Coyuntura actual y futuro próximo de las exportaciones de camarón en Ecuador
Ecuador First Class Shrimp, la imagen internacional del camarón ecuatoriano cumplirá 10 años de creación
DRONES: Nuevos aliados para espionaje y cometimiento de delitos
El secreto del camarón ecuatoriano= innovación + compromiso. Parte 2: Tecnificación en las fincas camaroneras
Síndrome de las Bolitas en criaderos de Penaeus vannamei en América Latina
Impacto de diversas condiciones de procesamiento en el nivel de sodio y en los atributos sensoriales del camarón
Primer registro de quistes de microalga Chattonella, un nuevo desafío para la producción de Penaeus vannamei
Bienestar Animal: un “nuevo término” largamente aplicado en la industria camaronera ecuatoriana
Guía práctica para el traslado de camarón vivo desde la finca hasta la planta procesadora
El Pacto Verde Europeo y sus implicaciones para el sector exportador camaronero
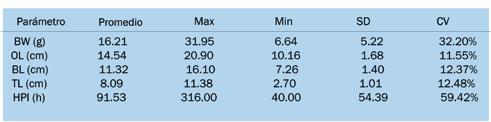
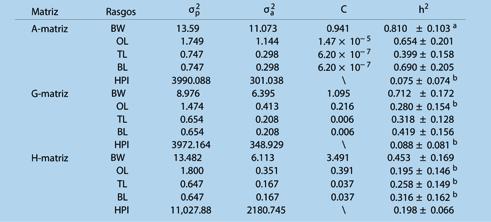
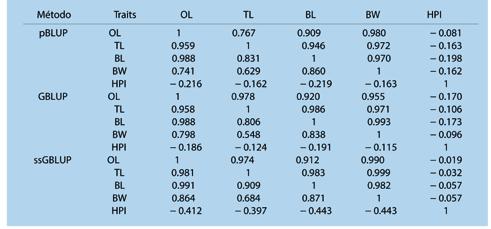
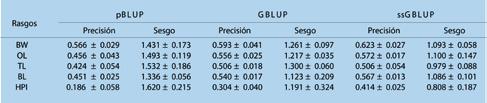
Estimación de parámetros genéticos para el crecimiento y rasgos de resistencia al WSSV en Penaeus vannamei
Presidente Ejecutivo Ing. José Antonio Camposano
Editora “AquaCultura”
MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita PhD. Leonardo Maridueña Ing. José Antonio Lince Ing. Alex de Wind
Exportaciones de camarón
Reporte de mercado de China
Reporte de mercado de EE. UU.
Noticias de interés
Noticias empresariales
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Corrección de estilo Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
Gabriela Nivelo gnivelo@cna-ecuador.com
Liderar significa estar dispuestos a aceptar el cambio como una constante y convertirlo en una oportunidad para impulsar nuestra competitividad. En lugar de ser simples observadores o víctimas de las circunstancias, en el sector camaronero ecuatoriano hemos optado por ser protagonistas del cambio. Las empresas que componen nuestra industria son ejemplos vivos de creatividad y capacidad de innovación, y han demostrado que, con la actitud correcta, los desafíos pueden convertirse en oportunidades.
Como Cámara Nacional de Acuacultura, asumimos el rol de líderes activos, no solo representando a nuestras empresas, sino también impulsando iniciativas que fortalezcan al sector. Tenemos el deber de fomentar un entorno en el que la colaboración, la innovación y el conocimiento compartido sean las piedras angulares del crecimiento.
En ese sentido, este año hemos fortalecido nuestra colaboración con Sustainable Shrimp Partnership (SSP) para introducir dos iniciativas que tienen como objetivo generar un cambio significativo a través de la transferencia de conocimiento y tecnología.
La primera de estas iniciativas es SustainED, un programa educativo diseñado para promover el desarrollo sostenible de nuestra industria. Este espacio, que integra conferencias especializadas, se convertirá en un espacio permanente para la discusión y el intercambio de ideas, con el objetivo de mejorar continuamente
EDITORIAL
José Antonio Camposano Presidente Ejecutivo
Colaboración e innovación: Claves para la sostenibilidad en la acuicultura
Como líderes del sector, estamos comprometidos a seguir impulsando el cambio, no como una respuesta a las circunstancias, sino como una decisión consciente de dirigir el futuro de nuestra industria. Porque el futuro de la acuicultura no solo se define por la capacidad de innovar, sino por la habilidad de formar alianzas sólidas y construir un camino sostenible para todos.
nuestras prácticas y avanzar hacia una acuicultura más responsable y eficiente.
La segunda iniciativa es Race to the Top, un foro que reúne a las empresas líderes del sector para compartir estrategias que han demostrado ser eficaces en áreas críticas, como la conservación de ecosistemas, la mitigación del cambio climático y el impacto social a gran escala. Este foro no solo nos permitirá aprender de las mejores prácticas, sino también identificar oportunidades de colaboración que potencien nuestras capacidades colectivas.
Estas acciones se suman a nuestra continua labor en la organización de AQUAEXPO 2024, el principal evento de la industria camaronera a nivel mundial. AQUAEXPO se ha consolidado como un espacio estratégico para quienes buscan estar al día con los avances científicos, tecnológicos y comerciales del sector. Es una oportunidad invaluable para compartir conocimientos, discutir las tendencias del mercado y fortalecer nuestras relaciones dentro de la cadena de valor.
Como líderes del sector, estamos comprometidos a seguir impulsando el cambio, no como una respuesta a las circunstancias, sino como una decisión consciente de dirigir el futuro de nuestra industria. Porque el futuro de la acuicultura no solo se define por la capacidad de innovar, sino por la habilidad de formar alianzas sólidas y construir un camino sostenible para todos•
DIRECTORIO
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Ing. Roberto Aguirre
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Blgo. Carlos Sánchez
Ing. Diego Puente
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Andrés Rivadulla
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León
Coyuntura actual y futuro próximo de las exportaciones de camarón en Ecuador
Autor: Daniel Pesantes
Director adjunto
Cámara Nacional de Acuacultura
dpesantes@cna-ecuador.com
Con una historia de más de 55 años, la industria camaronera ecuatoriana se ha convertido en un referente mundial, gracias al constante crecimiento e innovación de un sector productivo determinado a producir el mejor camarón del mundo, de una manera amigable con el ambiente y socialmente responsable.
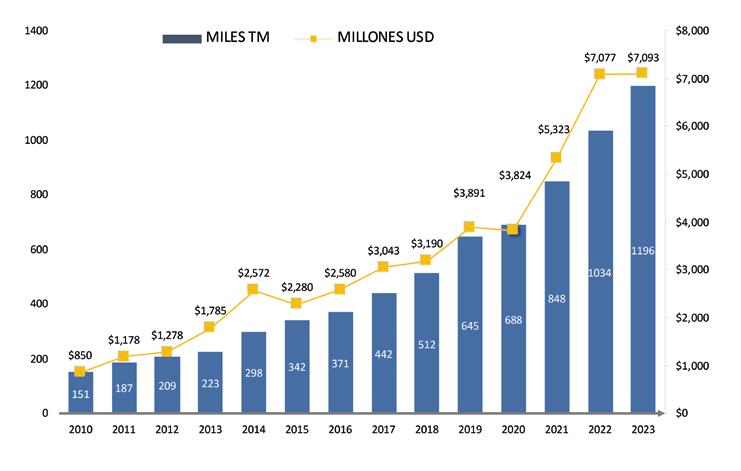
Hoy en día, Ecuador se mantiene como el principal exportador de camarón a nivel mundial, sosteniendo alrededor de 300 mil empleos directos e indirectos en el país. En 2023, esta industria aportó 7.2 mil millones de dólares en divisas, representando el 32.5% de las exportaciones no petroleras ecuatorianas y consolidándose como su principal producto de exportación.
Si bien las exportaciones de camarón ecuatoriano han tenido un impresionante crecimiento promedio en volumen del 19% en los últimos 10 años, es importante conocer hasta dónde podrán llegar esas cifras, tomando en consideración que existen muchas variables externas que sin duda afectan de manera directa este desempeño.
Situación actual de las exportaciones de camarón en Ecuador
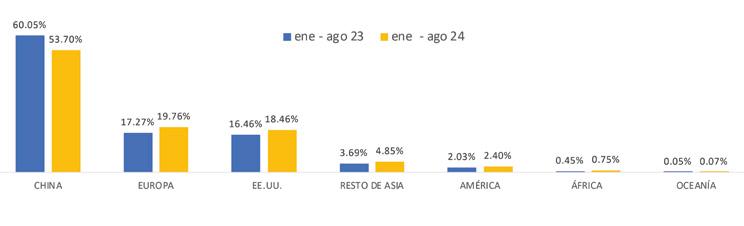
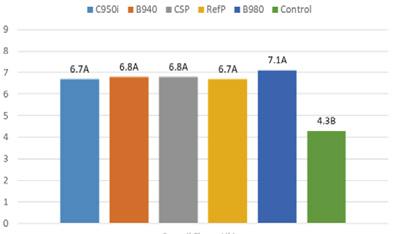
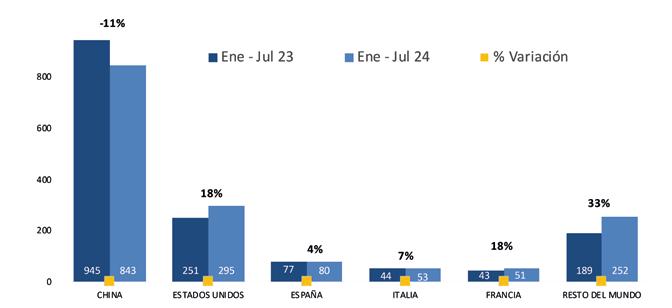
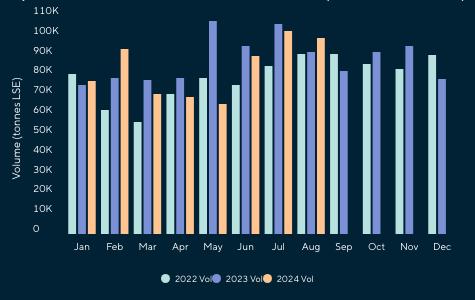
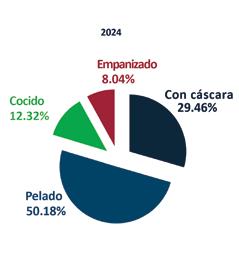
De acuerdo con cifras de la Cámara Nacional de Acuacultura, al mes de agosto del presente año, nuestro país ha exportado 818 mil toneladas de camarón, equivalentes a 4 mil millones de dólares. Si comparamos estos datos con el mismo período del año anterior, en volumen el crecimiento ha sido de apenas 1.63%, mientras que en divisas se presentó una caída del 6.25%.
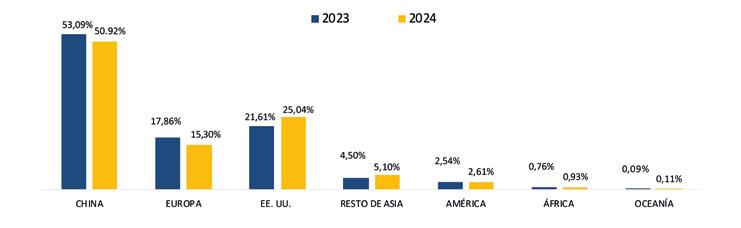
China abarcó el 53.70% de participación de las exportaciones, posicionándose como el principal destino de exportación en este período. A este mercado se enviaron un total de 439 mil toneladas, mientras que en 2023 las exportaciones fueron de 483 mil toneladas, lo que representa una caída del 9.12%.
Europa se ubicó como el segundo destino de las exportaciones de camarón con una participación del 19.76%, lo que representó un volumen de 161 mil toneladas de camarón al mes de agosto de 2024. En 2023, las exportaciones acumuladas a este mismo destino alcanzaron alrededor de 139 mil toneladas, lo que representa un crecimiento del 16.26%.
Estados Unidos, con una participación del 18.46%, se ubicó como el tercer principal destino recibiendo un volumen de 151 mil
toneladas de camarón ecuatoriano a agosto del 2024, valor que representa un incremento del 14% en comparación al 2023.
Situación de los mercados externos
El camarón ecuatoriano es un producto que depende de manera directa de muchos factores externos, como son las barreras técnicas de acceso a mercados, aranceles, problemas logísticos, así como de la posición de nuestros competidores como India y Vietnam, y actores en crecimiento como Indonesia, Tailandia, Arabia Saudita y Venezuela.
China se mantiene como nuestro principal mercado, con más del 53% de participación en las exportaciones en el período comprendido entre enero y agosto del 2024. No obstante, el volumen exportado representó una caída del 9% en comparación con el mismo período del 2023, valor muy lejano al crecimiento del 29% alcanzando en el período 2022 – 2023, y al 67% del comparativo 2021 – 2022.
Así mismo, desde el punto de vista de las importaciones chinas, Ecuador continúa siendo el principal proveedor de camarón
para este mercado, con una participación de más del 74% de sus importaciones al mes de agosto. Sin embargo, conforme cifras oficiales, alrededor del 75% del total del camarón consumido en China proviene de su propia producción local, situación que nos da una idea del potencial de tamaño de este importante mercado.
Por otro lado, y en lo que concierne al mercado estadounidense, las tasas de depósito en efectivo al ingreso del camarón ecuatoriano como resultado de la demanda por medidas de compensación y dumping impuestas a nuestro país, sumadas a los problemas logísticos por huelgas de estibadores en varios puertos podrían afectar el comportamiento de nuestras exportaciones en lo que queda del año.
En el caso de la Unión Europea, las alertas presentadas debido a la aplicación de un criterio erróneo para determinar la presencia de Vibrio cholerae en el último año generó una afectación en la dinámica de las exportaciones a este mercado; sin embargo, una vez superada esta situación, los volúmenes de exportación se han recuperado.
Proyección de las exportaciones de camarón
En los últimos años, la industria procesadora local ha venido ampliando y diversificando sus líneas de proceso, enfocándose en la agregación de valor. Así mismo, en fases de cultivo, se ha observado un cambio de densidades que ha afectado el comportamiento de la comercialización local del producto.
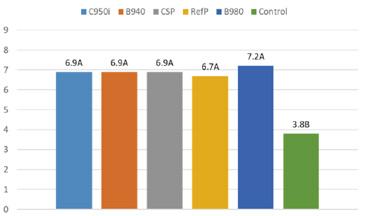
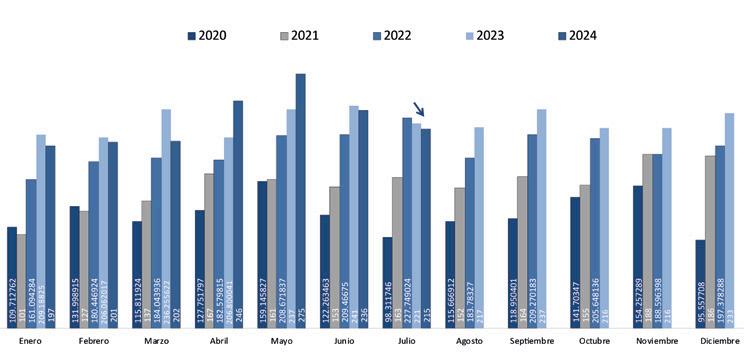
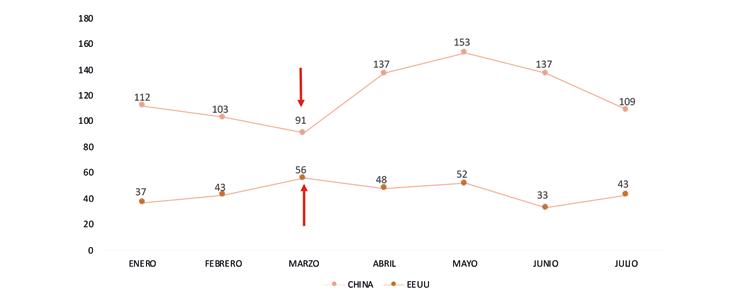
A pesar de estas situaciones, meses como abril y mayo de 2024 alcanzaron los niveles históricos más altos, con 111 mil y 125 mil toneladas respectivamente; no obstante, en el período acumulado entre enero y agosto se mostró un crecimiento de apenas el 1.6% comparado con el mismo período del año anterior, valor muy alejado del 15% de crecimiento en relación con el 2022 y más aún del 32% del 2021.
Estas cifras evidencian claramente una desaceleración en el crecimiento del volumen de exportaciones de camarón al cierre de este año. A pesar de los picos observados en el primer semestre, se proyecta que, al finalizar 2024, alcanzaremos un crecimiento aproximado del 3%, muy por debajo del 14% registrado el año anterior.
COYUNTURA
Factores
clave para el éxito futuro
Desde la Cámara Nacional de Acuacultura, promovemos una producción sostenible del sector, por lo que existen varias acciones que se realizan para alcanzar este objetivo. Aquí resalta la articulación con las instituciones públicas relacionadas con nuestra industria, que tiene como fin la implementación de políticas que sean favorables y que faciliten la dinámica de las empresas.
Así mismo, todos los actores de la cadena productiva deben plantear como prioridad la inversión en investigación y desarrollo, al igual que la aplicación de nuevas tecnologías para mejorar la producción y competir en calidad y sostenibilidad en los mercados globales.
No se puede ignorar la importancia de la necesidad de facilitar el acceso a los diferentes mercados internacionales. Es fundamental contar con una estrategia clara de diversificación y apertura de nuevos mercados, así como reducir la dependencia de los mercados tradicionales. En este sentido, hemos dado pasos adecuados pues hemos observado que, en los últimos 5 años, los destinos fuera de los mercados tradicionales han cobrado un papel más destacado en nuestras exportaciones.
Conclusiones
La industria camaronera de Ecuador ha experimentado un crecimiento impresionante en las últimas décadas, consolidándose como un pilar de la economía nacional y un líder en el mercado internacional. Sin embargo, los desafíos actuales, como la fluctuación en la demanda de mercados clave, las barreras regulatorias y la creciente competencia internacional, exigen estrategias de adaptación.
Los avances tecnológicos, la inversión continua y la diversificación de mercados, integrando la sostenibilidad como parte de
la estrategia, emergen como factores clave para garantizar la competitividad del sector en los próximos años.
Con un enfoque estratégico en estos aspectos, acompañado de una sólida promoción del consumo de camarón, no solo se mantendrá su posición en los mercados globales, sino que se fortalecerá su liderazgo como referente de calidad y sostenibilidad hacia el futuro•
LA IMAGEN INTERNACIONAL DEL CAMARÓN ECUATORIANO CUMPLIRÁ 10 AÑOS DE CREACIÓN
Autor: Shirley Suasnavas
Editora general de la Revista Aquacultura ssuasnavas@cna-ecuador.com
"Ecuador First Class Shrimp", imagen y marca registrada de la Cámara Nacional de Acuacultura, es el ícono del camarón ecuatoriano que se utiliza en ferias y eventos internacionales especializados en productos del mar en América, Europa y Asia. Su propósito es establecer una clara identificación de las empresas que forman parte del pabellón nacional y sus productos en las diversas actividades que a lo largo del año se llevan a cabo en los distintos mercados del camarón ecuatoriano.
La imagen fue creada por la CNA a finales del 2014 e hizo su aparición por primera vez en la Seafood Expo North America 2015 celebrada en Boston, representando al pabellón ecuatoriano.
Con una trayectoria de casi una década, ha logrado consolidarse como una imagen distintiva que no solo ha evolucionado, sino que también se ha convertido en un referente dentro de su sector. Su constante crecimiento y adaptabilidad han permitido que destaque, acogiendo a cada vez más empresas que se han integrado a la industria camaronera, así como acompañando a los exportadores tradicionales del país que empezaron sus activades de promoción en la década de los 90.
2024: UN PASO IMPORTANTE
Ecuador First Class Shrimp acaba de dar un paso más en el ámbito de la promoción del camarón ecuatoriano, al constituirse en un anfitrión para miembros de la alta cocina europea. La calidad y versatilidad excepcional del camarón ecuatoriano se evidenciaron en el primer encuentro “First Class Gala”, que se realizó en el reconocido Rooftop de la Maison de l’Alsace en París, el 26 de septiembre pasado.
El invitado de honor en este encuentro fue el chef ganador de estrella Michelin, Frédéric Simonin, conocido por su maestría en la cocina francesa y su enfoque en la simplicidad con sofisticación. Su arte culinario demostró a los asistentes cómo los productos
ecuatorianos pueden integrarse de manera magistral en la alta gastronomía europea, siendo el camarón su principal ingrediente a lo largo de la noche.
La embajadora de Ecuador en Francia, Ivonne Baki, fue la anfitriona de la gala, a la que también asistieron funcionarios del Ministerio de Producción, Comercio Exterior, Inversiones y Pesca, de la Cámara Nacional de Acuacultura y Sustainable Shrimp Partnership. A ellos se sumaron representantes europeos de cadenas de servicios de alimentación, mayoristas, supermercados, cadenas hoteleras y medios de comunicación especializados en cocina francesa, así como influencers gastronómicos que registraron fotos y videos que de inmediato compartieron en sus redes sociales, resaltando la exquisitez del camarón de nuestro país y subrayando el compromiso del sector con la calidad, la innovación y las prácticas sostenibles.
En un mercado altamente competitivo, el camarón ecuatoriano resaltó por sus atributos únicos como sus prácticas sostenibles y tecnologías avanzadas de producción.
La Cámara Nacional de Acuacultura ha jugado un papel clave en el posicionamiento internacional del camarón ecuatoriano con su distintivo Ecuador First Class Shrimp, mediante la participación en ferias y eventos de promoción comercial.
Además, ha facilitado que los productores y exportadores ecuatorianos exhiban su oferta, fortalezcan relaciones con clientes y atraigan a nuevos compradores. Estos espacios han sido esenciales
para consolidar la imagen de Ecuador como un referente de calidad en la industria camaronera.
La participación de los establecimientos exportadores ha permitido generar visibilidad y posicionamiento en un mercado altamente competitivo; conexión con clientes potenciales para generar ventas, acuerdos comerciales y relaciones de largo plazo; networking esencial para explorar nuevas oportunidades y fortalecer la red de contactos. También les brinda conocimiento de tendencias del mercado para obtener información sobre tecnologías innovadoras, prácticas sostenibles, regulaciones comerciales y demandas emergentes de los consumidores.
Estos espacios ofrecen la oportunidad de mostrar productos de manera tangible, lo que facilita la apreciación directa por parte de compradores, quienes pueden evaluar la calidad y diferenciación de los productos expuestos.
Participación en ferias internacionales recientes
En un permanente esfuerzo por continuar con la diversificación de mercados, Asia y Europa son objetivos clave para el camarón ecuatoriano. Es así que de forma simultánea a la consolidación de mercados como China y Estados Unidos, Ecuador ha buscado diversificar sus exportaciones hacia el resto de estos continentes.
EN ASIA
Ecuador First Class Shirmp participó en las ferias World Seafood Shanghai 2024 y Seafood Expo Asia en Singapur. Durante la feria de Shanghai, realizada del 28 al 30 de agosto, Ecuador contó con un pabellón de 360 m² en el que 17 empresas presentaron sus productos a compradores de China, el principal destino del camarón ecuatoriano.
La jornada continuó casi de inmediato en Singapur, del 4 al 6 de septiembre, donde el pabellón ecuatoriano de 81 m² acogió a cinco empresas participantes que buscaron expandir sus negocios en el sudeste asiático.
EN EUROPA
En la feria CONXEMAR, desarrollada en el Instituto Ferial IFEVI de la ciudad de Vigo, España, del 1 al 3 de octubre, el pabellón Ecuador First Class Shrimp tuvo un área de 341 m² y contó con la participación de 17 empresas. Cabe destacar que esta fue la vigésima participación de Ecuador en este evento, que cuenta con 25 ediciones. Durante mucho tiempo, el pabellón nacional estuvo conformado por solo 4 empresas, pero hoy en día más de una docena se han sumado, consolidando a CONXEMAR como la segunda feria más importante para el sector en el mercado europeo.
Para más información: www.ecuadorfirstclassshrimp.com
"EFCS nace en 2015 ante la necesidad de la CNA por participar en ferias internacionales con una imagen propia que nos diferencie de los demás, y que sobretodo lleve al mercado el mensaje de que, en los pabellones de la CNA, encontrarás un camarón de primera calidad diferente al de otros orígenes. Con el paso del tiempo, la presencia en ferias de EFCS, logró posicionar esta imagen como una marca de la CNA con la cual empresarios y la industria en general identifican al sector camaronero ecuatoriano. Desde su creación, y sin considerar los años que afectó la pandemia, hemos participado con EFCS en 39 ediciones de las ferias internacinonales más importantes del mundo.
“First Class Shrimp” es hoy una marca que lleva al mundo el mensaje del camaronero ecuatoriano. Somos una industria que se siente orgullosa de cultivar y ofrecer a los consumidores, el mejor camarón del mundo. Resultado del compromiso de Ecuador por mantener el liderazgo mundial no sólo en el volumen de producción y exportación, sino en la calidad, la atención y los cuidados necesarios para procurar producir de manera responsable con el ambiente y la sociedad (trabajadores, comunidades, consumidores)".
DRONES: Nuevos aliados para espionaje y cometimiento de delitos
Autor: Shirley Suasnavas
Editora general de la Revista Aquacultura ssuasnavas@cna-ecuador.com
En el 2024, el sector camaronero ecuatoriano ha comenzado a enfrentar una nueva amenaza tecnológica: el uso de drones para cometer delitos. Los reportes indican que estos vehículos aéreos no tripulados o también llamados aeronaves pilotadas a distancia (RPAs) se han convertido en herramientas de espionaje, robo y sabotaje en las granjas camaroneras, exacerbando aún más los problemas de seguridad que sufre la industria. Estos son algunos de los usos ilegales que están dando los delincuentes a estos equipos:
• Espionaje y robo de información: los drones se utilizan para sobrevolar y recopilar información sobre las operaciones de las granjas camaroneras, incluyendo datos sobre la ubicación de los estanques, equipos valiosos y patrones de producción. Esta información puede ser utilizada para planificar robos o sabotajes.
• Sabotaje de instalaciones: en algunos casos, los drones han sido empleados para lanzar sustancias contaminantes o dañinas en los estanques de cultivo, lo que puede afectar la salud del camarón y provocar pérdidas económicas significativas.
• Robo de productos: aunque menos común, se han recibido reportes de drones utilizados para transportar productos robados desde las instalaciones de procesamiento de camarón, aprovechando su capacidad para volar y evadir la detección.
• Vigilancia y acoso: se han registrado casos donde los drones son utilizados para vigilar a los trabajadores o propietarios de las granjas camaroneras, creando un ambiente de intimidación y preocupación constante.
• Interferencia de equipos: los drones pueden interferir con equipos de monitoreo o sistemas de control en las granjas camaroneras, afectando la operación normal y causando daños o mal funcionamiento de los mismos.
En Ecuador, la delincuencia ya ha adoptado el uso de drones como una herramienta tecnológica para cometer delitos, principalmente en actividades de espionaje, monitoreo de viviendas y ataques aéreos en prisiones.
Estos aparatos, equipados con cámaras de alta resolución, permiten a los criminales obtener información detallada de viviendas y negocios, identificando puntos vulnerables como accesos poco vigilados, horarios de ingreso y salida de los residentes, e incluso monitorear si las propiedades están desocupadas.
El país enfrenta una crisis de inseguridad sin precedentes, donde el crimen organizado, alimentado por el narcotráfico, ha encontrado en la tecnología una nueva forma de vulnerar la seguridad pública. El incremento en los índices de violencia, junto con el uso innovador de herramientas como los drones, plantea un desafío urgente para las autoridades y la sociedad.
Entre los tipos de drones existentes tenemos:
1. Drones de consumo con cámaras de alta definición: estos son los drones comerciales que se pueden comprar en tiendas de tecnología. Se utilizan para vigilar áreas residenciales, identificar puntos vulnerables de seguridad, horarios de actividad y obtener imágenes de casas para planificar robos.
2. Drones industriales o de carga: son vehículos aéreos más grandes y robustos, capaces de transportar objetos pesados. En Ecuador, estos han sido usados por bandas criminales para ingresar armas, drogas y otros objetos prohibidos en prisiones. También se han reportado casos de drones cargados con explosivos, como el ataque fallido en la cárcel La Roca de Guayaquil.
3. Drones modificados: son drones de uso comercial que han sido alterados por la delincuencia para realizar funciones específicas, como el transporte de paquetes ilegales o la colocación de explosivos. Estos drones pueden tener modificaciones en su estructura y capacidad de vuelo para evitar ser detectados por las autoridades.
El uso de estos vehículos por parte de la delincuencia ha generado preocupación entre las autoridades y los ciudadanos, especialmente en zonas de alta inseguridad y prisiones, donde se han reportado ataques con explosivos y otros delitos facilitados por esta tecnología. Casos reportados públicamente en Ecuador
Los habitantes del barrio La Armenia, en Quito, fueron víctimas de esta nueva modalidad delictiva. Según denuncian los moradores, días antes de sufrir un robo en su conjunto residencial, notaron la presencia de drones sobrevolando sus viviendas. Los dispositivos dejaron de aparecer tras el atraco, lo que confirma la preocupación de que hayan sido utilizados para obtener información y planificar el asalto.
Además del uso de estos equipos para perpetrar robos, se ha reportado su utilización en ataques a las cárceles de alta seguridad en Guayaquil. En uno de los eventos más impactantes, un dron equipado con explosivos fue neutralizado en el techo de la cárcel La Roca. Este incidente resalta cómo la tecnología está siendo empleada por organizaciones criminales no solo para robos, sino también para violentos enfrentamientos entre bandas rivales.
La Dirección General de Aviación Civil ha implementado reglamentos que obligan a los propietarios de drones a registrar sus equipos y a cumplir con restricciones de vuelo, especialmente cerca de áreas sensibles como aeropuertos, prisiones y zonas residenciales. Sin embargo, la capacidad de control sobre el uso de estos dispositivos sigue siendo limitada, dejando a los ciudadanos expuestos a nuevos riesgos tecnológicos.
COYUNTURA
Recomendaciones para evitar ser víctima de la delincuencia con drones
Para contrarrestar este riesgo, es fundamental implementar medidas preventivas. Una opción eficaz es la adquisición de inhibidores de señal, dispositivos que bloquean la presencia de drones en el perímetro aéreo de las viviendas. Estos aparatos son fáciles de obtener a través de compañías de seguridad o en línea, y no requieren permisos especiales.
Para evitar que un dron invada tu propiedad, se pueden implementar diversas medidas de seguridad. A continuación, algunas recomendaciones efectivas:
• Inhibidores de señal: estos dispositivos bloquean la señal de los drones, impidiendo que vuelen sobre tu propiedad. Son fáciles de adquirir en compañías de seguridad o tiendas en línea, y su uso no suele requerir permisos especiales en la mayoría de los casos, siempre que se utilicen de manera responsable para proteger el hogar.
• Instalación de sistemas de detección de drones: existen equipos que pueden detectar la presencia de drones en el área circundante. Estos sistemas alertan de manera temprana, permitiendo una respuesta rápida si un dron se aproxima a tu propiedad.
• Barreras físicas: aunque no es una solución definitiva, instalar redes o pérgolas sobre áreas vulnerables, como patios o jardines, puede dificultar que los drones accedan a tu espacio aéreo sin ser detectados.
• Organización comunitaria: mantener una red de comunicación con tus vecinos y establecer sistemas de vigilancia compartidos puede ayudar a identificar y reportar cualquier actividad sospechosa. Grupos en plataformas como WhatsApp pueden ser útiles para compartir alertas de seguridad.
• Comunicación con autoridades: en caso de que detectes un dron intrusivo, informa a las autoridades locales, ya que en muchos casos es ilegal sobrevolar propiedades privadas sin permiso. Las fuerzas de seguridad pueden tomar medidas para proteger tu privacidad y evitar futuras invasiones.
Ecuador debe implementar nuevas regulaciones para el uso de drones
Desde el 2015, a través de la Dirección General de Aviación Civil (DGAC), el país cuenta con un reglamento para el uso de drones; y aplica para las aeronaves cuyo peso máximo de despegue sea superior a 250 gramos y menor o igual a 150 kilogramos. Este reglamento exige a los propietarios de drones registrar sus aeronaves y contratar un seguro que cubra los daños a terceros causados por sus actividades de vuelo.
Además, la normativa exige que estos aparatos podrán volar solo entre “la salida y puesta del sol” (de 06h00 a 18h00), y su altura máxima de operación será de 400 pies (122 metros).
En 2024, este reglamento fue revisado para emitir nuevas normas generales que regulen la operación de aeronaves pilotadas a
distancia (RPAs), que incluyen reglas contra la operación negligente y temeraria, mantener responsabilidad sobre su manejo, garantizar la integridad de los bienes y la naturaleza; prohibir su uso cerca de aeropuertos, helipuertos, zonas de seguridad o áreas sensibles; restringir el lanzamiento o rociado y el transporte de armas, materiales peligrosos o mercancía.
El uso de drones ha ganado popularidad en Ecuador, con dispositivos que van desde los $50 hasta varios miles de dólares, y están disponibles en tiendas locales o importados desde Estados Unidos. La ley obliga a los propietarios a registrar sus drones y contratar una póliza de seguro que cubra daños por montos entre 3000 y 12 000 dólares, dependiendo del tamaño y capacidad del aparato.
Aunque los drones son una presencia habitual en parques y espacios públicos los fines de semana, estas nuevas regulaciones buscan garantizar un uso más seguro y controlado, evitando incidentes en zonas sensibles del país.Sin embargo, las autoridades enfrentan grandes desafíos para regular el uso de estos dispositivos, dejando a los ciudadanos expuestos a esta nueva modalidad delictiva.
El Gobierno y la sociedad civil deben coordinar esfuerzos para implementar medidas de seguridad más efectivas y prevenir que la tecnología sea utilizada para fines criminales, especialmente en sectores productivos vitales como el camaronero.
El espionaje y la extorsión
El espionaje con drones y los delitos de extorsión están estrechamente relacionados, ya que el uso de estos dispositivos permite a los delincuentes obtener información valiosa y detallada sobre sus víctimas, lo que facilita el acto extorsivo. Los drones con cámaras de alta resolución pueden sobrevolar propiedades y negocios sin ser detectados, capturando imágenes y videos que revelan aspectos clave como horarios, seguridad, bienes valiosos y vulnerabilidades.
Esta información recopilada puede ser utilizada por los extorsionadores para amenazar a las víctimas, exigiendo pagos a cambio de no divulgar información sensible o no llevar a cabo ataques físicos o
robos. Por ejemplo, en el caso de empresarios, los drones pueden ser utilizados para identificar los movimientos de la familia, el personal de seguridad o las áreas críticas de sus propiedades, lo que fortalece la posición de los extorsionadores al generar un ambiente de miedo y presión sobre las víctimas.
Cifras oficiales del delito
de extorsión
El incremento de los casos de extorsión en Ecuador durante 2024 ha alcanzado niveles alarmantes. Según datos oficiales de la Unidad Nacional de Investigación Antisecuestro y Extorsión (UNASE), se han registrado 2018 denuncias de secuestro y más de 10 mil casos de extorsión hasta la primera semana de septiembre. La cifra de extorsiones ha llegado a 10 771 denuncias, lo que representa un aumento del 95% en comparación con 2023, cuando se reportaron 8300 casos
Un ejemplo preocupante es la provincia de Santa Elena, donde se registran alrededor de 50 denuncias de extorsión mensuales. Sin embargo, esta cifra no refleja la totalidad del problema, ya que muchas víctimas optan por no denunciar debido al temor a represalias y/o por desconfianza en el sistema judicial.
En cuanto a los secuestros, la Policía Nacional ha logrado resolver 545 casos, mientras que 1563 denuncias permanecen bajo investigación por parte de las unidades especiales. Hasta el momento, se han atendido 821 denuncias, y 742 casos siguen en etapa de indagación. Según la UNASE, la intervención en estos 545 casos ha evitado el pago de aproximadamente $45,3 millones, que
COYUNTURA
habrían sido utilizados para financiar las actividades de los grupos de delincuencia organizada.
En la Zona 8 se han reportado 685 secuestros, de los cuales 71 personas han sido liberadas y 207 personas han sido detenidas en Guayaquil, Durán y Samborondón.
Las operaciones policiales en esta área han impedido que las bandas criminales obtengan $12 millones para los GDO.
En conclusión, el incremento de la delincuencia juvenil en Ecuador ha generado un debate urgente sobre la necesidad de reformar el Código Orgánico Integral Penal (COIP) para ajustar la edad de responsabilidad penal y eliminar la condición de inimputabilidad de los adolescentes que cometen delitos graves.
Esta medida busca hacer frente al creciente uso de menores por parte de organizaciones delictivas, quienes se aprovechan de su estatus legal para evadir sanciones. Sin embargo, cualquier reforma debe ir acompañada de un fortalecimiento del sistema judicial, garantizando que las decisiones de los jueces se ajusten a la legalidad y que no se otorguen medidas sustitutivas de manera indebida.
Solo a través de un sistema judicial íntegro, con reformas bien fundamentadas y un enfoque equilibrado entre prevención y sanción, se podrá enfrentar eficazmente la delincuencia juvenil y proteger la seguridad de la sociedad•
QPamela Nath Directora
ue desde el 2023 Ecuador sea el mayor exportador de camarón del mundo es un hecho, ya que las estadísticas a nivel global así lo demuestran. Pero lo que pocos saben, y que resulta verdaderamente sorprendente, es cómo lo ha logrado. En los últimos 23 años, la producción de camarón en el país ha aumentado casi 26 veces en volumen, sin expandir significativamente las áreas de cultivo, creciendo apenas un 0,85% por año durante el mismo período.
En la edición pasada de la Revista AQUACULTURA, compartí una anécdota que me gustaría retomar hoy. En esa ocasión, relaté cómo, desde que asumí mi rol en Sustainable Shrimp Partnership (SSP), he tenido la oportunidad de representar a la industria camaronera ecuatoriana en eventos internacionales, destacando los logros que han posicionado al país como líder mundial en producción de camarón y sostenibilidad. En varios de esos encuentros, surge una pregunta que parece inevitable: ¿Cuál es el secreto del éxito de Ecuador?
La respuesta, que ofrezco una y otra vez, es la misma: “La innovación tecnológica ha sido clave para nuestro desarrollo, pero existe un factor igual de importante para nuestro avance: el compromiso hacia la excelencia”. Esto es característico en toda nuestra cadena de valor.
El éxito ecuatoriano no se debe a una sola tecnología o a una única etapa del proceso de producción, sino al avance holístico e integral de toda la industria. Cada actor dentro de la cadena reconoce que un mejor desempeño en su área beneficia a todos. Comprenden que conformarse con lo ya probado y conocido sería una trampa que frena el avance hacia una producción más eficiente, rentable y sustentable.
En la primera edición de esta serie de artículos, ya exploramos innovaciones en los laboratorios de larvas, como los programas de mejoramiento genético, el uso de probióticos o las nuevas soluciones en la alimentación de las larvas que han sido vitales para el crecimiento sostenible de la industria camaronera en Ecuador. Estos avances han permitido incrementar la productividad desde el comienzo del ciclo de vida del camarón, sin comprometer el bienestar de las personas, fauna, flora y ecosistemas relacionados.
En esta segunda edición, el enfoque cambia hacia las fincas camaroneras, donde la adopción de diversas tecnologías como la automatización en la alimentación, las nuevas formulaciones de balanceado, el monitoreo de parámetros mediante sensores o la biorremediación están redefiniendo la gestión de recursos.
¿Es fácil adoptar estos cambios? No siempre. Pero quienes han decidido apostar por la innovación, ya están viendo resultados positivos y contribuyendo a un mejor futuro para la industria camaronera.
El secreto del camarón ecuatoriano:
innovación + compromiso
Parte 2: Tecnificación en las fincas camaroneras
Autores:
Pamela Nath
Sally Tabares
Sustainable Shrimp Partnership (SSP)
pnath@sustainableshrimp.org
¿Qué pensarías si te dijera que es posible cuadruplicar la producción de camarón sin comprometer la capacidad de carga de las piscinas, la calidad del producto y las buenas prácticas? ¿Y si además te contara que se pueden reducir significativamente los costos operativos mientras aumentan las ganancias? ¿Te sorprendería saber que esto no solo permitiría expandir tu negocio, sino también generar más empleos y posicionar tu producto como líder en los mercados más exigentes del mundo?
Estas no son solo ideas, sino realidades que se han logrado en la industria camaronera en Ecuador. ¿Cómo se han conseguido? A través de la aplicación de tecnologías y soluciones que han elevado el desempeño y transformado la forma de producir camarón.
En específico, estos son algunos de los beneficios que Omar Portugal, gerente técnico de Marfrisco, una camaronera del grupo Promarisco, ha experimentado desde que la finca bajo su gestión comenzó a tecnificarse e innovar.
El impacto de los alimentadores automáticos en las fincas
Una de las soluciones que ha permitido a Promarisco aumentar su densidad de siembra y mejorar su producción es el sistema de alimentación automática con tecnología acústica pasiva.
Estos dispositivos, diseñados para suministrar comida de manera precisa y controlada, permiten distribuir la ración exacta de alimento
que los camarones necesitan y demandan a tiempo real durante las 24 horas del día. Gracias a esta precisión, los ciclos de producción se vuelven más consistentes y predecibles, lo que facilita una mejor planificación y gestión de las operaciones en la finca.
“Antes de los alimentadores mecánicos, Marfrisco producía una tonelada de camarón por cada hectárea de piscina de engorde. Una vez que comenzamos a tecnificar y aprender a usar estos equipos, mejoramos nuestros indicadores productivos y logramos alcanzar hasta cuatro toneladas de camarón por la misma unidad de producción”, indica Portugal.
Las camaroneras, ¿automáticamente se nutren de beneficios?
De acuerdo con la directora técnica de AQ1 Systems, Keone Dodd, la automatización de alimento balanceado ha mejorado significativamente la eficiencia y la productividad de las fincas. A pesar de que requiere una mayor inversión inicial en infraestructura, con la adquisición de sensores y alimentadores, ofrece ahorros significativos a largo plazo al optimizar el uso y los costos del alimento, mejorar las tasas de crecimiento de los camarones, y prevenir la subalimentación y la sobrealimentación, reduciendo el desperdicio.
Además, Dodd explica que al distribuir el alimento con mayor frecuencia y en menores cantidades, los sistemas automáticos evitan que se acumule comida no consumida. Esto ayuda a reducir el riesgo de que se formen niveles altos de amoníaco, nitratos y otras sustancias dañinas en el agua. De esta manera, se previene que los nutrientes en
exceso contaminen el entorno, se mantiene una mejor calidad del agua y se reducen los problemas como las floraciones de algas nocivas. Todo esto contribuye a un ambiente más saludable y estable para los camarones.
La automatización de la alimentación también mejora la trazabilidad al registrar información detallada sobre los tipos de alimento y nutrientes requeridos, las cantidades y los tiempos de alimentación, lo cual es importante para el control de calidad, cumplir con los estándares de certificación y proporcionar transparencia a los consumidores preocupados por las prácticas sostenibles.
En Promarisco, los alimentadores funcionan completamente con energía solar. Gracias a los paneles y baterías, pueden operar todo el día sin necesidad de conectarse al sistema eléctrico nacional ni usar combustibles fósiles.
Sensorización de parámetros en la calidad de agua
Otro beneficio importante de los alimentadores automáticos es que integran tecnología de sensores y monitoreo para medir parámetros como los niveles de oxígeno disuelto en el agua, la temperatura, las precipitaciones y los acústicos. Los sensores proporcionan datos en tiempo real para predecir con precisión las necesidades de alimentación de la población de camarones dentro de la capacidad de carga del estanque.
“Estos sistemas recopilan datos extensos sobre el comportamiento de alimentación, las cantidades, las tasas de crecimiento de los camarones, la calidad del agua y las condiciones ambientales.
Luego, el análisis avanzado y el aprendizaje automático procesan estos datos para identificar patrones y correlaciones”, explica la directora técnica de AQ1 Systems.
El gerente técnico de Marfrisco complementa: “Gracias a los sensores, junto con el trabajo del equipo de monitoreo en la finca, podemos tomar decisiones acertadas en el momento adecuado. Esto reduce significativamente el riesgo de estrés, enfermedad y muerte en los camarones. Antes, en nuestra camaronera, la supervivencia alcanzaba, en el mejor de los casos, el 50% de los camarones sembrados. Ahora, estamos recuperando un 80%, incluso mientras sembramos cuatro veces más camarones que antes de implementar esta tecnología”.
Consejos al momento de adquirir nuevas tecnologías
Según el coordinador técnico de Sustainable Shrimp Partnership (SSP), Leonardo Maridueña, al adquirir un nuevo equipo para medir parámetros de calidad de agua o cualquier otro dispositivo, es importante asegurarse de que cumpla con las normas internacionales de calidad y esté avalado por un organismo de certificación reconocido. En el caso de las tecnologías de medición, el comprador debe investigar el rango de precisión e incertidumbre del equipo, comparando los resultados obtenidos con los de métodos tradicionales validados por laboratorios acreditados internacionalmente.
“Esto permitirá al comprador conocer el margen de error y los niveles de incertidumbre del equipo. Solo entonces podremos saber con certeza si un dispositivo es confiable en sus mediciones y si ofrece resultados en tiempo real. De lo contrario, no es recomendable invertir,
especialmente si no cuenta con respaldo técnico local, incluyendo el servicio de calibración”, señala Maridueña.
Se deben tomar en cuenta estos factores, ya que son los que garantizan la vida útil de los aparatos.
Expectativas vs. realidad: mitos de la tecnificación en las fincas
Las principales diferencias en las que ambos expertos coinciden entre los sistemas de alimentación manuales y los automatizados giran en torno a costos, beneficios, rendimiento predecible, replicación de los mejores resultados, cambio de roles y gestión de la finca.
“Los sistemas de alimentación manual tienen costos iniciales más bajos pero son intensivos en mano de obra y menos eficientes, lo que lleva a costos operativos más altos a largo plazo debido al desperdicio de alimento y al crecimiento subóptimo de los camarones”, describe Dodd.
Portugal cuenta que antes se creía que los camarones necesitaban tener acceso al alimento en toda la piscina para poder nutrirse bien. Sin embargo, la tecnología ha demostrado que esto no es necesario, ya que los camarones pueden adaptarse a comer en áreas más pequeñas dentro del estanque. “Ahora, con los alimentadores, el área de alimentación es mucho más
reducida, cubriendo solo entre el 3% y el 5% del área de la piscina”, comenta el gerente de Marfrisco.
Cambio cultural, el principal reto al implementar tecnologías La directora técnica de AQ1 Systems comparte que el mayor desafío que se encuentra en la implementación de sistemas de alimentación automatizados es el cambio cultural. “Es una forma completamente nueva de hacer las cosas, y el trabajo diario de los técnicos de la finca, los equipos de producción y los biólogos requiere de un cambio significativo que al inicio puede ser complicado”, expresa.
Portugal coincide con este punto de vista: “Cuando comenzamos a implementar los alimentadores mecánicos, al principio el personal de campo mostró cierta resistencia. Ellos pensaban que, al automatizar la alimentación, podrían perder su trabajo. Sin embargo, el resultado fue todo lo contrario”, revela.
Según indica, en lugar de eliminar puestos, los alimentadores mecánicos cambiaron las funciones. Ahora, en vez de distribuir el alimento manualmente, el personal se encarga de cargar los tanques de los alimentadores.
“Además, como la mejora tecnológica incrementó la producción aproximadamente cuatro veces más, se tuvo que contratar más personal, llegando a tener 2.5 veces más de nómina que al inicio del proyecto”, aclara. Extrusión, ¿uno de los avances más significativos en la formulación de los alimentos balanceados?
Otro de los avances que ha permitido mejorar sustancialmente el crecimiento y la productividad en las fincas es el desarrollo en la producción y formulación de los alimentos balanceados. Portugal destaca que la alimentación tiene un alto impacto en la producción, ya que el costo de estos alimentos puede representar hasta el 55% del gasto total en la producción de camarón. Gracias a la innovación, se han logrado reducir significativamente los costos de fabricación de alimento y desarrollar dietas que cubren de manera precisa las necesidades nutricionales de los camarones.
En el pasado, los alimentos para animales en forma de harina eran difíciles de manejar, se desperdiciaban fácilmente y tendían a producir más patógenos derivados del alimento no consumido. La peletización mejoró la eficiencia al convertirlos en gránulos (pellets), reduciendo el desperdicio. Sin embargo, el proceso no eliminaba todos los factores antinutricionales debido a las bajas temperaturas, lo que llevó a la extrusión a jugar un papel fundamental.
El gerente de Investigación y Desarrollo de Skretting Aquaculture Research Centre, César Molina, explica que gran parte del progreso en la fabricación de alimento se debe a los cambios que ha traído el proceso de extrusión, junto con mejoras en la molienda y en la adición de líquidos después de formar los pellets.
La extrusión es una técnica utilizada en la fabricación de piensos, que cocina los ingredientes a temperaturas muy altas, entre 120 y 150 °C, y con mucha presión. “Durante la extrusión ocurren transformaciones químicas que benefician la calidad nutricional de las materias primas
que conforman el producto final”, expone Molina.
A diferencia de la peletización, la extrusión permite una cocción más completa de los almidones y una desnaturalización de las proteínas que facilita el trabajo de las enzimas digestivas. Esto mejora la asimilación de proteínas, aminoácidos y carbohidratos de este tipo de alimentos.
“Aumentar la biodisponibilidad de estos nutrientes significó un impacto en campo, donde se pudieron evidenciar ciclos de cultivos más cortos y, por ende, un mayor rendimiento de producción con menor cantidad de alimento extruido por libra de camarón producido, lo que se refleja en una reducción del costo”, comparte el gerente de Investigación y Desarrollo.
Otros de los beneficios que Molina destaca sobre el alimento extruído son:
•Calidad del alimento: el extruido ofrece una excelente hidroestabilidad sin necesidad de aglutinantes sintéticos, y presenta una mayor homogeneidad en la longitud de los extrusos. Se obtienen más pellets por gramo, maximizando la disponibilidad del alimento en la piscina. Además, permite flexibilidad en la formulación y asegura la inactivación térmica de antinutrientes. Asimismo, la gelatinización del almidón durante el proceso de extrusión proporciona una mayor cohesión al alimento.
•Eficiencia y aprovechamiento: esto se traduce en una mayor ingesta de alimento por kilogramo comprado y suministrado, mejorando así el aprovechamiento de nutrientes y proteínas. El resultado es un mayor consumo de nutrientes con menos gasto energético y una mejor digestibilidad de aminoácidos. Finalmente, contribuye a una población de camarones mejor alimentados y tamaños más homogéneos, lo que resulta en una tasa de crecimiento más alta.
•Impacto ambiental y contaminación: el alimento extruido también disminuye la lixiviación de nutrientes en el agua de
cultivo, lo que reduce la contaminación de las piscinas y los efluentes. Además, se observan menores pérdidas de partículas de alimento en el agua y una reducción en las heces en suspensión. También se registra una menor demanda bioquímica de oxígeno por las heces.
El papel de los ingredientes alternativos en la sostenibilidad de los piensos
Los ingredientes alternativos son fundamentales en la formulación de alimentos para camarones, ya que buscan reducir la dependencia de harina y aceite de pescado, que conllevan un alto impacto ambiental y económico. A pesar de que existen alternativas, debemos ser conscientes de que no podemos prescindir de la proteína animal en la acuicultura de camarón, debido a que esta genera dos aminoácidos que aún no han podido ser reemplazados por las características presentes en la proteína animal. Incluso, las certificaciones orgánicas permiten producir camarón con un porcentaje determinado de proteína animal.
Molina expresa que sustituir productos por ingredientes alternativos disminuye la presión sobre los recursos marinos, contribuyendo a la conservación de las poblaciones de peces silvestres. Ingredientes como las harinas de insectos, biomasa microbiana, algas y subproductos agrícolas pueden producirse con menores recursos naturales, reduciendo las emisiones de gases de efecto invernadero y el impacto ambiental.
Además, señala que la diversidad en las fuentes de ingredientes mitiga el riesgo asociado con la dependencia de un solo recurso, como el colapso de pesquerías o las fluctuaciones en los precios de ingredientes tradicionales. Adicionalmente, el uso de subproductos agrícolas y de otras industrias ayuda a minimizar el desperdicio y fomenta una economía circular.
Un ejemplo destacado de tecnologías en ingredientes alternativos es una proteína de origen vegetal producida mediante una tecnología de fermentación. Este producto apoya a la sostenibilidad en la producción de alimento para camarones al reciclar harinas de semillas oleaginosas, subproductos
comunes del proceso de extracción de aceite. Según Stephanie Armstead, directora de sostenibilidad en Houdek (anteriormente Prairie AquaTech), la proteína fermentada ofrece una fuente de proteína más digestible, con factores antinutricionales reducidos y de mayor calidad, que puede reemplazar otras proteínas con un mayor impacto ambiental.
El impacto positivo de este ingrediente novel es aún mayor cuando se obtiene de proveedores responsables y trazables, como es el caso de Houdek. Su proceso de fermentación patentado hidroliza y concentra la proteína en el ingrediente, lo que aumenta la biodisponibilidad de los nutrientes, mejorando así la absorción y la tasa de digestión. Armstead señala que ha mejorado las tasas de conversión alimenticia (FCR), lo que resulta en una producción de camarón más eficiente, donde “los productores podrán ahorrar dinero, tiempo y proporcionar un producto de mayor calidad”.
Aditivos nutricionales en el alimento
En los últimos 30 años, el manejo de enfermedades ha sido uno de los principales desafíos en el sector camaronero y la acuicultura, dificultando su crecimiento y desarrollo. Estas enfermedades no solo limitan la capacidad de criar camarones a la densidad deseada y alcanzar las metas de producción, sino que también implican altos costos en tratamientos, afectando directamente la rentabilidad y sostenibilidad del sector. En este contexto, los aditivos nutricionales en las dietas balanceadas se han posicionado como una solución para enfrentar este desafío.
Molina señala que la adición de nutrientes específicos, como beta-glucano, ácidos grasos omega-3 y nucleótidos, puede mejorar la resistencia a enfermedades y la recuperación de los camarones enfermos. Además, menciona que la inclusión de agentes antimicrobianos naturales, como extractos de hierbas y especias, puede ayudar a controlar la carga de patógenos en el sistema digestivo de los camarones.
El profesor e investigador de la Universidad
Federal de Santa Catarina (Brasil), José Luiz Mouriño, destaca que añadir nucleótidos en las dietas contribuye a la síntesis de proteínas, promoviendo la formación de células en el animal, acelerando su crecimiento y fortaleciendo su robustez. También facilita el crecimiento de tejidos y cumple una función inmunoestimulante.
Asimismo, Mouriño, quien también es consultor internacional para la empresa dsmfirmenich, explica que para asegurar un buen crecimiento y un sistema inmunológico fuerte, es fundamental mantener niveles óptimos de vitaminas C y E en la dieta. Esto se debe a que, a medida que el camarón crece más rápido, realiza mudas de exoesqueleto más frecuentes. Durante este proceso de crecimiento, al producir proteínas y enzimas, también genera radicales libres, específicamente especies reactivas de oxígeno y nitrógeno. Las vitaminas C y E ayudan a neutralizar estos compuestos, que pueden afectar la salud del camarón.
Otro aditivo destacado es la astaxantina proveniente de algas, como la Haematococcus pluvialis. Mouriño resalta su poder antioxidante, que reduce el estrés oxidativo en los tejidos del camarón, mejorando su salud. Destaca que la astaxantina es un antioxidante mucho más efectivo que la vitamina C y, además, actúa como pigmento, dándole al camarón un color rojizo más intenso tras la cocción. Este color aumentará su valor en el mercado, ya que muchos mercados internacionales pagan más por camarones con mayor pigmentación, similar a lo que ocurre con el salmón o el atún.
Tecnologías enfocadas en la mejora del ambiente y salud
De la misma forma, el consultor expresa que otra tecnología que ha permitido el crecimiento sostenible de la acuicultura de camarón es la biorremediación.
De acuerdo con el coordinador técnico de SSP, Leonardo Maridueña, la biorremediación es vital porque ayuda a mantener la calidad del agua y del suelo en los sistemas de
producción. Esto es esencial para la salud de los organismos cultivados y el entorno natural.
Maridueña, quien es doctor en Ambiente, Acuicultura y Pesca, describe que a través de la biorremediación, microorganismos y otros organismos naturales se utilizan para descomponer o transformar contaminantes y desechos orgánicos que se acumulan en el suelo de las piscinas. Esto beneficia la calidad del agua y reduce el riesgo de toxicidad para los animales.
“La acuicultura genera residuos que pueden perjudicar los ecosistemas si no se gestionan adecuadamente. La biorremediación reduce la carga de contaminantes que podrían afectar cuerpos de agua cercanos, ayudando a mantener la biodiversidad y proteger los ambientes acuáticos”, expresa.
Spoiler
Alert: siguientes pasos
La industria camaronera sigue avanzando con nuevas tecnologías que están cambiando cada parte del proceso de producción. Estos avances han traído mejoras importantes en áreas clave. Un buen ejemplo es cómo se mide y controla el metabisulfito. Se han desarrollado nuevas metodologías que permiten hacer estas mediciones de manera más rápida, eficiente y precisa, sin el uso de químicos perjudiciales para la salud de los operadores y consumidores del producto. El beneficio de este avance se extienede no solo al momento de la cosecha, sino que también a las plantas donde se procesa el camarón y al consumidor final.
Asier Albizu, CEO de Biolan, la empresa que ha creado esta nueva metodología para la medición de metabisulfito certificada por la Association of Analytical Communities (AOAC), destaca la importancia de poder definir la calidad del producto en poco tiempo. En la tercera y última parte de esta serie de artículos, nos adentraremos más en este tema. Exploraremos cómo diversas mejoras tecnológicas están ayudando a las plantas de procesamiento de camarón y contribuyendo al secreto del éxito de la industria camaronera en Ecuador•
Para más información sobre este artículo, contactar a: pnath@sustainableshrimp.org
ARTÍCULOS TÉCNICOS ÍNDICE
Edición 161 - Octubre 2024
Síndrome de las Bolitas en criaderos de Penaeus vannamei en América Latina
Impacto de diversas condiciones de procesamiento en el nivel de sodio y en los atributos sensoriales del camarón
Primer registro de quistes de microalga Chattonella, un nuevo desafío para la producción de Penaeus vannamei
Bienestar Animal: un “nuevo término” largamente aplicado en la industria camaronera ecuatoriana
Guía práctica para el traslado de camarón vivo desde la finca hasta la planta procesadora
El Pacto Verde Europeo y sus implicaciones para el sector exportador camaronero
Estimación de parámetros genéticos para el crecimiento y rasgos de resistencia al WSSV en Penaeus vannamei
Síndrome de las Bolitas en criaderos de Penaeus vannamei en América Latina
Autores:
Pablo Intriago 1,2*
Bolívar Montiel 2
Mauricio Valarezo 2
Xavier Romero 3
Kelly Arteaga 2
Nicole Cercado 2
Milena Burgos 2
Andrew P. Shinn 4,5
Alejandra Montenegro 2
Andrés Medina 2
Jennifer Gallardo 2
1 South Florida Farming Corp., 13811 Old Sheridan St, Southwest Ranches, FL 33330, USA.
2 South Florida Farming Lab., Av. Miguel Yunez, Km 14.5 vía a Samborondón, Almax 3 Etapa 1
Lote 3 Bodega 2, Samborondón, Guayas, Ecuador.
3 Ficus 302 y Antonio Sánchez, Calle 11 N-O, Guayaquil, Ecuador.
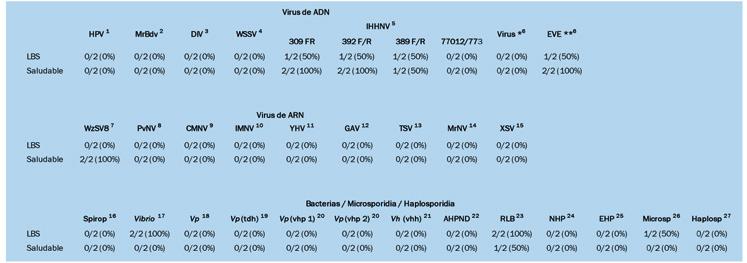
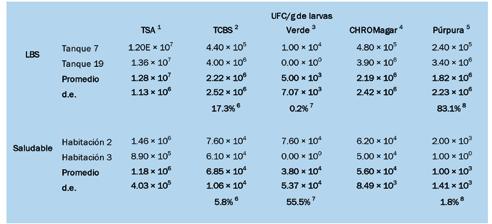
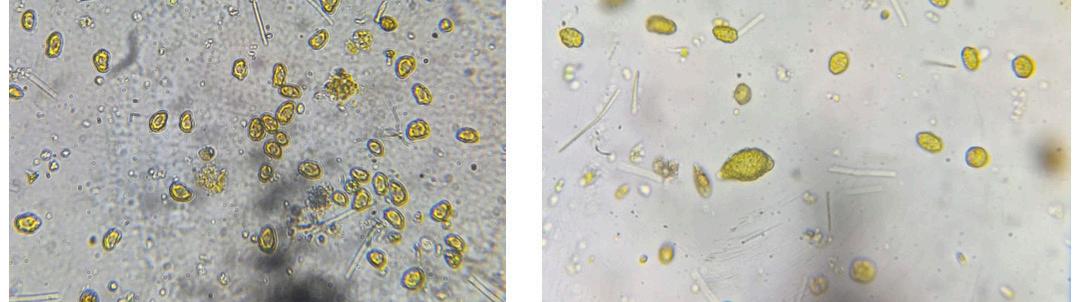
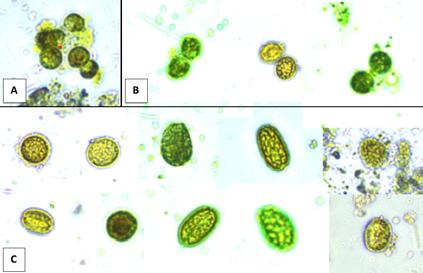
En septiembre de 2023, varios criaderos en América Latina experimentaron tasas de mortalidad significativas: hasta el 90% en el estadio de zoea 2 de Penaeus vannamei. Las observaciones de montajes frescos revelaron estructuras que se asemejaban a gotas lipídicas, similares a las que se observan en una condición conocida como “Síndrome de las Bolitas”. Los exámenes histopatológicos rutinarios identificaron células y tejidos desprendidos en los tractos digestivos de los zoeas afectados, en contraste con el contenido típico de células de algas en los zoeas saludables. Las pruebas de reacción en cadena de la polimerasa (PCR) para más de 20 patógenos conocidos de camarón indicaron diferencias mínimas entre los lotes enfermos y saludables. Ambos grupos dieron negativo para la enfermedad de Necrosis Hepatopáncreatica Aguda (AHPND), pero positivo para especies de Vibrio y bacterias similares a Rickettsia en las muestras enfermas. Los análisis histológicos de los zoeas afectados revelaron una degeneración característica del tejido en el hepatopáncreas, formando esferas que eventualmente migraron hacia el intestino superior, el intestino medio y las cecas del intestino medio, una patología identificada como Síndrome de las Bolitas (BS). Las evaluaciones microbiológicas revelaron especies de Vibrio en concentraciones de 10^6 UFC zoea/g en los zoeas afectados, aproximadamente dos órdenes de magnitud más alto que en los zoeas saludables.
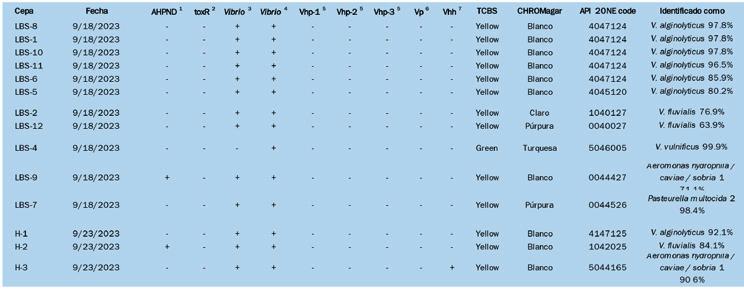
El aislamiento de bacterias tanto de zoeas saludables como afectados por BS en agar tiosulfato-citrato-sal de bilis-sucrosa (TCBS) y CHROMagar™ (París, Francia), seguido de la identificación utilizando API 20E, identificó seis cepas de Vibrio alginolyticus A pesar de las similitudes con el BS en montajes frescos, se observaron diferencias histopatológicas distintas, particularmente la presencia de células descamadas en los intestinos y variaciones en los lóbulos hepatopancreáticos. Este estudio resalta la necesidad crítica de investigar más a fondo para comprender completamente la etiología y patología de este síndrome en el estadio de zoea 2 de P. vannamei para desarrollar estrategias de mitigación efectivas para las operaciones de criaderos.
Durante finales de la década de 1980, la década de 1990 y principios de la de 2000, las patologías más predominantes en los criaderos de América Latina eran el Síndrome de Bolitas Larvales (LBS), el Síndrome de Zoea 2 y el Síndrome de Muda de Mysis [1–3]. Durante el mismo período, las principales patologías reportadas en postlarvas eran las bacterias luminiscentes [4–7]. Después de 2015, los brotes de enfermedades con altas tasas de mortalidad se vieron más comúnmente en la producción de postlarvas, por ejemplo, AHPND en postlarvas (PL-AHPND), enfermedad de postlarvas translúcidas (TPD), etc. [8–11]. El LBS es una condición caracterizada inicialmente por una patología distintiva del hepatopáncreas, donde se produce una descamación celular del epitelio del hepatopáncreas, formando esferas que eventualmente se trasladan al intestino superior [1]. La patología macroscópica del LBS normalmente se desarrolla en cuestión de horas, pasando de zoeas saludables y bien alimentados a animales moribundos y vacíos. Al mismo tiempo, las larvas se vuelven bioluminiscentes, lo que se acompaña de cambios en el comportamiento y pérdida de apetito [2]. Aunque el LBS puede ocurrir durante las etapas postlarvales (PL), se han observado mortalidades masivas, que alcanzan hasta el 90%, principalmente en las etapas tempranas de zoea y mysis. Una característica distintiva del LBS es la presencia de bolitas en el hepatopáncreas, que eventualmente migran al intestino.
LBS ha sido asociado con infecciones de Vibrio spp. [2,12]. El Síndrome de Zoea 2 solo se ha asociado con V. harveyi y V. alginolyticus [3,13], y la especie asociada con el Síndrome de Mysis nunca ha sido identificada. La bioluminiscencia nocturna observada en criaderos tanto en Asia como en las Américas se atribuyó típicamente a Vibrio luminiscentes [2,4–7]. En tiempos más recientes, los brotes de mortalidades súbitas y agudas en criaderos de camarones peneidos suelen comenzar en las etapas PL, pasando de un estado activo y aparentemente saludable a moribundo y muerto. La velocidad y virulencia con que ocurren estas mortalidades masivas se han observado en muchas instalaciones de producción, y en la mayoría de los casos se han asociado o vinculado con diferentes
cepas de Vibrio [8–10]. Intriago et al. [10] proporcionaron evidencia de que la causa de estos eventos de mortalidad rápida era una especie de Vibrio que portaba los mismos plásmidos que el VpAHPND reportado como causante de la enfermedad de Necrosis Hepatopancreática Aguda (AHPND) en estanques de cultivo en otras regiones. La condición fue tentativamente denominada AHPND en postlarvas (PL-AHPND) para diferenciarla de otras patologías que afectan a las larvas de camarón peneido. En Asia, la enfermedad de postlarvas translúcidas (TPD) ha sido la principal causa de enfermedades y mortalidades larvales, siendo el agente causante una cepa de V. parahaemolyticus portadora de un gen de hemolisina (aislamiento Vp-JS20200428004-2) [8,9,11]. En India, se describió una condición similar referida como Síndrome de Zoea 2 que afecta a los zoeas 2 de Penaeus vannamei [14].
En septiembre de 2023, algunas instalaciones de cultivo en la región de América Latina experimentaron altas mortalidades en los estadios de zoea 2 y 3. Dada la alta tasa de mortalidad, estos tanques fueron descartados. No se ha reportado el agente causal asociado con estas mortalidades. La observación microscópica de las larvas reveló la presencia de “bolitas” (esferas) en el hepatopáncreas. Generalmente, los indicadores clínicos y la apariencia macroscópica en montajes húmedos coincidían con lo que se había identificado previamente como LBS. El presente estudio informa sobre los hallazgos microbiológicos, las pruebas de reacción en cadena de la polimerasa (PCR) y los hallazgos histológicos en animales saludables y enfermos afectados por esta condición.
Razonamiento del estudio
Las muestras fueron enviadas por clientes para monitorear el estado de salud de los camarones o brotes de enfermedades en sus criaderos. Los animales (zoeas) utilizados como controles estaban saludables y se recolectaron como especímenes de referencia para comparar con los animales afectados. Este estudio no fue una investigación epidemiológica, sino un análisis simple de prevalencia de varios patógenos de camarón en muestras proporcionadas aleatoriamente de un criadero que reportó mortalidades.
Referencias
1. Morales, I. (1992) Observaciones sobre el síndrome de descamación del epitelio digestivo “Bolitas” en larvas de Penaeus vannamei en Ecuador. In: Memorias del Primer Congreso Ecuatoriano de Acuicultura; Escuela Superior Politécnica del Litoral: Guayaquil, Ecuador, 1992; pp. 203–207.
2. Robertson, P.A.W., Calderon, J., Carrera, L., Stark, J.K., Zherdmand, M. y Austin, B. (1998) Experimental Vibrio harveyi infections in Penaeus vannamei larvae. Dis. Aquat. Org., 32, 151–155.
3. Vandenberghe, J., Verdonck, L., Robles-Arozarena, R., Rivera, G., Bolland, A., Balladares, M., Gomez-Gil, B., Calderon, J., Sorgeloos, P. y Swings, J. (1999) Vibrios associated with Litopenaeus vannamei larvae, postlarvae, broodstock, and hatchery probionts. Appl. Environ. Microbiol., 65, 2592–2597.
4. Baticados, M.C.L., Lavilla-Pitogo, C.R., Cruz-Lacierda, E.R., de la Pena, L.D. y Sunaz, N.A. (1990) Studies on the chemical control of luminous bacteria Vibrio harveyi and V. splendidus isolated from diseased Penaeus monodon larvae and rearing water. Dis. Aquat. Org., 9, 133–139.
5. Lavilla-Pitogo, C.R., Baticados, M.C.L., Cruz-Lacierda, E.R. y de la Pena, L.D. (1990) Occurrence of luminous bacterial disease of Penaeus monodon larvae in the Philippines. Aquaculture, 91, 1–13.
6. Song, Y.L. y Lee, S.P. (1993) Characterization and ecological implication of luminous Vibrio harveyi isolated from tiger shrimp (Penaeus monodon). Bull. Inst. Zool. Acad. Sin., 32, 217–220.
7. Nithimathachoke, N., Pratanpipat, P., Thongdaeng, K., Withyachumnarnkul, B. y Nash, G. (1995) Luminous bacterial infection in pond reared Penaeus monodon Asian Shrimp News, 23, 1–4.
8. Zou, Y., Xie, G., Jia, T., Xu, T., Wang, C., Wan, X., Li, Y., Luo, K., Bian, X., Wang, X. et al. (2020) Determination of the infectious agent of Translucent Post-Larva Disease (TPD) in Penaeus vannamei Pathogens, 9, 741.
9. Yang, F., Xu, L., Huang, W. y Li, F. (2022) Highly lethal Vibrio parahaemolyticus strains cause acute mortality in Penaeus vannamei post larvae. Aquaculture, 548, 737605.
10. Intriago, P., Medina, A., Espinoza, J., Enriquez, X., Arteaga, K., Aranguren, L.F., Shinn, A.P. y Romero, X. (2023) Acute mortality of Penaeus vannamei larvae in farm hatcheries associated with the presence of Vibrio sp. carrying the VpPirAB toxin genes. Aquac. Int., 31, 3363–3382.
11. Yang, F., You, Y., Lai, Q., Xu, L. y Li, F. (2023) Vibrio parahaemolyticus becomes highly virulent by producing Tc toxins. Aquaculture, 576, 739817.
12. Intriago, P. y Jimenez, R. (1999) The effect of Vibrio parahaemolyticus, a luminescent bacteria isolated from grow out ponds in Penaeus vannamei larviculture. In: Book of Abstracts World Aquaculture ’99: Bridging the Gap, Proceedings of the Annual International Conference and Exposition of the World Aquaculture Society, Sydney, NSW, Australia, 26 April–2 May 1999, World Aquaculture Society: Sydney, NSW, Australia 1999. p. 353.
13. Wiradana, P.A., Sani, M.D., Mawli, R.E., Ashshoffa, F.N.D., Widhiantara, I.G. y Mukti, A. (2022) Monitoring the occurrence of Zoea Syndrome (ZS) in Pacific white shrimp (Litopenaeus vannamei) larval from several hatcheries in East Java, Indonesia. IOP Conf. Ser. Earth Environ. Sci., 1036, 012003.
14. Kumar, S.T., Vidya, R., Kumar, S., Alavandi, S.V. y Vijayan, K.K. (2017) Zoea-2 syndrome of Penaeus vannamei in shrimp hatcheries. Aquaculture, 479, 759–767.
15. Buller, N.B. (2004) Bacteria from Fish and Other Aquatic Animals: A Practical Identification Manual. CABI Publishing: Cambridge, MA, USA. eISBN 978-0-85199.
16. Phromjai, J., Boonsaeng, V., Withyachumnarnkul, B. y Flegel, T.W. (2002) Detection of hepatopancreatic parvovirus in Thai shrimp Penaeus monodon by in situ hybridization, dot blot hybridization and PCR amplification. Dis. Aquat. Org., 51, 227–232.
17. Gangnonngiw, W., Bunnontae, M., Kayansamruaj, P., Senapin, S., Srisala, J., Flegel, T.W. y Wongprasert, K. (2023) A novel ssDNA Bidnavirus in the giant freshwater prawn
Lamentablemente, los propietarios y gerentes no divulgan información sobre los protocolos utilizados ni sobre los parámetros físicos y químicos del cultivo, ya que creen que podría dañar la reputación del criadero. En consecuencia, no podemos realizar muestreos más extensos ni proporcionar información detallada relacionada con la cultura larval.
Materiales y métodos
Recolección de muestras
Se tomaron muestras de larvas de P. vannamei de dos criaderos en América Latina (los detalles precisos de ubicación se mantienen en reserva para respetar la solicitud de confidencialidad de las instalaciones). Se tomaron muestras para microbiología, PCR e histopatología de dos tanques de cada criadero. Los criaderos fueron seleccionados porque uno de ellos había reportado una alta mortalidad en los estadios de zoea 2–3, mientras que el otro criadero, con zoeas 2–3 aparentemente saludables, fue seleccionado como control. Cabe señalar que los camarones muestreados para PCR e histología eran individuos diferentes de las mismas poblaciones. Para proteger la privacidad del cliente, el país y la ubicación precisa de cada instalación de cultivo de donde se obtuvieron las muestras no serán divulgados.
Microbiología de las larvas
La concentración total de bacterias en las larvas se determinó sumergiendo primero las larvas (es decir, 1 g o aproximadamente >2000 zoea de estadio 3) en un tamiz de nylon de 200 µm durante 2–3 minutos en una solución de 50 ppm de cloro activo preparada con agua de mar a 35 ppt. Las zoeas se enjuagaron posteriormente con agua de mar estéril en exceso. El peso de las larvas se registró utilizando una balanza digital Metler con una precisión de 0.01 g y luego se colocaron en un mortero con aproximadamente 1 g de arena de playa esterilizada y 10 mL de agua de mar estéril, que se utilizó para facilitar la molienda de la mayoría de las larvas. Después de moler la muestra, se diluyó secuencialmente en tubos de ensayo con agua de mar estéril a 1 × 10^2, 10^3 y 10^4. Luego, se colocaron 100 μL de la dilución relevante en duplicado en placas de Petri que contenían agar de soja
tríptica (TSA, Difco) o agar tiosulfato-citratobilis-sucrosa (TCBS, Difco) o en CHROMagar™ Vibrio, y se incubaron durante 24–48 horas a 30 °C. Posteriormente, se registró el número de unidades formadoras de colonias (UFC) en cada placa. Las diluciones se basaron en obtener >20 y <200 colonias por placa. La identificación bacteriana se realizó y las características bioquímicas se examinaron utilizando un Kit API 20E [15].
Métodos de PCR
Se extrajo el ADN de larvas enteras fijadas en alcohol al 95% utilizando un kit de ADN de tejido E.Z.N.A. de Omega (Omega Biotek, Inc., Norcross, GA, EE. UU.) siguiendo el protocolo del fabricante. En resumen, cada muestra de 1 g se trituró utilizando un pistón de microcentrífuga. Aproximadamente 200 mg del tejido se transfirieron a un tubo Eppendorf limpio de 1.5 mL, luego se añadieron 500 μL de buffer de lisis de tejido (TL, Omega, Bio-Tek E.Z.N.A.) y 25 μL de solución de proteasa de Omega Biotek (OB, Omega, Bio-Tek E.Z.N.A). La muestra se agitó en vórtice y se incubó en un termobloque a 55 °C durante aproximadamente 3 horas, con agitación cada 30 minutos. El ARN se eliminó añadiendo 4 μL de RNasa A (100 mg/ mL), mezclando y manteniendo la muestra a temperatura ambiente durante 2 minutos. La muestra se centrifugó a 13,500 RPM durante 5 minutos y el sobrenadante se transfirió cuidadosamente a un nuevo tubo Eppendorf de 1.5 mL. A esto se le añadieron 220 μL de buffer BL (Omega, Bio-Tek E.Z.N.A.), se agitó en vórtex y se incubó a 70 °C durante 10 minutos. Luego, se añadieron 220 μL de etanol al 100% y se agitó; los contenidos se pasaron a través de una columna HiBind® DNA Mini en un tubo de recolección de 2 mL. Las columnas se centrifugaron a 13,500 RPM durante 1 minuto y el filtrado se descartó. Posteriormente, se añadieron 500 μL de buffer HBC (Omega, Bio-Tek E.Z.N.A.), diluido con isopropanol al 100%, a la columna, y la muestra se centrifugó a 13,500 RPM durante 30 segundos. El filtrado se descartó, la columna se lavó dos veces con 700 μL de buffer de lavado de ADN diluido con etanol al 100% y la muestra se centrifugó a 13,500 RPM durante 30 segundos. El filtrado se descartó y este paso se repitió. La columna se centrifugó a 13,500 RPM durante 2 minutos para secarla. La columna seca se colocó en un nuevo tubo Eppendorf libre de
nucleasas de 1.5 mL y se añadieron 100 μL de buffer de elución, previamente calentado a 70 °C. La muestra se dejó reposar durante 2 minutos antes de centrifugarla a 13,500 RPM durante 1 minuto. Este paso de elución se repitió. El ADN eluido se almacenó a −20 °C hasta su uso.
El ARN se extrajo de larvas enteras, tejidos u órganos fijados en alcohol al 90% siguiendo el protocolo del fabricante (Omega, Bio-Tek E.Z.N.A. Total RNA Kit (TRK)). Aproximadamente 200 mg de tejido se trasladaron a un tubo Eppendorf limpio de 1.5 mL. A esto se le añadieron 700 μL de Buffer de Lisis TRK (Omega, Bio-Tek E.Z.N.A) y el tubo se dejó a temperatura ambiente durante aproximadamente 3 horas con agitación vórtex cada 30 minutos. La muestra se centrifugó a 13,500 RPM durante 5 minutos y el sobrenadante se transfirió cuidadosamente a un nuevo tubo Eppendorf de 1.5 mL al que se le añadieron 420 μL de etanol al 70%. Después de agitar para mezclar bien, los contenidos se pasaron a través de una Columna HiBind® RNA Mini en un tubo de recolección de 2 mL. Las columnas se centrifugaron a 13,500 RPM durante 1 minuto y el filtrado se descartó. Posteriormente, se añadieron 500 μL de Buffer de Lavado de ARN I (Omega, BioTek E.Z.N.A) a la columna y la muestra se centrifugó a 13,500 RPM durante 30 segundos. El filtrado se descartó y la columna se lavó dos veces con 500 μL de Buffer de Lavado de ARN II diluido con etanol al 100%. La columna se centrifugó a 13,500 RPM durante 1 minuto para secarla. El filtrado se descartó y este paso se repitió. La columna se centrifugó a 13,500 RPM durante 2 minutos para secarla completamente. La columna seca se colocó en un nuevo tubo Eppendorf libre de nucleasas de 1.5 mL y se añadieron 70 μL de agua libre de nucleasas. La muestra se centrifugó a 13,500 RPM durante 2 minutos. Este paso de elución se repitió. El ARN eluido se almacenó a −70 °C hasta su uso. Los patógenos que se analizaron se enumeran a continuación.
Se realizaron análisis de PCR para los siguientes patógenos en muestras de animales: Hepanhamaparvovirus (DHPV) [16]; Macrobrachium bidnavirus (MrBdv) [17]; Virus Iridiscente de Decápodos 1 (DIV1) [18]; Virus del Síndrome de Mancha Blanca
(WSSV) [19]; Virus de Necrosis Hipodérmica e Hematopoyética Infecciosa (IHHNV) [20–23]; Virus de Camarón de Wenzhou 8 (WzSV8) [24]; Nodavirus de P. vannamei [25]; Nodavirus de Mortalidad Encubierta (CMNV) [26]; Virus de Mionecrosis Infecciosa (IMNV) [27]; Virus de la Cabeza Amarilla (YHV) [28]; Virus Asociado a Branquias (GAV) [28]; Virus del Síndrome de Taura (TSV) [29,30]; Nodavirus de Macrobrachium (MrNV) [31]; Virus Extra Pequeño (XSV) asociado generalmente con MrNV [32]; Spiroplasma [33]; Vibrio spp. (fragmento del gen 16S rRNA específico de Vibrio) [34]; V. parahaemolyticus (gen de colagenasa) [35]; V. parahaemolyticus (gen vhvp-1 y gen vhvp-2) [36]; V. parahaemolyticus (gen tdh) [37]; V. harveyi (gen vhh) [37]; bacterias similares a Rickettsia (RLB) [38]; bacterias de Necrosis Hepatopancreática (NHP-B) [39]; Ecytonucleospora (Enterocytozoon) hepatopenaei (EHP) [40]; Microsporidia no-EHP [41]; enfermedad de Necrosis Hepatopancreática Aguda (AHPND) [42]; y Haplosporidia [43].
Además, se realizó un cribado de los siguientes patógenos en bacterias aisladas: Vibrio spp. (fragmento del gen 16S rRNA específico de Vibrio) [34]; V. parahaemolyticus (gen de colagenasa) [35]; V. parahaemolyticus (gen vhvp-1 y gen vhvp2) [36]; V. parahaemolyticus (gen tdh) [37]; V. harveyi (gen vhh) [37]; y gen toxR [44].
Histopatología
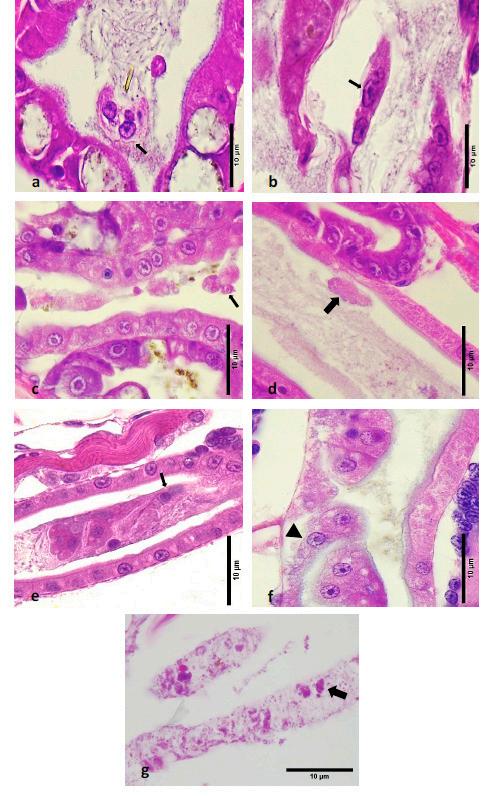
Para el análisis histológico, las muestras se prepararon siguiendo los procedimientos descritos por Bell y Lightner [45]. En resumen, las larvas se fijaron en alcohol de Davidson, formalina y ácido acético (AFA) utilizando al menos 1 g de larvas de cada tanque. Estas se fijaron durante al menos 24 horas antes de su procesamiento para la inclusión de tejidos y secciones histológicas de rutina. Las secciones de tejido de 5 μm de grosor se tiñeron con hematoxilina y eosina (H&E) y con la tinción de Gram de Twort para diferenciar bacterias Gram-positivas de Gram-negativas (CP Lab Chemicals, Novato, CA, EE. UU.). Para cada muestra de larvas recolectadas
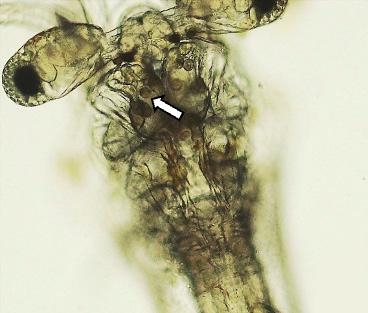
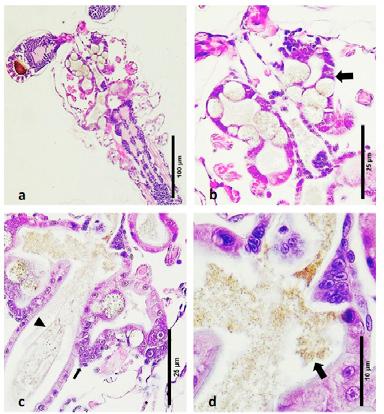
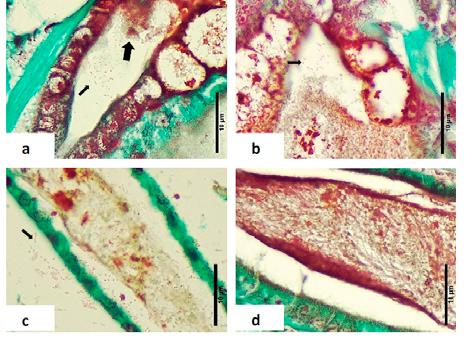
Figura 1. Montaje fresco de larvas de zoea 2. (a) Una larva saludable con un tracto digestivo normal. (b–d) Larva afectada, observe la falta de contenido alimenticio y la presencia de “bolitas” (flechas) en el hepatopáncreas, así como en el intestino superior (cabeza de flecha).
18. Qiu, L., Chen, M.M., Wan, X.Y., Li, C., Zhang, Q.L., Wang, R.Y., Cheng, D.Y., Dong, X., Yang, B., Wang, X.H., et al. (2017) Characterization of a new member of Iridoviridae, Shrimp Hemocyte Iridescent Virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci. Rep., 7, 11834.
19. Lo, C.F., Ho, C.H., Peng, S.E., Chen, C.H., Hsu, H.C., Chiu, Y.L., Chang, C.F., Liu, K.F., Su, M.S., Wang, C.H., et al. (1996) White Spot Syndrome Baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Organ., 27, 215–225.
20. Nunan, L.M., Poulos, B.T. y Lightner, D.V. (2000) Use of polymerase chain reaction (PCR) for the detection of infectious hypodermal and hematopoietic necrosis virus (IHHNV) in penaeid shrimp. Mar. Biotechnol., 2, 319–328.
21. Tang, K.F.J., Durand, S.V., White, B.L., Redman, R.M., Pantoja, C.R. y Lightner, D.V. (2000) Postlarvae and juveniles of a selected line of Penaeus stylirostris are resistant to infectious hypodermal and hematopoietic necrosis virus infection. Aquaculture, 190, 203–210.
22. Tang, K.F.J. y Lightner, D.V. (1999) A yellow-head virus gene probe: Application to in situ hybridization and determination of its nucleotide sequence. Dis. Aquat. Organ., 35, 165–173.
23. Tang, K.F.J., Navarro, S.A. y Lightner, D.V. (2007) PCR assay for discriminating between infectious hypodermal and hematopoietic necrosis virus (IHHNV) and the virus-related sequences in the genome of Penaeus monodon Dis. Aquat. Organ., 74, 165–170.
24. Srisala, J., Thaiue, D., Sanguanrut, P., Aldama-Cano, D.J., Flegel, T.W. y Sritunyalucksana, K. (2021) Potential universal PCR method to detect decapod hepanhamaparvovirus (DHPV) in crustaceans. Aquaculture, 541, 736782.
25. Tang, K.F.J., Pantoja, C.R., Redman, R.M. y Lightner, D.V. (2007) Development of in situ hybridization and RT-PCR assay for the detection of a nodavirus (PvNV) that causes muscle necrosis in Penaeus vannamei Dis. Aquat. Organ., 75, 183–190.
26. Zhang, Q., Xu, T., Wan, X., Liu, S., Wang, X., Li, X., Dong, X., Yang, B. y Huang, J. (2017) Prevalence and distribution of covert mortality nodavirus (CMNV) in cultured crustacean. Virus Res., 233, 113–119.
27. Poulos, B.T., Lightner, D.V. (2006) Detection of infectious myonecrosis virus (IMNV) of penaeid shrimp by reversetranscriptase polymerase chain reaction (RT-PCR). Dis. Aquat. Org., 73, 69–72.
28. Mohr, P.G., Moody, N.J., Hoad, J., Williams, L.M., Bowater, R.O., Cummins, D.M., Cowley, J.A. y Crane, M.S. (2015) New yellow head virus genotype (YHV7) in giant tiger shrimp Penaeus monodon indigenous to northern Australia. Dis. Aquat. Org., 115, 263–268.
29. Nunan, L.M., Poulos, B.T. y Lightner, D.V. (1998) Reverse transcription polymerase chain reaction (RT-PCR) used for the detection of Taura Syndrome Virus (TSV) in experimentally infected shrimp. Dis. Aquat. Organ., 34, 87–91.
30. Navarro, S.A., Tang, K.F.J. y Lightner, D.V. (2009) An improved Taura syndrome virus (TSV) RT-PCR using newly designed primers. Aquaculture, 293, 290–292.
31. Gangnonngiw, W., Bunnontae, M., Phiwsaiya, K., Senapin, S. y Dhar, A.K. (2020) In experimental challenge with infectious clones of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV), MrNV alone can cause mortality in freshwater prawn (Macrobrachium rosenbergii). Virology, 540, 30–37.
Senapin, S., Jaengsanong, C., Phiwsaiya, K., Prasertsri, S., Laisutisan, K., Chuchird, N., Limsuwan, C. y Flegel, T.W. (2012) Infections of MrNV (Macrobrachium rosenbergii nodavirus) in cultivated whiteleg shrimp Penaeus vannamei in Asia. Aquaculture, 338–341, 41–46.
33. Nunan, L.M., Pantoja, C.R., Salazar, M., Aranguren, F. y Lightner, D.V. (2004) Characterization and molecular methods for detection of a novel Spiroplasma pathogenic to Penaeus vannamei Dis. Aquat. Organ., 62, 255–264.
34. Yong, L., Guanpin, Y., Hualei, W., Jixiang, C., Xianming, S., Guiwei, Z., Qiwei, W. y Sun, X. (2006) Design of Vibrio 16S rRNA gene specific primers and their application in the analysis of seawater Vibrio community. J. Ocean. Univ. China, 5, 157–164.
35. Khimmakthong, U. y Sukkarun, P. (2017) The spread of
Tabla 1. Resultados de PCR de ADN, ARN, bacterias, Microsporidia EHP y no EHP, y Haplosporidia en 1 g de zoea de tanques tanto LBS como saludables.
1 Hepanhamaparvovirus (DHPV) [16]; 2 Macrobrachium Bidnavirus (MrBdv) [17]; 3 Virus Iridiscente de Decápodos 1 (DIV1) [18]; 4 Virus del Síndrome de Manchas Blancas (WSSV) [19]; 5 Virus de Necrosis Hipodérmica e Hematopoyética (IHHNV) [20–23]; 6 IHHNV * como infeccioso o ** EVE; 7 Virus del Camarón de Wenzhou 8 (WzSV8) [24]; 8 Nodavirus de P. vannamei [25]; 9 Nodavirus de Mortalidad Encubierta (CMNV) [26]; 10 Virus de Mionecrosis Infecciosa (IMNV) [27]; 11 Virus de Cabeza Amarilla (YHV) [28]; 12 Virus Asociado a Branquias (GAV) [28]; 13 Virus del Síndrome de Taura (TSV) [29,30]; 14 Nodavirus de Macrobrachium (MrNV) [31]; 15 Virus Extra Pequeño (XSV) generalmente asociado con MrNV [32]; 16 Spiroplasma [33]; 17 Vibrio spp. fragmento del gen 16S rRNA específico de Vibrio [34]; 18 V. parahaemolyticus. Gen colagenasa [35]; 19 V. parahaemolyticus. Genes vhvp-1 y vhvp-2 [36]; 20 V. parahaemolyticus. Gen tdh [37]; 21 V. harveyi. Gen vhh [37]; 22 Enfermedad Aguda de Necrosis Hepatopancreática (AHPND) [42]; 23 Bacterias Tipo Rickettsia (RLB) [38]; 24 Bacterias de Hepatopancreatitis Necrótica (NHP-B) [39]; 25 Ecytonucleospora (Enterocytozoon) hepatopenaei (EHP) [40]; 26 Microsporidia no EHP [41]; 27 Haplosporidia [43].
de cada tanque, se preparó un bloque, cada bloque conteniendo aproximadamente 1 g de zoea en los estadios 2–3. Se cortaron cuatro secciones de tejido de cada bloque de parafina.
Resultados
La observación microscópica de las larvas reveló la presencia de “bolitas” (esferas) en el hepatopáncreas (Figura 1a–d). En las preparaciones frescas de zoea 2–3 sanos, se observó material algal en el tracto digestivo, que le daba una coloración marrón-dorada (Figura 1a). En comparación, las larvas afectadas/enfermas presentaban un tracto digestivo vacío y material de forma redonda que aparecía como gotas de lípido (Figura 1b–d) y falta de alimento en el intestino. Este signo clínico general es el que usualmente describen los técnicos y biólogos de criaderos de camarones como el conocido “Síndrome de las Bolitas”.
Resultados de PCR
El análisis de PCR de las muestras de zoea tanto saludables como afectadas resultó negativo para diecisiete patógenos
conocidos de camarones, incluidos DHPV, MrBdv, SHIV, WSSV, PvNV, CMNV, IMNV, YHV, GAV, TSV, MrNV, XSV, Spiroplasma, NHPB, EHP, AHPND y Haplosporidia (Tabla 1). Sin embargo, las zoea aparentemente sanas fueron positivas para IHHNV EVE (2/2), WzSV8/PvSV (2/2), RLB (1/2) y Microsporidia (1/2). En comparación, las zoea afectadas por LBS fueron positivas para IHHNV EVE (1/2), para Vibrio spp. (2/2), para RLB (2/2) y para Microsporidia (1/2). En resumen, la única diferencia entre los dos conjuntos de muestras fue la detección de Vibrio en las zoea afectadas por LBS y WzSV8 en las zoea sanas.
Microbiología
Las concentraciones de bacterias heterotróficas totales (TSA) y Vibrio presuntivos (TCBS y CHROMagar™ Vibrio) se muestran en la Tabla 2. Las concentraciones de bacterias totales en las zoea afectadas fueron casi un orden de magnitud superiores a las determinadas para la muestra de zoea sanas. Los valores de Vibrio presuntivos fueron casi dos órdenes de magnitud más altos en las zoea afectadas que en las zoea
sanas. Los Vibrio presuntivos representaron el 17% y el 6% de la población total de bacterias en las zoea enfermas y sanas, respectivamente. Las colonias verdes (en TCBS) y las colonias moradas (en CHROMagar™ Vibrio) representaron el 0,2% y el 82% de las recuperadas de las zoea afectadas, mientras que se encontró una imagen inversa del 56% y el 1% en el análisis de las zoea sanas (Tabla 2).
Se aisló un total de once cepas de las zoea afectadas por LBS y tres de las zoea sanas (Tabla 3a). El perfil bioquímico de las once cepas recuperadas de las zoea afectadas identificó seis como V. alginolyticus, dos como V. fluvialis y una como V. vulnificus, una como Aeromonas (no determinada) y una como Pasteurella (no determinada). De las tres cepas aisladas de las zoea sanas (Tabla 3a), una cepa fue identificada como V. alginolyticus, una como V. fluvialis y una como Aeromonas. Curiosamente, las 14 cepas resultaron positivas para Vibrio por PCR, lo que sugiere que el 27% de los resultados devueltos por los perfiles bioquímicos API 20E fueron falsos negativos. Ninguna
Tabla 2. Resultados de microbiología de zoeas afectadas y saludables.
1 Agar Soytrypticase (TSA, Difco); 2 Agar tiosulfato-citrato-bilis-sacarro (TCBS, Difco); 3 Colonias verdes en TCBS; 4 CHROMagarTM Vibrio; 5 Colonias moradas en CHROMagarTM Vibrio; 6 Porcentaje de Vibrio presuntivos (TCBS) de la población bacteriana total; 7 Porcentaje de colonias verdes de la población total de Vibrio presuntivos (TCBS); 8 Porcentaje de colonias moradas de la población total de Vibrio presuntivos en CHROMagarTM Vibrio.
de las 14 bacterias dio positivo para los genes específicos de V. parahaemolyticus, incluidos ToxR, Vhp-1 y Vhp-2, que codifican las proteínas altamente virulentas de V. parahaemolyticus (Tabla 3a). Las cepas identificadas como Aeromonas y V. fluvialis por el API 20E (LBS-9 y H-2) pero como Vibrio por PCR resultaron positivas para Pir AB (AHPND). La cepa H-3 identificada por API 20E como A. hydrophila fue identificada positivamente como V. harveyi por PCR.
La caracterización bioquímica de las 13 cepas de Vibrio aisladas se muestra en la Tabla 3b. Esta tabla está dividida entre dos tipos de Vibrio aislados de las zoea afectadas por LBS (V. alginolyticus y Vibrio spp. identificados por API 20E) y tres especies de Vibrio aisladas de las zoea sanas. La disponibilidad de fermentación/oxidación de arabinosa fue la principal diferencia entre Vibrio spp. aislados de zoea con LBS y los otros dos grupos de bacterias, a saber, V. alginolyticus de LBS y Vibrio spp. de zoea sanas. Por otro lado, V. alginolyticus de zoea afectadas por LBS puede diferenciarse de los otros dos grupos por su incapacidad para fermentar.
Histopatología
Ocho diapositivas de microscopía teñidas con hematoxilina y eosina (H&E) fueron preparadas, cada una conteniendo aproximadamente 120 larvas de zoea 2–3 en estado normal y de tanques enfermos. Estas diapositivas fueron examinadas
exhaustivamente para identificar diferencias histopatológicas entre larvas saludables y enfermas.
Observaciones en larvas de zoea 2–3 saludables:
Contenido del tracto digestivo: las larvas de zoea 2–3 saludables exhibieron restos de alimento dentro de sus tractos digestivos. El sistema digestivo en esta etapa aún está en desarrollo, y la presencia de restos de alimento indica procesos normales de alimentación y digestión.
Desarrollo del hepatopáncreas: el desarrollo temprano del hepatopáncreas se evidenció a través de la presencia de lóbulos laterales, que son precursores del hepatopáncreas completamente funcional [47]. Estos lóbulos eran claramente visibles en las secciones de tejido (Figura 2a,b).
Integridad de la membrana peritrófica: se observó una membrana peritrófica intacta en las larvas saludables. Esta membrana juega un papel crucial en la protección del epitelio digestivo y en la facilitación de la digestión. Material algal: el material algal consumido por las larvas en esta etapa se vio como pequeños restos marrones dentro del tracto digestivo (Figura 2c,d). Esto indica que las larvas estaban alimentándose activamente de materia algal, lo cual es típico para esta etapa de desarrollo.
Vibrio parahaemolyticus in tissues of the Pacific white shrimp Litopenaeus vannamei analyzed by PCR and histopathology. Microb. Pathog., 113, 107–112.
36. Liu, S., Wang, W., Jia, T., Xin, L., Xu T-t Wang, C., Xie, G., Luo, K., Li, J., Kong, J. y Zhang, Q. (2023) Vibrio parahaemolyticus becomes lethal to post-larvae shrimp via acquiring novel virulence factors. Microbiol. Spectr., 11, e0049223.
37. Wang, Z., Aweya, J.J., Yao, D., Zheng, Z., Wang, C., Zhao, Y., Li, S. y Zhang, Y. (2022) Taurine metabolism is modulated in Vibrio-infected Penaeus vannamei to shape shrimp antibacterial response and survival. Microbiome, 10, 213.
38. Nunan, L.M., Poulos, B., Redman, R., Groumellec, M.L. y Lightner, D.V. (2003) Molecular detection methods developed for a systemic Rickettsia-like bacterium (RLB) in Penaeus monodon (Decapoda: Crustacea). Dis. Aquat. Organ., 53, 15–23.
39. Aranguren, L.F., Tang, K.F.J. y Lightner, D.V. (2010) Quantification of the bacterial agent of necrotizing hepatopancreatitis (NHP-B) by real-time PCR and comparison of survival and NHP load of two shrimp populations. Aquaculture, 307, 187–192.
40. Jaroenlak, P., Sanguanrut, P.,Williams, B.A.P., Stentiford, G.D., Flegel, T.W., Sritunyalucksana, K. y Itsathitphaisarn, O. (2016) A nested PCR assay to avoid false positive detection of the microsporidian Enterocytozoon hepatopenaei (EHP) in environmental samples in shrimp farms. PLoS ONE, 11, e0166320.
41. Pasharawipas, T. y Flegel, T.W. (1994) A specific DNA probe to identify the intermediate host of a common microsporidian parasite of Penaeus merguiensis and P. monodon Asian Fish. Sci., 7, 157–167.
42. Dangtip, S., Sirikharin, R., Sanguanrut, P., Thitamadee, S., Sritunyalucksana, K., Taengchaiyaphum, S., Mavichak, R., Proespraiwong, P. y Flegel, T.W. (2015) AP4 method for two-tube nested PCR detection of AHPND isolates of Vibrio parahaemolyticus Aquacult Rep., 2, 158–162.
43. Utari, H.B., Senapin, S., Jaengsanong, C., Flegel, T.W. y Kruatrachue, M. (2012) A haplosporidian parasite associated with high mortality and slow growth in Penaeus (Litopenaeus) vannamei cultured in Indonesia. Aquaculture 366–367, 85–89.
44. Aguilera-Rivera, D., Prieto-Davó, A., Rodríguez-Fuentes, G., Escalante-Herrera, K.S. y Gaxiola, G. (2019) A vibriosis outbreak in the Pacific white shrimp, Litopenaeus vannamei reared in biofloc and clear seawater. J. Invertebr. Pathol., 167, 17246.
45. Bell, T.A. y Lightner, D.V. (1998) A Handbook of Normal Penaeid Shrimp Histology. The World Aquaculture Society: Baton Rouge, LA, USA. ISBN 0-935868-37-2.
46. Thompson, J.R., Randa, M.A., Marcelino, L.A., TomitaMitchell, A., Lim, E. y Polz, M.F. (2004) Diversity and dynamics of a North Atlantic coastal Vibrio community. Appl. Environ. Microbiol., 70, 4103–4110.
47. Abubakr, M.A. y Jones, D.A. (1992) Functional morphology and ultrastructure of the anterior mid-gut diverticulae of larvae of Penaeus monodon Fabricius, 1798 (Decapoda, Natantia). Crustaceana, 62, 142–158.
48. Intriago, P., Medina, A., Cercado, N., Arteaga, K., Montenegro, A., Burgos, M., Espinoza, J., Brock, J.A., McIntosh, R. y Flegel, T. (2023) Passive surveillance for shrimp pathogens in Penaeus vannamei submitted from 3 Regions of Latin America. Aquac. Rep., 36, 102092.
49. Soto-Rodriguez, S.A., Lozano-Olvera, R., PalaciosGonzalez, D.A., Bolan-Mejia, C. y Rendon-Aguilar, K.G. (2019) Characterization and growth conditions of Vibrio parahaemolyticus strains with different virulence degrees that cause acute hepatopancreatic necrosis disease in Litopenaeus vannamei J. World Aquac. Soc., 50, 1002–1015.
50. Overman, T.L., Kessler, J.F. y Seabolt, J.P. (1985) Comparison of API 20E, API rapid E, and API rapid NFT for identification of members of the family Vibrionaceae. J. Clin. Microbiol., 22, 778–781.
51. Croci, L., Suffredini, E., Cozzi, L., Toti, L., Ottaviani, D., Pruzzo, C., Serratore, P., Fischetti, R. y Goffredo, E. (2007) Vibrio parahaemolyticus Working Group. Comparison of different biochemical and molecular methods for the identification of Vibrio parahaemolyticus J. Appl. Microbiol 102, 229–237.
Tabla 3. (a). Caracterización de bacterias aisladas de zoeas afectadas y saludables. (b). Caracterización de bacterias aisladas de zoeas afectadas y saludables mediante API 20E.
Las cepas identificadas como zoeas LBS fueron aisladas de zoeas afectadas por LBS, y las identificadas como saludables de zoeas aparentemente saludables o no afectadas. 1 Enfermedad Aguda de Necrosis Hepatopancreática (AHPND) [42]; 2 Gen toxR [44]; 3 Vibrio spp. [46]; 4 Fragmento del gen 16S rRNA específico de Vibrio [34]; 5 Gen de proteína altamente virulenta de Vibrio [36]; 6 V. parahaemolyticus [35]; 7 V. harveyi (vhh) [37]. - como negativo + como positivo. a Cepas positivas/total de cepas.
Ausencia de células desprendidas: no se observaron materiales celulares desprendidos en el lumen del sistema digestivo en desarrollo de las larvas saludables. La ausencia de células desprendidas indica una falta de degeneración tisular o patología dentro del tracto digestivo (Figura 2c,d).
Desprendimiento de células epiteliales y necrosis: la mayoría de las zoeas afectadas por LBS mostraron desprendimiento de células epiteliales dentro del lumen y pérdida estructural debido a necrosis en el hepatopáncreas (Figura 3a–c). Este desprendimiento varió de mínimo a severo y contrastó de manera pronunciada con los epitelios intactos en las larvas saludables (Figura 3d–f).
Epitelio del intestino medio: en las larvas afectadas por LBS, se observó un notable desprendimiento de células epiteliales del intestino medio hacia el lumen. Esta característica patológica estaba ausente en las larvas saludables, donde el epitelio del intestino medio permanecía intacto.
Membrana peritrófica: se observaron células dentro de la membrana peritrófica en
Figura 2. Secciones teñidas con hematoxilina y eosina de larvas normales y afectadas de zoea 2. (a) Larva de zoea 2 normal. (b) Vista más cercana de la imagen a, mostrando la formación temprana del hepatopáncreas (flecha gruesa). Observe el contenido alimenticio del sistema digestivo y la ausencia de restos de tejido. (c) Lóbulos que formarán el hepatopáncreas (flecha delgada). Note la membrana peritrofica intacta y el contenido alimenticio (cabeza de flecha). (d) Vista más cercana del tracto digestivo con partículas marrones de posibles microalgas (flecha gruesa).
las larvas afectadas por LBS (Figura 3g). Esto indica una disrupción en el papel protector de la membrana peritrófica.
Bacterias Gram-negativas: la tinción de Twort reveló la presencia de bacterias Gramnegativas, con o sin material desprendido, en las larvas afectadas (Figura 4a,b). Este hallazgo fue consistente con los análisis microbiológicos que mostraron concentraciones más altas de Vibrio spp. en las zoeas enfermas.
Diferencias intestinales: se observaron diferencias significativas en el área intestinal entre las larvas normales y las afectadas por LBS. En las larvas normales, los intestinos parecían saludables, sin sobrecrecimiento bacteriano y con membranas peritróficas intactas (Figura 2c). En contraste, las larvas afectadas por LBS presentaban una abundancia de bacterias Gram-negativas y membranas peritróficas comprometidas (Figura 4d).
Figura 3. Secciones teñidas con hematoxilina y eosina de larvas normales y afectadas de zoea 2. (a) Hepatopáncreas de larvas de zoea 2–3 afectadas. Observe el desprendimiento de células dentro del lumen (flecha delgada). (b) Pérdida estructural debido a necrosis en el hepatopáncreas (flecha delgada). (c) Área del hepatopáncreas y del intestino, presencia de material descamado (flecha delgada). (d) Descamación de células epiteliales del intestino medio en el lumen, observe el exterior de la membrana peritrofica (flecha gruesa). (e,f) Área del intestino con material descamado severo (flecha delgada), incluyendo la membrana basal (cabeza de flecha). (g) Heces con material celular descamado (flecha gruesa).
52. Fabbro, C., Cataletto, B. y Del Negro, P. (2010) Detection of pathogenic Vibrio parahaemolyticus through biochemical and molecular-based methodologies in coastal waters of the Gulf of Trieste (North Adriatic Sea). FEMS Microbiol. Lett 307, 158–164.
53. Lydon, K.A., Kinsey, T., Le, C., Gulig, P.A. y Jones, J.L. (2021) Biochemical and virulence characterization of Vibrio vulnificus isolates from clinical and environmental sources. Front. Cell Infect. Microbiol., 11, 637019.
54. Vivijs, B., Moons, P., Aertsen, A. y Michiels, C.W. (2014) Acetoin synthesis acquisition favors Escherichia coli growth at low pH. App. Environ. Microb., 80, 6054–6061.
55. Oh, Y.T., Kim, H.Y., Kim, E.J., Go, J., Hwang, W., Kim, H.R., Kim, D.W. y Yoon, S.S. (2016) Selective and efficient elimination of Vibrio cholerae with a chemical modulator that targets glucose metabolism. Front. Cell. Infect. Microbiol., 6, 156.
56. Muhammad, F., Zhang, Z.F., Shao, M.Y., Dong, Y.P. y Muhammad, S. (2012) Ontogenesis of digestive system in Litopenaeus vannamei (Boone, 1931) (Crustacea: Decapoda). Ital. J. Zool., 79, 77–85.
Sun, J.,Wang, X.Y., Yang, Q., Xie, G.S., Dong, X. y Huang, J. (2019) Case studies: Pathogenic agent and microbiome analysis of zoea of Litopenaeus vannamei suffering from an unknown disease. Progress. Fish. Sci., 40, 134–144. (In Chinese).
58. Wolkenfelt, B. y Blonk, R. (2021) Balanced shrimp breeding in Ecuador, for now and the future. In: Proceedings of the Aqua Expo 2021, World Aquaculture Conference and Trade Show, Cámara Nacional de Acuacultura, Guayaquil, Ecuador, 25–28 October 2021
59. Hameed, A.S.S. (1993) A study of the aerobic heterotrophic bacterial flora of hatchery-reared eggs, larvae and postlarvae of Penaeus indicus Aquaculture, 117, 195–204.
60. Garcia, A.T. y Olmos, J.S. (2007) Quantification by fluorescent in situ hybridization of bacteria associated with Litopenaeus vannamei larvae in Mexican shrimp hatchery. Aquaculture, 262, 211–218.
61. Intriago, P., Espinoza, J., Medina, A., Altamirano, L., Enriquez, X., Sanchez, A., Navarrete, A., Alvarez, E. y Forestieri, J. (2018) Microbiological dynamics of a Litopenaeus vannamei hatchery in Ecuador. In: Proceedings of the Aqua Expo 2018, World Aquaculture Conference and Trade Show, Cámara Nacional de Acuacultura, Guayaquil, Ecuador, 15–18 October 2018.
62. Kurmaly, K., Jones, D.A., Yule, A.B. y East, J. (1989) Comparative analysis of the growth and survival of Penaeus monodon (Fabricius) larvae, from protozoea 1 to postlarva 1, on live feeds, artificial diets and a combination of both. Aquaculture, 81, 27–45.
63. Soto-Rodriguez, S.A., Simoes, N., Jones, D.A., Roque, A. y Gómez-Gil, B. (2003) Assessment of fluorescent labeled bacteria (FLB) for evaluation of in vivo uptake of bacteria (Vibrio spp.) by crustacean larvae. J. Microbiol. Methods, 52, 101–114.
64. Aguirre-Guzman, G., Vazquez-Juarez, R. y Ascencio, F. (2001) Differences in the susceptibility of American white shrimp larval substages (Litopenaeus vannamei) to four Vibrio species. J. Invert. Pathol., 78, 215–219.
65. Intriago, P. y Jimenez, R. (1997) The effect of bacteria isolated from an outbreak of haemocytic enteritis in pond reared shrimp in Litopenaeus vannamei (Boone) larvae in Ecuador. Unpublished Internal Report Aqualab 1997, Ayangue-Peninsula de Santa Elena, Ecuador, 1997.
66. Carella, F. y Sirri, R. (2017) Editorial: Fish and shellfish pathology. Front. Mar. Sci., 4, 375.
67. Reyes, G., Andrade, B., Betancourt, I., Panchana, F., Solorzano, R., Preciado, C., Sorroza, L., Trujillo, L.E. y Bayot, B. (2023) Microbial profiles of Penaeus vannamei larvae in low survival hatchery tanks affected by vibriosis. Peer J., 11, e15795.
Ausencia de bolitas: durante este estudio, no se observaron bolitas (esferas) en ninguna de las diapositivas histológicas, y dichas estructuras no están documentadas en la literatura [1,2]. Se sugiere que las bolitas podrían ser de naturaleza lipídica, dado que la preparación de muestras de tejido para histología implica el procesamiento a través de varios solventes que podrían potencialmente disolver estructuras basadas en lípidos.
Discusión
El presente estudio describe las lesiones histopatológicas, los resultados de PCR y los análisis microbiológicos de muestras de zoea de P. vannamei recolectadas en dos criaderos diferentes: uno con un brote de LBS y otro con animales aparentemente sanos. De todos los patógenos analizados por PCR, las únicas diferencias clave entre las zoeas afectadas por LBS y las saludables fueron la detección de Vibrio y una mayor abundancia de bacterias similares a Rickettsia (RLB) en las zoeas enfermas. La presencia del virus WzSV8 en las zoeas saludables requiere más estudio; este virus de ARN se ha encontrado en una amplia gama de ambientes y regiones, incluyendo la población silvestre de reproductores, y su efecto en la producción de peneidos necesita ser objeto de estudios adicionales [48].
Los análisis microbiológicos mostraron que el número total de bacterias cultivables (TSA) era un orden de magnitud mayor en las zoeas afectadas, y el número de Vibrio presuntivos era casi dos órdenes de magnitud mayor en comparación con las zoeas saludables. Los Vibrio presuntivos representaron el 17% y el 6% de la población bacteriana total en las zoeas enfermas y saludables, respectivamente. Las colonias verdes (en TCBS) y las colonias color malva, que son presuntivamente V. parahaemolyticus (en CHROMagar™ Vibrio), representaron el 0.2% y el 82% del recuento total de Vibrio en las zoeas afectadas, mientras que se observó un patrón inverso en las zoeas saludables, con el 56% y el 2%, respectivamente (Tabla 2). Este hallazgo es interesante porque la suposición común es que las colonias verdes en TCBS y las colonias moradas en CHROMagar™ Vibrio típicamente representan V. parahaemolyticus. En este sentido, Soto-Rodríguez et al. [49], al
4. Secciones teñidas con Gram de Twort a través de larvas normales y afectadas de zoea 2. (a) Hepatopáncreas de zoea afectada con material descamado (flecha gruesa) y células bacterianas en el lumen (flecha delgada), color que indica bacterias Gram-negativas. (b) Células bacterianas normales en el lumen (flecha gruesa). (c) Zoea afectada, área del intestino con una presencia abundante de bacterias (flecha delgada). (d) Membrana peritrofica intacta de zoea normal. Note la ausencia de bacterias.
estudiar las características fenotípicas y la cinética de crecimiento de tres cepas de V. parahaemolyticus con diferentes virulencias y una cepa no patógena, encontraron que, independientemente de la virulencia de la cepa, había una alta diversidad metabólica, lo que resultaba en fenotipos de diferentes colores en el CHROMagar™ Vibrio
Es importante destacar que todas las 14 cepas fueron identificadas por PCR como miembros del género Vibrio, por lo que se puede concluir que los resultados del API 20E (21% de identificación errónea a nivel de género) no fueron confiables y deben tomarse con cautela (Tabla 3a). Aunque Overman et al. [50] afirmaron que el API 20E es un sistema válido para la identificación de los miembros más comunes de la familia Vibrionaceae, se ha reportado que el sistema puede resultar en falsos negativos [51,52]. La identificación por API se basa en perfiles bioquímicos, pero se ha encontrado que los perfiles bioquímicos y el genotipo no están necesariamente asociados con el potencial de virulencia [53]. La relación entre las variaciones o diferencias en las identificaciones del API 20E y la posible virulencia y variación de género de cada
cepa es algo que requiere una exploración más profunda.
La Tabla 3b proporciona detalles sobre la caracterización de las bacterias aisladas utilizando el sistema API 20E. Esta tabla está dividida en dos grupos: bacterias aisladas en zoea de LBS (V. alginolyticus y Vibrio spp., con seis y cinco cepas, respectivamente) y bacterias (tres cepas) aisladas de zoea saludables. La fermentación/oxidación de arabinosa y amigdalina fue diferente entre los tres grupos de Vibrio aislados de zoea afectada versus saludable (Tabla 3b). La producción de acetoína fue la principal diferencia entre V. alginolyticus y otras especies de Vibrio aisladas de zoea saludables y afectadas. La capacidad de producción de acetoína por Vibrio spp. aislados de animales afectados por LBS es interesante, ya que generalmente este metabolito se produce cuando los microorganismos utilizan la vía de 2,3-butanodiol para fermentar azúcares, y esta vía genera productos finales menos ácidos y más neutros, como acetoína y 2,3-butanodiol. Dado que la acetoína es un producto de fermentación neutro y esta reacción biosintética consume protones intracelulares, el crecimiento bacteriano
Figura
puede ocurrir en una fuente de carbono de glucosa sin disminución del pH [54,55]. Además, se ha sugerido que la inhibición de la producción de acetoína podría ser un mecanismo potencial para controlar V. cholerae, que se sabe es sensible al ácido [55]. La diferenciación de cepas de Vibrio por sus propiedades bioquímicas no es rara. Por ejemplo, V. anguillarum podría separarse principalmente en función de su reacción en la producción de indol y la fermentación de amigdalina y arabinosa [56].
Una comparación entre la histología de este estudio y el primer informe de LBS descrito por Morales [1] es imposible. Lamentablemente, solo se presentaron imágenes de microscopía electrónica de transmisión (TEM) y no se registraron secciones de tejido teñidas con H&E. La publicación de Robertson et al. [2] muestra la presencia de racimos necróticos melanizados en los túbulos hepatopancreáticos (Figura 2 en [2]). Esta característica nunca se observó durante la presente investigación. La razón más probable es que los animales utilizados para la histología no eran zoea, sino que estaban al menos en la etapa PL1 al comparar la anatomía de los que se presentaron con otras descripciones detalladas sobre el desarrollo larval de P. vannamei [57]. Las diferencias importantes entre la patología presentada aquí para LBS y para PL-AHPND [10] es que no hay una descamación masiva de células en el hepatopáncreas como se describió para PL-AHPND. Además, PLAHPND nunca se encontró ni se describió en zoea [10]. Además, no se detectaron PirAB ni otros genes patogénicos. Un material similar presente en larvas de camarón criadas en China y que sufren mortalidades durante la etapa zoea 2 ha sido descrito por otros autores; en su caso, esto se asoció con una cepa de V. alginolyticus [58].
Históricamente, el sector camaronero latinoamericano ha seguido el modelo ecuatoriano, donde las postlarvas se producen a partir de reproductores obtenidos de estanques de producción. Los reproductores se seleccionan en función de su peso y luego se trasladan a una incubadora para producir la siguiente generación de postlarvas. Este proceso ofrece poco o ningún control a largo plazo sobre la bioseguridad [59]. Además, altas concentraciones de Vibrio spp. pueden
ser muy comunes en incubadoras de peneidos no bioseguras o aquellas que no son libres de patógenos específicos, tanto libres como adheridos a las larvas. Desde la incubación hasta la cosecha, el entorno microbiológico es una sopa de bacterias y partículas similares a virus (VLPs) [60–62]. A medida que las larvas transitan de una dieta basada en algas como zoea a animales que obtienen proteínas como misis, las larvas sufren un cambio dramático en el volumen del hepatopáncreas y la bioquímica de las enzimas digestivas [63]. En zoea, la filtración de partículas es casi indiscriminada. A medida que las etapas de zoea se exponen a concentraciones muy altas de bacterias, tanto libres como adheridas, incluyendo Vibrio que son fácilmente ingeridos y capaces de pasar al tracto digestivo [64], se pueden observar los efectos perjudiciales de estos dependiendo de varios factores como el subestado larval involucrado, las especies de Vibrio presentes y su concentración [65].
Intriago y Jiménez [66] replicaron el Síndrome de las Bolitas en zoea de P. vannamei utilizando una cepa luminiscente de V. harveyi a concentraciones tan bajas como 10³ células/mL. Curiosamente, esta cepa fue aislada de P. vannamei enfermo cultivado en granja, afectado por enteritis hemocítica. Postularon que los patógenos podrían estar rebotando de las incubadoras hacia los reproductores o viceversa, y que las diferencias encontradas en la histopatología entre las larvas y los adultos podrían atribuirse, por un lado, a las diferencias en el grado de desarrollo de los órganos y, por otro, a las especies de patógenos, su virulencia y su concentración.
El evento clave en la aparición de enfermedades podría atribuirse al estrés (temperatura, salinidad, densidad, toxinas, etc.) debido a alteraciones en el entorno, lo que ejerce un cambio en la interacción huéspedpatógeno y la transmisión de bacterias entre especies. Tales modificaciones actúan sobre los patógenos para facilitar su mayor transmisión entre los huéspedes individuales y su mayor contacto con nuevas poblaciones o especies de huéspedes, así como sobre la presión de selección que lleva a la dominancia de cepas patógenas adaptadas a estas nuevas condiciones ambientales [46,67]. Desafortunadamente, no hay forma de comparar la histología de
este estudio con la de informes publicados anteriormente, aunque la histología de este estudio se asemeja al síndrome de zoea 2 reportado por Kumar et al. [14]. Sin embargo, las diferencias en la patología podrían ser el resultado de diferentes respuestas del huésped a una amplia dinámica de genotipos bacterianos y la concentración de patógenos bacterianos [12]. Además, dos patógenos también podrían producir la misma patología macroscópica (LBS), pero las diferencias en el daño a nivel tisular dependerán de todos los factores descritos anteriormente.
Conclusiones
A partir de la información anterior, queda claro que no se encontró ningún virus involucrado en esta enfermedad. Aunque algunos fueron detectados por PCR, no se encontraron cuerpos de inclusión ni lesiones asociadas. También podemos descartar patologías comunes de Vibrio como AHPND o la nueva y altamente letal V. parahaemolyticus descrita para larvas de P. vannamei que llevan los genes Vhp1y Vhp-2. Las 14 cepas bacterianas aisladas fueron identificadas como Vibrio por PCR. Es tentador sugerir que Vibrio jugó un papel en la patogenicidad; sin embargo, no podemos descartar la posibilidad de RLB o toxicidad del agua. Sin embargo, podemos concluir que las especies de Vibrio, ya sea como bacterias patógenas u oportunistas, juegan un papel importante tanto en los tanques afectados por LBS como en los saludables, y en general se caracterizaron por una alta diversidad metabólica, lo que dio lugar a fenotipos de diferentes colores en el CHROMagar™ Vibrio El examen histopatológico de las larvas de zoea etapas 2–3 afectadas por LBS reveló cambios patológicos significativos, incluidos el desprendimiento de células epiteliales, necrosis hepatopancreática, membranas peritróficas alteradas y la presencia de bacterias Gram-negativas. Estos hallazgos destacan el grave impacto de LBS en el sistema digestivo de las larvas de P. vannamei, contrastando marcadamente con las estructuras intactas y la ausencia de características patológicas en las larvas saludables. Se necesita más investigación para comprender completamente la etiología del Síndrome de las Bolitas y desarrollar intervenciones efectivas para mitigar sus efectos en los criaderos de camarones•
Impacto de diversas condiciones de procesamiento en el nivel de sodio y en los atributos sensoriales del camarón
Autores:
Kaylan Hayman, Prayon, Augusta Georgia 30906 EE.UU., KHayman@prayon.com
Caroline Mees, Prayon, Engis Bélgica, CMees@prayon.com
LProtocolo experimental realizado por el Dr. Paul Sarnosky del Departamento Marino de la Universidad de Florida, Gainesville, Florida 32611 EE. UU.
KHayman@prayon.com
os consumidores de todo el mundo exigen opciones cada vez más saludables en el sector de mariscos, especialmente en el camarón procesado. Entre estas demandas, los productos con menor contenido de sodio son especialmente atractivos, ya que el consumo excesivo de este elemento es un problema importante para la salud humana y está relacionado con muchas enfermedades no transmisibles, como la hipertensión, la obesidad y algunos tipos de cáncer (Jachimowicz-Rogowska y Winiarska-Mieczan, 2023). Muchos consumidores ingieren más del doble de la cantidad máxima de sodio recomendada por la Organización Mundial de la Salud (Rybicka et al., 2022), por lo que entidades como la OMS y la Administración de Alimentos y Medicamentos de los Estados Unidos han creado pautas para ayudar a las procesadoras a alcanzar los objetivos de reducción de este componente.
El camarón es una excelente fuente de proteína económica, bajo en calorías y grasas, y es fácilmente accesible en muchas partes del mundo, lo que lo convierte en un elemento importante de muchas dietas. Sin embargo, durante el procesamiento, el contenido de sodio del camarón puede aumentar hasta cien veces (Rybicka et al., 2022), debido a que aditivos utilizados en su manipulación, como el cloruro de sodio y el fosfato de sodio, desempeñan un papel importante en la mejora de la retención de humedad, la apariencia y el sabor del crustáceo. Por lo tanto, para muchas procesadoras, reducir el sodio significa que también se reduce el rendimiento, el sabor y la calidad general del producto (Pedro y Nunes, 2019).
El objetivo principal de este estudio fue medir la absorción de sodio en camarones tratados con cuatro mezclas de fosfato, incluyendo una mezcla baja en sodio, en comparación con una mezcla de fosfato de referencia utilizada en el mercado ecuatoriano. El segundo objetivo del estudio fue comparar la absorción de humedad, la pérdida de cocción y las características sensoriales de los camarones tratados, para determinar si la menor absorción de sodio resultó en una reducción del rendimiento del producto y el atractivo para el consumidor.
Diseño experimental Camarones
Se compró camarón blanco fresco producido (Litopenaeus vannamei a un proveedor local en Florida, EE.UU. Los camarones eran de tamaño grande (26-30) y estaban descabezados, pelados y desvenados.
Mezclas de tratamiento
Se evaluaron cuatro mezclas de fosfato y una mezcla de referencia de mercado:
Tabla 1: Mezclas de fosfato evaluadas en este estudio
Producto Aplicación objetivo
Mezcla 1 (CSP) Tiempo de tratamiento corto
Mezcla 2 (C950i) Formulaciones bajas en sodio
Mezcla 3 (B940) Mejora de calidad global Mezcla 4 (B980) Tiempo de tratamiento largo Mezcla 5 (RefP) Referencia del mercado
Cada mezcla se utilizó para preparar un tratamiento salmuera compuesto por 2% de NaCl, 3% de mezcla de fosfato y 10% de hielo en peso en agua destilada. Se agitó constante para disolver completamente los solutos antes de agregar el hielo. La temperatura de la salmuera se mantuvo a 4 °C.
Absorción de humedad
Los camarones se secaron y se pesaron, luego se agregaron a cada tratamiento de salmuera en una proporción de 1:2 (es decir, 150 g de camarones: 300 g de salmuera). Luego, las salmueras con camarones se transfirieron a un enfriador portátil ajustado a 4 °C durante 2 horas con agitación constante a 165 rpm. Después de la exposición a la salmuera, los camarones se retiraron de la solución, se secaron suavemente con papel absorbente para retirar el exceso de agua y se pesaron.
Pérdida de cocción por ebullición
Después de la exposición a la salmuera, una parte de los camarones tratados se hirvió a una temperatura interna de 63 °C durante aproximadamente 15 a 30 segundos (Torti et al. 2016). Los camarones se pesaron después de un breve período de enfriamiento.
Absorción de sodio
Las muestras de camarón tratado fueron pasadas por un sistema de digestión de microondas, de acuerdo con el método
AOAC 2011.14. El contenido de sodio de los camarones se determinó de acuerdo
con el método AOAC 984.27, que utiliza espectroscopia de emisión con plasma acoplado inductivamente (ICP) para medir el sodio elemental en una muestra dada. Se analizó una muestra compuesta homogénea de 25 g de varios camarones por cada uno de los tratamientos de salmuera.
Análisis sensorial
Los camarones se trataron y cocinaron el día anterior al panel. Los camarones cocidos se colocaron en hielo antes de distribuirlos a los panelistas. El panel sensorial estuvo compuesto por 75 panelistas no entrenados. A cada panelista se le dio una muestra de cada uno de los tratamientos, cada uno con 2 camarones, así como una muestra control de camarones que se remojaron solo en agua destilada. Se pidió a los panelistas que calificaran la apariencia, la textura, el sabor y el gusto general de los camarones utilizando una escala hedónica de 1 a 9. En la escala hedónica, una calificación de 1 indica “desagrado extremo” del atributo dado y una calificación de 9 indica “agrado extremo”. Después de evaluar la apariencia, se pidió a los panelistas que probaran cada camarón y calificaran la textura, el sabor y el gusto general. El orden de las muestras de camarón presentadas a los panelistas se asignó al azar utilizando un diseño Williams para evitar el sesgo de orden.
Resultados y discusión
Se utilizaron cuatro mezclas de fosfato y
una mezcla de referencia del mercado para tratar los camarones, en las que luego fueron evaluadas para saber sobre la absorción de sodio, el gusto por la apariencia, el sabor y la textura.
La absorción de sodio se muestra en la Figura 1, mientras que la absorción de humedad y la pérdida por cocción se muestran en la Figura 2.
Los camarones tratados con B940 lograron una absorción de humedad de 9.92 ± 0.18% después de 2 horas de tratamiento, que es similar a la absorción observada en los camarones tratados con la mezcla de referencia del mercado (9.84 ± 0.55%). Ambos tratamientos compartieron una mayor absorción de humedad que los camarones sin tratar, que solo tuvieron un aumento de 6.74 ± 0.74%. Los camarones cocidos tratados con B940 experimentaron una pérdida de 15.39 ± 5.19% de humedad, mientras que los tratados con la referencia del mercado perdieron 8.24 ± 0.59%; nuevamente, ambos tratamientos tuvieron un mejor desempeño que el camarón control que experimentó una pérdida de 29.51 ± 0.43%.
Naturalmente, el contenido de sodio del camarón control fue el más bajo a 135 ppm. El tratamiento con B940 resultó en un menor aumento en el contenido de sodio a 434 ppm en comparación con la referencia
Contenido de sodio del camarón después de 2 horas de tratamiento
Figura 1: contenido de sodio del camarón después de 2 horas de tratamiento en salmuera preparada con CSP, B940, C950i, B980 o RefP
del mercado, que aumentó el sodio en 461 ppm. Tanto el B940 como la mezcla de referencia del mercado están diseñados para impartir una mejora de la calidad global en el camarón y sus composiciones son relativamente similares, lo que explica el rendimiento similar observado en este estudio.
Comparativamente, los camarones tratados con B980 lograron una mayor absorción de humedad en 11.11±0.14%, con una pérdida de humedad de 13.14 ± 0.04% después de la cocción. Como esta mezcla está diseñada específicamente para tiempos de tratamiento prolongados, normalmente más de dos horas, la absorción de humedad probablemente seguiría aumentando a medida que aumenta el tiempo de tratamiento. El B980 proporcionó menos sodio al camarón que la mezcla de referencia del mercado, con un aumento de 444 ppm.
El CSP confirió una absorción de humedad de 10.19 ± 0.52% a los camarones tratados, con una pérdida de cocción de 14.96 ± 1.72%, y proporcionó la mayor absorción de sodio entre todas las mezclas con un aumento de 519 ppm. Curiosamente, esta mezcla tuvo un rendimiento similar al de otras en términos de ganancia y pérdida de humedad, a pesar de estar diseñada para usarse en tiempos de tratamiento mucho más cortos, generalmente de 30 minutos a una hora.
La mezcla baja en sodio, C950i, logró la mayor absorción de humedad entre todas las mezclas, con una absorción de 11.61±1.36% en los camarones tratados. La pérdida debido a la cocción fue comparativamente mediana, con 14.80±2.88%. Como se esperaba, C950i proporcionó la menor absorción de sodio en los camarones tratados en comparación con todas las demás mezclas, con un aumento de solo 311 ppm, que es un 33% menos que la mezcla de referencia del mercado.
La evaluación sensorial se presenta en las 3 figuras a continuación: El CSP, C950i, B940 y B980 obtuvieron puntajes tan buenos o mejores que la mezcla de referencia del mercado en todos los atributos sensoriales de los camarones tratados, incluyendo la apariencia, el sabor, la textura y el gusto general. Cabe destacar
Absorción de humedad y pérdida por cocción en el camarón después de 2 horas de tratamiento
Figura 2: porcentaje de absorción de humedad en el camarón después de 2 horas de tratamiento en salmuera preparada con CSP, B940, C950i, B980 o RefP
general de la apariencia
Figura 3: gusto general de la apariencia de los camarones tratados con CSP, C950i, B940, B980 o RefP. Las calificaciones que comparten la misma letra de sufijo (A o B) no difieren significativamente (HSD de Turkey α = 0.05)
Gusto general por el sabor
Figura 4: gusto general del sabor de los camarones tratados con CSP, C950i, B940, B980 o RefP. Las calificaciones que comparten la misma letra de sufijo (A o B) no difieren significativamente (HSD de Turkey α = 0.05)
Absorción de humedad Pérdida por cocción
Gusto
Gusto general de la textura
Figura 5: gusto general por la textura de los camarones tratados con CSP, C950i, B940, B980 o RefP. Las calificaciones que comparten la misma letra de sufijo (A o B) no difieren significativamente (HSD de Turkey α = 0.05).
que las calificaciones de todos los atributos en los camarones tratados con C950i no fueron significativamente diferentes a las de cualquier otra mezcla, a pesar de proporcionar alrededor de un tercio menos de sodio que todas las demás mezclas, lo que indica que esta mezcla permitió la reducción de sodio sin afectar negativamente la calidad sensorial de los camarones tratados.
Conclusiones
Este estudio demostró que es posible reducir el aumento de sodio en camarones tratados en más de un tercio en comparación con la mezcla de referencia del mercado que se usa comúnmente en el mercado de Ecuador, y que esta reducción no tiene un impacto significativo en el sabor, la textura o la apariencia del camarón. Esto indica que es posible que las procesadoras de camarón mantengan la alta calidad de sus productos y, al mismo tiempo, satisfagan la demanda de opciones más saludables para los consumidores en Ecuador y de todo el mundo•
Referencias
Jachimowicz-Rogowska, K., & Winiarska-Mieczan, A. (2023). Initiatives to reduce the content of sodium in food products and meals and improve the population’s health. Nutrients, 15(10), 2393. https://doi.org/10.3390/nu15102393
Luzuriaga, D. A., Balaban, M. O., & Yeralan, S. (1997). Analysis of Visual Quality Attributes of White Shrimp by Machine Vision. Journal of Food Science, 62(1), 113–118.
Official methods of analysis of the association of official analytical chemists (AOAC). 1990.
Official Method 967.13 1:15–69.
Pedro, S., & Nunes, M. L. (2019). Reducing salt levels in seafood products. Reducing Salt in Foods, 185–211. https://doi.org/10.1016/b978-0-08-100890-4.00008-1
Rybicka, I., Gonçalves, A., Oliveira, H., Marques, A., & Nunes, M. L. (2022). Salt reduction in seafood – a review. Food Control, 135. https://doi.org/10.1016/j. foodcont.2022.108809
Torti, M. J., Sims, C. A., Adams, C. M., & Sarnoski, P. J. (2016). Polysaccharides as Alternative Moisture Retention Agents for Shrimp. Journal of Food Science, 81(3), S728–S735.
Para más información sobre este artículo, contactar a: KHayman@prayon.com
vannamei
Autores:
Gladys Torres
Carlos Mora-Pinagorte1
Manuel Espinoza-Ortega1
Jaritza Sabando
César Molina-Poveda*1
Skretting LATAM
1Skretting Aquaculture Innovation
*cesar.molina@skretting.com
En las áreas costeras alrededor del mundo, ninguna otra actividad económica ha crecido tanto en los últimos años como la industria camaronera, convirtiéndose en una fuente crucial de proteínas y empleo para muchas comunidades (Páez-Osuna, 2001). Sin embargo, este desarrollo no está exento de desafíos, especialmente en lo que respecta a enfermedades emergentes que amenazan la salud del camarón y la productividad de la industria camaronera. Una de estas amenazas es la proliferación de microalgas, que genera preocupación en la acuicultura global debido al riesgo potencial sobre la vida marina y la economía de las costas (Borbor-Córdova et al., 2018; Lily, 2024; Paerl y Huisman, 2008). Dentro de los grupos de microalgas perjudiciales, el género Chattonella ha ganado atención dada su habilidad para producir potentes toxinas que pueden afectar a los organismos marinos (Khan et al., 1996).
Los afloramientos de Chattonella pueden verse estimulados por una combinación de varios factores, incluyendo un enriquecimiento de nutrientes en el agua, producto de una “eutrofización cultural” a partir de los desechos domésticos, industriales y agrícolas que la contaminan (Hallegraeff & Hara, 2003). Estos afloramientos pueden proliferar rápidamente y varios estudios los han asociado con altas mortalidades de la fauna marina, causando una gran pérdida económica (Barraza-Guardado et al., 2004; CortésAltamirano et al., 2006; Pérez-Morales et al., 2017). Las mortalidades se encuentran asociadas con la habilidad de la microalga de producir diferentes compuestos tóxicos, principalmente especies reactivas de oxígeno (ROS, por sus siglas en inglés) (Band-Schmidt et al., 2012; Oda et al., 1998; Shimada et al., 1993; Tanaka et al., 1994), que causan daño celular en las branquias y dificultan la respiración del camarón afectado (PérezMorales et al., 2017).
En Ecuador, los productores de camarón se enfrentan a varios retos con el afloramiento de Chattonella. Recientemente, se ha logrado detectar la presencia de quistes del alga en aguas de piscinas camaroneras que supondrían un riesgo para la producción. Este artículo examina el impacto de
Chattonella en la producción de camarones, sus mecanismos de toxicidad y las estrategias potenciales para mitigar sus efectos adversos.
Biología, ecología y caracterización de Chattonella Chattonella es un género de microalgas dinoflageladas de aguas marinas y estuarinas pertenecientes a la clase Raphidophyceae (Hallegraeff y Hara, 2003), conocidas por ser un agente altamente nocivo de las mareas rojas que causan grandes daños a las granjas acuícolas costeras y estuarinas (Imai y Itoh, 1988; Imai y Yamaguchi, 2012). Estas microalgas son unicelulares, de color marrón-dorado (contienen fucoxantina), flagelados desnudos que poseen un flagelo anterior dirigido hacia adelante con pelos tubulares (mastigonemas tubulares tripartitos) y un flagelo posterior liso que parte de un sirco flagelar más o menos pronunciado (Hallegraeff y Hara, 2003). El género Chattonella está compuesto por cinco especies: Chattonella subsalsa (Biecheler, 1936), Chattonella antiqua (Hada) (Ono, 1980), Chattonella marina (Hara y Chihara, 1982), C. minima (Hara y Chihara, 1994) y C. ovata (Hara y Chihara, 1994) (Imai y Yamaguchi, 2012). Debido a su falta de pared celular, estas especies tienen la capacidad de cambiar de tamaño y forma, haciendo poco exactas sus identificaciones por microscopia. Demura et al. (2009), mediante la aplicación de métodos moleculares, propone una revisión taxonómica que considere características morfológicas y genéticas (genes ribosomales, mitocondriales y regiones microsatélites), introduciendo a C. ovata y C. antiqua, como variaciones de C. marina
El ciclo de vida de Chattonella comprende dos fases: una fase móvil planctónica y una fase estacionaria en forma de quiste (Imai y Itoh, 1988). Bajo condiciones desfavorables, Chattonella entra en la fase de quistes que se sedimentan en el fondo del cuerpo de agua, permitiendo la supervivencia de la microalga. Una vez que las condiciones vuelven a ser óptimas, los quistes de Chattonella germinan y proliferan de manera acelerada, pudiendo ocasionar los afloramientos (Imai y Itoh, 1988; Imai y Yamaguchi, 2012). De acuerdo con observaciones en campo y estudios en laboratorio, se ha podido confirmar que
la temperatura es el principal factor que afecta la fisiología (latencia, maduración y germinación) de los quistes de Chattonella (Imai et al., 1991, 1989; Imai y Itoh, 1988, 1987; Imai y Yamaguchi, 2012; Yamaguchi et al., 2008). Esta habilidad de enquistarse le permite persistir en el ecosistema y contribuye a generar repentinos e intensos eventos de afloramientos cuando las condiciones lo permiten, lo que hace necesario llevar un control de las condiciones ambientales del cultivo de camarón para reducir las probabilidades de estos eventos.
Mecanismos de toxicidad
El mecanismo por el cual Chattonella spp. causa muerte a los organismos marinos aún no está claro; sin embargo, la asfixia suele ser la causa final de muerte (Hallegraeff y Hara, 2003; Imai y Yamaguchi, 2012). El taponamiento de las branquias por células de Chattonella y la secreción de una mucosidad fueron propuestos inicialmente como las causas de muerte en peces y camarones (Matsusato y Kobayashi, 1974). Actualmente se posee evidencia que apunta a la generación de ROS (e.g.: superóxido), que son los responsables de daños en los tejidos de las branquias y producción de la mucosidad (Ishimatsu et al., 1996; Marshall et al., 2005; Tanaka et al., 1994).
Otros compuestos relacionados con la toxicidad que han sido identificados en varias especies de Chattonella son brevetoxina (Band-Schmidt et al., 2012), óxido nítrico (Kim et al., 2008), ácidos grasos libres poliinsaturados (Band-Schmidt et al., 2012), hemaglutinina y hemolisina (de Boer et al., 2009; Dorantes-Aranda et al., 2009). Además, algunos autores han reportado la existencia de otros compuestos inestables que no han podido ser identificados, pero que podrían estar implicados en la toxicidad de estas microalgas. La presencia de estas toxinas en el ambiente acuático puede ser devastadora para las granjas de camarones, ya que los productores a menudo tienen poco tiempo para reaccionar ante un brote de marea roja.
Afloramientos de Chattonella en Ecuador
Ecuador ha reportado más de 150 eventos de marea roja entre 1968 y 2017 (BorborCórdova et al., 2019; Torres, 2017, 2015;
Bibliografía
Anderson, D.M., 2014. Prevention, Control and Mitigation of Harmful Algal Blooms: Multiple Approaches to HAB Management. Band-Schmidt, C.J., Martínez-López, A., Bustillos-Guzmán, J.J., Carreón-Palau, L., Morquecho, L., Olguín-Monroy, N.O., Zenteno-Savín, T., Mendoza-Flores, A., GonzálezAcosta, B., Hernández-Sandoval, F.H., Tomas, C., 2012. Morphology, biochemistry, and growth of raphidophyte strains from the Gulf of California. Hydrobiologia 693, 81–97. https://doi.org/10.1007/s10750-012-1088-y Barraza-Guardado, R., Cortés-Altamirano, R., SierraBeltrán, A., 2004. Marine die-offs from Chattonella marina and Ch. cf. ovata in Kun Kaak Bay, Sonora in the Gulf of California. Harmful Algae News 25, 7–8. Borbor-Córdova, M.J., Pozo-Cajas, M., CedenoMontesdeoca, A., Mantilla Saltos, G., Kislik, C., Espinoza-Celi, M.E., Lira, R., Ruiz-Barzola, O., Torres, G., 2018. Risk Perception of Coastal Communities and Authorities on Harmful Algal Blooms in Ecuador. Front Mar Sci 5. https://doi.org/10.3389/fmars.2018.00365 Borbor-Cordova, M.J., Torres, G., Mantilla-Saltos, G., Casierra-Tomala, A., Bermúdez, J.R., Renteria, W., Bayot, B., 2019. Oceanography of Harmful Algal Blooms on the Ecuadorian Coast (1997–2017): Integrating Remote Sensing and Biological Data. Front Mar Sci 6. https://doi.org/10.3389/fmars.2019.00013 Brown, A.R., Lilley, M., Shutler, J., Lowe, C., Artioli, Y., Torres, R., Berdalet, E., Tyler, C.R., 2020. Assessing risks and mitigating impacts of harmful algal blooms on mariculture and marine fisheries. Rev Aquac 12, 1663–1688. https://doi.org/10.1111/raq.12403 Cortés-Altamirano, R., Alonso-Rodríguez, R., Sierra, A., 2006. Fish mortality with Chatonella marina anf Ch. cf. ovata (Raphidophyceae) blooms in Sinaloa (Mexico). Harmful Algae News 31, 7–8. de Boer, M.K., Tyl, M.R., Fu, M., Kulk, G., Liebezeit, G., Tomas, C.R., Lenzi, A., Naar, J., Vrieling, E.G., van Rijssel, M., 2009. Haemolytic activity within the species Fibrocapsa japonica (Raphidophyceae). Harmful Algae 8, 699–705. https://doi.org/10.1016/j.hal.2009.02.001 Demura, M., Noël, M.-H., Kasai, F., Watanabe, M.M., Kawachi, M., 2009. Taxonomic revision of Chattonella antiqua, C. marina and C. ovata (Raphidophyceae) based on their morphological characteristics and genetic diversity. Phycologia 48, 518–535. https://doi.org/10.2216/08-98.1 Dorantes-Aranda, J.J., Parra, L.M.G. la, Alonso-Rodríguez, R., Morquecho, L., 2009. Hemolytic activity and fatty acids composition in the ichthyotoxic dinoflagellate Cochlodinium polykrikoides isolated from Bahía de La Paz, Gulf of California. Mar Pollut Bull 58, 1401–1405. https://doi.org/10.1016/j.marpolbul.2009.06.007 Hallegraeff, G.M., Hara, Y., 2003. Taxonomy of harmful marine raphidophytes, in: Hallegraeff, G.M., Anderson, D.M., Cembella, A.D. (Eds.), Manual on Harmful Marine Microalgae. pp. 511–522. Imai, I., Itakura, S., Itoh, K., 1991. Life cycle strategies of the red tide causing flagellates Chattonella (raphidophyceae) in the Seto Inland Sea. Mar Pollut Bull 23, 165–170. https://doi.org/10.1016/0025-326X(91)90668-I Imai, I., Itoh, K., 1988. Cyst of Chattonella antiqua and C. marina (Raphidophyceae) in sediments of the Inland Sea of Japan. Bulletin of Plankton Society of Japan 35, 35. Imai, I., Itoh, K., 1987. Annual life cycle of Chattonella spp., causative flagellates of noxious red tides in the Inland Sea of Japan. Mar Biol 94, 287–292. https://doi.org/10.1007/BF00392942
Imai, I., Itoh, K., Anraku, M., 1989. Dormancy and Maturation in the Cyst of Chattonella spp. (Raphidophyceae), Red Tide Flagellates in the Inland Sea of Japan. Red Tides: Biology, Environmental Science, and Toxicology. Imai, I., Yamaguchi, M., 2012. Life cycle, physiology, ecology and red tide occurrences of the fish-killing raphidophyte Chattonella. Harmful Algae 14, 46–70. https://doi.org/10.1016/j.hal.2011.10.014 Ishimatsu, A., Oda, T., Yoshida, M., Ozaki, M., 1996. Oxygen Radicals are Probably Involved in the Mortality of Yellowtail by Chattonella marina. Fisheries science 62, 836–837. https://doi.org/10.2331/fishsci.62.836 Jiménez, R., 1989. Red Tide and Shrimp Activity in Ecuador. Khan, S., Arakawa, O., Onoue, Y., 1996. A toxicological study of the marine phytoflagellate, Chattonella
PRODUCCIÓN
Torres-Zambrano, 2000), de los cuales, alrededor de 30 incidentes cuentan con registros de mortalidades en peces, camarones y larvas de camarón (BorborCórdova et al., 2018; Pérez-Morales et al., 2017). Los más notables ocurrieron en 1985, 2003 y 2007, con picos estacionales en marzo, abril y mayo (Borbor-Córdova et al., 2018; Torres, 2015). Los lugares de mayor concurrencia han sido el golfo de Guayaquil y la península de Santa Elena (Borbor-Córdova et al., 2019; Torres, 2015; Torres-Zambrano, 2000), identificando en el primero hasta 33 especies con potencial tóxico en los que se incluye a C. subsalsa y C. marina (Torres, 2015) (Torres, 2015). Los primeros reportes de Chattonella en Ecuador tuvieron lugar en el río Guayas en enero de 1997 y 2007, coincidiendo con el inicio de la época lluviosa.
Impacto de la Chattonella en la acuicultura de camarón
Uno de los primeros estudios sobre el efecto de la marea roja sobre la acuicultura de camarón en Ecuador fue realizado por Jiménez (1989). En este análisis se concluyó que las principales causas de mortalidades en piscinas camaroneras por marea roja es el bombeo de aguas contaminadas; a esto se le añade una alta concentración de fertilizantes que incentivaban la proliferación de microalgas nocivas. La marea roja puede resultar en una mortalidad considerable durante los cultivos de camarones, postlarvas y larvas, que se ven traducidas en pérdidas económicas hacía el productor. A pesar de que no todos los eventos han sido tóxicos o han generado afectaciones en la supervivencia del camarón, existen otros efectos secundarios: por ejemplo, a través
de estudios del contenido intestinal de camarones existe evidencia que relacionan el consumo de estas microalgas con un cambio negativo en sus condiciones organolépticas, otorgándoles un olor y sabor pastoso; este efecto podría causar el rechazo de camarones en las empacadoras, afectando directamente al productor (Jiménez, 1989) (Jiménez, 1989). Otro impacto sobre los ingresos de los camaroneros está relacionado con los costos económicos asociados con las medidas de mitigación, como la implementación de sistemas de filtración de agua y el monitoreo continuo de la calidad del líquido que pueden llegar a ser prohibitivos. La incertidumbre y la imprevisibilidad de los brotes de Chattonella también pueden disuadir la inversión en la acuicultura, afectando negativamente el crecimiento de la industria.
Casos de estudio
En mayo del 2024 se realizó un muestreo de agua superficial de una piscina camaronera ubicada en el golfo de Guayaquil. Las muestras fueron tomadas en condiciones de marea roja, es decir, zonas del cuerpo de agua con franjas de tonalidades ligeramente rojizas. Estas fueron narcotizadas mediante la adición de 2 gotas de 10% de Lugol, lo que disminuye la locomoción de las células (Fig. 1), permitiendo analizarlas con mayor detalle y pudiendo corroborar la presencia de Raphidophyceae del género Chattonella.
Adicionalmente, dos muestras de la misma camaronera con presencia de marea roja fueron analizadas tres días después in vivo (sin conservantes), observando células agrupadas y en proceso de lisis (Fig. 2A) con coloraciones amarillentas; otras células se encontraron formando
duplas de color amarillo o verde (Fig. 2B), posiblemente cursando un proceso de fusión o recombinación genética, característica de células con reproducción sexual durante la formación de quistes como Chattonella (Imai y Yamaguchi, 2012). Además, durante el muestreo se identificaron células solitarias, indicando que el evento se encontraba en proceso de enquistamiento (Fig. 2C). En cuanto a las formas solitarias, algunas registraron ligeros movimientos, la mayoría de forma circular o redonda con una pared fina, que permitió la visibilidad de cloroplastos amarillentos, cafés o dorados.
Con esta experiencia registrada en la formación de quistes, se llevaron a cabo contajes celulares de las formas de quistes según la Fig. 3 registradas desde el 20 de mayo al 12 de julio de 2024 en tres piscinas de engorde (23, 24, 25), Reservorio y Estero Canal, aplicando la misma metodología para dinoflagelados en las cercanías de Posorja (Torres et al., 2024). El resultado obtenido fue que la presencia de estos quistes no llegó a formar marea roja.
Hallegraeff y Hara (2003) indican que los quistes son esféricos, con pared celular trasparente sin ninguna ornamentación en la superficie del quiste, el protoplasma es de color verde amarillento a marrón oscuro, lo que coincide con lo observado en este estudio (Fig. 2). Sin embargo, para confirmar que la especie observada es una Chattonella spp. es necesario realizar estudios moleculares de los diversos procesos del ciclo de vida de estas Raphidophyceae, ya que como concluye Demura et al. (2009), dada la capacidad de cambiar de forma y tamaño que posee este taxón, existe una
Figura 1. Imágenes tomadas a partir de un microscopio de las muestras de marea roja colectadas en mayo de 2024.
alta probabilidad de cometer errores de identificación a nivel de especie. Además, es recomendable realizar estudios palinológicos (estudio de los quistes) para observar la viabilidad de los quistes en relación con otras variables ambientales que promueven los afloramientos y las condiciones para la germinación de los quistes (Imai y Itoh, 1988), y de esta manera prevenir los efectos nocivos de la marea roja.
En cuanto a los parámetros del agua, durante el tiempo de estudio la salinidad promedio fue de 21,4 ppt y con límites profundidad Secchi de 43 cm, cercano al límite máximo de transparencia del agua. Los nitratos y fosfatos juegan un papel importante en los afloramientos algales o mareas rojas. Durante el tiempo de estudio, los promedios de estos dos nutrientes se encontraron en unas concentraciones relativamente altas: 5,72 mg/L NO3 y 0,50 PO4. Entre estos nutrientes, Yamaguchi et al. (2008) observó mediante evaluaciones de cinética de crecimiento de una especie de Chattonella que las fuentes de nitrógeno eran el nutriente limitante para el crecimiento de esta microalga, y por ende para las afloraciones. Considerando lo expuesto anteriormente, es probable que en el área de estudio hayan existidos quistes de Chattonella spp u otras Raphidophyceae que hayan permanecido en latencia hasta que las condiciones del ambiente; en este caso, un nivel de nitrógeno disponible alto puede que haya incentivado la germinación y el crecimiento de la microalga.
Estrategias de mitigación y manejo
Para la minimización del impacto de afloramientos de Chattonella, la mitigación y el manejo juegan un control crucial. Para esto es necesario un enfoque integral que combine la vigilancia ambiental, las mejoras en la gestión de las granjas camaroneras y la investigación científica. Aunque no existe una solución única, varias medidas se han logrado implementar con cierto grado de éxito:
Monitoreo y detección temprana
Un monitoreo constante de las condiciones ambientales en las piscinas camaroneras relacionadas con las afloraciones de Chattonella y otras microalgas tóxicas puede marcar la eficacia de una estrategia. Parámetros claves en las afloraciones, como la temperatura, concentración de nutrientes, oxígeno disuelto y la turbidez del agua, son indicadores claves que permitirían tomar medidas preventivas antes de que los afloramientos alcancen niveles críticos. El uso de sensores remotos en las piscinas camaroneras han mostrado ser grandes aliados en la prevención de eventos, toma de decisiones y control de la calidad del agua de la piscina camaronera (Mirza Esmaeili et al., 2020).
Filtración y tratamiento del agua
Una de las respuestas inmediatas a la presencia de marea roja es el uso de tecnologías que permitan la filtración y tratamiento de las aguas, con el objetivo
Figura 2. Diversidad de quistes de Chattonella spp. en proceso de lisis (A), formando duplas (B) y células unitarias con cloroplastos amarillentos, cafés y dorados (C) observados en una muestra in vivo el 15 de mayo del 2024.
antiqua (Raphidophyceae). Phycologia 35, 239–244. https://doi.org/10.2216/i0031-8884-35-3-239.1 Kim, D., Kang, Y.S., Lee, Y., Yamaguchi, K., Matsuoka, K., Lee, K.-W., Choi, K.-S., Oda, T., 2008. Detection of nitric oxide (NO) in marine phytoplankters. J Biosci Bioeng 105, 414–417. https://doi.org/10.1263/jbb.105.414 Lily, 2024. Los criadores de camarones ecuatorianos se enfrentan a los retos del reciente brote de Chattonella. Roda Internacional 1–3. Marshall, J.-A., de Salas, M., Oda, T., Hallegraeff, G., 2005. Superoxide production by marine microalgae. Mar Biol 147, 533–540. https://doi.org/10.1007/s00227-005-1596-7 Matsusato, T., Kobayashi, H., 1974. Studies on death of fish caused by red tide, in: Nansei Reg. Fish. Res. Lab. pp. 43–67. Mirza Esmaeili, F., Mortazavi, M.S., Dehghan Banadaki, A.R., 2020. An overview of management and monitoring of harmful algal blooms in the northern part of the Persian Gulf and Oman Sea (Hormuzgan Province). Environ Monit Assess 192, 42. https://doi.org/10.1007/s10661-019-8002-2 Oda, T., Nakamura, A., Okamoto, T., Ishimatsu, A., Muramatsu, T., 1998. Lectin-induced enhancement of superoxide anion production by red tide phytoplankton. Mar Biol 131, 383–390. https://doi.org/10.1007/s002270050331 Paerl, H.W., Huisman, J., 2008. Blooms like it hot. Science (1979) 320, 57–58. https://doi.org/10.1126/science.1156721 Páez-Osuna, F., 2001. The Environmental Impact of Shrimp Aquaculture: Causes, Effects, and Mitigating Alternatives. Environ Manage 28, 131–140. https://doi.org/10.1007/s002670010212 Pal, M., Yesankar, P.J., Dwivedi, A., Qureshi, A., 2020. Biotic control of harmful algal blooms (HABs): A brief review. J Environ Manage 268, 110687. https://doi.org/10.1016/j.jenvman.2020.110687 Pérez-Morales, A., Band-Schmidt, C.J., Martínez-Díaz, S.F., 2017. Mortality on zoea stage of the Pacific white shrimp Litopenaeus vannamei caused by Cochlodinium polykrikoides (Dinophyceae) and Chattonella spp. (Raphidophyceae). Mar Biol 164, 57. https://doi.org/10.1007/s00227-017-3083-3 Piyatiratitivorakul, P., Lirdwitayaprasit, T., Thooithaisong, J., 2002. Laboratory Studies on Chemical Control of Red Tide Phytoplankton (Chattonella marina and Heterosigma akashiwo) for Black Tiger Shrimp (Penaeus monodon) Culture. Science Asia 9. Shimada, M., Kawamoto, S., Nakatsuka, Y., Watanabe, M., 1993. Localization of superoxide anion in the red tide alga Chattonella antiqua. Journal of Histochemistry & Cytochemistry 41, 507–511. https://doi.org/10.1177/41.4.8383714 Tanaka, K., Muto, Y., Shimada, M., 1994. Generation of superoxide anion radicals by the marine phytoplankton organism, Chattonella antiqua. J Plankton Res 16, 161–169. https://doi.org/10.1093/plankt/16.2.161 Torres, G., 2017. Evaluación del fitoplancton como un mecanismo preventivo a la ocurrencia de bloom algal frente a las costas de Esmeraldas, Manta, La Libertad y Puerto Bolívar en Ecuador 2013 -2015. Universidad Nacional Mayor de San Marcos. Torres, G., 2015. Evaluación de Mareas Rojas Durante 1968 - 2009 en Ecuador. Acta Oceanográfica del Pacífico 20. Torres-Zambrano, G., 2000. Mareas Rojas durante 1989 - 1999, en Aguas Ecuatorianas. Acta Oceanográfica del Pacífico,INOCAR 10. Yamaguchi, H., Sakamoto, S., Yamaguchi, M., 2008. Nutrition and growth kinetics in nitrogen- and phosphoruslimited cultures of the novel red tide flagellate Chattonella ovata (Raphidophyceae). Harmful Algae 7, 26–32. https://doi.org/10.1016/j.hal.2007.05.011 Yamaguchi, M., Yamaguchi, H., Nishitani, G., Sakamoto, S., Itakura, S., 2008. Morphology and germination characteristics of the cysts of Chattonella ovata (Raphidophyceae), a novel red tide flagellate in the Seto Inland Sea, Japan. Harmful Algae 7, 459–463. https://doi.org/10.1016/j.hal.2007.10.002 You, W., Zhi-Ming, Y., 2005. Biological Strategies in Controlling or Mitigating Marine Harmful Algal Blooms (HABs). Acta Phytoecologica Sinica 29, 665–671.
PRODUCCIÓN
de eliminar o neutralizar las toxinas de Chattonella y otras microalgas tóxicas antes de que ingresen a los sistemas de cultivo de camarón. Sin embargo, esta medida tiene limitaciones para las zonas con acceso limitado a una fuente de agua limpia. El uso de aireadores es otra estrategia muy común para contrarrestar el agotamiento de oxígeno dada la alta concentración de microalgas, pero este método no contrarresta el efecto de las toxinas secretadas al agua (Brown et al., 2020; Mirza Esmaeili et al., 2020).
Uso de químicos y alguicidas
Una de las prácticas utilizadas en camaroneras para controlar las afloraciones de Chattonella ha sido el uso de alguicidas y químicos como el sulfato de cobre. Sin embargo, aunque en ocasiones es efectivo, su uso puede conllevar efectos colaterales no deseados sobre el ecosistema y la salud de los camarones. Otras opciones han sido investigadas a nivel de laboratorio: Piyatiratitivorakul et al. (2002) logró obtener un buen control de C. marina usando peróxido de hidrogeno a 39 ppm; además, observó que la calidad del agua mostraba poca variación. Es así que se recomienda el peróxido de hidrogeno como una opción viable en el manejo de afloraciones de Chattonella
Gestión integrada de granjas
La colaboración entre productores de camarón, autoridades gubernamentales y las instituciones privadas y de investigación pueden generar esfuerzos coordinados para compartir datos de monitoreo, implementar regulaciones sobre el uso de fertilizantes en áreas agrícolas cercanas, y promover prácticas acuícolas sostenibles para reducir la frecuencia e impacto de las mareas rojas. Además, la adopción de este tipo de prácticas de gestión integrada ayudaría a los productores a incluir dentro de los procesos de producción la rotación efectiva de cultivos, tener un mejor control en la diversificación de especies planctónicas y el uso de biocontroladores naturales para reducir la vulnerabilidad a las mareas rojas (Anderson, 2014).
Investigación en toxicología y resistencia
Investigadores se encuentran explorando nuevos métodos más sostenibles para controlar las afloraciones de Chattonella. Por ejemplo, You y Zhi-Ming (2005) describen el uso de bacterias, virus y parásitos como agentes controladores de mareas rojas dada su rápida replicación y especificidad de huésped. Los autores también proponen estrategias basadas en interacciones entre las microalgas de marea roja y otras especies que podrían competir por la disponibilidad de nutrientes o secretando sustancias que sean tóxicas para las microalgas, limitando la proliferación de las algas nocivas (Pal et al., 2020; You y Zhi-Ming, 2005). Otras estrategias relacionadas con la investigación es el desarrollo de alertas tempranas que integren modelos predictivos basados en el clima y la calidad del agua para prever las condiciones que favorecen las afloraciones de microalgas tóxicas.
Conclusión
El género Chattonella representa una amenaza emergente para la industria de la acuicultura de camarones, con el potencial de causar graves pérdidas económicas y ecológicas. Este es el primer caso de estudio que reporta la presencia de quistes de Chattonella spp. en un sector de producción de camarón ubicado en el
golfo de Guayaquil. Referencias científicas indican que las especies de dinoflagelados y Raphidophyceae (Chattonella y Heterokwivo) tienen una etapa de quiste en reposo durante su ciclo de vida, pasan la mayor parte como quistes en el sedimento, los quistes recién formados se encuentran en un estado de latencia y necesitan un periodo de temperatura fría para que la maduración adquiera germinabilidad (Imai y Itoh, 1988; Imai y Yamaguchi, 2012).
Considerando que el golfo de Guayaquil no es únicamente esencial para la industria camaronera sino también para el sector turístico, la pesca y el comercio internacional, Borbor-Córdova et al. (2019) sugieren la necesidad de establecer programas de investigación sobre la salud de los ecosistemas, haciendo evidente la importancia del control de la distribución de las especies planctónicas cerca de las zonas de gran importancia económica y social•
Para más información sobre este artículo, contactar a: cesar.molina@skretting.com
Figura 3. Densidad de quistes de Chattonella spp. registrados en las cercanías de Posorja, mayo-julio del 2024.
BIENESTAR ANIMAL
Bienestar Animal: un “nuevo término” largamente aplicado en la industria camaronera ecuatoriana
Autor:
Yahira Piedrahita
Directora
Ejecutiva Cámara Nacional de Acuacultura
ypiedrahita@cna-ecuador.com
El término “Bienestar Animal” ha empezado a escucharse hace poco tiempo en Ecuador, motivado principalmente por situaciones adversas respecto a la fauna silvestre, así como a los animales de trabajo y compañía. Sin embargo, no se trata de un concepto nuevo: la industria camaronera ecuatoriana lo utiliza y aplica desde hace muchos años. Los principios de las Cinco Libertades (detalladas más adelante) se han venido empleando bajo la denominación de Buenas Prácticas de Cultivo y abarcan, en gran medida, lo que la literatura científica recoge sobre esta materia.
Ecuador siempre ha sido pionero en demostrar la búsqueda de la sostenibilidad de la industria camaronera mediante acciones que impulsan el cambio. Por ello, el propósito de este artículo es dar a conocer de dónde surgen los principios, cuáles son los indicadores y en dónde estamos respecto al nivel de bienestar proporcionado a los camarones que se cultivan en nuestras fincas, de modo que podamos cumplir con las regulaciones adicionales que se implementen sobre este aspecto, a la vez que se mantenga la productividad y rentabilidad de los cultivos y se garantice el suministro de una proteína de alta calidad a los consumidores del mejor camarón del mundo.
Las actitudes humanas hacia los animales han cambiado a medida que los animales se han incorporado más ampliamente a la categoría de agentes morales que merecen cierto respeto. Se han hecho paralelismos entre el funcionamiento de los humanos y animales durante miles de años, pero la idea de que los animales pueden sufrir se ha extendido recientemente. Una mejor comprensión de la motivación, la cognición y la complejidad del comportamiento social en los animales ha llevado en los últimos 30 años al rápido desarrollo de la ciencia del Bienestar Animal.
Los primeros intentos de definir el bienestar se referían a que los individuos estuvieran en armonía con la naturaleza, pero la primera definición utilizable incorporaba los sentimientos y la salud como parte de los intentos de lidiar con el medio ambiente. Otros consideraban que el bienestar se trata solo de sentimientos, pero se sostiene que, como los sentimientos son mecanismos que han evolucionado, son una parte del bienestar en lugar de ser su
totalidad. La mayoría de las revisiones del bienestar ahora comienzan con una lista de las necesidades del animal, incluida la necesidad de mostrar ciertas conductas. Este enfoque ha utilizado estudios sofisticados de lo que es importante para los animales y ha reemplazado las pautas generales anteriores, descritas como “libertades”. Ahora se utilizan muchas medidas de bienestar que indican cuán bueno o malo es el bienestar. La naturalidad no forma parte de la definición de bienestar, pero explica por qué existen algunas necesidades. En los últimos años, el bienestar se ha convertido en uno de los diversos criterios que se utilizan para decidir si un sistema es sostenible, porque los ciudadanos no aceptarán sistemas que provoquen un bienestar deficiente. El estudio de este concepto se ha convertido en parte de la base científica sobre la que se toman decisiones políticas importantes (Broom, 2011).
Considerando que en la actualidad la Asamblea Nacional ya debate una posible Ley de Bienestar Animal, y que los principales mercados de destino de nuestras exportaciones, incentivados por distintos grupos de usuarios, están implementando criterios específicos y medibles sobre el bienestar animal en los cultivos de camarón que se convierten en condiciones de acceso al mercado, es preciso que los actores de la industria se familiaricen con el término y los principios que lo rigen.
El concepto de las Cinco Libertades y del Bienestar Animal
Este concepto surgió en 1965, cuando el Gobierno británico designó un comité para investigar más sobre el bienestar de los animales de granja. En diciembre de ese año, el comité publicó un análisis técnico conocido como el Informe Brambell, que decía que los animales deberían poder “ponerse de pie, acostarse, darse la vuelta, acicalarse y estirar sus extremidades”. Este concepto fue refinado posteriormente por el Comité de Bienestar de los Animales de Granja del Reino Unido (actualmente Consejo de Bienestar Animal) para formar las Cinco Libertades tal como se las conoce hoy (Thiry, 2024). Las Cinco Libertades son un conjunto de principios rectores de bienestar animal que describen las libertades clave que todos los animales, incluidos los criados para la alimentación, deberían experimentar en su vida. Reconocidas por muchas personas y grupos en todo el mundo, estas se consideran el enfoque de referencia para proteger el bienestar mental y físico de los animales de granja al minimizar las experiencias y estados negativos, ocasionalmente con ligeras modificaciones.
Según el Comité de Bienestar Animal del Reino Unido, las Cinco Libertades son:
1. Libertad de pasar hambre o sed, mediante un acceso fácil al agua y una dieta para mantener la salud y el vigor completos.
2.Libertad de sufrir incomodidad,
proporcionando un entorno apropiado.
3.Libertad de sufrir dolor, lesiones o enfermedades, mediante la prevención o el diagnóstico y tratamiento rápidos.
4. Libertad de expresar un comportamiento normal, proporcionando espacio suficiente, instalaciones adecuadas y la compañía apropiada de animales de su misma especie.
5.Libertad de sufrir miedo y angustia, garantizando condiciones y tratamientos que eviten el sufrimiento mental.
En la década de 1980, surgió un consenso entre biólogos y veterinarios sobre los desafíos que enfrentan los animales debido a su entorno, como patógenos, daños en los tejidos, depredación y competencia social. La incapacidad para controlar estas interacciones genera problemas tanto para los humanos como para los animales. Aunque el Informe Brambell no definió el bienestar, Hughes (1982) lo vinculó a la armonía del animal con su entorno, lo que, aunque relevante, no permite mediciones científicas.
Broom (1986) definió el bienestar animal como el estado de un individuo en relación con su capacidad para afrontar su entorno. Afrontar implica mantener la estabilidad mental y física, y el bienestar puede medirse en un rango que va de muy bueno a muy malo. El bienestar es deficiente si hay dificultades para hacer frente a los desafíos, y las estrategias de afrontamiento incluyen componentes
conductuales, fisiológicos e inmunológicos coordinados por el cerebro. Las emociones, como el dolor y el placer, según el autor, son parte esencial de estas estrategias.
A principios de los años 90, los científicos coincidieron en que el bienestar animal es un concepto medible. Sin embargo, la definición de Broom ha sido considerada funcional y contrastada con las teorías de Ian Duncan, quien considera que el bienestar animal tiene que ver con los sentimientos que experimentan los animales (Duncan, 2005). Aunque algunos sostienen que el bienestar se centra únicamente en las emociones, Marian Dawkins argumentó que tanto las emociones como la salud son importantes para el bienestar. También se ha cuestionado la idea de que la salud abarca todo el bienestar (Dawkins, 2016).
A pesar de que algunos veterinarios comenzaron a involucrarse en la investigación del bienestar animal, la mayoría no lo consideraba como una disciplina científica. Las opiniones sobre este tema en la comunidad veterinaria y científica se centraron en el tratamiento de enfermedades, marginalizando el estudio del comportamiento animal y la función cerebral. Esto llevó a que la investigación sobre bienestar no se valorara adecuadamente en el ámbito científico. Hoy en día, los principios que sustentan las Cinco Libertades son utilizados globalmente por diversas personas en los sistemas alimentarios (incluidos agricultores y
ganaderos) en el desarrollo de protocolos y prácticas de bienestar animal, y para dar forma a la manera en que se crían animales para la alimentación. Por ejemplo, las Cinco Libertades han sido referenciadas en políticas y han influido en programas de acreditación y legislación con requisitos específicos, como estándares mínimos claros de espacio para que los animales de granja se tumben o estándares para la atención veterinaria de animales enfermos.
Bienestar Animal en acuicultura
El Código Sanitario para los Animales Terrestres de la Organización Mundial de Sanidad Animal (OMSA) indica que un animal está en buenas condiciones de bienestar si está sano, cómodo, bien alimentado, en seguridad, puede expresar formas innatas de comportamiento y si no padece sensaciones desagradables de dolor, miedo o desasosiego. Esta definición general se aplica para los sistemas acuáticos, al no existir una definición más específica en el Código Acuático. La OMSA ha elaborado normas internacionales para el bienestar de los peces de cultivo en el Código Acuático, e insta a “recurrir a métodos de manipulación que sean apropiados a las características biológicas del animal, así como un entorno adaptado a sus necesidades”.
No existe aún una regulación que determine con precisión los lineamientos del bienestar animal en camarones; sin embargo, el desarrollo de la industria y las presiones desde los grupos de conservación y ética animal vuelven necesario establecer
directrices al respecto. Las Cinco Libertades describen los estándares mínimos aceptables para los animales, y este enfoque propuesto continuaría mejorando su calidad de vida. No obstante, tendría implicaciones para los sistemas alimentarios, incluidos los recursos, los costos, la mano de obra y más. Los sistemas alimentarios deberán adaptarse para satisfacer las cambiantes necesidades de bienestar animal en el futuro (Thiry, 2024), pero también deberán tener en consideración el bienestar humano, y la necesidad de fuentes de proteína animal para la alimentación de una población en crecimiento, especialmente en los países pobres, donde existen altos índices de desnutrición infantil, enfermedades y mortalidad debido a la escasez de alimentos.
La industria camaronera y el Bienestar Animal
En términos generales, el bienestar de los cultivos de camarón se analiza desde los ámbitos de la nutrición, medio ambiente, salud y comportamiento. Criterios más específicos determinan recomendaciones respecto a la no ablación del pedúnculo ocular en las reproductoras, condiciones de calidad de agua, alimentación, manejo de enfermedades, densidades de siembra, transporte y manipulación, aturdimiento y sacrificio, entre otros. A continuación, se describen algunos de los criterios considerados por Pedrazzolli en su publicación sobre Métodos no invasivos para evaluar el bienestar de los camarones (Penaeus vannamei) en condiciones de cultivo (Pedrazzolli, et al., 2023).
BIENESTAR ANIMAL
Ablación del pedúnculo ocular: los camarones demuestran respuestas de comportamiento aversivas a la ablación del pedúnculo ocular, y la aplicación de anestesia antes de la ablación tiene efectos terapéuticos. Se considera que esto es un fuerte indicativo de que la ablación del pedúnculo ocular es una preocupación de bienestar.
Enfermedades: las enfermedades infecciosas causan importantes eventos de mortalidad. Es probable que esto cause sufrimiento antes de la muerte y aumente el número total de camarones que se cultivan y experimentan sufrimiento.
Aturdimiento y sacrificio: es importante establecer métodos adecuados para aturdir y matar camarones de manera efectiva. Aunque existen propuestas para el uso de dispositivos eléctricos, es necesario considerar los aspectos logísticos y operativos de las fincas y las implicaciones de seguridad humana en el personal. Hasta el momento, lo más aplicado es la inmersión en agua helada.
Densidad de población: existe alguna evidencia tentativa de que la densidad de población afecta directamente el comportamiento de los camarones y los biomarcadores de estrés medibles (por ejemplo, la serotonina). Por ello, es importante tomar en consideración la capacidad de carga de los sistemas de cultivo, de modo que se eviten las condiciones de estrés en los animales cultivados.
Enriquecimiento ambiental (EE): los enriquecimientos ambientales (por ejemplo, métodos de alimentación que imitan el comportamiento natural, escondites, diferentes formas y colores de tanques, plantas, sustratos y sedimentos) probablemente mejoran la supervivencia del camarón, pero hay poca evidencia sobre su impacto en el estrés del animal. Existen pruebas moderadamente sólidas de que el enriquecimiento físico (como la estructura física, la planta y el sustrato) mejora el bienestar de los animales acuáticos, incluidos los crustáceos.
•Transporte y manipulación: es probable que las prácticas deficientes de transporte y manipulación provoquen lesiones físicas y estrés, aunque la investigación sobre los efectos de las prácticas actuales de
cultivo de camarón en el bienestar es limitada. Sin embargo, se recomienda un adecuado manejo de los animales cuando son transportados o manipulados.
Alimentación: si bien algunos decápodos parecen resistentes a la falta de alimentos, una nutrición inadecuada conlleva el riesgo de sufrir enfermedades no infecciosas y puede provocar un comportamiento agresivo y anormal. Es importante el suministro de alimento de calidad y suficiente para evitar condiciones adversas de bienestar.
Calidad del agua
•Oxígeno disuelto (OD): varios estudios indican que unos niveles insuficientes de OD aumentan la mortalidad, que probablemente vaya precedida de sufrimiento debido a una mala condición física. Los niveles óptimos de OD también pueden ayudar a prevenir el comportamiento agresivo.
•Amoníaco no ionizado: las altas concentraciones de amoníaco no ionizado (NH3) son tóxicas para los camarones y perjudiciales para su bienestar. El alto nivel de amoníaco no ionizado dificulta la respuesta inmune y conduce a altas tasas de mortalidad.
•pH: las desviaciones del pH óptimo tienen efectos perjudiciales sobre la salud, la inmunidad y la susceptibilidad a las enfermedades. Las fluctuaciones repentinas (dentro del rango óptimo de pH) también pueden ser perjudiciales.
•Temperatura: los camarones se adaptan a las fluctuaciones de la temperatura del agua. Sin embargo, es muy probable que las altas temperaturas sean perjudiciales para el bienestar, ya que el agua retiene menos oxígeno, aumenta el amoníaco tóxico y empeora la supervivencia. Aunque pequeñas desviaciones por debajo de la temperatura óptima pueden ser menos dañinas, las temperaturas muy bajas también perjudican la salud física y provocan mortalidad.
•Salinidad: si bien es probable que los niveles o fluctuaciones extremas de salinidad sean perjudiciales para el bienestar y la supervivencia, la evidencia experimental no demuestra consistentemente efectos perjudiciales por pequeñas desviaciones del rango de salinidad recomendado.
Principios de Bienestar Animal aplicados en la industria camaronera ecuatoriana
El término “Bienestar Animal” ha empezado a escucharse hace poco en Ecuador, la industria camaronera nacional ha venido aplicando estos principios desde hace muchos años.
El bienestar animal en acuicultura contribuye a tener animales mejor adaptados durante todo el ciclo de cultivo y se evidencia en la calidad del producto final, obteniendo mayor porcentaje de producto de calidad premium
Para la industria ecuatoriana, el bienestar de los cultivos siempre ha sido una prioridad , ya que camarones mantenidos en condiciones inadecuadas tienen conversión alimenticia más alta, requieren mayor uso de productos, incluido antibióticos para mejorar la salud; tienen mayor riesgo de morir y, en general, representan un alto riesgo de pérdidas económicas.
A continuación, se detallan algunas prácticas concretas que demuestran la alineación de la industria a las Cinco Libertades y a los principios de bienestar.
A nivel de maduraciones de reproductores y criaderos de larvas:
• Progresivamente se está eliminando la técnica de ablación en las facilidades de reproducción y desove.
• Se aplican criterios de exclusión o reducción de la carga de patógenos para seleccionar reproductores, sin utilizar animales libres de patógenos específicos (SPF).
• El éxito de la industria se apoya en los programas de mejoramiento genético, que se basa en la cría de animales expuestos a todos los patógenos (APE). Los reproductores provienen únicamente de granjas locales y las postlarvas son cultivadas tomando en consideración las condiciones en las que completarán su ciclo de cultivo. Se aplican criterios de buenas prácticas y se está implementando un estándar de sostenibilidad (proyecto Sustainable Shrimp Partnership).
• La autoridad nacional ejecuta planes de vigilancia sanitaria para verificar
las condiciones en las que se mantienen los animales en producción.
A nivel de fincas:
• Las granjas operan principalmente a través de sistemas abiertos, con intercambio de 5-15% de agua desde cuerpos naturales. Algunas fincas han implementado sistemas de recirculación para asegurar una mejor calidad del agua de cultivo.
• Se utilizan nuevas tecnologías para mejorar la productividad, la eficiencia y la sostenibilidad, incluida la aireación, la alimentación automatizada, el seguimiento y control automatizados, y la inteligencia artificial.
• La industria emplea alimentos funcionales bajo el concepto de “nutrición de precisión”, utilizando materias primas sostenibles y aditivos que promueven una mejor asimilación del alimento suministrado, generando condiciones de salud adecuadas.
• Las densidades de siembra se ajustan a la capacidad de carga de los sistemas de cultivo y, de manera general es menor que en el resto de los países productores.
• Las condiciones de cosecha se han ido automatizando, de modo que el proceso
es más rápido. Los animales son sumergidos en agua helada (a 4° C) tan pronto salen del estanque, por lo que su muerte se produce rápidamente por hipotermia y minimizando el sufrimiento.
Regulaciones sobre la salud en los cultivos y el uso de antibióticos
Los notables progresos en la reducción del uso de antibióticos en la industria influyen de manera positiva no solo en el bienestar animal, sino también en el bienestar humano y ambiental. Los productores son cada vez más conscientes del riesgo de los antibióticos sobre el medio ambiente y la salud humana, y también sobre el riesgo de rechazo de productos y sanciones en los mercados finales. Por lo tanto, son más responsables y se preocupan más de decidir sobre el uso o no de antibióticos.
En 2017, la Cámara Nacional de Acuacultura (CNA) reportó exportaciones totales de 848,000 TM de camarón, mientras la autoridad sanitaria nacional, la Subsecretaría de Calidad e Inocuidad (SCI), reportó autorizaciones para la importación de 131.30 TM de antibióticos aprobados para uso en acuacultura, lo que indica que en promedio se aplicó antibióticos en una proporción de 297.0 g/TM de de camarón (sin considerar que la producción siempre es mayor que el
BIENESTAR
volumen de las exportaciones). Para el 2021, las exportaciones de camarón alcanzaron 848,000 TM, mientras que únicamente se importaron 29.57 TM de antibióticos para uso en acuicultura, lo que demuestra que se ha reducido la aplicación de antibióticos a menos de 35.0 g/TM. Diferentes productos son utilizados como sustitutos de los antibióticos (probióticos, ácidos orgánicos, aceites esenciales, etc.) y se han aplicado terapias de fagos a los cultivos con muy buenos resultados. Todos los tratamientos han sido probados en bioensayos, así como en el campo de producción.
Es muy importante tener en cuenta que las regulaciones locales sobre uso de antibióticos están alineadas con los requerimientos de acceso a los principales mercados del camarón ecuatoriano (Unión Europea, Estados Unidos y China), así como a otros de interés. La autoridad sanitaria nacional (SCI) es auditada periódicamente por las autoridades sanitarias de los mercados del camarón ecuatoriano con resultados satisfactorios. Las granjas que optan por una certificación de tercera parte no enfrentan inconvenientes respecto al cumplimiento de los estándares sobre antibióticos y trazabilidad.
La SCI y la CNA trabajan activamente en el Plan de Acción Nacional sobre
Resistencia a los Antimicrobianos, mediante una agenda con actividades e hitos a cumplir, lo que se ha demostrado con la reducción en el uso de estos agentes.
Bienestar Animal en los aspectos legales y los requisitos de acceso a mercados
La Constitución de la República del Ecuador determina en su artículo 71 que la naturaleza tiene derecho a que se respete integralmente su existencia, así como el mantenimiento y regeneración de sus ciclos vitales, estructura, funciones y procesos evolutivos. Por otro lado, el artículo 281 señala que la soberanía alimentaria constituye un objetivo estratégico y una obligación del Estado para garantizar que las personas, comunidades, pueblos y nacionalidades alcancen la autosuficiencia de alimentos sanos y culturalmente apropiados de forma permanente. Para ello, será responsabilidad estatal precautelar que los animales destinados a la alimentación humana estén sanos y sean criados en un entorno saludable. Tomando en consideración estos dos artículos de nuestra Constitución, es necesario que las normativas de menor jerarquía que se promulguen equilibren los principios del bienestar animal con los principios de soberanía alimentaria, y permitan el desarrollo sostenible de las industrias de producción de animales de consumo.
A nivel internacional, también es necesario tomar en consideración algunos tratados internacionales, como el Acuerdo sobre la Aplicación de Medidas Sanitarias y Fitosanitarias (MSF) de la Organización Mundial de Comercio (OMC), respecto a las condiciones bajo las que se comercializan los animales y sus productos (referido principalmente al manejo de la salud y los patógenos presentes en los animales, sus productos y subproductos). El Acuerdo MSF está alineado también con el Código Sanitario de los Animales Terrestres y al Código Acuático de la Organización Mundial de Sanidad Animal (OMSA), en lo referente a los patógenos de declaración obligatoria, métodos de manejo apropiados para las características biológicas de los peces y un entorno adecuado para satisfacer sus necesidades.
A nivel comercial también existen movimientos que exigen mejorar las
condiciones de cultivo de los camarones y se están realizando ajustes en las certificaciones privadas. Un ejemplo de ello es la organización denominada Shrimp Welfare Project (Proyecto de Bienestar del Camarón) que menciona como objetivo “mejorar las vidas de miles de millones de camarones de cultivo en todo el mundo”, impulsando la reducción del riesgo de enfermedades, las condiciones adecuadas de la calidad de agua y la eliminación de la ablación en las hembras. Un reflejo de las presiones ejercidas por los grupos de bienestar animal es la nueva versión de estándar para fincas que plantea la certificación que otorga Aquaculture Stewardship Council (ASC), cuyo borrador final ya ha sido sometido a consulta pública, y en el que se ha incluido un nuevo capítulo destinado al Bienestar Animal en el que incorporan varios indicadores relacionados con la no ablación de las reproductoras, las condiciones del ambiente de los estanques y los métodos de sacrificio dignos. Del mismo modo, en Latinoamérica ya existe una certificación de bienestar animal para empresas pecuarias y acuícolas que ofrece una serie de beneficios para aumentar la confianza de los usuarios, y busca entregar una ventaja competitiva para los establecimientos en un mercado altamente competitivo.
La Ley Orgánica para el Desarrollo de la Acuicultura y Pesca (LODAP), expedida en 2020, su Reglamento (2022) y el Plan Nacional de Control Sanitario para la Acuicultura y la Pesca (actualizado en 2023), así como el Código Orgánico del Ambiente (COA), contemplan de manera implícita y transversal el concepto de Bienestar Animal y los principios de las Cinco Libertades en varios de sus articulados. Adicionalmente, la Cámara Nacional de Acuacultura ha trabajado proactivamente aportando insumos a la Comisión de Biodiversidad de la Asamblea Nacional para que el proyecto de Ley de Bienestar Animal que está en elaboración tome en consideración los aspectos relevantes de nuestra industria y siga fomentando su desarrollo sostenible para fortalecer la economía del país y mejorar las condiciones de vida de los ciudadanos.
Conclusiones
Si bien es cierto que la industria camaronera de Ecuador ha venido aplicando los principios
de Buenas Prácticas de Cultivo, y que con ello abarca en gran medida los aspectos del Bienestar Animal, es importante entender que las preferencias del consumidor y los requisitos de acceso al mercado en adelante considerarán cada vez más las condiciones de bienestar bajo las cuales son producidos, cosechados y procesados los animales destinados al consumo humano.
Bibliografía
Broom, D. M. (2011). A History of Animal WelfareScience. Acta Biotheor, 59, 121–137. doi:10.1007/ s10441-011-9123-3
Dawkins, M. S. (2016). Animal welfare with and without consciousness. Journal of Zoology.(301 (2017) 1–10). doi:10.1111/jzo.12434
Duncan, I. (2005). Science-based assessment of animal welfare:. Rev. sci. tech. Off. int. Epiz., 24(2), 483-492.
Pedrazzolli, A. S., Cozer, N., Quintiliano, M. H., dos Santos Tavares, C. P., Teixeira da Silva, U., & Ostrensky, A. (2023). Non-Invasive Methods for Assessing theWelfare of Farmed. Animals. doi:https://doi. org/10.3390/ani13050807
Thiry, M. (2024, March 5). Academy of Nutrition and Dietetics. Retrieved from Animal Welfare: The Five Freedoms: https://acortar.link/ Me11Pv
A pesar de que los debates a nivel local hasta ahora se han centrado principalmente en los animales de compañía y en la producción terrestre, es importante mantenerse atentos a los cambios en las normativas y en las preferencias del mercado, ya que las condiciones pueden variar y hay que estar preparados para lo que ello represente. Lograr un crecimiento continuo y sostenible, asegurando el bienestar de los cultivos, es posible• Para más información sobre este artículo, contactar a: ypiedrahita@cna-ecuador.com
Guía práctica para el traslado de camarón vivo desde la finca hasta la planta procesadora
Autor: Julio Castañeda Aquapesca S. de R.L. aquapesca@hotmail.com
En la industria camaronera, uno de los desafíos más persistentes es la optimización de la rentabilidad sin sacrificar la calidad del producto final. El camarón blanco del Pacífico (Penaeus vannamei) es una de las especies más cultivadas en el mundo, y su éxito comercial depende en gran medida de sus características organolépticas, especialmente de la coloración, que es un indicador visual clave en la aceptación del producto por parte del consumidor. Un camarón de color más oscuro, como el clasificado en la categoría A4, es particularmente valorado en mercados internacionales alcanzando precios más altos y, por tanto generando mayores ganancias para los productores.
Los principales países importadores de P. vannamei en la presentación HOSO (Head-On Shell-On, o camarón con cabeza y caparazón completos) son:
• China: es uno de los mayores importadores de camarón HOSO debido a su alta demanda interna.
• Estados Unidos: aunque prefieren productos de valor agregado, también importan cantidades significativas de camarón HOSO.
• España: es uno de los principales importadores en Europa, especialmente de productos crudos o cocidos con cabeza y cáscara.
• Francia: otro gran importador europeo que consume una variedad de productos de camarón.
• Italia: importa camarón HOSO para su uso en la cocina tradicional y en la industria de alimentos.
Fuente: Cultivo de camarones vannamei: el papel de diferentes países en el mercado global (rodaint.com).
Sin embargo, no todas las regiones productoras permiten naturalmente la obtención de camarones con esta coloración. En muchos casos, los productores enfrentan el reto de mejorar la coloración de camarones que inicialmente presentan un color más claro, como el A2. Ante esta necesidad, se ha desarrollado un Protocolo de manejo de camarón vivo desde la finca de producción hasta la planta de proceso, que permite optimizar la coloración y mantener otras características organolépticas deseables,
como el sabor, la textura y la integridad del hepatopáncreas durante el proceso de cocción.
La importancia de la coloración en el camarón P. vannamei
Uno de los mayores retos que enfrenta actualmente la industria del camarón de cultivo es la falta de rentabilidad de esta actividad. Por eso, el manejo eficiente de la cosecha es crucial para mantener las características organolépticas del camarón, asegurando la mejor calidad durante el procesamiento y empaque hasta su despacho a destino.
Se han implementado diversas estrategias para mejorar las utilidades económicas en la producción camaronera, siendo una de ellas la diferenciación en las características organolépticas, especialmente el color del camarón, ya que un camarón de color más oscuro como el A4 alcanza un mejor precio en el mercado. Es fundamental que las demás características organolépticas también sean óptimas (olor, sabor, textura). En algunas regiones productoras, los camarones presentan naturalmente una coloración más clara; en estos casos, la aplicación de un protocolo viable para ganar color es de suma importancia, económicamente hablando (Foto 1).
El presente artículo detalla el procedimiento utilizado por empresas productoras y exportadoras de Centroamérica y Ecuador que logran los mejores resultados en la obtención de camarones con coloración A4, partiendo de una coloración A2. Esta tecnología se ha desarrollado a partir de la necesidad de exportar a mercados que demandan camarón HOSO de alta calidad (Figura 1).
Protocolo
A continuación, se describen los aspectos metodológicos del protocolo de manejo de camarón vivo:
a. Muestreos para determinar el momento óptimo de cosecha del camarón en la piscina de cultivo
Los muestreos de precosecha para el camarón cultivado son cruciales para evaluar la calidad y el estado de salud de los camarones antes de la cosecha (Foto 2).
• Frecuencia de muestreo: realizarlos de forma programada, generalmente cada 15 días, y aumentar la frecuencia de muestreos a medida que se acerque la fecha de cosecha.
• Tamaño de muestra: tomar una muestra representativa de la población, generalmente entre 50 y 100 camarones por estanque.
• Parámetros a evaluar:
-Peso y talla: valorar el peso y la longitud de los camarones para evaluar su talla y conteo empacado.
-Condición física: revisar la apariencia externa, buscando signos de enfermedades o deformidades.
-Análisis de salud: realizar pruebas para detectar enfermedades comunes que puedan afectar negativamente los resultados de empaque (por ejemplo, la flacidez patológica, variación de tallas y cualquier otro que demerite la calidad de camarón a procesar, empacar y exportar).
-Empaque objetivo: la meta es empacar la mayor cantidad de camarones de cada piscina en camarón entero (HOSO).
• Registro de datos: mantener un registro detallado de todos los datos obtenidos durante los muestreos para tomar decisiones acertadas sobre la cosecha.
Foto 1: Camarón A2 (pálido), amerita manejo vivo para ganancia de color.
Figura 1: Tabla de referencia de color de camarón Penaeus vannamei, crudo y cocido.
MANEJO
Estos pasos aseguran que los camarones estén en óptimas condiciones para el momento de la cosecha y así optimizar la calidad del producto final para poder maximizar los beneficios económicos al momento de la comercialización.
b. Preparación de la piscina para la cosecha Una vez definido el mejor momento para cosechar, se programa la cosecha y se realizan los preparativos necesarios. Esto se lleva a cabo en total coordinación con la planta de proceso. Es crucial revisar minuciosamente todos los aspectos logísticos, como el transporte, materiales, equipos, personal de apoyo y seguridad necesarios para el manejo del camarón vivo en la finca y su traslado a la planta de proceso.
Antes de iniciar la cosecha, uno de los procedimientos más importantes es bajar el nivel del agua en la piscina hasta el nivel operativo de cosecha.
Hay que tener mucho cuidado con bajar muy bruscamente el nivel de agua de cultivo a nivel operativo de cosecha, ya que esto puede provocar la muda. Así también, durante la cosecha, detener y volver a activar la salida de agua puede causar la entrada en muda del camarón y por ende, tener que suspender la cosecha.
c. Preparación del equipo e instrumentos de trabajo y envío a campo
Es esencial asegurar la correcta instalación de la máquina cosechadora y los equipos de apoyo, los tanques (tinas) de acondicionamiento, el abastecimiento de oxígeno, el hielo para bajar la temperatura, así como el personal y equipo de apoyo para permitir que el buen flujo vehicular.
La ubicación de todo el equipo es fundamental para permitir un flujo eficiente de camiones con bines isotérmicos que transporten el camarón vivo (Foto 3). Un factor muy importante es el equipo técnico. Es fundamental contar con personal técnico y colaboradores debidamente capacitados en la aplicación de protocolo de manejo de camarón vivo.
A lo interno de la tina de acondicionamiento de camarón, se debe colocar una malla sin
Foto 2: Técnico responsable de finca realizando muestreo rutinario de camarón conjuntamente con el supervisor de cosecha que reporta a departamento de Control de Calidad de planta de proceso. Evalúan peso promedio, flacidez, defectos, disparidad de tallas.
Foto 3: Instalación de los equipos en la borda de cosecha. De izquierda a derecha: Cosechadora de especies vivas, manguera de descarga, tina de acondicionamiento, cilindro de oxígeno con regulador, mangueras de presión y difusores, gavetas con red, tarimas de acero inoxidable forradas con malla acuícola de 6 mm, camión plataforma para traslado de bines con camarón vivo.
nudos tipo Raschel, para facilitar la recolecta del camarón que escapa sin dañar su caparazón (Foto 4).
Foto 4: Malla tipo Raschel, sin nudos.
Foto 5: Ubicación de gavetas dentro de tina de acondicionamiento.
La tubería de descarga de la cosechadora mecánica debe llegar hasta el punto donde sale el camarón y se llenen las gavetas ranuradas con camarón que se va a acondicionar para posteriormente ser trasladado en bines isotérmicos al área de mimetizado, ya sea dentro de la finca o a una planta de proceso (Foto 5)
Nota: En AQUAEXPO Guayaquil 2024 se presenta una propuesta de manejo en finca que logra la reducción de costos de producción con menor empleo de esfuerzo laboral y reducción de hasta un 30% en los costos de transporte de camarón a planta de proceso (Fotos 6 y 7).
d. Acondicionamiento
Los mejores resultados se logran realizando un acondicionamiento muy cuidadoso. Es necesario preparar al camarón para que su sistema respiratorio (branquias) esté limpio y apto para recibir la hipersaturación de oxígeno durante su traslado en bines.
Es fundamental reducir gradualmente la temperatura para que el camarón disminuya su metabolismo de manera controlada. Durante el acondicionamiento, los camarones se colocan en gavetas recubiertas con malla sin nudos (Raschel), en cantidades adecuadas para evitar daños por aplastamiento (35-50 libras) (Foto 5)
Los camarones se retienen en el agua de acondicionamiento dentro de cada gaveta durante 5 a 8 minutos, permitiendo así que asimilen el oxígeno a una concentración 8 miligramos por litro (saturación mecánica de O2) a una temperatura de 20 °C.
e. Llenado de bines con agua
Los bines de transporte de camarón vivo son una herramienta muy importante. Previo a su uso en planta de proceso, debe de seguirse una rutina de limpieza, sanitización y preparación de los bines para la siguiente corrida (ver procedimientos recomendados en la referencia bibliográfica No. 3 al final del artículo).
Los bines deben de armarse en planta de la siguiente forma: colocar las cerámicas de aireación al fondo con sus mangueras de abastecimiento de oxígeno líquido con
6 y 7: Muestran la colocación de la tina de acondicionamiento en el punto de cosecha, prestando atención a la ubicación de la manguera de descarga del camarón hacia las gavetas dentro de la tina. Esta manguera también se utiliza para llenar o reponer el agua de acondicionamiento.
codificación de color: rojo, lado izquierdo conectadas al manifold del lado izquierdo y, verdes del lado derecho, conectadas al manifold del lado derecho (Foto 9).
Sobre las cerámicas se coloca la primera tarima baja de acero inoxidable con su respectiva malla para impedir el paso de camarón a la cámara de aireación (Foto 10).
Sobre esta primera tarima se vacía el hielo
a granel, la cantidad depende del tiempo de traslado, usualmente para traslados de 6 a 8 horas unas 300 libras de hielo en escarcha. Sobre esta cama de hielo se colocan otros 12 sacos de hielo en su saco y se cierra el bin con su tapadera. Lo usual es utilizar camiones plataforma equipados con los recipientes de oxígeno líquido y sus manifolds (derecho e izquierdo) para abastecer 10 o 12 bines de transporte de camarón vivo (Foto 11).
Fotos
Foto 8: Muestras de distintas tonalidades de color que va adquiriendo el camarón con el protocolo de manejo vivo.
MANEJO
f. Traslado de camarón vivo a planta Luego del acondicionamiento, los camarones que se trasladan vivos a planta viajan dentro de bines isotérmicos de 1000 litros de capacidad. Es muy importante que los bines sean de excelente calidad y que resistan el trabajo pesado. Adicionalmente, tienen que ser sellados para evitar pérdida de agua durante el traslado.
En respuesta a la necesidad creciente de manejo de camarón vivo en la industria, se ha diseñado un nuevo modelo de bin y sus componentes. Estos se equipan con sistemas de aireación que consisten de cerámicas que producen burbujas muy finas y se logra una excelente difusión del oxígeno liquido, que se aplica para lograr una hipersaturación. Asimismo, en el interior de los bines se colocan parrillas construidas en acero inoxidable SS forradas con malla acuícola, para lograr separar a los camarones en pisos y que no se transporten aglomerados (Foto 9).
El primer piso no lleva camarón, se reserva para la colocación de las cerámicas de difusión de oxígeno (Foto 10).
Los otros pisos se forman con parrillas un poco más altas, y en cada uno se colocan libras de camarón preacondicionado, logrando trasladar hasta 700 libras de camarón vivo.
En cuanto a la malla acuícola, se recomienda utilizar una que contenga tratamiento anti UV, ya que su duración es mayor ante la continua exposición al sol; así también, que sean de grado acuícola para garantizar la inocuidad del producto y su trazabilidad (Figura 2).
11: Camión
en planta de proceso con bines isotérmicos preparados y sistema de abastecimiento de oxígeno líquido.
g. Llenado de los bines con agua para transporte Es de vital importancia la calidad de agua que se utiliza para el trabajo de acondicionamiento y transporte de camarón vivo. Se recomienda utilizar los bines con el primer piso con su parrilla y cerámica ya instalados. Asimismo, cargar con hielo a granel el contenido de unos seis sacos bien acomodados para bajar la temperatura en el agua de la tina de acondicionamiento.
Cuando el camión llega a la finca hay que descargar todos los sacos de hielo en el sitio de cosecha, justo a la par de donde se hará el acondicionamiento del camarón en las tinas preparadas para tal fin.
Paralelo al camión plataforma que transporta el camarón vivo, es necesario enviar otro camión plataforma que traslada todo el equipo de apoyo para trabajo en el sitio de cosecha (Foto 13).
El agua que se utiliza en todo el proceso de manejo de camarón vivo tiene que ser obligatoriamente de la mejor calidad. Para ello se busca en la finca camaronera el mejor punto para sacar el agua y trabajar el acondicionamiento, así como para llenar los bines con hielo a granel que servirán para transporte de camarón vivo (Fotos 14 y 15).
Factores físico-químicos del agua de acondicionamiento
Puesto que se trata de manejo de camarón vivo, lo más importante a considerar en todo momento es el estado del camarón; por lo tanto, durante el acondicionamiento debe mantenerse el camarón por 5 a 10 minutos en agua con 8 mg/lit de Oxígeno a 20 °C. Luego de esto, el camarón ya está acondicionado para continuar con el proceso de traslado.
Foto 9: Bin isotérmico para traslado de camarón vivo, equipado con cerámica de aireación y tarima con malla acuícola.
Foto
plataforma cargado
Foto 10: Cerámicas de difusión de oxígeno instaladas dentro del bin y parrilla de primer piso, manguera con código de color rojo (lado izquierdo).
Figura 2: Malla rómbica PEAD 6 X 6 mm.
Factores físico-químicos del agua de transporte
Hipersaturación de oxígeno en un rango de 25 a 27 mg/lit y temperatura 15 a 17 °C.
Foto 16: Multímetro digital para medir oxígeno (lectura de arriba) y temperatura (lectura de abajo).
Homeostasis
del camarón
Este factor tiene gran importancia, por lo que se busca la manera de mantener relativamente constante su equilibrio con un funcionamiento corporal óptimo. A continuación, analizaremos este proceso conocido como homeostasis.
Hay que tener en cuenta lo importante que es el monitoreo de los parámetros de oxígeno y temperatura en todo momento. Se recomienda realizar muestreos durante la cosecha a un intervalo de media hora a una hora de frecuencia (Foto 17).
17: Técnico de finca y supervisor de cosecha realizando chequeo rutinario del estatus de camarón durante la cosecha, cada media a una hora de intervalo. Finalidad: asegurar la calidad del camarón que se llevara a manejo vivo (flacidez, talla, homeostasis).
14: Agua limpia para tina de acondicionamiento.
Traslado de camarón vivo
El tiempo de transporte del camarón a la planta de proceso es muy importante, en el que se debe realizar el monitoreo de los parámetros de oxígeno y temperatura, y sobre todo, el estatus del camarón que debe ir en letargo mediante baja de temperatura.
Los tiempos pueden variar de acuerdo con la distancia y condiciones logísticas involucradas. Los mejores resultados se han alcanzado con tiempos de traslado de 6 a 8 horas, pero para tiempos de traslado mayores se debe emplear una técnica híbrida.
Resultados
La sobrevivencia mínima aceptable al final del proceso de manejo de camarón vivo es de 95%; por debajo de esto se compromete la viabilidad económica del protocolo.
La mezcla de factores preacondicionamiento, letargo e hipersaturación de oxígeno provoca un cambio en la coloración de los camarones, y los torna más oscuros (Fotos 18 y 19).
Tabla 1: Resultados de sobrevivencia en camarón manejado vivo
Foto 13: Camión plataforma de apoyo que traslada los utensilios.
Foto
Foto
Foto 12: Parrillas de acero inoxidable recubiertas con malla acuícola.
Foto 15: Bombeando agua limpia de la mejor calidad en canal reservorio de finca camaronera.
MANEJO
color A3 oscuro, manejado vivo a partir de color A2.
Conclusiones:
El protocolo de manejo de camarón vivo para ganancia de color de A2 a A4 es viable y factible económicamente.
La sobrevivencia debe reflejar porcentajes arriba de 95% para ser una operación viable. El protocolo de manejo de camarón vivo presentado es una alternativa real para reducir costos, incrementar calidad y generar rentabilidad.
Este protocolo es un lineamiento básico de manejo, y para su aplicación es necesario realizar las adaptaciones de acuerdo con las características particulares de cada finca•
Para más información sobre este artículo, contactar a: aquapesca@hotmail.com
Resultados comparativos
A continuación, se presentan las evidencias fotográficas del proceso:
Foto 18: Camarón
Foto 19: Camarón color A4.
Foto 20: Coloración A4 de camarón manejado bajo protocolo de camarón vivo.
Foto 21: Camarón manejado vivo, 0% hepatopáncreas destruido en cocción (misma fuente, la misma cosecha).
Foto 22: Camarón de pesca tradicional, 75% de hepatopáncreas destruido en cocción (misma fuente, la misma cosecha).
El Pacto Verde Europeo y sus implicaciones para el sector exportador camaronero
Autores:
Leonardo S. Maridueña
Simone Maridueña
Dirección de Ambiente
Cámara Nacional de Acuacultura lmariduena@cna-ecuador.com
El presente artículo resume las condiciones que en corto y mediano plazo limitarán las exportaciones al mercado de la Unión Europea.
El cambio climático es la crisis determinante de nuestro tiempo y afecta de manera desproporcionada a las personas más vulnerables del mundo.
En un escenario de calentamiento global, las condiciones meteorológicas extremas (fuertes lluvias, sequías, olas de calor, tormentas tropicales) son cada vez más impredecibles, más intensas y más frecuentes, al tiempo que aumenta el riesgo de nuevos fenómenos adversos, tanto repentinos como de evolución lenta, tales como inundaciones, deslizamientos de tierras, erosión, incendios forestales y desertificación. Al mismo tiempo, el aumento del nivel del mar está provocando mayores inundaciones costeras, erosión, salinización de los suelos y la amenaza de una inundación permanente de zonas bajas.
¿Qué es el cambio climático?
Se denomina cambio climático a la variación global del clima del planeta debido a causas naturales, pero principalmente por la acción humana, que se traduce en quema de combustibles fósiles, pérdida de bosques y otras actividades producidas en los ámbitos industrial, agrícola, transporte, entre otros, teniendo como principal consecuencia el colapso de los ecosistemas y por consiguiente, se alteran sus estructuras, funciones y biodiversidad.
Cuando hablamos de combatir el cambio climático, siempre nos referimos a reducir las emisiones de gases de efecto invernadero. Pero no se trata solo de eso.
Para lograr la neutralidad climática hasta el año 2050, la Unión Europea ha establecido un programa en que involucra no solamente a los Estados Miembros que la conforman, sino que lo hace extensivo a las naciones que proveen de productos a la UE.
¿Qué es el Pacto Verde Europeo?
Es un conjunto de iniciativas políticas cuyo objetivo es posicionar a los países que conforman la Unión Europea en el camino de la transición ecológica, con el objetivo de
alcanzar la neutralidad climática a partir de hoy hasta el 2050.
El Pacto Verde Europeo fue adoptado en diciembre del 2019, estableciendo el plan rector que se aplica a este cambio transformador e involucra factores como el clima, ambiente, energía, transporte, industria, agricultura, acuicultura y finanzas sostenibles, todos ellos estrechamente relacionados.
Los objetivos climáticos y la política exterior de la UE
El cambio climático no observa ni respeta fronteras, y ningún país está a salvo. Conscientes de esta problemática, los Estados Miembros de la UE colaboran con sus socios mundiales para impulsar y financiar una acción urgente por el clima. Para una profundización de la temática, se establecen los siguientes objetivos:
Ser “climáticamente neutro” significa reducir al máximo las emisiones de gases de efecto invernadero, pero también compensar las emisiones restantes. De esta manera, se puede lograr un balance de emisiones netas cero.
Este balance se alcanza cuando se neutraliza la cantidad de gases de efecto invernadero liberados a la atmósfera. Esto puede lograrse mediante el “secuestro de carbono”, es decir, eliminando carbono de la atmósfera, o mediante medidas de compensación, que generalmente implican el apoyo a proyectos orientados al clima.
Emitiendo menos…
Todos los sectores económicos pueden y deben contribuir a reducir las emisiones de gases de efecto invernadero. Por ejemplo, la industria debe seguir modernizándose y contaminar menos. Los sectores de la aviación y el transporte marítimo, que se encuentran entre las fuentes de emisiones de gases de efecto invernadero de mayor crecimiento, deberían volverse más eficientes energéticamente y pasar a combustibles alternativos más ecológicos.
Para garantizar la igualdad de condiciones para las empresas europeas, el nuevo Mecanismo de Ajuste en Frontera por Carbono garantizará que los productos
importados también paguen un precio del carbono en la frontera en los sectores cubiertos. Se trata de una herramienta valiosa para promover la reducción de las emisiones mundiales y aprovechar el mercado de la UE para perseguir nuestros objetivos climáticos mundiales.
“Solo podemos resolver la amenaza global del cambio climático trabajando con nuestros socios internacionales”.
…absorbiendo más
A pesar de las reducciones, algunas emisiones serán inevitables. Entonces, ¿cómo se pueden neutralizar las restantes? Tanto los océanos como el suelo absorben dióxido de carbono de la atmósfera, pero los bosques representan la forma más eficaz de marcar la diferencia.
Los ecosistemas naturales que tienen la capacidad de absorber más carbono del que emiten se denominan “sumideros de carbono”. Las acciones para proteger los océanos, el suelo y los bosques, especialmente los manglares, son vitales para absorber las emisiones.
Haciendo respetar la ley
Tomando en cuenta estos antecedentes, la Unión Europea no solamente aplicará medidas a sus Estados Miembros, sino que las extenderá a aquellos países de los que importa productos de diversa índole.
Para el efecto, la UE creó la Ley Europea del Clima, que convierte en norma el objetivo establecido en el Pacto Verde Europeo para que la economía y la sociedad europeas sean climáticamente neutras para 2050. La ley también establece el objetivo intermedio de reducir las emisiones netas de gases de efecto invernadero en al menos un 55% para 2030, en comparación con los niveles de 1990.
La neutralidad climática de ahora al 2050 significa lograr cero emisiones netas de gases de efecto invernadero para el conjunto de los países de la UE, principalmente mediante la reducción de emisiones, la inversión en tecnologías ecológicas y la protección del medio ambiente.
Los objetivos de la ley incluyen:
•Establecer la dirección a largo
plazo para alcanzar el objetivo de neutralidad climática para 2050 a través de todas las políticas, de manera socialmente justa y rentable.
•Establecer un objetivo más ambicioso de la UE para 2030, con el fin de situar a Europa en una senda responsable para alcanzar la neutralidad climática de aquí al 2050.
•Crear un sistema para monitorear el progreso y tomar medidas adicionales si es necesario.
•Proporcionar previsibilidad a los inversores y otros agentes económicos.
•Garantizar que la transición hacia la neutralidad climática sea irreversible.
¿Qué ocurre con Ecuador?
Para dar viabilidad a esta ley, una Comisión de la Unión Europea, en conjunto con delegados del Ministerio de Producción, Comercio Exterior, Inversiones y Pesca, elaboraron en marzo de 2023 un informe denominado “Plan de acción para la perfecta adecuación al Pacto Verde Europeo de las seis cadenas priorizadas”.
En este documento se incluye al sector acuicultor inmerso en la cadena de pesca, resaltando que el 48% de la producción exportable de camarón del país cuenta con certificaciones socioambientales (Global GAP, ASC) que incluyen la auditoría de criterios de sostenibilidad ambiental y social. Sin embargo, el documento manifiesta que “aunque no se identifican reportes o denuncias relacionadas con el incumplimiento de los derechos laborales, aproximadamente el 62% de las exportaciones de camarón no estarían preparadas para cumplir con los requisitos de debido cumplimiento con el Pacto Verde Europeo, al no contar con trazabilidad con alguna certificación voluntaria”.
El informe indica además que “los productores de pequeña escala tienen dificultad de llegar a los mercados que valoran las certificaciones voluntarias de sostenibilidad, porque son las empacadorasexportadoras las que tienen conexión con los mercados de exportación”. La comisión también identificó que los eslabones de la cadena desconocen qué es la huella de carbono y cómo disminuirla, y evidenció que el uso del diésel y refrigerantes
representan el 89% de los gases de efecto invernadero, incluyendo al sector pesquero. En este aspecto, el informe resalta “el no desarrollo de los esquemas de acreditación y respectivo reconocimiento internacional, con la excepción de una empresa en sector acuícola”
También enfatiza que no acceder a acreditaciones flexibles en alcances como pesticidas, antibióticos y metales pesados, y la falta de sostenibilidad de laboratorios públicos y privados acreditados, puede resultar en pérdidas de conectividad, por no transparentar los análisis de los resultados y la falta de una ley de fraude alimentario.
Al autor de esta publicación se le indicó que entre las solicitudes específicas para el sector acuicultor de camarón, se incluirá el tema de reforestación del manglar como
sujeto de compensación por la tala de esta formación vegetal y un mayor control al uso de plásticos, específicamente en los llamados “pañales” para las cajetas de exportación.
El primer aspecto se ha convertido en un asunto prioritario, debido a que la recuperación de bosques y manglares, según el Reglamento del Parlamento Europeo y el Consejo de la Legislación Europea, es una estrategia vital para conseguir la neutralidad climática.
Se espera que las regulaciones entren en vigencia a partir del segundo semestre del año 2025.
El autor ha contactado a la Agregada Comercial de la Unión Europea en la Delegación Diplomática en Quito y hasta el
cierre de la edición de la presente revista no ha obtenido respuesta•
Diario Oficial de la Unión Europea: Reglamentos por el cual se establece el marco para lograr la neutralidad climática. Junio 2021.
Plan de acción para la perfecta adecuación al Pacto Verde Europeo de las seis cadenas de producción priorizadas en Ecuador. Marzo 2023.
Para más información sobre este artículo, contactar a: lmariduena@cna-ecuador.com
Estimación de parámetros genéticos para el crecimiento y rasgos de resistencia al WSSV en Penaeus vannamei
Autores:
Juan Sui 1,2
Kun Sun3
Jie Kong1,2
Jian Tan1,2
Ping Dai1,2
Jiawang Cao1,2
Kun Luo1,2
Sheng Luan1,2
Qun Xing4
Xianhong Meng1,2,*
1Laboratorio Estatal Clave de Maricultura, Bioreproducción y Recursos Sustentables, Instituto de Investigación Pesquera del Mar Amarillo, Academia China de Ciencias Pesqueras, Qingdao 266071, China; suijuan0313@126.com (J.S.); kongjie@ysfri.ac.cn (J.K.); tannjian@163. com (J.T.); daiping54@163.com (P.D.); caobaoxiang401@126.com (J.C.); luokun@ ysfri.ac.cn (K.L.); luansheng@ysfri.ac.cn (S.L.)
2Laboratorio de Ciencias de Pesca Marina y Procesos de Producción de Alimentos, Centro de Ciencia y Tecnología Marina de Qingdao, Qingdao 266237, China
3Facultad de Pesca y Ciencias de la Vida, Universidad Oceánica de Shanghái, Shanghái 201306, China; sunkun6008@163.com
La producción mundial de camarón blanco del Pacífico (Penaeus vannamei) alcanzó 6.3 millones de toneladas en 2021 [1], consolidando su estatus como una de las especies más destacadas en el cultivo de camarón a nivel mundial. Se distribuye principalmente a lo largo de la costa del Pacífico de América Central y del Sur, y fue introducido en China en 1988. A lo largo de tres décadas de desarrollo, su producción aumentó a 2.09 millones de toneladas en 2022, lo que representa cerca de un tercio de la producción acuícola a nivel mundial [2]. Sin embargo, la escasez de recursos de germoplasma superior limita gravemente el desarrollo de la industria de la producción de camarón en China. Con la expansión continua del cultivo del camarón a escala y la adopción de modelos de cultivo intensivo de alta densidad, los brotes de enfermedades son cada vez más frecuentes durante el proceso de cultivo.
El virus del síndrome de la Mancha Blanca (WSSV, por sus siglas en inglés) ha sido particularmente devastador desde 1992, provocando mortalidades superiores al 95% en un plazo de 7 a 10 días [3–5]. Actualmente no se dispone de medidas eficaces de prevención y control para este patógeno. Contar con selectos reproductores es una potente estrategia para controlar la enfermedad [6,7]. Hasta ahora, el Comité Nacional de Certificación de Variedades Acuáticas de China ha aprobado doce variedades nuevas de P. vannamei, como “Zhongxing No. 1” y “Haixing Nong No. 2”. Sin embargo, el crecimiento sigue siendo el rasgo de mayor interés para la industria [8–10], y 11 de estas 12 variedades presentan características superiores de crecimiento. Existe una demanda urgente en el mercado de reproducir nuevas variedades que combinen tanto la tasa de crecimiento como la resistencia al WSSV.
La estimación precisa de los parámetros genéticos para rasgos objetivos es fundamental para los programas de reproducción, en particular para orientar sobre la formulación de índices de selección, mantenimiento del stock de reproductores y las estrategias de apareamiento. Muchos estudios han documentado los niveles de heredabilidad de la resistencia del camarón al WSSV, que van desde 0.03 a 0.53 [3,11–
14]. Las correlaciones genéticas entre el crecimiento y la resistencia al WSSV en diferentes poblaciones variaron de negativas a ninguna asociación significativa [15–17]. Dados los intrincados antecedentes genéticos de las diferentes poblaciones de P. vannamei, la evaluación de los parámetros genéticos está significativamente influenciada por la estructura de la población, los métodos de infección y los métodos de evaluación. La estimación tradicional de parámetros genéticos se basa principalmente en la mejor predicción lineal insesgada (pBLUP), que se basa en registros de pedigrí. Este método, ampliamente utilizado en la evaluación genética de rasgos económicos en especies acuícolas, estima los valores de reproducción a través de la construcción de una matriz A para tener en cuenta diversos factores influyentes [18]. Sin embargo, debido al riesgo de introducir patógenos en las poblaciones de reproductores, la selección familiar para una mayor resistencia se basa en pruebas de hermanos, donde los candidatos a ser reproductores están protegidos de estar expuestos al patógeno. Este enfoque utiliza solo el 50% de la variación genética aditiva aportada por el componente interfamiliar [19,20].
La mejor predicción lineal genómica imparcial (GBLUP) es muy reconocida por su precisión de selección mejorada, lo que permite la evaluación del muestreo mendeliano en todos los individuos, utilizando marcadores de todo el genoma [21]. Sin embargo, la secuenciación de una gran cantidad de individuos reproductores sigue siendo significativamente costosa. Para abordar esto, se ideó el método de predicción lineal imparcial genómica de un solo paso (ssGBLUP), que incorpora una matriz de relación genómica-pedigrí H mediante la fusión de la matriz A de todo el pedigrí con la matriz de relación genómica G que cubre todos los animales genotipados [22-24]. Durante la construcción de la matriz H, la matriz G se escaló para parecerse a una matriz de relación de pedigrí utilizando las frecuencias de alelos actuales, con coeficientes que van de 0 a 1 en los elementos diagonales y no diagonales que se aplican a 1 [25,26]. La GBLUP se ha utilizado para estimar parámetros genéticos relacionados con la resistencia contra Vibrio
parahaemolyticus en P. vannamei [27] y para el crecimiento y la resistencia a enfermedades en el camarón bananero Fenneropenaeus merguiensis [28]. Estos estudios han demostrado una considerable precisión predictiva en evaluaciones genéticas realizadas utilizando la GBLUP y ssGBLUP. Además, el ssGBLUP se ha empleado para estimar los parámetros genéticos asociados con rasgos de peso corporal en el camarón de agua dulce Macrobrachium rosenbergii [29] y para rasgos de eficiencia alimentaria en P. vannamei [30]. Sin embargo, cuando todos los animales se sometieron al genotipado, la inclusión de información de pedigrí en la matriz H podría disminuir la precisión de las relaciones genómicas realizadas en cierta medida debido a posibles errores en los pedigríes registrados [30].
En 2019, iniciamos un programa de mejoramiento para P. vannamei, importando ocho lotes de germoplasma de alta calidad de todo el mundo. Estos lotes incluyeron cuatro lotes de camarones parentales de rápido crecimiento provenientes de los Estados Unidos y cinco lotes de camarones parentales altamente resistentes provenientes de Ecuador. En este estudio, realizamos pruebas de crecimiento y resistencia al WSSV en linajes de progenie derivados de estas poblaciones importadas. Al combinar data fenotípica, genotípica y de pedigrí, realizamos una estimación de parámetros genéticos asociados con el peso corporal (BW), la longitud total (OL), la longitud corporal (BL), la longitud total (TL) y los rasgos de resistencia al WSSV utilizando las metodologías pBLUP, GBLUP y ssGBLUP. Esta investigación permitió una comparación de varios modelos sobre estimaciones de parámetros genéticos y los valores de reproducción estimados (EBV) o EBV genómicos (GEBV). Este estudio puede ser tomado como referencia para posterior desarrollo de programas de reproducción y como base experimental para la reproducción independiente de variedades de P. vannamei con alta resistencia al WSSV y crecimiento rápido.
Materiales y métodos
Camarones y recopilación de data Camarón
Los camarones experimentales utilizados en este estudio fueron de la población G1
establecida en Bangpu Seed Technology Co., Ltd., ciudad de Weifang, provincia de Shandong, China. Ocho lotes de poblaciones de P. vannamei comercial se introdujeron desde Estados Unidos y Ecuador en 2019, siendo cuatro lotes de grupos altamente resistentes y cuatro lotes de grupos de rápido crecimiento. Se produjeron veinte familias en seis días a través del apareamiento entre camarones machos y hembras seleccionados en una proporción de 1:1 dentro de cada grupo (Tabla 1), que se probaron de estar libres del síndrome de Taura (TSV), Necrosis Hipodérmica y Hematopoyética (IHHNV), enfermedad de Necrosis Hepatopancreática Aguda (AHPND) y Mancha Blanca (WSSV).
Prueba de rasgos de crecimiento
Las larvas de cada familia se cultivaron en condiciones ambientales y densidades estandarizadas [31]. Después de 2 meses de cultivo, el peso corporal promedio alcanzó los 5g. Se eligieron al azar 55 camarones de cada familia y se pesaron, luego se colocaron en baldes nuevos de 150 L. La temperatura del agua se redujo gradualmente de 2-3 °C por día durante tres días hasta alcanzar 20-22 °C, mientras que al mismo tiempo se agregó Vc, glucosa, etc., para el tratamiento de resistencia al estrés a una concentración de 2.5 ppm. Cada individuo fue envuelto en gasa empapada en agua de mar, con solo la cola expuesta. Se etiquetaron individualmente con diferentes combinaciones de colores de “elastómero de implante visible” (VIE) en los sextos segmentos abdominales para distinguir las diferentes familias. Después del marcado, los individuos se sumergieron en agua de mar que contenía 20 ppm de povidona yodada durante 30 s para su desinfección, luego se devolvieron a los baldes de 150 L para su cultivo temporal durante 3 días, con la temperatura aumentando gradualmente a 26-28 °C. Todos los camarones marcados se cultivaron en un estanque rectangular de 50 m2 durante 2 meses. La temperatura del agua de mar se controló entre 26 °C y 28 °C y la salinidad fue de 33‰. Todas las familias fueron alimentadas con pellets comerciales que contenían 42% de proteína, 12% de humedad y 17% de ceniza cruda. La cantidad de alimento se ajustó diariamente en función del crecimiento y condiciones restantes de alimento de los camarones. Al concluir el período de cultivo, se cosechó un total de
GENÉTICA
1040 camarones, lo que resultó en una tasa de supervivencia del 94.55%.
Los camarones fueron transferidos a cinco estanques de 10 m3 de un taller de prueba de rasgos en el Instituto de Investigación Pesquera del Mar Amarillo. Después de aclimatarse durante 3 días, el exceso de agua de los camarones se absorbió utilizando papel absorbente y se pesó utilizando una balanza electrónica con una precisión de 0.01g. Cada camarón fue marcado simultáneamente con un anillo de pedúnculo ocular. Se registró el número de etiqueta del pedúnculo ocular, el sexo, la combinación de colores VIE y el peso corporal de cada camarón. Se utilizó una cámara digital para tomar fotografías de la vista lateral y posterior del camarón (cada camarón fue fotografiado dos veces con una regla colocada al lado). Los rasgos de crecimiento medidos incluyeron longitud total (cm, OL), longitud corporal (cm, BL) y longitud de la cola (cm, TL). Cada rasgo se sometió a tres mediciones, y el valor promedio resultante se consideró como la medición final. El control de calidad del agua, la alimentación y el manejo diario durante el período transitorio se mantuvieron consistentes con los procedimientos estandarizados. Los sitios morfométricos para cada rasgo se muestran en la Figura 1.
Prueba de ensayo del WSSV
Después de la prueba de crecimiento, los camarones se aclimataron en estanques de 10 m3 durante una semana. Antes del ensayo del WSSV, la edad de los camarones era de 180-185 días post-eclosión (dph), con un promedio de 183 dph y un peso promedio de 16g. Los tejidos musculares de camarones infectados con WSSV se utilizaron para preparar cebo envenenado. La cuantificación de la carga del WSSV en el músculo se realizó mediante PCR en tiempo real (sistema de PCR en tiempo real ABI 7500). Los tejidos musculares se picaron y se mezclaron con tejidos musculares no infectados y colorante rojo comestible para hacer cebo con una concentración de 104 copias del genoma del WSSV por miligramo (mg). Después de la inanición durante 24h para asegurar la evacuación gastrointestinal, cada camarón fue alimentado con 10 mg de cebo. Luego, los camarones fueron devueltos al estanque y observados cada hora. Los
camarones recibieron dietas formuladas, divididas en cuatro raciones diarias iguales, que constituyeron una asignación diaria total del 5% de la biomasa en peso húmedo, y se ajustaron diariamente. Se retiraron los individuos muertos. Se registraron las horas posteriores a la infección (h, HPI), la combinación de colores y el número de etiqueta del pedúnculo ocular de todos los individuos de prueba. La prueba de ensayo duró 316h, hasta que no se produjeron muertes de camarones en un período de 24h, momento en el que concluyó el experimento. Para cada familia, se prepararon entre 8 y 10 individuos adicionales, que sirvieron como grupos control para el ensayo. En total, se recolectaron 1033 individuos para la adquisición de data de crecimiento y resistencia al WSSV (Tabla 1). Tras el
examen, todos los individuos muertos exhibieron características típicas del síndrome de la Mancha Blanca (WSS), y cinco camarones seleccionados al azar se sometieron a pruebas de PCR en tiempo real para confirmar la infección por el virus. Se recolectaron tejidos musculares de todos los individuos de prueba y se almacenaron a -80 °C.
Genotipado
Según el experimento preliminar, se encontró que los grupos R2, R3 y R4 tenían un parentesco cercano. Por lo tanto, se seleccionaron 50 individuos de estas tres poblaciones, mientras que entre 30 y 70 individuos se eligieron al azar de las poblaciones restantes, lo que dio como resultado un total de 300 individuos para
Figura 1. Diagrama esquemático de la posición morfométrica de P. vannamei. (a) muestra una vista dorsal; (b) muestra una vista lateral. OL: longitud total; BL: longitud del cuerpo; TL: longitud de la cola.
Nota: R: grupo altamente resistente; G: grupo de rápido crecimiento.
Tabla 1. Número de familias e individuos analizados por lote y grupo.
la secuenciación del genoma completo. El ADN genómico se extrajo del tejido muscular utilizando el kit de ADN para animales marinos TIANamp (Tiangen, Pekín, China). La pureza se verificó utilizando NanoDrop 1000 (Thermo Scientific, Waltham, MA, EE.UU.), la integridad se verificó utilizando una electroforesis en gel de agarosa al 1% y la cantidad se verificó utilizando Qubit (Thermo Scientific, MA, EE.UU.). La resecuenciación del genoma completo se realizó utilizando una plataforma Illumina HiSeq2500 con una profundidad de secuenciación de 10x y lecturas de extremos emparejados (PE) de 125 pb (Beijing Berry Genomics Biotechnology Co., Ltd., Pekín, China). Se eliminaron las lecturas sin procesar que contenían más del 10% de bases ambiguas (N), que mostraban una calidad deficiente (donde los nucleótidos con un valor de calidad Q ˂ 3 representan más del 50% de la lectura) o que albergaban adaptadores. Las lecturas limpias de cada muestra se alinearon con el genoma de P. vannamei (ASM378908v1) utilizando el Burrows–Wheeler Aligner 0.7 [32] con parámetros establecidos como “mem 4-k 32-M” para detectar SNPs. La llamada de variantes se ejecutó a través de UnifiedGenotyper de GATK. Los SNPs se sometieron a filtración empleando Variant Filtration de GATK con criterios apropiados (-Window 4, -filter “QD < 2.0||FS > 60.0||MQ < 40.0”, -G_filter “GQ < 20”), y se descartaron aquellos que demostraron distorsión de segregación o errores de secuenciación. Finalmente, se identificaron 103,745,370 loci. El control de calidad del genotipo se realizó utilizando el software Plink versión 1.9 [33], adhiriéndose a los criterios de exclusión posteriores: tasa de identificación de SNP < 90%, frecuencia de alelos menores < 0.05, equilibrio de Hardy-Weinberg con valor p < 1 x 10−6 y exclusión de camarones con una tasa de identificación de genotipo < 90%. Siguiendo estos pasos, se conservaron 18,137,179 SNP y 293 animales para análisis posteriores.
Análisis de data
Construcción de data y matrices
Se examinó la distribución normal de la data fenotípica y se eliminaron los valores atípicos, lo que dio como resultado un dataset final de 1017 individuos para el análisis estadístico. La evaluación genética para el crecimiento y la resistencia al WSSV
se realizó utilizando los métodos pBLUP, GBLUP y ssGBLUP. El método pBLUP utilizó una matriz A de parentesco basada en el registro de pedigrí, que incorporaba un pedigrí de dos generaciones que constaba de 293 individuos y 34 padres. Se supuso que los individuos parentales no estaban relacionados al pedigrí. El método GBLUP implicó la utilización de una matriz G que contenía información genómica de 293 individuos genotipados. El método ssGBLUP implicó la utilización de una matriz H que integraba el pedigrí de 1017 individuos y la información genómica de un conjunto común de 293 individuos genotipados.
La matriz A se construyó utilizando el paquete ASReml-R V4.1 [34]. La matriz G basada en la información del genotipo y la matriz H basada en la información combinada del pedigrí, se construyeron utilizando el programa preGSf90 en BLUPF90 1.70 [35]. A la matriz A se le asignó coeficientes de ponderación del 5%, mientras que a la matriz G se le asignó un coeficiente de ponderación del 95%.
Estimaciones de heredabilidad y componentes de varianza
Los componentes de varianza para los rasgos de crecimiento y resistencia se estimaron utilizando el paquete ASReml-R V4 [34] empleando el método de máxima verosimilitud restringida por información promedio. Los modelos lineales mixtos para cada rasgo se proporcionan de la siguiente manera:
Y ijk = µ + bwi + sexj + ai + fk + eijk (1)
Yil = µ + bAgei + Pondl + ai + eil (2) donde yijk es el valor de crecimiento observado del i-ésimo individuo, yil es el tiempo de supervivencia observado del i-ésimo individuo, b es el coeficiente de regresión, wi y Agei son las covariables de peso corporal medio y edad en el momento del etiquetado familiar, respectivamente, sexj es el efecto fijo del j-ésimo género, Pondl son los efectos fijos del l-ésimo pond, ai son los efectos genéticos aditivos del i-ésimo individuo, a~(0, Mσa2), donde M es la matriz A, G o H entre individuos, fk es el efecto aleatorio común a la k-ésima familia de hermanos completos, f ~(0, Iσ2 c), donde I es una matriz de identidad, eijk y eil son el efecto residual aleatorio del i-ésimo individuo, e~(0, Iσ2 e).
La heredabilidad se calculó de la siguiente manera: h2 = σ2 a/σ2 p (3) donde h2 es la heredabilidad, σ2 p =
+
2 e según el modelo utilizado.
1.2.3. Prueba Z
La prueba Z se empleó para evaluar la significancia de las diferencias en la heredabilidad de uno o cero [36], de la siguiente manera:
z = xi – xj (4)
donde xi y xj son las estimaciones de heredabilidad, σi y σj son sus respectivos errores estándar. Al probar si una estimación era significativamente diferente de uno, tanto xj como σj se establecieron en uno y cero; al probar si una estimación era significativamente diferente de cero, tanto xj y σj se establecieron en cero.
Correlación genética
Se empleó un modelo bivariado para estimar las correlaciones fenotípicas y genéticas entre diferentes rasgos. El modelo de análisis bivariado fue idéntico al modelo univariante. La matriz A, la matriz G y la matriz H se utilizaron en el modelo para construir las matrices de correlación genética aditiva respectivas.
Precisión de predicción y sesgo
Para comparar la precisión de predicción de GBLUP y ssGBLUP con pBLUP, se empleó un método de validación cruzada de cinco veces y diez veces. Aproximadamente, 293 individuos fueron secuenciados de manera uniforme y se dividieron aleatoriamente en cinco grupos. Se seleccionó aleatoriamente un grupo y su data fenotípica se enmascaró como un conjunto de validación, mientras que los cuatro grupos restantes y los fenotipos de los individuos no tipificados se utilizaron como un conjunto de entrenamiento para predecir los valores de reproducción estimados para la población de validación. Se realizaron diez repeticiones de validación cruzada, con precisión predictiva definida como el promedio del coeficiente de correlación de Pearson entre los valores fenotípicos del conjunto de validación y EBV/GEBV, y el sesgo predictivo como el coeficiente de regresión de los valores
GENÉTICA
fenotípicos contra EBV(GEBV).
Un sesgo de 1 indica que EBV (GEBV) teóricamente proporciona una estimación no sesgada del valor genético verdadero (BV), mientras que un sesgo menor o mayor que 1 sugiere subestimación o sobreestimación de EBV (GEBV), respectivamente.
Resultados
Estadísticas descriptivas
Las estadísticas descriptivas de BW, OL, BL, TL y HPI se muestran en la Tabla 2. El valor promedio de BW fue 16.21 ± 5.22 g, con un coeficiente de variación de 32.20%. Los coeficientes de variación para otros rasgos de crecimiento fueron más bajos que para el BW, oscilando entre 11.55% y 12.48%. El valor promedio del índice HPI fue de 91.53 ± 5.22 h, con el coeficiente de variación más alto entre los caracteres de prueba, alcanzando el 59.42 %.
Análisis de correlación genética molecular Los mapas de calor de coeficientes de parentesco por pares entre individuos para las matrices A, G y H se muestran en la Figura 2. Solo las relaciones de hermanos completos y medios hermanos entre individuos se reflejaron en función del parentesco de pedigrí, mientras que todos los individuos tenían coeficientes de parentesco específicos entre sí en función de la información del genotipo. La correlación de los coeficientes de parentesco fue de 0.614 entre los elementos triangulares inferiores de las matrices A y G, de 0.649 entre las matrices A y H, y de 0.999 entre las matrices G y H.
2. Estadísticas descriptivas de los valores fenotípicos de crecimiento y resistencia al WSSV.
Nota: BW: peso corporal, OL: longitud total, TL: longitud de la cola, BL: longitud corporal, HPI: horas posteriores a la infección, SD: desviación estándar, CV: coeficiente de variación.
Las correspondencias entre las matrices G y H con la matriz A se muestran en la Figura 3. Cuando los coeficientes de parentesco por pares entre individuos en la matriz A (eje x) fueron 0, el parentesco genómico entre estos individuos fue mayor en la matriz G y la matriz H (eje y), lo que indica que el parentesco entre individuos no fue lo suficientemente preciso basándose únicamente en la información del pedigrí. La misma situación se encontró en el parentesco genómico de 0.125, 0.25 y 0.5 en la matriz A. Esto demuestra además que las matrices G y H, construidas con base en marcadores moleculares, pueden describir con mayor precisión el parentesco entre individuos en comparación con la matriz A Figura 2. Mapa de calor de los coeficientes de parentesco entre pares para la
matriz A (a), G (b) y H (c).
Tabla
basada en un pedigrí de dos generaciones.
Heredabilidad
Los componentes de varianza y las estimaciones de heredabilidad del BW, OL, BL, TL y HPI de camarones de 6 meses de edad con base en las matrices A, G y H se muestran en la Tabla 3. De acuerdo con los criterios de clasificación de Cardellino et al. (1987), los valores de heredabilidad específicos se pueden dividir en cuatro niveles: heredabilidad baja (0.050.15), heredabilidad media (0.20-0.40), heredabilidad alta (0.45-0.60) y heredabilidad muy alta (>0.65). Basados en la matriz A, las estimaciones de heredabilidad de los rasgos relacionados con el crecimiento (BW, OL, BL y TL) fueron de 0.399 ± 0.158 a 0.810 ± 0.103, lo que indica niveles medios a altos. La heredabilidad estimada de HPI fue 0.075 ± 0.074, sin mostrar diferencias significativas con respecto a 0 (p > 0.05). Basados en la matriz G, las estimaciones de heredabilidad del BW, OL, BL y TL disminuyeron en un 12.10%, 57.19%, 39.28% y 20.30%, respectivamente. La heredabilidad estimada de la OL no se desvió significativamente de 0, mientras que otros rasgos exhibieron niveles moderados. La heredabilidad de HPI aumentó en un 17.33%, pero aun así no hubo diferencias significativas con respecto a 0 (p > 0.05). Basándonos en la matriz H, las heredabilidades de la OL, BL y TL no difirieron significativamente de 0 (p > 0.05). La heredabilidad del BW disminuyó en un 44.07% y un 36.38% en comparación con las matrices A y G, respectivamente, lo que indica una heredabilidad moderada. La del HPI aumentó en un 164% y un 125% en comparación con las matrices A y G, respectivamente, lo que también indica una heredabilidad moderada. Para los rasgos relacionados con el crecimiento, la heredabilidad estimada disminuyó de las matrices A y G a H, pero aumentó para los rasgos de resistencia al WSSV.
Correlación genética
Las correlaciones genéticas y fenotípicas entre los caracteres estimados utilizando diferentes métodos se muestran en la Tabla 4. Las correlaciones genéticas entre el BW, OL, BL y TL basadas en los métodos pBLUP, GBLUP y ssGBLUP variaron de 0.767 a 0.980, 0.920 a 0.993 y 0.912 a 0.999, respectivamente. Las correlaciones
Parentesco entre pares en la matriz G y /h
Parentesco entre pares en la matriz A
Parentesco entre pares en la matriz G y /h
Parentesco entre pares en la matriz A
Figura 3. Relaciones correspondientes en la matriz G, H y la matriz A de los individuos secuenciados. (a) la relación correspondiente entre la matriz G y A; (b) la relación correspondiente entre la matriz H y A.
Tabla 3. Componentes de varianza y heredabilidad del BW, OL, BL, TL y HPI de camarones de 6 meses de edad basadas en diferentes métodos.
Nota: BW: peso corporal, OL: longitud total, TL: longitud de la cola, BL: longitud corporal, HPI: horas posteriores a la infección, σ2 : varianza fenotípica, σ2 : varianza genética aditiva, C: efectos ambientales comunes, h2: heredabilidad. a : la estimación no es significativamente diferente de 1 (p < 0.05).b: la estimación no es significativamente diferente de 0 (p < 0.05).
fenotípicas variaron de 0.629 a 0.959, 0.548 a 0.988 y 0.684 a 0.991, lo que indica niveles medios a altos de correlaciones positivas. Las correlaciones genéticas de HPI con BW, OL, BL y TL basadas en pBLUP, GBLUP y ssGBLUP variaron de −0.198 a −0.081, −0.173 a −0.096 y −0.057 a −0.019, respectivamente. Las correlaciones fenotípicas variaron de −0.219 a −0.162, −0.191 a −0.115 y −0.443 a −0.397, respectivamente, lo que indica niveles bajos a moderados de correlaciones negativas. Se observaron disparidades menores en las correlaciones genéticas y fenotípicas de estos rasgos entre los diferentes métodos.
Precisión de predicción y análisis de sesgo Se analizó la precisión de predicción y el
sesgo de los tres métodos para el BW, OL, BL, TL y HPI mediante una validación cruzada de cinco y diez veces. Los resultados se muestran en la Tabla 5. En comparación con pBLUP, GBLUP mostró una mayor precisión de predicción para el BW, OL, BL, TL y HPI en un 4.77%, 21.93%, 19.73%, 19.34% y 63.44%, respectivamente, mientras que redujo el sesgo en un 11.88%, 18.49%, 15.94%, 15.14% y 26.48%, respectivamente. Además, el ssGBLUP mostró mayores mejoras en la precisión de predicción, con aumentos del 10.07%, 25.44%, 25.72%, 19.34% y 122.58%, y mayores reducciones en el sesgo en un 23.62%, 26.32%, 18.71%, 36.10% y 50.12%, respectivamente. Al comparar GBLUP con ssGBLUP, este último demostró mejoras adicionales en la precisión
GENÉTICA
de predicción en un 5.06%, 5.00%, 2.88%, 0% y 36.18%, respectivamente. Además, ssGBLUP mostró mayores reducciones en el sesgo de predicción para todos los rasgos en comparación con GBLUP. Entre los tres métodos, ssGBLUP mostró consistentemente una precisión de predicción superior y un sesgo menor, seguido de GBLUP, con pBLUP mostrando el rendimiento más bajo en ambas métricas.
Discusión
En la reproducción selectiva de P. vannamei, los rasgos de crecimiento como el BL y BW son de mayor interés, atrayendo la atención constante de los productores. Por lo tanto, la estimación precisa de los parámetros genéticos se vuelve fundamental para estimar los valores de reproducción, formular programas de reproducción y dilucidar mecanismos genéticos subyacentes a estos rasgos [37]. Sin embargo, el enfoque predominante en estos estudios ha sido la utilización del método pBLUP, basándose en información de pedigrí. Muchas investigaciones, tanto a nivel nacional como internacional, han profundizado en la estimación de parámetros genéticos para rasgos de crecimiento en P. vannamei. Por ejemplo, Xu et al. [38] analizaron data fenotípica de 3240 individuos en 18 familias de medios hermanos y 54 familias de hermanos completos, revelando valores de heredabilidad de 0.460, 0.392 y 0.303 para el BW, OL y BL, respectivamente. Luan et al. [39] ampliaron esto con data fenotípica de 19,199 individuos en 130 familias, estimando la heredabilidad del peso de la cosecha entre 0.19 y 0.43. Sui et al. [31] contribuyeron además con evaluaciones de heredabilidad del peso de cosecha, basados en data fenotípica y de pedigrí de dos generaciones de 24,072 individuos, que van desde 0.278 a 0.423. Hernández-Ruíz et al. [40] reportaron una heredabilidad del peso de cosecha de 0.24, basados en data de pedigrí de 2002 a 2019 y la data fenotípica de 12,440 individuos de 160 familias. En este estudio, el método pBLUP se combinó con la data fenotípica de 1017 individuos de 20 familias para evaluar las heredabilidades de los rasgos de BW, OL, BL y TL en P. vannamei, que se encontraron en 0.810 ± 0.103, 0.654 ± 0.201, 0.690 ± 0.205 y 0.399 ± 0.158, respectivamente. La heredabilidad del BW estimada, basada en
4. Correlaciones genéticas y fenotípicas del BW, OL, BL, TL y HPI basadas en diferentes métodos.
Nota: el valor en el triángulo inferior es la correlación fenotípica entre los rasgos, y el valor en el triángulo superior es la correlación genética entre los rasgos. OL: longitud total, TL: longitud de la cola, BL: longitud del cuerpo, BW: peso corporal, HPI: horas post infección.
Tabla 5. Precisión de predicción y sesgo de cada rasgo bajo diferentes métodos.
Notas: BW: peso corporal; OL: longitud total; TL: longitud de la cola; BL: longitud del cuerpo. HPI: horas post infección.
la matriz A, mostró un nivel considerable que puede estar significativamente influenciado por el conjunto de datos (estructura de la población) y el ambiente de prueba. El conjunto de datos es relativamente pequeño, con solo 1014 individuos finalmente incluidos en el análisis. La profundidad del pedigrí de los individuos de prueba es de solo dos generaciones, y los individuos parentales se obtienen de forma extensiva y se consideran no relacionados. Las variaciones entre familias se atribuyen principalmente a diferencias genéticas. Durante el período de cultivo comunitario de dos meses, la densidad de población es relativamente baja (22 ind/m2), lo que permite diferencias significativas en el crecimiento individual, con un alto coeficiente de variación en el peso individual (32.2%). Estos factores han impactado significativamente los resultados de los parámetros genéticos. Se podría realizar un estudio adicional aumentando el número de individuos de prueba para obtener un parámetro más preciso.
La integración de la información genotípica mediante el método GBLUP produjo distintos grados de disminución de las heredabilidades para todos los rasgos, especialmente fue evidente en los rasgos de tamaño corporal como OL, BL y TL, que disminuyeron moderadamente a niveles altos, oscilando entre 0.280 ± 0.154 y 0.419 ± 0.156. Estas fluctuaciones en las heredabilidades podrían atribuirse a factores como la estructura de la población, el número limitado de miembros de la familia, la documentación del pedigrí y la data fenotípica. La evaluación posterior con el método ssGBLUP, que combina la información del genotipo y el pedigrí, afectó aún más a las heredabilidades, en particular en el BW, mientras que los otros rasgos mostraron cambios mínimos, lo que dio como resultado valores de heredabilidad que oscilaron entre 0.195 y 0.453, lo que consistente con hallazgos anteriores. En particular, los rasgos de tamaño corporal mostraron una mayor precisión de predicción en comparación con el BW. El análisis comparativo reveló que tanto el método
Tabla
GBLUP como el ssGBLUP superaron al pBLUP en términos de precisión de predicción y reducción de sesgo para la evaluación de parámetros genéticos en P. vannamei. Además, la combinación de información de pedigrí y genotipo en el ssGBLUP demostró una precisión de predicción superior y una reducción de sesgo en comparación con el uso de información de genotipo sola. Específicamente, los rasgos OL, BL y BW evaluados a través del ssGBLUP mostraron una precisión 2.88%, 5.00% y 5.06% mayor, con un sesgo 9.61%, 3.29% y 13.32% menor, respectivamente, en comparación con GBLUP. Aunque los estudios similares sobre GBLUP y ssGBLUP en P. vannamei siguen siendo escasos, su aplicación exitosa en otras especies subraya su eficacia en la evaluación de parámetros genéticos para rasgos de crecimiento. Aunque la precisión del rasgo TL se mantuvo sin cambios, se observó una notable reducción del 24.69% en el sesgo.
El virus del síndrome de la Mancha Blanca (WSSV) siempre ha sido un foco importante de investigación sobre las enfermedades en P. vannamei, y la reproducción selectiva para rasgos de resistencia al WSSV, es el siguiente paso para los productores. En comparación con los rasgos de crecimiento, menos estudios han reportado sobre la evaluación de los parámetros genéticos para la resistencia al WSSV en P. vannamei Por ejemplo, Gitterle et al. [15], CaballeroZamora et al. [16] y Campos-Montes et al [12] adoptaron el método BLUP tradicional para evaluar la heredabilidad de los rasgos de supervivencia del WSSV en P. vannamei y encontraron estimaciones de heredabilidad bajas de 0.03 a 0.20 para los rasgos de supervivencia del WSSV. En este estudio, la heredabilidad del rasgo de resistencia al WSSV en P. vannamei evaluada con base en el método pBLUP no fue significativamente diferente de 0 (p > 0.05). Además, tuvo una precisión de predicción de 0.186 ± 0.058 y un sesgo de 1.620 ± 0.215, lo que sugiere una precisión de predicción relativamente baja y un sesgo relativamente alto en comparación con los rasgos de crecimiento. Cuando se combinó la información del genotipo para la evaluación basada en el método GBLUP, la precisión de la predicción aumentó en un 63.44%, el sesgo disminuyó en un 26.48% y la heredabilidad resultante fue de 0.088
± 0.081. Además, cuando se combinó la información del pedigrí y el genotipo para la evaluación basada en el método ssGBLUP, la precisión de la predicción aumentó en un 122.58%, el sesgo disminuyó en un 50.12% y la heredabilidad resultante fue de 0.198 ± 0.066. Campos-Montes et al. [12] documentaron las heredabilidades relativas al momento de muerte posterior a la infección por el WSSV y la supervivencia binaria utilizando GBLUP y ssGBLUP, que oscilaron entre 0.094 y 0.095 y entre 0.085 y 0.105, respectivamente, empleando un panel comercial de 50 K SNP en postlarvas. Las discrepancias en las estimaciones de heredabilidad pueden deberse a diversos factores, incluyendo la población experimental, la edad de la población y los métodos de infección. Los estudios antes mencionados subrayan colectivamente la eficacia de incorporar información genómica para mejorar la precisión de las estimaciones de heredabilidad. Se prevé que la integración de data genómica refine la estimación de los componentes de varianza al proporcionar una evaluación más precisa de las relaciones de parentesco, facilitando así una demarcación más clara entre la varianza genética aditiva y residual [30,41]. También se observaron resultados similares en evaluaciones genómicas de rasgos de resistencia a enfermedades en otras especies. Robledo et al. [42] utilizaron GBLUP para evaluar la resistencia a la enfermedad amebiana de las branquias en el salmón del Atlántico, y sus resultados mostraron una mayor precisión para GBLUP (0.62) que para pBLUP (0.51). Además, se encontró que el método ssGBLUP era superior al método GBLUP. Varios informes han explorado el uso de GBLUP y ssGBLUP para evaluar los parámetros genéticos de los rasgos de resistencia a las enfermedades en animales acuáticos. Sukhavachana et al. [43] también utilizaron el método GBLUP para estudiar la resistencia de Streptococcus agalactiae en la tilapia roja híbrida, con una precisión de GBLUP de 0.25, en comparación con pBLUP de 0.15. Tsai et al. [44] evaluaron la resistencia a los piojos de mar en el salmón del Atlántico bajo diferentes densidades de SNP (0.5 –33 k). Vallejo et al. [45] utilizaron el método ssGBLUP para evaluar los parámetros genéticos de resistencia a la enfermedad bacteriana del agua fría en la trucha arcoíris, mejorando la precisión de la predicción en
un 83.3 – 85.3%. Además, Lu et al. [36] utilizaron el método ssGBLUP para evaluar los parámetros genéticos de la resistencia a la Edwardsielosis en el lenguado japonés, logrando una precisión de predicción final de 0.65 para ssGBLUP, en comparación con 0.54 para pBLUP. Yoshida et al. [46] evaluaron los parámetros genéticos de resistencia al síndrome rickettsial de los salmónidos en la trucha arcoíris bajo diferentes densidades de marcadores (0.5 – 27k), mostrando una precisión de predicción mejorada en un 21–37% utilizando el método GBLUP y en un 23–40% utilizando el método ssGBLUP. Nuestros resultados coinciden con estos hallazgos, lo que indica que los métodos GBLUP y ssGBLUP producen una mayor precisión de predicción que el método pBLUP para evaluar los parámetros genéticos de los rasgos de resistencia a las enfermedades. Un problema que vale la pena señalar es que la densidad de marcadores y la cantidad de individuos secuenciados pueden ser factores críticos que afecten la precisión de las evaluaciones. Sin embargo, una densidad de marcadores excesivamente alta y una gran cantidad de individuos también pueden aumentar los costos de secuenciación. La elección de una estrategia de secuenciación adecuada es un asunto que requiere consideración.
Se ha demostrado que existe una fuerte correlación positiva entre los rasgos de crecimiento en P. vannamei (rg > 0.95), lo que sugiere un vínculo estrecho entre los genes que rigen el peso corporal (BW) y la longitud corporal (BL). En consecuencia, la selección simultánea del BW junto con el BL, puede producir resultados más favorables [47]. En este estudio, estimamos las correlaciones genéticas y fenotípicas entre los rasgos BL, BW, OL y TL utilizando varios modelos de evaluación que incorporan información genotípica. Nuestros hallazgos se hicieron eco de estudios previos, indicando efectos mínimos de los modelos en las correlaciones genéticas y los fenotipos. Además, investigamos las correlaciones genéticas y fenotípicas entre el crecimiento y los rasgos de resistencia al WSSV utilizando diferentes modelos. Nuestros resultados revelaron que los modelos tuvieron un impacto relativamente menor, con el crecimiento y los rasgos del WSSV mostrando correlaciones genéticas que van desde 0.198 a 0.019 y correlaciones
GENÉTICA
fenotípicas de 0.443 a 0.115. Los estudios de correlación genética entre el crecimiento y la resistencia a enfermedades virales a menudo muestran asociaciones negativas en animales acuáticos. En el camarón, Argue et al. [48] reportaron una correlación genética de 0.46 entre el crecimiento y la resistencia al síndrome de Taura. Fu et al. [17] encontraron correlaciones genéticas que van desde 0.034 a 0.573 entre el crecimiento y la resistencia al WSSV. En peces, Bangera et al. [49] reportaron correlaciones genéticas de 0.25 y 0.032 entre la tasa de crecimiento y la Vibriosis y la Necrosis Nerviosa Viral, respectivamente, y ambas no fueron significativamente diferentes de cero. Cock et al. [50] propusieron que los genes de resistencia pueden encontrar presiones selectivas antagónicas, lo que lleva a una frecuencia de equilibrio que no alcanza la fijación completa. Dada la prolífica capacidad reproductiva del camarón, la recolección extensiva de poblaciones diversas permite
la construcción de numerosas familias para pruebas de resistencia, lo que facilita la identificación de fuentes de resistencia incluso a bajas frecuencias. De todos modos, es imperativo incorporar coeficientes de ponderación económica o asignaciones porcentuales tanto para los rasgos de crecimiento como para los rasgos de resistencia al WSSV en los programas de reproducción de P. vannamei. El desarrollo de un índice de selección compuesto de múltiples rasgos puede facilitar la evaluación y selección de reproductores de alta calidad.
Conclusiones
Este estudio demostró que la heredabilidad de rasgos relacionados con el crecimiento obtenidos de GBLUP y ssGBLUP fue menor que la obtenida del pBLUP, mientras que la heredabilidad de la resistencia al WSSV fue mayor que la obtenida del pBLUP. La precisión predictiva de los rasgos de crecimiento y la resistencia al WSSV obtenida con los
métodos GBLUP y ssGBLUP fue mayor que la del método pBLUP, con menores sesgos predictivos que el pBLUP. Las correlaciones genéticas del crecimiento y la resistencia al WSSV obtenidas con los tres métodos fueron negativas bajas. Los hallazgos de este estudio ofrecen metodologías y conocimientos valiosos para evaluar los parámetros genéticos en P. vannamei, sentando las bases para futuros programas de mejoramiento y desarrollo de variedades•
Para más información sobre este artículo, contactar a: mengxianhong@ysfri.ac.cn
ESTADÍSTICAS ÍNDICE
Edición 161 - Octubre 2024
81 87 90 Exportaciones de camarón Reporte de mercado de EE. UU. Reporte de mercado de China
CAMARÓN Estadísticas de Comercio Exterior
EVOLUCIÓN DE EXPORTACIONES 2010 - 2023
La gráfica presentada muestra un crecimiento constante en el volumen y valor de las exportaciones de camarón, con un incremento notable durante los últimos cinco años. Este desempeño destaca el fortalecimiento del sector camaronero, evidenciando una mejora en su capacidad productiva y una consolidación en el mercado global.
La tendencia ascendente sugiere que la industria ha logrado adaptarse a las demandas internacionales, manteniendo su competitividad y contribuyendo de manera significativa a la economía exportadora del país.
Las exportaciones de camarón han mostrado un incremento constante desde 2020 hasta 2024 en casi todos los meses, con algunos meses alcanzando picos considerables.
Se observa que en el año 2020 el volumen de exportación mensual es significativamente más bajo en comparación con los años siguientes debido a los efectos de la pandemia de COVID-19. Sin embargo, para los años siguientes, se evidencia un aumento significativo en las exportaciones, reflejando una clara recuperación del sector.
En mayo de 2024, Ecuador alcanzó un récord histórico al exportar 275 millones de libras de camarón, un hito impulsado por la optimización de su producción para obtener camarones de tallas más grandes. Este logro refleja la capacidad del sector para adaptarse a las tendencias del mercado internacional, maximizando tanto la calidad como el valor del producto exportado. No obstante, para los meses de junio y julio de este mismo año, se produjo una caída importante debido, principalmente, a restricciones en los mercados internacionales.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
PARTICIPACIÓN POR MERCADOS DESTINO JULIO 2023 VS JULIO 2024
El siguiente gráfico muestra la participación porcentual de las exportaciones de camarón por destino en libras durante julio de 2023 y julio de 2024. Como se puede observar, aunque China sigue siendo el principal mercado, se observa una ligera disminución en su participación de 2.17 puntos, e igualmente Europa, cuya disminución es de 2.57 puntos. Por el contrario, Estados Unidos presenta un aumento significativo en su participación, incrementando 3.43 puntos porcentuales.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
PRINCIPALES PAÍSES DESTINOS (MILLONES DE LIBRAS)
ENERO - JULIO (2023 VS 2024)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
La tendencia general muestra crecimiento en todos los países/regiones, excepto en China que, aun siendo el principal destino de exportación, presenta una disminución significativa en el volumen exportado. Por otro lado, se puede observar un crecimiento importante en Estados Unidos, país que experimentó un aumento del 18%, pasando de 251 a 295 millones de libras entre el período Enero - Julio del 2024 versus el mismo período del 2023. En cuanto al volumen exportado al resto del mundo, cabe resaltar el notable esfuerzo del sector camaronero por diversificar sus mercados tras la pandemia COVID-19.
EVOLUCIÓN DE EXPORTACIONES (MILLONES DE LIBRAS)
CHINA Y EE. UU. ENERO - JULIO 2024
El gráfico presenta la evolución de las exportaciones de camarón ecuatoriano hacia China y Estados Unidos, representando más del 70% del volumen total exportado. En el caso de China, se observa una notable volatilidad a lo largo del período, notándose una caída pronunciada en marzo, seguida de una rápida recuperación en mayo. Posteriormente, el volumen de exportaciones vuelve a disminuir en junio y julio. Por otro lado, las exportaciones hacia Estados Unidos presentan un comportamiento más estable, con fluctuaciones menos abruptas. Durante el primer trimestre de 2024, se aprecia un crecimiento sostenido en los envíos hacia este mercado. Sin embargo, en junio ocurre una importante caída en el volumen exportado, antes de estabilizarse nuevamente en julio.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
En marzo, las exportaciones a China tocaron su punto más bajo, mientras que las dirigidas a Estados Unidos alcanzaron su nivel máximo. Esta tendencia opuesta sugiere no solo ajustes estratégicos en la asignación de los envíos, sino también variaciones en la demanda de estos mercados fundamentales.
Importación de camarón de China
Autor: Erwin Termaat
Kontali
Shrimp
sander@kontali.no www.shrimp.kontali.no
Los volúmenes de importación a China en agosto se mantuvieron estables en comparación con julio, en 86,970 toneladas. Esto significa que los déficits interanuales constantes que se han estado mostrando desde febrero se están desacelerando relativamente hasta un -2% para este mes, una tendencia que predijimos en nuestro reporte más reciente.
El precio de importación aumentó $0.05 a $4.97/kg. Aunque esto sigue siendo $0.33 más bajo que el precio del año pasado para el mismo mes, la tendencia al alza puede continuar el próximo mes. Esto se debe al aumento de la demanda durante las vacaciones en China, que es importante para el consumo de camarón. El festival de Medio Otoño, que tuvo lugar a mediados de septiembre, probablemente tuvo algo que ver con el aumento de precios de agosto, y el aumento de la demanda por el feriado del Día Nacional el 1 de octubre podría llevar a otro repunte en los precios de importación de septiembre. El valor de las importaciones alcanzó los $432 millones en agosto, totalizando $3.2 mil millones en lo que va de 2024.
El volumen de importación de vannamei en agosto disminuyó un 3% hasta 98,690 toneladas de LSE. Esto representó un aumento del 7% en comparación con el año anterior. La participación de las importaciones de vannamei de Ecuador alcanzó el punto más bajo en 2024, pero aún representó el 63%, o 62.210 toneladas de LSE. Según nuestro modelo de mercado Kontali, se espera que el volumen de importación de Ecuador en septiembre aumente aún más en otro 19%. La menor participación del camarón ecuatoriano en agosto se compensó principalmente con las importaciones de vannamei de India, que aumentaron un 18% con respecto a julio y un 26% más en comparación con el año anterior (YoY), hasta las 24,290 toneladas de LSE. Esto representó el 25% de las importaciones totales del mes. Las importaciones procedentes de Vietnam aumentaron una vez más hasta las 8,080 toneladas de LSE, lo que fue un 31% más que en agosto de 2023. Esta es una tendencia que esperamos que continúe también en septiembre.
El volumen de importación de monodon de China experimentó otra disminución en agosto, un 13% menos, hasta 1,050 toneladas de LSE. Vietnam siguió siendo el mayor proveedor de monodon a China, con 684 toneladas de LSE suministradas, mientras que las importaciones de India disminuyeron un 37% hasta 216 toneladas. Parece poco probable que los volúmenes de importación de monodon aumenten significativamente durante los próximos meses.
Muchos productores respondieron a la creciente demanda vendiendo sus productos antes del Festival de Medio Otoño, lo que provocó un breve aumento de los precios en origen. Sin embargo, los precios han vuelto a tender a la baja desde que terminó el festival. Anticipamos un ligero repunte de precios para finales de mes a medida que se intensifican los preparativos para el feriado del Día Nacional.
Además del camarón local, el camarón importado congelado se está volviendo cada vez más popular durante estos períodos festivos, gracias en parte a la conveniencia de las plataformas de comercio electrónico como TikTok, Kuaishou y Taobao. Esta mayor demanda se ha reflejado en los volúmenes de importación, que se han acercado
Importaciones de China en 2022, 2023 y 2024 (volúmenes y peso promedio/kg)
Importaciones de Vannamei al mercado chino (2022, 2023 y 2024)
a los niveles interanuales en julio y agosto en comparación con los meses anteriores. Se espera que esta estabilización continúe en el período previo a las celebraciones de fin de año, que suelen generar un pequeño aumento de la demanda, en particular cuando también comienzan los preparativos para el Año Nuevo chino.
Si bien la próxima temporada navideña puede ofrecer un impulso modesto a las importaciones de camarón, es poco probable que los volúmenes generales alcancen los niveles de 2023 debido a los desafíos económicos que enfrentan los consumidores chinos. Reportes recientes de la Oficina Nacional de Estadísticas destacan ventas minoristas más lentas de lo esperado, disminución del consumo de los hogares y debilitamiento de la seguridad laboral, lo que podría moderar el crecimiento del volumen de importación•
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
Importación de camarón de Estados Unidos
Autores: Jim Kenny
jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com
Urner Barry
Importaciones de todos los tipos, por tipo
En la data publicada a principios de este mes para dar inicio al segundo semestre, las importaciones de camarón de EE.UU. cayeron dos dígitos por segundo mes consecutivo en julio interanual. Esto se esperaba en general dado el contexto de factores externos, en particular retrasos en el transporte y aranceles/derechos, pero es impulsado principalmente por pérdidas descomunales de Ecuador. Las importaciones del mes fueron de 134.437 millones de libras, un 12.1 por ciento menos que en julio de 2023. Mientras que India ganó ligeramente (+0.3%), el resto de los top cinco cayeron drásticamente. En segundo lugar, Ecuador lideró las caídas (-30.5%), pero también se observaron en Indonesia (-9.7%), Vietnam (-15.7%) y Tailandia (-11.5%).
Los flujos comerciales generales mostraron pocos positivos netos interanuales. También se observaron pérdidas en las importaciones de Argentina (-1.6%), México (-30.5%) y Canadá (-39.1%).
La cifra acumulada en lo que va de año, de 905.702 millones de libras, ahora se sitúa un 4.4 por ciento por debajo de la del año pasado.
En términos de forma del producto, hubo pérdidas en todas las categorías, camarón sin cabeza y con cáscara, que incluye el pelado fácil (-28.3%), pelado (-1.6%), cocido (-21.8%) y empanizado (-2.8%).
Ciclos de importación mensual por país (todos los tipos)
India: Las importaciones de India aumentaron ligeramente después de una fuerte caída en junio (+0.3%). Fue esencialmente el único “brote verde” o recuperación económica, en el set de data de julio, con 58.928 millones de libras. La data acumulada ahora se sitúa solo un 0.8% por encima de los del año pasado para el mismo período. Hubo ganancias en el camarón con cáscara (+4.3%), pelado (+2.5%), pero mucho menos en el camarón cocido (-17.5%).
Ecuador: Nuevamente hubo una fuerte caída en las importaciones de Ecuador (-30.5%) debido a que las procesadoras/exportadores enfrentaron un clima comercial difícil en los EE.UU. Los precios en el extranjero y otras estructuras de costos han aumentado, pero la demanda en el país ha sido moderada en el mejor de los casos y es difícil trasladar el aumento de precios por completo. El país logró mantener la segunda posición en 28.468 millones de libras. Esto llevó a que las importaciones en lo que va del año, cayeran por debajo del año pasado en un 4.7 por ciento. Hubo grandes pérdidas en el camarón con cáscara (-48.6%) y empanizado (-81.8%) que perjudicaron una ligera ganancia en el camarón pelado (+2.9%).
Indonesia: Las importaciones de Indonesia (-9.7%) cayeron casi dos dígitos nuevamente en julio. Esto sitúa el total acumulado de 159.911 millones de libras en el nivel más bajo para el período desde 2019, y un 15.4 por ciento por debajo de 2024. Si bien hubo más camarón con cáscara (+8.7%), las grandes disminuciones en el camarón pelado (-18.4%) y cocido (-15.0%), y en menor medida en los empanizados (-3.5%), llevaron a las pérdidas.
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
Vietnam y Tailandia: Los dos países asiáticos se mantuvieron en línea con la mayoría del resto de los socios comerciales de EE.UU., ya que las dificultades de exportación tanto de Vietnam como de Tailandia obstaculizaron el intercambio. Las importaciones tanto de Vietnam (-15.7%) como de Tailandia (-11.5%) mostraron grandes disminuciones. Las primeras, aunque siguen siendo más altas, se redujeron aún más a un 4.8% más en lo que va del año, mientras que las últimas se ampliaron más a un 7.2 por ciento.
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones de camarón con cáscara, que incluye el pelado fácil, cayeron un 28.3 por ciento más en julio de 2024 en comparación con el año pasado.
La historia fue similar a la del mes pasado, con actores dispares. Las pequeñas ganancias en el segundo y tercer lugar, Indonesia (+8.7%) e India (+4.3%), fueron superadas por un claro primer lugar, Ecuador (-48.6%), así como por México (-36.9%) y Vietnam (-33.0%).
En cuanto a tamaños, todos mostraron pérdidas de dos dígitos, con la excepción de la talla 26-30 que perdió un solo dígito.
Los valores de reemplazo del camarón con cáscara (importación $/lb.) han continuado la tendencia al alza. El aumento de los costos en el exterior y precios bajos casi récord ha conllevado a ofertas más firmes. Los precios
se dispararon $0.12 por libra a $3.44 por libra.
Valor agregado, importación de camarón pelado
Las importaciones de camarón pelado, la categoría más grande por volumen, perdieron un 1.6 por ciento. Las importaciones de julio totalizaron 71.841 millones de libras. Si bien el número uno, India (+2.5%) y el número dos, Ecuador (+2.9%), ganaron, las grandes pérdidas de Indonesia (-18.4%), Vietnam (-17.4%) y Tailandia (-12.2%) derivaron a una disminución.
Los valores de reemplazo (importación $/ lb.) para el camarón pelado resistieron la tendencia del aumento de precios en muchas categorías y detuvieron dos meses de ganancias. Los precios fueron $0.02 más bajos por libra, a $3.42 por libra.
Las importaciones de camarón cocido (de agua tibia) también fueron en gran medida más bajas, con cifras un 21.8 por ciento por debajo de las del año pasado. Las pérdidas en las importaciones de camarón empanizado fueron más moderadas, con un 2.8 por ciento.
Importaciones de camarón cocido, empanizado y otros
Los precios de reemplazo del camarón cocido cayeron $0.12 por libra en julio después de un fuerte aumento en junio. La data más reciente se registra en $4.26 por libra.
Línea de tiempo del precio del camarón; anuncios minoristas
Minoristas: El movimiento de minoristas en agosto se retrasó significativamente el mes pasado, ya que cayeron un 31 por ciento con respecto a julio. También hubo una caída
Fuente: USDOC. Urner Barry
Detalle de importaciones de camarón YTD por tipo
Fuente: USDOC. Urner Barry
REPORTE DE MERCADO
de casi el 19 por ciento en comparación con el promedio de tres años. El péndulo precio/oportunidad puede haber sido un indicador. Después de que los minoristas llevaran los precios a la baja durante cuatro meses consecutivos, el precio promedio de los anuncios se disparó a $7.64 por libra, $0.32 por libra más, perdiendo todos los descuentos desde abril.
Suministro de camarón a EE.UU. y situación del Golfo
***La data de desembarques proporcionada por el NMFS ha sido inconsistente y hay lagunas en la data que hace que las comparaciones no sean confiables.
La data de desembarques de mayo de 2024 publicada por el NMFS mostró un aumento estacional continuo hasta los 8.246 millones de libras (total, todas las especies, sin cabeza) desembarcados. Todavía está por debajo del año pasado y es el mes de julio más bajo desde 2020. Hasta mayo, se reporta que los desembarques fueron de 19.298 millones de libras, o casi un 25 por ciento por debajo de los del año pasado.
Exportación del camarón ecuatoriano
El mercado de camarón tigre blanco y negro producido en granjas e importado, continuó oscilando entre completamente estable y firme, ya que los suministros siguen siendo limitados y los costos de reemplazo siguen un curso más difícil y de incremento. Los cambios en las evaluaciones siguen una tendencia al alza controlada. La demanda moderada continúa limitando el ritmo de
las ganancias. El camarón tigre negro muy grande, en su mayor parte, no ha tenido tan buen desempeño, pero sigue siendo el único lugar donde persiste la debilidad.
El mercado del camarón silvestre sigue bastante inestable. La oferta de camarón de origen estadounidense sigue siendo baja y los precios se ven ligeramente respaldados por aspectos positivos del mercado importado•
Mesa de trabajo con ministra de Producción para revisar mecanismo
tecnológico para la seguridad de carga de exportación
La Cámara Nacional de Acuacultura (CNA) logró agendar una reunión con la ministra de Producción, Comercio Exterior, Inversiones y Pesca, Sonsoles García, para conocer el nuevo sistema de perfilamiento de riesgo que el Gobierno busca implementar. La mesa de trabajo fue convocada por el ente público y contó con la participación de la CNA, como miembro del Directorio de la Corporación de Gremios Exportadores del Ecuador (CORDEX).
Este sistema busca minimizar las inspecciones intrusivas y garantizar que las exportaciones estén libres de contaminación, protegiendo a las empresas de posibles implicaciones legales en puertos
Revisión de procesos públicos para el sector productivo – exportador
La optimización de la logística de comercio exterior fue el tema central de la reunión entre la ministra de Producción, Comercio Exterior, Inversiones y Pesca, Sonsoles García, y directivos de la Cámara Nacional de Acuacultura.
En la mesa de trabajo, la CNA insistió en la necesidad de implementar mejoras en los servicios públicos, con el objetivo de simplificar y agilizar los procesos clave para el sector.
Se programó un próximo encuentro para evaluar los progresos y asegurar que las soluciones discutidas se traduzcan en resultados concretos para el sector.
Cámara Nacional de Acuacultura analiza proyectos de ley que afectan al sector camaronero
El equipo de la Cámara Nacional de Acuacultura (CNA) se reunió para examinar recientemente los diversos proyectos de ley en discusión en la Asamblea Nacional que podrían tener un impacto significativo en el sector camaronero. Durante la sesión, se evaluaron detalladamente las implicaciones legales y económicas de las propuestas legislativas, con el fin de identificar posibles efectos adversos para la industria. Con el objetivo de mitigar estos impactos, la CNA planea anticipar acciones frente al legislativo, en las que se incluye comparecer ante las comisiones correspondientes o ante el pleno de la Asamblea.
internacionales, reduciendo tiempos de espera y potenciales daños a la carga producidos por inspecciones físicas previas al embarque. La iniciativa fue bien acogida por el sector exportador, que la calificó como una herramienta positiva en el combate a la contaminación de la carga y en favor de las mejoras logísticas que requiere el país.
Reunión binacional
para fortalecer
los lazos comerciales con Argentina y Mercosur
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, participó en la reunión binacional organizada por el Comité Empresarial Ecuatoriano, la Cámara Argentina de Comercio y Servicios, y la Cámara de Comercio Ecuatoriano-Argentina.
El evento reunió a líderes empresariales y funcionarios clave del sector para discutir y promover nuevas oportunidades de cooperación. La ministra de Producción, Comercio Exterior, Inversiones y Pesca, Sonsoles García, estuvo presente en el encuentro, reafirmando el compromiso de Ecuador de ampliar su intercambio comercial con el Mercosur.
Esta reunión ha sido una plataforma crucial para fortalecer los lazos comerciales entre Ecuador y Argentina, y para explorar vías de colaboración que beneficien a ambos países en el ámbito comercial.
SUSTAINED: El nuevo programa de educación para el desarrollo sostenible de la industria camaronera ecuatoriana
Más de 120 personas vinculadas al sector camaronero participaron en SustainED, un programa educativo para impulsar el desarrollo sostenible de la industria camaronera, en un evento organizado por la Cámara Nacional de Acuacultura (CNA) y Sustainable Shrimp Partnership (SSP).
La primera jornada, realizada en el hotel Hilton Colón de Guayaquil, tuvo un enfoque clave en el uso responsable del metabisulfito de sodio, asegurando la calidad del camarón y el acceso a los mercados internacionales.
SustainED busca transformar el sector a través de la formación en los principios de sostenibilidad aplicados a la industria acuícola y comunicar la forma de afrontar los desafíos y riesgos relacionados a su aplicación. Esto incluye aspectos sociales, ambientales, económicos y de calidad, los cuales serán transmitidos mediante talleres, capacitaciones, redes sociales y actividades de campo.
Mesa de seguridad públicoprivada con el Ministerio del Interior
En la Gobernación del Guayas se llevó a cabo una reunión de trabajo con el subsecretario de Seguridad del Ministerio del Interior, para evaluar el protocolo de seguridad de la cadena logística de comercio exterior. Durante el encuentro, se discutieron diversas estrategias enfocadas en implementar requisitos de seguridad más estrictos para prevenir y combatir delitos como el contrabando, el tráfico de divisas y el tráfico ilícito de sustancias controladas. El objetivo de esta iniciativa es trabajar en conjunto para reducir las actividades ilícitas en la cadena logística de comercio exterior, mediante la implementación de procesos integrados y rigurosos requisitos de seguridad.
Ecuador presente en GLOBEFISH de la FAO
En Roma, la Directora Ejecutiva de la Cámara Nacional de Acuacultura, Yahira Piedrahita, participó en un diálogo abierto centrado en el futuro de la acuacultura y la pesca, en el marco del 40º aniversario de la iniciativa GLOBEFISH de la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO).
Este evento reunió a expertos y profesionales del sector para abordar desafíos y oportunidades en ambas industrias a nivel global.
Durante el encuentro, Piedrahita se reunió con destacados representantes de la FAO, como el Dr. Audum Lem, director adjunto de Pesca y Acuacultura; Mariana Toussaint, oficial de Pesca del equipo de Comercio y Mercados; Shirlene María Anthonysamy, coordinadora
CNA planteó los escenarios de riesgo por racionamientos eléctricos
La grave crisis energética que atraviesa el país fue el tema central abordado con la ministra de Producción, Comercio Exterior, Inversiones y Pesca, Sonsoles García, en varias mesas de trabajo que tuvieron el propósito de coordinar acciones emergentes entre el sector público y privado.
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, planteó los escenarios y su posible impacto en la industria camaronera. Fue enfático en dos puntos fundamentales: ser oportunamente informados sobre el plan de suspensión energética y analizar cortes diferenciados en zonas industriales como Durán y Yaguachi.
de GLOBEFISH, y Marcio Castro de Souza, oficial superior de Pesca. Juntos discutieron estrategias para fomentar prácticas sostenibles y resilientes que aseguren un futuro próspero para la acuacultura y la pesca.
Concluyó Primera etapa del SCALE UP: 17 Laboratorios participaron en el programa
En septiembre, el programa Scale Up del Sustainable Shrimp Partnership (SSP) registró importantes avances en su implementación en los laboratorios de larvas. Se realizaron preauditorías e inspecciones en 17 laboratorios que participan activamente en el programa, luego de haber capacitado a más de 85 personas en la Guía Técnica para la Sostenibilidad.
Estas auditorías permitieron a los laboratorios identificar y corregir no conformidades, lo que ha mejorado significativamente sus prácticas operativas y los ha acercado al cumplimiento de los estándares internacionales de sostenibilidad.
Los logros del SSP fueron posibles gracias a una colaboración estratégica con la Cámara Nacional de Acuacultura (CNA), que permitió organizar y llevar a cabo una serie de capacitaciones especializadas para los laboratorios.
Las inspecciones de campo, llevadas a cabo entre el 9 de julio y el 29 de agosto, ofrecieron retroalimentación constante a los laboratorios, permitiéndoles ajustar sus procesos a tiempo. Este esfuerzo coordinado involucró a representantes de los 17 establecimientos, el SSP y la CNA, marcando un avance crucial en la sostenibilidad de la producción de larvas.
Viceministro de Aduanas de China visitó Ecuador
En el marco del Tratado de Libre Comercio entre Ecuador y China, vigente desde mayo de 2024, una delegación encabezada por el viceministro de la Administración General de Aduanas, Zhao Zenglian, visitó Ecuador para fortalecer la colaboración bilateral.
Durante esta cumbre, la ministra de Producción, Comercio Exterior, Inversiones y Pesca, Sonsoles García, suscribió el Memorando de Entendimiento sobre Cooperación en Inocuidad de Alimentos entre las autoridades aduaneras del gigante asiático y varias entidades ecuatorianas.
La Cámara Nacional de Acuacultura informó oportunamente al sector sobre esta visita, brindando la asistencia técnica necesaria a sus empresas afiliadas en torno a los procesos de auditorías, destacando la importancia de cumplir con las normativas de bioseguridad y calidad que demanda el mercado chino.
AQUAYSISTEMAS abrirá nueva sucursal en Guayaquil
Aquaysistemas S.A., empresa manabita dedicada al diseño, construcción y operación de sistemas de purificación, tratamiento, desalinización y depuración de aguas para uso industrial, potable y humano, anunció la apertura de su nueva oficina técnica en Guayaquil, con el propósito de fortalecer su presencia en la región y optimizar la atención a sus clientes.
Esta nueva sede atenderá a las industrias que operan en Guayaquil, Durán y Santa Elena, zonas de alta incidencia productiva del sector acuícola.
Kampi llegó a Ecuador con inteligencia artificial de vanguardia
El 4 de septiembre, en el marco del ‘Global Shrimp Forum’ celebrado en Utrecht, Países Bajos, se presentó ‘Kampi’, una aplicación diseñada para transformar la gestión de las granjas camaroneras mediante el uso de tecnología avanzada. Desarrollada en colaboración entre Canadá y Ecuador, esta innovadora plataforma busca mejorar la toma de decisiones en la producción acuícola a través del monitoreo en tiempo real de las piscinas camaroneras desde cualquier parte del mundo.
Kampi incorpora inteligencia artificial y utiliza la cámara de un smartphone para capturar imágenes que permiten medir el tamaño y peso promedio de los camarones, generando histogramas automáticos para visualizar su distribución. Entre sus principales funciones están Optima Analytics, que integra la gestión de la granja en una sola pantalla; Optima Predictions, que permite realizar predicciones precisas sobre aspectos cruciales del negocio, como el momento óptimo para la cosecha, y también FarmBook, una herramienta que facilita la comunicación interna entre los equipos de producción.
Esta app busca optimizar la eficiencia y mejorar la rentabilidad en el sector camaronero global.