


Edición 154 - Agosto 2023
Inversiones del sector camaronero ecuatoriano.
La crisis del mercado afecta al sector camaronero ecuatoriano: pérdidas superiores a mil millones de dólares.
Sector camaronero ecuatoriano atraviesa grave crisis de inseguridad.
Aqua Expo El Oro 2023, intercambio de conocimientos e innovación.
¿Sabías que lo correcto es denominar al camarón blanco del Pacífico Penaeus vannamei y no Litopenaeus vannamei?.
Evaluación de la transmisión horizontal del virus del Síndrome de la Mancha Blanca en el camarón blanco (Litopenaeus vannamei) según el grado de severidad de la enfermedad y la tasa de desprendimiento viral.
Las características físicas y nutricionales de los alimentos microextruidos y su efecto en el rendimiento de larvas de Litopenaeus vannamei.
Acumulación de sólidos, un aspecto crítico en los sistemas acuícolas intensivos. Alternativas para una gestión sostenible.
Evolución del Fenómeno de El Niño. Recomendaciones para el sector camaronero.
Calidad del agua, calidad de vida y producción.
Presidente Ejecutivo Ing. José Antonio Camposano
Editora “AquaCultura”

Msc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita
PhD. Leonardo Maridueña Ing. José Antonio Lince Econ. Danny Vélez Ing. Alex de Wind
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Exportaciones de camarón y tilapia.
Reporte de mercado de EE. UU.
Reporte de mercado de China.
Noticias de interés.
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Corrección de estilo
Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
Gabriela Nivelo gnivelo@cna-ecuador.com
ÍNDICE
INFORMACIÓN DE COYUNTURA ARTÍCULOS TÉCNICOS
ESTADÍSTICAS
NOTICIAS 8 10 21 13 16 24 38 48 56 60 66 71 74 80
En el presente cierre editorial de la revista AquaCultura, observamos con notable interés el próximo escenario electoral en el Ecuador, en el cual se perfila la elección de un nuevo Presidente y otras dignidades por un corto período de alrededor de año y medio. Es, en cierta medida, sorprendente que el país se disponga a invertir recursos y a transitar por un sendero caracterizado por la inestabilidad, hasta que se logre instaurar un nuevo mandatario en el año 2025, momento en el cual se procederá a seleccionar las autoridades que regirán durante un período subsiguiente de cuatro años. A pesar de ello, aquel que emerja triunfante de este proceso democrático puede confiar en que nuestra labor respaldará aquellas iniciativas que propicien la generación de empleo y la promoción del bienestar de la ciudadanía.
El contexto político que nos convoca es uno que desafía a aquellos de nosotros que participamos activamente en la esfera gremial del Ecuador. En ausencia de agendas con enfoques a largo plazo y políticas de Estado consolidadas, nos encontramos en una constante oscilación en torno al modelo de desarrollo que anhelamos. Paralelamente, las empresas se ven abocadas a intentar vislumbrar el futuro con el fin de infundir cierto grado de certidumbre en las inversiones que proyectan ejecutar.
En tales circunstancias, los gremios emergen como interlocutores cruciales, cargados de la responsabilidad primordial de elevar a discusión aquellos asuntos prioritarios capaces de mitigar los riesgos inherentes y de capitalizar las oportunidades que puedan emerger en el ámbito de las actividades productivas. Por ende, se erige como requisito esencial la presencia de gremios sólidos que respalden eficazmente las iniciativas que favorecen al sector productivo y, al mismo tiempo, cuestionen con propiedad aquellas que lo perjudican. De esta manera, a cada iniciativa empresarial en el Ecuador debería corresponderle la compañía de un gremio solvente, dispuesto a reaccionar de manera ágil frente a la diversidad de desafíos que el gobierno de turno puede demorar en comprender y resolver.
En congruencia con este enfoque, los gremios que desempeñan su rol adecuadamente, respaldando las propuestas que redundan en beneficio del entorno productivo y ejerciendo cuestionamientos constructivos ante las decisiones que lo comprometen, se encuentran constantemente en proceso de innovación, desarrollando servicios nuevos con miras a satisfacer las demandas emergentes de sus afiliados.
José Antonio Camposano Presidente Ejecutivo

Enfrentando la inestabilidad política: El gremialismo como protagonista en Ecuador
No obstante, es innegable que Ecuador dista de consolidarse como un destino capaz de atraer inversiones significativas mediante la gestión gubernamental. La administración saliente, inicialmente concebida como proclive a un enfoque de desarrollo basado en el fomento del empleo privado, se focalizó de manera exclusiva en la búsqueda de soluciones al apremiante problema fiscal. Sin embargo, omitió abordar los desafíos en términos de competitividad inherentes a un país dolarizado que, hasta la fecha, no ha implementado acciones efectivas para subsanar los factores que generan costos crecientes constantemente.
En paralelo, gremios como la Cámara Nacional de Acuacultura han dirigido su atención a diversas problemáticas y han esbozado posibles soluciones, pese a que estas propuestas raramente han sido objeto de un análisis exhaustivo de parte de las autoridades. Paradójicamente, en ocasiones la negativa ha imperado como respuesta, mientras hoy se encuentra en desarrollo una nueva reforma tributaria que incidirá negativamente en el rendimiento de las empresas en diversos sectores, incluyendo el ámbito productor y exportador camaronero.
Resulta desalentador observar el desenlace para una administración que inicialmente auspiciaba una apertura económica enérgica mediante la concertación de acuerdos comerciales. Lamentablemente, careció de la perspicacia necesaria para reconocer que esta apertura conlleva la obligación de preparar y fortalecer a los agentes económicos privados, con miras a potenciar su capacidad competitiva en el ámbito internacional.
Hoy, lamentamos el evidente deterioro de nuestra capacidad para competir en los mercados, motivo por el cual no dejaremos de señalar las áreas en las que debemos concentrarnos para estimular la producción nacional y recobrar una senda de crecimiento que engendre bienestar para la ciudadanía, en particular aquellos individuos que anhelan oportunidades laborales en una nación que encara una seria crisis económica y social, alimentada por la decadencia de nuestro potencial para generar oportunidades; eslogan que ha perdido su brillo para aquellos que culminan sus días al frente de la administración•
EDITORIAL
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
DIRECTORIO
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
SEGUNDO VICEPRESIDENTE

Ing. Fabricio Vargas
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
Ing. Roberto Aguirre
VOCALES
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Miguel Uscocovich
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
Blgo. Carlos Sánchez
Ing. Diego Puente
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Carlos Miranda
Econ. Danny Vélez
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León

INVERSIONES
Banco Holandés y Grupo Almar:
Con el propósito de fortalecer su compromiso de construir un futuro sustentable y ecoeficiente, el Grupo Almar de Ecuador y el banco de desarrollo neerlandés Entrepreneurial Development Bank FMO anunciaron un acuerdo mediante el cual la institución financiera destinará $35 millones a las empresas Limbomar y Limbopack, para la ejecución de dos proyectos de fundamental importancia para la sostenibilidad acuícola.

El primer proyecto tiene como objetivo principal continuar con la electrificación de las granjas productoras de camarón, reemplazando el uso de diésel por fuentes de energía principalmente renovables. Esta transición no solo permite una operación más productiva, sino que además es altamente beneficiosa para el ambiente, pues se logra una mayor producción de camarón utilizando menos terreno. Adicionalmente, este proyecto fomenta la automatización de los procesos productivos, lo que se traduce en una reducción significativa de los costos operativos.
El segundo proyecto abarca la construcción de una nueva planta de procesamiento y empaque que contará con la certificación ASC (Aquaculture Stewardship Council). Esta instalación tiene previsto ofrecer cerca de mil plazas de empleo, de las cuales el 70% estarán destinadas a mujeres, lo que refleja un compromiso sólido del Grupo Almar con la equidad de género en el ámbito laboral.
Con estas acciones, el grupo acuícola ecuatoriano no solo busca consolidar su
presencia en la cadena de valor, desde el cultivo hasta el empaque y la exportación, sino que también se enfoca en ser un agente activo para el desarrollo de la comunidad y sus familias a través de la generación de empleo.

Ricardo Noboa, gerente financiero de Grupo Almar, destacó la relevancia de este acuerdo económico, enfatizando en los beneficios que trae no solo al desarrollo y crecimiento de la empresa dentro de la industria, sino también a las oportunidades tangibles que genera para las comunidades de influencia. “Estos son proyectos que nos permiten avanzar hacia el cumplimiento de objetivos comunes, y mirar con optimismo y entusiasmo hacia el futuro”, afirmó.
Estos dos proyectos, a desarrollarse en el mediano plazo, se enmarcan perfectamente en los Objetivos de Desarrollo Sostenible (ODS) 2 - Hambre Cero y 13 - Acción por el Clima, reafirmando así el compromiso de Grupo Almar y FMO con una agenda global de desarrollo sostenible.
La empresa ecuatoriana tuvo su origen en 1981 y actualmente agrupa a las empresas Produmar, Limbomar, Biogemar y Produpesada. Con más de 40 años de experiencia en la industria, Grupo Almar ha consolidado su compromiso y liderazgo de la sostenibilidad a través de las certificaciones Global GAP, Aquaculture Stewardship Council (ASC) y Sustainable Shrimp Partnership (SSP).
El FMO es el banco de desarrollo empresarial de los Países Bajos. Fundado en 1970, tiene como objetivo promover el crecimiento sostenible en países en vías de desarrollo y mercados emergentes mediante la inversión en proyectos empresariales. El banco es propiedad del Estado neerlandés (que posee una participación mayoritaria), y de bancos comerciales, empresas y sindicatos de Países Bajos.
FMO tiene un fuerte enfoque en el desarrollo sostenible y busca invertir en proyectos que tengan un impacto social y ambiental positivo. Esto incluye, por ejemplo, proyectos que generan empleo, fomentan la energía limpia y ayudan a combatir el cambio climático.
8
aliados en la ruta hacia la acuacultura sostenible
Multinacional japonesa Mitsui, interesada en comprar acciones de Santa Priscila
El 7 de agosto pasado, el CEO de la firma japonesa Mitsui & Co., Kenichi Hori, anunció en su portal que busca desarrollar una sociedad con Industrial Pesquera Santa Priscila (IPSP), a través de la adquisición de acciones minoritarias de la firma ecuatoriana por un valor de $360 millones.
“En Ecuador, epicentro global de la industria del cultivo de camarones para exportación, Mitsui y los fundadores de IPSP (Santa Priscila) trabajarán juntos para potenciar la productividad y eficiencia, optimizando la operatividad y satisfaciendo las demandas del cliente", indicó Mitsui en un comunicado oficial. Además, se resalta la intención de ambas partes de mantener un firme compromiso con la responsabilidad social, el respeto ambiental y la calidad del producto, con la visión de reforzar a IPSP en su liderazgo para modernizar esta industria en crecimiento. “Nuestro objetivo es fortalecer y expandir nuestra presencia en el mercado camaronero global, y vemos en Santa Priscila un socio estratégico ideal para lograrlo”, aseguró el CEO de la multinacional japonesa, reafirmando su confianza en este posible acuerdo.
Por su parte, para Industrial Pesquera Santa Priscila, esta alianza se percibe como una oportunidad de fortalecimiento y expansión, beneficiándose de la experiencia y el capital de una de las multinacionales más importantes de Japón, expresó en un comunicado emitido a la opinión pública.
La empresa ecuatoriana, liderada por el
empresario Santiago Salem Kronfle, fue fundada en 1976 en la ciudad de Machala, provincia de El Oro. Produce y exporta camarón ecuatoriano, cumpliendo los más altos estándares que exigen los mercados de destino. Durante la pandemia, la empresa demostró su capacidad de adaptación a las nuevas condiciones globales, fortaleciendo sus protocolos de bioseguridad.
Entre los hitos más destacados dentro de su trayectoria: IPSP hizo historia al convertirse en la primera entidad no petrolera de Ecuador en superar los US$ 1.005 millones en exportaciones, posicionándose además como el principal proveedor de camarón para Estados Unidos, superando incluso a distribuidores asiáticos.
Para finales de 2022, Santa Priscila continuó dominando el ranking de exportadoras de camarón, alcanzando ventas por US$ 1.422,9 millones. Además, se consolidó como la tercera industria más grande de Ecuador, según la clasificación de la Superintendencia de Compañías.
IPSP cuenta con seis plantas empacadoras en la costa ecuatoriana y cuenta con una fuerza laboral de más de 15 mil colaboradores, haciéndola uno de los más importantes creadores de fuentes de trabajo del sector privado. Sus principales destinos de exportación son China, Japón, Taiwán, Rusia, España y otros países europeos. Además, abastece a los mercados de Estados Unidos, Colombia y Chile.
José Antonio Camposano, presidente
ejecutivo de la Cámara Nacional de Acuacultura, destacó en redes sociales la importancia de la inversión extranjera en esta empresa ecuatoriana:
“La noticia nos llena de un profundo sentido de orgullo a todo el gremio camaronero. El acuerdo en cuestión constituye indudablemente un reflejo de la confianza depositada en empresas emblemáticas del sector camaronero ecuatoriano como Santa Priscila, por parte de inversionistas que comparten una visión común de desarrollo y, a su vez, creen fervientemente en el potencial del Ecuador”.
Por su parte, Mitsui es una reconocida compañía japonesa que opera en siete segmentos con 16 unidades de negocio. Se enfoca en el crecimiento a través de comercio, gestión y desarrollo de proyectos en diversos sectores, desde minería y energía hasta alimentación y agricultura. Su visión es contribuir al bienestar de la sociedad global a través de sus actividades comerciales.
La compañía reconoce la creación de ecosistemas de bienestar como una de sus principales iniciativas dentro de su Plan de Gestión a Mediano Plazo 2026. Mitsui aspira a mejorar la calidad de vida de las personas ofreciendo alimentos saludables. Con esta inversión, se propone añadir valor a su conglomerado en el sector de alimentos y nutrición, apuntando a promover la salud y el bienestar a través de la alimentación.
Sin embargo, para materializar este acuerdo, se espera obtener las aprobaciones regulatorias de diversos países hasta marzo de 2024, y se deberá implementar un plan de acción previamente establecido, conforme lo indicado por IPSP en su comunicado de prensa•

9
- AGOSTO 2023 COYUNTURA
La Cámara Nacional de Acuacultura (CNA), entidad que agrupa y representa al sector camaronero ecuatoriano, informó mediante un comunicado sobre la situación crítica que enfrenta actualmente la actividad, debido a que los precios internacionales del camarón han descendido a niveles alarmantes, situándose incluso por debajo de los mínimos registrados durante la pandemia de COVID-19 en 2020.
En los últimos meses, el precio de exportación del camarón ha disminuido a USD 0,70 por libra, provocando un fuerte revés en los ingresos de la industria.
La caída en los precios internacionales, atribuida en gran medida al debilitamiento del consumo en China y otros mercados clave, está ejerciendo una presión insostenible sobre la cadena del camarón ecuatoriano.
La contracción de la demanda en muchos países ha afectado negativamente la capacidad de los exportadores para encontrar mercados receptivos. Ante esto, se registra el crecimiento de apenas 1 dígito en las exportaciones en contraste con años anteriores, de acuerdo con los datos del Banco Central del Ecuador.
Por otra parte, el productor camaronero ecuatoriano ha enfrentado duros desafíos en términos de competitividad, ocasionados por el incremento sostenido de los costos operativos. Estos incluyen el precio del combustible, los salarios de la mano de obra, la inseguridad y el costo de las materias primas. En el último año, los costos de producción para la industria camaronera han escalado en USD 0.28 por cada libra producida.
Ante este escenario de descenso de precios y aumento de costos, el sector camaronero ha experimentado una merma de liquidez de USD 0,98 por cada libra producida. Esta situación se traduce en un grave impacto económico que supera los mil millones de dólares.

A pesar de las adversidades, el sector camaronero ecuatoriano reafirma su compromiso de continuar trabajando de manera unida y decidida para identificar soluciones viables, tomando en cuenta que la industria representa un pilar económico esencial y una fuente de empleo crucial para el Ecuador•

10
10
La crisis del mercado afecta al sector camaronero ecuatoriano:
- AGOSTO 2023 COYUNTURA
Pérdidas superiores a mil millones de dólares


Sector camaronero ecuatoriano atraviesa grave crisis de inseguridad
De enero a mediados de agosto de 2023 han sido reportados 61 incidentes delictivos que dejaron 44 heridos, dos muertos y una víctima de secuestro que finalmente fue rescatada por la Unidad Antisecuestros y Extorsión de la Policía Nacional (UNASE). De acuerdo con las estadísticas de la Dirección de Seguridad de la Cámara Nacional de Acuacultura (CNA), estas cifras son apenas una muestra de lo que ocurre en el sector, pues existe un 30% de “cifras negras”, correspondiente a personas que no denuncian por temor a represalias y por ende, al no ser judicializadas, se invisibilizan.

Ante esta grave situación de inseguridad, la CNA, a través de su presidente ejecutivo, José Antonio Camposano, ha manifestado públicamente que el sector privado con la ayuda de la Policía Nacional ha identificado las rutas fluviales y terrestres con mayor incidencia criminal. Estas zonas son: el kilómetro 26 de la vía Naranjal-Taura, y las vías Durán-Tambo y Playas. En cuanto a las rutas fluviales, se destacan el Golfo de Guayaquil (Puerto Roma y Punta Piedra) y el Archipiélago de Jambelí. No obstante, por falta de recursos económicos y logísticos de las entidades de control, no se realiza un patrullaje efectivo y no se cuenta con un eficiente trabajo de inteligencia; lo que impide la desarticulación de las bandas criminales que operan en estos sectores y están plenamente identificadas por los organismos de control.
Entre tanto, el sector camaronero ecuatoriano continúa invirtiendo cerca de 80 millones de dólares anuales en materia de seguridad, para contar con guardias en turnos rotativos y financiar la implementación de sistemas de vigilancia con inteligencia artificial.
Como medida urgente, la CNA solicitó, a través de la Corporación de Gremios Exportadores del Ecuador (Cordex), mediante oficio al presidente de la República, Guillermo Lasso, se decrete ‘Estado de excepción por grave conmoción interna' en las provincias de Santo Domingo, Manabí, Los Ríos, Guayas y El Oro. El objetivo de esta medida era recuperar el control en áreas del país que se han identificado como altamente peligrosas, y llevar a cabo operativos focalizados con labores de inteligencia para desarticular
COYUNTURA
13
44 personas heridas, dos muertos y una víctima de secuestro vinculadas al sector se registran en menos de 8 meses.
COYUNTURA
bandas delictivas que están sembrando el terror a través de extorsiones, amenazas, asaltos y robos cotidianos.
Sin embargo, el gremio advirtió que se deben abordar las raíces de esta problemática, que encuentra sus orígenes en la falta de atención a la profunda crisis de pobreza, tanto en el ámbito urbano como rural, con una alarmante tasa que alcanza el 44%. La inquietante confluencia de factores, incluyendo el índice de desempleo y el estancamiento económico, ha generado un deterioro social que favorece el reclutamiento de jóvenes hacia organizaciones criminales dedicadas al crimen y la violencia.
En tal sentido, el sector camaronero ecuatoriano, a través de boletines de prensa difundidos por CNA y Cordex, han planteado como necesaria la colaboración internacional especializada para enfrentar el crimen organizado, y reitera al Gobierno sus propuestas anteriores, que incluyen el fortalecimiento de la inteligencia policial en colaboración con las Fuerzas Armadas, intensificar patrullajes terrestres y fluviales, resguardar zonas de alta incidencia delictiva, autorizar armas de largo alcance para guardias de seguridad y tomar medidas judiciales, como la destitución de jueces que dicten medidas sustitutivas a delincuentes y el refuerzo del programa de víctimas y testigos.
En agosto pasado, el General de Distrito William Villarroel Trujillo, comandante de la Zona 8 de la Policía Nacional de Ecuador, participó de una reunión con el director de Seguridad de la CNA, Luis Herrera; el subsecretario de Acuacultura, Axel Vedani, y representantes del sector acuícola, con el objetivo de analizar y proponer nuevas estrategias de control del delito.

Posterior a esta reunión se evaluaron los resultados de los trabajos coordinados y articulados a nivel público y privado, que buscan fortalecer la colaboración entre estas entidades y mejorar la seguridad en zonas críticas.
En la reunión también se discutió la
"La situación actual exige una respuesta decidida y coordinada por parte de las autoridades para salvaguardar la seguridad de nuestros ciudadanos y garantizar un ambiente propicio para el desarrollo económico y social. Es esencial que las medidas implementadas aborden tanto las causas profundas como las circunstancias inmediatas que rodean este problema, de lo contrario no retomaremos el ambiente de paz que nos merecemos”.
conformación del grupo de la Policía Judicial (PJ) destinado a realizar labores investigativas específicas relacionadas con eventos delictivos en contra del sector camaronero. Este grupo tiene el objetivo de investigar y combatir las actividades ilícitas que afectan a este segmento económico clave.
La inseguridad que atraviesa Ecuador ha trascendido fronteras, generando un impacto grave a nivel económico y social. La CNA, como entidad representante del sector camaronero del país, continúa proponiendo ante el Gobierno nuevas estrategias; sin embargo, es fundamental el rol del Ejecutivo para la ejecución de acciones concretas y efectivas para restituir la seguridad ciudadana•

- AGOSTO 2023
14
José Antonio Camposano Presidente ejecutivo de la Cámara Nacional de Acuacultura

Aqua Expo El Oro 2023 fue inaugurada el 11 de julio pasado, en Machala con la presencia de autoridades provinciales y de gobierno. La mesa directiva estuvo integrada por Karina Torres, viceprefecta de la provincia de El Oro; Jenny Machuca, vicealcaldesa de Machala; Axel Vedani, subsecretario de Acuacultura; Luis Mejía, director de Habilitación y Certificación Sanitaria de la Secretaría de Calidad e Inocuidad (SCI); Marcelo Vélez, presidente del Directorio de la Cámara Nacional de Acuacultura (CNA), y José Antonio Camposano, presidente ejecutivo de la CNA.

Este año, el evento estuvo conformado por una feria comercial con más de 100 stands, en la que participaron 73 empresas ecuatorianas y extranjeras que mostraron lo más destacado de su cartera de productos y servicios.


Por otra parte, contó con un congreso técnico en el que participaron 18 conferencistas internacionales y ecuatorianos, quienes abordaron temas de interés e importancia para el sector acuícola en diversos aspectos: salud animal, nutrición, producción, manejo, tecnología, mercado, ambiente y prevención de riesgos.
INTERCAMBIO DE CONOCIMIENTOS E INNOVACIÓN
“La Cámara ha sido siempre la voz representativa de nuestra industria. Su papel fundamental es ser el referente técnico que vela por el desarrollo competitivo de toda la cadena de valor del mejor camarón del mundo”.
16
José Antonio Camposano Presidente ejecutivo de la Cámara Nacional de Acuacultura
“No quería perder la ocasión para rendir un homenaje de admiración y respeto a los pioneros de nuestro sector y a todos los que tomaron la posta después, y que, con su trabajo esforzado, han logrado que el sector acuicultor se haya convertido en un pilar de la economía ecuatoriana, generador de empleo y divisas para el país”.
Marcelo Vélez Presidente del Directorio de la Cámara Nacional de Acuacultura


El objetivo de Aqua Expo, que se realizó hasta el 13 de julio en el hotel Oro Verde de Machala, fue intercambiar conocimientos y reunir lo mejor de la industria camaronera en un solo lugar. Este evento técnico-comercial camaronero con más de dos décadas de trayectoria fue organizado por la Cámara Nacional de Acuacultura•





“El éxito de la producción acuícola no la darán las normativas ni impedimentos legales sobre el sector productivo en alguna de sus fases, sino la mejora productiva y la eficiencia en sus procesos. He ahí la importancia de eventos como este, donde la temática siempre se adecúa a la realidad que está atravesando el sector”.
Axel
“Es un espacio que propicia el networking, para que puedan venir clientes potenciales y enterarse sobre las mejoras para sus procesos productivos”.

“Es importante realizar este tipo de eventos, porque unen a todos los actores de la cadena y propician que se pueda avanzar más rápido a soluciones más productivas”.
“Aqua Expo El Oro nos ha permitido crear lazos más fuertes con algunos de nuestros clientes y eso es muy importante para nosotros”.
“Esta feria nos ha permitido exponer nuestros productos a nivel nacional e internacional”.

- AGOSTO 2023 COYUNTURA 17
Vedani Subsecretario de Acuacultura
Karen Moreira Expositora Aqua Expo El Oro
Gonzalo Bigeriego Expositor Aqua Expo El Oro
Carlos Merchán Expositor Aqua Expo El Oro
Manuel Martínez Expositor Aqua Expo El Oro
“Para nosotros es de gran beneficio el darnos a conocer como nuevos en este importante mercado”.
“Hemos escuchado a expertos de diversos países, quienes nos han dado tips importantes para que podamos ponerlos en práctica en nuestros predios camaroneros, así como conocer también sobre investigaciones científicas que han dado resultado en otros países”.
“He aprendido más aquí en Aqua Expo que lo que puedo aprender en el campo, porque hay expositores de otros países que nos han compartido sus experiencias de primera mano”.

Kevin Ocaña
Eduardo Heras
Expositor Aqua Expo El Oro
Joan Jaramillo
Asistente al Congreso Aqua Expo El Oro
Asistente al Congreso Aqua Expo El Oro
“Se ha producido un excelente intercambio de conocimientos, basado en la investigación científica que hay sobre la producción camaronera a nivel mundial”.
Estefanía Carvache
Asistente al Congreso Aqua Expo El Oro




- AGOSTO 2023
COYUNTURA

ARTÍCULOS TÉCNICOS ÍNDICE
Edición 154 - Agosto 2023 21 24 38 48 56 60
¿Sabías que lo correcto es denominar al camarón blanco del Pacífico Penaeus vannamei y no Litopenaeus vannamei?.
Evaluación de la transmisión horizontal del virus del síndrome de la Mancha Blanca en el camarón blanco (Litopenaeus vannamei) según el grado de severidad de la enfermedad y la tasa de desprendimiento viral.

Las características físicas y nutricionales de los alimentos microextruidos y su efecto en el rendimiento de larvas de Litopenaeus vannamei.
Acumulación de sólidos, un aspecto crítico en los sistemas acuícolas intensivos: alternativas para una gestión sostenible.
Evolución del Fenómeno de El Niño. Recomendaciones para el sector camaronero. Calidad del agua, calidad de vida y producción.
¿Sabías que lo correcto es denominar al camarón blanco del Pacífico Penaeus vannamei y no Litopenaeus vannamei?
Autores:
Arnaldo Figueredo1,2
Marcos De Donato3 César Lodeiros4
1 Escuela de Ciencias Aplicadas del Mar, Universidad de Oriente, Boca del Río, Isla de Margarita, Venezuela
2 Sociedad Venezolana de Acuicultura, Maracay, Aragua, Venezuela
3 The Center for Aquaculture Technologies, San Diego, CA, USA
4 Facultad de Acuicultura y Ciencias del Mar, Universidad Técnica de Manabí, Bahía de Caráquez, Manabí, Ecuador.
cesar.lodeiros@utm.edu.ec
El camarón blanco del Pacífico es el crustáceo más cultivado a nivel mundial, pero es una especie que tiene un conflicto taxonómico, encontrándose comúnmente en la literatura científica y popular tanto como Penaeus vannamei como Litopenaeus vannamei. Por ejemplo, una encuesta rápida en la base de datos SCOPUS, repositorio de revistas de alto impacto en la literatura científica, realizada recientemente (27 junio de 2023) sobre la frecuencia de aparición de ambos nombres de este camarón en los títulos de artículos científicos, indican 1.118 artículos (99 en lo que va del año) para Penaeus vannamei y 4.314 (211 en el 2023) para Litopenaeus vannamei. De hecho, todo el género Penaeus se encuentra en una controversia que ha durado más de 20 años.

Esta ambigüedad es inapropiada, además de prestarse a confusión o mala interpretación por parte de los lectores de la literatura que
no conocen el desacuerdo taxonómico. Así, nosotros nos planteamos dar respuesta a la pregunta “¿Cómo deberíamos llamar al camarón blanco del Pacífico, Penaeus vannamei o Litopenaeus vannamei?” en un ensayo que apareció en la revista Reviews in Aquaculture a principios de este año 2023 (Figueredo et al. 2023).
Todo comenzó cuando Pérez-Farfante y Kensley (1997) propusieron la reestructuración completa de la familia, elevando los subgéneros propuestos anteriormente por varios investigadores al estatus de géneros, basándose en características morfológicas. Esto desató gran controversia entre investigadores de diferentes especialidades, pero para aquella época la taxonomía molecular no se había desarrollado y los taxónomos aceptaron los argumentos de Pérez-Farfante y Kensley.
Sin embargo, con el transcurrir del tiempo, varios trabajos que utilizaron información genética empezaron a reunir evidencias que demostraban que las diferencias morfológicas no eran suficientes para dividir las especies que antiguamente se agrupaban en el género Penaeus en diferentes géneros. Recientemente, Katneni et al. (2021) realizaron un análisis filogenético de las secuencias completas
- AGOSTO 2023 DENOMINACIÓN
21
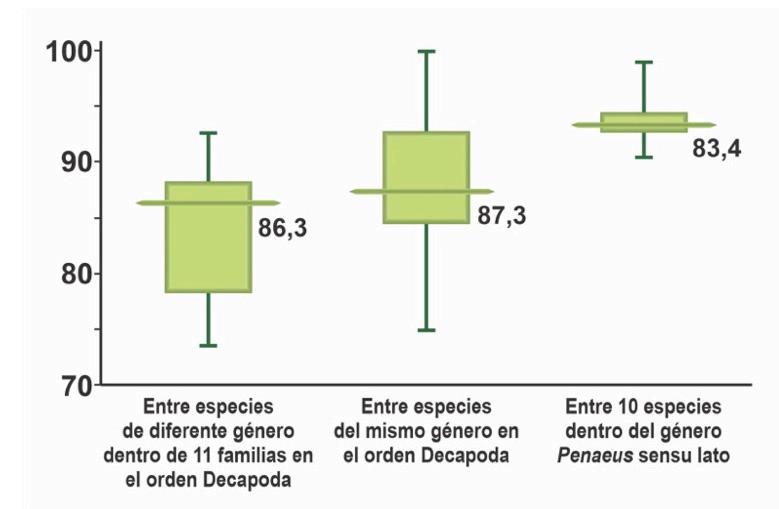
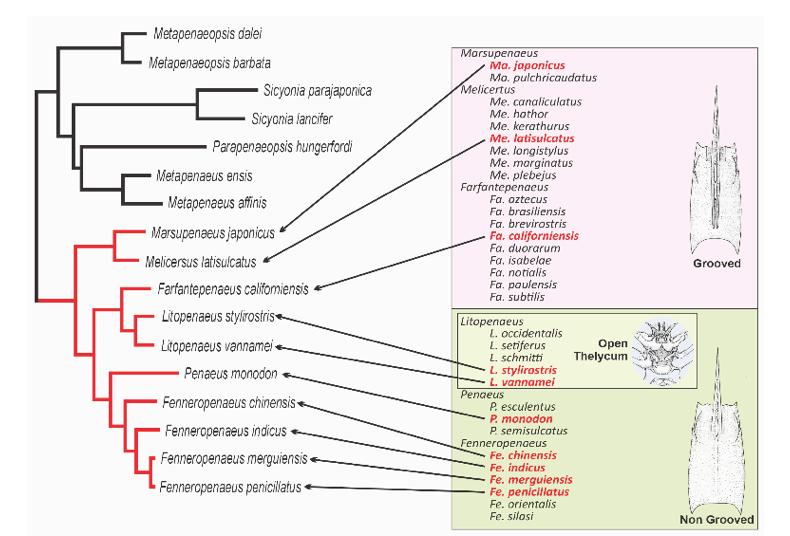
de las mitocondrias de 10 especies de los 6 géneros controversiales, así como de secuencias de otras especies relacionadas al género, demostrando el origen único (monofilético) de todas las especies del género (Fig. 1). Ellos también calcularon las diferencias promedio entre especies a nivel de los aminoácidos de las proteínas, demostrando que éstas eran mucho menores a las diferencias intergenéricas encontradas en otros géneros (Fig. 2).
En conclusión, hoy en día lo más correcto es utilizar Penaeus vannamei (Boone, 1931) como nombre científico del camarón blanco del Pacífico•
Para mayor información escriba a: cesar.lodeiros@utm.edu.ec
Referencias bibliográficas
Figueredo A, Lira C, Vera-Caripe J, De Donato M, lodeiros C. 2023. The Pacific white shrimp, the most cultivated shrimp species, is it Litopenaeus or Penaeus vannamei? Review in Aquaculture. 15(1): 7-13. Peréz-Farfante I, Kensley B. 1997. Penaeoid and Sergestoid Shrimps and Prawns of the World (Keys and Diagnoses for the Families and Genera). Memoires du Muséum National d'Histoire Naturelle, Tome 175 Zoologie. Paris, Francia.

Katneni VK, Shekhar MS, Jangam AK, Paran BC, Selvaraj A, Krishnan K, Kaikkolante N, Prabhudas SK, Gopalapillai G, Koyadan VK. 2021.Phylogenetic relations and mitogenomewide similarity metrics reveal mono-phyly of Penaeus sensu lato. Ecology and Evolution 11(5):2040-9
- AGOSTO 2023 DENOMINACIÓN
Figura 2. Algunos valores de identidad media de aminoácidos relevantes para la filogenia de Penaeus, siguiendo Katneni et al. (2021).
Figura 1. Árbol filogenético modificado a partir del realizado por Katneni et al. (2021), que presenta todas las especies de Penaeus que se muestran en la base de datos de taxonomía del NCBI (Centro Nacional para la Información Biotecnológica de EE.UU).

Evaluación de la transmisión horizontal del virus del Síndrome de la

El virus del síndrome de la mancha blanca (WSSV), que causa la enfermedad de la mancha blanca (WSD), es el patógeno de crustáceos más importante [1,2]. Desde su primer reporte en Taiwán en 1992, el WSSV se ha extendido gradualmente a otras regiones del mundo [3]. Las enfermedades causadas por el WSSV dan como resultado una mortalidad masiva en camarones peneidos dentro de los 10 días posteriores a la infección y causan pérdidas económicas sustanciales en la industria del cultivo de camarón [4,5].
Autores:
Min-Jae Kim1
Jae-Ok Kim2
Gwang-Il Jang3
Mun-Gyeong Kwon3
Kwang-Il Kim1,*
1Departamento de Medicina de Vida Acuática, Universidad Nacional de Pukyong, Busan 48513, República de Corea; manutd5274@pukyong.ac.kr
2Oficina Regional de Tongyeong, Servicio Nacional de Gestión de Calidad de Productos Pesqueros (NFQS), Tongyeong 53019, República de Corea; kimjaeok@korea.kr
3División de Control de Enfermedades Acuáticas, Servicio Nacional de Gestión de Calidad de Productos Pesqueros (NFQS), Busan 49111, República de Corea; gijang2@korea.kr (G.-I.J.); mgkwon@korea.kr (M.-G.K.)
kimki@pknu.ac.kr
Publicación original:
Edorial MDPI https://www.mdpi.com/2076-2615/13/10/1676
Al igual que la mayoría de patógenos en los invertebrados, la ruta de transmisión a través del agua es uno de los mecanismos de transmisión horizontal más importantes para el WSSV, junto con el canibalismo [6]. Por lo tanto, comprender el modelo de transmisión a través del agua es crucial para prevenir el brote y la propagación de enfermedades [7]. En particular, el desprendimiento viral (viral shedding) y la dosis de infección mínima son parámetros importantes para estimar y predecir las cargas virales en el agua de mar, determinar los niveles de exposición para individuos “ingenuos” (no infectados), y comprender el mecanismo de transmisión de enfermedades de animales acuáticos a través del agua [8,9]. Varios estudios han investigado la transmisión del virus de la septicemia hemorrágica viral (VHSV), el virus de la necrosis hematopoyética infecciosa, el virus de la necrosis nerviosa y el iridovirus del mar Rojo a través del agua en función de la tasa de desprendimiento viral de los peces infectados y la dosis de infección mínima del virus [7,10-12]. Durand y Lightner [13] determinaron la dosis infectiva mínima del WSSV a través de ensayos de inmersión de 4 h y reportaron que las cargas virales por debajo de 105 copias del genoma del WSSV/mL son insuficientes para inducir una infección en el camarón blanco (Litopenaeus vannamei). Por el contrario, Kumar et al. [14] y Qayoom et al. [15] reportaron que la exposición continua a 103 copias del genoma del WSSV/mL en agua de mar es suficiente para inducir una infección. Debido a la ausencia de una línea celular específica, es difícil determinar la dosis infectiva mínima del WSSV en condiciones naturales [16].
Los grados de severidad de las enfermedades infecciosas brindan información importante
- AGOSTO 2023 PATOLOGÍA
Mancha Blanca en el camarón blanco (Litopenaeus vannamei) según el grado de severidad de la enfermedad y la tasa de desprendimiento viral
sobre el potencial impacto de la enfermedad en las camaroneras. Lightner [17] estableció el grado de severidad del WSSV basado en la proporción (%) de cuerpos de inclusión intracelular determinados mediante análisis histopatológico. Aunque los hallazgos histopatológicos pueden reflejar el estado patológico de las células huésped, la clasificación histopatológica no es mayormente cuantitativa y puede variar entre las diferentes técnicas de diagnóstico [18]. Varios estudios han investigado la gravedad de la infección por WSSV mediante análisis cuantitativos, como los ensayos competitivos [18] y de reacción en cadena de la polimerasa (PCR) en tiempo real basados en el sistema SYBR green [19-21]. Aunque varios estudios han establecido el grado de severidad del WSSV e investigado la dosis infectiva mínima a través de ensayos de inmersión, ningún estudio ha investigado la transmisión a través del agua en función de la correlación entre el grado de severidad de la enfermedad y la tasa de desprendimiento viral.
En este estudio, investigamos las correlaciones entre los cambios clínicos, el grado de severidad de la enfermedad y las copias virales de camarones pleópodos infectados con WSSV utilizando camarón blanco artificialmente infectado con WSSV a diferentes temperaturas. Para determinar la dosis infectiva mínima del WSSV por vía acuosa, realizamos pruebas de inmersión a diferentes dosis de administración y períodos de exposición. Además, verificamos la dosis infectiva mínima del WSSV y su dinámica de transmisión a través del agua en función de las cargas virales en el camarón y agua de mar a través del ensayo de cohabitación.
Materiales y métodos Camarón y virus
Se obtuvieron camarones blancos juveniles (2.03 ± 0.85 g) de una granja en Geoje, Corea, y se confirmó que estaban libres del WSSV mediante PCR anidada, como lo describieron previamente Lo et al. [22] (Tabla S1). Los camarones se aclimataron en un tanque de 250 L a 25±0.5°C durante 1 semana y se alimentaron con una dieta comercial una vez al día. El virus utilizado en este estudio se extrajo de camarones blancos enfermos en Taean, Corea, en 2014 (WSSV-Te-14 aislado) y se purificó como se describió anteriormente
[23]. Brevemente, el tejido positivo para WSSV (pleópodos, branquias y músculos) se homogeneizó en una solución 1:10 de solución salina tamponada con fosfato (PBS). El tejido homogeneizado se centrifugó a 3000×g durante 30 min y el sobrenadante se almacenó a -80°C después del proceso de filtración. Para preservar la viabilidad del virus, el WSSV se purificó y propagó mediante inyección en camarones libres del WSSV antes de usarse en los experimentos (dentro de los 7 días antes de la infección experimental).
Determinación de copias del genoma del WSSV en camarón blanco
Para determinar las copias del genoma del WSSV en el camarón blanco, se extrajo el ADN total de 10 mg de tejido pleópodo utilizando un kit de preparación de plantillas de PCR de alta pureza (Roche). Para el análisis cuantitativo, se realizó PCR en tiempo real de acuerdo con los métodos descritos por Durand y Lightner [13]. Brevemente, cada mezcla de 20 μL de reacción en cadena de polimerasa cuantitativa en tiempo real contenía 1 μL de ADN, que se extrajo con el kit de preparación de plantillas de PCR High Pure (Roche, Mannheim, Alemania), 300 nM de primers reverso e inverso, 150 nM de sonda (Tabla S1), 10 μL de TaqManTM Universal Master Mix II con UNG (Thermo Fisher Scientific, Waltham, MA, EE.UU.) y 7.5 μL de agua libre de nucleasas. La amplificación se realizó utilizando el sistema de PCR en tiempo real StepOne (Applied Biosystems, Foster City, CA, EE.UU.). Las condiciones de ciclado fueron las siguientes: 50°C por 2 min y 95°C por 10 min, seguido de 40 ciclos a 95°C por 15 s (desnaturalización) y 60°C por 1 min (hibridación y extensión). La data cuantitativa por debajo del valor de corte se determinó por el LOD95% (límite inferior del nivel de confianza: 7.24 copias del genoma del WSSV/μL) utilizando ADN plasmídico codificado por el WSSV diluido en serie, se excluyeron del análisis en este estudio de acuerdo con nuestro estudio anterior [23]. Las copias del genoma viral por miligramo de pleópodo se determinaron utilizando la siguiente fórmula: Copias del genoma del WSSV/mg = (copias del genoma del WSSV por μL de ADN total extraído) × (volumen de ADN total extraído) Peso de los tejidos de pleópodos utilizados para extraer el ADN total (mg)
2.3. Ensayo de floculación de hierro para determinar la concentración del WSSV en el agua de mar
Para concentrar las partículas del WSSV en el agua de mar de cultivo, se realizó el método PC-ascorbato en el ensayo de floculación de hierro de acuerdo con un estudio previo [23]. Brevemente, el agua de mar de cultivo se filtró previamente usando un filtro de microfibra de vidrio (tamaño de poro, 1.6 μm; Whatman, Maidstone, Reino Unido) y un filtro de membrana MF-MilliporeTM (tamaño de poro, 0.45 μm; Merck, Darmstadt, Alemania) mediante una bomba de vacío (Gast, Benton Harbor, MI, EE.UU.). A continuación, se añadieron 50 μL de solución de FeCl3 (4.83 g por 100 mL de agua destilada de FeCl3·6H2O; 4.83 μg/mL de concentración final de FeCl3 en el agua de mar) al agua de mar de cultivo prefiltrada (500 mL) y se disolvió usando un agitador magnético (<120 rpm; Sigma-Aldrich, St. Louis, MO, EE.UU.) durante 1 hora a temperatura ambiente (aproximadamente 25°C). Los floculados de Fe-WSSV se recolectaron filtrando a través de una membrana de policarbonato (tamaño de poro: 1.0 μm; Whatman) utilizando una bomba peristáltica a <15 psi (Eyela, Tokio, Japón). Luego, la membrana como floculado de Fe-WSSV se transfirió a un tubo de fondo redondo de 5 mL, seguido de la adición de 1 mL de tampón de elución (0.1 M Tris-0.1 M ácido etilendiaminotetraacético 6H2O0.2 M MgCl2 · 6H2O-0.2 M L -ascorbato, pH 6.0; Sigma-Aldrich). Finalmente, la resuspensión viral se realizó durante la noche (aproximadamente 20 h) a través de un Bio RS-24 Mini-Rotator (30 rpm; Biosan, Riga, Latvia) en un cuarto oscuro a 4°C durante la noche. Para determinar el número de copias del genoma viral en el concentrado de agua de cultivo, se extrajo el ADN total de 200 μL del tampón viral resuspendido usando un kit de preparación de plantillas de PCR High Pure (Roche), y se realizó un análisis cuantitativo usando PCR en tiempo real como se describe anterior (Sección 2.2). La tasa de desprendimiento viral en camarones infectados con el WSSV se presenta como copias del genoma viral/L/g/día.
- AGOSTO 2023 25 PATOLOGÍA
Correlación entre el grado de severidad de la enfermedad y la tasa de desprendimiento viral en camarones infectados con el WSSV
Ensayo de inyección intramuscular
En el experimento 1 (Exp. 1), la infección experimental de camarones se realizó mediante la administración de diferentes dosis. Las dosis administradas se seleccionaron de acuerdo con la patogenicidad del WSSV en juveniles de camarón blanco (105 copias del genoma del WSSV/camarón: 100% de mortalidad inducida en 52 h; 103 copias del genoma del WSSV/camarón: infección inducida o estado moribundo en el plazo de 52 h; 101 copias del genoma del WSSV/camarón: dosis desconocida para inducir infección o mortalidad) [13,23]. Se crio un total de 120 camarones juveniles en 4 tanques (n = 30) y se les administró por vía intramuscular 0.1 ml del WSSV-Te-14 diluido en diferentes dosis (105, 103 o 101 copias del genoma del WSSV/camarón). Como control negativo, a los camarones sanos (n = 30) se les administró por vía intramuscular 0.1 ml de tampón fosfato salino (PBS; Sigma-Aldrich).
A partir de entonces, los camarones se mantuvieron a 25°C en tanques de 50 L (191.4 individuos/m2) durante 14 días para observar la mortalidad acumulada, y se recolectó 500 mL de agua de mar del cultivo (por triplicado) a los 1 y 3 días posteriores a la infección (dpi). Para obtener muestras de evolución temporal, se recolectó un total de 180 camarones de 6 tanques (n = 30 por grupo y días de muestreo) a los 1 y 3 días posteriores a la infección (dpi) en condiciones experimentales idénticas.
En el experimento 2 (Exp. 2), se llevó a cabo una prueba de ensayo a diferentes condiciones de temperatura (constante [20°C], constante [30°C], cambio ascendente [20 a 30°C] y cambio descendente [30 a 20°C]). Un total de 160 camarones juveniles (n = 20 por grupo) se aclimataron a 20 o 30°C durante 1 semana en 8 tanques de agua de 20 L y se les administró por vía intramuscular 0.1 ml del WSSV-Te-14 diluido (103 copias del genoma del WSSV/camarón). Como control negativo, a los camarones sanos (n = 20) se les administró por vía intramuscular 0.1 ml de PBS. Para el grupo de cambio ascendente (20 a 30°C), la temperatura del agua de mar del cultivo se incrementó en 1°C cada 12 h
desde 1 dpi hasta alcanzar los 30°C a los 7 dpi. Por el contrario, para el grupo de cambio descendente (30 a 20°C), la temperatura del agua de mar del cultivo se redujo en 1°C cada 12 h desde 1 dpi hasta alcanzar los 20°C a los 7 dpi. La temperatura del agua de mar del cultivo para los grupos constantes (20°C y 30°C) se mantuvo durante todo el experimento. Después de la infección experimental, los camarones se mantuvieron en tanques de 20 L (200.0 individuos/m2) durante 14 días para observar la mortalidad acumulada, y se recolectaron 500 mL de agua de mar del cultivo (por triplicado) a 1, 2 y 4 dpi. Para obtener muestras de evolución temporal, se recolectó un total de 240 camarones en 12 tanques (n = 20 por grupo y días de muestreo) a 1, 2 y 4 dpi bajo condiciones experimentales idénticas.
Durante los Exp. 1 y 2, el agua de cultivo se reemplazó una vez cada dos días. Para el análisis cuantitativo, las partículas del WSSV en el agua de mar del cultivo se concentraron utilizando un ensayo de floculación de hierro como se describe anteriormente (Sección 2.3). Las copias virales en camarones puestos a prueba y las partículas del WSSV concentradas se determinaron usando la PCR en tiempo real como se describe en la Sección 2.2. Los resultados de la PCR en tiempo real en los grupos de mortalidad acumulada se usaron para analizar el valor de corte para la mortalidad como se describe en la Sección 2.6. El breve diagrama esquemático de los ensayos realizados en este estudio se presenta en la Figura S1.
Análisis histopatológico y determinación del grado de severidad de la enfermedad Para el análisis histopatológico, los tejidos del epitelio subcuticular, las branquias, el hepatopáncreas, el intestino anterior y los tejidos de los órganos linfoides infectados con el WSSV se procesaron como se describió anteriormente [24]. Brevemente,
los camarones infectados con el WSSV de las muestras de evolución temporal en el Exp. 1 (n = 3 por grupo) y Exp. 2 (n = 5 por grupo) fueron seleccionados al azar y sus cefalotórax fueron fijados en solución de Davidson (Cancer Diagnostics Inc., Durham, NC, EE.UU.) durante 24 h. Luego, las muestras se deshidrataron en una serie graduada de alcohol (70-100%) y se embebieron en parafina. Las secciones de tejido (4 μm de espesor) se tiñeron con hematoxilina y eosina (H&E; Abbey Color, Filadelfia, PS, EE.UU.). Para el control negativo, se administró 0.1 ml de camarones con PBS en el Exp. 1 (n = 3 por grupo) y el Exp. 2 (n = 5 por grupo) se fijaron, procesaron, embebieron, seccionaron y se tiñeron con H&E, de manera similar a los grupos experimentales. El análisis histopatológico y las imágenes se obtuvieron utilizando un microscopio óptico (BX50; Olympus Ltd., Tokio, Japón).
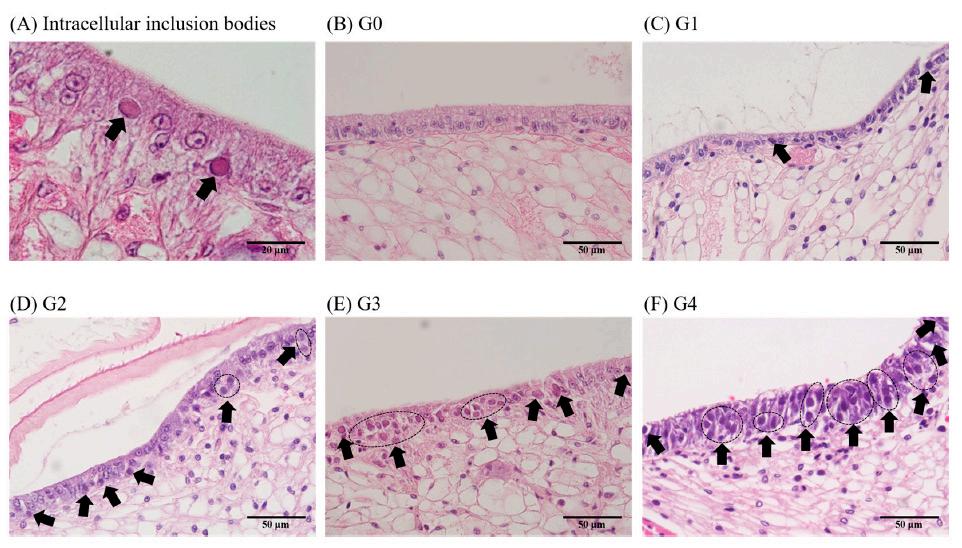
La severidad de la infección por el WSSV en los camarones se determinó evaluando el porcentaje de cuerpos de inclusión intracelulares observados a través del análisis histopatológico [17]. En resumen, las infecciones con una proporción de cuerpos de inclusión de menos del 10% del total de células se clasificaron como G1, 30–40% como G2, 40–50% como G3 y más del 80% como G4. Cualquier resultado negativo fue etiquetado como G0 (Tabla 1) (Figura 1). Se utilizó un total de 164 camarones y 6 secciones por individuo (2 bloques de parafina con 3 secciones por bloque) para evaluar los grados de severidad, y los grados de severidad se determinaron con base en el puntaje promedio de las 6 secciones.
Análisis de expresión génica del WSSV VP28
Para comparar los niveles de propagación del WSSV a diferentes temperaturas, se analizó la expresión del gen VP28 de WSSV utilizando un sistema de PCR en tiempo real
Grados de severidad de la infección por WSSV
Criterios de categorización
Todos los tejidos son normales sin ningún signo.
Los cuerpos de inclusión intracelulares se pueden ver en menos del 10% Los cuerpos de inclusión intracelulares se pueden ver en menos del 30-40% Los cuerpos de inclusión intracelulares se pueden ver en menos del 40-50% Se pueden ver cuerpos de inclusión intracelulares en más del 80%
- AGOSTO 2023 26
PATOLOGÍA
G0 G1 G2 G3 G4
Tabla 1. Categorización de grados de severidad de WSSV (Lightner [17])
StepOne (Applied Biosystems) de acuerdo con estudios previos [25,26]. Brevemente, el ARN total se extrajo de 50 mg de tejido muscular de los camarones muestreados a lo largo del tiempo (Exp. 2; n = 5 por grupo) utilizando el minikit de extracción de ARN total yesRTM (GenesGen, Busan, República de Corea) y el ARN se transcribió inversamente utilizando el 1st ADNc PrimeScriptTM Synthesis Kit (Takara, Japón) siguiendo los protocolos del fabricante. La diferencia de veces de la expresión del gen VP28 en relación con la β-actina usando conjuntos de primers específicos (Tabla S1) se determinó mediante el método 2−∆∆Ct como lo describen Rao et al. [27].
Transmisión del WSSV a través del agua bajo las condiciones naturales imitadas
Determinación de la dosis infecciosa mínima por agua
En los Exp. 3 y 4, para investigar la dosis infectiva mínima del WSSV a través del agua, se realizaron ensayos de inmersión a diferentes dosis y períodos de exposición. Las dosis administradas se seleccionaron con base en estudios previos (105 copias del genoma del WSSV/mL: infección inducida por ensayo de inmersión; 103 copias del genoma del WSSV/mL: infección inducida por exposición continua; 101 copias del genoma del WSSV/mL: cargas virales en el efluente de la camaronera con brote del WSSV) [13,14,23].
En el Exp. 3, se criaron 80 camarones juveniles en cuatro tanques de 20 L (n = 20) y se sumergieron en WSSV en tres dosis diferentes (105, 103 o 101 copias del genoma del WSSV/mL) con agua de mar esterilizada con UV a 25°C durante 24 h, y se recambió el 100% del agua de cultivo. Como control negativo, se sumergieron camarones sanos (n = 20) en agua de mar enriquecida con PBS. A los 1, 3, 5, 7, 9 y 11 ppp, se recogió 500 ml de agua de mar de cultivo (por triplicado) para investigar la tasa de desprendimiento viral de los camarones infectados. Bajo las mismas condiciones experimentales, se recolectaron pleópodos de camarón blanco (n = 3 por grupo) a 1, 3, 5, 7, 9 y 11 dpi para investigar el curso temporal de las copias virales.
En el Exp. 4, se criaron 60 camarones
juveniles en tres tanques de 20 L (n = 20) y se sumergieron en WSSV a dos dosis diferentes (103 o 101 copias del genoma del WSSV/mL) con agua de mar esterilizada con UV a 25°C durante 14 días. Como control negativo, se sumergieron camarones sanos (n = 20) en agua de mar enriquecida con PBS. Bajo las mismas condiciones experimentales, se recolectaron pleópodos de camarón blanco (n = 3 por grupo) a 1, 3, 5, 7, 9 y 11 dpi para confirmar la infección.
Para los ensayos de inmersión, se diluyó 0.1 ml de solución del WSSV-Te-14 en series de 10 veces en agua de mar esterilizada con UV para obtener la dosis final de ensayo con un volumen de 10 L, y los grupos control negativo se sumergieron en el mismo volumen de agua de mar enriquecida con PBS. Después de los ensayos de inmersión, los camarones se mantuvieron a 25°C en tanques de 20 L (200,0 individuos por m2) durante 14 días para observar la mortalidad acumulada. Durante el experimento, el agua del cultivo se reemplazó una vez cada dos días. En el Exp. 3, se usó agua de mar esterilizada con UV como reemplazo, mientras que en el Exp. 4, se usó agua de mar enriquecida con WSSV a la misma dosis de inmersión inicial. Para el análisis cuantitativo, las partículas del WSSV en el agua de mar del cultivo se concentraron utilizando el ensayo de floculación de hierro como se describe anteriormente (Sección 2.3). Las copias virales en el camarón puesto a prueba y las partículas del WSSV
concentradas, se determinaron usando PCR en tiempo real como se describe en la Sección 2.2.
Ensayo de convivencia
En el Exp. 5, para verificar la tasa de desprendimiento viral de los camarones infectados y las dosis infectivas mínimas del WSSV, se realizó un ensayo de cohabitación a diferentes dosis del WSSV y temperaturas del agua. Se aclimató un total de 180 camarones juveniles (n = 20 por grupo; donante) en tanques de agua de 9 50 L a 20, 25 o 30°C durante 1 semana y se les administró por vía intramuscular 0.1 mL del WSSV-Te-14 diluido a diferentes dosis (105 o 103 copias del genoma WSSV/camarón). Como control negativo, a los camarones sanos (n = 20) se les administró por vía intramuscular 0.1 ml de PBS. A los 3 ppp, se instalaron jaulas de plástico en cada tanque para albergar los camarones sanos.
Un total de 180 camarones sanos (n = 20 por grupo; receptores) se alojaron en jaulas de plástico en cada tanque y se mantuvieron durante 14 días (n = 40 por grupo; 250.0 individuos por m2) para observar la mortalidad acumulada. Durante el experimento, el 50% del agua del cultivo se reemplazó una vez cada dos días. Bajo las mismas condiciones experimentales, se recolectaron pleópodos de los recipientes (n = 3 por grupo) y 500 mL de agua de mar
- AGOSTO 2023 27 PATOLOGÍA
Figura 1. El grado de severidad del virus del síndrome de la mancha blanca se determinó en función de la proporción de cuerpos de inclusión (flecha) en las células del epitelio subcuticular mediante análisis histopatológico.
(A) Cuerpos de inclusión intracelulares
del cultivo (por triplicado) a los 3, 5, 7, 9 y 11 dpi. Se recolectó agua de mar de los tanques del grupo experimental para medir la mortalidad. Para el análisis cuantitativo, las partículas del WSSV en el agua de mar del cultivo se concentraron utilizando un ensayo de floculación de hierro como se describe anteriormente (Sección 2.3). Las copias virales en camarones del ensayo y las partículas del WSSV concentradas se determinaron usando PCR en tiempo real como se describe en la Sección 2.2.
Análisis estadístico
La significación estadística de las copias virales en pleópodos, agua de mar del cultivo y la expresión del gen VP28 entre los grupos administrados se determinó mediante ANOVA de dos vías. Se realizó un análisis de regresión lineal para verificar la correlación entre las copias virales de los pleópodos y la tasa de desprendimiento viral. Los valores de corte para el desprendimiento viral y la mortalidad de los camarones infectados con WSSV se determinaron mediante el análisis de la curva ROC. Los análisis estadísticos se realizaron utilizando SPSS ver. 27 (IBM) y la significación se fijó en p < 0.05.
Resultados
Correlación entre copias virales de pleópodos y tasa de desprendimiento viral
Mortalidad acumulada en grupos expuestos a inyecciones del ensayo
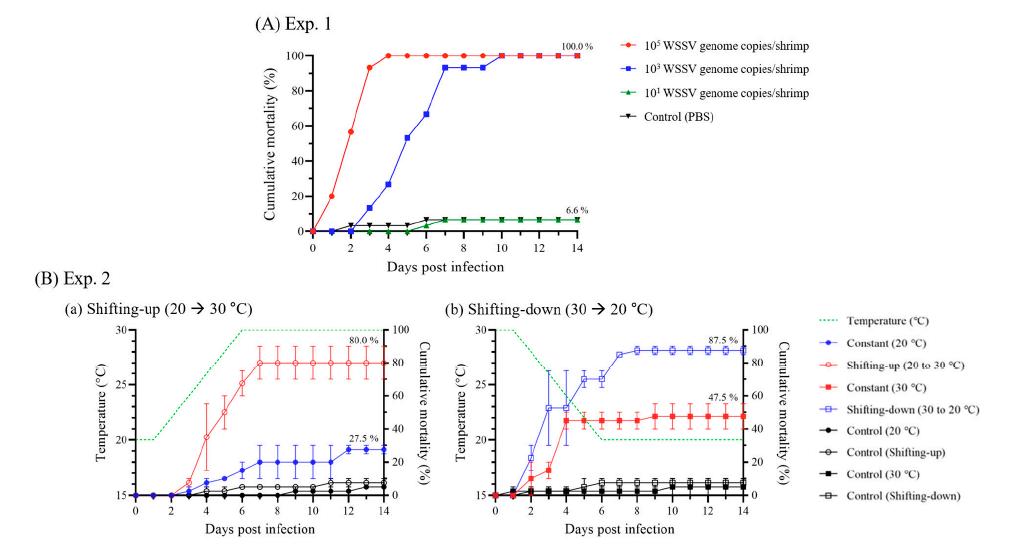
En el Exp. 1, a diferentes dosis de administración, los camarones a los que se administraron 105 y 103 copias del genoma del WSSV comenzaron a morir a 1 y 3 ppp, respectivamente, y exhibieron una mortalidad acumulada del 100.0 % a los 4 y 10 ppp, respectivamente. El grupo experimental al que se le inyectó 101 copias del genoma del WSSV/camarón exhibió una mortalidad del 6.6%, que fue la misma que la del grupo control negativo (6.6% de mortalidad), y el genoma viral no pudo identificarse mediante PCR en tiempo real (Fig. 2A).
En el Exp. 2, la infección experimental se realizó a 20°C (constante [20°C] y cambio ascendente [20 a 30°C]) y 30°C (constante [30°C] y cambio descendente [30 a 20°C]).
Si bien la mortalidad inicial de los camarones se observó en función de la temperatura administrada (20°C: 3 dpi; 30°C: 2 dpi),
Figura 2. Mortalidad acumulada por infección experimental del WSSV durante 14 días. (A) Experimento 1 (n = 30/grupo) se realizó una inyección intramuscular usando 105, 103 o 101 copias del genoma del virus del síndrome de la mancha blanca (WSSV)/camarón a 25°C. (B) Experimento 2 (n = 20/ grupo) se realizó una inyección intramuscular con 103 copias del genoma WSSV/camarón en cuatro condiciones de temperatura diferentes: constante (20°C), aumento (20 a 30°C), constante (30°C), y cambio descendente (30 a 20°C), con cada condición probada por duplicado. Las líneas punteadas indicaron la temperatura del agua cada día, y el cambio de temperatura se realizó a razón de 1°C/12 h. A los grupos de control negativo en los experimentos 1 y 2 se les inyectó solución salina tamponada con fosfato (PBS).
las tasas de mortalidad acumulada se mostraron de manera diferente según los cambios de temperatura (Fig. 2B). Las tasas de mortalidad acumuladas de los grupos constante (20°C) y de cambio ascendente fueron 27.5 y 80.0%, respectivamente, mientras que las de los grupos constante (30°C) y de cambio descendente fueron 47.5% y 87.5%, respectivamente (Fig. 2B).
Tasa de desprendimiento viral en grupos expuestos a las inyecciones Para comparar la tasa de desprendimiento viral entre los grupos expuestos a la inyección, las copias virales en el pleópodo se presentan como media (rango Inter cuartil [IQR]) y la tasa de desprendimiento viral se presenta como media ± desviación estándar (DE).
En el Exp. 1, las copias virales de pleópodos de los camarones aumentaron con el tiempo (1 y 3 dpi), con diferencias significativas según la dosis inicial administrada. La mediana de las copias del genoma viral del pleópodo después de la inyección con 105 y 103 copias del genoma WSSV/camarón fue 3.1 × 105 (IQR, 1.5 × 105 a 7.3 × 105) y 1.2 × 103 (1.5 × 102 a 7.6 × 103) copias del genoma WSSV/mg a 1 ppp, y 2.0 × 107
(1.2 × 107 a 2.9 × 107) y 1.2 × 106 (5.4 × 105 a 1.7 × 106) copias del genoma de WSSV/ mg a 3 ppp, respectivamente. Mientras tanto, las copias virales en los camarones muertos no fueron significativamente diferentes, independientemente de la dosis administrada. La tasa de desprendimiento viral también aumentó, con una tendencia similar a la de las copias virales de pleópodos, con una diferencia significativa según la dosis inicial administrada (Fig. 3A). La tasa media de desprendimiento viral de los camarones administrados con 105 y 103 copias del genoma de WSSV/camarón fue de 1.7 × 104 (SD, ±9.6 × 103) y 2.6 × 102 (±1.3 × 102) copias del genoma de WSSV/L/g a 1 ppp, y 3.8 × 106 (±1.9 × 106) y 3.1 × 105 (±1.8 × 105) copias del genoma WSSV/L/g a 3 ppp, respectivamente (Figura 3A).
En el Exp. 2, las copias virales de pleópodos de los camarones aumentaron con el tiempo (1, 2 y 4 ppp), con diferencias significativas según el cambio de temperatura (Fig. 3B). La mediana de las copias virales de pleópodos de camarón administradas a 20°C (constante [20°C] y ascendente) exhibieron diferencias significativas a 4 dpi, con 5.8 × 103 (IQR, 2.7 × 103 a 9.0 × 103) y 4.4 × 105 (1.4 × 105 a 1.3 × 106) copias del genoma del WSSV/mg.
- AGOSTO 2023 28 PATOLOGÍA
Mortalidad acumulada (%) Temperatura (°C) Temperatura (°C) Días post infección Cambio ascendente Cambio descendiente Días post infección Días post infección Mortalidad acumulada (%) Mortalidad acumulada (%)
Además, para los camarones administrados a 30°C (constante [30°C] y descendente), la mediana de las copias del genoma viral de pleópodo mostró diferencias significativas tanto a 2 como a 4 ppp, con 3.9 × 103 (3.5 × 102 a 1.8 × 104) y 6.0 × 104 (2.9 × 103 a 5.1 × 105) copias/mg del genoma de WSSV a 2 ppp y 3.5 × 104 (3.1 × 103 a 8.4 × 105) y 1.9 × 106 (2.1 × 105 a 4.9 × 107) copias del genoma del WSSV/mg a 4 ppp. Sin embargo, las copias virales en los camarones muertos no mostraron diferencias significativas, con medianas de copias virales de pleópodos superiores a 106 copias del genoma WSSV/ mg, independientemente de la temperatura del agua (Fig. 3B).
La tasa de desprendimiento viral también aumentó con una tendencia similar a la de las copias virales en pleópodos, y hubo una diferencia significativa según las condiciones de temperatura del agua. La tasa media de desprendimiento viral de los camarones administrados a 20°C (constante [20°C] y en aumento) mostró diferencias significativas a 4 dpi, con 2.9 × 103 (SD, ±2.1 × 103) y 2.0 × 106 (± 8.3 × 105) copias del genoma del WSSV/L/g, respectivamente (Fig. 4A). Además, para los camarones administrados a 30°C (constante [30°C] y decreciente), la mediana de las copias del genoma viral de los pleópodos mostró diferencias significativas tanto a 2 como a 4 ppp, con 5.4 × 103 (±2.5 × 103) y 5,2 × 105 (±3.2 × 105) copias del genoma de WSSV/l/g a 2 ppp y 1.6 × 105 (±1.2 × 105) y 4.5 × 107 (±2.7 × 107) copias del genoma de WSSV/l/g a 4 ppp, respectivamente. En particular, el desprendimiento viral no se detectó o estuvo por debajo del límite de detección a 1 ppp, independientemente de la temperatura del agua (Fig. 4A).
Niveles de propagación del WSSV en camarón de acuerdo con el transcurso del tiempo
En el Exp. 2, la viabilidad del WSSV en camarones (n = 5) se determinó midiendo la expresión relativa de VP28, que se normalizó del gen constitutivo β-actina (Fig. 4B). A 1 ppp, no hubo diferencias significativas en la expresión de VP28 entre los grupos, independientemente de la temperatura del agua. El grupo de cambio descendente mostró una regulación positiva significativa de la expresión del gen VP28 en comparación
Figura 3. Copias del genoma viral del pleópodo después de la inyección intramuscular en los experimentos 1 y 2. (A) En el experimento 1, a los camarones (n = 30/grupo) se les inyectó por vía intramuscular 105, 103 o 101 copias del genoma/camarón del virus del síndrome de la mancha blanca (WSSV) a 25°C. Copias del genoma viral del pleópodo recolectadas en diferentes intervalos de tiempo (1 y 3 días después de la infección y después de la muerte). (B) En el experimento 2, camarones (n = 20/grupo) recibieron una inyección intramuscular con 103 copias del genoma WSSV/ camarón en cuatro condiciones de temperatura diferentes (20°C: cambio ascendente; 30°◦C: cambio descendiente) a diferentes intervalos de tiempo (1, 2 y 4 d post infección, individuos muertos y sobrevivientes). Las barras horizontales indican el valor de la mediana de cada grupo. La línea punteada horizontal indica el valor LOD95% de este estudio, y U.D. indica indeterminado. Se realizó un análisis de varianza bidireccional para determinar las diferencias significativas en las copias del genoma viral de los pleópodos entre los grupos experimentales a p < 0.05 (*).
con los otros grupos a 2 ppp. Cabe destacar que la expresión relativa del gen VP28 se reguló significativamente tanto en los grupos de cambio ascendente como descendente en comparación con cada uno de los grupos constantes (20°C) y constantes (30°C), respectivamente, que fueron administrado a las mismas temperaturas (Fig. 4B).
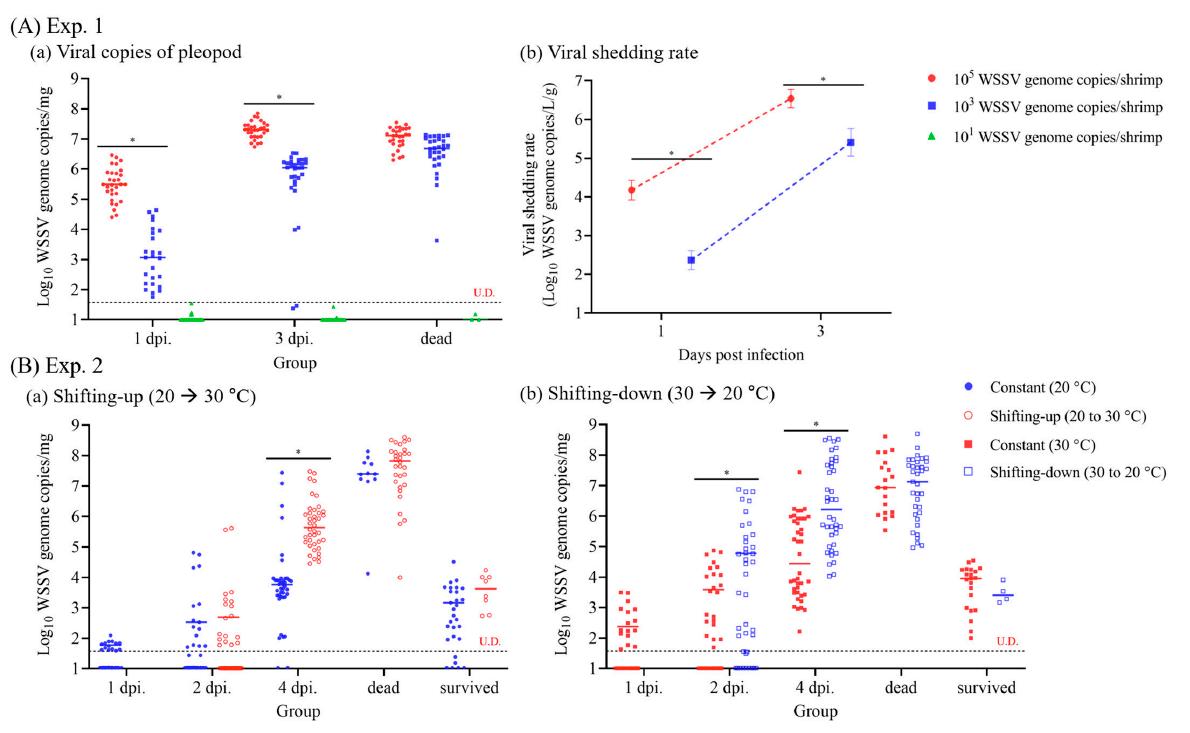
Correlación lineal entre copias virales de pleópodos y tasa de desprendimiento viral Basado en las tasas de desprendimiento viral en los Exp. 1 y 2, se llevó a cabo un análisis de regresión lineal para examinar la correlación entre las copias virales de pleópodos en camarones y la tasa de desprendimiento viral. El análisis reveló una correlación positiva (y = 0.7076x + 1.414) (p < 0.001) como se muestra en la Figura 4C.
Grado de severidad del WSSV y análisis cuantitativo
Grado de severidad de la enfermedad en camarones infectados con WSSV
Con base en los hallazgos histopatológicos, el grado de severidad de los camarones infectados con WSSV se determinó de acuerdo con Lightner [17], y los grados de severidad con el transcurso del tiempo se presentan en las Tablas 2 y 3.
En el Exp. 1, el grado de severidad de los camarones infectados con WSSV inyectados con 105 y 103 copias del genoma WSSV/camarón fueron G2 y G1–G2, respectivamente, a 1 ppp. A los 3 ppp, el grado de severidad aumentó a G3–G4 y G2–G3 en camarones inyectados con 105 y 103 copias del genoma WSSV, respectivamente. En ausencia de una infección inducida, no se observaron núcleos hipertróficos o lesiones patológicas en los camarones después de la inyección de 101 copias del genoma WSSV (G0) (Tabla 2).
- AGOSTO 2023 29 PATOLOGÍA
Copias
Grupo Grupo Grupo Tasa de eliminación viral Días post infección Cambio ascendente Cambio descendiente Log 10 copias del genoma del WSSV/mg Log 10 copias del genoma del WSSV/mg Log 10 copias del genoma del WSSV/mg Log 10 copias del genoma del WSSV/L/g)
virales de pleópodos
(A) Tasa de desprendimiento viral
Correlación lineal entre copias virales de pleópodos y tasa de desprendimiento viral
(B) Expresión del gen VP28 del WSSV
Figura 4. Correlación entre las copias virales de los pleópodos y la tasa de desprendimiento viral determinada a lo largo del experimento 1 y el experimento 2. (A) Tasa de desprendimiento viral (copias del genoma del WSSV/L/g) en el Exp. 2 se determinó en función del número de copias del genoma viral en agua de mar de cultivo y el peso de los camarones restantes. (B) Expresión del gen VP28 del WSSV en el Exp. 2 se determinó mediante la diferencia de veces en la expresión del gen VP28 en relación con la β-actina mediante el método 2−∆∆Ct. (C) La correlación entre las copias virales de los pleópodos y la tasa de desprendimiento viral se determinó mediante un análisis de regresión lineal utilizando los resultados de las copias virales de los pleópodos y la tasa de desprendimiento viral en los experimentos 1 y 2. La línea punteada horizontal indica el valor LOD95% de este estudio, y U.D. indica indeterminado. Se realizó un análisis de varianza bidireccional para determinar las diferencias significativas en la tasa de desprendimiento viral y la expresión del gen VP28 entre los grupos experimentales a p < 0.05.
En el Exp. 2, el grado de severidad de los camarones infectados con WSSV diferían según las condiciones de temperatura. A 1 ppp, el grupo constante (20°C) mostró G0, mientras que el grupo constante (30°C) mostró G0–G1. Aunque los dos grupos que se administraron a 20°C (constante [20°C] y ascendente) mostraron niveles de severidad G1-G2 a 2 dpi, debido al cambio de temperatura, los grupos constantes (20°C) y en aumento mostraron G0–G1 y G2–G3, respectivamente, a 4 ppp. Además, para los camarones administrados a 30°C (constante [30°C] y descendiente) mostraron diferentes grados de severidad tanto a 2 como a 4 dpi, con G0–G2 y G1–G2 a 2 dpi y G1–G2 y G3 –Grupos G4 a 4 dpi, respectivamente. Mientras tanto, los camarones sobrevivientes en cada grupo mostraron grados de severidad G0-G2, independientemente de las condiciones de temperatura del agua (Tabla 3).
Umbral de cambios clínicos en camarones infectados con WSSV
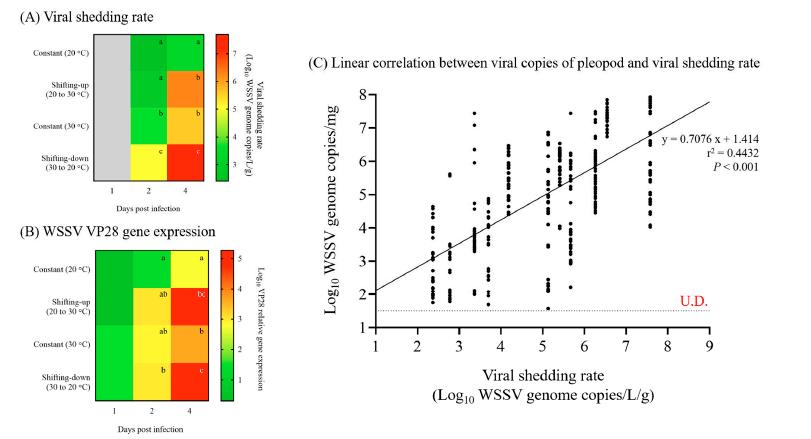
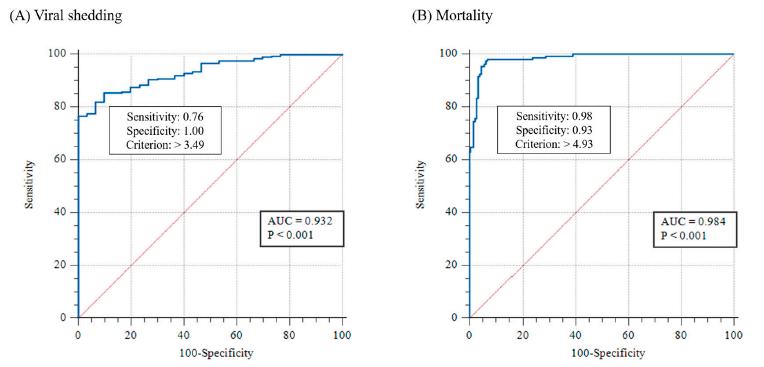
Los resultados de la PCR en tiempo real en los Exp. 1 y 2 se utilizaron para investigar el umbral de los cambios clínicos (desprendimiento viral y mortalidad) en camarones infectados con WSSV utilizando el valor de corte, que se determinó mediante el análisis de la curva ROC (Fig. 5). Los valores de corte para el desprendimiento viral y la mortalidad fueron 3.1 × 103 (103.49) y 8.5 × 104 (104.93) copias/mg del genoma del WSSV, respectivamente (Fig. 5).
Correlación entre el grado de severidad del WSSV y las copias virales de pleópodos
Con base en los resultados del análisis histopatológico y PCR en tiempo real en los experimentos 1 y 2, la correlación entre los grados de severidad y el análisis cuantitativo se presenta en la Tabla 4. Un total de 23 individuos mostraron un grado de severidad
de G0, con una mediana de copias virales de 7.9 × 101 (IQR, 4.3 × 101 a 2.6 × 103) copias del genoma del WSSV/mg observados en el pleópodo. Además, los grados de severidad de G1, G2 y G3 correspondieron a copias virales medianas de 3.1 × 103 (7.4 × 102 a 9.9 × 103), 7.5 × 104 (1.3 × 104 a 5.5 × 105) y 1.9 × 106 (1.4 × 105 a 1.5 × 107) copias del genoma del WSSV/mg, el umbral de desprendimiento viral correspondió al grado de severidad de G1 y la mortalidad correspondió al grado de severidad de G2 (Fig. 6).
Transmisión a través del agua del WSSV en condiciones que imitan condiciones naturales
Dosis infecciosa mínima del WSSV a través del agua
En el Exp. 3, a diferentes dosis de inmersión, los camarones a los que se les administró
- AGOSTO 2023 30 PATOLOGÍA
Log10 copias del genoma del WSSV/L/g)
Log 10 copias del genoma del WSSV/mg)
105 copias del genoma del WSSV/mL comenzaron a morir a los 3 dpi y mostraron un 65.0% de mortalidad acumulada durante 14 días. Mientras tanto, los camarones a los que se administró 103 y 101 copias del genoma WSSV/mL no mostraron diferencias significativas con respecto a los grupos control, y los genomas virales no pudieron identificarse mediante PCR en tiempo real (Fig. 7A). Las copias virales de pleópodos y la tasa de desprendimiento viral de los camarones a los que se administró 105 copias del genoma de WSSV/mL se detectaron por primera vez a los 3 ppp, y tanto el número máximo de copias virales de pleópodos (7.8 × 105 copias del genoma del WSSV/mg) como la tasa de desprendimiento viral (3.5 × 106 copias del genoma del WSSV/L/g) se observaron a 7 ppp (Fig. 7A). Después de 9 ppp, las copias virales de los pleópodos y la tasa de desprendimiento disminuyó durante el período experimental (9 y 11 ppp), y los camarones vivos en el grupo de mortalidad acumulada mostraron 7.5 × 103 (DE ± 8.2 × 103) copias del genoma del WSSV/mg.
En el Exp. 4, los camarones se sumergieron en diferentes dosis y se expusieron continuamente. Se encontró que la mortalidad acumulada de los camarones a los que se administró 103 y 101 copias/ml del genoma del WSSV fue del 85.0 y el 35.0 %, respectivamente, durante un período de 14 días (Fig. 7B). Además, los grupos de 103 y 101 copias del genoma WSSV/mL confirmaron la infección a los 3 y 7 ppp, respectivamente, de los camarones muestreados en el transcurso del tiempo.
Dinámica de patogenicidad y carga viral en el ensayo de cohabitación
En el Exp. 5, el ensayo de cohabitación se realizó con diferentes dosis de administración del WSSV y temperaturas del agua para verificar la desprendimiento viral de los camarones infectados y las dosis infectivas mínimas del WSSV en condiciones naturales similares.
En los grupos a los que se administró 105 copias del genoma del WSSV/camarón, la transmisión a través del agua se indujo con éxito en los grupos receptores a 5 ppp con una tasa de mortalidad acumulada superior al 85.0%, independientemente de la temperatura del agua (Fig. 8). A los
Tabla 2. Grados de gravedad del virus del Síndrome de la Mancha Blanca (WSSV) en el Exp. 1, de acuerdo con el transcurso del tiempo.
Dosis administrada (copias del genoma del WSSV/camarón)
Días de muestreo Nivel de gravedad de WSSV1 (n = 3)
1Niveles de infección del WSSV según Lightner (1996); 2dpi, días posinfección.
Tabla 3. Grados de gravedad del virus del síndrome de la mancha blanca (WSSV) en el Exp. 2, de acuerdo con el transcurso del tiempo
Grupo Grupo Días de muestreo Días
Cambio descendente 30 - 20
1Niveles de infección del WSSV según Lightner. (1996); 2dpi, días posinfección.
Figura 5. Umbrales para cambios clínicos de camarones infectados con WSSV, que se determinaron mediante análisis de curvas ROC. Los valores de corte de cada cambio clínico se analizaron en función de los resultados del análisis de la PCR en tiempo real en los experimentos 1 y 2.
- AGOSTO 2023 31 PATOLOGÍA 1 dpi 2 G0 1 dpi2 dpi G0–G1 2 dpi G0–G1 4 dpi G1–G2 4 dpi G2–G3 survived G0–G1 survived G0–G2
105 1 dpi 2 G2 3 dpi G3–G4 103 1 dpi G1–G2 3 dpi G2–G3 101 1 dpi G0 3 dpi G0
Grupo
de muestreo Nivel de
de
(n = 5) Nivel de gravedad de WSSV1 (n = 5)
gravedad
WSSV1
- 30 Constant (20 °C) Constant (30 °C) 1 dpi G0–G1 1 dpi2 dpi G0–G2 2 dpi G1–G2 4 dpi G1–G2 4 dpi G3–G4 survived G0–G2 survived G0–G1
Cambio ascendente 20
3 ppp, cuando los grupos receptores se alojaron en los tanques experimentales, aunque estaban presentes 104 a 105 copias del genoma WSSV/mL de cargas virales en todas las condiciones de temperatura, las copias virales de los pleópodos en los recipientes diferían dependiendo de la temperatura. Similar a los resultados de los ensayos de inyección, la infección progresó más rápidamente a 25°C. En los grupos de 20°C y 30°C, la mortalidad masiva de los grupos receptores se produjo a partir de los 7 dpi, con más de 105 copias del genoma del WSSV/mg de copias virales en pleópodos (Fig. 9).
En los grupos a los que se administró 103 copias de genoma del WSSV/camarón, se observó un 100.0% de mortalidad acumulada a 25°C tanto en el grupo de donantes como en el de receptores. Para los camarones alojados a 30 y 20°C, se observó una mortalidad acumulada de 40.0 y 30.0% en los grupos donantes, mientras que en los grupos receptores se observó una mortalidad acumulada de 60.0 y 35.0%, respectivamente (Fig. 8). A los 3 dpi, cuando los grupos receptores se acomodaron en los tanques experimentales, aunque tanto los grupos de 30°C como los de 25°C mostraron aproximadamente 103 copias del genoma WSSV/mL de carga viral en agua de mar, se observaron infecciones en los grupos receptores a los 7 y 5 ppp, respectivamente. Para el grupo de 20°C, la infección del grupo receptor se confirmó a los 9 dpi, mientras que las cargas virales en agua de mar se mostraron como 102 a 103 copias del genoma WSSV/mL (Fig. 9). Además, el máximo de copias virales de pleópodos mostradas en los grupos receptores coincidió con la observación de mortalidad masiva (30 y 25°C: 9 dpi; 20°C: 11 dpi; Fig. 9).
Discusión
La tasa de desprendimiento viral y la dosis infecciosa mínima del WSSV son algunos de los factores más importantes que deben investigarse en los modelos de transmisión de patógenos de animales acuáticos a través del agua. Además, debido a que los camarones infectados con WSSV muestran varios cambios clínicos (como desprendimiento viral y mortalidad) que varían según el grado de gravedad [19,20], es importante comprender la correlación
Grado de severidad del WSSV basados en la histopatología1
n2 Copias del genoma viral/mg (mediana [IQR3])
Umbral de cambios clínicos en camarones infectados con WSSV4
1Grado de infección del WSSV según Lightner [17]; 2 número de individuos correspondientes al grado de severidad en el Exp. 1 y Exp. 2; 3IQR, rango Inter cuartil; 4umbral de cambios clínicos en camarones infectados con WSSV determinado mediante análisis de curva ROC.
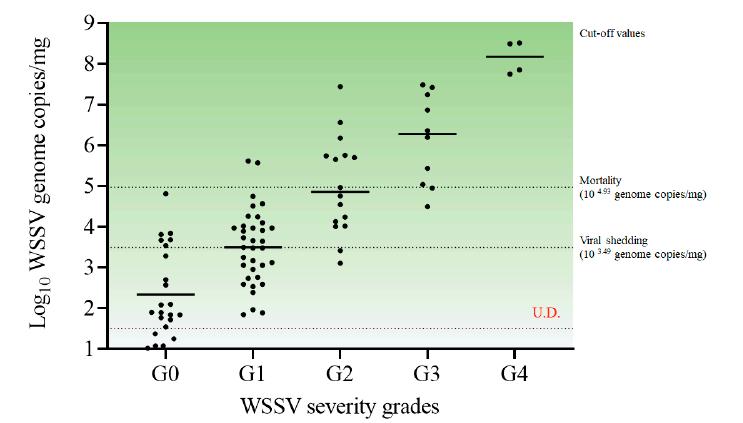

Figura
de
y umbrales de cambios clínicos en los experimentos 1 y 2. Los grados de severidad del WSSV se determinaron mediante hallazgos histopatológicos según Lightner [17]. Las barras horizontales indican el valor de la mediana de cada grupo. Las líneas punteadas horizontales indican los valores de corte de cada cambio clínico o el límite de detección del ensayo de PCR en tiempo real. UD indica indeterminado. Valores de corte
entre el desprendimiento viral y el grado de severidad para evaluar el impacto potencial de los camarones infectados en individuos ingenuos. Por lo tanto, este estudio investigó la correlación entre los niveles de severidad, las copias virales en los pleópodos y los cambios clínicos (desprendimiento viral y mortalidad) de los camarones infectados mediante ensayos de inyección en diferentes dosis de administración y condiciones de temperatura (Exp. 1-2). Con base en la correlación entre el grado de severidad y las tasas de desprendimiento viral, verificamos la transmisión del WSSV a través del agua con base en las dosis infecciosas mínimas y
las cargas virales en el agua de mar a través de ensayos de inmersión y cohabitación (Exp. 3-5).
Aunque la virulencia del WSSV varía según la especie de camarón y la temperatura, los camarones peneidos son generalmente muy susceptibles al WSSV. Por lo tanto, en este estudio, se investigó un modelo de transmisión horizontal del WSSV utilizando camarón blanco, según lo recomendado por la Organización Mundial de Sanidad Animal (WOAH) [28]. En el Exp. 1, con base en las copias virales determinadas por la PCR en tiempo real, según lo recomendado
- AGOSTO 2023 32 PATOLOGÍA
Tabla 4. Correlación entre los niveles de infección por WSSV, las copias del genoma viral y los cambios clínicos en camarones infectados con WSSV.
6. Correlación entre los grados de severidad del WSSV, copias virales
pleópodos
por la WOAH [28], a los camarones se les administraron diferentes dosis del genoma del WSSV (105 como infección temprana/ mortalidad, 103 como infección/mortalidad y 101 como no o infección de bajo grado). Como dosis inducibles de infección y mortalidad, los camarones a los que se administraron 105 y 103 copias del genoma del WSSV mostraron una mortalidad acumulada del 100% a los 4 y 10 ppp, respectivamente, mostrando diferencias significativas según la dosis inicial administrada (Fig. 2A). Del mismo modo, Pang et al. [29] reportaron que los camarones después de la administración de 104 copias del WSSV tenían 103 copias/ mg de copias virales a 1 dpi y mostraron una mortalidad acumulada del 100% en cuatro días. Con base en los resultados de los experimentos 1 y 2 se llevó a cabo una prueba de ensayo, administrando 103 dosis de copias del genoma del WSSV en un rango de temperatura del agua de 20 a 30°C, que es comúnmente reportado en estanques de cultivo de camarones [23].
Aunque el WSSV indujo la infección a temperaturas del agua de 20°C y 30°C, se reportó que el rango de temperatura óptimo para la propagación del WSSV era de 23 a 28°C [30]. Al igual que en un estudio anterior, la mortalidad acumulada de camarones infectados con WSSV aumentó significativamente en los grupos de cambio ascendente (80.0%) y descendente (87.5%) en comparación con los grupos constante (20°C; 27.5%) y constante (30°C; 47.5%), a los que se les administró la misma temperatura (Fig. 2B). Aunque hubo una diferencia significativa en la expresión del gen VP28 del WSSV, que indica los niveles de propagación del WSSV [31], dependiendo de los cambios de temperatura a 4 dpi (Fig. 4B), la reducción en la mortalidad de los grupos cambiantes después de 7 dpi podría sugerir que los niveles de propagación del WSSV se redujeron debido a que se completaron los cambios de temperatura y la temperatura del agua estaba fuera del rango óptimo para la propagación. Además, las copias virales de los pleópodos no mostraron diferencias significativas entre los camarones muertos y los sobrevivientes, independientemente de la dosis administrada y la temperatura del agua (Fig. 3). Además, aunque la tasa de desprendimiento viral también aumentó con una tendencia similar a la de
Figura 7. Resultados de los retos de inmersión (experimentos 3 y 4). (A) En el experimento 3, los camarones se sumergieron en WSSV en tres dosis diferentes (105, 103 y 101 copias del genoma del WSSV/mL) con agua de mar esterilizada con UV a 25°C durante 24 h. (a) Se observó mortalidad acumulada durante 14 días por duplicado. (b) La tasa de desprendimiento viral (copias del genoma del WSSV/L/g) se determinó en función del número de copias del genoma viral en agua de mar de cultivo y el peso de los camarones restantes. Las barras verticales indican copias virales de pleópodos en camarones muestreados a lo largo del tiempo (n = 3). (B) En el experimento 4, los camarones se sumergieron en agua de mar enriquecida con WSSV en dos dosis diferentes (103 y 101 copias del genoma del WSSV/mL) durante 24 h. Luego se usó agua de mar enriquecida con WSSV para mantener las cargas virales en el agua de mar una vez cada dos días con la misma dosis de inmersión inicial. (a) Se observó mortalidad acumulada durante 14 días. (b) La prevalencia en el transcurso del tiempo (n = 3) se determinó mediante PCR en tiempo real con un valor LOD95%. El número sobre cada diagrama de dispersión indica el resultado de la PCR en tiempo real para el grupo correspondiente. La línea punteada horizontal indica el valor LOD95% de este estudio, y UD indica indeterminado.
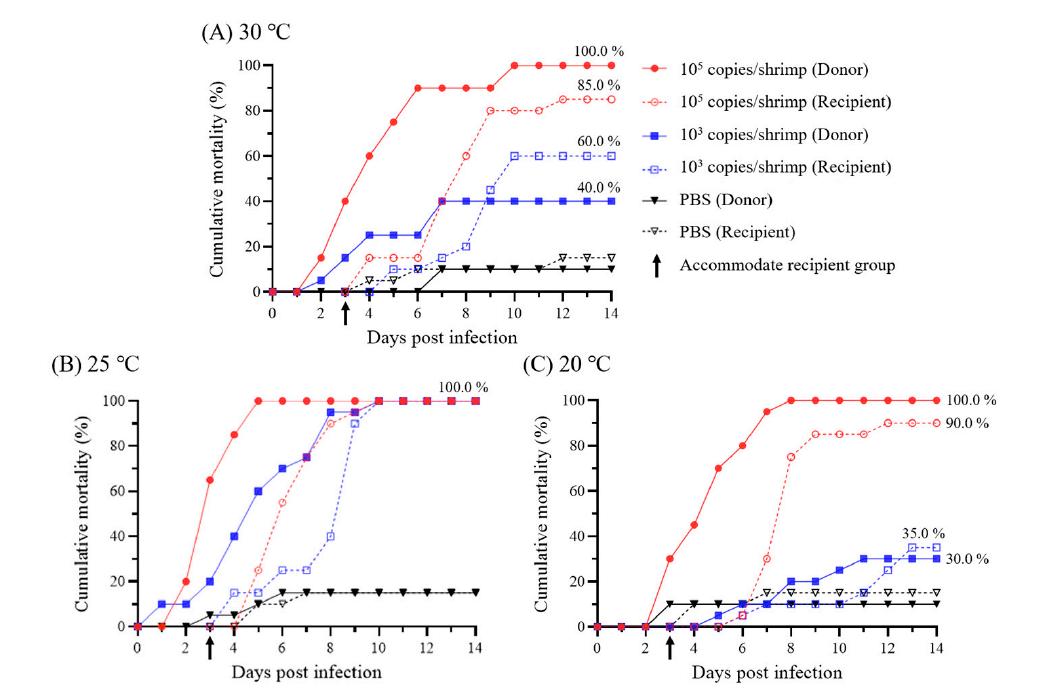
Figura 8. Mortalidad acumulada del experimento 5 (ensayo de cohabitación) durante 14 d. A los camarones se les inyectó vía intramuscular 105 o 103 copias del genoma del WSSV/camarón (donante; n = 20) a 30, 25 o 20°C. Los receptores (n = 20) en cada grupo se alojaron 48 horas después de la infección y se mantuvieron durante 14 días en jaulas separadas en el mismo tanque. A los grupos de control negativo se les inyectó solución salina tamponada con fosfato (PBS).
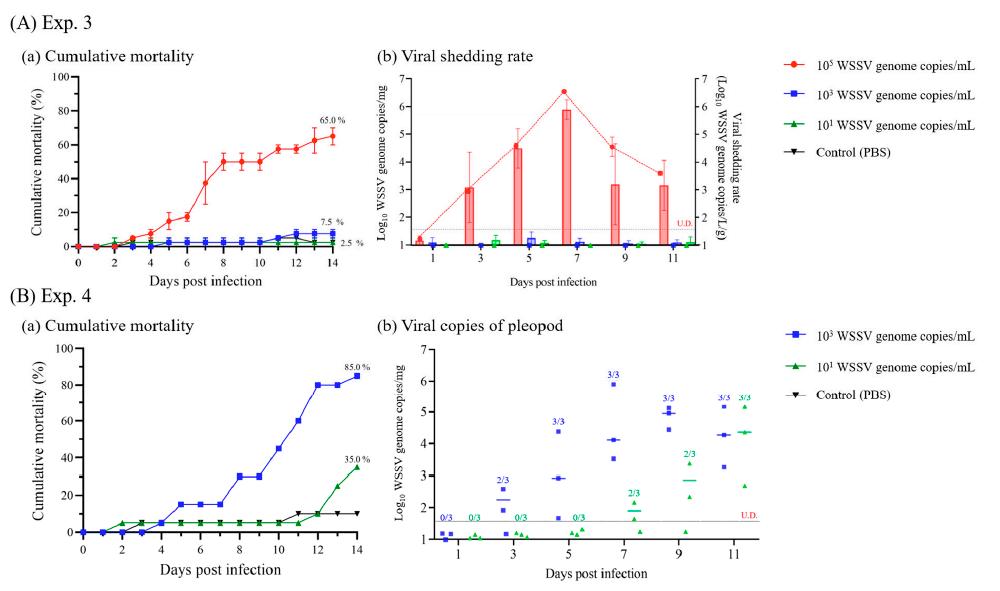
- AGOSTO 2023 33 PATOLOGÍA
las copias virales en pleópodos, con una diferencia significativa según la dosis inicial administrada y la temperatura del agua, el desprendimiento viral podría ocurrir a aproximadamente 103 copias del genoma del WSSV/mg (Fig. 4). Por lo tanto, clasificamos los grados de severidad de WSSV usando pruebas de ensayo de inyección (Exp. 1-2) y determinamos los grados de severidad de la infección del WSSV en camarones blancos (etapa juvenil) con base en hallazgos histopatológicos.
Los grados de severidad en los camarones infectados con WSSV se determinaron de acuerdo a Lightner [17] en función de los hallazgos histopatológicos, y se investigó su relación con los resultados de la PCR en tiempo real. Según los hallazgos histopatológicos y los resultados de la PCR en tiempo real de camarones infectados con WSSV, el número de copias virales en el grupo G1 fue de 10 a 103 copias/mg del genoma de WSSV. Además, los grados de severidad de G2, G3 y G4 correspondieron a 104 a 105, 105 a 107 y 107 a 108 copias del genoma WSSV/mg, respectivamente, en los pleópodos (Tabla 4). De manera similar, Tang y Lightner [18] evaluaron la correlación entre los análisis cuantitativos e histopatológicos y encontraron que los grados de severidad G2-G3 y G4 correspondían a 106 y 108 copias virales/mg en camarones infectados, respectivamente. Aunque el análisis histopatológico no es muy cuantitativo y puede variar entre las técnicas de diagnóstico [18], estos resultados respaldan la correlación entre el análisis histopatológico y la PCR en tiempo real (Tabla 4).
Para investigar la correlación entre los grados de severidad y los cambios clínicos en los camarones infectados con WSSV, determinamos los umbrales para el desprendimiento viral y la mortalidad como 3.1 × 103 (103.49) y 8.5 × 104 (104.93) copias/ mg del genoma del WSSV, respectivamente, utilizando el análisis de la curva ROC (Fig. 5). En el Exp. 1, la mediana de copias virales en pleópodos en la primera muerte de camarones (105 copias/grupo de camarón a 1 dpi; 103 copias/grupo de camarón a 3 dpi) fue de aproximadamente 105 a 106 copias del genoma del WSSV/mg. Mientras tanto, en el Exp. 2, aunque la primera muerte ocurrió a los 2–3 ppp (con 102 a
103 copias del genoma WSSV/mg de copias virales medianas), varios individuos en cada grupo mostraron cargas relativamente altas (aproximadamente 104 a 105 copias/mg) a los 2 ppp (Fig. 3). En comparación con Sun et al. [32], aproximadamente 105 a 106 copias del genoma del WSSV/mg de copias virales fueron suficientes para inducir la mortalidad en los camarones. Además, la mortalidad masiva de 105 copias/mL de camarones sumergidos en el Exp. 3 (inmersión durante 24 h) y grupos de destinatarios en el Exp. 5 (ensayo de cohabitación) cuando el promedio de copias virales en los pleópodos fue superior a aproximadamente 105 copias/ mg (grado de severidad estimado de G2; figuras 7 y 9). Existe información limitada sobre el desprendimiento del WSSV en el camarón, pero varios estudios previos han reportado que el WSSV se puede detectar en el agua de mar, incluso cuando el número de copias virales en los camarones es de aproximadamente 102 a 103 copias/mg [23,31]. Estos resultados sugieren que el
grado de severidad de G1 (umbral: 3.1 × 103 copias/mg) y G2 (umbral: 8.5 × 104 copias/ mg) en camarones induce suficientemente el desprendimiento viral y la mortalidad, respectivamente (Fig. 6). Dado que no se pudo determinar la viabilidad del virus mediante la PCR en tiempo real, se necesitan más estudios para aclarar la correlación entre el análisis histopatológico y los ensayos moleculares, incluyendo una combinación de hallazgos histopatológicos y viabilidad viral determinada mediante ensayos de PCR de viabilidad.
El desprendimiento viral es esencial para predecir las cargas virales en el agua de mar y los niveles de exposición de individuos sanos en contacto con individuos infectados (o expuestos a agua contaminada) y podría ser una herramienta útil para comprender la epidemiología de la enfermedad [7,9]. Por lo tanto, este estudio analizó las cargas virales en el transcurso de tiempo en pleópodos y agua de mar para investigar la correlación
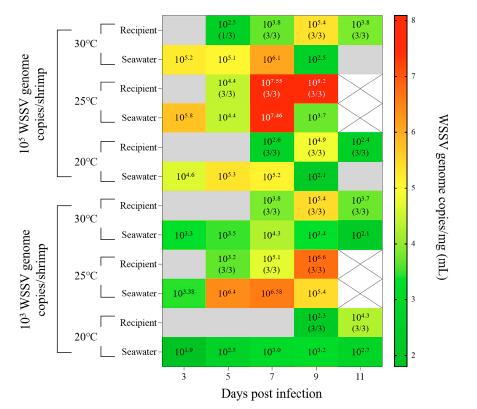
- AGOSTO 2023 34 PATOLOGÍA
Figure 9. Time-course mean viral copies of pleopods in recipients (n = 3) and viral loads of seawater in experiment 5 (cohabitation challenge). The values in each plot indicate the mean viral copies/mg (the prevalence of each sampled shrimp) and mean viral copies/mL. The gray block indicates values below the LOD95% value, and the white ‘x’ marked box indicates time points at which no measurements were carried out due to all shrimp being dead.
entre las etapas de infección del WSSV y la tasa de desprendimiento viral. Basado en los resultados de la PCR en tiempo real para copias virales en pleópodos y agua de mar de los Exp. 1 y 2, se observó que la tasa de desprendimiento viral aumentó de manera similar a las copias virales de los pleópodos. Además, el análisis de regresión (Fig. 4C) confirmó una correlación lineal entre la tasa el desprendimiento viral y las copias virales en pleópodos, lo que indica una relación positiva entre las dos variables.
En el Exp. 2, la tasa de desprendimiento viral mínima se observó en el grupo constante (20°C) a 2 ppp (2.0 × 102 copias del genoma del WSSV/L/g) con grados de severidad G0G1, mientras que la tasa de desprendimiento viral máxima se observó en el grupo de cambio descendiente a 4 ppp (4.5 × 107 copias del genoma WSSV/L/g) con grados de severidad G3–G4. Además, en el Exp. 5, los grupos a los que se les administró 103 copias/camarón mostraron cargas virales máximas en agua de mar dos días antes de la observación de mortalidad masiva en los grupos receptores (Figuras 8 y 9). Varios estudios han reportado una correlación entre el desprendimiento viral, la carga viral en la especie huésped y la mortalidad. Según Hershberger et al. [7], los picos de desprendimiento viral en arenques del Pacífico infectados con VHSV precedieron a los picos de mortalidad diaria en 1 a 2 días.
En estudios de campo, Jang et al. [33] llevaron a cabo el monitoreo del WSSV en 20 granjas de cultivo de camarón en Corea del Sur y encontraron una correlación positiva entre la carga viral tanto en los camarones como en el agua de mar. Además, nuestro estudio anterior [23] confirmó que la tasa de detección del WSSV en camarones aumentó en julio y septiembre, coincidiendo con un aumento en las cargas virales en el agua de mar. Estos hallazgos sugieren que la tasa de desprendimiento viral se correlaciona positivamente con la gravedad y viabilidad del huésped. Con base en estos resultados, los grados de severidad de los camarones y las cargas virales en el agua de mar, podrían ser factores útiles para determinar la gravedad de la enfermedad y rastrear los brotes de enfermedades en estudios epidemiológicos.
La dosis infectiva mínima es un parámetro importante para investigar el riesgo
de transmisión horizontal a especies susceptibles expuestas a patógenos [8]. Varios estudios epidemiológicos han reportado modelos de transmisión por agua basados en la dosis infectiva mínima y el desprendimiento viral en huéspedes infectados [7,10,21]. A pesar de varios estudios previos que investigaron las dosis infectivas mínimas del WSSV a través de ensayos de inmersión, la transmisión horizontal del WSSV está influenciada por factores ambientales (es decir, la temperatura del agua y la densidad de siembra), y debido a la ausencia de una línea celular para cultivar WSSV, la determinación de la dosis infectiva mínima del WSSV no es fácilmente accesible [16,34].
Por lo tanto, para comprender el modelo de transmisión del WSSV a través del agua, investigamos la dosis infectiva mínima del WSSV a través de ensayos de inmersión a diferentes dosis y períodos de exposición (Exp. 3-4). A partir de entonces, para verificar el desprendimiento viral en camarones infectados con WSSV y las dosis infectivas mínimas del WSSV, se llevaron a cabo ensayos de cohabitación (Exp. 5) a tres temperaturas diferentes (30, 25 y 20°C), que comúnmente se reportan en camarones cultivados en estanques. En el Exp. 3, el camarón blanco se sumergió en 105, 103 y 101 copias del genoma WSSV/ml de agua de mar durante 24 h. Se realizaron ensayos de inmersión continua en el Exp. 4 a dosis relativamente bajas (103 y 101 copias del genoma de WSSV/mL, respectivamente) durante 14 días (Figura 7).
En un estudio previo, Durand y Lightner [13] encontraron que 105 copias del genoma del WSSV/mL de agua de mar eran suficientes para inducir la infección y la mortalidad en el camarón blanco, mientras que menos de 104 copias del genoma del WSSV/mL de agua de mar eran insuficientes para inducir la infección por inmersión para 4 horas. Mientras tanto, un estudio previo reportó que poslarvas de camarón blanco eran susceptibles a 103 y 104 copias del genoma WSSV/mL de agua de mar a través de ensayos de inmersión con un 50% de condiciones de intercambio de agua [15]. Del mismo modo, en el Exp. 3, solo los grupos expuestos a 105 copias/mL de WSSV mostraron infección y mortalidad, mientras que los grupos expuestos a 103 y
101 copias del genoma del WSSV/mL se confirmó que estaban infectados a 3 y 7 dpi, respectivamente, bajo exposición continua (Fig. 7). Durante los ensayos de convivencia del Exp. 5, las cargas virales en el agua de mar de los grupos administrados con 105 copias del genoma del WSSV/camarón fueron de 105 copias del genoma del WSSV/ mL, y los receptores se infectaron en dos días, independientemente de la temperatura del agua (30, 25 y 20°C; Fig. 9). Mientras tanto, los grupos de administrados de 103 copias del genoma WSSV/camarón mostraron 103 copias/mL de carga viral en agua de mar a 30 y 25°C, y la infección de los receptores se observó después de 4 y 2 días después del alojamiento, respectivamente. Además, en el grupo de 20°C, los receptores confirmaron la infección a los seis días, mientras que las cargas virales del agua de mar se observaron en aproximadamente 101 a 102 copias del genoma del WSSV/mL (Fig. 9).
Con base en estos resultados, encontramos que, a pesar de la influencia de la temperatura del agua en la transmisión del WSSV a través del agua, 105, 103 y 101 copias del genoma del WSSV/mL en el agua de mar podrían inducir infecciones transmitidas por el agua en aproximadamente uno, tres y siete días, respectivamente. Estos resultados sugieren que la transmisión del WSSV a través del agua se ve afectada no solo por la carga viral en el agua de mar, sino también por la duración de la exposición. Por lo tanto, para comprender mejor la transmisión del WSSV a través del agua, es importante analizar el desprendimiento viral para estimar su impacto en individuos ingenuos.
En los sistemas de cultivo de camarón en estanques, la interacción entre los huéspedes infectados y los no infectados podría ser crucial para mantener la viabilidad del WSSV [35]. Un estudio previo investigó la viabilidad del WSSV en agua de mar y reportó que el WSSV podría conservarse hasta 14 días en agua de mar en condiciones de laboratorio [14]. Song et al. [36] reportaron que el WSSV se detectó consistentemente en agua de mar de estanques de camarón desde junio hasta septiembre de 2007, mientras que Kim et al. [23] reportaron que se detectó el WSSV durante hasta 55 días en agua de estanques de camarón durante el período de estudio.
- AGOSTO 2023 35 PATOLOGÍA
En el Exp. 5 de este estudio, se indujo la transmisión a través del agua en el reservorio y las cargas virales del agua de mar se mantuvieron a medida que se producía la infección, independientemente de la temperatura del agua (Fig. 9). Según estos resultados, las cargas virales en el agua de mar podrían mantenerse a través de interacciones con huéspedes o vectores, y estas interacciones ayudaron a que el WSSV siguiera siendo viable durante un período prolongado.
Se necesitan más estudios para aclarar el mecanismo de transmisión horizontal del WSSV en los sistemas de cultivo en estanques, para investigar las interacciones más complejas, como la influencia del canibalismo y la dinámica del WSSV entre especies huésped, vectores y reservorios.
Conclusiones
En este estudio, investigamos la correlación entre el grado de severidad de la enfermedad y los cambios clínicos en los camarones
infectados con WSSV. Nuestros resultados revelaron que un grado de severidad de G2 (valor de corte: 104,9 copias/mg) fue suficiente para inducir la mortalidad en el camarón blanco y los camarones infectados con un grado de severidad de G1 (valor de corte: 103.5 copias/mg) fue capaz de eliminar el virus. Sobre la base de la correlación entre el grado de severidad de la enfermedad y la tasa de desprendimiento viral, determinamos una correlación positiva entre las copias virales de los pleópodos y la tasa de desprendimiento viral (y = 0.7076x + 1.414; p < 0.001). Además, las pruebas de ensayo que imitan las condiciones naturales (ensayos de inmersión y cohabitación) revelaron que 105, 103 y 101 copias del genoma del WSSV/mL de agua de mar podrían inducir una infección transmitida por el agua en 1, 3 y 7 días, respectivamente.
Nuestros hallazgos sobre el modelo de transmisión por agua del WSSV pueden ayudar a comprender la dinámica del WSSV en un sistema de cultivo en estanques. Además,
nuestros resultados se pueden utilizar para dilucidar aún más las interacciones entre las especies durante los brotes de enfermedades y la propagación de WSD en los sistemas de cultivo en estanques.
Materiales complementarios: La siguiente información de apoyo se puede descargar en:
https://www.mdpi.com/article/10.3390/ ani13101676/s1 , Tabla S1: Primer utilizado en este estudio; Figura S1: Diagrama esquemático de las infecciones experimentales en este estudio•
Para mayor información escriba a: kimki@pknu.ac.kr

- AGOSTO 2023
PATOLOGÍA
Las características físicas y nutricionales de los alimentos microextruidos y su efecto en el rendimiento de larvas de Litopenaeus vannamei
Autores:
César Molina-Poveda
Xavier Zavala*
Felipe Ramón*
Josselyne Pizarro*
Francisco Campuzano*
Carlos Mora-Pinargote
Manuel Espinoza-Ortega
cesar.molina@skretting.com
Skretting Aquaculture Innovation
*Skretting LATAM
Unos de los pilares para el desarrollo y mantenimiento de la industria de camarón blanco Litopenaeus vannamei en el Ecuador es la obtención y disponibilidad de larvas de calidad. Por su naturaleza, las larvas de camarón requieren de dietas especiales que satisfagan sus necesidades nutricionales para promover un crecimiento saludable (García-Galano, 1998).
La acuicultura de camarón blanco continúa experimentando un crecimiento sin precedentes durante los últimos años. Sin embargo, existen una variedad de factores que pueden conducir a ineficiencias en la producción. Estos factores, a veces marginales, cuando se suman podrían contribuir a generar cuellos de botella que llevan a una producción reducida, y en el tiempo, a pérdidas económicas significativas (Neiland et al., 2001, Engle et al. 2017). Los problemas comunes incluyen una pobre producción de larvas (Juinio-Menez y Gontaco, 2004; Bardera et al., 2019).
Las larvas comúnmente son alimentadas con alimento vivo: Artemia, zooplancton y fitoplancton (García-Galano, 1998), cuyo mantenimiento representa un gasto elevado para los laboratorios (Eras-Agila y MeleánRomero, 2021). Esta actual demanda de alimentos larvales y su constante expansión ha fomentado la investigación y desarrollo de dietas inertes con características nutricionales más eficientes y equilibradas, que se vea complementado con procesos de manufactura innovadores y materias primas nóveles (Gamboa-Delgado y Le Vay, 2009) que apoyen dicha mejora física y nutricional.
En los últimos años, el uso de la tecnología de extrusión en la elaboración de alimento acuícola representa uno de los desarrollos más versátiles y energéticamente eficientes en cuanto a transformación de ingredientes (Sorense, 2012; Sorense et al., 2007; Tacon, 2017; Welker et al., 2018). La extrusión permite un control preciso de la textura y el tamaño del pellet, asegurando una alimentación más uniforme y evitando la segregación de nutrientes. Además, por la mayor temperatura y presión a las que son sometidos durante su manufactura, los alimentos extruidos pueden reducir la presencia de antinutrientes y patógenos. El

- AGOSTO 2023 NUTRICIÓN
38
aumento de la digestibilidad de los alimentos es otra ventaja asociada a los alimentos extruidos, al tener una mayor gelatinización de los almidones que permiten una mejor utilización de los nutrientes, resultando en un mayor crecimiento y eficiencia de conversión del alimento. Entre otros beneficios que otorga la extrusión de los alimentos destacan la aumentada hidroestabilidad que reduce la tasa de pérdida de nutrientes en el agua, lo que se traduce en una disminución del potencial de contaminación del medio acuático. Adicionalmente, la durabilidad de los alimentos es mejorada, lo que prolonga la vida útil, almacenamiento y transporte sin riesgo de deterioro.
En el presente trabajo se explorarán las ventajas a nivel físico y nutricional de alimentos microextruidos de última generación, con énfasis en las mejoras que se pueden lograr por medio del proceso de extrusión en frío aplicado a alimentos larvales.
Aspectos físicos
Forma y tamaño adecuados
La forma y tamaño adecuados en un alimento larval son características importantes ya que aseguraran la captura del pellet por parte del animal, permitiendo el consumo. Además, un número adecuado de pellets por unidad de peso asegura la máxima disponibilidad.
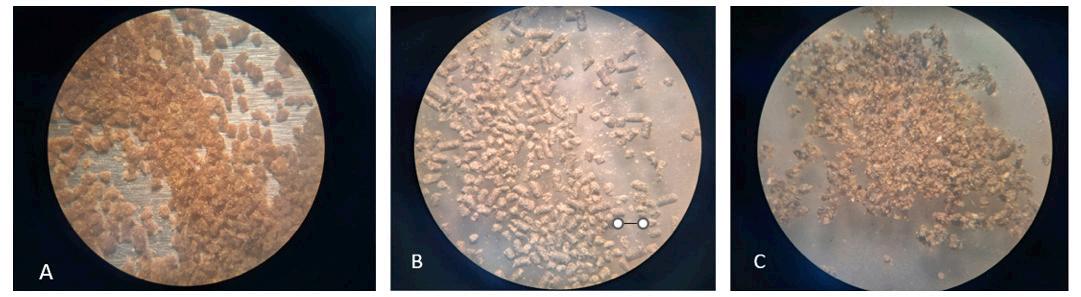
Muchos aspectos subyacentes a los procesos de alimentación son aclarados mediante la observación de la morfología de los apéndices de alimentación, el patrón de movimiento de estos apéndices y la forma de las dietas. Las observaciones del comportamiento de alimentación de las larvas de camarón ayudan a definir con precisión el tamaño de la partícula ingerida en los diferentes estadios hasta llegar al estadio postlarval, donde los hábitos de captura de presas son más evidentes. Esto sugiere que es posible simplificar los programas de alimentación en larvas de camarón a un solo tamaño, permitiendo un manejo simplificado de los esquemas de producción.
La fabricación de los alimentos larvales tiene varios desafíos. Durante el proceso de extrusión, a medida que la masa plastificada sale del molde bajo presión, se hincha, pierde humedad y se enfría,
produciéndose la consecuente expansión del producto. Estos eventos afectan el tamaño real y la forma de los gránulos extruidos. Sin embargo, a diferencia de los métodos convencionales de procesamiento, como el desmoronado de alimentos previamente peletizados, los alimentos microextruidos generan un producto menos irregular. Al observar el producto de los procesamientos mencionados: desmoronamiento (Figs. 1A y 1C) y microextruido (Fig. 1B), se hace evidente el impacto sobre dichas propiedades físicas en donde el producto del desmoronamiento pierde por completo la forma habitual de un pellet, y sus partículas son más irregulares en comparación al microextruido.
Densidad
La densidad aparente de un alimento es el cociente entre la masa de producto y el volumen aparente del mismo, incluyendo la porosidad existente. Este parámetro se mide generalmente utilizando un método adaptado de Wenger Manufacturing. Descrito brevemente, se vierten 30 mL de agua en una probeta de 100 mL y se registra el volumen exacto (v1). Posteriormente, una muestra de alimento es pesada (15
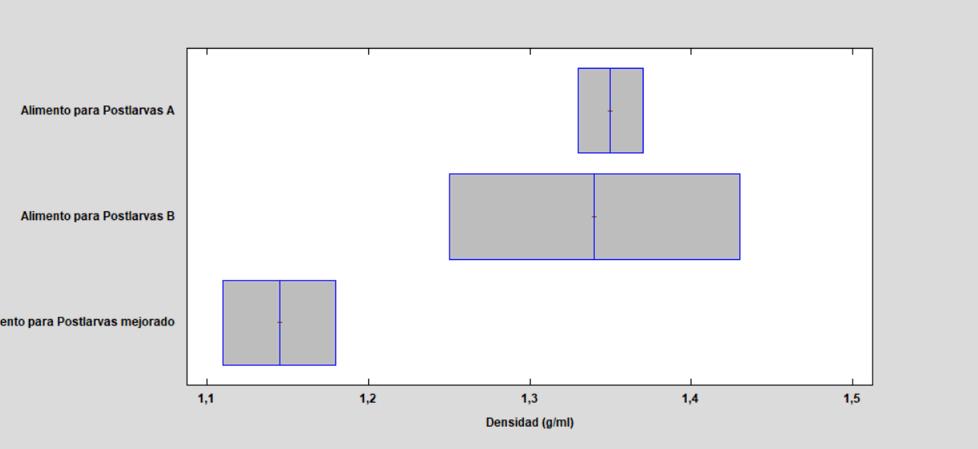
g), utilizando un embudo con una precisión de 0.01 g (Ms). Este alimento es colocado en la probeta con el v1 de agua, la cual se golpea aproximadamente 10 veces por segundo hasta eliminar las burbujas de aire. El volumen de agua es medido nuevamente (v2) y la densidad del alimento (g/ml) es calculada utilizando la fórmula δ = Ms (v2–v1).
La densidad en alimentos larvales está regida principalmente por los procesos de manufactura como los parámetros de elaboración y la operación del equipo de fabricación, por lo que la densidad apropiada se logrará al estandarizar los procesos según los requerimientos del animal o el estadio de vida a alimentar. En alimentos larvales, propiedades tan importantes como la velocidad y uniformidad de distribución durante el hundimiento a lo largo de la columna de agua, están directamente relacionadas con la densidad (Fig. 2). Una menor densidad, como en los alimentos larvales mejorados, hará que el alimento descienda lentamente en la columna de agua permitiendo a las larvas de camarón capturarlo durante el descenso. Por otro lado,
39 - AGOSTO 2023 NUTRICIÓN
Figura 1. Microfotografía tomada con 20X de 3 tipos de alimentos larvales (300 - 600 µm): A: alimento en polvo; B: alimento microextruido en frío; C: alimento desmoronado.
Figura 2. Densidad (g/ml) para diferentes tipos de alimentos destinados a postlarvas de camarón blanco (Tamaño de partícula: 300-500 µm).
densidades altas ( > 1,2 g/ml) resultan en un descenso brusco del alimento, causando que las larvas deban de desplazarse distancias mayores hasta alcanzar el fondo y poder alimentarse.
Dispersión uniforme en la superficie del agua
Las mejoras en las propiedades físicas del alimento pueden ayudar a hacer frente a diferentes desafíos que han sido una constante en la producción de larvas. La naturaleza del comportamiento de crustáceos tiene dos características muy marcadas: territorialidad y canibalismo (Akbari et al., 2010) que pueden generar dispersión de tallas y pérdidas en la producción de larvas (Su et al., 2020).´
El canibalismo se refiere a la mutilación entre individuos, un fenómeno generalizado en los animales acuáticos, e importante en términos de adaptabilidad biológica que generalmente trae asociada una considerable mortalidad, y que en acuicultura se traduce en bajas tasas de supervivencia y déficits económicos. Este fenómeno permite que ciertas especies alteren su densidad de población en función a su entorno y recursos fluctuantes. Así, el canibalismo puede determinar dominancia y subordinación intraespecífica, en la que el ganador obtiene beneficios que incluyen recursos y espacio (Su et al., 2020).
Con los avances en elaboración de dietas larvales, los alimentos pueden dispersarse al tocar la superficie del agua (ver video de la Fig. 3), lo que ayuda a que las partículas que componen las dietas lleguen a una gran cantidad de larvas. Con estas mejoras, las dietas se dispersan maximizando la disponibilidad del alimento, evitando la competencia concentrada en un área del tanque por acumulación del alimento.
Flujo uniforme
Los laboratorios de producción de camarones son ambientes muy húmedos por su cercanía al mar. Las microdietas tienden a ser higroscópicas, lo que genera grumos de alimento que pueden afectar la calidad de agua y reducir la uniformidad de la alimentación.
Con la formulación de los alimentos larvales de última generación se logra la eliminación
de grumos. En una prueba de laboratorio para evaluar las propiedades mecánicas de flujo usando un embudo troncocónico (Fig. 4), los resultados mostraron un flujo libre del alimento microextruido; mientras que usando una dieta convencional se produce atasco debido a la mayor cohesión entre las partículas. Estas propiedades se logran gracias a la encapsulación celular y los componentes clave; los alimentos microextruidos exhiben una característica más hidrofóbica, manteniéndose fluidos y sin grumos, favoreciendo una distribución uniforme después de entrar en contacto con el agua, y reduciendo la creación de aglomeraciones en el tanque que resultarán en una menor utilización del alimento


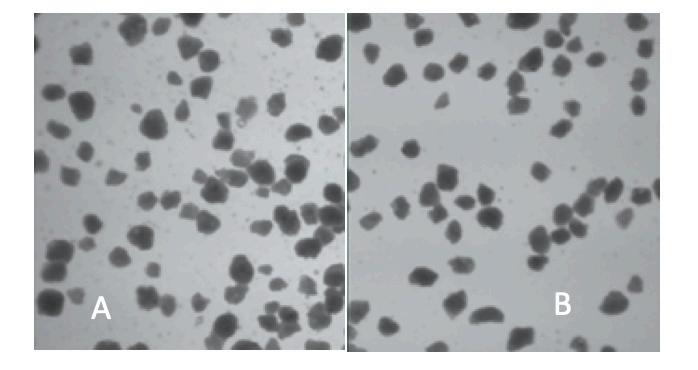


por parte del camarón, produciendo más desechos en los sistemas de producción.
Reducción de finos
Los finos son pequeños fragmentos de alimento que se desprenden de los gránulos fabricados, caracterizados por tener dimensiones inferiores al tamaño nominal del alimento. Los finos son generados en el mismo procesamiento o bien por la manipulación del alimento durante el transporte, el almacenamiento y la distribución, los cuales, en diferente medida, provocan la desintegración de los gránulos, lo que consecuentemente aumenta las cantidades de partículas de alimento
- AGOSTO 2023 40 NUTRICIÓN
que no nutren a los camarones y
Figura 4. Propiedades mecánicas de flujo mejoradas (escanear al código QR para observar el video).
Figura 3. Distribución del alimento en la superficie del agua. Nótese que con alimentos mejorados se maximiza la disponibilidad (escanear al código QR para observar el video).
Figura 5. Impacto del procesamiento en la calidad física de alimentos larvales: A) alimento procesado con métodos convencionales; B) alimento procesado por microextrusión en frío. Nótese la reducción en la cantidad de finos en el alimento procesado por microextrusión en frío (B).
que generalmente deterioran la calidad del agua. Por lo contrario, los alimentos extruidos tienen menos posibilidades de producir finos por manipulación o transporte, lo que implica una menor desintegración física. La extrusión produce unidades que son más duraderas y resistentes al daño mecánico y, en consecuencia, menos finos (Thomas y Poe, 1996). Además, la ausencia de finos en el alimento extruido reduce la contaminación ambiental y mejora la calidad del agua.
Generalmente, la tecnología de producción extruida resulta en la disminución de finos y reducción del impacto sobre la calidad de agua debido a su mayor estabilidad, permitiendo a los laboratorios de larvas intensificar la producción.
Aspectos nutricionales
Sustitución del aceite de pescado por aceite de algas
El aceite de pescado es un líquido que puede filtrarse y formar gotas de aceite que se fusionan, lo que da como resultado películas en la superficie del agua y reducción de oxígeno, incrementando la eutrofización del tanque. Por otro lado, las algas que contienen DHA tienen este ácido graso esencial encapsulado en su célula. El aceite está en esta «celda», protegido por una membrana que evita fugas pero que es digerible para los camarones, evitando su desperdicio en el agua.
La mejora en la formulación y sostenibilidad de alimentos acuícolas está estrechamente relacionada con la eliminación parcial o total del aceite de pescado. En los alimentos larvales mejorados, todos los ácidos grasos esenciales se encapsulan en las células de algas a través de un proceso conocido como encapsulación celular. Esto protege los ácidos grasos y reduce la cantidad de lixiviación y el impacto en la calidad del agua. Las células de algas siguen siendo digeribles, por lo que los lípidos están biodisponibles para las larvas.
Inclusión de péptidos

Los alimentos mejorados usan un amplio número de fuentes de proteínas hidrolizadas de origen marino. Estas proteínas incluyen péptidos y dipéptidos de bajo peso molecular que aumentan la atracción y la digestibilidad. Los péptidos son esenciales para el
crecimiento de las larvas debido a su primitivo sistema digestivo. Los alimentos mejorados incluyen una diversidad de proteínas marinas hidrolizadas que emulan lo consumido por las larvas en la naturaleza. Sin embargo, la fórmula de la dieta no es el único factor que promueve una nutrición efectiva. Una de las claves para una alimentación exitosa es la interacción del alimento con el medio. Inclusive, la mejor fórmula no consigue los resultados esperados si la estabilidad en el agua es baja o si el tamaño no es el adecuado para el estadio de vida del camarón.
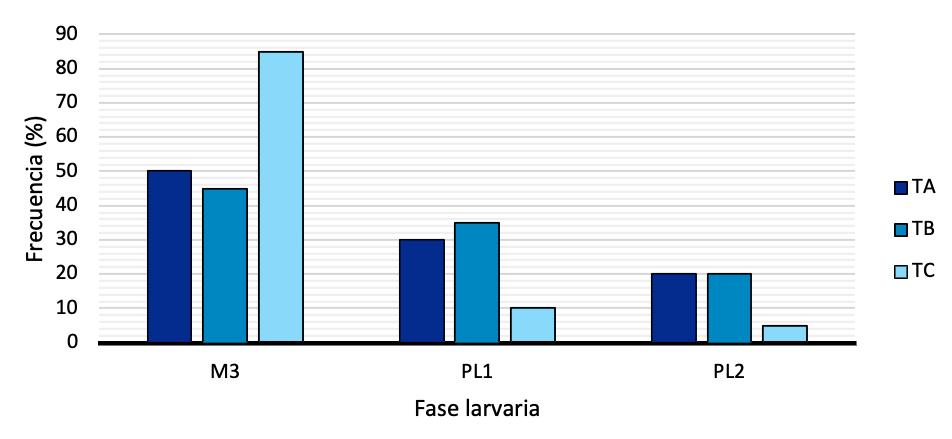
Otro punto importante es el proceso de manufactura. Las producciones de alimentos larvarios para camarón con métodos mejorados se realizan a temperaturas específicas, atendiendo a la naturaleza de los ingredientes usados y su grado de termolabilidad. Por el contenido de materias primas ricas en ácidos grasos y la adición de altas concentraciones de vitaminas usados en los alimentos larvarios, las temperaturas de procesamiento deben ser menores a
las usadas en alimentos de engorde. Un alimento larvario tiene casi el doble de contenido graso que un alimento de engorde y la velocidad de autooxidación aumenta con la temperatura, por tanto un proceso inadecuado a una temperatura excesiva puede acelerar la formación de peróxidos.
La combinación de proteínas hidrolizadas con procesos mejorados garantiza una máxima retención de nutrientes. La producción a temperatura y tiempo adecuados asegura una mayor digestibilidad y la disponibilidad de diferentes nutrientes como vitaminas y ácidos grasos.
Pruebas de campo
Prueba de tamaño de la dieta en Zoea/Mysis
Una pregunta común es la necesidad de dos o incluso tres tamaños de dieta para el período Zoea- Mysis y PL.
Un estudio llevado a cabo por nuestro equipo técnico
41 - AGOSTO 2023 NUTRICIÓN
evaluó en larvas de Litopenaeus
Figura 6. A) algas, y B) aceite celular encapsulado incluido en la dieta mejorada para larvas. Créditos de la imagen: Journal of Applied Phycology, 31 (2) (Jaseera, 2019).
Figura 7. Resultados en porcentaje de estadio larvario Mysis 3 (M3), Postlarva 1 (PL1) y Postlarva 2 (PL2), obtenidos luego de alimentar camarones con tres tratamientos: TA, TB y TC.
vannamei tres tratamientos dietéticos: (1) TA, una dieta mejorada con rango optimizado de partículas de 10 – 110 µm; (2) TB, la misma fórmula de la dieta mejorada con dos rangos diferentes para diferente estadio larvario (10 – 50 µm y / 50 – 100 µm); y (3) TC, una dieta comercial control con diferente formulación y con dos tamaños (20 – 80 µm / 70 – 150 µm).
Los resultados presentados en la Fig. 7 mostraron que al alimentar con un solo rango de tamaño de partículas (TA, 10-110 µm) se obtiene prácticamente la misma proporción de animales en M3, PL1 y PL2 comparado con un esquema que incluye dos tipos rangos de tamaños de una dieta experimental con la misma fórmula (TB, 10 – 50 µm y 50 – 110 µm). Por el contrario, cuando estos dos tratamientos se comparan con TC, el alimento comercial control en dos rangos de tamaños diferentes, se observa una mayor proporción de camarones en estadio M3. Esto sería contraproducente para cualquier productor que usará este esquema de alimentación con este tipo de alimento, tanto para su esquema de manejo como para su eficiencia productiva. La proporción de animal que creció desde M3 a PL2 no fue diferente entre TA y TB con alimento mejorado, sugiriendo que con una alimentación simplificada, es decir, de un solo rango de tamaño de alimento, es posible obtener los mismos resultados manteniendo una dieta nutricionalmente mejorada, simplificando así los protocolos de alimentación en laboratorios de producción de larvas.
Además, durante este experimento se examinó el efecto de los alimentos durante la metamorfosis de Zoea/Mysis observando rendimientos similares entre los tratamientos. Podemos concluir que el tamaño de la dieta de 0 – 110 µm es perfectamente adecuado para la alimentación de Zoea/Mysis y no hay ninguna ventaja en la alimentación con tamaños de partículas más finos durante el período de Zoea/Mysis. Esto abre la posibilidad de simplificar aún más el programa/gestión de alimentación dentro de los laboratorios de camarón, mejorar la economía y desafiar las normas aceptadas.
La tabla 1a muestra que no hubo beneficio en tener diferentes clases de tamaño para
Tabla 1a. Resultados de la prueba de tamaños de la dieta Zoea/Mysis realizada en L. vannamei con una densidad de siembra de 150 animales por litro, en tanques de 300 litros con 3 réplicas. Temperatura del agua de 28°C y salinidad de 30 ppt.
Tabla 1b. Resultado de la prueba de tamaños en PL 12 en L. vannamei
la alimentación de larvas en estadio Zoea/ Mysis. Todos los indicadores de desempeño fueron similares tanto en Z3, M3 e inclusive en PL12 (Tabla 1b).
Prueba de crecimiento y supervivencia en laboratorio
Los ensayos fueron llevados a cabo en sistemas de producción comerciales en tanques de polietileno circulares de 20 000 litros con aireación suministrada por un aireador, usando tubería de PVC con manguera porosa. La evaluación fue llevada a cabo en 4 tanques: Tq 5 y 6 se alimentó 100% con dieta extruida mejorada, mientras que los tanques 7 y 8 se alimentaron con una dieta control.
Se sembró con 60% de nivel y se fue suministrando agua durante los primeros 6 días de cultivo y a partir del octavo día se hizo recambio del 15% hasta PL4. A partir de PL5, el recambio fue de 30%/día. Los sistemas usan agua de mar filtrada con un filtro de piola de 1 micra para el llenado de los reservorios, y al pasar a los tanques con bolso de 10 micras. La temperatura media del agua estuvo en un rango de 32°C hasta 33°C y la salinidad a 30 ppt. La alimentación en todos los casos fue manual cada 3 horas en estadios de Zoeas y Mysis, y en estadios postlarvales cada 2 horas. Las pruebas fueron realizadas en duplicado.
Las observaciones realizadas al microscopio en estadios PL1 hasta PL3 revelaron que los tanques alimentados con dieta mejorada
tenían hasta el 70% de porcentaje de llenura; mientras que los tanques control solamente llegaron hasta el 60%. Este patrón se repitió en PL7, en donde el porcentaje de llenura llegó hasta un 80% con dieta mejorada y solamente hasta el 70% con el alimento control.
Otros parámetros como la actividad, porcentaje de larvas vacías y porcentaje de deformes y necrosis muscular no varió entre los dos tratamientos, permaneciendo similar a lo largo del experimento (Tabla 2a).
El grado de lípidos observados fue mayor en las dietas mejoradas. Adicionalmente, el porcentaje de llenura desde PL1 a PL3 estuvo entre el 60-70%, mientras que en las dietas control solamente llegó al 60%. Estas diferencias fueron más evidentes en PL7, en donde con dietas mejoradas se alcanzó un porcentaje del 70-80% comparado con un 60-70% con alimento control (Fig. 8).
Al término de la corrida de larvicultura, los resultados (Tabla 2b) revelaron en promedio una mayor supervivencia (+10%) en las postlarvas alimentadas con dieta mejorada (92% vs 82%, respectivamente).
Un incremento del 30% en eficiencia productiva (IEP) fue observada en favor de las larvas alimentadas con dieta mejorada. De igual manera, la biomasa cosechada fue mayor en el caso de los animales alimentados con dieta mejorada (+20%), con una larva más grande reflejada en un Pl/g de 165
184 con dieta control.
- AGOSTO 2023 42 NUTRICIÓN
frente a uno
Control Alimento mejorado Alimento mejorado Tasa de Supervivencia (%) 20-80/70-150 µm 0-50/50-110 µm 0-110 µm Z3 95 95 96 M3 80 83 80 Alimento mejorado Alimento mejorado PL 12 0-50/50-110 µm 0-110 µm Tasa de Supervivencia (%) 48 48 Longitud media (mm) 10 11 Peso medio (mg) 6,52 6,48
de
La velocidad de crecimiento en miligramos por día también se vio beneficiada por el alimento mejorado. Finalmente, la eficiencia alimenticia mejoró, lo que fue reflejado en un menor factor de conversión alimenticia (-20%).
De igual manera, la biomasa cosechada fue mayor en el caso de los animales alimentados con dieta mejorada.
Prueba de crecimiento y supervivencia con adaptación a baja salinidad
Para esta valoración se trabajó con 4 tanques (17 y 18 con alimento mejorado, mientras que el 23 y 24 fueron alimentados con la dieta control). Todos los animales correspondían a un mismo código genético.
Este ensayo también fue llevado a cabo en sistemas de producción comerciales en tanques de polietileno circulares de 15 000 litros con aireación suministrada por un aireador, usando tubería de PVC con manguera porosa. Al igual que en el primer experimento, la siembra fue con 60% de nivel y se fue suministrando agua fresca más algas durante los primeros 6 días de cultivo y a partir del octavo día se hacía recambio del 15% hasta llegar al estadio PL4.
A partir de PL5 en adelante, el recambio era de 30% diario. Los sistemas usan agua de mar filtrada con un filtro de piola de 1 micra para el llenado de los reservorios, y al pasar a los tanques el filtrado se realiza con bolso de 10 micras. La temperatura media del
Referencias bibliográficas
Akbari, S., Khoshnod, M.J., Rajaian, H., Afsharnasab, M., 2010. The Use of Eugenol as an Anesthetic in Transportation of With Indian Shrimp (Fenneropenaeus indicus ) Post Larvae 429, 423–429. https://doi.org/10.4194/trjfas.2010.0317
Bardera, G., Usman, N., Owen, M., Pountney, D., Sloman, K.A., Alexander, M.E., 2019. The importance of behaviour in improving the production of shrimp in aquaculture. Rev Aquac 11, 1104–1132. https://doi.org/10.1111/raq.12282
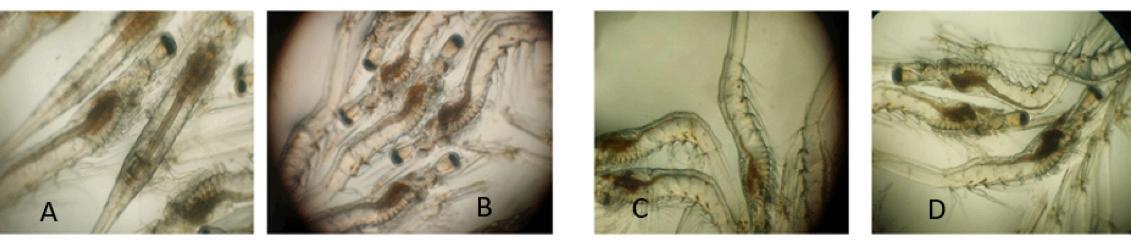
Cruz-suarez, L.E., Ruiz-díaz, P.P., Cota-cerecer, E., Guajardo-barbosa, C., Tapia-salazar, M., Ricque-marie, D., 2008. Revisión sobre Algunas Características Físicas y Control de Calidad de Alimentos Comerciales para Camarón en México. Avances en Nutrición Acuícola IX. 330–370. https://doi.org/10.14763/2017.1.452
Eras-Agila, R.D.J., Meleán-romero, R., 2021. Ecosistemas de producción camaroneros : Estudios y proyecciones para la gestión de costos Shrimp production ecosystems : Studies and projections for cost management 6, 41–59. Gamboa-delgado, J., Le Vay, L., 2009. Artemia replacement in co-feeding regimes for mysis and postlarval stages of Litopenaeus vannamei : Nutritional contribution of inert diets to tissue growth as indicated by natural carbon stable isotopes. Aquaculture 297, 128–135. https:// doi.org/10.1016/j.aquaculture.2009.09.009 García Galano, T., 1998. Nutrición de larvas de camarón. Jaseera, K. V., Kaladharan, P., Vijayan, K. K., Sandhya, S. V., Antony, M. L., & Pradeep, M. A. (2019). Isolation and phylogenetic identification of heterotrophic thraustochytrids from mangrove habitats along the southwest coast of India and prospecting their PUFA accumulation. Journal of Applied Phycology, 31, 1057-1068.Juinio-Menez, M.A., Gontaco, R., 2004. Spiny lobster ecology and exploitation in the South China Sea region. Neiland, A.E., Soley, N., Baron, J., Whitmarsh, D.J., 2001. Shrimp aquaculture : economic perspectives for policy development 25, 265–279. Sorense, M., 2012. A review of the effects of ingredient composition and processing conditions on the physical qualities of extruded high-energy fish feed as measured by prevailing methods. Aquac Nutr 18, 233–248. https://doi.org/10.1111/j.1365-2095.2011.00924.x
Sorense, M., Waagbo, R., Olsen, R.E., 2007. Feed Resources - Feed Technology, in: Thomassen, M., Gudding, R., Norberg, B., Jorgensen, L. (Eds.), Aquaculture Research: From Cage to Consumption. pp. 235–250. Su, Z., Xu, W., Shi, H., He, J., 2020. Research progress on the cannibalistic behavior of Aquatic Animals and The Screening of Cannibalism-Preventing Shelters Review. The Israeli Journal of Aquaculture. Tacon, A.G.J., 2017. Biosecure Shrimp Feeds and Feeding Practices: Guidelines for Future Development. J World Aquac Soc 48, 381–392. https://doi.org/10.1111/jwas.12406
Thomas, M., Poe, A.F.B. Van Der, 1996. Physical quality of pelleted animal feed 1 . Criteria for pellet quality 8401. Welker, T.L., Overturf, K., Snyder, S., Liu, K., Abernathy, J., Frost, J., Barrows, F.T., 2018. Effects of feed processing method (extrusion and expansioncompression pelleting) on water quality and growth of rainbow trout in a commercial setting. Journal of Applied Aquaculture 30, 97–124. https:// doi.org/10.1080/10454438.2018.1433095
43 - AGOSTO 2023 NUTRICIÓN
Tabla 2a. Resultados de las observaciones al microscopio realizadas en PL1- PL3 hasta PL7 en la valoración con larvas de Litopenaeus vannamei
Figura 8. Observaciones al microscopio del porcentaje de llenura y grado de lípidos de las larvas de camarón blanco del Pacífico en PL1 alimentadas con dieta mejorada (A y B) y dieta control (C y D).
Tabla 2b. Resultados del experimento con alimento mejorado versus alimento control realizado con larvas de L. vannamei en tanques de 20 TM con una temperatura media de 32.5°C.
PL1 hasta PL3 PL7 Criterio Tanque 5 Tanque 6 Tanque 7 Tanque 8 Tanque 5 Tanque 6 Tanque 7 Tanque 8 Actividad Buena Buena Buena Buena Buena Buena Buena Buena Porcentaje de llenura 60-70 60-70 60 60 70-80 70-80 60-70 60-70 Grado de lípidos G 2-3 G 2-3 G 2 G 2 G 2-3 G 2-3 G 2 G 2 Porcentaje de vacías 1 1 1 1 1 1 1 1 Porcentaje de deformes 1 1 1 1 2 2 2 2 Necrosis muscular 2 2 2 2 2 2 2 2 Alimento mejorado Alimento control Identificación de los tanques 1 y 2 3 y 4 Estadio PL 10 Días de cultivo 16 Biomasa (kg) 13,19 10,50 Pl/g 165 184 Crecimiento (mg/d) 0,38 0,34 Supervivencia (%) 92 82 FCA 1,61 1,80 IEP* 0,23 0,16
de eficiencia productiva (IEP) = ((Crecimiento semanal (g) x Supervivencia (%) )/ FCA)/100
*Índice
NUTRICIÓN
agua estuvo en un rango de 32°C hasta 33°C. La alimentación en todos los casos fue manual cada 3 horas en estadios de Zoea y Mysis, y en estadios postlarvales cada 2 horas. Las densidades de algas usadas fueron de aproximadamente 60 000 células por ml. La prueba validación fue realizada por duplicado.
Un conteo inicial de las fundas sembradas en los tanques fue realizado. Los resultados se presentan en la Tabla 3.
Los resultados productivos se presentan en la Tabla 4. Una mejora de aproximadamente 17% en supervivencia fue obtenida usando un alimento mejorado, así como también un 25% más de larvas fueron cosechadas con este mismo alimento en comparación con alimento control, aun cuando uno de los tanques control tenía 3,730,000 animales (3,5% más animales que su réplica alimentada con dieta mejorada).
El PL/gramo de tanques control y de prueba fueron similares hasta PL7 (que empieza la aclimatación a baja salinidad hasta 1 ppt). Después de PL7, los tanques control tomaron ventaja en crecimiento, pero considerando que había un 17% menos de población que los tanques con alimento mejorado.
Con el alimento mejorado se cosecharon
Tabla 3. Condiciones iniciales en cuanto a conteo y número de larvas iniciales en el ensayo.
Tabla 4. Resultados de supervivencia, PL/g, biomasa y número de larvas al enfrentar un alimento mejorado con un alimento control en larvas de Litopenaeus vannamei.
1´143.000 animales más en comparación con el control, manteniendo la uniformidad de tallas pese a tener una mayor cantidad de postlarvas. Estas mejoras en crecimiento y supervivencia fueron obtenidas usando solamente una dieta.
Conclusión
En resumen, mediante las mejoras nutricionales y de procesamiento de fabricación del alimento para larvas se puede ofrecer una dieta que sea más estable en el agua y nutricionalmente adecuada para las necesidades especiales en esta fase de crecimiento. Esto permite mejorar los parámetros zootécnicos, así como también
una mayor intensificación en las prácticas que conducen a una mayor capacidad de carga dentro del sistema sin desmejorar la calidad del medio•
Para mayor información escriba a: cesar.molina@skretting.com

- AGOSTO 2023
Número de larvas facturadas Conteo Diferencia Tratamiento Tq 17 3,333,000 2,500,000 3,334,000 +0.03% 100% con alimento mejorado Tq 18 3,333,000 2,500,000 3,590,000 +7.16% 100% con alimento mejorado Tq 23 3,333,000 2,500,000 3,345,000 +0.36% Control Tq 24 3,333,000 2,500,000 3,730,000 +10.64% Control Alimento mejorado Control Promedio de supervivencia (%) 70 53 Promedio PL/g a cosecha 264 172 Suma de biomasa cosechada (g) 17.600 22.200 Suma cantidad de larvas cosechadas 4´664.500 3´521.500
Tanque Número de larvas sembrado



Autor:
Sara Cristina Chaverra-Garcés1
schaverra@ucundinamarca.edu.co
Orinoquia, 26(1), 53–64. https://orinoquia.metarevistas.org/index. php/orinoquia/article/view/731
La acuicultura convencional desarrollada en tierra presenta una serie de debilidades relacionadas con el uso ineficiente del agua, la acumulación de materia orgánica y la liberación de nitrógeno y fósforo que, mal gestionados, terminan contaminando diversas fuentes hídricas. El problema se magnifica cuando se pretende mejorar la productividad aumentando las densidades de siembra por área. Adicionalmente, los sistemas intensivos son susceptibles a la presencia de enfermedades bacterianas, cuyo manejo y control representan grandes inversiones en tratamientos con agentes antimicrobianos.
La implementación de sistemas cerrados, con cero recambios de agua como RAS (Recirculating Aquaculture System) y los sistemas biofloc (BFT, Biofloc Technology) se proponen como una alternativa ambientalmente sostenible (Mansour et al., 2017; Adineh et al., 2019) para el cultivo de peces y camarones (Nguyen et al., 2021), solventando los principales cuellos de botella de la acuicultura de estanques en tierra. Ambas tecnologías utilizan comunidades microbianas para minimizar el exceso de nutrientes (para fines prácticos, el término nutrientes en este texto hace referencia a la concentración de nitrógeno y fósforo) y evitar la proliferación de patógenos en el agua de cultivo.
Estos sistemas cerrados permiten manejar con mayor facilidad parámetros de calidad del agua en comparación con estanques en suelo (Nguyen et al., 2021). Por medio de los procesos de metabolización y absorción de los compuestos nitrogenados, las bacterias quimiautotróficas y heterotróficas consumen el nitrógeno amoniacal excretado por los peces, lo trasforman en compuestos menos tóxicos y/o lo absorben, de tal forma que se presenta una mejoría en las condicionesde calidad del agua y a su vez, la biomasa microbiana aumenta. Sin embargo, en ambos sistemas de cultivo es preciso realizar una adecuada gestión de la materia orgánica, de los compuestos nitrogenados y fosforilados, ya que tienden a acumularse rápidamente convirtiéndose en un problema ambiental (Bao et al., et al., 2018; Jasmin et al., 2020). Asimismo, aumentan la susceptibilidad a enfermedades causadas por patógenos oportunistas, lo cual genera mayores costos operativos.
Las aguas residuales que provienen de la

- AGOSTO 2023 PRODUCCIÓN
Acumulación de sólidos, un aspecto crítico en los sistemas acuícolas intensivos: alternativas para una gestión sostenible
actividad acuícola pueden ser salinas, salobres o agua dulce; por tanto, precisan ser tratadas con tecnologías específicas para cada tipo. Adicionalmente, cada sistema de producción genera cargas de nutrientes y sólidos en cantidad y calidad variables, dependiendo de la especie y la tecnología de cultivo adoptada (Castine et al., 2013; Paniagua, 2020). El objetivo de este documento es brindar una revisión sobre el principal desafío que enfrentan los sistemas RAS y biofloc con relación a la acumulación de sólidos, así como las alternativas que se han propuesto recientemente para mitigar este inconveniente.
Sólidos totales
Los sólidos son partículas de tamaño variable generadas en el ambiente de cultivo y están compuestas por heces, bacterias, microorganismos vivos y muertos, alimento no consumido, metabolitos de peces y desprendimiento de perifiton y bioflóculos (Santaella et al., 2018, Baoet al., 2018; Paniagua et al., 2020). Generalmente, en sistemas de recirculación las principales fuentes de sólidos son el alimento no consumido, las heces y el biofilm(Bao et al., 2018). Se ha reportado que en los sistemas acuícolas intensivos, del alimento que es suministrado diariamente, entre el 25 y 30% es convertido en sólidos (Ebeling et al., 2006). El alimento no consumido, el nitrógeno, fósforo y demás compuestos de la dieta que no son asimilados, se convierten en desechos y rápidamente son transferidos al medio de cultivo (Castine et al., 2013; Jasmin et al., 2020).
En acuicultura, los sólidos generalmente se clasifican en Sólidos Suspendidos Totales (SST), Sólidos Disueltos Totales (SDT) (Arantes et al., 2016) y Sólidos Volátiles Totales (SVT) (Castine et al., 2013; Bao et al., 2018). Otra forma de clasificarlos es como sólidos y semisólidos (Jasmin et al., 2020). Aquellas partículas que se sedimentan y se quedan en el fondo de los estanques constituyen lo que comúnmente se denomina sedimento o lodo (Castine et al., 2013). Este es un tipo de residuo sólido rico en nitrógeno, fósforo, carbonos orgánicos y también contiene trazas de metales (Castine et al., 2013). Estos compuestos en altas concentraciones y mal gestionados ocasionan eutrofización, disminución del oxígeno y toxicidad, afectando así la vida en los ecosistemas acuáticos (Santaella et al., 2018
Bao et al., 2028; Jasmin et al., 2020; Paniagua, 2020).
En RAS, la mayoría de las partículas son menores de 100 µm; entre más pequeña sea la partícula, más complejo será el manejo (Bao et al., 2018). Las más grandes pueden ser removidas fácilmente (Ebeling et al., 2012) empleando filtros mecánicos o tanque de decantación. Los métodos empleados para la remoción de sólidos se basan en la Ley de Strokes o de separación gravitacional, la cual funciona basándose en el principio de sedimentación y velocidades de sedimentación. Partículas de mayor tamaño y de mayor densidad poseen mayor velocidad de sedimentación (Castine et al., 2013; Bao et al., 2018). Sin embargo, las características de los sólidos dependerán de factores como la especie, tamaño del pez, así como de la composición del alimento (Bao et al., 2018).
Manejo de sólidos
Para el manejo de sólidos en los sistemas RAS, generalmente se emplea una unidad de sedimentación que puede ser un sedimentador o filtro mecánico. El objetivo de la filtración mecánica es separar, removery retener tanto partículas en suspensión como sedimentables que provienen principalmente de las heces de los peces y el alimento no consumido. Nozzi et al. (2018) emplearon un filtro de tambor y filtro de flujo radial (RFS) en sistemas acuapónicos donde se evaluó el efecto de diferentes administraciones de nutrientes (efluentes de peces, Fe, P, K, N y Ca) sobre la productividad y calidad del cultivo de lechuga (Latuca sativa),menta (Mentha piperita) y setas (Rungia klossii).
En algunos estudios se han empleado otros dispositivos que desempeñan la misma función. Suhl et al. (2018) evaluaron en un sistema acuapónico de doble recirculación un dispositivo de succión (filtro de succión) en reemplazo de la unidad de sedimentación. El objetivo de los investigadores fue evaluar la implementación de este dispositivo para la reducción de las pérdidas de nitrógeno en el sistema, por medio de la optimización del flujo de nutrientes entre la unidad de producción de peces y la de plantas, mejorando el funcionamientode DRAPS (Double Recirculating Aquaponic System) ydisminuyendo el impacto ambiental generado por la acuicultura convencional.
El dimensionamiento y tipo de filtro mecánico dependerá del tamaño de partícula y de la biomasa de peces que tenga el sistema en acuaponía, de acuerdo con el tipo de cama de crecimiento de las plantas. Por otra parte, en los sistemas biofloc a parte de emplear una unidad de sedimentación, también es posible implementar sustratos artificiales o materiales fijadores para el biofilm con el objetivo de aumentar la superficie de fijación para las bacterias y realizar mayor control sobre los sólidos suspendidos (Lara-Rodrígues, 2017; Pinheiros et al., 2017). Por su parte Green et al. (2019) emplearon un sedimentador de fondo cónico. En otro estudio, Arantes et al. (2016) observaron que la implementación de un sedimentador para controlar el exceso de sólidos en suspensión resulta igual de eficiente en la remoción de sólidos (57-65%) que los recambios para eliminar de manera continua una parte de los bioflóculos.
De esta manera, la remoción de material particulado se da de manera más lenta y sin cambios bruscos. Asimismo, se obtiene un mayor ahorro de agua y mejor desempeño productivo del camarón blanco (Litopenaeus vannamei). Resultados similares obtuvieron Gaona et al. (2016) al evaluar la eficiencia de la remoción de sólidos aplicando un alto o bajo flujo de agua a través de un clarificador de flujo radial, encontrando una mayor velocidad de sedimentación y mejor retención de partículas cuando se empleó un bajo flujo de agua. Generalmente en sistemas biofloc y RAS, el fósforo y el nitrógeno se encuentran disueltos en el agua de cultivo, y son removidos por medio de diferentes tipos de filtros, clarificadores o decantadores.
De acuerdo con Zemor et al. (2019), el uso de clarificadores permite controlar las concentraciones de SST en los sistemas productivos de gran escala; igualmente reduce la cantidad de agua empleada para las renovaciones, así como la generación de efluentes.
Otra alternativa que surge en los sistemas biofloc consiste en cosechar los bioflóculos que están en exceso y utilizarlos como ingrediente en dietas balanceadas para otros organismos de cultivo.
En este sentido, Jung et al. (2020) evaluaron tres tecnologías para colectar los flóculos
- AGOSTO 2023 49 PRODUCCIÓN
provenientes del cultivo de juveniles de tilapia: sedimentación, centrifugación y filtración con membrana. Los mejores resultados de remoción de floc y menor uso de energía se obtuvieron empleando filtración por membrana. Con el fin de evitar la descarga de efluentes (ricos en nutrientes, materia orgánica, sólidos suspendidos y microorganismos) estos son tratados empleando dispositivos de espesamiento de lodos, que luego pueden ser incorporados como insumo en sistemas agrícolas para producción de compost para jardinerías (Van Rijn, 2013; Santaella et al., 2018; Bao et al., 2018; Paniagua, 2020), así como de biogás que puede emplearse como fuente de energía alternativa (Castine et al., 2013; Campanati et al., 2021).
Además de las unidades de espesamiento (reactores) también se emplean lagunas aerobias y anaerobias, así como humedales artificiales para aumentar la eficiencia de degradación biológica de los lodos. Dicha eficiencia está relacionada con el tiempo de retención y exposición a los microorganismos (aerobios y anaerobios) involucrados en la digestión de los compuestos orgánicos e inorgánicos (Van Rijn, 2013; Paniagua, 2020). Estas metodologías se aplican principalmente para el tratamiento de efluentes de RAS de agua dulce, ya que los desechos de los RAS marinos desafortunadamente suelen ser descargados en el ambiente y en muchas ocasiones sin un tratamiento previo (Van Rijn, 2013; Melo Filho et al., 2020).
Durante los últimos años han surgido algunos enfoques para evaluar el impacto ambiental de los sistemas de recirculación y de los sistemas con tecnología biofloc. Uno de los más empleados ha sido la Evaluación del Ciclo de Vida (LCA, por sus siglas en inglés); este enfoque, aparte de muchos otros componentes, contempla como indicadores del impacto ambiental las concentraciones de materia orgánica, nitrógeno y fósforo que se producen en el sistema (Van Rijn, 2013; Campanati et al., 2021). A medida que se desea incrementar la producción acuícola intensiva, aumenta también la necesidad de implementar prácticas que propendan a ahorrar recursos y minimicen la producción de desechos, o bien los recuperen integrándolos nuevamente a la cadena productiva (Campanati et al., 2021).
Manejo de compuestos nitrogenados
Para el manejo y control de compuestos nitrogenados, en RAS se emplean filtros biológicos (bioflitros) con diferentes tipos de sustratos para albergar las bacterias (Cerozi et al., 2017), mientras que en los sistemas biofloc, el biofiltro se encuentra in situ haciendo parte de los bioflóculos en suspensión, que además constituyen como una fuente permanente de alimento para los organismos cultivados (Ekasari et al., 2014). El objetivo del biofiltro, sea ex situ como en RAS o in situ como en los sistemas biofloc, es favorecer la remoción de amonio que servirá como sustrato para aumentar la biomasa de bacterias heterotróficas; asimismo, promover la oxidación del amonio a nitrito y posteriormente a nitrato por acción de las bacterias de amonio y nitrito oxidantes, principalmente Nitrobacter y Nitrosomonas.
En algunos casos se puede adicionar un inóculo comercial para acelerar el proceso de nitrificación y aumentar la biomasa bacteriana. En sistemas biofloc, usualmente se emplea una fuente de carbono externo para promover el crecimiento de las bacterias heterotróficas, las cuales han demostrado ser eficientes cuando el agua de cultivo presenta una relación de C:N entre 15-20:1. Se emplea comúnmente melaza. La adición de la fuente de carbono mejora la tasa de remoción de Nitrógeno Amoniacal Total (NAT) (Kuhn et al., 2009); sin embargo, en algunos casos puede ocurrir un aumento considerable en la concentración de sólidos en suspensión, comprometiendo la disponibilidad de oxígeno disuelto y generando lesiones como obstrucción en branquias, lo cual puede inducir al aparecimiento de enfermedades, así como poner en riesgo la estabilidad y funcionamiento del sistema de cultivo (Bao et al., 2018; Souza et al., 2019).
De manera resumida, el proceso de remoción de sólidos puede realizarse mediante tres fases o tratamientos (Castine et al., 2013). En una primera fase se remueven las partículas de mayor tamaño por medio de gravedad y centrifugación empleando diferentes dispositivos. En una segunda fase o tratamiento secundario, las partículas de menor tamaño (menor de 30 µm) que no pudieron ser retiradas en la fase anterior son removidas empleando filtros para sólidos más
finos (filtros de medios granulares, porosos), fraccionador de espuma, fraccionador de proteínas, filtro de tierra de diatomeasDiatomaceus Earth (DE). Y finalmente, se puede emplear un tratamiento terciario con el objetivo de eliminar fracciones de nitrógeno, fósforo, coloides y otras partículas disueltas de menor tamaño (0,001 -10µm) por medio de micro, ultra y nano filtración (Bao et al., 2018).
Melo Filho et al. (2020) diseñaron un sistema de bajo costo para la remoción de amonio en un cultivo intensivo de camarón blanco (350 camarones/m3) en biofloc, aplicando los procesos de nitrificación y desnitrificación. El sistema estuvo compuesto por un tanque de cultivo con una malla sintética en el interior para favorecer la fijación de bacterias y dos reactores, uno de nitrificación y otro de desnitrificación de lecho móvil, empleando como sustrato biobolas. Adicionalmente, acoplaron un reactor de UV para el control de patógenos. Bajo este esquema se logró obtener una eficiencia de remoción de nitrógeno del 71,3 %.
El éxito de la producción en ambos sistemas de cultivo (RAS, biofloc) consistirá en remover el exceso de desechos, primero del agua de cultivo y luego transformarlos a un costo operativo razonable y con cero o mínimo impacto ambiental.
Estrategias alternativas para la gestión de los sólidos en RAS y biofloc
En sistemas de recirculación a gran escala es común emplear para la desinfección y reacondicionamiento final del agua métodos fisicoquímicos tales como ozonización, desinfección con UV o remediación electroquímica, conocida también como desinfección electrolítica u oxidación electroquímica. Estos procedimientos permiten disminuir las concentraciones de nitrógeno disuelto por medio de procesos de oxidación y generan un efecto antimicrobiano (Van Rijn et al., 2013; Romano et al., 2020; Campanati et al., 2021). Otra alternativa consiste en la instalación de biorreactores de membrana de lodos activados (MBR, por sus siglas en inglés), los cuales poseen gran capacidad de retención de sólidos, incluyendo partículas finas y remoción de microorganismos (Davidson et al., 2019).
- AGOSTO 2023 50 PRODUCCIÓN
Así como los residuos provenientes de la acuicultura de agua dulce, los efluentes de sistemas marinos también deben tratarse, debido a que la inadecuada gestión es responsable en gran medida de la eutrofización en ecosistemas costeros, marinos y ribereños, así como de la salinización de los suelos. De modo tal que el diseño de estrategias eficientes para el tratamiento de efluentes marinos constituye también un desafío.
En ese sentido, los cultivos multitróficos integrados (IMTA, por sus siglas en inglés) surgen como una estrategia de biomitigación que busca generar sistemas equilibrados por medio de la articulación entre diferentes organismos acuáticos cuyos requerimientos son complementarios, ya que pertenecen a diferentes niveles tróficos (Ahmed et al., 2017). De esta manera se aumenta la productividad del sistema por medio de la integración de peces con diferentes hábitos alimenticios, y crustáceos y cultivos de plantas con interés comercial. Al mismo tiempo se logra obtener una reducción significativa de los desechos sin generación de efluentes (Campanati et al., 2021; van Rijn, 2013).
Las partículas en suspensión, fitoplancton y zooplancton pueden ser aprovechados por organismos extractivos orgánicos como los crustáceos y moluscos, mientras que compuestos inorgánicos como el nitrógeno y fósforo pueden ser aprovechados por especies extractivas inorgánicas como las plantas (Ahmed et al., 2017; Campanati et al., 2021). Li et al., (2019) evaluaron un sistema IMTA en tierra, asociando el cultivo de Lubina europea (Dicentrarchus labrax) en RAS, con el cultivo de microalgas (especie extractiva) y juveniles de ostras (Crassostrea gigas) (consumidor primario) en estanques.
En este sistema, el agua del cultivo de peces circuló a través de un filtro mecánico, posteriormente pasó por un tratamiento con rayos UV y finalmente por un biofiltro. Una parte de esta agua retornó al tanque de los peces y la otra fue distribuida al cultivo de algas en raceways. A su vez, el flujo de salida de esta unidad se unió a un tanque de mezcla con aireación y se distribuyó como alimento para las ostras. Los autores evidenciaron una remoción de nitrógeno y fósforo superior
al 96%. Por otra parte, a pesar de que el rendimiento productivo de las ostras no fue significativo, por medio de isótopos estables naturales lograron observar que las ostras consumieron las microalgas. Además, concluyeron que el sistema acoplado RASIMTA tuvo tasas de eliminación de nutrientes muy similares a los sistemas IMTA asociados con macroalgas.
En la misma línea, Brito et al. (2018) evaluaron la biorremediación de aguas residuales de camarón blanco en sistema biofloc usando como biorremediadores organismos de importancia comercial para la pesca y acuicultura: almejas (Anomalocardia brasiliana), algas (Gracilaria birdiae) y peces (Orechromis niloticus). La concentración de sólidos disminuyó hasta en un 74% cuando se usaron almejas. Tal reducción representa un cambio positivo en la calidad del agua y mayor posibilidad de reúso.
En otro estudio, Robinson et al. (2018) emplearon pepinos de mar (Holothuria scabra) como modelo de bioturbación para el tratamiento de aguas residuales de acuicultura. Se evaluó la suplementación con almidón soluble como fuente carbono para llegar a una relación C:N 20:1 con el objetivo de favorecer la fijación de nitrógeno. Se obtuvo un resultado positivo, ya que la fijación resultó ser mayor con respecto al tratamiento control sin adición de carbono (5:1).
La adición de una fuente de carbono externa puede estimular el crecimiento de bacterias heterotróficas que asimilan amonio, así como las rutas metabólicas de reducción disimilatoria de nitrato a amonio. Estos mismos autores en otro estudio (Robinson et al., 2019) reportan los hallazgos obtenidos en la cría intensiva de juveniles de Holothuria scabra alimentados únicamente con efluentes provenientes del cultivo de abulón (Haliotis midae) en RAS terrestre. En este estudio se evaluó la adición de glucosa, almidón y celulosa como fuente de carbono externa para aumentar la relación C:N en el efluente. Se observó que la tasa de crecimiento y la biomasa de juveniles de Holothuria scabra fue mayor cuando la proporción C:N se mantuvo en 20:1. Del mismo modo, como se evidenció en el trabajo anterior, la suplementación con una
fuente de carbono favorece la asimilación de amonio, promoviendo a su vez el aumento de biomasa microbiana la cual puede integrarse nuevamente a la cadena trófica.
Por otra parte, Pinho et al. (2017) utilizaron el efluente generado a partir del cultivo de Tilapia nilótica (O. niloticus) en sistema biofloc para la producción de tres variedades de lechuga (Latuca sativa) en sistemas acuapónicos. Los autores indicaron que las lechugas presentaron mayor crecimiento y mayor productividad (kg/m2) con biofloc en comparación con un sistema de aguas claras. Por lo tanto, el efluente de los sistemas biofloc puede ser usado como un fertilizante alternativo para el cultivo acuapónico de lechuga roja, mantequilla y crespa (crujiente).
En la misma línea, Legarda et al. (2021) evaluaron el efecto del cultivo integrado de lisa (Mugil liza), camarón blanco (Litopenaeus vannamei) y lechuga de mar (Ulva fasciata) en sistema biofloc sobre la calidad del agua, el rendimiento productivo, crecimiento de algas, recuperación de nitrógeno y fósforo y compuestos bioactivos de algas. Se compararon dos grupos, uno con alga, camarón y lisa, y el grupo control sin alga. Ambos grupos en sistema biofloc. El rendimiento final de todo el sistema, así como la recuperación de nitrógeno y fósforo, el contenido de clorofila y carotenoides resultaron ser más altos en el sistema biointegrado en comparación con el grupo control (sin alga). La generación de lodos al final del experimento fue similar en ambos tratamientos; sin embargo, la relación entre la cantidad de lodo producida (kg) y la biomasa final cosechada (kg) resultó ser menor en el sistema con alga, lo cual representa una ventaja desde el punto de vista ambiental.
Por su parte, Chu et al. (2021) evaluaron el efecto de la salinidad (10, 15 y 20 ppt) sobre el crecimiento del camarón blanco en sistema biofloc y de tres especies de plantas halófitas (Atriplex hortensis, Salsola komarovii y Plantago coronopus) en un sistema acuapónico. El agua proveniente del cultivo de camarón fue circulada primero través de un biofiltro y posteriormente se empleó como riego en el sistema acuapónico con halófitas. Los autores indicaron que la salinidad de 15
- AGOSTO 2023 51 PRODUCCIÓN
ppt fue la más adecuada para favorecer el rendimiento productivo tanto del camarón como de las plantas halófitas, y que las especies cultivadas poseen potencial comercial parar ser empleadas en sistemas biointegrados. En la misma línea, Poli et al. (2019) evaluaron un sistema multitrófico también con camarón blanco y tilapia nilótica en sistema biofloc acoplado a un sistema hidropónico con Sarcocornia ambigua. El agua del tanque de camarones se distribuyó al tanque de tilapias, posteriormente por gravedad fue transferida a la unidad de hidroponía y finalmente retornaba al cultivo de camarones. En este estudio, el sistema integrado con Sarcocornia exhibió una mayor remoción de nitrato, mayor remoción de SST y mayor rendimiento (Kg/m3) en comparación con un co-cultivo convencional de camarones y tilapias. Resultados muy similares obtuvieron Pinheiros et al. (2017) al evaluar el cultivo integrado de camarón blanco en biofloc y Sarcocornia ambigua en un sistema acuapónico.
Recientemente, Beyer et al. (2021) evaluaron la eficiencia de S. neei, una nueva especie de halófita, en la remoción de nitrógeno de aguas residuales derivadas de un RAS marino. Los autores emplearon dos sistemas hidropónicos, uno con sustrato de arena y el otro con cama flotante. Se observó 100% de sobrevivencia de las plantas y mayor productividad en el sistema de camas flotantes. Por otra parte, la tasa de remoción de compuestos nitrogenados y fósforo fue significativa. La remoción de amonio y fosfato fue más alta en el sistema con arena (0.68 ± 0.41 g m−2 día−1 y 0.44 ± 0.34 g m−2 día−1 respectivamente), mientras que la mayor absorción de nitrato (0.54 ± 0.91 g m−2 día−1) se observó en el sistema de camas flotantes. Los autores señalan que S. neei tiene potencial para ser usada como biofiltro en sistemas integrados.
Las plantas halófitas del género Sarcocornia y Salicornia han resultado una alternativa promisoria para el tratamiento de efluentes marinos y salobres en los sistemas acuapónicos. Adicionalmente han demostrado una buena remoción de compuestos nitrogenados y un buen rendimiento en sistemas integrados (Pinheiros et al., 2017). Del mismo modo, poseen potencial industrial para la agricultura y la
producción de biocombustibles (Peña et al., 2021).
En la misma línea, recientemente ha llamado la atención el uso de microorganismo debido a su potencial para biorremediación tanto de aguas residuales como para el tratamiento de lodos (Robinson et al., 2018; Jasmin et al., 2020; Paniagua, 2020; Campanati et al., 2021). Los principales desarrollos están orientados al uso de biofilm, esteras o tapetes microbianos y tecnología biofloc.
El mecanismo de acción se basa en el mismo principio: mejorar la calidad de agua por medio de la transformación de compuestos nitrogenados en proteína y biomasa microbiana (Martínez et al., 2014; Paniagua, 2020).
Uso potencial de microorganismos como biorremediador
El mecanismo más práctico para tratar los compuestos nitrogenados tóxicos es por medio de procesos de nitrificación (Melo Filho et al., 2020). Las bacterias que oxidan amonio (Nitrosomonas, Nitrosovibrio, Nitrosococcus y Nitrospira) y nitritos (Nitrobacter, Nitrococcus y Nitrospira) desempeñan un papel fundamental en la biorremediación (Melo Filho et al., 2020; Jasmin et al., 2020). Asimismo, las encargadas del proceso de desnitrificación (Pseudomonas, Bacillus, Rheinheimera, Pannonibacter, Rhizobium, Gordonia, Stenotrophomonas, Brevundimonas, Paracoccus, Rhodococcus, Pseudochrobactrum, Arthrobacter, Gemmobacter y Alcaligenes) (Jasmin et al., 2020) son fundamentales para convertir finalmente el nitrato a nitrógeno gaseoso.
Se ha evidenciado que la incorporación de Bacillus sp mejora tanto la calidad del agua de cultivo como la de los sedimentos por medio de la reducción de los niveles tóxicos de compuestos nitrogenados y fósforo. Asimismo, se ha observado con éxito la aplicación de bacterias fotosintéticas (Cromatiaceae y Clorobiaceae) capaces de degradar compuestos sulfurados. Por su parte, la biorremediación de lodos con poliquetos ha sido una estrategia que toma cada vez más fuerza, dado su potencial en la reducción de la materia orgánica, el nitrógeno y carbono (Jasmin et al., 2020; Campanati et al., 2021).
Manan et al. (2016) encontraron que tanto la variedad de fitoplancton, zooplancton, nematodos y protozoos que conforman el bioflóculo en los sistemas biofloc actúan de manera simbiótica como estabilizadores naturales del medio de cultivo, ya que regulan la cantidad de nutrientes en el agua, y descomponen la materia orgánica. Los mismos autores evidenciaron que bacterias del género Aeromonas y Pseudomonas tienen la capacidad para reducir la materia orgánica del fondo del tanque del cultivo de camarón blanco. De tal forma que el sistema biofloc en sí puede actuar como un mecanismo natural de biorremediación.
Por su parte, Syakti et al. (2016) consideran que es posible generar una valorización a partir de los lodos provenientes de la acuicultura por medio de la aplicación de la tecnología biofloc con fines de biorremediación. En este estudio, los investigadores emplearon lodos provenientes del cultivo de Bagre africano (Clarias glariepinus) en sistema biofloc como uno de los tratamientos para la biorremediación. Observaron que los microorganismos formadores de flóculos fueron capaces de remover sedimentos contaminados con petróleo hasta en un 60% (v/v) cuando este fue suministrado al 5% (v/v). En este sentido, Paniagua (2020) indicó que los microorganismos tienen la capacidad de degradar y mineralizar completamente los compuestos contaminantes debido a que utilizan hidrocarburos del petróleo como fuente de carbono y energía. Sin embargo, es preciso seguir realizando más pruebas donde se evalúe la eficiencia de biorremediación.
La eficiencia de la biorremediación está influenciada por diversos factores tales como el nivel o grado de contaminación y por variables de calidad como pH, temperatura, oxígeno, salinidad y alcalinidad, por mencionar algunos (Jasmin et al., 2020; Paniagua, 2020).
Uso de poliquetos como biorremediadores
Algunos microorganismos como el poliqueto Nereis virens han demostrado la capacidad de aprovechar sustratos sólidos provenientes de RAS marinos, al tiempo que se convierten en biomasa que puede ser utilizada como fuente de alimento para peces y crustáceos (Bao et al., 2018; Mandario, 2020; Mandario
- AGOSTO 2023 52 PRODUCCIÓN
et al., 2021). Asimismo, se ha observado que larvas del poliqueto Marphysa iloiloensis crecen y se alimentan de sedimentos salinos y salobres, y de material en descomposición de estanques de peces (Mandario, 2020; Mandario et al., 2021). Mandario (2020) evaluó la sobrevivencia, crecimiento y producción de biomasa del cultivo de Marphysa iloiloensis empleando dos tipos de dieta: biofloc (BF) y sobrantes de ingredientes secos empleados en la fabricación de alimento balanceado para acuicultura (FS). Este autor observó que los poliquetos alimentados exclusivamente con BF tuvieron mayor contenido de proteína cruda y menor contenido de lípidos (66%–70% proteína y 3.50%–4.50% grasa) en contraste con la dieta FS (61%–64% proteína y 12%–13% grasa). La mayor biomasa se obtuvo combinando las dos dietas. Asimismo, evidenció que altas concentraciones de fósforo y azufre no afectaron negativamente la sobrevivencia ni el rendimiento productivo de los poliquetos.
A su vez, Liu et al. (2020) combinaron el uso de un filtro de arena con poliquetos (Perinereis helleri) con el objetivo de favorecer la remoción de nutrientes del efluente del cultivo de camarón tigre (Penaeus monodon). Perinereis helleri resultó ser una especie eficiente debido a que posee hábitos alimenticios omnívoros y se adapta muy bien a altas concentraciones de materia orgánica. Los autores de este estudio indican que es aconsejable evacuar el contenido intestinal de los poliquetos antes de iniciar el proceso de biorremediación para evitar la transferencia de algunos virus, en este caso el de la necrosis hematopoyética infecciosa (IHN).
De acuerdo con Yarnold et al. (2019) el uso de poliquetos para el tratamiento de aguas residuales derivadas de la acuicultura constituye una alternativa para fomentar la economía circular por medio de la integración del cultivo de peces, camarones, crustáceos, moluscos y la producción de una fuente valiosa de proteína que puede ser empleada como ingrediente en la dieta de animales de producción.
Por su parte, Bao et al. (2018) consideran que existe aún mucho por hacer con respecto a la reutilización de los sólidos en la acuicultura,
especialmente hablan- do desde el enfoque del desarrollo sostenible.
Potencial de las algas como biorremediadores Tanto micro como macroalgas desempeñan un rol importante en la remoción de compuestos inorgánicos derivados de la acuicultura marina. Las algas exhiben altas tasas crecimiento y de asimilación de nutrientes. Es preciso seleccionar especies que se adapten a las condiciones ambientales variables y que posean valor industrial (Castine et al., 2013; Paniagua, 2020). Las algas tienen gran capacidad para biorremediar los efluentes de la acuicultura por medio de la remoción de nutrientes adicionalmente, producen compuestos de gran valor nutricional e industrial, y pueden ser integradas en policultivos, así como en cultivos multitróficos (Castine et al., 2013; Campanati et al., 2021). Mawi et al. (2020) evaluaron el potencial de biorremediación de Gracilaria edulis y Gracilaria changii usando los efluentes de camarón blanco cultivado en un sistema de recirculación outdoor. Estos autores observaron que la tasa de remoción de amonio y nitrato de ambas especies de macroalgas fue de 72,5% y 71% para amonio, y 58,8% y 56,8% para nitrato, en contraste con el tratamiento control (sin macroalgas) que presentó una tasa de remoción de amonio del 2% posiblemente debido a la actividad microbiana y fitoplanctónica. Con respecto al control de fosfatos, la tasa de remoción fue de 45,9% y 43,5 % respectivamente. En este estudio se evidenció que Gracilaria edulis y G. changii son dos especies que presentan un buen crecimiento, adecuada biofiltración y favorecen la productividad del camarón (Mawi et al., 2020).
Por su parte, Wu et al. (2015) emplearon G. chouae para evaluar el flujo de nutrientes del cultivo integrado con Sparus macrocephalus Se obtuvo una eficiencia de eliminación de nitrato (NO3) del 37,76%, la remoción del nitrito (NO2) fue 36,99%, amonio (NH +) 29,27% y fosfato (PO4) 40,64%. Asimismo, se evidenció una disminución en la concentración de clorofila y densidad planctónica. Los autores informaron que esta alga roja constituye una especie para la biorremediación eficiente en sistemas IMTA. En la misma línea, Pires et al. (2021) evaluaron dos medios de cultivo para el crecimiento del alga roja Kappaphycus
alvarezii. Uno consistió en una solución de 50% de von Stosch y agua de mar destilada y el otro fue una solución con 25% de biofloc (BFT) proveniente del cultivo intensivo de camarón y agua de mar destilada. Las algas fertilizadas con BFT presentaron una menor remoción de nitrato y fosfato (22,97% y 83,58%) en contraste con las de la solución de von Stosch (58,39% y 100%) Sin embargo, se observó una mejor calidad de la carragenina. Asimismo, los autores reportan que el efluente de biofloc para el cultivo de Kappaphycus alvarezii puede ser utilizado como fertilizante en reemplazo de la solución de von Stosch. Igualmente, mencionan que la integración de las algas, los microorganismos autotróficos y heterotróficos constituyen un biofiltro eficiente en los sistemas IMTA que se combinan sistemas biofloc.
En otro estudio, de Oliveira et al. (2019) analizaron el crecimiento de Chlorella vulgaris utilizando aguas residuales del cultivo de alevinos de tilapia nilótica a baja salinidad en sistemas biofloc. Se evaluaron tres proporciones del efluente como medio de cultivo: 0% (medio Provasoli), 50% y 100%. A pesar de que al final del experimento el crecimiento del alga resultó mayor en el tratamiento control (0% efluente), los primeros 4 días de cultivo se observó que la tasa de crecimiento y la densidad de células fue mayor en los tratamientos que contenía efluente de biofloc. Adicionalmente se evidenció que hubo una remoción del amonio (NH4) del 79,3%; nitrito del 80%, nitrato del 93,6% y 48, 3% de fosfato cuando se empleó como medio de cultivo 100% efluente. Los autores señalan que el uso de efluente de biofloc es favorable para el crecimiento del alga Chlorella vulgaris y representa un ahorro en el uso de reactivos; además, esta alga tiene potencial para emplearse en Acuicultura Multitrófica Integrada con alevinos de tilapia nilótica.
La capacidad de remoción de nutrientes por parte de las algas está influenciada por aspectos ambientales como luz, temperatura, flujo de agua, pH, etc.; fisiológicos y por los requerimientos de nutrientes (Wu et al., 2015; Mawi et al., 2020).
A parte de los beneficios del uso de algas como organismos biorremediadores, estas constituyen una fuente nutriente de gran
- AGOSTO 2023 53 PRODUCCIÓN
valor para dietas acuícolas y poseen gran potencial para ser usados como reemplazo de la harina y aceite de pescado. Tal como mencionan Yarnold et al. (2019) en su revisión.
Conclusiones
Durante los últimos años se ha evidenciado que existe un creciente interés por valorar de mejor manera los recursos, favorecer el reciclaje de nutrientes y subproductos y fomentar la economía circular con ánimos de aumentar la producción de alimentos acuáticos de manera sustentable (Campanati et al., 2021). No obstante, los trabajos sobre el tratamiento de efluentes provenientes de acuicultura marina son pocos en comparación con las investigaciones sobre el manejo de efluentes de acuicultura de aguas continentales.
La aplicación de la biorremediación aumenta cada vez más su popularidad debido a que se propone como una tecnología ambientalmente adecuada para minimizar los impactos generados por los efluentes que afectan los ecosistemas costeros, marinos y ribereños, lograr la sostenibilidad y la resiliencia (Paniagua, 2020).
Diversas técnicas para reusar los residuos provenientes de la acuicultura se han implementado con el objetivo de fomentar la economía circular; sin embargo, es preciso seguir avanzando en las investigaciones sobre el uso de microorganismos y microalgas con potencial para biorremediación y bioprospección. La construcción de humedales artificiales constituye una alternativa eficiente y ecológica para la gestión de cargas masivas de efluentes acuícolas; sin embargo, las grandes áreas que se requieren para su establecimiento y funcionamiento exitoso siguen siendo un gran desafío.

Asimismo, la obtención de biogás a partir de la digestión anaerobia de los lodos constituye una alternativa que puede satisfacer parte de la energía necesaria para el funcionamiento de los RAS y sistemas biofloc.
De acuerdo con Martínez et al. (2014), el uso de microorganismos como biorremediadores y como fuente de proteína representa una estrategia con gran potencial para avanzar hacia una acuicultura sostenible; sin embargo, es preciso adelantar investigaciones orientadas a la
caracterización taxonómica, mecanismos de acción y requerimientos ambientales que permitan aprovechar al máximo los beneficios que estos ofrecen. Existe un gran desafío con respecto al conocimiento sobre el tratamiento de compuestos de mayor toxicidad, metales pesados, gases, etc., y sobre dinámicas entre microorganismos.
Con respecto al uso de algas, es necesario avanzar en investigaciones a mayor escala, enfocadas en la obtención de compuestos a partir de microalgas cultivadas con efluentes de acuicultura de manera más eficiente y que sea económicamente viable.
Las entidades gubernamentales encargadas de formular las políticas ambientales deberán diseñar directrices para involucrar diferentes actores del sector acuícola para contribuir con el uso sostenible de los recursos naturales y la reutilización de los residuos en la cadena productiva•
Para mayor información escriba a: schaverra@ucundinamarca.edu.co
- AGOSTO 2023 PRODUCCIÓN

Evolución del Fenómeno de El Niño. Recomendaciones para el sector camaronero
Según el último reporte de la Oficina Nacional de Administración Oceánica y Atmosférica de los EE.UU. (NOAA), los pronósticos más recientes indican que el fenómeno de El Niño persistirá durante el 2023-2024 (Fig. 1).
La tendencia favorece el crecimiento continuo de El Niño, empezando en octubre - noviembre de 2023, alcanzando su punto máximo entre noviembre y febrero de 2024, con una intensidad de moderada a fuerte, y cuyas predicciones y calentamiento se mantendrán hasta después de abril del año próximo. Siendo este un evento que se vuelve “históricamente fuerte”, comparado con el evento de 1997-98, existe una probabilidad sobre el 90% de que El Niño continúe durante 2024, debido a que el océano es rápido en adquirir calor y lento para desprenderse del mismo; por eso, los modelos predicen que para abril de 2024 todavía tendríamos 1 grado más de temperatura que la normal para esa época del año.
Asimismo, podemos observar la más reciente imagen de la NOAA publicada el 6 de agosto (Fig. 2), donde se indica la temperatura superficial del mar a esa fecha y no se observa probabilidad de que la corriente de Humboldt ascienda hacia la línea ecuatorial, trayendo consigo aguas frías que contribuirían a disminuir los efectos del fenómeno.
En lo que respecta al calentamiento de las aguas, en cuanto a profundidad, la información gráfica disponible indica que el calentamiento frente a nuestras costas se mantiene hasta los 350m de profundidad al 1 de agosto de 2023 (Fig.3).
Es necesario indicar que, a más de los efectos que El Niño pueda generar, hay otros dos aspectos no posibles de pronosticar pero que estarían influenciando en el fenómeno, y estos son el calentamiento global, cuyos efectos los estamos evidenciando en el país, con temperaturas atmosféricas extraordinarias y una alta radiación ultravioleta; de allí el mensaje del secretario general de la ONU, António Guterres, quien indicó que “ya no estamos en etapa de calentamiento, sino de ebullición global”. Y el segundo factor es el cambio climático, que se está manifestando en otras regiones

- AGOSTO 2023
56
Autor: Leonardo S. Maridueña Director de Ambiente Cámara Nacional de Acuacultura
lmariduena@cna-ecuador.com
con fuertes lluvias o sequías, inundaciones, movimientos sísmicos, huracanes, tornados, deshielo de los casquetes polares y otros fenómenos climáticos que conducen a una elevación del nivel del mar.
A esta actualización de la información la complementamos con la siguiente figura (No. 4) de la Columbia Climate School, donde se muestran las áreas secas y lluviosas.
La tercera Ley de Newton o principio de
acción y reacción. Esta ley plantea que toda acción genera una reacción de igual intensidad, pero en sentido opuesto. Es decir, siempre que un objeto ejerza una fuerza sobre otro, este último devolverá una fuerza de igual magnitud, pero en sentido opuesto al primero. Igual sucede con El fenómeno del Niño, se debe esperar que a continuación del Niño se genere La Niña, termino con el cual se conoce al enfriamiento anormal de las aguas ecuatoriales del Océano Pacifico Tropical, la cual se caracteriza por bajas
temperaturas tanto del aire como en el agua provoca fuertes sequías en las zonas costeras del Pacífico.
Impacto en el sector camaronero ecuatoriano
Con lo mencionado anteriormente, podemos inferir que todas las áreas donde se asientan las camaroneras y laboratorios son susceptibles a inundación y erosión por estar situados en tierras bajas; esto incluye las provincias costeras de Esmeraldas,
57 - AGOSTO 2023 AMBIENTE
Fig. 3. Predicciones del desarrollo de El Niño 2023-2024. Fuente: NOAA
Fig. 2. Temperatura ecuatorial sub superficial al 1 de agosto. Fuente: NOAA
Fig. 1. Anomalías (°C) promedio de la temperatura de la superficie del océano (SST, por sus siglas en inglés) para la semana centrada el 2 de agosto de 2023. Fuente: NOAA
Manabí, Santa Elena, Guayas y El Oro, por lo que se les recuerdan las medidas a tomar en prevención a un fuerte El Niño:
- Mejorar las vías de acceso.
- Reforzar el borde costero, especialmente en áreas descubiertas de vegetación de manglar, con sacos reforzados de arena o sedimento, recordando que el nivel del agua se incrementa durante este período probablemente entre 30 y 50cm.
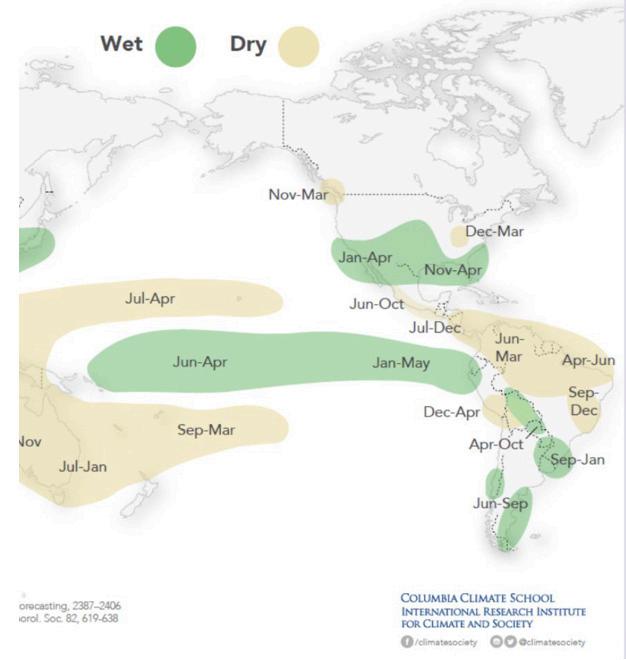
- Proteger los muros de las piscinas y compuertas, para evitar el escape de la producción.
- Evitar que a las bodegas de insumos ingrese agua.
- Revisar el estado de los muelles y corregir cualquier defecto; recordar que en muchos casos es el único sitio de acceso a la camaronera.
- Dotar al personal de campo de botas, ponchos de agua, sombreros, bloqueadores solares.
- Dotar al personal de repelente de insectos. Estos son vectores de muchas enfermedades, especialmente aquellas transmitidas por mosquitos.
- Dotar de agua purificada en ciertos puntos estratégicos de las camaroneras; no olvidemos que el excesivo calor produce deshidratación.
- No deben guardarse alimentos preparados. Preparar solo lo que van a consumir ese día. Las altas temperaturas degradan la composición de los alimentos.
- Dotar de ventiladores, especialmente en los dormitorios, además de mosquiteros.
- Revisar el estado de los pozos sépticos para reducir materia orgánica; pueden utilizar bacterias y carbonato de calcio.
- Incrementar el control de plagas; los roedores buscan áreas secas para sobrevivir.
- No acercarse a los playones. Los cocodrilos pueden estar en época de reproducción y son agresivos cuidando sus nidos.
- Los contenedores de combustible no deben llenarse al máximo de su capacidad.
- Tener cuidado de que las cubetas que contienen residuos de combustible no se vayan a rebosar y se presente un derrame de mezcla de agua con diésel que puede llegar a las piscinas camaroneras.
- Bajar en lo posible densidades de siembra; si un accidente o escape ocurre, la pérdida será menor.
- Profundizar los canales de drenaje e
implementar bombas de drenaje en lo posible.
- Proteger equipo caminero, estaciones de bombeo y otros equipos que normalmente permanecen al aire libre.
- Revisar las conexiones eléctricas y los cables que las conducen.
- Prevenir la entrada potencial de patógenos y depredadores. Después de cada El Niño fuerte se ha presentado una enfermedad
severa en el camarón debido al desequilibrio ambiental que se genera; por eso, se recomienda revisar la calidad de agua antes de bombearla, y si hay mucho sedimento, abstenerse de bombear lo más que se pueda.
- Utilizar carbonato de calcio en el canal de aducción. El incremento de temperatura en el agua genera la proliferación de virus y bacterias patógenas (Fig. 5)•
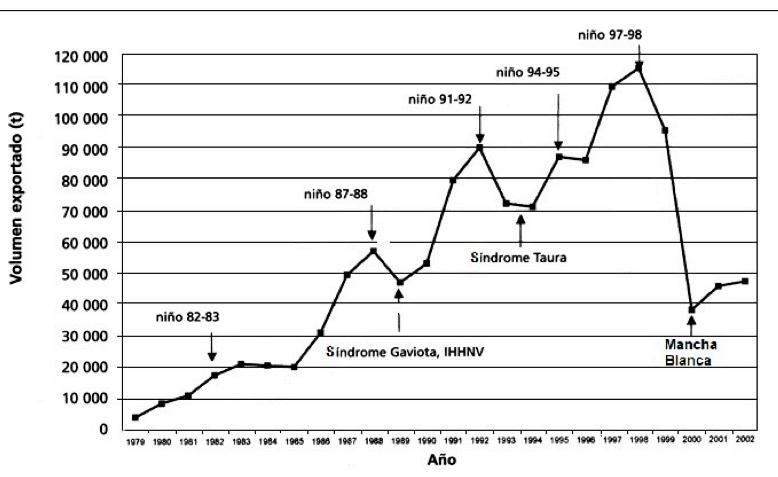
58 - AGOSTO 2023
AMBIENTE
Fig. 4. Áreas de alta precipitación (wet) y sequía (dry) pronosticadas para los próximos meses
Fig 5. Producción camaronera y El Niño Elaboración: CNA

Calidad del agua, calidad de vida y producción
EDITORIAL
Agua y sostenibilidad en la industria camaronera: equilibrando crecimiento y conservación
Amedida que la población global se expande, asegurar un suministro de alimentos suficiente y sostenible se ha convertido en una prioridad en todo el mundo. En este contexto, la industria acuícola camaronera ha experimentado un crecimiento significativo y emerge como una solución para satisfacer estas necesidades en alza.
Sin embargo, la expansión del sector no puede llevarse a cabo a expensas del medio ambiente. Este progreso plantea interrogantes sobre la capacidad de nuestros ecosistemas marinos para mantener una producción sostenible a largo plazo. Por ello, es crucial comprender la importancia de un recurso vital para la vida y para la industria: el agua.
La calidad del agua no solo afecta directamente a los camarones y a su producción, sino que también tiene un impacto en el ecosistema que lo rodea. Un ambiente acuático saludable es esencial para prevenir enfermedades y reducir el estrés que puede sufrir el animal. Esto, a su vez, asegura un cultivo exitoso y sostenible a largo plazo.
En esta edición de la revista, nos enorgullece presentar el artículo "Calidad del agua, calidad de vida y producción". Este análisis es un llamado a la reflexión y la colaboración. Al comprender cómo la calidad del agua y la acuicultura están intrínsecamente entrelazadas, podemos avanzar hacia un futuro sostenible.
Esperamos que este texto inspire conversaciones significativas y desencadene acciones informadas que tracen una ruta hacia adelante, que equilibren el crecimiento de la industria con la preservación ambiental. Juntos podemos garantizar un suministro de camarón que satisfaga las demandas futuras, sin comprometer la salud de los ecosistemas ni la prosperidad de las generaciones por venir.
Autores:
Pamela Nath
Sally Tabares

Sustainable Shrimp Partnership (SSP)
pnath@sustainableshrimp.org
Imagina que vives en una casa que está completamente cerrada, en donde las ventanas y puertas están herméticamente selladas y no existe la posibilidad de abrirlas ni de ventilar. Imagina que la única fuente de aire es una pequeña abertura en el techo.
Al principio, el aire en la casa estará limpio y fresco. Podrás respirar con tranquilidad. Pero, con el paso del tiempo, a causa de la falta de ventilación y a la acumulación de polvo, suciedad y contaminantes del interior, el aire se volverá cada vez más denso y tóxico. A medida que pasan los días, sentirás que respirar se hace más difícil y te empezarás a agobiar. Los malos olores y la falta de oxígeno dispararán tus niveles de estrés y ansiedad. Tu salud comenzará a deteriorarse y, como consecuencia, te volverás más débil. Ahora, si ese ambiente no mejora, imagina cuáles son tus probabilidades de sobrevivir.
En la acuicultura, el camarón se enfrenta a un escenario similar cuando existe una mala calidad del agua en las piscinas de producción. El agua proporciona un hogar y representa un recurso crítico para el camarón, porque es el medio donde come, respira y se desarrolla. Al igual que en el ejemplo, si el agua se contamina, el ambiente se vuelve tóxico para este crustáceo. De la misma manera, si el agua no contiene las sustancias necesarias para su crecimiento, reproducción y bienestar, no podrá sobrevivir.
Someter al camarón a una baja calidad de agua genera estrés en el animal. El coordinador técnico de Sustainable Shrimp Partnership (SSP), Leonardo Maridueña, explica que en este estado, un camarón presenta un comportamiento errático y una navegación desorientada, similar al que un humano presenta en caso de asfixia. Cuando los niveles de oxígeno disuelto en el agua son bajos, el camarón trata de conseguir oxígeno de la superficie, exponiéndose a los depredadores. Muchas veces, un estrés prolongado lo debilita y lo vuelve propenso a desarrollar enfermedades a causa de bacterias patógenas que pueden existir en el medio.
“La calidad del agua es la que da la sostenibilidad a la producción del camarón, porque de ella dependen todos los factores de desarrollo, crecimiento y bienestar del animal. Una mala calidad de agua con toda seguridad tendrá una mala producción”, indica Maridueña, quien también es doctor en Ambiente, Acuicultura y Pesca.
Es importante tener en cuenta que si una camaronera descarga agua de mala calidad, esto tendrá un impacto negativo en el cuerpo de
60 - AGOSTO 2023 SOSTENIBILIDAD
Pamela Nath Directora
60
agua receptor. Resulta imprescindible gestionar cuidadosamente la calidad del agua y adoptar prácticas de manejo sostenibles para mantenerla dentro de los límites máximos permitidos en cada parámetro. Esto implica realizar mediciones constantes y estar preparados para implementar acciones correctivas en caso de ser necesario.
El director de Producción de Grupo Almar, Eduardo Reyes, expone que entre todos los parámetros a evaluar, el oxígeno disuelto siempre es el más importante. “Si en una piscina los niveles de oxígeno son bajos, el camarón no podrá crecer adecuadamente y el productor enfrentará problemas de mortalidad”, declara, “aunque utilice el mejor alimento balanceado, con todos los nutrientes necesarios, si no hay suficiente oxígeno en el agua, no será efectivo”.

La importancia de mantener el oxígeno disuelto en equilibrio está estrechamente relacionada con el concepto de capacidad de carga del agua. Un cuerpo de agua es capaz de procesar y eliminar una cantidad específica de materia orgánica como alimento balanceado, organismos en descomposición, etc., antes de saturarse. Si se excede el límite natural de materia orgánica que el agua puede soportar, es decir, si se excede su capacidad de carga, se puede producir una eutrofización que agotará los niveles de oxígeno disuelto y contaminará el agua.
En este contexto, el aumento en la densidad de siembra y, por ende, el incremento en la producción de camarón, cobran una gran relevancia debido a su impacto directo en la capacidad de carga y calidad del agua. Reyes destaca que cuando se aumenta la densidad de siembra, se incrementa automáticamente la cantidad de alimento suministrado.

“El alimento es usualmente la carga de nutrientes más significativa que se introduce en la piscina. Todos los nutrientes que componen el balanceado, como el nitrógeno, carbono, fósforo, entre otros, son esenciales para el desarrollo del camarón. Sin embargo, si se introduce una mayor cantidad de este alimento en la piscina, el grado de contaminación del agua aumentará y se correrá el riesgo de que estos nutrientes se acumulen en el medio acuático”, continúa.
El aprovechamiento y la eficiencia del balanceado son limitados. Una porción significativa es asimilada por el camarón y el excedente se integra en el entorno, ya sea a través del proceso de digestión del camarón o disolviéndose en el agua.
“A medida que el sistema se carga de nutrientes, aumenta la demanda de oxígeno para llevar a cabo el proceso de oxidación y transformación de dichos nutrientes. Entonces, si no se monitorea ni se cuida adecuadamente el oxígeno disuelto y al mismo tiempo se aumenta la densidad de manera indiscriminada, la calidad del agua en la piscina se deteriorará”, expresa.
Para incrementar o corregir el nivel de oxígeno en el agua, en ocasiones se recurre al uso de peróxido de hidrógeno. Reyes señala
61 - AGOSTO 2023 SOSTENIBILIDAD
que este producto es efectivo y considera que toda camaronera debería contar con un stock del mismo. Sin embargo, advierte que debe utilizarse solo como una medida de emergencia.
“Tenerlo como un producto recurrente, donde todas las noches es necesario su uso, es una mala señal. Si una camaronera tiene problemas de oxígeno significa que algo está mal y hay que atenderlo. Hay dos opciones: bajar densidades de siembra o poner aireación”, comenta Reyes. “En el caso de que la camaronera no pueda invertir en aireadores, esto es un indicador de que no está preparada para las densidades con las que está trabajando y deberá reducirlas”. Reyes añade que otro de los parámetros de especial importancia que se deben cuidar son los elementos nitrogenados, como el amonio y el nitrito, que pueden volverse tóxicos para los organismos acuáticos. También menciona al pH, que requiere atención porque, si se altera, puede afectar el metabolismo y otros procesos fisiológicos del camarón.
En algunas camaroneras se están implementando nuevas tecnologías para medir el oxígeno y la temperatura en tiempo real y de manera más eficiente. Como es inevitable que en ciertos días los niveles de oxígeno disminuyan, “con la información en tiempo real, es posible anticipar estas caídas
y tomar decisiones cruciales, como reducir la cantidad de alimento en momentos críticos”, expone el especialista.
Sin embargo, también subraya que lo más importante en las mediciones de parámetros de calidad del agua es garantizar su precisión y confiabilidad. “La medición de parámetros implica poner atención en muchos detalles. No me preocupa tanto si un laboratorio tiene los equipos más modernos y avanzados para medirlos, sino que los esté midiendo con equipos debidamente calibrados, que los esté verificando regularmente, y que pueda dar confianza de que lo que está reportando es correcto. Medir mal puede llevar a tomar decisiones equivocadas, y eso es lo peor que puede pasar”, apunta.
En cuanto a otras tecnologías que se utilizan en las camaroneras, Reyes indica que cualquier herramienta que mejore la eficiencia en el manejo del alimento balanceado tendrá un impacto directo en la calidad del agua. “Al ser más eficientes en el uso del balanceado, se reduce la presión sobre el sistema y se requiere menos cantidad de alimento para producir la misma cantidad de camarones”, informa.

Al implementar estos instrumentos, la producción se vuelve más sostenible, ya que se logra mejorar las conversiones de alimentos. Según el Grupo de Trabajo de
Acuicultura sobre la Huella Ambiental, del cual SSP es miembro, uno de los puntos críticos más significativos en la cadena de suministro de la acuicultura es el alimento balanceado. Por lo tanto, si los productores se enfocan en optimizar su uso y lo convierten en un insumo utilizado de manera eficiente, reducirán considerablemente su huella de carbono y a su vez, disminuirán la cantidad de materia orgánica vertida en las piscinas.
La evaluación y certificación de una finca camaronera responsable involucra la consideración de diversos criterios, y entre ellos destaca la calidad del agua como uno de los más significativos. En el caso de SSP, la preservación del agua adquiere un carácter fundamental.
Desde sus inicios, uno de los enfoques principales de esta iniciativa ha sido la preservación de la calidad del agua, verificando que el líquido empleado para la producción sea devuelto al entorno natural en condiciones equivalentes a las que tenía al ingresar a las piscinas.
“Cuando creamos la Calificación SSP, en colaboración con nuestro consejo asesor, reflexionamos profundamente sobre la razón detrás de nuestra responsabilidad en la preservación del agua”, indica Pamela Nath, directora de SSP, “el agua es un recurso natural esencial para la producción, y esta
62 - AGOSTO 2023
SOSTENIBILIDAD
consideración nos llevó a cuestionarnos cómo asegurar su uso sostenible. Reconocimos que si empleamos el agua en nuestros procesos, es necesario garantizar que no tengamos un impacto negativo en este recurso vital”.
Dentro de los criterios generales para evaluar la calidad del agua, las fincas SSP deben asegurar un impacto ambiental mínimo, lo que se demuestra mediante monitoreos constantes en cada ciclo de producción. Estos muestreos sistemáticos comparan los parámetros fisicoquímicos del agua que entra en la finca con los parámetros fisicoquímicos del agua que se descarga.
“Para asegurar la transparencia y la calidad de los resultados, SSP elige de forma aleatoria las piscinas y las fechas en las que los laboratorios acreditados llevarán a cabo la recolección y el análisis de las muestras. Además, SSP recibe los resultados de estos análisis al mismo momento que los productores. Esto permite que ambas partes conozcan la calidad del agua en ese momento y brinda la oportunidad a SSP de notificar al productor para que tome medidas correctivas si es necesario”, explica la gerente de Proyectos de SSP, María Fernanda Vilches.
Entre los parámetros fisicoquímicos que se evalúan se encuentran el pH, la demanda química y bioquímica de oxígeno, el nitrógeno, el fósforo y los sólidos suspendidos totales, los coliformes fecales y los aceites.
Además, SSP reconoce el papel crucial desempeñado por los manglares en los ecosistemas y su función reguladora en la eutrofización, lo que a su vez contribuye a la preservación de la calidad del agua. Por esta razón, SSP colabora estrechamente con destacadas organizaciones de sostenibilidad a nivel mundial para establecer una hoja de ruta que fomente la conservación y restauración de los manglares a lo largo de la costa ecuatoriana. Entre las instituciones participantes se incluyen World Wildlife Fund (WWF), The Nature Conservancy (TNC), Conservation International, Earthworm Foundation y Fondagua.
Asimismo, otras certificaciones reconocidas destacan la importancia de la calidad del
agua para la sostenibilidad de la industria. El Aquaculture Stewardship Council (ASC) es un ejemplo de ello.
“La industria de la acuicultura proporciona más de la mitad de todos los mariscos consumidos cada año a nivel mundial. El cultivo de mariscos puede tener efectos adversos en los ecosistemas locales, los recursos hídricos y las condiciones laborales si no se realiza de manera responsable”, señala la jefa de Impactos de ASC, Jill Swasey. “En ASC reconocemos el papel que debe desempeñar la acuicultura responsable para mitigar estos efectos y contribuir a impactos positivos que son verificados de manera independiente a través de nuestros requisitos. La calidad del agua es una pieza fundamental del rompecabezas que se está abordando ahora y en el futuro”, continúa.
Como parte de su próximo Estándar para Fincas, ASC está desarrollando un conjunto más sólido de indicadores de calidad del agua para abordarla en todos los principales sistemas de producción. Los indicadores propuestos han sido desarrollados por un grupo de trabajo técnico compuesto por expertos, y se basarán en una forma de manejo por regiones. Esto permitirá entender cómo afecta la calidad del agua en las áreas
donde se encuentran las granjas certificadas por ASC.
En última instancia, este enfoque en la calidad del agua en la producción de camarones no solo garantiza la viabilidad económica de la industria acuícola, sino que también protege los ecosistemas acuáticos y la salud humana. Así como en la metáfora de la casa herméticamente cerrada, donde la falta de aireación afecta la calidad del aire y la calidad de vida, la mala calidad del agua en las piscinas de camarones puede tener efectos adversos en la salud y el bienestar de los crustáceos, así como en la sostenibilidad de la producción a largo plazo.
A través de prácticas responsables, monitoreo constante y colaboración entre la industria y las organizaciones de sostenibilidad, se puede mantener un equilibrio vital entre la producción de camarones, la calidad del agua y la salud de los ecosistemas marinos, asegurando así un futuro más saludable tanto para la industria acuícola como para el medio ambiente•
Para mayor información escriba a: pnath@sustainableshrimp.org

63 - AGOSTO 2023 SOSTENIBILIDAD


ÍNDICE Edición 154 - Agosto 2023 66 71 74 Exportaciones de camarón y tilapia. Reporte de mercado de EE. UU. Reporte de mercado de China.
ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: EVOLUCIÓN DE EXPORTACIONES 2010 -2022
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: COMPARATIVO MENSUAL (Millones de Libras) 2019 - 2023
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- AGOSTO 2023 66 ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: PARTICIPACIÓN POR DESTINO (Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: PRINCIPALES PAÍSES DESTINOS DE EXPORTACIÓN (Millones de Libras)
Fuente: Estadistic S.A.
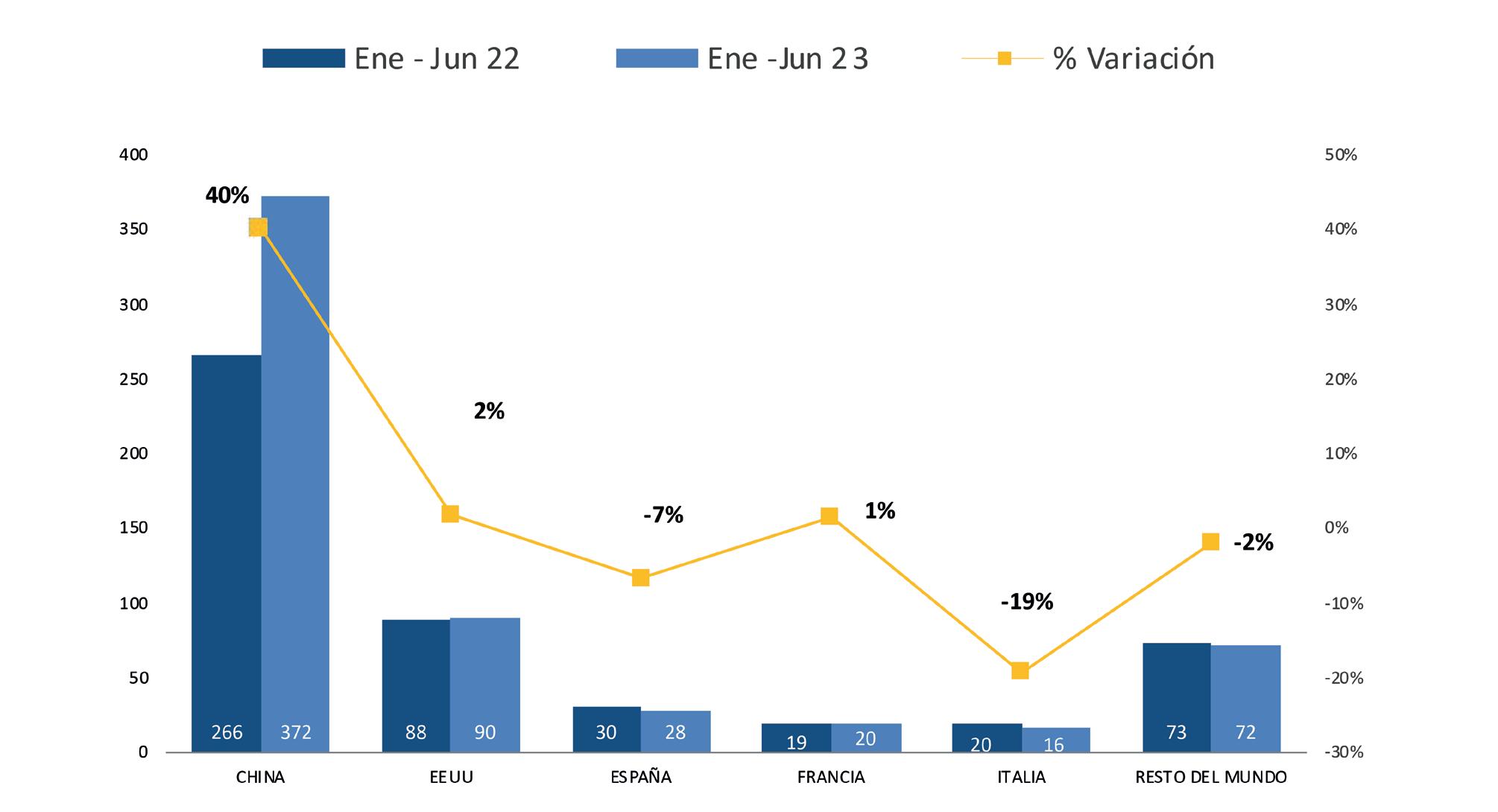
Elaborado por: Cámara Nacional de Acuacultura
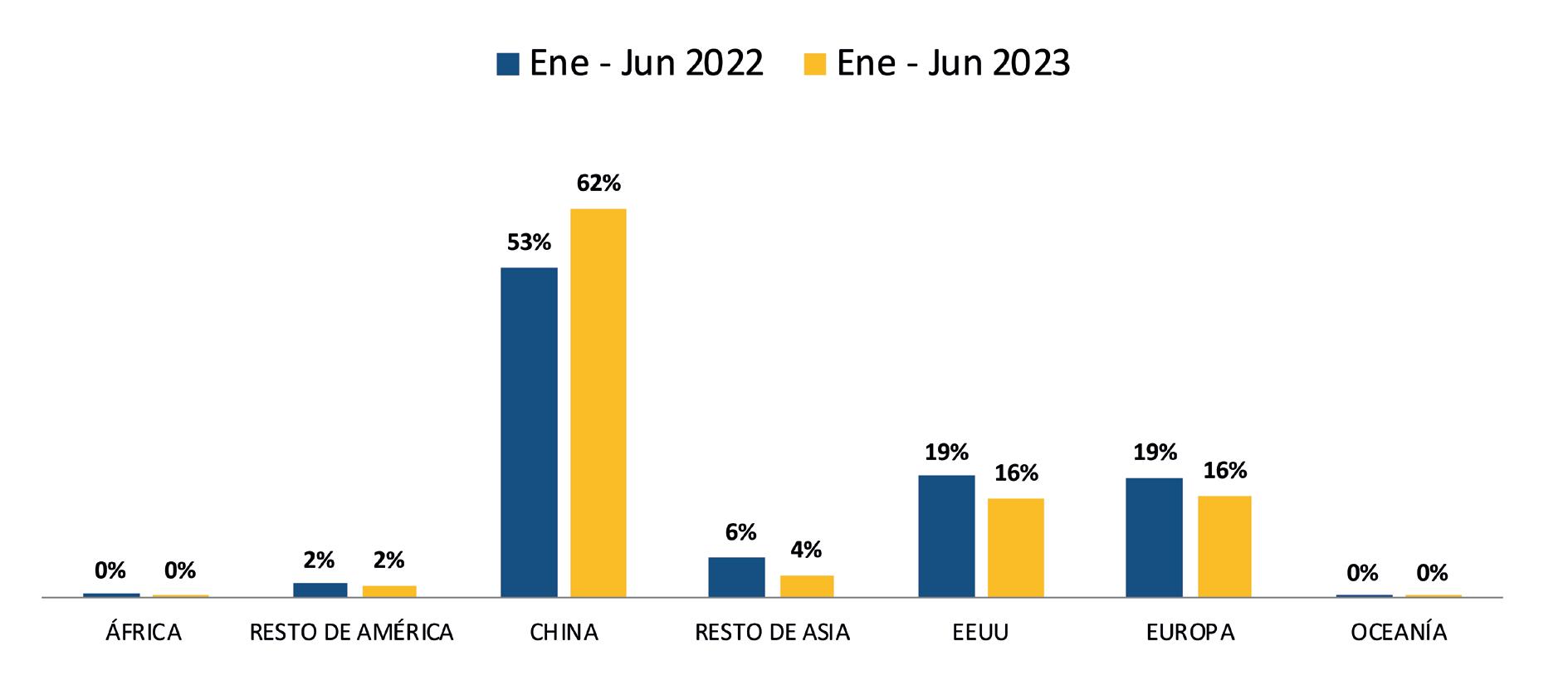
- AGOSTO 2023 67 ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: EXPORTACIONES POR PARTIDA ARANCELARIA
Enero - Junio 2023
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO MENSUAL DE EXPORTACIÓN (Libras)


Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- AGOSTO 2023 ESTADÍSTICAS
COMERCIO EXTERIOR
TILAPIA: EVOLUCIÓN DE EXPORTACIONES MENSUALES A EE. UU.
Fuente: National Oceanic and Atmospheric Administration - NOAA
Elaborado por: Cámara Nacional de Acuacultura
TILAPIA: EVOLUCIÓN DEL PRECIO PROMEDIO MENSUAL DE EXPORTACIÓN A EE. UU. (Libras)
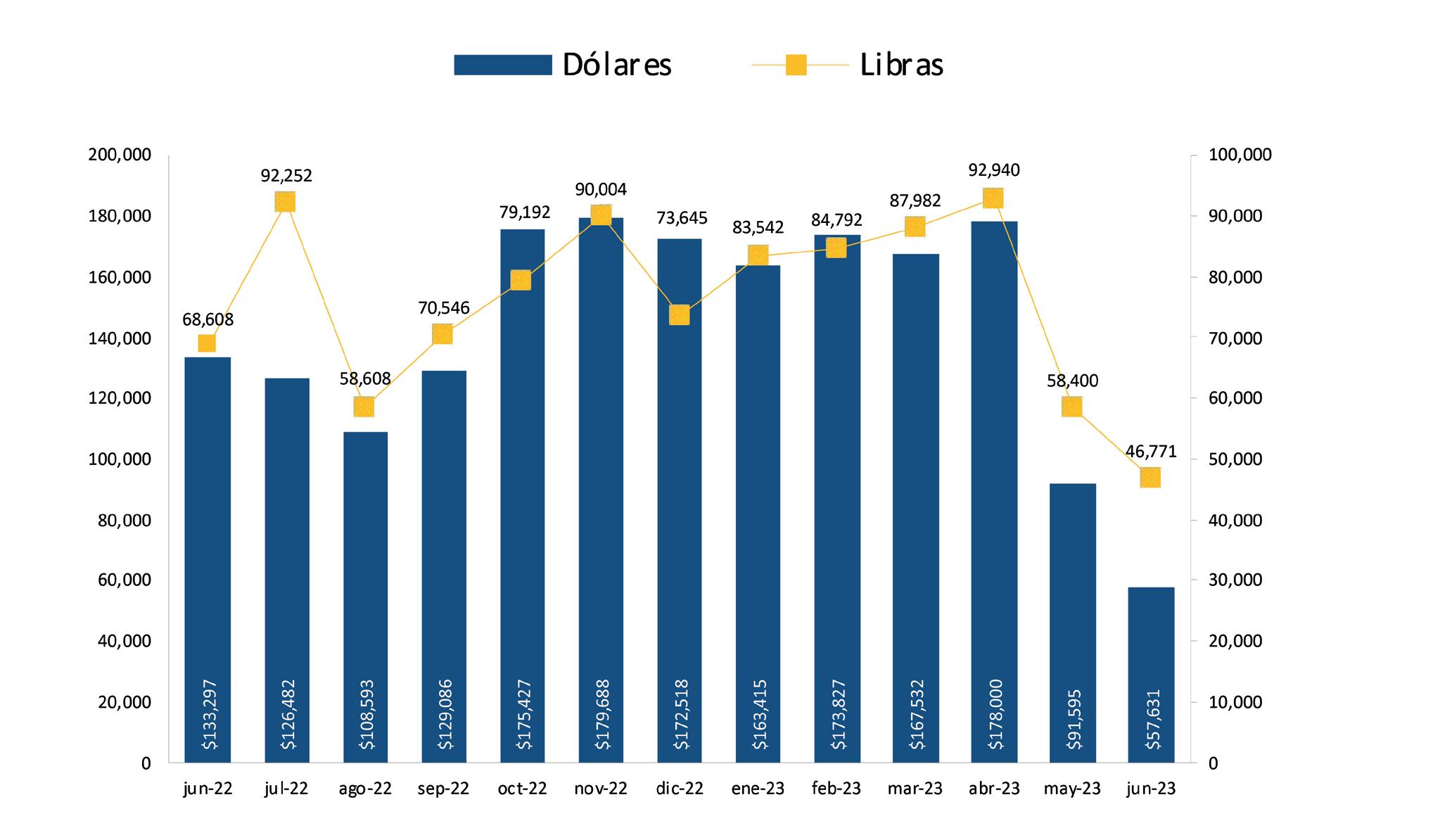
Fuente: National Oceanic and Atmospheric Administration - NOAA
Elaborado por: Cámara Nacional de Acuacultura
- AGOSTO 2023 69 ESTADÍSTICAS

Importación de camarón de Estados Unidos
Autores:
Jim Kenny jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com

 Urner Barry
Urner Barry
Importaciones de todos los tipos, por tipo
Las importaciones de junio se publicaron y revelaron una disminución interanual del 19.6%, una caída mucho más pronunciada de lo que habían anticipado la mayoría de los observadores del mercado. Las importaciones han caído durante 12 meses consecutivos, ya que los importadores buscaron reducir y mantener niveles de inventario más bajos dado el fuerte aumento en los costos de mantenimiento. Las importaciones de junio totalizaron 136.3 millones de libras frente a las 169.4 millones del año anterior. El total de seis meses es de 794.6 millones de libras en comparación con los 968.3 millones de enero a junio de 2022, una disminución del 18%. Sin embargo, el flujo de importación se mantiene un 3% por encima del promedio de los cinco años anteriores, y un 20% por encima del promedio de 2018-2019 antes de la pandemia.

Nuestros cinco principales países proveedores enviaron menos camarón en el mes de junio: India (26.2%), Ecuador (-13.1%), Indonesia (-3.7%), Vietnam (-22.9%) y Tailandia (-31.0%).
En términos de forma de producto, todos los tipos fueron más bajos en su comparación YOY (entre un período de tiempo y un año antes): camarón con cáscara (-25.5%), pelado (-9.8%), cocido (-30.9%) y empanizado (-22.5%), todos disminuyeron.
Ciclos de importación mensual por país (todos los tipos)
India: El país envió 51.3 millones de libras de camarón en el mes de junio, reduciendo envíos en un 26.2% o 18.2 millones de libras. India envió menos camarón con cáscara (-52.9%), pelado (15.0%) y cocido (-36.5%). Seis meses después, India sigue siendo el proveedor número uno de camarón a los EE.UU., al suministrar el 36% del total general, pero el país ha reducido los envíos en un 15.2% o 51 millones de libras. Esto representa 14.5 millones de libras menos de camarón con cáscara, 18.2 pelado y 13.7 cocido.
Ecuador: Durante cinco meses consecutivos, Ecuador redujo sus envíos a los EE.UU., y en junio, la disminución fue del 13.1% para todos los tipos de camarón. En este mes, Ecuador despachó menos camarón con cáscara (-15.8%), pelado (-7.7%) y empanizado (-59.5%). De enero a junio, los envíos de Ecuador a los EE.UU. disminuyeron un 3.6%, el margen más pequeño de cualquier nación proveedora importante. Como se evidencia por la pequeña disminución, EE.UU. sigue siendo un destino crítico para el camarón ecuatoriano.
Indonesia: Como uno de los tres principales proveedores de los EE.UU., Indonesia ha reducido los envíos al país durante 14 meses consecutivos. En junio, un 3.7% menos de camarón ingresó al país, pero hasta la fecha (YTD), esta disminución equivale a un fuerte 22.5% o 47.5 millones de libras. En el mes, Indonesia envió menos camarón con cáscara (-13.1%) y cocido (-25.3%), pero más camarón pelado (+16.5%) y empanizado (+9.2%). Es importante tener en cuenta que Indonesia en realidad envió un 1.2% más de camarón pelado de enero a junio, el único país proveedor importante que lo hizo.
Vietnam y Tailandia: Ambos países continúan enviando menos
REPORTE DE MERCADO
71
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
Fuente: USDOC. Urner Barry
camarón a EE.UU. Vietnam envió un 22.9% menos de camarón en el mes de junio y un 37.4% en lo que va del año (YTD). Tailandia envió un 31.0% menos en junio y un 44.6% menos de enero a junio. Vietnam ha reducido los envíos durante 13 meses consecutivos y Tailandia durante 12.
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones de camarón con cáscara, que incluyen el de fácil pelado, cayeron un 25.5% en comparación con junio de 2022.
Todos los países proveedores enviaron menos, excepto México y Perú. Ecuador (-15.8%), Indonesia (-3.1%), India (-52.9%), Tailandia (-18.7%) y Vietnam (-40.1%) registraron descensos de dos dígitos. México aumentó los envíos en un 9.7%.
Los envíos de todos los tamaños declinaron, excepto del tamaño 61-70.
Los valores de reemplazo del camarón con cáscara (importación $/lb.) disminuyeron 5% en junio, cayendo $0.17 a $3.54 por libra. En comparación con el año pasado, el valor de reemplazo es 9% más bajo.
Valor agregado, importación de camarón pelado
Las importaciones de camarón pelado, la categoría más grande por volumen, cayeron un 9.8% en junio, luego de una pequeña ganancia en mayo. India (-15.0%), Ecuador (-7.7%), Vietnam (-9.7%) y Tailandia (-18.6%) enviaron menos, mientras que Indonesia (+16.5%) aumentó sus envíos.
Los valores de reemplazo (importación $/lb.) para el camarón pelado se redujo en el mes, cayendo de $3.69 por libra a $3.67 por libra.
En comparación con el año pasado, el valor de reemplazo es un 12% más bajo.
Las importaciones de camarón cocido (agua tibia) disminuyeron un 31.6% en junio y empanizado, un 22.5%.
Importaciones de camarón cocido, empanizado y otros
Los precios de reemplazo para el camarón cocido bajaron $0.08 por libra en el mes de junio a $4.38 por libra. En el mismo punto del año pasado, los valores de reemplazo fueron casi un 14% más altos, a $5.08 por libra.
Línea de tiempo del precio del camarón; anuncios minoristas
Venta al por menor: Las oportunidades de compra aumentan y los precios bajan en comparación con el mes pasado y el año pasado. Las oportunidades de compra de camarón en los anuncios aumentaron un 29.5% en comparación con el mes pasado, y un 11.4% más en comparación con el año pasado. El precio cayó 1% desde mayo, pasando de $8.02 a $7.90 por libra, un 5% más bajo que el año pasado.
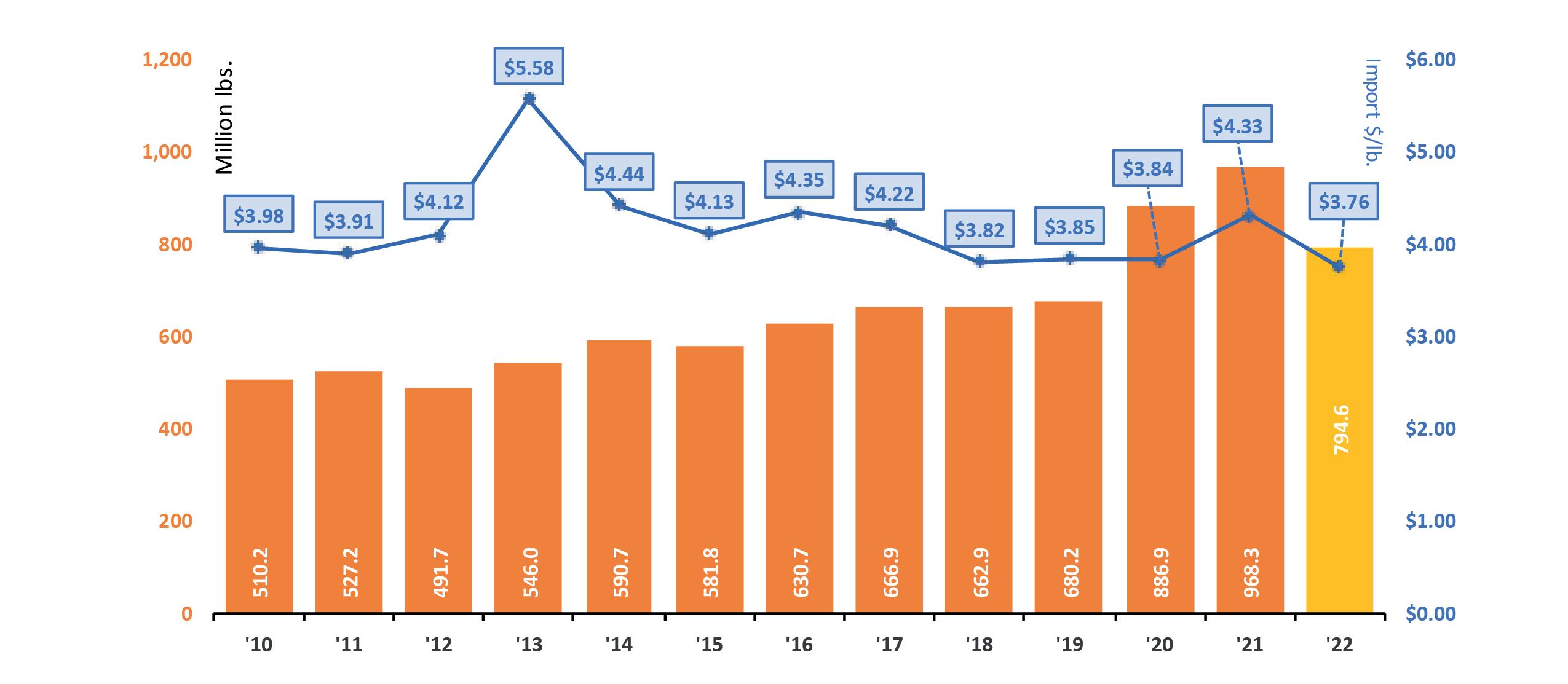
Suministro de camarón a EE.UU. y situación del Golfo
**El NMFS aún no ha publicado data de desembarques de mayo de 2023.
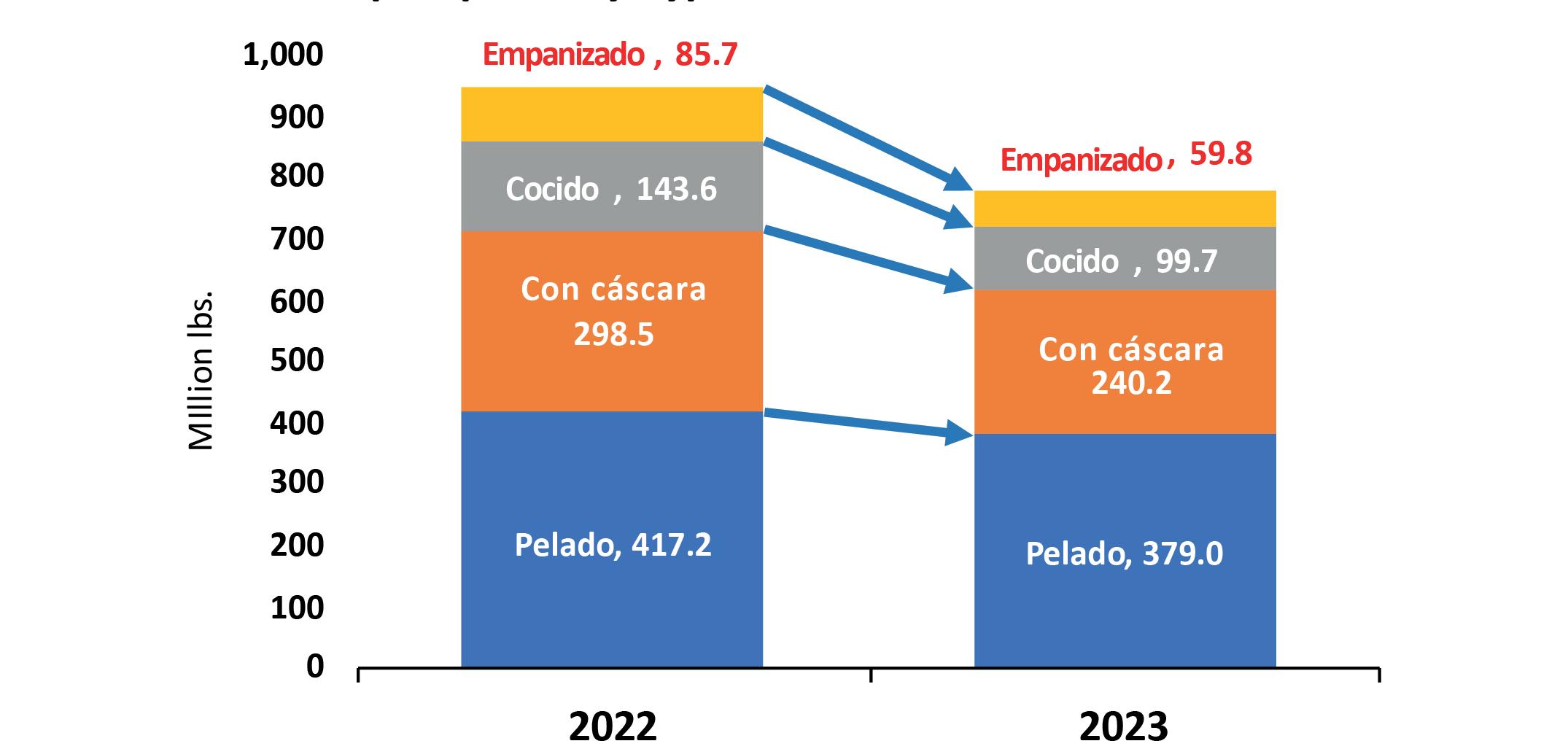
***La data de desembarques proporcionados por el NMFS ha sido inconsistente y hay brechas en la misma
Detalle de importaciones de camarón YTD por tipo
Fuente: USDOC. Urner Barry
- AGOSTO 2023 72 REPORTE DE MERCADO
que hace que las comparaciones no sean confiables.
La data de desembarques de abril de 2023 publicada por el NMFS muestra que se desembarcaron 2.553 millones de libras (total, todas las especies, sin cabeza); una fuerte reducción del 16% con respecto al mismo mes hace un año. Hasta abril, el total de 11.703 millones de libras está muy por debajo de los 16.740 millones de libras de 2022. El mercado de camarón de origen estadounidense del Golfo de México, así como de otras variedades de camarón silvestre de otras naciones proveedoras, ha estado bajo una presión constante. El interés se mantiene por debajo de las expectativas a medida que disminuyen muchos ítems comercializados por varios años.
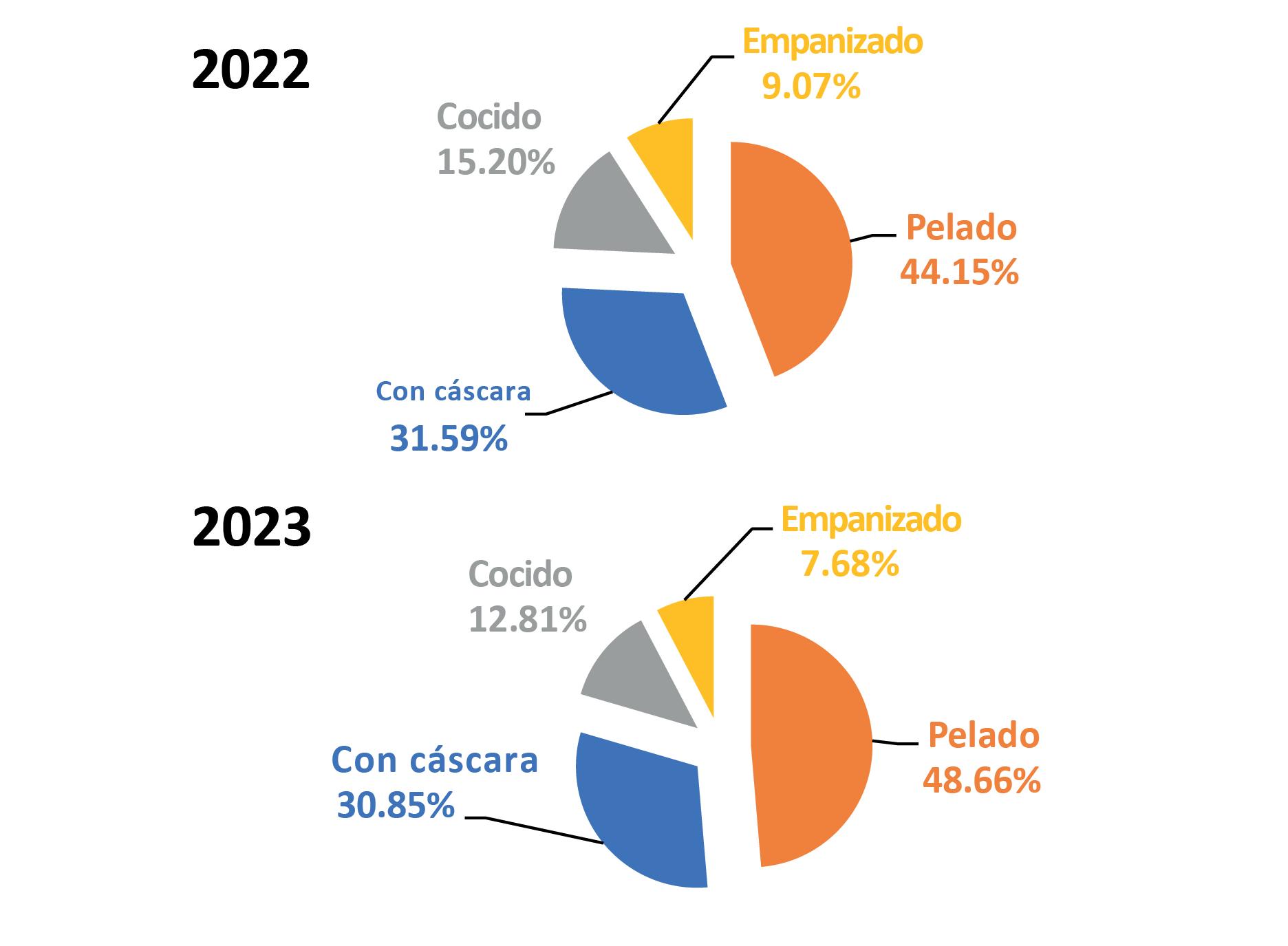
Exportación de camarón ecuatoriano
El mercado del camarón blanco ha adquirido un tono más estable en las últimas semanas, ya que el patrón prolongado de disminución de importaciones año tras año (YOY) ha ubicado los inventarios en los EE.UU. cerca de un equilibrio. Los inventarios se han reducido y la intención en toda la
cadena de distribución es mantener niveles de inventario más bajos dado el fuerte aumento en los costos de mantenimiento. Este cambio ha causado que se formen algunas brechas de suministro a corto plazo, lo que ha resultado en un aumento en el comercio de importador a importador.
Los precios de venta en el extranjero están aumentando, pero es un entorno competitivo y los descuentos ocasionales continúan apareciendo. La categoría de camarón tigre negro sigue siendo susceptible debido a la presencia de alternativas de camarón blanco de menor precio•

- AGOSTO 2023 REPORTE DE MERCADO
Importación de camarón de China
Autor: Sander Visch Kontali Shrimp sander@kontali.no www.shrimp.kontali.no
La aduana china ha publicado las estadísticas de importación de junio de 2023 para camarón. Después de un fortalecimiento de las importaciones en mayo, las importaciones disminuyeron un 13% a 87,510 toneladas. En comparación con el YOY, esto fue un 56% más alto. El volumen total de importación en el primer semestre de 2023 alcanzó las 503,624 toneladas, un aumento del 49% en comparación con el mismo período del año pasado.

Precio
Los precios de importación disminuyeron aún más en junio, en $0.04 a $5.75/kg en el mes. En comparación con el YOY, esta fue una disminución de $0.77. Con la caída tanto en el volumen como en los precios de las importaciones, el valor de las importaciones disminuyó un 13% desde el mes pasado a $503 millones. En comparación con junio del año pasado, el valor de importación fue un 38% superior. El valor de importación YTD en el primer semestre terminó en $2.85 mil millones, un 29% más que en el mismo período del año pasado.
Proveedores
Ecuador continuó siendo el mayor proveedor de camarón a China en junio, con 60,125 toneladas. Esta fue una disminución del 17% con respecto a mayo, pero aun así casi el doble que las importaciones en junio de 2022. El volumen total de importaciones hasta junio desde Ecuador alcanzó las 362,413 toneladas y, en comparación con el mismo período del año pasado, representó un aumento del 63%. Sin embargo, los precios de importación de Ecuador cayeron aún más en junio, solo $0.06, y terminaron en $5.35/kg. La caída tanto en el volumen como en los precios en junio significa una demanda algo debilitada de China, impulsada por los altos niveles de stocks reportados después de los altos volúmenes de importación de los últimos meses. En comparación con el YOY, los precios fueron más bajos, en $0.96.
Las importaciones de India se fortalecieron aún más en junio, alcanzando las 13,185 toneladas. Este fue un aumento del 21% con respecto al mes anterior y un 24% más alto en comparación con junio del año pasado. El aumento se debe en parte a que más materia prima estuvo disponible en la India después de que se llevaran a cabo las actividades de cosecha a lo largo de mayo. Los precios de importación de India disminuyeron $0.10 a $5.63/kg. En comparación con el YOY, esto fue $1.33 más bajo.
Para las importaciones de Tailandia, se observó una disminución del 4% desde el mes pasado, finalizando en 2,900 toneladas. A pesar de la caída respecto al mes anterior, Tailandia aún se convirtió en el tercer mayor proveedor de China en junio. En comparación con el mismo mes del año pasado, el volumen de importación fue un 90% superior. Al igual que en Ecuador e India, los precios de importación de Tailandia cayeron en junio. Los precios cayeron $0.57 a $10.38/kg. En comparación con el YOY, los precios fueron aún más altos, en $0.76.
Si bien las importaciones de Vietnam aumentaron ligeramente en el segundo trimestre debido a que hubo más materia prima disponible en el país, las importaciones en junio disminuyeron un 19% en comparación con mayo, alcanzando las 1,153 toneladas. Esto fue un 82% menos que el mismo mes del año pasado. A diferencia de los otros principales proveedores de China, los precios de importación de

- AGOSTO 2023
MERCADO 74
Vietnam se fortalecieron en junio, finalizando en $8.68/kg, $0.16 más que el mes pasado. Los precios de importación de Vietnam en junio terminaron en $2.18 más que durante el mismo mes en 2022. En comparación con junio del año pasado, los precios fueron más altos en $2.14/kg.
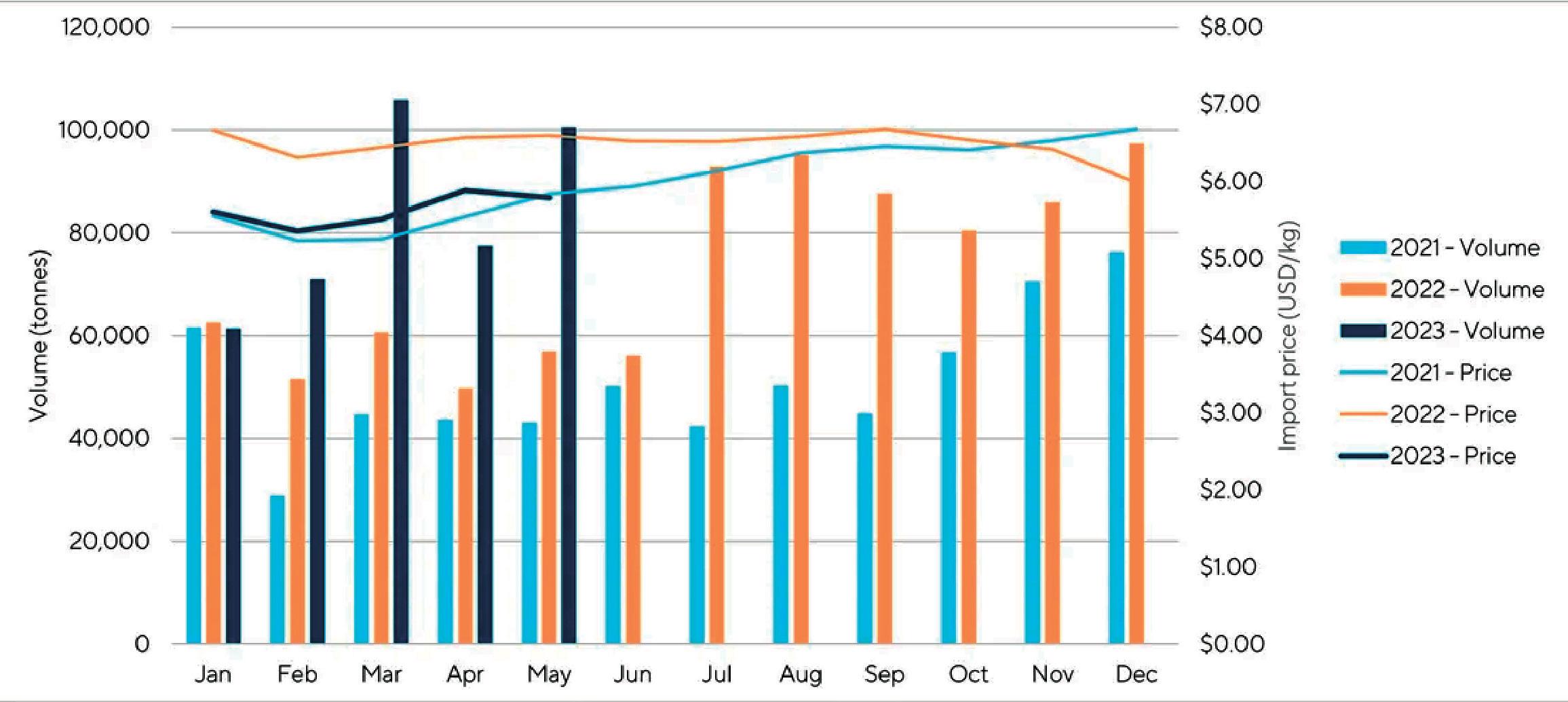
Perspectivas
Después de un fuerte repunte de la economía china durante el primer trimestre de este año, la recuperación ha sido más lenta de lo esperado durante el segundo trimestre. En la última semana, China ha presentado estadísticas económicas débiles para el último trimestre, y con una falta de confianza en el avance de su economía, la población china es más cuidadosa con sus gastos, ejerciendo más presión sobre su economía. Según fuentes de la industria, el mercado del camarón chino todavía enfrenta altos niveles de inventario. La presión sobre las instalaciones de almacenamiento en frío ha resultado en un exceso de contenedores de camarón, lo que nuevamente ha reducido un poco la disposición entre los importadores de camarón chinos para realizar nuevos pedidos. Sin embargo, a medida que nos acercamos al otoño, un período en el que las
importaciones tienden a aumentar con varias festividades importantes que se realizan en China, esto probablemente reducirá parte de la presión sobre el inventario.
Mirando el suministro local de camarón en China, las recientes condiciones climáticas en algunas partes de Guangdong, como lluvias seguidas de altas temperaturas, han afectado negativamente la salud del camarón blanco del Pacífico. Si bien esto ha resultado en una reducción de la oferta en esta parte del país, la demanda de los consumidores se ha mantenido baja, provocando que los precios en finca se mantengan bajos en Guangdong. Sin embargo, la situación ha sido un poco mejor en Fujian. Según fuentes del sector, el volumen de camarón ha ido aumentando paulatinamente; sin embargo, la demanda también ha comenzado a mostrar una tendencia al alza, lo que ha provocado que los precios en finca se hayan fortalecido en las últimas semanas.
Echando un vistazo más de cerca a los proveedores del mercado chino, la CNA en Ecuador ha expresado su preocupación por los bajos precios en finca, en combinación con el aumento de los costos de producción,
y sobre cómo esto afecta especialmente a los pequeños productores en la toma de decisiones con respecto a las actividades de siembra. Aunque la situación que reporta la CNA parece pesimista, hay un rayo de esperanza en el horizonte, con los precios en finca estabilizándose o incluso aumentando ligeramente. Mirando los patrones históricos de importación en China, en el tercer y cuarto trimestre podrían ver un aumento en la demanda de camarón ecuatoriano, a medida que los importadores chinos comienzan a prepararse para varias festividades, lo que ofrece cierto alivio para los productores ecuatorianos.
En India, los bajos precios en finca también preocupan a los productores del país. Estos precios bajos han llevado a reducir las actividades de siembra en comparación con el mismo período del año pasado. Especialmente en Andhra Pradesh, en donde no se está sembrando activamente en todo el estado. Sin embargo, algunos productores son un poco más optimistas y han reanudado las actividades de siembra•
- AGOSTO 2023 75 MERCADO
Ene Volumen Volumen Volumen Volumen (Toneladas) Precio de Importación (USD/kg) Precio Precio Precio Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Importaciones Chinas en 2021, 2022 y 2023 (volúmenes y precio promedio/kg)
Volumen
Ene
Comparación de volúmenes de importación de los principales países proveedores (2022 vs 2023)
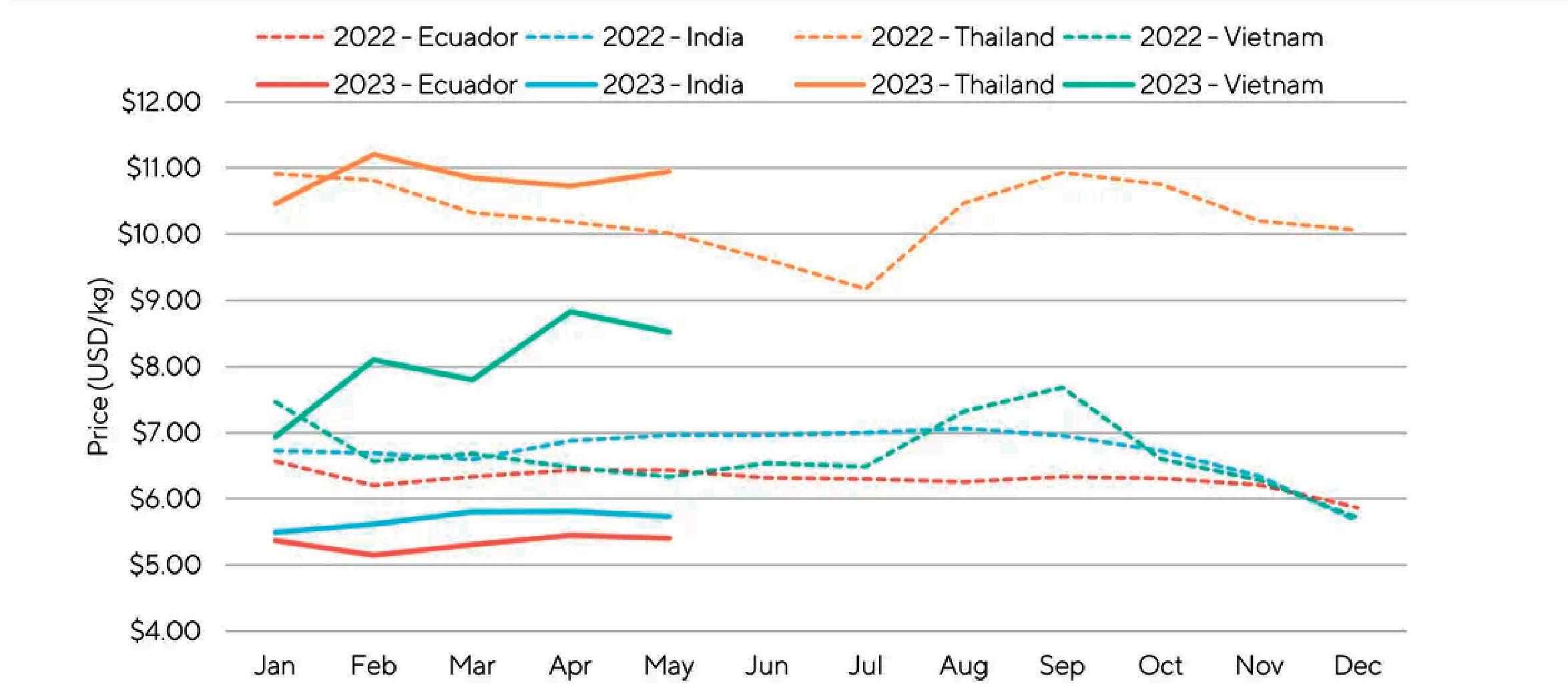
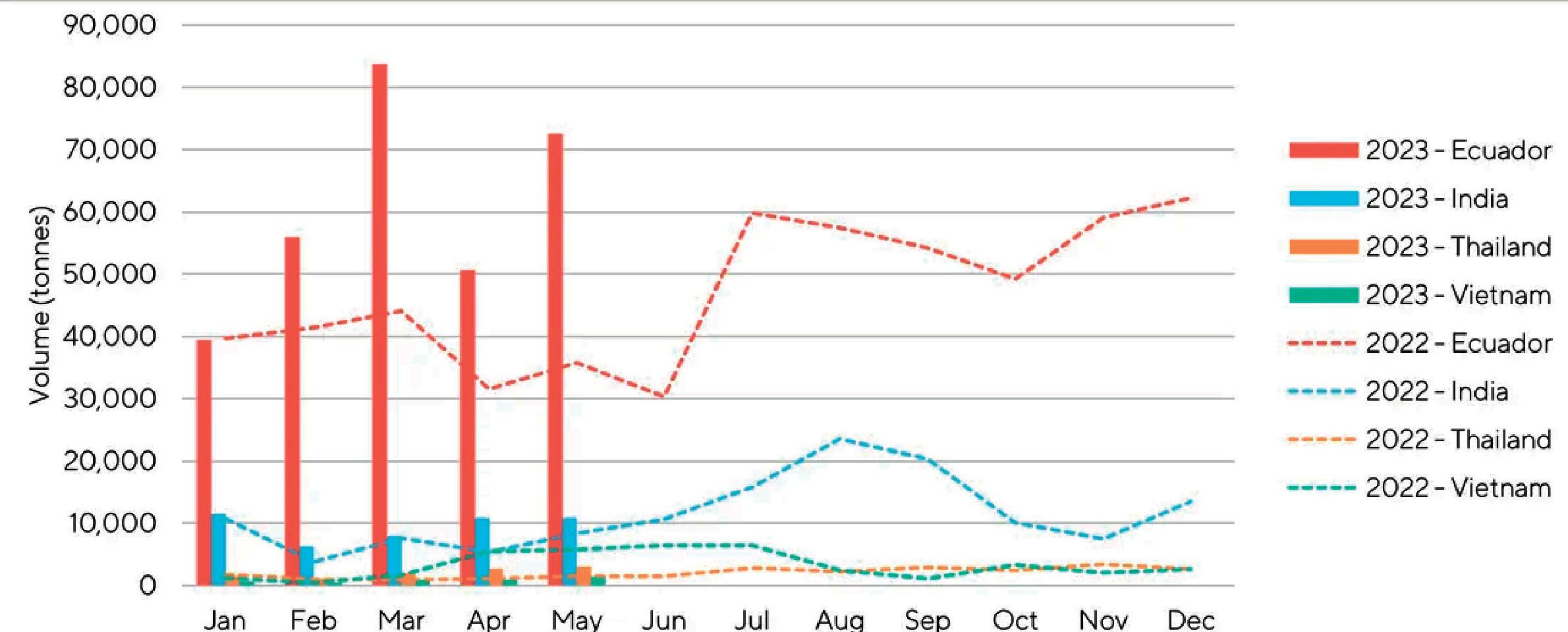
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
- AGOSTO 2023 76 MERCADO
Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
(Toneladas)
(USD/Kg)
Precios promedio de importación de los principales países proveedores (2022 vs 2023) Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Precio
Ene



Autoridades de control se reunieron con representantes del sector acuícola

El General de Distrito William Villarroel Trujillo, Comandante de la Zona 8 de la Policía Nacional de Ecuador, encabezó una reunión relevante con el Director de Seguridad de la Cámara Nacional de Agricultura (CNA), Luis Herrera; el Subsecretario de Acuacultura, Axel Vedani, y representantes del sector acuícola, con el objetivo de analizar y proponer nuevas estrategias de control del delito para garantizar la seguridad en la región.
La reunión se realizó en la sede de la CNA, donde se abordaron temas clave para analizar estrategias de control delincuencial.
Preparativos para próximas ferias internacionales
La Cámara Nacional de Acuacultura reunió a representantes de empresas exportadoras afiliadas, con el propósito de coordinar su participación en dos importantes ferias internacionales: la "Shanghai International Fisheries and Seafood Exhibition 2023" en China, y la "Seafood Expo Asia 2023" en Singapur.
Durante la actividad, se realizó el sorteo para la ubicación de cada empresa en las ferias antes mencionadas; además, se presentaron mapas de distribución y diseño de ambos eventos para conocimiento de los exportadores afiliados a la CNA.
Capacitación para verificadores sanitarios
Con el objetivo de fortalecer las habilidades y conocimientos de 40 verificadores sanitarios de la Subdirección de Calidad e Inocuidad, se llevó a cabo un curso de capacitación en “Técnicas de Auditoría”, dictado por el Dr. Leonardo Maridueña. La reunión se desarrolló en el Auditorio de la CNA, con una duración de cuatro días (4 al 7 de julio), combinando tres días presenciales y un día virtual.

La capacitación, que consistió en un curso intensivo de 16 horas con examen incluido, contó con la participación de 40 profesionales de diferentes provincias, entre ellas Manabí, El Oro y Guayas. Durante el evento, se abordaron temas relevantes relacionados con auditorías sanitarias, buscando dotar a los verificadores de las herramientas técnicas necesarias para llevar a cabo su labor de manera eficiente y precisa.

- AGOSTO 2023 80 NOTICIAS
Emisión de certificados de origen Reunión de Cuarto Adjunto
Representantes de empresas exportadoras afiliadas a la Cámara Nacional de Acuacultura participaron en una reunión virtual para conocer más detalles sobre el servicio de emisión de certificados de origen para exportadores, gracias a la reciente alianza firmada entre la CNA y la Cámara de Industrias de Guayaquil.
En la reunión se dieron a conocer los beneficios del servicio, como horario extendido de entrega: de lunes a viernes de 10h00 a 19h00 y sábados de 09h00 a 12h00; agilidad: entrega del certificado el mismo día si se hace el ingreso de solicitud al SIGCO hasta las 12h00, y entrega a domicilio: de acuerdo con el volumen mínimo de 50 certificados por semana.
Representantes del sector productivo participaron en una reunión del Cuarto Adjunto para conocer los avances en la agenda de comercio exterior, liderada por el Ministerio de Producción, Comercio Exterior, Inversiones y Pesca (MPCEIP).

Se dio a conocer la situación de los acuerdos comerciales vigentes y la Ley de Fomento Productivo, Atracción de Inversiones, Generación de Empleo y Estabilidad y Equilibrio Fiscal (Ley IDEA).
Los representantes gremiales recibieron información detallada sobre el estado actual de los acuerdos comerciales que involucran a Ecuador y la implementación de la Ley IDEA. El diálogo entre ambas partes permitió conocer el avance en materia de relaciones internacionales, y cómo esto puede impactar en el desarrollo económico y productivo del país.
- AGOSTO 2023 NOTICIAS
Medidas de prevención por Fenómeno de El Niño
La Cámara Nacional de Acuacultura lideró la reunión entre representantes del sector camaronero y funcionarios de CELEC U.N. HIDRONACIÓN, para conocer las acciones planificadas a corto, mediano y largo plazo para mitigar la llegada del fenómeno de El Niño.

En la reunión se abordó la importancia de contar con descargas planificadas del embalse Daule-Peripa. Se plantearon las inquietudes de los afiliados a la CNA y miembros del sector, para conocer de primera mano el alcance de estas medidas y el impacto que tendrían en sus operaciones y actividades.
De esta manera, los asistentes conocieron las razones detrás de las descargas y las acciones preventivas que buscan reducir los posibles efectos de El Niño en zonas identificadas por su alto riesgo de inundación.


SSP participó en programa
Hatch Blue
El quinto programa de aceleración de acuicultura de Hatch Blue se realizó en Hawaii, y contó con la participación de Pamela Nath, Directora de Sustainable Shrimp Partnership (SSP), quien compartió detalles sobre los logros alcanzados por SSP y estableció conexiones con nuevas empresas dedicadas al desarrollo de un futuro sostenible en la industria acuícola.

Esta colaboración refuerza el enfoque de sostenibilidad a nivel global y posiciona a Ecuador como un productor responsable y líder en el sector, atrayendo oportunidades comerciales e inversiones al país.
SSP se reunió con ONG europea
Pamela Nath, Directora de Sustainable Shrimp Partnership (SSP), y Leonardo Maridueña, Coordinador de Proyectos Técnicos de SSP, se reunieron con la ONG francesa Earthworm Foundation para abordar la sostenibilidad de las piscinas camaroneras ecuatorianas y mostrar su capacidad de carga durante el proceso de producción.
Al presentar la capacidad de carga de las piscinas, SSP y Earthworm Foundation pueden demostrar el compromiso de la industria con prácticas responsables y sostenibles. Esto ayuda a mejorar la reputación del sector y cultivar la confianza de los consumidores.
Houdek-Prairie
Aquatech, nuevo socio SSP
La empresa estadounidense Houdek-Prairie AquaTech se ha sumado como miembro asociado a Sustainable Shrimp Partnership (SSP), consolidando el compromiso conjunto de promover la producción acuícola sostenible y reducir el impacto ecológico en el sector. Esta inclusión refuerza el enfoque sostenible de la industria camaronera ecuatoriana, impulsando prácticas responsables en todas las etapas de producción y abriendo oportunidades para desarrollar proyectos de mitigación ambiental.
Tanto SSP como Houdek-Prairie AquaTech son un ejemplo inspirador para que otros productores y actores del sector adopten enfoques más responsables y respetuosos con el medio ambiente.
- AGOSTO 2023 82 NOTICIAS


