción de prácticas de bienestar animal en la acuicultura. De igual forma, las certificaciones han comenzado a incorporar criterios específicos de bienestar animal en sus estándares. Por ello, para acceder a estos mercados, los productores se ven obligados a cumplir con estas nuevas exigencias, lo que está llevando a una revisión de las prácticas tradicionales.
Este cambio no es un fenómeno aislado, pues forma parte de una tendencia mundial hacia una mayor responsabilidad en la producción de todo tipo de bienes. Los consumidores demandan cada vez más información y detalles sobre el origen y las condiciones en que se producen los alimentos que consumen, por lo que el bienestar animal se está convirtiendo en un criterio importante para asegurar la aceptación en estos mercados.
Aunque la adopción de prácticas de bienestar animal es un objetivo noble, su implementación presenta varios desafíos.
José Antonio Camposano Presidente Ejecutivo
Bienestar animal en la acuicultura de camarón: un debate en evolución
Si bien el bienestar animal continuará siendo un tema de creciente importancia, es crucial que el debate se aborde con realismo, reconociendo tanto los logros alcanzados como las limitaciones existentes.
La tecnología para monitorear y mejorar el bienestar animal en la cría de camarón aún está en desarrollo, y su aplicabilidad técnica y económica varía según la escala de producción. Los pequeños y medianos productores, en particular, pueden enfrentar dificultades para adoptar nuevas tecnologías o prácticas debido a limitaciones de recursos.
Además, la evaluación del bienestar en los camarones, que son animales con un comportamiento y biología únicos, es más compleja que en otras especies acuícolas. Esto recalca la necesidad de continuar invirtiendo en investigación y desarrollo para crear herramientas y prácticas que sean viables y efectivas en nuestro sector.
A pesar de los desafíos, la industria camaronera ecuatoriana ha realizado avances significativos en el mejoramiento de las prácticas de bienestar animal. Las mejoras en bioseguridad, manejo del agua, el uso responsable de antimicrobianos o su eliminación total, así como condiciones de cultivo han tenido un impacto positivo tanto en la sostenibilidad como en el bienestar de estos crustáceos.
Si bien el bienestar animal continuará siendo un tema de creciente importancia, es crucial que el debate se aborde con realismo, reconociendo tanto los logros alcanzados como las limitaciones existentes. No debemos apresurarnos a adoptar prácticas que, aunque bien intencionadas, no sean viables en el corto plazo bajo los esquemas de producción actuales.
El futuro de la industria camaronera depende de nuestra capacidad para equilibrar las demandas del mercado con la realidad técnica y económica de nuestras operaciones. Por ello, al seguir de cerca el desarrollo de prácticas de bienestar animal, estamos no solo respondiendo a las exigencias del mercado, sino también asegurando la sostenibilidad y competitividad de nuestra industria•
DIRECTORIO
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Ing. Roberto Aguirre
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Blgo. Carlos Sánchez
Ing. Diego Puente
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Andrés Rivadulla
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León
Marcó el camino hacia la acuacultura del futuro
Con la presencia de autoridades nacionales y provinciales se dio apertura al evento técnico comercial camaronero AQUAEXPO El Oro 2024, en el Hotel Oro Verde de Machala del 9 al 11 de julio pasado.
La mesa directiva estuvo conformada por Clemente Bravo, prefecto de la provincia de El Oro; Jenny Machuca, vicealcaldesa de Machala; Ana María Maldonado Carrasco, subsecretaria de Calidad e Inocuidad; Axel Vedani, subsecretario de Acuacultura; Marcelo Vélez, presidente del Directorio de la Cámara Nacional de Acuacultura (CNA); y Yahira Piedrahita, directora ejecutiva de la CNA, quien estuvo a cargo de dar la bienvenida a los participantes de esta actividad.
"En un momento donde nuestra industria enfrenta numerosos desafíos a nivel nacional e internacional, es imperativo que busquemos la eficiencia y controlemos los costos a lo largo de la cadena de valor del camarón. La Cámara Nacional de Acuacultura siempre será un aliado para tender puentes, fomentar el diálogo y ofrecer soluciones", expresó la directora de la CNA, entidad que organiza desde hace más de dos décadas este evento, quien destacó además la importancia de enfrentar los retos de la industria camaronera a través de la eficiencia y las mejores prácticas productivas.
Yahira Piedrahita, Directora Ejecutiva de la Cámara Nacional de Acuacultura
Autor: Shirley Suasnavas - Editora general de la Revista Aquacultura- ssuasnavas@cna-ecuador.com
Marcelo Vélez, Presidente del Directorio de la Cámara Nacional de Acuacultura
Por su parte, Marcelo Vélez, presidente del Directorio de la Cámara, destacó la resiliencia del sector acuícola frente a las dificultades del mercado: "El año pasado hablaba de la situación complicada de los mercados y de la confianza que tenía en la fortaleza del sector para reinventarse. La realización de este evento es una prueba de ello. Sabemos que hay productores que están encontrando maneras de producir de manera rentable, a pesar de la difícil situación actual del mercado".
Para finalizar, las palabras de inauguración estuvieron a cargo de Ana María Maldonado, subsecretaria de Calidad e Inocuidad, quien expresó su satisfacción al ver a tantos profesionales y empresarios reunidos con un propósito común: impulsar el desarrollo acuícola ecuatoriano. "Este evento AQUAEXPO El Oro es una plataforma clave para nuestro sector”, afirmó.
El principal propósito de los eventos AQUAEXPO es impulsar el desarrollo de toda la cadena de valor del camarón, a través de:
1.Fomento de innovación y tecnología:
AQUAEXPO fue un espacio clave donde se presentaron las últimas innovaciones y tecnologías en la producción acuícola. Esto permitió a los productores y técnicos del sector mantenerse actualizados y mejorar sus prácticas.
2.Intercambio de conocimientos: A través de conferencias, talleres y mesas redondas, AQUAEXPO facilitó el intercambio de conocimientos entre expertos, investigadores y profesionales del sector. Este intercambio es crucial para la adopción de nuevas técnicas y para enfrentar desafíos comunes.
3.Networking y colaboraciones: El evento reúnió a actores clave de la cadena de valor acuícola, incluyendo productores, proveedores, exportadores y reguladores. Este entorno propició el networking y la creación de alianzas estratégicas, lo cual es vital para el crecimiento y desarrollo del sector.
4.Actualización sobre normativas: Durante el evento, se abordaron temas relacionados con regulaciones y normativas tanto nacionales como internacionales. Esto es fundamental para que los productores ecuatorianos cumplan con los estándares exigidos por los mercados internacionales.
5.Promoción de la sostenibilidad: AquaExpo puso un fuerte énfasis en la sostenibilidad, promoviendo prácticas que buscan minimizar el impacto ambiental y asegurar la viabilidad a largo plazo del sector acuícola en Ecuador.
6.Impulso económico: El evento contribuyó al impulso del sector acuícola, que es uno de los pilares de la economía ecuatoriana. A través de la promoción de buenas prácticas y la mejora de la competitividad.
A continuación algunos testimonios de quienes fueron parte de este evento técnico comercial.
“Definitivamente, si no conocemos lo que está pasando en el mundo, no vamos a poder ser partícipes de esos beneficios. La Cámara Nacional de Acuacultura tiene un rol protagónico en la transferencia de información, para traer expositores de todas las partes del mundo y actualizar nuestros conocimientos”.
Walter Moncayo
Asistente del congreso
“En los stands hay bastante información importante, ya sea de los productos o servicios, para poco a poco ir innovando”.
Jimmy Quiroga
Visitante de la feria
“AQUAEXPO nos permite conocer directamente todo lo que ofrece la línea de acuicultura, y estar informados de las nuevas tendencias y procedimientos”.
Anthony Maldonado
Visitante de la feria
“Soy de Pedernales, Manabí, y curso la carrera de Biología y Trabajo en Larvicultura en una empresa familiar. En el congreso AQUAEXPO El Oro he adquirido conocimientos sobre nuevas líneas de alimentación, sobre todo para laboratorios, que fortalecerán mi desarrollo y el de mi compañía”.
Reynaldo Galarza
Asistente del congreso
AQUAEXPO El Oro 2024 contó con la participación de más de 50 empresas nacionales y extranjeras en su feria comercial, que se extendió sobre un área de exposición de más de 500 m2, donde se exhibió la última tecnología en productos y servicios para toda la cadena de valor del camarón.
El congreso contó con la participación de más de 15 expositores nacionales e internacionales, quienes abordaron temas cruciales como la reducción de costos,
"Estas charlas son muy pertinentes y relevantes para la producción acuícola en la provincia de El Oro".
Juan Garcés
Asistente del congreso
manejo de datos, salud animal, nutrición, producción, tecnología, mercado, ambiente y prevención de riesgos.
Durante su trayectoria, el evento AquaExpo de la Cámara Nacional de Acuacultura ha demostrado ser una valiosa oportunidad de aprendizaje y actualización para los actores del sector acuícola, reafirmando su condición de ser el espacio esencial para el crecimiento y desarrollo del sector en la región•
La sostenibilidad empieza en el primer eslabón de la cadena
Autores:
Leonardo S. Maridueña
Simone Maridueña
Dirección de Ambiente
Cámara Nacional de Acuacultura
lmariduena@cna-ecuador.com
Para ofrecer las bases necesarias que permitan al primer eslabón de la cadena productiva adoptar prácticas sostenibles en el tiempo, Sustainable Shrimp Partnership (SSP), en colaboración con la Cámara Nacional de Acuacultura (CNA), diseñó SCALE UP, un programa que les proporciona a los laboratorios una ruta clara hacia la sostenibilidad en la producción de camarón.
Esta iniciativa busca que los laboratorios de producción larvaria sean reconocidos por la aplicación de buenas prácticas, responsabilidad con el ambiente y compromiso con la sociedad.
El programa se estructura en torno a cuatro componentes clave:
- Aplicar los criterios de la guía técnica para la sostenibilidad de laboratorios de producción larvaria: contiene directrices específicas que abordan diversos aspectos de la producción y gestión, tanto ambiental como social.
- Capacitación del personal: se proporciona formación especializada al personal de los laboratorios sobre los principios de sostenibilidad y cómo aplicarlos en sus operaciones diarias. La capacitación asegura que todos los miembros del equipo comprendan y puedan implementar las prácticas recomendadas.
- Pre-auditorías o visitas técnicas: se realizan inspecciones en los laboratorios para identificar áreas de mejora y proporcionar recomendaciones específicas sobre los hallazgos encontrados, mediante la entrega de informes técnicos que los respalden.
- Auditoría final: tiene como propósito calificar al laboratorio en cuanto al cumplimiento de las medidas establecidas en la guía. Este será el paso final antes de la obtención del reconocimiento.
SCALE UP fue presentado en febrero pasado durante el evento técnico-comercial AQUAEXPO SANTA ELENA 2024. Desde entonces, ha atraído el interés de más de 40 laboratorios que han participado en las capacitaciones y están en proceso de obtener el reconocimiento dentro del proyecto piloto del programa.
Guía técnica para la sostenibilidad de laboratorios de producción larvaria
La guía técnica está basada en la normativa nacional y estándares internacionales relacionados con temas de ambiente, bienestar social, bienestar animal, sanidad, calidad acuícola y sostenibilidad.
A partir de estos estándares se desarrollaron medidas y criterios específicos, considerando el contexto bajo el cual se desarrollan las operaciones de los laboratorios de producción larvaria, lo cual incluye aspectos como:
- Lugar de aplicación (zona costera)
- Bienestar del personal con respecto a las actividades que se realizan en un laboratorio
- Necesidades específicas para la producción de Penaeus vannamei
- Insumos, desechos y tipo de efluentes generados
De esta forma, la guía puede ser aplicada en todos los laboratorios de producción de larvas de la especie Penaeus vannamei en cualquier parte del mundo.
Este documento destaca por su sostenibilidad, ya que integra aspectos fundamentales para que la producción se mantenga en el tiempo. Entre ellos se detallan los siguientes:
- Protección del ambiente: se establecen medidas que minimicen los posibles impactos negativos en el ecosistema, se asegure la calidad de los recursos y se optimice el uso de estos.
- Bienestar animal: se garantiza el adecuado tratamiento de las larvas en cuanto al no uso de antibióticos, la prohibición de la ablación o corte del péndulo ocular, y se promueve la decapsulación de la artemia sin el uso de sustancias tóxicas.
- Aspecto social: se establecen directrices que aseguren la responsabilidad del laboratorio tanto con el personal como con la comunidad circundante.
- Aspecto económico: se fomenta la inclusión de tecnologías avanzadas que no solo mejoren la eficiencia, sino que también reduzcan costos de producción a largo plazo.
Además, el desarrollo de la guía ha tenido en cuenta los Objetivos y Metas para el Desarrollo Sostenible de las Naciones
Unidas, asegurando que sus principios estén alineados con las metas globales para la sostenibilidad.
Criterios de evaluación
La guía abarca criterios claramente definidos en 10 secciones:
1.- Cumplimiento de requisitos legales: garantiza que el laboratorio cuenta con los permisos necesarios para su funcionamiento.
2.- Ubicación e infraestructura: asegura que el entorno no impacte negativamente en la producción y viceversa.
3.- Calidad del agua: establece criterios para asegurar que los efluentes sean de igual o mejor calidad que el agua que ingresa.
4.- Saneamiento: requiere procedimientos para garantizar la sanidad del medio en el que se desarrollan las larvas.
5.- Gestión de riesgo: busca reducir la posibilidad de ingreso de patógenos en el proceso de producción.
6.- Energía y combustibles: proyecta un uso eficiente de la energía y la reducción de impactos por contaminación.
7.- Almacenamiento: establece criterios para el manejo de insumos, materiales peligrosos y desechos dentro del laboratorio.
8.- Requisitos de producción: garantiza la trazabilidad y el bienestar animal en todas las etapas de producción.
9.- Responsabilidad ambiental: optimiza recursos, reduce la huella de carbono,
controla la contaminación y promueve la restauración del medio ambiente.
10.- Responsabilidad social: asegura el compromiso del laboratorio con la comunidad y el bienestar de los trabajadores.
Inspecciones
- Inspección de instalaciones: evaluación de todas las áreas del laboratorio, especialmente las zonas de almacenamiento, tratamiento de aguas residuales y áreas de producción.
- Revisión documental: verificación de los documentos legales, procedimientos y registros correspondientes a la operación del laboratorio.
Una vez obtenidos los resultados de la preauditoría, se programa una reunión con los responsables del laboratorio para su presentación a través de los siguientes documentos:
1. Matriz de conformidades y no conformidades: incluye observaciones y recomendaciones específicas para cada hallazgo.
2. Documento de hallazgos: contiene un registro fotográfico en el cual se explican las observaciones y se realizan las recomendaciones para subsanar las no conformidades.
Cabe resaltar que la información recopilada es utilizada únicamente con el propósito de
Imagen 2. Revisión de la documentación del laboratorio.
COYUNTURA
brindar las recomendaciones necesarias para el laboratorio, y esta no será de carácter público.
Hasta el momento, se han realizado 9 preauditorías de un total de 17 laboratorios que han presentado la solicitud correspondiente. Los resultados obtenidos hasta ahora han sido positivos. Entre los aspectos más destacados se pueden mencionar:
- La mayoría de los laboratorios cumple con todos los requisitos legales establecidos.
- Los laboratorios están ubicados en sitios aprobados por la normativa ecuatoriana para el establecimiento de laboratorios de larvas de camarón.
- El personal del laboratorio está consciente de la necesidad de estas medidas y está dispuesto a realizar las mejoras necesarias para reducir los posibles impactos que puedan generarse por el almacenamiento de combustible, la calidad de los efluentes y el establecimiento de los protocolos necesarios para mantener la calidad de la larva.
- En algunos de estos laboratorios ya se están realizando actividades de responsabilidad ambiental, como la limpieza de playas y áreas cercanas.
- Se ha identificado que existe un gran compromiso con la comunidad, pues no solo el personal pertenece al sector, sino que también aporta en muchas de las actividades que la comunidad más cercana solicite.
Una vez que los laboratorios demuestren el cumplimiento de la guía mediante la aplicación de las medidas correctivas, podrán solicitar la auditoría final. Esta se podrá realizar hacia finales de septiembre o entre los meses de enero-febrero de 2025.
Finalmente, se contempla que en el mes de octubre, durante AQUAEXPO GUAYAQUIL 2024, se entreguen los primeros reconocimientos a los laboratorios que cumplan con los requisitos establecidos dentro de SCALE UP.
Para participar en el programa, los laboratorios deben enviar una solicitud para la pre-auditoría que debe estar dirigida a la directora de Sustainable Shrimp Partnership, Pamela Nath, y demostrar su compromiso con el programa, incluyendo la disposición para recibir al personal técnico de SSP, participar en las reuniones requeridas y
aplicar las medidas correctivas que estén dentro de su alcance.
Primera capacitación: se realizó en el mes de abril en la Universidad Estatal de Santa Elena, y estuvo dirigida al personal técnico del laboratorio. Durante dos días, se abordaron temas como: bases de la guía, normativa, implementación de Buenas Prácticas y optimización en cuanto al uso de alimento.
Segunda capacitación: también se realizó durante abril pero en la ciudad de Guayaquil; tuvo una duración de un día y se centró en la explicación de las medidas, la aplicación de la guía y los procesos a seguir.
Tercera capacitación: se realizó a comienzos del mes de agosto y se desarrolló de manera virtual. Estuvo dirigida a quienes ya son parte del proceso, por lo que se enfocó en brindar recomendaciones para corregir los hallazgos encontrados y en la elaboración de procedimientos y planes necesarios para cumplir con la guía•
Para más información sobre este artículo, contactar a: lmariduena@cna-ecuador.com
Pamela Nath Directora
Desde que asumí mi rol en Sustainable Shrimp Partnership (SSP), he tenido la oportunidad de representar a nuestra industria en eventos internacionales, compartiendo los logros y avances de la acuicultura ecuatoriana de camarón. En estas plataformas, hemos demostrado cómo nuestro país no solo se ha posicionado como el principal productor de camarón a nivel mundial, sino también como un líder en responsabilidad social y ambiental.
En más de una ocasión me han preguntado: “¿Cuál es el secreto de Ecuador?, ¿qué les ha llevado a ser un referente en producción y sostenibilidad?”.
Para responder estas interrogantes, en SSP hemos decidido presentar una serie de tres artículos donde destacaremos uno de los elementos clave que ha impulsado nuestro progreso. A través del conocimiento de expertos en cada etapa de la cadena de valor, resaltaremos cómo la innovación tecnológica ha sido vital para nuestro desarrollo.
Sin embargo, creo que es importante subrayar que, además de la innovación, hay un factor igual de fundamental: el compromiso.
Existen muchas industrias con capacidad de innovar, pero lo que nos distingue es ese compromiso compartido por todos los actores del sector. Un compromiso que impulsa la mejora continua, la tecnificación y cada paso en esta carrera hacia la cima.
Hasta ahora, hemos alcanzado un rendimiento excepcional, pero nuestra misión no se detiene aquí. Mientras lees estas palabras, un sinnúmero de proyectos y tecnologías se están probando e implementando en Ecuador, todos con el objetivo de seguir mejorando el desempeño de nuestro camarón.
En esta primera edición, exploraremos las soluciones que han sido utilizadas en los laboratorios de larvas, el punto de partida para una producción sostenible.
El secreto del camarón ecuatoriano:
innovación + compromiso
Parte 1: Laboratorios de producción larvaria
Autores:
Pamela Nath
Sally Tabares
Sustainable Shrimp Partnership (SSP)
pnath@sustainableshrimp.org
Hace 26 años, los productores de camarón en Ecuador vieron cómo una industria en amplio crecimiento se desplomaba debido al virus de la Mancha Blanca, uno de los patógenos más devastadores para la acuicultura. Para el año 2000, esta enfermedad había arrasado los cultivos en el país, reduciendo las exportaciones en un 70% y limitando la producción a 46 mil toneladas.
Muchas de las piscinas que se encontraban en las 175 mil hectáreas destinadas a esta actividad quedaron vacías por la alta mortalidad que la Mancha Blanca ocasionó en los camarones. Las pérdidas económicas fueron tan significativas que varios productores se vieron obligados a cerrar de manera definitiva sus operaciones. Esta crisis estuvo a punto de colapsar totalmente el sector; sin embargo, también fue el punto de inflexión para dar inicio al crecimiento sostenible de una industria entera.
Quienes no conocen la historia de los camaroneros en Ecuador podrían imaginar que superar esta situación era una tarea imposible. Pero, a pesar de las dificultades y gracias a un factor clave que se destacará a lo largo de este reportaje, hoy en día Ecuador es el principal exportador de camarón a nivel mundial y un líder en prácticas sociales, ambientales y de calidad en su producción.
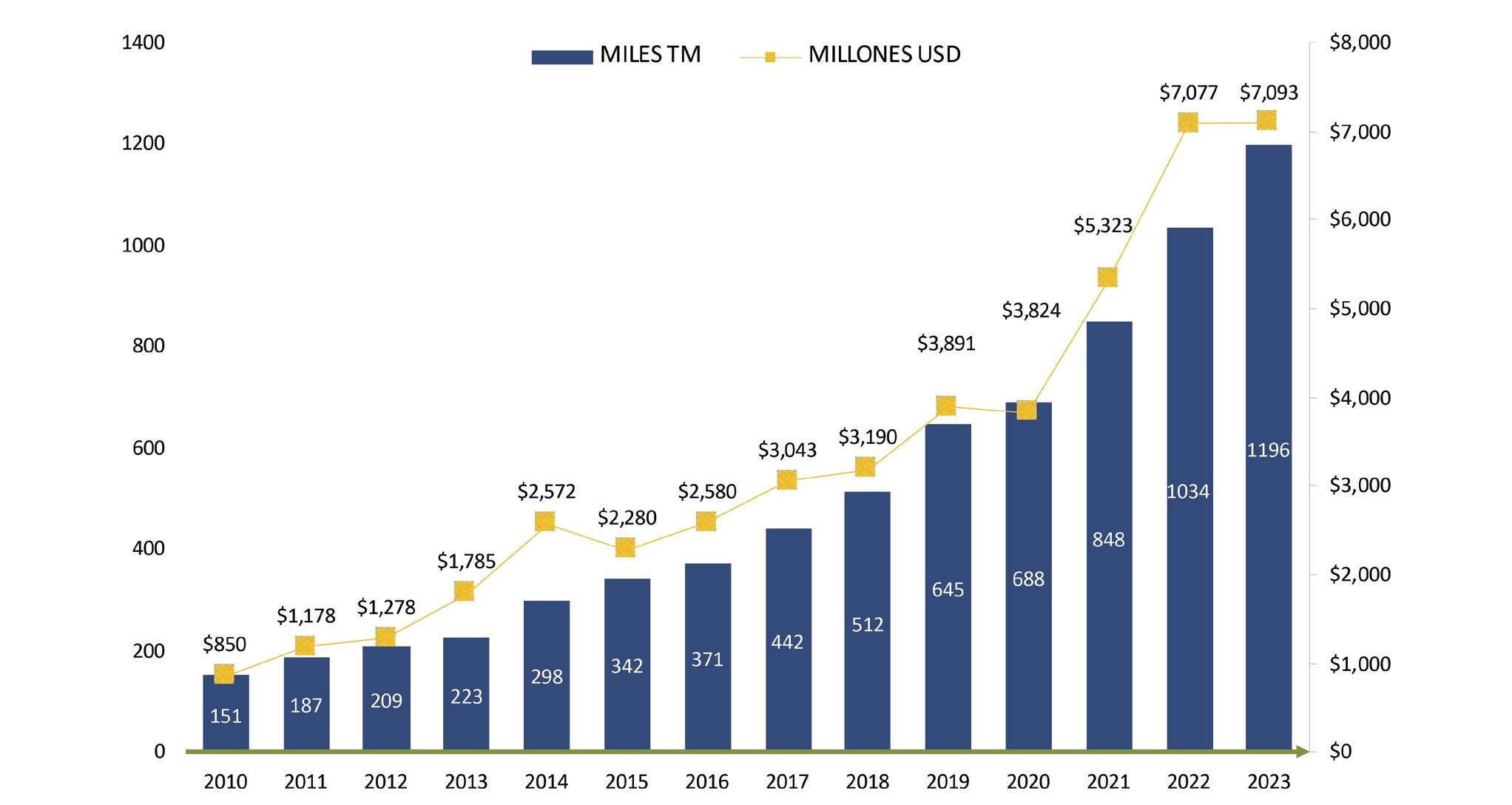
En 2023, desde sus 225 mil hectáreas de camaroneras, Ecuador llevó cerca de 1.2 millones de toneladas de camarón al resto del mundo, según datos de la Cámara Nacional de Acuacultura (CNA).
Comparando este escenario con la crisis del 2000, surgen una serie de preguntas: ¿Cómo, en un lapso de 23 años, Ecuador logró aumentar el volumen de su producción de camarón casi 26 veces? Y una interrogante aún más sorprendente: ¿Cómo se consiguió este aumento en la producción cuando el área destinada para cultivar camarones solo creció un 0,85% por año?
‘Spoiler Alert’: el factor clave fue y sigue siendo
la
innovación tecnológica, que
permite el incremento de la densidad de siembra, y que tiene una gran repercusión en la sustentabilidad de la acuicultura de camarón.
De acuerdo con la directora ejecutiva de la CNA, Yahira Piedrahita, “la tecnificación ha sido el promotor principal del crecimiento de la industria”. Actualmente, existen nuevas soluciones tecnológicas en cada etapa de la producción de camarón. Los avances en genética, biorremediación, formulación de alimentos, maquinarias y software son solo algunos de los recursos que ella enumera.
Pero, ¿cuál es el papel de las tecnologías y cómo ayudan al desarrollo sostenible de la industria? El coordinador de proyectos técnicos de Sustainable Shrimp Partnership (SSP), Leonardo Maridueña, expresa que “son herramientas que se usan para crear sistemas más eficientes que reduzcan los costos”. Este ahorro puede reflejarse a nivel económico, energético, en tiempo o en materias primas, influyendo en temas de contaminación y reducción de impactos ambientales y sociales.
Si regresamos a la historia del virus de la Mancha Blanca, encontraremos el despertar de una de las áreas tecnológicas que ha tenido mayor impacto en la industria y en las buenas prácticas de producción de camarón en Ecuador: los programas de mejoramiento genético.
Cuando esta temible enfermedad causó la muerte de millares de camarones, los animales que lograron sobrevivir en las piscinas levantaron el interés del sector. Fueron seleccionados y llevados a laboratorios de larvas para su reproducción, con el objetivo de crear una nueva generación de camarones que tengan la misma resistencia que presentaron los sobrevivientes. Así, se obtuvieron animales más robustos y de mayor calidad en comparación con los que se obtenían previamente del mar.
“El mejoramiento genético ha sido clave para la sostenibilidad de la actividad camaronera, ya que ha permitido eliminar la dependencia de la larva silvestre, proveniente del mar, eliminando una práctica de captura depredadora y protegiendo a otras especies y su ecosistema. Asimismo, el mejoramiento genético ha producido un animal más fuerte y adaptado a las condiciones de cultivo”, comparte Juan Manuel Afonso, responsable del programa genético de Biogemar, el laboratorio de producción de nauplios y larvas del Grupo Almar.
Afonso explica que la adaptación del camarón a las condiciones de cultivo reduce sus probabilidades de morir, lo que ayuda a reducir el desperdicio de alimento balanceado. De la misma forma, indica que cuando se mide la huella de carbono, se observa que los animales que crecen más rápido y tienen una menor tasa de mortalidad generan una menor huella ambiental.
Un programa genético es un plan para mejorar las características de una población de animales mediante la selección y reproducción. Este proceso implica identificar y elegir los organismos con las mejores cualidades genéticas y reproducirlos para que sus descendientes también posean esas características deseables. “No se está alterando la genética, solo se está escogiendo a la que sirve más”, aclara Afonso.
El genetista detalla que los programas de selección genética a partir de información familiar funcionan así: se toma un macho y una hembra, se obtiene su descendencia y esa descendencia constituye una familia. Luego, esa familia se compara con otras familias creadas de manera similar. Para que un programa genético sea efectivo, se necesitan entre 150 y 200 familias.
Luego, se coloca a todos juntos en un ambiente común, se observa cuáles se desempeñan mejor y se eligen a los que muestran mejores resultados. “Seleccionas las características que quieres mejorar para la siguiente generación. Por ejemplo, si estás enfocado en el peso, escogerás a los animales que tienen el mejor crecimiento”, describe Afonso.
El problema surge cuando se seleccionan a los dos animales que crecen más, una hembra y un macho, y descubren que podrían ser hermanos, pertenecientes a la misma familia. Aquí es donde entra en juego el programa con información familiar. “Para que funcione, es esencial identificar de qué familia proviene cada animal que se desea cruzar. Si los animales provienen de familias sin relación, se pueden cruzar con confianza”, enuncia. Para seleccionar a los mejores animales y conocer su familia, Afonso revela que es necesario realizar un análisis genético a cada animal.
En 2017, Biogemar lanzó un programa de mejoramiento genético en asociación con la Universidad de Las Palmas de Gran Canaria. Gracias a esta colaboración, se desarrollaron programas con información de la estructura familiar que potenciaron aún más las ganancias genéticas en sus productos. Por ejemplo, en cada generación nueva de camarón, obtienen un aumento de más del 10% en crecimiento.
Otra de las innovaciones tecnológicas que han contribuido al crecimiento y desarrollo de la industria son los probióticos, una solución enfocada en la salud y bienestar del animal y el ambiente.
El profesor e investigador de la Universidad Federal de Santa Catarina (Brasil), José Luiz Mouriño, expone que el camarón tiene naturalmente una microbiota, es decir, un conjunto de bacterias en su cuerpo. Algunas de ellas, como los Vibrios, pueden ser patógenas y causar enfermedades. Después de que las hembras desovan y los huevos se incuban en el laboratorio, se observa una alta presencia de Vibrios y otras bacterias dañinas.
“En cada cambio de fase de los estadios larvarios, el animal experimenta cambios enzimáticos y fisiológicos que son estresantes. Por esto, durante la larvicultura es muy importante dirigir la microbiota del camarón hacia una composición más saludable. Este es el objetivo de los probióticos”, indica el profesor.
Mouriño, quien también es consultor internacional para la empresa DSMFirmenich, señala que si se revisan los objetivos del sector camaronero y de la acuicultura en general durante los últimos 30 años, se notará que las enfermedades siempre han sido el principal obstáculo para el crecimiento de la actividad. Estas limitan la capacidad de producir a la densidad deseada y alcanzar las cantidades de producción que se buscan, además de generar gastos significativos en tratamientos.
El investigador indica que en los laboratorios de larvas se utilizan probióticos por tres razones fundamentales. Primero, para aumentar la diversidad microbiana. “Cuantas más cepas se utilicen y mayor sea la diversidad de microorganismos, tanto en el estanque como en el tracto intestinal del camarón, mejor será la salud general de los animales”, advierte.
En segundo lugar, los probióticos ayudan a inhibir los patógenos. Al aumentar la diversidad microbiana y la presencia de bacterias específicas y funcionales, los probióticos favorecen una colonización más eficiente. Esto reduce los puntos donde los patógenos pueden atacar y disminuye la cantidad de nutrientes disponibles para ellos.
Y en tercer lugar, los probióticos producen sustancias, como biocina y ácidos orgánicos. Estas sustancias tienen el efecto de ‘desplazar’ a las bacterias patógenas, como los Vibrios, al competir con ellas por espacio y nutrientes. Al crear condiciones desfavorables para los Vibrios y ocupar los lugares donde podrían haberse asentado, los probióticos reducen la cantidad de estos patógenos en el sistema.
El consultor destaca varios beneficios de usar probióticos. Por un lado, ayudan a usar menos agua y a mantenerla en mejor
estado. Por otro lado, disminuyen el riesgo de entrada de patógenos, lo que contribuye a reducir el uso de químicos en la acuicultura, como los antibióticos. Esto ayuda a prevenir la resistencia antimicrobiana.
“Mientras menos químicos se usen en el laboratorio y más se fomente una mayor diversidad bacteriana, las larvas se desarrollan más fuertes. Una buena alimentación, junto con este manejo de bacterias, mejora el crecimiento del animal, haciendo que se desarrolle más rápido y con menos problemas cuando se lleva a las fincas”, explica.
En la misma línea, otra área donde los avances tecnológicos han sido esenciales para hacer que la larvicultura tenga un mejor desempeño social, ambiental y económico, es la de las soluciones en alimentos de las larvas de camarón.
En los laboratorios de larvas, el ciclo de producción dura 15 días aproximadamente. A partir de la fase larval, los animales reciben alimentos vivos y alimentos artificiales. Entre los alimentos vivos se encuentran las microalgas y los nauplios de artemia, que son crustáceos microscópicos, mucho más pequeños que las larvas de camarón.
Para alimentar adecuadamente a las larvas, la artemia debe cumplir con ciertas características tanto en su composición nutricional como en su estadio naupliar. La composición nutricional de la artemia consiste en proteínas, lípidos, vitaminas y minerales, mientras que su estadio naupliar debe corresponder al primero después de la eclosión, conocido como Instar I.
Sin embargo, conseguir una suspensión pura de nauplios de artemia es todavía un desafío. Esto se debe a que, tradicionalmente, para descapsular los quistes de artemia y liberar los embriones, se usan productos químicos que en combinación generan una reacción exotérmica y provocan la desnaturalización de la capa coriónica de los quistes.
Estos químicos requieren precauciones especiales para garantizar la seguridad del operador durante el proceso de decapsulación. Para ello, es fundamental utilizar métodos de bioseguridad, como
guantes, mandiles impermeables, gafas protectoras, mascarillas con filtro de carbono, botas de caucho y cascos.
“Esta técnica es considerada peligrosa para los trabajadores, dañina para el medio ambiente y, como el uso de químicos afecta la calidad de la artemia, también tiene un impacto negativo sobre la relación costobeneficio”, expone Ermel Viteri, encargado regional de soporte técnico de ventas de INVE Aquaculture.
En investigaciones, “se ha descubierto que es posible sustituir la artemia en la alimentación de las larvas de camarón con otros microrganismos planctónicos”, indica Viteri. Estos microorganismos pueden proporcionar buenos resultados en los estudios, pero en la producción comercial, reemplazar la artemia no es práctico.
“Aunque no existe una única alternativa para reemplazar los alimentos vivos en las dietas de larvas, la artemia sigue siendo la opción más completa a nivel nutricional debido a su composición proteínica - lipídica y su disponibilidad en el medio natural”, comenta.
Además de los métodos químicos tradicionales para abrir los quistes de artemia, que son perjudiciales para la salud y el medio ambiente, Viteri comparte que se ha desarrollado una solución sostenible alternativa.
Esta consiste en quistes de artemia que están recubiertos con un material magnético no tóxico y que no afecta las características de eclosión globales de los quistes. Mediante el uso de una herramienta de separación especial con imanes, se atraen solo los quistes y cáscaras vacías, dejando libres los nauplios de artemia. Luego, los quistes se adhieren a los imanes y se pueden retirar fácilmente de la suspensión de nauplios.
“Esto permite la cosecha de nauplios de artemia en mayor cantidad y calidad, y en menor tiempo, sin comprometer la vitalidad de los nauplios, ya que no usa la fuerza física o una reacción química”, revela
Viteri, “en general, es una estrategia más eficiente que permite a los productores incrementar la calidad y sobrevivencia de los nauplios cosechados y apoyar el crecimiento saludable de las larvas de camarón”
En cuanto a las microalgas, estas son la primera nutrición de las larvas de camarón y se consumen desde Zoea 1 hasta Postlarva 1. El gerente de Investigación y Desarrollo de Skretting Aquaculture Research Centre, César Molina, explica que estas algas aportan diferentes nutrientes, dependiendo de la especie de alga que se añada al tanque de cultivo. Además, contienen compuestos como carotenoides y polisacáridos, que pueden fortalecer el sistema inmunológico de las larvas.
“La producción y el uso de algas también ayudan a mejorar la calidad del agua en los tanques de cultivo al reducir los niveles de amonio y otros compuestos tóxicos, creando un ambiente más saludable para las larvas”, declara Molina.
Biogemar, el laboratorio de larvas mencionado previamente, ha implementado nuevas tecnologías para mejorar la producción de algas. Están utilizando biorreactores, que son equipos avanzados que permiten cultivar una mayor cantidad de algas en un espacio reducido y en condiciones extremadamente limpias. Esto significa que en el mismo volumen obtienen muchas más algas, lo que les ayuda a
ahorrar espacio y a producir algas de mejor calidad. Al principio, Biogemar adquirió estos biorreactores como una prueba, pero ahora planean expandir su uso.
Por otro lado, entre los alimentos artificiales que se les provee a las larvas de camarón, se encuentran los alimentos balanceados.
“Una de las tecnologías que se ha usado en la producción de alimento balanceado para larvas es la microextrusion en frío, lo cual permite que los nutrientes no se dañen por efecto del proceso y se logran los tamaños que requiere la larva desde sus primeros estadios de vida”, relata Molina.
El gerente de Investigación y Desarrollo de Skretting indica que la tecnología avanzada permite la formulación de dietas que satisfacen de manera precisa las necesidades nutricionales de los camarones. Además, añade que una nutrición óptima fortalece el sistema inmunológico de los camarones, haciéndolos menos susceptibles a enfermedades.
(Nota del Autor: Las tecnologías utilizadas en la producción de alimentos balanceados son un tema amplio y de gran interés. Dado su impacto y relevancia, se abordarán con más detalle en la segunda edición de esta serie de artículos).
Esta búsqueda constante de mejorar el desempeño de los procesos en la industria camaronera ecuatoriana ha demostrado que, incluso en los momentos más críticos, es posible encontrar caminos hacia el éxito.
En la siguiente parte de este reportaje, presentaremos cómo las fincas camaroneras en Ecuador han adoptado soluciones que permiten producir más, y que aseguran que la industria perdure a largo plazo. Estas innovaciones son esenciales para entender cómo Ecuador sigue liderando la producción de camarón, cuidando tanto del medio ambiente como del bienestar de todos los involucrados.
La directora de SSP, Pamela Nath, expresa que el avance integral de la tecnología a lo largo de toda la cadena de valor ha sido clave para el éxito alcanzado como país. Asimismo, anuncia que este desarrollo ha permitido producir camarón de manera eficiente y liderar en buenas prácticas a nivel mundial.
“Sin embargo, la tecnología por sí sola no es suficiente. Lo que realmente hace la diferencia es el compromiso constante de cada uno de los actores en la cadena de producción. Este compromiso es lo que impulsa la mejora continua, la adopción de nuevas soluciones y la superación de los desafíos que enfrentamos”, concluye.
Así, el éxito de Ecuador, con un incremento en la producción de camarón que superó ampliamente la expansión del área de cultivo, no solo es un triunfo económico, sino una lección de que la sostenibilidad y la tecnología, guiadas por el compromiso de toda una industria, pueden y deben ir de la mano para asegurar el futuro de la acuicultura global•
Para más información sobre este artículo, contactar a: pnath@sustainableshrimp.org
TIPOS DE EXTORSIÓN
Unidad Antisecuestro y Extorsion (UNASE)
Extorsión virtual
En esta modalidad, los delincuentes suelen presentarse como líderes de organizaciones criminales nacionales o internacionales, aunque no proporcionan información precisa sobre la ubicación de la víctima. Las amenazas que emiten suelen ser vagas, afirmando que “han recibido órdenes” de atentar contra la integridad de la persona, pero sin aportar detalles concretos. Por lo general, la exigencia de dinero es inmediata y no se establecen plazos para el pago. Estos extorsionadores emplean técnicas de ingeniería social, buscando que la víctima revele información clave de manera voluntaria.
Por otro lado, existe la extorsión siciliana que se originó en el sur de Italia durante la década de 1950, con la aparición de los carteles de la mafia. Los delincuentes de la “Cosa Nostra” exigían pagos periódicos a los negocios a cambio de protección.
Extorsión real
En este tipo de delito, los delincuentes proporcionan datos específicos sobre la víctima, como su ubicación exacta o una descripción detallada de su vestimenta, e incluso le envían fotografías de su domicilio o lugar de trabajo. La información que manejan no proviene de redes sociales, lo que genera mayor temor en la víctima. Además, en muchos casos se emplea violencia directa o amenazas tangibles contra la persona o sus propiedades, como disparos o explosivos. Los delincuentes establecen plazos específicos para el pago a cambio de no causar daño.
Extorsión real vs. Extorsión virtual
En contraste, la extorsión virtual se distingue por la falta de precisión en la información proporcionada por los extorsionadores. Estos suelen presentarse como miembros de organizaciones criminales, pero sus amenazas son más generales y carecen de detalles específicos sobre la víctima. La demanda económica es inmediata, sin plazos claros, y los extorsionadores utilizan la manipulación psicológica para obtener información.
RECOMENDACIONES:
No proporcionar datos personales
•Los códigos QR ubicados en restaurantes, tiendas o estacionamientos pueden ser utilizados como trampas por estafadores. Es importante que las personas verifiquen la autenticidad de estos códigos antes de escanearlos. No deben, bajo ninguna circunstancia, compartir su información personal o bancaria con servicios en línea que no sean confiables. Mantenerse alerta ante este tipo de fraudes es fundamental para proteger su seguridad y privacidad.
•En un mundo cada vez más conectado, es crucial no compartir datos financieros ni personales a través de llamadas, correos electrónicos o mensajes. Además, el uso de contraseñas seguras y la habilitación de la autenticación de dos factores son medidas clave. Mantener precaución al navegar en internet también es fundamental; es aconsejable desconfiar de ofertas que parezcan demasiado buenas para ser verdad, y verificar la autenticidad de empresas y personas antes de compartir datos.
•Se recomienda no ingresar a ningún enlace ni abrir archivos adjuntos provenientes de remitentes desconocidos. Incluso, si el mensaje proviene de un contacto conocido, lo mejor es confirmar directamente con la persona antes de interactuar con el enlace. Las llamadas inesperadas representan otra táctica utilizada por los estafadores o vacunadores. Si alguien se comunica alegando pertenecer a una institución oficial, lo más seguro es colgar y llamar directamente a la entidad para confirmar la veracidad de la información. En redes sociales, es importante no compartir demasiada información personal y ser cauteloso con solicitudes de amistad o mensajes.
•Además, es fundamental realizar compras solo en sitios web seguros que cuenten con protocolos de seguridad, identificados por "https://", y revisar las opiniones de otros usuarios antes de completar una transacción. Para garantizar la protección de los dispositivos, se deben mantener actualizados el sistema operativo, el navegador y el software de seguridad. Los expertos también recomiendan desconfiar de
las solicitudes urgentes, una característica común en muchas estafas.
•Instalar identificador de llamadas
•Restringir acceso en dispositivos
•Mantener actualizado el software en todos los dispositivos
•Manejo cifrado de claves (cambiarlas constantemente)
•Evitar hacer clic en enlaces de dudosa procedencia
•No llenar formularios en línea
•Evitar llenar datos por medio de códigos QR
•Evitar dejar datos en ánforas de concursos
•Ser cauteloso con la información que se publica en redes sociales
Protegerse contra la extorsión e incluso el secuestro requiere una combinación de precauciones en la protección de la información personal y la seguridad física. Es fundamental mantener discreción en redes sociales, evitando compartir detalles sobre la rutina diaria, la ubicación o la situación financiera. La información personal y documentos sensibles deben resguardarse de manera segura, y es aconsejable evitar publicar contenido que revele demasiado.
Educarse y educar a los demás sobre las tácticas de extorsión, como el secuestro virtual o la extorsión telefónica, ayuda a identificar y prevenir amenazas. El apoyo de las autoridades es crucial en estos casos, y las personas deben seguir siempre las instrucciones de seguridad que estas proporcionen. También se debe tener especial cuidado en los entornos públicos, manteniendo discreción sobre temas sensibles y evitando la exhibición de objetos de valor que puedan atraer la atención de delincuentes.
¿Qué debe hacer una persona si es víctima de extorsión?
Se debe verificar si los datos proporcionados por el extorsionador coinciden con información publicada en sus redes sociales. Este análisis puede ayudar a determinar la gravedad y veracidad de la amenaza. Ante cualquier sospecha o peligro inminente, la víctima debe comunicarse con el ECU911 y solicitar asistencia especializada de autoridades como la Unidad Antisecuestros y Extorsión (UNASE) de la Policía Nacional, que están capacitadas para manejar este tipo de situaciones de alto riesgo.
LLAME AL ECU911
Para evitar ser víctima de extorsión en Ecuador, es crucial mantener la discreción tanto en la vida pública como en la virtual. Es importante no divulgar información personal o financiera en redes sociales y evitar compartir detalles sobre rutinas diarias. Además, se recomienda desconfiar de llamadas o mensajes sospechosos, no proporcionar datos sensibles y reportar cualquier intento de extorsión a las autoridades competentes. Implementar medidas de seguridad, como variar las rutas diarias y utilizar sistemas de vigilancia en el hogar o negocio, también puede ser efectivo para prevenir amenazas. La educación sobre los métodos más comunes de extorsión y mantenerse informados son claves para protegerse y evitar caer en este tipo de delitos.
Ecuador mantiene Estado de 'guerra interna' contra organizaciones criminales
Desde el 9 de enero de 2024, Ecuador enfrenta una 'guerra interna' contra más de 20 organizaciones criminales identificadas como terroristas, según un decreto del presidente Daniel Noboa. Esta declaración de conflicto armado se produjo un día después del establecimiento del estado de excepción, que tenía una duración prevista de 90 días, pero que se ha extendido indefinidamente debido a la persistencia de las amenazas. Aunque el estado de excepción concluyó, las Fuerzas Armadas (FF.AA.) continuarán operando sin una fecha límite para garantizar la soberanía y la integridad territorial del país. El conflicto armado se mantendrá mientras los grupos terroristas sigan operando y afectando significativamente al Estado, como se detalla en el decreto ejecutivo de Noboa. El Centro de Inteligencia Estratégica (CIES) emitió el 5 de abril de 2024 dos informes secretos que indican que los grupos armados siguen activos, con bandas terroristas operando en al menos 20 provincias de Ecuador. Por ello, los militares continuarán sus operaciones para prevenir y erradicar las actividades de estas agrupaciones, con una intervención que se prolongará de manera indefinida.
Durán | Fuerzas Armadas y Policía Nacional realizan operativos simultáneos diariamente
Fuerzas policiales y militares llevaron a cabo operativos antidelincuenciales simultáneos en varias zonas de Durán, en la provincia de Guayas. Se desplegaron diariamente en al menos 20 zonas críticas del cantón, con el objetivo de reforzar el control tras recientes episodios de violencia que se han producido en la localidad. Durán ha experimentado un incremento alarmante en homicidios, con 192 muertes violentas registradas desde enero, lo que representa un aumento del 300% en comparación con el mismo período del año pasado. Las acciones buscan contrarrestar la violencia asociada con disputas entre bandas por el control territorial para actividades criminales como narcotráfico y extorsión.
Cumbre Latinoamericana de Seguridad En Guayaquil se llevó a cabo la Cumbre Latinoamericana de Seguridad, que contó con la presencia de delegaciones de 13 países de América y Europa, incluyendo Italia, España, Francia, Reino Unido, Estados Unidos, Colombia, Costa Rica y Chile. El presidente Daniel Noboa inauguró este evento con un discurso en el que abordó la situación del país, acompañado por la ministra del Interior, Mónica Palencia. Durante el encuentro se firmó la Alianza para la Seguridad, Justicia y Desarrollo, con el propósito de buscar financiamiento
y asesoría técnica para combatir el crimen en la región. La delegación ecuatoriana también incluyó a la canciller Gabriela Sommerfeld; Félix Wong, ministro de Gobierno; Giancarlo Loffredo, ministro de Defensa, y Juan Carlos Vega, ministro de Economía y Finanzas. Esta cumbre reflejó un compromiso global por fortalecer la cooperación en la lucha contra el crimen, demostrando un esfuerzo integral para abordar tanto los desafíos internos como las colaboraciones regionales en la búsqueda de soluciones efectivas para la seguridad y estabilidad del país.
CNA activa red de videovigilancia público-privada para mejorar capacidad de respuesta en materia de seguridad Con el objetivo de enfrentar la creciente inseguridad en las rutas de transporte de productos acuícolas, la Comisión de Seguridad de la Cámara Nacional de Acuacultura (CNA) ha dado un paso crucial para el resguardo de la industria.
En colaboración con la Policía Nacional, ECU911, Prefectura del Guayas y el sector privado, se ha implementado una red de cámaras de videovigilancia en zonas georreferenciadas de alta incidencia delictiva. En esta primera fase, se han instalado cuatro cámaras: una móvil y tres fijas, en la zona de Taura (Guayas). El monitoreo de estas cámaras está a cargo del ECU911, mientras que las cámaras espejo se encuentran instaladas en el centro de vigilancia del sector privado.
Este sistema permite el monitoreo constante de las operaciones de transporte de camarón y otros productos acuícolas, mejorando la capacidad de respuesta ante incidentes que comprometan la seguridad de conductores y productos. Como resultado, ha habido una notable disuasión de grupos delictivos en la zona. Además, la CNA ha logrado un acuerdo clave con el ECU911 que permitirá a las empresas del sector camaronero acceder a dispositivos de radio inteligentes troncalizados, conectados directamente a la frecuencia del Sistema Integrado de Seguridad. Esta tecnología mejora los tiempos de respuesta, al facilitar una comunicación directa y ágil con los operadores de emergencia.
Actualmente, se están llevando a cabo estudios de factibilidad técnica para la instalación de más cámaras de videovigilancia, en coordinación con los sectores público y privado. Una vez que estén listos estos estudios, el sector privado se encargará de la instalación y mantenimiento de los equipos, conforme a lo estipulado en el convenio.
Es necesario precisar que el sector camaronero ha invertido más de 80 millones de dólares para contrarrestar la ola delictiva que afecta a la industria, contratando empresas de seguridad privadas y adoptando sistemas de vigilancia, equipos de comunicación y tecnologías de inteligencia artificial. A través de su Dirección de Seguridad, la Cámara Nacional de Acuacultura recopila información, coordina alertas y trabaja en conjunto con los equipos técnicos de la Policía Nacional y la Armada del Ecuador, manteniendo al sector camaronero informado sobre la incidencia delictiva en el país, mediante los boletines de seguridad CNA•
ARTÍCULOS TÉCNICOS ÍNDICE
Edición 160 - Agosto 2024
29 Síndrome de las Heces Blancas, un trastorno multifactorial del camarón cultivado: una breve revisión
36 La diversificación ecológica revela rutas de aparición de patógenos en poblaciones endémicas de Vibrio vulnificus
50
59
Evaluación de fitoesteroles como alternativa al colesterol en dietas prácticas sobre el crecimiento y la inmunidad no específica de Penaeus vannamei
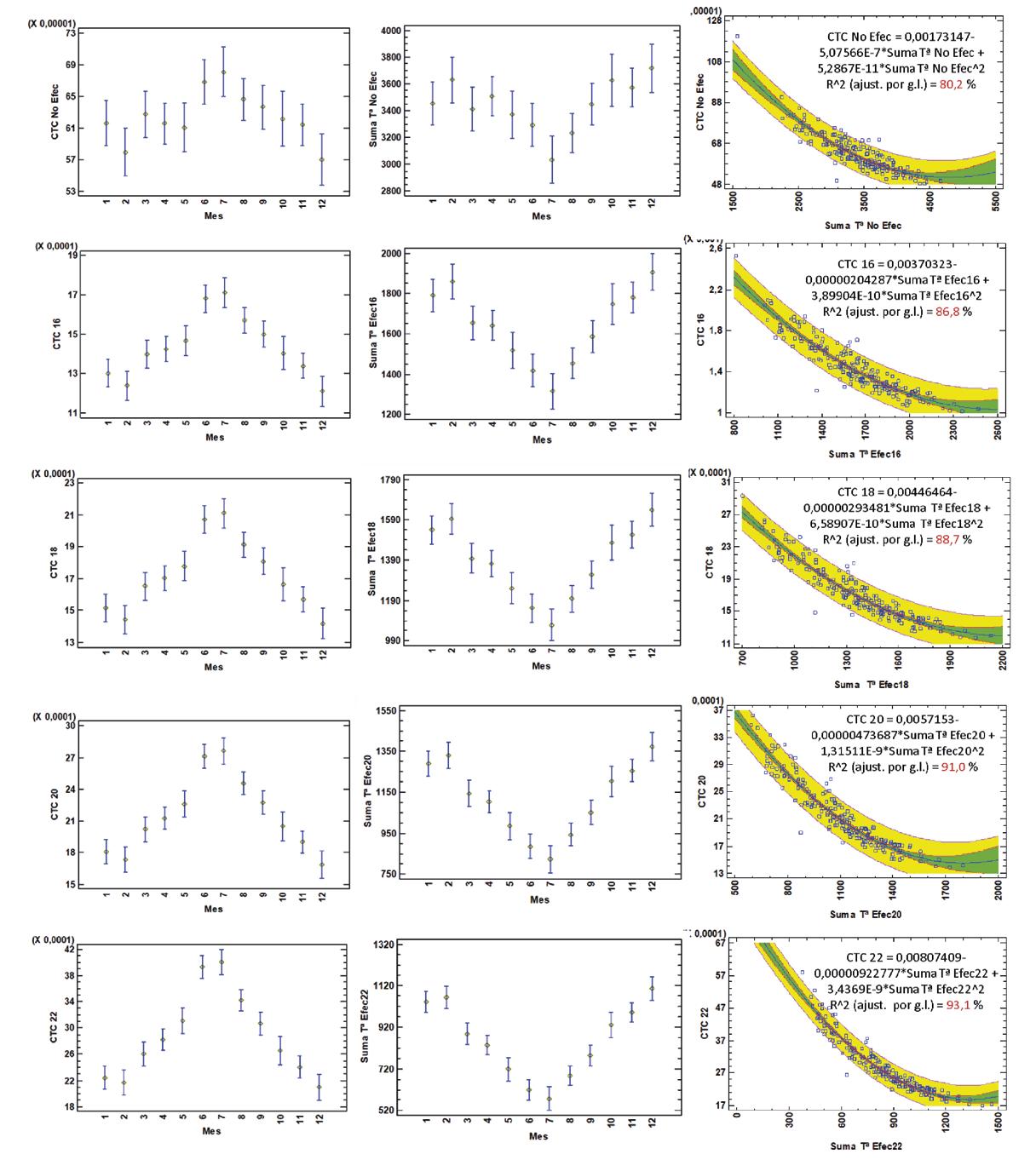
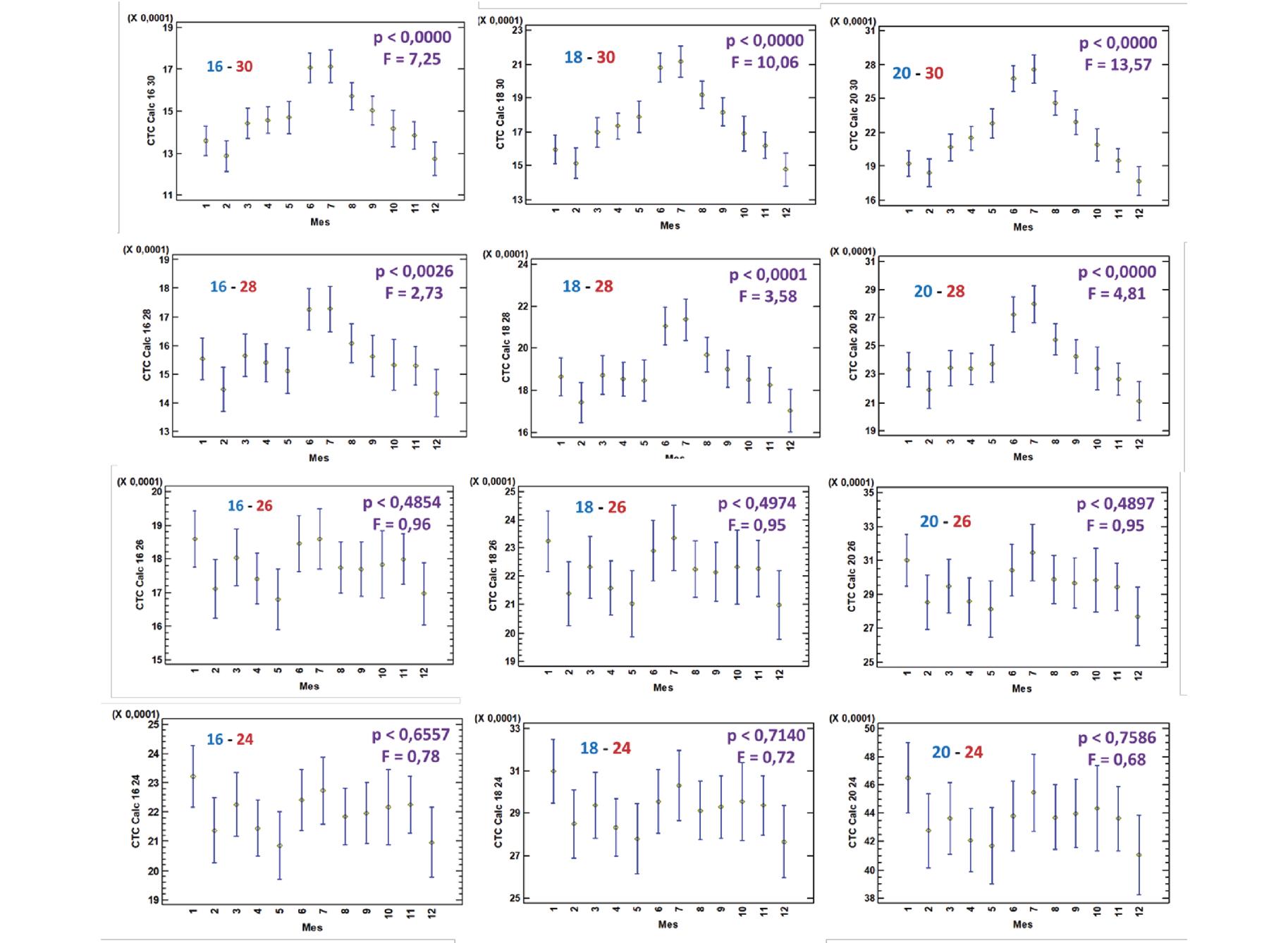
Comparación del coeficiente térmico de crecimiento en el camarón Penaeus vannamei en Ecuador con diferentes sistemas de alimentación
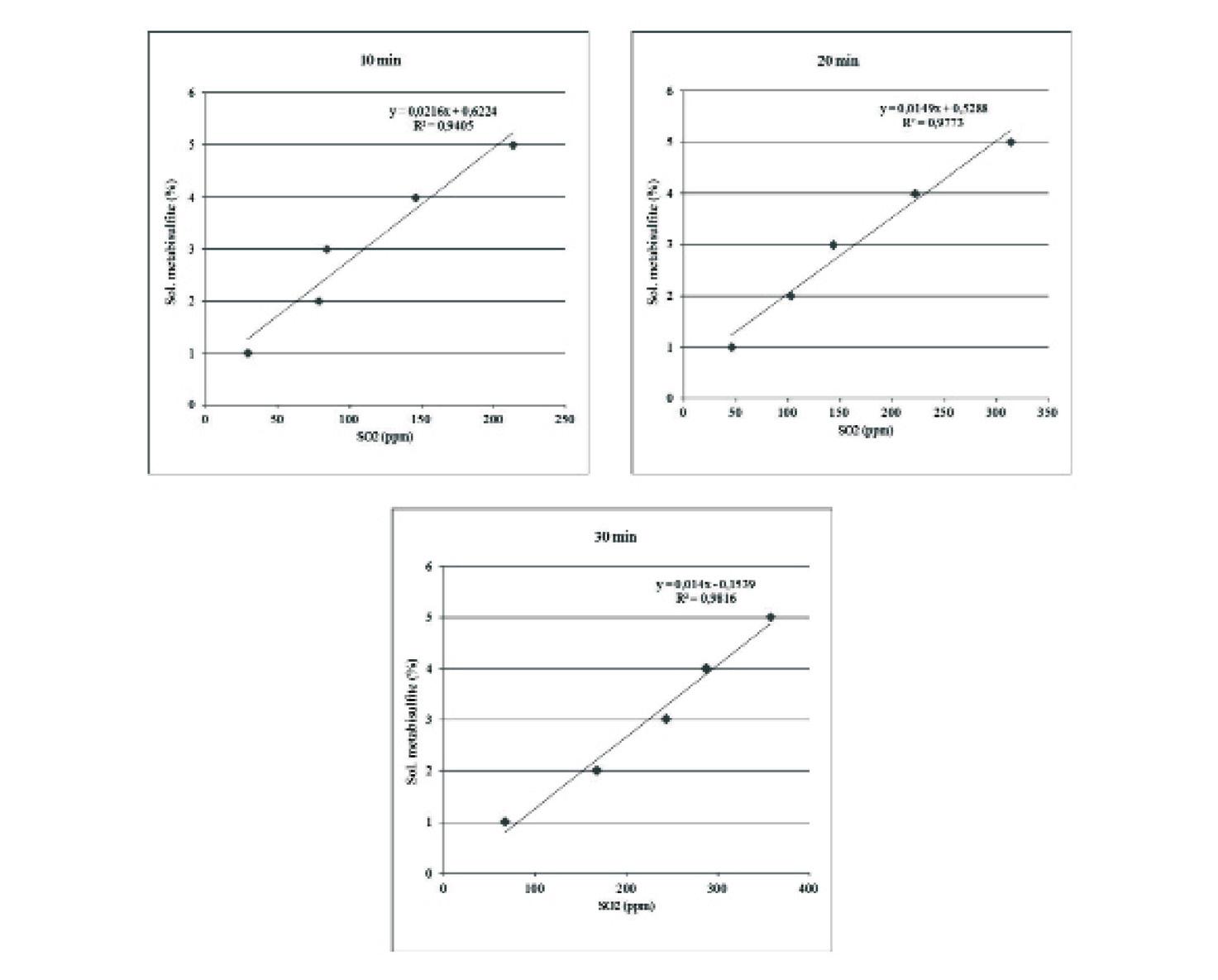
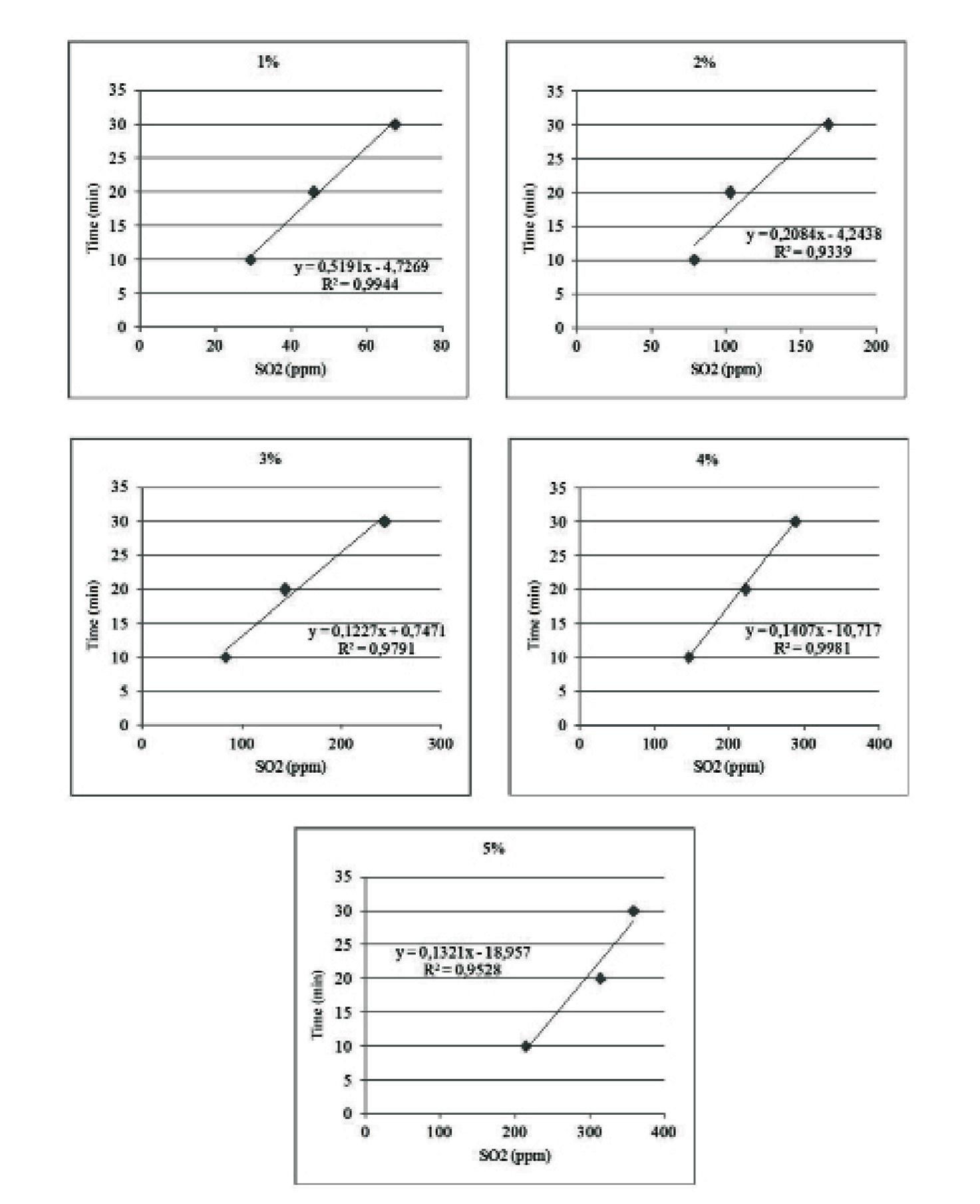
66 Estandarización de concentraciones de solución de metabisulfito de sodio y tiempo de inmersión para el camarón de cultivo Litopenaeus vannamei
Síndrome de las Heces Blancas, un trastorno multifactorial del camarón cultivado: una breve revisión
Autores: Patharapol Piamsomboon1,2
Jee Eun Han3,*
1 Departamento de Medicina Veterinaria, Facultad de Ciencias Veterinarias, Universidad de Chulalongkorn, Bangkok 10330, Tailandia.
2 Centro de Excelencia para la Investigación Veterinaria Médica de Animales Acuáticos, Universidad de Chulalongkorn, Bangkok 10330, Tailandia.
3 Facultad de Medicina Veterinaria, Universidad Nacional de Kyungpook, Daegu 41566, República de Corea.
El síndrome de Heces Blancas (SHB) es un trastorno gastrointestinal que afecta a los camarones peneidos cultivados en todo el mundo, que se caracteriza por la presencia de hilos fecales blancos flotando en la superficie del agua de los estanques de engorde. Los camarones infectados generalmente presentan intestinos medios blanquecinos a amarillentos, crecimiento retardado, alta variación de tamaño, crecimiento diario promedio reducido, tasas de conversión alimenticia elevadas, exoesqueletos sueltos y, a veces, mortalidad [1, 2]. La histopatología del intestino ha revelado una pared intestinal delgada combinada con el desprendimiento de células epiteliales intestinales y la reducción o desaparición de microvellosidades [ 3 ]. El SHB, que generalmente ocurre durante 50 a 60 días de siembra, debilita al camarón, lo que resulta en mortalidades persistentes y en la reducción del rendimiento de producción hasta en un 60% [3, 4].
La histopatología del camarón afectado por SHB también revela lesiones notables en el hepatopáncreas. Wang et al. [5] describieron afectación histológica de origen natural en el camarón blanco del Pacífico Penaeus vannamei, que se puede dividir en tres fases. Durante la Fase I, la pulpa y la apariencia suculenta del hepatopáncreas fueron visibles, lo que puede haber sido causado por la expansión de los túbulos hepatopancreáticos, mientras que las células B, R y F todavía estaban intactas.
Luego, el grosor del epitelio tubular se redujo en tamaño y el lumen se agrandó relativamente, a medida que la enfermedad progresó en la Fase II, acompañada de una infiltración grave de hemocitos en la parte proximal del seno. Además, se observó una reducción de células F y células R durante la Fase II. En la Fase III, las células B, F y R desaparecieron y la mayoría de los túbulos colapsaron, mostrando un epitelio adelgazado y un lumen grande.
La capa de microvellosidades desprendida se agregó en el lumen de algunos túbulos. El epitelio tubular hepatopancreático desprendido luego se agregó para formar un cuerpo vermiforme y alguna vez se entendió mal como el parásito gregarino del camarón, pero luego se reconoció como
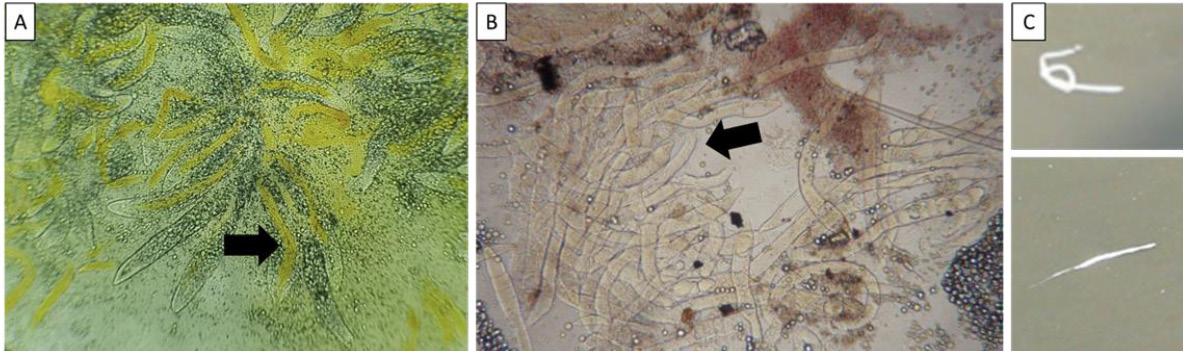
microvellosidades agregadas y transformadas (ATM) [2, 6]. Cuando la ATM se acumula en una cantidad muy alta combinada con composiciones variadas, como lípidos y componentes bacterianos mixtos, dan como resultado hilos fecales blancos [ 2 , 7 ]. Recientemente, las observaciones de la ATM bajo microscopía óptica de preparaciones de calabaza (Figura 1) o frotis teñidos de tejido HP fresco se han utilizado como indicadores para el monitoreo de SHB en estanques de camarones [8]. Además, se han informado grupos de microsporidios Enterocytozoon hepatopenaei (EHP), un parásito intracelular que causa el síndrome de crecimiento lento en camarones, adheridos a hilos fecales blancos [9, 10].
Hasta la fecha, la etiología de SHB se ha propuesto como la combinación de EHP con patógenos marinos oportunistas, incluyendo Vibrio spp. y Propionigenium sp. [1, 11], pero el desafío experimental utilizando el patógeno individual no puede cumplir el postulado de Koch. Además, se ha introducido el concepto de disbiosis intestinal (el desequilibrio de la microflora intestinal); sin embargo, es difícil discernir si la disbiosis ocurrió antes o después de la aparición de SHB.
La disbiosis intestinal puede estar influenciada por varias causas no infecciosas, como la alteración de la nutrición de una dieta y la calidad del agua [3], pero para inducir la formación de heces blancas, no podemos descuidar la participación de patógenos particulares, en particular EHP [1]. Muchos investigadores han discutido la participación de EHP en SHB. En conjunto, se plantea la
hipótesis de que esta es una enfermedad multifactorial, y se ha introducido el concepto de "patobioma". Esta revisión resume la información general y actualizada sobre SHB, la investigación de la participación de bacterias y EHP, el microbioma de los camarones afectados por el síndrome y las estrategias de tratamiento o control que se han propuesto.
Parámetros del agua y SHB
Hay pocos informes sobre la calidad del agua relacionada con la aparición del síndrome de las Heces Blancas. Se informaron casos de SHB en entornos de baja (<5 ppt) y alta salinidad (>30 ppt) [5, 12]. La mortalidad después de la incidencia de este trastorno en el camarón tigre negro Penaeus monodon se observó cuando se informaron niveles bajos de oxígeno disuelto (>3 mg/L) y baja alcalinidad (<80 ppm) en Tailandia [13].
Un estudio en Indonesia informó que los estanques afectados mostraron un pH del agua más bajo (7,71–7,84) y oxígeno disuelto (5,57–5,98 mg mL −1), en comparación con un pH del agua > 8 y oxígeno disuelto a >6 mg mL −1 en los estanques saludables [14]. Un pH del agua más bajo y un mayor desperdicio de materia orgánica promueven el crecimiento de bacterias heterotróficas, mientras que el deterioro de la calidad del agua también afecta la microbiota intestinal del camarón [14]. En general, la calidad del agua puede no tener un impacto directo en la aparición de SHB sino más bien un efecto colateral en la salud del camarón y las poblaciones microbianas del estanque.
Etiología del síndrome de SHB
Patógenos bacterianos asociados con el síndrome
Se han reportado infecciones bacterianas que causan exudados gastrointestinales blanquecinos o amarillentos no solo para SHB en camarones. La formación de excreción intestinal blanca en otras especies acuáticas también se encontró en el esturión siberiano Acipenser baerii y el barramundi
Lates calcarifer infectados con dosis letales bajas de Streptococcus iniae [15, 16]. Las materias gastrointestinales blancas fueron causadas por inflamación exudativa y producción excesiva de mucina inducida por las bacterias.
Estos informes muestran evidencia de participación bacteriana en la patología gastrointestinal de especies acuáticas. Para SHB de camarones, Vibrio, un patógeno oportunista marino, fue un punto de atención durante la investigación inicial. Vibrio spp. son bacterias anaeróbicas facultativas Gram-negativas de la familia Vibrionaceae, ubicuas en todo el mundo, de las cuales algunas de las Vibrio spp., como V. parahaemolyticus y V. harveyi , son los patógenos primarios en mariscos [17]. Vibrio spp. parece desempeñar un papel importante en la aparición de SHB, ya que se ha encontrado una carga significativamente mayor de Vibrio spp. en el agua del estanque, el intestino y la hemolinfa en los camarones afectados que en la población sana [8, 19]. El aislamiento bacteriano y la secuenciación del ARNr 16s del intestino de los camarones afectados por SHB cultivados en el delta del Mekong, Vietnam, revelaron cinco
Figura 1. Preparaciones de hepatopáncreas (A) y tejido intestinal (B) del camarón afectado por SHB observadas bajo un microscopio óptico, que muestran una acumulación de microvellosidades agregadas y transformadas (flecha). Hilos fecales blancos flotando en la superficie del agua del estanque afectado (C).
grupos dominantes de Vibrio, incluidos V. parahaemolyticus , V. haveryi , V. vulnificus , V. cholerae y V. alginolyticus [20]. Se informó un hallazgo similar en la India, donde V. parahaemolyticus y V. alginolyticus fueron las especies dominantes de Vibrio aisladas de los camarones enfermos [21]. Un estudio de estanques de engorde de P. monodon en Sri Lanka mostró una fuerte correlación entre el porcentaje de ocurrencia de SHB y el recuento total de Vibrio en el intestino, hepatopáncreas y hemolinfa, donde las especies dominantes de Vibrio fueron V. alginolyticus y V. fluvialis [ 22 ].
Se aisló una cepa de V. cholerae que contenía genes de virulencia (ompU, hlyA y toxR) de P. vannamei criado en agua dulce que exhibía SHB de China, y la formación de heces blancas también se reprodujo mediante desafío de inmersión utilizando esta cepa [12]. Sin embargo, este procedimiento nunca ha sido repetido por otros investigadores a pesar de que el SHB sigue siendo problemático en todo el mundo.
Es posible que el V. cholerae informado fuera una especie diferente de Vibrio, ya que el árbol filogenético de las especies de Vibrio basado en la secuencia del ARNr 16S es menos confiable [1]. La inducción de SHB en condiciones de laboratorio utilizando una bacteria individual nunca ha sido exitosa. Luego se propuso una teoría de la etiología del síndrome como una relación compleja entre más de un patógeno relacionado con la disbiosis intestinal del camarón.
Análisis del microbioma de los camarones afectados por el SHB
El análisis del microbioma proporciona detalles intensivos sobre las especies y la cantidad de microbios que residen en una determinada condición. Esta técnica nos ayuda a comprender los cambios en el microbioma intestinal que probablemente tengan un impacto en la salud del camarón. Una abundancia diferencial de taxones microbianos particulares o la reducción de la diversidad bacteriana dentro del intestino pueden estar relacionadas con el inicio de la patogénesis, ya que la microbiota intestinal y el sistema inmunológico del huésped están conectados [23]. Se ha sugerido que la disbiosis de la microbiota intestinal o el desequilibrio de la comunidad microbiana en
el tracto gastrointestinal están fuertemente asociados con la aparición de SHB.
El trasplante de microbiota intestinal a través de sonda inversa de donantes afectados indujo la formación de aguijón fecal blanco en el 36,7% de los camarones receptores sanos [3]. Aunque no se logró una tasa de replicación del 100%, este estudio proporcionó cierta evidencia de que la disbiosis intestinal estaba relacionada con SHB. Varios estudios han encontrado que el microbioma intestinal del camarón que sufre de SHB tiene baja α-diversidad y es menos homogéneo [24]. Además, los patógenos oportunistas, como Vibrio, Candidatus Bacilloplasma, Phascolarctobacterium, Photobacterium y Aeromonas estaban sobrerrepresentados en el camarón afectado, mientras que las bacterias beneficiosas del camarón, incluidas Shewanella, Chitinibacter, Rhodobacter, Paracoccus y Lactococcus eran más abundantes en camarones sanos [3 , 4 , 24]. Se informaron hallazgos comparables en un caso de enfermedad de Necrosis Hepatopancreática Aguda (AHPND), que mostró un aumento en la abundancia de Candidatus bacilloplasma junto con varios grupos de Vibrio [25].
Sin embargo, la abundancia relativa de C. bacilloplasma puede haber contribuido a la reducción de otros taxones en el intestino del camarón, ya que es una bacteria comensal que se encuentra comúnmente en varios invertebrados acuáticos, y este género está bien adaptado al ambiente intestinal [23, 26]. Se realizó un análisis metagenómico en P. vannamei afectado que fue criado en estanques de invernadero en China. V. tubiashii y V. harveyi se identificaron a partir del análisis de la unidad taxonómica operativa, y se encontró la expresión de vías inmunes innatas relacionadas con la infección bacteriana (receptores tipo Toll (TLR) y receptores tipo dominio de oligomerización de nucleótidos (NLR)), lo que sugiere la participación bacteriana durante la progresión de la enfermedad [27, 28].
Además, los autores también investigaron las interacciones depredador-presa intestinales y resumieron que la población de fagótrofos intestinales y Bdellovibrio y organismos similares (BALO; comensales intestinales del camarón que depredan
REFERENCIAS BIBLIOGRÁFICAS:
Aranguren Caro, LF; Mai, HN; Cruz-Florez, R.; Marcos, FLA; Alenton, RRR; Dhar, AK Reproducción experimental del síndrome de Heces Blancas en el camarón patiblanco, Penaeus vannamei PLoS ONE 2021 16 , e0261289. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Prachumwat, A.; Munkongwongsiri, N.; Eamsaard, W.; Lertsiri, K.; Flegel, TW; Stentiford, GD; Sritunyalucksana, K. Un posible patobioma procariota y microsporídico que puede causar el síndrome de Heces Blancas del camarón (SHB). bioRxiv 2021 [ Google Scholar ] [ CrossRef ] Huang, Z.; Zeng, S.; Xiong, J.; Hou, D.; Zhou, R.; Xing, C.; Wei, D.; Deng, X.; Yu, L.; Wang, H. Los postulados microecológicos de Koch revelan que la disbiosis de la microbiota intestinal contribuye al síndrome de Heces Blancas del camarón. Microbiome 2020 , 8 , 32. [ Google Scholar ] [ CrossRef ] [ PubMed ] [ Green Version ]
Durai, V.; Gunalan, B.; Johnson, PM; Maheswaran, M.; Pravinkumar, M. Efecto sobre la enfermedad del intestino blanco y las Heces Blancas en el sistema de cultivo semiintensivo de camarones Penaeus vannamei en el estado de Tamilnadu, en el sur de la India. Int. J. Mar. Sci. 2015 5 , 1–5. [ Google Scholar ] [ CrossRef ] Wang, H.; Wan, X.; Xie, G.; Dong, X.; Wang, X.; Huang, J. Perspectivas sobre la histopatología y el microbioma del camarón blanco del Pacífico, Penaeus vannamei, que padece el síndrome de las Heces Blancas. Acuicultura 2020 , 527 , 735447. [ Google Scholar ] [ CrossRef ] El síndrome de Heces Blancas del camarón surge de la transformación, desprendimiento y agregación de las microvellosidades hepatopancreáticas en cuerpos vermiformes que superficialmente se parecen a las gregarinas. PLoS ONE 2014 9 , e99170. [ Google Scholar ] [ CrossRef ] [ Green Version ] Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, PV; Sritunyalucksana, K.; Flegel, TW; Itsathitphaisarn, O. Revisión de las amenazas actuales de enfermedades para los camarones peneidos cultivados en Asia. Acuicultura 2016 452 , 69–87. [ Google Scholar ] [ CrossRef ] Vinod, B.; Haq, MB; Prathap, T.; Meetei, K.; Baharlooeian, M.; Sureandiran, B.; Tiwary, C. Vigilancia del síndrome de Heces Blancas (SHB) en camarones peneidos capturados en el grupo de islas Tuticorin cerca de un área de actividad de acuicultura semi-intensiva. JICR 2020 12 , 2051–2071. [ Google Scholar ] Tang, KF; Pantoja, CR; Redman, RM; Han, JE; Tran, LH; Lightner, DV Desarrollo de ensayos de hibridación in situ y PCR para la detección de Enterocytozoon hepatopenaei (EHP), un parásito microsporidio que infecta a los camarones peneidos. J. Invertebr. Pathol. 2015 130 , 37–41. [ Google Scholar ] [ CrossRef ] Tang, KF; Han, JE; Aranguren, LF; White-Noble, B.; Schmidt, MM; Piamsomboon, P.; Risdiana, E.; Hanggono, B. Poblaciones densas del microsporidio Enterocytozoon hepatopenaei (EHP) en heces de Penaeus vannamei que exhiben el síndrome de Heces Blancas y vías de su transmisión a camarones sanos. J. Invertebr. Pathol. 2016 , 140 , 1–7. [ Google Scholar ] [ CrossRef ] [ PubMed ] [ Versión verde ] Munkongwongsiri, N.; Prachumwat, A.; Eamsaard, W.; Lertsiri, K.; Flegel, TW; Stentiford, GD; Sritunyalucksana, K. Especies de Propionigenium y Vibrio identificadas como posibles causas componentes del síndrome de Heces Blancas del camarón (SHB) asociado con el microsporidio Enterocytozoon hepatopenaei J. Invertebr. Pathol. 2022 , 192 , 107784. [ Google Scholar ] [ CrossRef ] Cao, H.; Wen, L.; He, S.; Lu, L.; Yang, X.; Chen, B. Vibrio cholerae : un agente causal del síndrome de Heces Blancas en camarones patiblancos cultivados en agua dulce (Penaeus vannamei). Isr. J. Aquac. 2015 , 67 , 1–7. [ Google Académico ] Limsuwan, C. Enfermedad de las Heces Blancas en Tailandia. Revista Buletines Nicovita, Lima , abril-junio de 2012; 2–4. [ Google Scholar ] Alfiansah, YR; Peters, S.; Harder, J.; Hassenrück, C.; Gärdes, A. Estructura y patrones de coocurrencia de comunidades bacterianas asociadas con brotes de enfermedad de Heces Blancas en la acuicultura del camarón patiblanco del Pacífico Penaeus vannamei Sci. Rep. 2020 , 10 , 11980. [ Google Scholar ] [ CrossRef ] Chen, D.; Peng, S.; Chen, D.; Yang, F.; Liu, J.; Wang, J.; Liu, Q.; Huang, X.; Ouyang, P.; Wang, K. Dosis letales bajas de Streptococcus iniae causaron enteritis en el esturión siberiano ( Acipenser baerii ). Fish Shellfish Immunol. 2020 , 104 , 654–662. [ Google Scholar ] [ CrossRef ] [ PubMed ] Piamsomboon, P.; Tanpichai, P.; Wongtavatchai, J. Enteritis asociada con infección subclínica de Streptococcus iniae en juveniles de lubina asiática Lates calcarifer (Bloch, 1790). J. Fish Dis. 2021 , 44 , 1879–1882. [ Google Scholar ] [ CrossRef ] Dash, P.; Avunje, S.; Tandel, RS; Sandeep, KP; Panigrahi, A. Biocontrol de la vibriosis luminosa en la acuicultura del camarón: una revisión de los enfoques actuales y las perspectivas futuras. Rev. Fish. Sci. Aquac. 2017 , 25 , 245–255. [ Google Scholar ] [ CrossRef ]
bacterias Gram-negativas, especialmente Vibrio spp.) se redujeron significativamente en los camarones afectados por SHB [27].
En Indonesia, se evaluó la asociación entre las comunidades bacterianas en camarones (o estanques) afectados por SHB y el deterioro de la calidad del agua (disminución del pH, oxígeno disuelto y aumento de materia orgánica). En las granjas afectadas por este síndrome, el análisis del microbioma mostró similitudes en las comunidades bacterianas de las heces blancas y las fracciones de agua de los estanques afectados [14].
En el agua del estanque afectado, Alteromonas, Marinomonas, Photobacterium, Pseudoalteromonas y Vibrio fueron las poblaciones dominantes. Por el contrario, Acinetobacter y Pseudomonas fueron dominantes en camarones sanos, y las bacterias relacionadas con el proceso de nitrificación, incluidas Exiguobacterium, Halomonas, Psychrobacter, Salegentibacter y Sulfitobacte dominaron las comunidades bacterianas en el agua del estanque de individuos sanos [14].
Además del microbioma bacteriano, se han estudiado comunidades de eucariotas intestinales en granjas de camarones en China. La diversidad de los eucariotas intestinales también disminuyó, mientras que el número de eucariotas patógenos aumentó. Los taxones de parásitos intestinales, incluidos Pseudoperkinsidae, Prostapa parásito, Malassezia sp. y Acineta sp. están asociados positivamente con la exacerbación de la enfermedad [29]. Otro estudio sobre P. vannamei afectado por SHB mostró que los patógenos oportunistas Basidiomycota y Ascomycota (afiliados a hongos) estaban sobrerrepresentados, mientras que Desmodesmus, los comensales comunes en individuos sanos, estaban subrepresentados en los camarones afectados.
Los autores sugirieron que los camarones con SHB más débiles pueden tener menos capacidad para excluir a estos invasores externos. Alternativamente, la abundancia de estos patógenos oportunistas puede deberse al agua más eutrófica encontrada en los estanques afectados [30].
Estos estudios sugieren que la inspección de
SHB es posible observando cambios en la población eucariota intestinal.
En esencia, varios estudios del microbioma han indicado que la reducción de la diversidad bacteriana y el predominio de patógenos oportunistas en el intestino o en el agua de cultivo son características significativas de la SHB.
Los análisis del metagenoma y del metaboloma sugieren que las alteraciones en las comunidades microbianas afectan las vías funcionales y los perfiles metabólicos, lo que puede contribuir a la enfermedad [3]. Sin embargo, el aumento de microbios intestinales particulares, como Candidatus bacilloplasma o Bdellovibrio debido a la condición desequilibrada del entorno intestinal puede reflejar solo el estado no saludable de la salud intestinal en lugar de una indicación específica de SHB. Aunque se ha propuesto la hipótesis de la disbiosis intestinal, es difícil diferenciar entre la causa y el efecto de un brote porque faltan estudios de seguimiento que involucren la suplementación intestinal y/o organismos gnotobióticos [23]. Además el EHP, un patógeno importante asociado con SHB, no se probó en estos estudios.
Asociación entre EHP y vibriosis en la aparición de SHB
Cuando se investigó por primera vez el EHP en camarones cultivados, se sugirió que el microsporidio EHP era la única causa de SHB. Poco después, esta hipótesis no fue completamente aceptada porque también se detectó una alta prevalencia de EHP por PCR e hibridación in situ en muestras de camarones que no presentaban el síndrome [31]. Además, un desafío experimental con una dosis alta de EHP no produjo los signos clínicos que representan SHB [32]. Sin embargo, más evidencia apoya la asociación entre SHB y EHP.
De las granjas indonesias, P. vannamei que exhibía el síndrome presentó esporas de EHP empaquetadas en sus bacterias de heces, intestino medio y forma de bastón, presumiblemente Vibrio acompañadas de Necrosis Hepatopancreática Séptica (SHPN), que se observaron histológicamente en los túbulos HP [10]. En las granjas vietnamitas, P. monodon exhibió SHB
que se manifestó con EHP [33]. Además, todas las muestras intestinales de los camarones afectados por SHB recolectadas en India y Vietnam fueron EHP-positivas, y se detectaron esporas dentro y alrededor de las microvellosidades transformadas, mientras que aproximadamente el 90% de los camarones con intestino normal de las granjas afectadas por SHB fueron EHPpositivas [34]. Caro et al. [35] encontraron que todos los camarones afectados por SHB de dos ubicaciones geográficas diferentes (Indonesia y Venezuela) fueron EHP-positivas por PCR en tiempo real e histología y que las cargas de EHP fueron significativamente más altas en los estanques afectados que en los estanques sin SHB. Esto implica que SHB está asociado con un efecto sinérgico entre la infección por EHP y otros patógenos. Es posible que EHP debilite al camarón por la alteración de las células hepatopancreáticas infectadas.
Las células desprendidas y la membrana basal expuesta son favorables para la infección posterior de patógenos oportunistas, especialmente bacterias Vibrio ya presentes en el tracto gastrointestinal [36]. Por lo tanto, se introdujo el concepto de “patobioma”. Bass et al. [37] sugirieron que la patogénesis de una enfermedad puede no estar vinculada únicamente a la abundancia relativa de un taxón particular, sino más bien al cambio en las interacciones entre múltiples taxones, simbiontes asociados al huésped y/o el huésped mismo.
La complejidad del patobioma procariotaeucariota se ha sugerido como una causa de SHB en un estudio de P. vannamei cultivado en Tailandia. Se investigaron las poblaciones infectadas con EHP presentes o ausentes del SHB. La cantidad de EHP fue mucho mayor en el grupo SHB, y el análisis del microbioma reveló que las bacterias de los géneros Vibrio y Propionigenium eran predominantes en el tracto intestinal de los camarones afectados por EHP-SHB, mientras que el grupo sin SHB tenía una mayor diversidad bacteriana [2].
La SHB no se puede reproducir en el laboratorio utilizando un único patógeno, como EHP, pero puede estar asociada con múltiples patógenos, incluidas las especies de Vibrio. Un estudio desafiante con cepas específicas de V. sinaloensis o V.
parahaemolyticus en P. vannamei mediante alimentación por sonda inversa produjo lesiones histopatológicas del hepatopáncreas comparables a las SHB naturales, pero no logró inducir heces blancas [4]. Sin embargo, se indujeron heces blancas alimentando camarones con HP de camarones enfermos [34] o mediante trasplante de microbioma intestinal de donantes de SHB [3]. Además, los efectos sinérgicos de EHP y vibriosis fueron propuestos por Caro et al. [1], que indujeron la formación de heces blancas en camarones infectados con EHP producidos en laboratorio y desafiados con una cepa específica de V. parahaemolyticus aislada de camarones afectados por SHB.
Estrategias de control
Hasta la fecha, no se encuentran disponibles medicamentos o tratamientos específicos para SHB. El manejo se enfoca principalmente en implementar buenas prácticas de acuicultura para reducir el riesgo de ocurrencia de SHB, que son el uso de semillas no saludables, mala calidad del agua, disbiosis intestinal, infección por EHP y una alta carga de Vibrio en los tractos gastrointestinales de los camarones y el medio ambiente [2, 38]. En términos de exclusión de patógenos, las medidas preventivas para SHB apuntan a eliminar las esporas de EHP y limitar el número total de Vibrio en el sistema de cultivo. Se recomienda la combinación de control físico, químico y biológico para el manejo de SHB.
Control físico
Los métodos físicos implican la manipulación de las condiciones fisiológicas para eliminar o inactivar el agente patógeno y el uso de barreras físicas para evitar el contacto entre el agente patógeno y el huésped. Se recomiendan prácticas estándar, como el secado del estanque antes de la siembra y la eliminación manual de los sedimentos del estanque durante el cultivo para mejorar la calidad del agua y minimizar el crecimiento del patógeno.
Se necesitan medidas de bioseguridad estrictas para EHP, ya que no hay un fármaco terapéutico eficaz disponible y la prevención puede ser la única opción [39]. Las postlarvas (PL) negativas a EHP por PCR deben confirmarse antes de la siembra de camarones. También se ha informado que
el alimento fresco o vivo, por ejemplo, los poliquetos de los estanques afectados por SHB, son positivos a EHP por PCR [40]. El alimento fresco se congeló a -20 °C durante al menos 48 h o a 70 °C durante 15 min [39]. Munkongwongsiri et al. [41] afirmaron que las esporas de EHP son muy sensibles a la inactivación por calor a 75 °C durante un minuto.
Control químico y biológico
El objetivo general de los enfoques químicos y biológicos para prevenir SHB es limitar el número de patógenos, especialmente bacterias EHP y Vibrio en el sistema de cultivo. El uso de sustancias inorgánicas y orgánicas para reducir el número de esporas de EHP y la población de Vibrio en estanques o camarones se ha implementado ampliamente.
También se recomienda agregar cal viva (CaO) para aumentar el pH ambiental a 9, para inducir la germinación de esporas de EHP, y aplicar cloro o permanganato de potasio (KMnO 4) para inactivar las esporas antes del stock de camarones [42]. Una vez que se excluye EHP del sistema, Supono et al. [19] sugirieron que el recuento total de Vibrio en el agua de cultivo no debe exceder 10 4 UFC mL −1 para reducir el riesgo de ocurrencia de SHB. Varios estudios han mostrado resultados prometedores en el control de Vibrio spp. aislados de camarones afectados por SHB.
Los camarones que sufrieron SHB se recuperaron sin recurrencia hasta la cosecha, después de recibir lisozima extraída derivada de clara de huevo durante cinco días. Esta enzima inhibe el crecimiento de Vibrio spp. tanto in vitro como in vivo y ayuda a estimular la respuesta inmune del camarón [43]. La inmunización pasiva mediante la alimentación con inmunoglobulina Y (IgY) derivada de la yema de huevo puede reducir significativamente las cantidades de V. harveyi y V. parahaemolyticus en el camarón [44]. Posteriormente, Keetanon et al. [45] demostraron la implementación de IgY de yema de huevo para inhibir V. parahaemolyticus aislado del camarón afectado por SHB.
La gravedad del desprendimiento del hepatopáncreas después del desafío con
Somboon, M.; Purivirojkul, W.; Limsuwan, C.; Chuchird, N. Efecto de Vibrio spp. en camarones infectados con heces blancas en Chanthaburi, Tailandia. J. Fish. Environ. 2012 , 36 , 7–15. [ Google Scholar ] Supono, S.; Wardiyanto, W.; Harpeni, E. Identificación de Vibrio sp. como causa de enfermedades de Heces Blancas en el camarón blanco Penaeus vannamei y manipulación con ingredientes a base de hierbas en la regencia de East Lampung, Indonesia. AACL Bioflux 2019 , 12 , 417–425. [ Google Scholar ] Actividad antimicrobiana de extractos de hierbas contra bacterias Vibrio spp. aisladas del síndrome de las Heces Blancas en el camarón de patas blancas (Penaeus vannamei ) en algunas provincias del delta del Mekong. CTUJS 2021 , 13 , 61–68. [ Google Scholar ] [ CrossRef ] Mastan, S. Incidencias del síndrome de Heces Blancas (SHB) en camarones criados en granjas, Penaeus vannamei , Andhra Pradesh. IAJPS 2015 , 5 , 3044–3047. [ Google Scholar ] Kumara, K.; Hettiarachchi, M. Síndrome de Heces Blancas causado por Vibrio alginolyticus y Vibrio fluvialis en el camarón, Penaeus monodon (Fabricius 1798): estrategia multimodal para controlar el síndrome en estanques de engorde de Sri Lanka. Asian Fish Sci. 2017 , 30 , 245–261. [ Google Académico ] Holt, CC; Bass, D.; Stentiford, GD; van der Giezen, M. Entendiendo el papel del microbioma intestinal del camarón en la salud y la enfermedad. J. Invertebr. Pathol. 2021 , 186 , 107387. [ Google Scholar ] [ CrossRef ] [ PubMed ] Hou, D.; Huang, Z.; Zeng, S.; Liu, J.; Wei, D.; Deng, X.; Weng, S.; Yan, Q.; He, J. Firmas bacterianas intestinales del síndrome de Heces Blancas en camarones. Appl. Microbiol. Biotechnol. 2018 , 102 , 3701–3709. [ Google Scholar ] [ CrossRef ] [ PubMed ] Chen, W.-Y.; Ng, TH; Wu, J.-H.; Chen, J.-W.; Wang, H.-C. Dinámica del microbioma en un estanque de engorde de camarones con posible brote de enfermedad de necrosis hepatopancreática aguda. Sci. Rep. 2017 , 7 , 9395. [ Google Scholar ] [ CrossRef ] [ PubMed ] [ Versión verde ]
Kostanjsek, R.; Strus, J.; Avgustin, G. Candidatus Bacilloplasma,” un nuevo linaje de Mollicutes asociado con la pared del intestino posterior del isópodo terrestre Porcellio scaber (Crustacea: Isopoda). Appl. Environ. Microbiol. 2007 , 73 , 5566–5573. [ Google Scholar ] [ CrossRef ] [ PubMed ] [ Green Version ] Lu, J.; Li, X.; Qiu, Q.; Chen, J.; Xiong, J. Las interacciones depredador-presa entre reinos intestinales son determinantes clave de la salud del camarón. Acuicultura 2022 546 737304. [ Google Scholar ] [ CrossRef ]
Dai, W.; Yu, W.; Xuan, L.; Tao, Z.; Xiong, J. Integración de enfoques moleculares y ecológicos para identificar posibles patógenos polimicrobianos a lo largo de la progresión de una enfermedad del camarón. Appl. Microbiol. Biotechnol. 2018 102 , 3755–3764. [ Google Scholar ] [ CrossRef ] Xiong, J.; Yu, W.; Dai, W.; Zhang, J.; Qiu, Q.; Ou, C. Predicción cuantitativa de la incidencia de enfermedades del camarón a través de los perfiles de la microbiota intestinal eucariota. Appl. Microbiol. Biotechnol. 2018 , 102 , 3315–3326. [ Google Scholar ] [ CrossRef ]
Dai, W.; Qiu, Q.; Chen, J.; Xiong, J. Los taxones que discriminan enfermedades en los eucariotas intestinales son indicativos del síndrome de Heces Blancas del camarón blanco del Pacífico (Penaeus vannamei ). Acuicultura 2019 , 506 , 154–160. [ Google Scholar ] [ CrossRef ] Tangprasittipap, A.; Srisala, J.; Chouwdee, S.; Somboon, M.; Chuchird, N.; Limsuwan, C.; Srisuvan, T.; Flegel, TW; Sritunyalucksana, K. El microsporidio Enterocytozoon hepatopenaei no es la causa del síndrome de Heces Blancas en el camarón patiblanco Penaeus vannamei BMC Vet. Res. 2013 9 , 139. [ Google Scholar ] [ CrossRef ] [ Versión verde ]
Sritunyalucksana, K. Actualización de la investigación sobre patógenos emergentes del camarón en Tailandia. Actas del Taller internacional sobre la promoción de la acuicultura sostenible, la salud de los animales acuáticos y la mejora de los recursos en el sudeste asiático, Iloilo City, Filipinas, 25-27 de junio de 2019 ; Centro de Desarrollo Pesquero del Sudeste Asiático, Departamento de Acuicultura: Tigbauan, Filipinas, 2021; pág. 217. [ Google Académico ] Ha, N.; Ha, D.; Thuy, N.; Lien, V. Se ha detectado la parasitación de Enterocytozoon hepatopenaei en camarones tigre (Penaeus monodon) infectados mediante cultivo de heces blancas en Vietnam. Agric. Rural Dev. Sci. Technol. 2010 , 12 , 45–50. [ Google Scholar ] Kumar, TS; Makesh, M.; Alavandi, S.; Vijayan, K. Manifestaciones clínicas del síndrome de Heces Blancas (SHB) y su asociación con Enterocytozoon hepatopenaei en granjas de engorde de Penaeus vannamei : una investigación patobiológica. Acuicultura 2022 , 547 , 737463. [ Google Scholar ] [ CrossRef ] Caro, LFA; Mai, HN; Pichardo, O.; Cruz-Flores, R.; Hanggono, B.; Dhar, AK Evidencias que apoyan la asociación de Enterocytozoon hepatopenaei con el síndrome de Heces Blancas en Penaeus vannamei de granja en Venezuela e Indonesia. Dis. Aquat. Organ. 2020 , 141 , 71–78. [ Google Scholar ] [ CrossRef ] [ PubMed ]
V. parahaemolyticus estaba inversamente relacionada con la dosis de administración de IgY [45]. También se sugiere el uso de probióticos de Bacillus subtilis para reducir el riesgo de SHB [38]. Un estudio en la India reveló que la alimentación con probióticos comerciales que contienen Bacillus productor de ácido láctico, enzimas digestivas y ácidos orgánicos (Gut Probiotics, V Sthiraa Bioscience, India) a 5 g kg −1 ayudó al camarón a recuperarse de SHB [46].
Varios extractos de plantas también han mostrado efectos inhibidores contra Vibrio spp. aislado de camarones afectados por SHB. P. vannamei alimentado con extracto de galanga Alpinia galanga Linn. mostró un recuento total más bajo de Vibrio y una incidencia más baja de EHP encontrados en los tractos intestinales, lo que podría ayudar a reducir la incidencia de SHB en condiciones de cultivo [47]. Hoja flor Phyllanthus urinaria L. muestra actividad antibacteriana contra V. harveyi y V. alginolyticus aislados de camarones afectados por SHB [20]. Palanikumar, et al. [48] evaluaron la posibilidad de las medicinas a base de hierbas, incluido el limón Citrus limon, el
ajo Allium sativum, el jengibre Zingiber officinale, la palma doble Borassus flabellifer y el gramo negro Vigna mungo para reducir la infestación de EHP en camarones de cultivo y eventualmente tratar SHB. Se ha informado que los compuestos activos, como alcaloides, flavonoides, pigmentos, fenólicos, terpenoides, taninos y glucósidos que se encuentran en estas plantas tienen un efecto hepatoprotector, promueven el crecimiento, tienen capacidad antioxidante y tienen la capacidad de eliminar el parásito al interferir con su metabolismo. Sin embargo, estas medicinas a base de hierbas no se han evaluado específicamente en condiciones experimentales para el tratamiento de SHB.
En Tailandia, la estrategia para combatir el SHB generalmente se inicia con la interrupción de la alimentación durante unos días y el cambio del tipo de alimento a una fuente de proteínas de mayor calidad. Se pueden aplicar ácidos orgánicos, seguidos de la suplementación con extractos de hierbas, como galanga o extractos de curcumina para reducir la cantidad de EHP y Vibrio Posteriormente, se administran probióticos, como Bacillus o Lactobacillus para colonizar
Aranguren, LF; Han, JE; Tang, KF Enterocytozoon hepatopenaei (EHP) es un factor de riesgo para la enfermedad de Necrosis Hepatopancreática Aguda (AHPND) y la Necrosis Hepatopancreática Séptica (SHPN) en el camarón blanco del Pacífico Penaeus vannamei Acuicultura 2017 , 471 , 37–42. [ Google Scholar ] [ CrossRef ] [ Versión verde ] Bass, D.; Stentiford, G. D.; Wang, H.-C.; Koskella, B.; Tyler, CR El patobioma en enfermedades animales y vegetales. Trends Ecol. Evol. 2019 , 34 , 996–1008. [ Google Scholar ] [ CrossRef ] [ Versión verde ] Kurniasih, RA; Suwiryono, J.; Penataseputro, T. Síndrome de Heces Blancas. Volumen II. Enfermedades de los crustáceos y moluscos. En Fisiopatología de la acuicultura ; Kibenge, FSB, Baldisserotto, B., Chong, RS, Eds.; Academic Press: Cambridge, MA, EE. UU., 2022; págs. 235–242. ISBN 978-0-323-95434-1. [ Google Académico ] Chaijarasphong, T.; Munkongwongsiri, N.; Stentiford, G. D.; Aldama-Cano, DJ; Thansa, K.; Flegel, T. W.; Sritunyalucksana, K.; Itsathitphaisarn, O. El microsporidio del camarón Enterocytozoon hepatopenaei (EHP): biología, patología, diagnóstico y control. J. Invertebr. Pathol. 2021 , 186 , 107458. [ Google Scholar ] [ CrossRef ] Desrina, D.; Prayitno, SB; Haditomo, AHC; Latritiani, R.; Sarjito, S. Detección de ADN de Enterocytozoon hepatopenaei (EHP) en poliquetos de estanques de camarones que sufren brotes de síndrome de Heces Blancas. Biodiversitas 2020 , 21 , 369–374. [ Google Scholar ] [ CrossRef ] Munkongwongsiri, N.; Aldama-Cano, DJ; Suebsing, R.; Thaiue, D.; Prasartset, T.; Itsathitphaisarn, O.; Sritunyalucksana, K. Las esporas del microsporidio Enterocytozoon hepatopenaei (EHP) se inactivan en 1 minuto a 75 °C. Aquaculture 2021 , 533 , 736178. [ Google Scholar ] [ CrossRef ] Aldama-Cano, DJ; Sanguanrut, P.; Munkongwongsiri, N.; IbarraGámez, JC; Itsathitphaisarn, O.; Vanichviriyakit, R.; Flegel, TW; Sritunyalucksana, K.; Thitamadee, S. Bioensayo para la extrusión de tubos polares de esporas de camarón Enterocytozoon hepatopenaei (EHP). Acuicultura 2018 , 490 , 156–161. [ Google Scholar ] [ CrossRef ] Woraprayote, W.; Pumpuang, L.; Tepaamorndech, S.; Sritunyalucksana, K.; Phromson, M.; Jangsutthivorawat, W.; Jeamsripong, S.; Visessanguan, W. Supresión del síndrome de Heces Blancas en el camarón blanco del Pacífico, Penaeus vannamei , utilizando lisozima de clara de huevo de gallina. Acuicultura 2020 , 521 , 735025. [ Google Scholar ] [ CrossRef ] Gao, X.; Zhang, X.; Lin, L.; Yao, D.; Sun, J.; Du, X.; Li, X.; Zhang, Y. Inmunoprotección pasiva de Penaeus vannamei contra infecciones por Vibrio harveyi y Vibrio parahaemolyticus con alimento encapsulado con yema de huevo anti-Vibrio (IgY). Int. J. Mol. Sci. 2016 , 17 , 723. [ Google Scholar ] [ CrossRef ] [ PubMed ] [ Green Version ] Keetanon, A.; Chuchird, N. Efectos del anticuerpo IgY en el crecimiento, la supervivencia, las respuestas inmunitarias y la prevención de la infección por el virus del síndrome de las Heces Blancas y del síndrome de la Mancha Blanca causado por Vibrio parahaemolyticus en el camarón blanco del Pacífico en condiciones de laboratorio; Universidad de Kasetsart: Bangkok, Tailandia, 2020. [ Google Académico ] Pravinkumar, M.; Ponraj, JG; Ravi, AV Eficacia de la evaluación de probióticos intestinales contra la enfermedad del Intestino Blanco y las Heces Blancas en sistemas de acuicultura de camarones Litopenaeus vannamei en dos regiones geográficas diferentes de Andhra Pradesh. Int. J. Mar. Sci. 2020 10 1–12. [ Google Académico ] Chaweepack, T.; Muenthaisong, B.; Chaweepack, S.; Kamei, K. El potencial del extracto de galanga (Alpinia galanga Linn.) contra los patógenos que causan el síndrome de Heces Blancas y la enfermedad de Necrosis Hepatopancreática Aguda (AHPND) en el camarón blanco del Pacífico (Penaeus vannamei ). Int. J. Biol. 2015 , 7 , 8. [ Google Scholar ] [ CrossRef ] [ Versión verde ] Palanikumar, P.; Wahjuningrum, D.; Abinaya, P.; Babu, MM; Citarasu, T. Uso de productos naturales de origen vegetal para la prevención y el control del síndrome de las Heces Blancas (SHB) en el cultivo de camarón patiblanco del Pacífico Penaeus vannamei en la India. Aquac. Int. 2020 , 28 , 113–125. [ Google Scholar ] [ CrossRef ] Chaweepack, T. El uso de extracto de galanga en combinación con ácido orgánico reduce eficazmente la cantidad de Vibrio y la presencia de ATM en camarones cultivados ; Informe de investigación de la Agencia de Desarrollo de Investigación Agrícola (Organización pública); Agencia de Desarrollo de Investigación Agrícola: Bangkok, Tailandia, 2015. [ Google Scholar ]
y competir con los patógenos bacterianos que residen en el intestino.
Algunos granjeros también aplican tratamientos químicos para reducir la cantidad total de bacterias en la columna de agua [49]. Sin embargo, no existe una fórmula fija para este trastorno multifactorial en los cultivos de camarones.
Solo una bioseguridad estricta y una buena acuicultura, como proporcionar alimentos de buena calidad, un buen manejo de la alimentación y mantener una buena calidad del agua durante el cultivo, pueden reducir el riesgo de aparición de SHB.
La verificación rutinaria de la presencia de ATM en el tracto intestinal del camarón puede ayudar en el diagnóstico temprano, lo que lleva a un manejo rápido. Otros suplementos, como enzimas, probióticos o medicamentos
a base de hierbas, son opciones que no han garantizado completamente la curación del SHB.
Conclusiones
El SHB es un trastorno gastrointestinal del camarón, asociado con Enterocytozoon hepatopenaei (EHP) y otras bacterias, como Vibrio y Propionigenium
Se sugiere que la patogenia es que EHP induce la alteración de los túbulos hepatopancreáticos, y las células desprendidas que se mueven a través del tracto gastrointestinal ayudan al crecimiento de bacterias oportunistas. El cambio en la comunidad microbiana puede resultar en disbiosis intestinal, trastorno metabólico y, finalmente, formación de heces blancas.
En consecuencia, los camarones infectados muestran un consumo de alimento y una
tasa de crecimiento reducidos, junto con decoloración hepatopancreática.
Aunque la enfermedad conduce a graves pérdidas económicas y déficit de producción en varios países de cultivo de camarón, no se han encontrado tratamientos ideales y métodos de control para la SHB. Por lo tanto, se han sugerido sistemas de bioseguridad estrictos para eliminar las esporas de EHP y controlar el número total de Vibrio para la prevención de la SHB•
Para más información sobre este artículo, contactar a: jehan@knu.ac.kr
La diversificación ecológica revela rutas de aparición de patógenos en poblaciones endémicas de Vibrio vulnificus
La aparición de patógenos en humanos es uno de los temas de salud pública más preocupantes de los tiempos modernos (1–4). Según la Organización Mundial de la Salud, se han notificado más de 300 enfermedades infecciosas emergentes en el período de 1940 a 2004, una tendencia que ha continuado de manera constante con los recientes brotes de ébola en África occidental, cólera en Yemen y la pandemia mundial causada por COVID-19 (3–5). Si bien los enfoques moleculares clásicos han avanzado nuestra comprensión de la patogénesis bacteriana, hasta la fecha, las adaptaciones genéticas y los impulsores ecológicos que facilitan que cepas seleccionadas dentro de una especie emerjan como patógenos y colonicen con éxito al huésped humano siguen siendo poco comprendidos. Dada la magnitud y la complejidad de esta amenaza urgente, es fundamental desarrollar sistemas modelo organismales manejables y marcos teóricos que nos permitan diseccionar las adaptaciones moleculares y los factores ambientales que conducen a la aparición de dichos patógenos humanos.
Vibrio vulnificus, un patógeno humano emergente, es una de las principales causas de muertes no asociadas al cólera, Vibrio, a nivel mundial (6). A pesar de ser un habitante natural de aguas estuarinas, costeras y salobres (7), esta bacteria carnívora ha ganado particular notoriedad como uno de los patógenos que matan más rápido (8, 9). Los humanos generalmente se infectan con V. vulnificus a través de la ingestión de mariscos crudos contaminados o por exposición directa de heridas abiertas al agua de mar (6). Las infecciones por V. vulnificus a menudo resultan en septicemia fulminante con una tasa de mortalidad alarmante que excede el 50% (6, 10-13). La bacteria es particularmente letal en algunos huéspedes susceptibles, como pacientes inmunodeprimidos o aquellos con cirrosis hepática asociada al alcohol, diabetes mellitus o hemocromatosis (14). Los recuentos anuales de casos de infecciones por V. vulnificus han aumentado de manera constante durante los últimos 20 años en los Estados Unidos (15). Un aumento repentino de su distribución mundial durante las últimas tres décadas, en correlación con el cambio climático, ha provocado brotes de
enfermedades en regiones sin antecedentes de infecciones por V. vulnificus (16-18) Además, los modelos predicen que esta tendencia continuará, lo que dará como resultado una expansión constante de su área de distribución geográfica y el consiguiente aumento del riesgo de infecciones humanas (16, 19-21)
Con base en una serie de rasgos bioquímicos y fenotípicos, las cepas de V. vulnificus han sido históricamente clasificadas en tres Biotipos (BT): BT1, que se asocia principalmente con infecciones humanas (22, 23), BT2, que es principalmente patógeno para las anguilas (24, 25), y BT3, que está geográficamente restringido a Israel y posee características híbridas de BT1 y BT2 (26, 27). A diferencia de Vibrio cholerae, en el que todas las cepas capaces de causar cólera pertenecen a un solo clado, las comparaciones genómicas de V. vulnificus revelan un patrón más complejo en la distribución de sus cepas clínicas (28-30). Los análisis filogenómicos indican que la población de V. vulnificus está compuesta por cuatro grupos o clúster distintos (Clúster 1 a 4), que se superponen en gran medida con el sistema de clasificación clásico de BT (23, 26, 28, 31, 32). Nuestros análisis indican que los dos clúster más grandes, C1 y C2, exhiben una alta divergencia genómica y parecen estar especiándose (28), con cepas clínicas de BT1 pertenecientes predominantemente a C1 (22, 23), mientras que las cepas de C2 se asocian principalmente con BT2 (6, 24, 25). C3 es altamente clonal y se superpone completamente con BT3, y el raro C4 contiene solo cuatro cepas no clonales y pertenece a BT1 (28, 31). Curiosamente, a pesar de que los pacientes muestran síntomas clínicos conservados, las cepas clínicas C1 surgen de diferentes clados dentro del grupo, lo que sugiere eventos de emergencia independientes de este patógeno mortal (28, 31, 32). Hasta la fecha, los rasgos genéticos únicos que permiten que ciertas cepas C1 causen septicemia grave siguen siendo en su mayoría desconocidos, lo que plantea un riesgo desalentador para la salud pública, ya que dificulta nuestra capacidad para detectar V. vulnificus potencialmente patógeno (33).
Recientemente, utilizando una combinación de análisis bioinformáticos y fenotípicos
que examinaron más de 100 cepas de V. vulnificus, determinamos que V. vulnificus C1 parece estar asociado con un estilo de vida ecológico o ecotipo único (28). No obstante, hasta la fecha, los impulsores ecológicos que conducen a la aparición de V. vulnificus C1 clínico y sus rasgos patógenos siguen siendo poco conocidos. Para comenzar a desenredar las complejas interacciones in situ entre los genotipos y el medio ambiente que subyacen a la aparición de cepas clínicas, en este estudio, recientemente examinamos un gran estuario en el este de Florida, la laguna del río Indio (IRL), donde ocurren con frecuencia brotes de la enfermedad (7, 34). Encontramos poblaciones endémicas de V. vulnificus en el estuario y establecimos dos lugares de muestreo para estudiar la dinámica ambiental de esta bacteria en varios reservorios naturales como agua, sedimentos, ostras y cianobacterias. Curiosamente, los dos sitios de muestreo muestran diferencias importantes en la distribución de los grupos de V. vulnificus
Uno de ellos, Feller’s House (sitio A), parece estar significativamente enriquecido con cepas C1, mientras que en el segundo sitio de muestreo, Shepard Park (sitio B), recuperamos principalmente cepas de C2. Los análisis genómicos de estas cepas indican que, a pesar de estas diferencias importantes en la distribución, se producen altas tasas de recombinación, así como un intercambio frecuente de elementos genéticos móviles y factores de virulencia entre estas poblaciones de V. vulnificus. Los análisis de microdiversidad de estos genomas revelaron marcadores genómicos únicos entre las cepas C1 en forma de alelos asociados a la clínica (CAA) con un posible papel directo en la virulencia. Las cepas aisladas de V. vulnificus son resistentes a numerosos antibióticos de uso común independientemente del grupo o el sitio de aislamiento. Sin embargo, los análisis fenotípicos indican que las cepas del Sitio A exhiben rasgos asociados con resultados clínicos, incluyendo la capacidad de resistir el suero y catabolizar el ácido siálico, a diferencia de las del Sitio B. Finalmente, los análisis metagenómicos y fisicoquímicos de los sitios de muestreo indican que esta marcada distribución de grupos está fuertemente asociada con distintos factores bióticos y abióticos (por ejemplo, salinidad,
REFERENCIAS BIBLIOGRÁFICAS:
1. O. Faye et al., Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl. Trop. Dis. 8, e2636 (2014).
2. E. M. H. Wellington et al., The role of the natural environment in the emergence of antibiotic resistance in gram-negative bacteria. Lancet Infect. Dis. 13, 155–165 (2013).
3. K. A. Alexander et al., What factors might have led to the emergence of Ebola in West Africa? PLoS Negl. Trop. Dis. 9, e0003652 (2015).
4. S. Khan et al., The emergence of a novel coronavirus (SARS-CoV-2), their biology and therapeutic options. J. Clin. Microbiol. 58, e00187-20 (2020).
5. F. Qadri, T. Islam, J. D. Clemens, Cholera in Yemen—An old foe rearing its ugly head.N. Engl. J. Med. 377, 2005–2007 (2017).
6. K. E. Phillips, K. J. F. Satchell, Vibrio vulnificus: From oyster colonist to human pathogen. PLoS Pathog. 13, e1006053 (2017).
7. S. P. Heng et al., Vibrio vulnificus: An environmental and clinical burden. Front. Microbiol. 8, 997 (2017).
8. M. K. Jones, J. D. Oliver, Vibrio vulnificus: Disease and pathogenesis. Infect. Immun.77, 1723–1733 (2009).
9. S. R. Rippey, Infectious diseases associated with molluscan shellfish consumption.Clin. Microbiol. Rev. 7, 419–425 (1994).
10. P. A. Gulig, K. L. Bourdage, A. M. Starks,Molecular pathogenesis of Vibrio vulnificus. J. Microbiol. 43, 118–131 (2005).
11. W. G. Hlady, K. C. Klontz, The epidemiology of Vibrio infections in Florida, 1981- 1993. J. Infect. Dis. 173, 1176–1183 (1996).
12. N. A. Daniels, Vibrio vulnificus oysters: Pearls and perils. Clin. Infect. Dis. 52, 788–792 (2011).
13. M. S. Strom, R. N. Paranjpye, Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2, 177–188 (2000).
14. M. A. Horseman, S. Surani, A comprehensive review of Vibrio vulnificus: An important cause of severe sepsis and skin and soft-tissue infection. Int. J. Infect. Dis. 15,e157–e166 (2011).
15. C. Baker-Austin, J. D. Oliver, Vibrio vulnificus: New insights into a deadly opportunistic pathogen. Environ. Microbiol. 20, 423–430 (2018).
16. C. Baker-Austin, J. Trinanes, N. Gonzalez-Escalona, J. Martinez-Urtaza, Non-cholera vibrios: The microbial barometer of climate change. Trends Microbiol. 25, 76–84 (2017).
17. C. W. Kaspar, M. L. Tamplin, Effects of temperature and salinity on the survival of Vibrio vulnificus in seawater and shellfish. Appl. Environ. Microbiol. 59, 2425–2429 (1993).
18. S. Paz, N. Bisharat, E. Paz, O. Kidar, D. Cohen, Climate change and the emergence of Vibrio vulnificus disease in Israel. Environ. Res. 103, 390–396 (2007).
19. L. Vezzulli et al., Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. U.S.A. 113, E5062–E5071 (2016).
20. R. Deeb, D. Tufford, G. I. Scott, J. G. Moore, K. Dow, Impact of climate change on Vibrio vulnificus abundance and exposure risk. Estuaries Coasts 41, 2289–2303 (2018).
21. E. K. Lipp, A. Huq, R. R. Colwell, Effects of global climate on infectious disease: The cholera model. Clin. Microbiol. Rev. 15, 757–770 (2002). 22. J. D. Oliver, The biology of Vibrio vulnificus. Microbiol. Spectr. 3, VE-0001-VE-0014 (2015).
23. D. L. Tison, M. Nishibuchi, J. D. Greenwood, R. J. Seidler, Vibrio vulnificus biogroup 2:New biogroup pathogenic for eels. Appl. Environ. Microbiol. 44, 640–646 (1982).
24. J. D. Oliver, Wound infections caused by Vibrio vulnificus and other marine bacteria. Epidemiol. Infect. 133, 383–391 (2005).
25. C. Amaro, E. G. Biosca, Vibrio vulnificus biotype 2, pathogenic for eels, is also an opportunistic pathogen for humans. Appl. Environ. Microbiol. 62, 1454–1457 (1996).
26. N. Bisharat et al., Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Lancet 354, 1421–1424 (1999).
27. R. Zaidenstein et al., Clinical characteristics and molecular subtyping of Vibrio vulnificus illnesses, Israel. Emerg. Infect. Dis. 14, 1875–1882 (2008).
28. M. López-Pérez et al., Evolutionary model of cluster divergence of the emergent marine pathogen Vibrio vulnificus: From genotype to ecotype. MBio 10, e02852-18 (2019).
29. B. J. Shapiro, I. Levade, G. Kovacikova, R. K. Taylor, S. Almagro-Moreno, Origins of pandemic Vibrio cholerae from environmental gene pools. Nat. Microbiol. 2, 16240 (2016).
30. S. N. Sakib, G. Reddi, S. Almagro-Moreno, Environmental role of pathogenic traits in Vibrio cholerae. J. Bacteriol. 200, e00795-17 (2018).
31. F. J. Roig et al., Phylogeny of Vibrio vulnificus from the
nutrientes o biodiversidad) revelando cómo los ecosistemas podrían generar presiones selectivas que faciliten el surgimiento de cepas específicas en una población con potencial patógeno.
Resultados y discusión
El marcador genético thiF puede detectar V. vulnificus y distinguir entre grupos Antes de iniciar nuestro protocolo de muestreo, buscamos marcadores específicos para examinar rápidamente muestras ambientales a gran escala. Específicamente, necesitábamos marcadores genéticos confiables que pudieran: 1) detectar específicamente V. vulnificus, 2) caracterizarlos con precisión en función de su grupo y 3) discriminar entre cepas clonales y no clonales. El gen de hemolisina vvhA , que se usa típicamente para detectar V. vulnificus , aunque es específico de la especie, tiene un potencial limitado para clasificar claramente las cepas en grupos o discriminar entre cepas no clonales (35–37) (Fig. 1A y Apéndice SI , Fig. S1A). Otros enfoques, como la tipificación de secuencias de loci múltiples (MLST), aunque son efectivos para caracterizar cepas, requieren la amplificación por PCR, el ensamblaje y la concatenación de varios genes de mantenimiento (32, 37-40), lo que requiere muchos recursos y es poco práctico para la detección rápida de V. vulnificus en muestras ambientales.
Para identificar marcadores potenciales que cumplan con todos los requisitos mencionados, comparamos todos los genomas C1 y C2 disponibles en bases de datos públicas a través del análisis pangenoma. El número de familias de genes compartidas fue 978 (lo que representa aproximadamente el 22% del número promedio de genes en un genoma de V. vulnificus), que consideramos el genoma central. Realizamos árboles filogenéticos individuales para estos grupos de genes e identificamos, en ambos cromosomas, un total de 47 genes que diferenciaban claramente los grupos C1 y C2. Comparamos estos genes, utilizando cepas representativas de todos los grupos (Apéndice SI, Tabla S1), en función del porcentaje de identidad de secuencia, contra CMCP6, una cepa de referencia de V.
Fig. 1. Aislamiento de V. vulnificus del este de Florida. (A) Árbol filogenético de máxima verosimilitud del gen de hemolisina, vvhA, la proteína transportadora de azufre adenylyltransferasa, thiF, y la concatenación de los seis genes candidatos (yycF, pfeS, acuB, yqhD, uvrY, y thiF) para cepas representativas de los cuatro clústeres. Los miembros del mismo clúster (C1 a C4) se indican con el mismo color. Los árboles no están enraizados y se dibujan a escala. Las longitudes de las ramas indican el número de sustituciones por sitio. (B) Mapa de Florida que indica los sitios de muestreo: la casa de Feller y Shepard Park. (C) Árbol filogenético de máxima verosimilitud de aislamientos de V. vulnificus basado en thiF. Las ramas que contienen miembros que pertenecen al mismo clúster se indican con el mismo color: verde para los representantes de C1 y azul para C2. Los nombres de las cepas están coloreados en relación con la ubicación de origen. Los círculos de colores representan el lugar de aislamiento, y las estrellas rojas representan aquellas cepas que han sido secuenciadas.
vulnificus. Seleccionamos los genes que tenían el mayor porcentaje de identidad con cepas de C1 pero la menor identidad con las de C2 y viceversa y los comparamos con vvhA. Finalmente, seleccionamos un total de seis genes candidatos (yycF , pfeS , acuB , yqhD , uvrY y thiF), tres de cada cromosoma, como marcadores potenciales (Apéndice SI , Tabla S2). Aunque los seis genes marcadores candidatos diferencian claramente las cepas C1 de V. vulnificus de las C2 (Apéndice SI , Fig. S1 A), el regulador de respuesta uvrY y la proteína transportadora de azufre adenililtransferasa thiF, tuvo la máxima resolución para distinguir los cuatro grupos (C1 a C4) así como cepas individuales dentro de cada grupo, lo que sirve como un proxy para la discriminación de poblaciones clonales (Fig. 1 A). Además, las distancias relativas de los cuatro grupos en el árbol filogenético de thiF corresponden con mayor precisión
al árbol evolutivo de V. vulnificus construido utilizando polimorfismos de un solo nucleótido e identidades de nucleótidos promedio de todas las cepas conocidas de V. vulnificus (28).
Al probar la especificidad de especie de thiF con Vibrio parahaemolyticus RMID2210633 o V. cholerae O395, se encontró que thiF era específico para V. vulnificus (Apéndice SI, Fig. S1 D). Por lo tanto, thiF tiene el potencial de 1) detectar cepas de V. vulnificus, 2) separarlas por grupos y 3) discriminar entre cepas clonales y no clonales en función de su genoma completo. Además, la concatenación de los seis genes tuvo al menos el doble de resolución y poder discriminatorio para diferenciar los cuatro grupos en comparación con vvhA , lo que lo convierte en un conjunto preciso de genes para los análisis MLST de las cepas de V. vulnificus (Fig. 1 A).
Fig. 2. Estructura filogenómica y poblacional de V. vulnificus (A) Árbol filogenómico de máxima verosimilitud de las cepas de V. vulnificus obtenidas en este estudio (resaltadas en rojo) junto con todos los genomas de referencia disponibles utilizando el genoma central del cromosoma I. Las ramas que contienen miembros que pertenecen al mismo grupo (C1 a C4) (ANI > 97%) se indican con el mismo color. El gráfico de colores de los círculos del gráfico indica la fuente aislada y el huésped de las cepas correspondientes. El cuadro gris muestra las 15 poblaciones recombinantes detectadas entre todas las cepas. El cuadro naranja resalta las cepas que pertenecen a la subpoblación 15. (B) Representación esquemática de la isla genómica CPS. Las flechas codificadas por colores muestran las ubicaciones de las características genómicas importantes. Las regiones variables 1 y 2 están resaltadas en azul y verde, respectivamente.
Detección
de V. vulnificus a lo largo
del IRL
El IRL (este de Florida, Estados Unidos) es uno de los estuarios con mayor biodiversidad que abarca un amplio rango geográfico con ambientes contrastantes en Florida, donde ocurren con frecuencia brotes de la enfermedad (Fig. 1 B) (7, 34, 41). Recientemente, inspeccionamos este gran estuario y establecimos dos sitios de muestreo en ubicaciones ambientalmente distintivas a lo largo del IRL (Fig. 1 B). Recolectamos muestras en tres eventos de muestreo (15 de noviembre de 2018, 24 de julio de 2019 y 22 de agosto de 2019), incluidos reservorios bióticos como ostras y cianobacterias. V. vulnificus se aisló mediante siembra secuencial de las poblaciones enriquecidas en CHROMagar Vibrio (CaV) y sales biliares de tiosulfato citrato sacarosa (TCBS), como se describe en Materiales y métodos (42). De un total de 1.856 colonias examinadas, solo 245 fueron identificadas como posibles
aislamientos de V. vulnificus según el método de siembra cromogénica. Se detectó una mayor proporción general de V. vulnificus en el Sitio B (Apéndice SI , Fig. S1 B). En el Sitio A, se encontró que la distribución de V. vulnificus era más alta en ostras (45,3%) y agua (43,4%) en contraste con los sedimentos, que contienen en promedio solo 11,32% (Apéndice SI , Fig. S1 B). Además, se observó una mayor proporción de V. vulnificus durante el verano en ambos sitios (Apéndice SI , Fig. S1 B ; 96,3% en el Sitio A, 91,7% en el Sitio B), probablemente como consecuencia del aumento de las temperaturas del agua (>20 °C). Los 245 posibles aislamientos de V. vulnificus se confirmaron además utilizando el nuevo marcador genético thiF . La amplificación por PCR del gen thiF arrojó 141 aislamientos confirmados de V. vulnificus . Secuenciamos estos productos de PCR y construimos un árbol filogenético para determinar la afiliación de grupos. Para minimizar el examen posterior de cepas de origen clonal
analysis of the core-genome: Implications for intra-species taxonomy. Front. Microbiol. 8, 2613 (2018).
32. N. Bisharat et al., Hybrid Vibrio vulnificus. Emerg. Infect. Dis. 11, 30–35 (2005).
33. P. C. Thiaville et al., Genotype is correlated with but does not predict virulence of Vibrio vulnificus biotype 1 in subcutaneously inoculated, iron dextran-treated mice. Infect. Immun. 79, 1194–1207 (2011).
34. A. Newton, M. Kendall, D. J. Vugia, O. L. Henao, B. E. Mahon, Increasing rates of vibriosis in the United States, 1996-2010: Review of surveillance data from 2 systems. Clin. Infect. Dis. 54 (suppl. 5), S391–S395 (2012).
35. V. J. Harwood, J. P. Gandhi, A. C. Wright, Methods for isolation and confirmation of Vibrio vulnificus from oysters and environmental sources: A review. J. Microbiol. Methods 59, 301–316 (2004).
36. F. Han, B. Ge, Multiplex PCR assays for simultaneous detection and characterization of Vibrio vulnificus strains. Lett. Appl. Microbiol. 51, 234–240 (2010).
37. E. Sanjuán, B. Fouz, J. D. Oliver, C. Amaro, Evaluation of genotypic and phenotypic methods to distinguish clinical from environmental Vibrio vulnificus strains. Appl. Environ. Microbiol. 75, 1604–1613 (2009).
38. N. Bisharat et al., The evolution of genetic structure in the marine pathogen, Vibrio vulnificus. Infect. Genet. Evol. 7, 685–693 (2007).
39. H. J. Kim, J. C. Cho, Genotypic diversity and population structure of Vibrio vulnificus strains isolated in Taiwan and Korea as determined by multilocus sequence typing. PLoS One 10, e0142657 (2015).
40. N. González-Escalona, B. Whitney, L. A. Jaykus, A. DePaola, Comparison of direct genome restriction enzyme analysis and pulsed-field gel electrophoresis for typing of Vibrio vulnificus and their correspondence with multilocus sequence typing data. Appl. Environ. Microbiol. 73, 7494–7500 (2007).
41. D. J. Bradshaw II, N. J. Dickens, J. H. Trefry, P. J. McCarthy, Defining the sediment prokaryotic communities of the Indian River Lagoon, FL, USA, an Estuary of National Significance. PLoS One 15, e0236305 (2020).
42. A. Huq et al., Detection, isolation, and identification of Vibrio cholerae from the environment. Curr. Protoc. Microbiol. Chapter 6, 5 (2012).
43. J. M. A. Klapproth, The role of lymphostatin/EHEC factor for adherence-1 in the pathogenesis of gram negative infection. Toxins (Basel) 2, 954–962 (2010).
44. V. L. Barbosa-Jefferson, F. J. Zhao, S. P. McGrath, N. Magan, Thiosulphate and tetrathionate oxidation in arable soils. Soil Biol. Biochem. 30, 553–559 (1998).
45. S. E. Winter et al., Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 467, 426–429 (2010).
46. S. B. Korch, T. M. Hill, Ectopic overexpression of wildtype and mutant hipA genes in Escherichia coli: Effects on macromolecular synthesis and persister formation. J. Bacteriol. 188, 3826–3836 (2006).
47. P. Arevalo, D. VanInsberghe, J. Elsherbini, J. Gore, M. F. Polz, A reverse ecology approach based on a biological definition of microbial populations. Cell 178,820–834.e14 (2019).
48. M. López-Pérez, J. M. Haro-Moreno, F. H. Coutinho, M. Martinez-Garcia, F. Rodriguez-Valera, The evolutionary success of the marine bacterium SAR11 analyzed through a metagenomic perspective. mSystems 5, e00795-17 (2020).
49. G. S. Pettis, A. S. Mukerji, Structure, function, and regulation of the essential virulence factor capsular polysaccharide of vibrio vulnificus. Int. J. Mol. Sci. 21, E3259 (2020).
50. T. C. Williams, M. Ayrapetyan, H. Ryan, J. D. Oliver, Serum survival of Vibrio vulnificus: Role of genotype, capsule, complement, clinical origin, and in Situ incubation. Pathogens 3, 822–832 (2014).
51. H. Y. Kim, M. Ayrapetyan, J. D. Oliver, Survival of Vibrio vulnificus genotypes in male and female serum, and production of siderophores in human serum and seawater. Foodborne Pathog. Dis. 11, 119–125 (2014).
52. G. Gautreau et al., PPanGGOLiN: Depicting microbial diversity via a partitioned pangenome graph. PLOS Comput. Biol. 16, e1007732 (2020). 53. S. Almagro-Moreno, E. F. Boyd, Sialic acid catabolism confers a competitive advantage to pathogenic Vibrio cholerae in the mouse intestine. Infect. Immun. 77, 3807–3816 (2009).
54. S. Almagro-Moreno, E. F. Boyd, Insights into the evolution of sialic acid catabolism among bacteria. BMC Evol. Biol. 9, 118 (2009).
55. V. Bouchet et al., Host-derived sialic acid is incorporated into Haemophilus influenzae lipopolysaccharide and is a major virulence factor in experimental otitis media. Proc. Natl. Acad. Sci. U.S.A. 100, 8898–8903 (2003).
56. E. Severi, D. W. Hood, G. H. Thomas, Sialic acid utilization by bacterial pathogens. Microbiology (Reading) 153, 2817–2822 (2007).
57. J. B. Lubin, J. J. Kingston, N. Chowdhury, E. F. Boyd, Sialic acid catabolism and transport gene clusters are lineage
que podrían haber proliferado durante el enriquecimiento, solo analizamos una cepa dentro de un grupo si 1) la alineación de thiF parecía idéntica dentro del grupo, 2) las cepas provenían de la misma réplica y fracción, y 3) se aislaron durante el mismo evento de muestreo. Como resultado, 87 de los 141 aislamientos confirmados de V. vulnificus fueron seleccionados para análisis adicionales (39 aislamientos del Sitio A y 48 del Sitio B) (Fig. 1 C).
Sorprendentemente, el análisis filogenético utilizando el marcador genético thiFmostró que la mayoría de los aislamientos del Sitio A pertenecen a C1 (97,4%, 38/39), mientras que la mayoría de los aislamientos del Sitio B pertenecen a C2 (87,5%. 42/48) (Fig. 1 C). Esta clara separación ecológica entre los dos grupos proporciona un marco ideal para examinar los procesos evolutivos subyacentes a la aparición de rasgos patógenos dentro de una población y una plataforma para comprender cómo los ecosistemas generan presiones que facilitan la selección de cepas con potencial patógeno. Para abordar esto, primero diseccionamos los determinantes genómicos y la estructura poblacional de estas cepas ambientales de V. vulnificus, evaluamos su potencial patógeno y finalmente vinculamos estos resultados con factores ambientales (abióticos y bióticos) asociados con su marcada distribución de grupos.
Determinantes genómicos de la aparición de V. vulnificus
Preferencias ecológicas de los grupos de V. vulnificus
Para investigar los determinantes genómicos que potencialmente impulsan las preferencias de nicho ecológico de los grupos, seleccionamos varias cepas para la secuenciación del genoma para obtener una representación proporcional de cada grupo, reservorio, fracción, huésped y fecha de aislamiento. Esto resultó en un total de 27 aislamientos de V. vulnificus secuenciados (Apéndice SI , Tabla S3): 13 del Sitio A (2 sedimentos, 6 ostras y 5 aislamientos de agua, 1 de los cuales pertenecía a C2) y 14 del Sitio B (4 sedimentos, 4 cianobacterias y 6 aislamientos de agua, incluidos 2 de C1). Para una asociación filogenómica robusta, incluimos 74 genomas desreplicados de V. vulnificus (por ejemplo, genomas >99% de
Fig. 3. Análisis del pangenoma de cepas de V. vulnificus. (A) Análisis del pangenoma para los grupos: 1) nueve cepas C1 aisladas del Sitio A (C1 IRL), 2) nueve cepas C2 del Sitio B (C2 IRL), 3) nueve cepas de referencia C1 que son auténticamente clínicas (C1 Clinical), y 4) nueve cepas ambientales de referencia de C2 (C2 environmental). Las proporciones de familias de genes en el genoma persistente, de nube y de caparazón están destacadas en naranja, verde y azul, respectivamente. (B) Representación esquemática comparando la isla genómica del grupo de genes involucrados en el catabolismo del ácido siálico. (C) Comparación de la razón entre sustituciones no sinónimas y sinónimas (razón dN/dS) entre cepas clínicas de referencia y cepas C1 aisladas del Sitio A en el IRL. (D) Comparación de los valores de dN/dS de cada cepa individual versus el resto en el grupo C1 IRL para genes que codifican estas CAAs. Aquellas con un valor por encima del promedio han sido destacadas en rojo."
identidad de nucleótidos promedio [ANI]) actualmente disponibles en bases de datos públicas.
Utilizamos árboles filogenómicos y agrupamiento basado en ANI de ambos cromosomas por separado para agrupar los genomas en los grupos previamente definidos [C1 a C4 (28)] (Apéndice SI , Figs. S2 A y S3).
Con base en estos resultados, decidimos utilizar el ANI del cromosoma I como referencia para la clasificación taxonómica ya que la cobertura es alta (>70%), incluso entre los grupos más divergentes (C1 y C2). Curiosamente, encontramos evidencia de mezcla o transferencia de cromosomas entre grupos de V. vulnificus. Por ejemplo, mientras que el cromosoma I de FORC_037, una cepa ambiental aislada de almeja de caparazón blando, tuvo un ANI >98% con miembros de C2 y ca. 95% con C1, para el cromosoma II fue al revés (Apéndice SI , Fig. S2 A y B).
La filogenia del genoma completo confirmó las marcadas diferencias en la distribución de los clúster de V. vulnificus obtenidos con el gen thiF, corroborando el enriquecimiento de cepas C1 en el Sitio A (Fig. 2 A), excepto la cepa IRLE0015 que junto con NV22 se agrupó estrechamente con las cepas BT3 del brote de Israel (Fig. 2 A). Como se mencionó anteriormente, seleccionamos una cepa no clonal del Sitio A que pertenecía a C2 (IRLA0043) y dos del Sitio B que pertenecían a C1 (IRLE0056 e IRLE0004). Esto nos dio la oportunidad de investigar la presencia de determinantes genómicos potenciales asociados específicamente con cada sitio, es decir, si las cepas C1 y C2 del Sitio A tienen un grupo único de genes que está ausente en las cepas del Sitio B independientemente del cluster. La parte común del pangenoma de todas las cepas C2 del Sitio B se restó del genoma de la cepa IRLA0043, la única en este grupo aislada del Sitio A. Más de 500 genes eran específicos de esta cepa, y aparte de los genes de glicosilación de la
cápsula, encontramos un segundo grupo de genes (rtxB-rtxD-rtxE) que codifican un sistema de secreción de tipo I (T1SS) con una alta similitud (99%) con varias cepas de V ibrio coralliilyticus .
Específicamente, este sistema parece estar asociado con la excreción de una enterotoxina (Efa-1/LifA) (43). Dentro de estos genes específicos, también encontramos un segundo sistema de secreción de tipo VI (T6SS) (28) y un elemento conjugativo integrador. Por otro lado, las cepas C1 del Sitio B (IRLE0004 e IRLE0056) tenían solo 200 genes únicos en comparación con las cepas C1 del Sitio A. Entre los genes específicos de IRLE0004, encontramos un grupo de genes que confiere la capacidad de utilizar tetrationato como aceptor de electrones, un compuesto de azufre común presente en la mayoría de los suelos (44); curiosamente, esta cepa ha sido aislada del sedimento. La capacidad de utilizar tetrationato se ha asociado con la virulencia en Salmonella enterica al proporcionar una ventaja de crecimiento a la bacteria en el intestino inflamado (45). La anotación funcional asociada con la parte específica de IRLE0056 se limitó al uso de ramnosa, varios sistemas toxina-antitoxina y el gen que codifica la HipA involucrada en la latencia (46). Aunque resalta la presencia en el ambiente de algunos factores de virulencia que pueden ser fácilmente compartidos entre los dos clúster, nuestro análisis no identificó ningún determinante genómico específico que pueda explicar la distribución diferencial de estas cepas.
Estructura filogenómica y poblacional de V. vulnificus (A) Árbol filogenómico de máxima verosimilitud de las cepas de V. vulnificus obtenidas en este estudio (resaltadas en rojo) junto con todos los genomas de referencia disponibles utilizando el genoma central del cromosoma I. Las ramas que contienen miembros que pertenecen al mismo grupo (C1 a C4) (ANI > 97%) se indican con el mismo color. El gráfico de colores de los círculos del gráfico indica la fuente aislada y el huésped de las cepas correspondientes. El cuadro gris muestra las 15 poblaciones recombinantes detectadas entre todas las cepas. El cuadro naranja resalta las cepas que pertenecen a la subpoblación 15. (B) Representación
esquemática de la isla genómica CPS. Las flechas codificadas por colores muestran las ubicaciones de las características genómicas importantes. Las regiones variables 1 y 2 están resaltadas en azul y verde, respectivamente.
Poblaciones ecológicamente significativas de V. vulnificus
A pesar de las marcadas preferencias ambientales y la divergencia genómica entre los grupos C1 y C2, nuestros recientes estudios in silico indican un intercambio frecuente de elementos genéticos móviles (28). Aquí, tenemos la oportunidad de estudiar la recombinación potencial en poblaciones naturales de V. vulnificus en un área endémica. La recombinación es particularmente preocupante ya que las prácticas novedosas como la acuicultura pueden conducir a la aparición de cepas híbridas, como lo demuestra un brote mortal en Israel causado por un grupo completamente nuevo (C3) (27) y la presencia de una cepa similar a C3 aislada en este estudio (IRLE0015) (Fig. 2 A). Para evaluar este fenómeno, utilizamos un enfoque para evaluar eventos recientes de recombinación que permite la delineación de poblaciones ecológicamente relevantes, es decir, grupos con el potencial de intercambiar material genético (47). Nuestros análisis revelaron la presencia de 15 poblaciones recombinantes principales. Algunas de estas poblaciones coinciden con la clasificación de clúster indicativa de alta recombinación intragrupo, por ejemplo, C3 y C4 (Fig. 2 A). Sin embargo, C2 está formado por 12 poblaciones. Once de ellas formadas por un solo miembro y por lo tanto indicando que no hay eventos de recombinación que conecten estas cepas con el resto del clúster (48). Curiosamente, todos los miembros de C1 forman una sola población (P15) con la mayoría de los representantes de C2 indicando que, a pesar de la divergencia (ca. 95% ANI), estos clúster están conectados por eventos de recombinación recientes (Fig. 2 A).
El grupo de polisacáridos capsulares (CPS) es un factor de virulencia esencial de V. vulnificus (49). Nuestros análisis previos sugieren que la recombinación puede ser un mecanismo evolutivo importante que conduce a la alta diversidad del grupo de
specific in Vibrio vulnificus. Appl. Environ.Microbiol. 78, 3407–3415 (2012).
58. D. E. Kunkle, X. R. Bina, J. E. Bina, Vibrio cholerae OmpR contributes to virulence repression and fitness at alkaline ph. Infect. Immun. 88, e00141-20 (2020).
59. D. E. Kunkle, T. F. Bina, X. R. Bina, J. E. Bina, Vibrio cholerae OmpR represses the ToxR regulon in response to membrane intercalating agents that are prevalent in the human gastrointestinal tract. Infect. Immun. 88, e00912–e00919 (2020).
60. J. Liu, K. Duncan, C. T. Walsh, Nucleotide sequence of a cluster of Escherichia coli enterobactin biosynthesis genes: Identification of entA and purification of its product 2,3-dihydro-2,3-dihydroxybenzoate dehydrogenase. J. Bacteriol. 171, 791–798 (1989).
61. E. E. Wyckoff, J. A. Stoebner, K. E. Reed, S. M. Payne, Cloning of a Vibrio cholerae vibriobactin gene cluster: Identification of genes required for early steps in siderophore biosynthesis. J. Bacteriol. 179, 7055–7062 (1997).
62. S. I. Wu, S. K. Lo, C. P. Shao, H. W. Tsai, L. I. Hor, Cloning and characterization of a periplasmic nuclease of Vibrio vulnificus and its role in preventing uptake of foreign DNA. Appl. Environ. Microbiol. 67, 82–88 (2001).
63. I. Sepúlveda Cisternas, J. C. Salazar, V. A. García-Angulo, Overview on the bacterial iron-riboflavin metabolic axis. Front. Microbiol. 9, 1478 (2018).
64. J. Sivaraman et al., Crystal structure of Escherichia coli PdxA, an enzyme involved in the pyridoxal phosphate biosynthesis pathway. J. Biol. Chem. 278, 43682–43690 (2003).
65. A. Nakhamchik, C. Wilde, D. A. Rowe-Magnus, Identification of a Wzy polymerase required for group IV capsular polysaccharide and lipopolysaccharide biosynthesis in Vibrio vulnificus. Infect. Immun. 75, 5550–5558 (2007).
66. E. Severi et al., Sialic acid transport in Haemophilus influenzae is essential for lipopolysaccharide sialylation and serum resistance and is dependent on a novel tripartite ATPindependent periplasmic transporter. Mol. Microbiol. 58, 1173–1185 (2005).
67. B. P. Bougnom, L. J. V. Piddock, Wastewater for urban agriculture: A significant factor in dissemination of antibiotic resistance. Environ. Sci. Technol. 51, 5863–5864 (2017).
68. S. B. Zaman et al., A review on antibiotic resistance: Alarm bells are ringing. Cureus 9, e1403 (2017).
69. K. S. Shaw et al., Antimicrobial susceptibility of Vibrio vulnificus and Vibrio parahaemolyticus recovered from recreational and commercial areas of Chesapeake Bay and Maryland Coastal Bays. PLoS One 9, e89616 (2014).
70. M. A. Campos et al., Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides. Infect. Immun. 72, 7107–7114 (2004).
71. M. P. E. Slack, W. W. Nichols, Antibiotic penetration through bacterial capsules and exopolysaccharides. J. Antimicrob. Chemother. 10, 368–372 (1982).
72. G. Reddi, K. Pruss, K. L. Cottingham, R. K. Taylor, S. Almagro-Moreno, Catabolism of mucus components influences motility of Vibrio cholerae in the presence of environmental reservoirs. PLoS One 13, e0201383 (2018).
73. S. N. Sakib, G. Reddi, S. Almagro-Moreno, Environmental role of pathogenic traits in Vibrio cholerae. J. Bacteriol. 200, e00795-17 (2018).
74. C. Baker-Austin et al., Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 3, 73–77 (2012).
75. J. S. Mackenzie, M. Jeggo, The one health approach-why is it so important? Trop. Med. Infect. Dis. 4, E88 (2019).
76. G. C. Sigua, W. A. Tweedale, Watershed scale assessment of nitrogen and phosphorus loadings in the Indian River Lagoon basin, Florida. J. Environ. Manage. 67, 363–372 (2003).
77. P. J. Barile, Widespread sewage pollution of the Indian River Lagoon system, Florida (USA) resolved by spatial analyses of macroalgal biogeochemistry. Mar. Pollut. Bull. 128, 557–574 (2018).
78. B. M. Schuster et al., Ecology and genetic structure of a northern temperate Vibrio cholerae population related to toxigenic isolates. Appl. Environ. Microbiol. 77, 7568–7575 (2011).
79. C. A. Kaysner, J. Angelo DePaola, “Vibrio” in Bacteriological Analytical Manual, K. Jinneman, Ed. (AOAC International, 2004).
80. A. M. Bolger, M. Lohse, B. Usadel, Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
81. A. Bankevich et al., SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477 (2012).
82. D. Hyatt et al., Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 11, 119 (2010).
83. T. M. Lowe, S. R. Eddy, tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 25, 955–964 (1997).
84. E. P. Nawrocki, “Structural RNA homology search and alignment using covariance models,” PhD thesis, Washington
CPS (28). Por lo tanto, investigamos la diversidad genómica del CPS entre ambos grupos en estas poblaciones naturales. La cepa IRLA0152 (C1) aislada de la fracción de vida libre en el Sitio A tenía una variante similar del CPS encontrado en un aislado de paciente infectado (FDAARGOS_119) (Fig. 2 B). Una de las partes hipervariables del CPS del aislado de ostras OH0023 fue idéntica a la encontrada en la cepa clínica de referencia CMCP6, destacando el medio ambiente como un reservorio de estos genes de virulencia esenciales (Fig. 2 B). Además, ciertos grupos de CPS se distribuyen en la población independientemente del grupo de origen y la ubicación del muestreo. En concreto, encontramos el mismo CPS en una cepa C1 del Sitio B (IRLE0056) y tres cepas C2, una de ellas del Sitio A (IRLE0043) y dos del Sitio B (IRLE0062 e IRLE0057) (Fig. 2 B). La única variación fue una pequeña inserción en IRLE0043 debido a varios elementos de secuencia de inserción (IS), lo que sugiere que este puede ser otro mecanismo que puede introducir variabilidad dentro del cluster CPS (Fig. 2 B). En general, nuestros resultados indican que a pesar de la divergencia genómica y sus marcadas diferencias ecológicas, existe una amplia recombinación entre los clúster en un área endémica como el IRL, incluyendo la transferencia de importantes factores de virulencia dentro de su entorno natural. Los análisis del pangenoma revelan impulsores genéticos asociados con la aparición de virulencia.
La mayoría de las cepas clínicas de V. vulnificus pertenecen a C1, de manera similar a la mayoría de las cepas aisladas del Sitio A. Hasta la fecha, los determinantes genómicos específicos que permiten que algunas cepas C1 colonicen con éxito a los humanos siguen siendo en su mayoría desconocidos. Con el fin de dilucidar los factores genéticos asociados con la aparición de V. vulnificus C1 clínico a partir de acervos genéticos ambientales y para determinar si las cepas C1 del Sitio A codificaron rasgos asociados a la clínica, comparamos los genomas de las cepas aisladas en este estudio con los de cepas C1 clínicas auténticas y no patógenas (50 , 51). Específicamente, seleccionamos genomas de cuatro grupos distintos: 1) nueve cepas C1 aisladas del Sitio A y 2) nueve cepas C2 del Sitio B junto con 3) nueve cepas C1
Fig. 4. Evaluación del potencial patogénico de aislamientos de V. vulnificus IRL. (A) Patrones de resistencia a antibióticos de 27 aislamientos de V. vulnificus frente a 12 antibióticos comúnmente usados. Rojo: resistente; rosa: resistencia intermedia; blanco: sensible. (B) Resistencia al suero de V. vulnificus expuesto a suero humano normal durante 2 horas, evaluada en términos de supervivencia como UFC/mililitros. Cepas resistentes: UFC/mililitros similar a la entrada; cepas sensibles: UFC/ mililitros menor que la entrada; resistente y crecimiento en suero: UFC/mililitros mayor que la entrada. (C) Capacidad de catabolizar ácido siálico evaluada por el crecimiento de aislamientos de V. vulnificus en un medio mínimo M9 suplementado con NANA como única fuente de carbono en salinidades que representan las dos ubicaciones de muestreo. El crecimiento se midió en función del aumento de OD (OD595) de las culturas a lo largo del tiempo.
que son clínicamente bona fide, es decir, aisladas de pacientes con septicemia, así como 4) nueve cepas no patógenas de C2, es decir, aisladas de fuentes ambientales y susceptibles al efecto bactericida del suero y los monocitos (50, 51). La diversidad de especies microbianas se analizó mediante un Gráfico PanGenómico Particionado de Vecinos Vinculados [PPanGGOLiN (52)]. El tamaño estimado del “genoma persistente” (familias de genes presentes en casi todos los genomas) es similar para cada grupo individual así como para todos los grupos combinados juntos, ca. 3.700 familias de genes (ca. 52% del total de familias de genes por genoma). Esto es bastante notable dada la divergencia genómica entre los grupos (Fig. 3 A). La proporción de familias de genes que formaron el “genoma de concha” (familias de genes presentes en tres a siete genomas) fue solo del 1% del total para ambos grupos C1 y del 2% para los grupos C2. Las familias de genes restantes
presentes en baja frecuencia (uno a tres genomas) se clasificaron como el “genoma de la nube” (Fig. 3 A). Como se predijo, el porcentaje de familias de genes asignadas a categorías funcionales (base de datos de subsistemas SEED) para cada partición del pangenoma varió significativamente: desde el 64% asignado al genoma persistente hasta aproximadamente el 20% para la nube y la concha. Este último se asocia típicamente con diversas adaptaciones ambientales, incluida la patogénesis, lo que resalta la enorme plasticidad genómica que aún queda por abordar para estos organismos.
A continuación, comparamos las clasificaciones funcionales de las secuencias codificantes de genes de los genomas persistentes de las nueve cepas clínicas C1 de referencia contra las nueve cepas C1 analizadas del Sitio A. Encontramos que ambos grupos solo difieren en ˜2% del contenido total de genes de su
genoma persistente. La mayoría de estas diferencias se asociaron con la presencia de genes pertenecientes a la clasificación “Metabolismo del ácido siálico” en las cepas clínicas C1 (Fig. 3 B). Este grupo de genes codifica un sistema de transporte periplásmico independiente de ATP tripartito completo (TRAP) involucrado en el transporte de ácido siálico para las enzimas responsables de su catabolismo (N-acetilneuraminato liasa, N-acetilmanosamina quinasa y N-acetilmanosamina-6-fosfato 2-epimerasa) así como una mutarotasa de ácido siálico (familia YjhT) y un regulador de la utilización de ácido siálico, familia RpiR (53). La capacidad de hurgar, decorar su superficie y utilizar ácido siálico como fuente de carbono es un factor de virulencia importante para las bacterias patógenas y oportunistas, incluyendo V. vulnificus (54–57) . Utilizando el genoma de referencia clínica C1 CMCP6, encontramos que el grupo completo estaba ubicado en una isla genómica en el cromosoma II (Fig. 3 B). El mismo grupo de genes se puede encontrar en otras especies de Vibrio (aproximadamente 70% de identidad BLASTN) como V. cholerae O1, Vibrio mimicus o Vibrio anguillarum ; sin embargo, a diferencia de V. vulnificus , en estas especies, el grupo estaba flanqueado por elementos de secuencia de inserción.
Dada la frecuente transferencia horizontal de genes en las poblaciones de V. vulnificus , es poco probable que la presencia/ausencia de genes o grupos de genes sea suficiente para explicar la aparición de rasgos de virulencia que conducen a resultados clínicos en este patógeno. Nuestras investigaciones previas con V. cholerae sugieren que las variaciones alélicas de los genes centrales pueden ser impulsores principales de la aparición de virulencia (29). Por lo tanto, evaluamos los patrones de microdiversidad del genoma persistente estimando la relación de las tasas de sustitución no sinónima (dN) a sinónima (dS) en la comparación de genomas por pares. Encontramos seis genes dentro de las cepas clínicas C1 que mostraron una fuerte selección positiva en comparación con las cepas C1 IRL, que en promedio exhibieron una fuerte selección purificadora (Fig. 3 C y Apéndice SI , Tabla S4). Además, los valores promedio de dN/dS para estos genes dentro de los grupos C2, tanto en
las referencias ambientales como en los aislados del IRL, también exhibieron valores de dN/dS muy bajos (Apéndice SI , Tabla S4). Los genes que codifican estos CAA difieren entre cepas clínicas y están involucrados en procesos asociados a la virulencia y el metabolismo de nutrientes relacionado con el huésped (Apéndice SI , Tabla S4). Por ejemplo, uno de estos genes codifica el regulador de porina de membrana externa OmpR, que regula la virulencia en V. cholerae a través de aphB (58 , 59). Otro, que codifica la subunidad EntD, forma parte del complejo enzimático enterobactinasintetasa, un sistema de adquisición de hierro esencial para la virulencia en Escherichia coli (60), y se propuso que desempeña un papel en las últimas etapas de la biosíntesis de enterobactina en V. cholerae (61). La endonucleasa vvn , identificada como una nucleasa periplásmica en V. vulnificus , previene la captación de ADN extraño (62), dificultando así la introducción de plásmidos por transformación. La riboflavina sintasa, ribE , cataliza el paso final en la biosíntesis de riboflavina o vitamina B2. La riboflavina está involucrada en varias vías metabólicas [p. ej., biodisponibilidad y adquisición de hierro (63)] en muchos patógenos, incluido V. cholerae . El fosfato de piridoxal, PdxA, la forma catalíticamente activa de la vitamina B6 , es un cofactor importante para muchas vías enzimáticas que involucran la descomposición de aminoácidos (64) .) y el componente TusB del complejo de transferencia de azufre TusBCD. En promedio, estos genes tuvieron valores de dN/dS más bajos en las cepas C1 IRL en comparación con C1 clínica; sin embargo, dado que V. vulnificus clínica es endémica de esta área, es posible que algunas cepas C1 IRL individuales codifiquen CAA. Para determinar esto, analizamos su presencia identificando variantes alélicas individuales que se desvían de los valores promedio (Fig. 3 D). Curiosamente, aunque ninguno de los alelos de las cepas C1 IRL fue idéntico a los encontrados en las cepas clínicas, cada uno de ellos codificó al menos un gen con un dN/dS por encima del promedio. Estos variaron desde la cepa OH0003 que codifica uno (gen tus B) hasta IRLA0186 que codifica cuatro de ellos (omp R, rib E, ent D y pdx A) (Fig. 3 D). En general, nuestros resultados demuestran que 1) las cepas clínicas codifican CAA únicas y 2) las
University in St. Louis, St. Louis, MO (2009).
85. Y. Huang, P. Gilna, W. Li, Identification of ribosomal RNA genes in metagenomic fragments. Bioinformatics 25, 1338–1340 (2009).
86. B. Buchfink, C. Xie, D. H. Huson, Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60 (2015).
87. R. L. Tatusov et al., The COG database: New developments in phylogenetic classification of proteins from complete genomes. Nucleic Acids Res. 29, 22–28 (2001).
88. D. H. Haft et al., TIGRFAMs: A protein family resource for the functional identification of proteins. Nucleic Acids Res. 29, 41–43 (2001). 89. S. R. Eddy, Accelerated profile HMM searches. PLOS Comput. Biol. 7, e1002195 (2011).
90. S. F. Altschul et al., Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402 (1997).
91. M. Mirdita et al., Uniclust databases of clustered and deeply annotated protein sequences and alignments. Nucleic Acids Res. 45, D170–D176 (2017).
92. A. Gudy s, S. Deorowicz, QuickProbs 2: Towards rapid construction of high-quality alignments of large protein families. Sci. Rep. 7, 41553 (2017).
93. A. Criscuolo, S. Gribaldo, BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 10, 210 (2010).
94. L. T. Nguyen, H. A. Schmidt, A. von Haeseler, B. Q. Minh, IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274 (2015).
95. L. Pritchard, R. H. Glover, S. Humphris, J. G. Elphinstone, I. K. Toth, Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 8, 12–24 (2016).
96. R. Overbeek et al., The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic Acids Res. 33, 5691–5702 (2005).
97. H. G. Drost, A. Gabel, I. Grosse, M. Quint, Evidence for active maintenance of phylotranscriptomic hourglass patterns in animal and plant embryogenesis. Mol. Biol. Evol. 32, 1221–1231 (2015).
98. J. B. Lubin et al., Host-like carbohydrates promote bloodstream survival of Vibrio vulnificus in vivo. Infect. Immun. 83, 3126–3136 (2015).
99. C. Scarano et al., Antibiotic resistance of Vibrio species isolated from Sparus aurata reared in Italian mariculture. New Microbiol. 37, 329–337 (2014).
100. F. Han, R. D. Walker, M. E. Janes, W. Prinyawiwatkul, B. Ge, Antimicrobial susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus isolates from Louisiana Gulf and retail raw oysters. Appl. Environ. Microbiol. 73, 7096–7098 (2007).
101. C. Baker-Austin et al., Multi-site analysis reveals widespread antibiotic resistance in the marine pathogen Vibrio vulnificus. Microb. Ecol. 57, 151–159 (2009).
102. CLSI, Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria. CLSI Guideline M45. (Clinical and Laboratory Standards Institute, 2015).
103. R. W. Bogard, J. D. Oliver, Role of iron in human serum resistance of the clinical and environmental Vibrio vulnificus genotypes. Appl. Environ. Microbiol. 73, 7501–7505 (2007).
104. USEPA, “Methods for the determination of metals in environmental sample” (EPA 600/R-93/100s, 1993).
105. W. McDaniel, Environmental Monitoring Systems Laboratory, Methods for the Determination of Metals in Environmental Samples (Noyes Publications, 1996).
106. J. Sambrook, D. W. Russel, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Lab Press, 2000).
107. R. C. Edgar, Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461 (2010).
108. J. R. Cole et al., Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, D633–D642 (2014).
109. S. R. Eddy, Multiple alignment using hidden Markov models. Proc. Int. Conf. Intell. Syst. Mol. Biol. 3, 114–120 (1995).
variantes alélicas de estos genes circulan en poblaciones naturales.
Evaluación del potencial patogénico de cepas de V. vulnificus
Con el fin de evaluar el potencial patógeno de las cepas ambientales de la vida real y su asociación con la filogenia y la ubicación, probamos fenotípicamente 1) su perfil de resistencia a los antibióticos, 2) su supervivencia en presencia de suero humano y 3) su capacidad para utilizar ácido siálico como única fuente de carbono. Para estos ensayos, incluimos V. vulnificus CMCP6 (C1 clínico) y V. vulnificus SS108-A3A (C2 ambiental no patógeno) como cepas de referencia auténticas. Además, construimos tres cepas mutantes isogénicas en el fondo de V. vulnificus CMCP6, donde eliminamos los genes que codifican 1) la proteína de transporte CPS Wza (∆ wza), que ha demostrado desempeñar un papel en la supervivencia del suero y la producción de cápsulas (65), 2) N -acetilneuraminato liasa (∆ nanA), la primera enzima en la vía catabólica del ácido siálico (54), y 3) la permeasa grande del transportador TRAP del ácido siálico (∆ siaM), que está asociada con la captación de ácido siálico y también está involucrada en la resistencia del suero (66).
Resistencia antibiótica
Primero, examinamos el perfil de resistencia a antibióticos de las cepas IRL para determinar si había patrones asociados con la distribución diferencial de los grupos, ya que ambos sitios tienen una exposición muy diferente a perturbaciones provocadas por el hombre, incluidos los antibióticos (67, 68). Probamos varios antibióticos recomendados por los Centros para el Control y la Prevención de Enfermedades para el tratamiento de Vibrio spp. (69). Mientras que V. vulnificus CMCP6 mostró resistencia o resistencia intermedia a prácticamente todos los antibióticos probados (Fig. 4 A), ∆ wza , ∆ nanA y ∆ siaM mostraron una mayor sensibilidad a varios de ellos en comparación con el tipo salvaje (Fig. 4 A). La cápsula generalmente confiere resistencia a los antibióticos (70 , 71); sin embargo, los mecanismos por los cuales el catabolismo y la captación de ácido siálico están involucrados en la resistencia a los antibióticos aún deben dilucidarse. La mayoría de las cepas IRL son resistentes a la polimixina
Fig. 5. Factores ambientales asociados con la divergencia de clústeres. (A) Clasificación taxonómica basada en fragmentos del gen 16S rRNA (lecturas crudas) de los diferentes metagenomas obtenidos de agua de mar filtrada con un filtro de 0.22 μm. Solo se muestran aquellos grupos con valores de abundancia superiores al 1% en cualquiera de los metagenomas. El tamaño del diámetro de los círculos indica el porcentaje del total de lecturas para cada taxón. (B) Análisis de coordenadas principales entre parámetros fisicoquímicos y abundancia de los diferentes taxones basado en fragmentos metagenómicos del gen 16S rRNA. (C) Diagramas de caja que ilustran la diversidad de la comunidad microbiana medida usando el índice de Shannon."
B, gentamicina, sulfadiazina e imipenem, un antibiótico β-lactámico. Por el contrario, prácticamente ninguna cepa IRL fue resistente al cloranfenicol o la oxitetraciclina (Fig. 4 A). Siete cepas del Sitio B exhibieron resistencia intermedia al ácido nalidíxico y/o trimetoprima, mientras que solo dos de los aislados del Sitio A fueron resistentes a estos compuestos. Sorprendentemente, una cepa C1 aislada del Sitio B (IRLE0004) mostró niveles de resistencia variados a todos los antibióticos probados con la excepción de la oxitetraciclina. Curiosamente, dos cepas C1 del Sitio A (IRLA0161 e IRLA0152) que pertenecían al mismo marco clonal, es decir, ANI > 99%, mostraron diferentes patrones de resistencia a los antibióticos (Fig. 4 A). A diferencia de IRLA0152, IRLA0161 es resistente a oxitetraciclina, ácido nalidíxico y trimetoprima. El análisis del genoma mostró la presencia de un plásmido de 172 Kb en esta cepa, en el que identificamos un gen codificante para una familia de dihidrofolato reductasas resistentes a trimetoprima, DfrA. Aunque no se identificaron los genes directamente responsables de las otras dos resistencias, encontramos varios genes relacionados con bombas de eflujo codificados en el mismo plásmido.
Parece, a partir de nuestro análisis, que las presiones selectivas en el Sitio B, el sitio con mayor exposición antropogénica, favorecen la aparición de resistencia a antibióticos, particularmente al inhibidor de folato, trimetoprima, y a la quinolona, ácido nalidíxico (Fig. 4 A). Además, la presencia de plásmidos resistentes y su facilidad de transmisión entre los dos clúster (28) aumenta la probabilidad de que las cepas de C1 adquieran estos genes a través de transferencia horizontal de genes.
Resistencia del suero
Algunos estudios han informado previamente de la capacidad de las cepas clínicas de V. vulnificus para resistir el efecto bactericida del suero, mientras que la mayoría de las cepas ambientales probadas fueron susceptibles a él (50 , 51). Dado que la resistencia al suero es un rasgo de virulencia esencial para la patogénesis de V. vulnificus, analizamos la susceptibilidad de los aislamientos IRL a esta defensa primaria del huésped. Como se esperaba, la cepa clínica C1 de tipo salvaje fue resistente al suero, mientras que la cepa no patógena C2 fue sensible a su efecto bactericida (disminuciones de 3 a 4 logaritmos en las unidades formadoras de
colonias [UFC]) (Fig. 4 B). Solo 3 de las 12 cepas del Sitio A fueron sensibles al suero, mientras que en el Sitio B, encontramos el patrón opuesto, con la mayoría de las cepas (8 de 14) siendo sensibles (Fig. 4 B). Estas diferencias se asociaron fuertemente con la distribución de grupos y nos proporcionaron una oportunidad para examinar los posibles determinantes genómicos que conducen a la resistencia al suero en V. vulnificus. Primero comparamos el contenido genético entre cepas C1 resistentes al suero (OH0023 e IRLA0152) contra las sensibles (OH0012 e IRLA0153). Entre esos genes únicos en las cepas resistentes, encontramos varios relacionados con sistemas de restricciónmodificación de tipo I, síntesis de cápsula y aquellos involucrados en el metabolismo del ácido siálico. Posteriormente, analizamos la presencia del grupo de ácido siálico en los genomas de todos los aislados IRL en nuestro estudio. Encontramos que 12 de 15 cepas que eran resistentes al suero (8 Sitio A; 4 Sitio B) codificaban el grupo, mientras que solo 1 de 11 cepas sensibles lo hacía (Fig. 4 B). Dada esta clara asociación, probamos la resistencia al suero de ∆ wza y los dos mutantes de ácido siálico, ∆ nanA y ∆ siaM . Como se esperaba, ∆ wza era sensible al suero. Curiosamente, mientras que ∆ siaM exhibió una disminución de 2 logaritmos en UFC en comparación con el tipo salvaje, ∆ nanA no se vio afectado por el efecto bactericida del suero, y el mecanismo detrás de la diferencia en la supervivencia entre estos dos mutantes aún debe abordarse.
Catabolismo del ácido siálico
El ácido siálico, además de jugar un papel importante en las interacciones huéspedpatógeno (54 , 56), es crítico para la interacción de varios Vibrios patógenos con algunos de sus reservorios ambientales, como las cianobacterias, potencialmente vinculando diferentes estilos de vida de patógenos bacterianos (72 , 73). Tanto nuestro análisis pangenómico como fenotípico sugieren que el catabolismo de este aminoazúcar parece ser un factor esencial asociado con los resultados clínicos. Para probar inicialmente nuestros hallazgos, examinamos la capacidad de las cepas IRL para utilizar ácido N-acetilneuramínico (NANA) como única fuente de carbono. Probamos su crecimiento en medios
mínimos M9 suplementados con NANA en dos salinidades que reflejan los dos sitios de muestreo (1% y 3% NaCl; Apéndice SI , Tabla S5). Ni los mutantes ∆ nanA y ∆ siaM ni los aislados IRL que no codificaron el grupo de ácido siálico pudieron crecer en estos medios. Todas las cepas del Sitio A que poseían el grupo de ácido siálico (8 de las 12) exhibieron patrones de crecimiento similares a la referencia clínica CMCP6 en ambas salinidades. En el Sitio B, solo 6 de los 14 aislamientos pudieron crecer, todos conteniendo el grupo de ácido siálico (Fig. 4 C).
En conjunto, nuestros análisis genómicos y fenotípicos de las cepas IRL y sus comparaciones con cepas clínicas mostraron un potencial diferencial para la emergencia de patógenos en estas poblaciones naturales. Por ejemplo, la cepa IRLA0186 exhibe varios rasgos que indican su fuerte capacidad para la emergencia como cepa clínica, como su capacidad para resistir el suero, catabolizar el ácido siálico y resistir la mayoría de los antibióticos probados, así como codificar variaciones en cuatro de los seis CAA. Por otro lado, OH0008, aislado del mismo sitio IRLA0186 (ANI 98,3%), es sensible tanto al suero como a la mayoría de los antibióticos que probamos, pero no puede crecer en ácido siálico y solo codifica una variación alélica similar a los CAA, lo que sugiere una probabilidad limitada de resultados patogénicos.
Factores ambientales asociados con la divergencia de cúmulos
Nuestros análisis revelaron distintas firmas genómicas y fenotípicas vinculadas con la aparición de rasgos clínicos asociados en V. vulnificus ambiental. Con el fin de descubrir los impulsores ecológicos que conducen a la selección de estos rasgos y la distribución sesgada de los grupos de V. vulnificus, investigamos los parámetros abióticos y bióticos asociados con cada sitio. Primero, medimos varios factores abióticos de las muestras acuáticas recolectadas durante el aislamiento de la cepa, como la temperatura, el oxígeno disuelto, el pH, la materia orgánica disuelta, la salinidad y el fósforo, entre otros (Apéndice SI, Tabla S5). A continuación, las muestras de agua se filtraron secuencialmente a través de filtros de tamaño de poro de 20, 5 y 0,22 μm. El
ADN se obtuvo del filtro de 0,22 μm que contiene la fracción microbiana de vida libre para analizar la estructura de la comunidad microbiana (factores bióticos) asociada con cada sitio de muestreo (Fig. 5 A). Utilizamos el análisis de coordenadas principales para examinar posibles correlaciones entre la distribución de los grupos y los factores abióticos (parámetros fisicoquímicos) y bióticos (clasificación taxonómica a partir de fragmentos metagenómicos del gen del ARN ribosomal [ARNr] 16S) (Fig. 5 B). La estructura de la comunidad del Sitio A es muy similar a la encontrada en ambientes marinos donde los taxones principales fueron Cyanobacteria, SAR11, Bacteriodetes, Oceanospirillales o “Candidatus actinomarina” (Fig. 5 A). De hecho, la salinidad en esta ubicación fue de 29 ppm, que es ligeramente inferior a la del agua de mar (35 ppm) (Apéndice SI , Tabla S5). El porcentaje de lecturas de ARNr 16S asociadas con el género Vibrio representó un total de 1.8% de la población total (Fig. 5 A). Sin embargo, son indetectables en el Sitio B, donde la salinidad fue mucho menor que en el Sitio A (5 a 18 ppm), que son características de un ambiente salobre. También encontramos en el Sitio B concentraciones más altas de fosfatos, nitratos y materia orgánica disuelta en comparación con el Sitio A, lo que probablemente se deba a las escorrentías del cercano lago Okeechobee, que experimenta una afluencia de fertilizantes de las granjas agrícolas cercanas (Apéndice SI , Tabla S5). Estas variaciones en los factores ambientales probablemente cambian la comunidad microbiana al predominar microbios adaptados a la baja salinidad, como los géneros Polynucleobacter y Limnohabitans dentro de la familia Burkholderiales o las familias Microtrichal y Frankial dentro del orden Actinobacteria (Fig. 5 A). La diversidad microbiana, medida como el índice de Shannon, indicó que la diversidad era mayor en el Sitio A que en el Sitio B (Fig. 5 C). Estos datos sugieren que los miembros de C1 prefieren un ambiente más oligotrófico, parecido al marino, con mayor salinidad y mayor diversidad microbiana dominada por cianobacterias, mientras que los miembros de C2 parecen estar mejor adaptados a ambientes salobres ricos en nutrientes marcados por la presencia de varias familias de Actinobacteria (Fig. 5).
En general, nuestros análisis metagenómicos
y fisicoquímicos de los sitios de muestreo indican que la marcada distribución de grupos y los rasgos genéticos están fuertemente asociados con distintos factores bióticos y abióticos (por ejemplo, salinidad, nutrientes o biodiversidad), lo que revela cómo los ecosistemas pueden generar presiones selectivas que facilitan la aparición de cepas específicas con potencial patógeno en una población.
Conclusiones
Elucidar los factores asociados con la aparición y propagación de patógenos humanos es fundamental para desarrollar herramientas que permitan predecir posibles fuentes de brotes de enfermedades y establecer estrategias de vigilancia eficaces. La aparición de patógenos es un fenómeno complejo y multifactorial que requiere métodos y herramientas analíticas que puedan considerar grandes cantidades de datos muy diversos. Por lo tanto, es esencial desarrollar sistemas modelo manejables que nos permitan diseccionar los impulsores ecológicos, genéticos y evolutivos que fomentan la selección de rasgos de virulencia y clones patógenos dentro de una población ambiental.
En este estudio, utilizamos V. vulnificus, un patógeno costero emergente que causa sepsis mortal, como sistema modelo para investigar las fuerzas genéticas y ecológicas que conducen a la aparición de patógenos. La alta plasticidad genómica de V. vulnificus , junto con los resultados inesperados asociados con los cambios ambientales provocados por el hombre, hacen de esta bacteria una gran amenaza para la salud humana para la que no hay vacunas efectivas ni estrategias terapéuticas disponibles (16, 28, 74). En este estudio, implementamos un enfoque holístico que combina campos como la genómica, la metagenómica, la ecología, la biología molecular y la patogénesis bacteriana para abordar este problema. En general, encontramos una fuerte correlación entre los factores ecológicos (por ejemplo, el sitio de aislamiento, los parámetros fisicoquímicos y la estructura de la comunidad) y el potencial patógeno, como lo ejemplifica la distribución sesgada de los grupos y los rasgos genéticos y fenotípicos asociados con los resultados clínicos.
Las capas de selección impuestas por los
diferentes factores abióticos y bióticos probablemente actúan como una importante presión selectiva que impulsa el desarrollo de características patógenas en las poblaciones de V. vulnificus. A partir de nuestros análisis, existe una clara asociación entre la distribución de grupos y factores abióticos (p. ej., salinidad o nutrientes disueltos) y bióticos (estructura de la comunidad, ostras o cianobacterias). Dada su relevancia, investigar la asociación de V. vulnificus y el papel específico de estos y otros factores abióticos y reservorios bióticos, como protistas (p. ej., amebas) y otros metazoos (p. ej., peces y crustáceos), en la selección de grupos arrojará luz sustancial sobre el proceso de aparición de rasgos patógenos en V. vulnificus
Además, cada sitio de muestreo está expuesto a diferentes influencias antropogénicas. Por ejemplo, el Sitio A está ubicado en un área protegida con acceso limitado en Cabo Cañaveral. Por otro lado, el Sitio B experimenta un enriquecimiento excesivo de nutrientes debido a la urbanización y la expansión agrícola, así como a otra contaminación provocada por el hombre, como las descargas de desechos fecales. Dadas las diferencias drásticas en la exposición antropogénica entre los dos lugares, es probable que desempeñen un papel en la selección y distribución de los grupos. Sería interesante que los estudios futuros abordaran el papel de estas perturbaciones antropogénicas en la aparición de Vibrios patógenos.
En general, nuestros resultados indican cómo los ecosistemas pueden generar presiones selectivas que facilitan la aparición y selección de cepas específicas dentro de una población con potencial patógeno. Nuestro estudio se alinea estrechamente con la iniciativa One Health (75) al 1) centrarse en la conexión entre un agente patógeno y los factores ambientales que conducen a su aparición y 2) crear un enfoque combinado para comprender la aparición de enfermedades desde una perspectiva integrada y manejable. Nuestro enfoque puede servir para desarrollar marcadores ecológicos y genéticos para sistemas de vigilancia para predecir fuentes de brotes o identificar patógenos humanos emergentes. En general, ofrecemos un paradigma y una
metodología generales para estudiar y comprender la aparición de enfermedades que se pueden extender naturalmente a otros patógenos humanos.
Materiales y métodos
Cepas y condiciones de cultivo
Una versión extendida de los Materiales y Métodos se puede encontrar como parte del Apéndice SI . Las cepas de V. vulnificus (Apéndice SI, Tablas S1 y S3) se cultivaron rutinariamente en placas de agar Luria-Bertani (LB) suplementadas con 2% NaCl (peso/vol; LB-2%), inoculadas en caldo LB-2% y cultivadas durante 16 h aeróbicamente a 37 °C, a menos que se especifique lo contrario. Las cepas de V. vulnificus CMCP6 y SS108-A3A se utilizaron como controles clínicos C1 y ambientales C2, respectivamente, para todos los ensayos fenotípicos. E. coli β2155, un auxótrofo del ácido diaminopimélico (DAP), se utilizó para la construcción del mutante y se cultivó en LB suplementado con 0,3 mM de DAP (LBDAP).
Sitios de muestreo
Las muestras se recolectaron en dos lugares ambientalmente distintivos a lo largo de la IRL (este de Florida, Estados Unidos) en tres eventos de muestreo. El primer lugar, Fellers House Field Station (N28°54’25.315”; W80°49’15.017”; norte de la IRL; sitio A), se encuentra dentro de la Costa Nacional Cañaveral protegida por el gobierno federal. El segundo sitio de muestreo, Shepard Park, se encuentra en Port St. Lucie (N27°11’48.864”; W80°15’33.172”: sur de la IRL; sitio B), que, debido a la urbanización y la expansión agrícola, experimenta un enriquecimiento excesivo de nutrientes, lo que lleva a una floración excesiva de macroalgas (Fig. 1 B) (76 , 77).
Aislamiento de V. vulnificus de fuentes ambientales
Muestras de agua
V. vulnificus se aisló de muestras de agua utilizando un protocolo modificado de Huq et al. (42). Los 500 mL de cada muestra se filtraron sucesivamente a través de filtros de membrana de 20, 5 y 0,2 µm (Sterlitech) para separar las fracciones planctónicas y de vida libre. Los filtros se suspendieron en solución salina tamponada con fosfato (PBS), pH 7,5, se agitaron vigorosamente y se cultivaron en
agua de peptona alcalina (APW) durante la noche a 37 °C.
Muestras de sedimentos
V. vulnificus se aisló del sedimento utilizando un protocolo modificado de Schuster et al. (78). Las muestras se recogieron utilizando un sacabocados universal. Las muestras se suspendieron en PBS (1:1) y se homogeneizaron y enriquecieron en APW. Muestras de ostras.
El aislamiento de V. vulnificus de las ostras se llevó a cabo mediante un protocolo adoptado y modificado del Manual de análisis bacteriológico de Vibrio de la Administración de Alimentos y Medicamentos de los Estados Unidos (79). Brevemente, las ostras recolectadas de la casa de Feller se lavaron para eliminar los sedimentos o la suciedad. Cada ostra se descascaró individualmente, se homogeneizó en 30 ml de PBS utilizando el homogeneizador SCILOGEX D160 y se cultivó en APW.
Muestras de cianobacterias
Las cianobacterias recolectadas en Shepard Park se sedimentaron, se eliminó el sobrenadante y se cultivaron en agua destilada. Todas las muestras se recolectaron por triplicado. Los cultivos enriquecidos en agua destilada de muestras de agua, sedimento, ostras y cianobacterias se diluyeron en serie y se sembraron en CaV (CHROMagar), un agar selectivo para Vibrio spp. Las colonias de color azul turquesa se analizaron posteriormente en placas de agar TCBS (Sigma) en las que V. vulnificus aparece como colonias verdes. Las colonias que aparecieron de color azul turquesa en CaV y verde en TCBS se consideraron posibles aislamientos de V. vulnificus
Verificación de aislamientos de V. vulnificus
Los posibles aislamientos de V. vulnificus en la vida real se verificaron mediante PCR utilizando iniciadores para el gen marcador thiF (Apéndice SI , Tabla S2). Los productos de PCR de los aislamientos positivos para thiF se secuenciaron (GENEWIZ, AT, GA) para determinar la afiliación del grupo. Se seleccionaron varios aislamientos diversos de V. vulnificus de ambos grupos y de cada uno de los reservorios ambientales para la secuenciación del genoma completo. Secuenciación del genoma
Las bibliotecas de genomas completos se prepararon utilizando el kit de preparación de bibliotecas Nextera DNA Flex de Illumina siguiendo las instrucciones del fabricante y se secuenciaron utilizando el sistema de secuenciación iSeq100 de Illumina. Los genomas secuenciados se analizaron utilizando el concentrador de secuencias Illumina BaseSpace. Las lecturas obtenidas para cada biomuestra se ensamblaron en contigs y estructuras utilizando el ensamblador de genomas SPAdes versión 3.9.0 y el ensamblador de novo Velvet versión 1.0.0.
Ensamblaje, predicción genética y anotación
Las lecturas se recortaron utilizando Trimmomatic v0.36 (80) y se ensamblaron de novo con SPAdes versión 3.11.1 (81). Los ORFs de los contigs ensamblados se predijeron utilizando Prodigal versión 2.6 (82). Los genes de ARN de transferencia y ARNr se predijeron utilizando tRNAscan-SE versión 1.4 (83), ssu-align versión 0.1.1 (84) y meta-rna (85). Utilizando DIAMOND (86), las proteínas predichas se compararon con la base de datos no redundante del Centro Nacional de Información Biotecnológica, y se compararon con COG (87) y TIGFRAM (88) utilizando HMMscan versión 3.1b2 (89) para la anotación taxonómica y funcional.
Reconstrucciones filogenómicas
A los contigs ensamblados se les asignó un cromosoma por comparación con este grupo de genomas de referencia usando Blastn (90). Los genes se predijeron usando Prodigal (82) y se agruparon usando el software MMsEqs (91). Los grupos de proteínas resultantes que estaban presentes en todos los genomas analizados se dividieron en dos grupos según el cromosoma en el que están codificados, lo que resultó en un grupo de 257 y 62 proteínas para los cromosomas 1 y 2, respectivamente. Luego, los grupos de proteínas se alinearon con QuickProbs2 (92), se recortaron con BGME (93) y se concatenaron. Finalmente, se construyó un árbol filogenético usando iqtree (94) con selección automática de modelos y 1000 réplicas bootstrap.
Comparaciones genómicas por pares
Se realizaron búsquedas recíprocas BLASTN y TBLASTX entre genomas, lo que permitió identificar regiones de similitud, inserciones
y reordenamientos. El ANI y la cobertura entre pares de genomas se calcularon utilizando el software PYANI (95).
Análisis de pangenoma y recombinación
Para analizar la prevalencia de familias de genes en todos los genomas, utilizamos el software PPanGGOLiN para dividir las familias de genes en particiones persistentes/ de capa/de nube (52). Los genes que constituyen cada partición se anotaron luego en la base de datos del subsistema SEED (96) utilizando DIAMOND (86), manteniendo todas las coincidencias con E < 0,001 y longitud de alineamiento > 0,5 tanto para el sujeto como para la consulta. Finalmente, los valores de dN/dS para las diferentes particiones de proteínas se obtuvieron utilizando el paquete Orthologr en R (97). La tubería PopCOGenT (47) se utilizó para definir las poblaciones recombinantes basadas en el flujo de genes entre los diferentes genomas secuenciados. Construcción mutante
Las deleciones en marco de los genes de interés, wza , nanA y siaM , se construyeron mediante recombinación homóloga (98) (la lista de cebadores se puede encontrar en el Apéndice SI , Tabla S6). Brevemente, dos fragmentos de PCR de aproximadamente 500 pb que flanqueaban los genes de interés se clonaron en el plásmido contraseleccionable sacB , pDS132, y se electroporaron en la cepa donante de E. coli , β2155. Las cepas donantes que albergaban los vectores knockout se conjugaron con CMCP6 de V. vulnificus de tipo salvaje en LBDAP, y los transconjugantes se seleccionaron en placas LB-2% suplementadas con cloranfenicol (Cm) (25 μg/mL). Las colonias exconjugadas de Cm R se cultivaron en LB2% sin antibióticos y se realizaron diluciones seriadas en placas de LB-2% que contenían sacarosa al 10 % (peso/volumen). Se examinaron los posibles mutantes de deleción de doble cruce mediante PCR y las supuestas deleciones se confirmaron mediante secuenciación de ADN.
Resistencia antibiótica
Los aislamientos de V. vulnificus se examinaron para determinar su susceptibilidad a los antibióticos destacados en la Fig. 4 en las concentraciones más altas en el rango de concentración de punto de corte recomendado por el Clinical and Laboratory Standards Institute en M45-A (99-
102) (SI Apéndice , Texto complementario). Brevemente, las colonias individuales de cada cepa se transfirieron secuencialmente utilizando palillos de dientes estériles a placas LB-2% suplementadas con los respectivos antibióticos e incubadas a 37 °C durante la noche. Se midió el diámetro del crecimiento y la resistencia se definió como un crecimiento de al menos 2 mm en los respectivos antibióticos. Las cepas que no mostraron crecimiento se tomaron como sensibles y cualquier diámetro de crecimiento intermedio se consideró como resistencia intermedia. Los experimentos se realizaron en tres réplicas biológicas independientes.
Resistencia al suero
El ensayo de supervivencia del suero in vitro se adaptó de Bogard y Oliver (103). Brevemente, las células se subcultivaron durante la noche en LB-2% para obtener células en fase logarítmica a una densidad óptica (DO) de 0,15 a 0,25. Luego, las células se lavaron en PBS y se inocularon a una dilución de 100 veces en suero humano normal combinado (Fisher Bioreagents) y se incubaron a 37 °C durante 2 h. La resistencia al suero se evaluó comparando las UFC/ mililitros antes y después de la exposición al suero. Los experimentos se realizaron en tres réplicas biológicas independientes.
Catabolismo del ácido siálico
La capacidad de catabolizar el ácido siálico por los aislados de V. vulnificus se evaluó mediante el crecimiento en NANA, la forma predominante de ácido siálico en células humanas, como única fuente de carbono (57). Brevemente, los cultivos nocturnos de cada cepa se lavaron y resuspendieron en medio mínimo M9, y se realizó una dilución de 100 veces de las células en medio mínimo M9 suplementado con NANA (2 mg/ ml) (Chem-Impex International). Las alícuotas de 200 µL de cada muestra se agregaron por pocillo a una microplaca de 96 pocillos y se incubaron a 37 °C con agitación. La DO a 595 nm (OD595) se midió cada hora durante 24 h utilizando un lector de microplacas Tecan Sunrise (Tecan US), y los resultados se evaluaron utilizando el software de lectura de placas Magellan. Los ensayos de crecimiento se realizaron por triplicado en tres réplicas biológicas independientes.
Medición de parámetros fisicoquímicos Durante los aislamientos se realizaron mediciones de temperatura del agua (°C), salinidad (gramos/litros), oxígeno disuelto (%), pH, presión (milímetros de mercurio), materia orgánica disuelta, clorofila-a (microgramos/ litros) y algas totales (microgramos/litros). Las mediciones se registraron utilizando una sonda YSI EX02 desplegada en los sitios en el momento del muestreo que se calibró dentro de las 24 h previas a cada evento de muestreo. Las muestras de agua, recolectadas por triplicado, también se examinaron para determinar la concentración de fosfatos (o -Fosfato–P, método 365.1) y nitratos (Nitrato–N, método 353.2; Amoniaco–N, método 350.1) de acuerdo con los protocolos estándar descritos por la Agencia de Protección Ambiental de los Estados Unidos (104 , 105). Brevemente, las muestras de agua recolectadas se filtraron a través de un filtro de membrana de 0,2 µm y se acidificaron a un pH < 2 con H 2 SO 4 bidestilado y se almacenaron a 4 °C hasta su análisis. Las muestras se analizaron para detectar nitrato + nitrito (NO3−), amonio (NH4+) y ortofosfato (PO43−) en un analizador discreto automatizado SEAL AQ2 (Seal Analytical).
Análisis metagenómico
La extracción de ADN se realizó a partir del filtro de 0,22 μm. Las células adheridas se
rompieron utilizando un tampón de lisis de bromuro de cetiltrimetilamonio (CTAB) y perlas de vidrio seguido de un tratamiento con lisozima. Luego, los ácidos nucleicos se extrajeron utilizando el método de extracción de fenol-cloroformo (106). Los metagenomas se secuenciaron utilizando Illumina Hiseq-4000 (150 pb, lectura de extremos emparejados). Para analizar la clasificación filogenética de las muestras, las secuencias candidatas del gen 16S rRNA en los metagenomas sin procesar se identificaron utilizando USEARCH6 (107) (valor E < 10 a 5) contra una base de datos que contenía secuencias de 16S rRNA no redundantes descargadas del Ribosomal Database Project (108). Estas secuencias se alinearon luego con los modelos ocultos de Markov (HMM) del ARNr 16S de arqueas y bacterias (109) utilizando ssu-align para identificar secuencias verdaderas (84). Solo los resultados de las secuencias del ARNr 16S se clasificaron en un taxón de alto nivel si la identidad de secuencia era ≥80% y la longitud de alineación ≥90 pb•
Para más información sobre este artículo, contactar a: mario.lopezp@umh.es
Evaluación de fitoesteroles como alternativa al colesterol en dietas prácticas sobre el crecimiento y la inmunidad no específica de Penaeus vannamei
El colesterol es una de las fuentes de lípidos esenciales para los animales, junto con los fosfolípidos y los ácidos grasos, y conforma el componente integral de las membranas celulares [1]. Además, es un constituyente lipídico no polar que actúa como precursor de algunas hormonas cruciales de crecimiento y reproducción [2]. Más importante aún, el colesterol es un componente de las lipoproteínas que participan en el transporte y la absorción de lípidos en los animales [3]. Por lo general, los teleósteos nacen con la capacidad de sintetizar colesterol a partir de acetato a través del escualeno de novo. Sin embargo, aunque los crustáceos se consideran una fuente rica de colesterol, generalmente se acepta que los camarones u otros crustáceos no pueden sintetizar colesterol de novo debido a la falta de escualeno monooxigenasa y lanosterol sintasa [4]. Por lo tanto, los crustáceos deben obtener colesterol o esteroles de su dieta para satisfacer los requisitos de crecimiento, muda y reproducción.
Como una alternativa potencial para el colesterol, los fitoesteroles son esteroles de origen vegetal y constituyen principalmente β-sitosterol, campesterol y estigmasterol [5]. Se ha informado que los fitoesteroles pueden ser un sustituto parcial o completo del colesterol en el alimento de Penaeus vannamei [6]. En un estudio previo, la suplementación de 1,2 g/kg de fitoesteroles en dietas semipurificadas aumentó significativamente el peso final y la tasa de ganancia de peso de P. vannamei [7]. También se informa que 1,0 g/kg de fitoesteroles dietéticos satisface el requerimiento de esta especie [8].
En general, se cree que los fitoesteroles reducen el colesterol plasmático al inhibir la absorción de colesterol por las células intestinales y promover su excreción, lo que se considera el efecto reductor del colesterol [9]. Por otra parte, los fitoesteroles también funcionan como antioxidantes y juegan un papel positivo en la inmunidad [10, 11]. Según un estudio previo, los fitoesteroles dietéticos pueden aliviar el estrés oxidativo inducido por una dieta alta en lípidos en la corvina amarilla grande (Larimichthys crocea), activando las actividades de la catalasa y la superóxido dismutasa [12]. Además, los fitoesteroles dietéticos aumentaron los contenidos
totales de inmunoglobulina plasmática y regularon positivamente las expresiones genéticas de β-defensina , péptido antimicrobiano de hepcidina 1 , interleucina-10 e interleucina-18 del pez dorada (Sparus aurata) después de los 48 días de prueba de alimentación [13], lo que implica el efecto de inmunopotenciación del fitosterol.
El objetivo de este estudio fue evaluar preliminarmente el efecto sobre el crecimiento y la inmunidad no específica del camarón después de que se suplementaran en las dietas diferentes fuentes y niveles de esteroles. El camarón blanco del Pacífico (Penaeus vannamei), que ganó popularidad debido a su rápida tasa de crecimiento, alta supervivencia, sabor único y abundancia de nutrientes, se ha empleado en este estudio. Con el aumento de los componentes de origen vegetal en dietas prácticas, como la harina o el aceite de soja, el camarón de cultivo puede crecer lentamente debido a la falta de colesterol [5]. Además, los estudios sobre la respuesta inmune de los fitoesteroles dietéticos se han realizado principalmente en los teleósteos, mientras que los estudios
sobre la inmunidad no específica de P. vannamei aún son limitados. Dado que el camarón depende principalmente de la inmunidad innata en lugar de la inmunidad adaptativa, los fitoesteroles dietéticos pueden tener diferentes efectos sobre la respuesta inmune del camarón. Por lo tanto, se realizó un ensayo de alimentación para demostrar la eficacia de los fitoesteroles en dietas prácticas de camarones mediante la comparación de diferentes fuentes y niveles de esteroles. Este estudio buscó explorar preliminarmente el efecto del fitosterol sobre el crecimiento y la inmunidad no específica del camarón.
Material y métodos
Preparación de la dieta
El colesterol se adquirió de Sigma-Aldrich (EE.UU.) y el fitosterol (constituido por 28,1% de campesterol, 17,2% de estigmasterol, 43,9% de β-sitosterol) fue proporcionado por Guangdong Wei Lai Biotechnology Co., Ltd. (Guangzhou, China). Las principales fuentes de proteína estaban compuestas de harina de carne y huesos, harina de pescado y concentrado de proteína de soja. Las fuentes de lípidos se elaboraron con aceite
de pescado, aceite de soja y lecitina de soja. Con base en un estudio previo, la dieta de control se formuló para contener 1 g/kg de colesterol (LC (colesterol bajo)) [7] y se utilizó fitosterol para reemplazar el colesterol en la dieta LP (bajo en fitosterol). En las otras tres dietas experimentales, la suplementación de colesterol se incrementó a 2 g/kg (HC (colesterol alto)) y luego, el colesterol se reemplazó parcialmente (CP, 1 g/kg de colesterol + 1 g/kg de fitosterol) o totalmente (HP (fitosterol alto)) con fitoesteroles.
Para preparar las cinco dietas, todos los ingredientes se molieron y tamizaron a través de una malla de 80, se pesaron y se mezclaron hasta homogeneidad (M256, Universidad Tecnológica del Sur de China, China) [14]. Los pellets de 1,5 mm de diámetro se extruyeron utilizando un peletizador (Instituto de Ingeniería Química, Universidad Tecnológica del Sur de China, Guangdong, China), se calentaron a 60 °C durante 30 min y se almacenaron a -20 °C antes de su uso. La formulación y los niveles de nutrientes de las dietas se muestran en la Tabla 1
El fitosterol fue proporcionado por Guangdong Wei Lai Biotechnology Co., Ltd. (Guangzhou, China) y constituyó el 28,1% de campesterol, el 17,2% de estigmasterol, el 43,9% de β-sitosterol y otros2. Cada 1 kg de premezcla de vitaminas contiene vitamina A 4.200.000 UI, D3 2.200.000 UI, E 35 g, K3 10 g, B1 4,5 g, B2 16 g, B6 9 g, B12 0,02 g, pantotenato de calcio 28 g, ácido fólico 2,5 g, biotina 0,08 g, ácido nicotínico 38 g e inositol 145 g3. Cada 1 kg de premezcla mineral contiene MgSO 4 ·H2O 13 g, KCl 85 g, Met-Cu 3 g, FeSO 4 ·H2O 1 g, ZnSO 4 ·H2O 12 g, Ca(IO3) 2 0,06 g, Met-Co 0,16 g y NaSeO 3 0,003 6 g. 4 Los niveles de nutrientes fueron valores medidos.
Tabla 1. Composición y niveles de nutrientes de las dietas (base seca g/kg).
Cría de camarones y condiciones experimentales
Los camarones juveniles se obtuvieron de Doumen (Zhuhai, China). Los camarones fueron alimentados con alimento comercial y aclimatados a las condiciones experimentales antes de ser utilizados. Durante la distribución de los camarones, los 750 animales de tamaño uniforme y saludables (peso inicial 0,52 ± 0,008 g) se dividieron en 5 grupos por triplicado, con 50 camarones en un tanque de fibra de vidrio de 500 L. Normalmente, los camarones se alimentaron tres veces al día a las 8:30, 13:00 y 18:00, con una tasa de alimentación total del 3%-8% del peso del camarón por día [15]. En caso de clima extremo, como tifones y tormentas eléctricas, la alimentación puede reducirse o incluso suspenderse. La prueba de alimentación duró 60 días y se llevó a cabo en un sistema de cultivo de agua recirculante al aire libre. Durante el experimento, se detectó continuamente la calidad del agua. La temperatura del agua se mantuvo entre 25 y 32 °C, el oxígeno disuelto >5 mg/L, la salinidad al 5% y la ventilación fue continua. El pH fluctuó entre 7,7 y 8,5. La concentración de nitrógeno amoniacal fue inferior a 0,2 mg/L.
Recolección y análisis de muestras
Al final de la prueba de alimentación, los camarones se dejaron sin alimento durante 24 h para obtener perfiles metabólicos basales. Los camarones en cada tanque se contaron y pesaron para calcular los indicadores de crecimiento. Se recolectaron ocho camarones de cada tanque para el análisis de la composición corporal total. La hemolinfa se recolectó de 15 camarones por tanque utilizando una jeringa estéril de 1 ml y luego se colocó a 4 °C durante 1 h [16]. Después de la centrifugación a 4 °C, 8000 rpm durante 15 min, se obtuvo el sobrenadante y se almacenó a -80 °C antes de la determinación de los parámetros bioquímicos de la hemolinfa. El hepatopáncreas se extrajo de 4 camarones por tanque como un grupo de muestra y se almacenó a -80 °C antes del ensayo de actividad enzimática.
La humedad de las dietas y los camarones se determinó mediante secado en horno a 105 °C de acuerdo con el estándar de GB/T 6435-2014 en China (reducción de peso del
alimento después del secado). La proteína cruda se detectó mediante el analizador Primacs100 (Skalar, Países Bajos) como el estándar de GB/T 18868-2002 en China (proteína cruda = total − N × 6,25). Los lípidos crudos se detectaron mediante un extractor XT15 (ANKOM, EE.UU.) de acuerdo con el estándar de GB/T 6433-2006 en China (reducción de peso del alimento después de la extracción con éter de petróleo). Las cenizas se detectaron quemándolas a 550 °C de acuerdo con el estándar de GB/T 6438-2007 en China (reducción de peso del alimento después de la quema completa) [17]. La determinación del contenido de colesterol se siguió de acuerdo con GB 5009.128-2016 en China. Brevemente, se saponificaron muestras de 1,0 g con etanol anhidro-hidróxido de potasio al 60% en solución (3:1, v:v) y se extrajeron con éter de petróleo y éter anhidro (1:1, v:v). Después de concentrar los extractos a sequedad, se disolvieron en etanol anhidro y se detectaron mediante cromatografía de gases.
Parámetros bioquímicos de la hemolinfa Los indicadores bioquímicos de la hemolinfa incluyen triglicéridos (TG, artículo n.º A1101-1), colesterol total (T-CHO, artículo n.º A111-1-1), colesterol de lipoproteína de baja densidad (LDL-C, artículo n.º A113-
1-1), colesterol de lipoproteína de alta densidad (HDL-C, artículo n.º A112-1-1) y malonaldehído (MDA, artículo n.º A003-12) [18]. Se analizaron las actividades de la superóxido dismutasa de la hemolinfa (SOD, artículo n.º A001-3-2), la fenol oxidasa (PO, artículo n.º A136-1-1) y la lisozima (LZM, artículo n.º A050-1-1) [19]. Todos los parámetros de la hemolinfa se determinaron mediante kits comerciales (Nanjing Jiancheng Bioengineering Institute, China) siguiendo las instrucciones y monitoreando los cambios de absorbancia mediante un lector de microplacas de longitud de onda completa (Thermo, Multiskan GO 1510, EE. UU.) [20].
Parámetros bioquímicos del hepatopáncreas
Se determinaron las actividades de la aspartato aminotransferasa (AST, artículo n.º C010-2-1), la alanina transaminasa (ALT, artículo n.º C009-2-1) y la fosfatasa alcalina (AKP, artículo n.º A059-2-2) [21].
Se analizaron las actividades de la proteasa del hepatopáncreas (artículo n.º A080-2-2), la amilasa (artículo n.º C016-1-1) y la lipasa (artículo n.º A054-2-1). Todos los parámetros del hepatopáncreas se determinaron mediante kits comerciales (Nanjing Jiancheng Bioengineering Institute, China)
donde t es la duración experimental en días. Los resultados se presentaron como medias con SEM. Los datos se compararon entre tratamientos mediante análisis de varianza unidireccional (ANOVA unidireccional) utilizando el software estadístico SPSS 21.0 (SPSS, Chicago, IL, EE. UU.). Cuando las diferencias generales fueron significativas ( P < 0,05), se utilizó la prueba de Duncan para comparar los valores medios entre tratamientos individuales. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de Student-Newman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles. Un valor de probabilidad de P < 0,05 se consideró estadísticamente significativo.
siguiendo las instrucciones y monitoreando los cambios de absorbancia mediante un lector de microplacas de longitud de onda completa (Thermo, Multiskan GO 1510, EE. UU.) [22].
Cálculos y análisis estadístico
El rendimiento del crecimiento se calculó de la siguiente manera [23]: (1) Donde t es la duración experimental en días.
Los resultados se presentaron como medias con SEM. Los datos se compararon entre tratamientos mediante análisis de varianza unidireccional (ANOVA unidireccional) utilizando el software estadístico SPSS 21.0 (SPSS, Chicago, IL, EE.UU.). Cuando las diferencias generales fueron significativas
(P<0,05), se utilizó la prueba de Duncan para comparar los valores medios entre tratamientos individuales. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de Student-NewmanKeuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles. Un valor de probabilidad de P<0,05 se consideró estadísticamente significativo.
Resultado
Efectos de las fuentes de esteroles en la dieta sobre el crecimiento del camarón Como se muestra en la Tabla 2 , la suplementación con niveles más altos de esteroles ayudó a mejorar el rendimiento de crecimiento del camarón. Para ser específicos, el peso corporal final (FBW), la
2. Rendimiento del crecimiento de camarones alimentados con diferentes dietas.
tasa de ganancia de peso (WGR) y la tasa de crecimiento específico (SGR) del camarón aumentaron significativamente después de la suplementación del alimento con 2 g/kg de colesterol o fuentes de esteroles mixtas (P<0,05). No hubo diferencias significativas en los indicadores de rendimiento de crecimiento en los camarones alimentados con la dieta HP en comparación con los alimentados con las dietas HC y CP (P>0,05). Además, la tasa de conversión alimenticia (FCR) disminuyó significativamente después de la suplementación del alimento con 2 g/kg de fuentes de esteroles en los grupos HC, HP y CP (P<0,05). La supervivencia no se correlacionó con la fuente o el nivel de esteroles y no difirió significativamente entre los cinco grupos (P>0,05).
Los datos representan la media ± SEM de tres réplicas (n=3). Los valores en la misma línea con letras diferentes son significativamente diferentes (P<0,05) según la prueba de Duncan. La falta de letras en superíndice indica que no hay diferencias significativas entre los tratamientos. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de Student-Newman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles. FBW = peso corporal final; WGR = tasa de ganancia de peso; SGR = tasa de crecimiento específico; FCR = índice de conversión alimenticia.
Tabla 3. Composición corporal e indicadores físicos de camarones alimentados con diferentes dietas.
Los datos representan la media ± SEM de tres réplicas (n=3). Los valores en la misma línea con letras diferentes son significativamente diferentes (P<0,05) según la prueba de Duncan. La falta de letras en superíndice indica que no hay diferencias significativas entre los tratamientos. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de Student-Newman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles.
Tabla
Tabla 4. Parámetros bioquímicos de la hemolinfa de camarones alimentados con diferentes dietas.
Los datos representan la media ± SEM de tres réplicas (n=3). Los valores en la misma línea con letras diferentes son significativamente diferentes (P<0,05) según la prueba de Duncan. La falta de letras en superíndice indica que no hay diferencias significativas entre los tratamientos. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de Student-Newman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles. TG = triglicéridos; T-CHO = colesterol total; LDL-C = colesterol de lipoproteínas de baja densidad; HDL-C = colesterol de lipoproteínas de alta densidad; MDA = malonaldehído.
Efectos de las fuentes de esteroles en la dieta sobre la composición corporal y los indicadores físicos de los camarones
Como se muestra en la Tabla 3 , la proteína cruda del camarón disminuyó debido a los bajos niveles de esteroles en la dieta. El contenido de proteína cruda del camarón alimentado con las dietas HC y CP fue significativamente mayor que el de los grupos LC y LP (P<0,05). El tipo de fuentes de esteroles tuvo un efecto significativo en el contenido de lípidos crudos, con una disminución del cual en los grupos LP y HP en comparación con los otros tres grupos (P<0,05 en el ANOVA de dos vías).
El contenido de cenizas del camarón entero también aumentó cuando se complementó con niveles más altos de esteroles en la dieta, y las cenizas del camarón alimentado con las dietas HC, HP y CP aumentaron significativamente en comparación con los alimentados con las dietas LC y LP (P<0,05). Ni la fuente ni el nivel de esteroles tuvieron efectos significativos en el factor de condición y el contenido de carne del camarón (P >0,05).
Efectos de las fuentes de esteroles en la dieta sobre los parámetros bioquímicos de la hemolinfa del camarón
Los resultados de los parámetros bioquímicos de la hemolinfa del camarón se muestran en la Tabla 4 Tanto las fuentes de esterol como los niveles, así como sus interacciones, tuvieron efectos significativos en los triglicéridos (TG) y el colesterol total
Figura 1
(T-CHO) de la hemolinfa del camarón. Específicamente, los TG y T-CHO de la hemolinfa en los camarones alimentados con dietas HC fueron significativamente más altos que en otros grupos (P<0,05), mientras que los de los camarones suplementados con fitosterol dietético mostraron una tendencia a la reducción en comparación con los grupos LC y HC. La fuente de esterol tuvo efectos significativos en el contenido de colesterol de lipoproteína de alta densidad (HDL-C) de la hemolinfa y malonaldehído (MDA), con el primero aumentado y el segundo disminuido en los
camarones suplementados con fitosterol dietético (grupos LP, HP y CP) (P<0,05). Ni la fuente de esterol ni el nivel tuvieron efectos significativos en el contenido de colesterol de lipoproteína de baja densidad (LDL-C) de la hemolinfa del camarón (P>0,05).
Efectos de las fuentes de esteroles en la dieta sobre la inmunidad no específica y los indicadores de salud en la hemolinfa y el hepatopáncreas del camarón
Los resultados de las actividades enzimáticas relacionadas con la inmunidad no específica y la salud se muestran en la Figura 1. La
fuente de esterol tuvo efectos significativos en las actividades de la superóxido dismutasa (SOD) y la lisozima (LZM) de la hemolinfa, con la actividad de SOD de los camarones alimentados con las dietas HP y CP, así como la LZM de los camarones alimentados con la dieta CP, aumentada significativamente en comparación con los grupos de colesterol (P<0,05). Tanto las fuentes de esterol como los niveles, así como sus interacciones, tuvieron efectos significativos en la actividad de la fenol oxidasa (PO) de la hemolinfa, que fue impulsada por el fitosterol dietético y un mayor nivel de inclusión de esterol (P<0,05). En el hepatopáncreas, se demostró que la actividad de la fosfatasa alcalina (AKP) aumentó significativamente en los camarones suplementados con un nivel de esterol de 2 g/kg (P<0,05). La interacción de las fuentes y los niveles de esteroles tuvo un impacto en la actividad de la aspartato aminotransferasa (AST), que aumentó significativamente en los camarones alimentados con las dietas HC y LC en comparación con los otros tres grupos (P<0,05).
Efecto de diferentes fuentes y niveles de esteroles sobre la inmunidad no específica y los indicadores de salud en la hemolinfa y el hepatopáncreas de P. vannamei . Las barras verticales representan la media ± SEM (n=3). Los datos marcados con diferentes letras difieren significativamente (P<0,05) entre los grupos según la prueba de Duncan. La falta de letras en superíndice indica que no hay diferencias significativas entre los tratamientos. Se realizó un ANOVA de dos vías seguido de una prueba de comparación múltiple de Student-Newman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles. SOD = superóxido dismutasa (U/mL); PO = fenol oxidasa (U/mL); LZM = lisozima (U/ mL); AKP = fosfatasa alcalina (U/g); ALT = alanina transaminasa (U/g); AST = aspartato aminotransferasa (U/g).
Efectos de las fuentes de esteroles en la dieta sobre las actividades de las enzimas digestivas en el hepatopáncreas del camarón
Como se muestra en la Figura 2 , tanto las actividades de proteasa como de lipasa del camarón se vieron afectadas principalmente por los niveles de esterol, y podrían mejorarse suplementando con 2 g/kg de fuentes de
esterol (dietas HC, HP y CP) en la dieta en comparación con los grupos de inclusión de esterol más bajos (1 g/kg de fuentes de esterol, dietas LC y LP) (P<0,05). También se observó que la actividad de la lipasa fue significativamente mayor en el camarón alimentado con la dieta HC que en el alimentado con las dietas HP y CP (P<0,05). La actividad de la amilasa no se vio afectada por los factores de fuente y nivel de esterol en este estudio (P>0,05).
Efecto de diferentes fuentes y niveles de esteroles sobre las actividades de las enzimas digestivas en el hepatopáncreas de P. vannamei . Las barras verticales representan la media ± SEM (n=3). Los datos marcados con diferentes letras difieren significativamente (P<0,05) entre los grupos según la prueba de Duncan. La falta de letras en superíndice indica que no hay diferencias significativas entre los tratamientos. Se realizó un ANOVA de dos vías seguido de la prueba de comparación múltiple de StudentNewman-Keuls (SNK) para investigar los efectos de interacción de las fuentes y los niveles de esteroles.
Discusión
Para los crustáceos, el colesterol dietético es esencial para la síntesis de ecdisteroides
y sesquiterpenoides, que regulan las actividades vitales, incluyendo la muda, el crecimiento y la reproducción [24]. Sin embargo, generalmente se piensa que los camarones son incapaces de sintetizar suficiente colesterol y requieren una ingesta externa, lo que aumenta los desafíos en el cultivo de camarones: ya sea mayores costos de alimentación o menor crecimiento [4]. Afortunadamente, se ha informado que algunas fuentes de lípidos dietéticos, incluidos los fitoesteroles y los fosfolípidos, funcionan como colesterol [5]. Los fitoesteroles están constituidos principalmente por campesterol, estigmasterol y β-sitosterol y tienen una cadena de carbono hidrofóbica adicional en la posición C-22 o C-24 en comparación con el colesterol [25].
De manera similar al colesterol, los fitoesteroles forman micelas con grasa, ácidos biliares y colesterol en el lumen intestinal y son absorbidos por las células epiteliales intestinales a través del transportador de esteroles Niemannpick C1-like 1. Los esteroles son luego esterificados por la isoforma 2 de la acil-CoA: colesterol aciltransferasa y se combinan con quilomicrones para viajar en la linfa y finalmente ser absorbidos por el hígado a través del receptor dependiente de ApoE
Figura 2
ALIMENTACIÓN
[26]. Estudios previos han informado que el camarón puede ser capaz de convertir fitoesteroles en colesterol in vivo, pero el proceso bioquímico detallado y la eficiencia de biodisponibilidad siguen siendo desconocidos [6, 7].
Además, los fitoesteroles funcionan reduciendo el almacenamiento de colesterol, alteran el metabolismo de los lípidos y afectan la inmunidad innata, así como la capacidad antioxidante en algunos estudios [12, 27 – 29]. Por lo tanto, es importante tener una comprensión integral de los efectos de los fitoesteroles dietéticos en el camarón.
En el presente estudio, el rendimiento del crecimiento aumentó significativamente en el camarón suplementado con 2 g/ kg de colesterol o fuentes de esteroles mixtas, lo que indicó que el aumento de la ingesta del nivel de esterol fue beneficioso para el crecimiento del camarón y que los fitoesteroles pueden reemplazar parcialmente el colesterol. Además, el FCR en el camarón alimentado con las dietas HC, HP y CP fue menor que aquellos suplementados con 1 g/kg de nivel de esterol, lo que implica que un nivel de esterol más alto (nivel de esterol de 2 g / kg) podría facilitar el nutriente utilizado por el camarón y reducir el consumo de alimento en el mismo rendimiento de crecimiento. Además, aumentar el contenido de proteína cruda del camarón entero sugirió que el mayor contenido de esterol afectó positivamente la conversión de proteína del camarón, lo que podría explicarse por la mayor actividad de la proteasa en los grupos HC y CP.
En un estudio previo, el requerimiento óptimo de colesterol del camarón fue de alrededor de 1,7-2,7 g/kg según los diferentes indicadores. Sin embargo, cuando el colesterol dietético fue menor a 0,8 g/kg, el FBW y WGR del camarón mostraron una reducción significativa en comparación con aquellos alimentados con dietas de inclusión de colesterol más altas y óptimas [7], lo que fue similar a nuestros resultados.
Varios estudios han informado sobre la seguridad y el uso potencial de fitoesteroles en alimentos acuícolas, sin efectos adversos en el crecimiento de la corvina amarilla (Larimichthys crocea), salmón del
Atlántico (Salmo salar L), lubina europea (Dicentrarchus labrax) y dorada (Sparus aurata) [12, 30-32] y efectos promotores del crecimiento en besugo negro (Megalobrama amblycephala), tilapia (Oreochromis niloticus) y lubina negra de boca grande (Micropterus salmoides) [33-36] . Aunque los fitoesteroles pueden reemplazar el colesterol dietético en el alimento para camarones, los mecanismos moleculares por los cuales los fitoesteroles facilitan el crecimiento del camarón y los efectos sinérgicos y competitivos entre los fitoesteroles y el colesterol aún no están claros, y se necesitan más estudios para arrojar luz.
Dado que los fitoesteroles tienen la función de promover el metabolismo de esteroles, tienen efectos potenciales predecibles en el metabolismo de lípidos y la inmunidad no específica en camarones [26 , 37]. En el presente estudio, se observa que el contenido de lípidos crudos de camarones enteros fue influenciado principalmente por la fuente de esteroles, con una tendencia a una disminución en el contenido de lípidos causada por los fitoesteroles dietéticos.
Además, el contenido de TG y T-CHO de la hemolinfa se vio afectado por la interacción de la fuente y los niveles de esteroles. Para ser específico, suplementar el alimento con un alto nivel de fitosterol redujo significativamente el contenido de TG y T-CHO. Los estudios también han demostrado que los fitoesteroles son capaces de reducir las concentraciones de T-CHO, TG y LDL-C por la secreción hepática de lipoproteínas de muy baja densidad (VLDL) o la conversión a ácidos biliares [38 , 39]. Estos efectos reductores de lípidos pueden estar mediados por la inhibición de los niveles de traducción del receptor alfa activado por el proliferador de peroxisomas hepáticos (PPAR-α) y la sintetasa de ácidos grasos hepática (FAS) [18 , 40 , 41]. Por lo tanto, la suplementación de 2 g/kg de fitoesteroles ha tenido como resultado un efecto reductor del colesterol en Penaeus vannamei y aún se necesita más estudios para saber cómo este efecto afecta los procesos metabólicos lipídicos en el camarón.
Además del crecimiento y el metabolismo, el efecto de los fitoesteroles dietéticos sobre la inmunidad no específica de Penaeus
vannamei merece ser estudiado, ya que aún faltan investigaciones relevantes. A diferencia de los vertebrados, el camarón lucha contra los patógenos mediante la inmunidad innata, en la que la profenoloxidasa, la fosfatasa alcalina (AKP) y la lisozima (LZM) juegan un papel notable [42 , 43]. La profenoloxidasa es capaz de generar una gran cantidad de fenol oxidasa (PO) biológicamente activa a través de una reacción en cascada y además media la síntesis de melanina [44 , 45].
Luego, las funciones de la melanina inhiben el crecimiento y la transmisión de patógenos, y su metabolito intermedio quinona puede causar efectos tóxicos sobre los patógenos, protegiendo en última instancia al camarón de la invasión [46 , 47]. También se informa que LZM y AKP actúan como enzimas antibacterianas en el camarón [48-50]
En el presente estudio, la actividad de LZM se vio afectada principalmente por la fuente de esterol en lugar de los niveles de esterol, y las fuentes dietéticas altas de fitosterol y esterol mixto mostraron un efecto positivo en la inmunidad no específica del camarón. Si bien la actividad de AKP se vio influenciada principalmente por los niveles de esterol, la inclusión de niveles de esterol de 2 g/kg aumentó la actividad de AKP en los grupos HC, HP y CP en comparación con los grupos LC. Además, la actividad de PO se vio influenciada interactivamente por la fuente y los niveles de esterol, con un aumento significativo en los grupos HP y CP, similar a los resultados de LZM.
En un estudio previo, se informó que los fitoesteroles dietéticos mejoran la salud intestinal, la inmunidad y la actividad antiinflamatoria de los lechones [11]. Además, una dieta rica en fitoesteroles es capaz de reducir el riesgo de problemas cardiovasculares y cáncer [51].
En algunas especies acuáticas, una dieta suplementada con fitoesteroles no tuvo efectos sobre la respuesta inmune a la dorada y la lubina europea [31 , 32], lo que sugiere que la tolerancia y la utilización de los animales a los fitoesteroles es específica de la especie. Por otro lado, el sistema antioxidante también juega un papel vital en el apoyo a la respuesta inmune y la prevención del estrés oxidativo [52]. Por lo
general, el malondialdehído (MDA) se utiliza como un biomarcador de daño oxidativo en animales [53].
En el presente experimento, las fuentes de esteroles tienen un efecto principal en el contenido de MDA en la hemolinfa, siendo el MDA más bajo en los camarones suplementados con fitoesteroles (grupos HP, CP y LP) en comparación con los grupos LC y HC. En contraste con el contenido de MDA, la actividad de la superóxido dismutasa (SOD) en los camarones alimentados con dietas HP y CP aumentó significativamente en comparación con los grupos LC y HC. Estos resultados sugieren que los fitoesteroles dietéticos ayudan a reducir el daño por oxidación y a mantener la salud de los camarones activando la actividad de la SOD [54].
Un estudio previo ha demostrado que los fitoesteroles dietéticos podrían reducir el
estrés oxidativo en el suero y el hígado de lechones destetados al estimular la actividad de la SOD, y también se informaron resultados similares en ratas Sprague-Dawley [55, 56]. No es coincidencia que se haya informado que los fitoesteroles dietéticos aumentan la actividad de la glutatión peroxidasa sérica y hepática y reducen el contenido de MDA del músculo de la pechuga en los pollos Partridge Shank [57].
En un estudio anterior, los fitoesteroles y sus componentes principales se consideraron antioxidantes [58]. Esto puede atribuirse a la inducción del desacoplamiento mitocondrial por fitoesteroles que conduce a la producción de especies reactivas de oxígeno y la estimulación del ciclo redox del glutatión y el sistema antioxidante, que en última instancia protege a las células del daño oxidativo [59, 60]. En el futuro, se realizarán experimentos de transcripción y traducción de genes inmunes y de desafío bacteriano
para investigar más a fondo el efecto de los fitoesteroles en la inmunidad no específica.
Conclusión
La suplementación de 2 g/kg de fitoesteroles en la dieta no afectó el crecimiento de los camarones, pero tuvo un efecto reductor del colesterol en Penaeus vannamei. Las fuentes mixtas de esteroles podrían mejorar el crecimiento y la inmunidad no específica de los camarones. Este estudio reveló los efectos de diferentes fuentes y niveles de esteroles en el crecimiento y la inmunidad no específica de los camarones y proporcionó una base para una mayor exploración del mecanismo del fitoesterol•
Para más información sobre este artículo, contactar a: xswzsdx@163.com
Comparación del coeficiente térmico de crecimiento en el camarón Penaeus vannamei en
Ecuador con diferentes sistemas de alimentación
Autores: Miguel Jover-Cerdá
Juan Carlos Valle Masson
Patricia Aguilar Astudillo
Ignacio Jauralde García
Universitat Politécnica de Valencia. Instituto de Ciencia y Tecnología Animal. Grupo de Acuicultura y Biodiversidad. Camino de Vera s/n 46022 Valencia (España)
Publicación original AQUATECHNICA: https:// doi.org/10.33936/at.v6i2.6689
El camarón o langostino de patas blancas Penaeus vannamei (Boone, 1931) es la especie acuícola de más valor en la producción mundial, con ventas de 29.000 millones de dólares en 2021, y una producción de 6.8 millones de toneladas (FAO, 2024). Su producción es mayoritariamente semiintensiva en estanques de tierra y tradicionalmente, el sistema de alimentación más utilizado ha sido el manual por voleo, pero en los últimos años se están imponiendo los alimentadores automáticos, principalmente con mecanismos temporizados y con hidrófonos. Los resultados en estanques experimentales de 1000 m2 con aireación indican una mayor productividad utilizando hidrófonos (Jescovitch, et al. 2018, Ullman et al. 2019 a,b, Reis et al. 2020, 2021), pero existen pocos estudios en condiciones reales de producción con estanques de varias hectáreas y sin aireación, como el de Valle et al. (2023), trabajo en el que no se pusieron de manifiesto las ventajas de los comederos automáticos sónicos.
El crecimiento del camarón se ha estudiado tradicionalmente usando índices como la Tasa de Crecimiento Específico (TCE) o los gramos por día o semana, pero no se ha utilizado el Coeficiente Térmico de Crecimiento (CTC).
Este último coeficiente fue desarrollado por Cho y Bureau (1998) para peces, y ha sido ampliamente empleado por otros autores (Mayer et al. 2012, Estruch et al. 2017), y aunque Bureau et al. (2000) apuntaron la posibilidad de utilizar este coeficiente, tuvieron que pasar 20 años hasta que Powell et al. (2020) lo aplicaran al langostino (P. vannamei), encontrando dos patrones diferentes de crecimiento y un punto de ruptura a los 7,5 g, de forma que en la primera etapa (0,01-7,5 g) el CTC fue de 0,152 y en la segunda etapa (7,5-34 g) de 0,807, con un valor del exponente del peso de 0,416 y de 0,952, respectivamente, y obteniendo un mejor ajuste que con el TCE y el CTC tradicional.
Ruiz-Velazco et al. (2010a,b) modelizaron el crecimiento del camarón en diferentes períodos del año con distinta temperatura, y encontraron que en verano (con temperaturas más altas), el crecimiento fue
mayor que en invierno (con temperaturas más bajas). Asimismo, Araneda et al. (2020) incluyeron el efecto de la temperatura en el crecimiento del langostino P. vannamei usando el modelo de Gompertz, pero durante una misma estación, de noviembre a mayo. Otros modelos de crecimiento han sido propuestos (Carvajal y Nebot, 1998, Franco et al. 2006, Sánchez y González, 2021), pero de momento no han tenido atención por parte de investigadores ni productores.
El presente estudio tuvo como objetivo estudiar el comportamiento del modelo de crecimiento de CTC en el camarón a partir de resultados de camaroneras comerciales con tres sistemas de alimentación: manual, automático temporizado y a demanda con hidrófonos.
Material y métodos
Se utilizaron datos reales de producción comercial de 208 lotes de los años 20152017 procedentes de 8 fincas camaroneras ubicadas en Ecuador (esclusas en el golfo de Guayaquil y la isla Puná), donde la temperatura del agua osciló entre los 24 °C y los 32 °C, y la salinidad entre 22,85 y 34,69 ppm. Se seleccionaron aquellos lotes del trabajo de Valle et al. (2023) que disponían de datos de temperatura adecuados para calcular la suma de temperaturas y el CTC.
El tamaño de los estanques varió entre 3 y 23 ha, y el tipo de alimentación suministrada fue manual (desde barca dos veces al día), automática temporizada (1 comedero Madam® por hectárea, con distribución de alimento cada 15 min desde las 12:00 h hasta las 24:00 h) y con hidrófonos a demanda (1 comedero AQ1 cada 3 ha disponible 24 h al día). La densidad inicial de siembra fue de 10 camarones de 0,17 g por cada m2, independientemente del mes de siembra, y no se empleó ningún sistema de aireación.
El crecimiento se evaluó mediante el Coeficiente Térmico de Crecimiento (CTC) considerando una temperatura efectiva de 18 ºC, la Tasa de Crecimiento Específica (TCE), y el Crecimiento Diario (CD, en g por día) usando las siguientes expresiones: CTC = 1000 x (Peso medio final1/3 – Peso medio inicial1/3) / Suma Temperatura Efectiva TCE = 100 ln (Peso medio final / Peso medio inicial) / días
CD = (Peso medio final – Peso medio inicial) / días
El análisis de los resultados se realizó mediante ANOVA usando el programa Statgraphics (Statistical Graphics System, Version Plus 5.1., Herndon, VA, USA). La comparación a posteriori de las medias se hizo utilizando el test Student-NewmanKeuls. El Factor F, cociente entre el Cuadrado Medio entre Grupos y el Cuadrado Medio Intra Grupos, también se consideró para visualizar las diferencias entre las variables.
Resultados y discusión
El número de días promedio de producción fue de 131 d, el peso final de 22,4 g y la producción de 1,718 kg/ha.
No aparecieron diferencias en el crecimiento con los diferentes sistemas de alimentación cuando se consideró el CTC o el CD (g/día), pero sí al considerar el TCE, de forma que el mejor valor de este índice se obtuvo con los hidrófonos respecto a la alimentación automática (Tabla 1), aunque sin diferencia con la alimentación manual.
En la Tabla 2 se puede observar el efecto del mes de inicio del ciclo de crecimiento sobre los diferentes parámetros. Al observar la Tasa de Crecimiento Específico (TCE) no se encuentran diferencias estadísticamente significativas (p > 0,05) respecto al mes de inicio de la producción, aunque sí apareció un mayor Crecimiento Diario (CD) en los lotes iniciados en octubre respecto a los iniciados en mayo.
Por el contrario, para el CTC se encontraron diferencias estadísticamente significativas (p< 0,05), en los lotes que iniciaron su crecimiento en los meses con temperaturas medias más bajas, junio, julio y agosto (26,7, 26,7 y 27,3 ºC, respectivamente), siendo los valores de CTC más altos que en los meses de enero, febrero y diciembre, con temperaturas medias más altas (30,3, 29,8 y 30,1 ºC, respectivamente), lo que no corresponde con el valor de crecimiento en g/día, que no presentó diferencias en estos meses.
Aunque por definición el CTC debería ser constante con los cambios de temperatura, se constata que los lotes iniciados en diciembre, enero o febrero, con temperaturas altas (30 ºC) que anticipan temperaturas más frescas en los siguientes meses, obtienen CTC más bajos que los lotes iniciados en junio o julio, con temperaturas bajas (26- 27 ºC) que anticipan temperaturas más cálidas, de lo que se deduce que el modelo térmico podría no estar funcionando con temperaturas mayores a 27 °C.
Al hacer el cálculo del CTC sin Temperatura Mínima Efectiva, es decir con valor 0 ºC, y con los valores de 16, 18, 20 y 22 °C (Tabla 3), se observa un aumento del valor del CTT a medida que se incrementa la temperatura efectiva, como consecuencia de una reducción de la suma de temperatura efectiva.
Asimismo, las diferencias en el CTC entre los meses de temperaturas bajas (junio, julio y agosto) y altas (enero, febrero y diciembre), se incrementan a medida que se consideran
Tabla 1. Efecto del sistema de alimentación en los diferentes índices de crecimiento (CTC, TCE y CD del camarón Penaeus vannamei.
Manual al Voleo 93 0,001725 3,97 ab 0,1712 ± 0,000034 ± 0,07 ± 0,0025
Automática 90
Nivel Sig.
0,001743 3,82 b 0,1717 ± 0,000035 ± 0,07 ± 0,0026
No significativo p < 0,05 No significativo
Medias con distinta letra presentan diferencias significativas (p < 0,05). EE = Error Estándar
valores crecientes de la Temperatura Mínima Efectiva (0, 16, 18, 20 o 22 ºC) a la vez que aumenta el valor del Factor F, lo que apunta a que el modelo del CTC podría no estar funcionando bien en el camarón producido en condiciones reales de producción con oscilación mensual de la temperatura de los diferentes lotes a lo largo del año.
El crecimiento diferencial del camarón alimentado con sistemas acústicos de hidrófonos solo se detecta cuando se utiliza la Tasa de Crecimiento Específica (TCE), y tan solo respecto a la alimentación con equipos temporizados, pero no cuando se analizan los resultados mediante el Crecimiento Diario (CD) o el Crecimiento Térmico (CTC), lo que puede plantear errores de interpretación por falta de significación al considerar diferentes índices de crecimiento.
Así, Valle et al. (2023) no encontraron diferencias con los citados sistemas de alimentación al considerar el crecimiento semanal, mientras que la mayoría de trabajos (Jescovitch, et al., 2018; Ullman et al., 2019a,b; Reis et al., 2020, 2021) indican que los alimentadores acústicos son más efectivos (mayor peso final y crecimiento en gramos por semana), seguidos por los alimentadores automáticos y la alimentación manual. Estas diferencias entre autores podrían ser debidas al bajo número de alimentadores por hectárea en el estudio de Valle et al. (2023) y el presente trabajo.
Debido a que los crustáceos son animales poiquilotermos, es esperable que la temperatura tenga un efecto influyente sobre su crecimiento, por lo que la temperatura óptima para el desarrollo del camarón ha sido objeto de numerosos estudios.
En algunos trabajos se ha visto que la temperatura por encima de los 27 °C acorta el período de crecimiento tanto en larvas como en juveniles, alcanzando un tamaño mucho mayor en relación a camarones que han sido mantenidos en temperaturas inferiores (Ponce-Palafox et al., 1997; Bureau et al., 2000). Asimismo, Wyban et al. (1995) obtuvieron valores óptimos a 30 ºC para camarones de pequeño tamaño (< 5 g) mientras que para ejemplares mayores citaron una temperatura óptima de 27 ºC. Mansouri (2013) no citó diferencias en el
Tabla 2. Efecto del mes de inicio del ciclo de producción en los diferentes índices de crecimiento (CTC, TCE y CD) del camarón Penaeus vannamei.
Enero n = 18
Febrero n = 16
Junio n = 18 26,7
Julio n = 15 26,7
Agosto n = 21 27,3
Septiembre n = 19 27,8
Octubre n = 12 28,7
Noviembre n = 21 29,5
Diciembre n = 14
Nivel Sig.
Medias con distinta letra presentan diferencias estadísticamente significativas (p<0,05%).
EE = Error Estándar
TCE de camarones de 5 g en biofloc con 28 y 30 ºC, pero el crecimiento con 26 ºC fue menor, aunque las supervivencias fueron mejores con 26 y 28 ºC (72-76%) frente a 30 ºC (43%). Recientemente, Araneda et al. (2020) obtuvo que la mejor productividad se consiguió con temperaturas por encima de 26 ºC. Finalmente, Al-Masqari et al. (2021) obtuvieron mejores crecimientos de juveniles de 0,6-3,4 g con incrementos de temperatura de 22, 26 y 30 ºC, pero con una reducción de la supervivencia.
En sistemas semiintensivos de producción la temperatura no es constante, sino que varía a lo largo del día y del ciclo. En este sentido, Guo et al. (2010) encontraron un efecto significativo del cambio cíclico de la temperatura diaria en el crecimiento de juveniles del camarón P. vannamei, de forma que los ciclos 25 ±2 y 25 ±3 ºC originaron mayor crecimiento y eficiencia alimentaria que los ciclos 25 ±1 y 25 ±4 ºC, lo que evidencia el efecto negativo de las temperaturas bajas (25±1) y altas (25 ±4 ºC), y la necesidad de considerarlas en los modelos de crecimiento.
De igual forma, Abdelrahman et al. (2018) estudiaron el crecimiento del camarón P. vannamei en estanques de producción comercial, y encontraron que la mayor tasa de crecimiento semanal se produjo cuando las temperaturas del agua fueron de 26-27,5 ºC durante un mayor número de horas, lo que explicaría en parte los resultados obtenidos en el presente trabajo.
Estos indican claramente que los valores de CTC son mayores en los lotes que iniciaron su ciclo en los meses de junio y julio (con temperaturas menores) y menores en enero, febrero y diciembre (con temperaturas mayores) para todas las temperaturas mínimas efectivas ensayadas, y que cuanto más alta sea la temperatura mínima efectiva utilizada para calcular el CTC, menor es la suma de temperaturas efectivas y mayor será dicho valor, lo que se demuestra por la correlación negativa entre CTC y la suma de las temperaturas efectivas del ciclo de crecimiento (Fig. 1).
Los resultados con el resto de los parámetros de crecimiento fueron distintos, pues la TCE apenas presenta diferencias en el mes de agosto que es relativamente menor, y el CD en gramos/día son mayores en octubre, pero sin diferencia con los demás meses a excepción de mayo, lo que podría explicarse por una mayor temperatura durante los meses siguientes a la siembra en octubre (27,1 ºC) que en mayo (30,0 ºC).
En un trabajo similar al presente (Borbor, 2020), se mostró un comportamiento similar entre los valores de CTC y los meses con mayor y menor temperatura, aunque se obtuvieron valores más altos de CTC (con Temperatura Efectiva de 18 °C), en torno a 0,00297 frente a 0,00172 en el presente estudio, probablemente debido a diferencias en las condiciones de producción.
Tabla 3. Efecto del mes de inicio del ciclo de producción sobre el Coeficiente Térmico de Crecimiento (CTT) calculado considerando diferentes valores de temperatura mínima efectiva (0, 16, 18, 20 y 22 ºC) en el camarón Penaeus vannamei.
Medias con distinta letra presentan diferencias estadísticamente significativas dentro de la misma columna (p<0,05%)
Con base en lo mencionado, se podría decir que en este caso, para el camarón P. vannamei, que es una especie cuyo ciclo de producción dura de tres a cuatro meses, el CTC podría estar fallando por las temperaturas elevadas, ya que al hacer el sumatorio de temperaturas el valor es muy alto, lo que se ve reflejado en la obtención de un Coeficiente Térmico más bajo, pero sin que esto se traduzca en el crecimiento real, por lo que podría darse el caso de que exista una temperatura máxima que no es efectiva y que podría corregirse en este modelo de crecimiento.
Al estudiar el efecto de la Temperatura Mínima Efectiva (16, 18 y 20 ºC) y la Temperatura Máxima Efectiva (24, 26, 28 y 30 ºC) se obtiene que para 28 y 30 °C se mantienen los mayores valores de CTC en junio y julio, pero al considerar 24 y 26 °C, las diferencias entre los meses desaparecen (Figura 2), no presentando diferencias significativas para las temperaturas efectivas mínimas de 16, 18 y 20 ºC.
Considerando la reducción de los valores del Factor F a medida que disminuye la Temperatura Máxima Efectiva, se podría
Figura 1. Valores mensuales del CTC, suma de temperatura efectiva y regresiones matemáticas para el CTC y el sumatorio de temperatura, considerando temperaturas efectivas de 16, 18, 20 y 22 ºC y sin temperatura efectiva.
Figura 2. Valores mensuales del CTC, considerando temperaturas mínimas efectivas de 16, 18, 20 ºC y sin temperaturas máximas efectivas de 24, 26, 28 y 30 ºC
deducir que la modelización del CTC considerando temperaturas mínimas efectivas de 16-18 ºC y máximas de 24-26 ºC podrían ser las más adecuadas.
Se deduce del presente trabajo que los ciclos de crecimiento con mayor temperatura no mejoran el crecimiento real de los camarones, por lo que se justificaría el empleo de una temperatura máxima para los modelos de crecimiento.
Comparando los resultados de crecimiento obtenidos por Powell et al. (2020), considerando el exponente de peso clásico de 1/3 y sin temperatura mínima efectiva, 0,0012 y 0,0010 para las dos fases, con los del presente estudio, 0,00062, se observa un mayor crecimiento de los camarones en los sistemas intensivos analizados por Powell et al. (2020), con una temperatura media de 29,5 ºC y 0,34 g/d, frente a los 0,17 g/d en los sistemas semiintensivos y temperatura media de 28,5 ºC del presente estudio.
Finalmente, aunque las camaroneras utilizan
la TCE o el CD para evaluar el crecimiento de los camarones, estos no son apropiados para hacer predicciones de crecimiento, fundamentales en los planes de producción para el diseño de nuevas instalaciones y en la gestión de la biomasa, por lo que se necesita un modelo de crecimiento que permita tales predicciones en condiciones de temperaturas variables.
Así, los resultados obtenidos por Powell et al. (2020) sugieren que se puede utilizar el Coeficiente Térmico de Crecimiento (CTC) para modelar el crecimiento del camarón, pero considerando varios períodos y exponentes del modelo, a lo que habría que añadir que se debe tener en cuenta que la variación de temperatura puede ser muy limitada en condiciones tropicales, que es donde se encuentran la mayor parte de las granjas, ya que la diferencia de temperaturas medias mensuales extremas en muchas ocasiones no llega ni a 3 °C, como en el presente estudio, haciéndose necesaria la consideración de temperaturas efectiva mínima y máxima.
Conclusiones
El índice utilizado para evaluar el crecimiento del camarón es muy importante, pues la alimentación con hidrófono produjo una mejor Tasa de Crecimiento Específica, pero sin diferencias con el Crecimiento Diario o el Coeficiente Térmico de Crecimiento.
El modelo de crecimiento de CTC podría ser apropiado para el camarón si se consideran en ambas temperaturas efectivas mínimas y máximas, las diferentes fases de crecimiento y los exponentes del modelo, pero se requieren más estudios de campo con datos de pesos a lo largo del ciclo de producción que confirmen los resultados obtenidos en este trabajo•
Para más información sobre este artículo, contactar a: mjover@dca.upv.es
Estandarización de concentraciones de solución de metabisulfito de sodio y tiempo de inmersión para el camarón de cultivo Penaeus vannamei
Autores:
Luciana Trigueiro de AndradeI
Nkarthe Guerra AraújoII
Ana Paula Moraes VenturaIII
Andréa de Lucena LiraIV
Marciane MagnaniV
José Marcelino de Oliveira CavalheiroVI
IInstituto Federal de Educación, Ciencia y Tecnología de Paraíba (IFPB), Campus Cabedelo, Santa Rita de Cassia, s/n, Jardim Jerico, 58310-000, Cabedelo, PB, Brasil. Correo electrónico: luciana.andrade@ifpb.edu.br. Autor para correspondencia.
IIPrograma de postgrado en Ciencia y Tecnología de Alimentos, Universidad Federal da Paraíba (UFPB), João Pessoa, PB, Brasil.
IIIDepartamento de Nutrición, UFPB, João Pessoa, PB, Brasil.
IVDepartamento de Química, IFPB, João Pessoa, PB, Brasil.
VDepartamento de Ingeniería de Alimentos, UFPB, João Pessoa, PB, Brasil.
VICentro de Tecnología y Desarrollo Regional (CTDR), UFPB, João Pessoa, PB, Brasil.
El camarón marino Penaeus vannamei es actualmente la principal especie de camarón cultivada en Brasil debido a sus excelentes condiciones de crecimiento y adaptabilidad, fácil nutrición, manejo y altos niveles de productividad y rentabilidad (Pérez-Velázquez et al., 2012).
Uno de los factores limitantes para incrementar la comercialización del camarón, tanto a nivel nacional como externo, son las pérdidas de calidad a las que está sometido el animal después de su recolección, siendo la melanosis uno de los principales problemas y la responsable del oscurecimiento de su caparazón. Se produce debido a la acción de un complejo enzimático endógeno presente en el camarón, el polifenol oxidasa (PPO), en el que la tirosinasa es la principal enzima activa (Huang et al., 2010).
El método más utilizado para inhibir este oscurecimiento enzimático en el camarón es el uso de conservantes a base de sulfitos, ya que actúan eliminando el oxígeno y reduciendo el pH, condiciones esenciales para la reacción enzimática (Rocha, 2000). Su uso en concentraciones inferiores a las ideales puede resultar en una melanosis o provocar el rechazo por parte del comprador/importador, y en los casos en que la concentración excede el límite establecido por la ley, los conservantes a base de sulfito provocan náuseas, dolor abdominal, vómitos, reacciones cutáneas, así como atragantamiento y neumonitis química en los consumidores o el personal operativo. Además, el uso excesivo de estos compuestos puede ser perjudicial para el medio ambiente, ya que sus residuos al ser desechados alcalinizan el agua provocando la muerte de varias especies acuáticas (Araújo y Araújo, 2011; Lima, 2008; Pedale et al., 2012).
Varios estudios han reportado el uso de soluciones de metabisulfito en concentraciones entre 1.25% y 12% con el objetivo de controlar la melanosis en el camarón durante el almacenamiento, con tiempos de inmersión que varían de 1 a 20 minutos bajo temperaturas de enfriamiento (Cintra et al., 1999; Góes et al., 2006; Barbieri Jr. y Ostrensky, 2001; Ogawa et al., 2003; Araújo y Araújo, 2011). Por lo tanto, es evidente que existe una gran variación
entre los procedimientos y, en consecuencia, una gran diferencia de concentraciones residuales de SO2 encontradas en el producto final.
Dada la importancia de controlar los niveles de SO2 en el músculo comestible del camarón, el objetivo de este estudio fue determinar la correlación entre las diferentes concentraciones de solución de metabisulfito de sodio y los tiempos de inmersión del camarón entero para establecer el valor de estas variables que permitan obtener la concentración de dióxido de azufre (SO2) en el músculo comestible del camarón de cultivo (P. vannamei), de acuerdo con el límite máximo establecido por la ley (100ppm).
Material y métodos
El camarón Penaeus vannamei con peso promedio de 10 g, equivalente a una clasificación de 81/100 (individuos por kilogramo) fue obtenido de una camaronera ubicada en el municipio de Pilar - PB, en la que se realizó la recolección de manera aleatoria.
Inmediatamente después de la recolección, los camarones fueron sumergidos en agua potable a temperatura cercana a 0 °C durante 10 minutos, causando una muerte por choque térmico y fueron luego transportados al Laboratorio de Tecnología y Procesamiento de Carnes y Pescado de la UFPB en una caja térmica con hielo, utilizando una relación hielo/camarón de 2:1 (aproximadamente 4 °C), los cuales fueron colocados en bolsas plásticas LDPE de 500 g, para ser sometidos a congelación lenta en un congelador doméstico para posterior análisis.
Para caracterizar la muestra, se realizaron análisis de actividad de agua con un medidor electrónico AQUALAB modelo CX2 (Decagon Devices, Washington, USA); pH, según parámetros descritos por el método No. 947.05 de la AOAC (2000), y composición proximal con análisis de humedad, ceniza y proteínas realizados según lo descrito en los ítems 950.46.41, 920 153 y 928.08 de la AOAC (2000), respectivamente, mientras que el contenido de lípidos fue determinado por Folch, Lees y Slaon - Stanley (1957).
Se analizó el sulfito residual en músculo
Figura 1 - Correlación entre el valor promedio de la concentración residual de dióxido de azufre (SO2) en el músculo comestible de camarones de cultivo (Penaeus vannamei) y la concentración de soluciones de metabisulfito de sodio a diferentes tiempos de inmersión.
comestible de camarón para diferentes tiempos de inmersión (10, 20 y 30 minutos) y las concentraciones de soluciones de metabisulfito de sodio (1%, 2%, 3%, 4% y 5%). Las muestras de 100 g de camarón entero fueron sumergidas en soluciones de metabisulfito de sodio ajustadas a la concentración de ensayo y pre-enfriadas a 7 °C para los diferentes tiempos de inmersión del ensayo. Para evitar la pérdida por evaporación de sulfatos, las soluciones de metabisulfito de sodio se prepararon justo antes de los ensayos. Luego, el exceso de agua se drenó durante 3 minutos, se eliminó el caparazón y el exoesqueleto de los camarones enteros y el sulfito residual en el músculo comestible se determinó directamente de acuerdo con el procedimiento optimizado descrito por Monier-Williams en marco con la legislación brasileña (Brasil, 2011). Se utilizó la temperatura de 7 °C considerando que
esta es la temperatura habitual empleada en las camaroneras durante la inmersión en la solución de metabisulfito. Todos los análisis se realizaron por triplicado en dos experimentos diferentes.
El diseño experimental fue completamente aleatorio y los resultados de las tres réplicas se evaluaron mediante análisis de varianza, y las diferencias entre medias se trataron utilizando la prueba de Tukey (Cockran & Cox, 1957) con la ayuda del software SAS System (2001).
Resultados y discusión
Las características físicas y químicas del músculo comestible del camarón estudiado (Tabla 1) están acorde a caracterizaciones descritas previamente en estudios del camarón de cultivo de la misma especie (Sriket, 2007; Gonçalves Gomes, 2008; Araújo et al., 2012).
Tabla 1 - Características físicas y químicas del músculo comestible del camarón de cultivo (Penaeus vannamei).
El valor de pH encontrado (6.75) indica que el camarón ha cumplido con el límite máximo establecido por la RIISPOA, que es de 6.5 a 6.8 (Brasil, 1997). La actividad de agua encontrada (0.979) fue similar a la observada por Santos et al. (2011), al analizar Macrobrachium olfessi
Luego de analizar los niveles de SO2 en el músculo comestible de camarones que fueron previamente pelados e inmersos en concentraciones de soluciones de metabisulfito de sodio de 1%, 2%, 3%, 4% y 5% en tiempos de 10, 20 y 30 minutos a temperatura de 7 °C, fue posible construir las curvas que se muestran en la Figura 1.
Esta figura muestra que la correlación entre el valor medio de la concentración residual de SO2 en el músculo comestible de camarones de cultivo y la concentración de soluciones de metabisulfito de sodio es lineal en el rango de concentraciones utilizadas en este estudio para todos los tiempos de inmersión estudiados. Gómez-Guillén et al. (2005) estudiaron el efecto del SO2 en la inhibición de la melanosis en el camarón Penaeus longirostris luego de diferentes tratamientos con soluciones de metabisulfito y encontraron un aumento exponencial en los residuos de SO2 cuando se relacionaron con tratamientos de 0-9%. Sin embargo, se observó que hasta el 5% de concentración existe una tendencia de linealidad en sus puntos.
También se encontró un aumento significativo (P<0.05) en los contenidos de SO2 en el músculo comestible de los camarones con el aumento de la concentración de las soluciones de metabisulfito de sodio utilizadas en este estudio. Este aumento también fue observado por Vieira et al. (2008) para la mayoría de las concentraciones y métodos analíticos utilizados por los autores.
De manera similar, al analizar el aumento de la concentración de SO2 en el músculo comestible de los camarones con el aumento de los tiempos de inmersión, también se observó una variación significativa (P<0.05).
Góes et al. (2006) reportaron que para la concentración de soluciones de metabisulfito de sodio probadas en su experimento (1% y 10%), hubo una influencia significativa del tiempo de exposición en los niveles de SO2
Figura 2 - Correlación entre el valor promedio de la concentración residual de dióxido de azufre (SO2) en el músculo comestible de camarones de cultivo (Penaeus vannamei) y el tiempo de inmersión del camarón entero en solución de metabisulfito de sodio a diferentes concentraciones.
Estos resultados confirman los resultados encontrados por Wedzicha (1992), quien reportó la influencia de algunos factores como la concentración y el tiempo de inmersión en el uso de dióxido de azufre.
Cabe destacar que todas las concentraciones de soluciones de metabisulfito de sodio utilizadas en este estudio excedieron la concentración de 100 ppm de SO2 establecida por la legislación (Brasil, 1988), al menos en dos tiempos de inmersión (20 y 30 minutos), excepto la concentración de 1%, donde, incluso después de la inmersión durante 30 minutos, se alcanzó una concentración máxima de SO2 de 67.62 ppm. Según Vieira et al. (2008), en la práctica se observa el uso de concentraciones mucho más altas de metabisulfito de sodio (alrededor del 10%) para prevenir la melanosis, lo que puede sugerir que el camarón comercializado presenta contenidos residuales de SO2 por encima de los límites establecidos por la legislación brasileña (100 ppm) y europea (300 ppm).
Estudios de Ogawa et al. (2003) y Hardisson et al. (2002) con P. vannamei comprobaron este hallazgo, ya que se observaron excesivos niveles de este compuesto en más del 50% y 40% de las muestras analizadas, respectivamente. Este exceso
de SO2 presente en los camarones puede causar daños a la salud del consumidor, de los operarios y rechazo del mercado de consumidores.
Al analizar la correlación entre el valor medio de la concentración residual de dióxido de azufre (SO2) en el músculo comestible de los camarones de cultivo y el tiempo de inmersión de los camarones enteros en la solución de metabisulfito de sodio, fue posible construir curvas que se muestran en la Figura 2 para cada concentración de metabisulfito de sodio utilizada a la temperatura de 7 °C.
Por medio de ecuaciones que se muestran en la Figura 2, fue posible obtener el tiempo de inmersión necesario para alcanzar la concentración de SO2 de 100 ppm, establecida por la legislación brasileña para cada concentración de metabisulfito de sodio utilizada, excepto para la solución al 1%, que no alcanzó ese valor. Por lo
tanto, con el fin de optimizar las etapas de recolección del camarón, se recomienda sumergir a P. vannamei en una solución de metabisulfito de sodio al 3% por un tiempo de 13 minutos, tiempo que se calculó no extrapolando valores mínimos y máximos encontrados en este estudio, obteniendo así el menor tiempo de inmersión con la menor concentración posible, con el fin de reducir costos en reactivos y ahorrar tiempo a las empresas que trabajan en esta área.
Sin embargo, se deben realizar estudios adicionales con el fin de verificar la posibilidad de obtener una concentración umbral de SO2 en tiempos de inmersión más cortos, que permitan la aplicación y estandarización de estos parámetros por parte de las empresas productoras de camarón.
Conclusión
Con base en ecuaciones que relacionan la concentración residual de SO2 en el músculo
CALIDAD
comestible de camarones con una solución de metabisulfito de sodio y la concentración residual de SO2 en el músculo comestible de camarones con el tiempo de inmersión del camarón entero en solución de metabisulfito de sodio a diferentes concentraciones, es posible sugerir la inmersión del camarón en una solución al 3% durante un tiempo de 13 minutos para alcanzar una concentración de 100 ppm de SO2, como lo recomienda la legislación brasileña (Brasil, 1988)•
Para más información sobre este artículo, contactar a: luciana.andrade@ifpb.edu.br.
ESTADÍSTICAS
Edición 160 - Agosto 2024
72 77
80 Exportaciones de camarón Reporte de mercado de EE. UU. Reporte de mercado de China
COMERCIO EXTERIOR
CAMARÓN:
EVOLUCIÓN DE EXPORTACIONES 2010 -2023
Fuente: Cámara Nacional de Acuacultura
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: COMPARATIVO MENSUAL (Millones de Libras)
2020- 2024
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
COMERCIO EXTERIOR
CAMARÓN: PARTICIPACIÓN POR DESTINO (Libras)
Junio 2023 - 2024
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: PRINCIPALES PAÍSES DESTINOS DE EXPORTACIÓN (Millones de Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
COMERCIO EXTERIOR
CAMARÓN: EXPORTACIONES POR PARTIDA ARANCELARIA
Enero - Junio 2024
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO ANUAL DE EXPORTACIÓN (Libras)
Las importaciones de camarón a China en junio aumentaron un 12% con respecto a mayo, alcanzando las 83,700 toneladas. En comparación con el año anterior, esta cifra sigue siendo un 10% inferior, demostrando que la demanda de camarón en el mercado chino sigue siendo moderada.
En el primer semestre, las importaciones totales de camarón alcanzaron las 483,170 toneladas, un 11% menos que en el mismo período de 2023. Sin embargo, esto se ve ligeramente exagerado por el fuerte comienzo del año pasado, después del cual los volúmenes de importación cayeron un 3% en el segundo semestre, mientras que tradicionalmente, las importaciones durante el segundo semestre aumentan significativamente con respecto a la primera mitad del año. En junio, el precio de importación disminuyó $0.04 hasta los $4.91 por kg; esto fue $0.99 menos que en el mismo mes del año pasado. Con esto, el valor de las importaciones para el mercado chino alcanzó los 2.4 billones de dólares en el primer semestre, una disminución del 24% con respecto a 2023.
La demanda de camarón tigre (Penaeus monodon) importado se mantiene baja. En junio, el volumen importado se mantuvo estable en 1,280 toneladas LSE, un 34% por debajo del mismo mes de 2023. Vietnam vio aumentar su participación con un volumen de un 40% desde mayo a 792 toneladas, mientras que India disminuyó un 17% a 365 toneladas.
Perspectivas
Los meses de verano en China tradicionalmente resultan en un ligero repunte en la demanda de camarón, que es una de las razones por las que los volúmenes de importación de junio aumentaron un 12% con respecto a mayo. Sin embargo, la confianza de los consumidores en China ha estado en una tendencia a la baja desde marzo, poniendo a la vista la difícil situación económica en la que se encuentran los consumidores en ese país.
Debido a esta situación, los consumidores están dispuestos a gastar menos en proteínas más caras. Con el camarón vivo del mercado interno a un precio más alto, especialmente porque la disponibilidad de materias primas en este momento es relativamente baja debido a que los productores luchan con los efectos del clima y las enfermedades en los últimos meses.
El mercado está gravitando un poco más hacia el camarón importado congelado más barato. Esto significa que el aumento de las importaciones no se puede atribuir únicamente a un aumento de la demanda. Si bien la demanda normalmente crece hacia fines de año, así como también lo hacen las importaciones; este año, la expectativa es que los volúmenes se mantendrán relativamente estables. Especialmente si los productores se ven incentivados a abastecerse nuevamente durante estos tiempos con precios más altos en granja, y la oferta interna, por lo tanto, tendrá una mayor participación en el consumo hacia fines de año.
Las exportaciones de Ecuador a China aumentaron significativamente después de que se levantara la suspensión de envíos de los exportadores que habían excedido los niveles permitidos de
Importaciones de China en 2022, 2023 y 2024 (volúmenes y peso promedio/kg)
Importaciones de Vannamei al mercado chino (2022, 2023 y 2024) metabisulfito. Pero en julio, hubo una ligera disminución nuevamente. Se ha reportado que esto podría haber sido causado por la intensificación de controles aleatorios de las aduanas chinas sobre las importaciones de camarón, especialmente el camarón ecuatoriano. No ha habido ningún envío cancelado, por lo que solo está causando una interrupción en la "velocidad" a la que el camarón ecuatoriano está entrando a China, pero no una disminución del volumen, lo que puede llevar a que los exportadores tengan en cuenta tiempos de envío más largos, lo que resulta en un ligero cambio con respecto a los patrones tradicionales•
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
Importación de camarón de Estados Unidos
Autores: Jim Kenny jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com
Urner Barry
Importaciones de todos los tipos, por tipo
La data publicada a principios de este mes mostró que las importaciones de camarón de EE.UU. cayeron drásticamente en junio en comparación con el año pasado, con amplias pérdidas en casi todos nuestros principales socios comerciales. Las importaciones del mes cayeron un 14% a 117.241 millones de libras desde 136.278 millones de libras el año pasado. El impacto negativo de la data interanual se vio en todos los top cinco: India (-14.8%), Ecuador (-20.2%), Indonesia (-8.9%), Vietnam (-8.5%) y Tailandia (-14.0%).
También hubo pérdidas de Argentina (-6.6%) y México (-45.8%). El único ganador entre el top 10 fue Canadá (+6.6%).
La cifra anual hasta a la fecha de 771.265 millones de libras amplió el déficit del primer semestre a casi un 3%.
En términos de forma del producto, hubo fuertes caídas en la mayoría de las categorías de camarón sin cabeza y con cáscara, que incluye el pelado fácil (-17.1%), pelado (-14.4%) y cocido (-20.4%), pero nuevamente más empanizado (+15.1%).
Ciclos de importación mensual por país (todos los tipos)
India: las importaciones de India (-14.8%) cayeron significativamente en comparación con junio de 2023, con la decisión de los derechos compensatorios a la vista del mercado. Las 43.702 millones de libras fueron las más bajas en junio desde 2020, con las interrupciones iniciales del COVID-19. Las cifras del primer semestre totalizaron 287.489 millones de libras, o un 0.9% más que el año pasado. Hubo pérdidas en todas las categorías con menos camarón con cáscara (-8.9%), pelado (-11.8%) y cocido (-34.3%).
Ecuador: al igual que algunos de los otros socios comerciales, las resoluciones sobre derechos compensatorios (CVD) y anti-dumping (AD) han restringido los flujos comerciales de camarón a los Estados Unidos desde América Latina. Las importaciones de Ecuador (20.2%) cayeron en la mayoría de los top cinco socios comerciales. A pesar de la gran caída, las importaciones hasta junio son un 0.2% más altas en 218.108 millones de libras. Hubo menos camarón en las categorías más grandes de camarón con cáscara (-25.5%) y pelado (-13.8%), que superaron fuertemente al empanizado (+35.6%).
Indonesia: Indonesia (-8.9%), al igual que el resto del mundo, envió nuevamente menos camarón a los Estados Unidos. Con 20.264 millones de libras, este fue el total mensual más bajo desde agosto de 2017. El primer semestre está un 16.3% por detrás del año pasado, con un total de 137.197 millones de libras, y está continuamente muy por debajo del ahora consolidado número dos, Ecuador. Si bien hubo más envíos de camarón con cáscara (+6.8%), cocido (+9.3%) y empanizado (+5.25%), todos fueron eclipsados por las pérdidas en la categoría más grande: pelado (-29.0%).
Vietnam y Tailandia: ambos países revirtieron las pérdidas interanuales y se mantuvieron en la tendencia con los demás. Si bien Vietnam (-8.5%) y Tailandia (-14.0%) enviaron menos que en junio pasado, las cifras interanuales son mixtas. El primero sigue en la punta, pero se contrajo un 10.9% con respecto al año pasado,
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
mientras el segundo tuvo una pérdida del 6.2%.
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones de camarón con cáscara, incluido el de pelado fácil, cayeron un 17.1% más en junio de 2024 que el año pasado.
El segundo lugar de ganancias mínimas, Indonesia (+6.8 %), fue superada por las pérdidas de Ecuador (-25.5 %), India (-8.9 %), México (-57.2 %) y Vietnam (-46.5 %).
En cuanto a los tamaños, fue mayoritariamente una mezcla con ganancias en los tamaños 16-20, 26.30 y o/70 con pérdidas en otras categorías.
Los valores de reemplazo de camarón con cáscara (importación $/lb) rebotaron levemente de la gran caída del mes pasado. Los precios fueron de $3.32 por libra y $0.03 por libra.
Valor agregado, importación de camarón pelado
Las importaciones de camarón pelado, la categoría más grande por volumen, disminuyeron un 14.4%. Las adquisiciones de junio totalizaron 57.489 millones de libras. Las pérdidas fueron amplias y afectaron a todos los principales socios comerciales. Las cifras se reflejan aquí: India (-11.8%), Ecuador (-13.8%), Indonesia (-29.0%), Vietnam (-7.5%) y Tailandia (-39.1%).
Los valores de reemplazo del camarón pelado (importación $/lb.) continuaron recuperándose, con un aumento de $0.10 por libra a $3.44 por libra, el nivel más alto desde 2024.
Las importaciones de camarón cocido (de agua tibia) también fueron menores, con un 20.4%, mientras que las de camarón empanizado superaron el año pasado en un 15.1%.
Importaciones de camarón cocido, empanizado y otros
Los precios de reemplazo del camarón cocido reflejaron el repunte del camarón pelado. Los costos más altos y la confianza en la mejora de los valores llevaron a un aumento de $0.25 por libra a $4.38 por libra. Este
también fue el más alto en 2024 y el más alto desde septiembre de 2023.
Línea de tiempo del precio del camarón; anuncios minoristas
Comercio minorista: los anuncios minoristas de julio aumentaron con un 18.7% más de anuncios que en junio y un 12.5% por encima del promedio de tres años. Además de abril de este año, esta fue la mayor oportunidad de cantidad de compra minorista (o promociones) desde mayo de 2023. Ha habido oportunidades de obtener margen ya que los precios mayoristas se mantienen en mínimos en varios años; sin embargo, el desafío es llevar a los consumidores a los pasillos de mariscos. El precio promedio anunciado también aumentó significativamente a $ 7.76 por libra o $ 0.33 por libra.
Fuente: USDOC. Urner Barry
Detalle de importaciones de camarón YTD por tipo
Fuente: USDOC. Urner Barry
REPORTE DE MERCADO
Suministro de camarón a EE.UU. y situación del Golfo
La data de desembarques proporcionada por el NMFS han sido inconsistente y hay lagunas en la data, lo que hace que las comparaciones no sean confiables.
La data de desembarques de abril de 2024 publicada por el NMFS muestra que se desembarcaron 3.058 millones de libras (total, todas las especies, sin cabeza). Si bien esto fue estacionalmente más alto que marzo, todavía estuvo por debajo del año pasado. Durante los primeros cuatro meses, se reportó de desembarques de 11.052 millones de libras, un 24% por debajo de los 14.612 millones de libras del año pasado. Los pescadores siguieron enfrentándose a los desafíos económicos del camarón de menor precio y a los crecientes costos.
Exportación del camarón ecuatoriano
El mercado de importación de camarón tigre blanco y negro producido en camaroneras se caracteriza por precios de reemplazo y costos de envío más altos, tiempos de tránsito más prolongados, disponibilidad de contenedores más ajustada y mayores
retenciones de la FDA. Estos factores contribuyen a las brechas de suministro y a un sesgo generalmente constante de los importadores. Las consultas de importador a importador también están aumentando, pero la capacidad de satisfacerlas ha disminuido con la reducción de inventarios y las preocupaciones antes mencionadas.
Sin embargo, la aceptación amplia de precios es más moderada, ya que la demanda, si bien está mejorando, todavía está por debajo de las expectativas y se ve presionada aún más por las olas de calor en curso.
El mercado del camarón silvestre sigue inestable. La data de desembarques sigue siendo esporádica, pero muy inferior a los de años anteriores, lo que indica que las decisiones económicas en medio de la presión del producto importado siguen siendo pertinentes•
Reunión de seguridad fluvial con Armada del Ecuador
La Dirección Nacional de Espacios Acuáticos de la Armada del Ecuador (DIRNEA) convocó a representantes de los sectores productivos a una mesa de trabajo para mejorar la cooperación en la lucha contra las estructuras delictivas que operan en vías fluviales. Durante la reunión, Luis Herrera, director de Seguridad de la Cámara Nacional de Acuacultura, destacó las zonas peligrosas en el golfo de Guayaquil y se comprometió a proporcionar nuevamente esta información a las autoridades para impulsar su acción de control.
Declaración de origen en factura para exportar a Europa
La Cámara Nacional de Acuacultura participó en el foro público-privado
“Declaración de origen en factura para la exportación a Europa”, organizado por la Asociación de Exportadores de Banano del Ecuador (AEBE) y la Fundación PROTRADE. El evento abordó la introducción de una nueva herramienta que permite a los exportadores utilizar una "declaración de origen en factura" en lugar del tradicional "certificado de origen" para las exportaciones hacia la Unión Europea, con el objetivo de simplificar y agilizar el proceso comercial.
El foro contó con la presencia de importantes funcionarios del Gobierno Nacional, como Homero Larrea, viceministro de Comercio Exterior; Christian Correa, jefe de Calidad y Mejora Continua de la Secretaría Nacional de Aduanas, y Ma. Angélica Trujillo, subsecretaria de Políticas de los Sectores Estratégicos del Ministerio de Economía y Finanzas. Su participación destacó el compromiso gubernamental con la mejora y la facilitación del comercio internacional.
Mesa de trabajo con representante del Ministerio del Interior
En la Gobernación del Guayas se instaló una mesa de seguridad encabezada por Guido Núñez, subsecretario de Seguridad Pública del Ministerio del Interior, con el objetivo de fortalecer la colaboración público-privada contra la delincuencia. El director de Seguridad de la Cámara Nacional de Acuacultura, Luis Herrera, pidió intensificar y mantener de manera permanente los controles en ejes viales para combatir los delitos en áreas identificadas. Además, se acordó una reunión futura para socializar un nuevo protocolo de inspecciones intrusivas para productos de exportación con personal policial.
Reunión con Cónsul de China para fortalecer cooperación comercial
Daniel Pesantes, director adjunto de la Cámara Nacional de Acuacultura, participó en representación de la CNA y la Corporación de Gremios Exportadores del Ecuador (CORDEX) en una reunión con el cónsul general de la República Popular China en Guayaquil, Gao Zhenting. El encuentro se desarrolló en el marco del foro organizado por la Cámara Ecuatoriano-China, titulado "TLC: Nuevas rutas logísticas y oportunidades aduaneras".
Durante la actividad, se subrayó la importancia de fortalecer la cooperación y las relaciones entre los gremios ecuatorianos y las autoridades chinas, con el fin de potenciar las oportunidades comerciales y logísticas entre ambos países.
Taller sobre Certificado de Registro Sanitario Unificado
La Subsecretaría de Calidad e Inocuidad del Ministerio de Producción, Comercio Exterior, Inversiones y Pesca, en colaboración con la Cámara Nacional de Acuacultura, organizó un taller especializado para la obtención del Certificado de Registro Sanitario Unificado. Esta socialización, que estuvo dirigida a importadores y fabricantes de productos e insumos veterinarios acuícolas, buscó capacitar a los participantes en los procedimientos y requisitos para obtener este documento, asegurando que los productos cumplan con los estándares nacionales de calidad e inocuidad.
Durante el taller, los asistentes interactuaron con expertos que ofrecieron charlas y demostraciones sobre la documentación requerida, pruebas de laboratorio y protocolos de control de calidad.
CNA presentó nuevamente denuncias
sobre tala indiscriminada de manglar
La Cámara Nacional de Acuacultura presentó un informe de 36 casos de tala de manglar ocurridos entre 2017 y 2024 ante funcionarios de los ministerios del Ambiente y de Producción, Comercio Exterior, Inversiones y Pesca. El documento se realizó utilizando coordenadas y análisis de imágenes satelitales de las 25 áreas afectadas, identificando las camaroneras involucradas y delimitando las áreas establecidas en el acuerdo ministerial, para determinar su estado actual y la extensión de las áreas deforestadas.
El objetivo de esta denuncia es asegurar que no se concesionen zonas naturales de manglar para la acuicultura y aplicar las sanciones correspondientes a quienes intenten expandirse ilegalmente en estas áreas. La CNA busca que las autoridades tomen medidas contundentes para proteger los manglares y evitar futuras talas ilegales. Actualmente, se espera una respuesta firme y acciones concretas por parte de la Subsecretaría de Acuicultura y Pesca.
Denuncia de CNA motivó la limpieza de escombros en área de laboratorios de larvas
En el sector La Chueca de Mar Bravo, provincia de Santa Elena, donde se concentra más del 70% de los laboratorios de larvas de camarón del país, el Municipio de Salinas ha llevado a cabo una operación de remoción de escombros y basura. Esta acción fue emprendida tras una denuncia presentada por la Dirección de Ambiente de la Cámara Nacional de Acuacultura, ante las autoridades municipales y el director Zonal 5 del Ministerio del Ambiente, Agua y Transición Ecológica, Ronal Merino.
La CNA insiste ante las autoridades competentes la importancia del saneamiento externo de forma permanente en esta área, lo que contribuirá significativamente al cumplimiento de los protocolos sanitarios en el primer eslabón de la cadena de producción acuícola.
CNA da seguimiento a la ejecución de estudio sobre fluctuaciones en zonas de cultivo en el Golfo de Guayaquil
En la primera fase del estudio, los científicos están procesando las muestras de agua y sedimentos tomadas en semanas anteriores. Tres grupos de expertos iniciaron los muestreos a finales de junio pasado en 15 sitios específicos del golfo de Guayaquil, en un proceso que durará un año. El análisis ejecutado por la Cámara Nacional de Acuacultura busca conocer la calidad del agua y las condiciones ambientales y biológicas en las áreas de cultivo de camarón, así como evaluar las fluctuaciones espacio-temporales de variables físicas, químicas y biológicas. Directivos de la CNA mantuvieron varias reuniones virtuales para confirmar el avance de ejecución, acorde con lo planificado.
Nueva inversión: VITAPRO destina 2 millones de dólares para la modernización de su cadena logística
La multinacional Vitapro invierte en mejoras logísticas que prometen fortalecer su propuesta de servicio, a través de la trazabilidad de la cadena de suministro del alimento para camarón. Estas innovaciones se realizaron en la planta de una de sus marcas, Nicovita, en Milagro. La primera fase de este proyecto incluye la implementación de un software de gestión de inventarios y la optimización de sistemas de almacenamiento, lo que mejora la eficiencia operativa. Además, se ha dado un paso importante hacia una logística más sostenible mediante la incorporación de montacargas eléctricos, reduciendo un 30% las emisiones de CO2.
La inversión también ha sido dirigida al desarrollo de capital humano, mediante capacitaciones especializadas para garantizar altos estándares de calidad en el servicio al cliente y las operaciones. De esta forma, Vitapro refuerza sus procesos de trazabilidad, sostenibilidad, conservación y distribución en beneficio de su labor productiva.
Eco-Eficiencia:
GISIS S.A. obtiene la certificación ambiental "Punto Verde"
Gisis S.A. (Skretting Ecuador) es la primera empresa en la industria de alimentos balanceados de Ecuador en recibir la certificación ambiental "Punto Verde" con mención en Eco-Eficiencia. Este reconocimiento refleja el compromiso de la compañía con el crecimiento económico sostenible y la reducción del impacto ambiental.
La multinacional ha implementado tecnologías limpias y eficientes en sus plantas, optimizando el uso de recursos clave como agua, energía y combustibles, a la vez que ha disminuido significativamente la generación de desechos. Entre los principales logros se destacan la reducción del consumo de búnker en un 15.4% y una notable disminución del 61.3% en el consumo de agua en su planta “Galápagos”.
Estos avances consolidan el impulso de prácticas acuícolas más responsables y respetuosas con el medio ambiente, llevando al sector hacia un futuro más eficiente y sostenible.
Shirley Vásconez, asesora ambiental; Karina Briones, gerente de calidad de Skretting Latam; ministra del Ambiente, Agua y Transición Ecológica, Sade Fritschi; Gustavo Navarro, director financiero de Skretting latam y Ronald Heredia, director zonal 5 del Guayas.