
ANNO L numero 4







2024
Didattica & Cultura Dal Lucherino “agata” al Verdone agata…
Canarini di Colore Discutendo di agata pastello ad Arezzo
Ondulati ed altri Psittaciformi Becchi potenti ma sensibili
Estrildidi Fringillidi Ibridi La Tortora dal collare domestica
Rivista mensile di Ornitologia Scientifica - Tecnica - Pratica Organo Ufficiale della F.O.I.-Onlus









sommario


F.O.I.
Lucherino “agata” al Verdone agata…
di agata pastello


e
fattore opale - 2ª parte


AMMINISTRAZIONE E PUBBLICITÀ: Segreteria F.O.I.-Onlus Via Caorsana, 94 - Località Le Mose 29122 Piacenza Tel. 0523.593403 - Fax 0523.571613
Web: www.foi.it - E-mail: redazione@foi.it
Direttore Responsabile: Antonio Sposito
Caporedattore: Gennaro Iannuccilli
Collaboratori di Redazione: Giovanni Canali, Maurizio Manzoni, Francesco Rossini
Commissione Salute, Benessere animale e Ricerca Scientifica: Elena Circella, Giuseppe Marruchella, Gianluca Todisco
Coadiutore Editoriale: Lorenza Cattalani

Associato all’Unione Stampa Periodica Italiana ISSN 0391-254X (International Standard Serial Number) Autorizzazione del Tribunale di Bologna n. 4396 del 12-3-1975
Stampa: TEP s.r.l. - Strada di Cortemaggiore, 50 29122 Piacenza - Tel. 0523.504918
Inoltro postale in Italia: Effezeta srl Via Francesco Nicoli 10/G - 29122 Piacenza
ABBONAMENTI ANNUI:
Italia € 50,00 - Estero-Europa € 70,00
Estero-ExtraEuropa € 90,00
Un numero € 5,00 - Arretrato € 6,50
C.C.P. 53684957
Le quote abbonamento vanno versate, mediante vaglia o assegno, alla Segreteria. Le opinioni espresse dagli Autori non impegnano la Rivista e la sua Direzione.
La Redazione si riserva il diritto di non pubblicare o emendare gli articoli proposti. I collaboratori assumono piena responsabilità delle affermazioni e delle immagini contenute nei loro scritti.
Vietata la riproduzione, anche parziale, se non espressamente autorizzata. © F.O.I.
In copertina:
Tortora dal collare domestica mutazione onice Streptopelia risoria
Foto e allevamento: VINCENZO RALLO
ANNO L NUMERO 4 2024
Assemblea
2024 Gennaro Iannuccilli 3 Dal
Riccardo Rigato 7 Discutendo
ad
Giovanni Canali 13 Becchi potenti ma allo stesso tempo sensibili Rafael Zamora Padrón 17 La Tortora dal collare domestica - 1ª parte Francesco Faggiano, Vincenzo Rallo e collaboratori 21 Schemocromi:
Mimmo Alfonzetti 31 Canarino x Ciuffolotto Piercarlo Rossi 35 Pasto unico sì o no? Gaetano Campisi 42 Photo Show Le foto scattate dagli allevatori 44 L’Associazione
compiuto 50 anni Raffaele Cecere 45 Lo Storno rosa (Pastor roseus) Giovanni Musumeci 49 Centocchio (Stellaria media) Pierluigi Mengacci 51 La gabbia, tra ragionamento
giudizio morale Pasquale Leone 55 Spazio Club Lizard Canary Club Italiano 57 OrniFlash News al volo dal web e non solo 58 Il mondo degli uccelli “Allevare è proteggere” Nicola Grimaldi 61 60 anni di ornicoltura e passione ornitologica Mirko Masiero 62 Pensieri in libertà Micaela ed Elisa Mengacci 64
Arezzo
Ornitologica Vesuviana ha
Ondulati
Estrildidi Fringillidi Ibridi 7 13 17 35 Italia Ornitologica è la rivista ufficiale della F.O.I. - Federazione Ornicoltori Italiani, pubblicata in 10 (dieci) numeri annuali a cadenza mensile, 2 (due) dei quali in versione bimestrale nel periodo estivo (Giugno/Luglio e Agosto/Settembre). Il numero 4 - 2024 è stato licenziato per la stampa il 23/4/2024
Didattica & Cultura Canarini di Colore
ed altri Psittaciformi









Editoriale
Assemblea F.O.I. 2024
di G ENNARO IANNUCCILLI, foto F.O.I.


Come di consueto, nel mese di Aprile si svolge l’Assemblea Generale delle Associazioni FOI, momento fondamentale per la vita federale. Anche quest’anno abbiamo avuto modo di vivere questo evento nel weekend del 13-14/04 presso l’Auditorium Gatti di Piacenza Fiere. L’Assemblea è stata molto partecipata, con la presenza di 148 Associazioni rappresentate dai rispettivi Presidenti o Delegati, anche perché quest’anno ricorreva l’elezione del nuovo Consiglio Direttivo Federale, che resterà in carica per il prossimo quadriennio con la seguente composizione: Antonio Sposito (Presidente), Diego Crovace (Vice Presidente), Giovanni Nunziata (Segretario), Gennaro Iannuccilli, Francesco Badalamenti, Maurizio Adami e Giovanni Nisticò (Consiglieri).





Editoriale


Nella giornata di Sabato si è inoltre tenuta l’Assemblea Nazionale dei Giudici FOI che ha provveduto a rieleggere il Presidente dell’Ordine dei Giudici – Andrea Benagiano – nonché i Presidenti dei rispettivi Collegi di Specializzazione: Michele Laricchia (Colore), Alessandro Fuson (EFI), Claudio Berno (FPL), Antonio Altobelli (FPA), Giorgio Roccaro (O&aP), Roberto Gabriele (Canto).




Oltre alla trattazione dei temi assembleari all’ordine del giorno, tutti affrontati in maniera esaustiva, ci sono stati momenti emozionanti e finanche commoventi nel celebrare Associazioni e rappresentanti del nostro mondo ornitologico che si sono contraddistinti per traguardi raggiunti e collaborazioni prestate volontariamente al servizio della collettività.


Editoriale


Dall’Assemblea 2024 è emersa chiaramente la percezione di voler affrontare insieme le difficoltà a cui siamo esposti in questo periodo, evitando sterili polemiche ma concentrandosi sulla ricerca di soluzioni che possano – nonostante la recessione in atto in questi anni – favorire il mantenimento e il miglioramento dei servizi erogati per gli allevatori e degli elevati standard raggiunti in ambito federale.
Il nuovo Consiglio Direttivo auspica di poter cooperare con tutte le Associazioni e i loro rappresentanti affinché tali obiettivi possano essere centrati fin dalla prossima stagione, grazie all’entusiasmo e alla passione che ha sempre contraddistinto gli ornicoltori amatoriali e sportivi, confermando piena disponibilità verso un confronto costruttivo nell’ottica di rendere la FOI sempre più dinamica e al passo con i tempi.
Lasciamo alle immagini allegate il compito di trasmettere l’essenza di questo momento istituzionale, al quale abbiamo avuto l’onere e l’onore –ma soprattutto il piacere – di partecipare.
















Dal Lucherino “agata” al Verdone agata…
Passando per il Diamante zebrato
testo
di RICCARDO
RIGATO, foto R. RIGATO, WWW ORNITALIA COM, F.O.I. e E. DEL POZZO
Il vasto panorama delle cromie del mondo alato è il risultato di una selezione evolutiva operata dalla natura attraverso meccanismi di rimaneggiamento genetico frutto anche di condizionamenti ambientali che influenzano la sopravvivenza di specifici stipiti selettivi che meglio si adattano ai vari ecosistemi. Chi influenza dunque la sopravvivenza di una nuova variante cromatica o morfologica, risultato di mutazione, è il vantaggio che ne trae la specie in oggetto in termini




di adattamento. Emblematico l’esempio dei fringuelli delle Galapagos studiati da Darwin nei quali la struttura del becco si è evoluta in funzione della disponibilità di un determinato cibo. Nei nostri aviari, dove l’apporto trofico è garantito dall’allevatore e dove le condizioni ambientali sono controllate, si assiste ad un processo evolutivo delle popolazioni ornitiche che è il risultato di due grandi strade percorribili. Una prima possibilità, che finalizza

ad ottenere nuove cromie o varianti morfologiche, è quella della selezione operando una corretta scelta dei riproduttori con caratteristiche volte a raggiungere un risultato ipotizzato sommando caratteri quantitativi; un esempio può essere quello della modificazione della taglia. Selezionare, per esempio, un gloster presuppone un’attenta analisi dei riproduttori prestando molta attenzione ai minimi particolari strutturali, affinché la progenie
NUMERO 4 - 2024 7 DIDATTICA & CULTURA
Lucherino presunto Agata alla nascita, notare occhio depigmentato, foto per gentile concessione di Ornitalia
Lucherino presunto agata allo svezzamento, foto per gentile concessione di Ornitalia



si avvicini il più possibile al modello teorizzato. Il risultato che l’allevatore deve ottenere è frutto di una convenzione concordata dagli organi competenti per ciascuna razza, frutto di modifiche ed aggiustaggi influenzati sia dal buon gusto che dalle mode. La meta in questione è definita standard. Nelle razze di forma e posizione lo standard viene redatto da un pool di esperti che individuano un modello teorico a cui aspirare attraverso la selezione. La capacità dell’allevatore consiste nel raggiungere un modello strutturale il più vicino possibile all’ipotizzato riferimento standardizzato.
Tralasciando in questa sede attraverso quali meccanismi selettivi si possa raggiungere l’agognato risultato, è fuori dubbio che sia di vitale importanza la sensibilità e la competenza con cui opera l’allevatore affinché, nell’arco delle varie generazioni, migliori la struttura dei propri soggetti amalgamando sapientemente i caratteri quantitativi che gli permettano di rag-
giungere il risultato sperato. Ben altra cosa è la corretta selezione di soggetti interessati da mutazioni della livrea, siano esse del colore o della distribuzione dei pigmenti (mutazioni del di-
Alla comparsa di una modificazione dell’assetto cromatico, in qualsiasi specie, in prima istanza è giusto vagliare l’ipotesi che tale mutazione sia già presente in altri ceppi di allevamento appartenenti ad altre specie
segno). Alla comparsa di una modificazione dell’assetto cromatico, in qualsiasi specie, in prima istanza è giusto vagliare l’ipotesi che tale mutazione sia già presente in altri ceppi di allevamento appartenenti ad altre specie. La tonalità e la saturazione dei pigmenti non devono trarre in inganno chi opera il confronto in quanto ogni specie possiede un suo corredo melanico e lipocromico tipico e, come già precisato in una precedente nota, i geni mutanti, agendo su substrati differenti, possono determinare risultati fenotipici differenti.
Bisogna essere molto cauti nell’attribuire l’esatta nomenclatura ad un nuovo fenotipo. Comparando per esempio il canarino bruno pastello all’organetto bruno pastello, chi potrebbe darne la corretta collocazione genetica con il solo confronto cromatico? Il diamante zebrato agata (identificato correttamente solo attraverso prove di complementazione per ibridazione) è stato erroneamente, in
8 NUMERO 4 - 2024
Agata frutto di selezione, foto: R.Rigato
Agata non selezionato con disegno largo e ricco di feomelanina, foto: R.Rigato
Altra immagine di agata proveniente da ceppo di nero bruni, foto: R.Rigato
prima istanza, definito bruno in quanto i geni responsabili della mutazione agata, agendo su un substrato melanico ancestrale in possesso di un’importante quota di melanina bruna, produce un risultato ottico di tonalità bruno ridotto piuttosto che grigia come invece avviene in gran parte delle specie interessate dalla suddetta mutazione agata. Quali sono dunque i punti cardine per dare una giusta collocazione genetica alle nuove cromie? Quando è possibile, come già accennato, attraverso l’ibridazione con specie affini già codificate si può dare risposta ad alcuni quesiti, ma tale metodica possiede anch’essa dei limiti. Produrre progenie mutata accoppiando fenotipi simili appartenenti a specie diverse non certifica che trattasi della stessa mutazione. Può essere infatti motivo di un’errata interpretazione dei risultati quando vengono accoppiati soggetti appartenenti a mutazioni tra loro alleliche. Prendiamo
Produrre progenie mutata accoppiando fenotipi simili appartenenti a specie diverse non certifica che trattasi della stessa mutazione
in esame il caso di un ibrido mutato maschio frutto dell’accoppiamento di diamante modesto agata x femmina di diamante mandarino mascherato occhio rosso. Il soggetto in questione possiede un fenotipo mutato non perché i parentali sono in possesso della stessa mutazione ma perché le mutazioni in oggetto sono tra loro alleliche. Sappiamo infatti che nel diamante mandarino, oltre all’agata (definito in


questa specie dorso chiaro), sono presenti altre mutazioni alleliche le quali, accoppiate tra loro, producono prole mutata il cui fenotipo risulterà fortemente influenzato dal gene recessivo che, pur non esprimendosi in toto, riesce a modificare l’espressione del gene dominante. Un esempio pratico è il maschio diamante mandarino dorso chiaro portatore di ino dove il frutto del fenotipo dorso chiaro (agata) è il risultato della presenza, appunto, della mutazione agata su di un cromosoma sessuale x e della ino sull’altro cromosoma sessuale x che determina una inibizione dell’espressione melanica conferendo al soggetto una livrea più chiara rispetto al corrispettivo dorso chiaro (agata) omozigote. In merito a tale fenomeno Giovanni Canali ipotizza che tali geni si esprimano con effetto di co-dominanza. Per i cultori di fringillidi forse può essere più palese come un cardellino o un verdone agata, portatori di lutino, appaiano


NUMERO 4 - 2024 9
Bruno pastello intenso giallo avorio, foto: E. del Pozzo
Organetto pastello bruno, foto: F.O.I,



meno ossidati rispetto agli agata omozigoti.
Nella stesura di uno standard vanno dunque fatte tutta una serie di considerazioni affinché venga identificata con certezza la mutazione e venga rispettata la corretta espressione dei pigmenti non affidandosi al gusto personale, ma rispettando le caratteristiche espressive della mutazione in oggetto. Nel verdone, per esempio, si è assistito alla tendenza, in sede espositiva, a premiare verdoni lutini privi di carica eumelanica bruna preferendo soggetti apparentemente privi di soffusioni brune. Quando nel corso degli anni ci si è resi conto che la mutazione lutino non è in grado di azzerare le melanine, si è cominciato ad apprezzare la presenza del suddetto residuo melanico, giusta espressione di tale genotipo.
Rimanendo nella selezione del verdone, a mio avviso, anche nella mutazione agata sono stati commessi degli errori di interpretazione della corretta espressione. Nell’ultimo decennio sono stati preferiti verdoni agata particolarmente ossidati, a tal punto che anche gli annessi cutanei (becco, zampe e ed unghie) appaiono mela-
nizzati, il lipocromo risulta presente in minor quantità al fine di rendere più evidente il contrasto tra fondo e disegno. Tutto ciò in contrasto con quanto è contemplato nello standard sulla corretta manifestazione della mutazione agata. Essa infatti riduce l’eumelanina nera, inibisce la feomelanina e la eumelanina bruna, non permette il deposito melanico su becco e zampe, inoltre l’iride, fin dalla nascita, risulta palesemente pigmentata. Pertanto, in una mutazione che interessa il solo corredo melanico, il lipocromo deve potersi esprimere in tutta la sua totalità e non essere inibito attraverso una forzata pressione selettiva. Esiste dun-
Rimanendo nella selezione del verdone, a mio avviso, anche nella mutazione agata sono stati commessi degli errori di interpretazione della corretta espressione
que un’etica da rispettare sia nella stesura degli standard che nel giudizio dei soggetti interessati da mutazione; va rispettata l’originale espressione del fenotipo, facendo fedele riferimento al genotipo.
Ho più volte sottolineato quanto sia controproducente selezionare delle varianti cromatiche dove, oltre all’azione dei geni specifici della mutazione, sfruttiamo l’azione di geni additivi modificatori che migliorano, a nostro avviso, il risultato estetico ma che in realtà inquinano l’identità della mutazione. Una specie che rischia di essere preda di una non corretta interpretazione di un nuovo fenotipo è il lucherino. Come è noto in tale spinus è stato commesso l’errore di definire agata una variante fenotipica che invece, grazie a test ibridologici, si è poi rivelata essere pastello recessivo sesso-legato. In tempi recenti (2015), nell’allevamento del compianto Maestro Paolo Gregorutti è giunta dalla Croazia una femmina di lucherino dalla livrea in possesso di una carica melanica che ha fatto ipotizzare che ci trovassimo finalmente al cospetto della vera mutazione agata nel lucherino. Dopo aver ottenuto da tale femmina i primi maschi portatori, Paolo ha prodotto delle femmine mutate che stranamente, nei primi giorni di vita, presentavano una evidente depigmentazione delle melanine dell’occhio. Con lo sviluppo l’occhio progressivamente si pigmenta allo stesso modo di quanto avviene, per esempio, nel diamante mandarino mascherato ad occhio nero, nel cardellino aminet e nel verdone ambra e mascherato. In virtù di tale osservazione non è corretto definire agata tale cromia, né tantomeno è possibile definire uno standard. Ho avuto il piacere di osservare degli ibridi maschi ottenuti con tale mutazione del lucherino con canarina agata. In effetti le mutazioni non complementano e producono tutti i soggetti a fenotipo mutato perché si tratta di parentali appartenenti a mutazioni alleliche. Al fine di verificare tale ipotesi basta accoppiare un lucherino maschio in possesso della discussa mutazione con una canarina agata e verificare, fin dalla nascita, il colore degli occhi degli ibridi. Se le
10 NUMERO 4 - 2024
Diamante modesto bruno, foto: E. del Pozzo
femmine ibride manifestano gli occhi rossi che si scuriscono con il passare dei giorni, mentre i maschi, pur essendo mutati, possiedono l’iride pigmentata sin dalla nascita, sarà dimostrata l’allelicità delle mutazioni dei genitori. Invito inoltre i cultori di lucherini a riflettere su come selezionare il disegno di tale recente fenotipo.
Nel canarino, come sappiamo, si è giunti all’agata odierno attraverso una selezione che sfrutta in gran misura geni additivi. Attraverso un certosino lavoro è stata modificata l’espressione del disegno agata originario rendendolo più corto e fine. Tale risultato è frutto di un modello teorizzato cui si è giunti attraverso un percorso ispirato dal gusto estetico soggettivo di chi orienta la selezione e non rispettando invece l’identità della mutazione. La foto a corredo mette a confronto il disegno di un agata proveniente da un ceppo di nero bruni con un agata
Nel canarino, come sappiamo, si è giunti all’agata odierno attraverso una selezione che sfrutta in gran misura geni additivi
frutto di selezione. Questa considerazione non deve far pensare che alla base di tale precisazione ci sia una esasperata motivazione etica. Discostarsi dall’originaria espressione di una mutazione modellando il fenotipo con i geni additivi ci può far cadere nell’errore di produrre soggetti artefatti a volte simili a fenotipi frutto dell’espressione di un solo gene mutato.
Nel canarino di colore tale grave realtà è osservabile nel nero odierno dove non si può sapere se la mancata espressione della feomelanina dipenda da una spinta selettiva, dall’azione di un solo gene (come avviene per esempio nella mutazione petto bianco del diamante di Gould) o dalla sinergia di entrambi. Una facile soluzione sarebbe quella di riproporre in sede espositiva un canarino a corredo originario completo tipico della specie in natura utilizzando come corredo di base il nero bruno. Saremmo allora facilitati nella giusta interpretazione delle novità genetiche. Seguendo l’esempio della canaricoltura di colore si rischia di pensare che sia corretto eliminare la categoria ancestrale nell’organetto solo perché esistono gli organetti scuri o i becchi di piombo ventre scuro o il cardellino nero o il diamante mandarino charcoal. Pensiamoci bene!




NUMERO 4 - 2024 11









Discutendo di agata pastello ad Arezzo
di GIOVANNI CANALI, foto E. DEL POZZO e F.O.I.
Non capita spesso che io e Gennaro Iannuccilli ci si veda di persona, data la distanza delle nostre residenze, quindi bisogna ringraziare molto Meucci per la sua invenzione. Abbiamo quasi un filo diretto per le esigenze della rivista. Fortunatamente è capitato d’incontrarci ad Arezzo, alla cui ottima mostra abbiamo esposto entrambi. Nell’occasione abbiamo potuto discutere ampiamente su tanti soggetti, ovviamente anche i nostri, che fra l’altro hanno ben figurato. Non parlerò di tutto, ci vorrebbero troppe pagine, ma mi soffermerò su di un aspetto. È stata esposta una femmina agata pastello intenso giallo (allevamento Iannuccilli), con interessanti caratteristiche, fra l’altro ben valutata. (Tale canarina, premiata come “Campione Razza melaninici”, è nata da un accoppiamento tra maschio agata pastello intenso giallo e femmina agata brinato giallo, quest’ultima proveniente da selezione del proprio allevamento, ottenuta incrociando maschio agata/isabella e femmina isabella – Nota di Gennaro Iannuccilli).
L’effetto ali grigie, in bruni ed isabella, lo ricerchiamo per ridurre il disegno, “appastellare” come talora si dice, con un’espressione suggestiva



NUMERO 4 - 2024 13 CANARINIDI COLORE
Agata pastello intenso giallo, foto: E. del Pozzo





Il soggetto in questione, buono o ottimo in tutte le voci, nel tipo ha un disegno dorsale eccellente ed anche una diluizione del bruno fortissima, rara in una femmina. Disegno del giusto grigio ferro, sottilissimo e spezzato, ben diverso da certi disegni pesanti che talora capita di vedere. Inoltre ben rappresentato in tutte le parti del corpo, testa compresa, ma ulteriormente ridotto sulle ali. Questo è stato l’aspetto che ci ha suggerito diverse considerazioni. Sappiamo che l’effetto ali grigie, vale a dire geni modificatori, agisce o meglio può agire anche sugli altri tipi pastello e non solo sul nero, ove è classificato a parte. L’effetto ali grigie, in bruni ed isabella, lo ricerchiamo per ridurre il disegno, “appastellare” come talora si dice, con un’espressione suggestiva ma forse non del tutto esatta. Mentre negli agata pastello non vo-
Il soggetto in questione, buono o ottimo in tutte le voci, nel tipo ha un disegno dorsale eccellente ed anche una diluizione del bruno fortissima
gliamo tale effetto, per avere il disegno non dissimile da quello degli agata classici, ma non nero bensì grigio ferro, in altri termini solo il carattere prodotto dal gene maggiore che mutando produce il pastello. Una situazione particolare, quasi anomala, che rende diverse le selezioni per il pastello in agata ed isabella. Non a caso l’accoppiamento fra agata pastello ed isabella pastello non è consigliato poiché adatto per il tipo base, ma non per il tipo aggiunto pastello. Una situazione strana ed insolita, alla quale tuttavia bisogna adattarsi. In effetti non si potrebbero avere ottimi agata pastello ed ottimi isabella pastello con la stessa linea selettiva. Magari ottimi come tipo base, ma non come tipo aggiunto pastello. La ragione è che, se presenti i
14 NUMERO 4 - 2024
Isabella pastello mosaico giallo femmina, foto: E. del Pozzo
Bruno pastello bianco recessivo
geni modificatori, potrebbero essere ottimi gli isabella pastello, ma gli agata pastello avrebbero almeno le ali schiarite. Se non presenti i geni modificatori, potremmo avere ottimi agata pastello, ma non ottimi isabella pastello, poiché avrebbero disegno.
A questo punto vale la pena di ricordare una vecchissima esperienza (anni 70, allevamento Canali): accoppiando un maschio agata pastello buono con una femmina isabella pastello difettosa, per presenze di disegno specialmente sui fianchi, ma molto diluita, nacquero agata pastello di notevole bellezza (anche un 92). Certo solo un esempio ed un caso particolare, non una linea da seguire; ci sono metodi migliori. Tuttavia indicativa di certi meccanismi. Ora non volendo fare solo accoppiamenti in purezza, si può accoppiare l’agata pastello con agata
Non a caso l’accoppiamento fra agata pastello ed isabella pastello
non è consigliato poiché adatto per il tipo base, ma non per il tipo aggiunto pastello
classici, se maschi portatori di pastello, collaudati come non aventi geni modificatori del pastello occulti. Tali geni modificatori sono autosomici e possono essere presenti sia nel maschio come nella femmina, ma nei classici non hanno alcun effetto visivo. Volendo fare isabella pastello, potremmo accoppiare isabella pastello con agata classici, se maschi portatori di isabella e di pastello, collaudati come dotati di geni modificatori del pastello, al contrario del caso precedente. Di solito, dopo alcuni anni di accoppiamenti si hanno se non certezze alte probabilità sulla presenza o meno dei geni modificatori, rilevabili attraverso l’esame della progenie pastello.
Che fare di una femmina come quella descritta all’inizio? Un interrogativo im-






NUMERO 4 - 2024 15
Agata pastello intenso giallo, foto: E. del Pozzo
Bruno pastello mosaico rosso maschio, foto: E. del Pozzo
pegnativo. Certo, accoppiata con un classico isabella o agata, i risultati dovrebbero essere almeno buoni, probabilmente ottimi, per l’ottimo tipo base, ma sarebbe dubbia la presenza di geni modificatori occulti. Se accoppiata per andare verso l’agata pastello, ci vorrebbe un maschio agata pastello del tutto privo di tracce di ali grigie, sperando che tale caratteristica prevalga. Tuttavia non è detto, poiché le tracce di tale effetto si trasmettono abba-
stanza e se ben visibili in una femmina significa che sono notevoli, stante il fatto che l’ali grigie si evidenzia molto meglio nei maschi. Se si volesse andare verso l’isabella pastello, ci vorrebbe un maschio isabella pastello ottimo, sperando che le femmine isabella pastello ottenute siano ottime, anche se non è certo ma abbastanza probabile; non è invece affatto probabile che siano ottimi i maschi agata pastello, poiché molto probabilmente con evidenze di


ala grigia. Certo potrebbero avere un ottimo disegno, ma non basta per la piena tipicità.

Insomma, la selezione dei pastello ha i suoi problemi, in tutti i tipi. Si ricordi inoltre che in bruni ed isabella pastello a volte si seleziona anche contro il disegno in quanto tale e la cosa può fare cattive sorprese, qualora si usassero dei classici. Ci possono cioè essere anche due selezioni, una ortodossa per i geni modificatori, inefficaci sul classico, ed una espediente contro il disegno in quanto tale, che incide negativamente sui portatori classici.
La femmina agata pastello di Gennaro era comunque davvero interessantissima, per le notevolissime rare qualità sopra descritte, anche se in presenza di leggere riduzioni sulle ali.
Chi volesse approfondire gli aspetti del pastello e dell’effetto ali grigie, potrà
Se si volesse andare
verso l’isabella pastello, ci vorrebbe un maschio isabella pastello ottimo, sperando che le femmine isabella pastello ottenute siano ottime
vedere il mio testo “I colori nel Canarino” reperibile in FOI, oltre a diversi articoli su Italia Ornitologica.
Si noti che una tesi, mi dicono diffusa all’estero, secondo la quale l’ali grigie sarebbe una mutazione allelica al pastello, non regge. Se fosse una mutazione, nelle femmine sarebbero possibili solo 2 forme e cioè: pastello tradizionale ed ali grigie, mentre nei maschi al massimo 3 e cioè: pastello tradizionale, ali grigie ed eventualmente intermedio. Le varie espressioni possibili palesano ben diversa situazione. Semmai appare strana la difficoltà di espressione dell’effetto ali grigie nelle femmine; argomento su cui abbiamo disquisito Alfonzetti ed io. Anche qui da vedere il mio testo e diversi articoli.
16 NUMERO 4 - 2024
Agata pastello mosaico rosso, foto: E. del Pozzo
Becchi potenti ma allo stesso tempo sensibili
testo di RAFAEL ZAMORA PADRÓN (*), foto MOISÉS PÉREZ (LPF)



Alla Loro Parque Fundación stiamo attraversando il periodo primaverile. Per i pappagalli è un momento delicato poiché si verificano cambiamenti nell’ambiente che modificano il loro comportamento.


(*)Direttore Scientifico Fondazione Loro Parque
Per uccelli sensibili come questi, la fase riproduttiva comporta reazioni inaspettate alle quali dobbiamo essere preparati
Per uccelli sensibili come questi, la fase riproduttiva comporta reazioni inaspettate alle quali dobbiamo essere preparati.
È possibile che una coppia che ieri si lisciava le piume a vicenda, la mattina dopo può impegnarsi in un’instancabile battaglia che potrebbe porre fine alla vita di uno di loro.
Nella maggior parte dei casi, sono le femmine a farne le spese. Ed è una circostanza che obbliga gli allevatori di pappagalli a prestare particolare
NUMERO 4 - 2024 17 ONDULATIEDALTRI PSITTACIFORMI
Rosella di Stanley, Platycercusicterotis, foto: M Perez LPF






È possibile che una coppia che ieri si lisciava le piume a vicenda, la mattina dopo può impegnarsi in un’instancabile battaglia che potrebbe porre fine alla vita di uno di loro
attenzione in questo periodo dell’anno.
Qualcosa di simile accade con le uova appena deposte. La prima settimana la coppia può mostrare un ottimo comportamento e le uova vengono incubate perfettamente da una femmina ben assistita dal suo maschio. Ma nella seconda settimana le uova potrebbero apparire rotte o scomparire completamente senza che noi comprendiamo ap-
18 NUMERO 4 - 2024
Conuro dorato, Guaroubaguarouba, foto: M Perez LPF
Domicella ventre viola occident., Loriush.devittatus, foto: M Perez LPF
pieno quale degli uccelli sia responsabile di tale azione.
Tra le tante osservazioni che fa l’allevatore di pappagalli c’è quella di vedere attentamente lo stato dei becchi di ciascun esemplare. Un becco poco brillante, secco o difettoso durante la stagione riproduttiva può indicare carenze che possono compromettere i risultati della riproduzione. È per questo motivo che in questa fase dell’anno ai pappagalli bisogna rinnovare frequentemente la somministrazione di grit, osso di seppia e pietra calcarea. Anche i posatoi dovrebbero essere rinnovati in modo che i becchi siano in forma.
Quando una coppia rompe le uova, spesso è a causa della mancanza di calcio. La femmina produce uova morbide oppure il maschio cerca risorse alimentari nel guscio delle uova. Correggere il problema non è difficile ma a volte richiede un po’ di tempo. E bi-
Quando una coppia rompe le uova, spesso è a causa della mancanza di calcio.
La femmina produce uova morbide oppure il maschio cerca risorse alimentari nel guscio delle uova
sogna tenere conto della presenza del sole, che gioca un ruolo importante anche nel metabolismo del calcio dei pappagalli.
I becchi dovrebbero apparire lucenti e puliti. Se sono secchi, macchiati o con escrezioni biancastre, ciò è un indicatore di problemi da correggere. Alla Loro Parque Fundación stiamo osservando in questi mesi un tasso di natalità elevato. Un buon indicatore della vitalità dei genitori, della fertilità e dell’atteggiamento per l’inizio della primavera. Specie come i parrocchetti delle tane (Cyanolyseus patagonus) hanno già i loro primi pulcini e anche specie africane come i pappagalli del Senegal (Poicephalus senegalus senegalus), i pappagalli testa bruna (Poicephalus cryptoxanthus) o gli inseparabili di Lilian (Agapornis lilianae) sono in procinto di vedere i loro pulcini lasciare il nido. Nello stesso momento i Cacatua bianchi stanno preparando i loro nidi e deponendo le prime uova.
Entriamo in azione, in uno dei periodi più interessanti dell’anno.




NUMERO 4 - 2024 19









La Tortora dal collare domestica
Un orientamento comune tra compromessi e selezione
testo di
Prima parte
Il tipo classico, l’onice e il feo a dominanza incompleta Pur essendo uno degli uccelli storici della tradizione ornicolturale d’Europa, la Tortora ridente, o meglio definita oggi Tortora domestica, rimane sempre una specie marginale nelle nostre manifestazioni, per innumerevoli motivi. Nonostante questo, la sua longevità, associata alla prolificità, ha permesso di fissare nella popolazione domestica bellissime varietà di colore che purtroppo ancor oggi spesso non trovano la giusta collocazione genetica e, conseguentemente, la dovuta selezione e un giudizio oggettivo. Questo comporta da un lato l’impossibilità degli organi tecnici di dettare le giuste linee guida e gli orientamenti che consentano univocamente ad ornicoltori e giudici di proiettare
La sua longevità, associata alla prolificità, ha permesso di fissare nella popolazione domestica bellissime varietà di colore



verso un futuro selettivo la specie, dall’altro il perseverare nella confusione l’allevamento, basato paradossalmente su nomi di fantasia e convinzioni personali, il tutto sostenuto dalla fantasia e non dal tecnicismo di bravissimi appassionati stranieri che, diciamo, non condividono l’approccio tecnico scientifico a cui siamo abituati nel movimento F.O.I. A questo si deve
aggiungere il contributo genetico che purtroppo la Streptopelia decaocto e probabilmente anche la chinensis e la senegalensis hanno portato attraverso i numerosi ibridi (voluti e non) che la loro facile reperibilità ha permesso negli ultimi 25/30 anni di realizzare. In particolare la decaocto, passando da specie quasi occasionale a comune ed invadente e presentando
NUMERO 4 - 2024 21 ESTRILDIDI FRINGILLIDI IBRIDI
Tortora dal collare domestica “feo”, all. Chèradame
FRANCESO FAGGIANO, VINCENZO RALLO e COLLABORATORI, foto V. RALLO, F. VERONESE, F. FAGGIANOE V. CHÈRADAME
taglia e colorazioni più decise, ha notevolmente modificato il fenotipo della Tortora domestica. Personalmente ricordo che la mia nonna aveva una piccola voliera in giardino con tortore color caffelatte e bianche (erano probabilmente delle ancestrali e delle albine), di morfologia molto più minuta e gentile delle attuali. Solo all’età di 6 anni vidi per la prima volta in quel di La Spezia una decaocto selvatica e mia nonna mi fece notare che, se pur simile, era ben diversa dalle sue perché molto più grande e molto più scura! Solo qualche anno dopo osservammo, in un piccolo paese del Monferrato dove abitavamo, una coppia di decaocto nidificare per la prima volta su un grande platano. Da allora la diffusione di questa specie è stata frenata solo dalle occasionali epidemie, permettendo l’incrocio casalingo con la più gentile e domestica risoria, che oggi presenta diffusamente tratti non propri.
Spinto dall’entusiasmo vertiginoso del super appassionato Vincenzo Rallo e di altri numerosi amici cultori della Tortora domestica, con i quali condividiamo questi approfondimenti tecnici e questi orientamenti selettivi, tenteremo di creare i presupposti per una sistematicità selettiva che può evidentemente solo migliorare, cominciando a parlare
Con il termine “tipo classico” nella Tortora domestica intendiamo indicare il fenotipo base
tecnicamente del tipo base, ovviamente definibile meglio come “classico” e non “ancestrale”, sia per l’ingentilimento che secoli di allevamento hanno determinato sia per l’evidente condizionamento che la genetica della decaocto prepotentemente comporta.
Vi proponiamo così una base per un successivo inquadramento sistematico delle colorazioni su base genetica e una corrispondente proposta di denominazione tecnica delle varietà di colore di cui abbiamo sufficiente conoscenza per poter indicare una selezione che condivida anche l’orientamento generale di tutte le specie oggi protagoniste delle esposizioni FOI, sul principio che la stessa mutazione deve avere lo stesso nome in tutte le specie e che sia intuitivo e significativo per la selezione ed il giudizio.
Dal tipo classico che, come detto, presenta taglia e struttura fortemente ingentilita e aumentata sia per l’ingentilimento domestico, appunto,


che per l’apporto delle specie cugine, comunque senza privilegiare soggetti fuori taglia e di cui evidenziamo anche la richiesta di un fondo ricco di feomelanina, dai toni leggeri e dal petto rosaceo e non violetto, zampe rosso e non viola, passeremo a denominare e descrivere in questo primo lavoro la varietà onice, la feo dominante sia singolo che doppio fattore. In un secondo lavoro chiuderemo questa prima fase con la descrizione di altri fenotipi empiricamente certi, mentre crediamo opportuno dover aspettare ulteriori conferme per proporre nuove denominazioni e indirizzi selettivi per tutte le altre colorazioni diffusamente presenti e sovrapposte, di cui abbiamo pochi dati oggettivi.
Descrizione del fenotipo classico
Con il termine “tipo classico” nella Tortora domestica intendiamo indicare, come è in uso per altri uccelli in sede internazionale, il fenotipo base o, concedeteci il termine, “il fenotipo ancestrale domestico” della specie in assenza di mutazioni determinanti riduzioni, aumenti e/o modifiche della pigmentazione e/o aberrazioni del disegno e del piumaggio. Nella Tortora domestica il tipo classico deve essere un fenotipo delicato e sobrio che si realizza per la combinazione dell’eumelanina bruna dai toni cannella, beige e caffelatte (quindi tutti toni bruni e freddi, ma non scuri né grigi) con la feomelanina dalle tinte nocciola e fulvo (toni anche questi chiari, ma caldi); quest’ultima è presente soprattutto sullo scudo alare e sul petto, dove realizza un delicato e tipico fondo rosa. Se questo è il fenotipo ascrivibile ad una presunta Streptopelia risoria/roseipectus, non possiamo che ammettere che questo fenotipo sia oltremodo superato da un orientamento selettivo che ha privilegiato toni più saturi e ricchi di eumelanina scura, ma che comunque non devono essere grigi. Rimane fondamentale quindi mantenere un giusto equilibrio espressivo che consenta di distinguere bene la Tortora domestica dalla decaocto e per questo è importante sottolineare alcuni
22 NUMERO 4 - 2024
Tortora dal collare domestica fenotipo classico, all. Faggiano
tratti fenotipici come di seguito riportati, per poi tradurre gli stessi in presenza di determinate mutazioni.
Becco: di colore grigio scuro tendente al bruno (sarebbe bene avesse lo stesso colore delle remiganti).
Occhi: iride di color rosso intenso e pupille nere, con palpebra rosa.
Testa: color cipria saturo ed uniforme di tonalità opaca. La femmina presenta generalmente una saturazione maggiore e quindi un colore più scuro rispetto ai maschi.
Collare: di colore nero. È pregio il sovracollare bianco candido, non più alto di un paio di millimetri.
Gola e Petto: fondo di color beige chiaro infiltrato di rosa uniforme e luminoso, fianchi grigiastri.
Mantello: dorso di color beige nocciola scuro saturo e di tonalità opaca, caratterizzato da sfumature brunastre leggere che danno un effetto cipria.
Groppone: grigio cenere fumato di beige.
Remiganti: remiganti primarie di color bruno grigio più scuro possibile, con una leggera orlatura periferica beige chiaro, rachide scuro.
Scudo alare: beige caffelatte o nocciola uniforme e saturo, con lo stesso caratteristico tono opaco e l’effetto cipria del dorso.
Coda: la parte superiore e prossimale delle timoniere è grigio cenere, con rachide centrale scura, mentre il terzo distale delle remiganti è leggermente più chiaro, con estremità biancastra.
Zampe: di colore rosso scuro con unghie di color grigio.
Basso petto e ventre: rosa che tende a sfumare in un crema chiaro nella parte ventrale quasi bianca.
Pregi:vanno premiati i soggetti che presentano una colorazione uniforme e satura della melanina sempre ed assolutamente nei toni del beige e nocciola e del rosa, come descritto per ogni area. Sono da premiare remiganti e timoniere scure che staccano dal colore di fondo. È apprezzata la pigmentazione bruno scuro ed evidente del rachide centrale delle piume del dorso e dello scudo alare, così come delle remiganti.



Vanno premiati i soggetti che presentano una colorazione uniforme e satura della melanina
E particolarmente gradita un’evidente differenza di tonalità tra la parte superiore dell’animale, più calda e scura, opaca e caratterizzata dall’effetto cipria, mentre la parte inferiore deve essere chiara, delicata e luminosa. Apprezzato, se evidente e regolare, il “ciliare inferiore”, disegno sotto-oculare di color cipria molto chiaro, la cui espressività indica la buona ossidazione dei pigmenti melanici di testa, collo e petto.
Difetti: è difetto grave del colore un’espressione dell’eumelanina sbiadita, troppo fredda o di contro troppo scura e tendente al grigio. Anche schiarite, spesso localizzate su fronte, gola e petto sono difetti del colore, cosi come un fondo tendente al grigio sulla parte superiore e/o vio-
laceo su gola e petto. È difetto l’assenza dell’effetto cipria sull’intero piumaggio. Remiganti grigio chiaro o, viceversa, nerastre e assenza di stacco cromatico tra la porzione superiore e quella inferiore. Anche becco e unghie nere sono difetto perché espressione della decaocto.
Il fenotipo onice
Questa mutazione, autosomica recessiva, è presente da oltre trent’anni negli aviari d’Europa e d’Italia e ha riscosso da sempre un grande successo per l’effetto notevole che comporta su un fenotipo delicato e poco appariscente qual è quello della risoria, determinando un viraggio netto dai toni beige e cipria del tipo classico al grigio perla (chiaro o scuro a seconda della zona considerata). Dopo molti anni in cui si sono alternati pareri discordanti, anche determinati dalla variabilità espressiva della colorazione, che in casi estremi arriva ad un fondo grigio lavanda e ventre biancastro, siamo oggi arrivati a evidenziare che la mutazione non determina formalmente uno schiarimento, ovvero una riduzione delle melanine depositate
NUMERO 4 - 2024 23
Tortora dal collare domestica “onice”, All. Veronese
sul piumaggio, ma anzi ne migliora l’ossidazione e questo lo apprezziamo dal colore del collare, che rimane assolutamente nero, e dalle remiganti e timoniere che diventano nei soggetti migliori grigio antracite scuro, quindi possiamo affermare che non si tratti di opale! Ma determinando di contro la sostituzione del tipo di melanina, depositando cioè eumelanina grigio e grigio scuro al posto dell’eumelanina beige e della feo, è con buona probabilità una onice.
Descrizione del fenotipo onice
Becco: di colore grigio scuro, tendente al nero.
Occhi: iride di color rosso intenso e pupille nere, palpebra grigia.
Testa: grigio perla saturo ed uniforme di tonalità opaca. La femmina presenta generalmente una saturazione maggiore e quindi un colore più scuro rispetto ai maschi. La nuca è spesso interessata da sfumature grigio scuro.
Collare: di colore nero. È pregio il sovracollare bianco candido, non più alto di un paio di millimetri.
Gola e Petto:di color grigio perla chiaro, uniforme e luminoso.
Mantello:dorso di color grigio perla scuro saturo e di tonalità opaca, caratterizzato da sfumature nerastre che danno un effetto fuliggine (importante particolare). Le copritrici scapolari in quasi tutti i soggetti presentano una bordatura della piuma più scura che produce un effetto visivo quasi a scaglie leggero e discreto (importante particolare). Rachide centrale più scuro del vessillo. Groppone: grigio cenere chiaro.
Remiganti: remiganti primarie di color grigio antracite, più scuro possibile, con una leggera orlatura periferica grigio chiaro rachide scura.
Scudo alare: grigio perla scuro omogeneo e saturo, con lo stesso caratteristico tono opaco e l’effetto fuliggine del dorso.
Coda: la parte superiore e prossimale delle timoniere devono avere lo stesso colore grigio cenere del groppone, mentre l’estremità è leggermente più chiara, bordo distale biancastro.
Remiganti e timoniere che diventano nei soggetti migliori grigio antracite scuro
Zampe: di color rosso scuro, tendenti al porpora e unghie di color grigio scuro.
Basso petto e ventre: grigio ghiaccio più chiaro nella parte ventrale. Fianchi grigi.
Pregi:vanno premiati i soggetti che presentano una colorazione uniforme e satura della melanina sempre ed assolutamente nei toni del grigio come descritto per ogni area. Sono da premiare remiganti scure che staccano dal colore di fondo. È apprezzata la pigmentazione nera ed evidente del rachide centrale delle piume del dorso e dello scudo alare, così come delle remiganti e timoniere.
È particolarmente gradita una evidente differenza di tonalità tra la parte superiore dell’animale più scura, opaca e fuligginosa mentre la parte inferiore deve essere chiara e luminosa senza l’effetto fuliggine. Apprezzato se evidente e regolare il “ciliare inferiore”, disegno sotto oculare di colore grigio molto chiaro la cui espressività indica la buona ossidazione dei pigmenti melanici di testa, collo e petto.
Difetti: è difetto grave del colore un’espressione di eumelanina bruna. Anche schiarite spesso localizzate sulla fronte, gola e petto sono difetti del colore, cosi come un fondo tendente al grigio perla chiaro sulla parte superiore e/o l’assenza in queste parti dell’effetto fuliggine. Remiganti grigio chiaro e assenza di stacco
È difetto grave del colore un’espressione di eumelanina bruna
cromatico tra la porzione superiore e quella inferiore. Non è gradito un petto grigio scuro e opaco. Becco e unghie chiare.
Il fenotipo “feo a dominanza incompleta”: genetica e caratteri È questa una interessante colorazione data dalla mutazione a carattere dominante incompleto di un gene autosomico, che comporta così la possibilità di avere due fenotipi differenti tra i soggetti eterozigoti, ovvero singolo fattore per il mutante, che indicheremo come “feo singolo fattore” e i soggetti omozigoti, ovvero puri, che indicheremo come “feo”. La nomenclatura proposta tenta di organizzare l’identificazione tecnica e fenotipica delle tortore domestiche interessate da questa mutazione di colore, che sviluppa in omozigosi un disegno noto agli ornicoltori, dato dalla semplice disposizione periferica del pigmento feomelanico*… se pur la trasmissione ereditaria del carattere sia diversa, possiamo spingerci a dire che le richieste selettive del fenotipo omozigote debbano essere assimilabili ai feo tradizionali che interessano altre specie.
Descrizione del fenotipo feo singolo fattore
Becco: bruno
Occhi: iride di colore rosso scuro, pupille nere palpebre rosa
Testa/fronte guance: lilla intenso e caldo; nelle femmine la testa e la fronte tendono ad essere più sature quasi violetto, mentre nel maschio queste aree diventano cipria, così come appaiono speso le guance.
Collare:di color grigio perla ben evidente. È pregio il sovracollare bianco candido, non più alto di un paio di millimetri.
Collo/petto:di color lilla intenso uniforme e saturo, della stessa tonalità della nuca.
Ventre e regione perianale:basso petto e ventre di colore rosa tenue che schiarisce gradualmente verso la regione anale e di color biancastro, con fianchi grigiastri.
Mantello: fulvo, di color rosso rug-
24 NUMERO 4 - 2024
gine intenso e saturo, sul quale si apprezza una velatura di beige che opacizza leggermente l’espressione della feo. È apprezzata l’assenza di soffusione eumelanica.
Groppone: di color rosso ruggine intenso e saturo. È apprezzata l’assenza di soffusione eumelanica.
Scudo alare: scudo alare di color rosso-bruno intenso e il più possibile uniforme. È apprezzata l’assenza di soffusione eumelanica.
Remiganti: grigio perla uniforme ed espressivo, con rachide centrale scura.
Coda: le remiganti centrali di color rosso ruggine intenso e uniforme, mentre quelle laterali assumono colorazione grigio perla idealmente uguali alle remiganti.
Zampe: di color rosso. Unghie brune. Pregi: vanno premiati i soggetti con la massima espressione feomelanica, che deve dare un espressivo colore lilla/violetto su testa e petto, come descritto, e un colore fulvo sul dorso, scudo alare e groppone. L’espressione piena ed uniforme del colore in queste parti è un pregio. Anche l’uniformità di colore tra collarino, remiganti, fianchi e timoniere esterne è elemento di pregio. Apprezzato, se evidente e regolare, il “ciliare inferiore”, disegno sotto oculare di colore cipria chiaro la cui espressività indica la buona ossidazione dei pigmenti melanici di testa, collo e petto.
Difetti: colore scarico, freddo disomogeneo o infiltrato di toni beige grigiastri. Collarino troppo scuro oppure infiltrato di nero, così come può evidenziarsi su remiganti e timoniere a volte striate. Anche la presenza di perlatura è difetto. Becco ed unghie carnicini sono difetti che indicano un’ossidazione melanica carente.
Descrizione del fenotipo feo
Becco: color beige scuro
Occhi: iride di colore rosso scuro, pupille nere palpebre rosa.
Testa: lilla intenso; la nuca spesso tende al ruggine, con le guance, il mento e la fronte più tendenti al cipria soprattutto nei maschi (il viraggio dal lilla al cipria ci identifica l’azzeramento dell’eumelanina ed è


Anche la presenza di perlatura è difetto. Becco ed unghie carnicini sono difetti che indicano un’ossidazione melanica carente
quindi una condizione adeguata, da non penalizzare).
Collare: di colore bianco candido espressivo, alto e regolare.
Collo/Petto: di color lilla intenso, uniforme e saturo.
Ventre e regione perianale:basso petto e ventre di colore rosa tenue che schiarisce gradualmente verso la regione anale e di color biancastro, con fianchi bianco panna.
Mantello: di color rosso ruggine intenso e saturo, disegnato da perle bianco candido ampie e regolari*. Groppone: bianco, ma è apprezzata l’evidenza anche minima di orlature feomelaniche *.
Scudo alare: di color rosso-bruno intenso caratterizzato da un’orlatura regolare e precisa di ogni piuma, cosi da comporre un disegno a perle bianche *.
Remiganti e timoniere: totalmente bianche.
Zampe: di color rosso, unghie beige. Pregi:vanno premiati i soggetti con la massima espressione di saturazione feomelanica e che presentino un’orlatura ben definita e uniforme, così da realizzare un disegno a perle regolare. È pregio un’orlatura marcata su grandi copritrici alari e caudali. Apprezzato, se evidente e regolare, il “ciliare inferiore”, disegno sotto oculare di colore cipria chiaro. Difetti: colore della testa completamente ruggine e/o cipria. Collo e petto cipria, assenza di stacco tra petto e ventre. Orlature schiarite, irregolari, troppo sottili o cancellate. Difforme colorazione tra collarino, remiganti e timoniere. La presenza di orlature anche leggere o sottili sulla testa o su remiganti e/o timoniere è da considerare difetto. Becco e unghie carnicini.
Nota
Come in molti uccelli, anche nella Tortora domestica ritroviamo una disposizione zonale delle differenti melanine con la porzione centrale più interessata dall’eumelanina e la porzione periferica interessata dalla feomelanina. Nella Tortora domestica l’azione della mutazione feo in stato di omozigosi inibisce quasi totalmente il deposito delle eumelanine, che con la giusta selezione lascia un vuoto cromatico che vediamo bianco candido circondato da un orlo periferico rosso ruggine. Sarà cura dell’ornicoltore selezionare nell’espressione più satura, regolare ed uniforme possibile questo disegno.
NUMERO 4 - 2024 25
Tortora dal collare domestica “feo sf”, all. Rallo
Note della C.T.N.-E.F.I: a corredo dell’articolo: la Tortora dal collare domestica
Nel2021, dopo 72 anni di storia FOI (riconosciuta con D.P.R. 15 dicembre 1949 n. 1166) abbiamo avuto il piacere di assistere alla nascita del primo standard dedicato ad una specie appartenente alla Famiglia dei Columbidae, la Tortora Domestica dal Collare Africana - Streptopelia roseogrisea (cosiddetta risoria). Lo standard è stato concepito dalla Commissione Tecnica Nazionale EFI-FOI, composta da: Carmelo Montagno (Presidente), Gabriele Ragni (Commissario- Rappresentante della Specializzazione EST-Estrildidi e loro Ibridi), Riccardo Rigato (Commissario - Rappresentante della Specializzazione FRI-Fringillidi e loro Ibridi), con la collaborazione degli allevatori Massimiliano Melotti e Vincenzo Rallo.
Durante lo stesso anno 2021 è stato concepito un altro standard dedicato alla Famiglia dei Columbidae, quello della Tortora Diamantina (Geopelia cuneata) a firma della medesima CTN-EFI e con la collaborazione di Massimiliano Melotti e Vincenzo Rallo.
Ed a seguire sempre durante lo stesso anno 2021 è stato concepito un ulteriore standard dedicato alla Famiglia dei Columbidae, quello della Tortora Comune Europea – Streptopelia turtur, a firma della medesima CTN-EFI e con la collaborazione del Club Amatori Avifauna Autoctona e del Giudice FOI/OMJ Dott. Sebastiano Paternò.
Gli standard di riferimento concepiti per le tre specie elencate hanno descritto in maniera puntuale la loro rispettiva collocazione tassonomica ed a seguire i sei considerando presenti all’interno della scheda di giudizio EFI: STRUTTURA, COLORE, DISEGNO, PIUMAGGIO, PORTAMENTO, CONDIZIONI GENERALI, elencando pregi e difetti. Addirittura nel caso specifico della Tortora Domestica dal Collare Africana (Streptopelia roseogrisea -risoria), sono state evidenziate le principali differenze fenotipiche che la contraddistinguono con la Streptopelia decaocto (Tortora Dal Collare Orientale).
Gli standard sono stati regolarmente ratificati dal CDF-FOI. In data 21/08/2021 quello della Tortora Domestica dal Collare Africana - Strep-
topelia roseogrisea e l’altro della Tortora Diamantina - Geopelia cuneata. Ed in data 1819/Febbraio/2022 è stato ratificato quello della Tortora Comune Europea – Streptopelia turtur. E così come accade per tutti gli standard ratificati dalla FOI, il loro contenuto è stato reso di dominio pubblico e facilmente accessibile anche per gli allevatori, attraverso la loro pubblicazione all’interno del sito internet ufficiale della FOI.
Nel caso specifico dello standard della Tortora dal collare domestica, è stato precisato che nonostante vi fossero fra gli aviari diverse mutazioni del fenotipo, dalle caratteristiche genetiche comunque incerte, si preferiva rimandarne ad un successivo momento la loro standardizzazione. E pertanto si procedette a standardizzare soltanto il fenotipo classico. Leggere sulla nostra rivista ufficiale articoli sulla Tortora dal collare domestica è sempre piacevole, tenuto conto che trattasi comunque di una specie poco allevata e selezionata nelle varianti fenotipiche presenti. Addirittura, se si affrontano questioni tecniche che riguardano le mutazioni del piumaggio, l’argomento diventa maggiormente interessante.
Cionondimeno, lo scritto a firma di Francesco Faggiano è stato predisposto in una maniera tale che la CTN-EFI ha ritenuto doveroso redigere la presente Nota, al fine di evitare che i contenuti dell’articolo possano diventare regole selettive per la specie, in particolar modo laddove risultano essere in antitesi con i contenuti dello standard di riferimento ufficiale.
Lo scritto, infatti, non fa alcun cenno alla esistenza dello standard di riferimento per la specie, ratificato dalla FOI nel 2021 e citato in premessa, e detta regole selettive anche per il fenotipo classico non sempre in aderenza con lo standard ufficiale.
A tal proposito abbiamo ritenuto opportuno pubblicare, a corredo di questa Nota e coerentemente con i principi esposti, lo standard ufficiale della Tortora dal collare domestica (ratificato dal CDF-FOI il 21/08/2021), al fine di evitare fraintendimenti fra i lettori, circa gli indirizzi selettivi della specie.
Carmelo Montagno- Presidente C.T.N.-E.F.I.
26 NUMERO 4 - 2024
Standard Tortora Domestica dal Collare Africana Streptopelia roseogrisea (cosiddetta risoria)
Descrizione generale sistematica
Ordine: Columbiformes
Famiglia: Columbidae
Genere: Streptopelia
Specie: roseogrisea
Sottospecie: roseogrisea; arabica
La classificazione Tassonomica corrente, identifica due sottospecie della Tortora dal Collare Africana: la Streptopelia roseogrisea roseogrisea e la Streptopelia roseogrisea arabica
Al Genere Streptopelia, oltre alla Tortora dal Collare Africana (Streptopelia roseogrisea), appartengono altre 14 specie:
1)Streptopelia turtur (TORTORA COMUNE O SELVATICA)
2)Streptopelia lugens (TORTORA FOSCA)
3)Streptopelia hypopyrrha (TORTORA DI ADAMAWA)
4)Streptopelia orientalis (TORTORA ORIENTALE)
5)Streptopelia dusumieri (TORTORA DAL COLLARE DELLE FILIPPINE)
6)Streptopelia bitorquata (TORTORA DAL COLLARE INSULARE)
7)Streptopelia decaocto (TORTORA DAL COLLARE ORIENTALE)
8)Streptopelia xanthocycla (TORTORA
DAL COLLARE OCCIDENTALE)
9)Streptopelia reichenowi (TORTORA DAL COLLARE ALIBIANCHE)
10)Streptopelia decipiens (TORTORA DAL COLLARE DI MONTAGNA)
11)Streptopelia semitorquata (TORTORA DAL COLLARE OCCHIROSSI)
12)Streptopelia capicola (TORTORA DAL COLLARE DEL CAPO)
13)Streptopelia vinacea (TORTORA VINACEA)
14)Streptopelia tranquebarica (TORTORA DAL COLLARE DI BIRMANIA)

Brevi Note Introduttive La Tortora Domestica o Tortora dal Collare Domestica è la forma sviluppatasi in seguito all’ addomesticamento della Tortora dal Collare Africana (Streptopelia roseogrisea). In natura l’areale di distribuzione della sottospecie roseogrisea è collocato nell’Africa Sud-Sahariana, Sud-Ovest della Mauritania e ad Est del Senegambia, Mali, Niger, Ciad e Sudan fino ad Ovest dell’Etiopia. Mentre l’areale di distribuzione della sottospecie arabica è collocato a Nord dell’Eritrea, Nord Nord-Est dell’Etiopia e Nord della Somalia, e in Arabia da Jedda ad Aden (S Yemen).
In ogni caso si rinvengono popolazioni naturalizzate in maniera stabile, probabilmente originarie di tortore abbandonate o fuggite, in Nuova Zelanda, Porto Rico, New Providence (Bahamas), Messico (Guerrero), Stati Uniti SudOrientali (principalmente Florida), Spagna Costiera Orientale e Isole Canarie. L’addomesticamento di questa tortora è avvenuto alcuni secoli fa, tant’è che la prima forma descritta non è stata quella selvatica, ma quella sviluppatasi in ambiante domestico (Streptopelia risoria-Linneo 1758). Dalla quella data, ci sono voluti due secoli per capire che la Tortora, da sempre tanto comune in ambiente domestico e presente in quasi

tutto il mondo, non era altro che la discendente della Tortora dal Collare Africana. Ancora oggi, con estrema leggerezza viene confusa con la Tortora dal Collare Orientale (Streptopelia decaocto). Esistono diverse mutazioni del fenotipo, le cui caratteristiche ancora oggetto di studio, saranno definite e standardizzate successivamente.
Principali differenze fra la Streptopelia decaocto e la Streptopelia roseogrisea (risoria) La Tortora dal Collare Africana (Streptopelia roseogrisea -risoria), assomiglia molto alla Tortora dal Collare Orientale (Streptopelia decaocto), tuttavia è nettamente più piccola (lunghezza 26 cm contro 32 cm della decaocto), con la coda più corta e in gran parte marrone sabbia chiaro sopra, la testa e le parti inferiori sono di un delicato grigio rosato e il ventre e le copritrici sottocoda


NUMERO 4 - 2024 27
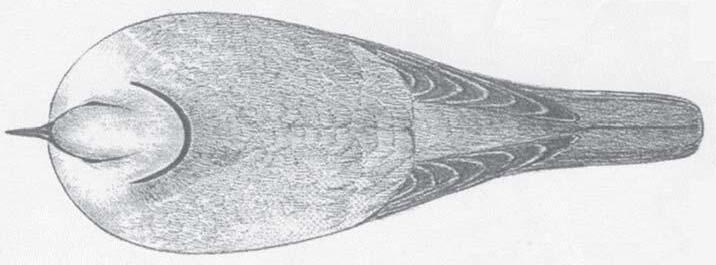
sono bianche. Al contrario, la testa e le parti inferiori della tortora dal collare orientale sono grigiastre e le parti superiori sono brune, più grigie sul dorso. La tortora dal collare africana mostra un bordo nero su tutto il sotto-ala; questa fascia è confinata alle remiganti primarie e alle remiganti secondarie nella tortora dal collare orientale. Sono altrettanto nette le differenze delle sottocaudali, che nella decaocto sono di colore grigio e invece nella roseogrisea (risoria) sono di un bianco sporco, si può notare anche che, nella decaocto, le timoniere presentano una bordatura di nero che nella roseogrisea (risoria) non è presente. Le piume soprastanti alle timoniere sono di color grigiobruno chiaro nella decaocto e invece sono di colore bruno nella roseogrisea (risoria).
Struttura
La lunghezza media del corpo deve essere, dalla punta del becco all’estremità della coda, di circa 28 cm (con un range accettabile compreso tra 26 e 30 cm). Il peso medio è di circa 170g. Il becco deve essere relativamente lungo (da 1,5 a 2 cm) però ben proporzionato ed armonico nell’insieme becco-fronte-testa del soggetto. L’estremità della parte superiore è leggermente ricurva e appuntita. Le due parti del becco, superiore (mascella) e la parte inferiore (mandibola) devono ben combaciare ed essere allineate e non presentare gibbosità o deviazioni. Gli occhi posizionati al centro della testa, contornati da un leggero cerchio oculare, di pelle nuda di colore grigio chiaro. La testa arrotondata in modo uniforme e naturale può essere tollerata anche se un poco allungata. La sommità dovrà curvarsi armoniosamente sia verso la fronte che verso la nuca, senza presentare spigoli o gibbo-
sità. Alla confluenza con il resto del collo e anteriormente con la gola, l’attacco e la curvatura devono essere armoniosi e continui, senza interruzioni o piume sporgenti.E’ da preferire un collo corto e armonioso, che si assottigli dal corpo in direzione della testa con una curvatura dolce e regolare, con un piumaggio liscio ed aderente. Deve essere ben proporzionato con il corpo del soggetto. Il corpo deve essere piuttosto lungo e di forma conica; dal punto più largo, all’altezza delle spalle, deve assottigliarsi gradualmente e regolarmente in direzione della coda. La forma conica la si osserva chiaramente, dall’alto. Il petto deve essere rotondo, pieno e largo visto dalla parte anteriore. Non deve presentare rigonfiamenti o piume sollevate o arricciate. Le ali non devono essere cadenti o troppo sollevate, ma in armonia con la forma affusolata assunta dal soggetto. Osservato di fianco deve presentare una curva del petto che degrada in maniera regolare e armonica in direzione del ventre e della coda. Le ali devono essere ripiegate regolarmente contro il corpo, rendendo uniforme la fisionomia del soggetto. Le remiganti primarie possono aderire alla coda nel suo mezzo senza però incrociarsi su di essa. La coda in posizione normale vista dall’alto non deve superare la larghezza

di una timoniera e mezzo (all’incirca 3-4 centimetri). La coda è composta da dodici timoniere, lunghe all’incirca 12 cm. Le zampe corte e robuste. Ciascuna zampa deve possedere tre dita rivolte in avanti e uno rivolto all’indietro. All’estremità di ciascun dito deve essere presente una unghia di forma compatta e regolare, piuttosto corta. Tutte le dita devono stringere il posatoio quando l’uccello sta sopra di esso.
Colore
I pigmenti responsabili del Colore del piumaggio nella Tortora dal Collare Africana si distinguono in Pigmento Melanico (EUMELANINE e FEOMELANINE). Nel piumaggio della Tortora non è presente Pigmento LIPOCROMICO. Le parti superiori sono fulvo grigiastro pallido, le copritrici secondarie esterne e copritrici primarie grigio-azzurre, le remiganti primarie nere. Le remiganti primarie devono essere ben ricoperte delle copritrici primarie. Le due timoniere centrali sono grigiastre. La collo e il petto sono rosa malva che sfumano nel bianco su mento e ventre. Il caratteristico collare di piume sul retro collo è nero bordato di bianco (dovrà essere delimitato sopra e sotto da una sottile linea di piume bianche). Questo deve estendersi sulla parte posteriore del collo, in maniera netta e regolare. Si deve arrestare con la proiezione verticale di ciascun occhio. La pagina inferiore delle copritrici alari è bianca. Le più esterne grigie superiormente con punte bianche, ma nere sotto con punte bianche. Non essendoci dimorfismo sessuale, i colori nei due sessi sono simili. Nei soggetti giovani i colori sono più chiari e più opachi, le piume di contorno con bordi più chiari e le zampe grigiastre opache; il colore dell’iride cambia dal giallo all’arancione al rosso dell’adulto. La sottospecie arabica, rispetto alla roseogrisea, è generalmente più scura con copritrici inferiori grigio chiaro. Le unghie devono essere dello stesso colore del becco del soggetto. Il colore delle zampe nel fenotipo classico è di colore rosso.
Il Becco: Grigio scuro tendente al nero. Gli Occhi: con palpebre biancastre, pupille nere e iride rosso molto scuro, la pelle orbitaria grigio chiaro.
La Testa (parte superiore, fronte, corona, nuca): Grigio Brunastro tenue, tollerato anche Grigio Bluastro, con evidenti riflessi Malva.
28 NUMERO 4 - 2024
La Faccia: Grigio Brunastro tenue, tollerato anche tendente al Grigio Bluastro, con riflessi Malva. Leggermente più chiara rispetto il resto della testa.
Il Mento: Appena al di sotto della mandibola (gnatoteca) è possibile trovare una piccola area di piumaggio più chiaro (Biancastro o Grigio molto chiaro)
Collo, Gola e Petto: Evidente il collare di piume nere di colore intenso, circondato da un sottile bordo di piume bianche. Tale bordura sarà più ampia sul lato superiore rispetto a quello inferiore. La gola e il petto sono di un colore tenue Grigio Brunastro con evidenti riflessi Malva intenso. Questo colore schiarisce gradualmente in direzione del ventre.
Mantello, Dorso, Groppone: Bruno Grigiastro con predominanza di Bruno intenso e caldo il più possibile.
Scudo Alare, Remiganti Secondarie e Terziarie: Bruno Grigiastro con Bruno predominante. E’ preferibile che il colore primario sia il più uniforme possibile. È tollerata l’ala con sottile bordura di Grigio Bluastro.
Remiganti Primarie: Di colore Nero il più intenso possibile. I margini di ciascuna penna possono presentare un sottilissimo tratto di colorazione più chiara, appena visibile.
Ventre e Regione Anale: Bianco.
Coda: La parte superiore è Bruno Grigiastro con predominanza di Bruno intenso e caldo il più possibile. Le timoniere nella loro pagina inferiore sono nere nella loro prima metà, ricoperte dalle sottocaudali, più chiare tendenti al Biancastro nella loro seconda metà che schiarisce ulteriormente fino all’estremità. Sottocaudali praticamente Bianche.
Zampe: Rosso tendente al porpora. Unghie Grigio scuro tendente al Nero, comunque più scure possibile.
Disegno
La Tortora dal Collare Africana presenta un mantello con disegni semplici e poco articolati. Essendo priva di dimorfismo sessuale, i disegni sono uguali sia per i maschi che per le femmine. Il caratteristico collare di piume sul retro collo, rappresenta il principale disegno del mantello, di colore nero, dovrà essere delimitato sopra e sotto da una sottile linea di piume bianche. Tale bordura sarà più ampia sul lato superiore rispetto a quello inferiore.
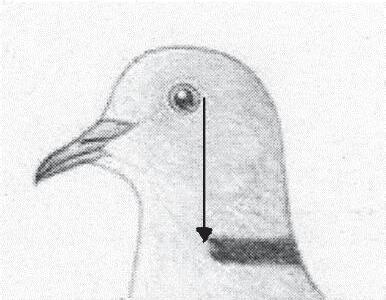
Questo COLLARE deve estendersi sulla parte posteriore del collo, in maniera netta e regolare e si deve arrestare in direzione frontale, con la proiezione verticale di ciascun occhio.
Pregi: l’espressione dei pigmenti più saturi possibile. Così come la tonalità del colore deve essere più calda possibile. Testa, collo e petto presentano i caratteristici riflessi rosei iridescenti (colore strutturale) da cui il nome scientifico (roseogrisea). Ventre e sottocaudali Bianchi, le remiganti primarie e il collarino Nere il più intenso possibile. Difetti: colore del mantello non omogeneo e non abbastanza intenso. Collare non abbastanza spesso o con mancanza di bordo Bianco. Colore del petto e della testa troppo chiari e senza riflessi Malva.
Piumaggio
Il piumaggio della Tortora dal Collare Africana deve presentarsi liscio, serico, composto e non arruffato. Non deve presentare piume e penne spezzate o mancanti. Una leggera usura alle estremità delle remiganti e delle timoniere può essere tollerata solo in animali adulti.
Portamento
In fase di giudizio qualora la posizione del soggetto sia sul fondo dell’alloggiamento, la Tortora deve rimanere calma, formando un angolo di 45° fra la linea del profilo dorsale e il piano di appoggio dell’alloggiamento. La coda può toccare il fondo sempre però se è allineata con il dorso dell’uccello; in questo caso il petto sarà prominente, dando l’impressione di portamento maestoso ed armonico. La coda non deve essere cadente ma allineata con il dorso. Sul posatoio la Tortora manterrà una posizione più orizzontale, rispetto alla postura assunta a terra. Ciò è dovuto al
fatto che la massa corporea è più concentrata sul davanti dell’animale e il peso grava sulle zampe e non viene distribuito anche sulle dita come invece avviene al suolo. L’animale risulterà quindi leggermente proteso in avanti. L’ottimale sarebbe che l’uccello tenga la posizione da terra mentre sta sul posatoio. La coda potrebbe protendere verso il fondo spezzando la linea con il dorso, in quanto l’animale cerca di stabilizzarsi sul posatoio. La posizione ideale è comunque con la coda allineata al dorso, quindi conviene attendere che l’animale sia in posizione stabile sul posatoio. A parità di giudizio e di tutte le considerazioni va preferito il soggetto dal portamento maestoso e dalle condizioni fisiche ottimali. Il corpo affusolato ma con il petto che trasmette condizioni di forza e robustezza, formando una curva armonica con il collo e che vada ad affusolarsi verso il ventre. L’animale si deve mantenere calmo, ma con un atteggiamento comunque vigile. Non deve presentare ali cadenti o coda troppo bassa. Portamento ben ritto sulle zampe e non disteso o accucciato, alcuni soggetti particolarmente confidenti, potrebbero essere in fase di riposo, in questi casi andranno sollecitati a rialzarsi durante il giudizio. I soggetti si debbono presentare calmi al giudizio. Troppo agitati o troppo statici, accovacciati sul posatoio sono da penalizzare.
Condizioni generali
Le zampe devono essere pulite, prive di piume, con la pelle lucida e dal colore adeguato. Tutte le unghie devono essere presenti, della giusta estensione, non presentare assottigliamenti o accenni di spirali per eccessiva lunghezza. Le zampe e il becco non devono presentare scaglie. Il piumaggio non deve risultare sporco o imbrattato di feci specialmente in prossimità della cloaca. Gli occhi devono essere rotondi e ben aperti. Qualsiasi difetto irreversibile dà luogo al non giudizio del soggetto.
Elaborato nell’anno 2021 dalla CTN-EFI: Carmelo Montagno (Presidente), Gabriele Ragni (Commissario-Rappresentante della Specializzazione EST-Estrildidi e loro Ibridi); Riccardo Rigato(Commissario-Rappresentante della Specializzazione FRI-Fringillidi e loro Ibridi)
Collaborazione: Massimiliano Melotti e Vincenzo Rallo
NUMERO 4 - 2024 29









Schemocromi: il fattore opale
testo di MIMMO ALFONZETTI, foto AUTORI VARI
Seconda parte
Diffusione o interferenza
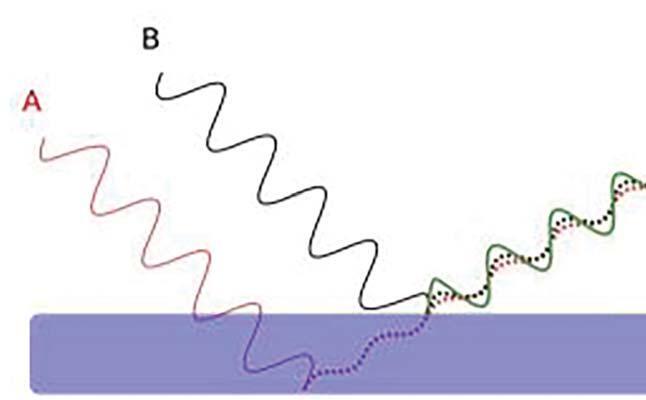
Non esiste, nelle strutture cheratiniche delle penne dei canarini opale, uno specifico pigmento blu in grado di isolare radiazioni luminose azzurrine; ne consegue che tali radiazioni possono trovare fondamento ricorrendo soltanto ad argomentazioni proprie della fisica, in particolare l’ottica ondulatoria. L’azzurro strutturale può originarsi in conseguenza di due precisi fenomeni dell’ottica fisica: la diffusione (scattering incoerente) e l’interferenza (scattering coerente). L’ipotesi avallata da gran parte della letteratura amatoriale (Veerkamp e Zingoni) suggerisce che tale azzurro sia conseguenza di fenomeni diffusivi (effetto Tyndall). Le ricerche più recenti in questo campo hanno dimostrato che i colori strutturali hanno tutti origine nella diffusione coerente. In particolare, si sup-
Tali caratteristiche, comuni a molte specie di volatili, non trovano riscontro nei canarini, nei quali le melanine si trovano addensate in particolari zone di convergenza
Non esiste, nelle strutture cheratiniche delle penne dei canarini opale, uno specifico pigmento blu in grado di isolare radiazioni luminose azzurrine

poneva che nei canarini tali colorazioni dovessero essere supportati da strati di melanine costituite da microgranuli, tutti di dimensioni inferiori alla lunghezza d’onda della luce, assemblati finemente tra loro in maniera uniforme ed omogenea. Tali caratteristiche, comuni a molte specie di volatili, non trovano riscontro nei canarini, nei quali le melanine si trovano addensate in particolari zone di convergenza e strutturate a forma di bastoncelli e addensate in blocchetti più o meno ordinati.
Ne consegue che l’unica spiegazione plausibile sia da ricercare in fenomeni di natura interferenziale. Il gioco dei colori che si osserva nelle bolle di sapone, nelle ali trasparenti delle mosche, nelle
ruote dei pavoni, nel piumaggio del Colibrì, trovano giustificazione proprio in fenomeni ottici di natura interferenziale. I riflessi iridescenti provocati dal movimento delle onde che scuotono il sottile strato di nafta sono un classico esempio di diffusione coerente e ci fanno individuare nell’interferenza costruttiva delle lamine sottili l’argomento della fisica che può aiutarci nei nostri intenti.
A supporto di quanto sopra detto, la foto di fig. 1 (vedi I.O. n. 3/2024) dispensatami dalla CTN di Colore mostra la penna di un canarino nero opale giallo dove è evidente, in maniera indiscutibile, che le radiazioni bluastre siano originate all’interno delle barbule e non delle barbe.
NUMERO 4 - 2024 31
CANARINIDI COLORE
Fig. 11 - Interferenza costruttiva su lamina. la radiazione luminosa riflessa è la sommatoria dei raggi incidenti con componente maggiore azzurra, fonte: SanchariChakraborti,lambdageeks.com
Le barbule del canarino opale che si illuminano di blu rafforzano il mio convincimento che il fattore opale sia la conseguenza di una colorazione ottica strutturale ma non generata da semplice diffusione (effetto Tyndall), bensì da interferenza costruttiva. Le barbule dei canarini opale sono scarsamente melanizzate e quindi manca il presupposto della presenza di melanina necessaria alla diffusione di Tyndall; d’altra parte è certezza scientifica che soltanto le barbule, in conseguenza della loro cheratina nanostrutturata, irradiano per interferenza costruttiva (scattering coerente)
Modello ottico
In ottica si considera lamina sottile qualunque pellicola trasparente il cui spessore sia non più grande di alcune lunghezze della luce; un raggio di luce incidente viene riflesso rispettivamente dalla faccia superiore ed inferiore della lamina. La differenza tra i cammini ottici e lo sfasamento di questi nel tempo fanno formare sulla lamina delle caratteristiche zone luminose, o frange di interferenza. Queste, osservate alla riflessione, appariranno variamente colorate, con valori massimi di intensità la cui lunghezza d’onda è determinabile da formula fisica [λ= lunghezza d’onda, d=spessore, n=indice di rifrazione]. Si tratta ora di individuare una particolare zona strutturale delle penne che possa essere assimilata ad una lamina sottile.
eterm
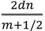
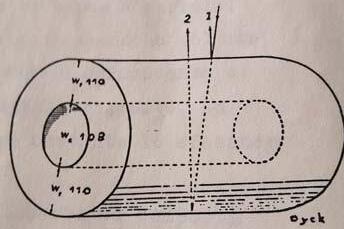
La sezione di una barba alla quale qualunque specie di uccelli può essere ricondotta presenta intorno alla zona midollare un sito, zona spugnosa, dove possono nascere i colori strutturali. Tale zona, costituita da cheratina e spazi di vuoto d’aria, circonda una zona midollare che nell’opale è ricca di melanine. In tale sito spugnoso è possibile circoscrivere quelle strutture cilindriche ipotizzate da J. Dyck (vedere IO n°1-2024) che possono essere assimilati a delle lamine ottiche, il cilindro di J. Dyck.
Nella figura successiva, invece, viene evidenziata opportunamente ingrandita una probabile lamina, estrapo-
Di tali cilindri l’autore ne fornisce anche le dimensioni. Personalmente ho utilizzato la formula della fisica ottica sopraindicataalfinedi prevederelafrequenzadelprimo massimo costruttivo: λ= = 2x1100x1,1/0,5 =4840 Angstrom.Talevalorerientrandonellagammadi frequenzadell’azzurro,lacuibandairradiatra4004-5.000 AngstrongconfermalavaliditàdelmodellodiJeanDyck. Ivaloricalcolatiperglialtrimassimidimuguale1,2… nonsidispongononellaregionedelvisibile.
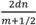
L’unicaincertezzanellecalcolazionisopraindicate èdovutaalvaloreipotizzatodellacheratinaottenuto medianteivaloridimaterialisimilariaquesti
In ottica si considera lamina sottile qualunque pellicola trasparente il cui spessore sia non più grande di alcune lunghezze della luce
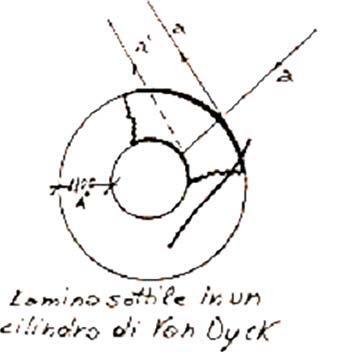
Fig. 13 - Schematizzazione cilindro cavo come lamina sottile, fonte: M.Alfonzetti,personalefigurazione
lata dal cilindro cavo di J. Dyck, con i due raggi riflessi a e a’ in interferenza costruttiva se in fase; in realtà, a causa dell’estrema esiguità delle dimensioni e della zona di riflessione, la lamina è da considerare a facce piane parallele, anche se in figura appare di aspetto diverso.
Variabilità dell’azzurro dell’opale Il canarino opale mostra radiazioni azzurre le cui tonalità, anche se variabili, si collocano in una banda abbastanza ristretta. La variabilità è associata a modeste variazioni delle dimensioni delle strutture ottiche ipotizzate (variazioni dello spessore delle microlamine), a variazioni dell’angolo di incidenza delle radiazioni, a variazioni dell’indice di rifrazione. Ci si può domandare come mai, nonostante tali variazioni, l’azzurro mantenga un tenore quasi costante. La risposta più ovvia è che il fine intreccio di barbe e barbule, nonché l’ultrastruttura interna di queste, consenta soltanto alle radiazioni che hanno un preciso angolo di incidenza di interessare le lamine; i raggi sono obbligati, pertanto, ad emergere da queste in maniera da mantenere costante lo sfasamento. D’altra parte, il reticolo di barbe e barbule si comporta come fonte di diffrazione ottica ed è proprio questo fenomeno che condiziona il tenore quasi costante dell’azzurro.
L’effetto cromatico del canarino opale è sempre la conseguenza della sovrapposizione degli effetti della interferenza e dell’azione di filtraggio operata dalla diffrazione della fitta trama del piumaggio.
Volendo con parole semplici descrivere il risultato della sovrapposizione, si può affermare che solo l’interferenza provoca frange nella quali l’energia luminosa è distribuita uniformemente nel range dell’azzurro, mentre la diffrazione opera sulla ridistribuzione dell’energia modificando l’intensità delle frange.
Disordine del piumaggio
La mutazione opale per diversi anni ha prodotto per un generico effetto epistatico, scompostezza nel piumag-
32 NUMERO 4 - 2024
Fig. 12 - Cilindro cavo di Jean Dick, fonte: J.DyckStructure and colour of the blue barbs.
gio, penne arricciate. La selezione di questi anni, mirata a disegni più melanizzati e la diversa disposizione delle melanine meno concentrata nelle pagine inferiori delle penne hanno portato ad un deciso miglioramento della qualità del piumaggio. In genere, la scompostezza del piumaggio può originare da diverse cause, la cui dipendenza è:
Genetica:
· Alcune linee di canarini arricciati hanno una predisposizione genetica per sviluppare piume contorte.
· Questo problema può essere trasmesso attraverso il DNA e si manifesta più frequentemente in soggetti con determinate caratteristiche genetiche.
· Il fenomeno arricciature anomale è limitato ai soli neri opale. Problemi di piumaggio:
· I canarini con un piumaggio troppo lungo e soffice sono più inclini a sviluppare piume contorte.
· La morbidezza eccessiva delle piume può ostacolare la loro crescita corretta, portando alla formazione di masse sottocutanee fatte di piume o penne attorcigliate su sé stesse.
In particolare per i canarini opale sono state date diverse giustificazioni, anche diverse da quelle precedentemente elencate, non tutte convincenti. È mia personale convinzione che la motivazione sia da ricercare nella disomogeneità della cheratina in relazione all’elasticità delle sue fibre. La presenza di melanina addensata nella pagina inferiore delle penne dei canarini opale ne irrigidisce le fibre. Non è un caso che le penne degli uccelli destinate a sostenere il volo siano maggiormente ricche di pigmenti melanici. Quindi le fibre inferiori risultano essere meno elastiche di quelle superiori; queste avranno un coefficiente di elasticità maggiore. L’evidente anisotropia (3) dovuta alla diversa elasticità delle fibre di cheratina è la causa delle piume arricciate. Le tensioni interne indotte dalle forze di espulsione dovute all’intensa proliferazione e corneificazione messe in atto dal germe della papilla provocano, a parità di

Fig. 14 - Bruno Mogno: a) parteinferioredellapennadiunacopritriceconmacromelanosomicomequellidegliuccelliopali - Nero Mogno: b) sezionetrasversalediunapennaforte;lapartesuperioreèpiùcosparsadimelaninarispettoallaparte inferiore, fonte: NorbertSchramm-UberdenMogno
tensione interna, deformazioni positivamente collegate ai coefficienti di elasticità: le fibre della pagina superiore si allungano più di quelle inferiori. La piuma dovrà necessariamente incurvarsi e quindi arricciarsi; non a caso, le piume deformate mostrano la concavità verso l’alto.
Melanosomi cavi
Una interessante ricerca sullo sviluppo della pigmentazione (4*) nelle penne degli uccelli asserisce che questa sia dovuta ad una particolare presenza di melanosomi cavi, forme di melanosomi uniche che non si ritrovano nel Metazoo (7). È questa una innovazione evolutiva limitata agli uccelli.
Non è ancora chiaro come avvenga l’appiattimento, ma l’evidenza suggerisce che alcuni melanosomi diventino cavi dopo la loro deposizione
I nuclei solidi dei melanosomi nei melanociti si disintegrano gradualmente e diventano cavi nelle cellule cheratiniche delle barbule
nelle cellule delle penne in via di sviluppo. I nuclei solidi dei melanosomi nei melanociti si disintegrano gradualmente e diventano cavi nelle cellule cheratiniche delle barbule.
Gli autori hanno scoperto che i melanosomi si formano come corpi solidi nei melanociti. Più tardi nello sviluppo, soprattutto dopo il posizionamento nelle barbe in via di crescita, i loro interni si dissolvono e lasciano i nuclei cavi. Questi melanosomi ormai cavi sono inizialmente disorganizzati nelle penne, ma diventano compatti man mano che vengono trascinati e distribuiti verso le aree di pigmentazione melaninica.
Una nota di relativa importanza riguarda le penne degli uccelli iridescenti, dove i melanosomi si dispongono nel bordo delle barbule, probabilmente per una combinazione di forze, tra cui l’attrazione e la deplezione.
Tali melanosomi cavi racchiudono una combinazione otticamente energica di indici di rifrazione alti e bassi (rispettivamente della melanina e dell’aria) che consente di produrre colori fisici più brillanti e più saturi rispetto ai melanosomi solidi. Non so dire se questa nanostrutturazione sia presente anche nei canarini opale ma mi attrae pensare che lo sia. Potrebbe essere una giustificazione che, nonostante la drastica rarefazione dei pigmenti melanici, le radiazioni bluastre fossero ascrivibili anche a questa modifica strutturale.
NUMERO 4 - 2024 33
Considerazioni
Sono giudice OMJ, da oltre quarant’anni giudico in mostre nazionali e internazionali; ho allevato con discreto successo bruno opale per diversi anni ma devo confessare che ho qualche perplessità nel giudizio dei canarini opale in questo periodo. I miei maestri mi avevano educato alla frase dirimente che semplificava il giudizio: “Ha l’azzurro? No! ...e allora al massimo sarà sufficiente”. Da alcuni anni a questa parte stiamo osservando canarini opale con maggiore ossidazione delle melanine, maggiore ampiezza delle striature del disegno, sensibile perdita di opalescenza e cambiamento della tonalità del colore dell’eumelanina, in particolare nel tipo bruno, dove sembra vetusta la tonalità bruno bluastro-grigiastro di cui si parla negli standard. Non so dire se sia solo conseguenza di selezione o l’intervento di una nuova mutazione. Personalmente ritengo valide entrambe le ipotesi.
Il gene melanofilina MLPH è estremamente instabile e multiallelico.
Una possibile trascrizione errata, anche minima, dell’RNA del codice genetico può aver avuto come conseguenza una diversa traslocazione dei melanosomi nelle strutture cheratiniche, interessando anche le parti mediane delle barbe. Più semplicemente, non un sollevamento dei melanosomi ma una distribuzione più omogenea di questi.
È mia convinzione che il tutto possa rientrare nei criteri di penetranza ed espressività della mutazione del gene MLPH. Espressività variabile è un concetto, nella genetica, che si riferisce alla variabilità qualitativa e quantitativa con cui si manifesta un genotipo. In altre parole, anche se due individui hanno lo stesso genotipo, possono esprimere caratteristiche diverse a livello fenotipico. In particolare, una espressività variabile non completa, che oscilla nel range compreso tra due estremi fenotipici, la mutazione opale e l’espressione mogno, potrebbe giustificare questi cambiamenti fenotipici. Al momento, il confine superiore di questa espressività variabile è proprio rappresentato dal
Una espressività variabile non completa, che oscilla nel range compreso tra due estremi fenotipici, la mutazione opale e l’espressione mogno, potrebbe giustificare questi cambiamenti fenotipici
fenotipo mogno e non escludo che in un prossimo futuro tale confine possa essere spostato più in alto. La ricerca del prof. Fontanesi ha accertato l’identica natura genetica dei mogno con il fattore opale. Nei canarini mogno è stata trovata la stessa variante allelica dell’opale. Si può quindi concludere che i mogno siano semplicemente dei canarini opale la cui espressione evolutiva è conseguenza dell’uso deliberato di uccelli ricchi di melanina e di un’adeguata selezione. Il mogno sarebbe una particolare selezione dell’opale e non è un caso che le indagini al microscopio ottico (Norbert Schramm) mostrino che i mogno formano macromelanosomi, come si verifica negli uccelli opale. Ad onor del vero, non manca chi esprime grosse perplessità (Canali e Crovace) sulla possibilità nei canarini mogno di rialzare l’eumelanina fino alla pagina superiore ed inibire totalmente la feomelanina solo con la selezione.
A margine, un po’ tutti i criteri di giudizio nel descrivere il fattore opale parlano di fattore di “riduzione” delle melanine. A rigore non esiste una riduzione delle melanine, anzi la produzione di melanina è aumentata, tanto che si formano macromelanosomi. Quello che cambia è invece il processo di traslocazione dei melanosomi nelle penne, dove in effetti provoca una sensibile riduzione della pigmentazione. I testi americani parlano invece di diluizione, termine da noi usato specificatamente per i lipo-
cromi, a mio avviso forse più consono perché risponde meglio alla distribuzione limitata e più omogenea diluita nelle barbe degli uccelli con mutazione lavanda.
Note
(1)Tecnica per la quale si ottengono immagini fotografiche con l’aiuto del microscopio, cioè immagini notevolmente più grandi dei rispettivi oggetti
(2)Il pleocroismo Ë un fenomeno osservabile in alcuni cristalli, che si manifesta con una variazione di colore osservando il cristallo da direzioni diverse. Se i colori diversi osservabili sono due, si parla di dicroismo
(3)Anisotropia: la proprietà per cui il valore di una grandezza fisica (durezza, resistenza alla rottura, velocità, indice di rifrazione, ecc.), in una sostanza o nello spazio, non è uguale in tutte le direzioni
(4)MC1R controlla lo switch dalla feomelanina alla eumelanina, di rilevante importanza nella melanizzazione della pantera nera
(5)MSH: Ormone melanoforo che stimola la produzione e il rilascio di melanina da parte dei melanociti, secreto dall’ipofisi.
(6)Ontogenesi, in biologia, è l’intero processo di sviluppo di un organismo
(7)Metazoo è l’insieme di tutti gli organismi pluricellulari
FONTI
Dirk Van den Abeele - Ornitho Genetics, VZW
· G. Bertarini - I.O. Onice e Opale, Gennaio 2022
· Bertrand Bed’hom, Mohsen Vaez, Jean-Luc Coville, David Gourichon, Olivier Chastel, Sarah Follett, The lavender plumage colour in Japanese quail is associated with a complex mutation in the region of MLPH that is related to differences in growth, feed consumption and body temperature
· Matthew D. Shawkey, Liliana D’Alba, Ming Xiao, Matthew Schutte and Richard Buchholz - Ontogeny of an Iridescent Nanostructure Composed of Hollow Melanosomes (4*)
· Mohsen Vaez, Sarah A Follett, Bertrand Bed’hom, David Gourichon, Michèle TixierBoichard & Terry Burke - A single point-mutation within the melanophilin gene causes the lavender plumage colour dilution phenotype in the chicken
· Norbert Schramm, Dresden (Germany) – Uber den Mogno
· Samuele Bovo, Anisa Ribani, Giuseppina Schiavo, Valerio Joe Utzeri, Luca Fontanesi: Analisi del genoma del canarino per la caratterizzazione delle basi genetiche e molecolari della colorazione del piumaggio conosciuta come “opale”
· Liliana d’Alba e Matthew D. Shake - Melanosomes: biogenesis, properties and evolution of an ancient organelle Xufeng Wu and John A. Hammer - Melanosome transfer: It is best to give and receive.
· M. Penzo - Mogno vs Opale
34 NUMERO 4 - 2024
Canarino x Ciuffolotto
di PIERCARLO ROSSI, foto D. CAUTILLO, P. SALDARINI, G. FUSI, P. ROSSI eG. BOMMARITO


Ed eccoci qui, nuovamente, a parlare degli ibridi con la ciuffolotta, regina indiscussa in questo campo.
Gli ibridi con il genere Pyrrhula hanno un fascino particolare: basti pensare a quello con il fanello (C. cannabina), senza ombra di dubbio uno dei più “eleganti”, con le sue calde tinte pastello; splendidi quelli realizzati con i Serinus esotici, sicuramente tra i più ambiti. Io, che sono un grandissimo appassionato del canarino del Mozambico, lo sogno con il piccolo Crithagra, ma fino ad oggi purtroppo ho potuto ammirarlo soltanto in foto, o con il cardellino, con una funzione perfetta dei colori dei due parentali.
Molti altri rappresentanti dell’avifauna italica hanno creato ibridi meravigliosi


La ciuffolotta risulta essere una femmina molto disponibile con maschi anche non conspecifici
con la ciuffolotta, ad esempio quello con l’organetto; interessanti anche quelli nelle versioni mutate, quello con il verdone con il suo aspetto fiero e la calottina nera tipica del Pyrrhula. Anche con questa specie abbiamo potuto ammirare soggetti a fenotipo mutato, sebbene a mio modesto parere il più bello è quello con il cardellino, l’ibrido per an-
tonomasia, la perfetta fusione tra due degli indigeni più belli. Mancano all’appello quello con il verzellino, il venturone, il fringuello, la peppola ed il verdone dell’Himalaya, che io definisco “il re Mida delle ibridazioni” per gli splendidi soggetti che scaturiscono ogni volta che viene utilizzato questo intrigante Chloris esotico.
La ciuffolotta risulta essere una femmina molto disponibile con maschi anche non conspecifici ed è in grado di conferire ai giovani ibridi un disegno netto, la forma tondeggiante ed aggraziata tipica di questa specie, la calotta nera, ed il rosso del petto ai soggetti di sesso maschile; gli F1 avranno un carattere docile e mite tipico del Pyrrhula. In questo mio scritto vorrei invece parlare dell’ibrido di canarino x ciuffolotto:
NUMERO 4 - 2024 35 ESTRILDIDI FRINGILLIDI IBRIDI
Canarino pastello x Ciuffolotto pastello, all.: P. Saldarini, foto: D. Cautillo
Canarino x Ciuffolotto, all.: F.Rigamonti
i più penseranno che, dovendo utilizzare il maschio di canarino, i giochi si semplificano, ma in realtà non è proprio così.
La ciuffolotta solitamente accetta di buon grado il maschio Serinus, che la conquisterà con il suo canto melodioso, ma a fronte di un numero elevato di uova deposte, quelle feconde che giungeranno alla schiusa, creando una nuova vita, saranno veramente poche. Andiamo ora ad analizzare le possibili cause di tali problemi.
Per prima cosa, analizzando il fenotipo delle due specie in oggetto, troveremo due soggetti molto diversi tra loro: il Serinus canaria, con alle spalle molti anni di allevamento in ambiente controllato, con un disegno dorsale molto marcato, nei soggetti melaninici, che viene sempre trasmesso alla prole ibrida; un disegno facciale non sempre evidente che ricorda quello del verzellino, il becco corto e conico.
Il ciuffolotto, dal canto suo, può essere considerato il gioiello dei nostri boschi: i vivaci colori del maschio contrastano con la sobrietà elegante della femmina.
Il suo corto becco di colore nero, forte e conico, dai bordi taglienti è un ottimo strumento per staccare gemme, rompere gusci e sbucciare semi, molto diverso da quello del Serinus canaria. Oltre a questa particolarità ve ne sono altre che dimostrano la grande differenza tra le due specie, come ad esempio la parata nuziale del maschio Pyrrhula, che è lunga, interrotta da piccoli graziosi saltelli ed inclinazioni laterali della testa, una corte estenuante, bella e romantica, che incanta la ciuffolotta, ma che scoraggia spesso le femmine non conspecifiche.
Ed in ultimo la dieta alimentare, molto diversa tra le due specie: il canarino è un granivoro “puro” che fa dei semi la parte preponderante della propria dieta, mentre il ciuffolotto è un frugivoro parzialmente insettivoro; una buona fetta della sua alimentazione è basata sulle bacche (biancospino, sorbo degli uccellatori, etc.), sui germogli di molte piante (in special modo quelle da frutto), un numero importante di insetti, afidi, oltre alle piccole chioccioline, lasciando che i semi siano soltanto una piccola parte degli alimenti consumati giornalmente.

Tutto questo ci fa capire che le due specie sono molto diverse tra loro e sicuramente lontane anche geneticamente, pertanto questo incrocio è tutt’altro che facile; infatti, a fronte di un numero importante di uova deposte, quelle feconde risultano essere relativamente poche, anche nel caso di ripetuti accoppiamenti.

Anche se non facile da realizzare, nelle mostre ogni anno è possibile ammirare soggetti scaturiti da questo accoppiamento; prima dell’avvento delle mutazioni, splendidi soggetti nero bruni di sesso maschile erano sempre molto ammirati nelle varie competizioni. Tra gli altri, come non ricordare quelli apparsi sulla copertina della rivista “Uccelli” agli inizi degli anni Novanta, realizzati da Patrizio Mariangelo Di Carmine: due soggetti strepitosi, visto che in quegli anni i ciuffolotti utilizzati erano quelli delle nostre Alpi, cosa che conferiva agli ibridi colori saturi, bellissimi.
Apparvero poi i primi soggetti mutati di sesso femminile: ne ricordo uno bellissimo del grande Piero Castellanza, nella mutazione satiné, una vera chicca che a quel tempo ci faceva sognare, viste le forme arrotondate del ciuffolotto ed i disegni del Serinus e gli occhi rossi, un ciuffolotto più piccolo e mutato, una favola.
Prima di chiacchierare dell’ibrido classico, ossia quello con la femmina di ciuffolotto, vorrei soffermarmi sull’utilizzo del maschio in ibridazione.
Il maschio Pyrrhula non viene quasi mai utilizzato in ibridazione; questo, come in altri casi, per una serie di problematiche: ad esempio parata nuziale e scarsa fecondità, che gli hanno conferito uno stigma che ha fatto sì che venisse escluso dai più a priori, ma andiamo ad analizzare nei dettagli.
Osservando la parata nuziale del ciuffolotto, descritta in precedenza, si può notare come la ciuffolotta, quando assume la classica posizione di invito all’accoppiamento, con dorso incurvato e ali tremolanti, mantenga questa posizione per un tempo superiore alla stragrande maggioranza degli altri fringillidi e si offra al maschio più frequentemente durante l’estro. Le canarine, ma ancor più altre femmine indigene (cardellina, verdona etc.) si offrono al maschio per pochi secondi alla volta e poche volte al giorno, quindi se il maschio non è pronto ad accoppiarsi difficilmente la fecondazione avrà successo. Ad onor del vero, diversi allevatori di questa specie mi hanno confermato che questa parata viene eseguita solamente all’inizio della stagione riproduttiva, mentre dalla seconda covata l’accoppia-
36 NUMERO 4 - 2024
Soggetto intenso, all.: G.Lamperti, foto: D. Cautillo
Canarino pastello x Ciuffolotto pastello, all.: P. Saldarini, foto: D. Cautillo


mento avviene “al salto”, come nella stragrande maggioranza delle specie. Anche il mito della monogamia, dovuto forse ad una “favola” tutta umana, è stato sfatato e diversi allevatori utilizzano maschi con determinate caratteristiche, o portatori di particolari mutazioni, con più femmine in una singola stagione riproduttiva.
Rispetto ad altre specie di fringillidi, analisi di laboratorio hanno rilevato che gli spermatozoi di questa specie sono sorprendentemente diversi, nella loro morfologia grossolana, da quelli di tutti gli altri fringillidi ed anche della maggior parte degli altri uccelli passeriformi, risultando essere poco mobili e scarsamente vitali.
Tuttavia, chi ha tentato questa strada, negli anni ha testimoniato di aver assistito ugualmente a numerosi accoppiamenti conclusisi con rarissime schiuse. Comprendere le basi biologiche dell’incapacità degli spermatozoi del ciuffolotto di fecondare le uova di altre specie fornirà utili spunti sui meccanismi di isolamento riproduttivo post-copulatorio, di cui non si sa quasi nulla negli uccelli.
A fronte di quanto appena riportato, mi sono chiesto se nel corso degli anni, o addirittura dei secoli, questo tipo di accoppiamento fosse stato tentato occasionalmente o in maniera più convinta, pertanto ho deciso di documentarmi spulciando vecchi scritti dei cugini d’oltremanica, grandi appassionati di ibridi, in special modo tra specie autoctone ed il canarino, ed ecco cosa ne è emerso. Gli allevatori di uccelli hanno prodotto
Gli allevatori di uccelli hanno prodotto ibridi tra diverse specie di fringillidi europei sin dall’inizio del 1600
ibridi tra diverse specie di fringillidi europei sin dall’inizio del 1600 (Birkhead, 2003).
Fino al 1887 solo cinque ibridi canarinociuffolotto erano stati dimostrati consolidati come identità.
Da quell’anno i casi si fecero sempre più frequenti, anche per il diffondersi dei bollettini e delle riviste ornitologiche a livello amatoriale.
Partridge ci ha inoltre informato che tra gli anni ‘40 e ‘60 gli allevatori di uccelli fecero migliaia di tentativi per allevare ibridi ciuffolotto-canarino utilizzando ciuffolotti maschi ma, a suo parere, senza prove convincenti di successo.
In un libro tedesco sugli usignoli (Luscinia megarhynchos) e sui canarini, l’autore anonimo discute anche degli ibridi dei canarini e fa riferimento a un ibrido ciuffolotto-canarino derivante da una giovane femmina di canarino e da un vecchio maschio di ciuffolotto sessualmente molto attivo (Anonimo, 1772).
Il marchese di Piolenc assicurava di aver visto un ciuffolotto accoppiato con una femmina di canarino e da quell’unione sarebbero nati dei piccoli verso i primi
di aprile; avevano becchi più grandi dei canarini della loro stessa età e cominciavano a ricoprirsi di una peluria nerastra, il che dava motivo di credere che somigliassero più al padre che alla madre; purtroppo, morirono tutti durante un breve viaggio.
Un caso riportato sulla rivista di birdkeeping Cage and aviary birds (Anonimo, 1919) registra che un maschio di ciuffolotto avrebbe prodotto prole con due femmine di cardellino nella voliera, nella stessa stagione riproduttiva. Il documento proveniva da un certo Sir Luke White ed era di particolare interesse perché, come affermò l’editore, si trattava di “un altro allevatore di uccelli con una notevole esperienza nell’allevamento di ibridi”.
J. Dunne tentò di allevare ibridi tra ciuffolotti maschi e canarini femmine, in risposta all’offerta fatta da P. Howe, segretario dell’allora sezione British Birds Breeders del British Mule and Hybrid Club, di una medaglia d’oro per la prima persona che avesse prodotto un ibrido da un maschio di ciuffolotto x canarino. Ogni anno per 22 anni Dunne ha accoppiato un diverso ciuffolotto maschio e una femmina di canarino in una grande gabbia senza altri uccelli. Durante questo periodo produsse sei uova fertili, quattro delle quali si schiusero ma in nessun caso i pulcini sopravvissero più di quattro giorni.
R. Caton ha accoppiato canarini femmine con ciuffolotti maschi che erano stati baliati da canarini in modo che subissero un imprinting. Sebbene tre cop-
NUMERO 4 - 2024 37
Soggetto nero Bruno, all.: F.Rigamonti, foto: P. Rossi Approcci iniziali, all. e foto: G. Bommarito
pie ciuffolotto-canarino formassero legami sessuali, evidenziati dal corteggiamento e dall’accoppiamento regolare, nessuna delle circa 40 uova totali mostrava segni di fertilità o sviluppo.
Tra il 2004 e il 2006 un allevatore, in collaborazione con due esperti allevatori di uccelli, S. Fitzpatrick e R. Drinkwater, ha creato 24 coppie ciuffolotto-canarino. In circa la metà di questi casi i ciuffolotti erano stati allevati da genitori adottivi di canarini e in cinque casi il ciuffolotto maschio aveva ricevuto un’impronta sessuale sul partner canarino, come dimostrato dal regolare corteggiamento e dalla reciproca alimentazione. Queste 24 coppie ciuffolottocanarino hanno prodotto un totale di 78 covate, per un totale di 291 uova. Esaminato in dettaglio il disco germinale di ciascuna di queste uova, nessuna ha mostrato alcun segno di sviluppo.
Tutto questo scoraggerebbe anche i più arditi, ma fortunatamente gli allevatori non demordono; infatti, nei giorni scorsi, il grandissimo appassionato di ibridi Francesco Coletti mi ha inviato un video, che ho condiviso sulla pagina Facebook del Club dell’Ibrido, dove all’interno di una comune volieretta si vede un maschio di ciuffolotto che imbecca ripetutamente una femmina di verdone, intenti nell’allevamento di un paio di pulli, che anche se molto piccoli si capisce che sono ibridi di ciuffolotto dalla forma del becco e dalla peluria scura. Questo ci fa ben sperare per futuri tentativi con il maschio Pyrrhula, sfatando forse un vecchio tabù che ci portiamo avanti ormai da troppi anni.
A conferma di tutto ciò, due anni fa l’amico Renzo aveva unito un maschio pastello bruno, rimasto vedovo, con una femmina di canarino rossa brinata: in una covata alcune uova risultano feconde, anche se purtroppo non si schiusero.
Inoltre, io credo che gli ibridi con il ciuffolotto siano sicuramente molto affascinanti, pertanto molto tentati ogni anno, a discapito di quelli che noi allevatori, nel tempo, abbiamo etichettato come “impossibili” quali ad esempio


quelli con il maschio Pyrrhula o fringuello x canarino; forse, se quest’ultimo fosse tentato con la stessa insistenza di quelli con il Pyrrhula... Rispetto al passato, un allevamento sempre più mirato alla selezione ci ha tolto il gusto di “giocare”: un tempo gli uccelli venivamo posti in voliera promiscua ed alcune volte qualcosa di interessante ed inedito usciva fuori.
Chiudiamo il cassetto dei ricordi e vediamo di tornare sul pezzo, andando ora ad analizzare gli ibridi con la femmina di ciuffolotto.
Prima dell’apparire delle nuove mutazioni nel canarino, i maschi utilizzati erano sicuramente dei nero bruni a fattore rosso. Questi ibridi appaiono ben proporzionati e presentano anche loro la calottina nera e la mascherina, sempre di colore nero intorno al becco. Quest’ultima giunge fino all’occhio e lo attraversa con una stria nera che si pone parallela alla calotta. Sulla fronte è sempre presente una macchia di colore rosso più o meno estesa.
Il parentale canarino dona a questi ibridi
uno splendido disegno dorsale che ben si evidenzia su un colore di fondo grigio. Tale disegno, appena accennato, è presente anche sui fianchi. Le ali e la coda sono nere. È presente una barratura alare che assume una colorazione rossa, più o meno intensa; anche il codione è di colore rosso. Il becco risulta essere di colore grigio piombo più o meno intenso nella parte superiore, con la valva inferiore di color carnicino e con l’apice scuro.
Nel tempo vennero usati canarini pastello, agata, isabella, satiné che diedero vita a rarissime femmine mutate: quando erano presenti in mostra, gli avversari non avevano scampo. Poi apparvero le mutazioni anche nei Pyrrhula ed allora come non ricordare lo splendido maschio pastello realizzato da Mauro Menenti, ma anche il maschio bruno di un maestro come Lino Clerici.
Vorrei terminare questo scritto con quattro big dell’ibridazione, che negli anni hanno realizzato soggetti che rimarranno per sempre nella storia di quest’arte.
Incominciamo da Giancarlo Lamperti, autore dello splendido ibrido canarino pastello bruno x ciuffolotto pastello bruno, che si aggiudicò ben 96 punti all’edizione dell’anno 2008 di Fringillia. “Mi chiamo Giancarlo Lamperti e mi diletto in questa nobile arte da tantissimi anni; io credo che come prima cosa, per ottenere questo ibrido, ci voglia una buona affinità di coppia.
La ciuffolotta produce circa 15/20 uova a stagione ma difficilmente sono feconde. Nel mio caso, in oltre 20 anni di tentativi, ho ottenuto un uovo non schiuso ma con l’embrione completamente sviluppato: sarebbe dovuta nascere una femmina satiné; in un caso, sono nati due maschi pastello bruno e nell’altro due ancestrali.
Purtroppo alcuni embrioni, anche se formati, non sono mai nati.
Credo che strategie vere e proprie non ve ne siano: ho usato solo tanta perseveranza e ho avuto tanta fortuna. Dell’ibrido menzionato, ricordo che dalla coppia un anno ottenni due ibridi ma l’anno successivo la stessa ciuffolotta
38 NUMERO 4 - 2024
Soggetto intenso, all.: G. Lamperti, foto: D. Cautillo
uccise il canarino. Tengo a precisare che la coppia non è mai stata divisa neanche in inverno. Da allora, la ciuffolotta non si è più voluta accoppiare con nessun altro maschio. Fortunatamente non ho mai avuto problemi di svezzamento con questi ibridi, in quanto utilizzo balie di canarino abituate ad alimentarsi con pinkies e buffalo, uova sode e piselli mischiati al pastoncino.
Credo che le mutazioni abbiano dato una bella svolta a questo ibrido, nonostante l’ancestrale rimanga sempre bello.
Non ho mai utilizzato il ciuffolotto maschio, però chissà, tentando in un futuro, qualcuno ce la farà. Di quelli ammirati in mostra ricordo con piacere l’ibrido di Lino Clerici tanti anni fa e l’ancestrale di Lino Robustelli. La difficoltà che ho come allevatore, nel mio caso, è la mancanza di spazio, in quanto allevo in gabbie da 120x50”.
Il secondo allevatore che ha deciso di regalarci le sue esperienze è Guido Fusi che da una coppia formata da satiné mosaico x ciuffolotto pastello bruno ottenne ben 4 soggetti: due maschi bruni e due femmine satiné su una quindicina di uova; uno vinse nel Mondiale a Bari nel 2014.
“Mi chiamo Guido Fusi ed allevo ibridi da diversi anni. Credo che questa ibridazione sia di media difficoltà in quanto la ciuffolotta si accoppia facilmente con i maschi di altra specie; l’unico requisito che deve avere il maschio Serinus è che deve essere un buon cantore e non deve essere irruento. Io di solito uso un maschio novello con ottime caratteristiche di tipo, mai accoppiato a canarine, e ciuffolotte anch’esse novelle. Formo la coppia sempre appena finita la muta, in modo che abbiano tempo di conoscersi e di avere un’alimentazione adeguata. La mia coppia era formata da canarino isabella satiné x ciuffolotta pastello bruno: era molto affiatata, con diverse covate annuali con una media di 4/5uova; purtroppo le uova fecondate erano sempre le ultime due e quasi sempre l’embrione non superava il sesto giorno di vita. Così al primo anno. Al secondo anno nel primo nido sono nati due maschi bruni e al terzo una femmina satiné; l’anno dopo è nata una
Credo che le mutazioni abbiano dato una bella svolta a questo ibrido, nonostante l’ancestrale rimanga sempre bello
femmina satiné con una malformazione a una zampa che non ho mai potuto esporre ma di cui ho un video nella mia pagina Facebook. Devo dire che il canarino era un ottimo padre e con la ciuffolotta ha allevato diversi canarini, mentre gli ibridi sono stati allevati da canarine provette, sempre soli o con massimo un soggetto per nido, in quanto erano sempre affamati, nonostante le nostre integrazioni giornaliere. Purtroppo, rispetto alle uova deposte il risultato pare esiguo e penso che ciò sia dovuto alla mutazione satiné del canarino, in quanto gli embrioni non nati sono stati veramente tanti, almeno una decina in tre anni. Penso che una mutazione del canarino meno spinta avrebbe dato risultati diversi. Non ho mai usato il maschio di ciuffolotto per i motivi chiari a
tutti; la possibilità che nasca qualcosa c’è sempre, però lascio ad altri i tentativi. I soggetti che mi hanno colpito sono parecchi: uno su tutti l’intenso di Lamperti e poi, dato che a Lurate Caccivio ero di casa, ho visto crescere quelli di Lino Clerici e di Patrizio Salandi. La nuova mutazione che si presume satiné penso apra nuove strade. La colorazione dei soggetti, per quanto mi riguarda, non è stata per niente semplice; infatti, al primo anno con solo colorante (carotenoidi e poco carophyll) nel pastoncino ha dato risultati scadenti, mentre al secondo anno ho usato carotene idrosolubile nell’acqua e colorante nel pastone e i risultati sono notevolmente migliorati. Spero di essere stato d’aiuto e di aver invogliato qualche lettore a tentare questa avventura”.
Il terzo allevatore ha realizzato, ed esposto, uno dei maschi più belli che io abbia mai visto nella mutazione pastello; si tratta di Pierangelo Saldarini.
“Mi chiamo Pierangelo Saldarini ed allevo uccelli fin dalla giovane età, appassionato da sempre di specie indigene e loro ibridi. Parlando di canarino x ciuffolotto, sicuramente è una delle ibridazioni che più ti prende perché il risultato



NUMERO 4 - 2024 39
Canarino bruno x Ciuffolotto bruno, all. e foto: G. Fusi


è quasi sempre sopra la media, ed anche perché quello con la ciuffolotta è uno degli ibridi più difficili. L’anno scorso ho formato quattro coppie di canarino e ciuffolotta, diciamo mirate, dall’ancestrale al pastello, al bruno e bruno pastello. Risultato: ho buttato nel secchio 86 uova, neanche uno fecondo. Quest’anno riprovo con solo due coppie. Ci vuole tanta fortuna: per esempio, l’anno dell’ibrido pastello ho acquistato un canarino pastello isabella da Lodato a febbraio, quindi non hanno svernato assieme, e ho messo la coppia in voliera esterna 80x160x200. La ciuffolotta aveva due anni e aveva già allevato in purezza. Hanno litigato fino alla prima deposizione che è risultata infeconda. Poi, andando d’amore e d’accordo, ha deposto ancora tre volte, ogni covata con due uova feconde, ma regolarmente nasceva un solo piccolo. Alla fine sono risultati due maschi pastello e una femmina pastello isabella. Non hanno dato nessun problema di svezzamento, come fossero canarini.
A proposito di fortuna, un anno mi era rimasta una femmina non accoppiata in purezza, così decisi di unirla ad un maschio canarino, ma ci vollero ben cinque covate per vedere un uovo fecondo, proprio all’ultima.
Ho avuto uova feconde sia in gabbia da 90 che in voliera da 100/200/200 ma più di qualche volta le uova non si schiudono
Sicuramente le mutazioni sono un valore aggiunto in questa ibridazione. Sinceramente non ho mai provato a usare il maschio di ciuffolotto in ibridazione perché ho sempre sentito dire che non è possibile avere uova feconde usando il maschio... mi sono fidato. Anche col cardinalino sono convinto che sia possibile, ma sono 40 anni che aspetto di vederlo esposto... speriamo. Nel corso degli anni ho visto parecchi ibridi in questa direzione veramente fantastici; tra gli altri, ricordo i due bruno pastello maschi di Lamperti, parecchi anni fa.
Vorrei chiudere con due aneddoti. Agli inizi della mia avventura nel mondo ornitologico, ricordo di essere stato a casa di Pontiggia negli anni Ottanta, dove ammirai un numero veramente importante di cardinalini svezzati: anche lui era un grande appassionato di ibridi ma, mentre io utilizzavo solitamente le femmine di canarino, lui aveva allestito già coppie al limite come cardellino x fringuella, che al mio arrivo era a nido; una volta scesa, il maschio la coprì immediatamente, lasciandomi completamente di gesso. In quegli anni lo stesso Pontiggia realizzò un maschio ibrido, stupendo, di canarino x ciuffolotto.


Il secondo ricordo è di aver visto a casa di Lino Clerici una voliera dove viveva una coppia formata da maschio di ciuffolotto e femmina di verdone che allevavano due ibridi di cardellino x verdone: i due formavano una coppia di balie eccezionali”.
In ultimo, ma solo in ordine di tempo, abbiamo un fresco vincitore di mondiale a Talavera in Spagna con un bellissimo maschio super ossidato: parlo di Felice Rigamonti che ad onor del vero a quel mondiale ha ottenuto ben due medaglie d’oro, con due soggetti esposti, la seconda con uno strepitoso ibrido di cardellino pastello bruno x ciuffolotto pastello bruno.
“Allevo da diversi anni e gli ibridi mi hanno sempre affascinato (ora un po’ meno, in quanto allevo insettivori), in particolare
40 NUMERO 4 - 2024
2 maschi di Canarino bruno x Ciuffolotto bruno, all. e foto: G. Fusi
Femmina satinè, all.: P. Saldarini
l’ibrido canarino x ciuffolotta, per me l’ibrido per eccellenza.
Negli anni ‘80 il primo ibrido maschio che vidi lo realizzò Franco Pontiggia con un canarino satiné rosso mosaico: ne rimasi molto colpito e da allora, quasi tutti gli anni, nel mio allevamento vado a formarne una o due coppie.
Nel corso degli anni qualche ibrido, 7/8 (pochi rispetto ai tentativi), l’ho realizzato e nell’anno 2018 un maschio di canarino x ciuffolotto vinse il mondiale in Olanda.
Nella stagione cove 2022 con una coppia (canarino nero mosaico rosso x ciuffolotta ancestrale) me ne nacquero ben tre, uno per covata, ma per sfortuna ne persi due dopo averli svezzati, il primo per una puntura di zanzara e l’altro si ruppe una zampa impigliandosi con l’anello.
Per fortuna il superstite, a muta ultimata, risultò essere un maschio e negli anni 2023 a Napoli e nel 2024 a Talavera si è aggiudicato il gradino più alto del podio regalandomi due belle medaglie d’oro.
Da quello che ho sperimentato, è una ibridazione molto difficile per le pochissime uova feconde e la fortuna gioca una parte molto importante.
Ho avuto uova feconde sia in gabbia da 90 che in voliera da 100/200/200 ma più di qualche volta le uova non si schiudono. Formo le coppie da inizio febbraio e solitamente non ci sono problemi di convivenza; inoltre, non ho riscontrato nessun problema nello svezzamento dei novelli.
Con l’avvento delle mutazioni si sono potuti apprezzare nuovi ibridi mutati e ricordo molto bene i pastello bruni di Lamperti, i bruni di Fusi e, soprattutto, i pastello di Saldarini.
Nel corso degli anni non ho mai usato il maschio in ibridazione”.
Ringrazio gli amici per questa bella chiacchierata e mi auguro di poter ammirare, nelle stagioni mostre che verranno, qualche soggetto in più di questa bellissima ibridazione.





NUMERO 4 - 2024 41
Pasto unico sì o no?
testo e foto di GAETANO CAMPISI
Subito una doverosa precisazione. Non citerò nessuna azienda e non farò nomi di alcun allevatore: quanto seguirà è frutto di esperienza personale, anche perché tutte le aziende del nostro settore, per fortuna, non producono solo il pasto unico ma anche tanti altri prodotti e quindi la libera scelta spetta solo ed esclusivamente al singolo allevatore. Ultimamente non si sente parlare di altro e

In molti mi hanno parlato di soli vantaggi dovuti a questa nuova tendenza nella conduzione di un allevamento
cioè: pasto unico o alimentazione tradizionale? Bene, dopo aver fatto anche il tentativo di passare al pasto unico, posso affermare che per quanto mi riguarda la risposta è: alimentazione tradizionale. Ma andiamo con ordine. In molti mi hanno parlato di soli vantaggi dovuti a questa nuova tendenza nella conduzione di un allevamento. Personalmente, io di vantaggi non ne ho visti; anzi, l’introduzione del pasto unico ha coinciso con la mia peggiore annata riproduttiva. Certo, direte voi, sarà stata solo una coincidenza, ma il peggio doveva ancora arrivare...
Avevo molte aspettative su quella nuova avventura, attirato dall’ottimizzazione dei tempi, dalla comodità di non dover preparare chissà quale “pozione magica”: niente più pappette, uova, pastoncini, integratori, vitamine etc. etc. Insomma, mi sono attenuto scrupolosamente alle indicazioni dell’azienda rispetto a preparazione, allevamento, muta. Partiamo da quest’ultimo aspetto, cioè la muta. Bene, non mi è servito in quanto, come accennato prima, i soggetti sono stati pochissimi e quindi il mio entusiasmo è andato scemando con la delusione in termini di numeri.
Quindi “preparazione”: poche uova e
chiare, morte embrionale. E già lì avrei dovuto allarmarmi. Però sono molto testardo e ho voluto continuare, tanto ormai avrei considerato l’annata, che si preannunciava disastrosa, come “laboratorio”. Faticosamente cominciano le prime schiuse e ovviamente passo al cambio prodotto e si somministra la versione “allevamento”. Disastro. Crescita lenta, che nella maggior parte dei casi si trasformava in morte; soggetti

42 NUMERO 4 - 2024 ALIMENTAZIONE
Scagliola e misto semi
Semi germinati e pastoncino secco
debilitati, tant’è che perdevo pure gli anellini; genitori poco propensi all’imbecco ed erano costantemente forastici, poco agili ed assolutamente annoiati. Nel mese di luglio, quando ormai tutto era perso, somministro dei semi di sesamo che normalmente uso per le panature in cucina e mi accorgo che non aspettano altro. Di colpo ho ricominciato ad assistere alla lite per la conquista del cibo, cosa che non vedevo dal mese di gennaio; non solo: somministro in contemporanea del cous-cous secco e l’allevamento riprende vita. Ormai la stagione è inevitabilmente compromessa e tutte le scorte che mi ero fatto sono rimaste lì in attesa di donarle a qualche amico allevatore. Certe volte avevo anche l’impressione che fosse rischioso quel metodo alimentare in quanto, se non controllavi costantemente le mangiatoie, i canarini avrebbero potuto svuotarle da un momento all’altro, non avendo altro da man-
giare… il rischio era altissimo. Però adesso è giunto il momento di trarre le conclusioni. Vantaggi inesistenti rispetto all’acquisto di semi, pastoncino e integratori. L’unico vantaggio che ho riscontrato è stato quello di non dover spazzare tutte le mattine il pavimento, oltre ad un gradevole odore in allevamento. Secondo il mio modesto parere, chiamarlo “pasto unico” porta a far credere che non occorra altro, quando invece è giusto far notare, non fosse altro per serietà, che da solo il prodotto in questione non è sufficiente, in partico-
Secondo il mio modesto parere, chiamarlo “pasto unico” porta a far credere che non occorra altro
lare nella fase più delicata che è quella della crescita dei piccoli. Quindi, prodotto che va usato contestualmente a quelli tradizionali, magari alternato al pastoncino. Mi piacerebbe tantissimo se da chi avrà la pazienza di leggere questo mio scritto scaturisse un dibattito costruttivo per scambiarci le nostre opinioni, anche perché dietro un prodotto finito c’è studio, ricerca, lavoro e la serietà delle aziende.
Adesso sono tornato alla cara vecchia e sicura scagliola, pastoncino secco e morbido, semi germinati (vedi foto) e finalmente mi è anche tornato il sorriso. Insomma, come spesso dico, se vinci: bene, se perdi: impari. Alla fine la morale è che gli uccellini sono granivori e quindi devono assumere alimenti che gli permettano di sgranare, di sbucciare...
Ah, dimenticavo: sono anche tornato con la cara vecchia scopa a spazzare il pavimento.




NUMERO 4 - 2024 43




•Invitiamo tutti gli allevatori a inviare foto di soggetti provenienti dai propri allevamenti, con descrizione della specie, razza e mutazione, all’indirizzo: redazione@foi.it
•All’autore della foto mensilmente prescelta da un comitato interno, verrà offerto in omaggio un libro edito dalla FOI in base alla preferenza e alla disponibilità.
(*)Tutte le foto inviate, anche quelle non pubblicate, rimarranno a disposizione della FOI a titolo gratuito e potranno essere utilizzate, senza alcun limite o vincolo temporale, per pubblicazioni, iniziative e scopi promozionali della Federazione
Questo mese, il protagonista di Photo Show è:
MATTEO CONTE R.N.A. 2BAT con la fotografia che ritrae il soggetto:
“Verdone maschio classico” (Chloris chloris) Complimenti dalla Redazione!



L’Associazione Ornitologica Vesuviana ha compiuto 50 anni
testo e foto di RAFFAELE CECERE
La Campania ornitologica è in festa. Doppio cinquantenario, insieme al Raggruppamento Regione Campania di cui è l’ispiratrice.
Congratulazioni all’Associazione Ornitologica Vesuviana per il suo cinquantesimo anniversario! Quante associazioni possono vantare questo primato?
È stata fondata nel 1973 da un gruppo di appassionati di uccelli. Instancabilmente si è dedicata alla promozione del loro allevamento, alla loro conservazione, alla loro protezione, alla diffusione della cultura ornitologica e alla promozione dell’aspetto sportivo di tale attività.
I festeggiamenti
I soci hanno festeggiato con un cortometraggio celebrativo e in libera distribuzione, visualizzabile e scaricabile dalla pagina Facebook dell’associazione, con una torta di cinque chili (un chilo per ogni decade), con un gadget per ricordare l’evento. Grazie a I.O., aggiungiamo questo articolo che porta le lancette dell’orologio indietro al 1973; per celebrare la nascita e il traguardo, per ricordare come è nata, il cammino, i successi, le soddisfazioni, ma anche tramandare la memoria storica a chi non ricorda, a chi non c’era, a chi non era ancora nato. Mi sono assunto io l’incarico di ricordare, poiché, per varie coincidenze, sono venuto in possesso di alcuni scritti che parlano della genesi di questa Associazione e del RR Campania, aggiungendo le lun-

ghe chiacchierate serali davanti a un buon caffè con il socio fondatore, il ragionier Enrico Palescandolo; ed ecco l’articolo, ricco di informazioni che nemmeno i soci conoscono o ricordano. De-
scriverò l’associazione parlando della sua storia, la cui nascita è un evento singolare; descriverò anche la nascita del Raggruppamento Regionale Campano, la cui storia è parallela alla storia dell’AOV, unica ispiratrice della sua nascita e crescita.
Lo stato dell’allevamento degli uccelli in Campania agli inizi degli anni ‘70 Nonostante la regione avesse da sempre una vocazione nell’allevamento degli uccelli, i tanti allevatori non erano coordinati, non erano associati, non conoscevano la FOI, non inanellavano gli uccelli e, quindi, non partecipavano alle mostre; eppure, nelle gabbie, avevano veri e propri gioielli. Le associazioni erano tre: l’Associazione Ornitologica Campana, l’Associazione Ornitologica Stabiese, l’Associazione Ornitologica Salernitana, lasciate sole per libero scio-

NUMERO 4 - 2024 45 CRONACA
Il logo dell’Associazione Ornitologica Vesuviana
Il banner realizzato in occasione dell’evento con i loghi dell’Associazione e della Federazione


glimento dall’Associazione Canaricoltori e Ornicoltori Frattesi, quest’ultima con sede in Frattamaggiore.
Le nuove regole della F.O.I.
Negli anni 1973-74 il nuovo Statuto della FOI prevedeva i Raggruppamenti Regionali e Interregionali tra le associazioni ornitologiche già esistenti. Le regioni potevano avere il raggruppamento regionale se esistevano almeno cinque associazioni, con un numero complessivo minimo di 400 soci. Se tali caratteristiche non fossero state soddisfatte, la FOI avrebbe dato la possibilità alle associazioni di regioni confinanti di potersi associare e, in questo caso, di formare il Raggruppamento Interregionale.
Primo obiettivo: la costituzione di un’associazione
Queste nuove regole furono illuminanti per alcuni allevatori; essi si resero conto che per far nascere il Raggruppamento Campano FOI era necessaria la costituzione di nuove associazioni ornitologiche che avrebbero colmato un secondo problema: la mancata diffusione dell’ornitofilia sportiva in alcune zone della regione, che non avevano nelle vicinanze un’associazione FOI.
Senza perdere tempo, animati dalla passione per gli uccelli, con atto per notaio Di Liegro, in data 27 marzo 1973, nella città di Torre Annunziata (NA), vide la luce l’Associazione Ornitologica Vesuviana, che fu affiliata alla FOI entro breve tempo.
Gli allevatori erano tutti dell’area vesu-
viana, dunque la dicitura Associazione Ornitologica e, poi, l’aggettivo Vesuviana erano scontati. Più tardi, in termini più colloquiali, si usò abbreviare con “la Vesuviana” per indicare l’associazione e “i vesuviani” per indicare i soci. Attualmente è l’unica associazione ad avere nel nome un riferimento al Vesuvio.
Il Primo Consiglio Direttivo dell’AOV fu così costituito:
Presidente:sig. Emilio Smaldone
Vicepresidente:sig. Antonio Auriemma
Vicepresidente: sig. Francesco Mangiala
Segretario: rag. Enrico Palescandolo
Consiglieri:il dott. Vincenzo Casella, l’ing. Arcangelo Balzano ed il Cav. Pellegrino Rizzo.
Tra i soci fondatori vi furono, altresì, i signori Francesco Ilardi, Antonio Fiorenza, Antonio Langella, Catello Lubrino ed il Reverendo Alfonso Cirillo.
L’entusiasmo fu l’autore del logo, definito da esperti di grafica un piccolo gioiello di tipografia artigianale (la grafica computerizzata era ancora lontana): una corona circolare con dentro il nome dell’associazione e al centro il golfo di Napoli con il Castel dell’Ovo, il Vesuvio sbuffante, un canarino bianco e uno Scotch fancy.
Secondo obiettivo: la costituzione del Raggruppamento Campano della F.O.I.
Nel 1974, con una robusta spinta dell’AOV, nacquero e vennero subito affi-
liate alla FOI l’Associazione Ornitologica Battipagliese e l’Associazione Ornitologica Forestale (delibera FOI del 14/12/1974). A quel punto c’erano tutti i numeri per chiedere alla FOI il riconoscimento delle associazioni campane come raggruppamento. In breve tempo la FOI riconobbe il Raggruppamento Campano delle Associazioni Ornitologiche. Il secondo obiettivo fu raggiunto rapidamente grazie alla perseveranza dei Soci dell’AOV. Il primo Presidente eletto fu il dott. Giacomo Ciampa che volle come segretario il rag. Enrico Palescandolo, il quale ricordava che i primi passi furono alquanto incerti. Era complicato gestire un nuovo assetto associativo senza un supporto tecnico e logistico, a cui si aggiungeva la distanza tra la Campania e la sede della FOI, allora situata a Brescia. Tale difficoltà fu anche degli altri raggruppamenti d’Italia (10 in tutto tra R.R. e R.I.). La FOI nominò un Coordinatore dei R.R. e R.I. che facilitò il compito ai raggruppamenti. I responsabili diventarono con il tempo dei veri esperti, al punto che furono chiamati a far parte del Direttivo nazionale. Il rag. Enrico Palescandolo fu eletto a larga maggioranza nel Consiglio direttivo della FOI, dove rivestì la carica di coordinatore dei R.R. e dei R.I. dell’Italia centro-meridionale ed insulare. Palescandolo rivestì, altresì, la carica di Vicepresidente della FOI per oltre un decennio subentrando all’ing. Giovanni Chillè di Messina.
I meriti dell’AOV a favore del R.R. della Campania? Tanti! Tutti hanno contri-
46 NUMERO 4 - 2024
Gruppo Soci A.O.V.
buito alla crescita del Raggruppamento Campano, chi in un modo, chi in un altro. Oltre alla creazione del Raggruppamento Campano, il merito di aver portato il Raggruppamento Campano a competere con i R.R. d’Italia più grandi, di aver portato l’ornitofilia là dove era sconosciuta, di averne diffuso l’aspetto sportivo.
E fu così che la FOI, in Rimini, in data 17 aprile 2004 per opera del suo Presidente geom. Salvatore Cirmi, in occasione dell’Assemblea annuale delle Associazioni Ornitologiche, encomiò il Raggruppamento Campano per lo sviluppo associativo promosso nella regione Campania, con chiaro riferimento al ruolo decisivo dei soci dell’AOV (nell’occasione Presidente del R.R.C. era sempre il rag. Enrico Palescandolo); lo stesso definì il R.R. Campania “una piccola FOI”. Chi partecipò all’assemblea ricorda che, subito dopo, partì un applauso interminabile che sottolineava la stima e la riconoscenza che tutta l’assemblea aveva nei confronti della AOV e del suo Direttivo.
Attualmente il Raggruppamento Campano FOI è costituito da 35 associazioni, distribuite sul territorio in modo piuttosto omogeneo, che dialogano tra loro in modo costruttivo e non competitivo.
Hanno organizzato il Campionato Italiano 2009 e 2016 in un centro fieristico a Ercolano, sul pendio del vulcano più conosciuto al mondo: il Vesuvio. Dopo, il Campionato Mondiale 2023 nella Mostra d’Oltremare a Napoli, facilmente raggiungibile con qualsiasi mezzo di trasporto, anche con l’alta velocità e l’aereo; infatti, molti allevatori stranieri hanno preferito proprio questi mezzi di trasporto. In entrambi i casi, gli ambienti erano luminosi e accoglienti, sia per gli uccelli sia per i visitatori.
Sono state organizzate mostre che sono diventate con il tempo appuntamenti fissi per gli allevatori, come l’Internazionale di Salerno, il Campionato Regionale e tante altre mostre più piccole, ma non per questo meno importanti.
Gli allevatori AOV
Tra le file ci sono stati allevatori che hanno selezionato il Gibber Italicus nella forma e posizione che conosciamo oggi e che hanno scritto la grammatica per

la gestione del suo allevamento, dando alla Campania il primato della competenza e della qualità in questa razza. Oggi, un folto gruppo di esperti allevatori porta nelle tasche dell’associazione, ogni anno, un ricco medagliere. Da noi e con noi si alleva di tutto e con competenza: esotici, psittacidi, indigeni e tutto ciò che ha due ali. Alcuni allevamenti sono dedicati al cardellino, a Napoli O cardill’. Un delizioso uccellino, dal canto unico, che da sempre è stato allevato dai napoletani. A questo uccellino i napoletani hanno dedicato canzoni, pitture, modi di dire e un posto nel presepe. Non è esagerato dire che esso faccia parte della cultura napoletana da secoli e da molti è considerato uno di famiglia, con un regolare nome. È bello incontrare gli allevatori che si dedicano a loro, puoi vedere tutta la maestria antica fusa con il moderno. Alcuni soci rivestono l’incarico di giudice e spendono la propria competenza anche a favore dei soci che necessitano di consigli per la selezione degli uccelli.
Tra i soci, vivono umilmente nella penombra, ma fanno sentire la loro competenza, alcuni esperti di specie rare, o di specie insolite per essere allevate in gabbia; non mi permetto di chiamarli “allevatori” perché sono ornitologi allo stato puro, fanno cose particolari, non

partecipano alle mostre ed hanno ragione, essi si confrontano solo con loro stessi. In quest’ultimo gruppo, posso dire che c’è qualche socio che presta consulenza ad alcuni produttori di mangimi, per i quali testa anche i prodotti prima della commercializzazione. Sempre in questo gruppo c’è un ornitologo che è il protagonista di una storia toccante che sarà raccontata più avanti.
L’aspetto umano dei soci Nel gruppo AOV è stato promosso l’aspetto umano degli allevatori, che si affidano l’un l’altro per risolvere i problemi che un allevamento pone giorno per giorno all’allevatore. È stata realizzata una premiazione interna che gratifica gli allevatori che hanno conseguito successi alle mostre. Non ultimo, l’AOV ha diffuso la filosofia che l’associazione non è altro che un fine unico a cui tutti devono contribuire; tradotto in termini pratici, tutti sono felici per i risultati che i soci raggiungono, come se fossero i propri.
L’attività di protezione e conservazione dell’ambiente e della fauna, ovvero la storia di Catello e del gabbiano L’associazione e il raggruppamento volevano far sentire il proprio peso anche nella protezione dell’ambiente. Così una task force AOV capitanata sempre da lui, il rag. Enrico Palescandolo, mosse verso i circoli didattici 1°, 3° e 4° e verso la Scuola Media Parini di Torre Annunziata. Il progetto si chiamava “UN
MONDO DI ALI – DIFENDIAMO LA NATURA”, patrocinato dal comune di Torre Annunziata. Gli incontri furono numerosi e la curiosità fu non solo degli alunni, ma anche del corpo docente. Fu descritta la fauna da proteggere e da salvare, focalizzando l’attenzione sulla fauna del Vesuvio, disegnando uno scenario floro-faunistico ben più ampio delle sole ginestre di leopardiana memoria.
Un socio dell’AOV mi raccontò una storia legata al progetto di cui sopra, che merita di essere riportata. I protagonisti sono un ragazzo liceale di nome Catello, all’epoca del progetto era un bambino della scuola elementare che conservò l’amicizia con Enrico, e un socio, che Pa-
NUMERO 4 - 2024 47
La torta preparata per i festeggiamenti
Gadget consegnato ai partecipanti
lescandolo chiamava “l’uomo che parla con gli uccelli”. Mi faccio da parte, sarà proprio quest’ultimo a raccontare la storia.
«Una sera d’autunno, io e il rag. Enrico Palescandolo eravamo seduti al bar davanti al nostro caffè; passò un ragazzo del liceo di nome Catello e lui lo invitò a sedersi con noi. Enrico mi presentò dicendo che ero il socio AOV che parlava con gli uccelli e rivolgendosi a me disse che un gabbiano si posava dove abitava Catello, all’ultimo piano del palazzo, ma non si lasciava avvicinare. Gli spiegai che gli uccelli hanno un loro idioma fatto di versi, sguardi, movimenti, apparentemente non dicono nulla, ma sono veri e propri discorsi. Come primo passo bisognava conquistare la sua fiducia offrendogli del cibo. Mi resi disponibile per questa impresa, a patto che procurasse del pesce.
Io ero entusiasta del mio allievo e il mio allievo era entusiasta di conoscere il linguaggio dei gabbiani. Ci volle tanta pazienza, ma alla fine il gabbiano prese il pesce dalle mani di Catello; erano diventati amici!
Dopo una decina d’anni viaggiavo sul treno della Circumvesuviana, un giovane mi chiamò e mi venne incontro: era Catello, lo invitai a sedere con me e gli domandai subito del gabbiano. Mi disse che non solo entrava in casa, ma saltava sulla scrivania e là gli faceva compagnia. Dopo diversi anni, smise di venire. Il volto di Catello diventò triste.
“Gli animali non tradiscono l’amicizia,” dissi io e poi chiesi “Le ultime volte che l’hai visto ti sembrava stanco, indifferente, non gli importava più di guardare intorno a sé?”
Catello rispose di botto: “Sì, era proprio così!”
Mi feci coraggio: “Il tuo gabbiano non ti ha lasciato, ma ci ha lasciato”. Al volto triste si aggiunsero gli occhi lucidi, fu proprio in quel momento che capii l’affetto che c’era fra loro due. “Tutto passa, restano solo i ricordi che il tempo fa più belli”. A questo punto una domanda scontata: “Perché fare amicizia con un gabbiano?” Catello prendeva tempo e sorrideva: “Non so, stava là, ci guardavamo; avere un amico che veniva volando era insolito e poi mi chiedevo se certe cose succedono solo nei film o anche nella vita reale. All’inizio è stato difficile da credere, ma poi ho dato ragione ad Enrico quando diceva che parlavi con gli uccelli; grazie a te ho parlato con un gabbiano”.
Ed io: “Hai parlato con il gabbiano, cosa ti ha detto?”
Catello: “Mi ha insegnato che la pioggia non bagna ed è bella come il sole; mi ha insegnato a vivere nel vento, mi ha insegnato ad ascoltare il silenzio… già, il silenzio, ha una poesia che nemmeno il mare d’inverno riesce a dare”.
Io: “Hai imparato a parlare con i gabbiani, hai imparato a guardare la natura con gli occhi giusti: fai di questo una religione e professa questa fede al mondo; non è difficile, gli uomini hanno nel cuore l’amore, non la guerra. Se pensi che ti ho insegnato qualcosa, questa è l’ultima lezione”.
Mentre mi raccontava della sua vita e della laurea in giurisprudenza, arrivammo alla stazione di Torre Annunziata.
Ci stringemmo la mano e contemporaneamente disse: “Salutami Enrico”.
Ed io: “L’hai già fatto, Enrico e il gabbiano ci guardano dall’alto”. Egli ridiventò triste, io gli strinsi la mano più forte e, guardan-

dolo negli occhi, gli feci un sorriso. Lui, fra sé, appena sussurrò: “Tutto passa, restano solo i ricordi che il tempo fa più belli”.
Scese alla stazione, venne vicino al finestrino, continuava a salutarmi con la mano, e quando il treno partì, rincorrendomi, mi volle salutare ancora una volta così: “Ciao, uomo che parla con gli uccelli”. Io non potevo che rispondere: “Ciao amico dei gabbiani”.
Il viaggio verso Sorrento continuò in religioso silenzio. Ricordi, fatti, persone, emozioni si affollarono dentro me. I soci: quanta strada hanno camminato e quanto hanno seminato! Senza risparmiarsi hanno dato all’Associazione Ornitologica Vesuviana tante facce e tanti motivi per esistere. Arrivati a Sorrento: “Hai finito di pensare?” disse la mia compagna e mi destai dal torpore.
Raggiungemmo il belvedere di Sorrento; seduti sulla panchina a guardare il panorama, un gabbiano si posò sul colonnino del parapetto a 2 metri da noi: io e lei ci guardammo negli occhi, uno sguardo che fu più eloquente delle parole, un sorriso e poi via».
Questa è la storia dell’Associazione Ornitologica Vesuviana, questa è la storia del RR Campania, questa è la storia di un gruppo di allevatori che tutti chiamano “i Vesuviani”; una storia unica che meritava di essere raccontata, una storia nata cinquant’anni fa in uno studio di un notaio e poi costruita giorno dopo giorno con tanto lavoro e dedizione. Alcuni soci fondatori, purtroppo, ci hanno lasciato pochi anni fa: tra loro, il rag. Enrico Palescandolo. Ci hanno donato un’eredità preziosa, a loro formuliamo una promessa: continuare la loro opera. Per questa ricca eredità diamo loro, con inchino, un commosso grazie. Il testimone è stato raccolto da un gruppo di allevatori che portano avanti il progetto AOV con lo stesso entusiasmo che ci hanno insegnato loro.
Cinquant’anni: quanta storia, quanti ricordi, tante cose fatte e tante da fare. Non guardiamo mai al tramonto, anzi, non siamo arrivati nemmeno a mezzogiorno, per noi è sempre un’alba radiosa. A tutti Voi che ci avete dato il nome che portiamo, in futuro continuate pure a chiamarci così, noi non nascondiamo l’orgoglio di essere “i Vesuviani”.
48 NUMERO 4 - 2024
Belvedere di Sorrento al tramonto. Un gabbiano riposa sul parapetto
Lo Storno rosa (Pastor roseus)
testo e foto di GIOVANNI MUSUMECI



Ha una taglia di circa 19-22 cm; gli adulti sono caratterizzati da un cappuccio nero molto esteso sino alle spalle, remiganti e timoniere nere con riflessi metallici. Nei maschi adulti è inconfondibile un ciuffo di penne lunghe che, quando il soggetto lo mette in evidenza durante il corteggiamento, forma una specie

di caschetto. Il resto del corpo, cioè petto e ventre, è rosa. Ad un attento

esame visivo si nota un leggero dimorfismo tra i sessi. Questo è il tipico piumaggio dei soggetti adulti, i giovani sono grigio bruni con remiganti e timoniere brunastre.
L’areale di distribuzione va dalla Turchia all’Asia minore e verso nord sino agli Urali. In Europa si hanno delle invasioni periodiche e le ultime han-

NUMERO 4 - 2024 49 ESTRILDIDI FRINGILLIDI IBRIDI
Piccoli accanto alle uova schiuse
Piccoli appena nati
Pullo di 3 giorni
Pullo di 8/10 giorni
Giovane
Maschio in abito nuziale

no toccato anche l’Italia, nel 20182020.
L’alimentazione in natura consta di diversi tipi di insetti, ma la predilezione va alle cavallette ed ai grilli; infatti, la stagione cove inizia con l’aumento della disponibilità delle cavallette. Negli altri periodi lo storno rosa non disdegna la frutta o anche semi.
Vive in grandi colonie, prevalentemente nelle steppe dove è più facile reperire l’alimento preferito, le cavallette, e nel periodo di riposo dalle cove va per zone boscose.
La stagione riproduttiva è influenzata dall’andamento climatico e dalla presenza delle cavallette, e nel momento
in cui le condizioni ottimali si verificano costruisce un nido frettolosamente in buchi su pareti rocciose o di alberi e qui depone 4/6 uova azzurrognole che cova per 11/12 giorni. I siti di nidificazione sono ubicati in prossimità di fonti di acqua, mentre per la ricerca del cibo arrivano a percorrere diversi chilometri. Effettuano una sola covata ed alla fine della stagione adulti e giovani si riuniscono alla ricerca del cibo.
È un uccello gregario, sia nel periodo delle cove che di riposo, durante il quale possono stazionare insieme molti esemplari.
Da due anni sono in possesso di tre coppie di questi uccelli, sistemati in una voliera di 100x250xh200 cm con diversi nidi a disposizione e già lo scorso anno si sono riprodotti.
Da quanto mi è stato possibile notare, nella piccola colonia si sono formate le coppie ma solo una, con il maschio dominante, si è riprodotta. Le altre, malgrado avessero costruito un nido, non hanno deposto. Così è stato anche quest’anno.
Questo non succede solo in voliera, ma ho potuto notarlo anche nelle piccole colonie di passeriformi all’esterno.
L’alimentazione che fornisco durante tutto l’anno è costituita da un pastone per insettivori del commercio integrato, saltuariamente, da frutta e insetti congelati. All’avvicinarsi della stagione cove aumento la quantità degli insetti.
Il misto degli insetti è formato da bigattini, tarme, grilli ed hermetia; a volte aggiungo delle camole della cera. Prima della cova vengono letteral-

I siti di nidificazione sono ubicati in prossimità di fonti di acqua, mentre per la ricerca del cibo arrivano a percorrere diversi chilometri
mente divorati e non resta nulla, ma al momento che nascono i pulli cominciano i problemi.
I primi ad essere imbeccati sono le tarme della farina e, se sostituiti da altre tarme, gli altri insetti rimangono nella mangiatoia o se ne cibano gli altri “ospiti” della voliera.
Questo è l’inizio dei nostri problemi, con pulli che stentano a crescere o con problemi alle ossa lunghe, quindi zampe storte, fratture ecc.
Negli anni, allevando insettivori, ho cercato di prevenire questo problema che ci penalizza molto. Ho utilizzato l’osso di seppia grattugiato sugli insetti e devo dire che dà una mano, ma l’uso di prodotti con vitamine e aminoacidi ha dato risultati migliori.
Per alcuni insettivori bisogna integrare il misto degli insetti con lombrichi.
Una piccola osservazione personale: in commercio vi sono tanti prodotti che dovrebbero sostituire gli insetti, ma vi posso assicurare, avendoli provati, che gli adulti li gradiscono ma non li usano per imbeccare i piccoli.

50 NUMERO 4 - 2024
Giovane
Femmina adulta (a sinistra) e maschio Femmina (a sinistra) con due mschi
Centocchio (Stellaria media)
Stellina selvatica dagli svariati utilizzi
testo di PIERLUIGI MENGACCI, foto P. MENGACCI, WIKIMEDIA.ORG, WWW.ACQUAPLANTARUM.COM, WWW.IN-VALGRANDE.IT,
Il centocchio comune (Stellaria media) è una piccola pianta biennale (alta fino a 40 cm, sottospecie del genere Stellaria), diffusa in tutto il mondo e comunissima su tutto il territorio italiano, appartenente alla famiglia delle Caryophyllaceae, genere Stellaria. Il nome del genere (Stellaria), di origine latina, fa riferimento alla forma stellata del fiore, mentre il nome della specie, media, si riferisce alla taglia intermedia rispetto a quella di altre specie dello stesso genere.
Delle170 specie che comprende il genere Stellaria, le più comuni e di maggiore interesse presenti in Italia che hanno tutte lo stesso utilizzo, sono (da Wikipedia)
-Stellaria media L. (chiamata volgarmente “erba paperina” o “gallinella”)
Dal libretto dei miei appunti orto-ornitofili e non solo
- Stellaria nemorum,L. - Centocchio dei boschi: in Italia è la specie, di questo genere, più alta (raggiunge i 60 cm); i fiori sono in posizione ascellare e si piegano dopo la fioritura; è presente in tutta l’Italia a quote elevate (1000 – 2100 s.l.m.).
-Stellaria negletta, Weihe - Centocchio a fiori grandi: sia i sepali che i petali sono lunghi fino a 6,5 mm. Si trova su tutta la penisola (isole comprese).

- Stellariacupaniana (Jordan & Fourr.), Nyman - Centocchio comune: presenta un fusto completamente pubescente su tutta la superficie. È una specie presente solo al sud, da non confondere con la Stellaria media
Il nome del genere (Stellaria) fa riferimento alla forma stellata del fiore, mentre il nome della specie, media, si riferisce alla taglia intermedia rispetto a quella di altre specie

NUMERO 4 - 2024 51 ALIMENTAZIONE
WWW.WEST-CRETE.COM
Centocchio Comune (Stellaria media), foto: P.Mengacci
Stellaria media, fonte: wikipedia.org



-Stellaria graminea L. - Centocchio gramignola: i sepali sono glauchi; il fusto e le foglie sono lisci. Si trova al centro-nord dai 1000 ai 2000 s.l.m.
Il Centocchio comune(Stellaria media) è una di quelle erbe selvatiche considerate “primizia primaverile”, ma è possibile trovarla fresca ed abbondante anche durante l’autunno-inverno; scompare però nei periodi maggiormente caldi e siccitosi. Data la sua natura, atta a moltiplicarsi velocemente e ad avvinghiarsi alle altre piante, è anche considerata un’erba infestante ed è presente nel mio giardino, dove invade perfino le aiuole ed i vasi dei fiori.
Predilige terreni freschi ed azotati e lo troviamo nei vigneti, negli orti, ai margini dei giardini e dei terreni coltivati e lungo i fossi. Generalmente assume una forma leggermente cespugliosa e strisciante, con piccole foglie ovali terminanti a punta di color verde chiaro lucide, molto vicine al gambo e con piccoli e caratteristici fiorellini bianchi in cima agli steli, a cinque petali (in realtà sembra averne dieci perché ognuno si divide in due) che lo rendono inconfondibile. Caratteristico è il gambo della pianta: un tubicino leggermente peloso ed elastico che, tirandolo lentamente con le mani per strapparlo, si rompe nella parte esterna del fusto ma all’interno rimane attaccato un filamento biancastro molto resistente (detto fascio vascolare). Altra caratteristica è quella dei petali: si aprono solo in pieno sole mentre restano serrati la sera e nelle giornate fredde e ombrose.
Ogni pianta produce in un anno circa 3000semini bruni piccolissimi che hanno un’alta capacità di germinazione e sono molto longevi; in caso di condizioni climatiche avverse, infatti, possono permanere nel terreno in uno stato di quiescenza fino a 50 anni ed oltre.
Secondo me, riconoscere il Centocchio è molto facile, però bisogna stare molto attenti a non confonderlo con il cosiddetto Centocchio dei campi o mordigallina, pianta del tutto simile, botanicamente nota come Anagallis arvensis, che viene considerata da molti esperti una pianta tossica.
Riporto schematicamente le differenze fra le due piante, evidenti anche nelle foto allegate:
-Il Centocchio (Stellaria media) ha il fusto a sezione circolare; i fiori sono bianchi; non presenta alcun segno nella pagina inferiore delle foglie.
-La mordigallina oanche Centocchio dei campi (Anagallis arvensis) ha fusto a sezione quadrangolare e fiori rosso-arancio o blu-celesti; a volte può presentare degli screzi o puntini violacei nella pagina inferiore delle foglie.
Proprietà del Centocchio ed utilizzi La saggistica che ho consultato attribuisce al Centocchio la presenza in percentuali variabili di minerali quali ferro, calcio, potassio, selenio, ma-
I semi sono inoltre ricchi di proteine e acidi grassi e sono di aiuto a disintossicare e purificare reni e fegato
gnesio, cromo, silicio, zolfo, zinco, manganese, cobalto e fosforo, vitamine A, C, D, E, B1, B2, B3 e beta-carotene. Inoltre, è stata rilevata la presenza in grande quantità di saponine e tannini, composti tuttora controversi, dal potere diuretico, espettorante, tonificante, antinfiammatorio e antibatterico. I semi sono inoltre ricchi di proteine e acidi grassi e sono di aiuto a disintossicare e purificare reni e fegato.
L’uso fitoterapico del Centocchio Storicamente non si trovano riferimenti medicinali e culinari del centocchio fino al Medioevo, dove comparve anche sulla tavola, ma non se ne apprezzavano le proprietà terapeutiche, fin quando un abate tedesco nel XIX secolo, Sebastian Kneipp,
52 NUMERO 4 - 2024
Stellaria media, fonte: www.actaplantarum.org, autore: Carlo Cibei
Stellaria dei boschi (S.nemorum), fonte: www.in-valgrande.it
Stellaria media, fonte: www.actaplantarum.org,
famoso fitoterapista bavarese, ne apprezza e ne vanta le proprietà calmanti delle irritazioni delle vie respiratorie. Da qui ne deriva anche la denominazione erba polmonaria. Il Centocchio in passato era utilizzato solo per curare le infezioni cutanee. Con il tempo, grazie ai numerosi studi effettuati, il Centocchio è stato definito un concentrato di sostanze che permettono di mantenere in ottima salute l’organismo e proteggerlo dai radicali liberi. Nelle fitofarmacie si trovano preparati in pillole, creme, lozioni e tinture che contengono gli estratti della pianta. Vengono consigliati nei casi di infiammazioni, eczemi, psoriasi, reumatismi, dolori articolari, varici ed emorroidi. Sono altresì consigliati per accelerare la cicatrizzazione di piccole ferite, forun-
L’uso alimentare del Centocchio va dosato con prudenza, data la presenza di saponine ed ossalati
coli e tumefazioni e anche per contrastare la ritenzione idrica e contro la stitichezza e disturbi legati alla digestione e per favorire il transito intestinale.
Infine, ho trovato molto interessante la dicitura in una confezione di semi di Centocchio comune posta in vendita da una nota ditta: “… Contiene Vitamine C, D, B6, B12 e sali minerali: magnesio, ferro, calcio, potassio, zinco, fosforo, manganese, sodio, rame e silicio. È un alimento rinfrescante, sfiammante e decongestionante. È un antico rimedio in caso di artrite o anemia alimentare. Ha proprietà diuretiche ed aiuta a disintossicare e purificare i reni e fegato”.
A mio avviso, per l’utilizzo a scopo medicinale è sempre bene attenersi ai consigli del proprio medico.
L’uso alimentare del Centocchio Va dosato con prudenza, data la presenza di saponine ed ossalati che, in dosi molto elevate, sono irritanti e tendono a sovraccaricare i reni, specialmente se si soffre di reflusso o di calcoli. Ad ogni modo le cime tenere sono deliziose in misticanze di insalata, oppure possiamo farne una salsa, dal sapore di cetriolo, per tartine, per spalmarla su formaggi teneri o per unirla alle patate lessate con aglio e prezzemolo. Anche un pesto con le mandorle è un ottimo condimento per gnocchi di patate, a cui mia moglie aggiunge anche la salsiccia. Inoltre, si può preparare una bevanda rinfrescante, estraendone il succo: ecco una ricetta tratta da “Il Prato è in tavola”*: “Estrarre il succo da un mazzetto di centocchio, 3 foglie di basilico e due di menta, una mela verde ed uno spicchio sottile di limone, poi miscelare con del ghiaccio tritato e del latte di riso. Per farne un cocktail, unitevi un cucchiaio di zucchero di canna e 60 ml di vodka”. Va comunque ricordato di raccogliere quest’erba quando è ancora giovane, così sarà più tenera, più digeribile e con una quantità minore di saponine.
L’uso in campo ornitologico del Centocchio
A mio avviso, da che mondo è mondo è sempre esistito. Animali da cortile come galline, anatre, oche, colombi ne sono ghiotti, così come è appetito da tutti gli uccelli silvani e da quelli di allevamento dove i giovani getti con foglie e capolini con fiori e semini sono consumati con avidità. Riferimenti al Centocchio comune nell’alimentazione dei canarini sono presenti anche nel testo “Canaricoltura”, dove il Prof. Zingoni, che ne sconsiglia però l’utilizzo nel periodo riproduttivo, scrive “… le coppie abituate ad allevare la prole con i semi di centocchio (ciò vale anche per altre piante ugualmente gradite) ...entrano in crisi se d’un tratto questi semi dovessero mancare”. Secondo la mia modesta esperienza, forse, con una turnazione di diverse erbe selvatiche stagionali il fenomeno non dovrebbe verificarsi. Inoltre, è risaputo che molti allevatori



NUMERO 4 - 2024 53
Stellaria neglecta, fonte: wikipedia.org
Stellariacupaniana: fonte: www.west-crete.com
Stellariagraminea, fonte: www.actaplantarum.org




Durante le cove, invece, stimola l’imbeccata e favorisce il buon funzionamento dell’apparato intestinale dei piccoli grazie alle sue proprietà toniche e diuretiche
di canarini e silvani considerino il Centocchio un alimento afrodisiaco che aiuta le femmine nell’estro riproduttivo.

Durante le cove, invece, stimola l’imbeccata e favorisce il buon funzionamento dell’apparato intestinale dei piccoli grazie alle sue proprietà toniche e diuretiche. Una nota negativa che ho riscontrato è la presenza di soffusioni alari aranciate nei novelli di canarini di colore a fattore rosso alimentati con il Centocchio durante il periodo implume, dovuta, a mio avviso, al beta-carotene che compare fra i nutrienti di questa pianta erbacea. Una nota positiva invece l’ho notata quando una mia femmina isabella, molto svogliata nell’imbeccare, appena le ho messo a disposizione un “ciuffo” di Centocchio pieno di semini immaturi si è alzata dal nido e, dopo averne fatta una “scorpacciata”, si è messa ad imbeccare di gran lena. Ho notato però che si è dedicata solo all’apice con i semini, lasciando le altre foglie e il gambo sulla griglia della gabbia. Una considerazione spontanea: sono forse i semini immaturi presenti nelle cime che hanno attivato l’imbecco? Chi lo ha già sperimentato me ne darà atto.
Ad maiora, semper.
ALCUNE FONTI:
-Il prato è in tavola – Dafne Chanaz – Terra Nuova Edizioni
-https://www.boscodiogigia.it/foodforest/centocchio-erba-spontanea#google_vignette
-https://it.wikipedia.org/wiki/Stellaria_(botanica)
-https://antropocene.it/2017/08/19/stellariamedia/
-Canaricoltura - Prof. U. Zingoni - edizioni F.O.I.
54 NUMERO 4 - 2024
Anagallisarvensismordigallina, fonte: wikimedia.org
Anagallisarvensisf.azurea, fonte: wikimedia.org
La gabbia, tra ragionamento e giudizio morale
testo di PASQUALE LEONE, foto PIXABAY


Nell’immaginario collettivo un uccello domestico che “evade” dalla gabbia si dirige verso la libertà. Nella realtà si dirige verso morte certa, fonte: pixabay
La gabbia, per ovvi motivi, è stata da sempre associata al concetto di detenzione.
Ciò prescinde dai soggetti che vi si trovano e dai motivi per i quali gli stessi siano alloggiati lì.
A causa di luoghi comuni, per talune ideologie o semplicemente per spirito di partito preso, si è creato nei confronti di noi ornicoltori, proprio per il fatto che noi teniamo i nostri soggetti

Se si dovesse effettuare una disamina quanto più obbiettiva possibile, uno dei primi parametri a dover essere preso in considerazione dovrebbe essere l’aspetto etologico
in gabbia, una sorta di astio basato più sulla falsa riga di una ratio abulica di questo genere che per delle motivazioni fortemente convinte.
Difatti, se si dovesse effettuare una disamina quanto più obbiettiva possibile, uno dei primi parametri a dover essere preso in considerazione dovrebbe essere l’aspetto etologico.
Purtroppo, viviamo in un contesto sociale dove il giudizio morale prevale sul
NUMERO 4 - 2024 55 DIDATTICA & CULTURA


ragionamento quando si parla di ornicoltura, per cui immagini come quella della foto 1 sembrano dare al canarino l’idea della libertà mentre, al contrario, lo condannano a morte certa. Questo poiché non viene mai preso in considerazione l’aspetto etologico della situazione e soprattutto non viene considerata per quello che è la stessa etologia, ossia una vera e propria scienza.
Tornando all’aspetto prettamente psicologico, diversi studi nell’ambito della neuroscienza e della psicologia morale hanno evidenziato come il ragionamento cosciente venga notevolmente ridimensionato da altri processi, innescati molte volte in maniera automatica e inconscia. È errato dunque pensare che il ragionamento abbia un ruolo esclusivo nella formazione delle nostre decisioni morali, in quanto condizione necessaria ma non sufficiente (*).
Pertanto, considerando la gabbia in sé, le persone avrebbero la tendenza a dare una risposta emotiva e non razionale quando messe davanti ad un uccello in gabbia.
La gabbia diverrebbe pertanto qualcosa di coercitivo e, dal punto di vista emozionale, quasi di cattivo, finanche malvagio nei confronti di un uccello.
Per quanto la cosa possa sembrare paradossale, viene inconsciamente igno-
rato il fatto che statisticamente (e parliamo dunque su basi scientifiche) un uccello domestico fuori dalla propria gabbia ha un’aspettativa di vita inferiore alle ventiquattro ore prima di essere predato o di morire di stenti.
Come dicevo, la stessa etologia viene surclassata da un modello utilitaristico del giudizio morale derivante da una serie di processi che portano alla formazione di giudizi morali.
Tali giudizi sono la risultante di alcuni processi decisionali per i quali le persone si comportano quali fossero degli alter ego (almeno dal punto di vista etico/morale) di veri e propri giudici in procinto di emettere il proprio verdetto. Difatti, quando poi si chiede di spiegare
in maniera razionale (intesa come valutazione dei pro e contro) quali siano state le motivazioni che hanno portato a tali scelte, non riescono a trovare spiegazioni convincenti.
Questo status quo viene strumentalizzato contro noi ornicoltori.
Sedicenti organizzazioni ambientaliste avallano una loro presunta etica Cicero pro domo sua (senza disdegnare attacchi contro il nostro movimento), avvalendosi proprio della mancanza di riflessione dell’opinione pubblica, surrogata da una serie di giudizi morali come sopra descritti.
Viene inconsciamente ignorato il fatto che un uccello domestico fuori dalla propria gabbia ha un’aspettativa di vita inferiore alle 24 ore prima di essere predato o di morire di stenti
Invero, senza avere nulla di concreto, al solo fine di muovere delle critiche verso il nostro movimento, forniscono dei dati che hanno il solo fine di creare una cortina fumogena che celi l’incapacità di evidenziare una motivazione persuasiva. Noi ornicoltori dobbiamo stare in allerta a quando ci troviamo di fronte a delle persone che parlano di noi e/o dei soggetti che noi alleviamo e che ci propongono una serie di dati semplicemente fini a sé stessi. Magari sono persone che svolgono anche mansioni importanti o che ricoprono ruoli istituzionali e che con qualche parola di uso poco comune, con delle performance di giochi di memoria, con dei dati presi un po’ qua e un po’ là, arrivano a delle conclusioni che nulla hanno a che vedere con noi o con i nostri uccelli. Memorizzare non vuol dire conoscere. La conoscenza è frutto dalla capacità dell’elaborazione, non della mera elencazione di dati. La conoscenza nasce da molteplici fattori: dal pensiero critico, dalla abilità di discernere le cose, dagli approfondimenti scientifici ecc. Quando vengono trattati alcuni argomenti delicati, i punti da prendere in considerazione vanno al di là di una serie di dati messi un po’ a casaccio e poi elencati da persone che vogliono farci credere di essere i primi della classe su una materia che è di nostra competenza e non loro, facendo sfoggio soltanto della memoria e rivelandosi poi incapaci di interloquire con persone che hanno idee diverse dalle loro.
NOTA
(*) Cognitive Science - Reflection and Reasoning in Moral Judgment, Paxton J. & Greene J.
56 NUMERO 4 - 2024
Per quanto la gabbia rappresenti per gli uccelli domestici il loro habitat, il giudizio morale prevale sul ragionamento e ci fa pensare alla gabbia come uno strumento di coercizione dell’animale
S pazio Club
Nuovo Consiglio direttivo per il Lizard Canary Club
Italiano
Ciè gradito comunicare che il giorno 2 marzo 2024 si è insediato il nuovo Consiglio Direttivo del Lizard Canary Club Italiano (L.C.C.I.) i cui membri, eletti dall’Assemblea dei Soci, hanno distribuito le cariche come segue:
Antonio Di Tillio - Presidente
Andrea Bartalini - vice Presidente
Stefano Canossi - Segretario
Nicola Giordano - Consigliere
Claudio Corallo - Consigliere
Pasquale Leo - Consigliere
Questo nuovo Consiglio intende operare, come sempre avvenuto anche in passato, con l’inderogabile intento di tutelare la razza e favorirne la diffusione. Affermiamo con forza che il nostro impegno e le nostre iniziative saranno condivisi con tutti i Soci, in un clima di amicizia e reciproco rispetto.
Intendiamo inoltre riprendere la consuetudine di diffondere notizie e divulgare la conoscenza della razza attraverso la riedizione del nostro giornalino periodico (“Lizard News”), effettuare frequenti meeting (il cui contenuto ci verrà suggerito dalle indicazioni dei Soci), mantenere contatti e relazioni con i Club di specializzazione di altri Paesi ed in particolare con la L.C.A. di Gran Bretagna (nazione detentrice dello standard del Lizard), affrontare vari argomenti riguardanti lo standard di perfezione (ad esempio taglia, ciglio e colore di fondo del Lizard, ecc.), favorire il dibattito su alcuni temi non ancora




sufficientemente sviscerati, come quello del Lizard c.d. “Blu”, monitorare l’evoluzione “corretta” e migliorativa dello standard, evidenziare e segnalare gli eventuali nuovi difetti, condividere il più possibile le varie esperienze di allevamento. Tutto ciò si renderà possibile se i Soci esprimeranno al Direttivo le loro opinioni ed eventuali necessità con chiarezza e spirito di collaborazione, utilizzando tutti i mezzi che la tecnologia offre, dalla nuova chat di gruppo su Whatsapp ai vari siti web (FB: LCCI Presidenza, LCCI Segreteria, in costruzione un nuovo sito web ufficiale).
Ci permettiamo di far presente che è sempre possibile iscriversi al Club; a nostro avviso, è più efficace operare “dal di dentro” che non “dal di fuori”. Un numero cospicuo di Soci conferisce certamente maggior potere e dignità a ciascun Club. Con questo sincero spirito di servizio, iniziamo pertanto il nostro lavoro.
Con i più cordiali saluti e auguri da parte del Consiglio Direttivo del L.C.C.I.
Presidenza L.C.C.I.: tel.: 3388803439 - e-mail: lcci.presidenza@gmail.com
Segreteria L.C.C.I.: tel.: 3465135941 - e-mail: lcci.segreteria@gmail.com
Club di specializzazione
NUMERO 4 - 2024 57
Premiazione dei Best alla specialistica di Cesena 2023
O rniFlash
Rondini, le migliori alleate delle aziende agricole
Se una rondine non fa primavera lo fa, invece, un articolo pubblicato in questi giorni sulla rivista internazionale Journal of Applied Ecology. Lo studio condotto da MUSE – Museo delle Scienze di Trento, Lipu e Università degli Studi di Milano ha indagato il “ruolo” di questi uccelli negli allevamenti bovini. La ricerca ha infatti valutato l’effetto che la presenza e l’abbondanza delle rondini all’interno delle stalle della Val di Non, in Trentino, possono avere sul tasso di attività delle mosche, loro potenziali prede e portatrici di diversi patogeni e stress per il bestiame allevato.

L’abbondanza di rondini e mosche è stata misurata per 16 settimane in nove diversi allevamenti in Val di Non, in Trentino, occupati o meno da rondini nidificanti. Da aprile ad agosto 2022, sono stati raccolti dati settimanali sul tasso di attività delle mosche e sulla presenza di rondini, sulle dimensioni della covata, sul numero e sull’età dei pulcini. Attraverso opportuni modelli statistici è stato quindi possibile quantificare l’effetto delle rondini su questi insetti. I risultati mostrano come il tasso di attività delle mosche aumenti, come atteso, con la temperatura e nel corso della stagione; tuttavia, tale incremento appare molto meno marcato in presenza di rondini e ulteriormente limitato quando le rondini sono relativamente abbondanti. A una temperatura di 22°, riporta lo studio, la presenza locale di 25 rondini corrisponde a una riduzione media di oltre il 60% nel tasso di attività delle mosche rispetto a quello che si avrebbe in una stalla senza rondini. La rondine (Hirundo rustica), secondo i dati del Farmland bird index, nel nostro continente ha subito un declino del 19% tra il 1980 e il 2021.
Fonte: Ufficio Stampa Università Statale di Milano - Immagine: www.kodami.it
Finalmente sappiamo cosa sognano gli uccelli
Ilconcetto che gli uccelli possano sognare è affascinante e le ricerche recenti hanno iniziato a svelare i misteri dei loro paesaggi onirici. Le scoperte scientifiche suggeriscono che, simile agli esseri umani, gli uccelli sperimentano fasi di sonno REM (Rapid Eye Movement) e NREM (Non-Rapid Eye Movement), con particolari attività cerebrali che suggeriscono la presenza di sogni. La ricerca su come gli uccelli sognano ha preso il volo negli ultimi anni, grazie all’avanzamento della tecnologia come la risonanza magnetica funzionale (fMRI) e le videocamere ad infrarossi, che hanno permesso di studiare in dettaglio l’attività cerebrale degli uccelli durante il sonno. In particolare, studi su piccioni e fringuelli zebra hanno mostrato che durante il sonno REM, si registra un’intensa attività nelle aree del cervello legate all’elaborazione visiva e alla navigazione spaziale, nonché movimenti oculari e attivazione di aree legate all’azione delle ali, suggerendo che questi uccelli potrebbero sognare di volare I sogni potrebbero aiutare gli uccelli a elaborare le informazioni e consolidare la memoria di eventi vissuti durante la veglia, come dimostrato negli studi sui fringuelli zebra, dove l’attività cerebrale durante il sonno sembra correlata all’apprendimento di nuove canzoni durante il giorno. In definitiva, le ricerche suggeriscono che il sonno e i sogni degli uccelli non sono solo fondamentali per la loro salute fisica ma potrebbero anche essere cruciali per la loro sopravvivenza e adattabilità. Questi studi non solo aprono nuove porte sulla comprensione del sonno e dei sogni nell’evoluzione animale, ma potrebbero anche avere implicazioni pratiche per la conservazione e la gestione degli uccelli

Fonte: https://www.24hlive.it/finalmente-sappiamo-cosa-sognano-gli-uccelli/ - Immagine: F.O.I.

News
al volo dal web e non solo
58 NUMERO 4 - 2024
O rniFlash
C’è un uccello che usa le ali per comunicare con il partner
Ilmetodo principale di comunicazione degli uccelli, non serve spiegarlo, è la voce: il loro canto serve per trasmettere ogni tipo di informazione e, diciamo così, tenersi in contatto. Non è l’unico, però: ci sono uccelli che comunicano con il colore delle loro piume, altri che danzano per conquistare una femmina… E poi c’è la cincia del Giappone, Parus minor, che come dice un nuovo studio pubblicato su Current Biology usa le ali in un modo che pensavamo fosse presente solo nei primati: quando devono indicare al partner “entra pure nel nido prima di me”, lo fanno frullando le ali in un gesto di invito, non dissimile da quello che facciamo noi con le mani.

Non è la prima volta che la cincia del Giappone finisce tra le news scientifiche: già nel 2016 avevamo scoperto che il loro canto è complesso e ha addirittura una sintassi. È stato proprio quello studio a convincere Toshitaka Suzuki dell’università di Tokyo a investigare altri possibili metodi di comunicazione in questo uccello. L’ha fatto osservando otto coppie di cince nel periodo in cui queste stavano allevando i loro pulcini, portando loro il cibo direttamente al nido. In questo modo, Suzuki e il suo team hanno potuto osservare un fenomeno interessante: ogni volta che c’era da entrare nel nido, uno dei due uccelli si metteva sulla soglia e muoveva le ali, “invitando” l’altro a entrare per primo, per poi seguirlo. Il gesto non è necessariamente legato al sesso della cincia, ma le femmine lo fanno più spesso, almeno stando alle osservazioni di Suzuki.
Fonte: https://www.focus.it/ambiente/animali/c-e-una-cincia-che-comunica-a-gesti
Evoluzione degli uccelli: avevamo sbagliato tutto
Gli
scienziati hanno passato secoli a classificare circa 10.000 specie di uccelli in albero genealogico per capire come gli ultimi dinosauri sopravvissuti avessero conquistato i cieli e il sequenziamento economico del DNA avrebbe dovuto rendere tutto più semplice, come è avvenuto per innumerevoli altre specie. Ma a quanto pare gli uccelli li hanno ingannati.

Infatti, due nuovi studi – “A region of suppressed recombination misleads neoavian phylogenomics” pubblicato su Proceedings of the National Academy of Sciences (PNAS) da un team di ricercatori guidato da Siavash Mirarab che insegna ingegneria informatica all’università della California San Diego (UCSD), e “Complexity of avian evolution revealed by family-level genomes”, pubblicato su Nature da un team di ricercatori guidato da Josefin Stiller della Københavns Universitet – rivelano che un altro evento avvenuto 65 milioni di anni fa ha fuorviato la ricerca scientifica dalla vera storia degli uccelli e hanno scoperto che «Una sezione di un cromosoma è rimasta congelata nel tempo per milioni di anni e si è rifiutata di mescolarsi con il DNA vicino come avrebbe dovuto». Questa sezione, appena il 2% del genoma degli uccelli, ha convinto gli scienziati che la maggior parte degli uccelli poteva essere raggruppata in due categorie principali, con fenicotteri e colombe come cugini evolutivi. L’albero genealogico più accurato, che tiene conto della sezione fuorviante del genoma, identifica 4 gruppi principali e identifica i fenicotteri e le colombe come imparentati più lontanamente.
Il biologo Edward Braun dell’università della Florida ha supervisionato il team internazionale di ricercatori guidati da Mirarab, che ha pubblicato le prove che questo pezzo “appiccicoso” di DNA ha confuso la vera storia dell’evoluzione degli uccelli.
Fonte: https://greenreport.it/news/aree-protette-e-biodiversita /evoluzione-degli-uccelli-avevamo-sbagliato-tutto/
News
al volo dal web e non solo
NUMERO 4 - 2024 59









Il mondo degli uccelli “Allevare è proteggere”
Progetto di Ornitologia presso Istituto Comprensivo Giannone - De Amicis Scuola primaria plesso Aldifreda (Caserta)
testo NICOLA GRIMALDI, foto di FLAVIO MORETTI
La A.O.C di Caserta (Associazione Ornicoltori Casertani) è un’organizzazione senza scopo di lucro ma di utilità sociale, che riunisce allevatori professionisti o semplici appassionati di uccelli con l’obiettivo di diffondere l’amore per i volatili, la conoscenza degli stessi e del loro habitat e soprattutto di far conoscere il loro corretto allevamento, sia a scopo “ornamentale” che espositivo, riproducendo in ambiente controllato anche soggetti altrimenti in via d’estinzione.
L’associazione è così composta: Presidente: Pasquale Abbate Vice Presidente: Giuseppe Di Lullo



Consigliere: Flavio Moretti
Segretario: Nicola Grimaldi Abbiamo organizzato due giornate di studio nel plesso di Aldifreda della Scuola Primaria “De Amicis” con gli esperti del settore e alla presenza delle istituzioni locali, il vicesindaco Dott. Emiliano Casale e la consigliera comunale Daniela Dello Buono, coinvolgendo gli alunni delle classi seconde, terze e quarte. I bambini hanno dapprima assistito alla proiezione di un video e poi hanno letto il racconto “Le avventure di PSIT e FRIN”; successivamente, hanno potuto osservare da vicino una serie di uccelli esotici tra cui il Diamante di
Gould ed il Passero del Giappone, oltre a diverse specie di pappagalli tra cui gli Psittacidi. Ne è seguito un avvincente dibattito con spunti ed interventi personali che hanno reso ancora più interessante l’intera attività.
L’A.O.C. è riuscita a condividere con i piccoli alunni della scuola primaria “De Amicis” l’importanza di tutte le attività che essa svolge a tutela della fauna volatile locale ma soprattutto la passione e la cura di questi delicati esserini che tanta compagnia donano a grandi e bambini con il loro canto tenero e melodioso, offrendo un ampliamento originale e costruttivo dell’offerta formativa.


NUMERO 4 - 2024 61 CRONACA
60 anni di ornicoltura e passione ornitologica
Associazione Ornitologica Arzignanese
testo e foto di MIRKO MASIERO
Al termine del sessantesimo anno, l’Associazione Ornitologica Arzignanese 025 è lieta di condividere con tutti i lettori un nostro momento di felicità.
Era il lontano 21 Febbraio 1963 quando un gruppo di 12 appassionati ornicoltori decide di costituire l’Associazione Ornitologica Arzignanese.
Per onor di cronaca erano: Povoleri Luigi (che sarebbe stato il primo Presidente), Capovin Alessandro, Spagnolo Giovanni, Baessato Henny, Mistrorigo Francesco, Fochesato Antonio, Canova Lodino, Bruttomesso Luigi, Cozza Guerrino, Tosolini Luciano, Turra Fabiano e Bertoli Italo.
Da allora molti direttivi si sono succeduti e, malgrado alti e bassi, l’A.O. Arzignanese ha sempre cercato di portare avanti quello che per statuto si era prefissata, ossia “riunire gli allevatori, gli amatori e gli uccellatori per incoraggiare

Era il lontano 21 Febbraio 1963 quando un gruppo di 12 appassionati ornicoltori decide di costituire l’Associazione Ornitologica Arzignanese


62 NUMERO 4 - 2024 CRONACA
Alcuni soci A.O.Arzignanese presenti alla cena
Diplomi consegnati ai Soci minorenni per aver partecipato alla 60ª mostra Contatto Natura 2023
L'attuale Consiglio Direttivo: al centro il nostro Presidente Marco Lovato, alla sua destra il Vicepresidente Eros Tregnaghi e il consigliere Marco Tagliapietra, alla sinistra del Presidente il Segretario Mirko Masiero e il consigliere Paolo Colalto

Presentazione del dolce ai presenti da parte del direttivo
e incrementare l’allevamento, migliorare la razza di uccelli (compresi gli ibridi) che possono essere allevati in cattività e contribuire nei limiti del possibile e del consentito a portare il proprio apporto alla scienza ornitologica”.
Ai giorni nostri numerose cose e sensibilità sono cambiate nella società, molte di più sono le difficoltà, ma malgrado ciò la nostra associazione cerca di man-

Il nostro dolce prima della degustazione
tenersi al passo con i tempi. Negli ultimi anni abbiamo unito le forze, collaborando con APOV, nel cercare di tenere viva la passione per l’ornicoltura.
Durante la nostra cena, siamo stati felici di omaggiare i nostri sette soci minorenni con un diploma di riconoscenza che possa essere per loro di stimolo nel continuare a portare avanti questa passione e per essere magari, un domani,
coloro che manterranno viva l’Associazione.
La serata è stata un ottimo momento per stare assieme, soci, famigliari ed amici, cosa che non guasta mai.
L’attuale Direttivo è costituito da Marco Lovato, Eros Tregnaghi, Paolo Colalto, Marco Tagliapietra e Mirko Masiero, che augurano all’ A.O. Arzignanese cento di questi giorni.




NUMERO 4 - 2024 63


P ensieri in libertà
A nostro padre, Micaela ed Elisa figlie di un ornitofilo
Può far sorridere questa frase, ma dietro

c’è tanta roba, tanti ricordi, valori e insegnamenti di vita che sono nati propri dalla passione di nostro padre.
Una passione, un hobby, quello di allevare canarini, tramandata dal nonno Timo, durata una vita. Coronata da importanti riconoscimenti, ma questo poco importa!
Correvano gli anni ‘70-‘80, eravamo bambine e all’epoca vivevamo a Pesaro. I nostri risvegli erano i più belli che si possano sognare! Aprivamo gli occhi al nuovo giorno in compagnia del cinguettio dei canarini che avevamo proprio sopra la nostra camera, in soffitta. Lì c’era il regno magico del babbo.
Siamo cresciute a pane, amore e canarini. Ogni anno arrivava il momento dell’accoppiamento, la schiusa delle uova (un’opportunità magica poter assistere all’inizio di una nuova vita), poi lo svezzamento, la preparazione e la selezione dei più belli per le mostre ornitologiche.

Abbiamo sempre respirato la passione, la dedizione, la costanza e pure il sacrificio che il babbo ha avuto per i suoi animali, ed è stato arricchente!
Ma il suo tempo non si limitava solamente a seguire l’allevamento, eh no, era troppo poco… e così, insieme ad altri amatori, diventati poi anche amici, ha fondato l’A.P.O., diventata poi negli anni punto di riferimento per tanti altri allevatori. Ha pure studiato per diventare Giudice F.O.I. Insomma, una passione a tutto tondo!

Poi c’erano i pranzi sociali, occasione di condivisione di una passione per grandi e piccini, e per le signore quella di sfoggiare il vestito nuovo e i gioielli più “luccicosi”; non poteva mancare l’amato gossip che io origliavo con piacere… Ah! Che belli quei ritrovi, quante amicizie fatte e perse.

E poi, le premiazioni alle mostre:
1° Premio Stamm Agata Rosso Mosaico: Mengacci Pierluigi!
1° Premio Singolo Isabella Rosso Mosaico: Mengacci Pierluigi!
1° Premio Campione Italiano Isabella Pastello: Mengacci Pierluigi!
E la lista continuò negli anni… Che bravo, babbo! Coppe e medaglie iniziavano a riempire la credenza di casa e la mamma, pazientemente, doveva spolverare.
Come dicevo, un babbo ornitologo è “tanta roba”! Ci si improvvisava anche prof. di Scienze, ti ricordi, ba’?
Non basterebbero pagine per ricordare aneddoti di vita.

Si faceva promozione nelle scuole, che molto volentieri partecipavano alle nostre mostre ornitologiche e io (mia sorella un po’ meno) ero sempre in prima fila, braccio destro di Gigi, la sua ombra. Devo dire che mi piaceva tanto.
Gli anni scivolano via velocemente, cambiano le cose, le situazioni, cresce la famiglia, arrivano i nipoti. E tu, figlia, assisti ora da mamma al ciclo della vita che si ripete e rivedi negli occhi di Sahumi, Niccolò, Federico, la meraviglia, lo stupore che tanti anni prima hai provato, quando per la prima volta, a mani unite a nido, con la paura di stringere troppo, tuo padre ti ha posato, con la grazia che solo in lui ho visto, un piccolo pulcino di canarino spennacchiato, caldo e morbido.
Che belle emozioni che ci hai fatto vivere, babbo, ricordi dal valore inestimabile!
Come tutte le più belle cose, anche questa esperienza di allevatore è giunta alla sua conclusione; i canarini del babbo sono “volati” altrove, ma a noi resta la cosa più preziosa:
resta il babbo, con il suo cuore speciale e con una passione in più, quella di scrittore orto-ornitofilo.

Ti vogliamo un mondo di bene: buona festa del papà, ba’!
Micaela ed Elisa


64 NUMERO 4 - 2024
Pierluigi Mengacci (babbo Gigi) Scrittore orto-ornitofilo


















