Italia € 30,00 - Estero-Europa € 50,00 Estero-ExtraEuropa € 70,00
Un numero € 5,00 - Arretrato € 6,50
C.C.P. 53684957
Le quote abbonamento vanno versate, mediante vaglia o assegno, alla Segreteria. Le opinioni espresse dagli Autori non impegnano la Rivista e la sua Direzione.
In copertina: Diamante codalunga di Hecki (Poephila acuticauda hecki) Foto: DANIELE ZANICHELLI
Canarini di Colore Canarini di Forma e Posizione Lisci
Editoriale
Una sentenza vale più di mille falsità
di D IEGO C ROVACE
Non avrei mai immaginato di subire un attacco indiscriminato e privo di qualsivoglia fondamento da parte di chi avevo supportato con entusiasmo, apportando tutto il mio impegno ed il mio bagaglio tecnico.
Mi riferisco alle vicende che mi hanno visto coinvolto, mio malgrado, in un procedimento disciplinare istruito dal Consiglio Direttivo della C.O.M., per difendermi dal quale ho dovuto adire le vie legali, rivolgendomi ad un giudice delle misure urgenti del Tribunale olandese dell’Aia, in quanto mi erano stati inflitti 4 anni di sospensione come giudice O.M.J. ed ero stato interdetto dallo svolgere il mio ruolo di Presidente dell’Ordine Mondiale dei Giudici.
Il tutto ha avuto inizio il 6 maggio dello scorso anno (data di ricezione dell’avviso di avvio di procedimento disciplinare nei miei confronti da parte della C.O.M.) ed ha avuto termine il 17 gennaio di quest’anno (data in cui è stata emessa la sentenza che sospende le sanzioni ingiustamente comminate e condanna la C.O.M. al pagamento delle spese legali).
In questo arco di tempo, all’estero e soprattutto in Italia, sono state diffuse molte falsità sulla vicenda in rassegna. Chi ricopre ruoli di vertice, in qualsiasi istituzione, deve sempre mettere in conto che potrebbe ricevere critiche, disappunti o maldicenze da parte di chi, per i più disparati motivi, non ne condivide l’operato, ha interessi contrastanti o è semplicemente invidioso.
La cosa che ritengo maggiormente indecente in tutta questa storia è la faziosità assunta da alcuni italiani i quali hanno raccontato una personale versione dei fatti, distorcendoli a seconda di come era maggiormente conveniente. Occorre in questo caso distinguere chi asserisce il falso per ignoranza della verità da coloro che affermano il falso pur essendo al corrente di come stanno realmente le cose.
I primi sono solitamente gli amanti del pettegolezzo, coloro i quali si trastullano con le chiacchiere da bar e quasi mai hanno finalità malevole. I secondi sono invece armati di sostanziale cattiveria, spesso mossi dall’invidia o da personali interessi per cui l’alterazione della realtà corrisponde all’esplicito desiderio di screditare l’altrui reputazione per raggiungere i propri abietti scopi. Questi ultimi sono spesso appellati con il sostantivo di infami. Tra costoro, spiace evidenziarlo, ci sono anche dei giudici, occul-
tamente schierati, privi di moralità e di senso di appartenenza, ai quali bastano meno di due denari e qualche promessa per ottenere in cambio misera piaggeria. Il tempo è galantuomo, ristabilisce la verità, ripara i torti, cura ogni cosa, dà ragione ai giusti. Ma veniamo ai fatti, che per necessità sintetizzo, ma che per precisione e rispetto della verità dovrò argomentare in maniera esaustiva negli aspetti più salienti della vicenda. In data 6 maggio 2024 ricevevo al mio indirizzo di posta elettronica una mail da parte del presidente della C.O.M., con la quale lo stesso mi comunicava la decisione assunta dal Comitato Esecutivo della stessa C.O.M. di sospendermi preventivamente, con effetto immediato, dall’esercizio delle mie funzioni sia come presidente dell’O.M.J. e sia come giudice O.M.J., Le motivazioni poste a base di tale decisione risiedevano nel mancato recepimento da parte della mia Federazione d’appartenenza, la F.O.I., di regole relativamente alle quali la mia persona non ha alcuna influenza né possibilità di modifica, atteso che la divergenza in rassegna è frutto del volere dell’Assemblea delle Associazioni, organo sovrano della F.O.I., espressasi nel corso dell’Assemblea Generale svoltasi lo scorso 14 aprile 2024. Il presidente Ramoa, presente all’Assemblea Generale, evidentemente disturbato da questa decisione, non curante dei principi generali su cui poggia un’istituzione giuridica senza scopo di lucro né delle regole proprie della C.O.M., invece di avviare un contenzioso con la F.O.I. per il tramite di C.O.M. Italia e porre in essere un arbitrato volto a stabilire se la regola in rassegna fosse obbligatoriamente da recepire oppure no (1), medita e pone in essere un’azione basata sostanzialmente sulla coercizione, come meglio evidenzierò in seguito.
Per cui riunisce il Comitato Direttivo della C.O.M. e, in qualità di testimone, riferisce gli accadimenti che pone a base della sua richiesta di sospensione nei confronti del sottoscritto.
Si riportano di seguito, in corsivo, i punti sostanziali meritevoli di essere esposti al fine di apprezzarne l’insussistenza, ed a seguire le mie osservazioni utilizzate nel corso dell’udienza svoltasi in olanda:
1. “Sono stato informato che NESSUN giudice OMJ della FOIItalia aveva potuto accettare l’invito per giudicare al concorso internazionale (si riferisce alla mostra di Salerno – mia nota) e
ciò per indicazione del Presidente dell’ordine dei giudici FOI, il giudice OMJ Andrea Benagiano”.
Oltre ad omettere di dire da chi è stato informato, il presidente Ramoa riferisce la prima notizia non vera, in quanto nessun diniego è pervenuto da parte del Presidente dell’Ordine dei Giudici italiano, ma è stata l’organizzazione della mostra di Salerno a comunicare ai giudici italiani di non voler attendere il nulla-osta di Andrea Benagiano revocando così l’invito, come comprovato dalle mail inviate al Presidente dell’Ordine dei Giudici da parte degli stessi giudici prima invitati e poi revocati.
2. “Immediatamente ho informato il presidente dell’OMJ signor Diego Crovace che questo fatto è grave e gli ho domandato di risolvere la situazione nel rispetto dei regolamenti della COM e dell’OMJ”.
Poiché è oggettivamente falsa la prima affermazione, di conseguenza non è veritiera anche la presente narrazione dei fatti.
3. “Io l’ho anche avvertito (con riferimento alla mia persona – mia nota) della gravità della situazione e delle possibili conseguenze istituzionali e disciplinari che si dovrebbero con ogni sforzo evitare”.
Per transitività è falsa anche questa circostanza in quanto nessun diniego era stato di fatto espresso dal Presidente dell’Ordine dei giudici italiano.
4.“ …. Quando il tema del sistema di convocazione dei giudici per i concorsi internazionali C.O.M. si è presentato, il signor Diego Crovace ha dichiarato“ah no, sui giudici non si può cambiare il sistema FOI, ciò porterebbe molti problemi a l’ordine dei giudici (traduzione letterale di dichiarazioni sentite da me stesso in prima fila dell’anfiteatro della sala)”
Per sfortuna di Ramoa l’intera assemblea è, come ogni anno, video-registrata e dalla visione della registrazione integrale si evince chiaramente che non solo non ho mai parlato durante la trattazione di questo punto, ma verso la fine della discussione mi sono alzato per andare via dalla sala per non farvi più ritorno.
5.“Sono rimasto veramente sorpreso dalla situazione, principalmente della condotta del Presidente dell’OMJ, il quale per inazione ma anche per azione si è dichiarato contro la discussione e l’aggiornamento dei regolamenti della FOI per renderli in linea e compatibili con quelli della C.O.M. e dell’O.M.J.”.
In sostanza mi viene attribuita la colpa di non essere intervenuto per indurre l’Assemblea delle Associazioni a modificare la propria decisione. Ritengo invece di aver assunto un comportamento assolutamente corretto, in qualità di vice-presidente FOI, lasciando che i singoli votanti decidessero liberamente, senza condizionamenti o imposizioni.
6.“La situazione è in tal modo inaccettabile anche per il giudice OMJ Riccardo Rigato, membro del Comitato Esecutivo dell’OMJ che s’è visto rifiutare l’invito per indicazione del Presidente dei giudici FOI Andrea Benagiano”.
Continua la sagra delle inesattezze narrate dal Presidente Ramoa, infatti il giudice Riccardo Rigato non ha mai ricevuto alcun diniego, semmai ha provveduto egli stesso a inviare un SMS al responsabile dell’organizzazione della mostra di Salerno con il quale comunica che probabilmente avrebbe ricevuto altra destinazione per quella data.
Per amore della verità devo dire che rispetto a questa vicenda ero a conoscenza delle pressioni fatte dagli organizzatori della mostra di Salerno ai giudici invitati già a dicembre dell’anno prima
per ottenere i primi di gennaio l’autorizzazione a partecipare alla mostra di Salerno, la quale si sarebbe svolta in ottobre, per cui non era giustificata l’urgenza. Di questo ne avevo parlato con il Presidente dell’OdG Benagiano che mi aveva assicurato il rilascio del nulla osta.
Il 29 maggio 2024, a Roma, veniva organizzato un incontro tra il Presidente della F.O.I., Antonio Sposito ed il Presidente della C.O.M., Carlos Fernando Ramoa, alla presenza del promotore della mediazione, il Presidente di una delle più prestigiose organizzazioni di eventi ornitologici internazionali a livello mondiale, Ivano Gualerzi.
Nel corso di tale incontro si addiveniva ad un accordo verbale vertente su tre punti:
-riproposizione della problematica relativa alle premiazioni speciali all’Assemblea delle associazioni della F.O.I.;
-redazione di norme di raccordo per armonizzare la disciplina della richiesta di giudici italiani direttamente da parte dei comitati organizzatori in Italia di mostre internazionali;
-revoca delle sospensioni nei confronti miei insussistenti.
Nei giorni immediatamente successivi il Presidente FOI Antonio Sposito convocava il Consiglio Direttivo della FOI e con esso conveniva:
-l’inopportunità di richiedere per la terza volta (2) all’Assemblea delle Associazioni di esprimersi sulla questione delle premiazioni;
-di proporre al Presidente Ramoa l’attivazione di una commissione d’arbitrato, come peraltro previsto per casi analoghi dall’art. 22 dello Statuto della C.O.M.;
-la necessità di armonizzare il sistema di chiamata dei giudici, così come regolamentata in Italia con il sistema previsto dalle mostre internazionali;
-di anteporre sopra ogni cosa la revoca delle arbitrarie ed insussistenti sanzioni disciplinari comminate ai due tesserati italiani Benagiano e Crovace;
-che lo scrivente conservava la facoltà di tutela dei propri diritti soggettivi dinanzi alle competenti Autorità.
Il Presidente FOI Sposito redigeva l’accordo e lo inviava per posta elettronica al mediatore Ivano Gualerzi.
Nel frattempo, dopo più di un mese dalla lettera in cui veniva comunicata la sospensione del sottoscritto, il Presidente della COM Ramoa faceva giungere tramite raccomandata la nota contenente le modalità attraverso le quali mi veniva data la possibilità di essere audito o di inviare scritti difensivi. Nella stessa lettera il Ramoa comunicava essere istruttore della procedura disciplinare intentata nei miei confronti.
A quel punto, ritenendo che si fosse raggiunto il colmo della vessazione nei miei confronti, essendo peraltro in essere un tentativo di conciliazione non ancora definito, nel quale si proponeva la revoca delle sanzioni per insussistenza nelle motivazioni, ho valutato di dovermi sottrarre da tale procedura, comunicando con mail le eccezioni che di seguito riassumo: -totale assenza di garanzia a tutela dei miei diritti, in quanto il Ramoa era al contempo testimone dei fatti posti a base della mia sospensione temporanea, promotore dell’attività accusatoria nei miei confronti ed ora si ergeva addirittura ad organo giudicante. In sostanza recitava tutti i ruoli nell’ambito di un medesimo procedimento disciplinare e ciò non è assolutamente compatibile in quanto non costituisce garanzia di imparzialità.
-a mia difesa avevo prove documentali e testimoniali che sconfessano le sue testimonianze; -la sua partecipazione a questo procedimento disciplinare è evidentemente viziata da un palese conflitto d’interesse, poiché da anni ha in essere reciproci accordi ornitologici ed amicali con gli organizzatori della mostra internazionale di Salerno, da cui ha origine la sua iniziativa di assumere provvedimenti disciplinari nei miei confronti, invece di utilizzare gli strumenti contenuti nello statuto della C.O.M., atti a risolvere in maniera pacifica le problematiche che vedono contrapposti la mia Federazione e la C.O.M.. Per tutto quanto innanzi esposto ho chiesto di indicarmi la persona o le persone che, in suo luogo, potessero procedere alla mia audizione alla quale mi dichiaravo senz’altro disponibile. La mia richiesta non ha avuto alcun esito, anzi alcuni giorni dopo giungeva la contro proposta di accordo da parte del presidente Ramoa dalla quale si evince chiaramente la natura coercitiva della mia sospensione poiché, oltre a non accettare nulla di quanto proposto dalla mia Federazione, venivano aggiunti argomenti che non erano stati concordati nell’incontro avvenuto a Roma e veniva richiesto che a firmare l’accordo fossero anche i due sospesi, i quali solo accettando le richieste fatte da Ramoa avrebbero ottenuto di far archiviare le sospensioni nei loro confronti.
In sostanza Ramoa chiedeva: «la F.O.I. si impegna a riformulare i suoi regolamenti sulla questione premiazioni, in quanto riguarda le mostre internazionali COM, adattandoli a quelli della COM e OMJ». In questo caso viene esercitata l’ennesima forma di coercizione al fine di ottenere l’introduzione di una norma che potrebbe essere adottata solo non tenendo conto della volontà dell’Assemblea delle Associazioni che l’ha votata. Ancora una volta Ramoa si sottrae all’arbitrato proposto dalla mia Federazione.
Ed inoltre lo stesso precisava: “Il Vicepresidente FOI e giudice OMJ, Diego Crovace, si dichiara d’accordo con la modifica delle regole e coopererà attivamente per la sua approvazione ed applicazione”, per poi concludere come segue: “Firmato questo documento, il Presidente della COM, nella sua qualità di istruttore della procedura disciplinare iniziata il 1° maggio 2024 nei confronti di Diego Crovace, proporrà, immediatamente, al comitato disciplinare della COM la sospensione degli ulteriori termini della procedura disciplinare”.
Da qui si evince con chiarezza la vera natura delle azioni poste in essere nei miei confronti, per cui la sospensione ed il conseguente discredito della mia persona sono funzionali ad ottenere l’applicazione della norma voluta da Ramoa e non accettata dall’Assemblea delle Associazioni.
Poiché non avevo alcuna intenzione di barattare la presidenza dell’O.M.J. con il voto della mia Assemblea ho espresso ai colleghi del C.D. della F.O.I. la mia contrarietà.
Per cui è stato valutato di non accettare la proposta del presidente della C.O.M. e, come conseguenza, il presidente Ramoa, ed il Comitato Direttivo della C.O.M. hanno emesso all’unanimità un dispositivo di sospensione di 4 anni nei miei confronti.
Tutte le affermazioni da me riportate nella narrazione suesposta sono supportate da prove testimoniali e documentali poste a corredo della citazione in giudizio della C.O.M. presso il Tribunale dell’Aia in Olanda.
A suffragio delle ragioni da me sopra esposte, riporto i passaggi più rilevanti della sentenza motivata emessa dal giudice delle misure urgenti del Tribunale olandese dell’Aia (3).
“Il giudice, in una valutazione marginale, ritiene che il consiglio
della COM non avrebbe potuto adottare le decisioni criticate con ragionevolezza, per cui la decisione di sospensione (e quella provvisoria) è annullabile”.
“Il giudice delle misure provvisorie concorda con Crovace nel ritenere che la COM non abbia sufficientemente supportato i fatti e le circostanze precedentemente menzionati, in relazione alle contestazioni motivate di Crovace”.
“Il giudice delle misure provvisorie comprende che vi sia delusione nella COM riguardo alla modalità in cui la FOI ha gestito la sua politica per quanto riguarda la designazione dei giudici per gli eventi della COM, e forse anche riguardo alla visione della FOI sulle politiche relative ai premi da assegnare ai vincitori. Tuttavia, queste circostanze non giustificano, nemmeno tenendo conto della prudenza con cui un giudice deve esaminare tali decisioni, la sospensione di Crovace, tanto meno per un periodo di quattro anni, dalle sue funzioni all’interno della COM”.
“Infine, la COM ha sostenuto che Crovace, durante il procedimento disciplinare, abbia ingiustamente rifiutato di farsi sentire, causando così indirettamente il fatto che il consiglio della COM abbia preso provvedimenti contro di lui senza il suo contributo. Il giudice delle misure provvisorie non condivide questa posizione. Il giudice ritiene che non sia incomprensibile che Crovace abbia richiesto, nella sua lettera del 20 giugno 2024, di essere ascoltato da un altro membro del consiglio della COM anziché da Ramoa. Non è irragionevole pensare che Crovace potesse nutrire dubbi sull’indipendenza di Ramoa, che nel procedimento disciplinare ricopriva sia il ruolo di testimone che quello di investigatore disciplinare e che era anche coinvolto nel processo decisionale del consiglio della COM”.
“Il giudice delle misure provvisorie conclude che la COM non ha potuto ragionevolmente decidere di imporre una sospensione di quattro anni”.
“La COM è risultata soccombente e dovrà quindi pagare le spese legali (compreso l’eventuale costo per l’ulteriore esecuzione)”.
In pratica, tutte le osservazioni addotte a mia difesa sono state avallate dal giudice e dallo stesso poste a motivazione della sentenza di condanna.
Ribadisco che per motivi di brevità ho esposto solo le questioni salienti afferenti l’incresciosa vicenda, ma ci sono molte altre circostanze che andrebbero rese note al fine di far comprendere la gravità delle decisioni assunte dalla C.O.M., l’insussistenza delle argomentazioni addotte per poterle sorreggere unitamente alle irregolarità procedimentali commesse.
Rivolgo un monito a coloro i quali definisco “collaborazionisti”, che in quanto appartenenti alla F.O.I. hanno vilmente fomentato queste situazioni, pur conoscendo la realtà dei fatti: dovreste vergognarvi, ma poiché non avete morale, sono certo che quando mi incontrerete avrete la sfacciataggine di salutarmi: risponderò al saluto solo per educazione.
Concludo citando una frase di Winston Churchill: “La verità è incontrovertibile. La malizia può attaccarla, l’ignoranza può deriderla, ma alla fine, eccola”.
Note
1)Tale procedura è prevista dall’art. 22 dello statuto della C.O.M.
2)La prima volta l’Assemblea si era espressa nel 2008, introducendo la regola in rassegna.
3)La versione integrale della sentenza è pubblicata sul sito della Federazione https://www.foi.it/
Stranezze
testo di GIOVANNI CANALI, FOTO A. J. SANZ e E. DEL POZZO
Da un po’ di tempo si notano stranezze in vari tipi. Una è la fortissima riduzione o totale inibizione della feomelanina in alcuni soggetti, ed è quest’ultimo l’aspetto da considerare. In passato non si poteva mai inibire totalmente la feomelanina ed in effetti, in certi tipi, si parlava giustamente della massima riduzione o diluizione. Il fatto però non è inspiegabile ed è forse fra i meno strani; si tratta o di selezione estrema, ma allora qualcosa rimarrebbe e si vedrebbe, oppure di aver inserito il monomelanico. Il monomelanico è una mutazione pare recessiva autosomica che inibisce la feomelanina. Sull’eumelanina alcuni dicono che il monomelanico non agisca, ma io ritengo, da osservazioni fatte, che un effetto modesto ma rilevabile ci sia anche sull’eumelanina. Direi che l’eumelanina non si esprima al massimo, si nota meglio sulla continuità e larghezza del disegno. Il problema che si pone è di evitare una presenza totale o comunque eccessiva del monomelanico. In effetti mi pare ovvio che la massima riduzione o diluizione basti ampiamente. Anche da considerare l’effetto sull’eumelanina non gradito. Ritengo senz’altro che sia da evitare un’ostilità eccessiva contro la feomelanina. Si badi che l’esistenza della feomelanina come pigmento naturale del Canarino è certa e tipica. Ci sono sati diversi studi nei Fringillidi in genere ed il Canarino non fa eccezione. Inoltre la risposta diversa a certe mutazioni la evidenzia, come pure il colore tipico. Per giunta le femmine canarine hanno più bruno da feomelanina dei maschi, i quali sono più ricchi di eumelanina e quindi
Nero rosso brinato, foto: A. J. Sanz
L’esistenza della feomelanina come pigmento naturale del Canarino è certa e tipica
sono in linea con una regola generale più volte descritta e della quale tenere ben conto (“La feomelanina nei canarini” I. O. n° 1/2021 di G. Canali e D. Crovace). Ricordo che la differenza chimica maggiore, fra le melanine, è la presenza dell’aminoacido cisteina nella feomelanina. Inoltre la feomelanina presenta anche diversità dal
punto di vista fisico: nella forma dei granuli oltre che a polimerizzare in modo differente. Direi che nei tipi diluiti si debba selezionare “contro” la feomelanina, per diluirla al massimo, ma senza farne un assillo. Nei neri che oggi si vogliono con minima (o nulla) feomelanina, ancor meno assillo per non finire col danneggiare anche il disegno. Inoltre bisogna tener conto che nelle penne giovanili il bruno da feomelanina è maggiore, rispetto al piumaggio adulto, per pregressa selezione naturale che intendeva rendere più mimetici i giovani e che, ovviamente, si nota nelle penne non mutate in prima muta. Un altro aspetto strano più complesso del precedente è dato da espressioni maggiori delle melanine rispetto al passato.
Mi pare di aver già accennato in qualche occasione dell’espressione nei bruni opale, ma direi anche nei bruni classici. In questi canarini si arriva ad un disegno così forte da lasciare perplessi. Di esperienza con gli opale ne ho avuta tantissima, sia come allevatore che come giudice. Ricordo che molto spesso gli opale li “rifilavano”
Anche i bruni classici di oggi talora hanno disegni così marcati da sembrare quasi neri
a me da giudicare, visto che non erano molto graditi per le loro difficoltà, e si sapeva della mia predilezione. Ebbene posso dire che di bruni opale con il disegno attuale non c’era traccia. Anche i bruni classici di oggi talora hanno disegni così marcati da sembrare quasi neri. In diversi casi sono carenti di feomelanina, questo è un difetto gravissimo che non può essere compensato dal disegno così spinto.
Il problema è: da cosa deriva tutto ciò? Mi risulta difficile pensare solo alla selezione. Mi pare che i discorsi che sono stati fatti su possibili fattori d’inscurimento possano essere fondati. Non facile però inquadrare bene tali fattori ed il loro comporta-
mento. Posso dire che i bruni opale di allora, se pigmentati, avevano sempre tono azzurrino evidente, gli attuali non sempre e comunque meno espresso. Direi fatto collegabile alla maggiore espressione di melanina che può ostacolare l’effetto azzurro. Del resto è fatto ben noto: basta ricordare che, anche in passato, i neri opale non avevano certo l’azzurro degli agata opale. Per evitare discorsi errati è comunque bene ricordare sempre che l’opale vero abbassa la melanina nella pagina inferiore della penna e, se tipico almeno in bruni ed agata pigmentati, produce sempre effetti azzurrini di natura strutturale. Ho ritenuto di precisare questo perché si sono visti troppi soggetti fatti passare per opale magari vincenti senza essere in effetti tali. Il gene opale, a mio avviso, è molto soggetto a mutazioni e quindi mi rifarei a questo. È stata accertata la mutazione onice, ma io ritengo ve ne siano anche altre, in particolare una da me battezzata “ossidiana”. C’è poi li problema del mogano che viene dato per fatto selettivo, ma a me sembra davvero singolare che una semplice selezione possa inibire totalmente la feomelanina, solo ridotta nell’opale, modificare radicalmente l’espressione dell’eumelanina rendendola maggiore nonché totalmente priva di azzurro e, soprattutto, riportarla nella pagina superiore della penna. Melanine che salgono e scendono francamente mi stupiscono molto. Quanto all’ossidiana, come mi sono permesso di chiamarla, ci sono aspetti diversi che ho valutato nei bruni ossidiana: l’eumelanina non è abbassata, è meno ridotta rispetto all’opale ed è priva di azzurro, mentre la feomelanina è meno ridotta rispetto all’opale. Quest’ultima è la differenza maggiore nei confronti del mogano che non esprime feomelanina. Oggi ne vedo molto meno, mi sembrava logico che questa palese mutazione venisse valutata, anche per un eventuale riconoscimento, non essendo disprezzabile esteticamente anche se molto meno affascinate dell’opale.
Nero mosaico rosso, foto: E. del Pozzo
L’aspetto più difficile da valutare rimane comunque il disegno più marcato sia nei bruni classici che opale. Si badi comunque che l’eumelanina bruna dei bruni non diventa veramente nera, anche se l’aspetto può far venire dubbi, l’occhio a volte si fa ingannare. Ricordo che non bisogna guardare solo i tipici o belli ma anche gli atipici o brutti per capire la natura di un tipo e non mi pare che vi sia dubbio che i bruni deboli palesino il bruno. Senza contare l’evidenza di becco e zampe che appaiono bruni in modo palese, anche nei bruni più spinti e tipici.
C’è poi la faccenda satiné di cui ho parlato in passato, ma che ritengo sia da rimarcare. Ribadisco che non vi è dubbio alcuno sulla condizione allelica con l’agata. Non mi soffermo, ritenendo le mie argomentazioni precedenti largamente più che sufficienti (“Domande sul Satiné” I. O. n° 1/2024 e “Discussioni sul Satiné” I. O. n° 3/2021) Si parla di satiné ossidati ed anche
Il disegno di eumelanina bruna si può dire che abbia subito una modificazione davvero drastica e singolare, visto che certi toni sono più da feomelanina che non da eumelanina, ancorché mutata
qui non mi soffermo avendo sviscerato il tema ampiamente in passato. Quello che vorrei valutare è una certa perdita di tipicità dell’espressione satiné che serpeggia. Il disegno satiné è sempre stato caratterizzato da to-
nalità rossicce o tendenzialmente arancio. Toni oggi compromessi come vedremo. Le descrizioni dei vari tecnici non sempre coincidevano, si definiva anche con il termine “beige chiaro” riferito alle melanine satiné. Di certo, il disegno di eumelanina bruna si può dire che abbia subito una modificazione davvero drastica e singolare, visto che certi toni sono più da feomelanina che non da eumelanina, ancorché mutata. Vero che si tratta di disegno di eumelanina bruna già mutata dalla mutazione di isabellismo che genera il bruno, tuttavia un effetto davvero forte. Ha sempre sorpreso il fatto che il disegno nero venisse inibito tranne tracce sulla punta delle remiganti. Su questo aspetto non mi soffermo, ma ho fatto ipotesi in passato. Dico solo che questa circostanza è molto eloquente sul fatto che l’eumelanina del disegno sia già mutata. Quello che desidero considerare ora è il fatto che si cominciano
Bruno intenso rosso avorio, foto: E. del Pozzo
Bruno opale mosaico giallo, foto: E. del Pozzo
a vedere disegni atipici, come anticipato. Si vedono disegni direi piuttosto sul marroncino chiaro, senza o molto carenti dei toni rossicci o arancio. A volte somigliano all’espressione dei portatori isabella. L’occhio appare più o meno rosso o rossiccio, di solito non molto rosso.
La questione occhio andrebbe trattata a parte. Il tutto necessita di approfondimenti capillari.
Il mal vezzo di accoppiare in purezza può aiutare a spiegare perdite di tipicità, ma non credo che basti. Si potrebbe pensare anche qui all’intervento di qualche fattore di inscurimento? Direi di non poterlo escludere, anzi...
Un’altra ipotesi, magari più complicata ancora, potrebbe indurci a pensare ad ulteriori mutazioni alleliche. In specie diverse dal canarino, si è verificata una mutazione detta “aminet”, allelica all’agata e quindi al satiné e forse anche altre in verifica,
Si vedono disegni direi piuttosto sul marroncino chiaro, senza o molto carenti dei toni rossicci o arancio
con forme intermedie ulteriori. Quindi non si può neppure escludere un’altra mutazione allelica anche nel canarino. Comunque sia, appare evidente che il gene che produce l’agata sia soggetto a diverse mutazioni. Non oso dire molto di più, ma questo aspetto va pure considerato, così come va ben valutato anche il fatto che il cosiddetto satiné nero abbia disegno sia pure debole. A complicare ulteriormente le cose c’è un vecchio ricordo, che ho già narrato e che risale a diversi anni or
sono. In effetti, molto tempo fa ho visto dei canarini che sembravano satiné scarsi, con disegno un poco debole, ma il fatto è che alcuni di loro avevano il cosiddetto sottopiuma (parte vaporosa) marroncino, non ricordo bene la tonalità comunque abbastanza chiara, altri nero o quasi. Si sarebbero detti isabella ed agata, anche perché il ceppo da cui provenivano era costituito da agata ed isabella buoni o ottimi. Dai dati genealogici forniti dall’allevatore, il comportamento genetico appariva non legato al sesso ma autosomico. Di conseguenza, se la valutazione fosse stata esatta, non potevano essere satiné e neppure mutazione allelica. Successivamente andarono dispersi, anche se ogni tanto qualche strano, troppo strano, satiné salta fuori e se ne sente parlare. Conclusione finale: un bel ginepraio! Da affrontare con rigore, senza nulla concedere ad idee preconcette.
Satiné intenso giallo, foto: E. del Pozzo
Satiné mosaico giallo femmina, foto: E. del Pozzo
Phaeo: puntualizzazioni
Bisognasempre tenersi aggiornati e riferire di cose insolite. In biologia talora vi sono imprevisti, evoluzioni od involuzioni che vanno considerate, certo senza perdere il senso dei fatti acquisiti. Recentemente sono andato in visita presso l’allevamento del mio amico e concittadino Nadino Ghillani, noto giudice internazionale, ove ho potuto ammirare una serie di tipi davvero di notevolissima qualità. Ebbene, ho notato anche qualcosa di molto insolito, vale a dire un paio di femmine nere phaeo mosaico rosso senza melanina centrale nella coda, fra l’altro ottime.
Come qualcuno ricorderà, recentemente - nel numero di I. O. 11/12 del 2024 nell’articolo sul “Nero phaeo” - ho parlato della persistenza della melanina centrale nelle timoniere, salvo casi rarissimi; ho parlato del 99,9% anche provocatoriamente ma dopo questa osservazione dovrò rivedere verso il basso tale percentuale. Mi sono posto dei perché ma non ne ho trovati molti. Ho dovuto pensare ad un nuovo equilibrio della situazione biologica, che forse consente una maggiore espressione phaeo almeno in quel ceppo. Comunque mi è stato riferito essere figlie di un maschio nero phaeo e di femmina bruna portatrice. Altri mi hanno detto di phaeo bruni con occhio apparentemente nero. Circostanza mai verificata da me in passato, anche recente. Nello scritto suddetto parlavo pure di questo aspetto. Non avendo verificato di persona devo necessariamente rimanere in una
posizione interlocutoria, anche se ritengo queste descrizioni come fondate. In questo caso però più che ad un nuovo equilibrio, come nel primo caso, penso che dovrei propendere più facilmente verso una perdita di tipicità, senza escludere l’aggiunta di qualcosa. In effetti un ulteriore pensiero che mi viene è che, come probabilmente accade nei Bruni opale, forse anche nei Phaeo potrebbe esserci stato un cambiamento per l’inserimento di qualcosa. Ho ipotizzato l’intervento di un fattore di inscurimento e rimango orientato verso questa ipotesi. In effetti i phaeo di oggi sono molto più spinti come bruno rispetto a quelli del passato, anche se la differenza è minore rispetto a quello che accade nei bruni opale di oggi rispetto a quelli di ieri. Inoltre nei phaeo si ha qualcosa di strano nel dicromatismo sessuale, almeno secondo mie osservazioni. Il dicromatismo, pur mantenendosi, appare un poco meno evidente rispetto a prima. La forte spinta verso il bruno ha portato ad avere maschi molto bruni. Inoltre, in alcuni casi, la concentrazione della feomelanina periferica rende i maschi con un orlo bruno più accentuato come tonalità, anche se meno ampio. Non parlo dei disastri indotti dai meticciamenti, tipo i lumps e non solo, poiché la circostanza è ben nota e da molti, me compreso, segnalata a diversi livelli. Stando così le cose spero in qualificati interventi dei cultori di questo tipo.
Giovanni Canali
Giallo, arancione e rosso… dall’ipotesi alla conferma
testo di FRANCESCO FAGGIANO, SIMONE OLGIATI con la collaborazione del CLUBDEL DIAMANTE CODALUNGA & POEPHILA,
Introduzione
Il tempo aggiusta molte cose, ma solo se seguiamo e perseguiamo le giuste strade, solo se all’intuizione facciamo seguire un approccio empirico, solo se alle convinzioni personali diamo prove inconfutabili e verificate possiamo dire di aver fatto il dovuto. Già nel numero 11 del 2022 scrissi un articolo abbastanza accurato sulla presumibile mutazione a “lipocromo arancio” del codalunga, che voleva mettere in evidenza come l’opinione comune sul fenotipo a lipocromo arancio in questa specie fosse sbagliata, frutto di opinioni e non di prove scientifiche e che in modo discriminante si accettava in alcune specie, vedi il guttato, e non si concepiva in altre, come appunto nel codalunga. Già allora era stato semplice ricercare articoli scientifici che riportavano scoperte e basi genetiche inconfutabili che non solo sostenevano la teoria e l’ipotesi formulata, ma che smentivano addirittura fondamenti della classificazione dei Poephila, una classificazione datata e basata sull’osservazione personale, sulla quale nel tempo fiumi di inesattezze erano state scritte da ornitologi D. codalunga Hecki a becco arancio e classico a confronto, all. Faggiano, foto: E. Mello, all. F.Faggiano
Oggi possiamo affermare con certezza ed orgoglio che come nel D. di Gould esista un polimorfismo del lipocromo
e poi da ornicoltori rispetto all’origine della colorazione rossa o arancio del becco del D. codalunga, rispetto al quale oggi possiamo affermare con certezza ed orgoglio che come nel D. di Gould esista un polimorfismo del lipocromo che ha permesso di identificare una forma tipo e una sottospecie, di cui Simone Olgiati ci fornisce di seguito accurata descrizione. A sostegno di tali affermazioni, oggi adduciamo alla ricerca precedente un risultato ibridologico che dimostra anche ai più reticenti che il fenotipo a lipocromo arancio nel Diamante codalunga presente in diversi aviari, come in tutti gli altri estrildidi, è il risultato di una mutazione di un gene maggiore, ubiquitario e responsabile, nella forma tipo, di produrre il pigmento rosso. Quando questo gene non fun-
foto EMANUELE MELLO
ziona, i carotenoidi non vengono ossidati e gli uccelli depositano sulla livrea, compreso il becco, pigmenti giallo-arancione. A conferma di ciò si adduce però una nuova ipotesi che in parte rimarrà nuovamente irrisolta: la mutazione a lipocromo arancio è una retromutazione, ovvero un tentativo della sottospecie Hecki di recuperare un’arcaica forma tipo, o è solo una mutazione a sé?
L’ibrido come prova inconfutabile
Se l’ibridazione per alcuni è un abominio, perché andrebbe ad inquinare il patrimonio genetiche delle specie, in realtà un’osservazione attenta ci rivela che questo è un fenomeno naturale che nell’evoluzione delle specie ha sempre avuto un ruolo fondamentale... Addirittura, gli ultimi studi sull’evoluzione della nostra specie Homo sapiens indicano l’ibridazione con altri ominidi o altre specie congeneri come il Neanderthal momenti fondamentali del percorso evo-
Ci si è accorti che l’ibridazione poteva diventare un vero e proprio testing per stabilire l’identità delle mutazioni
lutivo. In ornicoltura la pratica dell’ibridazione, ovvero dell’accoppiamento tra specie diverse, nasce come atto mistico, con la volontà di realizzare qualcosa di mitologico, risultato dalla fusione della bellezza di due diverse specie. Ben presto, però, si è capito che l’ibridazione indica in primo luogo la vicinanza genetica tra le diverse specie, anche se questo concetto è stato più volte rivisitato dalla scienza attraverso tecniche di ricombinazione genetica e l’analisi del DNA mi-
tocondriale, cioè quello più antico. Poi ci si è accorti che l’ibridazione poteva diventare un vero e proprio testing per stabilire l’identità delle mutazioni. Accoppiando specie diverse interessate da livree mutanti simili, l’ottenimento di progenie ibrida mutante (fatti salvi i rapporti di dominanza e la tipologia di ereditabilità del carattere), significa chiaramente e senza dubbi che entrambe le specie posseggono lo stesso gene e la stessa mutazione, altrimenti i caratteri complementano e da due mutati nascono figli con fenotipo classico, alias ancestrale. È questa una regola universale per tutti gli esseri viventi del nostro pianeta che ha solo un piccolo limite: identifica l’ubiquità del gene tra le specie ibridate, ma poco ci dice sui rapporti di allelia. Per essere chiari, se accoppio un canarino topazio con un messicano feo otterrò il 100% di figli ibridi mutati, tutti intermedi. Il gene interessato tra le due specie è lo stesso, ma la forma allelica no! E di forme alleliche ce ne possono essere molte… rimane però il fatto che nel dubbio e nell’esigenza di identificare se di mutazione si tratta e di quale mutazione si tratta, l’ibridazione della specie con specie affine di cui è già nota l’identità genetica è un percorso sicuro e inconfutabile. Ecco perché era necessario ottenere ibridi tra codalunga a lipocromo arancio e specie affini in cui era già nota la mutazione, che nel Poephila poteva essere stata identificata erroneamente dagli ornitologi del tempo come forma primitiva e quindi forma tipo di un presunto modello più evoluto a becco rosso descritta come sottospecie Hecki, mentre il modello originario sembrerebbe proprio quello a lipocromo rosso perché l’espressione giallo/arancio è, appunto, espressione di un mancato funzionamento del gene la cui funzione è proprio ossidare i carotenoidi. D’altronde, se una specie in natura presenta solo il fenotipo a lipocromo rosso, la comparsa di un fenotipo a lipocromo giallo-arancio sarà per forza di cose frutto di un evento mutante che crea una deficienza… come è successo per molte specie di selezione domestica. Per il codalunga, il Gould e addirittura il quadricolore la presenza nella popolazione ancestrale delle due espressioni lipocromiche è
Ruficauda mutato a becco arancio e figlio ibrido a becco dello stesso colore, foto: E. Mello, all. F.Faggiano
stata interpretata di volta in volta diversamente, ma, mentre nel Gould e nel quadricolore ci si era salvati parlando di polimorfismo (che sottintende la presenza di forme genetiche diverse), nel Poephila non è ben chiaro perché si sia parlato di sottospecie.
Il bias culturale
A fronte della documentazione e dell’elaborazione di un’ipotesi solida presentata nell’articolo di I.O. nel 2022, il mondo dell’ornicoltura non ha accolto tale proposta e addirittura, in sede di esposizione, alcuni soggetti esposti vennero declassati. Nulla di grave, certo, ma la necessità di dare conferma all’ipotesi e restituire dignità a questi soggetti diventò prioritaria, perché era evidente che la documentazione letteraria non fosse sufficiente a far comprendere la cosa. Ci stupiva il fatto che si desse più peso alla credenza popolare, basata sulla lettura di non si sa bene quali vec-
Ibrido Ruficauda a lipocromo arancio x Codalunga Hecki portatore di becco arancio, foto: E. Mello, all. F.Faggiano
chi libri di ornitologia in cui si sosteneva che in natura esistesse il fenotipo a lipocromo rosso, quello a fenotipo giallo ed uno intermedio a lipocromo arancione, eventuale condizione, questa, di totale eterogeneità di una specie difficilmente spiegabile scientificamente. Ma eravamo sicuri che le resistenze culturali e personali sarebbero potute sciogliersi al sole unicamente davanti a una prova inconfutabile che non desse adito a nessun dubbio: un ibrido con altre specie interessato dallo stesso fenomeno, ovvero dalla mutazione “lipocromo arancio”. Gli uomini funzionano in questo modo, a volte ipergeneralizzano e hanno pregiudizi… ad esempio, ritenere che tutti gli uccelli possano essere giudicati analiticamente con la stessa scala valori, perché giudicare un verzellino, una tortora o un codalunga è la stessa cosa; d’altro canto, se una specie come il D. Gould può avere un polimorfismo dei lipocromi su base mendeliana (trasmissione ereditaria del carattere con esatto rapporto tra dominante-recessivo), altre come il codalunga no!
La prova Ci siamo adoperati per riuscire a far nascere l’ibrido che confermasse la nostra ipotesi o, di contro, desse ragione a chi “ci faceva i sorrisini”… gli stessi che oggi ci dicono che lo sapevano, ma questa è un’altra storia. Abbiamo recuperato un Diamante mandarino portatore di becco a lipocromo arancio, un D. guttato e una femmina di ruficauda entrambi a lipocromo arancio; purtroppo, una serie di eventi sfortunati non ci ha portati ad ottenere nessun ibrido nel 2023. Poi nel 2024 la fortuna ha girato dalla nostra parte, anche grazie alle dritte del maestro Gabriele Ragni, grande ibridista che ringraziamo, e finalmente abbiamo avuto da una coppia di ruficauda formata da maschio a lipocromi arancio per codalunga femmina Hecki portatrice di lipocromi arancio (soggetto con becco perfettamente rosso) due ibridi: un maschio nero-bruno e una femmina agata entrambi a becco arancio.
Conclusioni e prospettive del Club dei Poephila
La gente funziona in questa maniera: crede nell’inverosimile e fa fatica a cre-
dere alla scienza, forse perché la scienza va compresa, altrimenti diventa un dogma di fede. Oggi abbiamo avuto la forza e la perseveranza di superare il pregiudizio e addurre all’ornicultura un piccolo sapere in più rispetto all’universo di cui godiamo facendo ornicoltura, sperando che gli addetti ai lavori valorizzeranno questo impegno al fine di perseverare nella mission di tutela e preservazione di ogni forma di biodiversità da noi allevata attraverso gli strumenti del nostro hobby, che si cristallizzano nel dare un’identità e un indirizzo selettivo ad ogni fenotipo. Come Club ci auguriamo in primis di riuscire a stabilizzare e diffondere questo genotipo, determinando le sue peculiarità e le sue potenzialità e dando il rispetto e il sostegno che meritano a questi soggetti e agli ornicoltori che se ne occupano. Poco importano nuove categorie in questo momento, ma la sicurezza che un nuovo fenotipo del Diamante codalunga venga identificato e trattato con il dovuto rispetto in sede espositiva è nostro dovere, per questo ringraziamo chi vorrà accogliere il nostro appello.
In un articolo del 2018 (Hooper et al.) si legge che tra le due sottospecie esistono alcune differenze sul cromosoma sessuale
Quale selezione
Riteniamo che un orientamento semplice e pragmatico sia possibile ed efficace, considerando che le evidenze di allevamento ci dicono che difetti della colorazione del becco degli Hecki sono indipendenti dalla presenza del gene mutante. Un Hecki avrà un becco con lipocromo difettato troppo chiaro o con attaccatura aranciata, indipendentemente dal fatto che sia portatore o imparentato con soggetti a lipocromo arancio. Mentre vanno premiati soggetti Hecki a becco rosso o becco arancio che presentino il becco con lipocromo saturo fino alla base, rosso co-
rallo nella forma classica, arancio ocra nella forma mutante.
Approfondimento scientifico sulla filogenesi del Codalunga (a cura di Simone Olgiati) Ho letto con molto interesse l’articolo dell’amico Francesco Faggiano riguardo l’avvenuto test di ibridazione tra il Diamante codarossa agata a lipocromo arancio e il Diamante codalunga becco rosso eterozigote per il fattore, così definito dall’autore, “becco arancio”. Il video che mi è stato inoltrato mostrava inequivocabilmente l’allelia tra i due fattori presi in considerazione. Nella ripresa veniva rappresentato un ibrido maschio caratterizzato dall’avere becco e maschera di color giallo-arancio, sull’ocra, molto simile a quello paterno e ai D. codalunga mutati “becco arancio”. Se la prova d’ibridazione si è rivelata inconfutabile e incontrovertibile, sulle ipotesi filogenetiche proposte tra specie tipo e sottospecie Hecki ho qualche perplessità. Questo anche alla luce di ricerche personali svolte sulla sistematica del genere Poephila e sul funzionamento del gene CYP2J19, codificante per il citocromo P450, nei viventi. Negli articoli consultati e citati in bibliografia non è stato trovato riscontro della teoria per cui nel Diamante codalunga non si hanno sottospecie e che il fenotipo a becco giallo sia dovuto ad un fenomeno di polimorfismo, come accade nel D. di Gould e nel D. quadricolore. Se la teoria del polimorfismo fosse vera, avremmo fenotipi a becco giallo e a becco rosso mescolati all’interno della stessa area e con una popolazione geneticamente omogenea, come accade nel Diamante di Gould. Al contrario, sussistono differenze genetiche, etologiche e geografiche nette ed evidenti che stabiliscono l’esistenza di due sottospecie nel Diamante codalunga: quella nominale, acuticauda, e la Hecki.
In un articolo del 2018 (Hooper et al.) si legge che tra le due sottospecie esistono alcune differenze sul cromosoma sessuale. In particolare, si fa riferimento al fatto che tra la sottospecie nominale e la Hecki circa il 75% del cromosoma Z è differente a causa di una o di due inversioni cromosomiche. L’inversione cromosomica è una mutazione che comprende
un lungo tratto di DNA. Tale mutazione consiste nella rottura del filamento di DNA in due punti; il frammento così ottenuto viene reincorporato, grazie alla riparazione ad opera di specifici enzimi (come le DNA ligasi), nel cromosoma, ma viene invertito di orientamento. Questo rende così più difficoltoso l’appaiamento di due cromosomi Z diversi e lo scambio di geni in queste regioni di DNA. In altre specie si è visto come le inversioni cromosomiche fungano da barriere efficaci per evitare l’incrocio tra popolazioni confinanti riducendo la fertilità della prole ibrida (Knief et al., 2024). Nella discussione del suddetto articolo si può leggere come le differenze di colore del becco tra le due sottospecie non riguardino solo geni autosomi ma anche geni collocati sul cromosoma Z, quindi sessolegati e non solo recessivi, come ipotizzato da Francesco Faggiano. In più paper (Jennings et al., 2005; Edwards et al., 2005) viene messo in evidenza come la speciazione, cioè la nascita di nuove specie o sottospecie, su base molecolare, tra la sottospecie acuticauda e quella Hecki sia avvenuta circa 340.000 anni fa, alla fine del periodo interglaciale Mindel-Riss e all’inizio della glaciazione Riss. Sebbene l’espansione dei ghiacciai sia stata visibile soprattutto nell’emisfero Nord, l’emisfero Sud ha comunque risentito degli effetti del raffreddamento climatico: temperature più basse, maggiore aridità, minori precipitazioni, abbassamento del livello del mare, spostamento dei monsoni. La separazione geografica tra le due sottospecie è netta lungo una linea ben evidente, rappresentata dal Kimberley Plateau e dalla Terra di Arnhem. Sulla linea di confine è possibile osservare qualche esemplare ibrido, così come accade in molte zone di confine tra sottospecie diverse in altre specie, per esempio negli Spinus sudamericani. Nel 2023 McDiarmid et al. hanno esaminato la preferenza sessuale delle femmine di Diamante codalunga. Si è scoperto che le femmine della sottospecie nominale tendevano a scegliere esclusivamente maschi conspecifici e lo stesso fenomeno si ripeteva con quelle della sottospecie Hecki. Quando i becchi dei maschi venivano colorati in modo diverso (acuticauda con becco colorato
Nel 2023 McDiarmid et al. hanno esaminato la preferenza sessuale delle femmine di Diamante codalunga
di rosso ed Hecki con becco colorato di giallo) le femmine tendevano a scegliere comunque il maschio appartenente alla propria sottospecie, sebbene il colore del becco fosse stato alterato. Tra i due taxa esistono quindi differenze non solo a livello di becco, ma anche in dettagli che visivamente non sono così immediati, come possono essere il canto, la posizione di corteggiamento o altro. Nello stesso articolo si può vedere, rappresentato nella figura 1, il processo di introgressione in atto dalla sottospecie acuticauda all’Hecki. Quello che sta avvenendo è che, piano piano, in una data regione, le due popolazioni
si stanno mescolando e alcuni geni di una sottospecie stanno entrando a far parte del genoma dell’altra. Inoltre, gli autori hanno evidenziato come la direzione di accoppiamento influisca sul fenotipo delle femmine F1. Tra i maschi e le femmine nati da maschio Hecki x femmina acuticauda non si hanno sostanziali differenze nel colore del becco, mentre le femmine F1 di acuticauda maschio x Hecki femmina hanno il becco decisamente giallastro piuttosto che arancio. Questo concorda con l’articolo di Hooper che indica differenze tra le due sottospecie anche sul cromosoma sessuale e non solo sugli autosomi e sottolinea come la genetica del becco del Diamante codalunga sia più complessa di quanto prospettata. Altro studio a sostegno dell’esistenza di due sottospecie è del 2017 (Griffith et al.), nel quale si evidenziano le dimensioni dell’area in cui si trovano fenotipi intermedi. Tale zona è larga appena 150 km, implicando delle barriere pre-zigotiche molto robuste tra le due sottospecie, che ne impediscono il rimescola-
Diamante codalunga Hecki classico e a becco arancio, foto: E. Mello, all. F.Faggiano
mento. Però sono avvenuti degli incroci tra le due sottospecie in un periodo compreso tra i 21.000 ed i 14.000 anni fa, quando le barriere fisiche tra i due gruppi sono state abbattute. Ciò è avvenuto perché si è formato un corridoio di ambienti più umido di quello di oggi, grazie allo spostamento dei monsoni in quella regione rispetto all’attuale. Si è ampliato l’habitat ideale delle due sottospecie e queste si sono mescolate, in parte, solo nella regione in cui si sono ritrovate in contatto. Questo intervallo di tempo corrisponde all’ultimo massimo glaciale in Europa, allo stadio Würm IV. Una volta che la barriera geografica si è ripristinata, le due popolazioni non si sono più mescolate se non sulla linea di confine tra gli areali delle due sottospecie. Non si è formato un enorme “sciame ibrido” ma gli acuticauda e gli Hecki hanno mantenuto la propria identità genetica.
L’Ibrido di Francesco Faggiano testimonia in modo inequivocabile che in alcuni ceppi italiani di Diamanti codalunga della sottospecie Hecki “serpeggia” una mutazione recessiva autosomica allelica al fattore “lipocromo arancio” del D. codarossa e, molto probabilmente, allelica alle altre forme “gialle” che abbiamo in
specie di Esotici a fattore rosso o a fattore misto come D. guttato, D. mandarino, Estrildidi africani, ecc. Sulla teoria del polimorfismo, alla luce di quanto riportato, non posso concordare perché non esiste bibliografia che sostenga questa ipotesi. Al contrario, molti paper e articoli supportano l’esistenza di due sottospecie distinte dei Diamante codalunga, le cui differenze non riguardano solo il colore del becco ma altri aspetti meno ovvi e visibili.
Le mie ipotesi sull’origine del fenotipo “becco arancio” dei Diamanti codalunga di Francesco Faggiano e di Daniele di Bella sono due. La prima si ricollega ai paper citati e riassunti in precedenza. È possibile che in cattività siano avvenuti, anni or sono, incroci tra alcuni acuticauda ed alcuni Hecki; nel corso degli accoppiamenti è avvenuta l’introgressione di uno dei geni codificanti per il lipocromo giallo dalla sottospecie nominale all’altra. In questo caso il gene che è “entrato” nell’Hecki è quello autosomico recessivo, allelico a quello del Diamante codarossa. Se questa ipotesi fosse vera, ci sarebbe da chiedersi perché gli altri geni non siano stati passati dall’acuticauda all’Hecki. Probabilmente quei soggetti che manifestavano un fenotipo giallo-arancio
sono stati scartati dalla selezione, mentre si sono riprodotti gli esemplari a becco rosso ma portatori di uno dei geni per il “becco arancio”. L’altra ipotesi consiste nel fatto che in un dato ceppo di Diamanti codalunga della sottospecie Hecki è comparsa quella mutazione autosomica recessiva comune alle specie nominate nel paragrafo precedente, probabilmente variante del gene CYP2J19. Concludo condividendo l’auspicio di studiare con maggiore attenzione questo fenotipo del Diamante codalunga Hecki in allevamento, cercando di capire come valorizzare tale fenotipo e, soprattutto, come eventualmente differenziarlo rispetto ad un D. codalunga acuticauda col becco “troppo carico”.
Replica (a cura di Francesco Faggiano) Ho apprezzato l’approccio sistemico e accademico di Simone Olgiati rispetto all’ipotesi dell’errore tassonomico da me sottoscritto e, pur adducendo lui tutta una serie di elementi ed indizi autorevoli, rimane in me un’incertezza. Su più fronti si considerano ere geologiche, inversioni genetiche e preferenze fenotipiche che confermano i processi di speciazione del codalunga, eppure nello stesso tempo non mi spiegano attra-
Da sinistra D. ruficauda a lipocromo giallo, Codalunga Hecki a lipocromo arancio e ibrido Ruficauda x Codalunga Hecki a becco arancio, foto: E. Mello, all. F.Faggiano
verso l’identificazione del mutante a lipocromo arancione che il test ibridologico ci certifica come in una specie la forma tipo non abbia un gene ubiquitario quale il CYP2J19, mentre la sottospecie non solo la possiede, ma ha espresso una forma allelica tipica di altre specie; non è più semplice pensare che i “becco giallo” siano la sottospecie che proprio a causa dei fenomeni di speciazione ha perso il gene del becco rosso e che le mutazioni sessolegate abbiano fatto il resto? Sicuramente in futuro Simone Olgiati potrà addurci altre conoscenze e risolvere egregiamente la questione, ma oggi è importante acquisire che il becco arancio anche nel codalunga Hecki è una mutazione!
BIBLIOGRAFIA:
•Evolution, 59(9), 2005, pp. 2033–2047, Speciational History Of Australian Grassfinches (Poephila) Inferred From Thirty Gene Trees, W. Bryan Jennings And Scott V. Edwards
•Hooper DM, Griffith SC, Price TD, Sex chromo-
Oggi è importante acquisire che il becco arancio anche nel codalunga Hecki è una mutazione
some inversions enforce reproductive isolation across an avian hybrid zone. Mol Ecol. 2019; 28: 1246-1262.
•Australian Journal of Zoology, 2011, 59, 270–272, Biogeographic barriers in north-western Australia: an overview and standardisation of nomenclature, Mark D. B. Eldridge, Sally Potter and Steven J. B. Cooper
•Proceedings of the National Academy of Sciences · June 2005, Speciation in birds: Genes, geography, and sexual selection, Scott V. Edwards, Sarah B. Kingan, Jennifer D. Calkins, Christopher N. Balakrishnan, W. Bryan Jennings, Willie J. Swanson, and Michael D. Sorenson
•BMC Evolutionary Biology (2018) 18:22, Evolution of CYP2J19, a gene involved in colour vision and red coloration in birds: positive selection in
the face of conservation and pleiotropy, Hanlu Twyman, Staffan Andersson and Nicholas I. Mundy
•Current Biology 32, 4201–4214.e1–e12, October 10, 2022, A mechanism for red coloration in vertebrates, Matthew B. Toomey, Cristiana I. Marques, Pedro M. Araùjo, Delai Huang, Siqiong Zhong, Yu Liu, Gretchen D. Schreiner, Connie A. Myers, Paulo Pereira, Sandra Afonso, Pedro Andrade, Malgorzata A. Gazda, Ricardo J. Lopes, Ivan Viegas, Rebecca E. Koch, Maureen E. Haynes, Dustin J. Smith, Yohey Ogawa, Daniel Murphy, Rachel E. Kopec, David M. Parichy, Miguel Carneiro, and Joseph C. Corbo
•McDiarmid C. S., Finch F., Peso M., van Rooij E., Hooper D. M., Rowe M. & Griffith S. C. (2023), Experimentally testing mate preference in an avian system with unidirectional bill color introgression, Ecology and Evolution, 13, e9812.
•EMU - AUSTRAL ORNITHOLOGY, 2017, Geographical variation in bill colour in the Long-tailed Finch: evidence for a narrow zone of admixture between sub-species, Simon C. Griffith and Daniel M. Hooper
•Molecular Biology and Evolution, Volume 41, Issue 6, June 2024, Evolution of Chromosomal Inversions across an Avian Radiation, Ulrich Knief, Ingo A. Müller, Katherine F. Stryjewski, Dirk Metzler, Michael D. Sorenson, and Jochen B.W. Wolf
Master sul Canarino Salentino
testo
e foto di SERGIO
PALMA
In data 6 Gennaio 2025, presso il Museo Civico di Storia Naturale di Calimera, si è tenuto il 1° Master sul Canarino Salentino. Calimera (buongiorno in greco) è un Comune in Provincia di Lecce, inserito nella Grecia salentina. La Grecia salentina è un'isola linguistica ellenofona del Salento, è consistente in nove comuni della stessa provincia in cui si parla un dialetto neo-greco noto come grico. La manifestazione è stata un vero successo tecnico, dove con la presenza del Giudice Internazionale Adriano Bertan (esaminatore) e il collega Vulzio Lazzarini quale moderatore e relatore, oltre a dare lustro al Master, ha dato modo di dibattere e confrontarsi sulle caratteristiche del Salentino e su quelle che sono le prospettive di selezione genetica, nonché gli accorgimenti da prendere a livello burocratico per far sì che la razza si migliori sempre di più. Durante la manifestazione si è avuta la gradita sorpresa della presenza di un gruppo di bambini che erano presenti al museo per uno dei tanti progetti che si svolgono all’interno e che prevedono la trattazione di argomenti quali la natura, la salvaguardia dell’ambiente e la divulgazione scientifica. Una vera e propria terapia alla frenesia dei nostri tempi, di cui proprio i più piccoli sono vittime. L’importanza dell’integrazione sociale e dell’educazione può passare anche attraverso la conoscenza degli uccellini.
Negli ultimi anni, l’integrazione e l’inclusione sociale sono diventate te-
matiche fondamentali in molte comunità. Un esempio significativo di questo approccio è rappresentato da un progetto che coinvolge i bambini nella conoscenza di simpatici uccellini, in particolare i canarini. Questo progetto non solo offre un’opportunità di apprendimento ma si inserisce
La manifestazione è stata un vero successo tecnico
anche in un contesto di turismo accessibile e di comunità, contribuendo alla fruizione dei servizi turistici, tra cui un museo dedicato. La partecipazione dei bambini a questo progetto va oltre la semplice osservazione degli uccellini. Essa rappresenta un’esperienza educativa che promuove valori di pazienza, passione e impegno. Questi valori sono essenziali non solo per gli allevatori, che dedicano tempo e cura per garantire il benessere degli animali, ma anche per i bambini stessi che imparano l’importanza di prendersi cura di un essere vivente. In un’epoca in cui la frenesia della vita moderna e l’in-
Gruppo di allevatori del Salentino
fluenza dei social media possono portare a una certa superficialità, questo progetto offre un’opportunità preziosa per riscoprire l’importanza della dedizione e della responsabilità. Inoltre, il progetto si distingue per il suo aspetto inclusivo. Coinvolgendo bambini di diverse provenienze e abilità, si promuove un ambiente di apprendimento collaborativo, dove ognuno può contribuire e apprendere dagli altri. Questo non solo favorisce l’integrazione sociale ma crea anche un senso di comunità, in cui i partecipanti si sentono parte di qualcosa di più grande. La conoscenza degli uccellini diventa così un mezzo per costruire legami e relazioni significative tra i bambini, incoraggiando la comunicazione e la comprensione reciproca. Il gruppo di infanti era guidato dal Dr. Potenza, Biologo presso l’Università del Salento, che a seguito di domande specifiche da me poste, così si è espresso sul Salentino: 1) “Estremamente caratterizzante con delle peculiarità uniche che la contraddistinguono”; 2) “Non è emerso nessun elemento di disturbo riguardo al fatto che oggi per moltissimi bambini l’unico modo di entrare in contatto con animali che non siano cani e gatti, sono proprio i musei e questo tipo di manifestazioni”; 3) “Domande particolari non ci sono state”. 4) L’ultima mia domanda, è stata quella di capire se l’Unisalento fosse interessata al progetto Salentino: “Nel dipartimento non ci sono ornitologi e l’ornitologia al momento non è molto “praticata”, tanto è vero che spesso si affida al Museo per sviluppare alcuni progetti o avere delle consulenze in tal senso. Diciamo che le due strutture, relativamente ai vari campi di studio, si compensano ed interagiscono vicendevolmente. Tuttavia posso indagare. A mio parere potrebbe essere interessante dal punto di vista genetico e filogenetico, ma anche per quel che concerne i tratti funzionali e gli adattamenti comportamentali”. Speriamo bene, attendiamo risposta. Infine, l’aspetto turistico del progetto non deve essere sottovalutato. La possibilità di visitare un museo dedicato a questi uccellini offre un’oppor-
tunità unica per le famiglie e i turisti di avvicinarsi a una realtà spesso trascurata. Attraverso visite guidate e attività interattive, il museo diventa un luogo di apprendimento e scoperta, dove la bellezza della natura e l’importanza della sua conservazione vengono messe in risalto. In conclusione, il progetto che coinvolge i bambini nella conoscenza degli uccellini rappresenta un esempio luminoso di come l’integrazione sociale, l’educazione e il turismo possano unirsi per creare esperienze significative. In un mondo in cui le caratteristiche fondamentali come pazienza, passione e impegno rischiano di essere trascurate, iniziative come questa ci ricordano l’importanza di coltivare valori autentici e di costruire comunità inclusive e consapevoli.
Durante la manifestazione si è avuta la gradita sorpresa della presenza di un gruppo di bambini che erano presenti al museo
Veniamo ora alla parte della manifestazione che vede sia l’aspetto divulgativo che quello strettamente tecnico. Sui vari caratteri del Salentino, si è discusso confermando molti punti che stanno dando i loro frutti nell’arte della selezione, uno in particolare la forma del corpo che, nella parte dello sterno non deve avere prominenze (moltissimi soggetti mancavano di petto prominente). Altra caratteristica che è ad un buon punto selettivo sono le spalle, che sono sufficientemente strette; qualche soggetto, però, aveva i carpi leggermente più alti di quanto desiderato. Si è dovuto però constatare come diversi soggetti, forse troppi, mostravano una taglia-lunghezza esagerata per i dettami dello standard.
Il Dr. Lazzarini, infatti, ha lanciato, prima all’Assemblea dei Presenti e quindi al Consiglio Direttivo del Club, di esaminare la possibilità di una modifica ed inversione delle voci sulla scheda di giudizio, portando al primo posto con punti 25 proprio la voce “taglia”. La proposta ha trovato unanime consenso tra i presenti, ma questo compito spetta prima ai dirigenti del Club e poi alla relativa CTN che era presente con il Giudice e Socio del Club Carmelo Caroppo.
Dopo quattro anni di soddisfazioni per quanto riguarda lo sviluppo e la divulgazione della nostra passione per il Salentino, il giorno 6 Gennaio 2025, in occasione del primo Master sul Canarino Salentino, si sono svolte le elezioni per il rinnovo del Consiglio Direttivo del Club Amici del Salentino. Lusingati della fiducia che ci è stata concessa, ringraziamo il consigliere uscente Sig. Carmelo Caroppo per il lavoro che ha svolto.
Il CAS, come sempre, darà supporto in materia di approfondimento tecnico a tutti gli amanti del Salentino, rispondendo a quesiti e chiarimenti da parte di tutti gli amici allevatori del Salentino e anche da parte degli amici che vogliano avvicinarsi a questa Razza.
Il Club Amici del Salentino comunica che, a seguito delle elezioni svoltesi il giorno 6/1/2025, il nuovo
Consiglio Direttivo è così composto:
Presidente
Alessandro Lezzi
E-mail: alessandrole@icloud.com
Cell. 338.1513999
E-mail Club: clubamicidelsalentino@gmail.com
Segretario
Marcello Iacovizzi
E-mail: marcelloiacovizzi@gmail.com
Cell. 338.8664702
Consigliere
Vulzio Lazzarini
E-mail: vulzio@gmail.com
Cell. 335.6363260
Facebook
Passione per il Canarino Salentino Il Presidente Alessandro Lezzi
F ocus C.T.N.
Nuove nomenclature
Aseguito di recenti studi e alla modifica nella classificazione di alcune specie, si porta all’attenzione degli allevatori il cambio di denominazione delle specie in elenco.
Per dare modo a tutti di recepire il cambiamento, si comunica che la nuova denominazione verrà riportata negli elenchi ufficiali FOI (vedi elenco per ordini anellini) a partire dalla stagione 2026.
Vecchie nomenclature Nuove nomenclature
Amazona xanthops. Alipiopsitta xanthops
Ara auricollis Primolius auricollis
Ara chloroptera Ara chloropterus
Ara couloni Primolius couloni
Ara Manilata Orthopsittaca manilatus
Ara Maracana Primolius maracana
Ara nobilis nobilis Diopsittaca n. nobilis
Ara severa Ara severus * * * * *
Aratinga acuticaudata Thectocercus acuticaudata a. acuticaudatus
Aratinga acuticaudata Thectocercus haemorrhous a. haemorrhous
Aratinga acuticaudata Thectocercus neumanni a. neumanni
Aratinga acuticaudata Thectocercus neoxenus a. neoxenus
Aratinga acuticaudata Thectocercus koenigi a. koenigi
Aratinga aurea Aurea Eupsittula aurea aurea
Aratinga brevipes Psittacara brevipes
Aratinga cactorum Eupsittula cactorum c. cactorum
Aratinga cactorum Eupsittula caixana c. caixana
Aratinga canicularis Eupsittula canicularis c. canicularis
Aratinga canicularis Eupsittula clarae c. clarae
Aratinga canicularis Eupsittula eburnirostrum c. eburnirostrum
Plumofagia nell’allevamento della Specie Serinus canaria
https://zenodo.org/doi/10.5281/zenodo.14610398
testo di PASQUALE LEONE, foto AUTORI VARI
Abstract
L’argomento della plumofagia inerente la specie Serinus canaria non trova al momento riscontri nell’ambito della letteratura scientifica.
La natura degli allevamenti della specie, di esclusivo interesse amatoriale e/o sportivo, porta come conseguenza il fatto di non porre le dovute attenzioni sulle peculiarità dell’allevamento, come succede invece per gli animali da reddito.
Questo scritto, pertanto, si pone l’obiettivo di creare una base sulla quale effettuare ulteriori approfondimenti a riguardo.
La poliedricità dell’assunto non permette al momento di valutare appieno tutte le singole ipotesi che sono causa del disturbo preso in esame.
Pertanto, gli aspetti patologici e quelli relativi alla genetica verranno soltanto menzionati, in quanto necessitano di evidenze e competenze non previste nell’ambito di questo studio. Del resto, va specificato che anche alcuni aspetti oggetto della disamina, considerata la carenza di evidenze scientifiche in materia, saranno supportati da studi comparati con specie affini.
In gergo, gli ornicoltori la chiamano “pica” anche se il termine più appropriato da usare è “plumofagia”.
In gergo, gli ornicoltori la chiamano “pica” anche se il termine più appropriato da usare è “plumofagia”
Il picacismo, altresì noto come allotriofagia o pica, nella scienza della nutrizione umana è considerato un disturbo del comportamento alimentare.
Questo disturbo è caratterizzato dall’ingestione di sostanze non nutritive ed è riconosciuto come tale quando si manifesta per un periodo di tempo uguale o superiore ad un mese.
Nel caso degli uccelli da gabbia e da voliera, le cause che provocano questo disturbo sono provocate da differenti motivi.
La piuma è la maggiore peculiarità di qualsiasi uccello. In particolare, per gli uccelli di affezione, un comportamento come quello della pica andrebbe preso in maggiore considerazione, fonte: PowerTemplate.com
Probabilmente l’unica correlazione che esiste tra il disturbo della pica diagnosticato negli esseri umani e quello individuato negli uccelli è inerente esclusivamente al termine col quale è stato identificato siffatto disturbo, ovvero il nome col quale è conosciuta la gazza, un uccello che ha l’abitudine di beccare degli oggetti non alimentari.
Definizione delle turbe comportamentali
Beccare è da considerarsi un comportamento naturale quando il fine è la ricerca di cibo o l’osservazione (in alcuni casi l’esplorazione) ambientale. Quando però un canarino inizia a beccare un conspecifico, è importante stabilire quali siano le motivazioni. Riuscire a capire la causa di questo comportamento è alquanto complesso, poiché le ragioni possono essere molteplici.
Va altresì posto in evidenza che, in queste circostanze, l’anamnesi fatta dallo stesso allevatore riveste un’importanza fondamentale per la susseguente diagnosi veterinaria poiché,
non necessariamente, le cause devono essere imputate ad una patologia intrinseca. Generalmente si ha la tendenza ad associare tali comportamenti a carenze alimentari, ma le motivazioni che sono alla base dell’anomalia comportamentale possono anche essere ricercate altrove.
Pertanto, nel caso in cui si manifestino in allevamento episodi di plumofagia, la raccolta di tutte le informazioni, le osservazioni o le eventuali note di allevamento sono fondamentali successivamente al medico veterinario per identificare una possibile patologia all’origine. Eliminare quindi tutte le ipotesi meno probabili, di fronte a situazioni in cui, nella quasi totalità, non si abbiano appositi esami diagnostici, è un compito del quale l’allevatore dovrebbe deontologicamente gravarsi.
Nel settore avicolo, la plumofagia viene suddivisa in due tipologie: dolce e grave.
Nella prima ipotesi, plumofagia dolce, può essere a bassa o a alta frequenza ed è mirata alle penne della coda. La plumofagia grave è quella
che, persistendo, cresce esponenzialmente fino ad arrivare a provocare ferite all’epidermide.
Per queste ragioni, è importante bloccare tale comportamento fin da quando viene osservato per la prima volta e separare i soggetti attaccati. La plumofagia, pur essendo una turba del comportamento, non va classificata nei cosiddetti Abnormal Repetitive Behaviour. Gli ARB si manifestano in due forme: quelli il cui scopo sembra essere il raggiungimento compulsivo di un obiettivo inappropriato e quelli la cui esecuzione è legata a una funzione motoria inappropriata. Dunque, entrambi i casi non rientrano in quello specifico della plumofagia.
Motivazioni della fenomenologia comportamentale
Tra i principali fattori eziologici e motivazionali che potrebbero essere scatenanti o causa originaria di questo comportamento si osservano:
• Densità e sovraffollamento Lo spazio territoriale nella specie Serinus canaria riveste particolare importanza, in special modo nelle fasi in cui i soggetti sono mantenuti in colonia e non in gabbie individuali o in coppia. La detenzione in spazi eccessivamente limitati, la carenza di mangiatoie, abbeveratoi, posatoi e, nel caso di nidi, una quantità non proporzionale al numero di soggetti, va adeguatamente valutata al fine di ridurre i motivi di attrito tra i singoli canarini.
• Emulazione
Può succedere che all’inizio la plumofagia possa essere praticata da un unico soggetto dominante o particolarmente vivace. A seguire, per emulazione, tale comportamento può dilagare nella voliera, causando le relative conseguenze dannose.
• Ordine di beccata
L’espressione “ordine di beccata” deriva dal fatto che in un gruppo o colonia di canarini esiste una gerarchia ben definita. I soggetti conviventi la conoscono bene e sanno esattamente chi comanda; da qui nasce l’etimologia del termine.
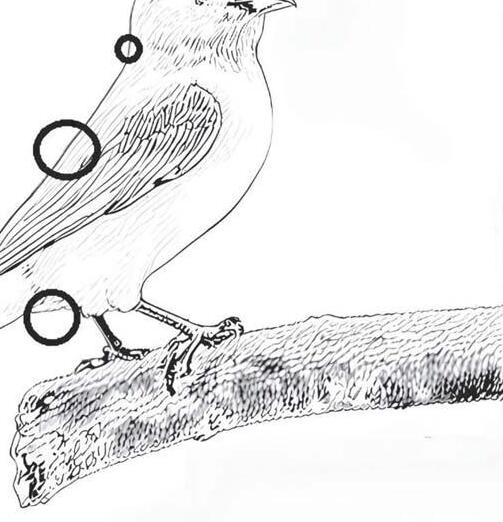
In evidenza, i punti in cui i soggetti più deboli vengono attaccati quando si verificano casi di plumofagia
Un comportamento comune in una voliera è l’aggressione o l’attacco ai nuovi soggetti da parte di quelli più anziani, anche se più grandi di pochi mesi o se presenti da maggior tempo nello spazio condiviso.
Questo poiché, in caso di nuove introduzioni nella voliera, è necessario che venga ristabilita la gerarchia tra i volatili che, come sopra menzionato, per indole sono molto territoriali. Emulazione e ordine di beccata sono fenomeni di interazione sociale. In genere riguardano un numero limitato di soggetti (due o tre) che attua tale comportamento. Soltanto successivamente la trasmissione si estende all’intera colonia.
• Fattori ambientali
È stato riscontrato che, ad una temperatura ambientale più elevata, la specie in questione è soggetta ad alterazioni comportamentali. Tali effetti possono portare a disfunzioni fisiologiche causate da un eccessivo sollecitamento del sistema nervoso. Questo porta ad una conseguente variazione del grado di tolleranza esistente tra i soggetti presenti nella colonia. Altro fattore da tenere in dovuta considerazione è la luminosità ambientale. L’esposizione dei soggetti ad un intenso grado di luce è uno dei motivi che possono causare questa anomalia comportamentale.
• Carenze nutrizionali
L’alimentazione è spesso il motivo principale della plumofagia poiché di fronte alla carenza di specifici nutrienti i canarini sono costretti a ricercarli nelle piume. Un regime alimentare sbilanciato porta sovente i soggetti alla ricerca delle proteine contenute nella piuma.
Carenze di amminoacidi quali la metionina, la lisina, la cistina o l’arginina, che tra le altre cose svolgono una funzione chelante, sono la causa primaria dell’apparire del disturbo. Strettamente correlata a questi elementi è la carenza di vitamine, minerali e oligoelementi che permettono la sintesi degli amminoacidi succitati e che coadiuvano il piumaggio ad acquisire la corretta resistenza ed elasticità tipica dei soggetti in salute.
Gli ARB si manifestano in due forme: quelli il cui scopo sembra essere il raggiungimento compulsivo di un obiettivo inappropriato e quelli la cui esecuzione è legata a una funzione motoria inappropriata
• Genetica
Da studi comparati su altre specie si evincono fattori genetici quali cause della plumofagia.
I primi risultati mostrano che la plumofagia è una caratteristica di tipo poligenico, influenzata cioè da più geni, ognuno dei quali apporta tanti piccoli effetti.
A seconda dei modelli statistici utilizzati, è risultato che l’ereditabilità può variare tra lo 0,11 e lo 0,20 in percentuale.
Non è tuttavia parte di questo studio approfondire questo aspetto.
• Malattia
Non solo le carenze nutrizionali provocate da errati regimi alimentari, ma anche il mancato assorbimento degli
stessi nutrienti per motivi patologici è causa del consequenziale disturbo. Anche questo aspetto non rientra però negli obiettivi di questo studio.
• Cannibalismo
È errato considerare a priori la plumofagia quale forma di cannibalismo. Il cannibalismo è da considerarsi tale soltanto quando l’atto di beccare inizia ad intaccare la parte dei tessuti cutanei. Pertanto, anche se la plumofagia è legata ad un primo stadio di cannibalismo, in virtù delle molteplici cause da cui può scaturire, non necessariamente vi va associata.
• Autodeplumazione
L’atto di un soggetto che becca le proprie piume viene identificato talvolta come l’inizio della plumofagia. Sebbene questo possa essere un segnale d’allarme, va considerato che alcuni canarini possono dedicarsi a un eccessivo preening. Nella maggior parte dei casi, si tratta di casi di overpreening ossia di un eccessivo tempo dedicato alla pulizia del piumaggio, con ragioni correlate per lo più a parassiti. Le restanti ipotesi rientrano nel patologico, ma non sono inerenti la plumofagia.
• Stress
Lo stress è un elemento scatenante la plumofagia.
Un esempio di featherpicking della zona cloacale, dove si riscontra un’ampia area rimasta deplumata
Diversi punti presi finora in esame sono da considerarsi motivo stressogeno per i canarini.
Discussione
Le cause scatenanti la plumofagia possono essere numerose e di diversa natura. Legate a motivi di salute, a questioni di genetica, a problemi ambientali o provocate da errori durante la gestione dell’allevamento.
Nella raccolta dei dati anamnestici è importante riuscire a stabilire il grado di aggressività della beccata.
La beccata leggera è la conseguenza di un normale comportamento d’interazione tra soggetti conspecifici o con lo stesso ambiente. Questo genere di beccata non deve essere contemplata come turba comportamentale e ricollegata alla plumofagia. Diverso invece è il caso della beccata grave (altresì conosciuta come feather picking), che è comportamento tipico della plumofagia.
In questa seconda ipotesi, i soggetti più deboli vengono attaccati sulle remiganti, sulla nuca, sul dorso e nella zona cloacale.
Nei casi più gravi, si arriva a beccare a sangue le cloache e l’attacco si può protrarre fino alle zampe, giungendo a provocare anche la perdita di qualche dito.
La plumofagia è di per sé un fattore di rischio per il benessere dei soggetti nell’aviario.
Il beccarsi nocivo può dunque arrivare a portare lesioni ed è quindi necessario evitare degenerazioni di tali comportamenti, apportando dovute misure correttive tempestivamente.
La beccata (anche quando si è di fronte a dei casi di plumofagia), considerata nel contesto dell’attitudine sociale, va individuata sia nella capacità del canarino nell’evitare di essere beccato (effetto vittima o effetto genetico indiretto) che in quello di beccare gli altri soggetti (effetto attore o effetto genetico diretto).
La plumofagia genera un aumento del consumo di cibo da parte dei soggetti deplumati, i quali provano in tal modo a stabilizzare il proprio metabolismo a seguito della dispersione
Nei casi più gravi, si arriva a beccare a sangue le cloache e l’attacco si può protrarre fino alle zampe, giungendo a provocare anche la perdita di qualche dito
del calore corporeo causato dall’assenza di piumaggio in alcune parti del corpo.
Alcuni test mirati a fornire arricchimenti alimentari hanno avuto come risultato la riduzione dell’anomalia comportamentale, ovviamente nei casi in cui le cause erano dovute a carenze nutrizionali.
Recenti studi hanno approfondito siffatto comportamento avvalendosi di tecnologie sensoriali. Si è quindi ricorso a dei sensori applicati ad alcuni soggetti per raccogliere informazioni sulle attività comportamentali collettive e sulle reazioni individuali.
L’identificazione tramite radiofrequenza dei soggetti, il monitoraggio dei movimenti e la creazione di modelli comportamentali potrebbe essere il futuro per capire meglio la plumofagia.
Conclusioni
Nei casi in cui si verifichino episodi di plumofagia negli allevamenti sportivi/amatoriali della specie, riveste un ruolo cruciale identificarne la causa. Non necessariamente le cause sono da attribuire a disfunzioni patologiche o a carenze nutrizionali. Le azioni correttive vanno attivate il prima possibile.
La maggior parte degli studi comparati riportati nel presente scritto fanno riferimento ai lavori di HuberEicher e Weschler del 1997, i quali riconoscono come causa primaria della plumofagia l’incapacità dei soggetti di svolgere un comportamento naturale. Dal punto di vista etologico, il
fatto che nella specie Serinus canaria, apportate le dovute azioni correttive, tale anomalia comportamentale scompaia può considerarsi una ulteriore dimostrazione che la specie in questione sia diventata nel corso dei secoli completamente domestica.
BIBLIOGRAFIA REFERENCES
-Huber-Eicher, B., & Wechsler, B. (1997). Feather pecking in domestic chicks: its relation to dustbathing and foraging. Animal Behaviour, 54 (4), 757–768.
https://doi.org/10.1006/anbe.1996.0506
-Van Zeeland, Y. R. A., Spruit, B. M., Rodenburg, T. B., Riedstra, B., van Hierden, Y. M., Buitenhuis, B., Korte, S. M., & Lumeij, J. T. (2009). Feather damaging behaviour in parrots: A review with consideration of comparative aspects. Applied Animal Behaviour Science, 121 (2), 75–95. https://doi.org/10.1016/j.applanim.2009.09.006
-Mason, G. J. (2010). Species differences in responses to captivity: stress, welfare and the comparative method. Trends in Ecology & Evolution, 25(12), 713–721.
https://doi.org/10.1016/j.tree.2010.08.011
-Rose, P. E., Nash, S. M., & Riley, L. M. (2017). To pace or not to pace? A review of what abnormal repetitive behavior tells us about zoo animal management. Journal of Veterinary Behavior: Clinical Applications and Research: Official Journal of: Australian Veterinary Behaviour Interest Group, International Working Dog Breeding Association, 20, 11–21.
https://doi.org/10.1016/j.jveb.2017.02.007 - Using sensor technology against feather pecking in laying hens. (2019, July 17). Zootecnica International.
-Ghelfi, E. (2019, October 5). Plumofagia o pica delle piume: perché le galline si strappano le penne? Animali da compagnia - Il portale per i proprietari di pet.
https://www.animalidacompagnia.it/plumofagia-o-pica-delle-piume-perche-le-galline-sistrappano-le-penne/ -Zootecnica. (2021, October 13). Il benessere animale negli allevamenti di ovaiole – Seconda parte. Zootecnica; Zootecnica International.
https://zootecnica.it/2021/10/13/il-benessereanimale-negli-allevamenti-di-ovaiole-secondaparte/ - Limitare la necessità di taglio del becco. (n.d.). Compassionsettorealimentare.it Retrieved January 14, 2025, from https://www.compassionsettorealimentare.it/ri sorse/galline-ovaiole/limitare-la-necessita-ditaglio-del-becco/ -Galline ovaiole in un sistema cage-free, D.S.C. e. S.Q.D.U. (n.d.). Effetto dell’arricchimento alimentare con zucca. Unipd.It. Retrieved January 13, 2025, from https://thesis.unipd.it/retrieve/85eb4001-f19e48ff-b6b8-0c313cb3bd/Zeccagno_Silvia.pdf
Cardellino Giada
testo e foto di FRANCESCO CIRAUDO
Durante la stagione riproduttiva del 2020, da una linea di “sciarpati” sono nati due soggetti con un fenotipo particolare che chiameremo Giada (foto 1). A prima vista e a seconda dei pregiudizi di ciascun osservatore, potevano essere scambiati per degli atipici Damasco (il Jaspe nel cardellino) o Aurora atipici (ex Phaeo), nel senso che l’espressione della diluizione manifestata ricordava vagamente più il Damasco che l’Aurora, ma con una tonalità diversa da entrambi.
Nel 2020 da una linea di “sciarpati” sono nati due soggetti con un fenotipo particolare che chiameremo Giada
La successiva selezione, dopo aver eliminato i pochissimi soggetti Giada “sciarpati”, ha permesso di replicare senza alcuna pezzatura il fenotipo ap-
parso. Il taglio di sangue con un ancestrale non ha prodotto mutati ma solo dei portatori che l’anno successivo, accoppiati fra loro, hanno generato un maschio Giada, il che, quindi, sembrerebbe dimostrare che la trasmissione genetica di questo fenotipo sia recessiva, non legata al sesso. Questo comportamento genetico porterebbe sia ad escludere che si tratti di una variante del Damasco, che si trasmette in modo semi-dominante, sia a considerare la possibilità che si tratti di una variante dell’Aurora oppure di un’altra mutazione mai
Foto 1
Foto 2
apparsa finora, come sembra suggerire la particolare tonalità delle melanine e il moderato albinismo oculare dei neonati (occhi rossicci alla nascita – foto 2) che scompare già dopo i primi 10/12 giorni e negli adulti non è più visibile, se non con particolari condizioni di illuminazione.
L’accoppiamento mutato x mutato, che teoricamente dovrebbe dare il 100% di mutati, non è stato ancora effettuato ma questa importante verifica è prevista per il prossimo anno.
Ai fini della replicazione del fenotipo mutato, è indifferente che il genitore mutato della coppia riproduttrice sia il maschio o la femmina, per quello che si è potuto osservare fino ad oggi.
Le ali e la coda (foto 3) sono colpite da una riduzione drastica, ma rimangono melanine grigie sia centralmente nel ra-
chide sia in rari granuli sparsi nel resto del vessillo delle penne che manifestano bordi nocciola, che sembrerebbero presenti anche nei portatori. Il resto del piumaggio, schiena e “fungo”, manifesta una riduzione del bruno che può ricordare vagamente il pastello bruno.
C’è una leggera differenza di tonalità della “croce” fra femmine e maschi (leggermente più scuri – foto 4 e 5) e il sottopiuma è grigio azzurrino (foto 6).
Quasi tutti, chi più chi meno, manifestano qualche fuga melanica nera in qualche penna che sfugge all’azione della diluizione (foto 1). Le fughe melaniche scure non scompaiono alla muta seguente né vengono trasmesse nello stesso punto alla prole, per quanto si è potuto vedere per il momento. Seguendo il suggerimento dell’esperto di colore
Foto 3
Foto 4 - Croce femminaFoto 5 - Croci maschi
Ai fini della replicazione del fenotipo mutato, è indifferente che il genitore mutato della coppia riproduttrice sia il maschio o la femmina
Bertarini, l’indirizzo della prossima selezione sarà di cercare di ottenere e stabilizzare la massima tipicità del Giada su un’ottima base ancestrale, evitando di inserire e mischiare ogni altra mutazione conosciuta, compresa quella bruna che forse potrebbe nascondere/inibire l’espressione del Giada, come succede per la mutazione alagrigia dei canarini che nel Bruno non si manifesta.
Foto 6 - Sottopiuma
•Invitiamo tutti gli allevatori a inviare foto di soggetti provenienti dai propri allevamenti, con descrizione della specie, razza e mutazione, all’indirizzo: redazione@foi.it
•All’autore della foto mensilmente prescelta da un comitato interno, verrà offerto in omaggio un libro edito dalla FOI in base alla preferenza e alla disponibilità.
(*)Tutte le foto inviate, anche quelle non pubblicate, rimarranno a disposizione della FOI a titolo gratuito e potranno essere utilizzate, senza alcun limite o vincolo temporale, per pubblicazioni, iniziative e scopi promozionali della Federazione
Questo mese, il protagonista di Photo Show è: NICOLÒ GIORDANENGO R.N.A. 3AUZ con la fotografia che ritrae il soggetto:
“Kakariki fronte rossa femmina” (Cyanoramphus novaezelandiae novaezelandiae)
Complimenti dalla Redazione!
Fenotipo bianco: recessivo vs dominante
testo e figure
MIMMO ALFONZETTI, FOTO E.DEL POZZOE F.O.I.
Seconda parte
SCARB1
Scavenger receptor class B
Una pubblicazione scientifica(Dorrestein, Schrijver, A genetic disorder of vitamin A metabolism in recessively white canaries) chiarisce in modo inequivocabile, attraverso analisi genomiche e biochimiche, l’esistenza di un difetto ereditario nell’assorbimento dei carotenoidi nella razza canarino Bianco Recessivo. I colori giallo, arancione e rosso del piumaggio degli uccelli sono prodotti attraverso la deposizione di pigmenti carotenoidi in piume e pelle. Il gene SCARB1, presente nel cromosoma 15 dei canarini, è un recettore delle lipoproteine legate ai processi vitali delle cellule attraverso i meccanismi del metabolismo dei carotenoidi. Saggi funzionali hanno ulteriormente dimostrato che SCARB1wild-type promuove l’assorbimento cellulare dei carotenoidi, ma che questa funzione viene persa nell’isoforma (e) mutante che predomina nei canarini bianchi recessivi. La sua espressione ad alta densità è richiesta per la colorazione dei carotenoidi negli uccelli. Orbene, in questa razza è stata identificata una mutazione nel gene SCARB1 che interrompe la funzione di trasporto dei carotenoidi: una mutazione del sito di giunzione in SCARB1 (splicing) spiega quindi il fenotipo recessivo bianco.
Lo studio definisce il canarino bianco recessivo un uccello “debole”(forse
anni fa). Gli autori hanno determinato le concentrazioni di vitamina A, dei carotenoidi totali e del betacarotene nel siero, nel fegato e anche nel mangime dei canarini sia colorati che bianchi recessivi. La conclusione è che i carotenoidi non sono praticamente riassorbiti dall’intestino dai canarini bianchi recessivi. La vitamina A viene anche assorbita in misura decisamente minore da questi uccelli rispetto ai canarini pigmentati.
L’espressione della colorazione dei carotenoidi negli uccelli coinvolge diverse fasi fisiologiche distinte; il loro trasporto nei sistemi circolatorio e linfatico è di fondamentale importanza. Il gene SCARB1 sostiene l’assimila-
zione dei carotenoidi nelle cellule, è un regolatore essenziale dell’espressione della colorazione a base di carotenoidi negli uccelli e svolge un ruolo importante nel processo sopra descritto. L’assorbimento dei carotenoidi avviene nel tratto intestinale dove, sotto l’azione degli enzimi biliari, vengono sciolti in finissimi aggregati chiamati micelle per essere secreti nella linfa, per poi entrare nel sistema circolatorio e nel fegato. L’intero compito di trasporto è assolto da SCARB1. I carotenoidi, messi in circolazione successivamente dal fegato, hanno bisogno di legarsi alle proteine lipidiche presenti nella circolazione sanguigna (Parker, 1996).
Isabella opale bianco recessivo, foto: FOI
Il trasporto di carotenoidi nel plasma viene correlato alle lipoproteine a bassa densità (LDL) mentre i carotenoidi con legami covalenti polari (d) tendono a legarsi alle lipoproteine ad alta densità (HDL) (Erdman et al., 1993). Ad esempio la luteina, carotenoide principale responsabile della colorazione gialla dei canarini, si lega alle lipoproteine HDL proprio perché la sua catena enzimatica contiene legami covalenti con alterata polarità a causa di una distribuzione elettronica asimmetrica.
Anche Toomey et al. (2017) hanno scoperto che i canarini bianchi recessivi hanno livelli molto bassi di carotenoidi nei loro tessuti rispetto alle varietà rosse o gialle. A seguito di mappatura genetica, si è scoperto che l’allele bianco recessivo è dovuto ad una mutazione puntiforme del gene SCARB1 che opera una scissione anomala degli
isomeri (g). SCARB1 perde la funzione di trasportare carotenoidi, con conseguente incapacità dei carotenoidi di accumularsi nei tessuti superficiali del corpo e quindi di colorarli.
SCARB1 può essere considerato quindi come recettore delle proteine lipidiche (lipoproteine) ad alta densità HDL per armonizzare la colorazione dei carotenoidi nei tessuti bersaglio (pelle, follicoli piume, retina, ecc.). Le mutazioni del gene codificante SCARB1 possono interrompere il processo di trasporto dei carotenoidi nelle cellule (Sakudoh et al., 2013).
Bianco dominante
Si ritiene che il bianco dominante sia apparso per la prima volta nell’anno 1667 ad Augsburg, nell’allevamento del Dr. Lucas Scrock (1646-1730) in Baviera. Risulta, inoltre, che solamente intorno al 1924 questa varietà incon-
trò notevole favore e stimolò l’interesse degli allevatori, cominciando a diffondersi con una rapidità mai prima conosciuta.
Specialmente dalla Germania molti di questi canarini si diffusero in tutto il mondo. Dagli stessi allevatori tedeschi erano già denominati bianchi dominanti, ma furono anche indicati da altri allevatori, data la loro provenienza, come “bianchi tedeschi”.
Bianco, cioè, come la stessa parola dice, senza colore: si trattava effettivamente di soggetti apigmentati. Il fattore genetico responsabile della pigmentazione del loro mantello, anni or sono, fu chiamato fattore inibitore del giallo (Veerkamp); tutto ciò si è dimostrato vero: non soltanto inibiva il giallo, bensì qualsiasi tipo di lipocromo. Una causa plausibile della mancanza di pigmentazione in questo esemplare, stante la corretta assimilazione e trasporto dei carotenoidi, potrebbe essere correlata ad un arresto in qualsiasi fase del processo di deposizione delle xantofille nei follicoli. Tuttavia, i processi coinvolti nella deposizione dei carotenoidi nell’insieme degli uccelli erano e sono ancora poco conosciuti (McGraw et al., 2003), anche perché tale deposizione è quasi sempre un processo pleiotropico (Brush, 1990).
Il relativo fattore genetico ha comportamento dominante nei riguardi del suo allele ancestrale ma, come si è detto, il fenotipo mostra piuttosto un processo genetico di inibizione limitato: il canarino bianco dominante presenta, infatti, delle soffusioni lipocromiche in particolari zone di concentrazione cromatica del mantello.
Queste soffusioni gialle si rilevano come carattere tipico sul bordo esterno delle remiganti primarie (1a e 2a). Se sono troppo estese o presenti in altri parti del piumaggio, sono considerate difetti. Spesso si notano sulle timoniere, sulle spalline, nella zona periorbitale e nei casi di gravissimo difetto anche altrove, mai nel cosiddetto sottopiuma emai sul codione. Quest’ultima è una caratteristica singolare ma utile, in particolare ai giudici, perché permette il discernimento
Intenso avorio giallo, foto: E.delPozzo
corretto rispetto a certi canarini mosaici (quasi sempre avorio); nei casi dubbi a visione superficiale, si può avere quella certa, osservando il codione.
Le soffusioni appaiono essere superficiali, limitate alle estremità delle piume, rilevando quindi una costruzione proprio nelle prime fasi di abbozzo della piuma, validando quella sensazione di apigmentazione profonda del fattore bianco dominante. La pelle è di colore normale (giallastro) come quella di tutti i lipocromici. La pigmentazione, quindi, interessa l’estremità distale della piuma, che è prevalentemente pennacea ed è assente nella parte prossimale (di natura plumacea). L’estremità distale di una piuma si forma prima dell’estremità prossimale e la struttura e la forma della piuma cambiano dinamicamente dall’estremità distale all’estremità prossimale.
Abbiamo cognizione da recenti ricerche che tra le 40 coppie di cromosomi del canarino, in particolare sul microcromosoma 25, è presente un gruppo di geni denominati EDC, Epidermal differentiation complex. Tale cluster di geni, presente in tutto il regno animale, codifica proteine strutturali che, tra le altre cose, sono implicati nella differenziazione delle cellule del tegumento, in particolare il processo di corneificazione dei cheratinociti; la pelle, i peli ma anche le piume degli uccelli sono coinvolti. Le mutazioni di questi geni, nell’uomo, inducono difetti nella cheratinizzazione del tegumento che compromettono la funzione protettiva della pelle. Alcuni esempi di queste anomalie includono ittiosi, psoriasi, eczema, cheratosi pilare.
Non solo sul cromosoma 25 ma anche sul cromosoma 8 sono presenti altri geni importanti per la pigmentazione del piumaggio. Il gene CYP2J19 che qui risiede è coinvolto nella trasformazione dei carotenoidi in chetocarotenoidi, che sono quelli che consentono i fenotipi con varietà rossa. [A margine il cariotipo del Serinus canaria è composto da 40 coppie di cromosomi (2n=80, macro=18, micro=62, Z e W metacentrici].
EDC
Epidermal Differentiation Complex
I canarini bianchi dominanti portano una mutazione che impedisce la deposizione di pigmenti carotenoidi nelle piume durante la muta (o alla nascita). Sebbene questa mutazione non sia ancora stata ricondotta a un gene specifico, i suoi effetti fenotipici sono ben noti: i canarini BD assorbono i carotenoidi e li fanno circolare agli stessi livelli dei canarini gialli, ma non assegnano carotenoidi all’ornamento (tranne le soffusioni) e hanno un piumaggio bianco. Questi uccelli, che evitano l’assorbimento di pigmenti carotenoidi circolanti, hanno chiaramente sviluppato una capacità di evitare una eventuale ridondanza funzionale dei pigmenti, verosimilmente regolando l’espressione genica follicolare. Tale espressione genica è governata da un gruppo di geni, recentemente scoperti: il complesso di
differenziazione epidermicaECD. Il complesso di differenziazione epidermica è un composito genetico comprendente oltre cinquanta geni che codificano per proteine coinvolte nella differenziazione terminale e nella corneificazione dei cheratinociti, il tipo cellulare fondamentale dell’epidermide degli uccelli. I geni EDC sono espressi prevalentemente negli strati cellulari dell’embrione, mentre altri sono confinati negli strati epidermici che si mantengono nella pelle post-natale. È anche responsabile dello sviluppo e della riparazione del tessuto epiteliale, regolando il programma di differenziazione terminale dei cheratinociti. Anche i geni della cheratina, fondamentali nell’evoluzione delle piume aviarie, hanno avuto origine all’interno di EDC, il cui locus non ha alcuna relazione apparente con i geni della cheratina. Delle due forme strutturali della cheratina le
Bianco dominante, foto: E.delPozzo
piume degli uccelli sono in prevalenza costituite da β-cheratina che permette un processo di corneificazione più efficiente delle α-cheratina, conformando strutture con proprietà fisiche uniche e meccanicamente resistenti ma flessibili. L’aggiunta di proteine EDC alle cheratine ha trasformato la morfogenesi delle piume; il processo di cheratinizzazione epidermica presente negli amnioti (h) subisce un diverso e nuovo processo di corneificazione dell’epidermide in particolare nelle piume.
Un interessante articolo di un ornitologo tedesco (Norbert Schramm), degno di particolare attenzione, giustifica la comparsa dei bianchi dominanti con una mutazione di questo complesso di geni, pregevole intuizione di una genesi fin adesso sconosciuta. La crescita e il differenziamento dei cheranociti sono meccanismi epigenetici (e) regolati da questa cassetta di geni che, attraverso l’induzione o la repressione dei geni EDC, svolge un ruolo cruciale nello sviluppo epidermico.
La famiglia di geni EDC ha avuto origine da un singolo o da un piccolo numero di geni ancestrali in un antenato degli arcosauri primitivi e da allora si è espansa rispettivamente negli uccelli, nei coccodrilli e nelle tartarughe. L’adattamento ad una vita completamente terrestre supporta l’ipotesi che i geni ancestrali dell’EDC si siano evoluti principalmente attraverso processi di duplicazione e diversificazione. La duplicazione genica è il processo che porta alla formazione, all’interno di un genoma, di due o più copie di uno stesso gene, con sequenze nucleotidiche identiche o molto simili, mentre la diversificazione è la caratteristica di un pool di geni comunemente ritenuta vantaggiosa per la sopravvivenza. Le piume degli uccelli mostrano un’ampia gamma di proprietà fisiche che hanno permesso loro di espandersi e sopravvivere in diversi ambienti in ogni continente, incluso l’Antartide.
È possibile, quindi, spiegare come nei canarini BD ad un piumaggio totalmente bianco si contrappongano limitate zone, le soffusioni, pigmentate da xantofille tramite meccanismi di differenziazione. Nei bianchi dominanti ete-
È possibile, quindi, spiegare come nei canarini BD ad un piumaggio totalmente bianco si contrappongano limitate zone, le soffusioni, pigmentate da xantofille tramite meccanismi di differenziazione
rozigoti si può ipotizzare che le cellule cheratiniche delle piume, tramite induzione e differenziazione dei geni EDC, inibiscano, forse, l’immagazzinamento dei lipocromi già circolanti nel sangue oppure, con grande attendibilità, che il trasferimento alle piume sia consentito solo in uno stadio giovanile iniziale. Ciò spiega perché le punte delle piume (le parti distali sono le aree che si formano per prima) possano presentare ancora depositi di lipocromo, mentre le parti prossimali, la cui formazione è succes-
siva, risulteranno non pigmentate. L’evoluzione nel tempo dei diversi tegumenti aviari e della diversità della pigmentazione lungo il vessillo delle piume è stata associata alla attività di regolazione epigenetica di EDC; l’informazione genetica di tali geni viene trascritta in modo temporale e specifico per ogni regione della pelle.
La tempistica del processo di implumazione è quindi variabile tra le diverse regioni del corpo degli uccelli, ma anche tra le specie. Le remiganti sono tra le prime ad emergere dalle papille dermiche (nei rondoni a 10 giorni dalla nascita) e hanno la possibilità di pigmentarsi. L’importanza ai fini del volo delle remiganti è indiscutibile e quindi il loro irrobustimento necessita di una nascita prematura. Al contrario, il codione con le sue piume, che non appaiono mai colorate, trova giustificazione in una conformazione ritardata nella nascita rispetto alle altre piume.
Gli uccelli BD a fattore singolo sono pienamente fertili e vitali perché solo i depositi di grasso nelle piume sono interessati dalla mutazione.
Degna di nota è comunque la tesi (G. Canali) che la mutazione BD non sia perfettamente mendeliana per la presenza di geni modificatori, anche se monogenica come gene principale o maggiore. La concezione mendeliana “un carattere >> un gene” è ormai abbandonata. Nella grande maggioranza dei casi i caratteri sono determinati da più geni, le cui azioni si completano o interferiscono in vario modo. I geni modificatori, dotati di scarsa espressione singola, alterano l’espressione genica esaltando o riducendo l’intensità del carattere principale. Con un volo pindarico, non sarei sorpreso se ritrovassi questi geni modificatori nel cluster di geni EDC.
La mutazione dominante-bianco e la letalità
La maggior parte dei libri che trattano l’allevamento dei canarini sostengono che l’uccello bianco dominante a doppio fattore non esista perché, in concomitanza (omozigosi), gli alleli dominanti hanno un effetto letale. Non ci sono prove evidenti a sostegno di questa credenza ampiamente diffusa, quindi la letalità forse è un altro esem-
Mosaico rosso avorio, foto: E.delPozzo
pio del “sapere del popolo” che persiste nella fantasia degli allevatori proprio perché sembra non avere basi scientifiche Ricordiamo che i geni letali sono geni che portano al decesso in una fase precoce dello sviluppo embrionale, prima della nascita. Una ricerca sperimentale, (Geoff Walker, Colour, type and song canaries) descrive un programma di riproduzione che fornisce prove concrete che la mutazione bianco dominante non è in realtà letale e che l’uccello BD a doppio fattore esiste davvero. La procedura utilizzata è stata quella di re-incrociare (test cross) un buon numero di bianchi dominanti e poi, la stagione successiva, di controllare la progenie da accoppiamenti con gialli per verificare l’esistenza di doppi fattori. Se da un incrocio bianco dominante per giallo viene generato un pulcino giallo, significa, senza alcun dubbio, che il genitore bianco era monofattoriale e quindi può essere escluso da ulteriori indagini. D’altra parte, se l’uccello bianco genera solo giovani bianchi consecutivi, la probabilità che abbia il fattore in doppia dose aumenterebbe per ogni pulcino bianco schiuso. Questa procedura naturalmente non dimostra assolutamente che un uccello sia a doppio fattore, ma ha il potenziale per fornire una probabilità convincente che lo sia.
I primi bianchi auto-incrociati, secondo il programma di allevamento previsto, hanno prodotto 12 novelli e questi sono stati incrociati con i gialli l’anno successivo. Dei dodici, nove hanno avuto figli di pulcini gialli e sono stati messi da parte, mentre tre hanno avuto rispettivamente 11, 8 e 6 giovani bianchi consecutivamente. Questi tre genitori erano di notevole interesse perché i loro risultati riproduttivi tendevano a contraddire il genotipo a loro attribuito (eterozigote). Il calcolo numerico della probabilità (c) che tali genitori fossero omozigotici per il fattore dominante era prossimo alla certezza matematica, 99, 95 % per la variabile progenie 11 novelli. In maniera più semplice, è come se si lanciasse una moneta 11 volte e uscisse sempre testa… si potrebbe pensare che la moneta non sia a due facce distinte.
La prova fornita dal genitore degli un-
dici pulcini bianchi consecutivi, supportata dai 6 e 8 degli altri due genitori, è un argomento convincente a favore dell’esistenza dell’uccello a doppio fattore. Bisogna dire che non c’è un grande vantaggio nel mantenere i doppi fattori. Non hanno nulla da offrire all’allevatore da esposizione oltre al singolo fattore e mantenere una linea di doppi fattori probabilmente coinvolgerebbe l’allevatore medio in un livello inaccettabile di consanguineità. Per tanto tempo il fattore bianco dominante è stato considerato, senza eccezioni, letale; successivamente, la letalità è stata sostituita dal concetto di subletalità. Infatti, oltre ai geni letali, esistono i geni subletali che determinano la comparsa di anomalie compatibili con la vita. Gli individui muoiono, tuttavia, nei primi periodi di vita perché non sono in grado di sopravvivere. Una distinzione delle anomalie genetiche dei vertebrati rispetto alla gravità riportata (Hutt, 1985) è la seguente: •disvitali o semiletali: non sono causa di morte per tutti gli individui ma soltanto per alcuni e la percentuale di
letalità è fortemente influenzata dalle condizioni esterne. Questi geni riducono l’efficienza fisiologica temporaneamente o permanentemente;
•subletali: tutte quelle che non siano letali alla nascita o immediatamente dopo, ma che inducono il decesso prima dell’età riproduttiva. Altri autori chiamano i geni che causano la morte in età più avanzate dopo la nascita letali ritardo.
•letali: dovute a geni che causano la morte prima della nascita o immediatamente dopo.
Possiamo quindi concludere che i termini “letalità” e “subletalità” non siano consoni per definire i geni del bianco dominante, ma sarebbe più corretto parlare di “semiletalità”o “disvitalità”. La teoria dell’evoluzione della dominanza di Fisher afferma che in condizioni naturali ogni mutazione dominante dannosa riesce a mantenersi nei soggetti selvatici e si trasmette alla generazione successiva più facilmente negli individui dotati di geni modificatori che ne riducono o ne eliminano gli effetti nocivi. La sopravvivenza dei bian-
Mosaico avorio giallo femmina, foto: E.delPozzo
chi dominanti omozigoti, rispetto a questa teoria, non dovrebbe essere impossibile.
R. A. Fisher nel 1928 introdusse la sua teoria dell’evoluzione della dominanza integrando quella di Darwin. A grandi linee, la teoria afferma che in presenza di selezione naturale il fitness (l) medio di una popolazione tende ad aumentare. Più semplicemente, se un gene sfavorevole viene continuamente prodotto per mutazione, alla fine verranno selezionati altri geni modificatori che correggono l’azione dannosa di questo gene, il che darà agli individui portatori del gene un fenotipo favorito. Poiché il gene si trova inizialmente in larga misura nello stato eterozigote piuttosto che omozigote, è l’eterozigote che è principalmente soggetto a questa modificazione. Si dovrebbe prevedere per la sopravvivenza dei BD in omozigosi un processo molto lento, poiché la frazione di canarini esposta a questa selezione è molto piccola. Teoria da prendere con le pinze, proprio perché è una teoria.
Bianchi ad eredità avorio?
Quando un carattere è controllato da due o più coppie di alleli, la loro interazione genica può far comparire fenotipi completamente nuovi. La polimeria, detta anche interazione genica, è un meccanismo di determinazione di un carattere fenotipico da parte di più geni. È quello che succede quando si incrociano canarini Bianchi dominanti con canarini Avorio. La mutazione bianco dominante si discosta da quella recessiva lasciando soffusioni lipocromiche che nei soggetti migliori sono limitate al bordo esterno delle remiganti primarie. Ora, dall’incrocio in oggetto ci si aspetta soggetti bianco dominante con soffusioni avorio, ma ciò non avviene. Nascono bianchi, nuovo fenotipo, che la consuetudine definisce bianco dominante puro per l’avorio, non distinguibili dai bianchi recessivi se non per la pelle carnicina e non violacea. L’interazione genica non è una semplice sovrapposizione di fattori ma qualcosa
di più complesso. Il mantello dei cani Labrador (nero, cioccolato, biondo) e la cresta dei galli (a rosa, a pisello, a noce e singola) sono esempi conclamati di interazione genica. In particolare, l’interazione tra geni non allelici che determina una situazione in cui un gene modifica la manifestazione di un altro gene, quindi situato in un locus differente, prende nome di interazione genica epistatica. Credo che i bianchi dominanti ad eredità avorio siano ascrivibili a questa tipologia di interazione genica dominante. Infatti, il fenotipo di questi ultimi differisce da quello che si otterrebbe se i geni si esprimessero in maniera indipendente. A mio avviso, ma potrei sbagliare, forse sarebbe più consono definirli avorio ad eredità bianco dominante. Il fattore Avorio non ha mai mostrato attitudine all’epistasi, cosa che si può affermare per il carattere Bianco dominante. La logica di quanto sopra affermato riviene dal processo di corneificazione del pool genico EDC risultante dall’interazione. Durante il differenziamento terminale, i cheratinociti epidermici intraprendono una serie di eventi cellulari programmati che consentono loro di entrare a far parte dello strato cornificato di corneociti non vitali; questi sono stati cheratinizzati dallo strato basale dei cheratinociti “attivi” del fattore avorio. I geni EDC influenzano questa pro-
grammazione modulando i fattori epigenetici finali. Le piume avorio affrontano la fase di cheratinizzazione con dosi abbondanti di cheratina, che porta ad un loro rapido indurimento, consentendo ai lipocromi di diffondersi nella corteccia solo in quantità limitate. D’altra parte, però, le cellule pigmentarie delle remiganti primarie, localizzate in una particolare zona embrionale del derma, la cui colorazione è legata alla tempistica di pigmentazione imposta da EDC, non hanno possibilità di differenziarsi. Il complesso EDC impedisce ai carotenoidi avorio, già in minime concentrazioni, di diffondersi nella corteccia a causa di un processo di corneificazione molto rapido.
Il processo di diffusione dei carotenoidi avorio, quindi, è negato anche alle piume che si sviluppano per prime (remiganti, timoniere, spalline...) perché il tutto si compie in tempi ristretti con il sopravento degli effetti inibitori del complesso EDC.
In maniera più sintetica, EDC agisce sulla papilla (di origine mesodermica), depositaria dei caratteri ereditari, stimolando l’epitelio a produrre un intenso accumulo di cheratina nelle piume formando materiale corneo delle barbe e del rachide di maggiore densità e durezza. Il processo rapido di corneificazione imposto da EDC impedisce a qualunque xantofilla di diffondersi. C’è quindi sinergia tra cheratinizzazione del fattore avorio e corneificazione di EDC in tempi rapidi, prima dell’espulsione della piuma dalla papilla.
Per tutte le piume che si conformano successivamente, la differenziazione delle cellule della cresta neurale sarà inibita dal complesso EDC e quindi sarà impedita la diffusione dei lipocromi. Il piumaggio finale sarà bianco senza lipocromi avorio e quindi senza soffusioni.
Non esiste alcun accertamento scientifico di quanto esposto in questo paragrafo, sono solo ponderazioni (probabilmente fantastiche) mie personali.
Brinato avorio giallo, foto: E.delPozzo
Note
a)Diffusione luminosa, la cui proprietà è l’indipendenza dalla direzione (isotropia)
b)La riflettanza misura, in ottica, la capacità di riflettere parte della luce incidente su una data superficie o materiale; è il rapporto tra intensità del flusso riflesso e del flusso incidente
c)Probabilità di eventi disgiunti, ossia che alla schiusa (successiva) di un uovo possa nascere un bianco dom. omozigote (evento incognito)
d)Illegame covalente polare si forma tra atomi diversi, con differenza di elettronegatività compresa tra 0, 4 e 1, 9, che mettono in comune delle coppie di elettroni
e)L’epigenetica studia come fattori ambientali ed ereditari possono cambiare l’espressione dei geni senza cambiare il DNA (Airc)
f)Molecole proteiche dotate della stessa funzione, seppur differiscano parzialmente per struttura
g)Composti con la stessa formula ma una diversa disposizione di atomi nelle molecole
h)Gli amnioti generano un uovo fornito di guscio
i)Numero adimensionale che indica il rapporto tra la velocità della luce nel vuoto e la velocità di questa nel mezzo; è una caratteristica ottica del materiale
l)Il fitness di un genotipo è una misura della capacità di un individuo di sopravvivere e riprodursi
Fonti
-Rebecca Koch, Molly Staley, Andreas Kavazis, Dennis Hasselquist, Matthew Toomey, Geoffrey Hill - Testing the resource tradeoff hypothesis for carotenoid-based signal honesty using genetic variants of the domestic canary
-Chen Siang Ng, Wen-Hsiung Li - Genetic and Molecular Basis of Feather Diversity in Birds
-Ricardo J. Lopes, James D. Johnson, Matthew B. Toomey, Geoffrey E. Hill, Joseph C. Corbo, Miguel Carneiro- Genetic Basis for Red Coloration in Birds
-W. Fox and J.R. Smyth Jr - The Effects of Recessive White and Dominant White Genotypes on Early Growth Rate
-Norbert Schramm - Farbenkanarien des JahresDominantweiß
-Francisco Grimalt Salvá - La herencia genética del canario blanco dominante
-Rosalyn Price-Waldman and Mary Caswell Stoddard - Avian Coloration Genetics: Recent Advances and Emerging Questions
-Chen Siang Ng, Wen-Hsiung Li- White Canary Genetics, Home - Spanglefish
-U. Zingoni - Canaricoltura, FOI
-G. Canali - Il canarino bianco, Adop Parma
-Matthew B. Toomey, Ricardo J. Lopes, Pedro M. Araújo and Miguel Carneiro - High-density lipoprotein receptor SCARB1 is required for carotenoid coloration in birds
-H.J. Veerkamp - Manuale sull’allevamento dei canarini di colore
-G.M. Dorrestein, J. Schrijver - A genetic disorder of vitamin A metabolism is recessively white canaries
-Toomey M.B., Lopes R.J., Araújo P.M., Johnson J.D., Gazda M.A., Afonso S., Mota P.G., Koch R.E., Hill G.E., Corbo J.C., et al - High-density lipoprotein receptor SCARB1 is required for carotenoid coloration in birds.
-V.A. Olson, I.P.F. Owens - Interspecific variation in the use of carotenoid-based coloration in birds: diet, life history and phylogeny
-Anthony Davis and Matthew J. GreenwoldEvolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds
-Tania Tanfoglio - dal blog www.scienceforpassion.com
-H.K. Garg - Chromosomes in birds
-J.A. Sved and O. Mayo - The Evolution of Dominance
-Summa Gallicana - I geni modificatori
Nella città della Musica risuonano i canti dei canarini Malinois Watersalger
testo e foto di ROBERTO GABRIELE, STEFANO GABRIELE e ROBERTA PUGLIESI
Il Concorso Canoro per Canarini da Canto Malinois Waterslager si è svolto a Isola del Liri, la “Città della Musica e della Cascata”, nei giorni 29, 30 novembre e 1 dicembre 2024 ed è stato un evento unico ed emozionante. L’iniziativa, organizzata dalle associazioni ornitologiche G.O.R. (Gruppo Ornitologico Romano) e A.O.V.D.L. (Associazione Ornitologica Valle del Liri), aderenti alla Federazione Ornicoltori
Italiani, è stata ospitata presso il Teatro Stabile Comunale, con il patrocinio del Comune di Isola del Liri.
Una manifestazione che ha unito passione e amicizia L’evento ha richiamato appassionati di ornitologia da tutta Italia e dall’estero, con una rappresentanza significativa dall’Albania. Sono stati tre giorni all’insegna della sana competizione, dell’ami-
cizia e della condivisione, dove l’amore per i canarini Malinois è stato il punto di unione più forte.
Gli organizzatori e i partecipanti hanno voluto sottolineare il ruolo fondamentale del Comune di Isola del Liri nel garantire il successo della manifestazione. Un sentito ringraziamento è stato rivolto al Sindaco Massimiliano Quadrini, all’Assessore Stefano Vitale e all’intera Giunta Comunale per il prezioso sup-
Organizzatori e partecipanti al concorso
Giudici del canto italiano insieme agli allievi giudici albanesi
Consegna della targa di ringraziamento al Sindaco Massimiliano Quadrini, presente con la consorte, per il suo prezioso sostegno all’evento
La maestosa Cascata Grande di Isola del Liri incorniciata dal fascino del centro storico
porto e per la calorosa accoglienza riservata agli ospiti.
Il Malinois Waterslager: una storia emozionante
Durante l’evento, il Sindaco Quadrini ha voluto sottolineare quanto affascinante sia la storia di questi canarini. Originari del Belgio, i Malinois Waterslager venivano utilizzati dai minatori per individuare eventuali fughe di gas nelle gallerie sotterranee. Grazie alla loro sensibilità e al loro canto, questi uccelli si sono rivelati veri e propri alleati nella sicurezza dei lavoratori.
Il canto dei Malinois è armonico e caratterizzato da toni unici, che variano dal “suono dell’acqua che scorre” ai richiami dell’usignolo, passando per note rullate e particolari armonie. Il loro nome, Waterslager, significa proprio “suoni d’acqua”, un riferimento all’eco delle gocce nelle miniere, che gli uccelli imparavano a riprodurre.
“Il borgo di Isola del Liri ha dimostrato ancora una volta di essere la cornice ideale per eventi che celebrano la musica e la bellezza in tutte le sue espressioni”
Isola del Liri: una cornice ideale per la musica
Non c’era luogo più appropriato di Isola del Liri per ospitare questa manifestazione. Conosciuta come la “Città della Musica” grazie alle sue tradizioni artistiche e culturali e per il fascino unico della sua cascata nel cuore del centro storico, la cittadina ha confermato la
sua vocazione a celebrare la bellezza in tutte le sue forme.
“Il borgo di Isola del Liri ha dimostrato ancora una volta di essere la cornice ideale per eventi che celebrano la musica e la bellezza in tutte le sue espressioni” ha dichiarato il Sindaco Quadrini. E ha aggiunto: “Non si smette mai di imparare. La storia di questi canarini è molto particolare, triste ma emozionante”.
Il Concorso Canoro per Canarini Malinois Waterslager è stato molto più di una competizione: è stato un momento di incontro, cultura e scoperta. Gli organizzatori e i partecipanti hanno già manifestato il desiderio di rinnovare l’appuntamento, continuando a celebrare la bellezza del canto e la passione per l’ornitologia. Isola del Liri, con il suo spirito accogliente e la sua atmosfera unica, ha saputo dare un tocco magico a un evento che rimarrà nel cuore di tutti.
Consigli per l’allevamento dei pappagalli
testo di RAFAEL ZAMORA PADRÓN (*), foto LPF
La stagione riproduttiva sta iniziando per i pappagalli della Loro Parque Fundación, ed è giunto il momento di affinare l’istinto dei custodi per fare molto meglio rispetto alla stagione precedente. Offrire il nido che piace di più a una coppia. Incoraggiare i soggetti che non hanno avuto successo riproduttivo l’anno scorso. Vedere cosa sanno fare le coppie appena formate. E godersi, in generale, uno dei momenti più emozionanti nell’allevamento e nella conservazione dei pappagalli. Nell’allevamento dei pappagalli è normale che l’apprendimento non finisca mai. Non sappiamo mai tutto, ci sono sempre nuove sorprese e nuove sfide. Ecco perché l’addestramento è così importante. Specie diverse hanno comportamenti diversi, ogni coppia è diversa, gli individui hanno le loro particolarità e in ogni centro di allevamento del mondo le situazioni sono diverse. Forse è questa variabilità e le sfide che presenta ad attrarre maggiormente gli appassionati che entrano in un ciclo di passione sfrenata per i pappagalli che si rinnova ogni anno.
Oggigiorno, le informazioni condivise tramite i social network e la possibilità di avere questi contenuti immediatamente a portata di mano tramite i nostri smartphone ci permettono di apprendere apparentemente più facilmente che in passato. Nonostante ciò, conti-
(*)Direttore
Scientifico Fondazione Loro Parque
nuiamo talvolta a subire grandi delusioni, commettendo errori e fallendo in dettagli che possono costare la vita a un animale. Per questo motivo l’esperienza personale basata sull’accesso ai libri, sia antichi che moderni, deve essere completata dalla partecipazione a
congressi, seminari e discussioni dirette nelle associazioni di allevatori. Questa via è ancora la più efficace per aggiornare le conoscenze e per poter decidere sui cambiamenti da apportare nella gestione quotidiana dei pappagalli. Un sistema che ora è completato dalla po-
Cacatua leadbeateri, foto: R. Zamora LPF
tenza della comunicazione digitale: più veloce, più breve e più immediata, ma con rischi associati a interpretazioni errate.
Brevi commenti scritti o informazioni visive sui social media sono un prezioso supplemento per gli allevatori esperti. Tuttavia, per i nuovi allevatori comportano rischi a causa della mancanza di esperienza accumulata e perché ricevono opinioni diverse che non sempre si basano sulla conoscenza. Si crea confusione e il destinatario è tentato di apportare ai propri animali cambiamenti che spesso non sono appropriati. La verità è che c’è un alto rischio di fare tutto per sentito dire da un altro allevatore che, magari, sta andando bene con qualche integratore vitaminico o altro accorgimento. Ma tutti questi cambiamenti devono essere apportati con attenzione e dando tempo per vedere i risultati particolari in ogni caso. In qualsiasi associazione di avicoltura, è
facile riscontrare come coloro che hanno i risultati migliori sono le persone che si sono ritirate dal loro lavoro a causa della loro età. Sono la voce dell’esperienza, coloro che hanno imparato dai propri successi e fallimenti. Sanno come identificare cosa funziona e cosa no. E sono anche aperti a nuovi suggerimenti per applicarli e vederne l’effetto
con il rigore e la cautela richiesti. Dall’inizio di settembre 2024, l’allevatore Has Jürgen Geil ci supporta con la sua conoscenza di una vita sulla gestione dei pappagalli. Questo è un aiuto importante, perché i suoi occhi esperti rendono più facile avere maggiori opzioni sul tavolo per cambiare eventualmente un tipo di gestione.
Alla Loro Parque Fundación abbiamo lavorato con molti esperti mondiali di pappagalli, ognuno con le proprie opinioni e i propri approcci particolari. Il loro contributo all’avicoltura e alla conservazione mondiale è stato immenso. Grazie a loro siamo riusciti a far sì che specie poco conosciute venissero affidate alla cura dell’uomo. E i loro protocolli di gestione sono stati resi disponibili agli hobbisti tramite congressi e pubblicazioni.
Da Loro Parque Fundación auguriamo a tutti voi una buona stagione riproduttiva.
Hans Jürgen Geil (a destra)
Ancora un successo a Lanciano
testo e foto DOMENICO MAIONE (Presidente Raggruppamento Ornitologico Abruzzo-Molise)
Dopo un riuscitissimo Campionato Italiano, Lanciano ha raddoppiato a settembre, nei giorni dal 25 al 29, con l’annuale Mostra di Erythrura, fiore all’occhiello per il raggruppamento che mi onoro di rappresentare: specialistica dedicata agli esotici e organizzata da un Comitato inter-associazione capitanato dal gentilissimo Luciano Di Biase e dal Presidente dell’A.O.C.A. Donato Sabatini.
Come ormai da tradizione, anche quest’anno l’evento ha avuto un grandissimo successo, con animali e allevatori giunti da ogni parte d’Italia (Lombardia e Sicilia i due estremi): la mostra ha infatti potuto vantare la partecipazione di 91 espositori con 1200 Diamanti di Gould, 40 Erythrura, 70 Codalunga, 25 Passeri del Giappone.
Come negli anni scorsi, gli animali sono stati ospitati nel padiglione
dell’Ente Fiera, luminoso e di buon impatto scenografico. Gli allevatori sono stati accolti con la proverbiale gentilezza e cortesia che sono ormai un marchio di fabbrica del Comitato Organizzatore, composto da un gruppo di amici uniti e spinti dalla comune passione che li anima. L’organizzazione praticamente perfetta è stata ulteriormente impreziosita da un’ottima cerimonia di inaugurazione, con la presenza del Sindaco
della città dott. Filippo Paolini, del presidente dell’ente fiera sig.ra Ombretta Mercurio e con la benedizione del parroco, tutto ripreso da Abruzzo WebTV.
Inoltre, quest’anno si è avuta la presenza fisica del sig. Gary Fitt, presidente dell’associazione Onlus “SAVE THE GOULDIAN FUND”, che si occupa da anni di salvaguardare il Gould nei suoi territori naturali dell’Australia del nord. La Onlus svolge progetti di ricerca sul campo, come ad esempio la posa di centinaia di nidi artificiali per lo sviluppo e la crescita delle colonie di Gould. L’inaugurazione è culminata con un buffet veramente gustoso e variegato. Il sig. Fitt è venuto a Lanciano per sincerarsi dei nostri allevamenti ed è stato molto soddisfatto dal nostro interessamento al Diamante di Gould.
Un Grazie a Luciano, Donato e a tutti i collaboratori e partecipanti.
Dissertazioni sul Fanello e Standard proposto
testo di SEBASTIANO
PATERNÒ, foto: S. PATERNÒ eF.O.I.
Introduzione
Archiviato il 55° Campionato Italiano di Ornitologia svoltosi a Montichiari, Brescia e memorizzati i giudizi di tutti gli esemplari presenti, in totale 12.000 di cui 2.391 EFI (avifauna europea – esotica – ibrida), quest’anno in costante aumento numerico (19%) rispetto alle altre categorie e agli anni precedenti (all.n.1), vogliamo toccare il tasto della presenza in mostra di un uccello, il Fanello, ancora in numero limitato per via di un’assenza di standard che penalizza il giudizio singolo in quanto abbinato in unica categoria con le Peppole (Fringilla montifringilla), maggiormente colorate e più calme rispetto al Fanello (Linaria
In presenza di uno Standard specifico sul Fanello i numeri incrementerebbero sicuramente
cannabina). Quest’anno, al Campionato, solo 7 soggetti e tale numero giustifica le remore della C.T.N.-EFI ad una categoria autonoma. Per altro verso, gli ibridi col Fanello di sesso maschile sono stati ben rappresentati e in totale erano
ben 59, senza tener conto che altri erano presenti fra i 34 ibridi generici di sesso femminile.
Premesso quanto sopra, appare chiaro che in presenza di uno standard specifico sul Fanello i numeri incrementerebbero sicuramente verso una specie che merita certamente una maggiore attenzione ed è per seguire tale linea di indirizzo che sono state fatte riunioni e valutazioni verso la stesura di uno standard.
La Commissione Tecnica Nazionale EFI è venuta incontro alle istanze dell’Amatori Avifauna Autoctona – Onlus che, nel tempo, con varie riunioni, fra i tanti argomenti da trattare, ha accettato l’in-
All. n.1: Diagramma - Camp.Italiano 2024, foto: F.O.I.
All. n. 2: Copertina Libro “Il Fanello”, foto: S.Paternò
serimento di discussioni sul Fanello ed in particolare ha preso atto di quanto segue:
1)10.01.2013: Viene dato alle stampe il primo ed unico libro – monografia sul Fanello (all.n.2)
2)01.04.2013: il Presidente della F.O.I. Salvatore Cirmi fa dono del libro a tutti i Giudici-EFI-FOI;
3)26 agosto 2018: riunione in Mascali – Fondachello presso il locale Brezza di Mare; uno dei tre argomenti all’ordine del giorno era proprio la prima proposta/ipotesi per uno standard sul Fanello (all.n.3);
4)25 agosto 2024: in Letojanni, presso il Museo civico Durante, uno dei tre argomenti posti all’ordine del giorno
è stato propriamente “Presentazione standard: Il Fanello (Linaria cannabina), relatore Sebastiano Paternò” (all.n.4);
Dopo ampia discussione, il Presidente della CTN-EFI, Ing. Carmelo Montagno, ha precisato che avrebbe inviato il lavoro al Consiglio Federale FOI per la delibera finale di accettazione e in tal modo renderlo di dominio pubblico. Lo standard si conclude con la precisazione che il piumaggio estrale, in ambiente domestico, è totalmente diverso da quello che si vede in natura. E in ambiente naturale tale è la differenza fra i due piumaggi che i primi tassonomisti considerarono l’uccello in piumaggio estrale specie diversa dall’uccello con piumaggio eclissale. In effetti, in primavera assume toni di colore estremamente diversi sia nel dorso, che varia in un colore castagna molto vivo, sia nella fronte e petto, il cui colore vira, nel maschio, dal rosso mattore ad un rosso cremisi molto vivo ed appariscente, che non si riesce ad ottenere in ambiente controllato.
Prove con alimenti pigmentanti
In ambiente domestico molte prove sono state fatte per pervenire a quel rosso cremisi così caratteristico. Si sono usati pastoncini con additivi pigmentanti sia per uccelli a piumaggio melaninico come anche per uccelli a vestito lipocromico. Carophil, Xantophil, Betacarotene, more, gelsi neri, carote, radici di Rubia tinctoris ecc. Il colore viene magnificamente acquisito più o meno intensamente e si avvicina molto al colore pettorale naturale dei soggetti con piumaggio invernale. Ma è totalmente diverso dal rosso cremisi primaverileestivo naturale. È per tale motivo che la specie risulta particolarmente interessante e in parecchi le dedicano maggiore attenzione rispetto ad altri uccelli della stessa famiglia.
Anche il sottoscritto, nelle osservazioni naturalistiche, è partito dal fatto che i Fanelli nel periodo di fine inverno-inizio primavera prediligessero alimentarsi di semi immaturi di Mercorella comune (Mercurialis annua) e in primavera di quelle che restavano più piccole e ingiallite rispetto a quelle che avevano uno sviluppo più pronunciato. Effet-
All. n. 3: Riunione Fondachello/Mascali - Proposta Standard, foto: S.Paternò
All. n. 4: Riunione Letojanni - Presentazione Standard, foto: S.Paternò
tuando ulteriori ricerche, è stato constatato che trattasi di piantina considerata velenosa e “pigmentante” per via di un alcaloide presente, chiamato hermidina, che si ossida facilmente all’aria dando origine ad una tipica colorazione blu. Anche Charles Darwin si interessò al fenomeno per via dell’interesse, nell’Ottocento, sui coloranti. Peccato che il risultato fu una colorazione blu per tingere i blue-jeans. Quindi pigmentante sì, ma verso il blu, non verso il rossocremisi.
Un’altra caratteristica di questa pianta è il fenomeno dell’androdoicia, ovvero presentare all’interno di una stessa popolazione piantine maschili e piantine più sviluppate ma ermafrodite. Aumenta quindi l’attenzione e il rebus, se c’è, verso questa pianta “pigmentante” blu.
Negli anni, in ambiente domestico, insieme alla consueta alimentazione sono stati forniti semi immaturi di tale pianta, presenti in coppia ascellare (all.n.5) e da non confondere coi fiori maschili riuniti in spiga. In definitiva, senza alcun risultato positivo.
Le osservazioni continuano e resta un vero enigma come le piume pettorali ed anche frontali non in crescita ma già a crescita completa, formata e pigmentata già in agosto, possano virare di colore in seguito, da febbraio/marzo, proprio nel periodo in cui la Mercorella comincia a offrire i suoi semi immaturi, tanto apprezzati per l’imbecco dei nidiacei, nel cui gozzo si vedono in trasparenza già a metà marzo e anche prima in Sicilia. Se c’è una relazione di causa-effetto prima o poi verrà fuori.
All. n. 5: Semi di Mercurialisannua, foto: S.Paternò
testo di MAURO CANDIOLI, foto Scuola materna G. B. Panizza di Lizzana
L’interesse per il mondo degli uccelli è sempre vivo e mantiene il suo fascino. Questa volta i piccoli pennuti sono stati richiesti alla Scuola Materna “Don G.B. Panizza” di Lizzana, in provincia di Trento, dove sono stati accolti dagli altrettanto piccoli e curiosissimi bambini. Le maestre della scuola, già entusiaste dell’attività da noi promossa durante l’anno scolastico 2022/2023, ci hanno chiesto di ripetere l’esperienza presso la loro scuola. L’As-
I piccoli pennuti sono stati richiesti alla Scuola Materna di Lizzana
sociazione Ornitologica Trentina di cui faccio parte non poteva, pertanto, esimersi da questo importante invito. Così io e il nostro instancabile segretario, Gianni Bonatti, compatibilmente con gli impegni di lavoro e le mostre ornitologiche che in quel periodo abbondano, siamo riusciti ad organizzare una mattinata all’insegna dei nostri amici pennuti. Mercoledì 20 novembre, di buon mattino, abbiamo preparato i nostri beniamini nella sala dedicata alle attività artistico-espressive, dividendoli principalmente in due categorie: gli indigeni e i non indigeni. Negli indigeni abbiamo presentato cardellini, lucherini, verdoni e fringuelli, spiegando le
loro principali caratteristiche riguardanti l’alimentazione, il loro areale abituale e soprattutto il dimorfismo sessuale. Nel secondo gruppo erano presenti i canarini gialli, rossi e mosaico giallo, un ciuffato tedesco ed alcuni messicani. I vivaci colori e gli acrobatici atteggiamenti dei nostri piccoli beniamini hanno subito attirato l’attenzione dei piccoli visitatori, che hanno posto diverse domande per soddisfare le loro più svariate curiosità.
Gianni Bonatti e Mauro Candioli
Il Lucherino ventre giallo
(Spinus xanthogastrus)
testo di FRANCO FORMISANO, foto F. FormisanoeB. Zamagni
Introduzione
Più o meno un trentennio fa, le mie preferenze ornitologiche virarono decisamente verso gli Spinus, dopo aver allevato per anni Canarini domestici (i sassoni - così veniva chiamato allora il Canarino di colore -) nonché numerose specie di Estrildidi presenti all’epoca sul mercato. Gli Spinus stazionavano da tempo in cima alla lista delle mie preferenze, grazie anche al debole che ho sempre avuto, oggi come allora, per il Lucherino eurasiatico; un debole che, come ho già avuto modo di dire in altre occasioni, data sin dall’infanzia.
Iniziai così a “raccogliere” quello che la piazza offriva di compatibile con le mie, allora, scarse finanze e oltre allo Spinus indigeno, da sempre presente, entrarono man mano a far parte della mia “scuderia”, con altalenanti risultati riproduttivi, il Lucherino di Magellano, il Cardinalino del Venezuela, il Lucherino dal mento nero e da un paio di lu-
Gli Spinus stazionavano da tempo in cima alla lista delle mie preferenze, grazie anche al debole che ho sempre avuto, oggi come allora, per il Lucherino eurasiatico
stri a questa parte anche quello che tra gli Spinus mi ha sempre intrigato più di ogni altro, non fosse altro per il nome comune col quale veniva indicato: lo “xanthogaster”. Con alcuni amici, coi quali a quel tempo condividevo quella passione, due giorni fissi a settimana, il lunedì e il venerdì, ci si vedeva presso un’uccelleria qui in provincia di Napoli, famosa per la vasta gamma di prodotti e per la
grande varietà di specie di volatili disponibili. Ricordo che di tanto in tanto capitava un tizio, sulla cinquantina, napoletano di Napoli - quartieri alti - non come noi provinciali..., il quale si distingueva perché si dava delle arie tipo snob e per la “erre” pronunciata moscia, una sciccheria che ben calzava al personaggio. Che fosse facoltoso lo si intuiva dal mezzo col quale viaggiava secondo stagione, auto di lusso e moto di grossa cilindrata, e dai capi d’abbigliamento firmati, indossati, tra l’altro, con naturale eleganza.Nessuno conosceva il suo nome, ma per noi, quando lo si citava, era “‘o francese” a causa di quella erre particolare di cui era dotato; nemmeno si sapeva quale fosse la sua professione, ma dalla dialettica e dalla sintassi corretta eravamo dell’idea che fosse un libero professionista, oppure un dirigente o un professore, comunque un intellettuale. Persona alquanto simpatica (nonostante l’aria da snob), linguaggio for-
Uova gallate
Pullus appena nato
Gusci d'uova portati lontano dal nido dalla femmina
bito, parlantina fluida, si lasciava ascoltare ben volentieri ed io, essendo alle prime armi, prestavo la massima attenzione alle sue parole, anche perché il tizio allevava Spinus e, a suo dire, anche di un certo livello. Volendo trovargli un difetto, non vi era discorso in cui non ficcasse, fra i suoi tanti blah blah blah, il termine xanthogaster: “I miei xanthogaster di qua…i miei xanthogaster di là…”diceva, e noi tutti in religioso silenzio ad ascoltare e annuire. Fatta la spesa, sempre semi di qualità e “strani”, salutava stringendo la mano a tutti gli astanti e io, come del resto gli altri, anche se nessuno per pudore osava ammetterlo, lo guardavo andar via chiedendomi come al solito "che xa..o fossero 'sti xanthogaster" che allevava, ripromettendomi con un sussurro: “La prossima volta che viene, giuro, glielo chiedo...”. Ma, a forza di giurare e rimandare, purtroppo non ci fu mai “una prossima volta” perché “‘o francese” tutto ad un tratto sparì. Pare si fosse trasferito al Nord per motivi di lavoro. Qualcuno ipotizzò addirittura che avesse smesso di allevare; personalmente ho sempre ritenuto valida la prima ipotesi, vista la passione che palesava per i suoi Spinus. Fatto sta che lo xanthogaster rimase per me un autentico mistero ancora per un bel po’, fino a quando non entrai in possesso del libro “I Fringillidi”, edito dalla F.O.I., afirma di uno dei più bravi e preparati tecnici italiani del settore nonché allevatore, che risponde al nome di Renzo Esuperanzi.
Biologia generale
Piccolo grazioso fringillide descritto come Chrysomitris xanthogastra nel 1855 da Bernard Aimè Léonard Du Bus De Gisignie, meglio noto nel mondo accademico, in maniera breve e concisa, come Du Bus (e meno male!), il Lucherino ventre giallo è originario del continente americano; infatti, vive in Costa Rica, a Panama e nella parte nord delle regioni settentrionali dell’America Latina come la Colombia ed il Venezuela oltre che nell’Ecuador occidentale (irregolare), con laforma nominale Spinus x. xanthogastrus;nelleestreme regioni sudorientali del Perù fino alla Bolivia
centrale compare la sottospecie Spinus x. stejnegeri
Taglia 10 cm; netto il dicromismo tra i sessi: il maschio presenta le parti superiori completamente nere, dalla fronte alla coda; quelle inferiori sono caratterizzate per metà dal nero, che senza soluzione di continuità dalla testa si estende fino al petto, formando una sorta di grande cappuccio; le restanti parti dall’addome fino al sottocoda sono di un color giallo (da qui il nome comune); becco grigio antracite, codione giallo.
La femmina presenta una colorazione verde oliva quasi uniforme e, come il maschio, lo specchio alare di colore giallo, ma di minore ampiezza.
Specie sedentaria o stanziale, erratica in alcune aree, ha fatto registrare da anni una notevole contrazione numerica in natura causa la persecuzione operata dai trappolatori ai fini com-
La femmina presenta una colorazione verde oliva quasi uniforme e, come il maschio, lo specchio alare di colore giallo, ma di minore ampiezza
merciali, stante la richiesta del mercato come uccello da gabbia e il degrado ambientale. La fascia geografica dove alligna è tipicamente tropicale e subtropicale; in questo contesto il biotopo è composto in prevalenza da foresta umida, ai margini della quale vi sono radure con pascoli ed alberi radi o arbusti, e piantagioni e pascoli di altura tra i 1400 e i 2000 metri, con punte estreme comprese tra gli 800 e i 3700 metri slm. Gregario, si sposta in piccoli gruppi al di fuori del periodo riproduttivo, spesso in compagnia del Negrito di Bolivia, là dove gli areali di distribuzione sono condivisi.
Prevalentemente arboricolo, ama sostare sulle cime degli alberi e come tutti gli altri Spinus si alimenta prele-
Pulli alla schiusa
Pulli a 3gg. dalla schiusa
Pulli a 5 gg. dalla sciusa
vando direttamente sulla pianta i semini immaturi o secchi, secondo la stagione, e si porta al suolo solo per l’abbeverata e per le abluzioni. Le preferenze alimentari vanno dai semi degli alberi endemici della regione a quelli delle composite; durante il periodo riproduttivo, quando ci sono i piccoli nel nido, larve di insetti e piccoli invertebrati sono cacciati con tenacia. Come detto poc’anzi, il clima dell’areale di distribuzione è tipicamente tropicale, quindi mite e con temperature che non presentano grosse variazioni termiche, se non in quota; praticamente assenti quelle riferite alla durata delle ore luce, pertanto a “scatenare” l’estro è solo la disponibilità del cibo, abbondante al termine della stagione piovosa. Due, eccezionalmente tre, le covate annue; il nido, costruito dalla sola femmina a forma di coppa, esternamente di pregevole fattura e ben rifinito all’interno con pelo animale o lanugine vegetale, viene assemblato nel folto dei rami degli alberi o arbusti ad altezza variabile ma comunque sempre considerevole. Mediamente quattro le uova deposte (2/6 gli estremi), incubate dalla sola femmina, si schiudono dopo 12 giorni di cova costante.In natura, alla schiusa
Prevalentemente arboricolo, ama sostare sulle cime degli alberi e come tutti gli altri Spinus si alimenta prelevando direttamente sulla pianta i semini immaturi o secchi
delle uova, la femmina preleva i gusci vuoti dal nido e li trasporta lontano da dove quest’ultimo è situato; questa operazione (comune a tutte le specie) ha lo scopo, non secondario, di depistare eventuali predatori quali rettili e piccoli roditori inprimis, ma anche i domesticissimi gatti.I pulli alla schiusa sono ricoperti da fitto piumino grigio; il maschio nei primi giorni nutre la compagna al nido mentre essa continua ad accudire i piccoli, ma dopo alcuni giorni entrambi i genitori provvedono al loro sostentamento. L’involo avviene intorno al 18° giorno di età e ne occorreranno più o meno altrettanti prima che si rendano indipendenti. Considerato quanto sono precoci i piccoli dei conspecifici
nell’emanciparsi dai genitori, gli juveniles sono da ritenersi degli autentici “bamboccioni”. Comunque, è compito del maschio completare lo svezzamento della nidiata, mentre la femmina cova la successiva deposizione. Emancipatisi, i novelli si imbrancano con coetanei conspecifici e non, formando piccoli stormi erranti alla ricerca delle zone di pastura. Questo è quanto accade, più o meno, allo stato brado.
Allevamento
Il Lucherino ventre giallo presente oggi sul mercato è solo di provenienza domestica, ma già prima della chiusura delle importazioni decretata dalla U.E. nel 2005causa influenza aviaria, la specie raramente veniva importata nel nostro Paese. Tutt’altra storia, però, sul finire degli anni ‘80 del secolo scorso, quando l’importazione era stata massiccia, data la grande richiesta da parte dei canaricoltori del colore, i quali, visti i lusinghieri risultati ottenuti molti anni prima con il Cardinalino del Venezuela, nel travasare i geni per il rosso dall’esotico al Serino domestico, grazie alla parziale fecondità degli ibridi maschi ottenuti tra le due specie, ben pensarono di sfruttare
Controllo anellinoNidiacei
la forte “negritudine” dello xanthogaster nella corsa al canarino nero, considerata la buona percentuale di fertilità palesata anche in questo caso dagli F1 maschi. Ma il nostro beniamino, purtroppo, di nero ne trasmetteva ben poco; così, dopo molti e infruttuosi tentativi, il “progetto” venne accantonato e l’allevamento (in purezza) del Lucherino ventre giallo, intrapreso solo da pochi e previdenti appassionati, divenne di nicchia. Grazie a questi signori oggi si può, con relativa facilità, acquisire coppie nate in ambiente controllato, esenti da ogni forma patologica che purtroppo accompagnava tutte le specie importate, dovuta sia allo stress per la cattura, sia alle pessime condizioni igieniche degli angusti contenitori nei quali viaggiavano, stabulati in numero eccessivo se rapportato alla capienza.
Esperienza personale
Allevo questa interessante specie da una decina di anni con alterne fortune riproduttive. La prima coppia la reperii presso l’amico e conterraneo Giuseppe Mellonesul finire del 2014: femmina novella, maschio adulto di due anni. Ricordo che li alloggiai, non appena giunto a casa, contrariamente
A partire da gennaio, inizio a fornire a tutti gli ospiti del mio piccolo allevamento le tarme della farina (Tenebrio molitor) come preparazione (proteine nobili) all’approssimarsi della stagione riproduttiva
al solito, benché fossimo già a novembre inoltrato, in una voliera esterna poiché si presentavano robusti e rustici. L’alimentazione fin da subito fu un misto composto in prevalenza dalla scagliola e poco altro, adeguato, a mio parere, allo stato captivus dei volatili; un mix leggero e spartano che uso da sempre per tutti i miei fringillidi. Praticamente li tengo a “pane e acqua”, come osserva qualcuno tra il serio e il faceto, ricordandomelo ad ogni occasione in cui si parla di alimentazione dei pennuti, mentre fornisco 2/3 volte a settimana e in contenitore a parte
un cucchiaino a testa dei cosiddetti semi condizionanti o della “salute”, che miscelo personalmente acquistando i semini in singole quantità e sottoponendoli al test di germinazione (solo se superano la prova verranno poi somministrati), oggi meglio conosciuti, revisionati e corretti con l’affascinante e suadente nomignolo di “pance chiare”.
A partire da gennaio, inizio a fornire a tutti gli ospiti del mio piccolo allevamento le tarme della farina (Tenebrio molitor) come preparazione (proteine nobili) all’approssimarsi della stagione riproduttiva; devo dire che la coppia di xanthogaster li accettò subito e di buon grado, nonostante fosse un’autentica novità per loro (Giuseppe non ne fa uso). Ne fornisco una/die a testa a settimana, sia alle coppie alloggiate all’interno nel locale dove allevo sia a quelle poste all’esterno in gabbioni e voliere modulari con fondo in terra naturale e in modo esponenziale man mano che si va verso la primavera, fino ad un massimo di trevolte a settimana. Il portanido fornito per questa specie è uno dei più piccoli, del tipo usato per il Cardinalino del Venezuela e, ai bei tempi che furono, anche per i piccoli Serinus africani quali il Cantore
Nido con pullo mutato BrunoFemmina Bruna
d’Africa, il Canarino del Mozambico, il Canarino gola nera... La riproduzione del Lucherino ventre giallo non si discosta molto da quella concernente gli altri Spinus, stimolata dal corteggiamento serrato del maschio, che mostra tutto il suo ardore alternando furiosi inseguimenti all’ultimo respiro a offerte di cibo e coccole varie, a cui segue l’esecuzione del repertorio canoro, sciorinato dal posatoio più in alto ad ali abbassate per meglio mostrare il codione; la femmina, in capo a pochi giorni, inizia a girare con sfilacci di juta o steli essiccati di Poa comune stretti nel becco. In poco tempo viene assemblato un nido di pregevole fattura nel cestino ed entro un paio di giorni avviene l’ovodeposizione (di solito tre uova); è buona norma sostituire con l’uovo finto, così, rimesse poi tutte assieme, si avrà una schiusa simultanea che normalmente avviene al 12° giorno di incubazione. Alla schiusa, la femmina traporta i gusci d’uovo lontano dal luogo dove è situato il nido, intendendo con ciò confondere eventuali predatori, così come avviene in natura. Sono alimentati nei primi giorni di vita dalla sola femmina, mentre il maschio, se presente, si limita ad imbeccare la com-
pagna al nido; alcuni giorni più tardi entrambi i genitori provvedono al loro sostentamento e i pulli hanno poi un buono sviluppo corporeo, pertanto è bene inanellare verso il sesto giorno dalla schiusa (per questa specie è previsto l’anellino del tipo A).
Per tutte le specie di fringillidi che allevo, di solito lascio fare non più di due covate annue, ed il Lucherino ven-
La riproduzione del Lucherino ventre giallo non si discosta molto da quella concernente gli altri Spinus, stimolata dal corteggiamento serrato del maschio, che mostra tutto il suo ardore alternando furiosi inseguimenti all’ultimo respiro a offerte di cibo e coccole varie
tre giallo non sfugge a questa regola. Come per diverse specie di Spinus, quasi sempre bisogna separare la coppia una volta che la femmina ha deposto, anzi spesso ciò è la regola, perché in gabbia il maschio si rivela un pessimo partner: focoso fino all’eccesso, cerca di copulare anche mentre la compagna è sul nido e ciò le causa stress e tante volte la scondiziona al punto tale da indurla all’abbandono del nido con le uova, con tutte le ovvie conseguenze. Inoltre, coppie che l’anno prima sembravano affiatate e i due partners fatti l’un per l’altro, in capo a pochi mesi, appena dopo la muta, iniziano a mostrare insofferenza reciproca, mandando in fumo tutti i sogni e i progetti fatti da chi con tanta passione si dedica a loro. Spesso, però, i ruoli sono invertiti; infatti, la femmina rincorre il maschio costringendolo all’angolo e gliele “suona di santa ragione”; in presenza di questo evento è opportuno separarli senza indugio (ogniqualvolta ho procrastinato questa mossa è capitato di ritrovare uno dei due passato a miglior vita, sul fondo del contenitore). Conviene separare tramite divisorio in fil di ferro, ma, a volte, può darsi che anche questo accorgimento non basti; quando
Foto e allevamento: B.Zamagni
Foto e allevamento: B.Zamagni
cercano di “darsele” anche attraverso le sbarrette, personalmente li separo e allontano definitivamente il maschio ponendolo in una gabbia e sistemandolo in un’altra stanza. La femmina si fa carico di provvedere a nutrire i nidiacei fino alla loro indipendenza (che avviene un po’ più tardi rispetto agli altri Spinus di comune allevamento).
In questo periodo fornisco pastone morbido all’uovo del commercio (in verità poco gradito, eccezion fatta per i primi giorni di vita dei pulcini), perilla, girasole piccolo nero, semi di lattuga e chia, somministrati alternandoli in
gesta cuticola (esoscheletro). Col passar dei giorni e man mano che i nidiacei crescono, ortaggi coltivati personalmente con metodi biologici vengono aggiunti alla dieta, come fettina di cetriolo e zucchina, foglia di radicchio, di lattuga e pomodorino tagliato a spicchio; le erbe prative sotto forma dei capolini o spighe con i semini allo stato ceroso, sia selvatiche, sia coltivate.Purtroppo, non tutti i Lucherini gradiscono: alcuni si specializzano su un alimento, rifiutando tutto il resto e per evitare il peggio do un piccolo aiuto con imbeccate supple-
contenitore a parte;fettina di mela due volte al giorno, capolini di dente di leone stracarichi dei semini allo stato ceroso, particolarmente per le prime nidiate (dal sesto giorno di vita fornirò anche le foglie di questa salutare pianta); larve di Tenebrio molitor (da me allevate curando in maniera maniacale l’igiene dei contenitori),dal secondo giorno dalla schiusa e fino all’impiumo in misura di una/die per pullus in rapporto alla loro crescita, fino ad un massimo di 3/die, somministrate quando sono bianche, appena mutate, quindi prive della dura e indi-
mentari tre volte al giorno, usando un apposito prodotto per l’imbecco a mano dei nidiacei.
Da alcuni anni,dietro suggerimento di un amico, uso durante la riproduzione un recente (per me) ritovato: le perle morbide. Confesso che sono sempre stato restio a queste “novità”, ma come si dice“Mai dire mai!” . Infatti, nonostante il mio scetticismo, le suddette perle sono bene accette e ad ogni controllo ritrovo i pulli sempre belli sazi, col gozzo pieno e un fondoschiena ricco di adipe,anche se continuano ad emanciparsi sempre intorno
ai quaranta giorni di età. Resisi indipendenti, lialloggio all’interno nel locale dove allevo, in un contenitore da 120 cm; nel gabbione possono effettuare brevi e salutari voli. In questo contesto, la fornitura del contenitore con l’acqua per le abluzioni (affinché la muta sia veloce e regolare) fissato ad una delle porte laterali è quotidiana.
Quest’anno (2024), in sintonia con la maggioranza, per non dire con la totalità delle specie allevate e sulla falsariga degli anni scorsi (sempre peggio), i miei Lucherini ventre giallo mi hanno dato tanti “ dolori” e pochissime “gioie”; infatti, dall’unica coppia in mio possesso ho ottenuto unicamente due piccoli, di cui uno solo portato all’indipendenza (femmina) e, come a voler rimarcare l’annata particolare, verso la metà di agosto, nel giro di pochi giorni e senza alcun segno apparente, sono morti entrambi i riproduttori. Nel corso di questi anni, per formare le coppie, sono stato costretto a inserire diversi soggetti esterni in prevalenza provenienti dall’allevamento degli amici Saverio Sirignano e Bruno Zamagni. Confesso che ancora oggi osservare lo xanthogaster nel quotidiano del mio allevamento scatena in me particolari emozioni, mentre la mente corre, non senza nostalgia, a quelle lontane spensierate serate passate tanti anni fa ad ascoltare, attento e incantato, le parole “d’o’ francese...”
Mutazioni
Diverse mutazioni hanno interessato il piumaggio di questo Spinus: fissate al momento sono la Bruno, la Silice, la Diluito e la Pastello. Personalmente, diversi anni fa, mi son ritrovato tra le mani (pardon, in un nido) una femminuccia mutata Brunonata da una coppia di ancestrali, composta da femmina del mio allevamento e un maschio frutto di uno scambio con un amico, il quale era senz’altro portatore di tale mutazione. L’amicoBruno Zamagni da un po’ di tempo ha travasato, fissandoli, in questo Spinus i geni della mutazione Silice (Giallo cecoslovacco), comparsa alcuni anni fa nel Lucherino eurasiatico. La denominazione Silice,
Novelli 2024
ufficialmente riconosciuta, si deve all’intuizione di quell’autentico fuoriclasse dell’ornitologia nostrana quale èRenzo Esuperanzi, il quale, dietro mia esplicita richiesta, mi ha suggerito (e non è la prima volta!) questo piccolo paragrafo concernente le mutazioni. Senza il suo aiuto, le mie conoscenze sarebbero state limitate alla mia Bruna di qualche anno fa e alla più recente Silice di Bruno.
A tal proposito, per chi fosse interessato: Bruno ha, in modo chiaro ed esaustivo, esplicitato questo suo lavoro in un articolo apparso su I.O. n° 3/2021,partendo da meticci ottenuti con il Lucherino eurasiatico Silice per Lucherino ventre giallo ancestrale. Da questo accoppiamento si ottengono già in prima generazione soggetti mutati (la mutazione Silice è dominante); successivamente, ha ottenuto degli R1 e nel mentre ha constatato la fertilità totale anche dei meticci sia F1
Diverse mutazioni hanno interessato il piumaggio di questo Spinus: fissate al momento sono la Bruno, la Silice, la Diluito e la Pastello
che R1 femmine, ottenendo definitivamente, nell’arco di otto anni circa, Lucherini ventre giallo Silice puri.
Conclusioni
Che dire? Atteso che allevare lo xanthogaster (amo chiamarlo ancora alla vecchia maniera) è un vero piacere, come per tutti gli altri Spinus d’altronde, questo piccolo fringillide ab-
bina alla sobrietà delle cromie della livrea la taglia minuta e una vivacità innata, oltre che una certa eleganza, doti che gli consentono di avere molti estimatori tra gli allevatori di E.F.I. Sento il dovere di ringraziare quei pochi “pionieri” che, decenni or sono, quando fu chiaro che il nostro beniamino nella corsa al canarino nero non sarebbe stato di alcuna utilità, si cimentarono nel suo allevamento in purezza. È grazie al loro appassionato lavoro se oggi il Lucherino ventre giallo è disponibile in numeri abbastanza rilevanti, considerato anche come è andata a finire con le importazioni dei volatili esotici dal 2005 in poi, con la faccenda dell’influenza aviaria.
BIBLIOGRAFIA
-I Fringillidi, R. Esuperanzi, Edizioni F.O.I., Piacenza, 1996
-Finches & Sparrows, P. Clement, A. Harris, J. Davis, C. Helm Guides, Ltd. 1993, London
R ecensioni
Gli ibridiAllevare opere d’arte
Autori:Renzo EsuperanziAntonio Javier Sanz
Edizioni:Engonari, Valencia 2024.
Pagine:428
Lingue:Italiano + Francese
Spagnolo + Inglese
Costo:55 euro
Formato:19 x 24
Corredato da oltre300 foto a colori
di PIERCARLO ROSSI
Questo splendido libro nasce dalla collaborazione tra un luminare degli ibridi come Renzo Esuperanzi ed un grandissimo fotografo come Antonio Javier Sanz. Il risultato che scaturisce da tale connubio è magnifico, sia nel testo sia nelle illustrazioni.
Il testo, che si apre con una raffinata prefazione a cura di Ivano Mortaruolo, racconta in maniera semplice e discorsiva tutto quanto c’è da sapere per favorire le ibridazioni degli Estrildidi e dei Fringillidi, per realizzare ibridi di grande interesse estetico e per portarli al top delle condizioni per le esposizioni. Il tutto passando per la classificazione delle specie, per la storia, le regole e le motivazioni di base dell’ibridazione, senza tralasciare l’alimentazione, la scelta dei riproduttori, l’alloggiamento, la preparazione dei soggetti alle mostre e gli accorgimenti antistress da adottare al loro rientro in allevamento.
Ognuno dei nove capitoli, con innumerevoli suggerimenti, aiuta il lettore a districarsi nella scelta dei soggetti e delle combinazioni da tentare per ottenere ibridi belli, rari e scientificamente interessanti. In particolare, nel sesto capitolo vengono approfondite le ibridazioni con il Canarino domestico, mentre nel settimo capitolo si parla delle ibridazioni con i Fringillidi e nell’ottavo ci si focalizza sulle ibridazioni tra Estrildidi. Tutti i capitoli sono copiosamente illustrati con fantastiche foto di ibridi bellissimi,
ripresi con grande maestria nei minimi particolari e nella posizione migliore, onde poterne ammirare ogni singola caratteristica. In conclusione, il libro è fantastico ed irresistibile da sfogliare, ma anche ammaliante e trascinante nella lettura.
Il libro è prenotabile accedendo al sito dell’editore, sul quale è anche possibile prendere visione di alcune pagine dello stesso: https://it.libros-engonari.com/product-page/gli-ibridi-les-hy brides
R ecensioni
Border - storia di un’amicizia
Autore: Luigi Malagoli
Editore: Etabeta - Pagine: 172
Acquistabile su Amazon e in libreria (su prenotazione € 17,50)
di MARA ZANICHELLI
“Border - storia di un’amicizia” è un romanzo in cui l’autore narra la storia dell’amicizia nata tra Luigi e Bruno, i due protagonisti principali dell’opera. Il primo è un ragazzino di dodici anni che frequenta la seconda media, mentre Bruno è un adulto ormai in pensione. Il mondo dell’ornitologia fa da sfondo all’intera vicenda. Bruno, che è un vecchio allevatore di border, una razza di canarini originaria del Regno Unito, conosce
Luigi e poco alla volta, durante l’intero ciclo riproduttivo dei propri beniamini, gli fa conoscere tutti i rudimenti dell’hobby che coltiva ormai da cinquant’anni. Luigi, andando a trovare il suo maestro quasi quotidianamente, segue dalla nascita, allo svezzamento, alla muta, fino alla partecipazione al campionato italiano di ornitologia il percorso di vita di “Capinera”, un border novello che Bruno gli ha dato in dono. Leggendo quest’opera traspare tutta la passione, la dedizione e la competenza dell’autore per la canaricoltura, da lui considerata un’arte. Consiglio la lettura di questo romanzo a tutti coloro che, come il suo autore, sono appassionati di ornitologia e credono nel sentimento dell’amicizia.
La consacrazione di Fringillia -Valconca
Con l’alto patrocinio del Parlamento Europeo
testo di PIETRO RUSSO, FOTO A.A.A. E A.O.P.
Edizione da record a Morciano di Romagna, che si conferma tra le mostre più importanti d’Europa: 3332 soggetti, di cui oltre 2200 tra fringillidi esotici, fringillidi europei ed ibridi e l’alto patrocinio del Parlamento Europeo (per la prima volta nella storia
3332 soggetti, di cui oltre 2200 tra fringillidi esotici, fringillidi europei ed ibridi
delle associazioni ornitologiche). Si può senza dubbio riassumere così la vincente accoppiata tra Fringillia e Valconca, le oramai storiche esposizioni ornitologiche frutto di una splendida e proficua collaborazione tra l’Associazione Adriatica Allevatori e l’Associazione Pesarese
I Presidenti dell'A.A.A. e A.O.P. Valentini e Mazzanti premiano Pietro Agostini
Tangaramexicana, Best in Show con 95 punti
Il riconoscimento ad Aldo Minardi
La mostra scambio
Uno dei trofei col patrocinio del Parlamento Europeo
Ornicoltori, che hanno fatto registrare il record assoluto di partecipazioni. Ma parlare solo di expo e record di partecipanti sarebbe restrittivo, poiché la realtà è ben più rosea. Fringillia-Valconca va infatti ben oltre l’essere l’esposizione specialistica con una qualità eccelsa di soggetti in gara. Fringillia-Valconca è, anno dopo anno, l’emblema dell’aggregazione, dell’amicizia, della vita conviviale, tanto che oramai la frequentano anche i non allevatori, con le mogli e i figli (e chi alleva sa benissimo cosa significhi coinvolgere le famiglie in questo hobby). Non è stata facile per i 33 giudici che si sono trovati a giudicare il meglio del meglio dei soggetti europei e che avranno certamente faticato per sancire il podio in ogni categoria, vista l’eccellenza esposta. Vedere soggetti come le Tangare, vedere ibridi rari e difficili come Canarino Ventre Bianco x Crociere Fasciato, o Crociere Fasciato x Ciuffolotta, contemplare le decine di varietà di Cardellini, tra cui quello nero (esposto per la prima volta in assoluto in una mostra ornitologica), le mutazioni dei Verdoni europei e la qualità di quelli asiatici, l’innumerevole quantità di Spinus sudamericani e doverli giudicare non è stato per niente facile. Posizionarsi al centro del padiglione della Fiera di Morciano di Romagna e guardarsi intorno è stata una delle sensazioni più appaganti. Non si erano mai visti tanti Diamanti di Gould, tanti Canarini selvatici; non si era mai vista una coppia ‘non cartacea’ di Fanello dello Yemen. Tutti i colori del mondo in un padiglione adibito ad expo e in un altro, che ha ospitato una altrettanto valida mostra scambio con soggetti inediti come i Verzellini d’Arabia (Crithagra rothschildi).
In vetta alle preferenze degli appassionati, sicuramente il classico Cardellino e tutte le varietà di Spinus sudamericani, Xantogaster e Cardinalini su tutti. Interessantissimo ed educativo il dibattito sul Lucherino e sul Verdone nelle mutazioni agata e isabella, tenuto dal presidente della C.T.N. - E.F.I. Carmelo Montagno Bozzone, che ha confermato l’altissima qualità dei soggetti a con-
corso appartenenti a queste due specie, con un’evoluzione continua delle varietà fenotipiche. Il riconoscimento del Parlamento Europeo ha contribuito a consacrare l’expo, che in realtà gode da anni di una bella fama. Ma oltre i pennuti, Fringillia-Valconca ha offerto i paesaggi, l’enogastronomia, le rocche e la storia di una terra bellissima che, tra mare e colline, ha pochi rivali. Una cena di gala sontuosa e soprattutto divertente, con 150 commensali provenienti da tutta Europa, ha coronato questa “tre giorni” che oramai rappresenta un appuntamento fisso per chiunque ami il connubio cultura-ornitologia. Oltre agli espositori italiani, Fringillia-Valconca ha visto appassionati provenienti da Slovenia, Polonia, Germania, Spagna, Grecia, Belgio e Olanda. Uno spettacolo nello spettacolo potersi confrontare con le differenti culture europee e i metodi d’allevamento. Fringillia-Valconca ha infatti stretto partnership con il Dutch Carduelis Club dall’Olanda, l’Internationaler Cardueliden Club e.V. dalla Germania, il Polskie Stowarzyszenie Hodowców Awifauny Europejskiej dalla Polonia e i gruppi di allevatori dalla Sardegna, il club degli amici provenienti dalla Croazia, sempre presenti da anni oramai.
La qualità è stata confermata dalla specialistica del Club Italiano del Diamante di Gould e da quella del Club del Canarino Arricciato Gigante Italiano, con soggetti strepitosi.
Una expo in continua crescita che ha soddisfatto davvero tutti e che, visto il trend, continuerà ad affermarsi tra le più importanti mostre del Vecchio Continente.
Il giudizio della specialistica A.G.I.
La serata conviviale tra i partecipanti
La delegazione di allevatori polacchi con Massi Esposto
Mario Vetere e Marcin Cichy
O rniFlash
Questo stupendo uccello esotico ha un segreto: si illumina al buio
Questa
straordinaria scoperta, documentata in uno studio recentemente pubblicato su Royal Society Open Science, è il risultato della ricerca condotta dalla biologa Rene Martin dell’Università del Nebraska-Lincoln.
La scienziata, incuriosita dalla rarità del fenomeno della biofluorescenza tra gli uccelli, ha deciso di esplorare in modo sistematico questa possibilità negli uccelli del paradiso, esaminando una vasta collezione di esemplari conservati all’American Museum of Natural History, alcuni dei quali raccolti fin dal 1800. I risultati hanno superato ogni aspettativa: 37 delle 45 specie conosciute di uccelli del paradiso possiedono piume che emettono luce quando esposte a radiazioni ultraviolette o blu. Questo fenomeno, noto come biofluorescenza, si verifica quando onde luminose ad alta energia vengono assorbite e poi riemesse a lunghezze d’onda più lunghe e con energia inferiore. Nel caso degli uccelli del paradiso, la luce ultravioletta o blu assorbita viene trasformata principalmente in tonalità di verde o giallo-verde, creando un effetto luminoso spettacolare.
La biofluorescenza, sebbene diffusa in natura tra funghi, pesci, rettili e anfibi, è stata documentata raramente negli uccelli, rendendo questa scoperta particolarmente significativa. Gli uccelli del paradiso possiedono nelle loro piume delle molecole chiamate fluorofori che consentono questa trasformazione della luce. Inoltre, il loro sistema visivo presenta filtri specializzati nelle cellule sensibili alla luce che rendono i loro occhi particolarmente reattivi a questo fenomeno luminoso.
Particolarmente interessante è la distribuzione della biofluorescenza tra i sessi. I maschi presentano una maggiore quantità di piumaggio biofluorescente, concentrato soprattutto nella zona della testa, del collo, del petto e dell’addome. Non solo le piume, ma anche la pelle delle zampe e l’interno della bocca di questi uccelli brillano quando esposti a luce blu o ultravioletta. Le femmine, al contrario, mostrano meno piume biofluorescenti, generalmente limitate al petto e all’addome. Questa differenza non sorprende i ricercatori, considerando l’alto dimorfismo sessuale di queste specie, dove maschi e femmine hanno aspetti molto diversi. …
Elevati livelli di microplastiche nei polmoni degli uccelli
Ipolmoni degli uccelli contengono notevoli quantità di microplastiche, il che dimostra quanto l’aria sia oggi contaminata dall’inquinamento causato dalla plastica. Secondo un nuovo studio cinese, il tessuto polmonare degli uccelli contiene centinaia di particelle di materiali plastici di uso comune, come il polietilene. «Gli uccelli sono importanti indicatori delle condizioni ambientali», ha affermato uno degli autori dello studio. «Ci aiutano a comprendere lo stato dell’ambiente e a prendere decisioni informate sulla conservazione e sul controllo dell’inquinamento». Gli scienziati cinesi hanno studiato oltre 50 specie diverse di uccelli selvatici nella Cina occidentale, prelevando campioni di tessuto polmonare dagli uccelli e li hanno analizzati utilizzando la tecnologia a infrarossi per rilevare e contare le microplastiche. È stata utilizzata anche la gascromatografia-spettrometria di massa con pirolisi per identificare nanoplastiche più piccole, che possono entrare nei polmoni attraverso il flusso sanguigno. Centinaia di microplastiche sono state trovate nei polmoni degli uccelli, con una media di 221 particelle per specie e 416 particelle per grammo di tessuto polmonare. I tipi più comuni identificati erano il polietilene clorurato, utilizzato per isolare tubi e fili, e la gomma butadiene, un materiale sintetico negli pneumatici. Attualmente non esistono livelli «sicuri» stabiliti di esposizione alle microplastiche e alle nanoplastiche. …
Ipannelli solari nei terreni agricoli (o destinati all’allevamento) non sono un danno né un ostacolo alle attività economiche. Ma contribuiscono addirittura a migliorare l’ambiente circostante. Lo dimostra una ricerca condotta dagli scienziati della Royal society for the protection of birds (Rspb) e dell’Università di Cambridge e appena pubblicata.
Nell’ambito del Centre for Landscape Regeneration, gli esperti hanno rilevato attorno agli impianti solari nell’East Anglia, una zona dell’Inghilterra zona prevalentemente agricola, ospitano un numero maggiore di specie e un numero complessivo di uccelli maggiore rispetto ai terreni agricoli circostanti. Ovviamente, nulla avviene per caso. Nel caso di impianti solari il cui progetto ha tenuto conto del contesto naturale – e in aree con un maggiore mix di habitat – i risultati sono stati ancora più performanti. In questo caso, nelle aree che ospitano gli impianti fotovoltaici sono state rilevate specie fino a tre volte il numero presente nei terreni circostanti ad uso agricolo. … I ricercatori hanno esaminato due tipi di impianti fotovoltaici: quelli con un mix di habitat all’interno del parco solare e quelli con un habitat “associato” più semplice. “L’habitat semplice era gestito in modo intensivo, senza siepi lungo i confini ed era costantemente frequentato dai greggi di pecore. I parchi solari con habitat misto avevano siepi lungo i bordi, senza pecore al pascolo o taglio dell’erba, il che ha portato a una maggiore diversità di piante da fiore”. La più alta abbondanza di specie di uccelli minacciate dall’avanzare delle attività umane (come strilloni, zigoli gialli e fanelli) è stata riscontrata attorno a impianti solari con habitat misto. …
Ilgipeto è il più grande uccello delle Alpi e per anni è stato oggetto di diverse leggende: chiamato “avvoltoio degli agnelli”, si diceva che rapisse i bambini, oltre a un concorrente dell’uomo nella caccia di caprioli e camosci. Una nomea che ha portato al suo sterminio: nel 1913 l’ultimo gipeto barbuto delle Alpi fu abbattuto in Italia e per decenni non ce ne fu traccia. Negli anni ‘80 è iniziato un programma di reintroduzione, prima in Austria, poi in Francia e in Italia. Nel 1991 è stata la volta della Svizzera. Oggi la Stazione ornitologica svizzera di Sempach stima che ci siano circa 350 esemplari che vivono nelle Alpi. In particolare, l’Engadina è considerata uno degli hotspot per i gipeti. Ma non ovunque la situazione è altrettanto positiva, afferma Livio Rey, portavoce della Stazione ornitologica svizzera di Sempach.
La reintroduzione del gipeto sembra quindi una storia di successo, e destinata a continuare. Secondo uno studio della Fondazione Pro Gipeto e della Stazione ornitologica svizzera, nei prossimi dieci anni la popolazione di questi uccelli potrebbe raddoppiare. Una previsione molto incoraggiante, secondo Rey, che si basa su riproduzione, rilasci e sopravvivenza dei gipeti registrati attualmente. Una crescita del genere, spiega, è possibile solo perché la popolazione è ancora distante dalle sue possibili dimensioni. …
Non mancano però i pericoli e lo studio mostra che già il decesso di qualche animale in più all’anno, oltre a quelli che muoiono per cause naturali, potrebbe portare la popolazione di gipeti a una contrazione. I rischi, elenca Rey, sono le collisioni con cavi sospesi, ma anche avvelenamenti, caccia di frodo. Anche il turismo può essere un fattore negativo. Le zone di nidificazione possono essere disturbate da chi vuole fotografare gli uccelli, chi fa arrampicata o parapendio, così come dai droni. …
Bisogna riconoscere che nel nostro hobby il ruolo più scomodo è quello rivestito dai Giudici. Certo, vi sono mansioni e cariche nella struttura FOI per altri versi molto più impegnative, ma quello che il mondo degli allevatori percepisce e che tocca con mano è sicuramente il giudizio, e quindi i Giudici. Non vi è mostra piccola o grande in cui non ci siano polemiche, lamentele, a volte contestazioni, spesso avanzate anche da chi non raggiunge quei traguardi che riteneva essergli dovuti “per grazia divina”. Ma cosa succede, forse i nostri Giudici non sono tecnicamente preparati? Purtroppo le ragioni di vedute diverse, di incongruenze vere o presunte dipendono da molti fattori, a volte anche indipendenti dalla volontà di chi giudica. Sarà capitato, assistendo alle fasi di giudizio, di veder premiato meritatamente un canarino che in quel momento si presentava molto bene, ma che dopo due o tre giorni in mostra, per i motivi più vari, non appariva più lo stesso, così come pure il contrario. Quindi viene da chiedersi quale sia la reale caratura di un Padovano: quella del momento del giudizio del mercoledì/giovedì espresso dal giudice o quello della domenica sancito dagli allevatori? Questo è un esempio dei fattori che influiscono nella mancata corrispondenza dei valori tra i punteggi e i relativi soggetti, anche se in questo caso trascendono dalla responsabilità del Giudice, sebbene egli abbia ben operato. Chi alleva Padovani o Arricciati in genere sa che basta
poco per far sì che un canarino in un certo momento non mostri appieno le proprie potenzialità e se questo conta negli Arricciati di forma, analogamente vale per quelli di posizione.
Quello che, a nostro parere, mette il più delle volte in crisi alcuni Giudici ed allevatori è la mancanza inconsapevole, ripetiamo inconsapevole, di oggettività, sovrastata inconsciamente dalla soggettività, dall’idea personale su come debba essere un Padovano o un qualunque altro soggetto da giudicare.
L’oggettività non dà spazio ad alcuna interpretazione. Un Padovano deve avere le giuste caratteristiche, tutto deve corrispondere a canoni e misure prescritte dallo standard; il resto è soggettività, ovvero giudicare (e non ci riferiamo solo ai Giudici) secondo un modo personale di intendere o interpretare lo standard. Tuttavia, non crediamo affatto ci sia dietro malafede, bensì il nostro (anche senza rendercene conto) particolarissimo e personalissimo modo di percepire l’oggettività richiamata dai criteri di giudizio. Siamo certi che se lo stesso Padovano venisse valutato da un insieme di Giudici e allevatori otterrebbe altrettanti punteggi diversi e, qualora uguali, difficilmente corrisponderebbero nei valori dei considerando, in quanto, come già detto (ma ciò vale anche nella vita di tutti i giorni), ciascuno di noi possiede una percezione propria dell’oggettività e della sua applicazione. Tale può essere verosimilmente il meccanismo psicologico per cui si verificano le diversità di vedute anche nel nostro campo, perché le riteniamo insite nell’uomo e, tranne nei casi di manifesta incapacità, appaiono di difficile soluzione ma comunque meritevoli di studio e di un serio approfondimento. Un metodo che potrebbe aiutare sarebbe quello di inserire anche queste tematiche nella preparazione degli allievi Giudici e, in ogni caso, all’atto del giudizio avere la forza di distaccarsi quanto più possibile dalle proprie convinzioni, aprendo e consultando i criteri ed il relativo standard… cosa che vorremmo vedere più di frequente.