

Guía de campo de las ciperáceas de España y Portugal Resérvaloen
www.jolube.es
Prólogo
Resérvaloen www.jolube.es
A lo largo de mi carrera como taxónoma vegetal he tenido la satisfacción de estudiar la maravillosa familia de las ciperáceas. Fueron la fascinación por la biodiversidad y la evolución de la vida en la Tierra las que me impulsaron a estudiar Biología en la Universidad de Gante (Bélgica). Allí tuve ocasión de conocer al profesor Paul Goetghebeur, eminente especialista de la familia, quién ha transmitido su pasión por las plantas a varias generaciones de estudiantes, entre los que me cuento. El profesor Goetghebeur fue responsable de que me decantara por desarrollar mis estudios de postgrado en Botánica y me convenció para trabajar con un grupo de ciperáceas africanas para mi Trabajo Fin de Máster. Desde aquel día del año 2005 he dedicado mis esfuerzos científicos al estudio de la diversidad, la evolución y el estado de conservación de las ciperáceas, disciplinas que han constituido el núcleo de mi carrera científica. Aproximadamente por la misma época que el equipo de la Universidad de Gante centraba sus investigaciones en la sistemática de ciperáceas tropicales y subtropicales, el profesor Modesto Luceño y sus estudiantes de la Universidad Pablo de Olavide de Sevilla (España) concentraban sus esfuerzos en el megadiverso género Carex que, con algo más de 2000 especies, es el más rico en especies de todos los géneros de angiospermas que habitan en las regiones templadas del hemisferio norte. Los profesores Goetghebeur y Luceño no solo se dedicaron con entusiasmo a la formación de sus estudiantes, sino que fundaron grupos de investigación que pronto establecieron una red de colaboraciones científicas a nival mundial. No tardé mucho en conocer al equipo de la Universidad Pablo de Olavide. Recuerdo claramente el primer encuentro de nuestros grupos de investigación, que tuvo lugar en el Real Jardín Botánico de Kew hace unos 15 años. Desde entonces, nuestra intensa colaboración ha generado numerosos artículos científicos, entre los que destaco los publicados conjuntamente en el número especial dedicado a las ciperáceas por la prestigiosa revista Journal of Systematics and Evolution. También hemos creado juntos la joven Sociedad Internacional de Ciperólogos (International Sedge Society).


Las ciperáceas son un grupo rico en especies que tienen un aspecto similar a las gramíneas y que habitan en lugares donde muchas otras plantas tendrían serias dificultades para sobrevivir, como es el caso de humedales, rocas o suelos pedregosos pobres en nutrientes y zonas muy elevadas de las montañas. Gracias a herramientas genéticas, como la secuenciación masiva del ADN, hemos empezado a entender el origen y evolución de este grupo de plantas tan espectacularmente diverso. Sin embargo, para comprender realmente la diversidad morfológica y ecológica de sus especies y evaluar con precisión su estado de conservación, debemos ser capaces de identificar las especies de ciperáceas sobre el terreno, que es el objetivo principal de esta guía de campo que tengo la satisfacción de prologar.

5
Es importante destacar que muchos botánicos, ecólogos y aficionados a las plantas evitan con frecuencia prestar atención a los grupos de plantas que presentan dificultades para identificar sus especies. Las ciperáceas y las gramíneas, con sus poco llamativas flores dispuestas en inflorescencias complejas, suelen pasarse por alto, a pesar de figurar entre las 10 familias de plantas más ricas en especies de toda la flora mundial. Por ello, disponer de una guía de campo fácil de usar y bien ilustrada que facilite la identificación de las especies vegetales es una herramienta muy valiosa para obtener los datos necesarios que permitan evaluar el estado de conservación y llevar a cabo las políticas adecuadas para su protección.


El profesor Modesto Luceño ha reunido un equipo de referencia a nivel internacional en botánica que llevan muchos años estudiando a fondo las ciperáceas; entre ellos los profesores Marcial Escudero y Santiago Martín-Bravo, así como los doctores Pedro Jiménez-Mejías, Enrique Maguilla, José Ignacio Márquez-Corro y Tamara Villaverde. La presente guía de campo está magníficamente ilustrada con fotografías y láminas de dibujos, y presenta una clasificación actualizada que incluye claves dicotómicas fáciles de usar y numerosos datos que facilitan enormemente la identificación de los géneros y especies de ciperáceas que crecen en España y Portugal, incluyendo los biodiversos archipiélagos de Baleares, Canarias, Azores y Madeira. Se proporcionan además descripciones diagnósticas de los diferentes táxones, así como mapas de distribución y datos sobre la época de floración, las preferencias edáficas y el estado de conservación en los casos que han sido evaluados. Por todas estas razones, esta guía de campo permitirá a cualquier persona identificar las especies de forma rápida y sencilla.
No me cabe duda de que esta guía de campo abrirá el mundo de las ciperáceas a muchos científicos y botánicos aficionados, y empujará a las nuevas generaciones a interesarse por este maravilloso grupo de plantas. Además, gracias a aplicaciones para teléfonos móviles como iNaturalist o el Global Biodiversity Information Facility, las observaciones de los táxones de ciperáceas de España y Portugal, correctamente identificadas con la ayuda de esta guía de campo, supondrán una contribución clave para su supervivencia en este mundo cambiante.
Resérvaloen www.jolube.es
31 de enero de 2023
6
Dra. Isabel Larridon. Investigadora principal en el Real Jardín Botánico de Kew (Reino Unido) (Senior Research Leader Integrated Monography at Royal Botanic Gardens, Kew) Profesora visitante de la Universidad de Gante, Bélgica.
Rhynchospora modesti-lucennoi
IntroducciónResérvaloen www.jolube.es
La familia Cyperaceae, con cerca de 5700 especies, es la octava con mayor biodiversidad dentro de las angiospermas y la tercera de las monocotiledóneas, tras las orquidáceas (=orquídeas; ca. 26000 especies), las asteráceas (=compuestas; ca. 25000), las rubiáceas (ca. 13500), las poáceas (=gramíneas; ca. 11400), las lamiáceas (=labiadas; ca. 7300), las euforbiáceas (ca. 6750) y las mirtáceas (ca. 5900). Está compuesta por 96 géneros, cuya distribución comprende la totalidad del planeta, a excepción del continente Antártico. La mayor diversidad de géneros se encuentra en las regiones tropicales, si bien Carex, el género que contiene un mayor número de especies (más de 2000), habita principalmente en las regiones templadas y frías del hemisferio norte. Le sigue Cyperus, con más de 950 especies, principalmente tropicales.

Los registros fósiles disponibles y relojes moleculares (que ayudan a estimar tiempos de divergencia mediante estudios del ADN) muestran que la familia se remonta al Paleoceno (Cenozoico temprano), hace más de 60 millones de años, y que los grandes géneros, como Carex y Cyperus, ya se habían establecido en el Eoceno tardío, unos 35-40 millones de años atrás. Por otra parte, estudios de reconstrucción de áreas ancestrales basados en filogenia molecular indican que las ciperáceas se originaron, probablemente, en Sudamérica.
Aunque la mayoría de las especies viven sobre suelos húmedos, e incluso existen algunas que realizan su ciclo vital casi completamente sumergidas, como la tropical Eleocharis confervoides, las ciperáceas medran en una amplísima gama de hábitats. Un claro ejemplo lo encontramos en Cyperus conglomeratus, capaz de desarrollarse sobre las tórridas dunas del desierto del Sahara. Suelen ser comunes en bosques y prados, pero muchas especies toleran también los suelos raquíticos de pedregales y fisuras rocosas. Desde el punto de vista de sus preferencias edáficas, se conocen especies que prefieren valores de pH muy elevados, como las que viven sobre suelos salinos, y otras que se desarrollan bien sobre turberas extremadamente ácidas, pero no son raras las que toleran rangos muy amplios de pH. Su espectro altitudinal comprende desde el nivel del mar hasta los 5700 m que alcanza Carex moorcroftii en las laderas del Himalaya.
Caracterización morfológica
La mayoría de las especies que forman parte de las ciperáceas son hierbas con aspecto graminoide, es decir, superficialmente similar al de las gramíneas (familia Poaceae) y juncos (familia Juncaceae). De hecho, las tres familias están emparentadas y clasificadas en el mismo orden (Poales) dentro de las monocotiledóneas, y a veces puede resultar confuso identificarlas visualmente entre aficionados, principiantes o no tanto. No obstante, ciperáceas, gramíneas y juncáceas son distinguibles de manera relativamente sencilla por una serie de caracteres fácilmente observables si se presta un poco de atención.


11
CyperaceaePoaceae Juncaceae
TallosDe sección frecuentemente trígona, macizos (rellenos de médula)
De sección más o menos cilíndrica, huecos De sección cilíndrica, macizos (rellenos de médula)
VainaDe márgenes cerradosDe márgenes abiertosDe márgenes abiertos o cerrados
Glumas por flor Una, que suele ocultar los órganos sexuales Dos, al menos una de las cuales oculta los órganos sexuales
Ausentes; a veces la flor está protegida por 1 o 2 bractéolas que no ocultan los órganos sexuales ni el perianto
FloresUnisexuales o hermafroditas Unisexuales o hermafroditas Hermafroditas
PeriantoAusente o reducido a cerdas o escamas que no suelen ocultar los órganos sexuales
Ausente o formado por tres escamas (lodículas) que no ocultan los órganos sexuales
Anteras Basifijas Dorsifijas Basifijas
FrutoIndehiscente (aquenio)Indehiscente (cariópside)Dehiscente (cápsula)

Resérvaloen www.jolube.es
Presente, sepaloideo, actinomorfo, formado por seis piezas que ocultan los órganos sexuales cuando la flor está cerrada
A efectos prácticos, en la presente guía hemos preferido simplificar la terminología y usar las siguientes expresiones para indicar el hábito de las plantas: 1) plantas anuales, 2) plantas perennes no cespitosas, 3) plantas perennes laxamente cespitosas y 4) plantas perennes densamente cespitosas.
Aunque de su apariencia graminoide se pudiera deducir que el aspecto de las ciperáceas es uniforme, la familia contiene especies con hábitos de crecimiento muy diversos: desde plantas anuales de ciclo efímero, como Bulbostylis glaberrima de Kenia y Uganda o Isolepis inconspicua de la región de El Cabo (Sudáfrica), que apenas alcanzan 2 y 3 cm de altura máxima, respectivamente, hasta formas arbustivas de larga vida -caso de Microdracoides squamosa (originaria de Camerún, Guinea, Nigeria y Sierra Leona), Afrotrilepis pilosa (África tropical) y del género Chamaedendron (endémico de Nueva Caledonia)- o lianas que pueden alcanzar 12 m de longitud (Scleria boivinii, nativa de África tropical y Madagascar). No obstante, la mayoría de las especies son hierbas anuales o perennes a través de rizomas subterráneos que, fundamentalmente, responden a dos tipos:
a) Rizoma simpódico, cuya yema terminal tiene crecimiento limitado; dicha yema genera un brote aéreo, de cuya base surgen rizomas horizontales laterales que producen, a su vez, nuevos brotes aéreos (fértiles o estériles) verticales y rizomas horizontales. Ello da como resultado plantas más o menos cespitosas (en ocasiones formando macollas) en función de la longitud de los entrenudos.
b) Rizoma monopódico, aquel cuya yema terminal tiene crecimiento horizontal indefinido y presenta yemas laterales muy separadas, de las que surgen tallos aéreos fértiles (con flores) y estériles (sin flores) a intervalos más o menos regulares. El resultado es que los tallos están muy distanciados unos de otros, por lo que no forman césped.
Los tallos fértiles son generalmente macizos y tienen los nudos frecuentemente concentrados en la parte inferior. Son habitualmente trígonos, aunque existen numerosos ejemplos de tallos cilíndricos y, en especies extraibéricas, poligonales o más o menos comprimidos e incluso planos. Los ángulos de los tallos trígonos pueden ser de redondeados u obtusos hasta muy agudos, e incluso prolongarse en forma de ala. La ausencia o presencia más o menos densa de expansiones epidérmicas (papilas y/o acúleos) resulta a menudo de utilidad para distinguir especies.
Las hojas suelen ser sésiles, raramente -en especies no ibéricas- estrechadas en la base formando un falso pecíolo (pseudopecíolo) en algunas especies de hojas anchas de los géneros Carex, Cyperus y Mapania. Su aspecto es similar al de las gramíneas, aunque en general de consistencia más rígida que las de estas. Constan de una vaina cerrada que rodea al tallo y de un limbo (=lámina). En el punto de separación entre el limbo y la vaina, en la base de la cara superior del limbo, se encuentra adherida la lígula, una membrana translúcida cuya longitud, anchura, bordes y

12 13
Tipos de ramificación del rizoma en ciperáceas: A, simpódico; B, monopódico.
Lígula y antelígula en Carex laevigata.
Principales diferencias morfológicas entre ciperáceas, gramíneas y juncáceas.
© M. S. Villegas
ápice pueden ser de interés para caracterizar especies. La cara de la vaina opuesta a la inserción del limbo es hialina en su tramo superior y, en ocasiones, su ápice se prolonga formando la denominada antelígula. El limbo foliar es entero y habitualmente linear, si bien existen especies con hojas filiformes o -especialmente en los trópicos- lanceoladas, ovadas, elípticas, obovadas o ensiformes (=isobilaterales; en forma de espada en las que no se distingue entre el haz y el envés). En ocasiones las hojas están reducidas a vainas o tienen el limbo muy corto, como es el caso de los géneros Eleocharis, Trichophorum, Schoenoplectus, Schoenoplectiella, etc. Según el corte transversal, las láminas foliares pueden ser, a grandes rasgos, planas, aquilladas (=carenadas), canaliculadas, revolutas, involutas, plegadas, conduplicadas o enrolladas. Los estomas se encuentran con más frecuencia en el envés (hojas hipostomáticas), pero también numerosas especies los muestran en ambas superficies (hojas anfistomáticas) y, más raramente, solo en la cara superior (hojas epistomáticas). Las hojas pueden ser glabras o más o menos pelosas (pelos siempre unicelulares), lisas o con acúleos y/o papilas en la superficie, en los bordes y/o en el nervio medio.
Resérvaloen www.jolube.es
La inflorescencia puede ser simple o variablemente ramificada; su unidad básica es la espiguilla o espícula, constituida por una a numerosas flores, cada una en la axila de una escama (gluma) que nace del eje de la espiguilla (raquis). El modo de agrupación de las espiguillas determina los tipos principales de inflorescencias, de manera que podemos distinguir espiguillas solitarias, como en Eleocharis y Trichophorum, o agrupadas formando espigas o panículas. Las espigas pueden ser solitarias o múltiples, en cuyo caso se disponen, a su vez, en espigas (inflorescencia espiciforme) o racimos (inflorescencia racemiforme). Por otra parte, las panículas (o inflorescencias paniculiformes) pueden ser simples (un solo orden de ramificación) o compuestas (varios órdenes de ramificación) y más o menos densas. Cuando forman grupos aproximadamente esféricos, estos reciben el nombre de glomérulos, mientras que si constituyen estructuras umbeliformes se denominan antelas. Cuando la inflorescencia paniculiforme forma un solo grupo más o menos apretado a modo de capítulo o cabezuela, se conoce con el nombre de inflorescencia capituliforme. Los tipos descritos pueden aparecer mezclados formando inflorescencias mixtas.
Tipos de lámina foliar según su corte transversal: A. plana; B. aquillada o carenada, C. canaliculada; D. revoluta; E. involuta; F. plegada; G. conduplicada; H. enrollada.


Diferentes tipos de inflorescencias en ciperáceas (ver texto): A. Cladium mariscus (paniculiforme con glomérulos laxos; Guadalajara, España); B. Scirpoides holoschoenus subsp. holoschoenus (paniculiforme con glomérulos densos; Valladolid, España); C. Cyperus eragrostis (antela; Cáceres, España); D. Cyperus capitatus (capituliforme; Alto Alentejo, Portugal); E. Eleocharis quinqueflora (espiguilla solitaria; Gerona, España); F. Carex rupestris (espiga solitaria; Gran Sasso, Italia); G. Carex canescens (espiciforme; Tierra de Fuego, Argentina); H. Carex limosa (racemiforme; sierra de Gredos, España).
La inflorescencia está acompañada por una a numerosas brácteas, que pueden ser foliáceas (con apariencia de hoja), setáceas o glumáceas (escuamiformes), la más inferior de las cuales puede proporcionar caracteres de importancia taxonómica. En ocasiones, cuando la bráctea inferior es foliácea y se dispone verticalmente, asemejando una prolongación del tallo, se habla de inflorescencia pseudolateral; cuando no es así, se denomina inflorescencia terminal
14 15
© M. S. Villegas
SubfamiliaTribu Género
Mapanioideae (186)
Hypolytreae (172)
Chysitricheae (14)
Hypolytrum (63), Mapania (100), Paramapania (7), Principina (1), Scirpodendron (1)
Diplasia (1), Capitularina (1), Chorizandra (6), Chrysitrix (4), Exocarya (1), Lepironia (1)
Trilepideae (16) Afrotrilepis (2), Coleochloa (8), Microdracoides (1), Trilepis (5)
Cladieae (3) Cladium (3)
Sclerieae (258) Scleria (258)
Bisboeckelereae (28)
Becquerelia (6), Bisboeckelera (4), Calyptrocarya (8), Diplacrum (10)
Carpheae (18) Carpha (15), Trianoptiles (3)
Cryptangieae (50)
Cephalocarpus (20), Cryptangium (1), Didymiandrum (1), Exochogyne (2), Koyamaea (1), Krenakia (10), Lagenocarpus (15)
Ciperáceas ibéricas y macaronésicas
Las ciperáceas están representadas en la península ibérica y Macaronesia por una subfamilia, 14 tribus y 18 géneros, que incluyen 210 táxones (197 especies y 13 subespecies), de los cuales 174 (163 especies y 11 subespecies) son nativos y 36 (34 especies y 2 subespecies) probablemente introducidos por el ser humano. Por territorios, en la península ibérica habitan 188 (160 nativas y 28 introducidas) especies y subespecies, mientras que en Macaronesia viven tan solo 66 (53 nativas y 13 introducidas).
Cyperoideae (5508)
Schoeneae (466)
Ammothryon (1), Anthelepis (4), Capeobolus (1), Caustis (7), Chaetospora (3), Chamaedendron (5), Costularia (15), Cyathochaeta (5), Cyathocoma (3), Evandra (2), Gahnia (41), Gymnoschoenus (2), Lepidosperma (80), Machaerina (55), Mesomelaena (5), Morelotia (6), Neesenbeckia (1), Netrostylis (11), Oreobolus (17), Ptilothryx (1), Reedia (1), Schoenus (149), Tetraria (39), Tricostularia (11), Xyroschoenus (1)
Rhynchosporeae (399) Rhynchospora (399)
Dulichieae (5) Blysmopsis (1), Blysmus (3), Dulichium (1)
Khaosokiaeae (1) Khaosokia (1)
Calliscirpeae (2) Calliscirpus (2)
Scirpeae (73)
Amphiscirpus (1), Eriophorum (18), Phylloscirpus (3), Rhodoscirpus (1), Scirpus (47), Zameioscirpus (3)
Trichophoreae (19) Trichophorum (19)
Sumatroscirpeae (4) Sumatroscirpus (4)
Cariceae (2010) Carex (2010)
Eleocharideae (302) Eleocharis (302)
Abildgaardieae (571)
Abildgaardia (9), Actinoschoenus (2), Arthrostylis (2), Bulbostylis (227), Fimbristylis (320), Nelmesia (1), Scleroschoenus (6), Trachystylis (1), Trichoschoenus (1), Zulustylis (2)
Bolboschoeneae (15) Bolboschoenus (15)
Fuireneae (55) Fuirena (55)
Schoenoplectieae (17) Actinoscirpus (1), Schoenoplectus (16)
Peudoschoeneae (64) Pseudoschoenus (1), Schoenoplectiella (63)
Resérvaloen www.jolube.es
El género más numeroso es Carex, que acumula más de la mitad de todas las ciperáceas, con 112 especies y 9 subespecies. De la península ibérica se aceptan 102 especies y 6 subespecies, todas nativas; mientras que de Macaronesia se conocen únicamente 29 especies y 2 subespecies. Le sigue el género Cyperus, con 38 especies y 1 subespecie, de las que únicamente 12 son nativas. En la península ibérica viven 32 especies de Cyperus, 21 de las cuales son introducidas, mientras que Macaronesia cuenta con 20 táxones (19 especies y 1 subespecie), de los cuales solo 9 son nativos. El resto de los géneros contienen menos de 6 especies y subespecies, a excepción de Eleocharis, con 12, 9 nativas y 3 naturalizadas. Blysmus, Fuirena, Schoenus, Scirpus y Trichophorum están representados por un solo taxon.
La riqueza de especies y subespecies por provincias es muy heterogénea, siendo notablemente más elevada en las provincias pirenaico-cantábricas, en muchas de las cuales se superan los 80 táxones, y disminuye en zonas del tercio sur y el oeste, donde algunas provincias no alcanzan los 35. La riqueza de los archipiélagos balear y macaronésicos es también baja, siendo las Islas Azores las que exhiben el valor más alto (37).

Cypereae (1131)
Afroscirpoides (1), Cyperus (964), Dracoscirpoides (3), Erioscirpus (2), Ficinia (87), Hellmuthia (1), Isolepis (69), Scirpoides (4)
Incertae sedis (posición incierta) (1) Rhynchocladium (1)
Sinopsis de la clasificación de las ciperáceas siguiendo la propuesta de Larridon et al. (2021). Las cifras entre paréntesis indican el número de especies aceptadas. Los táxones cuyo nombre aparece en negrita son nativos de la península ibérica y/o Macaronesia. La ordenación de las tribus sigue el esquema filogenético obtenido en los trabajos sobre filogenia de la familia, mientras que los géneros de cada tribu aparecen ordenados alfabéticamente.
de ciperáceas en la península ibérica y Baleares.
22 23
Mapa de riqueza de táxones (especies + subespecies)
De las 174 ciperáceas nativas, 23 son endémicas del territorio, todas pertenecientes a Carex. Esto supone una tasa de endemicidad de algo más del 13% de la familia y de poco más del 20% del género Carex. Además, algunos taxones no endémicos tienen su área principal de distribución en el territorio, entre los que destacamos Carex macrostyla, endemismo pirenaico-cantábrico que vive también en el lado francés de los Pirineos, y Carex helodes, distribuida por el suroeste de la península ibérica y extremo norte de Marruecos, de donde se conoce hasta el momento una sola población.
Península
ibérica y Baleares CanariasAzoresMadeira
Carex asturica
Carex camposii subsp. camposii
Carex camposii subsp. tejedensis
Carex canariensis
Carex caudata
Carex demissa subsp. cedercreutzii
Carex durieui
Carex furva
Carex hochstetteriana
Carex laevicaulis
Carex lainzii +---
Carex leviosa --+-
Carex lowei ---+
Carex lucennoiberica
Carex nevadensis
Carex paniculata subsp. calderae
Carex perraudieriana
Carex pilulifera subsp. azorica
Península
ibérica y Baleares CanariasAzoresMadeira
Carex quixotiana +---
Carex reuteriana subsp. reuteriana +---
Carex rorulenta +---
Carex sequeirae ---+
Carex vulcani --+-
Objetivos y estructura de la guía
El presente libro pretende ser un instrumento para el conocimiento y la identificación de las ciperáceas de la península ibérica (incluyendo en ella a las Islas Baleares y Andorra) y Macaronesia (archipiélagos de Azores, Canarias y Madeira). La familia tiene una inmerecida reputación de “difícil”, debido a la supuesta dificultad para identificar sus especies, lo que puede explicarse, a nuestro entender, porque sus flores y frutos son, en general, poco vistosos y su observación requiere frecuentemente del empleo de una lupa. Sin embargo, a diferencia de otros grupos con estructuras reproductivas más aparentes, las ciperáceas muestran numerosos caracteres que ayudan a separar con cierta sencillez unos táxones de otros.
Nuestra intención es que la guía pueda ser útil, tanto para profesionales de la botánica no especializados en la familia como para aquellos naturalistas aficionados al apasionante mundo de las plantas, con independencia del nivel de conocimientos botánicos que posean. Por ello hemos primado las ilustraciones, sean fotografías o dibujos, que ayuden a la identificación visual y a la comprensión de los caracteres diagnósticos. También con ese objetivo hemos evitado, en la medida de lo posible, descripciones largas y el uso de términos y disquisiciones académicas difíciles de asimilar por los no avezados en la materia, y hemos recurrido a símbolos intuitivos para reflejar aspectos como el rango altitudinal, las preferencias edáficas y el periodo de floración.
Hemos considerado todas las especies y/o subespecies de cuya introducción por parte del ser humano hay evidencias, tanto las que forman poblaciones silvestres estables (invasoras o no), como aquellas cuya presencia y espontaneidad está o ha estado confirmada en alguna ocasión. Hemos procedido de este modo porque no disponemos de información sobre la mayor o menor estabilidad poblacional de muchas de ellas.
El contenido de la obra comienza por las claves de identificación, la primera correspondiente a los géneros y las siguientes a las especies de cada género. A continuación, se presentan fichas descriptivas de las especies. Cuando se aceptan varias categorías infraespecíficas (únicamente admitimos aquí el rango de subespecie) hemos añadido una clave para distinguirlas y, en los casos necesarios, ilustraciones diagnósticas. La presentación de los géneros sigue el orden filogenético de las tribus. Dentro de cada género, las especies se exponen por subgéneros, y dentro de cada uno de estos por secciones. No obstante, cuando las filogenias recientes no han resuelto la ubicación de ciertos taxones en una determinada sección, estos se incluyen en el clado (linaje o rama del árbol filogenético) donde anidan, cuyo nombre alude al epíteto específico de una de las especies incluidas en dicho clado. Dada la rareza o el carácter esporádico de muchas de las especies introducidas a las que hemos hecho alusión, y con objeto de evitar en la medida de lo posible un volumen excesivo de esta obra, únicamente hemos confeccionado fichas completas de los taxones no nativos especialmente abundantes y de amplia distribución, debido a su carácter potencialmente invasor.

24 25
+---
+---
+---
-+-+
+---
--+-
+---
+---
--+-
--++
+---
+---
-+--
-+--
--+-
Mapa de riqueza de táxones (especies + subespecies) de ciperáceas en Azores, Canarias y Madeira.
Ciperáceas endémicas de la península ibérica y/o Macaronesia.
Resérvaloen www.jolube.es
Cada ficha está encabezada por el nombre latino de cada especie o subespecie, puesto que la mayoría de las especies del territorio de la guía no tienen nombres vulgares oficiales en español. Dichos nombres aparecen en cursiva y negrita cuando se trata de táxones nativos y solo en cursiva cuando son alóctonos. Seguidamente se presenta una descripción esencialmente diagnóstica, intentando resaltar únicamente los caracteres de utilidad para determinar el taxon. Al final de cada diagnosis se indica el o los números de cromosomas conocidos, de los cuales, los recuentos cromosomáticos llevados a cabo sobre poblaciones ibéricas se indican en negrita. Cuando aparece el símbolo de interrogación (?) tras el número, queremos indicar que el recuento en cuestión nos ofrece dudas. Finalmente, en cursiva, se describe el hábitat o hábitats donde vive el taxón en concreto. A continuación, se muestra el mapa de distribución por provincias y archipiélagos (véase más adelante), así como los símbolos correspondientes a la época de floración, las preferencias edáficas, el rango altitudinal y, en su caso, la categoría de conservación a nivel regional (España y Portugal), según las categorías y criterios de la Unión Internacional para la Conservación de la Naturaleza (UICN). Las ilustraciones de cada taxon nativo y de la mayoría de los introducidos incluyen una lámina fotográfica con una imagen principal y, frecuentemente, uno o dos recuadros que contemplan detalles (generalmente el utrículo y la gluma femenina en el caso de Carex, y los aquenios en el del resto de los géneros). Asimismo, se muestra también una lámina de dibujo en la que se representan el porte general y los principales órganos diagnósticos (glumas, frutos, utrículos, etc.).
El mapa de distribución se ha elaborado coloreando en verde la provincia o archipiélago donde el taxon está presente. En los pocos casos donde la presencia de un taxon en una determinada provincia
Indicación de la época de floración.
Resérvaloen

Símbolos empleados para indicar el grado de amenaza regional siguiendo las categorías y criterios de la UICN.
o archipiélago es dudosa, hemos coloreado dicho territorio en azul celeste. Por último, se inluyen un glorario terminológico (Apéndice I), que recoge los vocablos técnicos citados en el texto, y los créditos fotográficos (Apéndice II) de aquellas imágenes cuya publicación nos ha sido gentilmente autorizada..

Bibliografía
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 14, July 2017 [and more or less continuously updated since].” http://www.mobot.org/MOBOT/research/APweb/
Arechavaleta, M.; Rodríguez, S.; Zurita, N. & García, A. (coord.) (2010). Lista de especies silvestres de Canarias. Hongos, plantas y animales terrestres. 2009. Gobierno de Canarias. 579 pp.
Larridon, I.; Zuntini, A.R.; Léveillé-Bourret, É; Barret, R.L.; Starr, J.R.; Muasya, A.M.; Villaverde, T.; Bauters, K.; Brewer, G.E.; Bruhl, J.J.; Costa, S.M.; Elliott, T.M.; Epitawalage, N.; Escudero, M.; Frailie, I.; Goetghebeur, P.; Hipp, A.L.; Jiménez-Mejías, P.; Kikuchi, I.A.B.S.; Luceño, M.; Márquez-Corro, J.I.; Martín-Bravo, S.; Maurini, O.; Pokorny, L.; Roalson, E.H.; Semmouri, I.; Simpson, D.A.; Spalink, D.; Thomas, W.W.; Wilson, K.L.; Xanthos, M.; Forest, F. & Baker, W.J. (2021). A new classification of Cyperaceae (Poales) supported by phylogenomic data. J. Syst. Evol. 59(4): 852-895.
Global Carex Group: Waterway, M.J.; Roalson, E.H.; Ford, K.A.; Luceño, M.; Martín-Bravo, S.; Starr, J.R.; Wilson, K.L.; Yano, O.; Zhang, S.R.; Alverson, W.S.; Bruederle, L.P., Bruhl, J.J.; Chung, K.-S.; Cochrane, T.S.; Escudero, M.; Fored, B.A.; Gebauer, S. Gehrke, B.; Hann, M.; Hipp, A.L.; Hoffmann, M.H.; Hoshino, T.; Jiménez-Mejías, P.; Jin, X.-F.; Jung, J.; Kim, S.; Maguilla, E.; Masaki, T.; Míguez, M.; Molina, A.; Naczi, R.F.C.; Reznicek, A.A.; Rothrock, P.E.; Simpson, D.A.; Spalink, D.; Thomas, W.W. & Villaverde, T. (2015). Making Carex monophyletic (Cyperaceae, tribe Cariceae): a new broder circumscription. Bot. J. Linn. Soc. 179: 1-42.
Goetghebeur, P. (1998). Cyperaceae. In: Kubitzki, K. (ed.): The families and genera of vascular plants. Berlin: Springer-Verlag. 4: 141–190.
Símbolos indicadores de las preferencias edáficas de las ciperáceas ibérico-macaronésicas. A: taxon indiferente edáfico; B: taxon acidófilo; C: taxon basófilo.
Luceño, M.; Castroviejo, S. & Jiménez-Mejías, P. (2008). Cyperaceae, in Castroviejo, S.; Luceño, M.; Galán, A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora ibérica 18: 4-250.
Márquez-Corro, J.I.; Martín-Bravo, S.; Pedrosa-Harand, A.; Hipp, A. L; Luceño, M. & Escudero, M. (2019). Karyotype Evolution in Holocentric Organisms. In: eLS. John Wiley & Sons, Ltd: Chichester.

Márquez-Corro, J.I.; Martín-Bravo, S.; Spalink, D.; Luceño, M. & Escudero, M. (2019). Inferring hypothesis-based transitions in clade-specific models of chromosome evolution in sedges (Cyperaceae). Mol. Phyl. Evol. 135: 202-209.
Menezes de Sequeira, M.; Espirito Santo, D.; Aguiar, C.; Capelo, J. & Honrado, J. (2011). Checklist da Flora de Portugal (Continental, Açores e Madeira). Associação Lusitana de Fitossociologia Lisboa. 41 pp.
Schäfer, H. (2002). Chorologie und Diversität der Flora der Azoren. Ph. D. These. Universität Regensburg. 130 pp.
Simpson, D.A. & Inglis, C.A. (2001). Cyperaceae of economic, ethnobotanical and horticultural importance: a checklist. Kew Bull. 56: 257-366.
26 27
Ejemplo de símbolo indicador del rango altitudinal. El sombreado en verde indica el rango.
www.jolube.es
Clave de géneros
1. Todas las flores unisexuales; frutillos (aquenios) envueltos por una escama de bordes soldados (utrículo) o, más raramente, abiertos (periginio glumiforme) Carex

- Al menos alguna flor hermafrodita; aquenios desnudos 2
2. Inflorescencia de aspecto algodonoso en la fructificación Eriophorum


- Inflorescencia de aspecto no algodonoso ................................................................................................ 3
3. Espiguillas sésiles, dispuestas en dos filas a ambos lados del eje de la inflorescencia ................................................................................................................................ Blysmus
- Espiguillas sésiles o pedunculadas, no dispuestas en dos filas 4
4. Inflorescencia formada por una espiguilla terminal solitaria sin brácteas foliáceas ...... 5
- Inflorescencia compuesta por varias espiguillas, raramente formada por una sola, pero entonces aparentemente lateral, protegida por una bráctea foliácea que parece la prolongación del tallo .......................................................................................................................................... 6
5. Hojas sin limbo, reducidas a vainas . Eleocharis
- Hojas con limbo de (1,5)3-6(8) mm ................................................................................ Trichophorum
6. Espiguillas agrupadas formando varias cabezuelas esféricas muy densas; tallos cilíndricos, brillantes; inflorescencia con una bráctea cilíndrica que parece una prolongación del tallo; hojas reducidas a vainas; glumas denticuladas, dispuestas en espiral, provistas de pequeñas espinas Scirpoides
-
.......................................................................................
7.
.........................................................................................................................
8. Inflorescencia formada por una cabezuela densa de color negro, con las dos brácteas inferiores que abrazan al conjunto de las espiguillas; vainas basales brillantes; plantas perennes Schoenus
- Inflorescencia frecuentemente umbeliforme, si capitada, entonces de color diferente o sin dos brácteas inferiores que abrazan el conjunto de las espiguillas o vainas basales mates o plantas anuales .................................................................................................................... Cyperus 9.
29
Sin la combinación precedente de
7
caracteres
Glumas dispuestas en dos filas 8
Glumas dispuestas en espiral
9
-
Base del estilo muy ensanchada, persistente
en la madurez
10
Base del estilo no ensanchada 14
o caediza
.............................
-
Resérvaloen www.jolube.es
SCHOENUS L.
La última clasificación de la familia Cyperaceae, basada en estudios filogenómicos, incluye Schoenus en la tribu Schoeneae, como único género integrante de la subtribu Schoeninae. Se aceptan actualmente cerca de 150 especies, distribuidas principalmente por Australasia, aunque unas pocas habitan en Europa, Japón, oeste y sureste de Asia, así como el sur de Norteamérica y Sudamérica, y América Central. En el territorio está representado por una sola especie, Schoenus nigricans. El género se caracteriza por ser plantas en general densamente cespitosas, con tallos casi siempre cilíndricos, inflorescencia por lo común terminal, glumas caedizas de disposición dística, solo las superiores fértiles, perianto casi siempre presente y estilo con 3 estigmas.



Bibliografía
Castroviejo, S. (2008). Schoenus L. in Castroviejo, S.; Luceño, M.; Galán, A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 105-106.

Goetghebeur, P. (1998). Cyperaceae. In: Kubitzki, K., ed. The families and genera of vascular plants. Berlin: Springer-Verlag. 4: 141–190.
Jermy, C.; Simpson, D.; Foley, M. & Porter, M. (2007). Sedges of the British Isles. B.S.B.I. Handbook No. 1, ed. 3: 1-554. Botanical Society of the British Isles, London.

Musili, P.M.; Gibbs, A.K.; Wilson, K.L. & Bruhl, J.J. (2016). Schoenus (Cyperaceae) is not monophyletic based on ITS nrDNA sequence data. Austral. Syst. Bot. 29: 265-283.
Schoenus nigricans L.
Densamente cespitosa, formadora de macollas. Tallos fértiles de 15-65(85) cm, cilíndricos. Hojas de hasta 1,2(1,4) mm de anchura, canaliculadas o conduplicadas, rígidas, comúnmente más cortas que los tallos; vainas basales sin limbo desarrollado, negruzcas, brillantes. Bráctea inferior glumácea con lámina setácea o foliácea, por lo común mucho más larga que la inflorescencia. Inflorescencia densa, capituliforme, formada por (2)4-8(9) espiguillas de hasta 15 mm, lanceoladas o estrechamente elípticas, comprimidas. Glumas lanceoladas o estrechamente elípticas, fuertemente correosas, de color pardo muy oscuro o negruzcas, brillantes, con la quilla escábrida, frecuentemente con margen escarioso estrecho, las 2-3 inferiores estériles, agudas o acuminadas, las restantes hermafroditas y obtusas. Cerdas periánticas en número de 3-6, mucho menores que el aquenio, con acúleos retrorsos. 3 estambres. 3 estigmas. Aquenios de 1,4-2,1 mm de longitud, obtusamente trígonos, más o menos blanquecinos, brillantes. 2n= 44, 54. Prados húmedos y otros medios encharcados.

Resérvaloen www.jolube.es
38 39
TRIBU SCHOENEAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU SCHOENEAE
Schoenus nigricans
RHYNCHOSPORA Vahl
Único género encuadrado en la tribu Rhynchosporeae, de la subfamilia Cyperoideae, que cuenta con aproximadamente 400 especies distribuidas por todos los continentes a excepción de la Antártida, si bien su mayor diversidad se encuentra en el continente americano, especialmente en Brasil. En el territorio habitan exclusivamente tres especies que se distribuyen por la península ibérica. Se trata de plantas con tallos obtusamente trígonos o cilíndricos, inflorescencia muy variable -por lo común con espiguillas muy numerosas-, cuyas glumas se disponen en espiral, las inferiores estériles y las superiores hermafroditas, con (0)1-13 cerdas periánticas, estilo con 2 estigmas y aquenios biconvexos, con base del estilo persistente, triangular o deltoidea. Numerosas especies americanas y la europea R. alba son entomófilas.
Resérvaloen www.jolube.es
© M. S. Villegas
Schoenus nigricans: A) hábito; B) aquenio; C) gluma estéril; D) aquenios y raquidio; E) inflorescencia.
Bibliografía
Goetghebeur P. (1998). Cyperaceae. In: Kubitzki, K., ed. The families and genera of vascular plants. Berlin: Springer-Verlag. 4: 141–190.
Jermy, C.; Simpson, D.; Foley, M. & Porter, M. (2007). Sedges of the British Isles. B.S.B.I. Handbook No. 1, ed. 3: 1-554. Botanical Society of the British Isles, London.
Luceño, M. & Martín, J. (2008). Rhynchospora in Castroviejo, S.; Luceño; M., Galán; A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 99-102.

Thomas, W.W.; Araujo, A.C. & Alves, M.V. (2008). A preliminary molecular phylogeny of the Rhynchosporeae (Cyperaceae). The Botanical Review 75: 22-29.
Villa-Machío, I.; Zamora, J.C.; Sandoval-Sierra, J.V.; Blanco-Pastor, J.L.; Fernández-Mazuecos, M. & Jiménez-Mejías, P. (2020). Insect pollination in temperate sedges? A case study in Rhynchospora alba (Cyperaceae). Plant Biosystems. https://doi.org/10.1080/112635504.2020.1845846
CLAVE DE ESPECIES
1. Espiguillas blancas o blanquecinas en la floración ................................................................. R. alba
- Espiguillas pardas o negruzcas en la floración 2
2. Cerdas periánticas (0)1-3(5), más cortas que el aquenio; aquenios con finas arrugas transversales; tallos fértiles de (35)40-70(90) mm ................................ R. modesti-lucennoi
- Cerdas periánticas (4)5-6, la mayoría más largas que el aquenio; aquenios lisos; tallos fértiles de (10)15-25 cm .................................................................................................................... R. fusca
Rhynchospora alba (L.) Vahl
Densamente cespitosa. Tallos fértiles de (10)15-40(46) cm. Hojas de 0,5-1,5(2) mm de anchura, canaliculadas o más o menos involutas, más cortas que los tallos. Bráctea inferior más corta que la inflorescencia. Inflorescencia formada por (1)2-4 panículas corimbiformes de espiguillas, agrupadas en la parte superior del tallo o las panículas inferiores separadas; espiguillas de 4,3-5,3 mm de longitud, fusiformes, no o poco comprimidas. Glumas ovadas, de ápice mucronado o acuminado, más raramente agudo, blancas durante la floración, aunque pueden amarillear con la edad, las inferiores estériles, las superiores con una flor hermafrodita en la axila. Cerdas periánticas 8-13, generalmente más largas que el aquenio, con acúleos dirigidos hacia atrás. 2 estambres. 2 estigmas. Aquenios de 1,3-1,5 mm de longitud, de contorno
40 41
TRIBU SCHOENEAE
TRIBU RHYNCHOSPOREAE
elíptico u obovado, lisos, raramente con minúsculas estrías transversales apenas perceptibles, verdosos o de color pardo claro; base del estilo de 0,8-1,5 mm, de contorno triangular-subulado, liso. 2n=26. Turberas y prados higroturbosos


Resérvaloen www.jolube.es




42 43
TRIBU RHYNCHOSPOREAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU RHYNCHOSPOREAE
Rhynchospora alba
TRIBU RHYNCHOSPOREAE
Rhynchospora alba: A) hábito; B) aquenio; C) gluma; D) inflorescencia.
© M. S. Villegas
SCIRPUS L.

Bajo este género se han venido incluyendo hasta hace unas décadas numerosos y muy diferentes linajes que, tras los estudios de embriología y filogenia molecular, se consideran actualmente como varios géneros, en ocasiones filogenéticamente muy alejados. Entre ellos, en lo que afecta al territorio de esta guía, señalamos Bolboschoenus, Isolepis, Schoenoplectus, Schoenoplectiella, Scirpoides y Trichophorum. No obstante, hasta el momento, los estudios mencionados no han resuelto completamente la delimitación del género Scirpus, puesto que recientes trabajos filogenéticos apuntan a que Scirpus es parafilético si no se consideran en él las especies -aparentemente tan distintas- del género Eriophorum. Se han apuntado dos alternativas a este conflicto: (1) la inclusión de Eriophorum en Scirpus, lo que implicaría numerosos cambios nomenclaturales que afectarían a nombres ampliamente consagrados entre la comunidad botánica, o (2) mantener Eriophorum como género independiente, lo que conllevaría la división de Scirpus en al menos 6 géneros diferentes y también requeriría llevar a cabo importantes modificaciones de la nomenclatura comúnmente usada. Dado que la resolución del problema necesita estudios filogenómicos que incluyan un muestreo mucho más amplio que el realizado hasta ahora, adoptamos aquí provisionalmente una posición conservadora y parsimoniosa, manteniendo Eriophorum y Scirpus como géneros independientes. Según este concepto, Scirpus consta de unas 47 especies nativas en su mayoría del hemisferio norte, con unas pocas distribuidas por Sudamérica y el sureste de Asia. Dichas especies se caracterizan por la presencia de hojas caulinares con limbo desarrollado, la frecuente existencia de perianto formado por cerdas cortas que no confieren aspecto algodonoso a la inflorescencia, y lígulas no densamente ciliadas. En nuestro territorio solo habita la especie tipo del género: S. sylvaticus.
Bibliografía
Martín-Bravo, S. & Luceño, M. (2008). Scirpus in Castroviejo, S.; Luceño; M., Galán; A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 34-36.

Jermy, C.; Simpson, D.; Foley, M. & Porter, M. (2007). Sedges of the British Isles. B.S.B.I. Handbook No. 1, ed. 3: 1-554. Botanical Society of the British Isles, London.
Larridon, I.; Zuntini, A.R.; Léveillé-Bourret, É; Barret, R.L.; Starr, J.R.; Muasya, A.M.; Villaverde, T.; Bauters, K.; Brewer, G.E.; Bruhl, J.J.; Costa, S.M.; Elliott, T.M.; Epitawalage, N.; Escudero, M.; Frailie, I.; Goetghebeur, P.; Hipp, A.L.; Jiménez-Mejías, P.; Kikuchi, I.A.B.S.; Luceño, M.; Márquez-Corro, J.I.; Martín-Bravo, S.; Maurini, O.; Pokorny, L.; Roalson, E.H.; Semmouri, I.; Simpson, D.A.; Spalink, D.; Thomas, W.W.; Wilson, K.L.; Xanthos, M.; Forest, F. & Baker, W.J. (2021). A new classification of Cyperaceae (Poales) supported by phylogenomic data. J. Syst. Evol. 59(4): 852-895.
Léveillé-Bourret, É.; Gilmour, C.N.; Starr, J.R.; Naczi, R.F.C. & Spalink, D. (2014). Searching for the sister to sedges (Carex): resolving relationships in the Cariceae-Dulichieae-Scirpeae clade (Cyperaceae). Bot. J. Linn. Soc. 176: 1-21.

Léveillé-Bourret, É.; Starr J.R.; Ford, B.A.; Moriarty Lemmon E. & Lemmon A.R. (2018). Resolving rapid radiations within angiosperm families using Anchored Phylogenomics. Systematic Biology 67: 94–112.
Léveillé-Bourret, É. & Starr J.R. (2019). Molecular and morphological data reveal three new tribes within the Scirpo-Caricoid Clade (Cyperoideae, Cyperaceae). Taxon 68: 218–245.
Scirpus sylvaticus L.

No o laxamente cespitosa, con rizoma de entrenudos más o menos largos. Tallos fértiles de 30-70(120) cm, trígonos. Hojas de 7-18 mm de anchura, basales y caulinares, plegadas, carenadas o, más raramente, planas. Brácteas inferiores 3, foliáceas, frecuentemente más largas que la inflorescencia. Inflorescencia de (9)11-22 x 10-20(26) cm, paniculiforme, multiespiculada; espiguillas de 2,4-4(5) mm de longitud, ovoides o elipsoides, con (10)12-20(22)
flores hermafroditas. Glumas de ovado-lanceoladas a obovadas, de ápice obtuso, redondeado o apiculado, negruzcas con una banda central verde, con margen escarioso o sin él, dispuestas en espiral. Cerdas periánticas 6, poco más cortas o largas que el aquenio, con numerosos acúleos dirigidos hacia atrás. 3 estambres. 3 estigmas. Aquenios de 0,8-1,1(1,3) mm de longitud, de contorno obovado, elíptico o, más raramente, suborbicular, inflado-trígonos, pajizos o pardoamarillentos en la madurez. 2n= 44. Turberas y prados muy húmedos en hayedos, sotos ribereños y otros bosques frescos.
52 53
TRIBU SCIRPEAE
TRIBU SCIRPEAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Scirpus sylvaticus
Resérvaloen www.jolube.es
Resérvaloen www.jolube.es



54 55
TRIBU SCIRPEAE
Scirpus sylvaticus
TRIBU SCIRPEAE
Scirpus sylvaticus: A) hábito; B) aquenio; C) gluma; D) espiguilla; E) radio terminal de la inflorescencia.
© M. S. Villegas
ERIOPHORUM L.
El género Eriophorum consta de 18 especies distribuidas por casi todas las regiones templadas y frías del hemisferio norte. En nuestro territorio habitan únicamente 4 especies que se distribuyen por las turberas de las montañas de la mitad norte de la península ibérica. En el área de esta guía el género es fácilmente identificable por sus inflorescencias similares al algodón, de ahí sus nombres populares de junco lanudo, junco de algodón o hierba algodonera. Ese aspecto es debido a la presencia de numerosas cerdas periánticas aplanadas y blancas (rojizas en ciertas especies boreales). Hojas basales y caulinares. Flores hermafroditas, con 3 estambres y 3 estigmas. La inflorescencia, dependiendo de la especie, puede constar de una o varias espiguillas.

Bibliografía
Villar, L. (2008). Eriophorum in Castroviejo, S.; Luceño, M.; Galán, A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 34-36.
Jermy, C.; Simpson, D.; Foley, M. & Porter, M. (2007). Sedges of the British Isles. B.S.B.I. Handbook No. 1, ed. 3: 1-554. Botanical Society of the British Isles, London.
Larridon, I.; Zuntini, A.R.; Léveillé-Bourret, É; Barret, R.L.; Starr, J.R.; Muasya, A.M.; Villaverde, T.; Bauters, K.; Brewer, G.E.; Bruhl, J.J.; Costa, S.M.; Elliott, T.M.; Epitawalage, N.; Escudero, M.; Frailie, I.; Goetghebeur, P.; Hipp, A.L.; Jiménez-Mejías, P.; Kikuchi, I.A.B.S.; Luceño, M.; Márquez-Corro, J.I.; Martín-Bravo, S.; Maurini, O.; Pokorny, L.; Roalson, E.H.; Semmouri, I.; Simpson, D.A.; Spalink, D.; Thomas, W.W.; Wilson, K.L.; Xanthos, M.; Forest, F. & Baker, W.J. (2021). A new classification of Cyperaceae (Poales) supported by phylogenomic data. J. Syst. Evol. 59(4): 852-895.
Léveillé-Bourret, É.; Gilmour, C.N.; Starr, J.R.; Naczi, R.F.C. & Spalink, D. (2014). Searching for the sister to sedges (Carex): resolving relationships in the Cariceae-Dulichieae-Scirpeae clade (Cyperaceae). Bot. J. Linn. Soc. 176: 1-21.

Léveillé-Bourret É.; Starr J.R.; Ford, B.A.; Moriarty Lemmon E. & Lemmon A.R. (2018). Resolving rapid radiations within angiosperm families using Anchored Phylogenomics. Syst. Biol. 67: 94–112.
Léveillé-Bourret É. & Starr J.R (2019). Molecular and morphological data reveal three new tribes within the Scirpo-Caricoid Clade (Cyperoideae, Cyperaceae). Taxon 68: 218–245.
CLAVE DE ESPECIES
1. Inflorescencia formada por una sola espiguilla; bráctea inferior glumácea 2
- Inflorescencia constituida por varias espiguillas generalmente pedunculadas; bráctea inferior foliácea ........................................................................................................................................................... 3
2. Plantas densamente cespitosas; vaina de la hoja superior inflada; anteras de aproximadamente 2,5(3) mm de longitud E. vaginatum
- Plantas no o laxamente cespitosas; vaina de la hoja superior aplicada al tallo; anteras de aproximadamente 1 mm de longitud ................................................................................ E. scheuchzeri
3. Pedúnculos de las espiguillas ásperos; aquenios de color pardo claro o medio E. latifolium
- Pedúnculos de las espiguillas lisos; aquenios de color pardo oscuro o negro E. angustifolium
Eriophorum angustifolium Honck. subsp. angustifolium
No o muy laxamente cespitosa, con rizoma de entrenudos largos. Tallos fértiles de (10)2060(90) cm, subcilíndricos u obtusamente trígonos hacia el ápice. Hojas de 2-6(8) mm de anchura, carenadas o canaliculadas, más raramente casi planas. Bráctea inferior glumácea con punta foliácea, de más corta a casi igual que la inflorescencia. Inflorescencia umbeliforme, formada por 3-5(7) espiguillas pedunculadas, con pedúnculos de hasta 8 cm, lisos, arqueados; espiguillas de contorno obovado y colgantes en la fructificación. Glumas de ovado-lanceoladas a ovadas, de ápice obtuso, agudo o acuminado, de pardo-rojizas a negruzcas, con margen escarioso, dispuestas en espiral. Cerdas periánticas numerosas, acrescentes, mucho más largas que el aquenio, no aculeoladas, lisas en la zona apical. 3 estambres. 3 estigmas. Aquenios de (1)23 mm de longitud, de contorno obovado, inflado-trígonos, de color pardo oscuro o negruzcos, mates o ligeramente brillantes. 2n= 54, 58, 60. Turberas ácidas.
56 57
TRIBU SCIRPEAE
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU SCIRPEAE
www.jolube.es
Eriophorum angustifolium subsp. angustifolium
Resérvaloen
Eriophorum latifolium Hoppe



Laxamente cespitosa, con rizoma de entrenudos más o menos cortos. Tallos fértiles de 2060(90) cm, obtusamente trígonos. Hojas de (2)3-5(6) mm de anchura, planas o ligeramente carenadas. Bráctea inferior glumácea con punta foliácea, de más corta a poco más larga que la inflorescencia. Inflorescencia umbeliforme, formada por (2)4-10(12) espiguillas pedunculadas, con pedúnculos de hasta 9 cm, fuertemente escábridos, arqueados; espiguillas de contorno obovado y colgantes o inclinadas en la fructificación. Glumas de ovado-lanceoladas a ovadas, de ápice más o menos agudo, de color verde oliva oscuro a negruzcas, con margen escarioso, dispuestas en espiral. Cerdas periánticas numerosas, acrescentes, mucho más largas que el aquenio, no aculeoladas, papilosas en la zona apical. 3 estambres. 3 estigmas. Aquenios de 3-3,5 mm de longitud, de contorno ligeramente obovado, inflado-trígonos, de color pardo o pardo-rojizo, mates o ligeramente brillantes. 2n= 54, 58, 72. Turberas sobre sustrato silíceo o calizo.



58 59 Eriophorum angustifolium subsp. angustifolium TRIBU SCIRPEAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU SCIRPEAE Eriophorum latifolium
Resérvaloen www.jolube.es
Eriophorum scheuchzeri Hoppe



No o muy laxamente cespitosa, con rizoma de entrenudos largos. Tallos fértiles de 15-30(40) cm, más o menos cilíndricos. Hojas de 1-2 mm de anchura, basales salvo una caulinar con vaina adosada al tallo y limbo muy reducido, de carenadas a canaliculadas, raramente algo planas. Bráctea inferior glumácea, mucho más corta que la inflorescencia. Inflorescencia reducida a una espiguilla situada al final del tallo fértil, más o menos subglobosa, erecta. Glumas de lanceoladas a ovado-lanceoladas, las inferiores estériles, con estambres abortados, agudas o acuminadas, de color pardo muy oscuro a negruzcas, con margen escarioso, dispuestas en espiral. Cerdas periánticas numerosas, acrescentes, mucho más largas que el aquenio, no aculeoladas, lisas en la zona apical. 3 estambres. 3 estigmas. Aquenios de (2)2,5-3(3,5) mm de longitud, obovoides, inflado-trígonos, de color pardo oscuro, mates. 2n= 58. Turberas en el borde de lagos alpinos.

60 61
TRIBU SCIRPEAE © M. S. Villegas 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU SCIRPEAE Eriophorum scheuchzeri
Eriophorum angustifolium: A) hábito; B) aquenio; C) gluma; D) aquenio junto con las cerdas periánticas; E) espiguilla. Eriophorum latifolium: F) espiguilla.
Resérvaloen www.jolube.es
CAREX L.

Origen, distribución y hábitat.
Con poco más de 2000 especies aceptadas, Carex es el tercer género más biodiverso de las angiospermas, únicamente superado por Bulbophyllum (Orquidaceae; ca. 3200) y Astragalus (Fabaceae; ca. 3000). Tiene una distribución prácticamente cosmopolita, y solo está ausente de la Antártida continental, algunos desiertos cálidos y selvas ecuatoriales de zonas bajas. No obstante, su riqueza en especies no está uniformemente distribuida, de manera que cuenta con un mayor número en regiones templadas y frías del hemisferio norte, especialmente en el Este de Asia y el NE de Norteamérica. En el oeste del Paleártico, donde se incluye el área cubierta por esta guía, el número de especies es considerablemente menor que en Asia y Norteamérica, aunque superando ampliamente las 200. Por su parte, en el hemisferio sur, aunque el número de especies es menor que en el norte, también hay zonas de elevada diversidad relativa, como Patagonia y Nueva Zelanda. Este patrón general de distribución de la riqueza de especies se ajusta, a grandes rasgos, a un gradiente latitudinal, con más especies cuanto mayor latitud (exceptuando las latitudes polares extremas). Ello no deja de ser sorprendente porque resulta contrario al que muestran la mayoría de los grandes grupos de seres vivos, incluyendo muchos otros géneros de ciperáceas, cuya diversidad de especies es mayor en regiones cercanas al Ecuador.
Resérvaloen www.jolube.es
Evidencias fósiles y análisis de datación de filogenias moleculares nos indican que el género se originó probablemente en el este de Asia en el Eoceno tardío, hace unos 3540 millones de años. Allí permaneció probablemente durante un largo período (al menos 10 millones de años) en el que diversificaron sus principales linajes, antes de iniciar la colonización del resto del mundo, que debió de tener lugar desde las etapas finales del Oligoceno (hace ca. 23-24 millones de años). Investigaciones recientes han arrojado luz sobre la historia evolutiva que explica la apasionante biogeografía de Carex. Así, la distribución actual del género es el resultado de una enorme diversificación en el hemisferio norte, propiciada por una extraordinaria habilidad para dispersarse a larga distancia. Ello le ha permitido alcanzar múltiples territorios a pesar de no contar con mecanismos especializados para dicha dispersión (excepto el subgénero Uncinia, no presente en el territorio cubierto por esta guía -véase la Introducción general-, y alguna que otra especie de varias secciones: C. collinsii, que muestra los dientes del pico del utrículo en forma de gancho; C. vesicaria, C. rostrata y otras, con utrículos inflados y flotantes; y C. leporina, C. praecox, etc., cuyos utrículos alados facilitan su dispersión por el viento). Así, Carex ha conseguido colonizar en múltiples ocasiones independientes las principales masas de tierra del hemisferio sur (excepto la Antártida, aunque sí está presente en diversas islas subantárticas), así como los principales archipiélagos oceánicos de origen volcánico (excepto Galápagos).
A pesar de que Carex muestra una gran variabilidad morfológica y ecológica a lo largo de los territorios en los que está presente, en general presenta afinidad por hábitats con alta disponibilidad de agua de zonas templadas y frías. De hecho, a lo largo de su historia evolutiva, la diversificación de Carex parece haber sido favorecida durante períodos geológicos caracterizados por enfriamientos globales del clima, como el Mioceno tardío. Estas circunstancias llevan a pensar que, con toda probabilidad, el actual calentamiento global ocasionado por la actividad antropogénica no es positivo para la diversificación y conservación del género. Muchos hábitats en los que hay gran abundancia de especies de Carex, o en los que incluso éstas juegan un papel ecológico clave por su dominancia, están
66 67
TRIBU TRICHOPHORUM TRIBU TRICHOPHORUM
Trichophorum cespitosum: A) hábito; B) aquenio; C) bráctea inferior; D) gluma; E) inflorescencia; F) hoja superior.
© M. S. Villegas
TRIBU CARICEAE
ya sufriendo los efectos negativos del cambio climático, entre ellos los ecosistemas de alta montaña (turberas, comunidades quionófilas, prados alpinos, etc.). Este hecho resulta especialmente preocupante en las zonas más vulnerables al cambio climático, como las montañas de regiones tropicales o de la cuenca mediterránea, incluidas las principales cordilleras ibéricas.
Carex en la península ibérica, Baleares y Macaronesia.
En la península ibérica y los archipiélagos de Azores, Baleares, Canarias y Madeira habitan 112 especies y 9 subespecies de Carex. El territorio continental, junto con las Islas Baleares, alberga 102 especies y 6 subespecies, todas nativas; mientras que en Macaronesia se conocen únicamente 27 especies (9 endémicas) y 2 subespecies (endémicas), todas nativas salvo quizás C. debilis Michx., que podría haber sido introducida por el ser humano desde Norteamérica. Estos táxones se encuadran en 4 subgéneros (de los 6 aceptados) y 47 secciones y/o clados. El emplazamiento exacto de unas pocas especies está aún pendiente de estudios sistemáticos más profundos, por lo que en dichos casos hemos preferido indicar el nombre del linaje (clado) al que pertenecen, según los últimos estudios filogenéticos.
Subgénero Sección /Clado (nº especies)
Psyllophorae (6)
Especies
Phsyllophorae (5) C. distachya, C. macrostyla, C. oedipostyla, C. peregrina, C. pulicaris
Baldenses (1) C curvula
Clado Kobresia (2) C. myosuroides, C. simpliciuscula
Subgénero Sección /Clado (nº especies)
Albae (1) C alba
Aulocystis (1) C. sempervirens
Clado Ericetorum (1) C. ericetorum
Secalinae (1) C. hordeistichos
Clado Liparocarpos (1) C. liparocarpos
Especies
Eutyceras (4)
Clado Capitata (1) C. rupestris
Clado Pauciflora (1) C. pyrenaica
Stellulatae (1) C. echinata
Foetidae (1) C. foetida
Glareosae (4) C. canescens, C. furva, C. lachenalii, C. lucennoiberica
Paniculatae (1) C. paniculata
Divisae (1) C. divisa
Phaestoglochin (7) C. canariensis, C. divulsa, C. leersii, C. muricata, C. otrubae, C. pairae, C. spicata
Digitatae (2) C digitata, C ornithopoda
Vignea (25)
Resérvaloen www.jolube.es
Mitratae (3) C. basilaris, C. caryophyllea, C. depressa
Clado Conica (1) C umbrosa
Rhomboidales (1) C. brevicollis
Chlorostachyae (1) C. capillaris
Clandestinae (1) C. humilis
Hallerianae (2) C. halleriana, C. rorulenta
Acrocystis (2) C. montana, C. pilulifera
Clado Carex (6)
Phacocystis (7)
C. hirta, C. lasiocarpa, C. pseudocyperus, C. riparia, C. rostrata, C. vesicaria
C. acuta, C. cespitosa, C. elata, C. nigra, C. quixotiana, C. reuteriana, C. trinervis
Carex (77)
Limosae (1) C. limosa
Hispanicae (1) C. durieui
Clado Tomentosa (1) C. tomentosa
Paniceae (3) C. bicolor, C. olbiensis, C. panicea
Clado Flacca (8) C. asturica, C. austroalpina, C. brachystachys, C. caudata, C. depauperata, C. flaca, C. grioletii, C. hispida
Racemosae (2) C. atrata, C. parviflora
Frigidae (1) C. frigida
Rhynchocystis (3) C. leviosa, C. pendula, C. sequeirae
Ceratocystis (6)
C. demissa, C. flava, C. hostiana, C. lepidocarpa, C. nevadensis, C. oederi
Strigosae (1) C. strigosa
Sylvaticae (3) C. paui, C. sylvatica, C. vulcani
Ammoglochin (4) C. arenaria, C. brizoides, C. pseudobrizoides, C. praecox
Remotae (1) C remota
Clado Annectens (1) C diandra
Physoglochin (1) C. davalliana
Holarrhenae (1) C. disticha
Cyperoideae (2) C. bohemica, C. leporina
Spirostachyae (13) C. binervis, C. camposii, C. distans, C. extensa, C. helodes, C. hochstetteriana, C. laevicaulis, C. laevigata, C. lainzii, C. lowei, C. mairei, C. perraudieriana, C. punctata
Clado Castanea (1) C. debilis
Clado Mollicula (1) C. pallescens
Paludosae (1) C. acutiformis
Encuadramiento taxonómico de las especies de Carex presentes en el territorio. Entre paréntesis se indica el número de especies.
68 69
TRIBU CARICEAE
TRIBU CARICEAE
Características morfológicas.
Todas las especies de Carex son plantas perennes, aunque en ocasiones, como el caso de C. oederi, de vida corta. En función de la longitud de los entrenudos del rizoma (véase Introducción general), el hábito puede ser densamente cespitoso (entrenudos muy cortos), a veces formando incluso voluminosas macollas, como en C. paniculata y ciertas especies de la sección Phacocystis (C. reuteriana y C. acuta, por ejemplo). Sin embargo, otras especies (vg. C. arenaria y C. divisa) exhiben tallos aéreos muy espaciados unos de otros, debido a que los entrenudos del rizoma son muy largos. Los tallos fértiles son trígonos (más raramente cilíndricos o casi) y, en las especies ibéricas, oscilan entre los 2 cm de altura en ciertos ejemplares alpinos de C. capillaris y los más de dos metros de algunos individuos robustos de C. pendula y C. leviosa. La forma y consistencia de las hojas se ajustan a lo descrito para el conjunto de la familia.
Las flores son siempre unisexuales y aclamídeas (sin perianto), las masculinas reducidas a tres estambres dispuestos en la axila de una gluma y las femeninas a un ovario sincárpico bi o tricarpelar (excepcionalmente con 4 carpelos en algunas especies no presentes en el territorio). La forma, color, ápice y número de nervios de las glumas proporcionan interesantes caracteres taxonómicos.
La unidad básica de la inflorescencia es la espiguilla, cuyo conjunto forma 1-numerosas espigas, agrupadas a su vez en espigas, racimos o panículas. Las espigas pueden ser masculinas, femeninas, andróginas (con flores masculinas en la parte superior de la espiga y femeninas en la inferior) o ginecandras (flores femeninas en la parte apical y masculinas en la basal). En raras ocasiones pueden observarse espigas con flores masculinas en la zona apical, femeninas en la media y masculinas en la basal (androginecandras) o con flores femeninas en el ápice, masculinas en la parte media y femeninas en la base (ginandróginas); mucho más raramente, ciertas especies presentan una distribución irregular del sexo de las flores en las espigas (vg. C. bequaertii, ausente del territorio cubierto por esta guía).
Resérvaloen www.jolube.es
La espiguilla femenina, muy reducida, se compone de un eje rudimentario o corto (raquidio) del que surge lateralmente el ovario y cuyo conjunto se encuentra rodeado por un profilo que, cuando tiene los bordes soldados y forma de pequeña botella, se denomina utrículo, y cuando es escuamiforme y de bordes libres recibe el nombre de periginio glumiforme, como es el caso del que rodea al ovario de ciertas especies de Carex antiguamente incluidas en el género Kobresia. El profilo está también presente con frecuencia en ramas de órdenes inferiores, en cuyo caso suele ser hialino y tubular. En resumen, la presencia de profilo es indicadora de que se ha producido una ramificación. El utrículo es quizás el órgano que más caracteres aporta para la separación de los distintos táxones de Carex y en él se distinguen tres partes: estípite o parte inferior, frecuentemente muy corto o rudimentario; cuerpo o parte media, que es la principal; y pico o parte superior, a veces muy reducido o ausente y del que, a grandes rasgos, se distinguen tres tipos: de ápice truncado, bidentado o bífido. Cuando el ovario tiene tres carpelos el cuerpo del utrículo es trígono, mientras que cuando está formado por dos su forma es lenticular (planoconvexo o biconvexo). El contorno, el número de nervios y su mayor o menor prominencia, así como la presencia o ausencia (utrículo liso) de expansiones epidérmicas (papilas, acúleos o pelos) son también caracteres importantes para la clasificación de las especies.
Partes del utrículo de Carex lucennoiberica
El fruto es un aquenio de contorno variable, útil para separar grupos de especies. Su ápice puede ser columnar, truncado, ciatiforme (en forma de copa) etc., carácter este de gran importancia taxonómica.



70 71
TRIBU CARICEAE
Tipos de pico del utrículo según su ápice. A. Carex panicea; B. Carex extensa; C. Carex vesicaria
TRIBU CARICEAE
A. Aquenio de Carex praecox con ápice de tipo columnar. B. Ápice del aquenio ciatiforme de Carex basilaris C. Ápice del aquenio truncado de Carex caryophyllea. D. Ápice del aquenio trígono-columnar de Carex umbrosa subsp. umbrosa
Bibliografía
Global Carex Group: Waterway, M.J.; Roalson, E.H.; Ford, K.A.; Luceño, M.; Martín-Bravo, S.; Starr, J.R.; Wilson, K.L.; Yano, O.; Zhang, S.R.; Alverson, W.S.; Bruederle, L.P.; Bruhl, J.J.; Chung, K.-S.; Cochrane, T.S.; Escudero, M.; Ford, B.A.; Gebauer, S.; Gehrke, B.; Hann, M.; Hipp, A.L.; Hoffmann, M.H.; Hoshino, T.; Jiménez-Mejías, P.; Jin, X.-F.; Jung, J.; Kim, S.; Maguilla, E.; Masaki, T.; Míguez, M.; Molina, A.; Naczi, R.F.C.; Reznicek, A.A.; Rothrock, P.E.; Simpson, D.A.; Spalink, D.; Thomas, W.W. & Villaverde, T. (2015). Making Carex monophyletic (Cyperaceae, tribe Cariceae): a new broder circumscription. Bot. J. Linn. Soc. 179: 1-42.
Jiménez-Mejías, P.; Hann, M.; Lueders, K.; Starrr, J.R.; Brown, B.H.; Choulnard, B.N.; Chung, K.-S.; Escudero, M.; Ford, B.A.; Ford, K.A.; Gebauer, S.; Gehrke, B.; Hoffmann, M.H.; Jin, X.-F.; Jung, J.; Kim, S.; Luceño, M.; Maguilla, E.; Martín-Bravo, S.; Míguez, M.; Molina, A.; Naczi, R.F.C.; Pender, J.E.; Reznicek, A.A.; Villaverde, T.; Waterway, M.J.; Yang, J.-C.; Zhang, S.; Hipp, A.L. & Roalson, E.H. (2016). Megaphylogenetic specimen-level approaches to the Carex (Cyperceae) phylogeny using ITS, ETS, and matK sequences: implications for classification. Syst. Bot. 41: 500-518.
Jiménez-Mejías, P.; Luceño, M.; Wilson, K.L.; Waterway, M.J. & Roalson, E.H. (2016). Clarification of the Use of the Term Perigynium and Utricle in Carex (L.) (Cyperaceae). Syst. Bot. 41(3): 519-528.
Luceño, M. (1994). Monografía del género Carex en la península ibérica e Islas Baleares. Ruizia 14: 1-144.
Luceño, M. (2008). Carex, in Castroviejo, S.; Luceño; M.; Galán, A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 109-250.
Maguilla, E.; Escudero, M.; Hipp, A.L. & Luceño, M. (2017). Allopatric speciation despite historical gene flow: Divergence and hybridization in Carex furva and C. lucennoiberica (Cyperaceae) inferred from plastid and nuclear RAD-seq data. Mol. Ecol. 26: 5646-5662.
Martín-Bravo, S.; Jiménez-Mejías, P.; Villaverde, T.; Escudero, M.; Hahn, M.; Spalink, D.; Roalson, E.H.; Hipp, A.L.; and the Global Carex Group: Benítez-Benítez, C.; Bruederle, L.P.; Fitzek, E.; Ford, B.A.; Ford, K.A.; Garner, M.; Gebauer, S.; Hoffmann, M.H.; Jim, X.-F.; Larridon, I.; Lévéille-Bourret, É.; Lu, Y.-F.; Luceño, M.; Maguilla, E.; Márquez-Corro, J.I.; Míguez, M.; Naczi, R.F.C.; Reznicek, A.A. & Starr, J.R. (2019). A tale of worldwide success: Behind the scenes of Carex (Cyperaceae) biogeography and diversification. J. Syst. Evol. 57: 695-718.
Roalson, E.H.; Jiménez-Mejías, P.; Hipp, A.L.; Benítez-Benítez, C.; Bruederle, L.P.; Chung, K.-S.; Escudero, M.; Ford, B.A.; Ford, K.A.; Garner, M.; Gebauer, S.; Gehrke, B.; Hahn, M.; Hayat, M.Q.; Hoffmann, M.H.; Jim, X.-F.; Larridon, I.; Lévéille-Bourret, É.; Lu, Y.-F.; Luceño, M.; Maguilla, E.; Márquez-Corro, J.I.; Martín-Bravo, S.; Míguez, M.; Naczi, R.F.C.; Reznicek, A.A.; Spalink, D.; Starr, J.R.; Uzma; Villaverde, T.; Waterway, M.J.; Wilson, K.L. & Zhang, S.-R. (2021). A framework infrageneric classification of Carex (Cyperaceae) and its organizing principles. J. Syst. Evol. 59: 726-762.
Villar, L. (2008). Kobresia in Castroviejo, S.; Luceño; M., Galán; A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 107-109.
Villaverde, T.; Jiménez-Mejías, P.; Luceño, M.; Roalson, E.H.; Hipp, A.L.; and the Global Carex Group: Wilson, K.L.; Larridon, I.; Gebauer, S.; Hoffmann, M.H.; Simpson, D.A.; Naczi, R.F.C.; Reznicek, A.A.; Ford, B.A.; Starr, J.R.; Park, J.; Escudero, M. & Martín-Bravo, S. (2020). A new classification of Carex subgenera supported by HybSeq backbone phylogeny. Bot. J. Linn. Soc. 194: 141-163.
CLAVE DE ESPECIES
1. Envoltura del aquenio de bordes libres (periginio glumiforme)
- Envoltura del aquenio cerrada formando una estructura más o menos con forma de pequeña botella (utrículo o periginio utriculiforme) 3
2. Tallos cilíndricos en toda su longitud; inflorescencia aparentemente no ramificada; vainas de las hojas viejas acumuladas en la base de la planta, muy brillantes, de color pardo claro ............................................................................................................................................... C. myosuroides
- Tallos más o menos trígonos bajo la inflorescencia; inflorescencia claramente ramificada; vainas de las hojas viejas poco visibles, apenas acumuladas en la base, mates, generalmente de color pardo oscuro ....................................................................... C. simpliciuscula
3. Inflorescencia formada por una sola espiga terminal; bráctea inferior escuamiforme (glumiforme), sensiblemente menor que la espiga 4
- Inflorescencia formada por dos o más espigas, muy raramente algún tallo aislado con una sola, pero entonces la bráctea inferior es más larga que la espiga 9
4. Individuos que generan solo flores masculinas y otros únicamente femeninas (plantas dioicas); pico del utrículo curvado; plantas densamente cespitosas .............. C. davalliana
- Todos los individuos generan flores masculinas y femeninas (plantas monoicas); pico del utrículo recto; plantas cespitosas o no ...................................................................................................... 5
5. La gran mayoría de las flores femeninas con 3 estigmas 6
- Todas las flores femeninas con 2 estigmas ........................................................................................... 7
6 Utrículos suberectos, de anchamente elipsoides a obovoides, bruscamente estrechados en un pico de hasta 0,4 mm; espiga con hasta 8(15) flores femeninas; plantas no cespitosas, con rizomas de entrenudos largos; glumas persistentes sobre el eje de la espiga tras la caída de los utrículos .................................................................................... C. rupestris
- Utrículos finalmente patentes o ligeramente reflejos, de contorno lanceolado o estrechamente ovado o elíptico, suavemente atenuados en un pico de 0,81 mm; espiga con (12)20-45(67) flores femeninas; plantas densamente cespitosas, con rizomas de entrenudos muy cortos; glumas y utrículos que caen simultáneamente
C. pyrenaica
7. Pico del utrículo de 0,3-0,5 mm; hojas setáceas; plantas laxamente cespitosas; glumas masculinas inferiores agudas ................................................................................................... C. pulicaris
- Pico del utrículo de 1-2 mm; hojas canaliculadas, no setáceas; plantas densamente cespitosas; glumas masculinas inferiores obtusas 8
8. Utrículos más o menos bruscamente estrechados en el pico, con al menos dos nervios resaltados; glumas femeninas con margen escarioso ancho en toda su longitud; tallos (10)20-40 cm; espiga (20)25-30 mm de longitud C. peregrina
- Utrículos gradualmente atenuados en el pico, con nervios poco o nada perceptibles; glumas femeninas con margen escarioso estrecho hacia el ápice; tallos (3)8-20(30) cm; espiga (11)15-26 mm C. macrostyla
9. Espigas de apariencia similar, la terminal siempre con flores de ambos sexos o solo femeninas (raramente solo masculinas) 10
- Espiga(s) superior(es) de apariencia diferente a la(s) inferior(es), la terminal masculina o con escasas flores femeninas en la base y/o en el ápice ............................................................ 40
11. Utrículos densamente ornamentados con papillas evidentes 12
- Utrículos sin papilas 13
72 73
2
TRIBU CARICEAE
10. Estigmas 3 11
- Estigmas 2 .............................................................................................................................................................. 16
TRIBU CARICEAE
Resérvaloen www.jolube.es
Carex oedipostyla Duval-Jouve



Cespitosa. Tallos fértiles de 11-20(27) cm, trígonos. Hojas de 1-2,6(3) mm de anchura, más o menos planas, más largas que los tallos. Bráctea inferior foliácea, más larga que la inflorescencia, envainadora. Inflorescencia formada por 2-5 espigas de apariencia similar, más o menos separadas, las (1)2(3) superiores subsésiles, las (1)2 inferiores basilares o casi, con las flores masculinas densamente agrupadas en la parte superior y las femeninas laxamente agrupadas en la inferior. Glumas femeninas (sin contar el mucrón) más cortas que los utrículos, hialinas o verdosas, largamente mucronadas. Utrículos de 3-4(4,5) mm de longitud, suberectos, con los nervios muy prominentes, sin pico o con él rudimentario, de hasta 0,1 mm. Raquidio desarrollado. Base del estilo piramidal. 3 estigmas.
2n=68 Sotobosque de encinares, alcornocales, pinares y brezales

Resérvaloen www.jolube.es
96 97
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic TRIBU CARICEAE
TRIBU CARICEAE
Carex oedipostyla
Carex oedipostyla
Laxamente cespitosa. Tallos fértiles de (4)1025(40) cm, de obtusamente trígonos a subcilíndricos. Hojas de 0,2-0,9 mm de anchura, setáceas, de estrechamente canaliculadas a casi enrolladas, más cortas, raramente más largas, que los tallos. Bráctea inferior glumácea que protege a un utrículo. Inflorescencia formada por una espiga solitaria de 1-3 cm, con la parte masculina en la zona superior, densa, y la femenina en la inferior, laxa. Glumas femeninas prontamente caedizas, de color pardo, agudas u obtusas, con margen escarioso. Utrículos de 4-5 mm de longitud, reflejos en la madurez, estrechamente fusiformes, de color pardo obscuro o ferrugíneos, con los nervios poco o nada perceptibles, atenuados en un pico de 0,3-0,5 mm, de ápice hialino, truncado y hendido. 2 estigmas. 2n=58, 60. Turberas.




98 99 TRIBU CARICEAE
Carex oedipostyla: A) hábito; B) utrículo; C) aquenio; D) gluma femenina; E) gluma masculina; F) espiga andrógina.
© M. S. Villegas
3600 2400 1800 1200 600 3000 Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic pH TRIBU CARICEAE
L.
www.jolube.es
Carex pulicaris
Carex pulicaris Resérvaloen
No cespitosa, rizomas largos y gruesos. Tallos fértiles de (10)15-40(70) cm, más o menos pubescentes. Hojas de (1,6)3-5(7) mm de anchura, planas, más cortas o largas que los tallos, con pubescencia más o menos densa en ambas caras. Bráctea inferior foliácea, igual o poco más larga que la inflorescencia, envainadora. Inflorescencia en racimo de espigas de aspecto diferente, las (1)2-3(4) superiores masculinas, estrechamente fusiformes; las (1)2-3 inferiores femeninas, raramente la superior con algunas flores masculinas en el ápice, cilíndricas, largamente oblongas, erectas, distantes, a veces cortamente ramificadas en la base. Glumas femeninas lanceoladas o estrechamente ovadas, largamente aristadas, verdosas o de color pardo claro. Utrículos de (4,5)5,5-6,5(7,2) mm de longitud, suberectos, membranosos. densamente pubescentes, con los nervios muy prominentes, gradualmente atenuados en un pico de (1,5)2-2,5(3,2) mm, bífido. 3 estigmas. 2n=112, 113, 114 Prados y herbazales medianamente húmedos, frecuentemente nitrificados.





250 251 CLADO CAREX Carex hirta L. TRIBU CARICEAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Carex hirta
TRIBU CARICEAE
Carex hirta
www.jolube.es
Carex hirta
Resérvaloen
Carex lasiocarpa Ehrh.



Resérvaloen www.jolube.es
No cespitosa, rizomas largos, no muy gruesos. Tallos fértiles de 60-80(90) cm, glabros. Hojas de 1-2(2,5) mm de anchura, canaliculadas o carenadas, algo más cortas que los tallos, glabras. Bráctea inferior más larga que la inflorescencia, cortamente envainadora. Inflorescencia en espiga de espigas de aspecto diferente, las (1)2 superiores masculinas, estrechamente fusiformes; las (1)2(3) inferiores femeninas, de estrechamente ovoideas a cortamente cilíndricas, erectas, aproximadas o distantes. Glumas femeninas ovadas, de agudas a largamente aristadas, de color pardo-rojizo a pardo-purpúreo oscuro. Utrículos de (3,5)4-4,5(4,8) mm de longitud, suberectos, membranosos, densamente tomentosos, con los nervios bien marcados, más o menos bruscamente estrechados, con menor frecuencia suavemente atenuados, en un pico de (0,3)0,5-0,8(1) mm, bífido. 3 estigmas. 2n=56, 78, 80-82, 84, 85 Turberas muy ácidas en áreas de montaña.

252 253 TRIBU CARICEAE
Carex hirta: A) hábito; B) utrículo; C) aquenio; D) gluma femenina; E) gluma masculina; F) espiga femenina.
© M. S. Villegas
TRIBU CARICEAE 3600 2400 1800 1200 600 3000 pH VU Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Carex lasiocarpa
Carex lasiocarpa
Eleocharis acicularis (L.) Roem. & Schult.




No o laxamente cespitosa, con rizoma de entrenudos largos y filiformes. Tallos fértiles de (1)4-11(22) cm, más o menos capilares. Vainas membranosas, blanquecinas o variablemente teñidas de púrpura, la superior frecuentemente algo inflada, de ápice oblicua u horizontalmente truncado, obtuso o redondeado. Espiguillas de (1,5)3-5(6) mm, ovoides o elipsoides, de ápice variable, laxas, con (2)5-9(14) flores. Glumas de (1)1,5-2(2,5) mm, ovadas, agudas, con ancho margen escarioso, todas fértiles, la inferior rodea casi completamente la base de la espiguilla. Cerdas periánticas 0-3. 3 estambres. 3 estigmas. Aquenios de 0,7-1,2(1,5) mm de longitud, estrechamente obovoides, obtusamente trígonos o de sección transversal circular, pajizos o amarillentos, más o menos brillantes, con costillas longitudinales muy marcadas unidas por estrías transversales más tenues; base del estilo de cónica a piramidal, de relación longitudanchura variable, generalmente contraída en la base dejando una fina ranura en la zona de unión con el aquenio, no decurrente sobre este. 2n=10, 18, 20, 36-38, 50-58. Turberas, prados muy húmedos, bordes de lagunas y charcas de poca profundidad.

Resérvaloen www.jolube.es
446 447 TRIBU ELEOCHARIDEAE
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic Eleocharis acicularis TRIBU ELEOCHARIDEAE
Eleocharis acicularis
Eleocharis parvula (Roem. & Schult.) Bluff, Nees & Schauer
Laxamente cespitosa, con rizoma de estolones muy finos que rematan en pequeños bulbos ovoides. Tallos fértiles de (1,5)2-8(12) cm. Vainas escariosas, a veces rojizas en la base, la superior no inflada, de ápice oblicuamente truncado. Espiguillas de (2,8)3-3,5(3,8) mm, más o menos ovoides, de ápice agudo u obtuso, laxas, con 4-5 flores. Glumas de (1,1)1,5-2 mm, dispuestas casi en dos filas, ovado-lanceoladas, de agudas a redondeadas, con margen escarioso, todas fértiles excepto, generalmente, la inferior, que rodea como mucho la mitad de la base de la espiguilla y es mayor que el resto. Cerdas periánticas 3 o 6, escábridas. 3 estambres. 3 estigmas. Aquenios de aproximadamente 1 mm de longitud, obovoides, trígonos, pajizos o parduscos, algo brillantes, finamente punteados o reticulados; base del estilo piramidal, de relación longitudanchura variable, sin solución de continuidad con el ápice del aquenio, pero con un delgado reborde blanquecino, rojizo o negruzco en la zona de unión. 2n=10. Marismas, estuarios, campos de arroz y otros suelos salinos húmedos cercanos a la costa.
BULBOSTYLIS Kunth
El género Bulbostylis comprende aproximadamente 230 especies que se distribuyen principalmente por las regiones tropicales y subtropicales, con unas pocas especies en las regiones temperadas de América del Norte, sur de Europa y oeste de Asia. Después de Fimbristylis, es el más biodiverso de los 10 géneros que integran la tribu Abildgaardieae. Bulbostylis está representado en el territorio exclusivamente por dos especies, una de ellas esporádicamente naturalizada. El género se caracteriza por la presencia de largos pelos blancos en el ápice de las vainas, inflorescencia generalmente antelada, perianto ausente y estilo glabro, cuya base es casi siempre gruesa y persistente sobre el aquenio en la madurez.
Bibliografía
Larridon, I.; Zuntini, A.R.; Léveillé-Bourret, É; Barret, R.L.; Starr, J.R.; Muasya, A.M.; Villaverde, T.; Bauters, K.; Brewer, G.E.; Bruhl, J.J.; Costa, S.M.; Elliott, T.M.; Epitawalage, N.; Escudero, M.; Frailie, I.; Goetghebeur, P.; Hipp, A.L.; Jiménez-Mejías, P.; Kikuchi, I.A.B.S.; Luceño, M.; Márquez-Corro, J.I.; Martín-Bravo, S.; Maurini, O.; Pokorny, L.; Roalson, E.H.; Semmouri, I.; Simpson, D.A.; Spalink, D.; Thomas, W.W.; Wilson, K.L.; Xanthos, M.; Forest, F. & Baker, W.J. (2021). A new classification of Cyperaceae (Poales) supported by phylogenomic data. J. Syst. Evol. 59(4): 852-895.
Luceño, M. & Escudero, M. (2008). Bulbostylis in Castroviejo, S.; Luceño; M.; Galán; A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 97-99.

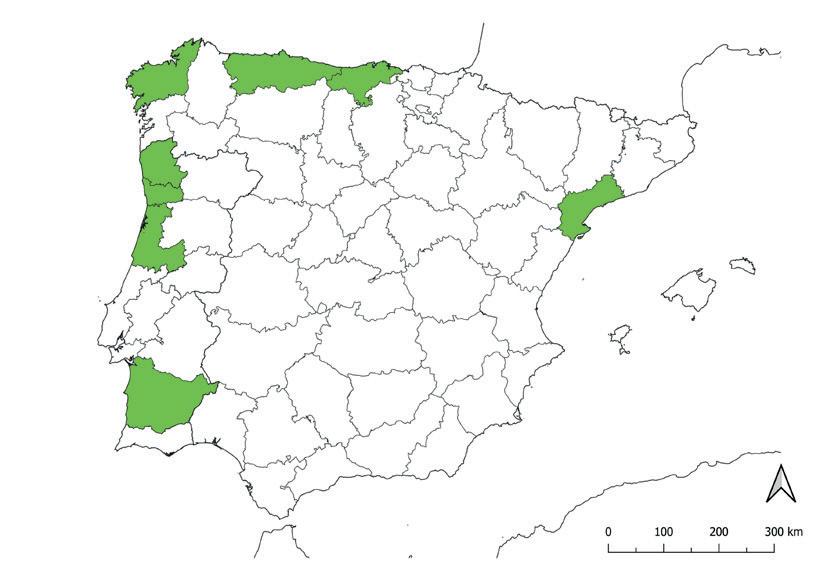
Verloove, F.; Laguna Lumbreras, E. & Ferrer Gallego, P.P. (2014). Some potentially weedy Cyperaceae new to Spain. Fl. Med. 24: 197-205.
CLAVE DE ESPECIES
1. Tallos pelosos; inflorescencia laxa; glumas de 2,5-3,5(4,5) mm; aquenios de (0,8)11,3 mm ................................................................................................................................................................ B. cioniana
Tallos glabros; inflorescencia bastante densa; glumas de 1,7-2,3 mm; aquenios de 0,7-1 mm .............................................................................................................................................. B. thouarsii
Bulbostylis cioniana (Pi. Savi) Lye
Anual o perenne de vida corta, cespitosa. Tallos de 5-10 cm, pubescentes, con costillas claramente prominentes. Hojas de 0,4-0,5 mm de anchura, filiformes, más cortas que los tallos, con frecuencia ciliadas hacia el ápice; vainas basales más o menos enteras, de color pardo pálido. Inflorescencia


456 457
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic EN TRIBU ELEOCHARIDEAE Eleocharis parvula TRIBU ABILDGAARDIEAE
-
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic EN Resérvaloen www.jolube.es
SCHOENOPLECTIELLA Lye

Género descrito por el ilustre ciperólogo noruego Kåre Arnstein Lye (1940-2021) para incluir ciertas especies antiguamente encuadradas en Schoenoplectus y/o Scirpus Se clasifica actualmente en la tribu Pseudoschoeneae, junto con el género Pseudoschoenus. La tribu se diferencia de Schoenoplecteae, filogenéticamente próxima y que contiene Schoenoplectus, por caracteres microanatómicos del embrión, así como porque los bordes de las glumas en Schoenoplectiella son glabros y los aquenios con frecuencia transversalmente rugosos. En general se trata de plantas anuales o perennes, con tallos trígonos o cilíndricos, hojas frecuentemente reducidas a vainas, inflorescencia pseudolateral (la bráctea inferior parece una prolongación del tallo) y aquenios frecuentemente con arrugas transversales y células epidérmicas más largas que anchas. Su área de distribución es subcosmopolita y se aceptan 62 especies, de las cuales solo cinco viven en el territorio, cuatro de ellas nativas.


Bibliografía
Luceño, M. & Jiménez-Mejías, P. (2008). Schoenoplectus in Castroviejo, S.; Luceño, M.; Galán, A.; Jiménez-Mejías, P.; Cabezas, F. & Medina, L. (eds.): Flora iberica 18: 42-59.

Larridon, I.; Zuntini, A.R.; Léveillé-Bourret, É; Barret, R.L.; Starr, J.R.; Muasya, A.M.; Villaverde, T.; Bauters, K.; Brewer, G.E.; Bruhl, J.J.; Costa, S.M.; Elliott, T.M.; Epitawalage, N.; Escudero, M.; Frailie, I.; Goetghebeur, P.; Hipp, A.L.; Jiménez-Mejías, P.; Kikuchi, I.A.B.S.; Luceño, M.; Márquez-Corro, J.I.; Martín-Bravo, S.; Maurini, O.; Pokorny, L.; Roalson, E.H.; Semmouri, I.; Simpson, D.A.; Spalink, D.; Thomas, W.W.; Wilson, K.L.; Xanthos, M.; Forest, F. & Baker, W.J. (2021). A new classification of Cyperaceae (Poales) supported by phylogenomic data. J. Syst. Evol. 59(4): 852-895.
Starr, J.R.; Jiménez-Mejías, P.; Zuntini, A.R.; Léveillé-Bourret, É; Semmouri, I.; Muasya, A.M.; Baker, W.J.; Brewer, G.E.; Epitawalage, N.; Fairlie, I.; Forest, F.; Kikuchi, I.A.B.S.; Pokorny, L. & Larridon, I. (2021). Targeted sequencing supports morphology and embryo features in resolving the classification of Cyperaceae tribe Fuireneae s.l. J. Syst. Evol. 59: 809-832.
CLAVE DE ESPECIES
1. Rizoma con entrenudos más o menos largos; aquenios con arrugas transversales poco marcadas; espiguillas frecuentemente pseudovivíparas ..................................... S. corymbosa
- Rizoma con entrenudos muy cortos; aquenios con arrugas transversales muy o escasamente marcadas; espiguillas nunca pseudovivíparas .............................................................................................. 2
2. Tallos agudamente trígonos ................................................................................................. S. mucronata
- Tallos cilíndricos, raramente (S. juncoides) de sección ligeramente angulosa 3
3. Aquenios de (1,9)2-2,5 mm de longitud, con arrugas transversales poco marcadas S. juncoides
- Aquenios de 1,2-1,6 mm de longitud, con arrugas transversales gruesas y muy marcadas 4
4. Todas las hojas insertas en la base de la planta; tallos sin flores basales S. supina
- Hoja superior inserta por encima de la base; tallos frecuentemente con flores en la base de la planta, escondidas entre las vainas foliares S. erecta
Schoenoplectiella corymbosa (Roth ex Roem. & Schult.)
J.R. Starr & Jim.Mejías
Perenne, con rizoma de entrenudos más o menos largos, aunque a veces pueden formar macollas. Tallos fértiles de (38)62-150 cm, cilíndricos, que se curvan hasta alcanzar el suelo en la fructificación, dando origen a nuevos individuos producidos por pseudoviviparismo. Hojas reducidas a vainas, las inferiores de color púrpura oscuro. Inflorescencia en antela pseudolateral, con entre 11 y 90 espiguillas, unas pocas sésiles y varios radios desiguales que rematan en un fascículo de 2-10 espiguillas; muy raramente la inflorescencia se reduce a un fascículo de espiguillas sésiles. Espiguillas de 4-8 mm de longitud, de contorno ovado u ovado-lanceolado, con (3)12-20(30) flores. Glumas de 2,7-3,5 mm de longitud, ovadas, rematadas en un mucrón de hasta 0,3 mm, pardas, hialinas hacia la base, con margen escarioso estrecho o sin él. Perianto ausente. 3 estambres. 3 estigmas. Aquenios de (1,3)1,4-2(2,2) mm de longitud, de contorno obovado o, más raramente, elíptico, obtusamente trígonos, con finas arrugas transversales de distribución irregular, de color pardo oscuro en la madurez, brillantes. 2n=desconocido. Bordes de lagunas de agua dulce sobre arenas o margas.
476 477
TRIBU PSEUDOSCHOENEAE
TRIBU PSEUDOSCHOENEAE 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Resérvaloen www.jolube.es
Schoenoplectiella corymbosa
Schoenoplectus lacustris (L.) Palla




No o laxamente cespitosa; entrenudos del rizoma de hasta 18 mm de diámetro. Tallos fértiles de (30)50-200(300) cm, cilíndricos. Hojas reducidas a vainas, a veces con lámina de hasta 30 cm de longitud. Inflorescencia terminal o pseudolateral, en antela simple o compuesta, con (5)2749(60) espiguillas dispuestas en glomérulos más o menos laxos. Espiguillas de (9)11-18(23) mm de longitud, ovoides u ovoide-lanceoladas, a veces subglobosas, con (12)18-36(40) flores. Glumas de (2,5)3-4,3(4,5) mm de longitud, ovadas, de ápice bífido, con un mucrón de (0,2)0,40,8(1,1) mm, lisas o con escasas papilas o acúleos pardo-rojizos hacia el ápice, especialmente las inferiores, pardo-rojizas, raramente hialinas, de bordes glabros o escasamente pelosos y margen escarioso o no. Cerdas periánticas 4-6(7), retrorsamente escábridas. 3 estambres. (2)3 estigmas. Aquenios de (1,7)2-2,8(3) mm de longitud, de contorno obovado o, con menos frecuencia, elíptico o subborbicular, obtusamente trígonos o, más raramente, planoconvexos, lisos, generalmente de color pardo oscuro en la madurez, mates o brillantes. 2n=38, 40, 42, 54, 76, 77, 80. Bordes de lagunas permanentes y cursos de agua remansados.

Resérvaloen www.jolube.es
492 493
3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
TRIBU SCHOENOPLECTEAE
Schoenoplectus lacustris
TRIBU SCHOENOPLECTEAE
Schoenoplectus lacustris
Anual, cespitosa. Tallos fértiles de (19)30-60(65) cm, agudamente trígonos, de caras cóncavas. Hojas de (2,6)3-5(6) mm de anchura, más o menos carenadas. Brácteas 2-3, la inferior más larga que la inflorescencia. Inflorescencia en antela simple o compuesta, más raramente capituliforme, con espiguillas dispuestas en glomérulos subesféricos, en número de hasta 50(60) por cada glomérulo. Espiguillas de (5,3)7-15 mm de longitud, de contorno lanceolado, con raquidio áptero y 12-30(40) flores. Glumas de 0,5-0,8 mm de longitud, anchamente obovadas o suborbiculares, obtusas o diminutamente mucronadas, de color púrpura oscuro, uninerviadas. 1(3) estambres. 3 estigmas. Aquenios de (0,5)0,6(0,7) mm de longitud, obovoides, trígonos, de color verde pálido o pajizos, finamente punteados. 2n=18, 26, 28, 32, 34, 36. Arenales húmedos, arrozales y suelos que se inundan temporalmente.


Observaciones: especie nativa de las regiones tropicales del globo, que se considera naturalizada en el sur de Europa y Macaronesia.

Resérvaloen www.jolube.es
526 527 SECCIÓN FUSCI Cyperus difformis L. 3600 2400 1800 1200 600 3000 pH Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
TRIBU CYPEREAE
Cyperus difformis
TRIBU CYPEREAE
Cyperus difformis
Apéndice I.
Glosario terminológico
A Abaxial

Dícese de la superficie más alejada del eje en un órgano de origen foliar. En las hojas es sinónimo de la cara inferior o envés.
Acidófilo
Con afinidad por los suelos ácidos.
Aclamídea
Flor que carece de perianto (cáliz y corola).
Acrescente
Órgano, frecuentemente el cáliz, que continúa creciendo durante el proceso de maduración.
Actinomorfa (flor)

Flor que posee simetría radiada, es decir, que tiene más de un plano de simetría.
Acúleo
Espinita corta y rígida.
Acuminado
Referido al ápice de un órgano foliar (hoja, pétalo, etc.) que se estrecha suavemente en una punta (acumen).
Adaxial
Resérvaloen www.jolube.es
Superficie de un órgano foliar más cercana al eje donde se inserta. En las hojas es sinónimo de la cara superior o haz foliar.
Adpreso
Adosado al eje.
Afilo
Tallo sin hojas o con ellas reducidas a vainas.
Ala
Prolongación membranosa de un órgano (tallo, utrículo, aquenio, etc.).
Alóctono
Dícese del taxon que no se considera nativo de un determinado territorio.
Androceo
Parte masculina de la flor formada por los estambres.
Andrógina (espiga)
Con flores masculinas en la parte superior y femeninas en la inferior.
Androginecandra (espiga)
Con flores masculinas en la parte superior, femeninas en la media y masculinas en la inferior.
Anemocoria
Dícese de la dispersión por el viento de propágulos: semillas y frutos, fragmentos de rizomas, etc.
Anfistomática
Hojas con estomas en ambas superficies (haz y envés).
Antela
Tipo de inflorescencia cimosa con aspecto de umbela, cuyos radios son frecuentemente desiguales.
Antelígula
Membrana hialina opuesta a la lígula.
Antera
Parte fértil de un estambre que contiene el polen.
Antrorso
Dirigido hacia delante.
Antrorso-escábrido
Tallo, rama, cerda periántica, etc. que presenta acúleos dirigidos hacia delante.
Apiculado/a
Acabado en una punta corta.
Aquenio
Tipo de futo seco indehiscente con una sola semilla más o menos soldada a la pared del fruto.
581
