
14 minute read
Marco Baldanzi


Cromosomi sessuali negli uccelli
testo e disegni di Marco Baldanzi
Partendo dal chiedermi come mai ci fosse un minor numero di femmine rispetto ai maschi nel mio aviario all’aperto per canarini, ho colto l’occasione per fare questa concisa rassegna bibliografica degli studi effettuati sui cromosomi sessuali negli uccelli. Non entro in merito alla cosiddetta Regola di Haldane (1). Per chi fosse interessato si veda la review: Haldane’s rule in the 21st century pubblicata da Schilthuizen, Giesbers & Beukeboom nel 2011 su Heredity. Mi preme però ricordare le due principali opere di Charles Darwin Origine delle specie e selezione naturale e Origine dell’uomo e selezione sessuale: questa, meno insegnata della prima, si incentra sull’importanza del dimorfismo sessuale. Negli Uccelli i cromosomi sessuali dei maschi sono indicati come ZZ e per le femmine ZW; viceversa, nei Mammiferi i maschi sono XY e le femmine XX. Ciò porta a un’altra differenza tra Mammiferi e Uccelli: nei primi, sono gli spermatozoi a portare il cromosoma sessuale X o Y, ereditato rispettivamente dalla madre e dal padre, mentre nei secondi sono le uova a portare il cromosoma sessuale Z, ereditato dal padre, o quello W, ereditato dalla madre. Tralasciamo qui ipotetiche interazioni tra i cromosomi sessuali e il citoplasma dell’uovo, che è di origine materna, p. es. mitocondri (mtDNA), seppur oggetto di studi scientifici. Cominciamo con l’ipotesi Ohno: nel suo libro del 1967 Sex Chromosomes and Sex-Linked Genesil biologo nipponico-statunitense Susumu Ohno propose che i cromosomi sessuali fossero in origine omologhi(2) e la successiva differenziazione in X e Y avvenne solo a spese del cromosoma Y. Nel 1974, su BioScience, Bloom pubblicò Current Knowledge About the Avian W Chromosome. I
Sterno carenato: la carena permette l’inserzione dei grossi muscoli toracici o pettorali atti al volo. Al contrario, lo sterno piatto è tipico degli Uccelli che non volano (Ratiti: struzzi, casuari, emù). cromosomi degli Uccelli sono difficili da studiare perché sono molti (range = 52-92) e di diverse misure. Si precisa che, secondo la specie, le femmine possono essere ZW oppure Z0. Negli anni ’60 si è confermato in numerose specie di Uccelli il meccanismo ZZ ( ) – ZW ( ) di determinazione sessuale. Come spesso accade in biologia, non mancano le eccezioni: varie specie di Uccelli mancano del cromosoma W. Infatti, nella rassegna di Bloom si evidenzia che il cromosoma W non è altrettanto automatico nel determinare il sesso femminile quanto lo è il cromosoma Y nel determinare il sesso maschile nei Mammiferi. Nel 1997, sulla rivista Science Progress, L. Stevens pubblicò Sex chromosomes and sex determining mechanisms in birds, evidenziando che nella maggior parte degli Uccelli il cromosoma Z è grosso, il 4° o 5° in ordine di grandezza, e contiene quasi tutti i geni sesso-legati. Il cromosoma W generalmente è un micro-cromosoma, cioè molto piccolo, contenente sequenze di DNA molto ripetute. La determinazione del sesso negli Uccelli avverrebbe tramite un meccanismo di equilibrio genico, in cui il fattore cruciale è il rapporto tra autosomi e cromosomi Z. Nel 1999, su Biocell, Maria Ines Pigozzi pubblicò Origin and evolution of the sex chromosomes in


Cromosomi sessuali Z e W in varie specie di Uccelli: Rhea americana è un Ratite, a sterno piatto; le altre specie sono, in ordine, il germano reale, la faraona, il gallo, il piccione e il diamante mandarino, fonte: Pigozzi, 1999
birds. Basandosi su evidenze citologiche, propose che la mancanza di ricombinazione nell’ancestrale omomorfico paio ZW causò la degenerazione della regione non-ricombinante del cromosoma W, portando agli eteromorfici cromosomi sessuali degli Uccelli carenati. Sia nei Rettili sia nei Mammiferi e Uccelli, nelle specie più primitive i cromosomi sessuali sono omomorfici, mentre nelle specie più moderne divengono eteromorfici, arrivando al cromosoma Y e al W. Questi cambiamenti deriverebbero dall’isolamento meiotico, con perdita di geni attivi, aumento di trasposoni e sequenze ripetute. Negli Uccelli, il primitivo cromosoma W differiva dall’omologo Z per almeno due geni caratterizzanti il sesso femminile. Questi geni erano in una zona cromosomica con poca ricombinazione, per esempio vicino al centromero, prevenendo l’eventuale rottura dell’associazione genica mediante crossing-over. Con la progressiva speciazione della classe Uccelli, nuovi geni sesso-determinanti o legati alla fertilità si sono evoluti, aumentando la zona non-ricombinante del cromosoma W e la sua degenerazione. Rimane un corto segmento di ricombinazione col cromosoma Z che permette la formazione dei bivalenti in meiosi e quindi il mantenimento della coppia cromosomica ZW. Nel 2002, sulla rivista Cytogenetic and Genome Research, Shetty et al. hanno pubblicato DMRT1 in a ratite bird: evidence for a role in sex determination and discovery of a putative regulatory element. Preciso che i Ratiti (struzzi, nandù, emù, casuari, kiwi) sono un raggruppamento artificiale di Uccelli che non volano, perché hanno lo sterno piatto anziché carenato. In questi antichi Uccelli, i cro-
I NOSTRI LUTTI
In ricordo di Antonio Romagnoli
Abbiamo appreso della tua scomparsa in questo 2021 come un fulmine a ciel sereno, non eravamo ancora pronti a ciò… Eri una persona generosissima, buona e sempre disponibile; eri da quasi sessant’anni l’anima di ogni nostra esposizione, e di ogni nostra iniziativa; ogni qual volta ci trovavamo, si sentiva immancabilmente la tua inconfondibile voce allegra svettare al di sopra di tutte le altre; eri presenti in ogni fase dell’organizzazione delle mostre e non facevi mai mancare, con un motto di spirito, una battuta od una giusta critica, il tuo fattivo e sagace apporto a rafforzare il morale di tutti i soci, negli alti come nei bassi che capitano in ogni evento umano… Per tutto questo ti Ringraziamo e ci mancherai tantissimo, “Rùmagna”!
Il Consiglio ed i Soci tutti della Società Ornitologica Ferrarese - APS
In memoria di Rinaldo Montagnoli
Ècon grande tristezza che a distanza di qualche mese i soci e amici della Associazione Ornitologica Legnaghese salutano per l’ultima volta un altro amico e socio del gruppo. Rinaldo Montagnoli uno dei primi fondatori dell’associazione ornitologica e per moltissimi anni vice presidente, presidente emerito, e stimatissimo giudice canarini di colore, a 86 anni ci ha lasciato. Da un paio di anni non allevava più, ma quando le forze glielo consentivano, era sempre presente in associazione. Ascoltava e consigliava, ma senza presunzione di sapere tutto. L’umiltà era una delle sue grandi qualità, oltre a un grande istinto paterno che lo portava ad accogliere tutti i nuovi soci come dei figli da accompagnare alla scoperta di un mondo tutto nuovo. Ciao Rinaldo riposa in pace, la tua bontà di animo sarà un grande ricordo per tutti noi.
Soci e amici dell’Associazione Ornitologica Legnaghese.
mosomi sessuali sono omomorfici, cioè Z e W hanno stesse dimensioni e il sesso sarebbe determinato da un gene. Gli autori hanno clonato parte del gene DMRT1dall’emù e hanno visto che si esprime anche durante la differenziazione delle gonadi nel pollo. DMRT1si trova sul cromosoma Z ma non in W e quindi potrebbe essere coinvolto nella determinazione del sesso di tutti gli Uccelli. Si fa notare che la sequenza di DMRT1dell’emù è omologa per l’88% con quella del pollo (Gallus gallus domesticus) (3) e al 65% con quella umana. Nel 2007, sulla stessa rivista, Smith et al.pubblicarono una reviewdal paradigmatico titolo Avian sex determination: what, when and where?Intanto, puntualizzano che negli Uccelli il meccanismo, seppur ignoto, sia determinato da un gene sesso-legato portato da uno o entrambi i cromosomi sessuali. Come nei Mammiferi dovrebbe agire nelle gonadi embrionali, facendo formare o testicoli o ovario; da queste, formazioni di ormoni steroidi sessuali e, di conseguenza, organi e tessuti tipici dei due sessi. Nei Mammiferi il gene chiave è SRY, portato dal cromosoma Y. Il testosterone rilasciato produce tessuti e organi maschili, altrimenti si segue un programma di sviluppo femminile. SRYè assente negli Uccelli ma si pensa che il principio sia lo stesso e i geni candidati sono DMRT1 sul cromosoma Z oppure HINTWsul cromosoma W, entrambi espressi nelle gonadi ben prima della differenziazione sessuale. Ancora nel 2007, sempre su Cytogenetic and Genome Research, Stiglec et al.hanno pubblicato A new look at the evolution of avian sex chromosomes. Mentre il gene HINTWsul cromosoma W è il candidato più forte come fattore determinante la formazione dell’ovario negli Uccelli, il gene DMRT1, sul cromosoma Z, è l’unico gene coinvolto nella determinazione sessuale in tutti i Metazoi(4), facendo nascere i maschi di Mammiferi, Uccelli, Anfibi, Rettili, Insetti e Nematodi. Nei Mammiferi Euteri(5) o Euplacentati, DMRT1è autosomico, cioè non su cromosoma sessuale, e ne occorrono due copie (omozigote) per la formazione dei testicoli. Negli Uccelli invece è portato sul cromosoma sessuale Z e i maschi sono ZZ. Alla meiosi, la ricombinazione tra Z e W è assente nella regione distale del braccio contenente DMRT1. Negli Uccelli carenati, dove i cromosomi sessuali sono eteromorfici, il cromosoma Z è simile nei vari gruppi sistematici, eccetto che per la posizione del centromero, mentre il cromosoma W mostra sensibili differenze tra le specie. Addirittura, i dati suggeriscono che il progenitore dei Mammiferi possedesse cromosomi sessuali ZZ/ZW simili a quelli degli Uccelli. Forse il sistema XX/XY dei Mammiferi e quello ZZ/ZW degli Uccelli si sono evoluti in modo diverso da cromosomi autosomici di un comune progenitore Rettile avente determinazione sessuale temperatura-dipendente. Nel 2014, su BMC Evolutionary Biology, autori cinesi hanno pubblicato Temporal genomic evolution of bird sex chromosomes, con l’analisi di 45 genomi di Uccelli e i trascrittomi (6) di quattro specie. I cromosomi sessuali di Uccelli e Mammiferi si sono originati indipendentemente da differenti coppie di autosomi ancestrali, seguendo una graduale e parallela soppressione della ricombinazione. I cromosomi sessuali, nel corso dell’evoluzione, sono andati incontro a mutazioni e regimi di selezione indipendentemente dagli autosomi. Interessante, per esempio, ricordare che le linee germinali maschili (spermatogenesi), proprio per il loro gran numero di divisioni cellulari, accumulano molte più mutazioni, in unità di tempo, rispetto alle femmine (cellule uovo) nella maggior parte degli animali. Le differenze dei cromosomi sessuali tra Mammiferi e Uccelli derivano
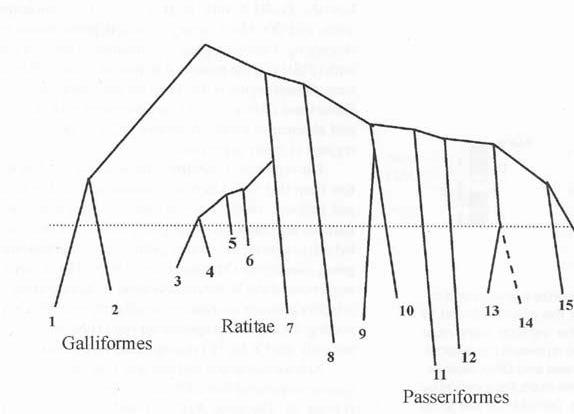
Albero filogenetico della classe Uccelli mostrante la relazione tra 15 ordini: 1) Gallus domesticus; 2) Numida meleagris (faraona); 3-7) varie specie di Ratiti; 8) Psittaciformi (pappagalli); 9) Pelecaniformes; 10) Charadriiformes (p.es. gabbiani); 11) il maggior numero di specie di Uccelli, riunite nell’ordine Passeriformi; 12) Strigiformes (p.es. civetta, barbagianni, gufo, ecc.); 13) Sphenisciformes (pinguini); 15) Procellariformes (p.es. albatro e uccelli marini); 16) Gaviiformes (alcuni uccelli acquatici). Manca l’ordine Anseriformes (p.es. anatre, oche, cigni) che, insieme ai Galliformi, sono gli uccelli neorniti più primitivi. Per neorniti si intendono tutti gli uccelli moderni, evoluti negli ultimi 66 milioni di anni, fonte: Pigozzi, 1999

Cromosomi sessuali Z e W di Gallus domesticus, fonte: Pigozzi, 1999
anche dal fatto che il cromosoma X è emizigote nei maschi, presente cioè in singola copia (XY), mentre il cromosoma Z è emizigote nelle femmine (ZW) degli Uccelli (e anche di Rettili, Pesci e farfalle), risentendo quindi della opposta selezione sessuale, come accennato nell’introduzione. Ancora nel 2014, su Chromosome Research, Jennifer Graves ha pubblicato Avian sex, sex chromosomes, and dosage compensation in the age of genomics. La scienziata considera il cromosoma W degli Uccelli una reliquia degenerata del cromosoma Z (eccetto Ratiti e Tinamidi(7), proprio come nei Mammiferi la Y è una reliquia della X. In quanto cromosoma specifico delle femmine, la maggior parte dei suoi geni si esprime nell’ovario. Al contrario, la maggior parte dei geni sul cromosoma Z si esprime nei maschi: questa “mascolinizzazione” è il risultato di una forte selezione (sessuale) del successo riproduttivo maschile. Questi geni sul cromosoma Z si sono evoluti a una velocità doppia rispetto ai geni autosomici, probabilmente sempre per la sua maggiore attivazione nei testicoli (spermatogenesi). Sono specifici geni a determinare il sesso negli Uccelli. Com’è risaputo, la temperatura d’incubazione delle uova condiziona l’espressione fenotipica, incluso il rapporto fem mi ne/maschi alla schiusa, che probabilmente deriva anche da una differenza di mortalità tra i due sessi ed è comunque sbilanciato in molte specie di Uccelli. Ma quali sono questi specifici geni portati dai cromosomi sessuali W e Z? Sul cromosoma W non sono stati per ora (2014) individuati. Invece, tra i geni portati sulla coppia cromosomica ZZ, il gene DMRT1 è quello essenziale per lo sviluppo di un maschio, sia negli Uccelli che nell’uomo. Necessario ma non sufficiente: negli Uccelli, esso avvia la formazione delle gonadi maschili ma gli ormoni sessuali che queste producono nonattivano tutte le differenze somatiche dei due sessi. Nel 2015, su Nature Communications, un’estesa collaborazione di ricercatori scandinavi, slavi e spagnoli ha pubblicato Evolutionary analysis of the female-specific avian W chromosome. La specie studiata è stata Ficedula albicollis, balia dal collare, fam. Muscicapidi. I risultati dimostrano punti in comune e differenze nell’evoluzione del cromosoma W e del cromosoma Y. La trasmissione attraverso l’oogenesi, piuttosto che la spermatogenesi, implica che i cromosomi W sono esposti a un diverso ambiente mutazionale ed epigenetico della linea germinale rispetto ai cromosomi Y dei Mammiferi. I mitocondri si trovano nel citoplasma; questo è ereditato tramite la cellula uovo e quindi è matrilineare. È lecito chiedersi: possono esserci negli Uccelli associazioni cito-nucleari tra mtDNA e cromosoma W? Bene, questi scienziati hanno dimostrato una stabilità co-evolutiva di questi due cromosomi. Nel 2018, sulla rivista Molecular Ecology, lo zoologo canadese Irwin Darren ha pubblicato Sex chromosomes and speciation in birds and other ZW systems. Concentrandosi sulla determinazione del sesso ZW (tipico di uccelli, molti serpenti e lepidotteri), ha esaminato come le dinamiche evoluzionistiche differiscano tra Z, W e gli autosomi, discutendo di come queste differenze possano portare a un ruolo maggiore dei cromosomi sessuali nella speciazione e utilizzando i dati di un’ampia varietà di specie di uccelli per confrontare i tassi evolutivi relativi di cromosomi sessuali e autosomi. Il più veloce accumulo di vari tipi di mutazioni e le complesse dinamiche evolutive di tratti sessualmente antagonisti contribuiscono a un certo ruolo per il cromosoma Z nella speciazione. Pure il cromosoma W ha un forte potenziale per essere coinvolto nella speciazione, anche per la sua co-ereditarietà con il genoma mitocondriale (mtDNA)(8). Teoria ed evidenza empirica suggeriscono un ruolo sproporzionato per i cromosomi sessuali nella speciazione, ma la loro importanza è moderata dalle loro piccole dimensioni rispetto all’intero genoma. Nel 2020, sulla rivista Genes, Luohao Xu e Qi Zhou hanno pubblicato The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized. Gli autori premettono che i cromosomi sessuali Y e W sono molto eterocromatici e poveri di geni, al contrario degli X e Z, molto eucromatici e ricchi di geni(9). Rispetto al sistema XY, ci sono meno studi riguardo alla selezione sessuale sul contenuto genico dei cromosomi Z e W: i cromosomi Z degli Uccelli sembrano essere mascolinizzati, cioè recanti geni espressi nei maschi, mentre la “femminilità” dei cromosomi W non è così netta. Questi autori, esa-
minando i genomi femminili di 27 specie nei tre principali cladi (10) della classe Uccelli, hanno rivelato una forte conservazione genica nei cromosomi W, ma poca evidenza circa la sua “femminilità” (trad. feminization). In effetti, la selezione sessuale è indirizzata verso i maschi, per i colori, il canto o il comportamento.
NOTE
(1)J.B.S. Haldane (1892-1964) nel 1922 affermò che “quando nella progenie di due diverse razze animali un sesso è assente, raro o sterile, quel sesso è il sesso eterozigote (eterogametico)”. Poiché la maggior parte degli alleli dannosi portati in singola dose (emizigosi) da un sesso non risultano bilanciati da altrettanti geni non mutati presenti sul cromosoma omologo, il sesso eterogametico risulta geneticamente più vulnerabile. (2)I cromosomi omologhi sono cromosomi morfologicamente identici presenti negli organismi eucarioti. Possono essere definiti come cromosomi che in locicorrispondenti presentano gli stessi geni con le
stesse informazioni. (3)Nel pollo, il grosso cromosoma Z ha oltre 350 geni, mentre il cromosoma W ne ha forse meno di venti. (4)Raggruppamento sistematico comprendente tutti gli animali che presentano una condizione monocellulare allo stadio di germe ma che raggiungono, durante lo sviluppo, una condizione pluricellulare con un numero più o meno elevato di elementi somatici variamente differenziati; si contrappongono ai Protozoi. (5)Il nome significa “vere bestie”. In greco zoon significa animale od essere vivente (come il latino animal), mentre therionsignifica bestia(cfr. latino fera, belua, bestia).
Gli Euteri hanno una temperatura corporea più elevata e stabile degli altri Mammiferi: a causa di ciò posseggono testicoli esterni, in quanto la spermioistogenesi viene disturbata da temperature troppo elevate. (6)Viene definito “trascrittoma” la totalità degli RNA trascritti a partire da un genoma. (7)Famiglia di Uccelli Neorniti, diffusi dal Messico alla Patagonia, scarsi volatori, di dimensioni fra quelle di una quaglia e quelle di un pollo, dei quali hanno pressappoco anche l’aspetto. (8)Il DNA mitocondriale, a volte abbreviato
in mtDNA, è il DNA collocato nei mitocondri. Esso codifica e produce gli enzimi (proteine funzionali), necessari alla corretta realizzazione del delicato processo di fosforilazione ossidativa. Le istruzioni per la sintesi di questi enzimi risiedono nei 37 geni che compongono il genoma del DNA mitocondriale umano. (9)La cromatina è la forma in cui gli acidi nucleici si trovano nella cellula. Si trova negli eucarioti ed è costituita da DNA, proteine acide e basiche. Si distinguono due tipi di cromatina: eucromatina, meno condensata, corrisponde a zone in cui vi è un’intensa attività di trascrizione per la sintesi proteica (ossia di copia delle molecole di
DNA in molecole di RNA messaggero, mRNA); eterocromatina è invece la componente più condensata, costituisce circa il 10% del genoma e non sembra presentare attività di trascrizione. (10)In tassonomia, si definisce “clade” un gruppo costituito da un antenato singolo comune e da tutti i discendenti di quell’antenato. Qualsiasi gruppo che corrisponde alla definizione viene considerato monofiletico e può essere rappresentato o da un’analisi filogenetica o da un cladogramma.











